| Главная | Неоцен |
Неоцен:
портрет Земли |
Эта страница – попытка нарисовать общий портрет нашей планеты через 25 миллионов лет с учётом нынешнего состояния и известных тенденций изменения планеты. Дан обзор географии, растительного и животного мира материков и островов.
Европа |
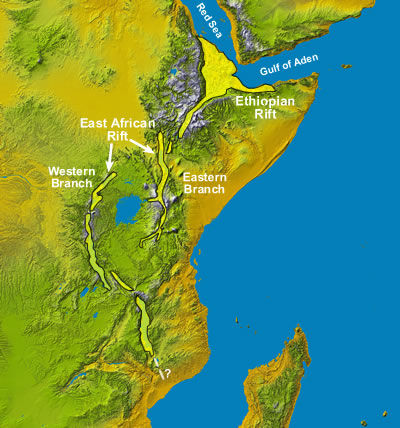
Территория Европы в неоцене по-прежнему испытывает влияние
со стороны Африки, которая на протяжении всего кайнозоя двигалась на север.
В неоцене движение
Африки полностью закрыло Гибралтарский пролив, служивший физической границей
между двумя материками, и последствия этого были поистине катастрофическими.
Самое главное из них – полное высыхание Средиземного моря. На севере Европы
постепенное поднятие северного края литосферной плиты вызвало обмеление и
отступление Северного моря. Британские острова соединились с материком, а Балтийское
море
превратилось в Венедское озеро – пресноводное и постепенно заполняющееся
отложениями, которые приносят реки.
В неоцене к югу от Альп, между Африкой и Европой, раскинулась пустынная котловина,
которая в эпоху человека была дном Средиземного моря. Здесь отложились пласты
солей, когда-то растворённых в воде, и на значительном протяжении Средиземноморская
котловина совершенно непригодна для жизни. Здесь высокие дневные температуры
сменяются ночным холодом, а недостаток пресной воды делает невозможной жизнь
растений и животных. В центре котловины разбросаны отдельные гипергалинные
озёра, а ближе к Альпам – обширные солёные болота, время от времени подпитываемые
дождями и реками, стекающими с Альп. Края Средиземноморской котловины представляют
собой полупустыню с отдельными оазисами, которые получают воду из рек. Практически
все реки, стекающие с южных склонов Альп, образуют дельты и превращаются
в сеть мелководных болот и озёр, чаще всего пересыхающих летом и осенью.
В Средиземноморской
котловине существуют также «оазисы», образованные вершинами гор, некогда
бывших островами Средиземного моря. Они существуют за счёт конденсации атмосферной
влаги на камнях из-за значительных суточных перепадов температуры.
К северу от горных массивов Альп климатические условия совершенно иные. Климат
Северной Европы в значительной степени определяют Северная Атлантика и Северный
Ледовитый океан. Северное побережье Европы представляет собой обширные равнины,
сложенные мягкими речными отложениями. Практически все реки текут на север,
образуя обширные дельты и болота, испытывающие влияние океанских приливов
и отливов.
Движение Африки на север и её медленное столкновение с Европой стало причиной
повышения тектонической активности в Альпах и соседних горных массивах. В
Южной Европе существует несколько действующих вулканов, регулярно извергающихся.
Вершины Альп покрыты ледниками, площадь которых меньше, чем в эпоху человека,
из-за общего потепления климата.
Практически все горные озёра Альп, существовавшие в эпоху человека, были
заполнены речными отложениями в течение нескольких тысяч лет и превратились
в болота,
ставшие затем лугами и лесами. Ледниковый период на рубеже голоцена и неоцена
оставил свои озёра, которые также исчезли в раннем неоцене. В альпийских
долинах есть небольшое количество крупных озёр, постепенно заполняющихся
речными отложениями.
Горные системы Балкан и Кавказа отличаются большим количеством пещер, образующих
в некоторых случаях протяжённые пещерные системы.
Скандинавия – ещё одно место в Европе, где можно встретить ледники. Они находятся
в горах на севере Скандинавии, и оттуда стекают короткие порожистые реки,
питающие Венедское озеро. Также в это озеро впадает часть равнинных рек северо-востока
Европы. Из Венедского озера вода течёт в Атлантику широким заболоченным потоком,
немного похожим на Эверглейдс (Флорида) из эпохи человека.
Центральная и северо-восточная Европа – это место, где господствует континентальный
климат и выражена смена времён года. Здесь тёплое лето, но снежные и морозные
зимы.
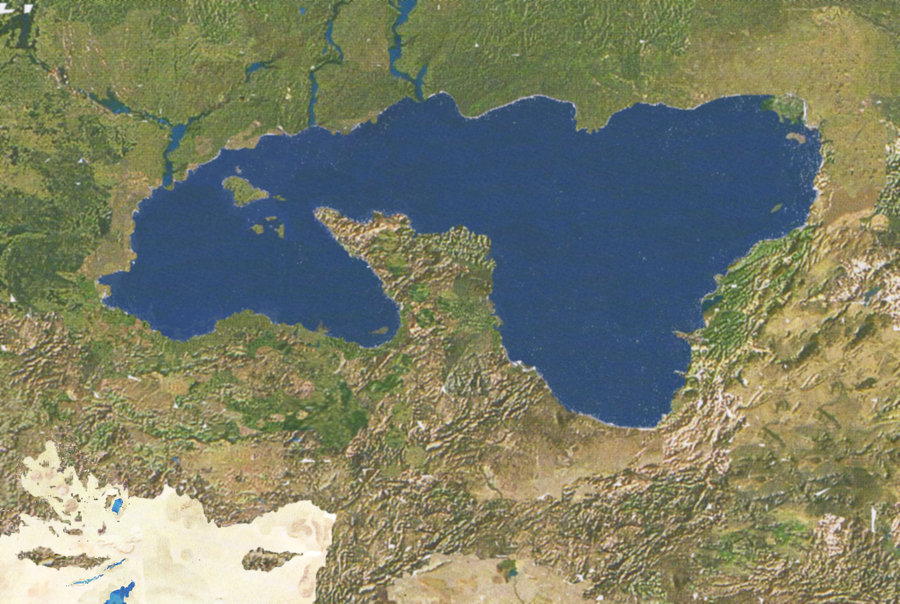
Карта Четвероморья.
На севере видно Трёхречье, на юго-западе находится Средиземноморская котловина
с
«оазисами» и гипергалинными озёрами. Рисунок Carlos Pizcueta (Electreel)
В Евразии есть один крупный внутриконтинентальный водоём – Четвероморье.
Оно образовалось после заполнения водой котловин Чёрного, Азовского, Каспийского
и Аральского морей. В ледниковую эпоху, через 5 миллионов лет после исчезновения
человечества, эти моря высохли, но позже вновь появились. Из-за горообразовательных
процессов проливы Босфор и Дарданеллы закрылись, и Четвероморье не имеет
связи
с океаном. Его уровень повысился, и в неоцене Крым представляет собой остров.
Восточная часть Четвероморья более мелководная; вблизи берегов имеется множество
отмелей и недолго существующих островов. Реки Средней Азии, впадающие в Четвероморье,
образуют дельты. Западная часть Четвероморья более глубокая, медленнее прогревается
летом, но также медленнее остывает зимой. Кавказ в неоцене представляет собой
крупный гористый полуостров, вдающийся в Четвероморье с юга.
На северо-востоке берега Четвероморья могут замерзать, западный и южный берега
никогда не замерзают. Четвероморье оказывает большое влияние на климат юго-востока
Европы, смягчая и увлажняя его. Местность вдоль северного берега Четвероморья
носит название Трёхречья: реки с равнинной части Восточной Европы текут по
древним руслам низовий Волги, Дона и Днепра.
Древние и сильно выветренные Уральские горы служат естественной границей
Европы на востоке. В ледниковую эпоху Полярный Урал был в значительной степени
сточен
ледниками и разрушен.
Климат на побережье Европы определяется влиянием холодного течения Антигольфстрим,
идущего из Северного Ледовитого океана на юг. Из-за него Гольфстрим сильно
отклонился на запад и климат на побережье Европы стал более влажным и прохладным.
По ночам на побережье обычны холодные туманы и обильная роса.
Климат неоцена в целом более влажный и тёплый, чем в эпоху
человека, а на юго-востоке Европы в котловинах четырёх морей образовалось Четвероморье
– гигантское озеро
с солоноватой водой. Вокруг этого водоёма условия стали более благоприятными
для роста растений – климат значительно более ровным и влажным. Растения
из лесов, окружающих Четвероморье с юга и запада, несут явные особенности
обитателей
влажных тропических лесов. У них широкие листья с вытянутыми кончиками,
образующими «капельницы». Но истинно тропические виды не проникали на
север
– им мешают
саванны Сахары, Альпы и горы Малой Азии, а также Средиземноморская котловина,
болота и солончаки которой резко неблагоприятны для роста деревьев. Поэтому
флора северных субтропиков сформировалась за счёт местных видов, которые
самостоятельно приобрели признаки адаптации к жизни в тёплом климате.
Большое количество эндемиков
растительного мира встречается на Кавказском полуострове, который вклинивается
в Четвероморье: здесь сказывается явление высотной поясности, а также
наличие большого количества изолированных местообитаний. Меньше их на острове
Крым,
который расположен сравнительно близко к материковой Европе.
Леса, особенно на Кавказском полуострове и на Балканах, относительно
молодые, многоярусные (некоторые виды европейской флоры ещё только «становятся»
деревьями), потому лесной полог «рыхлый», составлен из деревьев разной
высоты. Это позволяет
проникать достаточному количеству света в подлесок, где развивается флора
из тенелюбивых и влаголюбивых растений – мхов и папоротников. В среднем
ярусе леса процветают лианы – многолетние потомки плюща и винограда,
достигающие
гигантских размеров. Потомки плюща стали холодостойкими вечнозелёными
лианами с прочными кожистыми листьями и расселились дальше на север –
практически
по
всей Западной Европе. На севере они становятся более мелколистными, некоторые
виды с рассечёнными листьями. Южные потомки плюща – вечнозелёные деревянистые
лианы с толстым стволом, грубой корой и корнями-прицепками. Их стволы
часто ползут по земле, переплетаясь с корнями деревьев и делая участки
леса,
где
они растут, труднопроходимыми. Потомки винограда стали листопадными растениями
с крупными нежными листьями, зимой впадающими в состояние частичного
покоя. Они предпочитают влажные местности вблизи водоёмов. Для субтропических
лесов Европы (особенно на юго-востоке, по побережью Четвероморья) характерны
эпифитные
растения, представленые разными папоротниками и мелкими цветковыми растениями.
Рядом с ними на ветвях развиваются «эпифитные корзины» мхов и лишайников,
формирующих метровые «бороды», как лишайник уснея в эпоху человека.
На земле растут орхидеи – более крупные потомки видов эпохи человека,
клубневые и корневищные многолетники с крупными одиночными цветками (одиночный
цветок
орхидеи – это редуцированная кисть). Также на стволах и досковидных корнях
крупных деревьев, на высоте до двух метров над землёй, растут причудливые
лишайники и папоротники.
Особенной роскоши растительный мир достигает вблизи берегов Четвероморья.
Дальше вглубь Европы черты субтропической растительности постепенно стираются,
и флора
Центральной Европы (севернее Альп) более похожа на ту, что существовала
в эпоху человека. Основу европейских лесов составляют дуб, бук и каштан;
южнее
к ним
добавляется платан. Севернее распространяются мелколистные леса из берёзы
и тополей. Влажные местообитания у водоёмов занимает ива разных видов.
В субтропических
лесах также распространены потомки клёна. Севернее клён встречается лишь
как второстепенная порода в широколиственных лесах.
Своеобразный комплекс растительности сложился на северном побережье Западной
Европы. Здесь вдоль берега тянется обширная литоральная полоса, сменяемая
сетью солоноватых болот. Значительные пространства переувлажнённых земель
заняты
солеустойчивыми травами. Кроме того, из Венедского озера через бывшие
проливы Скагеррак и Каттегат течёт некое подобие флоридского Эверглейдса
– широко
разливающаяся и сильно заболоченная река, через которую озеро избавляется
от избытка воды.
На литорали Северного моря береговая линия очень изменчива, а в прибрежной
полосе постоянно возникают небольшие острова, намытые приливами. Древесная
растительность присутствует в таких районах в виде отдельных зарослей
влаголюбивых деревьев типа ивы. Другие деревья произрастают только на
возвышенностях,
где почва менее влажная и не ощущается влияние солёной морской воды.
Из-за холодного
Антигольфстрима лето на побережье прохладное и сырое, с моря постоянно
идут туманы.
Растительность Альп демонстрирует явление высотной поясности. По мере
подъёма в горы лиственные леса сменяются смешанными, а выше в горах распространены
хвойные леса. На вершинах гор господствуют альпийские луга, а у края
ледника
растительность похожа на тундровую.
Южнее Альп климат совершенно иной – ощущается жаркое и сухое дыхание
Северной Африки и Средиземноморской котловины. Здесь растения вынуждены
бороться
за жизнь на узкой полосе полупустынь между Альпами и солончаками Средиземноморья,
получая с южным ветром порции солёной пыли и совсем немного дождя с тучами,
что шли с Северного моря и смогли перевалить через Альпы.
Западная Европа (Гибралтарский перешеек) и области близ Средиземноморской
котловины (южнее Альп) покрыты сухими лесами и зарослями кустарников
(маквис). Здесь
господствуют лавровые, древовидные и кустарниковые миртовые, засухоустойчивые
виды дубов, оливковое дерево. Ближе к котловине Средиземноморья редколесья
и маквис сменяются полупустыней с отдельными участками солончаков, оставленных
отступавшим морем. Здесь произрастает лишь чахлая древесная растительность,
но появляются и местами господствуют солеустойчивые травы, не образующие
сплошного травяного покрова. Также для окраин Средиземноморской котловины
характерны
суккулентные растения, в том числе потомки завезённых кактусов. Весной
в области южнее Альп наступает период кратковременного, но обильного
увлажнения почвы,
когда на вершинах гор тают снеговые шапки и в полупустынные районы стекают
ручьи, образующие временные водоёмы. В это время начинается быстрый рост
клубневых и луковичных растений, которые большую часть года пребывают
под землёй в покоящемся
состоянии. За несколько недель они успевают отцвести, завязать семена
и накопить
запас питательных веществ, после чего их надземная часть отмирает.
На атлантическом побережье Европы на широте Гибралтарского перешейка
и Иберийского полуострова засушливый климат сочетается с обильными утренними
туманами,
и травянистые растения обзавелись приспособлениями для улавливания атмосферной
влаги.
Острова Средиземноморья превратились в разобщённые солёной пустыней «оазисы»,
питаемые водой, которая конденсируется на вершинах гор. Здесь существует
древесная растительность, под покровом которой процветает мир мхов и
папоротников. Растения
удерживают воду, как губка, и таким образом сами поддерживают собственное
существование по аналогии с тропическим лесом.
Север Европы (от Скандинавии до Полярного Урала) находится в области
чётко выраженного сезонного климата с холодной зимой, полярным днём и
полярной
ночью. Зимы снежные и часто морозные, поэтому преобладающий тип растительности
– хвойный
лес с отдельными вкраплениями холодостойких мелколиственных лесов. На
крайнем севере многолетняя мерзлота постепенно растаяла, дав возможность
роста
травам. Кратковременный период существования заболоченной тундры сменился
эпохой
северных лугов с преобладанием злаков и осок. Общее потепление в неоцене
на севере Европы
«сбивается» существованием холодного Антигольфстрима, который появился
после закрытия Берингова пролива, когда Северный Ледовитый океан стал
непроточным «ледяным мешком». Тем не менее, летнего тепла оказывается
достаточно для
обильного роста травянистых растений. Характерная тундровая растительность,
включающая
низкорослые стелющиеся деревья и вечнозелёные кустарнички, отступила
в болота
и к горным вершинам.
Южнее Северного полярного круга раскинулись хвойные и смешанные леса,
плавно сменяемые дубравами на юге. Для лесной зоны восточной Европы типичны
сфагновые
болота, особенно на севере. При движении с севера на юг болота становятся
более редкими, древесная растительность постепенно начинает тяготеть
к речным долинам,
а на водоразделах появляются участки кустарников и степной растительности.
На северных берегах Четвероморья распространена растительность степного
типа – злаки и другие травянистые растения, с небольшими вкраплениями
деревьев и кустарников в долинах рек. Ледники оставили нетронутыми долины,
образовавшиеся
в низовьях трёх рек – Волги, Днепра и Дона, и участки степей вдоль северного
берега Четвероморья получили общее название Трёхречье. Полоса степей
простирается вдоль берегов Четвероморья на восток, в Среднюю Азию. В
долинах рек и возле
стариц образуются отдельные островки древесной растительности, а в дельтах
рек образуются заросли деревьев и кустарников, способных выдерживать
затопление во время половодья. Климат в этих местах сохраняет сезонность:
прохладная
влажная
зима и сухое тёплое лето.
Солёность воды в Четвероморье невелика, и это позволяет болотным и водным
растениям обильно разрастаться. Берега Четвероморья во многих местах
поросли густыми
тростниками, а на глубине обильно разрастаются многолетние водные травы.
Животный мир Европы принадлежит к Голарктической фаунистической
области. Для этой части света характерен в целом низкий процент эндемичных
групп
животных. На западе Европа соединяется с Африкой через Гибралтарский
перешеек, и это
способствовало появлению в Западной Европе некоторых африканских
групп животных: крупных потомков даманов, массивных представителей виверровых,
а также мартышек
и страусов.
Эндемиками Европы являются сравнительно немногие группы млекопитающих.
Одна из них – кроличьи норцы, семейство роющих зайцеобразных, появившееся
уже
после исчезновения человечества. Очень своеобразным эндемиком Европы
является карликовый
выдротюлень – последний на Земле вид ластоногих, обитатель Венедского
озера. Ещё одна группа европейских эндемиков – крупные хищники, являющиеся
потомками
ежей. Крупные плотоядные ежи – это скорее «креодонты» на фоне более
продвинутых настоящих хищников, не слишком разнообразная ветвь реликтов
ранненеоценовой
радиации мелких млекопитающих. Активный хищник среднего размера,
добившийся некоторого успеха в борьбе за существование – европейский
ежеволк,
зверь размером с крупную собаку. В южноевропейском маквисе водится
его уменьшенный
аналог,
ежешакал – падальщик и охотник на мелких животных. Ещё один вид крупных
ежей – южноевропейский чагрин, крупный всеядный с уклоном в хищничество
вид из
лесов Южной Европы (Балканы, берега Четвероморья). Это приблизительный
аналог барсука,
но не имеющий постоянных жилищ и не впадающий в спячку.
Группой, характерной для Голарктики, являются зайцелопы, мелкие и
средних размеров травоядные млекопитающие, являющиеся экологическими
аналогами
оленей и мелких
антилоп, но произошедшие от зайцев эпохи голоцена. В областях, где
господствует континентальный климат, зайцелопы соседствуют с титанолагидами,
ещё одной
группой потомков зайцев, объединяющей крупные формы травоядных. Наряду
с такими специализированными
формами травоядных в Европе обитают настоящие зайцы и кролики, а
также многочисленные грызуны (белки, мыши и пр.). Среди грызунов
появилась группа плотоядных потомков сонь, конкурирующая с мелкими
хищными млекопитающими.
Южнее Альп, в маквисе по окраинам Средиземноморской котловины обитают
свистороги – представители азиатской по происхождению группы копытных,
потомки полорогих.
В «оазисах» Средиземноморской котловины млекопитающие представляют собой большую
редкость. Здесь нет крупных форм, и встречаются только представители грызунов
и насекомоядных. Грызуны в процессе эволюции независимо друг от друга вырабатывали
приспособления, позволяющие получать метаболическую воду, мелкие виды насекомоядных
просто слизывают росу или довольствуются влагой, получаемой из добычи. Кроме
того, вершины гор этих «оазисов», покрытые лесом или кустарником, являются своеобразными
«конденсаторами» влаги, и там изредка образуются небольшие водоёмы, чаще всего
в защищённых от солнца карстовых пещерах. Грызуны и насекомоядные «оазисов»
- сравнительно немногочисленные мелкие животные, уступающие по разнообразию
птицам и рептилиям.
В Европе встречаются крупные представители другой характерной группы
неоценовых травоядных – хоботковые свиньи. Напоминающие слонов и
тапиров, они питаются
преимущественно мягкой растительной пищей – листьями и травами, а
также кореньями, клубнями и плодами. В отличие от зайцелоп и титанолагид,
они предпочитают
более влажные местообитания, хотя некоторые их виды встречаются в
маквисе.
Семейство порциппул – это итог попытки свиней приспособиться к жизни
на открытых равнинах и питанию жёсткой травой – злаками и осоками.
Порциппулы родственны
хоботковым свиньям и населяют степи Трёхречья и открытые местности
к востоку
от Четвероморья. Некоторые их виды заходят в тайгу и предгорья горных
массивов Центральной Азии. В целом порциппулы являются эндемиками
умеренных и теплоумеренных
районов Голарктики, точнее, Палеарктики.
В Европе обитают также настоящие свиньи, но они встречаются реже,
чем в голоцене.
Эндемичных семейств хищников в Европе нет. По сравнению с эпохой
человека состав семейств почти не изменился, за исключением повсеместного
вымирания
медвежьих.
Мелкие и средние хищники представлены куньими и мелкими кошками (потомками
одичавшей домашней кошки). Это большей частью древесные хищники.
Крупные хищники Европы – это массивные представители куньих (север
Европы,
умеренные и полярные
районы), кошачьи и псовые (большая часть территории Европы), а также
крупные виверровые (запад и юг Европы). Наличие виверровых в европейской
фауне
является итогом соединения Африки с Европой – европейские виверровые
близки с североафриканским
видам. Представитель виверр азиатского происхождения, цибетоникс,
населяет степи на юго-востоке Европы, в Трёхречье. Потомки завезённого
в эпоху
человека североамериканского енота отчасти заменили медвежьих в неоценовой
европейской
фауне – это всеядные животные с уклоном в растительноядность. Самый
крупный вид среди них – ложная панда. Кроме того, несколько мелких
древесных
видов обитают в лесах Центральной Европы и в европейских субтропиках
по берегам
Четвероморья. Эти виды меньше отклонились от облика и образа жизни
своего предка, но реже
спускаются на землю.
Птицы Европы демонстрируют меньшую степень эндемизма по сравнению
с млекопитающими, что связано с их большими возможностями расселения
и отсутствием естественных
преград для этого. Семейство бегающих пастушковых, представленное
страусовым
пастушком в степях Трёхречья, общее для Европы и Азии.
Влияние Антигольфстрима привело
к расселению некоторых северных форм на юг. Для атлантического побережья
Европы характерны крупные нелетающие гагары – птицы арктического
происхождения.
Сухопутный мост между Евразией и Африкой способствовал появлению
в Западной Европе страусов. Также страусы заселили окраины Средиземноморской
котловины,
а особые карликовые формы страусов населяют некоторые «оазисы» Средиземноморской
котловины. Некоторые виды птиц приспособились к жизни среди соляных
равнин и болот. На солёных болотах обитают наземные нелетающие фламинго.
В остальном эндемизм европейских птиц проявляется на родовом уровне.
Фауна лесных птиц на уровне семейств остаётся примерно такой же,
как в эпоху
человека. Ряд характерных для голоцена групп птиц вымер ещё в эпоху
человека – это
благородные соколы, орлы, грифы, а также журавли. Крупные плотоядные
птицы представлены
разными видами орлиных воронов – представителей хищноврановых. Это
семейство птиц эндемично для Голарктики и распространено в холодных
и умеренных
областях Евразии и Северной Америки. Среди хищноврановых есть виды,
обитающие на
побережье Северного Ледовитого океана, в лесах умеренной зоны Евразии,
а также в степях
Трёхречья. Род пернатых хищников и падальщиков Tanatopterus распространён
по всему горному поясу вдоль Евразии – от востока до запада, образуя
непрерывную цепочку нечётко отграниченных друг от друга видов – соседние
виды связаны
промежуточными
формами. Виды из более тёплых и сухих районов имеют голую кожу на
передней части головы и шеи в отличие от обитателей Альп и Гималаев.
Куриные птицы представлены в Европе разнообразными потомками серой
куропатки, фазанов и перепелов. Европейские перепела выселились на
некоторые острова Атлантики, образовав там ряд эндемичных форм. Тетеревиные
как более холодолюбивые птицы встречаются в северных и горных районах Европы.
Это
преимущественно
потомки
белой куропатки
и рябчика.
Среди козодоев характерен крупный вид из степей юго-востока Европы
– мерлетта, короткоклювый козодой размером с орла, питающийся птицами
и летучими мышами.
Рептилии Европы по разнообразию на уровне семейств
остались примерно на том же уровне, что в эпоху человека. Большинство
видов рептилий
имеет широкий
ареал, на севере их разнообразие резко снижается, и до полярных
областей доходят лишь
единичные виды. Разнообразие рептилий резко возрастает в южных
районах Европы:
на Балканах и Апеннинах, по окраинам Средиземноморской котловины.
На юге и западе герпетофауна Европы испытывает явное переднеазиатское
и африканское
влияние.
Черепахи Европы не слишком разнообразны. Южные берега Четвероморья
населяет несколько видов водных черепах, и единичные виды встречаются
в Центральной
и Западной Европе. Северная граница распространения водных черепах
– Венедское озеро. Разнообразие сухопутных черепах очень велико
по окраинам
Средиземноморской
котловины. В «оазисах» встречается один крупный вид черепах и несколько
мелких. Также сухопутные черепахи обитают в субтропических лесах
Балкан.
На значительной территории Западной, Центральной, Восточной и Северной
Европы распространены представители семейства настоящих ящериц
(Lacertidae); в долинах
Альп они образуют несколько узкоареальных эндемичных видов. На
юге Европы распространены также представители семейства веретениц,
крупный
вид которых
водится в субтропических
лесах Балкан. В маквисе на южных склонах Альп встречаются представители
других семейств – гекконы, сцинки и агамы. Эти рептилии образуют
несколько эндемичных
видов в «оазисах» Средиземноморской котловины, а один вид сцинков
приспособился к жизни в гипергалинных водоёмах. Верховные хищники
в «оазисах» – ящерицы
и змеи: существа, способные расти большими, жить долго и иметь
умеренный аппетит,
что позволяет им образовать большую популяцию на скудных пищевых
ресурсах «оазисов». Агамы и гекконы обитают в субтропических лесах
по южному
и западному берегам
Четвероморья, и несколько видов агам обитает в степях Трёхречья.
Вараны обитают только на западе Европы и являются несомненными
переселенцами из Африки.
Среди змей имеется эндемичное семейство водноужовых, к которому
принадлежит атаргатис – постоянноводная неядовитая змея из Четвероморья.
Прочие
змеи представлены семействами гадюковых и ужеобразных. На Гибралтарском
перешейке
встречаются
представители аспидовых африканского происхождения.
Земноводные Европы представлены только хвостатыми
и бесхвостыми. Некоторые представители хвостатых земноводных, обитающие
в умеренных
областях
Восточной Европы, достигают очень крупных размеров, и их ближайшими
родственниками
являются гигантские виды из болот Западной Сибири. Большинство
хвостатых земноводных
Европы представлено мелкими формами – это тритоны и саламандры,
в неоцене более разнообразные, чем в эпоху человека. Бесхвостые
земноводные
представлены
настоящими
лягушками, жабами и жерлянками. Среди жерлянок встречается несколько
видов, обладающих очень сильным ядом. Реликтовые узкоареальные
виды жаб обитают
в некоторых «оазисах» Средиземноморской котловины, превратившись
в мелких жителей
карстовых расселин и роющие виды лесной подстилки. В пещерах Альп
существует несколько узкоареальных эндемичных видов. На крайнем
севере Европы
обитают только немногочисленные тритоны, жабы и настоящие лягушки.
Разнообразие рыб внутриконтинентальных водоёмов
Европы на уровне семейств также не отличается от такового в эпоху
человека. Значительно
сократилась
численность
и разнообразие осетровых – они остались только в Четвероморье,
но зато к ним относится один из крупнейших видов рыб Европы. В
пресных
водах
встречаются представители карповых и вьюновых рыб, а также бычки,
окунёвые и щуки.
В
ряду
крупнейших рыб Европы также стоят сомы, среди которых есть один
вид хищников, способный утащить под воду наземное животное весом
до 30
– 40 кг. Следом
эпохи человека является наличие в пресных водах Европы элеотрисовых
рыб, потомков
ротана, завезённого людьми. В неоцене в пресные воды Европы переселились
бельдюговые рыбы, которые смогли сделать это благодаря способности
к живорождению.
Среди проходных рыб Европы типичны горбылёвые, населяющие реки
Западной Европы. В литоральных болотах на побережье Северного моря
обитают
морские собачки
и бельдюги, приспособившиеся к переменной солёности и временному
пребыванию на
суше. В постоянных водоёмах, сообщающихся с морем, водятся камбалы,
нагуливающиеся в пресной воде и нерестящиеся в море.
Богатая ихтиофауна Средиземного моря исчезла вместе с морем, но
по северным окраинам Средиземноморской котловины есть несколько
реликтовых
солёных
озёр, существование которых поддерживается реками, стекающими с
Альп. Здесь обитают
настоящие морские рыбы – последние представители средиземноморской
фауны (карликовые горбыли, бычки и морские собачки).
Несколько узкоареальных видов рыб обитает в пещерах Балкан и «оазисов»
Средиземноморской котловины.
Среди беспозвоночных Европы примечательно развитие
бокоплавов в Четвероморье, где эти ракообразные представляют собой
пример адаптивной радиации,
дав целый ряд форм – от мелких планктонных фильтраторов до активных
хищников.
Также
в Четвероморье появилось несколько видов водных насекомых, постоянно
живущих вдали от берегов.
 |
 |
 |
Aepierinaceus tchagrin |
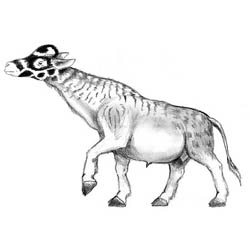
Embolohyrax brontotherioides |
Musticattus erminoides |
 |
 |
 |
Probosciaper mammutoides |
Tapirocephalus velox |
Ursicyon pseudoailuropoda |
 |
 |
 |
Eualle baltica |
Otiocrex struthiopus |
Subterraneotestudo talpina |
Северный Урал |
|
Земля Франца-Иосифа, Северный Ледовитый океан |
|
Атлантическое побережье Южной Европы |
|
Солёные болота Средиземноморской низменности |
|
Пещеры Балкан |
|
Солоноватое бессточное море-озеро в Юго-Восточной Европе |
|
Юго-Восточная Европа, северный берег Четвероморья |
|
Степи Южной Европы |
|
Степи Южной Европы (подземные животные) |
|
Венедское озеро (северо-запад Европы) - рыбы и ракообразные |
|
| Несколько минут солнца | Окраина Кипрского горного массива в Средиземноморской котловине; жители пустыни и подземного водоёма. |
| Последний вздох моря | Южная Европа, котловина Адриатического моря. Реликтовое солёное озеро и его обитатели - рыбы и беспозвоночные. |
| Охотники за росой | Средиземноморская котловина, мелкие животные и растения соляной пустыни. |
| Хибинское криволесье | Северная Европа, Хибины: звери и птицы северных лесов. |
| Северная Европа. Симбионты и паразиты сложного желудка крупного травоядного - лагоморопуса. | |
| Европейский маквис | Юго-западная Европа, окраины Средиземноморской
котловины. (автор - Bhut) |
Азия
|
Азия в неоцене по-прежнему представляет собой крупнейшую часть
света. Движение Африки на север оказывает меньшее влияние на географию Азии
по сравнению с Европой. Африканская литосферная плита своим движением на север
прижимает Аравийскую плиту к Иранской и препятствует раскрытию Красноморского
рифта. Итогом этого является высокая тектоническая активность в районе Аравии
и Ближнего Востока. Красное море потеряло связь с Индийским океаном и превратилось
в глубокую впадину, на дне которой отложились соли, когда-то растворённые в
воде. В него не впадало ни одной реки, и оно высохло за несколько веков.
Персидского залива в неоцене не существует, а на его месте поднялся молодой
и ещё невысокий горный хребет. Большие залежи нефти были исчерпаны в эпоху
человека, но тектоническая активность вызвала поднятие на поверхность глубинных
слоёв горных пород с остаточными залежами нефти, и в некоторых местах близ
Персидского хребта существуют естественные асфальтовые озёра, похожие на асфальтовые
озёра Калифорнии.
Огромная Индо-Австралийская литосферная плита продолжила движение на северо-восток,
оказывая давление на Китайскую литосферную плиту. Итогом этого стало продолжение
роста Гималаев и высокая вулканическая активность в цепи Больших Зондских островов.
Крупные острова постепенно объединились в узкую и длинную полосу суши – Берег
Джакарта.
Континентальная часть Азии составлена четырьмя литосферными плитами: Евроазиатской
(продолжающейся на восток за Уралом), Китайской (включающей Дальний Восток
и Юго-Восточную Азию), Индо-Австралийской (Индостан) и Североамериканской (Северо-Восточная
Азия – часть Восточной Сибири, Камчатка и Чукотка). Таяние ледников после голоцен-неоценового
оледенения вызвало поднятие северного края Евроазиатской плиты, что привело
к изменению течения рек и появлению обширных болот на равнинах Западной Сибири,
до Урала. В результате раскрытия Атлантического океана и движения Североамериканской
плиты на запад усилилась тектоническая активность в Северо-Восточной Азии,
и вместо Берингова пролива образовался Берингийский перешеек. Северо-Восточная
Азия в неоцене по-прежнему представляет собой горный район, отделённый от низменностей
Западной Сибири обширным плоскогорьем.
Дальний Восток в неоцене также является гористой местностью; основная область
тектонической активности здесь – прибрежные районы, место, где Тихоокеанская
литосферная плита уходит под край Китайской плиты. Видимый результат этого
– вулканическая активность на островах вдоль тихоокеанского побережья – от
Японии до Камчатки. Курильские острова в неоцене значительно увеличились по
площади и превратились в Большие Курилы. Они почти целиком отделили Охотское
море от Тихого океана – его с океаном связь поддерживается лишь через узкие
проливы между островами, составляющими Большие Курилы и Японию. Филиппинская
плита приблизилась к материку и образовалась связь между Филиппинским архипелагом
и материковой Азией через цепочку новообразовавшихся вулканических островов.
В Центральной Азии в неоцене продолжается раскрытие Байкальского рифта. Это
привело к расширению озера Байкал – глубочайшего и древнейшего озера на планете
в эпоху неоцена.
Азия, представляющая собой гигантский массив суши, является местом господства
континентального климата, несмотря на общее потепление в неоцене. На значительной
части Азии севернее Гималаев и горных массивов Ближнего Востока установился
чётко выраженный сезонный климат с холодной снежной зимой и жарким летом. Озеро
Байкал немного смягчает климат в своих окрестностях. На побережье Тихого океана
климат значительно мягче, чем в центре материка, но здесь часты дожди и ураганы.
В результате на побережье, особенно в областях с субтропическим климатом, часты
наводнения. Южное побережье Азии имеет муссонный климат, а Юго-Восточная Азия
(Берег Джакарта и острова, лежащие к востоку от него) находится в области экваториального
климата – обилие осадков и тепло круглый год.
Западная Сибирь представляет собой обширную заболоченную равнину, где в долинах
крупных рек образуется множество озёр и стариц. Со временем русла рек сильно
меняют направление течения, намывая новые берега и разрушая уже существующие.
В Восточной Сибири направление течения рек определяется главным образом особенностями
ландшафта, сложенного твёрдыми породами. Реки Западной Сибири достигают морей
бассейна Северного Ледовитого океана, разделяясь на множество рукавов, а восточносибирские
реки часто текут по узким ущельям и иногда достигают океана, низвергаясь в
него водопадами. В горной местности существует множество озёр, образовавшихся
при изменении течения рек. После отступления ледника в этих местах существовало
множество олиготрофных озёр с прозрачной водой и относительно бедной жизнью,
но в течение последующих тысячелетий они почти все пропали: заполнившись речными
отложениями, они превратились в болота и заросли лесом. На фоне недолго существующих
озёр ледникового происхождения величайшим исключением выглядит озеро Байкал.
За миллионы лет существования этого водоёма его глубина уменьшилась совсем
немного, зато ширина и длина увеличились.
На побережье Тихого океана (Камчатка, Большие Курилы, Япония) в горных долинах
существует большое количество геотермальных источников и даже небольших озёр,
подогреваемых подземным теплом.
Побережье Южной и Юго-Восточной Азии, а также островов Индонезии сложено сравнительно
рыхлыми породами, и реки в этих районах петляют по равнине, образуя множество
проток и стариц. В гористых районах Индокитая реки текут по горным долинам
и промыли за миллионы лет глубокие ущелья. В горных долинах существуют озёра,
и возраст некоторых из них насчитывает несколько миллионов лет.
Морские течения оказывают влияние преимущественно на климат тихоокеанского
побережья Азии. Тёплые южные течения значительно смягчают климат прибрежных
районов материка и способствуют продвижению теплолюбивых субтропических видов
на север. Закрытие Берингова пролива и значительный перепад температур между
Северным Ледовитым океаном и северной оконечностью Тихого океана приводит к
сильным северным ветрам, несущим в Берингию холод, а зимой ещё и значительное
количество снега.
Потепление, следующее за голоцен-неоценовым оледенением, вызвало
таяние многолетней мерзлоты на севере Евразии. Это привело к фактическому исчезновению
тундр и смене их растительными сообществами на основе злаков и осок, напоминающими
степи. Как и в Европе, растительность тундрового типа осталась только в горах,
у границы ледников.
По мере увеличения количества осадков при движении на юг северные приполярные
злаково-осоковые сообщества сменяются криволесьем из холодостойких лиственных
и хвойных пород, которые постепенно образуют обширный пояс северных лесов,
тянущийся от Западной Сибири через Берингию до востока Северной Америки. В
отличие от тайги эпохи голоцена, северные леса неоцена отличаются несколько
большим видовым разнообразием и не образуют «монокультурного» древостоя из-за
наличия в фауне Северной Азии крупных растительноядных млекопитающих, которые
прокладывают в хвойных лесах миграционные тропы, зарастающие травой, кустарниками
и лиственными деревьями, что способствует появлению разнообразных местообитаний
и обогащению видового состава лесной флоры.
Болота Западной Сибири – место обильного роста и особого развития болотной
флоры. Короткое, но жаркое и влажное лето способствовало эволюции крупных быстрорастущих
травянистых растений с многолетними клубнями и корневищами. Некоторые однодольные
из числа осоковых в условиях жаркого лета и обилия влаги развились в низкорослые
древовидные формы, напоминающие панданусы и драцены из областей тропического
климата, но листопадные. Также в болотах и реках Западной Сибири распространены
многолетние водные и болотные травы.
Центральная Азия находится в области ярко выраженного континентального климата,
и лишь в районе Байкала его особенности несколько смягчаются. Южнее пояса хвойных
и смешанных лесов лежит зона степной растительности, переходящая на юге в зону
полупустынь. Потепление и увлажнение климата в неоцене привело к тому, что
природные условия в пустынях Центральной Азии смягчились по сравнению с эпохой
человека. Реки с горных массивов Центральной Азии оживляют земли Центральной
Азии и способствуют росту обширных зарослей кустарников и невысоких лиственных
деревьев. Благодаря размножению корнеотпрысками и укореняющимся ветвям эти
растения хорошо переносят песчаные бури и другие стихийные бедствия, быстро
восстанавливаясь после них.
Горные массивы, тянущиеся через весь материк и соединяющиеся с горами Европы,
демонстрируют чёткую картину высотной поясности. На южных склонах гор встречаются
растения субтропиков и тропиков Южной Азии, сменяющиеся ближе к вершинам выходцами
с севера. Многочисленные горные долины, изолированные друг от друга, и хребты,
разделённые долинами, являются местами обитания разнообразных эндемичных растений,
преимущественно травянистых.
На Дальнем Востоке благодаря тёплым морским течениям лиственные леса субтропического
типа продвигаются дальше на север. Характерными растениями таких лесов являются
аралиевые, жизненные формы которых варьируют от многолетних трав до небольших
быстрорастущих деревьев. Леса Дальнего Востока образованы разнообразными представителями
преимущественно южных семейств, в том числе разнообразными видами ореха и каштана.
Также разнообразны виды дуба и клёна, отличающиеся крупными листьями причудливой
формы. Подлесок образован крупными широколистными травами и небольшими кустарниками.
На Японских островах обитает «дерево смерти» – один из крупнейших видов ядовитых
растений Земли. Родственные ему виды семейства сумаховых распространены в лесах
Дальнего Востока и образуют жизненные формы от кустарников до невысоких деревьев.
В горах распространены рододендроны, размером от невысоких кустарников до небольших
деревьев. Леса Японии, Больших Курил и южного берега Охотского моря – северный
край распространения эпифитных растений. Сообщества эпифитов представлены здесь
исключительно некрупными папоротниками и мхами, но южнее к ним прибавляются
орхидеи и другие типичные эпифиты, а папоротники становятся более разнообразными. Здесь
же распространены различные виды бамбука, от небольших растений высотой до
2 метров до гигантских, высотой около 10 метров.
Передняя Азия и области вокруг Средиземноморской котловины отличаются сухим
климатом со значительными суточными перепадами температуры. Здесь обитают различные
суккулентные растения, а также клубневые и корневищные эфемероиды, вегетирующие
лишь несколько недель в году. Древесная растительность представлена своеобразными
дубами с ползучим стволом, а на островах-«оазисах» Средиземноморской котловины
растут карликовые засухоустойчивые дубы. Для окраин Средиземноморской котловины
и Передней Азии характерны различные потомки кактусов опунций, завезённых человеком
в историческое время.
Восток Азии в области субтропического и тропического климата – место, где влажные
ветра с океана встречают барьер из гор и проливаются дождями. Равнины сложены
легко размываемыми осадочными породами, и реки часто меняют своё течение, разливаясь
и затапливая обширные пространства. Древесная растительность равнин способна
выдерживать затопления. В горах, где нет опасности наводнений, произрастают
теплолюбивые цитрусовые деревья и представители семейства чайных, а также фикусы
и пальмы.
Характерными представителями лесной флоры юга Азии являются разнообразные фикусы
и бобовые. Эти растения образуют широкий спектр жизненных форм – от травянистых
растений и кустарников до лиан и крупных деревьев. Кроме того, в этих лесах
встречаются пальмы и древовидные формы различных семейств: молочайные, сумаховые,
виноградные, сложноцветные и др. Травянистые растения представлены растениями
семейств имбирные, ароидные, орхидные, бегониевые и многими другими. Среди
бегоний в
неоцене появились
быстрорастущие бегониевые деревья – удачная попытка освоить новую жизненную
форму. В горных районах растёт бамбук разных видов, поднимающийся до высот,
где зимой ненадолго выпадает снег. В лесах на равнинах бамбук разрастается
на участках повреждённого леса, но постепенно вытесняется высокорослой древесной
растительностью.
По берегам Индийского океана в области тропического и экваториального климата
встречаются мангровые леса, а в устьях рек – заросли пандануса. Среди водных
растений сохранился и достаточно обычен лотос (возможно, благодаря культивированию
в эпоху человека), широко распространены кувшинковые, часто достигающие крупных
размеров.
Экваториальный климат лесов Берега Джакарта и островов Индонезии способствует
формированию настоящего влажного тропического леса. На уровне семейств многообразие
флоры осталось прежним, но значительное количество видов вымерло на рубеже
голоцена и неоцена, либо позже, вытесненное новыми растениями-поселенцами в
восстанавливающихся лесах. Характерными растениями тропического леса являются
гигантские травы, в том числе потомки банана, культивировавшегося в эпоху человека.
Среди травянистых растений богато представлены имбирные и ароидные, среди которых
есть многочисленные болотные и водные травы, а также клубневые травы. Ещё одно
характерное семейство растений юга и юго-востока Азии – непентовые, животноядные
растения со специализированными листьями-кувшинчиками. Среди них есть как некрупные
травянистые растения и эпифиты, так и большие лианы.
Фауна Азии принадлежит к двум зоогеографическим областям:
Голарктической и Восточной. Фауны практически не смешиваются на протяжении
почти всей Азии с запада на восток благодаря наличию горных массивов, разделяющих
материк вдоль. Лишь на Дальнем Востоке происходит смешение фаун и проникновение
северных видов на юг, а южных на север. Обитатели гор Центральной Азии принадлежат
преимущественно к выходцам из Голарктики, но здесь же встречается некоторое
количество видов южного происхождения.
Севернее горных массивов Центральной Азии складывается характерный комплекс
фауны Голарктической области. Типичными представителями фауны млекопитающих являются
разнообразные потомки свиней. В степях и редколесьях по южному краю лесной
зоны обитают порциппулы, более крупные, чем европейский степной вид.
К ним присоединяется крупный представитель хоботковых свиней – сибирская шурга.
Самой нетипичной группой свиней является малочисленное семейство хищных свиней,
представители которого являются чем-то вроде уменьшенных аналогов ископаемых
энтелодонов. Это падальщики и неспециализированные хищники в степях и редколесьях
Центральной Азии; один из видов обитает в лесах Дальнего Востока и расселяется
далеко на юг, до северных границ Восточной области. Для азиатского сектора
Голарктики типичны титанолагиды: здесь водится обда, крупнейший
вид
этой группы.
В районах
Крайнего
Севера водится
умингмак
– единственный
титанолагид, проникший в Новый Свет через Берингию. Также для лесных районов
Азии типичны зайцелопы, более крупные, чем виды из Европы и Африки. В лесах
Дальнего Востока обитает целый комплекс мелких видов зайцелоп, проникающих
на Японские острова. Также в Азии обитают типичные зайцеобразные – зайцы, кролики
и пищухи.
Полорогие в фауне Голарктики представлены немногочисленными специализированными
видами, такими, как сайгохения и лысый быкобаран из Центральной Азии. Видами
южного происхождения в фауне Дальнего Востока являются крупные олени тяжёлого
сложения – потомки оленя мунтжака. К их числу принадлежат оленебык из Китая
и шишигами с Японских островов.
Хищники азиатской Голарктики – преимущественно кошачьи, куньи и псовые. Среди
кошек имеются крупные формы, ведущие своё происхождение от одомашненных и впоследствии
одичавших кошек эпохи человека. Особняком среди них стоит массивный вид – сибирский
саблезуб, потомок рысей. Псовые представлены разнообразными потомками лисиц
и песцов. Некоторые из них занимают экологическую нишу волка охотятся на крупных
травоядных. Для азиатской Голарктики эндемично небольшое семейство медведеобразных
собак – потомков енотовидной собаки. В отрыве от основного ареала семейства
в степях близ Четвероморья обитает цибетоникс – бегающий вид виверровых. Среди
куньих есть как мелкие древесные и наземные формы, так и крупные наземные хищники
(берл) и всеядные формы тяжёлого сложения (арктомелес). Виверровые – нетипичные
обитатели Голарктики, разнообразные лишь на границе с Восточной областью. На
Дальнем Востоке встречается несколько видов всеядных древесных виверровых,
а южнее, на болотах Китая, появляются полуводные формы виверровых, всеядные
и рыбоядные, а также специализированные виды, поедающие ракообразных и моллюсков.
Приматы заходят в Голарктику лишь северным краем ареала. Представителями приматов
здесь являются крупные виды особого семейства парапонгид, аналоги человекообразных
обезьян. Кроме древесных видов, среди парапонгид есть горные, приспособленные
к наземному существованию в холодном климате. На Японских островах обитает
каппа – специализированный полуводный вид макак.
Озеро Байкал является
местом обитания реликтового вида китообразных, байкальского твердоклювого дельфина.
В озёрах
Китая обитает
второй
реликтовый
вид китообразных
неоцена, карликовый представитель морских свиней, потомок беспёрой морской
свиньи.
Грызуны азиатского сектора Голарктической области – это разнообразные беличьи
(белки, суслики, сурки), мыши и хомяки. В Средней Азии, по берегам Четвероморья,
на Балканах и в окрестностях Средиземноморской котловины обитают дикобразы.
В засушливых местностях многообразны тушканчики. Насекомоядные, как и в эпоху
человека, представлены ежами, кротами и землеройками. На Дальнем Востоке встречаются
представители семейства водяных землероек – относительно крупных животноядных
форм, по образу жизни сходных с выхухолью.
Среди птиц азиатского сектора Голарктики процент эндемичных групп невелик.
Характерно родственное пастушковым семейство журавельников, представленное
несколькими видами в умеренной области северного полушария. Семейство бегающих
пастушков, представленное страусовым пастушком, общее для Европы и Азии. Также
разнообразны собственно пастушковые птицы, образовавшие ряд видов в болотах
Западной Сибири. Их миграционные пути проходят через горные массивы к берегам
Индийского океана, а часть видов зимует на южном берегу Четвероморья.
Настоящие страусы обитают в Азии только в окрестностях Средиземноморской котловины.
На побережье Северного Ледовитого океана обитают представители своеобразной
группы цеторнитид, или олуш-китов – крайне специализированные веслоногие птицы,
являющиеся экологическими аналогами тюленей и мелких дельфинов. Они выводят
потомство преимущественно на островах, удалённых от берега. Эта группа птиц
распространена также на островах Атлантического океана.
Гусеобразные птицы разнообразны в северной и восточной частях Азии, и значительное
количество видов гусей и уток обитает на Крайнем Севере. В сухих степных районах
Центральной Азии водится дрофогусь – крупный травоядный вид птиц. Для субтропиков
Дальнего Востока характерны райские утки – птицы, отличающиеся яркостью оперения.
Эндемичное для Азии семейство тератанатид включает крупных нелетающих гусеобразных,
питающихся водорослями и высшими водными растениями. Северный вид семейства
встречается по всему северному побережью Тихого океана, а южный характерен
для Восточной области.
В лесной зоне Азии обычны голуби, но их наибольшее разнообразие приходится
на Дальний Восток. Здесь же проходит северная граница распространения фруктоедов
– особых голубеобразных птиц, больше характерных для тропиков Старого Света.
В лесах Дальнего Востока обитают крупные совы особого подсемейства грифовых
сов, одни из самых крупных летающих птиц, превышающие размерами орлов эпохи
человека. Прочие совы мало отличаются от видов, известных в эпоху человека.
Другая группа плотоядных птиц, орлиные вороны, широко распространена по лесам
Восточной Сибири и Дальнего Востока. Один из крупных видов обитает вдоль побережья
Северного Ледовитого океана. Лесные и горные виды нечётко отличаются друг от
друга, и особняком среди них стоит сравнительно мелкий и длинноногий вид, обитающий
на побережье Японских островов.
На уровне семейств многообразие воробьинообразных птиц азиатской Голарктики
осталось сравнимым с эпохой человека. Эндемичное для Азии семейство пилоклювов
– полуводные птицы, потомки врановых. На Дальнем Востоке обитает несколько
видов этих птиц, и некоторые из них заходят в горы Центральной Азии, а один
вид обитает на Японских островах. Эти птицы заходят южным краем ареала в Восточную
область.
Рептилии северной части Азии не слишком разнообразны из-за континентального
климата с холодной зимой, что препятствует расселению на север теплолюбивых
групп этих животных. На большей части азиатского сектора Голарктики черепахи
отсутствуют; они появляются лишь в полупустынях и кустарниковых зарослях Центральной
Азии. На Дальнем Востоке существует несколько видов водяных черепах. Сухопутные
черепахи, в том числе сравнительно крупные, населяют Юго-Западную Азию, области
аридного климата. Ящерицы в холодных и умеренно тёплых районах представлены
только семейством настоящих ящериц с немногими мелкими или среднеразмерными
видами. Южнее к ним прибавляются агамы и вараны. Варан-бегун из кустарников
Центральной Азии обладает способностью бегать на задних лапах. Юго-Западная
Азия обладает более богатой фауной рептилий. Эндемичной группой этого региона
являются пустынные попрыгушки – нетипичные агамиды, обладающие способностью
прыгать и лазать по растениям с помощью хорошо развитых задних лап. Здесь же
обитают гигантские растительноядные сцинки панголинии. Среди змей примечательна
очень крупная ядовитая змея иллуянка, многочисленны гадюки, аспиды и ужеобразные,
в том числе ядовитые.
Среди земноводных очень характерными являются трясинники – гигантские хвостатые
земноводные из болот Западной Сибири, имеющие родственников в умеренных областях
на востоке Европы. Хвостатые земноводные более разнообразны на Дальнем Востоке,
хотя и не достигают таких гигантских размеров, как в Западной Сибири. Разнообразны
бесхвостые земноводные – преимущественно жабы и настоящие лягушки. Среди лягушек
Сибири характерной группой являются карликовые лягушки-мошки, у которых взрослая
особь живёт только один сезон. На Байкале обитает постоянноводный вид настоящих
лягушек.
Рыбы северной и восточной Сибири не слишком разнообразны.
Среди них характерны карповые, окунёвые, щуки, лососевые и сиговые. Разнообразие
двух последних
групп значительно ниже, чем в эпоху голоцена – это итог антропогенного воздействия.
Характерны холодостойкие представители элеотрисовых – потомки ротана. Они достигают
большого разнообразия в озере Байкал, вытеснив группы, существовавшие в озере
в эпоху человека. В теплоумеренной зоне Азии встречаются сомы нескольких семейств
(настоящие сомы, косатковые, в горных районах багариевые).
Среди новых групп пресноводных рыб характерны колюшкообразные, часто достигающие
крупного размера, и перешедшие к жизни в пресной воде некрупные акулы.
Среди беспозвоночных богатством и обилием эндемиков отличается фауна Байкала.
Также в раннем неоцене произошёл переход в пресные воды некоторых групп десятиногих
ракообразных. На Дальнем Востоке разнообразны пресноводные десятиногие ракообразные
– речные раки, крабы и креветки. Среди насекомых многообразие на уровне семейств
осталось прежним, и видимым итогом общего потепления климата является появление
в фауне Дальнего Востока крупных видов, в том числе принадлежащих к тропическим
группам. В болотах Китая появились богомолы, приспособившиеся к полуводному
образу жизни.
Южнее горных массивов, на Индостане и в Индокитае, находится Восточная зоогеографическая
область.
В фауне млекопитающих заметны изменения по сравнению с Голарктикой. Для Восточной
области характерно разнообразие парнокопытных. Семейство громорогов, потомков
полорогих,
эндемично
для Евразии,
и большинство
его представителей
обитает в Восточной области. На Индостане обитает нандитерий – крупнейший вид
семейства. Мелкие и легко сложенные свистороги заходят в Голарктическую область:
они обитают в Передней Азии и проникают в маквис Европы. Потомки мелких азиатских
оленей мунтжаков превратились в крупные массивные быкообразные формы и населяют
сухие и заболоченные леса. Некоторые виды заходят далеко на север – до Японских
островов. Немногочисленное семейство оленьков населяет болота, лесные реки
и мангровые заросли в тропической зоне континента.
Для Восточной области характерны разнообразные узконосые обезьяны, образовавшие
здесь особое семейство парапонгид. Некоторые парапонгиды живут в Голарктической
области, но большинство их видов населяет тропики Азии. К их числу принадлежит
конг – крупнейший примат неоцена. Также в Восточной области распространены
другие обезьяны – потомки лангуров и макак эпохи человека. Человекообразные
обезьяны Азии – гиббоны и орангутанги – вымерли в эпоху человека или вскоре
после неё, в эпоху глобального экологического кризиса.
Хищники Восточной области – это преимущественно кошачьи и виверровые; псовые
и куньи менее разнообразны. Бегающие потомки мангустов обитают в горах Центральной
Азии. В тропических лесах и предгорьях живут массивные древесные и наземные
виверровые. Своеобразной попыткой конкуренции с хищниками является живущая
в лесах хищная кошачья землеройка – реликт ранненеоценовой радиации насекомоядных,
единичная попытка занять экологическую нишу мелких хищников. Среди насекомоядных
распространены лишь землеройки и ежи (отсутствуют на Берегу Джакарта). Среди
землероек примечательна крупная водная форма – фисония, обитающая в реках Индостана;
также в горных районах имеются представители семейства водяных землероек.
С эпохи человека сохранились панголины, среди которых появился один подземный
вид.
Среди околоводных птиц богато представлены аистообразные: различные виды аистов
и цапель. Среди гусеобразных птиц появились своеобразные фильтраторы – черпоклювы.
Эти птицы успешно расселились по тропикам Старого Света от Африки и Земли Зиндж
через тропическую Азию до Меганезии. Обычны и широко распространены утки разных
родов – от крупных до карликовых.
Среди лесных птиц разнообразны попугаи, относящиеся к числу настоящих попугаев.
На юго-востоке области, в месте контакта с Австралийской областью, появляются
мелкие виды какаду. В Восточной области обитают лорито, представляющие особое
семейство попугаев, сосущих нектар. Они эндемичны для островов Индонезии и
Берега Джакарта, представляя собой сильно уклонившееся семейство в отряде попугаеобразных,
конвергентно сходное с нектарницами и колибри. Стрижи обитают преимущественно
в горных районах, а в лесах их заменяют усатки – специализированные насекомоядные
потомки стрижей. Один из видов усаток проник в Голарктику и населяет горы Центральной
Азии.
Многообразие голубей сравнимо с таковым в эпоху человека; в неоцене в отряде
появилось новое семейство фруктоедов. Его представители – крупные курро, многочисленные
и разнообразные лесные плодоядные птицы тропиков Старого Света.
В Восточной области сохранились птицы отряда ястребообразных – ястребы и несколько
видов орлов. Один крупный орёл, перешедший к почти наземному образу жизни, встречается
на островах Индонезии и Берега Джакарта. Среди ястребов примечателен карликовый
вид, живущий в пологе леса. Другая группа плотоядных птиц Восточной области
– крупные потомки сорокопутов, ставшие аналогами соколов и ястребов. Орлиные
вороны в Восточной области отсутствуют.
В неоцене из-за раскола Африки облегчилась связь Восточной и Эфиопской областей.
Наземных нелетающих животных, общих для этих областей, в неоцене пока нет,
но среди птиц имеется семейство ежеголовых скворцов, обитающих на Индостане,
в Индокитае и на Земле Зиндж. Среди нектароядных птиц Восточной области характерно
эндемичное семейство цветоклюев. Из прочих воробьинообразных птиц характерны
примитивные рогоклювы и питты из числа непевчих. Певчие воробьинообразные представлены
вьюрками, трясогузками, мухоловками, синицами, ткачиковыми, астрильдовыми,
врановыми и многими другими.
Рептилии Восточной области значительно разнообразнее, чем
в Голарктике, что связано с более благоприятным климатом. Сухопутные черепахи
достигают большого
разнообразия и среди них часто встречаются крупные формы весом до 50 кг. В
реках обитают очень крупные водяные черепахи из числа пресноводных и мягкотелых,
также разнообразны мелкие виды. Кроме черепах, в реках живут представители
семейства водноварановых, общего с Африкой и Землёй Зиндж. Наземные вараны
также разнообразны, и в Юго-Восточной Азии среди них встречаются относительно
крупные виды. Многообразны гекконы, агамы, сцинки; настоящие ящерицы редки. Змеи
Восточной области представлены многочисленными семействами. Среди них имеются
слепуны, крупные питоны, многочисленные ужеобразные змеи. Среди ядовитых змей
встречаются гадюковые и многочисленные аспидовые. Некоторые змеи обитают в
пресноводных водоёмах – в озёрах и на равнинных участках рек.
Фауна земноводных отличается бедностью видового состава отряда
хвостатых (имеется несколько видов тритонов и саламандр в горах Центральной
Азии). Бесхвостые
земноводные отличаются высоким видовым разнообразием. Некоторые виды полностью
перешли к жизни в пологе леса и не нуждаются в водоёмах для размножения. Некоторые
островные виды жаб достигают крупных размеров, тогда как отдельные представители
квакш относятся к числу самых мелких земноводных. Есть роющие формы, постоянно
обитающие в лесной подстилке. Также очень характерной группой являются безногие
земноводные – роющие и полуводные червеобразные формы. Представители этого
консервативного отряда мало изменились с эпохи человека.
Среди пресноводных рыб характерны карповые, вьюновые и разнообразные
сомы (настоящие, косатковые, шильбовые, мешкожаберные, клариевые, в горных
районах также багариевые). Другой характерной группой пресноводных рыб являются
лабиринтовые; большинство их
видов мелкие,
но есть
несколько крупных.
Потомки
цихлид, служивших
объектами
рыбоводства в эпоху человека, также стали характерным компонентом ихтиофауны
Восточной области. Змееголовы могут достигать гигантских размеров, отчасти
замещая крокодилов в качестве верховных хищников речных экосистем. Большого
разнообразия достигают элеотрисовые и пресноводные бычки. Также в реках и озёрах
Восточной области обитают хоботнорылообразные рыбы, представленные роющими
бентофагами и хищниками-засадчиками, охотящимися на беспозвоночных и мелкую
рыбу. Потомки завезённой гуппи расселились по рекам тропиков Азии, дав начало
большому количеству
видов мелких живородящих рыб. Характерны сарганы, пресноводные иглобрюхи и
иглы-рыбы. В низовьях рек встречаются акулы, скаты, камбалы, терапоны и горбылёвые. В
мангровых зарослях обитают илистые прыгуны.
Фауна беспозвоночных разнообразна благодаря жаркому климату.
Насекомые разнообразны, среди них часто встречаются крупные виды. Необычны
водяные парусники – бабочки,
чьи
гусеницы
ведут водный
образ
жизни.
Эндемиками
Восточной области являются крупные наездники, паразитирующие на позвоночных
(в т. ч. на млекопитающих). Среди двукрылых характерны пиявкомухи, паразитирующие
на позвоночных, и инфекциомухи, питающиеся падалью. Среди десятиногих ракообразных
обычны пресноводные и сухопутные крабы; некоторые виды достигают
очень крупных
размеров.
Часть
пресноводных крабов перешла к древесному образу жизни. Креветки разнообразны,
большинство их видов мелкие; некоторые креветки встречаются в холодной воде
горных рек.
Характерными обитателями влажных тропических лесов являются крупные наземные
пиявки. Эти черви – хищники, питающиеся в том числе мелкими позвоночными.
 |
 |
 |
Cervibos robustus |
Heterolopa niveophila |
Arctotitanolagus umingmak |
 |
 |
 |
Nanditherium altissimus |
Ursivetta aromatica |
Nechlorus dendrobatoides |
 |
 |
 |
Ammobatrachus saltatus |
Miniranula crystallodermata |
Orchidocimex platypes |
| Дождевой лес Юго-Восточной Азии | |
| Обитатели Гималаев | |
| Монголия, кустарниковые заросли | |
| Остров в Индонезии, дождевой лес | |
| Равнинная часть Китая, болота | |
| Болота Западной Сибири | |
| Персидский хребет - горы на месте Персидского залива | |
| Остров Курильской гряды | |
| Горные тропические леса Южного Китая | |
| Озеро Байкал и его обитатели от мелководий до дна. | |
| Влажный тропический лес Юго-Восточной Азии и его обитатели - животные и растения. | |
| Восточная Сибирь. Представители мегафауны, их паразиты, симбионты и хищники. | |
| Каракумская дельта | Четвероморье, дельта среднеазиатской реки Узбой; жители дельты - птицы и их соседи. |
| Долина горячих озёр | Обитатели геотермальных водоёмов Камчатки, зимовка перелётных птиц. |
| Отшельники из джунглей | Обитатели джунглей Индостана - представители мегафауны и их окружения. |
| На склонах древних гор | Обитатели гор и долин Кавказского полуострова. |
| Приёмыш из Эдема | Степи Средней Азии к востоку от Четвероморья, животные и растения. |
| Большой и маленькие | Индостан - жители тропического леса, симбионты и паразиты нандитерия. |
| Жители царства холода | Мелкие обитатели тайги Восточной Сибири
- летучие мыши, птицы, рептилии и беспозвоночные. (автор - Bhut) |
| Зелёный отравитель и его свита | Япония, лиственный лес на склонах гор в области субтропического климата - растения и животные. |
| Чаша жизни | Мелкие обитатели лесов в южной зоне таёжного пояса Азии, район озера Байкал. |
Африка
|
В неоцене Африка составляет единое целое с Евразией. Постепенное
движение Африканской литосферной плиты на север началось ещё в раннем кайнозое,
и в итоге это привело к соединению Африки с Евразией и исчезновению древнего
моря Тетис. В эпоху человека ещё существовал его остаток, Средиземное море.
Но дальнейшее движение Африки вызвало закрытие Гибралтарского пролива в раннем
неоцене, и в итоге Средиземное море высохло, оставив после себя Средиземноморскую
котловину – пожалуй, самое негостеприимное место для жизни на неоценовой Земле.
Наличие огромного безводного массива суши во многом определяет климат северной
части Африки в неоцене.
Местность вдоль южного края Средиземноморской котловины представляет собой пустыню,
по которой разбросаны горные плато.
Дожди здесь бывают крайне редко – ветры с Атлантики редко доносят дождевые
тучи
так далеко вглубь суши, а высокие Альпы задерживают дождевые облака с Северного
моря.
Северную границу Африки отмечают горы Атлас. Они задерживают часть влаги, которую
несут ветры с Атлантики, поэтому в горах берут начало короткие реки, впадающие
в озёра и болота у подножия гор.
Крупнейшей рекой Северной Африки является Сахарский Нил. Эта река не такая полноводная,
как её великий предшественник эпохи человека: после великого раскола Африки истоки
Белого Нила остались на Земле Зиндж, и Сахарский Нил существует только за счёт
Голубого Нила и некоторых других притоков. Сахарский Нил – это река, петляющая
по Ливийской пустыне и сливающаяся на западе с рекой Нигер. Долина Сахарского
Нила сложена легко размывающимися породами, поэтому река много раз меняла своё
русло. На короткое время Сахарский Нил даже покидал своё нижнее течение, стекая
водопадом в Средиземноморскую котловину. Но в дальнейшем река пробивала новое
русло выше по течению и продолжала свой путь по Сахаре, широко разливаясь
и
распадаясь
на
множество
рукавов.
Сахарский Нил протекает между плато Эннеди и нагорьем Тибести, включает в своё
русло впадину озера Чад, где образует сеть болот и озёр, и продолжает течь на
запад,
сливаясь
с
Нигером. В сухой сезон часть болот и озёр в долине Сахарского Нила пересыхает.
Северо-восточная
часть Африки представляет собой гористую местность. Горы принимают на себя часть
осадков с океана, и в долинах существуют
короткие реки
и небольшие
озёра. Здесь же находятся истоки Сахарского Нила – Голубой Нил.
Западное побережье Африки находится в области влияния прохладного течения
и влажных ветров с Атлантики. Крупных рек здесь нет, но низменности вдоль
побережья
покрыты болотами и с нагорий по границе Сахары стекают короткие реки. Здесь
летние температуры не слишком высокие, а зима влажная и прохладная. Но такой
микроклимат складывается лишь на узкой прибрежной полосе. Наличие огромного
массива пустынь, быстро нагревающегося и быстро остывающего, приводит к резкому
чередованию высоких и низких температур: в начале дня нагрев воздуха над
Сахарой вызывает движение влажных воздушных масс с океана, несущих туманы
и дожди.
А ночью пустыня быстро остывает, и на побережье дует сухой и холодный ветер
из Сахары. Дальше вглубь материка климат становится более засушливым.
Восточное побережье Африки – это горная местность, результат раскола материка
в раннем неоцене. В эпоху человека здесь располагалась Великая Рифтовая долина,
в которой находились озёра Ньяса, Танганьика, Виктория и некоторые другие.
В неоцене озёра пропали, а их берега стали берегами пролива Танганьика. Дожди
проливаются на гористую местность, и значительная часть воды течёт на запад
– в долины Центральной Африки, где берёт своё начало большинство притоков
крупной реки Конго.
Центральная Африка представляет собой низменность, окружённую невысокими
горами. После отделения Земли Зиндж река Конго сохранила большую часть притоков
и является
самой полноводной рекой неоценовой Африки. Кроме того, несущие дождь ветры
с пролива Танганьика достигают её бассейна намного быстрее, чем ветры с Индийского
океана в эпоху человека. В Центральной Африке господствует довольно ровный,
влажный и жаркий экваториальный климат.
Южная часть Африки – относительно сухая гористая местность. Реки здесь короткие
и часто пересыхают, а озёра существуют сравнительно недолгое время. Южнее
экватора крупнейшей рекой является Замбези. Горы небольшой высоты, тянущиеся
вдоль побережья,
отделяют внутренние районы материка от влажных ветров с океана, и внутренние
районы Южной Африки представляют собой саванну и полупустыню. В некоторых
горных долинах образуются небольшие озёра и болота. Климат этих мест сезонный,
с прохладной
зимой, короткой дождливой весной и жаркими летом и осенью. Пустыня Намиб
осталась бесплодной местностью; условия в ней стали ещё суровее из-за смещения
Антарктиды
в сторону Меганезии и ослабления холодного Бенгельского течения. В результате
контраст температур между морем и сушей стал меньше, и количество влаги,
приносимой туманами, уменьшилось.
Типы растительности Африки находятся в прямой зависимости
от климатических условий. Север материка – это край Средиземноморской котловины,
крайне засушливая местность с резкими суточными перепадами температур. Здесь
растительность крайне скудная: деревья отсутствуют, а травянистые растения
представлены засухоустойчивыми многолетними травами. Также в Северной Африке
встречаются многочисленные виды луковичных эфемероидов из семейств луковых,
лилейных и амариллисовых. Деятельность человека оставила во флоре Африки свой
след в виде наличия кактусов – потомков опунции, завезённой человеком. В пустынях
на севере материка кактусы представляют собой невысокие травянистые растения.
Северо-запад Африки богаче растительностью, чем север. Это область Гибралтарского
перешейка и гор Атлас. Здесь осадки, приносимые с океана, способствуют росту
кустарников и засухоустойчивых деревьев – дубов и лавров. В горах Атлас растут
сосны, близкородственные европейским, а во флоре Африки и Западной Европы существует
много общих видов растений.
Вдоль атлантического побережья Африки южнее Гибралтарского перешейка складывается
особый тип растительности, обладающей способностью впитывать и запасать большое
количество влаги из туманов, приносимых с океана. Основу растительных сообществ
здесь составляют толстянковые и некрупные суккулентные молочаи, внешне напоминающие
кактусы.
На северо-востоке Африки складываются сообщества засухоустойчивых горных растений.
Виды растений этих мест близки к аравийским и ближневосточным. В ближайших
к океану районах растут пальмы и акации, а также другие засухоустойчивые деревья
(в частности, из семейства анакардиевых), образующие редколесья. Полоса густых
лесов существует лишь на побережье, где
проливается большая часть дождей, и в долинах гор Эфиопии. Здесь же произрастает
несколько видов эвкалиптов, происходящих от завезённых в эпоху человека растений.
В местах с отлогими берегами растут мангровые леса, но их площадь очень мала
по сравнению
с лесами
Южной Азии, поскольку восточный берег Африки большей частью скалистый, а крупные
реки встречаются лишь южнее экватора (Замбези).
Южнее пустынь Северной Африки тянется область с умеренным количеством осадков,
позволяющих расти засухоустойчивым деревьям. Главнейшей водной артерией этих
мест является Сахарский Нил – крупная река, берущая начало в горах Эфиопии.
Вокруг реки складывается комплекс влаголюбивых болотных растений – папируса
(не родственного виду эпохи человека), рогоза и крупных тростников. Также разнообразны
крупные виды наземных трав, произрастающие в местах с достаточным увлажнением
и образующие густые заросли на месте упавших деревьев. На островках среди русла
реки произрастают влаголюбивые деревья, образующие небольшие рощи. В мелководных
заводях развиваются разнообразные виды кувшинок, апоногетонов, рдестов и других
водных растений.
Вдали от вод Сахарского Нила господствует растительность саваннового типа с
преобладанием многолетних злаков и отдельными группами деревьев, в том числе
сбрасывающих листву в сухой сезон. Один из видов деревьев саванн Северной Африки
– сахарное дерево, потомок баобаба. Также широко распространены древовидные
суккулентные молочаи, имеющие характерную канделябровидную форму кроны.
Берега Гвинейского залива – это область, где начинаются сплошные вечнозелёные
леса. На северном берегу залива их составляют породы деревьев, приспособленные
к сезонным изменениям количества осадков, а ближе к экватору их сменяют виды,
типичные для влажного тропического леса.
Центральная Африка (бассейн реки Конго) – это царство влажных тропических лесов,
болот и озёр. Лесная флора Центральной Африки сохранилась несколько лучше,
чем в Азии и Новом Свете, поскольку страны этого региона в эпоху человека были
отсталыми в экономическом отношении и промышленность не смогла нанести столько
крупного ущерба природе, как в других местах. Кроме того, сведению лесов препятствовал
нездоровый климат этих мест, благоприятный для развития и распространения заболеваний.
Поэтому тропические леса Центральной Африки демонстрируют большую сохранность
и сходство с природными сообществами эпохи голоцена. Среди деревьев африканских
тропических лесов характерны гигантские фикусы, а также очень крупные пальмы
с перистыми листьями длиной более 10 метров. В лесах встречаются потомки растений,
культивировавшихся в эпоху человека: дерева какао, гевеи каучуконосной, некоторых
тропических плодовых деревьев и кофейного дерева. Среди травянистых растений
широко распространены потомки культурного банана и его гибридов с дикими видами
банана. Лианы представлены крупными видами семейства ароидных, часто с причудливо
изрезанными лопастными листьями. Очень характерны растения семейства диоскорейных,
образующие крупный наземный каудекс, покрытый опробковевшей корой, с длинными
однолетними травянистыми стеблями, поднимающимися по соседним деревьям. В горных
районах встречаются бамбуковые заросли, а во влажных тенистых долинах растут
древовидные папоротники. Эпифитные растения характерны для верхних ярусов влажного
тропического леса. Это папоротники, плауновидные (селагинеллы) и мхи, а также
орхидеи и ароидные.
На побережье Гвинейского залива растут мангровые леса. Их массивы особенно
велики по берегам реки Конго.
Южнее области влажных тропических лесов располагаются территории летнезелёных
лесов и тропических саванн. В южном полушарии они занимают меньшую площадь,
чем в северном, из-за характера местности: значительная часть юга Африки представляет
собой сухую гористую местность. Древесная растительность распространена здесь
локально, в долинах рек и возле более-менее долго существующих озёр. Калахари
в неоцене по-прежнему является пустыней, но получает с дождями немного больше
воды, чем раньше, поэтому локальные водоёмы существуют здесь значительную часть
года. Для этой местности характерна сезонная пустынная растительность, преимущественно
злаки. Редколесья здесь образованы растениями семейства бобовых – акациями
и родственными видами, способными расти на бедных почвах благодаря клубеньковым
бактериям на корнях.
Территория пустыни Намиб сократилась из-за расширения полосы саванн на севере.
Уменьшение количества влаги, поступающей с туманами, привело к оскудению пустынной
флоры и вымиранию более требовательных к влаге видов. Аизооновые Южной Африки
имеют разнообразный видовой состав и образуют жизненные формы от суккулентных
трав до невысоких полукустарников. В пустыне Намиб выживают только суперсуккулентные
виды, отличающиеся медленным ростом и высокими способностями к запасанию и
сохранению влаги. Истинное количество растений в пустыне Намиб можно оценить
лишь после сильных дождей, примерно раз в два-три года. Тогда все растения
дружно зацветают, и на однообразном фоне пустыни появляются яркие пятна – куртины
ранее незаметных растений раскрывают цветки броской расцветки, заметной издалека.
Южная Африка – это сухая гористая местность, а также пустыня Карру. Доминирующим
типом растительности здесь являются редколесья из древовидных видов алоэ, как
в эпоху человека. Благодаря промышленному культивированию часть видов алоэ
получила возможность выжить, и в неоцене их потомки восстановили утраченное
видовое разнообразие. Некоторые виды этих растений похожи на американскую агаву.
Также распространены другие суккулентные растения семейства лилейных. Аизооновые
горных районов представляют собой кустарниковые растения с толстыми, иногда
даже шарообразными листьями, покрытыми желёзками и воском. На восточном побережье
Южной Африки встречаются потомки культурного ананаса.
Крайний юг Африки занимает особая Капская флористическая область. Благодаря
влиянию океана климат здесь более ровный и влажный, хотя и прохладный по сравнению
с пустынными районами к северу. Протейные и древовидные вересковые образуют
леса и кустарниковые заросли по склонам Капских гор вдоль побережья Африки.
Из травянистых растений характерны различные виды многолетних гераней, в том
числе полукустарниковых, и различные виды аспарагуса (спаржи). В пустынных
и полупустынных районах богато представлены многолетние эфемероидные растения
– ирисовые, амариллисовые
и
лилейные.
Хвойные растения Южной Африки – виды сосны, потомки завезённых в эпоху человека.
Они произрастают главным образом в горных районах. Ещё один род завезённых
растений – эвкалипты.
Животный мир Африки почти целиком принадлежит к Эфиопской
зоогеографической области, кроме севера материка, который входит в Голарктическую
область. В неоцене связь между Африкой и Европой усилилась после образования
Гибралтарского перешейка, что позволило представителям разных групп животных
свободно мигрировать между материками; своеобразие африканской фауны сохраняется
на уровне семейств. Ряд африканских групп животных проник в Западную Европу,
и в целом африканских мигрантов в Европе больше, чем европейских в Африке.
Хищные млекопитающие Африки представлены разнообразными видами кошачьих и виверровых,
другие семейства представлены беднее. Виверровые распространены по всему материку
и образуют наземные и древесные формы, рацион которых варьирует от исключительно
мясного до смешанного с преобладанием растительной пищи. Один из крупных хищников
северной части Африки – генетта-убийца, саблезубое животное тяжёлого сложения,
приспособленное к охоте на крупную добычу. По соседству с ней обитают более
мелкие генетты-пантеры, являющиеся экологическими аналогами леопардов. В тропических
лесах возрастает количество древесных виверровых, а один вид обладает летательной
перепонкой и населяет вершины деревьев. В Южной Африке обитают всеядные наземные
виверровые, способные питаться падалью и листьями суккулентных растений, а
также пардиния – вид, приспособленный к преследованию быстроногой добычи. В
Северной Африке встречается эндемичное семейство муравьятников, происходящее
от виверровых (потомки мангуст). Эти животные поедают насекомых и примечательны
своей способностью бегать на двух ногах.
Южная Африка – дом для последнего на Земле вида гиен, марафила устрашающего.
Это один из крупнейших видов наземных хищных зверей, питающийся крупной добычей.
Самцы и самки марафила отличаются ярко выраженным половым диморфизмом.
Кошачьи Африки представлены потомками мелких видов кошек (последний вид крупных
кошек, нунда, обитает на соседней Земле Зиндж). Независимо друг от друга некоторые
из них увеличились в размерах, но такие виды немногочисленны. В саваннах Северной
Африки живёт кошка-герцог, один из таких видов. Большинство мелких кошек обитает
в лесах Центральной Африки, в горах Восточной и Южной Африки.
Куньи Африки представлены немногочисленными видами, обитающими в горах Атлас
и распространёнными по западному побережью материка. В реках Западной Африки,
в том числе в бассейне Конго, водится несколько видов полуводных куньих.
Семейство псовых представлено разнообразными лисицами, особенно многочисленными
в Южной Африке. Эти животные охотятся на мелких млекопитающих, но есть всеядные
и насекомоядные виды. К псовым относится чути – крохотный вид лисиц, обитающий
в пустынных районах Северной Африки и питающийся насекомыми.
Насекомоядные Африки довольно разнообразны, а некоторые их представители достигают
крупных размеров. Большинство африканских насекомоядных – различные виды землероек,
а также ежи типичного облика, которые живут по всему материку. Из средиземноморского
маквиса в предгорья Атласских гор заходит ежешакал – представитель своеобразной
европейской группы хищных ежей. Центральная Африка – родина эндемичного семейства
червеязычников. Эти массивные потомки землероек являются специализированными
аналогами муравьедов, а один вид обитает в воде, хорошо плавает и питается
водными беспозвоночными.
Африканские рукокрылые разнообразны и населяют практически весь материк, кроме
самых засушливых районов Средиземноморской котловины. Среди них имеются насекомоядные
и растительноядные формы, а также несколько хищников, питающихся мелкими позвоночными.
Самым явным признаком влияния Голарктики на африканскую фауну является наличие
в северной половине материка разнообразных видов зайцелоп. Это животные открытых
пространств, предпочитающие сухой климат. Среди них есть пустынные виды, способные
долго обходиться без воды, а большинство видов – степные травоядные. На западном
побережье обитают листоядные формы зайцелоп, предпочитающие мозаичный ландшафт:
открытую местность с отдельными группами деревьев и кустарников. Элементом
влияния фауны Восточной области являются свистороги – своеобразные парнокопытные,
потомки полорогих, обитающие в полупустынях по окраинам Средиземноморской котловины.
Южнее экватора водится немногочисленное семейство высокорослых листоядных парнокопытных
– костнорогие, похожие на примитивных жирафов. Это также потомки полорогих,
эндемичные для Африки.
Наиболее характерными обитателями саванн севера Африки являются представители
отряда даманов. В неоцене потомки мелких зверей сильно увеличились в размерах
и стали более разнообразными по образу жизни. Саванну населяют крупные четвероногие
травоядные – плоскороги и эмболохираксы. «Полудвуногие» ндипинотерии предпочитают
редколесья и встречаются в южной полосе саванн, а наиболее примитивный вид,
ксенондипина, вовсе населяет заболоченные леса Центральной Африки и умеет лазать
по деревьям. Полуводные потомки даманов – ипопо: тростниковый ипопо населяет
долины рек, а морской ипопо селится в мангровых лесах и может поедать морские
травы. Попыткой даманов стать аналогами лошадей и антилоп является семейство
гираколоп, эндемичных для севера и востока Африки.
Среди копытных свиньи составляют конкуренцию гигантским даманам. В болотах
и реках севера Африки обитает свино-бегемот – потомок кистеухой свиньи. В южной
части Африки распространена наземная группа потомков кистеухой свиньи – свинорогие,
отличающиеся крупными размерами, тяжёлым телосложением (напоминают быков или
лошадей-тяжеловозов) и крупными костными выростами на морде. Свинорогие распространены
и на Земле Зиндж, но собственно африканские формы обладают одним непарным рогом
на морде, а у видов с Земли Зиндж рога более тонкие и расположены симметричной
парой.
Группой, получившей в неоцене возможности для эволюции, являются оленьки. Несколько
видов этих копытных обитает в тропических лесах Центральной Африки, и один
крупный вид оленьков обитает в долине Сахарского Нила. В южной части Африки
появилось происходящее от них особое семейство бегающих оленеобразных копытных
– ложноолени, отличающиеся разветвлёнными неопадающими рогами, представляющими
собой костные выросты черепа.
Грызуны Африки столь же разнообразны, как в эпоху человека. Очень распространены
представители семейства беличьих: они населяют все лесные местности материка,
в том числе влажный тропический лес Центральной Африки. В Южной Африке беличьи
представлены в основном наземными формами, но на крайнем юге, в Капской флористической
области, большинство их видов – древесные. Суслики встречаются только на северо-западе
Африки, в районе Атласских гор. Мыши и крысы распространены по всему материку,
в тропических лесах много древесных форм. Среди крыс имеются крупные лесные
виды, питающиеся твёрдыми орехами. Эти грызуны разнообразны в пустынях юга
Африки, а также в горных районах на востоке и северо-востоке. В саваннах и
редколесьях обитают дикобразы, похожие на виды эпохи голоцена, а в пустынных
районах обитают тушканчики. Очень крупные грызуны (крысы-траворезы и кенгуровые
прыгуны) встречаются в саваннах Северной Африки, где сложился своеобразный
комплекс «новых травоядных», крупных растительноядных млекопитающих из новых
систематических групп, появившихся в неоцене. Грызуны юга Африки более консервативны.
В видовом составе приматов также произошли изменения. Человекообразные обезьяны
вымерли в эпоху человека или вскоре после неё, и их место в экосистемах Африки
занимают павианы. Среди павианов появились лесные формы, несколько некрупных
древесных видов тропического леса, а также формы с уклоном в плотоядность.
Представитель последних, павиан-фурия, обладает интеллектом, приближающимся
к уровню человекообразных обезьян, и обладает зачатками орудийной деятельности.
Мартышки расселились дальше на север, в Западную Европу и на Гибралтарский
перешеек. Они ведут большей частью древесный или наземно-древесный образ жизни
и обладают хорошо заметными участками яркоокрашенной безволосой кожи на голове
и различными волосяными «украшениями». Среди мартышек есть специализированные
древесные формы, например, карликовые беличьи мартышки, которые обитают в тропическом
лесу в кронах деревьев первой величины.
В Африке также встречаются представители других отрядов млекопитающих: панголины
и прыгунчики.
Фауна птиц Африки в эпоху неоцена отличается многообразием, на материке имеется
несколько эндемичных групп.
Страусы были одними из самых крупных животных, выживших на рубеже голоцена
и неоцена. Возможно, они выжили благодаря тому, что люди разводили их искусственно
и расселили там, где в дикой природе они вымерли. Неоценовые страусы достигают
гигантских размеров – страус-жираф из саванн Северной Африки является крупнейшей
птицей Земли. Другие страусы немного меньшего размера обитают в зоне полупустынь
вокруг Средиземноморской котловины – это однопалый страус и целый комплекс
родственных карликовых видов в «оазисах» Средиземноморской котловины.
Гусеобразные птицы распространены по всей Африке, встречаясь в местах, где
имеются постоянные или временные водоёмы. Это разнообразные виды уток, различающиеся
рационом, и преимущественно растительноядные гуси. Среди уток есть виды, фильтрующие
микроводоросли в поверхностных слоях воды. Прочие водные птицы представлены
разнообразными видами цапель, пастушковыми, болотными курочками. Некоторые
аисты достигают очень большого размера. Также для пресных водоёмов и морских
побережий типичны чайки и крачки, а на крайнем юге Африки, на побережье, обитают
колонии нелетающих чаек-пингвинов. Эти птицы стали экологическими аналогами
пингвинов эпохи голоцена, которые в неоцене практически полностью вымерли.
Кулики распространены по побережьям пресных и солоноватых водоёмов.
Куриные птицы представлены исключительно семействами цесарок и фазановых. Фазановые
включают потомков перепелов и франколинов, но среди них есть также потомки
примитивных пород домашних кур, одичавших в эпоху заката человечества. Потомки
кур населяют саванны, но встречаются также в лесах Центральной Африки. Цесарковые
населяют сухие пространства южной и восточной частей Африки, предпочитая саванну
с отдельными участками кустарниковой растительности и деревьями.
Среди голубеобразных птиц широко распространены голуби, а в тропических лесах
Центральной Африки и по побережью Гвинейского залива живут представители семейства
фруктоедов – несколько видов курро.
Попугаи неоценовой Африки происходят от сравнительно немногочисленных предков,
в том числе от попугаев-неразлучников и некоторых длиннохвостых попугаев. В
саваннах Северной Африки среди попугаев имеются плотоядные виды, способные
разгрызать кости и являющиеся своеобразными аналогами грифов. Кроме них, в
лесистых местностях обитает много видов, ведущих более типичный образ жизни.
Один из таких видов – попугай-бортник, приспособившийся к питанию жалящими
перепончатокрылыми.
Среди дятлообразных появилась целая группа плотоядных видов, к которой относятся
самые крупные дятлы Африки – большой грифовый дятел и его родственники. Один
из таких дятлов приспособился к питанию черепахами, панцири которых разбивает
ударами клюва. Наряду с этими нетипичными дятлами есть большое количество более
мелких видов, питающихся насекомыми. Дятлы распространены преимущественно в
лесистых местностях, но среди них есть также виды, обитающие в безлесной саванне
и гнездящиеся в термитниках. Родственная группа медоуказчиков включает виды
с нетипичным рационом – воскоед питается восковыми постройками диких пчёл,
а трупная птица (кадаверорнис) – мёртвыми животными.
В Африке богато представлены ракшеобразные и кукушки. Среди ракшеобразных имеется
много видов зимородков – от мелких до относительно крупных. Кукушки представлены
видами, находящимися на разных стадиях приспособления к гнездовому паразитизму.
Среди них есть насекомоядные виды, а также несколько хищников, охотящихся на
мелких млекопитающих, птиц и рептилий.
Среди певчих птиц очень разнообразны ткачиковые; некоторые из них гнездятся
очень большими колониями или собираются в стаи, насчитывающие десятки тысяч
особей. Среди ткачиков есть виды с необычным рационом – частичные гематофаги,
питающиеся кровью крупных копытных параллельно с извлечением паразитов из их
кожи. В тропических лесах и редколесьях обитают птицы-венценосцы – эндемичное
для Африки семейство, родственное ткачикам. Также разнообразны врановые птицы.
Некоторые из них заняли экологическую нишу рогатых воронов эпохи голоцена:
это крупные наземные птицы, питающиеся насекомыми и мелкими позвоночными.
В фауне рептилий основным изменением является смена крокодилов в качестве доминирующих
водных хищников другими группами рептилий. Крокодилы в Африке вымерли почти
повсеместно, и лишь в тропических лесах сохранилась небольшая группа потомков
карликового тупорылого крокодила – наземные хищники подлеска.
Роль крупного водного хищника в бассейне Сахарского Нила занимает черепаха-крокодил
– гигантский вид мягкотелых черепах, нападающий даже на водных птиц и небольших
млекопитающих. Наземные черепахи представлены очень богато, наибольшее их многообразие
приходится на пустынные районы. В реках обитают разнообразные водяные черепахи
из семейства эмидид; один из их представителей, чернильная черепаха, обладает
особым способом защиты от хищников. На восточном побережье Африки (побережье
пролива Танганьика) обитает водорослевая черепаха – представитель наземных
черепах, приспособленный к жизни в морской воде.
Тропический климат способствует существованию разнообразных ящериц. Для лесистых
местностей характерны хамелеоны, преимущественно мелкие или средних размеров.
Настоящие ящерицы, представленные только наземными формами, встречаются в большинстве
своём на севере, проникая из Европы через Гибралтарский перешеек, и по окраинам
Средиземноморской котловины. Некоторые настоящие ящерицы по западному берегу
материка расселились до Центральной Африки. Более типичны для Африки семейства
тропического распространения – агамовые, сцинки, поясохвосты. Агамы очень разнообразны
в тропических лесах, где обитают в основном в кронах деревьев, а также в пустынях.
Своеобразная «травяная рыба» – это агама с высоким и сжатым с боков телом,
живущая среди трав саванны. Сцинки – преимущественно роющие или просто наземные
формы. Типичны представители гекконов; как правило, это мелких или средних
размеров ночные ящерицы, питающиеся насекомыми или мелкими рептилиями и земноводными.
Некоторые из них представляют собой прекрасный пример мимикрии.
Вараны очень разнообразны и часто достигают крупных размеров. Среди пустынных
видов встречаются активные хищники и падальщики. В бассейне Конго и южнее роль
крупных водных хищников перешла от крокодилов к водноварановым – особому семейству
чешуйчатых, родственных типичным варанам. Водноварановые обитают также на соседней
Земле Зиндж.
Змеи Африки принадлежат к тем же семействам, что и в эпоху человека. Среди
ужеобразных змей появились виды, отличающиеся узкой специализацией. Сверлозмей
питается исключительно содержимым яиц птиц и крупных рептилий, а слепая пещерница
– змея-гермафродит, населяющая пещеры и питающаяся детёнышами летучих мышей
и пещерными насекомыми. В саваннах и редколесьях на севере и северо-востоке
Африки обитают эндемичные широкохвостые змеи – змеи грацильного сложения, питающиеся
насекомыми. Среди семейства гадюк примечательна гадюка-термовизор, способная
видеть инфракрасное излучение. В тропических лесах Центральной Африки обитают
различные представители семейства удавов, часто отличающиеся очень крупными
размерами. Среди них червехвостые питоны выработали оригинальный способ охоты
с помощью хвоста-приманки.
На территории Африки обитают только бесхвостые и безногие земноводные.
В Северной Африке бесхвостые многочисленны только в горах Атлас и в Эфиопии,
а по краю Средиземноморской
котловины их численность исчезающе мала и встречаются лишь отдельные представители
семейства жаб. Наибольшее многообразие земноводных приходится на Центральную
Африку. Здесь распространены и весьма обычны безъязыкие лягушки, родственные
южноамериканским пипам – разнообразные потомки шпорцевой лягушки. В более сухих
местностях обитают узкороты. Крупная лягушка-капкан из семейства настоящих
лягушек встречается в речной системе Сахарского Нила и Нигера. Её головастики
– частично неотенические хищные формы. В лесах широко распространены квакши.
Среди них появилось несколько ядовитых видов, использующих для защиты «заимствованный»
яд насекомых.
Безногие земноводные населяют исключительно лесистые местности с влажной почвой
и толстым слоем лесной подстилки.
Рыбы Африки наиболее разнообразны в центральной части материка, в бассейне
реки Конго. Раскол материка вызвал исчезновение многочисленных видов эндемичных
рыб Великих Африканских озёр. В реках бассейна Индийского океана встречаются
пресноводные и проходные акулы, а некоторые их популяции постоянно живут в
реках. Нижнее течение Конго населяет крупный пресноводный электрический скат.
Также в низовьях рек бассейна Индийского океана обычны скаты-хвостоколы.
Протоптерусы сохранились и процветают, несмотря на древность этой группы рыб;
они обитают к северу от бассейна Конго, но один из видов встречается в болотах
Конго и утратил способность пережидать засуху в коконе. Также для Африки характерны
многопёры и каламоихты, сохранившиеся с эпохи человека и очень мало изменившиеся.
Для Эфиопской области эндемичны мормировые (рыбы-слоны) из отряда костеязычных,
часто достигающие крупных размеров. Отряд хараковидных, общий с Южной Америкой,
ещё
в эпоху человека не включал ни одного общего семейства. В реках Центральной
и Западной Африки обитают многочисленные хараковидные рыбы нескольких семейств.
Большинство из них – мелкие всеядные и насекомоядные формы, но есть также крупные
растительноядные виды. Крупные харациниды, фаго-барракуды, являются активными
рыбоядными хищниками. Среди харациновых в неоцене появилось особое семейство,
эндемичное для Центральной Африки – гребнегубые. Это специализированные растительноядные
формы, питающиеся плавающими растениями, в основном ряской. На юге и востоке
материка распространены карповые – в большинстве случаев мелкие всеядные формы.
Сомы представлены несколькими семействами: бахромчатоусыми (синодонтисы), клариевыми,
косатковыми и электрическими сомами. Из окунеобразных рыб очень характерны
цихлиды, встречающиеся в постоянных водоёмах. Цихлиды Великих озёр почти все
вымерли при расколе Африки, и в ихтиофауне встречаются лишь немногие потомки
этих рыб. Большинство цихлид Африки – потомки тиляпий и родственных форм из
бассейнов Конго и Нила-Нигера. В областях с сезонной засухой водятся рыбы,
приспособленные к воздушному дыханию – змееголовы и лабиринтовые. Для рек тропической
Африки характерны хоботнорылообразные – угревидные роющие формы среднего и
крупного размера, питающиеся беспозвоночными и рыбами.
На крайнем юге Африки обитают галаксиевые – это наследие древней антарктической
фауны.
Беспозвоночные представлены очень богато, особенно в лесных областях. Среди
хищных жуков встречаются разнообразные жуки-скакуны, мягкотелки (в том числе
очень ядовитые), жужелицы, бомбардиры и разнообразные светляки. Разноядные
жуки часто достигают крупного размера. В лесах особенно распространены жуки-усачи
и жуки-олени, часто с причудливыми рогами, а также яркоокрашенные бронзовки.
Среди древогрызущих жуков распространены златки. В саванне и редколесьях, где
обитают крупные травоядные, водятся навозные жуки, в том числе довольно причудливые.
Один из их представителей, зеркальный жук, обитает в пустынных районах севера
Африки.
Тараканы населяют преимущественно лесные местности и принадлежат к тем же семействам,
что и в эпоху человека. Разнообразны термиты, в том числе питающиеся навозом.
Своеобразные комплексы видов беспозвоночных складываются вокруг постоянно подновляемых
навозных куч крупных травоядных. Среди бабочек типичны парусники, белянки и
нимфалиды. Муравьи разнообразны по размерам и образу жизни. Один из крупнейших
видов, гроза тропического леса – крупный муравей адумбулу. В болотистых местностях
обитают многочисленные комары. Среди мух есть паразитные виды – пиявкомухи
(некоторые виды общие с Восточной областью и даже с Голарктикой). Хищные и
падалеядные мухи достигают крупных размеров, среди них примечательны ядовитые
ктыри самбио, нападающие на мелких позвоночных.
Скорпионы обычны по всей Африке. В аридных районах они очень разнообразные,
но большей частью мелкие, а в лесных влажных местообитаниях встречаются крупные
виды. Часто встречаются большие пауки, в том числе пауки-скакуны, нападающие
на ящериц, и кругопряды, строящие сети диаметром несколько метров. В пустынях
Северной Африки обитает перекатипаук – особый вид пауков-волков. Также характерны
крупные многоножки, в том числе ядовитые сколопендры.
Разнообразны наземные улитки; пустынные виды отличаются приспособлениями, позволяющими
пережидать засуху. В тропических лесах обитают наземные пиявки, нападающие
на позвоночных. Пресноводные пиявки разнообразны, среди них есть крупные хищные
виды, питающиеся рыбой.
 |
 |
 |
Behemohyrax calamophilus |
Xenondipina agilis |
Hyracolopa velocipes |
 |
 |
 |
Vermiglossorex formicivora |
Breviardea pendula |
Carnopsittacus ferox |
 |
 |
 |
Afrodendrobates xanthonigrum |
Arachnosynodontis inconspicuus |
Speculunitor audax |
| Саванны Северной Африки, крупные травоядные и хищники | |
| Саванны Северной Африки, мелкие животные | |
| Нил-Нигер - крупнейшая речная система Северной Африки | |
| Саванны Северной Африки | |
| Североафриканская саванна - падальщики и пожиратели навоза. | |
| Цитадель на дереве | Влажный тропический лес Экваториальной Африки, полог леса |
| Ядовитое братство | Влажный тропический лес Экваториальной Африки; мелкие обитатели леса. |
| Между солнцем и солью | Миграция обитателей саванн Северной Африки к Мальтийскому горному массиву. |
| Край последних гиен | Южная Африка, крупные млекопитающие саванны и другие жители саванны и реки. |
| Река чудовищ | Экваториальная Африка, наземные и водные животные и растения в бассейне реки Конго. |
| Южная Африка, животные и растения пустыни Калахари | |
| Пульс реки | Северная Африка. Сахарский Нил - река в сезон дождей, и её обитатели. |
| Замбези в неоцене | Обитатели рек и речных берегов востока
Африки. (автор - Bhut) |
Земля Зиндж
|
Система рифтов Восточной Африки в эпоху человека. |
Фактически, Земля Зиндж (Восточно-Африканский микроконтинент)
представляет собой наиболее гористую часть Африки, отколовшуюся от основного
материка. Не исключено, что пролив Танганьика через много миллионов лет станет
океаном вроде Атлантического, но пока это довольно узкий пролив (ширина около
100 км), полностью разделяющий две части некогда единого материка. На дне пролива
Танганьика вулканические процессы проявляются в полной мере: со дна бьют горячие
источники, а в некоторых местах извергаются подводные вулканы. Восточная ветвь
Восточно-Африканского рифта глубоко врезалась с севера в массив Земли Зиндж,
образовав узкий залив Туркана, отделяющий северо-западную часть микроконтинента
(около половины его площади). Раскол материковой плиты продолжается – через
миллионы лет уже Земля Зиндж разломится по восточной ветви Восточно-Африканского
рифта надвое. Здесь часты землетрясения, а время от времени случаются извержения
вулканов.
Преобладающая форма рельефа Земли Зиндж – горные плато; большая часть микроконтинента
находится на высотах до 1 – 2 тысяч метров над уровнем моря и представляет собой
сильно выветренные плато со сглаженным рельефом. Кроме того, Земля Зиндж унесла
высочайшие горы Африки – Кению и Килиманджаро. Вулканические процессы, сопровождающие
раскол Африки, проявляются не только под водой, но и на суше: вдоль берега пролива
Танганьика появилась целая цепь молодых действующих вулканов. Раскол Африки
пробудил также Карисимби и другие вулканы, активность которых в эпоху человека
была низкой. Тектонические процессы вызвали небольшой наклон восточного осколка
Африканской литосферной плиты, что вызвало изменения в направлении течения рек.
Из-за большой высоты над уровнем моря на значительной части микроконтинента
господствует сухой климат. Все влажные ветры с океана оставляют влагу в прибрежных
районах, составляющих сравнительно небольшую часть территории. По степени увлажнения
на Земле Зиндж можно выделить две области: западную и восточную. Западная часть
микроконтинента – узкая полоса между цепью вулканов и побережьем пролива Танганьика.
Горные склоны достаточно круты, а берега в некоторых местах обрывисты. Здесь
мало рек, а те, что есть, короткие и порожистые, с многочисленными водопадами.
Лишь в области экваториального климата на западном побережье Земли Зиндж выпадает
достаточное количество осадков для роста деревьев.
Восточная часть Земли Зиндж – это область, подверженная влиянию ветров с Индийского
океана. Здесь рек значительно больше, и сами они полноводнее. Они текут в горных
долинах, и потому их русла остались узнаваемы даже несмотря на время, прошедшее
с эпохи человека. В горных долинах существуют озёра, а в центральной части микроконтинента,
где ранее располагалось озеро Виктория, раскинулась заболоченная местность –
остатки этого озера. Это место, между берегом пролива Танганьика и заливом Туркана,
достаточно хорошо увлажняется.
Северо-восточная часть Земли Зиндж более засушлива, чем центральная, находящаяся
в области экваториального климата. Благодаря обильным дождям в долинах центральной
и южной части Земли Зиндж текут реки и разливаются озёра.
В общих чертах растительный мир Земли Зиндж и Африки очень
сходен. Разница обусловлена особенностями рельефа Земли Зиндж – здесь явно
преобладают растительные сообщества, характерные для горных районов, и есть
несколько уникальных сообществ высокогорной флоры тропического происхождения.
Северо-западная часть Земли Зиндж – это область саванн и редколесий. Здесь
богато представлены злаки и засухоустойчивые деревья – акации, баобабы и эвкалипты.
Последние являются потомками видов, завезённых человеком в историческую эпоху,
успешно натурализовавшихся и эволюционировавших. Влаголюбивые виды растений
встречаются в этой части микроконтинента только в долинах рек. Ближе к берегам
залива Туркана саванны сменяются редколесьями, где доминируют акации.
Северо-западное побережье (берег пролива Танганьика) – место с наименее благоприятными
условиями для роста растений. Здесь выпадает меньше дождей, чем на восточном
побережье на той же широте, и во флоре явно преобладают засухоустойчивые виды.
В этой области произрастают суккулентные молочаи с редуцированными листьями
и колючими канделябровидными стеблями, а также крупные виды алоэ.
Значительная часть территории Земли Зиндж имеет сложный рельеф, сочетающий
низины и возвышенности, и результатом этого стали многообразие и мозаичность
ландшафтов. На возвышенностях условия более сухие, и здесь преобладают различные
виды злаков, образующие высокотравную саванну, а также отдельные виды деревьев.
Травянистые растения, не относящиеся к злакам, встречаются в таких местах реже
и населяют преимущественно склоны, где злаки не образуют цельной дерновины.
В ущельях и речных долинах складывается более влажный микроклимат, который
благоприятствует развитию древесной растительности. По речным долинам некоторые
виды деревьев тропического леса проникают с восточного побережья дальше на
запад, где на возвышенностях доминируют виды, более устойчивые к сухости. Для
таких мест характерны пальмы различных видов; нередко один и тот же вид в разном
количестве встречается и в лесах, и в саванне.
В высокогорных районах складывается уникальный комплекс видов тропической по
происхождению, но холодостойкой высокогорной флоры с листьями, покрытыми воском
или волосками для защиты от ночных холодов. В горных районах произрастают очень
крупные растения семейства сложноцветных – в большинстве своём кустарники,
но некоторые древовидные. Среди травянистых растений и одноствольных одревесневающих
«травяных деревьев» преобладают геснериевые и лобелиевые. Во время цветения
заросли этих видов украшены яркими цветками, привлекающими птиц и насекомых.
От горных лугов до низинных болот распространены виды рода сансевьера. Среди
них есть низкорослые растения, образующие плотные дерновины, и крупные виды
с хрящеватыми листьями.
Крупные участки влажных тропических лесов есть только по побережью Индийского
океана и в южной части микроконтинента. Для них характерно многообразие видов,
сравнимое с лесами Центральной Африки; разница проявляется на уровне видов
в пределах общих родов. В горных лесах на юге микроконтинента произрастает бамбук
разных видов, образующий сплошные заросли. В высокогорье складывается особый
тип мохового леса, где очень богат видовой состав мхов и лишайников. Также
в горных лесах растут древовидные папоротники, образуя подлесок в районах разреженного
древостоя. Итогом воздействия человека на флору является наличие в лесах Земли
Зиндж эвкалиптов, происходящих от завезённых в историческое время видов. Эвкалипты
особых видов, приспособленные к жизни в условиях влажного тропического леса,
составляют самый верхний ярус, возвышаясь над пологом леса поодиночке или небольшими
группами. Новой для лесов микроконтинента является группа бегониевых деревьев
– древесных растений, эволюционировавших на островах Индийского океана и заселивших
также побережья Азии и Мадагаскара.
На пологих восточных берегах Земли Зиндж в устьях рек разрастаются мангровые
заросли. На побережье пролива Танганьика мангровые заросли встречаются редко,
и лишь в глубине залива Туркана мангры занимают обширные площади, но их видовой
состав значительно беднее, чем на азиатском побережье.
Фауна Земли Зиндж относится к Эфиопской области и наследует
многие черты африканской фауны.
Знаковой группой неоценовой фауны млекопитающих Африки являются
даманы. Их эволюция на материке породила разнообразные формы крупных растительноядных,
занимающих различные экологические ниши. Даманы Земли Зиндж более консервативны:
они не образовали таких причудливых форм и сохранили больше черт сходства с
предками. В лесах и горах обитают различные виды рогатых жиряков, отличающихся
костяными выростами на переносице. Эндемично семейство ашкоковые, включающее
лазающие растительноядные формы, в том числе очень крупные (нголоко) и высокоспециализированные
(вентрохиракс). Из континентальных потомков даманов на Земле Зиндж обитает
морской ипопо – вид, способный пересекать морские проливы. Он обитает по всему
побережью Земли Зиндж, особенно на восточном побережье микроконтинента, где больше
распространены мангровые заросли.
Особого разнообразия на Земле Зиндж достигли прыгунчики. Некоторые их виды
освоили новые среды обитания: есть один вид из горных ручьёв, похожий на выхухоль.
Большинство видов прыгунчиков – некрупные формы из сухих местностей северо-востока
Земли Зиндж, но есть крупный всеядный вид из тропических лесов юга и юго-востока.
Насекомоядные млекопитающие представлены ежами, обитающими преимущественно
в области саванн, и землеройками, распространёнными повсеместно. На Земле Зиндж
сохранились представители отряда трубкозубов. Очень крупный вид этих животных
– мегаардварк, бипедальное животное, питающееся насекомыми и падалью.
Травоядные представлены многочисленными копытными. Жвачные – это различные козолопы,
потомки одичавших домашних коз. Они сильно варьируют в размерах от мелких до
очень крупных видов, питаются травой и листьями, и наиболее многообразны в саваннах
и редколесьях. В тропических лесах обитают разнообразные свинорогие, размерами
от мелких до средних. Свинорогие Земли Зиндж – формы с двумя крупными парными
рогами, в отличие от однорогих африканских. Некоторые свинорогие заходят в саванну.
Зайцелопы не смогли проникнуть на Землю Зиндж, и это позволило сохраниться первичной
фауне, включающей копытных. Среди них домашние козы быстро стали доминировать
и вытеснили потомков других групп.
По всему микроконтиненту распространены крупные грызуны, среди которых самым
большим является «панцирный дикобраз» катафрактерий. Большинство грызунов,
однако, представлено мелкими и средних размеров видами. Среди них есть роющие,
наземные и лазающие формы, принадлежащие к семействам мышиных, беличьих, хомяков
и дикобразов.
Приматы Земли Зиндж – это разнообразные полуобезьяны, являющиеся потомками
галаго, и мартышки, как наземные, так и древесные. Потомки галаго преимущественно
животноядные, обитают как на деревьях, так и на земле; в тропических лесах
есть несколько почти полностью наземных видов. Мартышки наиболее разнообразны
в мозаичном ландшафте северо-востока микроконтинента.
Хищники Земли Зиндж на уровне семейств сходны с африканскими. В фауне присутствуют
псовые (потомки лисиц и шакала), куньи, виверровые и кошачьи. Псовые наиболее
разнообразны на открытых пространствах – на возвышенностях северо-восточной
части микроконтинента. Лесные формы более редки – их заменяют представители других
семейств. Среди виверровых разнообразны мангусты и циветты – наземные формы,
обитающие в лесах. Большинство кошачьих и все куньи – мелкие хищники (среди
куньих есть водные). Среди крупных хищников микроконтинента примечателен последний
представитель крупных кошек (род Panthera) – нунда, потомок леопарда. Роль
более мелких хищников играют не кошки, а крупные кошкоподобные виверровые (маладомини).
Рукокрылые наиболее богато представлены в лесах, где встречаются многочисленные
плодоядные и нектароядные формы. Кроме них, есть отдельные виды хищных рукокрылых,
охотящихся на мелких позвоночных. На северо-востоке микроконтинента обитают преимущественно
мелкие насекомоядные виды, истребляющие кровососущих насекомых возле стад крупных
травоядных. Высокогорные летучие мыши существуют в условиях низких ночных температур,
поэтому они летают днём и впадают в оцепенение ночью. В верхнем ярусе тропических
лесов обитает солнечная летучая лисица – крупный, медленно летающий и преимущественно
лазающий вид летучих лисиц.
Фауна птиц Земли Зиндж несёт много сходства с африканской: общность проявляется
на уровне отрядов и большинства семейств. В орнитофауне Земли Зиндж, однако,
существуют эндемичные семейства, появившиеся после отделения её от африканского
континента.
В лесах микроконтинента обитают карликовые лесные страусы, анатомически сходные
с видами из Африки. Также есть некрупный всеядный вид страусов в саваннах северо-востока.
Водные птицы наиболее разнообразны в южной и центральной частях микроконтинента,
а также в низинных районах вдоль побережья Индийского океана. Фламинго, сохранившиеся
с эпохи человека, почти не изменились, в отличие от вида из Средиземноморской
котловины, и населяют озёра на северо-востоке (в области саванн) и на юго-западе,
вблизи болот Виктория. Богато представлены аистообразные птицы: аисты, в том
числе довольно крупные, цапли, а также своеобразный колониальный молотоглав,
образующий большие скопления в прибрежных зарослях и заболоченных лесах. На
болотах обитают разнообразные пастушковые птицы, часть их видов приспособилась
к жизни в высокотравной саванне. Также на болотах обитают различные султанки
и лысухи, многочисленные кулики. Мелкие кулики обитают на морских побережьях.
Гусеобразные птицы – различные виды уток и гусей, в том числе разнообразные
древесные утки, обитающие в заболоченных лесах южной части микроконтинента. Некоторые
виды уток обитают в мангровых лесах на океанском побережье.
Среди наземных птиц большого разнообразия достигли ракшеобразные и родственные
отряды. Представители ракшеобразных – различные виды зимородков, щурок и сизоворонок.
Зимородки наиболее разнообразны на болотах Виктория и в бассейнах рек центральной
и восточной частей Земли Зиндж. Щурки населяют преимущественно саванны северо-восточной
части микроконтинента, а сизоворонки и родственные им виды встречаются в лесных
районах микроконтинента.
Разнообразие хищных птиц заметно ниже, чем в эпоху человека. Крупные орлы и
грифы, а также соколы исчезли, но остались ястребы и коршуны. Также есть потомок
луня, конвергентно сходный с птицами-секретарями эпохи человека.
Совы многочисленны в лесных районах, где встречаются крупные виды, размерами
сходные с филинами эпохи человека. В саваннах совы – преимущественно мелкие
виды, обитающие в группах деревьев. Некоторые виды сов, обитающие в заболоченных
районах Земли Зиндж, ловят рыбу и лягушек. Один мелкий вид сов питается древесными
лягушками, охотясь на них на слух, по голосу. Семейство сипуховых разнообразно
и представлено несколькими мелкими видами.
Родственные совам козодоеобразные – представители семейства настоящих козодоев,
разнообразные в саваннах и горных районах с разреженной растительностью.
Кукушки наиболее разнообразны в лесах, где имеется большое количество крупных
видов. Они все являются гнездовыми паразитами, и их птенцов выкармливают преимущественно
различные воробьинообразные птицы. В саваннах обитают лишь единичные виды,
для размножения мигрирующие в лесные районы.
Дятлообразные обитают главным образом в лесных районах микроконтинента. Некоторые
виды общие для Африки и Земли Зиндж – воскоед (в тропических лесах) и трупная
птица (в саваннах и редколесьях). В лесах разнообразны настоящие дятлы, а в
саваннах северо-востока Земли Зиндж обитают дятлы-костоломы.
Голубеобразные птицы микроконтинента – различные виды голубей и курро. Голуби
распространены повсеместно, в том числе в засушливых районах западного побережья
микроконтинента, где они совершают регулярные миграции. В саваннах живут мелкие
стайные виды горлиц, питающиеся семенами, а в лесах – крупные голуби яркой
окраски, питающиеся семенами и плодами. В лесах южной части микроконтинента водится
несколько крупных видов наземных голубей, не утративших способностей к полёту.
Исключительно лесными обитателями являются курро, обитающие в южной и восточной
части микроконтинента. Они избегают конкуренции с голубями за счёт древесного
образа жизни.
Крупнейшим по количеству видов на Земле Зиндж является отряд Воробьинообразные.
На уровне семейств фауна певчих воробьиных сходна с фауной Африки, но существует
некоторое количество уникальных, в том числе эндемичных семейств. Одним из
них являются ежеголовые скворцы, потомки настоящих скворцов, распространённые
также в Южной и Юго-Восточной Азии и на островах Индийского океана. Настоящие
скворцы также обитают в лесах микроконтинента. Эволюция врановых на Земле Зиндж
привела к появлению новых групп птиц. Одной из них являются резонаторнисы –
лесные птицы, отличающиеся очень громкими голосами. Также на Земле Зиндж и
близлежащих островах существует эндемичное семейство гребнеклювов – лесных
птиц с ярким вертикальным выростом надклювья. Среди них имеются как древесные,
так и наземные формы, преимущественно всеядные и плодоядные. Из мелких певчих
птиц на Земле Зиндж обитают синицы и мухоловки, а также вьюрковые, ткачики
и астрильды. Необычным компонентом орнитофауны являются оляпковые – обитатели
быстрых ручьёв в горах. Очевидно, их расселение произошло во время ледникового
периода через Северную и Северо-Восточную Африку. На юге Земли Зиндж водятся
береговые оляпки, обитающие на морском побережье и добывающие корм на мелководьях.
В условиях тропического климата на Земле Зиндж очень разнообразны рептилии.
Среди наземных черепах появились виды, обладающие причудливыми выростами на
переднем краю панциря – клювоспинная и клыкастая черепахи. Наряду с ними в
саваннах Земли Зиндж обитает много видов наземных черепах более типичного облика.
По берегам пролива Танганьика обитает водорослевая черепаха – вторичноводный
растительноядный вид наземных черепах. Водные черепахи (семейств Emydidae и
Trionychidae) распространены преимущественно в восточной и южной частях микроконтинента.
Ящерицы очень разнообразны. Среди них богато представлены гекконы, обитающие
в лесах и среди скал; некоторые лесные виды обладают маскировочными приспособлениями
и являются активными хищниками. Также разнообразны агамы и хамелеоны, а в открытых
местностях – вараны. Характерное для тропиков Старого Света семейство водноварановых
представлено обитающим в реках крокодиловым вараном, который на Земле Зиндж
немного мельче, чем материковая форма.
Среди змей разнообразны питоны – от мелких до очень крупных, длиной несколько
метров. Некоторые питоны обитают в воде и покидают её только для кладки яиц.
Среди ужеобразных змей примечательна змея-копейщик. Среди наземных и древесных
змей распространены гадюки.
Земноводные Земли Зиндж принадлежат только к числу бесхвостых и безногих. Безногие
земноводные – различные виды червяг, в том числе очень мелкие и яркоокрашенные.
В лесной подстилке обитает несколько крупных видов червяг. Бесхвостые земноводные
– представители настоящих лягушек, жаб, узкоротов и квакш. Среди постоянноводных
лягушек характерны пиповые – различные виды шпорцевых лягушек.
Рыбы Земли Зиндж демонстрируют явное родство с африканскими. В реках встречается
несколько видов акул и скатов; одни из них населяют только мангровые заросли
и устья, где ощущается влияние морских приливов, а другие могут высоко подниматься
вверх по течению. Из двоякодышащих обитает лишь один вид протоптерусов, а другие
примитивные рыбы представлены многопёрами, сходными с видами эпохи человека.
Из пресноводных рыб африканского происхождения встречаются рыбы-слоники, среди
которых смертоносный филломормирус выработал способность испускать очень сильные
электрические разряды. Хараковидные рыбы немногочисленны по сравнению с обитателями
африканского континента. Самый крупный вид этих рыб на Земле Зиндж – узамбарский
хараколосось, обитатель горных рек. Карповые во многом замещают хараковидных
в реках микроконтинента. Среди них есть множество мелких яркоокрашенных барбусов
и расбор, а также растительноядные виды вроде потамолабео. Сомы нескольких
семейств водятся в пресных водах: это перистоусые, клариевые, косатковые сомы
и некоторые другие. В низовьях рек обитают морские ариевые сомы. Карпозубообразные
богато представлены на северо-востоке, в районах с сезонным климатом, где они
обитают во временных водоёмах. Потомки завезённой человеком гуппи обитают повсеместно
в постоянных водоёмах. Среди окунеобразных рыб характерны робаловые (стеклянные
окуни и латесы – потомки нильского окуня) и цихлиды. Часть цихлид является
потомками видов из Великих Африканских озёр, отступивших в пресные воды при
расколе Африки и затоплении озёр водами Индийского океана. Среди цихлид интересны
диспропорцихла – рыба с резко выраженным половым диморфизмом, и хромис-кукушка,
подбрасывающий свою икру этим рыбам. Из рыб, близких к окунеобразным, встречаются
хоботнорылообразные – разные виды мастацембелюсов. В устьях рек обитают рыбы
из морских семейств и отрядов – камбалы, монодактилюсы и скатофаги. Также для
рек Земли Зиндж типичны тетраодоны – как морские виды, заходящие в пресные
воды, так и полностью пресноводные виды.
Фауна беспозвоночных очень богата. Для саванн и лесов характерны разнообразные
термиты и муравьи, в том числе бродячие, представляющие опасность для большинства
животных мелкого и среднего размера. В различных ландшафтах распространены
общественные пчёлы – потомки медоносной пчелы, широко расселённой человеком.
Из двукрылых характерны крупные ядовитые ктыри самбио и паразитические пиявкомухи,
а также разнообразные комары, особенно многочисленные в болотистых районах.
В лесах обитают разнообразные бабочки, включая крупных парусников и бражников.
Среди жуков характерны навозники, разнообразие которых связано с разнообразием
копытных в саваннах.
Многочисленны и разнообразны пауки и скорпионы, среди которых встречаются крупные
виды. Некоторые виды многоножек достигают очень крупного размера и ядовиты.
Наземные моллюски часто достигают крупных размеров и обладают ярко окрашенной
раковиной. Среди пресноводных моллюсков появился роющий вид улиток-меланий,
обитающий в болотах на переувлажнённых почвах.
Среди червей характерны крупные дождевые черви, играющие важную роль почвообразователей
в саванне. В пресных водах и на суше обитают разнообразные пиявки.
 |
 |
 |
Ceratohyrax prolongocornis |
Gravicaper magnus |
Porceratella bovina |
 |
 |
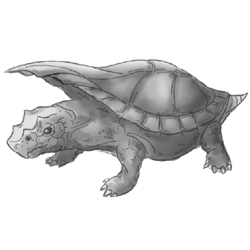 |
Ctenorhamphus sp. |
Thalassotestudo algophaga |
Ramphotestudo dorsognatha |
| Восточно-Африканский микроконтинент (Земля Зиндж), дождевой лес |
Мадагаскар
|
В неоцене география Мадагаскара почти не подверглась изменениям,
поскольку сам остров остался почти на прежнем месте по отношению к восточному
побережью Африки, лишь переместившись немного дальше на северо-восток, когда
произошёл раскол Африки и образование Земли Зиндж. Расстояние между Мадагаскаром
и некоторыми островами Индийского океана (Маскаренскими, Сейшельскими и Коморскими)
осталось прежним: они также «отплыли» дальше на северо-восток, оставшись в
прежнем положении относительно Мадагаскара. Это связано с тем, что все эти
острова образуют единое целое с восточной частью Африканской литосферной плиты,
расколовшейся в неоцене.
Мадагаскар является островом континентального происхождения, осколком Гондваны.
Его геология очень стабильна: остров находится далеко от очагов тектонической
активности, ближайшим из которых является «свежий» раскол, отделивший Землю
Зиндж от Африки. Вулканическая активность на острове практически отсутствует,
и лишь иногда случаются землетрясения, являющиеся отголосками геологических
процессов, протекающих на материке. География и климат острова тоже напоминают
эпоху голоцена: центр острова по-прежнему гористый (его занимает Высокое плато),
на западе раскинулись редкольесья и саванны с разной степенью сухости, а на
восточном побережье острова климат более влажный и там растут т. н. переменно-влажные
леса. Тем не менее, в неоцене склоны Высокого плато стали более пологими в
связи с эрозией, и западные и центральные области Мадагаскара стали более влажными,
чем прежде; переменно-влажные леса проникли и на северо- и юго-запад острова.
Это также связано с небольшим смещением острова к северу, поэтому климат северной
части острова стал более влажным.
На востоке острова образовалось большое количество болот и озёр, существующих
относительно недолго – на протяжении тысячелетий и десятков тысяч лет. Но благодаря
достаточному количеству осадков они полноводны на протяжении всего года. Большинство
водоёмов на западной части острова, напротив, зависят от сезонных изменений
погоды и могут даже пересыхать в сезон засухи или превращаться в грязевые ванны-ловушки.
Крупные озёра на равнинах отсутствуют совсем, но несколько озёр есть в горных
районах острова. Реки Мадагаскара берут начало на Высоком плато. В восточной
части острова они неширокие и короткие, а в западной значительно длиннее.
Обзор написан при участии Bhut'а, участника форума.
Флора Мадагаскара в неоцене сохранила своеобразие, которым
отличалась в эпоху человека, хотя из-за деятельности человека в ней появились
чужеродные элементы, а часть уникальных видов вымерла в результате разрушения
местообитаний и сведения лесов. Некоторые из представителей местной флоры занимают
доминирующее положение в лесах острова. Среди деревьев на западе острова встречаются
различные акации, а также баобабы – потомки тех немногочисленных видов, которые
смогли пережить антропогенное воздействие и климатические перемены, сопутствовавшие
смене голоцена неоценом. Акации процветают в сухих районах острова и являются
одними из доминантных видов деревьев Мадагаскара; некоторые виды закрепились
даже в предгорьях.
На западном побережье острова встречаются участки летнезелёных лесов, почти
полностью теряющих листву в сухой сезон. Но в саваннах есть также своеобразные
сообщества древовидных суккулентов. Их образуют суккулентные молочаи, крупные
виды алоэ и потомки завезённых человеком кактусов опунций. Они сохраняют свежий
зелёный цвет даже в засуху, а съедобные плоды кактусов привлекают в эти леса
обитателей окрестных саванн. В начале сезона дождей алоэ и кактусы зацветают,
привлекая множество опылителей из числа птиц и насекомых. Кроме того, в саваннах
острова, а также на горных склонах, обитает много видов невысоких травянистых
суккулентов – толстянок и молочаев.
На восточной половине острова, во влажных лесах, очень разнообразны пальмы
(среди них подавляющее большинство эндемично). Бобовые растения здесь – крупные
деревья, выделяющиеся яркими цветами на фоне зелени крон соседних деревьев.
Ещё одно типичное растение острова – равенала, мало изменившаяся с эпохи человека.
Веера листьев этого растения поднимаются в местах, где реки прорезают сплошные
лесные заросли. Яркими цветами выделяются деревья и кустарники семейства кутровых,
среди которых немало ядовитых видов. Интродуцированные в эпоху человека эвкалипты
также успешно натурализовались и дали начало новым видам, расселившимся по
острову и занявшим новые экологические ниши. Новая группа растений – бегониевые
деревья, быстрорастущие и разнообразные виды, не достигающие крупных размеров.
На морском побережье во многих местах растут мангры. Некоторые их виды смогли
приспособиться к пресной воде и проникнуть вглубь острова, где вместе с панданусами
они образуют на реках и в болотах густые заросли. Другие виды мангровых деревьев
доминируют как раз на морском побережье острова, образуя даже своеобразные
растительные аналоги рифов голоцена – они размножаются корневыми отпрысками,
которые отрастают от материнского растения и заходят далеко в море, на мелководья,
постоянно залитые морской водой.
На Мадагаскаре нет папируса, но есть некоторые его родственники семейства осоковых,
которые образуют густые заросли по берегам пресных водоёмов. С ними соседствуют
эндемичные виды камышей (потомки как коренных видов, так и завезённых сюда
челевеком), а на востоке к ним присоединяются виды мангровых деревьев, приспособившихся
к пресной воде. Среди водных растений характерны апоногетоны, среди которых
много эндемиков. Некоторые их виды живут в пересыхающих водоёмах и переживают
сухой сезон в виде клубней. Также в пресных водоёмах обитают разнообразные
пузырчатки, кувшинки, валлиснерии и другие растения семейства водокрасовых.
Нижние склоны мадагаскарских гор покрыты кустарниковым лесом-скрэбом – на восточных
склонах преимущественно акациевым, на западных – бамбуковым. Также на склонах
и в долинах встречаются эвкалипты. Вершины гор покрыты преимущественно травянистой
растительностью, образующей там своеобразную саванну с немногочисленными деревьями
и кустарниками – в основном всё теми же акациями, но в более влажных местах
растут также пальмы и бамбук.
Из травянистых растений наибольшим видовым разнообразием в восточной части
острова обладают орхидеи и папоротники-эпифиты. На западе, в саваннах, виды
цветковых растений более разнообразны на уровне семейств; большинство из них
опыляется ветром. Цветковые растения и папоротники разнообразием и оригинальной
внешностью не уступают видам, растущим в джунглях Африки и Азии. Среди них
велика доля эндемичных растений.
Обзор написан при участии Bhut'а, участника форума.
Фауна Мадагаскара в неоцене по-прежнему сохраняет свою необычность,
хотя в эпоху человека она обеднела в отношении местных эндемиков и приобрела
целый ряд несвойственных ей элементов – большей частью домашних животных.
Млекопитающие Мадагаскара в неоцене представлены, в первую очередь, разнообразными
лемурами. Среди них наиболее разнообразны семейства настоящих лемуров; также
существуют представители мышиных, индриевых и карликовых лемуров. Среди лемуров
появились аналоги наземных обезьян и даже кенгуру, освоившие жизнь на открытых
равнинах. Другие лемуры остались в лесах; среди них возникли формы, похожие
на ленивцев, и прыгающие виды, обладающие зачаточной летательной перепонкой.
Большинство видов лемуров является всеядными или растительноядными животными,
но среди них появились специализированные насекомоядные виды, а также несколько
видов с уклоном в хищничество.
Другие типично мадагаскарские млекопитающие – это тенреки, необычные насекомоядные
жители мадагаскарских лесов и саванн. Некоторые из них стали очень крупными
по сравнению со своими предками – размером со свинью. Большинство тенреков
осталось некрупными животноядными или всеядными видами; среди них есть наземные,
роющие, лазающие и водоплавающие формы. Грызуны представлены главным образом
семействами Nesomyidae (эндемичное мадагаскарское семейство) и мышиными (потомки
завезённых мышей и крыс). Среди них есть некрупные древесные виды, а также
много обитателей горных районов и саванн на западе острова.
Хищники острова представлены четырьмя семействами – кошачьими, псовыми и виверровыми
(потомки завезённых человеком видов), а также малагасийскими хищниками (более
редкие и специализированные потомки местных видов). Среди них есть экологические
аналоги леопардов, гепардов и куньих. Псовые и длинноногие виверровые обитают
в основном на равнинах, а кошачьи, большинство виверровых и малагасийские хищники
населяют лесные районы, кустарники и горы центральной части острова.
Рукокрылые представлены очень богато и разнообразно – их численность и многообразие
заметно увеличились после восстановления экосистем острова. Это различные летучие
мыши (мешкокрылые, молоссы, гладконосы и пр.) и крыланы, как эндемики, так
и родичи видов, которые живут на Земле Зиндж.
Птицы Мадагаскара также демонстрируют своеобразие, как и млекопитающие, но
степень эндемизма авифауны острова меньше, чем в эпоху человека. На болотах
и в саваннах живут различные виды голенастых птиц (аисты, цапли) и фламинго,
которые выжили в эпоху экологического кризиса. Фламинго Мадагаскара – общий
вид с Землёй Зиндж, образовавший устойчивые популяции на водоёмах близ западного
побережья острова. Другие водные птицы острова – бакланы, родственные видам
с Земли Зиндж, поганки, кулики и ибисовые. Чайки малочисленны и обитают на
морском побережье, а на пресных водоёмах селятся разнообразные крачки.
Гусеобразные птицы острова – это различные виды утиных птиц, общие с Землёй
Зиндж (различаясь на уровне подвидов) или родственных, но эндемичных видов.
Гусей и лебедей нет, но разнообразны утки, в том числе лесные, гнездящиеся
на деревьях влажных тропических лесов восточной части острова. Разнообразны
лысухи и родственные им птицы.
Самые примечательные птицы Мадагаскара – это крупные нелетающие куриные птицы
нумидорнисы, потомки завезённых цесарок. Они заняли нишу вымерших в плейстоцене-голоцене
эпиорнисов и успешно конкурируют с лемурами и тенреками в качестве крупных
растительноядных обитателей острова. Другие куриные птицы – несколько видов
перепелов.
Среди более мелких лесных птиц примечательны попугаи; некоторые из них могут
сравниться своими размерами и окраской с некоторыми птицами эпохи голоцена,
вроде попугаев ара из Южной Америки. Это в основном потомки попугаев-неразлучников
эпохи человека, но среди попугаев также есть несколько видов, являющихся потомками
более поздних мигрантов с Земли Зиндж – каких-то длиннохвостых попугаев. С ними
соперничают в яркости красок голубиные птицы, среди них местные виды курро.
Хищные птицы представлены потомками ястребов и коршунов, а также совами. Среди
сов самые крупные – местные виды семейства сипух; представители настоящих сов
мелкие, в основном насекомоядные или питающиеся мелкими позвоночными. На Мадагаскаре
очень разнообразны кукушки (в том числе крупные плотоядные виды) и ракшеобразные
птицы (сизоворонки, зимородки, удоды и некоторые другие). Воробьинообразные
птицы острова представлены ограниченным количеством семейств, среди которых:
ткачиковые и астрильдовые, врановые, мухоловки, ванги, скворцы (потомки завезённой
майны), нектарницы.
Рептилии на Мадагаскаре также демонстрируют высокую степень эндемизма, несмотря
на ущерб, понесённый в эпоху человека. Большинство их видов относится к отряду
чешуйчатых: это змеи и ящерицы, в первую очередь хамелеоны. На Мадагаскаре
живёт самый крупный хамелеон неоцена, хамелеон-стрелок, а также один из самых
маленьких – хамелеон-листочек. Хамелеоны соперничают в видовом многообразии
с гекконами, среди которых есть не только насекомоядные, но и растительноядные
виды. Гекконы тропических лесов часто вырабатывают искусные способы маскировки,
а виды из горных местообитаний отличаются способностями к лазанию по отвесным
скалам. Некоторые широко распространённые виды образуют много узкоареальных
подвидов. Также на Мадагаскаре обитают сцинки и поясохвосты; некоторые из них
достигают крупных размеров. Ядовитых змей нет, есть некрупные удавы.
Кроме ящериц и змей на Мадагаскаре обитают черепахи, как сухопутные, так и
пресноводные. Они происходят от видов, переселившихся с Земли Зиндж уже после
исчезновения человека; местные виды быстро вымерли в эпоху экологического кризиса.
Амфибий на Мадагаскаре очень мало, все они являются представителями отряда
бесхвостых – это лягушки (мантеллы, узкороты, настоящие лягушки).
Рыбы Мадагаскара сравнительно немногочисленны, а ихтиофауна
острова отличается бедностью, несмотря на расположение острова в тропиках: это
связано с давней изоляцией острова. Среди рыб имеются немногочисленные потомки
местных рыб: атеринообразные (бедоции), бычки, ариевые сомы и икромечущие карпозубые.
Эти рыбы связаны в своём происхождении с морскими формами. Эндемичные мадагаскарские
цихлиды были редкими в эпоху человека, а в эпоху экологического кризиса они
вымерли полностью. Их заменили потомки мозамбикской тиляпии, широко распространённой
в эпоху человека как объект коммерческого рыбоводства. Влияние деятельности
человека оставило свой след в ихтиофауне острова в виде потомков змееголовов,
завезённых из Азии, и большеротого форелеокуня из Северной Америки. Они явились
причиной исчезновения многих групп мадагаскарских рыб, а их потомки заселили
большинство пресных водоёмов острова: потомки форелеокуня преобладают в проточных
водах, а потомки змееголова населяют стоячие, часто пересыхающие водоёмы. Среди
растительноядных рыб на острове обитают потомки завезённых промыслового гурами
и карпа. Также на острове обитают живородящие рыбы – потомки гуппи и меченосцев,
завезённых человеком. Но их разнообразие не сравнимо с разнообразием живородящих
рыб в Карибском море и Центральной Америке.
Фауна беспозвоночных богата и отличается своеобразием. Насекомые
на Мадагаскаре представлены бабочками (среди них – урании, павлиноглазки, бражники),
мухами, жуками,
термитами,
богомолами
и т. д.
Эти
виды в
большинстве
своём
эндемичны, но есть и сравнительно недавние переселенцы с Земли Зиндж. Примечательны
разнообразные безжалые пчёлы. Палочники, тараканы и богомолы иногда достигают
очень крупных размеров. Среди других беспозвоночных особенно многочисленны
паукообразные. Некоторые пауки обладают роговидными выростами на брюшке,
есть гигантские пауки-кругопряды. В тропических лесах обитают многоножки –
крупные кивсяки и ядовитые сколопендры. В лесах широко распространены крупные
брюхоногие моллюски. Некоторые улитки являются потомками африканской ахатины
и достигают гигантских размеров.
Обзор написан при участии Bhut'а, участника форума.
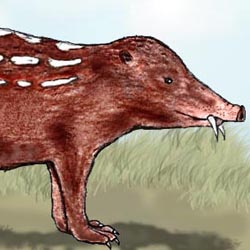 |
 |
 |
Scrofogale titan |
Floromungo apiphilus |
Pseudarara titan |
| Западная часть Мадагаскара, влажные саванны | |
| Крик лемура | Дождевой лес на востоке Мадагаскара |
Меганезия
|
В неоцене география Австралии претерпела существенные изменения,
связанные с продолжающимся движением Индо-Австралийской плиты на северо-запад.
Это движение сдерживается краем Китайской плиты, что привело к усилению вулканической
активности в районе Больших Зондских островов. Главным итогом этого явилось
объединение австралийского материка с островом Новая Гвинея; образовавшийся
в ходе этого процесса материк получил название Меганезия. Причиной объединения
стали движение Австралии на северо-восток и поднятие части литосферной плиты
на севере континента. Также в состав Меганезии вошли некоторые более мелкие
острова у северного побережья Австралии и близ Новой Гвинеи. Как следствие
образования Меганезии, на севере этого континента образовалось два очень больших
озера: Арафурское (бывшее Арафурское море) и Карпентария (бывший залив Карпентария).
Озеро Карпентария отделилось от океана раньше, и вода в нём стала почти пресной.
Арафурское озеро остаётся солёным, поскольку имеет связь с океаном через сложную
систему проток в обширных мангровых зарослях, но его солёность ниже океанической.
Оба озера окружены обширной полосой болот.
Если на севере Меганезии море отступило, то на юге сложилась прямо противоположная
ситуация. Здесь в сушу глубоко врезается мелководный морской залив Эйр. Он
отделяется от океана отмелями и песчаными косами, а сообщается с ним через
узкое горло. Мелководный и хорошо прогреваемый солнцем, залив Эйр беден кислородом,
а вода в нём может опресняться за счёт стока рек и осадков. Колебания солёности
могут быть довольно значительными – от половины океанической солёности до почти
пресной воды. Всё это создает определённые трудности для жизни населяющих его
животных. В центре входа в залив расположен небольшой остров Флиндерс, который
в эпоху человека был горным хребтом. В залив впадают короткие равнинные реки,
а также самая полноводная река Меганезии – Муррей. Остров Тасмания и в неоцене
остаётся изолированным участком суши, но в ледниковый период он несколько раз
соединялся с материком.
Меганезия в неоцене значительно сдвинулась на северо-восток и сблизилась с
островами Индонезии, однако сохранила географическую изоляцию. Несколько небольших
островов Индонезии ушли под воду и стали отмелями вблизи побережья Меганезии.
Движение тектонических плит спровоцировало активное горообразование на севере
материка: горы Новой Гвинеи, уже в эпоху человека бывшие сравнительно высокими,
в неоцене выросли ещё сильнее. На востоке материка расположена другая горная
цепь – Большой Водораздельный хребет. Здесь процессы горообразования уже не
идут, и эти древние горы постепенно разрушаются под воздействием эрозии. Однако
именно на этом хребте берут начало крупнейшие реки Меганезии. Часть рек, однако,
берёт начало в болотах и лесах, окружающих озеро Карпентария, и впадает в залив
Эйр.
Климат Меганезии в эпоху неоцена стал более влажным как за счёт географических
изменений, так и за счёт общего повышения влажности на всей планете. Север
материка лежит близко к экватору, здесь жаркий климат и почти ежедневно идут
обильные дожди. Впрочем, несмотря на близость к экватору, в горах северной
Меганезии может быть довольно холодно, а на горных вершинах круглый год лежит
снег. Большая часть Меганезии находится в зоне тропического климата с чередованием
сухих и влажных сезонов. За исключением Муррея и его притока Дарлинга, реки
здесь короткие. Многие из них существуют лишь в сезон дождей, а в сухое время
года представляют собой сухие русла. Некоторые реки круглый год текут под землёй.
Озёра небольшие и зачастую пересыхающие. Залив Эйр смягчает климат равнин,
лежащих вокруг него. На юго-западе материка климат становится более сухим и
жарким. Здесь большинство водоёмов – временные, за исключением родников и ключей.
На востоке Меганезии, восточнее Большого Водораздельного хребта, лежит зона
влажного климата: здесь воздушные массы с Тихого океана встречаются с горами,
и, как следствие, выпадают обильные осадки. Крайний юг Меганезии и Тасмания
– области субтропического климата. В горах Тасмании климат умеренный, с прохладной
зимой.
Обзор написан Семёном, участником форума.
Флора Меганезии является прямой преемницей австралийской и новогвинейской флоры, но её своеобразие в определённой степени сгладилось благодаря деятельности человека, который завёз в эти места ряд растений, ранее не встречавшихся в Австралии и на Новой Гвинее. Древесная растительность Меганезии представлена по большей части растениями трёх семейств: миртовые (главным образом эвкалипты), бобовые (акации) и протейные. Также в формировании лесов Меганезии участвуют нотофагусовые (южные буки) и араукарии.
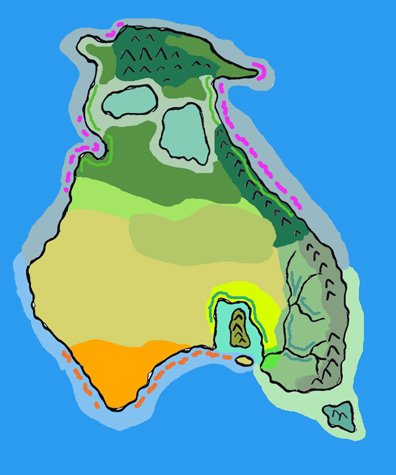
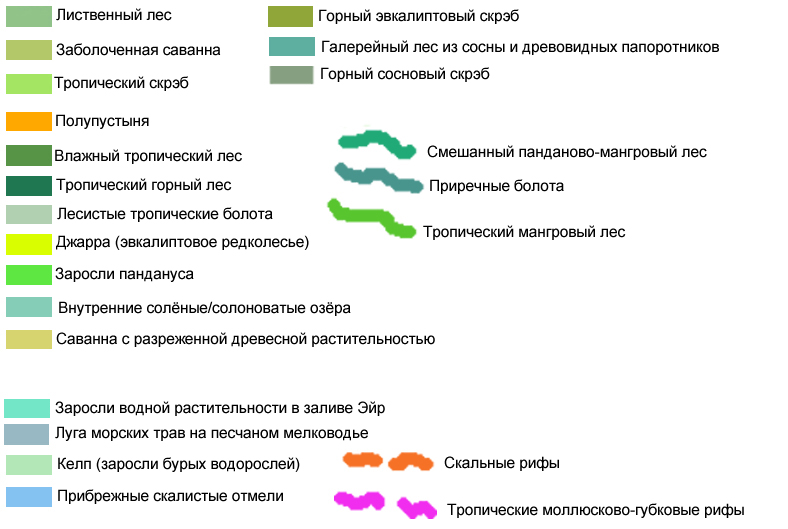
Схема природных зон Меганезии
Рисунок Тимоти Дональда Морриса (Австралия)
Северная часть Меганезии находится в области экваториального
и субэкваториального климата. Здесь ежедневно идут дожди, а постоянно высокая
температура способствует
росту влажных тропических лесов. Среди характерной растительности этих лесов
встречается много групп, общих с Азией: древовидные фикусы, пальмы с перистыми
листьями, сандаловые, лавровые и магнолиевые. Растения австрало-новогвинейского
происхождения здесь – некоторые виды эвкалиптов и других миртовых. Крупные
деревянистые лианы
представлены ротанговыми пальмами, а травянистые – различными видами семейства
перечных, часто с узорчатыми гладкими листьями. Среди эпифитов богато представлены
лишайники, мхи и папоротники. Очень характерны эпифитные папоротники «олений
рог» и асплениумы, часто с цельными листьями. Из эпифитных цветковых растений
обильно представлены орхидеи, демонстрирующие сходство с восточноазиатскими
родами. Среди деревьев влажных тропических лесов примечательны лапортеи – древовидные
представители семейства крапивных, имеющие очень жгучие листья.
Область Арафурского озера примечательна крупнейшими массивами мангровых лесов
на Земле эпохи неоцена. Многокилометровая полоса мангровых лесов отделяет озеро
от океана, ослабляя движение приливных волн и образуя изменчивую сеть проток
и отмелей. Берега озера также поросли солеустойчивыми мангровыми деревьями.
Подводные растения Арафурского озера – представители «морских трав», способные
расти в солёной воде. Лишь в устьях рек встречаются отдельные представители
пресноводной флоры, способные выдерживать кратковременное осолонение воды. Озеро
Карпентария более древнее, и вода в нём значительно опреснена (имеется лишь
остаточная солёность, около 1‰). Соответственно, меняется состав окружающей
растительности: озеро окружено зарослями пандануса, пальм и других деревьев,
способных выдерживать затопление или расти на мелководье, а гидрофлора озера
значительно богаче.
В результате движения материка горы, ранее принадлежавшие Новой Гвинее, стали
ещё выше, и в северной части Меганезии наблюдается явление высотной поясности:
влажные тропические леса низин по мере увеличения высоты над уровнем моря сменяются
менее теплолюбивыми горными лесами, которые постепенно переходят в высокогорные
заросли кустарников и альпийские луга. Для горной флоры севера Меганезии характерны
казуарина, нотофагусы и древовидные папоротники, образующие густой подлесок
в тени деревьев первой величины. На вершинах самых высоких гор Северной Меганезии
лежит снег, а ниже границы снегов располагается пояс растительности тропического
происхождения, приспособленной к резким суточным перепадам температуры.
Восточная окраина Меганезии получает больше влаги, чем западная, и пояса растительности
смещены соответствующим образом. Южнее пояса влажных тропических лесов располагается
пояс скрэба – тропической кустарниковой растительности, который с запада на
восток плавно переходит в область заболоченных саванн. На протяжении года эта
область материка получает различное количество влаги – от обильного увлажнения
в сезон дождей до крайне скудного в засушливый сезон. В результате этого растительность
в области скрэба представлена низкорослыми кустарниками (преимущественно протейными
и эвкалиптами), а в заболоченной саванне растут отдельные группы высоких деревьев,
а значительная часть растительности – многолетние корневищные травы, способные
выдерживать засуху. В этой области встречается много водоёмов, высыхающих в
сухой сезон (на 2 – 3 месяца), и в них произрастают преимущественно корневищные
и клубневые водные травы, способные переносить сезонное высыхание водоёма –
кувшинки и апоногетоны.
Особый комплекс растительности складывается на обращённой к Тихому океану стороне
Большого Водораздельного хребта. Здесь выпадает значительная часть дождей,
идущих с Тихого океана, и складываются благоприятные условия для произрастания
древесной растительности. Леса восточных склонов Большого Водораздельного хребта
образованы преимущественно эвкалиптами, араукариями и древовидными папоротниками.
Пальм в таких лесах почти нет, а по мере подъёма в горы встречаются сосны,
потомки видов, завезённых человеком.
На западных склонах Большого Водораздельного хребта образуется переходная зона
между горной растительностью и растительностью, типичной для равнин на соответствующей
широте.
Южнее пояса влажных тропических лесов условия, наиболее благоприятные для лесной
растительности, складываются в юго-восточной части Меганезии. Здесь произрастают
частично листопадные лиственные леса, приспособленные к существованию в условиях
сезонного изменения количества осадков. Большинство деревьев в таких лесах относятся
к миртовым (в основном, эвкалипты) и протейным. Кроны эвкалиптов пропускают
к земле достаточное количество солнечного света, чтобы в таких лесах образовывался
густой подлесок. Доминирующими видами подлеска становятся кустарниковые растения
семейства розоцветных – потомки завезённой и одичавшей ежевики. Как и в эпоху
человека, в лесах обильно представлены представители семейства росянковые –
многолетние травы, часто достигающие крупных размеров. Южнее влажных тропических
лесов эпифитные орхидеи отсутствуют, но разнообразны их наземные виды.
Вдоль рек бассейна Муррея тянутся заболоченные участки с типичной болотной
растительностью – крупными широколиственными травами. Водные растения представлены
пузырчатками, плавающими папоротниками, кувшинками и апоногетонами. На болотах
в условиях временного затопления произрастают некоторые орхидеи.
Большой Водораздельный хребет не пускает во внутренние районы значительное
количество осадков, поэтому значительная часть материка в центре и на западе
– сухие саванны с небольшими участками древесной растительности. Здесь доминируют
злаки, но встречаются представители других семейств растений. В сухой сезон
саванны имеют однообразный желтовато-коричневый цвет, а растения, произрастающие
здесь, совсем немного отличаются друг от друга по форме и цвету листвы. Но
после дождей облик саванн резко меняется: большинство растений раскрывает цветки
и равнины расцвечиваются яркими красками.
Залив Эйр представляет собой тёплый мелководный водоём, вокруг которого складываются
более благоприятные условия для роста растений. По берегам залива образуется
джарра – особое растительное сообщество, главными составляющими которого становятся
кустарниковые эвкалипты с примесью небольшого количества растений других семейств.
В заливе переменная солёность воды, зависящая главным образом от количества
осадков, поэтому на отмелях здесь образуются заросли водных растений, устойчивых
к переменам солёности – главным образом, «морских трав» особых видов, а также
устойчивых к солёной воде потомков пресноводных растений. В устьях рек, впадающих
в залив Эйр, вода обладает более стабильной низкой солёностью, поэтому здесь
формируются заросли пандануса, немного похожие на мангровые. Сплетения ходульных
корней этих растений дают убежище многочисленным видам рыб и беспозвоночных.
Особенно большие заросли пандануса развиваются в дельте Муррея, на юго-восточном
побережье залива Эйр.
Юго-запад материка – самая засушливая часть Меганезии. Здесь раскинулись полупустыни,
а растительность отличается скудностью и наличием приспособлений для выживания
в аридных условиях. Из древесной растительности в пустынях Меганезии встречаются
протейные с очень толстым водозапасающим стволом, а также «бутылочные деревья»
с высокими утолщёнными стволами, растущие небольшими группами. Древесная растительность
тяготеет к временным водоёмам, где вода задерживается на несколько недель.
Суккулентов, подобных кактусам, алоэ и молочаям, в Меганезии мало, и лишь некоторые
виды обладают отдельными признаками суккулентов. Зато в пустынях Меганезии
много однолетних растений-эфемеров, пережидающих засуху в виде семян и быстро
разрастающихся после дождя. Некоторые растения образуют водозапасающие подземные
клубни, а на поверхности земли заметны лишь листья и цветы.
В горных районах Меганезии и на острове Тасмания климат близок к умеренному,
но безморозный. Здесь господствуют папоротники разных видов, не требовательные
к теплу, в том числе древовидные. Древесная растительность представлена нотофагусом,
а также хвойными – различными видами араукарии и потомками завезённой человеком
сосны. В целом флора Тасмании отличается большим процентом эндемичных видов.
Фауна Меганезии относится к Австралийской области. Однако многие
животные неоценовой Меганезии являются потомками видов, завезённых человеком
и происходящих из иных зоогеографических областей.
Млекопитающие представлены различными систематическими группами.
Эндемиками Меганезии являются однопроходные: утконосы и ехидны. Они более разнообразны,
чем в голоцене. Потомки ехидн сильно различаются по размеру – от гигантской
стегоехидны до небольших форм, в числе которых древесный и роющий виды. От утконоса
ведут свой род специализированные водные млекопитающие.
Сумчатые Меганезии очень разнообразны, несмотря на то, что часть их видов вымерла
на рубеже эпох (сумчатые волки, сумчатые дьяволы, коала). Представители примитивного
семейства хищных сумчатых и семейств, «отпочковавшихся» от него в раннем неоцене,
занимают ниши насекомоядных и хищников. Неоценовые хищные сумчатые являются
потомками сумчатых крыс, сумчатых мышей, сумчатых тушканчиков и сумчатых куниц.
Среди мелких видов есть крайне специализированные формы – такие, как сумчатые
кровососы, сумчатые куторы и эмбриотерии. Некоторые из них стали подземным видами.
Другие – представители семейства иглохвостых сумчатых крыс – стали отчасти травоядными
и зерноядным формами. Крупные хищные сумчатые напоминают кошачьих, но среди
них есть и виды, не похожие ни на одно плацентарное млекопитающее – например,
сумчатый острозуб.
Кенгуру, своеобразная «визитная карточка» Австралии, продолжили эволюцию и в
неоценовой Меганезии. Они населяют как равнины, так и леса. Помимо типичных
кенгуру, появились необычные виды: массивные крючкопалые кенгуру, приспособленные
к быстрому бегу страусовые кенгуру. Семейство носатых кенгуру объединяет виды
с необычной внешностью, часть которых стала всеядными животными. Среди настоящих
кенгуру в неоцене встречаются и падальщики, и даже хищные виды.
По всей Меганезии широко распространены сумчатые из семейства кускусов, или
поссумов. Многие из них ведут свой род от голоценового лисьего кузу, очень многочисленного
вида, приспособившегося к жизни рядом с человеком. В тропических лесах и редколесьях
обитают древесные виды кускусов и близких к ним семейств, например, сумчатых
летяг. От поссумов происходят два новых семейства. Это крупные хищные поссумы,
напоминающие медведей и успешно конкурирующие с дазиуридами, и сумчатые лемуры,
потомки кускусов с более развитым мозгом и сложным социальным поведением – они
являются аналогами приматов.
На равнинах Меганезии обитают гигантские вомбаты ямути, питающиеся травянистой
растительностью. В целом, однако, ветвь вомбатов малочисленна и угасает.
За исключением летучих мышей и части грызунов, плацентарные обязаны своим появлением
в Меганезии человеку. До неоцена дожили далеко не все интродуценты: так, хищные
млекопитающие – кошки, собаки и куньи – вымерли из-за эпизоотий бешенства и
чумы плотоядных. Зато копытные Меганезии занимают важное место в экосистемах.
На севере материка обитают свиньи и олени, а в озере Карпентария – быкопотам,
полуводный представитель семейства полорогих, потомок азиатского буйвола. Южнее,
на равнинах и в редколесьях, распространены различные верблюдовые – от сравнительно
небольших верблюжьих антилоп до крупных кабаллокамелюсов и гигантского верблюда-жирафа
долихокамелюса.
Рукокрылые Меганезии разнообразны – это и настоящие летучие мыши, и крыланы.
Настоящие летучие мыши относятся к нескольким семействам, и их рацион сильно
варьирует – от нектара до насекомых и мелких позвоночных. Самый крупный вид
– летучий волк, хищная летучая мышь из семейства ложных вампиров, обитающая
в редколесьях. Большинство видов рукокрылых населяет тропические леса на севере
материка.
Все многочисленные грызуны Меганезии относятся к семейству мышиных. Они происходят
как от аборигенных видов, попавших в Австралию ещё в плейстоцене, так и от крыс
и мышей, случайно привезённых человеком. Эти звери населяют все типы ландшафтов
и демонстрируют разнообразие форм, среди которых выделяются водные и полуводные
виды, а также прыгающие грызуны саванн и пустынь. В фауне Австралийской области
также представлены млекопитающие отряда зайцеобразных. Все они ведут свой род
от европейских кроликов, завезённых человеком. Меганезийские кролики внешне
похожи на кроликов и зайцев голоценовой Евразии, но один их вид освоил водную
среду.
Птицы Меганезии стали более многочисленными, чем в голоцене.
В отличие от млекопитающих, интродуцированные виды птиц не смогли добиться успеха
на этом континенте: лишь несколько их видов оставили потомков в неоцене.
Самые большие птицы Меганезии не летают и относятся к отряду казуарообразных.
От эму произошли очень крупные ложные моа, похожие на страусов и населяющие
различные биотопы. Некоторые потомки казуаров стали животноядными видами: один
из них ловит рыбу, а другой, ягил, является одним из верховных хищников в тропических
лесах на севере Меганезии. Но большинство видов этих птиц (ложные моа) является
всеядными с уклоном в растительноядность.
Куриные птицы Меганезии происходят в основном от местных видов перепелов. На
севере материка обитают немногочисленные потомки сорных кур и одичавших домашних
кур.
На берегах пресных и солоноватых водоёмов обитают гусеобразные птицы – утки,
лебеди и черпоклювы, а также кулики, чайки, крачки, поганки, бакланы и пастушковые.
Аистообразные представлены цаплями и ибисами, среди которых выделяются гаруспики
– ибисы, ставшие специализированными падальщиками. Чайки, крачки, бакланы и
некоторые трубконосые населяют также морское побережье Меганезии. На юго-восточном
побережье обитают чайки-пингвины – представители группы птиц, характерной для
Антарктики и Субантарктики.
Большинство хищных птиц Меганезии относится к отряду ястребообразных, например,
ястребы, коршуны, канюки и луни. Крупнейшие пернатые хищники и падальщики Меганезии
являются потомками австралийского клинохвостого орла – одного из немногих орлов,
переживших эпоху человека. Имеются соколы, происходящие от мелких видов. Помимо
дневных хищных птиц, в Меганезии также обитают ночные хищники – сипухи и настоящие
совы (иглоногие совы), причём среди последних есть очень крупные виды.
Многочисленны голуби (некоторые из них ведут наземный образ жизни в тропических
лесах), кукушки, щурки, зимородки (в том числе потомки кукабарр) и стрижи. Дятлы
отсутствуют, и их экологическими аналогами являются животные других групп (например,
кускусы). Попугаи разнообразнее голоценовых и относятся к семействам какаду
и настоящих попугаев. Отряд козодоев представлен несколькими семействами и распространён
повсеместно, от лесов до пустынь.
Воробьинообразные птицы Меганезии весьма разнообразны. Некоторые семейства общие
для Меганезии и Евразии: это врановые, жаворонки, скворцы (в том числе потомки
завезённого обыкновенного скворца), дрозды, мухоловки (и родственные семейства),
трясогузковые, ласточки, личинкоеды, тимелии, питты, славковые, белоглазки и
другие. Широко распространены также астрильдовые, среди которых выделяется группа
меганезийских ткачиков и собственно ткачиковые, представленные потомками завезённого
домового воробья. Многие семейства воробьиных эндемичны для Меганезии и близлежащих
островов: это флейтовые птицы, шалашники, райские птицы, пардалоты, малюры и
многие другие. От малюров, или австралийских славок, в неоцене произошло семейство
бегающих малюров.
Фауна рептилий Меганезии отличается большим богатством. Здесь
обитают одни из немногих оставшихся на Земле видов крокодилов – крокодил-пилонос
(озёра Карпентария и Арафурское) и кархарозух (тропические морские воды). Пресноводные
черепахи представлены змеиношейными черепахами, разнообразие которых несколько
сократилось по сравнению с эпохой человека. Однако крупнейшая черепаха Меганезии
– бронтохелис – происходит от завезённой красноухой черепахи, из семейства эмидид.
Другие виды эмидид распространены в пресных водах Меганезии, в том числе в пересыхающих
водоёмах. В морях, омывающих материк, встречаются морские черепахи, относящиеся
к семействам, появившимся уже в неоцене, в частности, крупные черепахи-нетопыри.
Наиболее разнообразны чешуйчатые рептилии. Ящерицы относятся к семействам гекконов,
сцинков, агам и к другим, более мелким семействам. Наиболее характерно для Меганезии
семейство варанов, среди которых выделяется драковаран – верховный хищник пустынь
на западе континента. Большинство варанов Меганезии населяет открытые пространства,
хотя есть несколько лесных видов, обладающих способностями лазить по деревьям.
Все вараны питаются мелкими позвоночными, но среди них есть нетипичный вид,
питающийся общественными насекомыми. Змеи принадлежат к семействам питоновых
(среди них эйнгана, крупнейшая змея на планете), аспидовых, ужеобразных, слепозмеек
и некоторых других. Как и в эпоху человека, в неоценовой Меганезии обитает очень
большое количество видов ядовитых змей. В морях встречаются морские змеи, но
их разнообразие ниже, чем в эпоху человека: пелагические виды вымерли в эпоху
глобального экологического кризиса, и остались лишь потомки менее специализированных
прибрежных форм.
Амфибии Меганезии относятся только к отряду бесхвостыx. Наиболее
характерны квакши, узкороты и представители семейства австралийских жаб; настоящие
лягушки немногочисленны. Важное место в экосистемах занимают потомки завезённой
жабы-аги.
Разнообразие пресноводных рыб Меганезии невелико. В пресноводных
водоёмах обитают представители отрядов галаксиеобразные, атеринообразные и окунеобразные.
Галаксии образовали значительное количество пресноводных и проходных форм, отличающихся
друг от друга образом жизни и особенностями строения. В пустынных районах живут
рыбы из этого отряда, способные при пересыхании водоёмов выживать долгое время
в защитной капсуле. Атеринообразные представлены главным образом радужницами
и их потомками. В связи с конкуренцией с потомками завезённых рыб их многообразие
снизилось по сравнению с эпохой человека, но в реках обитают эндемичные малочисленные
виды этих рыб, отличающиеся яркостью окраски. Среди окунеобразных рыб в пресных
водах Меганезии обитают терапоны и горбылёвые. Пресноводные виды есть и среди
хрящевых рыб – акул и скатов. В реках водится некрупный вид пресноводной пилы-рыбы.
Эндемична гигантская двоякодышащая рыба буньип – потомок рогозуба, который смог
выжить благодаря его искусственному расселению и охране в эпоху человека. Многие
пресноводные рыбы происходят от морских видов – это камбалы, угрехвостые сомы
и другие. Они часто проводят в пресной воде лишь часть жизненного цикла, уходя
на нерест в море. Некоторые рыбы ведут свой род от завезённых человеком видов
– карпов, тиляпий, форелей. Потомки форелей населяют верховья рек Большого Водораздельного
хребта. Цихлиды заселили север и восток Меганезии, избегая солёной воды Арафурского
озера. Карповые предпочитают равнинные реки востока Меганезии (бассейн Муррея).
В мангровых зарослях водятся разные виды илистых прыгунов.
Беспозвоночные очень многочисленны, особенно в тропических
лесах. Наиболее разнообразны насекомые, а среди них жуки и бабочки. Новая для
Меганезии группа насекомых – жуки-навозники, происходящие от завезённых человеком
африканских видов. Они населяют главным образом равнины, где обитают копытные.
По всему континенту обитают термиты, многие виды которых строят внушительные
сооружения. Среди тараканов имеются очень крупные виды с долгим жизненным циклом.
Широко распространены паукообразные: пауки и скорпионы; среди пауков встречается
много ядовитых видов. Из ракообразных наибольших успехов достигли десятиногие
раки: крабы обитают у воды и в тропических лесах, а креветки отчасти замещают
пресноводных рыб в озёрах и реках. Речные раки населяют реки от холодных истоков
до тёплых заболоченных устьев. Моллюски достигают наибольшего числа видов в
лесах и в пресных водоёмах, но среди них есть и виды, приспособившиеся к жизни
в пустыне. Некоторые пресноводные двустворчатые моллюски способны переносить
пересыхание водоёма в течение нескольких лет подряд.
Характерной группой тропических беспозвоночных являются наземные пиявки, обитающие
в лесистых местностях. Они особенно обильны во влажных тропических лесах на
севере материка и в болотах долины Муррея.
Обзор написан Семёном, участником форума
(с дополнениями)
 |
 |
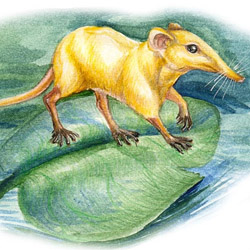 |
Bradyphalanger scandens |
Carnopossum heliarctiodes |
Nanantechinus aquadromeus |
 |
 |
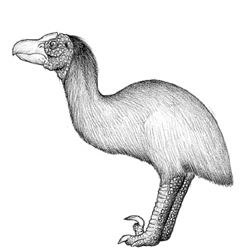 |
Phalangeropsis lemuroides |
Ancistrotherium radulus |
Carnocasuarius yagil |
 |
 |
 |
Dracovaranus horridus |
Carcharhinus viridis |
Dormicyprinus robustus |
| Меганезия, фауна саванны | |
| Меганезия, река Муррей | |
| Пустыни юго-запада Меганезии | |
| Голоса в ночи | Дождевые леса на севере Меганезии. Ночные виды животных разных ярусов леса. |
| Сон об ушедшем море | Мангровый лес на севере Меганезии, берега Арафурского озера. |
| Озеро Карпентария в Северной Меганезии, животные побережья, открытой воды и обитатели окружающего леса. | |
| Жизнь Зоораптора | Мегафауна саванн Меганезии: млекопитающие и птицы. |
| Сто дней для жизни | Сезонный водоём на юго-западе Меганезии, животные и растения. |
| Мангровые леса Арафурского озера в Меганезии - наземные и водные улитки, птицы и другие животные. | |
| Запретный плод | Тропический лес на севере Меганезии, взаимоотношения между местными животными и растениями. |
| Сад радостей земных | Леса Южной Меганезии, их обитатели - хищники и их добыча. (автор - Тим Моррис) |
| Драконы залива Эйр | Крупные рептилии залива Эйр на юге Меганезии
и их путешествие для кладки яиц. (автор - Bhut) |
Новая Зеландия
(Ао-Теа-Роа) |
Неоценовая география Новой Зеландии мало отличается от голоценовой.
Ао-Теа-Роа продолжает оставаться изолированным архипелагом из двух крупных
островов, Северного и Южного, разделённых проливом Кука. Их береговая линия
сильно изрезана, на Южном острове есть фьорды. У берегов располагается множество
более мелких островков. В ледниковый период они соединялись с главными островами,
но после его окончания вновь отделились от них. Вдали от берегов Новой Зеландии
располагаются несколько довольно крупных островов: архипелаг Чатем на востоке,
острова Антиподов на юго-востоке, острова Окленд на юге и некоторые другие.
Из-за движения литосферных плит Новая Зеландия немного сместилась на северо-восток.
Архипелаг в полной мере ощущает на себе процесс сжатия Тихого океана: в Новой
Зеландии периодически случаются землетрясения, а из-под земли в некоторых местах
вырываются гейзеры. Есть несколько действующих вулканов.
Рельеф Новой Зеландии гористый. Большая часть Новой Зеландии лежит на высоте
более 100 метров над уровнем моря. Самая крупная горная цепь, пересекающая
об острова – Южные Альпы. На вершинах гор Южного острова располагаются ледники,
но их площадь намного меньше, чем в голоцене – это следствие более тёплого
климата. Помимо гор и возвышенностей, рельеф Ао-Теа-Роа представлен обширными
долинами. Характерно наличие большого количества пещерных систем.
Климат архипелага более тёплый, чем в эпоху человека. Северный остров Новой
Зеландии и большая часть Южного лежат в зоне субтропического климата. На юге
Южного острова располагается зона умеренного климата. Огромное смягчающее влияние
на климат оказывает океан. Благодаря его воздействию в Новой Зеландии почти
не бывает заморозков (кроме юга Южного острова) и летней жары, а температура
воздуха меняется в течение года незначительно. В горных районах климат становится
суровым – здесь лежит зона альпийского климата. Для Новой Зеландии характерна
повышенная влажность: влагу приносят западные ветра. Южные Альпы преграждают
путь части воздушных масс, поэтому восток Новой Зеландии получает меньше влаги,
чем запад.
Реки Новой Зеландии в основном короткие и полноводные, они спускаются с гор.
Также на островах много озёр, образующихся в горных долинах.
Обзор написан Семёном, участником форума.
Растительность Новой Зеландии продолжает сохранять черты уникальности,
несмотря на огромное воздействие, которое оказала деятельность человека в историческую
эпоху. Число эндемичных таксонов в новозеландской флоре заметно снизилось в
эпоху человека, что связано с изменениями в фауне островов: появились растительноядные
наземные млекопитающие, и преимущество в борьбе за существование получили виды,
лучше приспособленные к вытаптыванию и поеданию ими.
На Северном острове сложился комплекс более теплолюбивой растительности. На
западной стороне островов, получающей больше влаги, господствуют леса из голосеменных
растений – каури (агатиса) и подокарповых. Эти растения выживают за счёт долговечности
и большого количества семян – значительная часть подроста гибнет из-за млекопитающих,
а цветковые растения конкурируют с ними на нарушенных участках леса. На ветвях
крупных деревьев разрастаются многочисленные эпифитные папоротники и мхи. Из
потомков туземных растений в лесах встречаются крупные древовидные представители
рода Pseudopanax, а также фуксии – потомки местного вида Fuchsia excorticata.
В небольшом количестве встречаются представители рода Hebe – древовидные вероники;
их многообразие, однако, заметно снизилось. В нижнем ярусе леса, особенно на
участках повреждённого древостоя, разрастаются клематисы – лианы, достигающие
большой высоты. Эти потомки завезённого человеком вида делают лес непроходимым,
а их листву могут поедать лишь некоторые насекомые. Пальмы представлены единичными
видами.
На более сухих восточных сторонах островов в лесах меньше разнообразие эпифитов,
а древостой более разреженный. Здесь развивается довольно высокий подлесок,
состоящий из кустарников.
Выше в горах леса с преобладанием потомков туземных видов сменяются сосновыми
лесами, образованными потомками интродуцированной монтерейской сосны. В подлеске
таких лесов разрастается ежевика местных видов – также потомок вида, нехарактерного
для исконной флоры. На сухих и солнечных северных склонах гор образуются леса
из акаций, являющихся потомками завезённых из Австралии видов. Во влажных долинах
и в руслах рек преобладают сообщества, образованные видами нотофагуса (южного
бука) и древовидными папоротниками.
В высокогорьях господствует растительность альпийского типа – низкорослые подушковидные
растения и ползучие кустарники. Здесь произрастают карликовые сосны с ветвями,
распростёртыми по земле, и выносливые лиственные растения, включая виды Hebe,
вытесненные сюда потомками интродуцированных видов. На альпийских лугах преобладают
злаки и многолетние виды люпина (потомки Lupinus polyphyllus). Многообразие
и успех в борьбе за существование интродуцированных бобовых связан с наличием
опылителей из числа общественных ос, завезённых человеком.
Болотистые местности Северного острова заселяет кордилина местных видов, успешно
конкурирующая с потомками завезённых видов растений. В тени на влажных местах
образуются густые заросли местных видов традесканции, потомков завезённой Tradescantia
fluminensis. Некоторые виды традесканции с Ао-Теа-Роа стали водными и полуводными
растениями. Южнее и в горах, в области теплоумеренного климата, обычны заросли
ив, потомков завезённых человеком видов.
В устьях рек и по берегам островов распространены заросли пандануса, в целом
обычные для тропиков Старого Света и Океании.
Южный остров отличается несколько более прохладным климатом: в целом субтропический
климат на крайнем юге переходит в теплоумеренный. Соответственно, состав растительности
несколько иной. В частности, в лесах значительно меньше эпифитных папоротников,
но разнообразнее мхи. В горах встречаются реликтовые растительные сообщества
эпохи голоцена, растительность умеренного климата – леса из нотофагуса и древовидных
папоротников. На юге острова такие леса встречаются повсеместно и образуют
обширные массивы, но древовидные папоротники предпочитают более прохладный
горный климат и обильно разрастаются в тенистых влажных ущельях, менее доступных
для травоядных млекопитающих. Также для лесов характерны древовидные Hebe с
красивыми яркими цветками, собранными в крупные соцветия. Рядом с ними произрастают
кустарниковые и древесные виды дрока с цветками белого и жёлтого цвета. Для
лесов Южного острова характерны заросли вьющихся видов жимолости с толстыми
одревесневающими стволами. В подлеске часто произрастают виды ежевики и барбариса
(потомка барбариса Дарвина, завезённого из Южной Америки).
В высокогорных районах Южного острова развиваются холодостойкие вечнозелёные
злаковники, образующие плотную дернину.
В реках по всей Новой Зеландии обычны потомки завезённых водных растений: крупные
виды валлиснерии, образующие обширные заросли путём вегетативного размножения,
эгерия и лагаросифон. Уже после человека на Новую Зеландию попал водяной папоротник
цератоптерис, но с ним успешно конкурирует местный вид мелкого плавающего папоротника
азоллы. Среди туземных гидрофитов Новой Зеландии в неоцене богато представлены
водяные мхи.
Фауна Новой Зеландии на протяжении всего кайнозоя была почти
полностью лишена млекопитающих и развивалась в изоляции. Здесь обитали многочисленные
эндемичные птицы, а также уникальные рептилии и амфибии. Человек существенно
нарушил экосистему Новой Зеландии: уже полинезийцы завезли первых интродуцентов
(крыс) и начали истреблять местных животных, но апогея этот процесс достиг
в эпоху колонизации архипелага европейцами. Неоценовая фауна Ао-Теа-Роа сохраняет
следы влияния давно вымершего человечества: некоторые эндемики полностью исчезли,
а многие животные эпохи неоцена происходят от интродуцированных видов. После
вымирания человечества продолжилось заселение островов новыми видами из Австралии/Меганезии
и Полинезии, в основном птицами. Таким образом, фауна неоценовой Новой Зеландии
состоит из трёх компонентов: малочисленные потомки аборигенных видов, обычные
и распространённые потомки интродуцированных видов и немногие виды, происходящие
от неоценовых мигрантов.
Все млекопитающие неоценовой Новой Зеландии, за исключением нескольких видов
летучих мышей, происходят от завезённых человеком видов. Крупные млекопитающие
заселяют главные острова архипелага и острова у побережья, а на отдалённых
островах они или представлены только потомками крыс, или отсутствуют вовсе.
Новозеландские сумчатые в основном являются потомками завезенного из Австралии
лисьего кузу, или щеткохвостого поссума. Одни из них являются травоядными животными,
а другие стали хищниками, образующими эндемичное семейство новозеландских хищных
сумчатых. Две эти эволюционные линии венчают гигантский медвежий кускус и сумчатый
барс. Впрочем, среди новозеландских потомков кузу есть и более мелкие, в том
числе древесные, виды, сохранившие образ жизни, близкий к предковому. Также
на островах обитают мелкие кенгуру.
Все насекомоядные Ао-Теа-Роа являются потомками европейских ежей. Среди них
есть как крошечные существа, напоминающие землероек, так и довольно крупные
животные, специализированные на питании общественными насекомыми.
Новозеландские рукокрылые представлены несколькими видами, происходящими в
основном от австралийских мигрантов и принадлежат к семейству обыкновенных
летучих мышей. Крыланов на архипелаге нет.
Из хищных млекопитающих в Новой Зеландии обитают куньи – потомки хорьков, горностаев
и норок. В основном это сравнительно некрупные звери (размером до барсука),
древесные и наземные. Один вид освоил реки и морское побережье. Самое необычное
кунье островов – таранга, аналог крота, полностью подземное млекопитающее,
приспособившееся к рытью. Среди псовых есть виды, происходящие от одичавших
домашних собак и являющиеся аналогами волков и лисиц. Стайные волкообразные
виды, однако, не достигают размеров континентального волка Голарктики и больше
похожи на коротконогого японского волка. Кошки являются потомками одичавшей
домашней кошки; это хищники небольшого размера, населяющие леса и кустарники.
Копытные Ао-Теа-Роа в основном происходят от одичавших домашних овец. Они довольно
разнообразны; самый крупный вид, тауровис, достигает размеров быка. Также в
новозеландской фауне есть потомки разных видов оленей, в то числе очень крупные.
Свиньи, потомки одичавших домашних свиней, образуют несколько видов: один полуводный
на Северном острове, лесной и горный на обоих островах, различающиеся на уровне
подвидов каждый.
Грызуны относятся исключительно к семейству мышиных (и эндемичным семействам,
производным от него). Это потомки завезённых полинезийских, серых и чёрных
крыс и домовых мышей. Они очень разнообразны: среди них есть древесные, наземные,
подземные и полуводные виды. Близкие к грызунам зайцеобразные, ведущие свой
род от кроликов и зайцев, также довольно многочисленны. Среди кроликов появился
общественный вид с развитой кастовой системой.
Новозеландские птицы отличаются большим разнообразием, они даже более разнообразны,
чем до прихода на острова европейцев. В основном это связано с тем, что завезённые
виды породили многочисленных потомков.
В горных реках и озёрах обитает реликтовый пингвин – один из последних представителей
этого отряда на Земле. К другим водяным и околоводным птицам Новой Зеландии
относятся цапли, крачки, чайки, бакланы, кулики и утки (потомки туземных и завезённых
видов). Помимо уток, на Ао-Теа-Роа имеются другие представители отряда гусеобразных,
происходящие от завезённых канадских казарок и чёрных лебедей. Чёрный лебедь
является предком очень крупной травоядной птицы с островов Чатем – бароцигнуса.
Широко распространёнными пресноводными птицами являются поганки. Один из их
видов с островов Чатем населяет морские воды. Более типичные морские птицы,
населяющие побережье архипелага – трубконосые нескольких семейств, бакланы,
олуши, крачки, чайки и несколько видов чаек-пингвинов. Также в Новой Зеландии
встречаются нырцы – морские птицы из отряда воробьинообразных, родственники
оляпок. Наиболее многочисленные колонии морских птиц расположены на отдалённых
островах, где отсутствуют хищники из числа млекопитающих.
У воды и в лесах живут многочисленные пастушковые; в отличие от островных видов
эпохи человека, они не утрачивают способность летать. Уникальное семейство
дейнораллид («ужасных пастушков») включает один вид – руакапанги, происходящий
от аборигенного пастушка уэки. Эти очень крупные птицы являются активными хищниками,
успешно конкурирующими с млекопитающими.
Куриные птицы Новой Зеландии происходят от завезённых видов, в основном от
перепелов, фазанов и американских зубчатоклювых куропаток. Некоторые из них
отличаются крупными размерами и ярким оперением.
Отряд соколообразных представлен одним видом луня, несколькими видами ястребов
(предки которых попали на архипелаг уже в неоцене из Австралии) и несколькими
видами соколов, потомков эндемичного вида и неоценовых мигрантов из Австралии.
Другие хищные птицы Новой Зеландии – совы. Неоценовые виды ведут свой род от
аборигенной иглоногой совы и завезённого домового сыча. Помимо настоящих сов,
уже после вымирания человека на острова проникли сипухи, образовав эндемичный
вид.
Полностью отсутствуют представители таких групп, как козодои, дятлы и стрижи.
Есть несколько видов кукушек и зимородков. Голуби происходят от одного аборигенного
и нескольких завезённых видов – в основном это лесные древесные птицы, питающиеся
фруктами и семенами. Попугаи довольно многочисленны, они представлены сразу
тремя семействами: настоящих попугаев, какаду и несторов. Свой род они ведут
от немногих аборигенных попугаев, а также от завезённых видов и видов, заселивших
архипелаг уже после вымирания человечества. Большинство из них – типичные попугаи
по облику и образу жизни, однако есть и исключения. На островах появился ночной
попугай, сохранивший способность к полёту. Несколько видов замещают дятлов,
приспособившись к питанию насекомыми, извлекаемыми из-под коры. Самый крупный
попугай неоценовой Новой Зеландии – орлиный кеа, потомок кеа, ставший настоящей
хищной птицей. Некоторые попугаи обитают на мелких островах вдали от побережья
главных островов. Так, на Чатеме живёт крупный наземный попугай.
Воробьиные птицы представлены наибольшим числом видов. Среди них выделяются
потомки эндемичного семейства новозеландских крапивников. Эпоху человека пережил
лишь один вид из этого семейства, но в неоцене он продолжил эволюцию и дал
начало нескольким узкоареальным видам. Часть семейств новозеландских птиц являются
общими с Меганезией, Полинезией и отчасти Азией. Это мухоловки (включая близкие
семейства, например, монархи, веерохвостки и петроики), медососы, белоглазки,
трясогузковые (в частности, коньки), славковые и некоторые другие, потомки
местных видов и неоценовых мигрантов. Ласточки достигли островов самостоятельно
ещё в XX веке.
С деятельностью человека связано существование в неоценовой фауне Новой Зеландии
таких семейств, как жаворонки, дрозды, скворцы, завирушки и флейтовые птицы.
К потомкам завезённых видов относятся также многочисленные местные вьюрковые
и ткачиковые (потомки домового воробья). Почти все врановые архипелага ведут
свой род от завезённого грача: они дали целый веер видов, напоминающих врановых
«большой земли»: воронов, соек, сорок и других. Впрочем, на Северном острове
обитает один вид врановых, происходящий от вида-мигранта из Полинезии.
По причине мягкого климата подавляющее большинство видов птиц Новой Зеландии
оседлы. Лишь единичные виды совершают ежегодные миграции. Новая Зеландия является
местом зимовки некоторых видов птиц из северных районов Азии и Северной Америки.
Рептилии Новой Зеландии относятся исключительно к отряду чешуйчатых. Черепах
и крокодилов нет, как и в эпоху человека. Прекратили своё существование клювоголовые
(гаттерии), не выдержавшие прессинга со стороны завезённых видов. Ящерицы представлены
семействами сцинков и гекконов. Новозеландские ящерицы ведут свой род от аборигенных
видов. Среди них есть как наземные, так и древесные формы. Самая крупная ящерица
Новой Зеландии – геккон-мышеед, достигающий 70 см в длину. В неоцене в Новой
Зеландии появились змеи – это потомки малоспециализированных плоскохвостых
морских змей, вторично вышедшие на сушу. Небольшое число их видов составляет
эндемичное семейство аотеарофисов.
Все новозеландские амфибии эпохи неоцена являются потомками завезённых человеком
австралийских квакш литорий. В условиях влажного и тёплого, но не жаркого климата
эти земноводные породили множество разнообразных видов. Среди них есть древесные,
водные и наземные формы, напоминающие обликом типичных квакш, жаб и настоящих
лягушек. От литорий происходит семейство новозеландских лжесаламандр – по сути,
«взрослых головастиков», утративших взрослую стадию. Лжесаламандры внешне схожи
с хвостатыми и безногими земноводными (хотя есть и совершенно уникальные формы,
такие, как батрахогимнотус) и населяют все ландшафты Новой Зеландии. Большинство
из них – не очень крупные животные. Самый крупный представитель семейства –
неоханазаки.
Примитивные аборигенные лягушки из семейства лейопельм сильно пострадали от
деятельности человека и влияния завезённых видов и не оставили потомков в неоцене.
Видовое разнообразие пресноводных рыб Ао-Теа-Роа невелико. В основном это рыбы
отряда галаксиеобразных и элеотрисовые рыбы, а также потомки морских видов,
например, камбал, иглообразных и других. В реках также обитают потомки интродуцированных
рыб: форелей и карпов, линя, речного окуня, американского кошачьего сомика.
На севере архипелага в реках обитают живородящие рыбы – потомки завезённых
гуппи и меченосцев.
Беспозвоночные Новой Зеландии довольно разнообразны. Наибольшим числом видов
представлены насекомые: бабочки, жуки, прямокрылые и другие. Многочисленны
муравьи, среди которых преобладают потомки завезённых видов. Из потомков других
завезённых насекомых следует отметить уховёрток, жуков-навозников, кровососущих
клопов и потомков сверчков. Эндемичные вета – новозеландская группа гигантских
нелетающих прямокрылых – исчезли на главных островах и на большинстве островков
вблизи их берегов. Эти насекомые остались только на отдельных небольших островках,
надёжно изолированных от проникновения наземных млекопитающих. Потомки завезённых
европейских ос образуют большие колонии, существующие в течение нескольких
лет. Пауки Новой Зеландии представлены большим числом видов. Из числа ракообразных
многочисленны десятиногие: крабы на побережье, во влажных лесах и в пресных
водах, креветки в реках и озерах. Речные раки семейства Parastacidae также
обитают в реках и озёрах, в том числе в горных районах. Во влажном климате
процветают наземные брюхоногие моллюски – улитки и слизни, потомки как туземных,
так и завезённых видов. В реках обитают многочисленные двустворчатые моллюски
и есть несколько видов пресноводных губок.
В пещерах Новой Зеландии существуют изолированные экосистемы, представленные
беспозвоночными-троглобионтами. Среди них чаще встречаются тараканы и сверчки
(в том числе мелкие реликтовые вета), но есть также жуки и пауки, многоножки
и улитки. Поставщиками пищи для них являются колонии летучих мышей; также эти
беспозвоночные питаются животными, случайно упавшими в пещеры и погибшими там.
Обзор написан Семёном, участником форума
(с дополнениями)
 |
 |
 |
Microerinaceus minutissimus |
Taranga partenogenetica |
Ursiphalanger marsupialis |
 |
 |
 |
Antipodornis ruacapangi |
Aquilopsitta horrida |
Macrogekko myophagus |
 |
 |
 |
Batrachogymnotus rivularis |
Cavernogyrinus caecus |
Scorodonodora foetidissima |
| Новая Зеландия, лесные животные | |
| Охотник из Ао-Теа-Роа | Новая Зеландия, крупные местные животные - хищники и травоядные. |
| Рыбы, лягушки и пингвины | Обитатели пресных вод Новой Зеландии - река и озеро. |
Северная
Америка |
В неоцене Северная Америка несколько сменила свое положение
из-за расширения Атлантического океана. Североамериканская литосферная плита
немного развернулась на месте (северный край – в северо-западном направлении,
южный – в юго-восточном), в результате чего разорвалась сухопутная связь с
Южной Америкой, которая снова стала материком-островом. Но на другом своём
конце материк объединился с Евразией, что привело к восстановлению Берингийского
перешейка, впервые с конца ледникового периода. На юго-западе Северной Америки
этот «перекос» отколол от континента бывшую Калифорнию, превратив её в длинный
узкий остров и образовав крутой скалистый берег, а на северо-востоке часть
бывших островов Канадского архипелага присоединилась к Гренландии, сделав этот
крупный остров ещё больше. Также Гренландия стала немного ближе к Северному
полюсу, и в северной оконечности острова господствует полярный климат с полярной
ночью и холодной снежной зимой.
После таяния ледников, образовавшихся на рубеже голоцена и неоцена, северная
оконечность Североамериканской плиты «всплыла», освободившись от колоссальной
тяжести ледника, и часть островов Канадского архипелага слилась друг с другом.
На юге материка по-прежнему существует Карибское море, усеяное островами разного
размера и отделённое от океана цепью крупных островов. Некоторые небольшие
острова, известные в эпоху человека, исчезли совсем (например, острова Флорида-Киз),
а другие объединялись и увеличивались за счёт вулканической активности и движения
малых литосферных плит. Также в Карибском море появились и новые острова. Кроме
того, в результате смещения литосферной плиты Северной Америки её узкая южная
часть, Центральная Америка, теперь направлена в акваторию Карибского моря,
сделав его ещё более мелководным. Благодаря обилию мелководий воды Карибского
моря хорошо прогреваются, что способствует процветанию прибрежной фауны и флоры.
Океанские течения образуют в Карибском море медленный водоворот; водная масса
Карибского моря относительно изолирована от океана барьером из островов и подводных
гор.
На юго-востоке материка из-за потепления климата и некоторого увеличения количества
осадков расширились территории болот, особенно ближе к побережью Карибского
моря. Переход между сушей, пресноводными болотами, солёноводными болотами и
морем в некоторых местах весьма неявный, и сложно указать, где кончается материк
и начинается Карибское море. Это территория господства тропического климата
с большим количеством осадков. Как и в эпоху человека, здесь нередки ураганы
и сильные наводнения.
Восток Северной Америки стабилен в геологическом отношении: здесь с севера
на юг тянется старый хребет Аппалачей и отсутствуют источники тектонической
активности. На востоке Северной Америки установился типичный умеренный климат,
но северная граница всех климатических зон вблизи побережья несколько смещена
к северу. Причиной этого является Гольфстрим, который в неоцене течёт практически
параллельно американскому побережью, оттеснённый к западу холодным Антигольфстримом,
текущим с севера на юг вдоль европейских берегов. Восток и запад североамериканского
материка разделены длинной полосой открытой равнинной местности – от тундры
на севере к прериям в центре материка и пустыням на юге. Там практически нет
гор, поэтому ветер с Северного Ледовитого океана может легко достичь Мексиканского
нагорья.
В центральной части материка установился типичный континентальный климат с чётким
чередованием сезонов – холодной зимы и влажного лета. Количество осадков наиболее
велико на востоке материка, постепенно снижается в центральной части материка,
резко падает на участке суши к востоку от Скалистых гор, и затем повышается
на побережье Тихого океана.
На открытых пространствах есть реки и озёра. Некоторые из них достаточно глубокие,
другие существуют лишь весной и летом, во время дождей. Из-за неравномерного
распределения осадков крупные реки имеются лишь на востоке Северной Америки,
где в историческое время располагался бассейн Миссисипи. Петляя по равнинам,
реки меняют своё течение на протяжении тысячелетий, но впадают в Мексиканский
залив, образуя дельту с множеством мелких островов. Реки с восточных склонов
Аппалачей беспрепятственно достигают Атлантики. Великие Озёра эпохи человека,
крупные водоёмы ледникового происхождения, постепенно заполнились осадком,
приносимым реками, и превратились вначале в заболоченную равнину, а затем в
сухую местность, поросшую лесом. В ледниковый период на рубеже голоцена и неоцена
в разных местах материка по окраинам ледникового щита существовало несколько
крупных озёр, но они постепенно исчезли, высохнув или превратившись в болота
в течение немногих миллионов лет раннего неоцена. На западе Северной Америки
реки появляются в Скалистых горах. Реки с восточных склонов Скалистых гор достигают
Мексиканского залива главным образом весной, когда они более полноводные. Летом
они могут почти полностью пересохнуть. Севернее, в глубине материка, реки с
восточных склонов Скалистых гор часто заканчиваются небольшими озёрами или
вовсе пропадают в пустыне. Короткие реки с западных склонов быстро достигают
Тихого океана.
Большинство озёр Северной Америки существует сравнительно недолгое время –
несколько десятков тысяч лет. Единственное крупное и долго существующее озеро
Северной Америки – огромное озеро Мише-Нама, бывший Гудзонов залив. На протяжении
миллионов лет ледники и океан вели борьбу за это место. Ледники полностью закрывали
это место на многие тысячи лет, уничтожая и вытесняя в реки всё живое, но затем
при отступлении льдов океан вновь заливал это место, принося с собой морскую
фауну. Но со временем устье залива обмелело из-за поднятия литосферной плиты
и накопления ледниковых отложений, и в неоцене Гудзонов залив разделил судьбу
Балтийского моря: он превратился в почти пресноводное озеро Мише-Нама (соленость
около 1 г/л, местами меньше), но сохранил часть потомков морской фауны.
Северо-запад неоценовой Северной Америки – это Берингия, место, где Северная
Америка сомкнулась с Евразией, образовав массивную горную гряду, которая отрезала
Северный Ледовитый океан от Тихого. В этих горах (а также в горах западной
Северной Америки и восточной Азии) течёт множество коротких порожистых горных
рек, которые впадают в северную часть Тихого океана, слегка опресняя прибрежные
воды. На равнинных участках севера материка образуется множество озёр и болот,
существующих в среднем лишь по несколько десятков тысяч лет.
Североамериканское побережье Тихого океана довольно сильно изменилось по сравнению
с эпохой человека: вдоль берегов материка тянется новый вулканический архипелаг,
немного похожий на Курильские острова. Это результат быстрого движения маленькой
литосферной плиты Хуан-де-Фука вдоль американского побережья на север.
К юго-западу горные массивы Берингии сменяются Скалистыми горами. Как и в эпоху
голоцена, там господствует достаточно мягкий и влажный климат. Горные гряды
преграждают ветрам с Тихого океана доступ в сердце материка, и значительное
количество дождей проливается в узкой полосе между берегом и горами, превращая
короткие реки в бушующие потоки. Эти же горы преграждают путь холодным ветрам
с севера. Из-за наличия Скалистых гор дождевые облака не проникают далеко вглубь
материка, и вдоль горной гряды располагаются засушливые земли – пустыни «ветровой
тени». В горных районах часты землетрясения и существует большое количество
горячих источников. Некоторые районы горных хребтов фактически представляют
собой спящие вулканы.
Дальше на юго-западе материка берега Северной Америки снова становятся крутыми
и скалистыми – это результат отделения от основного материка Калифорнии. В
этих местах более тёплые поверхностные воды смешиваются с более глубинными
и холодными, в результате чего в этих местах обычны дожди и туманы.
Местная экосистема напрямую зависит от дождей и туманов, которые приходят с
моря, и дальше на юг, где поднимается Мексиканское нагорье, которое лежит выше
полосы осадков, раскинулась настоящая пустыня, ставшая преградой для миграции
южных видов на север. Остров Калифорния также лежит в области сухого климата;
его растительность получает значительную часть влаги из туманов. Ещё дальше
к югу, где находится полуостров Юкатан, климат снова становится очень
влажным и тёплым.
Обзор написан при участии Bhut'а, участника форума.
Североамериканский материк наглядно демонстрирует смену типов
растительности в широтном направлении. На севере материка Гренландия представляет
собой царство полярного климата и соответствующего типа растительности. В северной
оконечности Гренландии сохранился ледниковый покров, вокруг которого располагается
зона тундровой растительности – вечнозелёные кустарнички и травянистые растения,
а также карликовые ползучие деревья. Южнее располагается область сухих северных
лугов, образованных осоками и злаками. Как и в Евразии, в Северной Америке почти
не осталось районов, где встречается многолетняя мерзлота – своеобразным «заповедником»
этого явления стала Гренландия, но и здесь влияние тёплого Гольфстрима отодвигает
область вечной мерзлоты от берегов.
На островах Канадского архипелага в прибрежных районах распространены холодные
северные болота с соответствующим типом растительности, а на возвышенностях
произрастают леса из выносливых и холодостойких хвойных деревьев. Значительную
часть побережий занимает литоральная полоса с зарослями крупных бурых водорослей.
Особенно обильно они разрастаются вдоль атлантического побережья материка,
где тёплый Гольфстрим поддерживает нужную температуру и препятствует замерзанию
моря. Обширные заросли бурых водорослей распространены также на тихоокеанском
побережье Северной Америки.
Южнее протягивается пояс хвойных лесов. В Северной Америке меньше крупных травоядных
млекопитающих, чем в Азии, поэтому леса более однородны по составу и в них
меньше вкраплений лиственных пород. Лиственные деревья встречаются главным
образом вдоль берегов рек и на болотах, где почва слишком влажная для роста
хвойных.
Пояс смешанных и лиственных лесов очень узок на северо-западе материка – сказывается
влияние цепи Скалистых гор, образующих «дождевую тень» и препятствующих проникновению
дождей с Тихого океана вглубь материка. Здесь же произрастают самые холодостойкие
виды деревьев. В направлении с северо-запада на восток и юго-восток пояс лиственных
лесов расширяется. Побережье озера Мише-Нама окружено смешанными лесами с преобладанием
лиственных пород – его водная масса немного смягчает и выравнивает климатические
условия в окрестностях, а также создаёт повышенную влажность воздуха, благоприятную
для развития травянистых растений. Берега озера Мише-Нама поросли тростниками,
образующими непроходимые заросли.
Своеобразный тип растительности складывается на тихоокеанском побережье, на
узкой полосе между Скалистыми горами и океаном. Здесь большое количество осадков
и ровный климат способствовали образованию настоящих северных «джунглей» –
лесов, составленных более теплолюбивыми видами деревьев, чем те, что растут
за горами. Здесь произрастают тсуга, пихта, ель, а из лиственных деревьев –
разнообразные виды клёнов и тополей. На крупных деревьях целыми массами развиваются
эпифитные мхи и папоротники, а растительность подлеска представлена кустарниками
и немногочисленными травянистыми растениями. Леса такого типа могут расти только
под прикрытием гор, которые заслоняют их от холодных северных ветров. Дальше
на юг, где горы не такие крутые, а их восточные склоны более открыты, климат
гораздо более сухой и умеренный, а зимы более холодные, чем на территории бывшей
Британской Колумбии. Здесь произрастают разреженные леса из листопадных видов
дубов, постепенно сменяемые кустарниками (скрэбом).
Вдоль хребтов Скалистых гор протянулся пояс равнин, лишённых древесной растительности.
Это североамериканские прерии, места произрастания многочисленных видов злаков
и травянистых растений других семейств. Весной прерии расцвечиваются яркими
красками, когда зацветают многолетние корневищные или луковичные эфемероиды,
но к лету яркость красок сменяется многообразием оттенков зелени злаков. На
юго-западе материка между горами и прериями вклиниваются участки полупустынь
и сухих бесплодных пустынь, где произрастают лишь немногочисленные виды засухоустойчивых
трав, а также суккулентные растения. В долинах рек, периодически заполняемых
водой, образуются редкостойные колючие леса из пустынных древовидных растений
– преимущественно из кактусов и древовидных потомков некоторых травянистых
растений.
Скалистые горы демонстрируют типичную высотную поясность: близ ледников на
их вершинах встречаются растения типа тундровых, ниже их тянется пояс альпийских
лугов с разнообразным видовым составом, сменяемый хвойными лесами. В умеренных
широтах в предгорьях произрастают смешанные и лиственные леса, а в субтропиках
и тропиках – чаппараль и даже вечнозелёные леса.
Аппалачи – старые горы на востоке материка. Они не столь высокие, и высотная
поясность здесь выражена слабее. Склоны Аппалачей покрывают широколиственные
леса, которые составляют преимущественно клён, орех, платан, липа и каштан.
Выше в горах произрастают дубы различных видов. Дубравы также характерны для
равнинных лесов, особенно в области теплоумеренного климата с достаточным увлажнением.
На юго-востоке материка во флоре появляются теплолюбивые растения: кипарисы,
магнолии и пальмы. В некоторых местах южных лиственных лесов сохранились реликтовые
популяции гинкго, являющиеся несомненными потомками деревьев, культивировавшихся
человеком. Травянистые растения юго-востока Северной Америки разнообразны,
во флоре юга Флориды присутствует некоторое количество форм, являющихся потомками
растений, культивировавшихся как декоративные (гладиолусы, бегонии). Здесь
растут разнообразные болотные растения, в том числе насекомоядные растения
семейства саррацениевых.
В долинах рек южной части Северной Америки произрастают влаголюбивые виды деревьев,
выдерживающие затопление в течение нескольких недель во время разливов и наводнений.
Мексиканское нагорье – это родина различных суккулентных растений, преимущественно
агав и кактусов. Среди кактусов часто встречаются древовидные формы, но большинство
видов – мелкие виды с неодревесневающими стеблями. Также в пустынях встречаются
многочисленные эфемеры и эфемероиды, многие из которых обладают яркими цветками
(преимущественно представители семейства амариллисовых).
Южнее Мексиканского нагорья начинается область лесов разнообразных типов –
от редколесий, образованных вечнозелёными деревьями, до настоящего влажного
тропического леса в южной части Центральной Америки, вдоль северного берега
Панамского пролива. Здесь многочисленны лианы и эпифиты, а количество видов
древесных растений составляет сотни на гектар – два соседних дерева одного
вида могут быть разделены несколькими сотнями метров. Полог леса связан воедино
лианами, и основная биомасса такого леса сосредоточена на высоте нескольких
десятков метров. Характерными травянистыми растениями тропических лесов являются
гилоцереусы и родственные им лесные кактусы.
Для южной части Северной Америки характерно семейство бромелиевых, распространённое
до северной границы субтропической зоны. На северных границах ареала это главным
образом эпифиты, а в тропических районах среди них появляются также крупные
наземные виды. Некоторые виды встречаются в составе высокогорных растительных
сообществ.
Фауна Северной Америки целиком относится к Голарктической
области (Неарктический отдел Голарктики). Тем не менее, здесь заметно значительное
влияние Неотропической области, к которой относится территория Южной Америки.
Среди млекопитающих есть несколько групп неотропического происхождения,
одна из которых – сумчатые. Несмотря на бедность видового состава, североамериканские
сумчатые не стоят на грани вымирания. В эпоху человека на материке существовал
лишь один вид опоссумов, который получил значительные преимущества в выживании
благодаря способности сосуществовать с человеком. Неоценовые сумчатые более
разнообразны – они занимают различные экологические ниши и распространены гораздо
шире, чем в голоцене. Сумчатая гиена, крупный наземный вид сумчатых, обитает
в пустынях на юге материка, а другой наземный вид, геопоссум, обитает в приполярных
районах на севере материка, на островах Канадского архипелага и в Гренландии.
Наземные опоссумы также обитают в лесах на востоке материка, но более обычны
лазающие виды; некоторые из них сравнительно мало изменились по сравнению с
предком эпохи человека.
Кроты типичного облика обычны в лесах и на равнинах материка; в неоцене среди
кротов появился крупный полуводный вид – кондилютра. Ежей в Северной Америке
по-прежнему нет, несмотря на наличие Берингийского перешейка. Землеройки обычны
и разнообразны. Во время ледникового периода на рубеже голоцена и неоцена некоторые
их виды проникли через Флориду на острова Карибского моря. На тихоокеанском
побережье Северной Америки встречается несколько видов эндемичного семейства
землероек-прыгунов – мелких плотоядных млекопитающих. Другие представители
этого семейства населяют пустыни и прерии материка.
Северная Америка – родина последних представителей семейства лошадиных. Появившись
на этом материке, лошади покинули свою прародину и вновь вернулись в Новый
Свет лишь благодаря деятельности человека. В прериях и полупустынях Северной
Америки обитают азиногиппусы, потомки одичавших ослов.
Полорогие Северной Америки постепенно исчезли, вытесненные другими животными.
Потомки пекариевых в неоцене достигают значительного видового разнообразия
и занимают различные экологические ниши. Один из видов достиг гигантских размеров
и стал экологическим аналогом бизонов. Родственные виды стали всеядными двойниками
кабанов в лесах умеренного и холодного климата. Среди пекариевых Северной Америки
также появились полуводные растительноядные формы, обитающие в болотах юго-востока
материка, и один вид неспециализированных падальщиков, аналог хищных свиней
Евразии.
С пекариевыми соседствуют другие копытные. В прериях обитают бегающие олени
рапидоцервусы; некоторые их виды приобрели грацильное телосложение, как у газелей,
и населяют полупустынные районы материка. На севере материка в полярных районах
обитает косорог, специализированный потомок северных оленей, приспособившийся
к жизни в условиях холодного климата.
Результатом связи Старого и Нового Света через Берингийский перешеек является
обитание на северо-западе и севере материка умингмака – единственного представителя
титанолагид в Америке. Зайцелопы в Северную Америку не проникли.
Неполнозубые являются типичной группой млекопитающих Нового Света. Потомки
девятипоясного броненосца распространились к северу от Мексиканского нагорья,
занимая разнообразные экологические ниши, вплоть до полуводных форм на юго-востоке
Северной Америки. На севере ареала североамериканские броненосцы впадают в
спячку. Южнее Мексиканского нагорья, в лесах Центральной Америки, броненосцы
обычны и разнообразны.
Фауна грызунов Северной Америки представляет собой результат причудливого смешения
голарктической и неотропической фаун. Голарктический элемент фауны грызунов
– разнообразные белки, бурундуки, суслики и сурки, а также мыши и крысы, в
том числе завезённые человеком. Разнообразны хомяки (в том числе мышевидные)
и гоферы. Некоторые североамериканские грызуны достигли гигантских размеров
– потомки древесных дикобразов иглошерстов стали наземными и расселились по
всему материку, от Мексиканского нагорья до Гренландии, образовав несколько
видов, отличающихся размерами и экологией. Нишу водных хищников на юге материка
занимают неоихтиомисы – крупные водные грызуны, аналоги выдр и каланов. Они
более разнообразны в Неотропической области, а южнее Мексиканского нагорья
проходит северная граница их ареала. Карибское море является северной границей
распространения своеобразного отряда водных растительноядных млекопитающих
альгоцетов, также группы южноамериканского происхождения. Элементом южноамериканской
фауны среди бегающих травоядных является род мар-оленей, близкородственный
южноамериканским формам. Эти животные проникли на север по Антильскому мосту
и заняли экологическую нишу антилоп и вилорогов в экосистеме прерий. На островах
Карибского моря обитают родственные формы. Материковые мары-олени населяют
прерии и полупустынные районы материка, а также пустыни Мексиканского нагорья.
Хищники Северной Америки принадлежат главным образом к семействам кошачьих,
псовых, енотов и куньих. Одними из самых крупных плотоядных являются берлы
– представители общей для Голарктики группы хищников. Куньи занимают экологическую
нишу водных хищников в областях умеренного и холодного климата, а также являются
древесными и наземными хищниками мелкого и среднего размера. На востоке материка
наземных всеядных куньих (типа барсука) отчасти вытеснили наземные опоссумы.
Многие псовые Северной Америки являются потомками койота и серой лисицы, живших
в эпоху человека. Псовые достигли разнообразия в лесах и на равнинах, среди
них есть специализированные формы (люпард). На севере обитает вахила – потомок
песца, общий вид для Евразии и Северной Америки. Еноты широко распространены
и разнообразны главным образом в южной и юго-восточной части Северной Америки,
в составе теплолюбивой фауны; на севере встречаются немногие виды. Древесные
формы хищников представлены специализированным семейством енотовых лемуров,
которое отчасти замещает в тропиках юга и юго-востока Северной Америки приматов.
Приматы обитают на территории Северной Америки в неоцене, но это потомки не
широконосых обезьян Неотропической области, а зелёных мартышек, завезённых
в эпоху человека на острова Карибского моря. С образованием Антильского моста
потомки мартышек расселились по юго-восточной части Северной Америки и образовали
несколько видов во Флориде. Реки препятствуют расселению узконосых обезьян
на запад и выходу их в тропические районы Центральной Америки.
Фауна птиц Северной Америки разнообразна и богата, и в ней прослеживается явное
сходство с фауной Евразии. Морские птицы с северного побережья Северной Америки
и Евразии сходны друг с другом, а часть видов совместно зимует на островах
Тихого океана. На островах Канадского архипелага выращивают своё потомство
олуши-киты, которые во время миграций часто заходят в мелководные бухты и кормятся
в стаях рыб, идущих в реки на нерест. На атлантическом побережье материка встречаются
плезиогагары, гнездящиеся на островах в центре Атлантики.
Тихоокеанское побережье – место гнездования многочисленных колоний морских
птиц: чистиковых, гусей и уток, а также нелетающих водорослеядных тератанатид.
Чайки обычны на побережьях всех океанов, омывающих материк; на атлантическом
побережье обитают грифовые поморники – крупные плотоядные птицы.
На озере Мише-Нама исторически сложился комплекс видов птиц морского происхождения:
карликовый вид олуш-китов, чайки, морские орлиные вороны и мелкие чистиковые.
Хищноврановые встречаются на побережье Северного Ледовитого океана (общий с
Евразией вид), в хвойных и смешанных лесах (особые североамериканские виды).
На равнинах нет аналогов евразийскому степному виду орлиных воронов, поскольку
в Неарктике их замещают бегающие катартиды. В целом катартиды – характерная
группа плотоядных птиц Нового Света, получившая особое развитие в Южной Америке.
Наряду с нелетающим видом, на территории Северной Америки живут несколько видов
катартид более типичного облика: в лесах Аппалачей, в Скалистых горах, во Флориде
и на южном побережье Северной Америки и в лесах Центральной Америки.
Среди сов Северной Америки (2 семейства: настоящие совы и сипухи) большинство
видов имеет типичный «совиный» облик и образ жизни. На Аляске и тихоокеанском
побережье водится крупный вид – сова-валькирия. В полупустынях на юге встречается
очень крупная нелетающая равнинная сова.
Среди птиц болот и открытых пространств характерно общее с Палеарктикой семейство
журавельников, включающее несколько родственных видов, различающихся по облику.
Через Гренландию эти птицы заселили острова Атлантического океана. Также для
болот Северной Америки характерны цапли различных видов и пастушковые; некоторые
цапли заселили открытые пространства и полупустыни. Особого разнообразия пастушковые
достигают в болотистых районах на юго-востоке материка и в долинах крупных
рек. В тропических болотах Центральной Америки встречается несколько видов
аистов и ибисов.
Курообразные птицы Северной Америки разнообразны и широко распространены по
всему материку. Разнообразные куриные птицы на равнинах относятся к семейству
тетеревиных. Также тетеревиные птицы населяют леса на севере и северо-западе
материка. Популяции полярных видов (потомков белой куропатки) распространены
в Гренландии и на островах Канадского архипелага. В полупустынях и пустынях
юго-запада материка, включая Мексиканское нагорье, обитает крупный нелетающий
потомок индейки, приспособившийся к быстрому бегу и занимающий экологическую
нишу страусообразных птиц. Индейковые также водятся в Центральной Америке и
на юго-востоке Северной Америки. Фазановых птиц (в узком смысле этого понятия)
в Северной Америке замещают зубцеклювые куропатки. Разные виды этих птиц достигают
веса от 150 – 200 г до 2 кг и встречаются в лесах и редколесьях умеренного
пояса, в горных лесах Аппалачей, в тёплых лесах по побережью Мексиканского
залива, а также в Центральной Америке и Флориде.
Среди голубей Северной Америки многообразие осталось приблизительно на уровне
эпохи человека. Появилось несколько эндемичных видов, в том числе на острове
Калифорния и во Флориде. Наибольшее многообразие голубей – в лесах Центральной
Америки.
Дятлообразные широко распространены в лесных районах материка; наибольшее видовое
разнообразие характерно для юго-востока материка, в области субтропических
лиственных лесов. Северные виды продвигаются на юг по хребтам Скалистых гор.
Попугаи в неоцене Северной Америки представлены значительно богаче, чем в эпоху
человека. В лесных областях на востоке материка распространено несколько перелётных
видов клинохвостых попугаев явного центральноамериканского происхождения. Некоторые
попугаи обитают в пустынях Мексиканского нагорья. В Центральной Америке попугаи
значительно более разнообразны.
Характерной группой птиц Нового Света являются колибри. В Северной Америке
существует несколько перелётных видов, один из которых долетает до Аляски.
Несколько перелётных видов встречается на востоке Северной Америки, особенно
в долинах рек. Южнее Мексиканского нагорья и во Флориде живёт несколько оседлых
видов. Стрижи особенно разнообразны в Центральной Америке, а на значительной
части материка обитает лишь несколько перелётных видов. Один из видов стрижей
пересекает Северный полярный круг и гнездится северным летом в Гренландии и
на островах Канадского архипелага.
Воробьинообразные по-прежнему являются самым разнообразным отрядом птиц, в
том числе в Северной Америке. Потепление климата позволило широко распространиться
по материку ряду групп неотропического происхождения. В северных районах материка
распространены вьюрковые, трясогузки, синицы, овсянки,
поползни, разнообразные врановые. Южнее появляются представители более теплолюбивых
семейств
– тиранновые, древесницы, танагры и трупиалы. Также с севера на юг увеличивается
видовое разнообразие птиц. Элемент влияния человека на авифауну Северной Америки
–
наличие представителей семейства скворцовых, происходящих от европейского скворца,
завезённого человеком. Их разнообразие невелико по причине конкуренции с трупиалами.
Другие потомки завезённых птиц – это американские ткачиковые, потомки воробьёв.
Они распространены преимущественно в редколесьях центральной части Северной
Америки, восточнее области прерий.
Среди рептилий Неарктики наибольшего распространения
достигли ящерицы семейства тейид (большей частью наземные формы). На юге материка
появляются представители игуановых (преимущественно древесные формы). По равнинным
и лесным районам
в области
умеренного и субтропического климата распространены веретеницевые, а южнее
– сцинки и
немногочисленные гекконы. Ящерицы эндемичного семейства ядозубов достигают
крупного размера и принадлежат к числу верховных хищников в пустынных экосистемах;
есть также несколько мелких видов ядозубов.
Змеи Северной Америки принадлежат к семействам ужеобразных змей, гадюковых
и аспидов. Южнее Мексиканского нагорья в тропических лесах обитают удавы.
На территории Северной Америки в неоцене нет крокодилов и аллигаторов. Пресноводные
черепахи Северной Америки принадлежат к нескольким семействам. К семейству
грифовых черепах принадлежит очень крупная черепаха-ловчий – массивная рептилия,
являющаяся экологическим аналогом крокодилов, которая охотится на наземных
млекопитающих и крупных птиц. Также многочисленны и разнообразны мелкие водяные
черепахи. Мягкотелые черепахи очень малочисленны и населяют только юг материка.
Хвостатые земноводные очень разнообразны и продвинулись далеко на юг. Представители
семейства скрытожаберников вымерли, но их место занимают крупные представители
семейства саламандровых. За полярным кругом водится серпентина – крупное гренландское
земноводное. Разнообразные саламандры и амбистомы обитают во влажных лесах
на тихоокеанском побережье. Южнее многообразие хвостатых земноводных падает,
но на территории Флориды водится много видов карликовых саламандр. В реках
умеренного пояса распространены представители семейства сиренов. Среди хвостатых
земноводных есть несколько неотенических форм, преимущественно горных и пещерных.
Бесхвостые земноводные Неарктики представлены настоящими лягушками, жабами
и квакшами. В Центральной Америке обитают также некоторые представители свистунов.
Рыбы северной части Северной Америки близки к азиатским. Характерны щуки, небольшое
количество лососевых рыб и корюшки. В умеренной зоне также встречаются пресноводные
колюшкообразные. Карповые рыбы Северной Америки представлены потомками карпа,
завезённого человеком и разводившегося в промышленных масштабах, а также небольшим
количеством мелких местных родов. Разнообразие чукучановых рыб меньше, чем
в эпоху человека – это связано с вымиранием ряда видов рыб в эпоху антропогенного
прессинга. Среди немногих выживших видов чукучановых, однако, имеется гигантский
растительноядный вид. В неоценовой фауне Северной Америки обычны харациновые
– потомки астианакса, распространившиеся на север после окончания оледенения
на рубеже голоцена и неоцена. По сравнению с неотропическими хараковидными
североамериканские виды менее разнообразны. Южнее Мексиканского нагорья, в
водоёмах Центральной Америки, обитают многочисленные хараковидные рыбы. Сомы
аборигенной фауны представлены разнообразными видами кошачьих сомов, варьирующими
по размерам от гигантов длиной до 2 метров до карликов длиной около 10 см.
Во Флориде имеются потомки завезённых человеком лорикариевых и клариевых сомов.
Центральноамериканские сомы немного разнообразнее, среди них имеются лорикариевые
и малочисленные непаразитические ванделлиевые. В устьях рек встречаются ариевые
сомы, один крупный вид этого семейства населяет Карибское море.
Среди окунеобразных рыб характерны центрарховые (ушастые окуни) и окунёвые.
На юге материка разнообразны цихлиды, часть из которых является потомками рыб,
обитавших в естественных условиях в Центральной Америке. В отрыве от общего
ареала цихлид несколько видов этого семейства обитает в реках Флориды и на
островах Карибского моря – это потомки цихлид, завезённых человеком. Характерными
обитателями Центральной Америки и южной части Северной Америки являются разнообразные
живородящие рыбы. Они особенно многочисленны южнее Мексиканского нагорья, где
сильнее сказывается влияние Неотропической области. Здесь, в бассейне Карибского
моря, находится центр разнообразия этой группы и появилось несколько семейств
живородящих. Среди живородящих рыб возникли специализированные формы – хищники,
растительноядные, фильтрующие потребители фитопланктона. В прибрежных водах
обитают специализированные представители семейства вертикалихтов, обитающие
как в морской, так и в пресной воде. Среди прочих рыб южной части материка
характерны акулы и скаты, заходящие в пресную воду, а также камбалы и иглобрюхообразные.
Среди беспозвоночных Северной Америки характерны разнообразные насекомые, сходные
с видами из Евразии (особенно на северо-западе материка). Фауна насекомых юга
и юго-востока материка включает большое количество видов субтропического и
тропического происхождения. Среди пресноводных ракообразных характерны представители
семейства американских речных раков (Cambaridae). В реках южной и юго-восточной
части Северной Америки обитают пресноводные креветки, а на болотах и в устьях
рек – крабы, в том числе лишь временно живущие в пресной воде. Для озера Мише-Нама
характерно значительное количество пресноводных потомков морских ракообразных
и моллюсков. Влажные леса тихоокеанского побережья Северной Америки населены
многочисленными видами улиток и слизней, часто достигающими крупного размера.
В Центральной Америке обитают пресноводные улитки семейства ампулляриевых.
 |
 |
 |
Aepythizon ursinus |
Condylutra ferox |
Hydrotatu potamophilus |
 |
 |
 |
Neohyaenognathus macrodens |
Asinohippus pseudoequinum |
Heliocervus heliocornis |
 |
 |
 |
Deinostrix sphinga |
Helodermopsis monstrosus |
Virolimax stridor |
| Пустыня Мексиканского нагорья, хищники и травоядные | |
| Равнины центральной части Северной Америки | |
| Гренландия, равнины | |
| Мелкие обитатели влажного субтропического леса на тихоокеанском побережье Северной Америки. | |
| Дом Зелёного Пера | Северо-восток Северной Америки, смешанные леса в области умеренного климата. |
| Лес в море | Мангровые заросли Флориды, водные и лесные животные. |
| Тайные связи | Мангровые заросли Флориды, паразиты местных видов животных. |
| Закат династии | Северная Америка, Каскадные горы - обитатели горных долин, травоядные и хищники. |
| Год гуся-путешественника | Северная Америка - обитатели различных природных зон. Миграция перелётных птиц. |
| Тропические леса Центральной Америки, Юкатан. Колонии муравьёв и их взаимодействие с окружающим миром | |
| Взаимодействие между разными видами муравьёв, их симбионтами и паразитами в тропических лесах Юкатана. | |
| Таёжная зона Северной Америки: крупные животные, их паразиты и симбионты. |
Южная
Америка |
В неоцене Южная Америка отделилась от Северной, и Панамский
перешеек перестал существовать – это произошло в результате расширения Атлантического
океана. Этот материк образован двумя литосферными плитами – Южноамериканской
(образующей большую часть территории материка) и Наска (находящейся под водами
Тихого океана и уходящей краем под Южноамериканскую плиту). На связь Северной
и Южной Америки оказывает влияние движение двух небольших литосферных плит
в Центральной Америке – плиты Кокос, образующей значительную часть Панамского
перешейка, и Карибской плиты, несущей крупнейшие острова Центральной Америки
и образующей дно Карибского моря. К крайнему югу Южной Америки примыкает небольшая
плита Скотия, границы которой отмечены несколькими субантарктическими островами.
В неоцене южноамериканский континент сдвинулся к юго-западу, но в целом его
география осталась вполне узнаваемой, несмотря на время, прошедшее с окончания
эпохи человека. Значительная часть материка представляет собой равнинную местность
с немногочисленными старыми горными массивами, подвергшимися сильному выветриванию,
и лишь на западе тянется высокий массив Анд – молодых и геологически активных
гор. Здесь извергаются вулканы, а горы постоянно поднимаются и в неоцене их
средняя высота больше, чем в эпоху человека. Это связано с процессами движения
литосферных плит – плита Наска погружается под Южноамериканскую, образуя глубокий
Перуанский жёлоб, и это является основной причиной вулканической активности
в Андах.
На севере ещё существует небольшое Гвианское плоскогорье, но оно сильно разрушено
выветриванием. В восточной части материка более крупное Бразильское плоскогорье
является водоразделом между двумя крупнейшими речными системами материка.
Севернее Бразильского плоскогорья располагается крупный речной бассейн, бывший
ранее бассейном реки Амазонки. Эта местность сложена легко размываемыми речными
наносами с Анд, и реки постоянно меняют своё течение. В неоцене самая полноводная
река Южной Америки, Амазонка, стала намного меньше из-за разделения её бассейна.
Северная часть бассейна Амазонки теперь образует самостоятельную реку – Гипполиту.
Обе реки завершают свой путь к океану примерно на одном участке побережья,
где образуется обширная болотистая дельта, выдающаяся в море. Между бассейнами
этих рек существует зыбкий водораздел, сложенный легко размываемыми отложениями,
поэтому между Амазонкой и Гипполитой время от времени образуется водообмен,
приводящий к смешению фаун водных животных. Южнее Бразильского нагорья располагается
бассейн Параны. Он не сообщается с бассейнами Амазонки и Гипполиты.
На западе Южной Америки рек мало и они очень короткие. На значительной части
тихоокеанского побережья Южной Америки реки, стекающие с гор, теряются в пустынях
у их подножия.
В Южной Америке существует большое количество озёр, но все они достаточно молоды
и невелики по размерам. Эти озёра представляют собой бывшие участки речных
русел, которые быстро (в геологическом смысле) заполняются осадком, заболачиваются
и бесследно исчезают. Многочисленные озёра в Андах также существуют сравнительно
недолго. Исключением является высокогорное озеро Тотора, расположенное на плато
Альтиплано. Это озеро ледникового происхождения, крупнейшее в Южной Америке,
было значительно «омоложено» ледниками, которые уничтожили существовавшее на
этом месте озеро Титикака и в дальнейшем сгребли пласты отложений с плато Альтиплано.
Обширные заболоченные территории имеются на севере материка, в бассейне Амазонки
и Гипполиты, а также на крайнем юге Патагонии и на Огненной Земле.
Области сухих равнин располагаются в бассейне Параны и далее на юге, в Патагонии.
Образованию таких областей всецело способствует горная цепь Анд, образующая
области «дождевой тени», перекрывая путь дождевым облакам с Тихого океана.
К западу от Анд протягивается узкая полоса суши, где влажный и прохладный климат
сменяется с юга на север жарким и засушливым.
Южную Америку окружает большое количество островов. На севере материка крупнейшими
островами являются Тринидад и острова из группы Малых Антильских. К западу
от материка лежат Галапагосские острова, площадь которых заметно увеличилась
в связи с вулканической активностью. Но наибольшее количество островов располагается
в южной части материка. В Тихом океане это архипелаг Хуан-Фернандес и многочисленные
острова вдоль западного побережья материка. В Атлантике острова немногочисленны,
и лишь на крайнем юге располагаются Фолклендские острова. На юге материка по-прежнему
существует архипелаг Огненная Земля, а также появилось немало вулканических
островов в проливе Дрейка – это след перемещения Антарктиды в сторону Меганезии.
В связи с перемещением материка климатические пояса сдвинулись с юга на север,
но это в значительной степени компенсируется общим потеплением в неоцене. На
севере материка влажно и жарко, как в эпоху человека – здесь господствует экваториальный
климат. Крайний юг материка, напротив, отличается сезонным и умеренно холодным
климатом. Лето здесь стало жарче, чем в эпоху человека – это связано со смещением
Антарктиды из полярного положения. Но жара сочетается с высокой влажностью
воздуха, образующейся из-за преобладания западных ветров. Зимой на юге материка
выпадает снег и несколько недель держится устойчивый снеговой покров, но сильных
морозов не бывает.
На территории Южной Америки располагаются два растительных
царства – Неотропическое и Голантарктическое. Граница между ними определяется
рельефом: по горным районам холодостойкие южные виды продвигаются дальше на
север.
В неоцене значительные территории Южной Америки покрыты лесами. Это относится
главным образом к северным районам материка, к области экваториального климата.
Здесь массивы влажного тропического леса (сельвы) оказываются нарушенными только
реками, протекающими сквозь них. Влажные тропические леса отличаются высочайшим
видовым разнообразием – два дерева одного вида могут быть разделены десятками,
и даже сотнями метров сплошного леса. Характерными семействами растений здесь
являются стеркулиевые, лецитисовые и бобовые, а также пальмы. Во флоре севера
Южной Америки меньше представителей древних, реликтовых семейств, хотя встречаются
виды семейств анноновые, лавровые и миртовые. Некоторые группы деревьев сравнительно
молоды в эволюционном плане (например, древовидные сложноцветные) – это является
свидетельством деградации лесов в эпоху человека и восстановления видового
многообразия за счёт появления новых таксонов. В ледниковую эпоху на рубеже
голоцена и неоцена площадь лесов была наименьшей – они оставались только в
виде небольших участков в долинах рек, откуда в дальнейшем началось восстановление
лесных массивов.
Влажные тропические леса отличаются многообразием лиан и эпифитов. Среди эпифитных
растений характерны орхидеи, ароидные, бегонии, папоротники и мхи. Многие эпифиты
принадлежат к семейству бромелиевых, характерному для южноамериканской флоры.
Ещё одна группа эпифитов, уникальная для южноамериканской сельвы – лесные кактусы
(гилоцереусовые и рипсалисы). Среди ароидных немало лиан, в том числе крупных,
и есть очень крупные многолетние наземные травы. К числу лиан принадлежат бигнониевые,
хорошо заметные благодаря крупным ярким цветкам. Травянистые растения влажного
тропического леса включают представителей семейств ароидных, марантовых, канновых,
геснериевых, паслёновых и многих других. Эндемичными являются семейства настурциевые,
включающее разнообразные виды травянистых растений, и канновые – многолетние
корневищные травы. Среди деревьев и травянистых растений часто встречаются
мирмекофильные виды, образующие полости в стеблях и/или съедобные образования
на листьях для привлечения муравьёв, защищающих эти растения от растительноядных
насекомых. В реках экваториальной области богато представлены высшие цветковые
растения – частуховые
(эхинодорусы),
кувшинки,
в т. ч. крупные,
а
также понтедериевые (водяной гиацинт, сохранившийся с эпохи человека).
Небольшие участки тропических редколесий имеются на Гвианском нагорье.
Южнее располагается область тропических редколесий, где доминируют пальмы,
древовидные кактусы цереусы и некоторые виды деревьев из числа бобовых. Такой
тип растительности преобладает на Бразильском нагорье, чередуясь с участками
сомкнутых лесов и кустарников в долинах рек. На солнечных сухих склонах произрастают
араукарии. Здесь начинают появляться элементы южной флоры: леса араукарии произрастают
на самых высоких участках нагорья; также встречаются заросли нотофагуса.
Область тропических саванн в Южной Америке отличается от обширных безлесных
пространств северного полушария. Здесь, в бассейне Параны, по саванне протекают
реки, берущие начало в Андах и на Бразильском нагорье. Вдоль рек произрастают
кактусы, достигающие крупных размеров, а также пальмы. В областях с сезонным
увлажнением, вдали от крупных рек, разрастаются травянистые растения – разнообразные
злаки и разнотравье. Для этих мест характерны амариллисовые, переживающие засуху
в виде подземных луковиц. Также в саванне произрастают крупные одиночные бромелиевые
и агавы, являющиеся аналогами африканских алоэ. Из-за смещения материка к югу
область сухих степей и полупустынь занимает большую площадь, чем в эпоху человека,
а площадь более влажной пампы сократилась до небольшого участка южнее Бразильского
нагорья. Реки бассейна Параны, берущие начало в горах, смягчают условия жизни
в сухих степях, давая возможность расти деревьям.
Вслед за сухими степями расширилась область, занимаемая лесами теплоумеренного
климата. Обширные лесные участки появились в Патагонии. Здесь господствуют
араукарии и нотофагусы с примесью некоторых других растений (например, фуксиевого
дерева и большого количества кустарников). В связи с потеплением климатом некоторые
виды пальм продвигаются далеко на юг, достигая области кратковременных зимних
заморозков. Кактусы также расселяются на юг, причём дальше, чем пальмы. Некоторые
мелкие кактусы могут даже зимовать под снегом.
Крайний юг материка – область сезонного климата с холодной снежной зимой. Здесь
растут листопадные деревья и холодостойкие араукарии, а также вечнозелёные
кустарники, способные зимовать под снегом. Они образуют многоярусные леса с
густым подлеском.
Горная цепь Анд чётко демонстрирует явление высотной поясности. В экваториальной
области (Северные Анды) по мере увеличения высоты над уровнем моря тропический
лес сменяется полулистопадными лесами, в которых доминируют кустарники и бамбук.
Ещё выше располагается область парамос – ксерофитной растительности. А ближе
к вершинам, до границ вечных снегов, располагается пояс растительности, похожей
на тундровую – вечнозелёные многолетние травы, способные переносить сильный
холод. В Центральных Андах вместо парамос складывается комплекс растительности
пуна, где доминируют засухоустойчивые травы (злаки и осоки). Такая растительность
характерна для плато Альтиплано и других высокогорных местностей. Характерными
представителями высокогорной флоры являются кактусы, покрытые белой «шерстью»,
а в экваториальных широтах агавы и крупные бромелиевые с восковым покровом
на листьях. В Южных Андах растут араукариевые леса с примесью нотофагуса и
кустарников, с подлеском из папоротников. Выше их располагается область низкорослых
трав и ползучих вечнозелёных кустарников, а у края ледников распространены
торфяные болота. Характерными растениями этих мест являются росянки.
Фауна Южной Америки принадлежит к Неотропической зоогеографической
области.
На протяжении нескольких миллионов лет Северная и Южная Америка были соединены
Панамским перешейком, который позволял Неарктической и Неотропической областям
свободно обмениваться видами флоры и фауны. Этот перешеек существовал несколько
миллионов лет в течение раннего неоцена, но позже разорвался в результате движения
тектонической плиты Кокос со стороны Тихого океана и расширения Атлантики.
Некоторое время связь между Северной и Южной Америкой поддерживалась за счёт
существования Антильского островного моста, и некоторые виды североамериканской
фауны (преимущественно птицы) смогли расселиться таким путём в Южную Америку
(в противоположном направлении по Антильскому мосту на север проникли не только
птицы, но и различные млекопитающие).
Фауна млекопитающих Южной Америки на уровне семейств сохраняет примерно тот
же состав, что и в эпоху человека, хотя появились новые группы – семейства
и даже отряды.
Сумчатые процветают, хотя их разнообразие осталось примерно таким же, как в
эпоху человека. Среди сумчатых доминируют две группы: ценолесты и опоссумы;
кроме того, есть несколько семейств, производных от опоссумов, например, американские
куньи опоссумы. Сумчатые расселились по всей Южной Америке и обитают, в том
числе, в холодных высокогорьях и на Огненной Земле, в условиях сезонного климата
с прохладным сырым летом и холодной снежной зимой. Большинство сумчатых относится
к насекомоядным или всеядным видам, но есть хищные древесные формы (американские
куньи опоссумы) в сельве и в горных лесах, активно охотящиеся на позвоночных.
Ценолесты, как и в эпоху человека, являются аналогами землероек, но среди них
появился один вид, питающийся паразитическими беспозвоночными в шерсти крупных
млекопитающих.
Благодаря тёплому климату на значительной территории Южной Америки процветают
рукокрылые. Среди них имеются хищные, насекомоядные и растительноядные виды.
В тропических лесах водятся очень мелкие виды, питающиеся нектаром. На равнинах
обитают виды, питающиеся кровососущими насекомыми, собирающимися вокруг стад
крупных травоядных млекопитающих.
Многообразие характерного для Неотропической области отряда неполнозубых на
рубеже голоцена и неоцена значительно сократилось. Ленивцы и муравьеды вымерли
из-за изменений в окружающей среде – сокращения площади лесов и деградации степей.
Лишь броненосцы сохранили некоторое разнообразие в переходный период, и это
позволило сохраниться нескольким самым распространённым видам, которые стали
предками неоценовых форм. В неоцене среди них появилось несколько специализированных
видов. Некоторые насекомоядные броненосцы отчасти занимают экологическую нишу
муравьедов; появились крупные формы броненосцев, питающихся падалью. Есть также
роющий броненосец – аналог кротов.
Насекомоядные практически не представлены в неотропической фауне неоцена: лишь
на крайнем севере обитает несколько видов землероек. Кротов в Южной Америке
нет.
Значительные изменения произошли в отряде приматов. В Новом Свете в эпоху человека
приматы были представлены исключительно представителями широконосых обезьян
и были большей частью древесными жителями. В неоцене многие специализированные
древесные группы вымерли (например, паукообразные обезьяны), и широконосые
обезьяны эпохи неоцена являются потомками мелких видов – капуцинов, беличьих
обезьян и игрунок. Среди них появились крупные формы, проводящие значительную
часть времени на земле: часть широконосых обезьян переселилась на землю и стала
аналогами павианов (южноамериканский барбудо) и горилл (мапингуари). Узконосые
обезьяны с Антильских островов не попали на материк.
Копытные Южной Америки относительно немногочисленны. Это связано главным образом
с их низким разнообразием в эпоху человека; к концу эпохи человека значительное
количество их видов находилось под угрозой исчезновения. Тем не менее, к числу
крупнейших травоядных материка принадлежат тапиротерии – потомки реликтовых
тапиров, представителей непарнокопытных. Отряд в целом угасает, но тапиротериев
несколько видов, распространённых в лесах и на равнинах материка. Парнокопытные
несколько более разнообразны. Это представители семейства пекариевых – виды,
сохранившие характерный свиноподобный облик и обитающие в горах, редколесьях
и на равнинах.
Хищные млекопитающие – представители семейств куньих, псовых, кошачьих и енотов,
а также родственные семейства, появившиеся в неоцене. Куньи представлены преимущественно
мелкими формами и охотятся на грызунов, но в семейство входит куница-ягуар
– очень крупный и массивный вид куньих, являющийся активным хищником. Среди
енотовых распространены древесные формы, но один из их представителей – пампаназуа,
наземный вид, обитающий в саваннах. Лесные формы енотовых в раннем неоцене
дали начало особому семейству южноамериканских цепкохвостов, представители
которого конвергентно сходны с приматами и отчасти замещают их. Мелкие кошки
(главным
образом потомки домашних кошек) разнообразны и населяют леса и редколесья,
а также горы. Среди кошачьих есть группа крупных видов – кошки-рапторы, отличающиеся
особенно развитыми когтями на задних лапах. Это потомки пумы, охотники на крупную
добычу. Псовые Южной Америки – различные виды лисиц и уктен (потомков койотов).
Они распространены главным образом в горных районах, редколесьях и пампасах.
Некоторые виды лисиц живут на крайнем юге материка, в области сезонного климата
с холодной зимой.
Грызуны Южной Америки отличаются колоссальным разнообразием. С эпохи человека
их разнообразие увеличилось, но потомки завезённых крыс и мышей являются очень
малой частью общего разнообразия этой группы. Кавиоморфные грызуны достигли
своего расцвета и значительно увеличились в размерах. Некоторые из них (мары-олени,
барокавии, гигантская пака) стали аналогами копытных; очевидно, их расцветом
объясняется невысокое разнообразие местных копытных. Гигантские грызуны-ленивцы,
очень крупные виды из умеренных широт материка, демонстрируют явление конвергенции
с ископаемыми халикотериями и неоценовыми ндипинотериями. Из древесных видов
распространены древесные дикобразы, среди которых иглобок является анатомическим
и экологическим аналогом ленивцев.
Грызуны в полной мере освоили водные местообитания: ложновыдры являются представителями
особого семейства неоихтиомисов. Это средних размеров и крупные водные формы,
населяющие пресные воды и прибрежные морские воды. Один из их видов, айяпух,
обитает на крайнем юге материка, на морских побережьях.
Наряду с нетипичными грызунами существует большое количество мелких форм, чаще
всего норных или древесных.
В раннем неоцене от грызунов (нутрий) произошёл целый отряд альгоцетов – водных
растительноядных млекопитающих, похожих на ламантинов. Некоторые альгоцеты
населяют пресные воды, а другие виды – морские. К морским формам принадлежит
фолклендский паральгоцетус – крупнейший вид этого отряда.
Птицы Южной Америки демонстрируют высокое видовое разнообразие, что связано
с расширением территорий, занятых лесами. На крайнем юге материка встречается
большое количество перелётных видов, в том числе улетающих в Северную Америку
и на острова Карибского моря.
К числу эндемиков Южной Америки относятся разнообразные тинаму. Представители
этого отряда практически не покидали материк, а попытки их интродукции в эпоху
человека в итоге окончились безрезультатно. В неоцене на территории материка
обитает очень крупный страусообразный вид тинаму. Нанду вымерли по окончании
эпохи человека, и их близкие родственники тинаму заместили их.
Очень характерной группой южноамериканских хищных птиц являются катартиды.
Они стали экологическими аналогами грифов и кондоров эпохи человека, а некоторые
некрупные виды стали активными охотниками на небольших птиц. К катартидам принадлежит
акату – крупнейшая летающая птица мира, живущая на пределе лётных возможностей.
Катартиды родственны аистообразным, и некоторые аистообразные в неоцене отличаются
такими же хищническими повадками. Крупные нелетающие цапли-охотники, охотящиеся
небольшими стаями, являются экологическими аналогами волков в саваннах Южной
Америки. Большинство цапель и выпей, однако, ведёт более типичный образ жизни:
они населяют тростниковые заросли и затопляемые леса, и охотятся на рыбу, земноводных
и беспозвоночных. Также по тропическим, субтропическим и горным районам материка
распространены ибисы. Среди водных птиц характерны поганки, особенно многочисленные
на горных озёрах в Андах, а также на юге материка. Некоторые их виды утратили
способность к полёту.
Журавлеобразные птицы представлены главным образом пастушковыми
и потомками серием; некоторые из последних стали активными хищниками, экологическими
аналогами
птиц-секретарей Африки.
Хищные птицы отряда соколообразных – многочисленные виды каракар, населяющие
главным образом равнины и редколесья материка. Несколько крупных видов есть
в лесных районах материка. Есть несколько видов настоящих соколов, среди которых
волчий сокол, у которого сложилось некоторое подобие общественного образа жизни.
Куриные птицы представлены семействами фазановых и зубцеклювых куропаток;
краксы вымерли в эпоху человека. Тинаму составляют значительную конкуренцию
куриным, и зубцеклювые куропатки населяют преимущественно кустарники, редколесья
и подлесок тропических лесов. Семейство фазановых представлено потомками одичавших
кур примитивных пород.
Птицы отряда гусеобразных достигают особенного многообразия в связи с большим
количеством мест обитания. Различные виды уток распространены от тропических
болот на севере до крайнего юга материка и питаются различным кормом: от рыбы
до планктонных рачков и водорослей. На побережье материка в области умеренного
климата распространены морские лебеди – крупные водорослеядные утки.
Кулики обитают по всем морским побережьям и на берегах рек. Некоторые перелётные
виды селятся на временных водоёмах в областях с сезонно-засушливым климатом.
На горных озёрах обитает несколько эндемичных узкоареальных видов. Чайки и
крачки распространены повсеместно, в южных районах материка встречаются представители
чаек-пингвинов – род кавескаров, представленный несколькими видами. Реликтовый
вид пингвинов сохранился в Южной Америке в единственном месте – на
озере Тотора.
В связи с восстановлением и расширением площадей, занятых лесами, многообразие
лесных птиц особенно велико.
Эндемичные для Южной Америки гоацины выжили в эпоху человека, но их разнообразие
не слишком велико из-за узкой специализации. Несколько видов этих листоядных
птиц обитает в пологе леса, отчасти заменяя исчезнувших краксов. Родственные
им кукушки отличаются многообразием и преимущественно выращивают потомство
сами. Некоторые из них строят крупные общие гнёзда и совместно выращивают молодняк.
Голубеобразные птицы Южной Америки принадлежат только к семейству голубиных
и распространены преимущественно на равнинах, в горных лесах и в лесах юга
материка. Часть видов происходит от одичавшего сизого голубя, завезённого человеком.
Попугаи являются представителями только семейства настоящих попугаев. Все попугаи
Южной Америки – длиннохвостые виды различных размеров, от мелких до крупных.
В эпоху человека численность и разнообразие попугаев значительно сократились,
и практически
все крупные попугаи исчезли или их численность сильно сократилась. Неоценовые
попугаи происходят от мелких видов, сохранившихся в остатках тропических лесов
и на Антильских островах. Они разнообразны и ярко окрашены, и в неоцене попугаев
можно найти на всей территории материка. Некоторые виды, происходящие от попугая-монаха
(Myiopsitta monachus), освоили области холодного климата: в горных районах
обитает снежный попугай, а несколько других видов освоили леса крайнего юга
материка и регулярно появляются летом на Огненной Земле.
Характерной группой птиц Нового Света являются колибри. Их численность и разнообразие
восстановились после изменений в природе, которые произошли на рубеже голоцена
и неоцена. От немногочисленных малоспециализированных предков произошло много
необычных и причудливых специализированных видов (например, кривоносые колибри
бакороро и итубори). Осовидный колибри из влажных тропических лесов севера
материка для защиты от хищников точно имитирует ядовитых ос-корсаров. В лесных
и горных районах встречаются стрижи.
Тропический отряд трогонов сохранил достаточное разнообразие в Южной Америке.
Большинство видов трогонов – мелкие птицы с длинными хвостами, но есть несколько
видов размером с ворону. Область их распространения ограничена исключительно
влажными тропическими лесами, в том числе на Антильских островах.
Козодои особенно разнообразны в открытых местностях, и встречаются даже в высокогорных
районах, в пустынях «дождевой тени» вдоль хребта Анд. В тропических лесах обитают
крупные козодои (потто, семейство исполинских козодоев), питающиеся мелкими
позвоночными.
Совы принадлежат к двум семействам: настоящие совы и сипухи. Распространение
сипух ограничено тропическими и теплоумеренными районами материка, а настоящие
совы обитают в лесах до самого юга материка, а также в горных районах. Некоторые
совы гнездятся на земле, выкапывая норы или занимая норы крупных грызунов.
На равнинах обитает мелкий вид сов, питающийся насекомыми, а в тропических
лесах есть виды, специализирующиеся на охоте на лягушек и летучих мышей.
Ракшеобразные представлены многочисленными зимородками, обитающими на реках
и озёрах по всему материку (на юге зимородки перелётные). Некоторые из них
приспособились к жизни в пологе тропического леса: бромелиевый зимородок добывает
головастиков и личинок насекомых из пазух листьев эпифитных бромелий.
Дятлообразные птицы представлены разнообразными настоящими дятлами, часть из
которых родственна видам с Антильских островов. Есть отдельные специализированные
виды, питающиеся общественными насекомыми. Имеются немногочисленные
виды других семейств дятлообразных птиц – туканы, якамары, пуховки и бородатки.
Среди воробьинообразных птиц господствуют разнообразные непевчие воробьиные
нескольких семейств: котинговые, печники, тапаколовые, тиранны, муравьеловки
и некоторые другие. Они демонстрируют явление конвергенции с певчими воробьиными,
доминирующими на других материках. В эпоху глобального экологического кризиса
сохранилось достаточно большое количество групп этих птиц, что позволило им
в дальнейшем восстановить многообразие и успешно конкурировать с певчими воробьиными,
расселяющимися с севера. Среди печников интересны степные свистуны – птицы,
строящие массивные толстостенные глиняные гнёзда на земле. Тапаколовые распространены
в южной части материка, предпочитая безлесные местности; в прошлом представители
этих птиц успешно расселились в Антарктиду. Котинговые населяют тропические
леса; их громкие, часто мелодичные, но однообразные крики являются характерной
чертой влажных тропических лесов. Из певчих воробьиных в Южной Америке распространено
около двадцати семейств, в том числе танагровые, трупиалы, вьюрковые, виреоны
и прочие. Результатом деятельности человека является наличие в орнитофауне Южной
Америки ткачиковых (потомков завезённого воробья). В неоцене на территории Южной
Америки появилось семейство ласточниц – мелких хорошо летающих насекомоядных
птиц, предпочитающих жить в прохладных районах материка. Некоторые их виды на
лето мигрируют в Антарктиду.
Рептилии Южной Америки разнообразны и многочисленны, что связано
с благоприятным климатом на большей части материка. Здесь встречаются представители
отрядов
черепах, чешуйчатых и крокодилов. Среди черепах большинство видов составляют
бокошейные, ведущие водный образ жизни. Подавляющее большинство черепах мелкие,
но в бассейне Амазонки и Гипполиты обитают крупные виды с длиной панциря около
100 см. Одним из крупнейших видов черепах является гидромедуза-эласмозавр,
очень крупная рыбоядная рептилия с очень длинной шеей, почти постоянно живущая
в воде. Настоящие наземные черепахи в Южной Америке немногочисленны по сравнению
с видами Старого Света, также имеется несколько видов, происходящих от водных
черепах и вторично перешедших к наземной
жизни.
Чешуйчатые рептилии представлены несколькими семействами ящериц и змей. Как
и в эпоху человека, наиболее распространённым семейством являются игуаны; это
преимущественно древесные, наземные и полуводные формы. Очень крупная водная
ящерица аквагуана питается исключительно водными растениями и плодами, падающими
с деревьев в воду. Другое семейство ящериц – тейиды. Оно включает главным образом
наземные формы, обитающие в засушливых районах и в области умеренного климата.
Также в Южной Америке встречаются гекконы, сцинки, веретеницы.
Среди змей характерны разнообразные удавы – от мелких, обитающих на деревьях,
до крупных наземных. Среди ядовитых змей наиболее распространены гремучники
и родственные формы. В тропических районах материка распространены слепозмейки
– консервативная группа мелких роющих рептилий, обитающих в почве тропических
лесов.
Наземные кайманы, представители угасающего отряда крокодилов, являются аналогами
варанов и населяют пампасы и редколесья.
Благодаря влажному климату в Южной Америке очень богата фауна земноводных.
Хвостатые земноводные богато представлены в Северной Америке, но в Южной они
отсутствуют. Наиболее разнообразны в Южной Америке бесхвостые земноводные (семейства
жабы, свистуны, древолазы, квакши), но имеется значительное количество видов
безногих. Пиповые сохранились в небольшом количестве в бассейне Параны. Большинство
бесхвостых обитает в пологе леса, где их представители демонстрируют огромное
разнообразие окрасок и размеров. Мелкие древолазы менее разнообразны, чем в
эпоху человека, и принадлежат к нескольким чётко различающимся родам. Среди
жаб имеются крупные ядовитые формы и большое количество мелких, похожих на
древолазов яркостью расцветки и малыми размерами. В неоцене появилось несколько
новых групп бесхвостых. Представитель неотенических лягушек, жаберный головастик,
является необратимо неотенической амфибией, полностью утративший способность
превращаться во взрослую особь. Он ведёт паразитический образ жизни на жабрах
крупных рыб. Эндемичное семейство шаровидных лягушек – ядовитые амфибии из
тропических районов материка, способные сильно раздуваться.
Безногие земноводные представлены роющими и водными формами, мало изменившимися
внешне с эпохи человека.
Рыбы Южной Америки демонстрируют разнообразие, сравнимое с таковым в эпоху
человека. Значительная часть видов рыб обитает в реках бассейна Атлантического
океана; ихтиофауна рек западной части материка отличается бедностью и включает
главным образом рыб морского происхождения.
В Южной Америке сохранились двоякодышащие – один вид лепидосиренов, который
приобрёл в процессе эволюции способность образовывать защитный кокон подобно
африканским протоптерам. Сохранились араваноидные рыбы; крупные ситожаберники
являются обитателями озёр и тихих рек, питаясь фито- и зоопланктоном с помощью
развитого жаберного цедильного аппарата. Есть небольшой вид плотоядных араван,
питающийся насекомыми и мелкой рыбой.
Среди более прогрессивных отрядов рыб разнообразны сомы, среди которых есть
паразитические формы (виды из различных семейств в озере Тотора и в бассейне
Амазонки и Гипполиты). Наиболее разнообразны лорикариевые сомы, включающие
много узкоареальных видов. Представитель броняковых сомов, сом-червяк, перешёл
к роющему образу жизни во влажной лесной подстилке. Необычный сом-усоход (семейства
пимелодовых сомов) населяет глубоководные участки рек.
Хараковидные рыбы – наиболее разнообразная по количеству видов группа. Представители
хараковидных варьируют от мелких безобидных всеядных видов до очень крупных
хищников и населяют не только тропические водоёмы, но также холодные горные
реки и реки крайнего юга Южной Америки. Крупнейшим их видом является тираннохаракс,
охотящийся на крупных позвоночных животных. Центром многообразия хараковидных
рыб является северная часть материка, где встречаются представители нескольких
семейств и многочисленные узкоареальные виды.
Эндемичной группой рыб являются гимноты, распространённые в тропических реках
и озёрах. Это хищные рыбы с удлинённым телом, размерами от мелких до сравнительно
крупных, использующие электрические разряды при поиске и поимке добычи. Некоторые
гимноты ведут роющий образ жизни на илистом дне рек.
Живородящие рыбы представлены многочисленными мелкими видами, похожими на своих
предков эпохи человека; они не достигают таких размеров и разнообразия, как
в Центральной Америке и Карибском море. Несколько крупных видов живородящих
рыб карибского происхождения встречаются в реках северной оконечности Южной
Америки. В центральной и южной части материка обитают икромечущие карпозубые
– мелкие рыбы, часто однолетние, переживающие сезон засухи в виде покоящейся
икры.
Сарганы являются рыбами морского происхождения. В реках северной части Южной
Америки встречается несколько видов этих рыб, но они не являются истинно пресноводными
и для размножения уходят в море.
Цихлиды были характерными представителями южноамериканской ихтиофауны в эпоху
человека. В неоцене многообразие этих рыб осталось прежним, но бурная эволюция
харациновых вытеснила цихлид в иной размерный класс: в неоцене это преимущественно
мелкие, и даже карликовые рыбы.
Другие окунеобразные рыбы пресных вод Южной Америки – бычки и горбыли. Они
продолжительное время живут в пресной воде, но для размножения уходят в море.
Среди бычков есть несколько настоящих пресноводных видов.
В реки Южной Америки в области тропического и умеренного климата заходят морские
языки – представители камбал. Как и все камбалы, они размножаются исключительно
в морской воде достаточной солёности. На крайнем юге Южной Америки в пресных
водах живут камбалы-скалярии – своеобразная группа камбал антарктического происхождения,
которые сохранили с личиночной стадии вертикальное положение тела.
Иглобрюхи населяют главным образом реки тропических широт; это исключительно
небольшие виды. Часть их проводит в пресной воде лишь часть жизненного цикла,
но имеются настоящие пресноводные виды.
Хрящевые рыбы – скаты и акулы – также встречаются в южноамериканских реках.
В тропиках это скаты семейства речных хвостоколов и некоторые виды из числа
настоящих хвостоколов. Один из представителей речных хвостоколов обитает в
холодном высокогорном озере Тотора.
Беспозвоночные демонстрируют исключительное разнообразие, особенно в тропических
лесах. Среди насекомых примечательны очень крупные жуки – усачи и пластинчатоусые.
Часто у них очень длинные усы или причудливые выросты на голове и переднеспинке.
Очень характерными жуками тропических лесов являются светляки. Крупных размеров
достигают кузнечики и кобылки, часто отличающиеся криптической окраской и причудливой
формой тела. Среди тараканов имеются ярко окрашенные виды, а также виды, способные
плавать. Термиты строят крупные глиняные гнёзда на равнинах и в редколесьях,
а в тропическом лесу искусно маскируют свои постройки на стволах деревьев или
выгрызают полости внутри ствола. Крупные богомолы обитают преимущественно в
пологе тропических лесов и демонстрируют богатство окрасок и маскировочных
приспособлений. В лесах обитает большое количество общественных насекомых,
часто жалящих и ядовитых. К таким видам относится оса-корсар, нападающая даже
на мелких позвоночных и строящая в кронах деревьев большие гнёзда. Имеются
потомки медоносной пчелы, завезённой человеком. Муравьи включают много крупных
бродячих видов, а также виды, вступающие в симбиоз с растениями. Также имеются
многочисленные муравьи-листорезы. Бабочки демонстрируют разнообразие форм,
окрасок и размеров – от невзрачных и мелких молей до очень крупных морфид,
парусников и нимфалид. Некоторые их виды питаются исключительно жидкостью,
выделяющейся из разлагающихся трупов позвоночных. На равнинах встречается много
видов кровососущих двукрылых, особенно вблизи водоёмов. В лесах юга материка
многообразие насекомых ниже, но некоторые из них достигают высокой численности:
мошки и комары размножаются в холодных моховых болотах и небольших озёрах.
Крупные многоножки сколопендры обитают в тропической части материка. Крупные
размеры и сильный яд позволяют им нападать на мелких летучих мышей, птиц и
рептилий. Некоторые сколопендры отличаются очень яркой окраской.
Среди пауков характерны наземные птицееды и пауки-волки. В пологе леса обитают
крупные кругопряды, ловящие в огромную паутину даже мелких птиц.
Ракообразные в пресных водах Южной Америки представлены многочисленными видами
креветок и пресноводными крабами яркой расцветки. На юге материка в реках обитают
крупные изоподы, очевидно, морского происхождения. В тропических лесах в почве
водятся многочисленные мокрицы.
Пресноводные улитки тропических рек достигают крупных размеров. Некоторые их
виды перешли к хищничеству и поедают других улиток, мелких рыб, головастиков
и падаль. Среди двустворчатых моллюсков южной части материка есть несколько
видов морского происхождения.
 |
 |
 |
Cervimara saltata |
Dromarmadillo asellinus |
Postpichisiego trachycephalus |
 |
 |
 |
Vespimimornis flammeocarbunculoides |
Bromelialcedo pseudotrochilus |
Teratopotoo venator |
 |
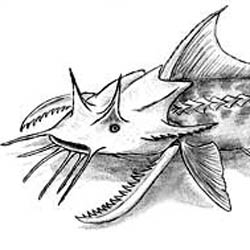 |
 |
Ziphocaiman terrestris |
Astacidoras chelopterus |
Barbambulus verticalis |
| Огненная Земля, равнины и горы | |
| Пампасы Патагонии | |
| Обитатели рек Амазонка и Гипполита (бассейн Амазонии) | |
| Сельва Южной Америки | |
| Южная Америка: неоценовая мегафауна Гранд-Чако | |
| Зеркало неба | Высокогорное озеро в Южной Америке на плато Альтиплано, и его обитатели от берегов до дна. |
| Долгий век | Южная Америка, фауна бассейна рек Амазонки и Гипполиты. |
| Пернатые эстеты | Южная Америка, птицы и другие жители полога влажного тропического леса. |
Южная Америка, беспозвоночные и позвоночные
животные влажного тропического леса. |
|
| Южная Америка, верховья реки Шингу. Обитатели полога сезонного тропического леса. |
Антарктида
и острова Субантарктики |
За время, разделяющее эпоху человека и неоцен, Антарктида
претерпела грандиозные изменения. В первую очередь они коснулись геологии:
материк сдвинулся из полярного положения в сторону Меганезии, вклинившись одним
из своих побережий в Индийский океан. Движение тектонических плит привело к
повышенной вулканической активности: вдоль побережья материка, обращённого
к Меганезии, из океана поднялись многочисленные острова, в том числе довольно
крупные. Другим очагом вулканической активности является Антарктический полуостров
(место соприкосновения Южноамериканской и Антарктической литосферных плит),
где существует несколько крупных действующих вулканов. В других местах материка
существуют потухшие вулканы.
Потепление климата и продвижение материка в умеренные широты привело к тому,
что значительная часть Антарктиды (в первую очередь её прибрежные районы) освободилась
ото льда. Впрочем, в полярной области продолжает существовать огромный Центрально-Антарктический
ледник, оказывающий существенное влияние на погодные условия. Также ледники
имеются в горах.
Из-за таяния ледяного панциря Антарктида несколько уменьшилась в размерах –
появились морские заливы, ранее скрытые шельфовыми ледниками, а некоторые участки
суши отделились от материка морскими проливами. Земля Мэри Бэрд представляет
собой крупный остров, отделённый мелководным морским проливом от Трансантарктических
гор, расположенных на сильно вытянутом участке суши, тянущемся от Антарктического
полуострова до острова Росса. Основной массив суши Антарктиды представляет
собой несколько участков плоскогорий с возвышающимися на них отдельными горными
вершинами (например, на Земле Королевы Мод). Плоскогорья окружены низменными
участками суши, в которые вдаются несколько морских заливов.
Береговая линия Антарктиды сильно изрезана, а рельеф в целом пологий, сглаженный
ледниками. Средняя высота материка над уровнем моря меньше, чем в голоцене,
из-за таяния ледяного покрова. Тем не менее, на территории Антарктиды имеются
высокие горы (высотой до 4 тысяч метров) и обширный горный массив Трансантарктических
гор, который отделяет Землю Мэри Бэрд от прочего материка. В связи со смещением
Антарктиды из полярного положения южный полюс в неоцене находится у подножия
Трансантарктических гор.
Хотя климат неоценовой Антарктики стал мягче, чем в эпоху человека, он всё
равно остаётся сезонным и очень суровым. Большая часть материка существует
в условиях смены полярного дня и полярной ночи. Вслед за коротким солнечным
и довольно тёплым летом (полярным днём) приходит суровая зима, сопровождаемая
сильными ветрами и обильными снегопадами, которые покрывают весь материк толстым
слоем снега. Летом осадки выпадают в виде дождя, но даже в середине лета холод
иногда возвращается и может ненадолго выпасть снег. Летние температуры достигают
20 градусов тепла и выше, а зимой могут опускаться ниже 40 градусов мороза.
В Антарктиде существует большое количество пресных водоёмов. Многочисленные
реки и ручьи имеют снеговое и дождевое питание, или спускаются с Центрально-Антарктического
ледника. Многие из них существуют временно, лишь в период интенсивного таяния
весной и летом. В низинах и на равнинах существуют обширные болота. Также в
Антарктиде много небольших ледниковых и термокарстовых озёр. Зимой большинство
пресных водоёмов промерзает до дна.
Очерк написан Семёном, участником форума
(с дополнениями)
Флора Антарктиды отличается бедностью – в неоцене она восстанавливалась
практически с нуля. В связи с оледенением богатая антарктическая флора раннего
кайнозоя исчезла в самой Антарктиде и лишь её остатки сохранились на территории
субантарктических островов, Южной Америки, Новой Зеландии, Австралии и Тасмании.
В неоцене, когда материк стал постепенно освобождаться от ледникового щита,
часть растений древней антарктической флоры вернулась на материк вместе с многочисленными
новыми поселенцами.
Древесная растительность в том виде, какая она существует на других материках,
в Антарктиде отсутствует. На материке произрастают виды нотофагуса (южного
бука) южноамериканского происхождения, но их жизненная форма – кустарники (в
том числе карликовые) и стланик. Также в Антарктиде произрастает несколько
карликовых видов ивы, происходящей от растений, завезённых в Новую Зеландию
в эпоху человека. Антарктические ивы имеют мелкие размеры и обильно ветвятся,
а весной их цветы служат важным источником нектара для местных насекомых. Ивы
образуют особые «ивовые луга», в период цветения сплошь покрытые пушистыми
жёлтыми соцветиями. Ивы произрастают преимущественно на влажных низинных участках
суши, тогда как нотофагусы предпочитают возвышенности.
Травянистые растения Антарктиды немногочисленны, но образуют достаточно большую
биомассу. Основу травянистой растительности составляют осоки и злаки. Из других
семейств встречаются сложноцветные, росянки, пузырчатковые. Среди росянок имеется
крупные виды, способные ловить и переваривать даже мелких птиц. Пузырчатковые
представлены не только водными видами с характерными ловчими листьями, но и
разнообразными наземными формами, в том числе достаточно крупными. Редким компонентом
антарктической флоры являются представители семейства гераниевых, южноафриканского
происхождения. Очевидно, они попали на материк в раннем неоцене, когда на побережье
Антарктиды сложился умеренный климат. Среди антарктических растений преобладают
анемохорные виды, и лишь некоторые из них утратили эту особенность уже после
того, как их предки попали на этот материк.
Для Антарктиды характерно многообразие споровых растений. Папоротники чаще
всего мелкие, с однолетними вайями и зимующим корневищем. Есть несколько крупных
видов папоротников, произрастающих на индо-тихоокеанском побережье материка
– в самых тёплых районах.
Характерной особенностью флоры Антарктиды является многообразие моховидных.
Мхи различных групп благодаря неприхотливости и способности расселяться спорами
были одними из первых растений, поселившихся на свободных ото льда местах.
На скалах ниже края ледников складываются сообщества, где главную роль играют
печёночники и листостебельные мхи. Сфагновые мхи поселяются на болотах, а в
реках разнообразны водяные мхи.
Разнообразны антарктические лишайники, образующие сообщества причудливо выглядящих
видов на голых камнях, прогреваемых полярным летом.
Бедная фауна Антарктиды эпохи человека была полностью уничтожена
ледником в конце голоцена. В неоцене, когда климат стал мягче, животные вновь
заселили Антарктиду – потомки морских животных и мигранты с других материков.
Однако разнообразие антарктической фауны невелико по сравнению с другими континентами.
В Антарктиде полностью отсутствует фауна млекопитающих. Единственным исключением
являются альгоцеты – морские млекопитающие, происходящие от южноамериканских
грызунов. Один из видов, фолклендский паральгоцетус, расселился по побережью
Антарктики. Однако эти звери никогда не выходят на сушу.
Наиболее разнообразными обитателями Антарктики из числа позвоночных являются
птицы. Отсутствие наземных хищников и богатые ресурсы моря позволяют огромным
колониям морских птиц существовать на побережье Антарктиды и на островах Субантарктики.
Основу этих колоний составляют чайки, крачки, поморники, олуши и трубконосые
(несколько семейств, в том числе лжеальбатросы). Также здесь гнездятся бакланы
разных видов, в том числе узкоареальные нелетающие. Антарктика и Субантарктика
– центр разнообразия семейства чаек-пингвинов, крупных нелетающих ржанкообразных,
являющихся экологическими аналогами пингвинов. Многочисленны морские утки,
питающиеся водорослями или беспозвоночными. На литорали добывают корм кулики,
напоминающие зуйков, песочников, камнешарок и кроншнепов. Другие кулики либо
узкоспециализированные (например, представители семейства куликов-сорок), либо
приспособившиеся к питанию падалью и отбросами. Помимо типичных морских птиц,
Антарктику населяют различные виды нырцов – морских воробьиных птиц, потомков
южноамериканских оляпок.
Наземные птицы Антарктиды уступают по численности морским. Они ведут свой род
от южноамериканских, новозеландских и меганезийских видов. Абсолютное большинство
антарктических птиц относятся к перелётным видам: они зимуют в Новой Зеландии,
Меганезии, Патагонии, на Огненной Земле и на островах Субантарктики. Единичные
виды проводят зиму в Антарктиде.
У рек и ручьёв обитают чайки и крачки, которые питаются рыбой и беспозвоночными,
в том числе стрекозами. Здесь же живёт один вид поганок, потомок тасманийской
популяции чомги. Многочисленны кулики разных семейств (в первую очередь бекасовые
и ржанковые), добывающие корм в реках и на моховых болотах. Необычными обитателями
Антарктиды являются растительноядные кулики, внешним видом и образом жизни
напоминающие куропаток – потомки южноамериканских зобатых бегунков.
Хищные птицы Антарктиды немногочисленны. На материке летом встречаются один
вид канюка и один – каракары. Они зимуют в Южной Америке и прилетают в Антарктиду
вместе со стаями водных птиц, когда побережье материка освобождается ото льда.
Берега водоёмов населяют утки разных видов, питающиеся водными растениями и
животными. Некоторые гусеобразные используют иной источник корма – вегетативные
части наземных растений. К ним относится и самое крупное травоядное животное
Антарктиды – антарктический луговой лебедь.
Наибольшего успеха в Антарктиде добились воробьиные птицы. Здесь обитают представители
подотряда кричащих воробьиных, характерного для Нового Света. Они относятся
к семействам печников и тираннов (и производному от тираннов семейству ложных
ласточек). Эндемиками Антарктики являются мелкие нелетающие птицы из семейства
мышевидок, потомки южноамериканских тапаколо. Они ведут оседлый образ жизни
и замещают отсутствующих на материке грызунов. Из певчих воробьиных Антарктику
населяют коньки, овсянки, крапивники и древесницы. На некоторых субантарктических
островах живут птицы из семейств славковых и мухоловковых (и близких семейств),
происходящие из Меганезии и Новой Зеландии. Самые крупные воробьиные Антарктиды
– представители семейства кассиков, или трупиалов. Они занимают ниши врановых
и даже хищных птиц (например, соколиный кассик). Также в Антарктиде летом гнездится
один вид жаворонков – это потомок европейского вида, завезённого человеком
в Новую Зеландию. На Новой Зеландии он зимует.
В Антарктике полностью отсутствуют рептилии и амфибии,
что связано с изоляцией и холодным климатом этого материка.
Рыбы пресных вод Антарктики немногочисленны; они представлены исключительно
потомками морских видов. Это пресноводные галаксии и камбалы, а также представители
особого семейства вертикальных камбал, встречающихся также в водах Южной Америки,
Новой Зеландии и Меганезии. Большинство рыб проводит в реках лишь тёплое время
года, а на зиму мигрирует к устью или в море. Для некоторых видов пресные воды
вообще служат лишь местом нагула, а нерест происходит в море. Впрочем, единичные
виды приспособились проводить в реках Антарктиды весь год, например, вмерзая
на зиму в лёд или закапываясь глубоко в грунт.
Антарктические беспозвоночные представлены большим числом видов, хотя их разнообразие
очень невелико по сравнению с другими частями света. Наиболее многочисленны
среди них насекомые. В условиях экстремального климата процветают примитивные
насекомые мелких размеров, например, ногохвостки. В Антарктике встречается
большое число видов стрекоз из разных семейств, причём некоторые достигают
значительных размеров. Их личинки занимают разнообразные экологические ниши
в пресных водоёмах, а личинки особой группы рыбострекоз из семейства красоток
отчасти заменяют мелких пресноводных рыб. Кроме стрекоз, в пресных водах Антарктиды
обитают водяные клопы – плавты и гребляки. Наземные клопы малочисленны, но
обладают различными пищевыми пристрастиями: есть растительноядные виды и хищники,
а также несколько паразитических кровососущих клопов, обитающих в птичьих гнёздах.
Самые массовые виды насекомых Антарктики относятся к двукрылым: комары и мошки
в
огромных
количествах
размножаются
в
пресных водах. Большинство из них является кровососущими. Среди комаров Антарктиды
появились нетипичные виды, напоминающие слепней, а некоторые стали активными
хищниками, схожими с клопами. Личинки комаров-долгоножек замещают отсутствующих
на материке дождевых червей и являются активными почвообразователями. В Антарктике
обитает несколько семейств жуков, в основном растительноядных, в частности,
листоедов. Антарктические чешуекрылые – моли и дневные бабочки – отличаются
мелкими размерами и неяркой окраской. Из общественных насекомых на материке
встречаются лишь несколько видов пчёл, образующих небольшие колонии, существующие
один сезон.
Паукообразные представлены несколькими видами пауков и клещей.
В пресных водоёмах обитают десятиногие ракообразные – креветки, успешно конкурирующие
с рыбами. От креветок происходит эндемичное семейство антарктических ложных
раков, ведущих наземный образ жизни. Фауна наземных крабов бедна: лишь на побережье
и на островах Субантарктики встречается несколько видов. В пресных водах встречаются
представители изопод.
Брюхоногие моллюски очень немногочисленны и представлены потомками морских
видов, приспособившихся к жизни в пресной воде. В реках обитают двустворчатые
моллюски.
Очерк написан Семёном, участником форума.
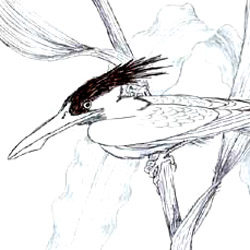 |
 |
Alcyosterna acutiptera |
Cryopodiceps australissimus |
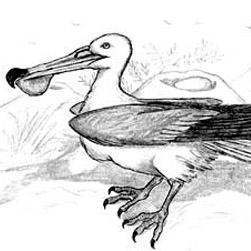 |
 |
Thalassocrator magnificus |
Barocygnus chathamensis |
| Субантарктические острова | |
| Обитатели лугов Антарктики | |
| Обитатели лугов Антарктики | |
| Завоевание юга | Природа Фолклендских островов и прибрежных вод Антарктиды. |
| Острова туманного солнца | Архипелаг Чатем, лесные и прибрежные жители. |
| Моря Южного Креста | Антарктида, обитатели морей в прибрежной зоне и вдали от берегов. |
| Острова Оркадас в Субантарктике, птицы и другие обитатели архипелага. |
Острова
Атлантического океана |
| Атлантический океан |
Исландия
|
Система разломов, пересекающая Исландию. |
Исландия является островом вулканического происхождения и
представляет собой одну из вершин Середино-Атлантического хребта, тянущегося
вдоль всего Атлантического океана в меридиональном направлении. Это остров
достаточно древнего происхождения, существовавший ещё в эпоху человека, но
расширение Атлантического океана является фактором, способствующим поддержанию
вулканической активности. На территории Исландии имеется большое количество
потухших и спящих вулканов, а наискосок острова, с юго-запада на северо-восток,
тянется участок сравнительно молодых пород – результат расширения Атлантического
океана. Видимыми проявлениями вулканической активности являются периодические
извержения вулканов и многочисленные горячие источники в долинах.
В ледниковую эпоху на рубеже голоцена и неоцена Исландия была полностью накрыта
ледниковым щитом, что привело к сглаживанию старых гор и образованию элементов
рельефа, сложенных моренными отложениями. Даже в неоцене на территории острова
заметно большое количество следов деятельности ледников, существовавших около
20 миллионов лет назад, на рубеже голоцена и неоцена. Молодой горный хребет
образовался уже после окончания ледниковой эпохи, и не несёт следов воздействия
ледников. Центральная часть острова очень гористая, а ближе к берегам рельеф
становится более сглаженным, с отдельными участками сильно выветренных скал.
Береговая линия острова сильно изрезана бухтами и скалистая.
Климат острова морской, сезонный, с высокой влажностью воздуха. Климат определяется
влиянием двух течений: тёплого Гольфстрима на западе (у побережья Гренландии)
и холодного Антигольфстрима на востоке (у европейских берегов). Лето на острове
прохладное и сырое, жарких дней очень немного. Зима морозная, снежная и холодная,
но сильных морозов не бывает из-за влияния океана.
Реки острова неглубокие, холодные и короткие; в некоторых долинах они могут
разливаться, образуя серию мелководных заболоченных озёр. На горных участках
реки бурные, часто образуют водопады. Также обычны моховые болота и небольшие
озёра в долинах рек.
Растительность Исландии имеет много общего с европейской и
американской. Во время ледникового периода на рубеже голоцена и неоцена вся
растительность острова была уничтожена ледниками, и после таяния льдов она
начала восстанавливаться с нуля. Самыми древними, пионерными представителями
исландской флоры являются мхи и лишайники. Пятна лишайников располагаются на
скалах, в том числе обращённых к морю. Заросли мхов обильны в тенистых и влажных
местах.
Леса Исландии в эпоху неоцена образованы видами деревьев, способными расселяться
при помощи ветра. Основу лесов на сухих почвах составляет сосна, близкая к американским
видам (благодаря преобладающим ветрам западного направления). В высокогорных
районах сосны образуют стланиковую форму. В более влажных местах из хвойных
произрастает ель, а также лиственные деревья – берёзы, ивы, вяз и холодостойкие
виды тополей. На хорошо прогреваемых южных склонах растут клёны американского
происхождения. Обычно клёны разрастаются в виде невысоких многоствольных кустарников,
и лишь в южной части острова они вырастают в невысокие деревья. Все лиственные
деревья на острове листопадные. Кустарниковая растительность – разнообразные
виды вереска, часто образующие сплошные заросли.
Травянистые растения представлены разнообразными видами осок и злаков с примесью
прочих видов разнотравья (семейства кипрейные, сложноцветные и другие). Весной
зацветают разнообразные луковичные растения – виды подснежников и лука. В болотах
и по берегам рек растут рогоз и тростник. Очень распространены сфагновые мхи,
образующие мощные торфяные залежи.
Фауна Исландии эпохи неоцена отличается бедностью на уровне
классов и отрядов – в ней отсутствуют или представлены лишь единичными видами
многие группы животных.
Млекопитающие Исландии представлены исключительно несколькими
видами летучих мышей. Среди них выделяется исландская крылоножка – плотоядный
вид, который плохо летает, но хорошо бегает и лазает. Это рукокрылое замещает
отсутствующих на острове мелких плотоядных млекопитающих. Прочие отряды млекопитающих
в исландской фауне отсутствуют.
Птицы представлены в фауне Исландии очень богато: в этом плане
остров немного напоминает Новую Зеландию до колонизации её человеком – птицы
занимают значительную часть экологических ниш, которые на материках занимают
млекопитающие.
Крупнейшими обитателями Исландии являются китовые олуши. Лежбища этих крупных
морских птиц имеются на нескольких пологих участках побережья, и отдельные птицы
или небольшие группы встречаются на скалистых побережьях.
Хищниками морского побережья являются грифовые поморники, или морские стервятники
– очень крупные поморники, аналоги мелких орлиных воронов с побережий Тихого
океана. На берегах острова также гнездятся бореевы фрегаты, разнообразные чайки
и крачки. Чайки обитают также на пресных водоёмах, но здесь они малочисленны.
Кулики разнообразны и населяют морские побережья, реки и болота. Некоторые виды
куликов обитают на лугах и в вересковых зарослях, питаются наземными насекомыми.
Крупные совы утбурды являются главными хищниками на острове. Они утратили способность
летать, но благодаря крупному размеру способны убить любого обитателя Исландии.
Также на острове есть несколько мелких видов сов, питающихся только птицами,
насекомыми и рукокрылыми.
Аналогом травоядных млекопитающих и новозеландских моа в Исландии является ксеномоа,
крупная куриная птица, происходящая от белой куропатки и утратившая способность
летать.
Другими растительноядными птицами Исландии являются толстоклювые гуси – крупные
перелётные птицы. Также летом на остров прилетает несколько мелких видов уток.
Часть их улетает на зиму в Европу, часть – в Северную Америку через Гренландию.
Характерными летними обитателями Исландии являются журавельники – перелётные
птицы, зимующие на островах Макаронезии (Канарские острова, Новая Азора).
Характерные воробьинообразные птицы Исландии – представители исландских крапивников,
наглядно демонстрирующие явление адаптивной радиации. Также на острове имеются
представители семейств оляпковых, трясогузок, ласточек, вьюрковых, врановых.
Часть их постоянно живёт на острове, а большинство насекомоядных видов улетает
на зимовку в Европу и Северную Америку, а также на острова Макаронезии. Также
на острове встречаются представители отряда длиннокрылых - два вида стрижей.
Один из них улетает на зимовку в Африку, другой на острова Карибского моря.
Пресмыкающиеся и земноводные отсутствуют в
фауне Исландии.
Пресноводные рыбы крайне малочисленны – в ледниковый период
на рубеже голоцена и неоцена вся пресноводная ихтиофауна, включая завезённые
виды, исчезла из-за обширного оледенения на острове. В реках острова обитают
только бычки, зимующие и нерестящиеся в морской воде. Также на откорм в реки
заходят камбалы.
Беспозвоночные заселили остров двумя путями: по воздуху и из
моря. Среди насекомых на острове многочисленны стрекозы, личинки которых являются
главными хищниками мелководных водоёмов. Среди прочих насекомых разнообразны
двукрылые: мухи, слепни, комары, долгоножки и мошки. Многие из них питаются
кровью позвоночных и массами нападают на крупных птиц острова. Бабочки представлены
семействами белянок, нимфалид, разнообразными совками и молями, преимущественно
некрупными видами довольно тусклой расцветки. Жесткокрылые Исландии – это жуки-дровосеки,
листоеды и некоторые другие семейства. Крупнейшее насекомое острова – жук твердозуб-великан,
принадлежащий к семейству листоедов. На территории Исландии нет дождевых червей
и основными почвообразователями являются роющие личинки насекомых.
В пресных водах Исландии много ракообразных морского происхождения. В устьях
рек обитают морские жёлуди, фильтрующие речной осадок. Бокоплавы живут не только
в пресных водах, но и на суше: по всему острову встречается прыгающий рачок
травяная блоха. Представитель равноногих раков речной древоточец точит стволы
деревьев, упавшие в воду.
Фауна наземных моллюсков очень бедна – имеется только несколько видов мелких
наземных улиток. Пресноводные моллюски отсутствуют, лишь некоторые морские виды
заходят в устья рек.
 |
 |
 |
Manuptera cursoria |
Xenomoa borealis |
Mundariornis subulatorostris |
| Сага о пернатых | Исландия и её обитатели |
Остров
Локи
|
Остров Локи – один из новых островов в Атлантическом океане;
он появился над поверхностью воды уже в неоцене, и его возраст составляет около
15 миллионов лет. Он является самой южной точкой подводного хребта Рейкьянес,
в северной точке которого находится Исландия. Исландия также является ближайшей
к острову Локи сушей (остров располагается к юго-западу от Исландии). По своему
положению по отношению к материкам остров Локи находится ближе к Северной Америке,
чем к Евразии.
Остров Локи – вулканического происхождения, его площадь – около 400 квадратных
километров. Он молод, и свидетельства продолжающейся вулканической активности
встречаются на нём
повсюду.
Остров
представляет собой
вулкан с несколькими кратерами – крупный основной кратер потух и одна из его
сторон обвалилась, открыв доступ океанской воде. Но на его наружных склонах
имеются несколько небольших активных кратеров, периодически извергающихся.
Побережья острова Локи скалистые, а вход в центральную бухту острова представляет
собой узкий пролив с несколькими островками. Много островков разбросано вдоль
побережья острова Локи, а окрестности действующих вулканов по окраинам острова
представляют собой отмели, сложенные чёрным песком вулканического происхождения.
Во внутренней части острова Локи имеется несколько постоянно действующих горячих
источников и небольших горячих озёр.
Климат острова Локи во многом определяется наличием Гольфстрима. Благодаря
тёплому океанскому течению он значительно теплее климата континентальной Европы
на той же широте. Лето здесь тёплое и влажное, а зима мягкая и бесснежная;
зимние температуры не опускаются ниже +10°C.
Благодаря мягкому климату остров Локи является благоприятным
местом для развития растений. Флора острова относительно молода и развивалась
преимущественно благодаря заносу семян с материков. На первых этапах расселение
растений происходило с ветром, но в дальнейшем новые виды появлялись на острове
благодаря птицам, поскольку остров находится на пути миграции птиц между Исландией
и Макаронезией.
Древесная растительность на острове – преимущественно лиственные деревья и
кустарники. Клёны и вязы – виды северного происхождения (главным образом из
Северной Америки), а деревья семейства вересковых происходят с юга, из Макаронезии.
Хвойные деревья, в отличие от соседней Исландии, довольно редки – на вершинах
острова есть небольшие группы сосен, относящихся к тому же виду, что и исландские.
На внешних склонах острова растительность более низкорослая и густая, способная
выдерживать штормовую погоду. Здесь древесные растения образуют низкорослые
формы с густой кроной, а их стволы часто полегают и укореняются.
Внутренняя часть острова, защищённая стенами кратера, поросла густым широколиственным
лесом, сохраняющим зимой значительную часть листвы. В тенистом подлеске формируется
влажный микроклимат, способствующий развитию комплекса тенелюбивой флоры. В
подлеске господствуют папоротники и многолетние травы. Благодаря влажности
и низкой конкуренции папоротники достигают почти двухметровой высоты. Возле
горячих источников они сохраняют листья круглый год. Цветковые растения представлены
главным образом сложноцветными, имеющими летучие семена с хохолками. Также
на острове произрастают несколько видов злаков (в основном на сухих склонах)
и орхидей (во влажной внутренней части острова).
На острове встречается несколько видов мхов, образующих обширные заросли во
влажных местах. Некоторые мхи являются эпифитами. Также богато представлены
лишайники; некоторые из них обладают причудливой формой.
На берегах в приливно-отливной полосе складываются сообщества галофильных травянистых
растений, выдерживающих регулярное затопление.
Настоящая островная фауна, изначально развивавшаяся без вмешательства
со стороны человека. Значительное число видов животных на острове имеет американское
происхождение – очевидно, расселение происходило через Гренландию. Мелкие виды
беспозвоночных (пауки, двукрылые, бабочки) были занесены с материка ветром.
Млекопитающие на острове Локи представлены только одним видом насекомоядных
летучих мышей.
Фауна птиц значительно богаче. На острове обитает мелкая цапля американского
происхождения, которая ищет корм в прибрежной полосе и во внутреннем заливе
острова. Своеобразным жителем острова является нелетающий кулик прибрежный
шилоклюв, обитающий главным образом на внешних склонах острова. В кратере обитает
небольшой наземный вид кулика, живущий в лесу. На берегах острова живут чайки
и чистиковые, время от времени появляются бореевы фрегаты (главным образом,
в зимнее время). В прибрежной полосе встречаются грифовые поморники. Китовые
олуши появляются редко, только во время зимних миграций, и не образуют постоянных
лежбищ.
Воробьинообразные птицы на острове Локи – наиболее разнообразный отряд, представленный
несколькими семействами. Во внутренних районах острова обитает крупный плохо
летающий вид врановых птиц, питающийся наземными животными. В противоположность
Исландии, на острове Локи существует только один вид крапивников: здесь эти
птицы не дали обширной радиации наподобие исландских видов. Разнообразны вьюрковые
– существует несколько насекомоядных и растительноядных видов. Среди перелётных
насекомоядных птиц имеются ласточки и стрижи.
Из рептилий в фауне острова имеется крупный представитель настоящих ящериц
(Lacertidae) – фафнирелла, один из немногих видов европейского происхождения.
Амфибии и настоящие пресноводные рыбы на острове Локи отсутствуют; во внутреннюю
бухту острова заходят морские рыбы.
Климат на острове Локи теплее, чем в Исландии, и фауна беспозвоночных здесь
богаче. Стрекозы очень разнообразны, их личинки обитают в ручьях и небольших
болотах. В переувлажнённой и болотистой почве живут личинки нескольких видов
слепней и мух. В лесу острова Локи обитают разнообразные жуки, преимущественно
усачи, листоеды и пластинчатоусые. Из хищных жуков имеются светляки и два мелких
вида плавунцов; один из плавунцов вторично приспособился к жизни на суше, и
его личинка также развивается вне воды.
Пауки представлены исключительно кругопрядами, родственными американским видам.
Наиболее вероятным способом их расселения на остров был разнос ветром молодых
особей на нитях паутины.
Из видов ракообразных встречаются мелкие наземные крабы семейства Grapsidae.
Они обитают в прибрежных районах и вокруг внутренней бухты острова, проводят
на суше только лето.
Наземные улитки на острове – несколько мелких видов американского происхождения.
Возможно, их предки были занесены на остров в виде яиц, приставших к лапам
птиц.
Новая
Азора
|
В неоцене из-за вулканической деятельности, сопровождающей
расширение Атлантического океана, часть Азорских островов (Пику, Терсейра,
Фаял, Сан-Жоржи и Грасьоза) превратилась в единый остров Новая Азора. Образование
острова происходило в несколько этапов
и сопровождалось
повышением вулканической активности. Последний этап формирования острова завершился
примерно через 15 миллионов лет после вымирания человечества.
Новая Азора эпохи неоцена – это гористый остров, сложенный породами вулканического
происхождения. По краям острова располагаются участки более старых, сильно
выветренных пород с отложениями вулканического пепла в низинах, отмечающими
очередные периоды вулканической активности. Центральная часть острова – высокий,
молодой и достаточно активный вулкан. На склонах этого вулкана имеется несколько
горячих источников, а отдельные участки склонов представляют собой лавовые
поля с отдельными участками пионерной растительности.
Остров Новая Азора располагается в тропических широтах, и сезонные изменения
климата выражены слабо – лишь зимой количество осадков меньше. Климатические
условия на Новой Азоре в значительной степени определяют Гольфстрим и Антигольфстрим.
Благодаря Антигольфстриму на острове очень часты туманы, особенно в зимнее
время. Летом благодаря Гольфстриму на острове высокая влажность воздуха.
Горы острова служат местами конденсации влаги, и по склонам стекают ручьи,
сливающиеся в небольшие реки. В горных долинах имеются озёра и болота.
Новая Азора как единое целое – достаточно молодой остров,
но Азорские острова как его предшественники существовали ещё в эпоху человека.
Из-за этого в островной флоре в неоцене заметно влияние деятельности человека
– среди растений острова существуют виды, которые не могли бы появиться на
Новой Азоре естественным путём.
В горах, где климат немного прохладнее, растут кипарисы, потомки завезённых
человеком видов, и можжевельники, родственные видам с Канарских островов. Они
образуют густые леса на вершинах, предпочитая влажный воздух. Ниже их располагается
пояс теплолюбивых и влаголюбивых лесов, демонстрирующих некоторые особенности,
характерные для тропического леса. Их образуют крупные древовидные вересковые
и некоторые другие деревья тропического происхождения – потомки видов, расселившихся
из Африки. Для этих лесов характерны лианы (различные виды плюща и клематиса),
а также эпифиты – орхидеи, мхи и папоротники.
Травянистые растения демонстрируют в основном европейское происхождение – их
расселение происходило благодаря Антигольфстриму. В меньшей степени в островной
флоре представлены африканские виды (их предки попали на остров благодаря птицам
или ветру). Злаки и осоки относительно немногочисленны, но образуют протяжённые
заросли, выдерживая конкуренцию с другими растениями благодаря устойчивости
к вытаптыванию и поеданию. В островной флоре богато представлены орхидеи, в
том числе эпифитные. Среди сложноцветных имеются крупные травы с колючими листьями,
а также невысокие кустарники с одревесневающим стеблями.
В горных районах, где меньше растительноядных животных, развиваются крупные
травянистые растения. Одно из них – гигантский вид подорожника, так называемый
«азорский банан».
Ближе к побережью, в более сухих районах предгорий располагается пояс вечнозелёных
лесов средиземноморского типа, образованных потомками некоторых культурных
деревьев – лавра, оливкового дерева, мирта, лимона. Здесь влаголюбивые папоротники
и мхи встречаются только в тенистых ущельях и долинах рек. В сухих лесах островками
встречаются заросли финиковой пальмы, потомка вида, культивировавшегося в эпоху
человека. Финиковая пальма благодаря способности образовывать корнеотпрыски
способна образовывать почти чистые заросли, тянущиеся на сотни метров. Ещё
одним свидетельством человеческой деятельности является местный древовидный
вид кактусов опунций, распространённый на значительной части прибрежных низменных
районов острова.
Прибрежные районы острова, отличающиеся сухостью, занимают сообщества засухоустойчивых
растений. Типичной растительной формацией на берегах Новой Азоры является
скрэб (мелколистные засухоустойчивые кустарники, часто колючие). Здесь обитает
небольшое количество суккулентных растений – молочаев (появившихся на острове
естественным
путём),
агав и алоэ (потомки завезённых человеком видов). Крупные агавы образуют большие
группы, над которыми возвышаются многометровые цветоносы.
В приливно-отливной полосе складывается комплекс солеустойчивых видов цветковых
растений. Здесь морская безлистная астра образует большие заросли, но цветёт
достаточно редко, размножаясь главным образом вегетативно.
К югу от Новой Азоры раскинулись обширные водорослевые поля. Здесь ветвь морского
течения образует локальный медленный водоворот, в котором растут плавающие
бурые водоросли, образующие довольно плотный покров на поверхности моря.
В фауне Новой Азоры влияние человека сказывается даже через
миллионы лет после его исчезновения. В первые миллионы лет на Азорских островах
обитали одичавшие козы, образовавшие комплекс низкорослых видов, но на различных
этапах формирования Новой Азоры они вымерли в результате вулканических извержений.
В неоцене фауна Новой Азоры образована исключительно потомками мелких видов,
сохранившихся с эпохи человека, и потомками более поздних вселенцев на остров.
Млекопитающие в фауне Новой Азоры – результат деятельности человека. На острове
обитает несколько видов ежей (потомки европейского ежа), среди которых один
вид умеет лазать по деревьям. Несколько видов летучих мышей, принадлежащих
к разным семействам, являются самостоятельными переселенцами на остров. В связи
с наличием наземных хищников ни один из их видов не показывает склонностей
к утрате навыка полёта.
Крупнейшими травоядными животными Новой Азоры являются тардолагусы – крупные
потомки завезённых европейских кроликов. Также на острове обитает несколько
мелких видов кроликов, обитающих в скрэбе и в лесах различных типов.
Главным хищником острова является мартилла – крупный потомок завезённого хорька.
Грызуны являются потомками мышей и крыс, завезённых на острова в эпоху человека.
Несколько видов этих грызунов населяют практически все биотопы острова.
Фауна птиц Новой Азоры очень богата. Несмотря на наличие хищников
из числа млекопитающих, на Новой Азоре существует несколько видов нелетающих
птиц. Из отряда гагарообразных на острове живёт плезиогагара – крупная морская
птица, приспособленная к длительным миграциям в океане. Ещё один вид нелетающих
гагар – похожая на небольшого тюленя гагара-хохотунья, обитающая также на Канарских
островах и атлантическом побережье Европы. Гагарообразные в тропических широтах
– выходцы с севера, свидетельство обширного оледенения на рубеже голоцена и
неоцена.
Ещё один вид нелетающих птиц – новоазорский псевдодронт, нелетающий потомок
глупыша. Ранние этапы его эволюции, связанные с утратой способности к полёту,
проходили на одном из островов, позже ставших частью Новой Азоры. Этот вид
оказался достаточно конкурентоспособным в окружении хищников из числа млекопитающих.
На пляжах Новой Азоры устраивают лежбища тропические китовые олуши – родственники
северного вида, появляющегося в этих местах крайне редко.
Среди околоводных птиц на острове обитают мелкие цапли, ищущие корм в лесной
подстилке и предпочитающие гнездиться на озёрах и болотах в горных долинах.
Также на болотах острова зимует макаронезийский журавельник и постоянно живёт
два вида пастушковых. Мелкие пастушковые, частично утратившие способность к
полёту, обитают на небольших островах возле Новой Азоры, где отсутствуют млекопитающие.
Отряд ржанкообразные представлен на Новой Азоре грифовыми поморниками, различными
видами чаек и
куликов. Своеобразным эндемичным видом является призрачный странник – птица,
которая ищет корм на водорослевых полях в море.
Также на водорослевых полях встречается саргассовая гага – вид, происходящий
с севера. На болотах острова встречается несколько видов перелётных уток (гнездящихся
в Европе и Исландии) и один оседлый.
Из ястребообразных на острове обитает морской лунь – хищная птица, приспособившаяся
охотиться на водорослевых полях. Другой вид хищных птиц, мелкий вид ястребов,
водится в лесах острова. Также на острове водится два вида сипух – крупный,
специализирующийся на питании позвоночными животными, и мелкий, охотящийся главным
образом на насекомых.
Куриные птицы представлены в фауне острова семейством фазановых. Несколько
видов азорских куропаток являются потомками завезённого в эпоху человека перепела.
Некоторые группы наземных птиц отсутствуют на Новой Азоре – например, козодои,
дятлы и ракшеобразные. Стрижи представлены одним видом эндемичного рода Azorapus.
Отряд воробьинообразные включает много мелких видов нескольких семейств. Среди
трясогузковых появились виды эндемичных родов – землешарки и водяная короткохвостка,
конвергентно сходная с оляпками. В лесах острова обитает несколько видов вьюрков,
эволюционировавших от предков европейского происхождения. Синицы представлены
несколькими насекомоядными видами, один из которых сильно специализирован на
поиске насекомых под корой деревьев. Из врановых птиц на острове живут азорская
сорока и мелкий всеядный вид ворон.
Рептилии Новой Азоры немногочисленны и включают одного крупного представителя
настоящих ящериц (Lacertidae) и несколько родственных друг другу видов мелких
гекконов.
Амфибии и пресноводные рыбы на Новой Азоре отсутствуют.
Фауна беспозвоночных демонстрирует богатство видов, которые произошли в ходе
длительной эволюции от немногих предковых форм.
Большинство насекомых происходит от хорошо летающих предков, самостоятельно
добравшихся до острова. Стрекозы типичны для пресных водоёмов, где их личинки
замещают мелких пресноводных рыб. Среди стрекоз есть и равнокрылые, чьи личинки
главным образом пелагические, и разнокрылые с донными ползающими личинками.
Своеобразны эндемичные виды клопов. Клоп азорский иглолаз является симбионтом
и чистильщиком местных видов ежей. Вероятнее всего, он попал на остров вместе
с перелётными птицами, поскольку родствен африканским видам. Среди растительноядных
клопов примечательна гигантская кружевница, имитирующая лист дерева. На острове
живёт несколько мелких видов щитников и хищнецов.
Среди бабочек есть несколько видов белянок и нимфалид европейского происхождения.
Несколько видов нимфалид достигают крупных размеров, сравнимых с тропическими
бабочками.
Жуки включают представителей листоедов, пластинчатоусых и мелких стафилинов,
происходящих из Европы. К видам американского происхождения относится несколько
представителей светляков.
Фауна двукрылых разнообразна и включает несколько видов мух, комаров и мошек.
Особый вид мошки обитает на водорослевых полях вблизи острова, развиваясь в
морской воде и никогда не появляясь в пресных водах или на суше.
Характерными обитателями острова являются наземные крабы семейства Grapsidae.
Несколько видов этих ракообразных занимают различные экологические ниши на
острове, играя большую роль в почвообразовании.
Наземные улитки представлены несколькими видами одного рода лёгочных улиток
и являются потомками виноградной улитки Helix pomatia, попавшей на острова
в эпоху человека и успешно пережившей вулканические извержения, сопровождавшие
формирование Новой Азоры.
 |
 |
 |
Dendrogalerix scandens |
Tardolagus novazorae |
Azopica collector |
 |
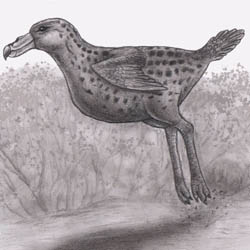 |
 |
Georimarornis psammophilus |
Terricolofulma pseudoraphoides |
Megalotingis phyllomorpha |
| Остров Новая Азора, морские птицы и другие обитатели побережья |
Большая
Антигуа
|
Остров Большая Антигуа образовался на окраине Карибского моря
в результате слияния островов Гаити, Пуэрто-Рико и нескольких мелких островов
из группы Антильских, лежащих к востоку от них. Это является результатом тектонических
процессов, происходящих в Центральной Америке и напрямую связанных с расширением
Атлантического океана. Большая Антигуа лежит на окраине Карибской литосферной
плиты и представляет собой сильно вытянутый узкий остров, на котором различимы
несколько потухших и действующих вулканов разного возраста, отражающих историю
образования и изменения острова. В процессе своего становления Большая Антигуа
была частью сухопутного Антильского моста, по которому какое-то время происходил
обмен флорой и фауной между Северной и Южной Америкой.
Значительную часть площади Большой Антигуа составляют горы разного возраста,
и лишь прибрежные районы представляют собой узкую полосу равнин.
Климат острова исключительно тропический, круглый год тёплый и влажный. Он
во многом определяется морями, окружающими остров, поэтому сезонные изменения
климата выражены очень неявно – лишь зимой выпадает немного больше дождей.
В горных районах прохладнее и суше, чем на побережье, но и здесь температура
редко опускается ниже +20°С. Дожди идут почти ежедневно, меняется лишь их продолжительность.
С весны до осени нередки тропические ураганы.
Реки на острове полноводные и не пересыхающие, но довольно короткие. Источником
воды для них являются дожди. В горных долинах много небольших озёр и болот;
болотистые участки имеются в низовьях рек.
Основной тип растительности на Большой Антигуа – тропические
леса, занимающие в общей сложности около трёх четвертей площади острова. В
горах тропический лес уступает место другому типу растительности – редколесьям
с элементами тропической саванны. Завезённые в историческую эпоху сосны образуют
редколесья на сухих и хорошо дренированных почвах, а в подлеске сосновых лесов
обитают засухоустойчивые кустарники и кактусы. На сухих горных склонах кактусы
образуют сплошные заросли вместе с агавами разных видов.
Тропические леса отличаются высоким видовым разнообразием деревьев. Очень характерными
являются крупные лианы и различные эпифиты – папоротники, бромелии и орхидные,
а также геснериевые и кактусы из группы гилоцереусов. На нарушенных участках
леса, в долинах рек, разрастаются крупные корневищные травы, в том числе гибридогенные
виды бананов, потомки одичавших культурных форм. Одно из своеобразных растений
тропических лесов Большой Антигуа – древовидный табак, чьи трубкообразные цветки
узко приспособлены к опылению одним видом бабочек.
На равнинах в прибрежных районах в лесу появляется большое количество пальм.
Одни из них растут в одиночку, возвышаясь над пологом леса, а другие образуют
протяжённые одновидовые заросли.
На побережьях пальмы являются преобладающей группой древесных растений. На
морских мелководьях растут обширные мангровые леса.
Тропические условия являются благоприятными для поддержания
многообразия фауны острова. В эпоху человека на острова Карибского моря были
завезены животные различных групп, не встречавшиеся здесь ранее, и их потомки
являются частью неоценовой фауны Большой Антигуа. В целом фауна острова принадлежит
к Неотропической зоогеографической области, но в её составе заметно сильное
влияние Голарктики.
В раннем неоцене острова Карибского моря представляли собой прерывистый, но
действующий «мост» для расселения южноамериканской фауны на север. Из-за неравномерного
осушения и затопления островов лишь часть североамериканских видов попала по
Антильскому мосту в Южную Америку, но значительно больше южноамериканских видов
проникло на север.
Несмотря на то, что Большая Антигуа является островом, фауна млекопитающих несёт много черт, характерных для материков. В частности, здесь присутствуют
животные типично наземных групп.
Сумчатые млекопитающие Большой Антигуа – это исключительно опоссумы североамериканского
происхождения, древесные всеядные животные разного размера. Один из видов опоссумов
напоминает континентальных североамериканских геопоссумов – он плохо лазает
по деревьям и предпочитает искать корм на земле.
Разнообразие насекомоядных невелико и включает несколько видов землероек североамериканского
происхождения; эндемичные щелезубы вымерли ещё в конце исторической эпохи.
Рукокрылые Большой Антигуа исключительно разнообразны и включают виды нескольких
семейств. Среди них имеются многочисленные насекомоядные виды, несколько фруктоядных,
хищников и один карликовый вид, питающийся кровью позвоночных.
В антигуанской фауне имеются представители отряда приматов. Завезённые в историческую
эпоху узконосые обезьяны (зелёные мартышки из Африки) успешно пережили экологический
кризис, эволюционировали в новые виды и даже мигрировали в Северную Америку.
Также на острове присутствует несколько видов мелких широконосых обезьян южноамериканского
происхождения – игрунок, населяющих кустарники и полог леса. Один вид игрунок
обитает в мангровых лесах.
Из отряда хищных на острове обитают карликовые виды псовых – уктены, потомки
койотов. Есть несколько небольших лесных видов кошек, американские енотовые
лемуры и настоящие еноты. Результатом деятельности человека является наличие
на Большой Антигуа виверровых – потомков завезённых в историческое время мангустов.
Отряд грызунов представлен многочисленными представителями как голарктических,
так и неотропических групп. Из голарктических грызунов обычны беличьи и белки-летяги.
Также встречаются различные виды мышей и крыс – преимущественно потомки видов,
завезённых человеком. Эндемичное семейство хутиевых вымерло в конце эпохи человека.
Чешуйчатые дикобразы, ведущие наземный и древесный образ жизни – вселенцы из
Южной Америки. С эпохи человека сохранились и эволюционировали представители
семейства агутиевых. Одной из самых необычных групп грызунов является семейство
мар-оленей южноамериканского происхождения. Его представителем на Большой Антигуа
является мазамара – карликовый вид, размером не превышающий собаку.
Фауна птиц Большой Антигуа демонстрирует богатство, характерное
для тропических областей. Здесь отсутствуют некоторые группы птиц, характерные
для Северной и Южной Америки, но в целом видовое разнообразие очень высокое.
Влажный климат и обилие пресных водоёмов способствуют разнообразию околоводных
птиц. На болотах и реках обитают ибисы и аисты, а также многочисленные цапли.
Среди цапель есть виды, обитающие в мангровых зарослях на морском побережье.
Среди морских птиц обычны различные виды чаек и тропических крачек. Большинство
видов куликов – это птицы, прилетающие на зимовку из Северной или Южной Америки.
На Большой Антигуа зимуют журавельники из Северной Америки. Пастушковые встречаются
повсеместно по берегам рек и в кустарниковых зарослях, не поднимаясь высоко
в горы.
Утки встречаются повсеместно, демонстрируя разнообразие размеров и окрасок.
Среди них есть вид, приспособившийся к гнездовому паразитизму.
Куриные птицы Большой Антигуа – потомки домашних кур и зубцеклювых перепелов.
Значительная часть этих видов встречается в лесах острова, но несколько видов
обитает в горных редколесьях.
Хищные птицы Большой Антигуа очень разнообразны. Крупными плотоядными птицами
являются катартиды, предпочитающие редколесья и прибрежные области. В лесных
районах обитают коршуны и южноамериканские по происхождению каракары. Совы
представлены разными видами сипух и настоящих сов.
В тропических лесах острова многочисленны попугаи – в основном небольшие виды,
но есть несколько крупных, питающихся орехами с твёрдой скорлупой.
На Большой Антигуа обитают разнообразные дятлы. Среди них имеются виды, ведущие
необычный образ жизни: обитатели мангровых зарослей и добывающие корм на рифах
во время отлива. Крупный нелетающий дятел с Большой Антигуа – представитель
особого семейства антигуанских дятлов.
Характерной группой лесных птиц Большой Антигуа являются трогоны. Кукушки представлены
большим количеством видов; крупнейшие из них питаются мелкими позвоночными,
а мелкие строго насекомоядны и могут поедать даже пчёл с ядовитым жалом.
Самым разнообразным отрядом птиц являются воробьинообразные. Среди них имеются
различные кричащие воробьиные южноамериканского происхождения – котинговые
и тиранны. Певчие воробьиные представлены значительно большим количеством семейств:
мухоловки, дрозды, ткачиковые (потомки воробья), вьюрковые, скворцы (потомки
завезённого в Северную Америку европейского скворца), врановые.
Разнообразие рептилий очень велико благодаря тропическому климату. Среди черепах
имеются представители семейства наземных черепах (Testudinidae), а также виды
семейства Emydidae – коробчатые и несколько видов пресноводных черепах. Среди
чешуйчатых примечательна очень крупная игуана террагуана – крупнейший хищник
острова. Кроме неё на острове обитает много видов мелких игуан, а также тейиды.
Гекконы встречаются в тропических лесах, среди них есть несколько очень мелких
видов. Среди змей наиболее типичны мелкие удавы, а также гремучие змеи и слепуны.
Земноводные представлены только отрядом бесхвостых. На Большой Антигуа есть
представители семейств квакш и короткоголовов, расселившиеся на островах Карибского
моря естественным образом. Кроме них, в фауне острова имеются представители
семейства жаб – потомки жабы аги, завезённой человеком. Большинство лягушек
обитает в пологе тропического леса.
Фауна рыб Большой Антигуа сравнительно бедна по сравнению с материковой. Здесь
отсутствуют представители многих групп рыб, имеющихся на соседних материках.
Сомообразные рыбы представлены несколькими видами морских ариевых сомов (Ariidae),
заходящих в реки. Также есть клариевые сомы, потомки вида, завезённого человеком
во Флориду и расселившиеся оттуда на юг через Кубу. Карповые рыбы были завезены
в историческую эпоху, но постепенно вымерли из-за конкуренции с другими группами
рыб. Среди окунеобразных рыб имеется несколько видов горбылей морского происхождения,
а также цихлид: местные виды цихлазом и потомки завезённой мозамбикской тиляпии.
Особого разнообразия на Большой Антигуа достигают живородящие рыбы, составляющие
большую часть пресноводной ихтиофауны. В реки заходят представители различных
семейств живородок Карибского моря, которые благодаря способности рождать живых
мальков могут проходить весь жизненный цикл в пресной воде. Также в низовьях
рек встречаются рыбы-иглы, несколько видов тетраодонов и своеобразные живородящие
рыбы вертикалихты. В реки острова часто заходят акулы и скаты. В пещерных системах
Большой Антигуа обитает несколько узкоареальных видов рыб – живородящих и карликовых
цихлид.
Беспозвоночные Большой Антигуа исключительно разнообразны, среди них имеется
много узкоспециализированных видов. Для тропических лесов характерны различные
тараканы, в том числе мелкие летающие и крупные роющие формы. Среди клопов
много крупных растительноядных видов причудливой формы и окраски; имеются хищные
виды, в том числе несколько водных из различных семейств. Бабочки демонстрируют
богатство видового состава. Некоторые из них отличаются крупными размерами:
гигантский антигуанский бражник обладает хоботком полуметровой длины. Очень
крупными размерами и яркой окраской отличаются местные бабочки-парусники. Общественные
перепончатокрылые острова являются потомками как южноамериканских видов, так
и завезённых медоносных пчёл. Одиночные осы на острове демонстрируют разнообразные
приспособления к паразитизму – независимо от наездников Юго-Восточной Азии
они приспособились к паразитизму на мелких позвоночных. На острове обитает
множество видов муравьёв. Жуки очень разнообразные, среди них имеются многочисленные
хищные (жужелицы, мягкотелки, светляки и др.) и растительноядные формы. Среди
двукрылых большого разнообразия достигают комары и мухи; среди мух имеются
очень крупные виды.
Для влажных мест характерны крупные многоножки – кивсяки и сколопендры. Некоторые
хищные многоножки поедают мелких птиц и летучих мышей, убивая их одним укусом.
Фауна паукообразных включает крупных скорпионов, разнообразных пауков (в том
числе крупных птицеедов) и клещей.
Среди ракообразных очень характерны наземные крабы семейства Grapsidae. Некоторые
крабы освоили жизнь в пресной воде, но для размножения мигрируют в море. Свидетельством
связи островов Карибского моря с Северной Америкой является наличие на Большой
Антигуа (а также на соседней Кубе) речных раков американского семейства Cambaridae,
среди которых имеется несколько мелких и ярко окрашенных видов. Также на острове
встречаются пресноводные креветки.
Благодаря регулярным связям с материком на острове имеются разнообразные лёгочные
моллюски. Виды из тропических лесов обладают разнообразно окрашенными раковинами
и образуют много местных подвидов и цветовых форм в изолированных долинах и
вулканических кратерах. В реках водятся различные виды семейства катушек, а
также крупные ампуллярии.
 |
 |
Mazamara sylvatica |
Antiguapicus apterus |
| Рыбы и другие обитатели Карибского моря | |
| Остров Большая Антигуа, тропический лес и горы | |
| Остров Большая Антигуа, водные и околоводные беспозвоночные и мелкие позвоночные. |
Новая
Тортуга
|
В процессе расширения Атлантики остров Сан-Паулу, находящийся
в тропических широтах Атлантики, затонул, но в дальнейшем на его месте в раннем
неоцене из океанских глубин неоднократно поднимались острова. Последним из
них был остров Новая Тортуга, который существует на протяжении приблизительно
десяти миллионов лет. Этот остров вулканического происхождения представляет
собой несколько вулканических конусов, самые старые из которых частично разрушены.
В окрестностях Новой Тортуги есть несколько небольших островов, также вулканических.
Площадь Новой Тортуги – около 1000 квадратных километров.
Признаки вулканической активности имеются на самом острове – есть несколько
мест, где бьют горячие ключи. Иногда в разных местах острова бывают выбросы
вулканических газов, удушающих на своём пути всё живое.
Рельеф острова разнообразен, но большую часть территории занимают горы. Пологие
участки – в основном узкая полоса побережья, сложенная главным образом вулканическим
песком.
Новая Тортуга лежит в области экваториального климата. Здесь круглый год господствует
жаркий климат, а количество осадков незначительно меняется по месяцам. Благодаря
дождям на острове существует большое количество водоёмов – коротких рек и небольших
озёр. Постоянные озёра есть в кратерах спящих вулканов.
На Новой Тортуге произрастают тропические леса, сходные с
южноамериканскими, но образованные главным образом деревьями, семена которых
разносятся ветром. Семена некоторых деревьев утратили эту способность, но сохраняют
рудиментарное крыло или хохолок, указывающие на способ появления предков этих
растений на острове. Некоторое количество растений было занесено с материка
птицами.
Видовое разнообразие деревьев на острове невелико; это главным образом представители
древовидных молочайных и сложноцветных. Ряд характерных для Южной Америки групп
растений (семейства лецитисовые, стеркулиевые, бобовые и пр.) на острове не
встречается. Пальмы, произрастающие на Новой Тортуге, являются потомками видов,
семена которых были принесены водой. На побережье произрастают мангровые леса,
образованные одним видом мангровых деревьев.
Травянистые растения представлены очень богато, среди них существует много
эндемиков. Очень многочисленны бромелиевые американского происхождения, среди
которых есть эпифиты и несколько крупных наземных видов, произрастающих главным
образом в горах. У всех бромелиевых имеются ёмкости для воды в середине розетки
или в пазухах листьев, всегда полные воды благодаря дождям.
В тропическом лесу произрастают многочисленные эпифиты (орхидеи, бегонии и
другие) и лианы. На острове встречаются различные папоротники, наземные и эпифитные.
Большинство их имеет американское происхождение, есть один вид африканского
происхождения.
Поскольку Новая Тортуга образовалась после исчезновения человека,
фауна этого острова формировалась исключительно естественным путём и в ней
отсутствуют многие группы наземных животных. Фауна Новой Тортуги формировалась
главным образом за счёт неотропической фауны. Выносимые реками на растительных
«плотах» животные должны были проделать долгий путь, попав в кольцо Бразильского
течения, идущего на юг вдоль берегов Южной Америки, и вернуться на север с
Южным Пассатным течением.
Млекопитающие Новой Тортуги очень малочисленны – это только несколько видов
мелких летучих мышей. Среди них есть насекомоядные виды и один нектароядный.
Птицы, населяющие остров, намного разнообразнее. На побережье острова встречаются
многочисленные морские птицы – чайки и кулики. Среди чаек есть один своеобразный
вид – земляная чайка, которая является наземной птицей и заменяет отсутствующих
на острове врановых. Из журавлеобразных на Новой Тортуге живёт куриный пастушок
– всеядная наземная птица, утратившая способность летать. Пернатые хищники
Новой Тортуги малочисленны: есть эндемичная новотортугасская сипуха и в редколесьях
встречается мелкий вид ястребов. На острове обитают разнообразные мелкие воробьиные
птицы, как певчие, так и непевчие. Среди них есть виды американского и африканского
происхождения. Среди американских видов имеется по одному виду птиц-печников
и котинг, а африканские представлены несколькими вьюрковыми.
Благодаря тропическому климату на острове процветают рептилии. Остров получил
своё название благодаря разнообразию черепах, которые создали уникальную фауну.
Черепахи семейства пеломедузовых, происходящие из Южной Америки, стали доминирующими
наземными животными. Благодаря отсутствию конкуренции среди них появились хищные,
травоядные и морские формы, и даже один древесный лазающий вид. Ящерицы острова
представлены несколькими видами гекконов африканского происхождения, а также
мелкими тейидами, происходящими из Южной Америки.
Амфибии и пресноводные рыбы на острове отсутствуют.
Фауна беспозвоночных демонстрирует разнообразие и высокую
степень эндемизма. Насекомые представлены разнообразными жуками, часть из которых
происходит из
Африки (бронзовки, златки). Среди американских по происхождению жуков есть
светляки и мягкотелки. Благодаря преобладанию восточных ветров фауна бабочек
формировалась почти исключительно африканскими по происхождению видами. На
острове обитает несколько видов нимфалид и парусников, а также многочисленные
мелкие моли. Бабочки американского происхождения – несколько видов семейства
геликонид. Также на острове обитает несколько видов двукрылых, в том числе
кровососущая
черепашья
муха.
Пауки Новой Тортуги имеют американское происхождение. На острове живут несколько
родственных видов пауков-скакунов, явно происходящих от единого предка.
Для островной фауны характерны наземные крабы, которые играют роль падальщиков
и мелких хищников, конкурируя с пауками и заменяя наземных пауков типа птицеедов.
Среди моллюсков на острове встречается несколько видов улиток американского
происхождения, родственных друг другу и отличающихся по экологии.
 |
 |
 |
Cheloniraptor ferox |
Gravichelonia giraffocervix |
Apterogallus melanoleucus |
| Каменные спины | Остров в тропиках Атлантического океана; его обитатели - птицы и рептилии. |
Фолклендские
острова
|
Фолклендские острова расположены примерно в 700 километрах
от атлантического побережья Южной Америки. Они находятся на той же материковой
плите, вдалеке от областей вулканической активности, поэтому геология островов
практически не изменилась с эпохи человека. Только во время оледенения на рубеже
голоцена и неоцена на значительной части островов появился ледниковый покров,
сильно сгладивший рельеф местности и оставивший валы моренных отложений.
Берега островов скалисты и сильно изрезаны бухтами, глубоко вдающимися в сушу.
Вокруг двух главных островов разбросано много отмелей и мелких островков.
Климат Фолклендских островов практически полностью зависит от океана. Острова
омываются холодным течением, а воды с экватора достигают островов сильно остывшими.
Кроме того, вместе с Южной Америкой острова сдвинулись на юг относительно своего
положения в эпоху голоцена. Из-за этого климат островов остаётся холодным, несмотря
на общее потепление в неоцене. Зимние температуры часто опускаются ниже нуля
и на островах выпадает снег. Лето прохладное, жаркая погода держится лишь несколько
недель в середине лета. Значительную часть года на островах стоят туманы, а
ночью и утром бывают заморозки.
Постоянных пресных водоёмов на Фолклендских островах очень мало. В основном
это озёра, питаемые снеговой и дождевой водой. Реки на островах короткие, узкие
и мелководные, с холодной водой.
Растительность Фолклендских островов очень скудная. Это связано
с суровостью климата, а также с деятельностью травоядных млекопитающих. Характерные
для эпохи человека заросли высокой травы (т. н. туссок) в неоцене исчезли на
главных островах, сохранившись как реликты только на удалённых островках. Одичавшие
домашние животные в конце эпохи человека уничтожили значительное количество
видов эндемичных растений и разрушили естественные растительные сообщества.
В неоцене травяной покров на крупных островах представляет собой плотную дернину
сравнительно невысоких злаков с примесью разнотравья. Травянистые растения
из числа двудольных (зонтичные, сложноцветные и пр.) произрастают главным образом
среди камней, в местах, менее доступных для травоядных млекопитающих. Кустарниковая
растительность очень скудная – два вида вереска. Деревьев на островах нет.
На болотах растут различные виды осоки и другие травянистые растения. На островах
есть сфагновые болота и торфяники.
Среди споровых растений очень характерны папоротники. Мхи представлены большим
количеством видов, произрастают главным образом среди скал.
На прибрежных мелководьях разрастаются бурые водоросли, образующие протяжённые
заросли (келп).
Фауна Фолклендских островов эпохи неоцена формировалась несколькими
путями. Часть обитателей островов попала на них по воздуху или из моря естественным
путём; также есть много потомков видов наземных животных, завезённых в эпоху
человека. На рубеже голоцена и неоцена оледенение закрыло острова ледниками
почти полностью, оставив лишь несколько «оазисов» на их окраине, где сохранились
немногочисленные наземные животные. После отступления ледника животные заселили
острова целиком.
Из млекопитающих на Фолклендских островах живут хищники – потомки
южноамериканской серой лисицы, завезённой в эпоху человека. Кошки, также завезённые
человеком, вымерли из-за похолодания, не выдержав конкуренции с псовыми и исчезновения
привычных местообитаний. Главными травоядными являются крупные потомки завезённых
кроликов. Также на островах обитают мелкие потомки завезённых человеком крыс.
В прибрежных водах обитает фолклендский паральгоцетус – крупнейший местный вид
млекопитающих и одно из самых крупных млекопитающих неоцена. Этот гигант питается
бурыми водорослями и не покидает море. Ещё одно морское млекопитающее островов
– айяпух, питающийся рыбой и похожий по образу жизни на выдру.
Фауна птиц значительно разнообразнее. Многие птицы являются
перелётными, и значительная часть птиц связана в своей жизни с морем. На реках
и озёрах обитает один мелкий вид поганок, встречающийся также на юге Патагонии
и на Огненной Земле. Также на пресных водоёмах гнездятся различные утки и кулики.
Фауна морских птиц значительно разнообразнее. Эндемичным видом является фолклендский
кавескар – представитель чаек-пингвинов. Кроме него на островах встречаются
патагонский кавескар и собственно чайки-пингвины нескольких видов; чайки и крачки
также разнообразны. В прибрежной полосе обитает много видов куликов, специализированных
к питанию различными видами морских беспозвоночных. Изредка на островах гнездится
альбатрос-кочевник, но этот вид предпочитает селиться на мелких островках и
не образует крупных поселений. Наземные птицы представлены лишь несколькими
видами тираннов, перелётными видами кассиков, коньков и одним видом южноамериканских
оляпок, а также оседлыми представителями овсянок. Главными пернатыми хищниками
островов являются крупная оседлая каракара и мелкий вид сов.
Рептилий и амфибий на Фолклендских островах
нет.
Пресноводные рыбы представлены на островах исключительно потомком
форели, завезённой в эпоху человека. Этот вид, карликовый фолклендский смолт,
демонстрирует значительное разнообразие местных форм по рациону, окраске и образу
жизни – от мелких насекомоядных разновидностей до среднеразмерных хищников-каннибалов.
Фауна беспозвоночных Фолклендских островов скудна. Из насекомых
на островах встречается несколько видов эндемичных жуков, утративших способность
летать, а также несколько видов молей и вид нимфалид, почти полностью утративший
способность летать. В пресные воды островов перешло несколько видов равноногих
ракообразных. В прибрежных районах в почве обитает несколько видов роющих изопод.
На берегах обитают наземные виды бокоплавов, конвергентно сходные с исландскими.
В низовьях рек обитает несколько видов двустворчатых моллюсков и усоногих ракообразных.
| Завоевание юга | Природа Фолклендских островов и прибрежных вод Антарктиды. |
Острова
Тихого океана |
| Пелагические животные Тихого океана | |
| Планктонные обитатели Тихого океана | |
| Тихий океан, глубина в 2500 метров | |
| Дно Тихого океана | |
| Тихий океан. Гигантский вид морских черепах, его паразиты и симбионты. | |
| Подводная твердыня | Жители рифов тропической зоны Тихого океана - рыбы и беспозвоночные. |
Гавайские острова
|
Гавайские острова отмечают своим местоположением поток магмы,
бьющий из земных недр и прожигающий тонкую океаническую кору. В результате
движений литосферных плит поток магмы прожёг в океанической коре цепочку островов.
Архипелаг составляют гористые острова, чётко различающиеся возрастом и рельефом.
Самый западный из крупных островов – Кауаи, один из самых старых в архипелаге,
с самой низкой вулканической активностью. У этого острова сильно сглаженный
рельеф и сильнее выветренные породы, а тектоническая активность ограничивается
землетрясениями. Более молодые острова, лежащие к юго-востоку от Кауаи – это
активные вулканы, на которых часто происходят излияния жидкой лавы (вулканическая
активность усиливается по направлению от северо-запада к юго-востоку). В неоцене
к цепочке островов прибавился Хекеуа – самый молодой и вулканически активный
остров. В целом с эпохи человека архипелаг несколько сдвинулся к северо-западу.
В отложениях островов осталось свидетельство крупного извержения, сопровождавшего
образование острова Хекеуа.
Помимо крупных островов, в архипелаге существует много мелких островков близ
побережья, облегчающих расселение представителей флоры и фауны.
Архипелаг лежит в области тропического морского климата, во многом определяемого
Тихим океаном. Сезонные изменения климата не выражены, климат влажный и тёплый
круглый год. Благодаря высоте над уровнем океана вершины гор служат естественными
конденсаторами влаги, обеспечивающими практически ежедневные дожди.
Высокогорные районы являются областями умеренного климата, а на высочайших
вершинах лежит снег, несмотря на тропическое местоположение островов.
Благодаря большому количеству осадков на Гавайских островах существует много
пресных водоёмов. Реки на Гавайских островах короткие, мелководные и порожистые;
в ущельях и горных долинах образуются озёра и болота.
Человек нанёс сильный вред туземной флоре, которая складывалась
на протяжении миллионов лет до эпохи колонизации островов. В неоценовой флоре
островов существует много потомков завезённых растений, но практически нет
потомков видов, существовавших до исторической колонизации. Однако, существуют
виды, попавшие на острова естественным путём уже после исчезновения человека.
Причинами разнообразия неоценовой гавайской флоры являются высотная поясность,
наличие большого количества сравнительно изолированных местообитаний и изоляция
островов друг от друга морскими проливами.
В низинных частях Гавайских островов господствует тёплый и влажный климат.
В долинах и предгорьях произрастают обширные леса из влаго- и теплолюбивых
растений. Среди пионерных растений в нарушенных участках древостоя развивается
сообщество быстрорастущих одревесневающих растений, среди которых имеются потомки
папайи и равеналы, завезённых в историческую эпоху. Во влажных лесах доминируют
акации местных видов (среди которых имеется несколько орнитофильных видов),
фикусы и веерные пальмы. Характерными для этих лесов являются древовидные сложноцветные,
являющиеся эндемиками этих островов. Разнообразие древесной растительности
вызвало появление интересного вида паразитических деревьев – сандалового дерева
незеленеющего. Это дерево полностью утратило хлорофилл и питается исключительно
за счёт окружающих деревьев. В пологе леса произрастает большое количество
эпифитных растений – орхидей (нескольких эндемичных родов), папоротников и
бегоний. Среди эпифитов также имеются хищные растения – росянковые. На поврежденных
участках леса и в пологе леса произрастает большое количество лиан.
На склонах гор леса становятся ниже, а выраженная ярусность пропадает. Появляется
большое количество растений, способных укореняться стволом и ветвями, образуя
сплошные и почти непроходимые заросли на склоне. Здесь господствуют потомки
завезённых в эпоху человека мирики, гуаявы и пуансеттии (пламенеющее ядовитое
дерево).
В тенистых ущельях складывается комплекс тенелюбивых лесов из древовидных папоротников
(также потомков завезённых видов) с отдельными высокорослыми растениями из
числа цветковых. В этих лесах складывается влажный микроклимат, образуются
заросли мхов различных видов и мелких травянистых папоротников. Из цветковых
растений в папоротниковых лесах живут орхидеи и бегонии.
Дальше вглубь суши, в районах, защищённых горными хребтами от ветров с океана,
произрастают более сухолюбивые леса из потомков маслины, пальм и юкки (реликт,
вымерший на континенте, но сохранившийся на архипелаге). На влажных бедных
почвах при обилии солнечного света развиваются бамбуковые леса – обычно они
являются одной из стадий восстановления растительности после извержений вулканов,
или захватывают участки повреждённого леса.
На крупных и относительно молодых островах складывается комплекс высокогорной
лесной растительности, обитающей в условиях относительной сухости и избыточного
солнечного освещения. Своеобразным типом гавайской растительности являются
«серебряные леса», образованные светолюбивыми растениями с серебристым опушением
и восковым налётом на листьях – эвкалиптами, гревиллеями, пальмами и акациями.
Эти леса разреженные, с густым кустарниковым подлеском. Выше по склонам гор
лиственные леса сменяются хвойными, образованными араукарией, сосной и можжевельником.
В высокогорье произрастает стланиковая араукария, а также рододендроны, потомки
азиатских видов, завезённых в историческую эпоху. На вершинах гор, вдоль линии
снегов, сложился комплекс растительности, приспособленной к суровым условиям
высокогорья. Здесь обитают отдельные крупные виды трав и многочисленные низкорослые
и подушковидные растения, конвергентно сходные с тундровой растительностью.
В горных районах остались реликты ледниковой эпохи, представители флоры умеренного
климатического пояса: пушица и росянка. Также в горах обитают высокоспециализированные
травянистые растения, выходцы из более тёплого климата, вторично приспособившиеся
к холоду и обилию солнечного света.
Характерными местообитаниями вулканически активных островов являются лавовые
поля. Здесь складывается комплекс пионерной растительности, способной существовать
на бедных органикой почвах. Характерным видом таких растений является «серебряная
пена», низкорослый вид сложноцветных. По мере накопления органики на уничтоженных
лавой участках поселяются мирика и бамбук, сменяемые настоящими лесными растениями.
На болотах развиваются заросли циперуса крупных видов, отдалённо напоминающих
африканский папирус. На переувлажнённых почвах произрастают гигантские потомки
таро, а мелководные участки рек с быстрым течением занимают местные виды кувшинок
и другие потомки водных растений, завезённых человеком.
На побережьях, в условиях обилия солнечного света и засоленности почвы, образовался
особый тип растительных сообществ. Прибрежные мелководья покрыты зарослями
морского кокоса – пальмы особого вида, образующей аналог мангровых лесов. Второй
линией растительности являются заросли пандануса и гигантской травы укропника
(азиатского происхождения). Дальше вглубь суши прибрежные сообщества постепенно
сменяются тропическими лесами.
Своеобразие гавайской фауны, сложившееся естественным образом
в доисторическую эпоху, было в значительной степени утрачено после колонизации
островов человеком. Деятельность человека в значительной степени изменила характер
островной фауны на уровне классов.
В неоценовой фауне Гавайских островов присутствуют многочисленные виды млекопитающих – это во многом результат деятельности человека. До людей на островах обитали
исключительно рукокрылые, но в неоцене на островах водится большое количество
наземных млекопитающих. Достаточно нетипичный представитель млекопитающих –
лазающее сумчатое млекопитающее, обезьяний валлаби. В горных лесах также водится
несколько видов валлаби, ведущих более типичный наземный образ жизни. Все они
являются потомками щеткохвостого валлаби Petrogale penicillata, завезённого
из Австралии и успешно натурализовавшегося. Плацентарные млекопитающие представлены
на Гавайях хищниками, парнокопытными, грызунами и рукокрылыми. Среди хищных
млекопитающих интересен илиохолокаи – крупный тюленеобразный потомок мангустов,
обитающий на морском побережье. Прочие потомки мангустов ведут наземный и древесный
образ жизни, являясь экологическими аналогами горностаев, куниц и лисиц. Парнокопытные
Гавайских островов – это несколько видов оленей, которые произошли от азиатского
оленя аксиса, завезённого в эпоху человека. Наиболее многочисленной группой
гавайских млекопитающих являются грызуны, являющиеся потомками завезённых на
острова крыс и мышей. Самый крупный среди них – капо, являющийся распространителем
семян многих видов местных деревьев. Адаптивная радиация вызвала появление
на островах многочисленных крыс, отличающихся образом жизни, рационом и поведением.
Среди них характерны всеядные бамбуковые крысы и лазающие растительноядные
цветочные крысы. Особое эндемичное семейство составляют роющие грызуны, утратившие
зрение – тихоокеанские ложные слепыши. Самостоятельно расселились на острова
лишь рукокрылые, среди которых есть хищники и насекомоядные формы.
Несмотря на присутствие на Гавайях млекопитающих, на островах очень разнообразны
птицы. На небольших островках близ побережья главных островов
архипелага гнездятся морские птицы – чайки и кулики. Разнообразные трубконосые
птицы обитают на побережьях островов, но предпочитают гнездиться на островках
вдали от побережья, где нет млекопитающих. Среди веслоногих птиц встречается
несколько мелких видов олуш. Также на островах водятся оседлые чистиковые,
явно расселившиеся на юг в ледниковый период и
образующие
эндемичный
род, ограниченный
в распространении только Гавайями. Среди прочих околоводных птиц для островов
характерны цапли нескольких видов, оседлые, но не утратившие способностей к
полёту. Также имеется крупный вид цапель эндемичного рода,
ищущий корм главным образом на земле.
Своеобразен
гелициорнис
–
ночная моллюскоядная птица, крупный кулик, конвергентно сходный с пастушковыми.
Собственно пастушковые на крупных островах населяют главным образом болота
и не утрачивают способности к полёту. На мелких островах, где меньше млекопитающих,
имеются лесные виды пастушковых. Гусеобразные птицы на Гавайях достаточно
многочисленны; большинство их видов составляют
утки,
ведущие
традиционный
околоводный образ
жизни. На
островах зимует
несколько
видов уток и гусей из Северной Америки и Восточной Азии. Оседлыми являются
крупные нелетающие травоядные птицы – гавайские лесные гуси. Куриные птицы
также присутствуют на островах: это разнообразные «ложные фазаны», потомки
американских зубцеклювых перепелов. Они обитают в редколесьях и кустарниках,
а также на горных склонах. Среди плотоядных птиц есть своеобразный нелетающий
вид луней – куахана, охотящийся на крупных наземных позвоночных. На морских
побережьях гнездятся эфирные коршуны, добывающие корм в море и в приливно-отливной
полосе. Интересным примером конвергенции с совами является неопуэо, потомок
луней, ведущий сумеречный образ жизни и питающийся насекомыми и мелкими позвоночными.
Для островов характерны совы: несколько мелких видов сов, питающихся почти
исключительно насекомыми. Они происходят от одного североамериканского вида,
попавшего на острова после окончания эпохи человека. Все голуби Гавайских островов
происходят от завезённых в эпоху человека видов. Среди них обычны разнообразные
виды, обитающие в пологе леса – эти виды происходят от китайской горлицы. В
высокогорных районах существуют виды голубей, кормящиеся на земле и гнездящиеся
в пещерах и нишах скал – это представители другой родословной линии голубей,
происходящей от сизого голубя. Также на островах имеется эндемичное семейство
ложнодронтовых, которое представляет крупный нелетающий парадодо, обитающий
в кустарниковых зарослях; это также потомок сизого голубя. Певчие птицы представлены
на островах большим количеством видов, часть которых происходит от предков,
завезённых человеком. От японской белоглазки происходит особое новое семейство
– медовые птицы (Melisugornitidae), специализированные на питании нектаром.
Насекомоядные птицы представлены главным образом видами семейства скворцовых.
Среди них имеются специализированные формы – моллюскоядная гавайская крючкоклювка
и муравьиный скворец, питающийся наземными насекомыми. Потомки завезённой индийской
майны занимают экологическую нишу врановых – это всеядные птицы средних размеров,
питающиеся насекомыми и мелкими позвоночными. Вьюрковые птицы и ткачики – основные
группы растительноядных птиц полога леса. Среди них клестовидка – специализированная
форма, питающаяся семенами хвойных деревьев. Пальмовый твердоклюв – другой
пример специализации к питанию семенами. Своеобразным пернатым хищником на
архипелаге является орлиный ворон эндемичного вида и рода, родственный азиатским
формам.
Благодаря тропическому климату рептилии хорошо чувствуют себя на архипелаге
и их видовое разнообразие велико. Наземные рептилии островов происходят исключительно
от видов, ввезённых человеком в историческую эпоху. Наземные черепахи на Гавайях
отсутствуют, но на островах гнездятся крупные морские черепахи-нетопыри, которые
для кладки яиц поднимаются в реки островов и углубляются в лес. Также в пресных
водах обитают мягкотелые черепахи, происходящих от китайского трионикса, завезённого
в эпоху человека. Среди них есть мелкие насекомоядные формы и крупный вид,
конвергентно сходный с африканской черепахой-крокодилом и питающийся крупными
позвоночными. Среди ящериц разнообразны гекконы и потомки анолисов. Гекконы
– это главным образом древесные и скалолазающие формы, преимущественно насекомоядные.
Среди потомков анолисов есть несколько лазающих растительноядных видов и одна
специализированная форма, питающаяся наземными улитками. Фауна змей на Гавайях
достаточно бедна, несмотря на тропический климат. Для лесных почв характерен
подземный коронохвост, крупный вид слепозмеек, питающийся мелкими позвоночными.
На разных островах существует несколько мелких видов слепозмеек, питающихся
насекомыми и незначительно отличающихся друг от друга внешне.
Земноводные Гавайских островов представлены исключительно потомками завезённых
видов: до появления человека земноводные не водились на этих островах. На архипелаге
водятся представители трёх родословных линий, происходящих от жабы аги, североамериканской
лягушки-быка и кубинской квакши. Гавайские жабы обитают главным образом на
суше – в лесной подстилке, среди кустарников и в горных районах, в том числе
в условиях прохладного климата. Квакши – исключительно лесные теплолюбивые
формы, обитающие в пологе леса и на стволах крупных деревьев. Потомки лягушки-быка
– это водные и полуводные формы, населяющие пресные водоёмы и влажные наземные
местообитания.
Пресноводные рыбы на Гавайских островах в неоцене представлены намного богаче,
чем до эпохи человека. Практически все они являются потомками пресноводных
рыб, завезённых человеком. Наиболее многочисленны мелкие живородящие рыбы,
происходящие от пецилий и меченосцев путём естественной гибридизации завезённых
видов. Благодаря способности жить в морской воде они преодолели проливы между
островами и заселили весь архипелаг. Потомок завезённой гамбузии превратился
в активного хищника, обитающего у поверхности воды. В пресных водах также живут
мелкие сомы из семейства Ictaluridae (североамериканского происхождения) и
несколько видов лорикариевых сомов (потомки вида Hypostomus watwata, южноамериканского
происхождения). Распространены цихлиды – потомки африканских тиляпий и южноамериканских
цихлазом. Морские рыбы заходят в реки и какое-то время живут в пресной воде.
Среди них бычки и некоторые элеотрисовые заселили реки островов естественным
путём после формирования нового комплекса пресноводных рыб.
Тропический климат и обильная растительность сделали возможным существование
богатой фауны наземных беспозвоночных. Насекомые на архипелаге очень разнообразны,
и часть их видов происходит от предков, завезённых человеком в историческую
эпоху. Стрекозы очень разнообразны, в равной степени представлены равнокрылые
и разнокрылые. Эти насекомые практически все происходят от местных видов эпохи
человека, или попали на острова уже после вымирания человека. Характерной группой
насекомых являются пилоноги, крупные хищные кузнечики. На Гавайях они мельче,
чем на других островах, где меньше млекопитающих, но видовой состав намного
богаче. Среди гавайских пилоногов есть нелетающие формы, эндемичные для разных
островов и горных долин. Клопы представлены многочисленными хищными и растительноядными
видами, среди которых есть формы, способные переваривать латекс. В пресных
водах обитают клопы-водомерки. Характерными представителями насекомых являются
муравьи многих видов – все они потомки завезённых видов. Чудовищный муравей
– один из крупнейших видов на архипелаге, опасный даже для мелких позвоночных.
Общественные осы, потомки завезённых на острова складчатокрылых ос, образуют
гигантские многолетние колонии. Некоторые виды этих ос нападают на мелких позвоночных
и собирают падаль. Среди плотоядных жуков встречаются мелкие жуки-плавунцы,
попавшие на острова уже после эпохи человека. Светлячки многочисленны и особенно
разнообразны в лесах; среди них появились экологические аналоги жужелиц, стафилинов
и жуков-мертвоедов. Среди разноядных жуков разнообразны жуки-дровосеки и листоеды.
Разнообразны ночные бабочки и бражники, достигающие крупных размеров и часто
специализированные к опылению строго определённых растений. Дневные бабочки
Гавайских островов представлены немногочисленными нимфалидами, белянками и
данаидами (потомками североамериканского монарха, расселившегося на острова
естественным путём). Благодаря человеку на островах появились комары, которые
пережили эпоху массового вымирания и образовали много новых видов в неоцене.
Среди них появились специализированные виды, в том числе питающиеся кровью
улиток и продуктами разложения трупов.
В фауне паукообразных произошли большие перемены. Большинство эндемиков, обитавших
на островах до человека, вымерло, не оставив потомков. Основу фауны паукообразных
составляют потомки завезённых на острова видов, и видов, попавших на острова
естественным путём в раннем неоцене. Среди гавайских пауков есть пауки-волки
(наземные и полуводные виды) и пауки-скакуны (преимущественно древесные формы).
Очень разнообразны пауки-кругопряды, обитающие в лесах и кустарниках.
Благодаря тропическому климату на Гавайях разнообразны наземные крабы – обитатели
лесов, болот и речных долин. Среди них есть падальщики, хищники, и даже пожиратели
помёта местных позвоночных. Большинство крабов уходит для размножения в море,
есть лишь несколько видов, способных проходить полный цикл развития в пресной
воде. Речные раки отсутствуют, но разнообразны креветки. Большинство их видов
встречается в устьях рек и уходит в морскую воду для размножения, но есть несколько
мелких узкоареальных видов, которые стали настоящими пресноводными жителями
с прямым развитием.
До эпохи человека на Гавайских островах обитало много видов наземных брюхоногих
моллюсков. После появления на островах человека многие эндемичные виды исчезли
из-за разрушения среды обитания или конкуренции со вселёнными видами улиток.
Неоценовые наземные улитки Гавайских островов – исключительно потомки форм,
завезённых человеком, или видов, появившихся на островах уже после исчезновения
человека. Некоторые из гавайских улиток ведут хищный образ жизни, и имеются
даже формы, вступившие в симбиотические отношения с муравьями. Существуют вторичноводные
«гавайские прудовики» – мелкие формы, происходящие от завезённой улитки ахатины.
В устьях рек встречаются неритины – брюхоногие моллюски морского происхождения,
способные временно жить в пресной воде. В низовьях рек встречаются немногочисленные
виды двустворчатых моллюсков, способные существовать в опреснённой воде.
 |
 |
Bambusiphagomys tector |
Kapo hawaiiensis |
 |
 |
Araucaricola loxirostra |
Horrohelix ovisuga |
| Гавайские острова, характерные типы растительных сообществ, различные виды растений. | |
| Гавайские острова - животные влажного тропического леса. | |
| Гроза серебряного леса | Сообщество высокогорных тропических лесов Гавайских островов - беспозвоночные и мелкие позвоночные. |
| Зелёная жемчужина Тихого океана: играющие в волнах | Гавайские острова: обитатели океанского побережья и рифов. |
Океания
|
В неоцене южная часть Тихого океана продолжает оставаться
крупнейшим на Земле скоплением островов – здесь их насчитывается более 15 тысяч.
Острова Океании группируются в три большие области: Меланезия на западе Тихого
океана, Микронезия на северо-западе и Полинезия на востоке. В каждой из этих
областей с голоцена произошли определённые изменения. Так, Меланезия лишилась
части островов (а именно северных Соломоновых) за счёт их присоединения к Меганезии.
Микронезия и Полинезия в эпоху человека включали множество островов кораллового
и вулканического происхождения, разбросанных по просторам Тихого океана, главным
образом, в тропиках, где могли расти кораллы-рифостроители. На рубеже голоцена
и неоцена кораллы-рифостроители вымерли из-за глобального экологического кризиса,
и значительное число коралловых островов исчезло. Другой причиной исчезновения
части низких островов стало их затопление вследствие повышения уровня Мирового
океана, связанного с неоценовым потеплением климата и таянием полярных льдов.
В неоцене, когда сложилась новая фауна животных-рифостроителей, мелкие острова
рифового происхождения вновь появились примерно в тех же местах, где до этого
были коралловые острова, и даже наросли на их старых известняковых фундаментах.
Вулканические острова располагаются главным образом в местах стыка литосферных
плит – Тихоокеанской, Индо-Австралийской и Филиппинской. Вулканические острова
более высокие и обладают большей площадью. На них часто имеются действующие
или потухшие вулканы.
Хотя большинство островов Океании имеет коралловое и вулканическое происхождение,
существует одно исключение: остров Новая Каледония в Меланезии, являющийся
осколком древнего материка. Климат островов Океании в целом одинаковый, тропический,
различающийся лишь количеством дождей. Лишь на некоторых южных островах тропический
климат сменяется субтропическим. Для многих островов Океании характерны разрушительные
тайфуны и ураганы.
Пресноводные водоёмы на островах чаще всего отсутствуют или представляют собой
мелководные ручьи и озёра на самых крупных островах. На мелких островах единственным
источником пресной воды являются дожди.
Очерк написан при участии Семёна, участника форума.
Флора Океании относительно бедна по сравнению с континентальной.
Здесь отсутствуют многие группы растений, особенно распространяющиеся с помощью
наземных животных. Большинство видов местной флоры расселяется с помощью
ветра или птиц, а также с помощью океанских течений. Немалую роль в формировании
островной флоры сыграла деятельность человека в далёком прошлом – часть местных
растений является потомками культурных растений эпохи человека.
Наиболее характерным типом растительных сообществ являются мангровые заросли,
образованные деревьями нескольких видов, относящимся к нескольким различным
семействам. Значительного распространения в Океании (особенно в Полинезии)
достиг особый вид пальм – морской кокос, обладающий ползучими стволами и ходульными
корнями.
Этот вид
растений образует большие заросли на морских мелководьях. В прибрежных районах
островов обширные заросли образуют пальмы нескольких видов и панданус.
На низких островах рифового происхождения грунтовые воды солёные; в таких местах
могут расти лишь солеустойчивые растения и небольшие кустарники и травянистые
растения, корни которых располагаются выше уровня грунтовых вод. Флора высоких
островов вулканического происхождения намного богаче. В отдалении от берега
на возвышенных участках образуются густые вечнозелёные леса. Их образуют виды,
попавшие на острова естественным путём, а также потомки завезённых видов. На
повреждённых участках леса пионерной растительностью являются быстрорастущие
виды банана и папайи. Их сменяют многолетние кустарники семейств никтагиновых
(пизония, известная с эпохи человека) и мальвовых (гибискусы нескольких местных
видов). Некоторые из кустарников разрастаются в низкорослые деревья. Деревья
туземной флоры принадлежат к семействам молочайных, кутровых и некоторых других.
Среди древесных растений Океании есть представители семейств сложноцветных
и колокольчиковых. Потомки завезённой человеком флоры в полинезийских лесах
– манго, казуарины, виды-потомки хлебного дерева и одичавших цитрусовых. Очень
обычны на островах виды миконии (семейство меластомовых), широко распространившейся
в эпоху человека и трудно искореняемой. Больше потомков туземной флоры среди
кустарников и травянистых растений.
На старых островах вулканического происхождения встречаются растения семейства
бромелиевых – потомки культурного ананаса с мелкими соплодиями и розетками
длинных листьев. Повсеместно в Океании встречаются потомки таро (Alocasia),
достигающие особенного развития на болотах некоторых островов. Также обычны
потомки батата – ползучие клубневые растения. Среди травянистых растений встречаются
несколько видов орхидей, в том числе потомки ванили – крупные лианы и полуэпифиты.
Папоротники особенно многочисленны на островах, где растут густые влажные леса.
Большинство видов является эпифитами, а некоторые произрастают в подлеске,
образуя обширные заросли благодаря теневыносливости. В южной части Океании
есть несколько видов древовидных папоротников новозеландского происхождения.
Фауна островов Океании бедна наземными животными из-за географических
особенностей этого региона. Удалённость от материков и большие расстояния,
разделяющие острова, препятствуют заселению островов большинством групп животных.
Характерной особенностью зоогеографии Океании является то, что по мере движения
на восток уменьшается количество таксонов животных.
Млекопитающие на островах очень редки. На большинстве островов
обитают исключительно летучие мыши и крыланы, способные расселяться самостоятельно.
На старых прочных островах вулканического происхождения живут грызуны – многочисленные
потомки крыс, завезённых в эпоху человека. Из хищников на некоторых островах
обитают кошки, а на островах Фиджи – мангусты, потомки завезённых человеком
видов. На некоторых островах Меланезии сохранились мелкие потомки одичавших
свиней.
Фауна птиц Океании гораздо богаче, чем млекопитающих. Богаче
всего на островах представлены птицы морского происхождения – трубконосые,
веслоногие, чайки и крачки. Необычной группой морских птиц являются коршуны-серафимы,
или эфирные коршуны – длиннокрылые морские птицы, приспособленные к длительным
перелётам и ловле рыбы на лету. Кулики в основном представлены пролётными видами,
но на островах, лишённых млекопитающих, живут эндемичные виды куликов из семейств
ржанковых и бекасовых. На островах Океании встречается много зимующих видов
морских птиц из Северной Америки и Азии.
Наземные птицы более разнообразны на островах, где отсутствуют наземные млекопитающие.
Султанки и пастушковые вновь расселились по островам, особенно по вновь образовавшимся,
где нет наземных хищников. В отличие от эпохи человека, на большинстве островов
они сохранили способность летать и навыки самозащиты. Некоторые пастушковые
приобрели способность охотиться на крыс. Голенастые представлены широко распространёнными
видами цапель. На некоторых островах цапли образовали эндемичные виды. На многих
островах обитают утки, в том числе узкоареальные виды. Из куриных птиц на западе
Полинезии и в Меланезии встречаются потомки одичавших домашних кур, в том числе
довольно крупные, и потомки перепелов. В некоторых частях Меланезии сохранились
потомки сорных кур. Хищные птицы немногочисленны: в лесах Микронезии, Меланезии
и на западе Полинезии живут эндемичные виды ястребов и мелких соколов, несколько
шире распространены луни. На островах обычны голуби; есть несколько эндемичных
мелких курро. На некоторых островах встречается несколько видов очень крупных
плохо летающих наземных голубей. Ещё одной характерной группой полинезийских
птиц являются попугаи. В эпоху человека вымерло много видов этих птиц, но в
неоцене часть азиатских и австралийских видов вновь заселила острова, образовав
много эндемичных видов. На островах Меланезии обитают иглоногие совы, дальше
на восток распространены сипухи. Ракшеобразные представлены несколькими видами
зимородков, образующих эндемичные формы видового и подвидового ранга на отдалённых
островах. В Микронезии помимо зимородков встречаются щурки и широкороты. Представители
отряда козодоев встречаются в Меланезии, Микронезии и западной Полинезии. До
самого востока Полинезии распространены стрижи и кукушки.
Острова населяют многочисленные певчие птицы. Наиболее широко распространены
ласточки, скворцы (в том числе потомки завезённых человеком майн и обыкновенных
скворцов), камышевки, белоглазки, астрильды, медососы, мухоловки (и близкие
к ним семейства, такие как веерохвостки, монархи, австралийские малиновки и
другие). Менее широкое распространение имеют птицы из семейств личинкоедов,
дронго, дроздов и некоторых других. Потомками завезенных человеком видов являются
бюльбюли и ткачиковые (происходящие от воробьёв). Врановые распространены довольно
широко, они происходят от южноазиатских и австралийских видов, а также от новокаледонской
вороны. Азиатские сороки также смогли распространиться по некоторым островам
Океании и образовать эндемичные виды.
Разнообразие рептилий на островах не очень велико, что связано
главным образом с трудностями расселения по островам. На большинстве островов
обитают мелкие ящерицы – гекконы и сцинки. По соседству с млекопитающими обитают
главным образом подвижные мелкие виды, а на островах, где они отсутствуют,
обитают более крупные виды этих ящериц. Эндемичные игуаны островов Фиджи не
оставили потомков в неоцене. На некоторых островах Микронезии живут вараны.
Среди змей многочисленны морские, а на севере Меланезии живут змеи, происходящие
от местных видов. Существование на некоторых других тихоокеанских островах
наземных змей связано с деятельностью человека: на Гуам были завезены бойги,
а на Фиджи и некоторые другие острова – слепозмейки. Крупнейшей рептилией Полинезии
является кархарозух – один из последних видов крокодилов, крупнейший неоценовый
представитель этого отряда. Он гнездится на островах Тонга, где грунт подогревается
вулканическим теплом. Также на островах южной части Тихого океана находятся
места размножения морских черепах-нетопырей.
Земноводных на островах Океании почти нет. На крупных островах
(например, на Фиджи) живут потомки видов, завезённых человеком: квакши (потомки
австралийских видов) и жабы (потомки жабы-аги). На некоторые острова земноводные
попали уже после вымирания человека, например, в виде икры, прилипшей к лапкам
птиц, или на плотах из растительности.
Пресноводные рыбы отсутствуют, но в пресные воды заходят некоторые виды морских
рыб.
Благодаря тропическому климату фауна беспозвоночных богата
и разнообразна. Среди островных видов примечательны крупные хищные кузнечики
пилоноги, способные нападать даже на мелких млекопитающих. Есть несколько видов
этих насекомых, способных перелетать с острова на остров, а также ряд нелетающих
островных видов, достигающих более крупных размеров. На островах, находящихся
ближе к материкам, встречаются палочники и клопы, часто причудливой формы.
Частыми обитателями островов Полинезии являются жуки и бабочки, среди которых
много эндемичных видов. Двукрылые разнообразны и встречаются практически на
всех островах. Личинки ряда видов играют большую роль в почвообразовании.
Часть беспозвоночных является выходцами из моря: это крабы и другие ракообразные.
Наземные крабы и раки-отшельники очень широко распространены благодаря наличию
в жизненном цикле стадии плавающей личинки. Крабы являются мелкими хищниками
и охотятся на местных насекомых, грызунов и мелких птиц, а раки-отшельники
подбирают гниющие органические остатки растительного и животного происхождения.
Наземные улитки представлены большим количеством эндемичных видов. На каждом
острове или архипелаге имеется целая группа видов, родственных друг другу и
представляющих собой результат единственной удачной попытки колонизации данной
территории. Очевидно, их расселение происходило на плавающих стволах деревьев
или на ногах и оперении птиц. На крупных островах обитают потомки улиток-ахатин,
завезённых человеком. Пресноводные улитки отсутствуют.
Очерк написан при участии Семёна, участника форума.
 |
 |
Dryorattopsis mustelinus |
Seraphimornis gracilissimus |
 |
 |
Carcharosuchus deinodontus |
Serratipodus dracocephalus |
| Тропики Тихого океана, острова Тонга |
Галапагосские
острова
|
На протяжении голоцена и неоцена Галапагосские острова оставались
источником вулканической активности из-за постепенного движения плиты Наска,
уходящей краем под Южноамериканскую плиту. Частые извержения вулканов и землетрясения
вызвали изменение облика островов. Несколько маленьких островков архипелага
утонули, а часть молодых вулканически активных островов (Исабела, Фернандина,
Сантьяго и Санта-Крус) слилась воедино. В неоцене на островах часто происходят
землетрясения, а некоторые вулканы регулярно извергаются. Горы островов стали
значительно выше и выполняют важную функцию конденсаторов влаги.
Климат островов остаётся тропическим, но, в связи с перемещением Южной Америки
к югу он стал несколько более засушливым. Климат в значительной степени зависит
от холодного Перуанского течения. Контраст температур между морем и сушей приводит
к образованию утренних туманов в прибрежных районах островов. На островах нет
рек, а неглубокие пруды в низинах наполняются только дождевой водой.
В некоторых горных долинах, защищённых от ветра, складывается влажный микроклимат
благодаря находящимся там болотам и небольшим озёрам.
В связи с засушливым климатом островов местная растительность
несёт черты приспособления к интенсивному поглощению и сохранению воды. Растительность
прибрежных районов приспособлена к улавливанию влаги туманов: прибрежные деревья
и кустарники (главным образом из семейства бобовых) имеют листья, способные
складываться вдоль. По утрам такие листья раскрыты и поглощают капли росы,
а на день складываются, защищая верхнюю сторону от солнца. У других растений
розетка листьев раскрывается ночью, поглощая влагу, и складывается днём, находясь
под защитой грубых и колючих наружных листьев. Прибрежные виды кактусов впитывают
воду густым опушением на стебле.
В сухих областях крупных островов, а также на небольших низких островах складывается
комплекс тропических редколесий, где преобладают бобовые растения (потомки
завезённого рожкового дерева), древовидные сложноцветные и кактусы. На участках
с бедной почвой образуются сплошные заросли злаков, потомков завезённой в эпоху
человека слоновой травы (Pennisetum).
Бобовые попали на острова с помощью человека (рожковое дерево) или расселились
естественным путём с материка по морю уже в раннем неоцене. Они достигли успеха
в борьбе за существование благодаря наличию грызунов, ставших распространителями
их крупных семян. Древовидные бобовые Галапагосских островов часто образуют
жизненную форму «бутылочного дерева» с рыхлой водозапасающей сердцевиной. Сложноцветные
отличаются многочисленными колючками, покрывающими стебель и главные жилки
листьев, особенно в молодом возрасте. Кактусы (цереусы и опунции) образуют
крупные заросли благодаря способности к вегетативному размножению.
Высокогорные районы островов более влажные; здесь произрастают густые леса
с сомкнутым пологом. Виды растений здесь часто родственны видам из сухих редколесий,
но обладают менее развитыми водозапасающими тканями, крупными листьями и мелкими
колючками. Среди местных кактусов примечательна эквадопунция – древовидный
кактус с опадающими однолетними побегами. В лесах обитает очень ядовитое манцинелловое
дерево рода Hippomane – потомок вида, произраставшего на островах в эпоху человека.
Во влажных горных долинах произрастают папоротники. Эпифитные растения немногочисленны
– это несколько видов папоротников и бромелиевых.
Важным компонентом галапагосских лесов являются пальмы, расселившиеся на острова
по морю. Они образуют небольшие заросли или встречаются одиночными экземплярами.
На прибрежных мелководьях произрастают мангровые леса. На берегах также встречаются
заросли морского кокоса (архипелаг находится на восточной оконечности ареала
этой пальмы).
В эпоху человека эндемичной флоре и фауне был нанесён сильный
удар – человек завёз на острова большое количество видов наземных млекопитающих,
растительноядных и хищных. Благодаря природоохранным мероприятиям на островах
были истреблены одичавшие козы и собаки, но крысы остались, сильно размножились
и в дальнейшем эволюционировали, образовав несколько новых видов.
Среди млекопитающих Галапагосских островов наиболее разнообразны потомки крыс,
ведущие различный образ жизни и приспособившиеся к различным условиям обитания.
Крупный медлительный вид крыс питается твёрдыми семенами пальм и бобовых, являясь
аналогом гавайского капо. Мелкие наземные и древесные виды питаются семенами
и насекомыми. Из хищников на островах остались потомки домашних кошек, образовавшие
несколько видов. Один из них мелкий, лазающий, похожий на куницу, а другой
более крупный наземный. На разных островах виды образуют местные формы, отличающиеся
размерами и пропорциями тела.
Свиньи остались в фауне островов, образовав несколько мелких островных форм,
сходных с карликовыми видами островов Крым и Маврикий.
Фауна птиц архипелага очень разнообразна, но отличается от
той, что существовала в эпоху человека. На островах обитают цапли нескольких
видов, в том числе один мелкий и коротконогий лесной вид, похожий на пастушковых
и питающийся грызунами и крабами. Эти птицы сдерживают размножение грызунов
и способствуют поддержанию биологического равновесия на островах. Крупные цапли
питаются главным образом насекомыми и грызунами. Ещё один вид цапель обитает
в мангровых зарослях и ищет корм в приливно-отливной полосе.
Морские птицы разнообразны: на островах встречаются разные виды чаек, в том
числе ночная чайка, скрывающаяся днём в убежищах среди скал. Многие виды крачек
для защиты от наземных хищников гнездятся на деревьях. На островах гнездятся
крупные олуши, утратившие способность к полёту. Они образуют плотные колонии
на островах, где нет млекопитающих. На побережьях разнообразны кулики, в том
числе перелётные, мигрирующие на зиму из Северной Америки.
Другими мигрантами, с юга, являются тихоокеанские морские лебеди. Утки редко
встречаются на островах; в основном это случайно залетающие и мигрирующие виды.
Это связано с малым количеством пресных водоёмов, пригодных для их жизни.
Из хищных птиц на островах встречаются один вид настоящих сов и каракара местного
эндемичного вида.
Попугаи, трогоны, ракшеобразные и дятлообразные птицы на Галапагосских островах
отсутствуют.
Воробьинообразные птицы немногочисленны по сравнению с Южной Америкой. Среди
непевчих воробьиных имеется несколько видов тираннов. Певчие воробьиные более
многочисленны, представлены несколькими семействами. Как и в эпоху человека,
острова населяют разнообразные вьюрки, прямые потомки земляных вьюрков эпохи
человека. Они так же занимают разные экологические ниши и различаются строением
и поведением. Среди них есть вид, являющийся гнездовым паразитом, а также ночной
вампир – своеобразная птица-гематофаг. Из других певчих птиц на островах обитают
пересмешники и танагры.
Фауна рептилий претерпела значительные изменения по сравнению с эпохой человека.
Слоновых черепах, знаковой группы обитателей Галапагосских островов, осталось
очень мало – значительное количество их подвидов вымерло в эпоху человека или
вскоре после неё из-за климатических изменений и конкуренции с наземными млекопитающими.
Один из немногих представителей слоновых черепах – шипоголовая черепаха. Кроме
неё, на островах обитает ещё несколько видов, отличающихся размерами и формой
панциря. Чешуйчатые представлены гекконами и несколькими видами тейид, а также
мелкими наземными игуанами, поселившимися на островах по окончании эпохи человека.
Крупные наземные игуаны конолофы вымерли, а морские игуаны стали более редкими.
Эти рептилии обитают на многих островах архипелага, но мигрируют для размножения
на мелкие острова, лишённые млекопитающих.
Земноводные и настоящие пресноводные рыбы на архипелаге отсутствуют.
Беспозвоночные Галапагосских островов не очень разнообразны по сравнению с
фауной Южной Америки. Острова никогда не сообщались с материком, и их фауна
складывается из двух компонентов: из потомков видов, попавших на острова в
разное время естественным путём, и потомков завезённых в историческую эпоху
видов.
Среди насекомых архипелага много потомков завезённых видов. Большинство местных
тараканов составляют мелкие виды, потомки завезённого рыжего таракана. Разнообразие
жуков невелико; среди них встречаются несколько видов жуков-усачей и листоедов.
В эпоху человека на острова попали жуки-водолюбы, населяющие стоячие водоёмы
островов. Есть несколько видов божьих коровок, окраска которых варьирует от
острова к острову. В эпоху человека на острова попали с культурными растениями
различные червецы и щитовки; их потомки составляют пищу некоторых островных
божьих коровок. Среди клопов есть несколько видов щитников и хищнецов. В береговой
полосе вблизи колоний птиц обитают паразитические клопы – потомки постельного
клопа эпохи человека. Бабочки сравнительно немногочисленны, среди них отсутствуют
специализированные виды. Есть несколько видов данаид, совок, пядениц и сатурний.
Некоторые бабочки вступили в симбиоз с муравьями, которые выкармливают и защищают
их личинок. Перепончатокрылые представлены осами и муравьями. Осы-полисты,
потомки завезённого в эпоху человека вида, образуют небольшие колонии на деревьях.
В лесах также встречаются крупные колонии бумажных ос, также потомков завезённого
в эпоху человека вида. Повсеместно на островах распространены хищные виды муравьёв
– потомки завезённых людьми огненных муравьёв. Разнообразие двукрылых невелико;
среди них имеются кровососущие виды (слепни и комары), питающиеся на теплокровных
животных, и несколько видов, питающихся нектаром и органическими остатками.
Из прочих членистоногих на островах живут крупные сколопендры, потомки местного
вида, населявшего острова в эпоху человека. Также есть несколько видов пауков-кругопрядов.
Из членистоногих морского происхождения на островах обитает несколько видов
крабов семейства грапсид, уходящих для размножения в море.
Наземные улитки представлены несколькими мелкими видами.
 |
Cuculispiza parasitica |
Острова Эонезии
|
В результате продолжающейся вулканической активности по линии
Восточно-Тихоокеанского поднятия (между плитами Тихоокеанской и Кокос, а далее
на юг – между Тихоокеанской и Наска) в неоцене появилась новая, пока ещё прерывистая
цепочка островов. Она протянулась от Калифорнии на юг в сторону острова Пасхи
почти в меридиональном направлении. В эпоху человека этот район был местом
высокой вулканической активности: вдоль него располагалось большое количество
подводных гидротермальных источников и «чёрных курильщиков». В неоцене подводные
горные хребты во многих точках достигли поверхности воды и образовали острова,
получившие название Эонезия. “Eos” по-гречески означает «заря», а также «ранний»,
что одновременно указывает на расположение островов в восточной части Тихого
океана, а также на их молодость в геологическом плане. Из-за быстрого движения
океанических плит образование островов происходило сравнительно медленно, а
сами острова разделены на три группы, достаточно далеко отстоящие друг от друга.
На большей части архипелага между ними и побережьем Америки другой суши нет.
Южнее Калифорнии (ставшей в неоцене островом) располагается группа островов
Атарапа. Это сравнительно небольшие вулканические острова, между которыми есть
отмели и подводные горы. Название «Атарапа» происходит от имени полинезийской
богини рассвета, поскольку острова находятся на востоке Тихого океана.
Большая группа островов, острова Фисага (в полинезийской мифологии – имя бога-морского
бриза), находится чуть севернее экватора, на стыке трёх литосферных плит: Тихоокеанской,
Кокос и Наска. Ближайшей к ним сушей являются Галапагосские острова. Вулканическая
активность здесь наиболее высокая.
Несколько небольших островов, острова Буроту (райские острова в мифологии фиджийцев),
разбросаны цепочкой в Тихом океане, протянувшись от экватора до острова Пасхи,
который является самой южной точкой островов Эонезии.
Поскольку все острова имеют вулканическое происхождение, на них в большей или
меньшей степени проявляется вулканическая активность: землетрясения, горячие
источники и более или менее регулярные извержения вулканов. Все острова расположены
в тропических широтах, поэтому климат на них везде тропический, с небольшими
различиями в количестве осадков. Северная часть островов Эонезии находится
вблизи американского побережья, и близость массива суши оказывает влияние на
климат островов: здесь количество осадков меняется по сезонам, и зимой с суши
дует прохладный сухой ветер. Проходя над океаном, он не успевает получить достаточно
влаги, поэтому зимой погода на островах Атарапа более сухая, чем летом.
Группа островов Фисага располагается значительно дальше от материка, и их климат
всецело зависит от океана. Он ровный, с большим количеством осадков, способствующих
развитию растительности.
Острова Буроту также отличаются тропическим морским климатом с высокой влажностью
воздуха и большим количеством осадков.
Все острова Эонезии окружены полосой рифов, образованных моллюсками, червями
и губками. На севере рифы образованы небольшим количеством видов, и там важную
роль играют водоросли и морские травы, а на экваторе и в южном полушарии рифы
образованы почти исключительно животными-рифостроителями, и острова вулканического
происхождения окружены широкой полосой отмелей и островков рифового происхождения.
Видовое разнообразие растений, произрастающих на островах,
напрямую зависит от расстояния до ближайшей суши, а также от широты.
Группа островов Атарапа демонстрирует видовой состав флоры, близкий к североамериканскому.
Здесь присутствуют потомки лиственных и хвойных деревьев, способные расселяться
с помощью ветра и птиц, а также переносить зимнюю сухость. На возвышенностях
островов произрастают сосны, а сухие прибрежные районы занимают заросли кактусов
нескольких видов, попавших сюда, очевидно, с острова Калифорния благодаря птицам.
Лиственные деревья и вечнозелёные кустарники растут главным образом в горных
долинах, где локально складывается влажный микроклимат.
Острова Фисага демонстрируют богатство растительного мира. Это связано с расположением
островов в области экваториального климата, способствующего развитию вечнозелёных
широколиственных лесов. На побережье островов произрастают морской кокос, образующий
естественный волнолом, и несколько видов мангровых деревьев, поселяющихся между
сушей и зарослями морского кокоса. Благодаря птицам на острова попали семена
деревьев нескольких семейств, а ветром были принесены споры папоротников. В
видовом составе флоры островов Фисага заметно влияние Галапагосских островов
– ближайшей суши. В горных районах островов на сухих почвах и каменистых осыпях
произрастают кактусы, а в лесах растут деревья видов и родов, близких к галапагосским.
Влияние флоры Океании проявляется в наличии ряда общих видов, включая потомков
миконии.
Флора островов Буроту в целом близка к флоре Океании, отличаясь на уровне близких
видов и родов. На островах доминируют пальмы и панданус; некоторые виды растений
появились на островах благодаря летучим мышам. Среди травянистых растений необычно
наличие тростника, являющегося потомком вида, произраставшего на озере Титикака
в эпоху человека. Этот тростник расселился по островам Буроту на север с острова
Пасхи, куда, скорее всего, был завезён человеком.
Поскольку острова Эонезии сформировались уже после окончания
эпохи человека, их заселение животными происходило естественным образом. По
таксономическому составу фауна островов сходна с типичной фауной островов голоцена
до их колонизации человеком.
Млекопитающие на островах Эонезии очень малочисленны. На островах
Атарапа обитает несколько видов грызунов и один вид кроликов; их предки попали
сюда с острова Калифорния. Также здесь обитают рукокрылые североамериканского
происхождения, как оседлые, так и перелётные. На архипелагах Фисага и Буроту
млекопитающие представлены только рукокрылыми центрально- и южноамериканского
происхождения, расселение которых происходило с севера на юг. Большинство видов
этих животных осталось насекомоядными или фруктоядными. Один из видов на островах
Фисага стал крупным и в значительной степени утратил способность летать. Этот
вид лазает по веткам и лишь немного перепархивает с дерева на дерево, но хорошо
бегает и ходит по земле, опираясь на крупный большой палец крыла.
Благодаря отсутствию конкуренции с млекопитающими фауна птиц
островов Эонезии включает некоторое количество крупных и нелетающих видов. На
всех островах распространены морские птицы: чайки и кулики, а также трубконосые
– буревестники и ложные альбатросы, родственные крупному субантарктическому
виду, но более мелкие. Благодаря отсутствию млекопитающих ряд видов птиц утратил
способность к полёту: на островах Буроту обитает несколько подвидов нелетающих
олуш, а на архипелаге Фисага водится крупный нелетающий баклан. На островах
Атарапа обитает цапля, утратившая способность летать и собирающая корм на суше.
Там же живут кукушки и несколько видов клинохвостых попугаев центральноамериканского
происхождения. Хищные птицы островов Атарапа представлены мелким видом настоящих
сов, крупной сипухой и крупным коршуном, который часто ищет корм на земле. В
прибрежных зарослях гнездится один вид эфирных коршунов. На этих островах обитают
куриные птицы – зубчатоклювые перепела, расселившиеся сюда с острова Калифорния.
Певчие птицы островов Атарапа разнообразны; их фауна на протяжении долгого времени
пополнялась за счёт перелётных видов или случайных переселенцев из Северной
Америки. Здесь живут кардиналы, танагры, вьюрковые, трупиалы и врановые. Из
перелётных видов характерны ласточки и стрижи.
Благодаря более мягкому и влажному климату растительность островов Фисага значительно
обильнее, что способствует видовому разнообразию птиц. На островах Фисага морские
птицы образуют многочисленные колонии; здесь есть несколько эндемичных видов
куликов, чаек и крачек. Есть местный вид цапель, распространённый на всех островах
архипелага и сохранивший способность к полёту. На островах обитают крупные утки,
родственные кряквам эпохи человека, которые утратили способность летать и являются
самыми крупными растительноядными птицами островов. Среди лесных птиц островов
Фисага есть трогоны, происходящие от видов Центральной Америки; несколько видов
этих птиц близкородственны друг другу и представляют собой результат одной удачной
попытки колонизации островов. Также на островах Фисага обитает несколько видов
попугаев и крупных лесных голубей. Воробьинообразные птицы представлены несколькими
видами танагр, демонстрирующих явление адаптивной радиации – среди них появились
специализированные плодоядные, насекомоядные и нектароядные виды. Кроме них
на островах обитает несколько видов пересмешников и дроздов. На островах Фисага
обитает один эндемичный оседлый вид ласточек и несколько перелётных.
Острова Буроту, наиболее удалённые от материков, обладают типичной островной
фауной, включающей высокоспециализированные эндемичные виды. Помимо морских
птиц, на островах живут крупные наземные голуби, питающиеся плодами и семенами.
В лесах островов живут крупные нелетающие (но хорошо лазающие по деревьям) попугаи,
являющиеся своеобразными экологическими аналогами обезьян. Также на островах
Буроту водится несколько видов нелетающих куликов, похожих на киви; это виды
американского происхождения, предки которых прилетали на острова на зимовку.
Главным хищником островов является плохо летающая наземная чайка, утратившая
перепонки между пальцами. Певчие птицы островов Буроту немногочисленны и относятся
к семействам танагр и вьюрковых (близкородственных видам с Галапагосских островов).
Рептилии островов Эонезии демонстрируют снижение разнообразия
по мере удаления от материка и движения с севера на юг, что связано с возрастанием
трудности расселения по островам.
Острова Атарапа обладают наиболее богатой фауной рептилий, что объясняется близостью
материка. Здесь живут мелкие игуаны и гекконы, а также крупная змееподобная
веретеница. Также на этом архипелаге живут наземные черепахи, потомки коробчатых
черепах (Terrapene), попавшие сюда из Северной Америки. Из-за конкуренции с
птицами и млекопитающими фауна островов Атарапа не содержит гигантских представителей
рептилий, характерных для островных фаун.
На островах Фисага обитает крупный наземный сцинк центральноамериканского происхождения,
а также мелкие анолисы нескольких родственных видов. Кроме них, на островах
обитают крупная всеядная наземная игуана и крупный варанообразный представитель
тейид, главный наземный хищник острова.
Острова Буроту отличаются бедной фауной рептилий. Здесь обитают только несколько
мелких партеногенетических ящериц семейства тейид южноамериканского происхождения,
расселившихся через Галапагосские острова.
Амфибии и пресноводные рыбы отсутствуют на
всех островах.
Фауна беспозвоночных островов Эонезии сравнительно бедная,
что связано с труднодоступностью островов для наземных беспозвоночных.
Наиболее разнообразна на уровне отрядов и семейств фауна островов Атарапа, но
на островах Фисага и Буроту более разнообразен видовой состав насекомых, относящихся
к немногим семействам. Характерным для островов Тихого океана в целом является
семейство пилоногов – крупных хищных прямокрылых. Они распространены повсеместно,
образуя на островах ряд нелетающих видов. Стрекозы встречаются на крупных островах,
где есть постоянные источники пресной воды. Они образуют ряд эндемичных видов
на островах Атарапа и Фисага. Из других хорошо летающих насекомых на островах
распространены бабочки семейств нимфалид и парусников, а также разнообразные
совки и моли. Жуки многочисленны, на островах часто встречаются крупные формы,
утратившие способность к полёту. Встречаются представители семейств долгоносиков,
щелкунов, дровосеков и пластинчатоусых. Личинки щелкунов играют важную роль
в почвообразовании. Из хищных жуков на островах встречаются плавунцы, стафилины
и светляки. На всех островах Эонезии обитают муравьи; они более многочисленны
на архипелагах Атарапа и Фисага, откуда происходило расселение этих насекомых
на юг.
Паукообразные на островах представлены пауками и клещами. Многоножки обитают
только на островах Атарапа (кивсяки и сколопендры) и Фисага (только сколопендры).
Острова Буроту недоступны для этой группы членистоногих из-за удалённости от
материка и друг от друга.
Крабы – важный компонент островной фауны наземных членистоногих. Большинство
видов этих ракообразных общие для всех островов, поскольку личинки этих животных
переносятся течениями, но на островах Атарапа фауна наземных ракообразных значительно
беднее, чем на юге. Очевидно, это связано с сухой зимой и богатой фауной наземных
позвоночных. На островах Фисага и Буроту, кроме крабов, обитают наземные раки-отшельники.
Улитки многочисленны на островах Атарапа, наиболее близких к материку и острову
Калифорния. По мере движения на юг разнообразие улиток на уровне родов падает.
Острова Буроту населены наземными улитками только одного рода с несколькими
узкоареальными видами, эндемичными для каждого из островов.
Дождевые черви имеются только на островах Атарапа, куда попали с материка, очевидно,
с помощью птиц.
 |
 |
 |
Taaroa involans |
Pezolarus miru |
Infantopsitta raphomima |
| Пленники рая | Острова Буроту в Тихом океане, птицы и другие животные тропического леса |
Острова
Индийского океана |
 |
 |
Pseudonoctilio laroides |
Thalassagrion maritimus |
 |
 |
Cancerotalpa cryptobia |
Venatocancer terrestris |
| Индийский океан - острова и их обитатели | |
| Индийский океан - морские мелководья |
Маврикий и Маскаренские
острова
|
В неоцене Маскаренские острова немного сдвинулись к востоку
из-за движения тектонических плит, но в целом с точки зрения географии этот
архипелаг мало изменился по сравнению с эпохой человека. Он по-прежнему состоит
из нескольких десятков островов, крупнейшими из которых являются Маврикий,
Реюньон и Родригес. Будучи вулканическими по происхождению, эти острова не
разрушились, подобно большинству небольших островов Индийского океана, известных
в эпоху человека. Однако вулканическая активность на них не проявлялась со
времени вымирания человека. Некоторые острова, входящие в состав архипелага,
построены организмами-рифостроителями уже после исчезновения человечества.
Рельеф островов сглажен эрозией, для крупных островов характерны обширные внутренние
плато и невысокие горные вершины. Внутренние плато окружены прибрежными равнинами.
Климат Маскаренских островов тропический, с большим количеством осадков. Сезонность
климата выражена слабо (существует длительный влажный и непродолжительный относительно
сухой сезон), но на погоду оказывают большое влияние циклоны, зарождающиеся
в Индийском океане в начале года.
С возвышенностей крупных островов стекают многочисленные небольшие ручьи и
реки, есть мелководные озёра и болота. На многих мелких островах существуют
лишь временные водоёмы, наполняющиеся водой только после дождя.
Очерк написан Семёном, участником форума
Флора островов претерпела резкие изменения, начиная с эпохи
человека. В историческое время произошло масштабное вымирание видов местной
флоры, связанное с деятельностью завезённых видов травоядных (копытных) и завозом
большого количества чужеродных видов растений. На одном из этапов становления
новой флоры произошло полное вымирание старой флоры, сложившейся до исторической
колонизации. В это время острова были покрыты редколесьями, среди которых бродили
потомки завезённых животных, не позволявшие развиваться сплошному тропическому
лесу. Позже, когда травоядные сильно измельчали, флора островов постепенно
восстановилась, но в ней присутствовали исключительно потомки завезённых видов
растений, которые смогли выдержать вытаптывание и объедание со стороны копытных.
Маврикийской флоре придают своеобразие древовидные потомки травянистых растений;
один из таких видов – ложноягодник маврикийский. Характерными представителями
флоры островов являются древовидные молочайные с едким соком, а также древовидные
сложноцветные. На островах встречается несколько видов бегониевых деревьев.
На возвышенностях крупных островов Маскаренского архипелага господствует тропический
лес с сомкнутыми кронами. Вершины гор занимает лес с густым папоротниковым
подлеском. Побережья островов окаймлены обширными зарослями кустарников с небольшими
группами пальм. Пальмы являются доминирующими растениями на небольших островах.
На побережьях крупных островов и почти на всех мелких островках в составе архипелага
имеются участки мангровых лесов.
Травянистые растения Маскаренских островов – это преимущественно сложноцветные
и орхидеи, среди которых имеются эпифитные виды. Также на островах произрастают
злаки, особенно многочисленные в горных районах, где они образуют пояс саванноподобной
растительности в сухих местах.
Фауна Маскаренских островов не отличается большим количеством
видов, однако подавляющее их большинство являются эндемиками. Почти вся аборигенная
фауна вымерла из-за охоты, уничтожения мест обитания и негативного влияния
завезённых человеком видов. После исчезновения человечества восстановление
фауны происходило за счёт потомков завезённых видов, а также более поздних
мигрантов из Африки, с Мадагаскара и отчасти из Южной Азии. Лишь некоторые
виды рептилий и единичные виды птиц ведут свой род от аборигенных видов.
Единственными млекопитающими Маскаренских островов, которые добрались до архипелага
самостоятельно, являются рукокрылые. Крыланы эндемичных видов образуют многочисленные
колонии в лесных районах. Также на архипелаге живут летучие мыши, в том числе
необычные очень крупные летучие мыши-чайки, охотящиеся на рыбу в море.
Все остальные млекопитающие Маскаренских островов происходят от видов, намеренно
или случайно завезённых человеком. На острове Маврикий нишу насекомоядных млекопитающих
занимают потомки обыкновенного тенрека, причём самый крупный из них, мавригистрикс,
напоминает скорее дикобраза. Немногочисленные хищные млекопитающие представлены
потомками домашних кошек и азиатских мангустов. Кошачьи занимают в экосистемах
верховное положение, а мангусты являются мелкими специализированными хищниками,
например, рыболовами.
Потомки крыс и мышей заселили все Маскаренские острова, за исключением самых
мелких и новообразованных. Они очень разнообразны и занимают различные экологические
ниши. Наиболее необычны древесные потомки крыс, среди которых появилось эндемичное
семейство Maurirattidae. Среди крыс имеются плотоядные формы, а также виды,
способные к планирующему полёту.
Копытные млекопитающие Маскаренских островов немногочисленны и отличаются карликовыми
размерами. Они ведут свой род от домашних свиней и коз.
На Маврикии обитает единственный на Маскаренских островах вид приматов – маврикийский
карликовый макак, потомок макака-крабоеда, отличающийся очень мелкими размерами.
Птицы Маскаренских островов очень разнообразны, число их видов больше, чем
в конце эпохи человека. Наиболее многочисленны морские птицы, гнездящиеся на
побережье главных островов или на мелких островках, свободных от наземных млекопитающих.
В основном это трубконосые, веслоногие, чайки и крачки. Также в море охотятся
необычные хищные птицы из семейства коршунов-серафимов. Основная масса куликов
относится к перелётным видам, останавливающимся на архипелаге лишь на несколько
дней в году, но отдельные виды (в первую очередь зуйки) живут здесь круглый
год.
Пресноводных птиц на Маскаренском архипелаге сравнительно немного. Это несколько
мелких видов цапель и уток, некоторые из которых являются эндемиками. Пастушковые
более разнообразны, однако они, за исключением нескольких видов, сходных с
камышницами и лысухами, очень скрытны. Среди них нет ни одного нелетающего
вида. В прибрежных районах обитают многочисленные чайки и кулики.
Куриные птицы относятся к семейству фазановых и происходят от африканских видов
перепелов, а также от одичавших домашних кур. К куриным относится самая крупная
птица Маскаренского архипелага – эпигаллус, достигающий 150 см в высоту.
Из хищных птиц характерны мелкие соколы африканского происхождения и один вид
луня.
Лесные птицы отличаются высокой степенью эндемизма на видовом и родовом уровне.
Кукушки, стрижи и ракшеобразные представлены несколькими видами. Совы являются
потомками мигрантов из Африки и с Мадагаскара; иглоногие совы в основном насекомоядны,
а более крупные сипухи эндемичного рода охотятся на позвоночных. Голуби и попугаи
ведут свой род от завезённых видов, а также от мигрантов раннего неоцена. Хотя
некоторые виды голубей ведут наземный образ жизни, среди них нет видов, подобных
вымершим дронтам. Мелкие виды древесных голубей отличаются яркой окраской,
для каждого из крупных островов характерны свои эндемичные виды. Среди попугаев
выделяется очень нетипичное семейство попугаев-рыболовов. Они питаются рыбой
и водными беспозвоночными и не встречаются за пределами Маскаренских островов.
«Типичные» виды попугаев также представлены в фауне архипелага, но немногочисленны.
Воробьиные птицы Маскаренских островов весьма разнообразны. Наибольшим количеством
видов представлены семейства ткачиковых, астрильдовых и белоглазок. Один из
видов ткачиков приспособился добывать насекомых из-под коры деревьев и из мягкой
древесины. Также на островах обитают представители таких семейств, как ласточки,
личинкоеды, бюльбюли, славковые, монархи и вьюрковые. Скворцы представлены потомками
завезённой индийской майны, среди которых один крупный вид занимает экологическую
нишу врановых. За исключением белоглазок, певчие птицы Маскаренских островов
происходят от завезённых человеком видов или от мигрантов раннего неоцена.
Маскаренские рептилии принадлежат к отрядам черепах и чешуйчатых. Наземные
черепахи нескольких видов происходят от мадагаскарских и африканских предков.
Среди черепах, обитающих на архипелаге, встречаются настоящие гиганты, такие,
как маврикийская черепаха-латник с длиной панциря до 2 м. Ящерицы относятся
к семействам гекконов и сцинков: их число довольно велико, причём многие виды
происходят от аборигенных. Несколько видов ложноногих змей являются потомками
змей с Мадагаскара; местные змеи вымерли в эпоху человека, не оставив потомков.
На архипелаге практически полностью отсутствуют земноводные: исключение составляет
один вид лягушки с Маврикия, проникший на остров, вероятно, в виде икры на
птичьих лапках.
Настоящие пресноводные рыбы Маскаренских островов относятся к единичным видам
из семейств карповых и лабиринтовых. Это потомки завезённых рыб – золотой рыбки
и настоящего гурами. Помимо этих видов, в реки архипелага заходят морские рыбы.
В условиях тропического климата беспозвоночные достигают значительного разнообразия.
Среди насекомых выделяются бабочки и жуки, в том числе яркие и крупные виды,
характерные для мелких островов. На крупных островах насекомые, как правило,
мельче из-за конкуренции и хищничества со стороны млекопитающих и более многочисленных
птиц. Эндемиками архипелага являются некоторые виды муравьёв. Многочисленны
пауки и скорпионы. Характерно большое количество эндемичных видов наземных
улиток. Пресные водоёмы населяют креветки и крабы морского происхождения, образующие
жилые пресноводные подвиды, а наземные крабы встречаются также в лесах и на
побережье.
Очерк написан Семёном, участником форума
 |
 |
 |
Maurosmilus furiosus |
Melirattula nectarophaga |
Nanosus mauritianus |
 |
 |
|
Aepygallus ambularis |
Psittacohamiota longipes |
| Остров Маврикий и жители леса |
| Главная | Неоцен |