Все натуралисты, насколько я знаю, за одним лишь исключением, принимают, что разные домашние породы кролика произошли от обыкновенного дикого вида; поэтому я опишу здесь домашние породы более тщательно, чем в предыдущих случаях. Профессор Жерве1 говорит, что «настоящий дикий кролик меньше домашнего; пропорции тела не вполне одинаковы, хвост у него меньше, уши короче и гуще одеты волосами; эти признаки, не говоря уже об окраске, составляют доводы против того мнения, которое соединяет обоих животных под одним видовым названием». Немногие лишь натуралисты согласятся с этим автором, что такие мелкие различия достаточны для того, чтобы разделять дикого и домашнего кролика как отдельные виды. Было бы нечто необыкновенное, если бы содержание в тесном помещении, полное приручение, неестественная пища и заботы о разведении, и все это в продолжение многих поколений, не оказали бы хоть некоторого влияния. Кролик был одомашнен с древних времен. Конфуций причисляет кроликов к животным, которые достойны быть приносимы в жертву богам; так как он советует размножать кроликов, то они, вероятно, были уже одомашнены в Китае в это давнее время. О кроликах упоминают некоторые из классических писателей. В 1631 году Джервез Маркгэм писал: «Вы не должны смотреть на форму их, как у других домашних животных, а на богатство шерсти; только выбирайте себе самцов из самых крупных и лучших кроликов, каких только можете достать; что касается достоинства шкурки, то самой лучшей считается та, у которой самым ровным образом перемешаны белые и черные волосы, но белый несколько затенен черным; мех должен быть плотный, длинный, гладкий и блестящий...; телом они гораздо жирнее и крупнее, и если шкурка другого сорта стоит два или три пенса, такие шкурки стоят два шиллинга». Из этого подробного описания мы видим, что серебристо-серые кролики в это время существовали в Англии, и, что гораздо важнее, мы видим также, что к разведению или отбору кроликов тогда относились с большим вниманием. Альдрованди в 1637 году, ссылаясь на различных более давних писателей (например, на Скалигера в 1557 г.), описывает кроликов разного цвета, иные «как заяц», и прибавляет, что П. Валерианус (который умер в 1558 г. в глубокой старости) видел в Вероне кроликов вчетверо крупнее наших2.
1 М. P. Gervais, «Hist. Nat. des Mammiferes», 1854, т. I, стр. 288.
2 U. Aldrovandi, «De Quadrupedis digitatis», 1637, стр. 383. Относительно
Конфуция и Дж. Маркгэма, см. статью одного автора, изучавшего этот предмет,
в «Cottage Gardener», 22 января 1861 г., стр. 250.
92
Исходя из того, что кролики были приручены в древности, мы должны искать
коренную прародительскую форму в северном полушарии Старого Света и
только в более теплых умеренных странах, так как кролик не может жить без
защиты
жилья в таких холодных странах, как Швеция, и, хотя он одичал в тропической
Ямайке, никогда сильно не размножался там. Кролик существует теперь
в теплых умеренных странах Европы; и существует издавна, так как ископаемые
остатки
его были находимы в различных странах3. Домашний кролик легко дичает
в этих же странах, и если выпущены кролики различной окраски, они обыкновенно
возвращаются к обычной серой окраске4. Дикие кролики, если их взять
молодыми,
могут быть одомашнены, хотя это обыкновенно бывает очень затруднительно5.
Разные домашние расы часто скрещивают и считают вполне плодовитыми
при скрещивании, и можно указать ряд совершенно постепенных переходов от
самых
крупных домашних отродий с огромными ушами и до обыкновенного дикого.
Прародительская форма должна была принадлежать к животным, роющим норы,—
привычка, насколько
я мог узнать, не существующая ни у какого другого вида в обширном роде
Lepus. Только один дикий вид достоверно известен для Европы; кролик
с горы Синая (если только это настоящий кролик) и алжирский представляют
мелкие
отличия; эти формы считаются некоторыми авторами за отдельные виды6.
Но
такие мелкие отличия мало помогут нам в объяснении более значительных
различий, характерных для разных домашних рас. Если они происходят от двух
или большего
числа близко родственных видов, то эти, за исключением обыкновенного
кролика, истреблены в диком состоянии; это весьма невероятно по тому упорству,
с
каким кролик противостоит истреблению. По этим причинам мы смело можем
заключить, что все домашние породы происходят от обыкновенного дикого
кролика. Судя по известиям об удивительном успехе, достигнутом во Франции
при выводе
помеси от зайца и кролика7, возможно, хотя и невероятно по великой
трудности, с какою достигается первое скрещивание, что некоторые из более
крупных
рас, окрашенных как заяц, могли быть изменены скрещиванием с этим последним.
Тем не менее, главные различия в скелете разных домашних пород, как
мы вскоре увидим, не могли произойти благодаря скрещиванию с зайцем.
Существует много пород, которые передают свои признаки потомству более
или менее точно. Всякий видел на наших выставках огромных вислоухих
кроликов;
разные близкие к ним подпороды разводятся на континенте, например, так
называемая андалузская; по описаниям, у нее большая голова с округлым
лбом, и она достигает
большего роста, чем все другие; еще одна крупная, парижская порода, с
четыреугольной головой, называется руанской (rouennais); у так называемого
патагонского
кролика замечательно короткие уши и большая, круглая голова. Хотя я всех
этих пород и не видел, я несколько сомневаюсь, чтобы у них было какое-либо
резкое отличие в форме черепа8. Английские вислоухие кролики часто весят
8 или 10 фунтов, а один был выставлен 18 фунтов весом; между тем, совершенно
взрослый дикий кролик весит лишь около 3 1/4 фунтов. У всех больших вислоухих
кроликов, которых я рассматривал, голова, или череп, значительно длиннее
относительно своей ширины, чем у дикого кролика. У многих из них есть
под горлом вися-
3 Owen, «British Fossil Mammals», стр. 212.
4 Bechstein, «Naturgesch. Deutschlands», 1801, т. I, стр. 1133. У меня есть
подобные же сведения относительно Англии и Шотландии.
5 Е. S. Delamer, «Pigeons and Rabbits», 1854, стр. 153. Сэр
Дж. Себрайт (Sir J. Sebright, «Observations on Instinct», 1836, стр.
10) говорит о трудностях
более решительно. Но эта трудность не неизменна: мне прислано два известия
о вполне удачном приручении и разведении диких кроликов. См. также Dr.
P. Broca, «Journal de la Physiologie»,. т. II, стр. 368.
6 Gervais, «Hist. Nat. de Mammiferes», т. I, стр. 292.
7 См. интересную статью Брока, в «Journ. de Phys.» Броун-Секара, т. II, стр.
367.
8 Череп этих пород кратко описан в «Journal of Horticulture», 7 мая 1861
г., стр. 108.
93
чие поперечные складки кожи или подгрудок, который можно оттянуть так,
что он хватает почти до конца челюстей. Уши чудовищно развиты и свешиваются
вниз по обе стороны головы. У одного кролика, выставленного в 1867
году,
длина обоих ушей вместе, от кончика одного до кончика другого, равнялась
22 дюймам, а ширина уха — 5 3/8 дюйма. В 1869 году был выставлен кролик,
у которого уши, измеряя таким же образом, были 23 1/8 дюйма в длину
и 5 1/2 дюймов в ширину, так что этот кролик «превосходил всех других, когда-либо
выставленных на премию». У одного обыкновенного кролика я нашел длину
обоих
ушей, от кончика до кончика, равной 7 5/8 дюйма, а ширину — всего в
1
7/8 дюйма. Вес тела у крупных кроликов и степень развития ушей — это качества,
которые награждаются премиями и подвергались старательному отбору.
Кролик заячьего цвета, или, как его иногда называют, бельгийский кролик,
не отличается от других крупных пород ничем, кроме цвета; но м-р Дж.
Йонг (Young) изСаусгэмптона, у которого большой завод этой породы,
сообщил мне,
что у самок — всех, которых он рассматривал,— только шесть сосков; так
было и у двух самок, которых получил я. М-р Б. П. Брент, впрочем, уверял
меня,
что число сосков у других домашних кроликов изменчиво. У обыкновенного
дикого кролика всегда десять сосков. Ангорский кролик замечателен по
длине и нежности
своего меха, который даже на подошвах лап достигает значительной длины.
Эта порода есть единственная, отличающаяся от других и по умственным
свойствам;
говорят, что она гораздо общительнее других и самец не выказывает желания
убивать молодых9. Из Москвы мне привезли двух кроликов, ростом приблизительно
с дикого, но с длинным, мягким мехом, не таким, как у ангорской породы.
Глаза у этих московских кроликов были красные, сами они снежно-белые,
а уши, два
пятна около носа, верхняя и нижняя поверхность хвоста и зад плюсны черновато-бурые.
Вообще они были окрашены очень похоже на так называемых гималайских кроликов,
которые дальше будут описаны, отличаясь от них только свойством меха.
Есть еще две другие породы, которые характеризуются окраской, но не
различаются
в других отношениях, именно серебристо-серые и шиншилловые. Наконец,
можно упомянуть никардского, или голландского, кролика; он изменчив
по цвету и
замечателен своим малым ростом; некоторые экземпляры весят только 1 1/4
фунта; кролики этой породы дают отличных кормилиц для других, более
нежных отродий10.
Некоторые признаки домашних кроликов замечательно непостоянны или плохо
передаются по наследству. Так, один заводчик говорил мне, что у более
мелких отродий
он едва ли хоть один раз получил весь помет одного цвета. Относительно
крупных вислоухих пород один большой знаток говорит: «невозможно вести
породу одной
окраски, но, разумно пользуясь скрещиванием, многое возможно сделать
в этом направлении. Любитель должен знать своих самок, то-есть окраску
их родителей».
Тем не менее, некоторые цвета, как мы вскоре увидим, передаются точно.
Подгрудок не строго наследуется. Вислоухие кролики, у которых уши висят
вниз, плотно
прилегая по сторонам лица, передают этот признак не вполне устойчиво.
М-р Деламер говорит: «у любительских кроликов, даже если оба родителя
совершенны
по складу, с типичными ушами и красивой окраски, потомство не постоянно
выходит таким же». Если у одного из родителей или даже у обоих «уши веслом»,
то-есть
торчат под прямым углом, и также если один родитель или оба полувислоухи,
то-есть с одним висячим ухом, то вероятность, что у потомства оба уха
будут вполне висячие, почти столь же велика, как в случае, если оба родителя
имеют
этот признак11. Но я имею сведения, что если у обоих родителей
уши вертикальные, то едва ли есть вероятность получить вполне вислоухое
потомство.
9 «Journ. of Horticulture», 1861, стр. 380.
10 «Journal of Horticulture», 28 мая 1861 г., стр. 169.
11 «Journal of Horticulture», 1861, стр. 327. Относительно ушей
см. Delamer, «Pigeons. and Rabbits», 1854, стр. 141; также «Poultry Chronicle»,
т. II,
стр. 499, и 1854, стр. 586.
94
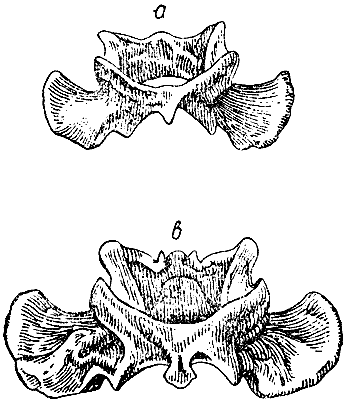
У некоторых полувислоухих кроликов висячее ухо шире и длиннее, чем стоячее12, так что здесь мы имеем необычный случай несимметричности обеих сторон. Это различие в положении и величине ушей, вероятно, указывает на то, что вислоухость получается благодаря большей длине и тяжести уха, и этому, без сомнения, способствует слабость мышц благодаря неупотреблению. Андерсон13 упоминает о породе с одним ухом, а профессор Жерве — о другой породе, вовсе без ушей.
Рис. 5. Полувислоухий кролик (из книги Э. С. Деламера). |

Переходим теперь к гималайской породе, которая иногда также называется китайской, польской или русской. Эти красивые кролики белого или изредка желтоватого цвета, кроме ушей, носа, лап и верхней стороны хвоста; эти все части буровато-черные; так как у этих кроликов красные глаза, то они могут считаться за альбиносов. Я имею различные сведения о постоянстве этой породы. Из-за симметричных отметин эти кролики сначала были признаны за отдельный вид и провизорно названы L. nigripes14. Некоторые хорошие наблюдатели думали, что могут найти отличие и в привычках, и решительно утверждали, что эти кролики образуют новый вид. Происхождение этой породы настолько интересно как само по себе, так и потому, что оно проливает некоторый свет на сложные законы наследования, что о нем стоит рассказать подробно. Но прежде этого необходимо описать кратко еще две породы. Серебристо-серые или серебристо-крапчатые кролики обыкновенно с черною головой и ногами, а прекрасный серый мех их усеян многочисленными черными и белыми длинными волосками. Они образуют совершенно постоянную породу, и их давно уже содержали на огороженных местах. Если они уйдут и спарятся с обыкновенными кроликами, то получается, как мне сообщил м-р Уирлей Берч, из Bretham Hall, не смесь двух цветов, а около половины выходят в одного из родителей, другая половина — в другого. Во-вторых, шиншилловые или ручные серебристо-серые (я буду употреблять первое название), с коротким, более бледным мехом мышастого или аспидного цвета, усеянным длинными волосками, черноватыми, аспидными и белыми15. Порода этих кроликов также вполне постоянна. В 1857 году один автор сообщил16, что он получил гималайских кроликов таким образом. У него была шиншилловая порода, и ее скрестили с обыкновенным черным кроликом; детеныши были черные или шиншилловые. Эти последние были снова скрещены с другими шиншилловыми (которые, в свою очередь, были скрещены с серебристо-серыми), и от этого сложного скрещивания получились гималайские кролики. Это сообщение и другие подобные побудили м-ра Бартлетта17 тщательно поставить опыты в лондонском зоологическом саду; оказа-
12 Delamer, «Pigeons and Rabbits», стр. 136; см. также «Journal
of Horticulture», 1861, стр. 375.
13 «An Account of the different Kinds of Sheep in the Russian
Dominions», 1794, стр. 39.
14 «Proc. Zool. Soc.», 23 июня 1857, стр. 159.
15 «Journal of Horticulture», 9 anp. 1861 г., стр. 35.
16 «Cottage Gardener», 1857, стр. 141.
17 Bartlett, «Proc. Zool. Soc.», 1861, стр. 40.
95
лось, что и простым скрещиванием серебристо-серых кроликов с шиншилловыми
всегда можно было получить немного гималайских; последние, несмотря
на внезапность своего появления, совершенно постоянны при размножении, если
их держать отдельно. Но недавно меня уверяли, что и чистокровные серебристо-серые
кролики любой подпороды случайно производят гималайских.
Гималайские кролики, только что родившиеся, сплошь белого цвета и,
следовательно, настоящие альбиносы, но в течение нескольких месяцев
они постепенно приобретают
темную окраску ушей, носа, лап и хвоста. Иногда, впрочем, как сообщили
мне м-р В. А. Уулер и преп. У. Д. Фокс, молодые родятся очень бледного,
серого
цвета; первый из названных джентльменов прислал мне образцы такого меха.
Однако серый тон исчезает по мере того, как животное приближается к зрелости.
Таким образом, у этих гималайских кроликов есть наклонность возвращаться
к окраске взрослой серебристо-серой прародительской формы, строго приуроченная
к ранней молодости. С другой стороны серебристо-серые и шиншилловые кролики
представляют замечательный контраст с гималайскими по окраске, пока они
совсем молоды; они родятся совершенно черными, но вскоре принимают
характерный для
них серый или серебристый тон. То же самое происходит и с серыми лошадьми;
будучи еще жеребятами, они обыкновенно почти черного цвета, но быстро
становятся серыми и все более и более белеют по мере того, как становятся
старше. Для
гималайских кроликов, таким образом, общее правило, что они родятся белыми,
и потом на некоторых частях тела является темная окраска, а серебристо-серые
родятся черными и потом становятся как бы с белыми блестками. Впрочем,
в обоих случаях иногда являются исключения, притом совершенно противоположного
свойства. Именно, как я слышал от м-ра У. Бёрча, в загонах иногда родятся
у серебристо-серой породы молодые кролики палевого цвета, и такие молодые
в конце концов становятся черными. С другой стороны, гималайские кролики,
как утверждает один опытный любитель, иногда производят на целый помет
одного
черного детеныша, и он, прежде чем пройдет два месяца, становится совершенно
белым18.
Резюмируем весь этот интересный вопрос. Диких* серебристо-серых кроликов
можно рассматривать, как черных кроликов, которые седеют в раннем возрасте.
При скрещивании их с обыкновенными кроликами потомство, как говорят,
не представляет смешения родительских окрасок, но принимает масть того
или другого из родителей
и в этом отношении они похожи на черные и альбиносовые разновидности
большинства четвероногих, которые часто передают потомству свою окраску
таким же образом.
При скрещивании с шиншилловыми кроликами, то-естъ с бледной подразновидностью,
молодые — сначала полные альбиносы, но скоро получают темную окраску
известных частей тела и тогда называются гималайскими. Молодые гималайские
кролики,
впрочем, иногда бывают сначала бледносерыми или сплошь черными и в обоих
случаях через несколько времени белеют. В одной из следующих глав я приведу
большое количество фактов, показывающих, что при скрещивании двух разновидностей,
отличающихся по окраске от их родоначальника, у молодых существует сильное
стремление возвратиться к первоначальной окраске; что очень замечательно,
этот возврат иногда происходит не до рождения, а во время роста животного.
Отсюда, если бы можно было доказать, что серебристо-серые и шиншилловые
кролики произошли от скрещивания черной и альбиносовой разновидности,
у которых окраски
родителей тесно смешаны,— предположение само по себе отнюдь не невероятное,
и в пользу его говорит то, что серебристо-серые кролики, содержимые в
загонах, иногда производят палево-
18 «Phenomenon in Himalayan Rabbits»; «Journ. of Horticulture»,
27 января 1865, стр. 102.
* То-есть коренную разновидность, содержимую в загонах, в отличие от «ручных»
серебристо-серых или шиншилловых. Прим. перев.
96
белых молодых, которые в конце концов чернеют,— тогда все вышеприведенные
парадоксальные факты изменения окраски у серебристо-серых кроликов
и их потомков, гималайских, подошли бы под закон возврата или к первоначальной
черной или к первоначальной альбиносовой прародительской разновидности,
причем возврат происходит в различные периоды роста и в разной степени.
Замечательно также, что гималайские кролики, несмотря на внезапное
свое появление, составляют прочную породу. Но так как в молодости они
являются альбиносами,
то этот случай подходит под очень распространенное правило; альбинизм,
как известно, хорошо наследуется, чему примером могут быть белые мыши
и многие
другие четвероногие, и даже белые цветы. Но почему спрашивается, к черной
окраске возвращаются уши, хвост, нос и лапы, а не другие части тела?
Это, очевидно, зависит от одного закона, применимого в большинстве
случаев, именно,
что признаки, свойственные многим видам данного рода (под чем подразумевается
долговременное наследование от древнего прародителя рода), более упорно
противостоят изменениям или, будучи утрачены, легче появляются снова,
чем признаки, характеризующие
отдельные виды. В роде Lepus у значительного большинства видов
уши и верхняя сторона хвоста окрашены черным; постоянство этих отметин
лучше
всего видно
у тех видов, которые на зиму белеют; так, в Шотландии L. variabilis в
зимнем наряде имеет цветной оттенок на носу и кончики ушей черные;
у L. tibetanus уши черные, верхняя сторона хвоста
серовато-черная, а подошвы ног бурые; у L. glacialis зимний
мех чисто белый, кроме подошв и кончиков ушей. Даже у разноцветных
любительских кроликов мы часто можем наблюдать
склонность
этих же частей к оттенку более темному, чем окраска остального меха.
Таким образом, становятся понятными различные отметины у гималайских
кроликов,
появляющиеся с возрастом. Я могу привести еще один близко сходный случай;
у любительских кроликов очень часто бывают белые звезды на лбу, и у обыкновенного
английского зайца, как я сам наблюдал, в молодости обыкновенно есть на
лбу такая же белая звезда19.
Когда кроликов различной окраски выпускают на свободу в Европе и они
таким образом попадают в естественные условия, они обыкновенно возвращаются
к первоначальной
серой окраске; это может отчасти зависеть от склонности всех скрещенных
животных возвращаться, как сейчас было сказано, к первобытному состоянию.
Но эта склонность
не всегда преобладает; так, серебристо-серые кролики содержатся в загонах
и сохраняют признаки породы, хотя живут почти в диком состоянии; только
не следует населять загон и серебристо-серыми и обыкновенными кроликами
вместе,
иначе «через немного лет останутся только обыкновенные серые»20.
Когда кролики дичают в чужих странах при новых условиях существования,
они
отнюдь не всегда
возвращаются к своей первоначальной окраске. В Ямайке одичалые кролики,
судя по описаниям, были «аспидного цвета, с значительной примесью белых
волосков
на шее, плечах и спине; окраска эта постепенно переходит в голубовато-белую
окраску нижней стороны груди и брюха»21. Но на этом тропическом
острове условия были неблагоприятны для размножения кроликов; они не
расселялись
широко и
теперь, как сообщил мне м-р Р. Хилл, исчезли вследствие большого лесного
пожара. Уже много лет одичалые кролики живут на Фаль-
19 G. R. Waterhouse, «Natural History of Mammalia: Rodents»,
1846, стр. 52, 60, 105.
20 Delamer, «Pigeons and Rabbits», стр. 114.
21 Gosse, «Sojourn in Jamaica», 1855, стр. 441, со слов превосходного
наблюдателя, м-ра Р. Хилла. Это единственный известный случай, что кролики
одичали в жаркой
«траке. Впрочем, их можно держать в Лоанда (см. Livingstone, «Travels»,
стр. 407). В некоторых частях Индии, как сообщил мне м-р Бляйсз, кролики
плодятся
хорошо
97
кландских островах; в некоторых местах они многочисленны, но не расселяются
широко. Большинство их обыкновенного серого цвета; немногие, как сообщил
мне адмирал Сюливэн, окрашены как заяц, и много кроликов черных, часто
с почти симметричными белыми отметинами на морде. Поэтому Лессон описал
черную разновидность, как отдельный вид, под именем Lepus magelanicus,
но это, как я показал в другом месте, неверно22. В недавнее
время промышленники за тюленями населили кроликами некоторые из мелких,
отдельно лежащих
островков; на Пеббль-Айлет, как я слышал от адмирала Сюливэна, значительная
часть
окрашены как зайцы, а на Рэббит-Айлет значительная часть кроликов голубоватого
цвета, который не был встречен в других местах. Какого цвета были кролики,
выпущенные на эти островки, неизвестно.
Кролики, одичавшие на острове Порто-Санто, близ Мадейры, заслуживают
более полного описания. В 1418 или 1419 году у X. Гонзалеса Зарко23 на
корабле случайно была беременная крольчиха, которая родила детенышей
во время
путешествия; все они были выпущены на остров. Кролики вскоре столь
размножились, что стали
вредными и, действительно, заставили бросить поселение на острове. Тридцать
семь лет спустя Када Мосто писал, что кролики стали неисчислимы; это
и неудивительно, так как на острове не было никаких хищных зверков
и вообще никаких наземных
млекопитающих. Мы не знаем признаков прародительницы этих кроликов; вероятно,
это была обыкновенная домашняя порода. Испанский полуостров, откуда отплыл
Зарко, с самых давних исторических времен, как известно, изобиловал обыкновенным
диким видом кроликов; так как кролики были взяты на корабль в качестве
провианта, то невероятно, чтобы они были какой-либо особенной породы.
Что эта порода
была вполне одомашнена, об этом свидетельствует то, что крольчиха родила
во время пути. М-р Уолластон, по моей просьбе, привез с собой два экземпляра
этих одичалых кроликов в спирту; потом м-р Гейвуд прислал мне еще три
экземпляра в рассоле и два живьем. Эти семь экземпляров, хотя и были
пойманы в разное
время, были близко сходны между собою. Они были вполне взрослые, как
это видно по состоянию костей. Хотя условия жизни на Порто-Санто, очевидно,
в
высокой степени благоприятны для кроликов, как это видно по необычайно
быстрому распространению их, все же они сильно отличаются от диких
английских кроликов
по малому росту. У четырех английских кроликов длина, от резцов до заднего
прохода, колебалась между 17 и 17 3/4 дюйма, а длина двух кроликов из
Порто-Санто была только 14 1/2 и 15 дюймов. Вес лучше всего показывает
уменьшение роста;
четыре диких английских кролика весили в среднем по 3 фунта 5 унций,
а один из кроликов с Порто-Санто, живший четыре года в лондонском зоологическом
саду и похудевший, весил только 1 фунт 9 унций. Еще лучший показатель
дается
сравнением хорошо очищенных костей конечностей, взятых от кролика, убитого
на Порто-Санто, с соответствующими костями дикого английского кролика
средней величины; разница между ними оказалась в отношении несколько
меньше пяти
к девяти. Таким образом, кролики на Порто-Санто убавились почти на три
дюйма в длину и почти вдвое в весе тела24. Длина головы
не убавилась в той же пропорции,
как длина туловища,
22 Darwin, «Journal of Researches», стр. 193; «Zoology of the
Voyage of the Beagle, Mammalia», стр. 92.
23 Kerr, «Collection of Voyages», т. II, стр. 177; относительно
Када Мосто — стр. 205. В книге «Historia Insulana», написанной одним иезуитом
и напечатанной
в Лиссабоне в 1717 году, говорится, что кролики были выпущены в 1420 году.
По мнению некоторых авторов, остров был открыт в 1413 году.
24 Нечто в таком роде имело место на острове Липари, где, по
словам Спаланцани («Voyage dans les deux Sicilies», цитировано у Годрона
«De l'Espece», стр.
364), один его соотечественник выпустил несколько кроликов; размножились
они удивительно, но, говорит Спаланцани, «кролики на острове Липари мельче,
чем те, которых разводят в домашнем состоянии».
98
а емкость черепа, как мы позднее увидим, удивительно изменчива. Я отпрепарировал
четыре черепа; сходство между ними было больше, чем это обыкновенно наблюдается
между черепами английских кроликов, но единственная особенность в строении,
которую они представляли, состояла в том, что надглазничные отростки
лобной кости были уже.
По окраске кролик с Порто-Санто значительно отличается от обыкновенного.
Верхняя сторона рыжее и по ней редко рассеяны волоски черные или с черными
кончиками. Горло и некоторые части нижней стороны обыкновенно бледносерые
или свинцового цвета вместо чисто белого. Самую замечательную разницу
представляют уши и хвост; я пересмотрел много свежеубитых английских
кроликов и большую
коллекцию шкурок из разных стран в Британском музее, и у всех их верхняя
поверхность хвоста и кончики ушей одеты черновато-серым мехом; в большинстве
руководств это приводится как один из видовых признаков кролика. У семи
кроликов с Порто-Санто верхняя поверхность хвоста была рыжевато-бурая,
а кончики ушей
не имели и следа черной каемки. Но тут мы встречаем замечательное обстоятельство;
в июне 1861 года я осмотрел двух из этих кроликов, недавно присланных
в лондонский зоологический сад, и у них хвост и уши были окрашены, как
сейчас было описано;
но когда один из них был прислан мне мертвым, в феврале 1865 года, то
на ушах у него была заметная каемка, верхняя сторона хвоста была покрыта
черновато-серым
мехом, и все туловище гораздо менее яркого рыжего цвета; таким образом,
в английском климате этот экземпляр кролика вернулся к характерной окраске
меха несколько скорее, чем в четыре года!
Эти два маленьких кролика из Порто-Санто, когда их держали в зоологическом
саду, имели совершенно иной вид, чем обыкновенный сорт. Они были необыкновенно
дики и проворны, и многие, видя их, говорили, что они больше похожи на
крупных крыс, чем на кроликов. По своим привычкам это были ночные животные
в степени,
необычной для кроликов, и дикость их так и не уменьшилась нисколько;
заведующий садом м-р Бартлетт уверял меня, что более диких животных у
него и не бывало.
Это очень странно ввиду того, что кролики эти произошли от домашней породы.
Я настолько был удивлен этим, что просил м-ра Гейвуда разузнать на месте,
не охотятся ли очень много за этими кроликами местные жители или не преследуют
ли их ястреба, кошки или другие животные; но этого не оказалось, и мы
не можем угадать причины этой дикости. Кролики эти живут и в центральной,
более
высокой и скалистой, местности острова и около береговых утесов и по
своей осторожности и робости редко появляются в более низменных культурных
местах.
Как говорят, они производят от четырех до шести детенышей зараз, и время
размножения приходится на июль и август. Наконец, и это в высшей степени
замечательно, м-ру Бартлетту ни разу не удалось спарить этих двух кроликов
— оба они были самцы — с самками разных пород, которых несколько раз
сажали с ними.
Если бы история этих кроликов на Порто-Санто не была известна, большинство
натуралистов возвели бы их в отдельный вид, принимая в соображение очень
малый рост, окраску рыжеватую сверху и серую снизу и отсутствие черного
на хвосте и ушах. Этот взгляд получил бы значительное подтверждение наблюдением
над живыми кроликами в зоологическом саду и сведениями, что эти кролики
не
желают спариваться с другими. А между тем, этот кролик, которого, несомненно,
возвели бы, таким образом, в особый вид, возник после 1420 года. Итак,
из этих трех случаев одичания кроликов на Порто-Санто, Ямайке и Фалькландских
островах мы видим, что при новых условиях существования у этих животных
не
наблюдается ни возврата к первоначальным признакам, ни сохранения их,
что должно бы иметь место по обычному утверждению большинства авторов.
99
СКЕЛЕТНЫЕ ПРИЗНАКИ
(здесь и далее по тексту
при оцифровке книги красным цветом добавлены примечания,
указывающие
величину рисунков в оригинальном издании. «Ширина
рисунка»
в данном случае означает расстояние между крайними
точками видимого изображения -
В. П.)
Если припомнить, с одной стороны, что важные части организации, как часто утверждают, никогда не варьируют, а с другой стороны, на каких мелких отличиях скелета зачастую основаны ископаемые виды, то изменчивость черепа и некоторых других костей у домашнего кролика весьма заслуживает внимания. Не следует предполагать, что более важные отличия, которые сейчас будут описаны, строго характеризуют какую-либо породу; все что можно сказать, это то, что они обыкновенно наблюдаются у известных пород. Мы должны помнить, что отбор не был прилагаем к упрочению какого-либо признака скелета
 |
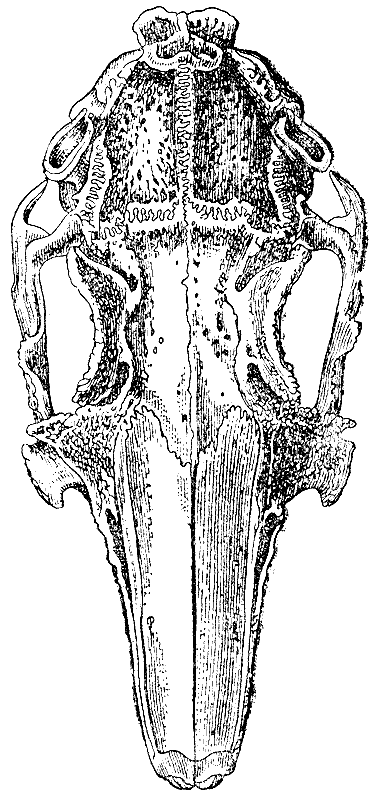 |
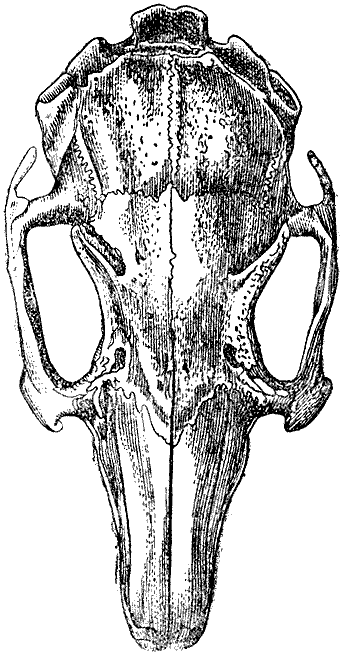
Рис. 6. Череп дикого кролика в естественную величину. (высота оригинального рисунка 80 мм) |
Рис. 7. Череп крупного вислоухого кролика в естественную величину. (высота оригинального рисунка 98 мм) |
и что этим животным не приходилось поддерживать себя своими силами при однородных условиях существования. Большинство из отличий скелета мы не можем объяснить, но мы увидим, что увеличение размеров тела вследствие тщательного кормления и постоянного подбора особым образом повлияло на голову. Даже удлинение и свисание ушей в некоторой степени повлияло на форму всего черепа. Отсутствие упражнения, очевидно, изменило относительную длину конечностей, по сравнению с туловищем.
В качестве мерки для сравнения я отпрепаровал скелеты двух диких кроликов
из Кента, одного с Шетландских островов и одного из Антрима, в Ирландии.
Так как у этих четырех экземпляров из столь удаленных друг от друга мест
все кости были близко сходны, без какой-либо уловимой разницы, то можно
заключить,
что кости дикого кролика вообще одинаковы по признакам.
Череп. Я тщательно исследовал черепа десяти крупных вислоухих кроликов
и пяти обыкновенных домашних; эти отличались от вислоухих только тем,
что уши
и туловище были не так крупны, но крупнее, чем у дикого кролика. Сначала
о десяти вислоухих кроликах. У всех них череп очень удлинен, в сравнении
с шириною. У одного дикого кролика длина черепа была 3,15 дюйма, а у
крупного любительского кролика 4,3, ширина же мозговой капсулы черепа
была у обоих
почти совершенно одинакова. Даже если взять мерой для сравнения наибольшую
ширину в области скуловых дуг, черепа вислоухой
100
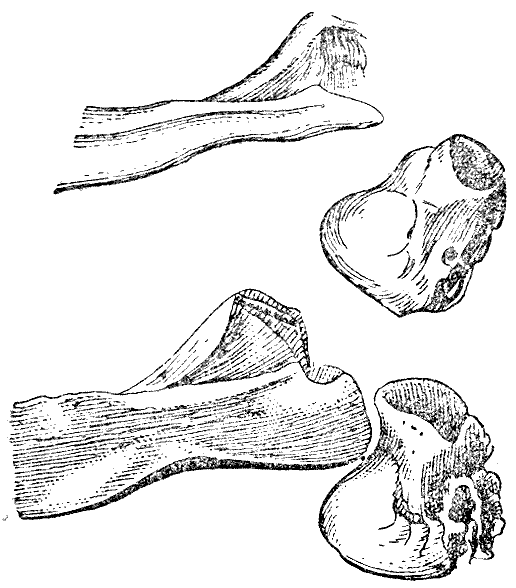
Рис. 8. Часть скуловой дуги с выдающимся концом скуловой кости и слуховой проход в естественную величину. Верхний рисунок — дикий кролик. Нижний рисунок — вислоухий кролик заячьего цвета. (ширина оригинального рисунка 42 мм) |
породы на три четверти дюйма длиннее, по сравнению с шириной. Высота головы
увеличилась почти в той же пропорции, как и длина, так что лишь ширина
не увеличилась. Теменные и затылочные кости, окружающие мозг, и в продольном
и в поперечном направлении менее выпуклы, чем у дикого кролика, так
что форма крыши черепа несколько отличается. Поверхность более шероховатая,
с менее тонким рельефом, и линии швов сильнее выдаются.
Хотя череп крупных вислоухих кроликов, сравнительно с черепом диких
кроликов, сильно удлинен относительно своей ширины, он отнюдь не удлинен
по сравнению
с величиной туловища. Вислоухие кролики, которых я исследовал, были слишком
вдвое тяжелев диких, хотя и не были жирны, но череп их далеко не был
вдвое длиннее. Даже если мы возьмем более подходящую меру сравнения,
длину тела
от носа до заднего прохода, то череп в среднем оказывается на треть дюйма
короче, чем можно было ожидать. У мелкого одичалого кролика с Порто-Санто,
напротив, голова по отношению к длине тела оказывается длиннее, чем можно
было ожидать, приблизительно на четверть дюйма.
Это удлинение черепа, по сравнению о его шириною, оказывается, по моим
наблюдениям, общим признаком не только для крупных вислоухих кроликов,
но и для всех искусственных
пород; оно хорошо видно и на черепе ангорской породы. Сначала я был
очень удивлен этим явлением и не мог представить себе, каким образом одомашнение
могло дать этот однородный результат; объяснение, повидимому, заключается
в том,
что искусственные расы в течение многих поколений жили в тесном заключении
и имели мало поводов упражнять свои чувства, умственные способности
и
произвольные мышцы; вследствие этого мозг, как мы вскоре узнаем более
подробно, не увеличился
пропорционально с ростом тела. Так как мозг не увеличился, не увеличилась
и вмещающая его костная капсула, а это, очевидно, в силу соотношения
повлияло на ширину всего черепа от одного конца до другого.
У всех черепов крупных вислоухих кроликов надглазничные пластинки или
отростки лобных костей гораздо шире, чем у дикого кролика, и обыкновенно
сильнее
выда-
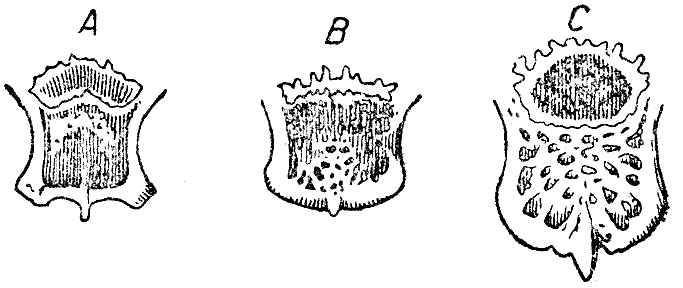
Рис. 9. Задний конец черепа в естественную величину с межтеменной костью. А — дикий кролик, В — одичалый кролик с Порто-Санто, близ Мадейры С — крупный вислоухий кролик. (ширина оригинального рисунка 55 мм) |
Рис. 10. Затылочное отверстие в естественную величину. (ширина оригинального рисунка 55 мм) |

ются вверх. В скуловой дуге задний, выдающийся конец скуловой кости шире
и тупее, и у экземпляра на рисунке 8 это выражено в замечательной степени.
Конец этот подходит ближе к слуховому проходу, чем у дикого кролика, как
это лучше всего видно на рисунке 8, но это обстоятельство зависит преимущественно
от перемены в направлении слухового прохода. Межтеменная кость (рис. 9) сильно
разнится по форме в различных черепах; обыкновенно она более овальна, то-есть
более вытянута в направлении продольной оси черепа, чем у дикого кролика.
Задний край «четырехугольной возвышенной площадки»25 затылка у дикого кролика
как бы обрублен или с маленьким выступом, а у большинства вислоухих выдается
острием, как на рисунке 9, С. Боковые отростки затылочной кости (рядом с
сосцевидным отростком височной*) пропорционально величине черепа обыкновенно
гораздо толще, чем у дикого кролика.
Затылочное отверстие (рис. 10) представляет некоторые замечательные отличия.
У дикого кролика нижний край между затылочными мыщелками изогнут (значительно,
25 Waterhouse, «Nat. Hist., Mammalia», т. II, стр. 36.
* Processus paramastoidei.— Прим. перев.
101
Рис. 11. Череп полувислоухого кролика в естественную величину. Видно различие в направлении слухового прохода правой и левой стороны и причиняемое этим общее нарушение симметрии черепа. Свешивалось вперед левое ухо (на правой стороне рисунка). (высота оригинального рисунка 97 мм) |
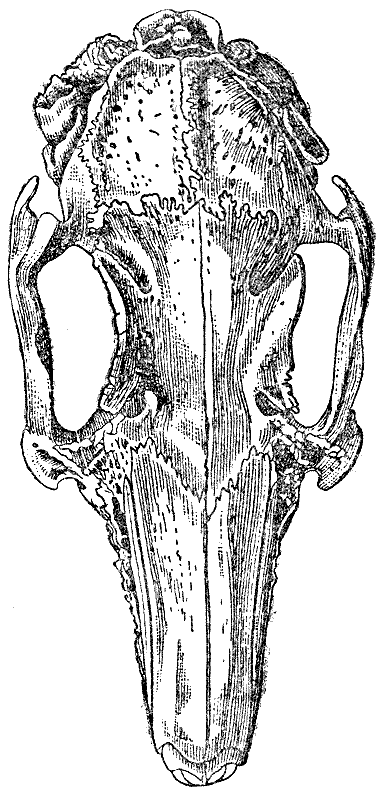
почти углом, а верхний край с глубокой четырехугольной вырезкой,
так что продольная ось длиннее поперечной. В черепе вислоухого кролика
поперечная
ось больше продольной; ни в одном из этих черепов нижний край отверстия
между мыщелками не был так сильно изогнут: в пяти черепах не было
четырехугольной вырезки верхнего края, в трех был след этой вырезки, и
только в двух
она была хорошо развита. Эти различия в форме затылочного отверстия замечательны,
ввиду того что чрез него проходит такой важный орган, как спинной
мозг, хотя, невидимому, на форме последнего не сказывается форма затылочного
отверстия.
Во всех черепах больших вислоухих кроликов костный слуховой проход
значительно шире, чем у дикого кролика. В одном черепе в 4,3 дюйма
длиною, который был
едва шире черепа дикого кролика (длина этого была 3,15 дюйма), больший
диаметр отверстия слухового прохода был ровно вдвое больше, чем у дикого
кролика.
Отверстие более сжато, и край его на стороне, ближайшей к черепу, более
высок, чем наружный край. Весь проход направлен более вперед. Так как
при разведении
вислоухих кроликов длина ушей и в связи с этим, свисание и прилегание
вплотную к лицу составляют главные черты достоинства, то едва ли может
быть сомнение
в том, что значительная перемена в величине, форме и направлении костного
слухового прохода, в сравнении с тем же органом дикого кролика, зависит
от постоянного отбора особей все с более и более крупными ушами. Влияние
наружного
уха на костный слуховой проход хорошо видно на черепе (я их исследовал
три) полувислоухих кроликов (рис. 8), у которых одно ухо стоит, а другое,
более
длинное, сисит: в этих черепах есть явная разница в форме и направлении
слухового прохода той я другой стороны. Еще более интересно, что изменение
в положении
и увеличение размеров костного слухового прохода несколько отразилось
на строении соответствующей стороны всего черепа. Я привожу рисунок
(рис. 11)
черепа полувислоухого кролика; на нем можно видеть, что шов между лобной
и теменной частями не идет в точности под прямым углом к продольной оси
черепа; левая лобная кость выдается вперед дальше правой; и передний
и задний концы
левой скуловой дуги на стороне висячего уха лежат немного впереди соответствующих
пунктов другой стороны. Даже нижняя челюсть подверглась изменению, и
сочленовные отростки ее не вполне симметричны: отросток левой стороны
лежит немного впереди
правого. Это, по моему мнению, замечательный случай соотношения в развитии.
Кто предположил бы, что, содержа животное в течение многих поколений
в неволе, что ведет к неупотреблению мускулов уха, и постоянно отбирая
особи с
самыми длинными и широкими ушами, возможно таким путем косвенно повлиять
почти на
каждый шов черепа и на форму нижней челюсти!
У больших вислоухих кроликов единственное отличие нижней челюсти, по
сравнению с диким кроликом, состоит в том, что задний край восходящей
ветви шире и
сильнее загнут внутрь. Зубы обеих челюстей не представляют никакой разницы,
за тем исключением, что малые резцы, сидящие за большими, пропорционально
немного длиннее. Коренные зубы увеличились в размерах пропорционально
увеличению ширины черепа, как она определяется измерением в области скуловых
дуг, а
не пропорционально возрастанию длины черепа. Внутренний край лунок коренных
зубов верхней челюсти у дикого кролика образует совершенно прямую линию:
на некоторых из самых крупных черепов вислоухих кроликов эта линия была
заметно вогнута внутрь. У одного экземпляра было по добавочному коренному
зубу на
обеих сторонах верхней челюсти, между ложнокоренными и коренными; однако
эти два зуба были неравной величины, и так как ни у одного грызуна нет
семи коренных, то это есть лишь уродство, хотя и интересное.
Пять других черепов обыкновенных домашних кроликов, из которых одни по
величине приближаются к вышеописанным наиболее крупным черепам, а другие
лишь незначительно
превосходят черепа диких кроликов, заслуживают внимания лишь в том отношении,
что во всех перечисленных различиях представляют полный ряд переходов
между черепами самых крупных вислоухих кроликов и диких. У всех, однако,
надглазничные
102
пластинки несколько шире, и у всех, сообразно увеличенным размерам наружного
уха, слуховой проход шире, чем у дикого кролика. Кривизна нижнего края
затылочного отверстия у некоторых была не так крута, как у дикого кролика,
но верхняя вырезка во всех пяти черепах была хорошо развита.
Череп ангорского кролика, как и последние пять черепов, по общим пропорциям
и большинству других признаков занимает середину между черепами самых
крупных вислоухих кроликов и диких. Он представляет лишь один своеобразный
признак:
хотя он значительно длиннее черепа дикого кролика, ширина между правым
и левым задним надглазничным отверстием почти на треть меньше, чем
у дикого.
У серебристо-серого, шиншиллового и гималайского кролика череп более
удлинен, чем у дикого, с более широкими надглазничными пластинками,
но мало отличается
в других отношениях, за исключением того, что вогнутость верхнего и нижнего
края затылочного отверстия не столь глубока или не столь развита. Череп
московского кролика едва ли чем отличается от черепа дикого кролика.
У одичалых кроликов
с Порто-Санто надглазничные пластинки обыкновенно уже и более заострёны,
чем у наших диких кроликов.
 |
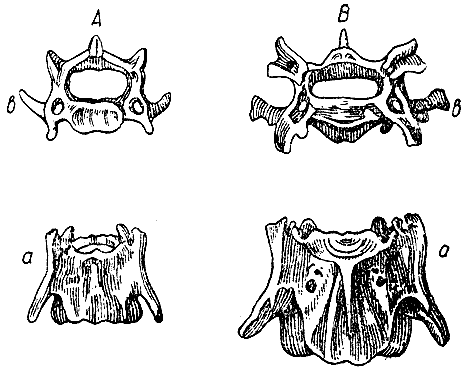 |
Рис. 12. Атлант в естественную величину снизу и спереди. Верхний рисунок — дикий кролик, нижний рисунок — крупный вислоухий кролик заячьего цвета, а — верхний непарный отросток, b — нижний непарный отросток. (ширина оригинального рисунка 43 мм) |
Рис. 13. Третий шейный позвонок в естественную величину. А — дикий кролик, В — крупный вислоухий кролик заячьего цвета; а,а — нижняя поверхность, b,b — передние сочленовные поверхности. (ширина оригинального рисунка 54 мм) |
Так как некоторые из самых крупных вислоухих кроликов, скелеты которых я
приготовил, были окрашены почти как зайцы, а последние, как утверждают, во
Франции недавно были скрещиваемы с кроликами, то можно было бы думать, что
некоторые из описанных выше признаков произошли благодаря скрещиванию в давнее
время с зайцем. Вследствие этого я исследовал черепа зайца, но это не пролило
света на особенности черепа более крупных кроликов. Интересен, впрочем, следующий
факт как иллюстрация закона, по которому разновидности одного вида часто
принимают признаки других видов того же рода; сравнивая в Британском музее
черепа десяти видов зайцев, я нашел, что они отличаются между собою в тех
же чертах, которые подлежат изменению у домашних кроликов, именно в общих
пропорциях, в форме и величине надглазничных пластинок, в форме свободного
конца скуловой кости и в очертаниях шва, разделяющего теменные и лобные кости.
Сверх того, два признака, особенно изменчивые у домашнего кролика, именно
очертание затылочного отверстия и форма «приподнятой площадки» затылка, были
в двух случаях также изменчивы в пределах одного вида зайцев.
Позвонки. Количество их одинаково во всех скелетах, которые я исследовал,
за двумя исключениями: одного из мелких одичалых кроликов с Порто-Санто и
у одного из кроликов самой крупной вислоухой породы; у обоих этих кроликов
было, как обыкновенно, семь шейных позвонков и двенадцать грудных с ребрами,
но у обоих было восемь поясничных позвонков вместо семи. Это замечательно,
так как Жерве приводит семь как характерное число для всего рода Lepus. В
числе хвостовых позвонков, невидимому, бывает разница на два или на три,
но я. не обращал на них внимания, и их трудно сосчитать точно.
В первом шейном позвонке, или атланте, передний край верхней дуги у диких
экземпляров немного изменчив; он или почти гладкий или с маленьким непарным
отростком; я даю рисунок экземпляра (рис. 12) с наиболее развитым отростком
(а), какой я только видел, но следует заметить, насколько он меньше и иной
формы, чем у крупного вислоухого кролика. У этого последнего нижний непарный
отросток (b) пропорционально также гораздо толще и длиннее. Очертания крыльев
несколько более квадратные.
103
Третий шейный позвонок. Если смотреть на этот позвонок снизу, то у дикого кролика (рис. 13, А, а) он имеет поперечный отросток, направленный косо назад и состоящий из одного заостренного стержня; у четвертого позвонка этот отросток слегка раздвоен в середине. У крупных вислоухих кроликов этот отросток (В, а) раздвоен на третьем позвонке, как у дикого кролика на четвертом. Но различие между третьими шейными позвонками у дикого и у вислоухого кролика (А, b и В, b) обнаруживается еще резче при сравнении их передних сочленовных поверхностей, потому что концы передних косых отростков у дикого кролика просто закруглены, а у вислоухого они тройчатые, с глубокой выемкой в середине. Спинномозговой канал у вислоухого кролика (В, b) более вытянут в поперечном направлении, чем у дикого кролика, и отверстия для артерий несколько иной формы. Мне кажется, что указанные различия этого позвонка заслуживают внимания.
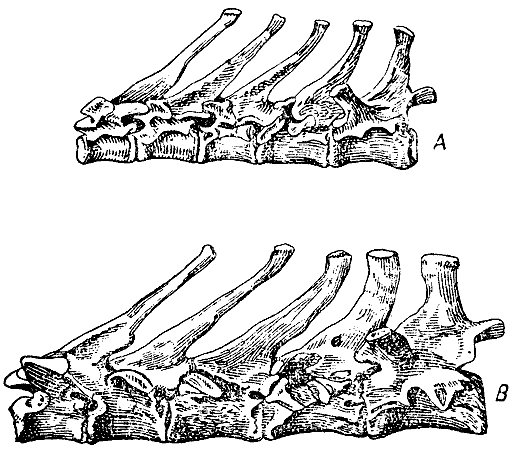 |
Рис. 14. Спинные позвонки с шестого по десятый включительно в естественную величину сбоку. А — дикий кролик, В — крупный заячьего цвета, так называемый испанский. (ширина оригинального рисунка 64 мм) |
Первый спинной позвонок. Длина остистого отростка изменчива у дикого
кролика; иногда он очень короток, но обыкновенно длиннее половины остистого
отростка
второго спинного позвонка; у двух крупных вислоухих кроликов этот отросток
был в три четверти длины остистого отростка второго спинного позвонка.
Девятый и десятый спинные позвонки. У дикого кролика остистый отросток
девятого позвонка едва заметно толще остистого отростка восьмого позвонка;
остистый
отросток десятого позвонка заметно толще и короче, чем остистые отростки
всех предшествующих спинных позвонков. У крупных вислоухих кроликов остистые
отростки десятого, девятого и восьмого позвонков, а в слабой степени
и остистый отросток седьмого позвонка толще и несколько иной формы, чем
у дикого кролика.
Вследствие этого эта часть позвоночника значительно разнится по виду
от соответствующей части дикого кролика и, что интересно, сильно походит
на соответствующие
позвонки некоторых видов зайцев. У ангорского, шиншиллового и гималайского
кролика остистые отростки восьмого и девятого позвонков незначительно
толще, чем у дикого. Напротив, у одного из одичалых кроликов с Порто-Санто,
которые
в большинстве признаков уклоняются от обыкновенного дикого кролика в
направлении, прямо противоположном тому, в котором уклонились крупные
вислоухие кролики,
остистые отростки девятого и десятого позвонков были нисколько не массивнее,
чем на предшествующих позвонках. У этого же экземпляра с Порто-Санто
на девятом позвонке не было и следа передних косых отростков (рис. 14),
которые хорошо
развиты у всех британских диких кроликов и еще сильнее у крупных вислоухих.
У одного полудикого кролика из Сэндонского парка26 на нижней
стороне двенадцатого спинного позвонка был довольно сильно развит нижний
отросток
тела; я этого
больше не видал ни у одного экземпляра.
Поясничные позвонки. Я уже сказал, что в двух случаях было восемь вместо
семи поясничных позвонков. Третий поясничный позвонок в одном скелете
дикого английского кролика и в одном скелете одичалого кролика с Порто-Санто
имел
гемальный отросток тела; между тем, в четырех скелетах большого вислоухого
кролика и у гималайского кролика нижний отросток тела на этом позвонке
был хорошо развит.
Таз. У четырех диких экземпляров форма таза была почти тождественна,
но у различных домашних пород можно было уловить намек на разницу. У
крупных вислоухих
кроликов вся верхняя часть подвздошной кости прямее или менее скошена,
чем у дикого кролика; бугристость на внутренней губе передней и верхней
части
подвздошной кости выдается относительно больше.
Грудина. Задний конец заднего отдела грудины у дикого кролика (рис. 15,
А) тонкий и слабо расширен; у некоторых из крупных вислоухих кроликов
(В) эта
часть значительно
26 Эти кролики довольно давно одичали в Сэндонском парке и в других местах в Стаффордшире и Шропшире. Как говорил мне смотритель, они произошли от выпущенных на волю домашних кроликов разного цвета. Окраска этих кроликов изменчива, но многие окрашены симметрично: белого цвета, с полосой по хребту, ушами и отметинами на голове черновато-серого оттенка. Туловище у них несколько длиннее, чем у обыкновенных кроликов.
104
сильнее расширена к вершине, а у других экземпляров (С) она почти одинаковой
ширины от одного конца до другого, но вначительно толще у вершины.
Лопатка. От акромиона отходит под прямым углом отросток, кончающийся
косым утолщением; у дикого кролика (рис. 16, А) незначительно изменяется
форма
и величина этого утолщения, и также заострение акромиона и ширина части
его тотчас под отростком. Но эти изменения у дикого кролика очень слабы,
тогда
как у крупных вислоухих кроликов они значительны. Так, у одних экземпляров
(B) косое вершинное утолщение отростка акромиона само развивается в
короткий отросток, стоящий под тупым углом к прямоугольному отростку. У других
экземпляров эти два отростка лежат почти по прямой (С). Ширина и заострение
вершины акромиона
сильно изменчивы, как можно видеть, сравнив рисунки В, С и D.
Конечности. В конечностях я не мог найти изменений; впрочем, тщательное
сравнение костей лапы потребовало бы слишком много работы.
 |
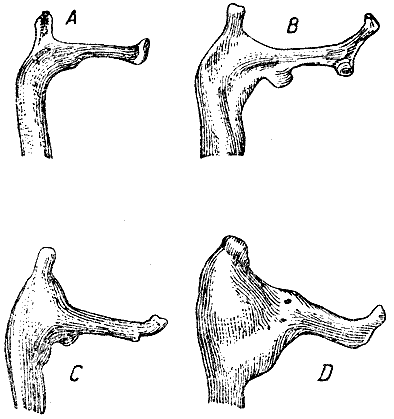 |
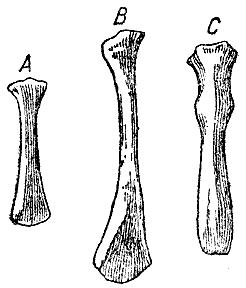
Рис. 15. Мечевидный отросток грудины в естественную величину. А — дикий кролик, В — вислоухий кролик заячьего цвета, С — испанский кролик (у В левый угол сочленовного конца был обломан, и кость так и нарисована). (ширина оригинального рисунка 28 мм) |
Рис. 16. Акромион лопатки в естественную величину. А — дикий кролик, В, С, D — крупные вислоухие кролики (ширина оригинального рисунка 49 мм) |
Я описал все различия в скелете, которые я наблюдал. Нельзя не удивляться высокой степени изменчивости или пластичности многих костей. Мы видим отсюда, насколько ошибочным является часто повторяемое утверждение, что лишь гребни костей, служащие для прикрепления мышц, изменяют свою форму и что лишь маловажные органы изменяются под влиянием одомашнения. Никто не скажет, например, что затылочное отверстие, или атлант, или третий шейный позвонок — маловажные части. Если бы различные позвонки диких и вислоухих кроликов, изображенные здесь, были найдены в ископаемом состоянии, палеонтологи без всякого колебания заявили бы, что эти позвонки принадлежат равным видам.
Действие употребления и неупотребления органов. У крупных вислоухих кроликов
относительная длина костей данной ноги и отношение длины между костями
передних и задних ног остались почти те же, что у дикого кролика; в весе
кости задних
ног, видимо, не прибавились в должном соотношении с передними ногами.
Вес всего тела больших кроликов, которых я исследовал, превышал вес диких
кроликов
от двух до двух с половиной раз; вес костей передних и задних ног вместе
(кроме лап, так как трудно очистить такое множество мелких костей) возрос
у крупных вислоухих кроликов почти в таком же отношении, следовательно,
в должном соответствии с весом тела, который эти кости поддерживают. Если
мы
берем как меру для сравнения длину тела, то конечности больших кроликов
против должного соотношения не доросли в длину на полтора дюйма. Также
если мы примем
ва меру длину черепа, который, как мы раньше видели, не прибавился в
длину в должном соответствии с длиною тела, то конечности окажутся, по пропорциям
дикого кролика, короче на полдюйма или три четверти дюйма. Следовательно,
какую меру сравнения ни взять, кости конечностей у больших вислоухих
кроликов
не увеличились в длину, хоть и увеличились в весе, в полном соответствии
с другими частями скелета; это, я предполагаю, может быть объяснено недеятельной
жизнью в течение многих поколений. Также и лопатка не увеличилась в длину
в должном соответствии с увеличением длины тела.
Более интересный пункт представляет емкость костной капсулы мозга; я
обратил на него внимание, найдя, как уже было указано, что у всех одомашненных
кроликов
105
длина черепа значительно увеличилась относительно ширины в сравнении с
дикими кроликами. Если бы иметь большое количество одомашненных кроликов
почти
такого же роста, как дикие, то измерить и сравнить емкость их черепов
было бы просто. Но этого нет на самом деле: почти все домашние породы крупнее,
чем дикие кролики, и вислоухие сорта тяжелее в два с лишком раза. Так
как
мелкому животному приходится упражнять свои чувства, ум и инстинкты в
одинаковой мере с крупным животным, то мы не имеем основания ожидать, чтобы
у животного,
которое вдвое или втрое больше другого, мозг был двойной или тройной
величины27. Свесив четырех диких кроликов и четырех крупных, но
не откормленных, вислоухих,
я нашел, что в среднем вес диких относится к весу вислоухих, как 1 к
2,17; среднее отношение длины тела есть 1 к 1,41; между тем емкости, черепа
относятся,
как 1 к 1,15. Отсюда мы видим, что емкость черепа и, следовательно, величина
мозга увеличились лишь незначительно, в сравнении с увеличением размеров
тела; это объясняет нам узость черепа по отношению к его длине у всех
домашних кроликов.
В верхней половине приложенной таблицы я дал измерения черепа десяти диких
кроликов, а в нижней половине — одиннадцати вполне одомашненных. Так как
эти кролики столь значительно различаются в росте, то необходимо иметь
какую-либо мерку для сравнения с нею емкости их черепа. Я избрал как лучшую
меру длину
черепа, так как она у крупных кроликов не увеличилась в такой степени,
как длина всего тела; но так как и череп, подобно всякому другому органу,
изменяется
в длине, то ни он, ни какой другой орган не дают совершенной мерки.
В первом столбце цифр дана наибольшая длина черепа в дюймах и десятых долях.
Я знаю, что эти цифры претендуют на большую точность, чем это возможно,
но я считаю самым простым записывать длину, как ее дает циркуль. Во втором
и
третьем столбцах даны длина и вес тела, если эти измерения были сделаны.
Четвертый столбец дает емкость черепа, выраженную весом мелкой дроби, которою
наполняли череп; точность этих взвешиваний в пределах нескольких гран не
гарантируется. В пятом столбце дается емкость, которую должен бы иметь
череп по вычислению, соответственно длине его и по сравнению с черепом
дикого кролика
№ 1; в шестом столбце — разница между наблюденной и вычисленной емкостью,
и в седьмом — прибавка или уменьшение в процентах. Например, так как у
дикого кролика № 5 тело короче и легче, чем у дикого кролика № 1, то мы
могли бы
ожидать, что его череп имеет меньшую емкость; действительная емкость, выраженная
весом дроби, 875 гран, на 97 гран меньше, чем у первого кролика. Но сравнивая
этих двух кроликов по длине черепа, мы находим, что у № 4 длина черепа
3,15 дюйма, а у №5 — 2,96; согласно этому отношению, мозг № 5 должен иметь
объем
913 гран дроби, что превышает наблюдавшийся объем, но только на 38 гран.
Или, рассматривая этот случай иным образом (столбец VII), мозг этого маленького
кролика (№ 5) на каждые 100 гран веса оказывается только на 4 грана легче,
то-есть он должен был бы быть на 4% тяжелее, судя по взятому за образец
кролику № 1. Я взял кролика № 1 за меру для сравнения потому, что из всех
черепов,
достигших полной средней длины, этот имел наименьшую емкость; следовательно,
он наименее благоприятен для результата, который я хочу показать, именно,
что мозг у всех давно одомашненных кроликов, в сравнении с мозгом дикого
кролика, уменьшен в размерах или абсолютно или относительно длины головы
и тела. Если бы я в качестве мерки взял ирландского кролика (№ 3), то приведенные
ниже результаты были бы несколько резче.
Обращаемся теперь к таблице. У первых четырех диких кроликов череп одинаковой
длины, и разница в емкости незначительна. Сэндонский кролик (№ 4) интересен
в том отношении, что, хотя он теперь дикий, произошел он от одомашненной
породы, как это доказывается его своеобразной окраской и длинным туловищем;
однако череп вернулся к нормальной длине и полной емкости. Следующие три
кролика дикие, но мелкого роста, и у всех них череп с несколько уменьшенной
емкостью. Три одичалых кролика из Порто-Санто (№ 8 — 10) представляют затруднительный
случай. Тело у них значительно уменьшилось в размерах, и также уменьшились,
но в меньшей степени, длина и абсолютная емкость черепа, в сравнении с
черепом диких английских кроликов. Но у трех кроликов из Порто-Санто, при
сравнении
их между собою по емкости черепа, мы находим неожиданную разницу, которая
не стоит ни в каком отношении с небольшой разницей между ними в длине черепа,
ни, как я думаю, с какой-либо разницей в величине их тела; я, однако, упустил
из виду отдельно взвесить их. Едва ли можно предположить, что вещество
мозга у этих трех кроликов, живущих при сходных условиях, может разниться
настолько,
как это указывает пропорциональная разница в емкости черепа; и я не знаю,
возможно ли, чтобы один мозг содержал гораздо больше жидкости, чем другой.
Поэтому я никак не могу объяснить этого случая.
Обращаясь к нижней половине таблицы, где даны измерения одомашненных кроликов,
мы видим, что у всех них, хотя в очень различной степени, емкость черепа
меньше, чем можно было бы ожидать по длине черепа, сравнивая с диким кроликом
№ 1. В строке 22 даны средние измерения семи крупных вислоухих кроликов.
Теперь возникает вопрос:
27 См. замечания проф. Оуэна в его статье «Zoological Significance of the Brain, etc. of Man. etc.», доложенной заседанию британской ассоциации в 1862 г.; относительно птиц см. «Proc. Zoolog. Soc.», 11 января 1848 г.. стр. 8.
106
| Название породы. Дикие и полудикие кролики | I Длина черепа |
II Длина тела от резцов до заднего прохода |
III Вес всего тела |
IV Емкость черепа, измеренная мелкой дробью |
V Емкость, вычисленная по длине черепа, по пропорциям № 1 |
VI Разница между действительн. и вычисленной емкостью черепа | VII Показывает, на сколько процентов мозг тяжелее или легче вычисленного по длине черепа и пропорциям № 1 |
|
1. Дикий кролик, Кент |
дюймы 3,15 |
дюймы 17,4 |
фунты 3 |
унции 5 |
граны 972 |
граны — |
граны — |
— |
| 2. ---»--- ---»--- Шотланд. о-ва | 3,15 |
— |
— |
— |
979 |
— |
— |
(На 2% тяжелее сравнительно с № 1) |
| 3. ---»--- ---»--- Ирландия | 3,15 |
— |
— |
— |
992 |
— |
— |
|
| 4. Домашний кролик одичалый Сэндон | 3,15 |
18,5 |
— |
— |
977 |
— |
— |
— |
| 5. Дикий обыкновенн. разновидн. мелкий экземпляр, Кент | 2,96 |
17,0 |
2 |
14 |
875 |
913 |
38 |
4% легче |
| 6. Дикий, красноватая разновидн. Шотландия | 3,1 |
— |
— |
— |
918 |
950 |
32 |
3% » |
| 7. Серебристо-серый, маленький экземпляр, Тетфорд | 2,95 |
15,5 |
2 |
11 |
938 |
910 |
28 |
3% тяжелее |
| 8. Одичалый кролик, Порто-Санто | 2,83 |
— |
— |
— |
893 |
873 |
20 |
2% » |
| 9. ---»--- ---»--- ---»--- | 2,85 |
— |
— |
— |
756 |
879 |
123 |
16% » |
| 10. ---»--- ---»--- ---»--- | 2,95 |
— |
— |
— |
835 |
910 |
75 |
9% » |
| Средние цифры кроликов из Порто-Санто | 2,88 |
— |
— |
— |
828 |
888 |
60 |
7% » |
Домашние кролики |
||||||||
| 11. Гималайский | 3,5 |
20,5 |
— |
— |
963 |
1080 |
117 |
12% » |
| 12. Московский | 3,25 |
17,0 |
3 |
8 |
803 |
1002 |
199 |
24% » |
| 13. Ангорский | 3,5 |
19,5 |
3 |
1 |
697 |
1080 |
383 |
54% » |
| 14. Шиншилловый | 3,65 |
22,0 |
— |
— |
995 |
1126 |
131 |
13% » |
| 15. Крупный вислоухий | 4,1 |
24,5 |
7 |
0 |
1065 |
1265 |
200 |
18% » |
| 16. ---»--- ---»--- | 4,1 |
25,0 |
7 |
13 |
1153 |
1265 |
112 |
9% » |
| 17. ---»--- ---»--- | 4,07 |
— |
— |
— |
1037 |
1255 |
218 |
21% » |
| 18. ---»--- ---»--- | 4,1 |
25,0 |
7 |
4 |
1208 |
1265 |
57 |
4% » |
| 19. ---»--- ---»--- | 4,3 |
— |
— |
— |
1232 |
1326 |
94 |
7% » |
| 20. ---»--- ---»--- | 4,25 |
— |
— |
— |
1124 |
1311 |
187 |
16% » |
| 21. Крупный, заячьего цвета | 3,86 |
24,0 |
6 |
14 |
1131 |
1191 |
60 |
5% » |
| 22. Средние цифры для семи крупных вислоух. кроликов | 4,11 |
24,62 |
7 |
4 |
1136 |
1268 |
132 |
11% » |
| 23. Заяц (L. timidus) английский экземпл. | 3,61 |
— |
7 |
0 |
1315 |
— |
— |
— |
| 24. ---»--- ---»--- германский ---»--- | 3,82 |
— |
7 |
0 |
1455 |
— |
— |
— |
107
возросла ли средняя емкость черепа у этих семи крупных кроликов настолько,
как этого можно было бы ожидать по их значительно увеличившимся размерам
тела? Мы можем попытаться ответить на этот вопрос двумя путями. В верхней
половине таблицы мы имеем измерения черепов шести мелких диких кроликов
(№ 5 — 10); мы видим, что в среднем череп у них на 0,18 дюйма короче
и емкость на 91 гран меньше, в сравнении с средней длиной и емкостью черепа
трех первых диких кроликов в таблице. У семи крупных вислоухих кроликов
в среднем длина черепа 4,11 дюйма, а емкость 1 136 гран; таким образом
эти черепа прибавились в длину более чем впятеро против того, насколько
убавились в длину черепа шести мелких диких кроликов; отсюда мы могли
бы
ожидать, что черепа крупных вислоухих кроликов прибавятся в емкости
на величину впятеро большую той, на какую убавилась емкость черепа шести
мелких кроликов; это дало бы в среднем увеличение емкости на 455 гран,
тогда как
действительный средний прирост — только 155 гран. С другой стороны,
у больших вислоухих кроликов тело почти того же веса и величины, как у обыкновенного
зайца, но голова длиннее; следовательно, если бы вислоухие кролики
были
в диком состоянии, то можно было бы ожидать, что череп у них будет
той же емкости, как у зайца. Но этого далеко нет, ибо средняя емкость двух
черепов зайца (№ 23, 24) настолько больше средней емкости семи черепов
вислоухих кроликов, что последняя должна возрасти на 21%, чтобы дойти
до
того уровня, как у зайца28.
Раньше я заметил, что если бы у нас было много домашних кроликов той
же средней величины, как дикий кролик, то было бы легко сравнить емкость
их черепа.
Гималайский, московский и ангорский кролик (№ 11, 12, 13 таблицы) лишь
немного крупнее телом и череп незначительно длиннее, сравнительно с
диким кроликом;
и мы видим, что наблюденная емкость их черепов меньше, чем у дикого животного,
и значительно меньше вычисленной соответственно разнице в длине черепа
(столбец VII). Узость мозговой капсулы у этих трех кроликов ясно видима
и доказуема
измерением снаружи. Шиншилловый кролик (№ 4) — животное значительно более
крупное, чем дикий кролик, однако емкость его черепа лишь незначительно
превосходит емкость дикого кролика. Ангорский кролик (№ 13) представляет
в высшей степени
замечательный случай, чисто белая окраска и длинный шелковистый мех кладут
на это животное печать долговременного одомашнения. Голова и тело у него
значительно длиннее, чем у дикого кролика, но действительная емкость
черепа меньше, чем даже у маленьких диких кроликов с Порто-Санто. Приняв
за меру
длину черепа, емкость (см. столбец VII) составляет лишь половину того,
что должно было быть! Я держал этот экземпляр живым, и он не был ни
болезненным,
ни идиотом. Я был настолько удивлен этим случаем, что повторил все измерения
и нашел их верными. Я сравнил также емкость черепа ангорского кролика
с емкостью черепа дикого, руководясь иными мерками, именно длиной и
весом тела и весом
костей конечностей; и по всем этим меркам сравнения мозг оказывается
чрезвычайно малым, хотя и в меньшей степени, если за мерило взять кости
конечностей;
это последнее обстоятельство вероятно, может быть объяснено тем, что
кости конечностей у пород, издавна одомашненных, сильно уменьшились
в весе от продолжительной
недеятельной жизни. Отсюда я заключаю, что у ангорской породы, которая
отличается от других пород, как говорят, более спокойным и общительным
нравом, емкость
черепа действительно претерпела замечательно сильное уменьшение.
Из различных вышеприведенных фактов — во-первых, из того, что действительная емкость черепа у гималайской, московской и ангорской пород меньше, чем у дикого кролика, хотя сами они по всем измерениям несколько крупнее; во-вторых, из того, что емкость черепа у больших вислоухих кроликов не возросла, хотя бы приблизительно, в том же отношении, в каком уменьшилась емкость черепа у более мелких диких кроликов; и в-третьих, из того, что емкость черепа у тех же крупных вислоухих кроликов значительно ниже, чем у зайца, который сам почти такого же роста,— я заключаю (несмотря на замечательные различия в емкости черепов у мелких кроликов с Порто-Санто и также у больших вислоухих), что у всех давно одомашненных кроликов мозг или отнюдь не увеличился в должном соответствии с возросшей длиной головы и величиной тела или что он даже убавился в величине сравнительно с тем, что было бы, если бы эти животные жили в естественном состоянии. Памятуя, что кролики, благодаря тому, что они были одомашнены и содер-
28 Здесь этот уровень, повидимому, слишком низок. Д-р Крисп (Dr. Crisp., Proc. «Zoolog. Soc.» 1861, стр. 86) дает 210 гран как действительный вес мозга зайца, весившего 7 фунтов, и 125 гран, как вес мозга кролика, который весил 3 фунта 5 унций, то-есть столько же, сколько кролик № 1 моей таблицы. Череп кролика № 1 по моей таблице вмещает 972 грана дроби; согласно отношению 125 к 210, указанному д-ром Криспом, череп зайца должен содержать 1 632 грана дроби вместо (как у самого крупного зайца моей таблицы) 1 455 гран.
108
жались взаперти в течение многих поколений, не могли упражнять свой ум,
инстинкты, чувства и произвольные движения с целью ли избежать разных опасностей
или в поисках пищи, мы можем заключить, что мозг их должен был слабо упражняться
и вследствие этого развитие его пострадало. Мы видим таким образом,
что
самый важный и сложный орган во всем организме подчиняется закону уменьшения
в размерах вследствие неупотребления.
В заключение подведем итоги более важных изменений, которым подверглись
домашние кролики, и в то же время рассмотрим причины этих изменений,
насколько мы
можем хотя бы смутно видеть их. Доставлением обильной и питательной пищи
вместе с малым упражнением и постоянным отбором наиболее тяжелых особей
вес крупных пород более чем удвоился. Кости ног, взятые вместе, прибавились
в
весе в должном соответствии с увеличившимся весом тела, но задние ноги
прибавились меньше; в длину кости ног не увеличились в должной пропорции,
и это может
быть вызвано недостатком надлежащего упражнения. С увеличением размеров
тела третий шейный позвонок принял черты, свойственные четвертому шейному
позвонку,
а восьмой и девятый спинные позвонки подобным же образом приняли черты,
свойственные десятому и дальнейшим позвонкам. Череп у крупных пород
прибавился в длину,
но не в должном соответствии с возросшей длиной тела; мозг не увеличился
в размерах должным образом или даже фактически уменьшился, а вследствие
этого костная капсула мозга осталась узкой и чрез соотношение повлияла
на кости
лица и на всю длину черепа. Таким образом череп приобрел характерную
узость. По неизвестным причинам надглазничный отросток лобной кости
и свободный конец
скуловой кости стали шире; у более крупных пород затылочное отверстие
вообще с менее глубокими выемками краев, чем у диких кроликов. Некоторые
части лопатки
и задний отдел грудины стали очень изменчивы по форме. Уши, благодаря
постоянному отбору, до чрезвычайности увеличились в длину и ширину;
вес их, вероятно,
в соединении с неупотреблением мышц, повел к свисанию; это повлияло на
положение и форму костного слухового прохода, а это, в свою очередь,
чрез соотношение
повлияло в слабой степени на положение почти всех костей в верхней части
черепа и даже на положение сочленовных отростков нижней челюсти.
ОГЛАВЛЕНИЕ
| Стр. | ||||
Основы учения Дарвина и обстоятельства, сопровождавшие появление его главных работ |
||||
| — |
|
........................................................................................... | 5 | |
ИЗМЕНЕНИЕ ЖИВОТНЫХ И РАСТЕНИЙ В
ДОМАШНЕМ СОСТОЯНИИ
|
||||
| Предисловие ко второму (английскому) изданию ................................................................. |
26 | |||
| ВВЕДЕНИЕ ................................................................................................................................. | 27 | |||
Глава I. ДОМАШНИЕ СОБАКИ И КОШКИ ............................................................................. |
35 — 66 | |||
|
||||
Глава II. ЛОШАДИ И ОСЛЫ ..................................................................................................... |
57 — 67 | |||
|
||||
Глава Ш. СВИНЬИ.— РОГАТЫЙ СКОТ.— ОВЦЫ.— КОЗЫ .............................................. |
68 — 90 | |||
|
||||
| Глава IV. ДОМАШНИЕ КРОЛИКИ .......................................................................................... |
91 — 108 | |||
|
||||
| Глава V. ДОМАШНИЕ ГОЛУБИ .............................................................................................. |
109 — 139 | |||
|
||||
| Глава VI. ГОЛУБИ (продолжение) ........................................................................................... |
140 — 167 | |||
|
||||
| Глава VII. КУРЫ ......................................................................................................................... |
168 — 198 | |||
|
||||
Глава VIII. УТКИ. ГУСИ. ПАВЛИНЫ. ИНДЕЙКИ.
ЦЕСАРКИ. КАНАРЕЙКИ. ЗОЛОТЫЕ РЫБКИ. ПЧЕЛЫ. ШЕЛКОВИЧНЫЕ ЧЕРВИ .....................................................................
|
199 — 217 | |||
|
||||
| Глава IX. ВОЗДЕЛЫВАЕМЫЕ РАСТЕНИЯ: ЗЛАКИ И ОВОЩИ ........................................ |
218 — 235 | |||
|
||||
| Глава X. РАСТЕНИЯ (продолжение).— ПЛОДОВЫЕ ДЕРЕВЬЯ.— ДЕКОРАТИВНЫЕ
ДЕРЕВЬЯ.— ЦВЕТЫ .................................................................................................................. |
236 — 262 | |||
|
||||
| Глава XI. О ПОЧКОВОЙ ВАРИАЦИИ И О НЕКОТОРЫХ АНОМАЛИЯХ ПРИ
ВОСПРОИЗВЕДЕНИИ И ПРИ ИЗМЕНЧИВОСТИ .................................................................. |
263 — 291 | |||
|
||||
| Глава XII. НАСЛЕДСТВЕННОСТЬ ........................................................................................... |
292 — 308 | |||
|
||||
| Глава XIII. НАСЛЕДСТВЕННОСТЬ (продолжение).— РЕВЕРСИЯ,
ИЛИ АТАВИЗМ ......... |
309 — 329 | |||
|
||||
| Глава XIV. НАСЛЕДСТВЕННОСТЬ (продолжение).— ПОСТОЯНСТВО ПРИЗНАКОВ.— ПРЕИМУЩЕСТВЕННАЯ ПЕРЕДАЧА (ПЕРЕДАЧА ДОМИНИРУЮЩИХ ПРИЗНАКОВ).— ОГРАНИЧЕНИЕ ПОЛОМ.— СООТВЕТСТВИЕ ВОЗРАСТА ........... | 330 — 344 | |||
|
||||
| Глава XV. СКРЕЩИВАНИЕ ...................................................................................................... | 345 — 354 | |||
|
||||
| Глава ХVI. ПРИЧИНЫ, ПРЕПЯТСТВУЮЩИЕ СКРЕЩИВАНИЮ РАЗНОВИДНОСТЕЙ.
ВЛИЯНИЕ ОДОМАШНЕННОГО СОСТОЯНИЯ НА ПЛОДОВИТОСТЬ ........................ |
355 — 363 | |||
|
||||
| Глава ХVII. ПОЛЬЗА СКРЕЩИВАНИЯ И ВРЕД ОТ СКРЕЩИВАНИЯ БЛИЗКИХ
РОДИЧЕЙ ............................................................................................................................... |
364 — 384 | |||
|
||||
| Глава XVIII. ПРЕИМУЩЕСТВА И НЕВЫГОДЫ ИЗМЕНЕНИЯ УСЛОВИЙ
ЖИЗНИ; БЕСПЛОДИЕ ОТ РАЗНЫХ ПРИЧИН .............................................................................. |
385 — 402 | |||
|
||||
| Глава XIX. ОБЗОР ПОСЛЕДНИХ ЧЕТЫРЕХ ГЛАВ И ЗАМЕЧАНИЯ О ГИБРИДИЗМЕ |
403 — 414 | |||
|
||||
| Глава XX. ОТБОР, ПРОИЗВОДИМЫЙ ЧЕЛОВЕКОМ .......................................................... |
415 — 434 | |||
|
||||
| Глава XXI. ОТБОР (продолжение) ........................................................................................... |
435 — 451 | |||
|
||||
| Глава XXII. ПРИЧИНЫ ИЗМЕНЧИВОСТИ ............................................................................. |
452 — 465 | |||
|
||||
| Глава XXIII. ПРЯМОЕ И ОПРЕДЕЛЕННОЕ ДЕЙСТВИЕ ВНЕШНИХ УСЛОВИЙ
ЖИЗНИ |
466 — 470 | |||
|
||||
| Глава XXIV. ЗАКОНЫ ИЗМЕНЧИВОСТИ — УПРАЖНЕНИЕ И НЕУПОТРЕБЛЕНИЕ ОРГАНОВ И ПР. ........................................................................................................................ | 480 — 496 | |||
|
||||
| Глава XXV. ЗАКОНЫ ИЗМЕНЧИВОСТИ (продолжение).— СООТНОСИТЕЛЬНАЯ (КОРРЕЛЯТИВНАЯ) ИЗМЕНЧИВОСТЬ ................................................................................. | 497 — 509 | |||
|
||||
| Глава XXVI. ЗАКОНЫ ИЗМЕНЧИВОСТИ (продолжение).—
ОБЗОР .................................. |
510 — 519 | |||
|
||||
| Глава XXVII. ВРЕМЕННАЯ ГИПОТЕЗА — ПАНГЕНЕЗИС .................................................. |
520 — 550 | |||
|
||||
| Глава XXVIII. ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ ............................................................... | 551 — 567 | |||
|
||||
| АЛФАВИТНЫЙ УКАЗАТЕЛЬ .................................................................................................. | 568 — 611 | |||