ГЛАВА 11
ВТОРАЯ ФАЗА:
ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
Слово deseado по-испански означает «желанный» или «вожделенный» — название,
кажущееся слишком романтическим для гавани и реки на суровом, холодном,
открытом штормовым ветрам побережье южной Патагонии. Его происхождение,
едва ли известное сейчас кому-нибудь из местных жителей, по прошествии
столь многих лет может показаться романтическим, но вряд ли оно казалось
таким тем людям, которые непосредственно участвовали в событиях 1586 г.
Достопочтенный капитан Томас Кендиш, или, как иногда пишут, Кавендиш, плыл
тогда с востока на запад вокруг света на трех кораблях: флагмане «Дизайе»
водоизмещением 120 тонн и двух совсем маленьких судах — «Контент» и «Хью».
Летописец этой «малой Армады», некий Френсис Притти, записал это так: «Декабря
семнадцатого дня после полудня мы вошли в гавань, причем наш флагман шел
первым; по этой причине наш командующий поименовал упомянутую гавань порт
Дизайе» («Желание», англ.).
Разумеется, название было дано в честь флагмана. Попутно интересно отметить,
хотя это и не имеет отношения к нашей теме, что в этой гавани экспедиция
впервые увидала южных морских слонов и магеллановых пингвинов, тогда еще
не имевших этих названий (названия были им даны позднее), но четко описанных
магистром Притти.
Когда пришли испанцы, они превратили «дизайе» в «десеадо», и с тех пор
город и река называются Пуэрто-Десеадо и Рио-Десеадо. Более удивительно
другое, чего, конечно, не могли ожидать ни достопочтенный капитан Кендиш,
ни испанские первопоселенцы, а именно что названию «десеадо» суждено было
стать одним из важнейших понятий в анналах геологии и палеонтологии Южной
Америки.
Во время седьмой экспедиции в Патагонию, летом 1893 — 1894 гг., Карлос
Амегино собрал ценную коллекцию ископаемых остатков млекопитающих в пункте,
называемом Ла-Флета («стрела», напоминание о границе с индейцами), в северной
части провинции Санта-Крус («святой крест») — самой южной из четырех административных
единиц обширной области, называемой в просторечии Патагонией. Это местонахождение
расположено около Пуэрто-Десеадо и устья реки Рио-Десеадо. В 1895 г. Флорентино
Амегино описал происходящую оттуда ископаемую Фауну, которую назвал фауной
Pyrotherium. Впоследствии Андрэ Турнуэр под руководством дона Карлоса собрал
ископаемые остатки этой фауны в Ла-Флеча и в других местах и отправил в
Париж. Там, как уже отмечалось в гл. 2, Альбер Годри, сам никогда не бывавший
в Южной Америке, изучал материалы, собранные Турнуэром. Здесь особенно
уместно отметить, что он применил название «десеадо» для обозначения этой
фауны и геологических слоев, в которых она встречается. В более точном
современном употреблении название десеадий означает век развития наземных
млекопитающих, южноамериканскую фауну млекопитающих, которая определяет
этот век, и ярус, к которой относятся породы, сформировавшиеся на протяжении
этого века.
Здесь следует привести исключительно четкий и поучительный пример того,
как можно в полевых условиях объективно оценить последовательность фаун,
направление эволюции и течение геологического времени. К югу от выраженной
в рельефе центральной впадины Патагонии, в которой лежат два озера -Мастэрс
и Колуэ-Уапи (у местных жителей это обычно звучит как «колуапи»), имеется
большой уступ или крутой склон, который палеонтологи теперь часто называют
Большим Уступом (the Great Barranca). Как геологическое местонахождение
он был открыт — можно было бы сказать «конечно же» — Карлосом Амегино.
Обнажающиеся в этом уступе геологические слои лежат почти горизонтально,
и, поднимаясь вверх по такому обнажению, вы поднимаетесь также во времени
— от более древних эпох к более поздним.
Здесь и, насколько мне известно, только здесь на всем земном шаре человек,
взбирающийся вверх по сплошному непрерывному крутому склону, проходит через
четыре совершенно различных геологических века, каждый из которых имеет
свою, характерную для него одного фауну млекопитающих. По современной,
так сказать «постамегинской», терминологии эти века (снизу вверх, т. е.
от более древнего к более позднему) называют касамайорий, мастэрсий, десеадий
и колуапий. В процессе своей работы дон Карлос начал постепенно постигать
эту последовательность; тем самым дон Флорентино получил непоколебимую
основу для значительной части всей последовательности фаун млекопитающих
Южной Америки. Хотя Годри, не мудрствуя, назвал древнейшую из этих четырех
фаун «касамайор» по местонахождению, на самом деле не опорному и не типичному
для нее, понятие «нотостилопия» братьев Амегино и наше современное понятие
касамайория основаны на фауне, которая в наиболее типичном выражении действительно
представлена в низах Большого Уступа. Это — типовое местонахождение; несколько
отклоняясь в сторону, отметим, что оно послужило также источником используемых
нами названий мастэрсий и колуапий; каждый из этих веков назван по одному
из двух озер, расположенных в этой области. (Мастэрс был английским исследователем,
посетившим Патагонию в XIX в.; Колуэ-Уапи — искаженные слова арауканских
индейцев, означающие «красный остров».) Десеадская фауна также присутствует
на положенном ей месте в Большом Уступе, однако, как говорилось в начале
этой главы, ее типовое местонахождение, от которого произошло ее современное
название, находится в другом месте.
Десеадий в ряде аспектов представляет собой ключевое время и ключевую фауну
для геологической и зоологической истории Южной Америки. Прежде всего в
настоящее время это древнейший век южноамериканских наземных млекопитающих,
для возраста которого в абсолютном летоисчислении имеются надежные радиометрические
данные. Довольно хорошо установлено, что слои с десеадскими млекопитающими
отлагались около 35 млн. лет назад, но что примерно 34 млн. лет назад формирование
этих пород прекратилось. Это позволяет провести достаточно обоснованную
корреляцию десеадия со стандартными европейскими эпохами и веками североамериканских
наземных млекопитающих. Таким образом было установлено, что десеадий соответствует
раннему олигоцену по европейским понятиям и примерно соответствует чадронию
Северной Америки, типом которого служит древнейшая часть отложений знаменитых
Больших бедлендов в Южной Дакоте. Подобный вывод обнадеживает, поскольку
на протяжении некоторого времени большинство палеонтологов, занимающихся
этим вопросом, исходя из данных, считавшихся недостаточными, называли десеадий
«приблизительно» или «условно» соответствующим по возрасту раннему олигоцену.
Оказалось, что это верно.
Другой интересный геологический момент заключается в том, что примерно
в это время поднятие Анд, пусть еще умеренное, начинает становиться все
более заметным. Уже происходят местные или региональные изменения высоты
суши над уровнем моря и вторжения и отступания океана на территории современной
суши, которые в дальнейшем продолжаются вплоть до современности. В раннем
олигоцене в результате прерывистого или эпизодического, но неуклонного
поднятия начали воздыматься холмы, которые впоследствии превратятся в Анды;
этот процесс продолжается и по сей день, судя по землетрясениям на западном
побережье и наличию в Андах действующих вулканов. В десеадский век вулканы
и лавовые потоки впервые появляются среди относительно ровно залегающих
слоев к востоку от холмов, которые в конце концов превратятся в высокие
плоскогорья и горы.
Десеадские осадки Патагонии, как и осадки касамайория и мастэрсия до того
и колуапия после, по-прежнему состоят почти целиком из обломочных пород
вулканического происхождения. Теперь, однако, это не только мелкий пепел,
принесенный ветром с далеких вулканов, а затем переработанный потоками
и измененный выветриванием. В патагонском десеадий есть и мелкий пепел,
и более крупный материал, грубые пеплы и шлаки, обычно также переработанные
водой. В одном особенно интересном местонахождении, найденном моей второй
экспедицией в Патагонию, богатое скопление остатков десеадских млекопитающих
было сконцентрировано главным образом в кармане, который некогда был кратером
маленького вулкана; мы назвали его Карман Скаррит. После того как извержение
вулкана прекратилось, кратер был глубоко погребен, а гораздо позже в результате
эрозии был снова вскрыт в почти поперечном разрезе. В том, что некогда
было мелким кратерным озером, погибло множество леонтиниид (см. гл. 10);
возможно, они были отравлены вулканическими газами, а затем погребены под
пеплом, происходившим из других вулканических выходов этого района.
Десеадские отложения сильно разбросаны по территории южной Аргентины и,
как правило, залегают на размытой поверхности, что указывает на региональное
изменение высоты суши над уровнем моря. Например, в знаменитом местонахождении
Кабеса-Бланка («белая голова»), названном так потому, что оно представляет
собой холм со светлоокрашенными породами на вершине, десеадские слои залегают
непосредственно на касамайорских, т. е. между ними нет ни мастэрских, ни
каких-либо других отложений промежуточного возраста. Такие случаи заставили
на некоторое время ряд палеонтологов, по большей части не аргентинцев и
притом менее опытных и знающих, чем Карлос Амегино, усомниться в существовании
самостоятельного мастэрского века и фауны. Один североамериканец, который
провел в Кабеса-Бланка целое лето, оказавшееся весьма плодотворным (был
найден единственный известный череп Pyrotherium), отрицал присутствие там
даже касамайория; касамайорий-то там есть, а вот мастэрсия нет. Хотя в
других местах мастэрсий существует, но все же непрерывность между мастэрсием
и десеадием нарушена, т. е. в последовательности датированных отложений
и фаун существует перерыв.
В связи с этим возникает вопрос о так называемом дивисадерии, о котором
раньше упоминалось лишь мимоходом. На окраине Мендосы, аргентинского города,
расположенного у подножья Анд, далеко на север от Патагонии, известна геологическая
формация, названная дивисадеро-ларго («место далекого обзора») по названию
холма, сложенного ее породами. Породы эти состоят не из обломочных пород
вулканического происхождения, как в десеадии Патагонии, а из пестроцветных
глин и песков водного происхождения. Они содержат небольшую, чрезвычайно
своеобразную фауну млекопитающих, представители которой относятся к 7 семействам
(два из них сомнительны), 9 родам (один под вопросом) и 10 видам. Все виды
этой фауны, 8 родов наверное и один под вопросом и 2 семейства — одно наверное
(Groeberiidae), а другое — под сомнением (в настоящее время оно не имеет
названия), совершенно неизвестны ни из какой другой фауны, независимо от
ее возраста. Три рода нотоунгулят, по одному виду в каждом, представлены
примитивными формами, очевидно сравнимыми по эволюционному уровню с некоторыми
из касамайорских и мастэрских нотоунгулят. Однако остальные четыре представителя
копытных, два из которых относятся к литоптернам, а два — к нотоунгулятам,
определенно более продвинуты и находятся на той же стадии эволюции, что
и роды, известные в других местах в заведомо десеадских фаунах. Примитивные
и продвинутые копытные бесспорно встречаются здесь вместе, а не в сменяющих
друг друга фаунах.
Фауна формации дивисадеро-ларго по возрасту почти наверное приблизительно
соответствует десеадию, поскольку выживание относительно примитивных форм
гораздо более вероятно, чем раннее появление специализированных животных,
определенно более продвинутых, чем любой вид из хорошо изученных более
ранних фаун. Нельзя полностью исключить возможность того, что эта явно
своеобразная фауна в геологическом смысле чуть древнее или чуть моложе
известных десеадских фаун Патагонии. Паскуаль — ведущий аргентинский исследователь
ископаемых млекопитающих — считает несколько более вероятным, что эта фауна
немного древнее типичной десеадской; на этом основании он предложил выделить
дивисадерский век развития наземных млекопитающих, частично охватывающий
перерыв, выявленный между мастэрсием и десеадием. Я не думаю, чтобы имеющиеся
сейчас данные оправдывали такой предварительный вывод.
Эта разница во мнениях на самом деле не имеет большого значения, но данный
случай представляет собой интерес и заслуживает дальнейшего обсуждения
по другой причине. По единодушному мнению всех, кто знаком с рассматриваемой
фауной, она не является промежуточной между известной мастэрской и типичной
десеадской. Эта фауна аберрантна и никак не укладывается в ту упорядоченную
последовательность, которую образуют почти все остальные южноамериканские
фауны. Имеет ли она в точности тот же возраст, что и десеадская фауна в
других местах, чуточку древнее она или чуточку моложе — не в этом дело;
она — особая, не будучи ни предковой для любой другой известной фауны,
ни происходящей в целом от какой-либо из них. Паскуаль (Pascual, Odreman
Rivas, 1971, с. 386) пишет, что эта фауна «относится к особой географической
области, где архаичные листоядные формы, к тому времени почти совершенно
исчезнувшие из Патагонии, обитали совместно с некоторыми травоядными формами,
подобными хегетотериидам десеадского века». Помимо некоторой географической
обособленности или изоляции такая фауна должна иметь определенные экологические
отличия, хотя мы и не можем установить во всех деталях вызывавшие эти отличия
экологические факторы.
Геологический возраст — это не точка во времени. Это некий определимый
интервал, в геологическом смысле довольно короткий, выделяемый в непрерывном
потоке времени. Тот или иной век развития наземных млекопитающих определяется
не каким-либо постоянным экологическим сообществом, а всеми локальными
сообществами, которые, как можно с достаточным основанием считать, существовали
в одном и том же интервале геологического времени. Как мы уже видели, изученный
риочикий Аргентины и Бразилии служит примером, пусть несколько неполным,
географически разобщенных и экологически различных фаун наземных млекопитающих;
так, например, в Аргентине жили нотоунгуляты, но было мало дидельфид (они
оттуда неизвестны), а в Бразилии было мало нотоунгулят, но много дидельфид.
В десеадии, впервые в излагаемой здесь истории, мы сталкиваемся с многочисленными
локальными фаунами, разбросанными по очень обширной географической области.
Некоторые из них имеют лишь небольшие экологические отличия, однако локальная
фауна Дивисадеро-Ларго, возраст которой, по крайней мере приблизительно,
соответствует десеадию, обладает резко выраженным экологическим своеобразием.
Наиболее важным географическим дополнением к нашим знаниям о десеадских
фаунах служит открытая совсем недавно богатая фауна этого возраста в формации
сайя в Боливии; она имеет некоторое, хотя и менее очевидное, экологическое
своеобразие. Эта фауна собрана в нескольких разных обнажениях невулканических
красноцветных слоев впадины Сайя-Лурибай. В настоящее время эти обнажения
находятся на высоте 3500 — 4000 м; столь большая высота над уровнем моря
влечет за собой значительные отличия местной фауны млекопитающих от фауны
низменностей, но поднятие Анд происходило главным образом в постдесеадское
время. Во времена существования этой десеадской фауны данная территория
уже возвышалась над уровнем моря, но значительно меньше, чем теперь. В
отличие от фауны Дивисадеро-Ларго фауну Сайя можно рассматривать как нормальную,
т. е. соответствующую общему течению эволюции фаун Южной Америки. Полностью
эта фауна еще не описана, но большая часть составляющих ее родов, уже вошедших
в публикации, идентична или близко родственна известным ранее из десеадских
фаун Патагонии. Имеются, однако, некоторые отличия, обусловленные, вероятно,
главным образом экологическими (связанными с окружающей средой) причинами.
Например, токсодонты, леонтинииды и астрапотерии, обычные для большинства
десеадских локальных фаун Патагонии, в фауне Сайя редки, а некоторые наземные
ленивцы и большая часть грызунов, о которых мы подробнее поговорим позже,
представлены иными родами.
Другая и в сущности главная причина нашего особого интереса к десеадию
заключается в том, о чем уже говорилось, особенно в двух последних главах,
и о чем здесь можно сказать в более общем плане. К десеадскому времени
старожилы, т. е. первые поселенцы Южной Америки, довольно хорошо дифференцировались,
и каждая группа значительно продвинулась по пути соответствующей специализации
к тому или иному конкретному образу жизни. Последующая история этих групп
сводилась в основном к двум процессам. Некоторые из них продолжали довольно
медленно развиваться дальше, в тех направлениях, которые уже наметились
в десеадии. Остальные одна за другой исчезали, и в конце концов большинство
их вымерло. Некоторые потомки старожилов, неожиданно появляющиеся на поздних
этапах истории как совершенно новые эволюционные ветви, немногочисленны
и, быть может, только кажутся исключениями. Наиболее поразительным примером
служат аргиролагиды (см. гл. 7), которые неожиданно появляются в палеонтологической
летописи в плиоцене и ничем, за исключением своей принадлежности к сумчатым,
даже отдаленно не напоминают ни одну из известных нам более древних форм.
Самое разумное объяснение заключается в том, что их предки, должно быть,
развивались (начиная, вероятно, с десеадия или с более раннего времени)
в каких-то частях тех обширных регионов, из которых ископаемого материала
пока мало или нет вовсе. Сходный случай в палеонтологической летописи —
появление в самом десеадии рода Groeberia (см. также гл. 7), не имеющего
известных предков или потомков, причем в этом случае мы знаем, что единственная
фауна, в которой обнаружен этот род и это семейство, несомненно исключительна
как в географическом, так и в экологическом аспектах.
Внезапное появление в десеадских фаунах двух групп свидетельствует о совершенно
иных исторических событиях, нежели отмеченные в предыдущем абзаце. С ними
связана еще одна причина нашего особого интереса к этому веку и его фауне.
Имеется в виду вторжение в десеадскую летопись отрядов Rodentia и Primates
(рис. 28), а также вообще их появление на континенте, которое несомненно
произошло в геологическом смысле незадолго до десеадия. Столь же несомненно,
что эти формы не имели предков в Южной Америке в те времена, когда там
обосновались старожилы, и что они вместе со своими потомками составляют
второй фаунистический пласт сформировавшегося в конечном итоге комплекса
южноамериканской Фауны. Когда эти животные достигли Южной Америки, она
уже была островным континентом. Первые иммигранты, принадлежащие к этим
двум отрядам, по всей вероятности, попали в Южную Америку, переселяясь
по цепочке островов или преодолев морское пространство сразу, дрейфуя на
плавнике. При таких способах переселения шансы на успешное достижение другого
материка или острова весьма невелики, но при наличии Достаточно длительного
времени могут быть реализованы. Животные, о которых идет речь, названы
ранними переселенцами по цепочкам островов (поздние переселенцы по цепочкам
островов будут рассмотрены в гл. 14). Эти два отряда необычайно преуспели
в расселении с острова на остров, о чем, помимо всего прочего, свидетельствует
тот факт, что в конце концов представители обоих отрядов «перепрыгнули»
на различные острова Карибского бассейна.
В настоящее время считается общепризнанным, что большая часть, а быть может,
и все многочисленные древние местные грызуны второго фаунистического пласта
Южной Америки произошли на этом континенте от одного предка. Возможное,
хотя и маловероятное исключение составляет семейство североамериканских
дикобразов (Erethizontidae), которое в принципе более своеобразно, чем
остальные семейства, и могло иметь отдельного предка-иммигранта. Таким
образом, следует постулировать существование одного, самое большее двух
стволов случайных иммигрантов. По теории одного ствола весь южноамериканский
комплекс грызунов объединяется в самостоятельную группу, называемую Caviomorpha.
Откуда пришел их предок (или предки) — остается спорным; в сущности это
один из тех вопросов истории и зоогеографии млекопитающих, по которым ведутся
самые острые дискуссии. Все согласны с тем, что вероятной родиной предка
кавиоморф могут быть лишь Северная Америка или Африка. Вопрос в том, какой
именно из этих двух континентов. На обоих континентах существуют грызуны,
которые могли бы, гипотетически или теоретически, дать начало Caviomorpha.
Однако эоценовые формы, несомненно близкие к кавиоморфам, пока не известны
ни на одном их этих континентов. Не известен и тот предок, который в конце
концов высадился на берег Южной Америки. Среди сторонников как той, так
и другой точек зрения есть столь хорошо осведомленные и столь авторитетные
исследователи, соотношение сил с обеих сторон столь сбалансировано, что
едва ли кто-нибудь со стороны может выступить за или против одной или другой
концепции. По А. Э. Вуду (А. Е. Wood) и Б. Паттерсону (В. Patterson), предок
кавиоморф пришел с юга Северной Америки, «перепрыгивая» с острова на остров.
По Р. Хоффстеттеру (R. Hoffstetter) и Р. Лавока (R. Lavocat), он совершил
более длинный (хотя в те времена и не такой длинный, как теперь) трансатлантический
«прыжок» из Африки. То, что Вуд и Паттерсон — североамериканцы, а Хоффстеттер
и Лавока — французы, не имеет отношения к делу. В былые годы я стоял на
стороне североамериканцев; теперь я занимаю нейтральную позицию, ожидая
появления с той или другой стороны каких-нибудь действительно убедительных
доказательств.
А |
Б |

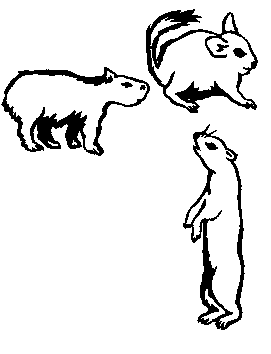
Рис. 28. Ныне живущие потомки животных, занесенных в Южную Америку из-за моря в позднем эоцене или самом начале олигоцена. А. Обезьяна из надсемейства Ceboidea. Б. Несколько кавиоморфных грызунов. Эти примеры приведены здесь для того, чтобы подчеркнуть значение прибытия их предков в Южную Америку. Эти две большие группы — южноамериканские приматы и кавиоморфные грызуны — рассматриваются в нескольких следующих главах.
Откуда бы ни происходил предок кавиоморф, все-таки он добрался до Южной
Америки. К десеадскому времени эта группа уже дифференцировалась на все
четыре основных ствола (надсемейства) и по меньшей мере семь семейств,
представленных примитивными, но различимыми формами. По-видимому, они уже
прожили некоторое время в Южной Америке, но в геологическом смысле это
время не обязательно было очень длительным. Наиболее правдоподобная версия
состоит в том, что предок кавиоморф прибыл в Южную Америку во время перерыва
между мастэрсием и десеадием, для которого в настоящее время не известно
почти никаких палеонтологических данных. На этой обширной и разнообразной
территории прежде не было никаких грызунов. В таких условиях адаптивная
радиация иммигрировавшей группы может происходить очень быстро. Предковые
формы кавиоморф несомненно интенсивно размножались, как это свойственно
грызунам, и за вполне приемлемый промежуток времени, порядка одного, двух
или трех миллионов лет, в десеадии предстают перед нами уже весьма преуспевшими.
Между прочим, один из аргументов в пользу того, чтобы рассматривать фауну
Дивисадеро-Ларго как непосредственно предшествующую десеадию, заключается
в том, что в ней грызуны не известны, хотя они присутствуют во всех изученных
в равной мере десеадских фаунах, Однако десеадские находки отчетливо указывают
на присутствие грызунов непосредственно перед десеадием. Таким образом,
отсутствие их в фауне Дивисадеро-Ларго все-таки следует приписать ее своеобразной
экологии, а не тому, что грызуны попали в Южную Америку позднее, чем существовала
эта фауна, На протяжении многих лет самые древние южноамериканские приматы
были известны из колуапия, приблизительно соответствующего по возрасту
самому позднему олигоцену, но в 1969 г. Хоффстеттер опубликовал описание
одной формы, Branisella, из десеадия Боливии. Сейчас распространение обезьян
Нового Света ограничено тропическими или самое большее субтропическими
лесами; по всей вероятности, так всегда и было, а поэтому их отсутствие
в десеадии Аргентины, если оно не объясняется просто отсутствием находок,
вполне может указывать на то, что климат и растительность Боливии в те
времена были более подходящими для этой группы. Десеадские местонахождения
Боливии находятся и находились примерно на 20° ближе к экватору, чем аргентинские,
а высота Боливии над уровнем моря была в то время невелика.
Здесь снова возникает та же самая биогеографическая проблема, что и с грызунами,
и она снова привела к тому же спору, который сейчас, очевидно, зашел в
тупик. Снова возможные родичи имеются по обе стороны Атлантического океана.
И тем не менее снова ни в Северной Америке, ни в Африке неизвестны эоценовые
формы, которые можно было бы с уверенностью принять за предков южноамериканских.
Было бы заманчиво предположить, хотя на самом деле в этом нет нужды, что
грызуны и приматы, которые, очевидно, попали в Южную Америку приблизительно
в одно и то же время, происходят из одного и того же места. А поэтому неудивительно
снова обнаружить, что, по мнению почти всех американских исследователей
этого вопроса, приматы пришли в Южную Америку из Северной Америки, тогда
как, по мнению французских исследователей, они пришли из Африки. Один американский
исследователь (F. S. Szalay, 1976) сделал забавное предположение, что южноамериканские
обезьяны происходят, возможно, от североамериканских предков, а африканские
обезьяны — от южноамериканских. Снова я соблюдаю нейтралитет, ожидая убедительных
доказательств с той или другой стороны.
Первые южноамериканские приматы вели, вероятно, древесный образ жизни,
как и все их современные потомки. Когда они попали на этот континент, там,
возможно, уже существовали некоторые древесные ксенартры, но, по всей вероятности,
приматы не встретили серьезной конкуренции. Первые южноамериканские грызуны
были преимущественно, если не полностью, наземными (рис. 29, А); все они
имели мелкие размеры и низкокоронковые коренные зубы, функционально (но
не структурно) сравнимые с зубами крыс и мышей. Возможно, они в какой-то
степени конкурировали с несколькими видами мелких сумчатых, но если такая
конкуренция и имела место, то она была не слишком интенсивной, и в общем
примитивные кавиоморфы имели доступ к многочисленным и разнообразным экологическим
нишам, остававшимся незанятыми до их прибытия.

Рис. 29. Некоторые характерные раннеолигоценовые (десеадские) млекопитающие.
А. Реконструкция Platypittamys — одного из древнейших известных нам кавиоморфных
грызунов. Б. Реставрированный череп (по Паттерсону) и реконструкция головы
Pyrotherium. В. Propachyrucos — травоядный нотоунгулят из хегетотериев,
похожий на грызуна. Г. Prosotherium — еще один хегетотерий.
Здесь необходимо уделить внимание в другом контексте одному эволюционному
направлению, прослеженному среди древних местных копытных и частично рассмотренному
в предыдущих двух главах. Размеры примитивных копытных вообще, и примитивных
южноамериканских копытных в частности, варьировали от мелких до средних,
коренные зубы (премоляры и моляры) были у них низкокоронковые (брахиодонтные),
что указывает на большую или меньшую всеядность или питание листьями. В
десеадии еще сохранилось несколько (относительно немного) примитивных неспециализированных
форм, Более характерны для него, однако, крупные копытные, обладавшие различными
типами специализации; они, несомненно, были листоядными и питались довольно
сочной растительной пищей. Среди наиболее ярких примеров последних — пиротерии
и леонтинииды. С экологической точки зрения важно указать на то что, хотя
в десеадии Боливии и встречаются пиротерии и некоторые другие листоядные
формы, судя по опубликованным до настоящего времени данным, они были там
значительно менее обильны, чем в десеадии Патагонии.
Другое общее направление адаптации среди копытных — развитие высококоронковых
(гипсодонтных, а затем гипселодонтных) коренных зубов, что указывает на
питание абразивной растительностью, в особенности растениями, богатыми
кремнием. В миоцене Северной Америки существовало много различных линий
в семействе лошадей. Некоторые из них по-прежнему оставались листоядными,
какими были некогда предки всех лошадей, но другие к концу миоцена стали
травоядными, В Северной Америке все они в конце концов вымерли; листоядные
формы вымерли вообще повсеместно, но травоядные сохранились в Евразии и
Африке, где они представлены настоящими лошадьми, ослами и зебрами. Выше
было показано, что у некоторых нотоунгулят еще в эоцене (мастэрсий) наблюдалась
тенденция к травоядности. В раннем олигоцене (десеадии) существовало несколько
различных нотоунгулят, полностью приспособленных к питанию травой (см.
рис. 25).
Некоторые палеонтологи, знакомые с лошадьми, но не знакомые с нотоунгулятами,
утверждали, что появление жестких травянистых растений, образовавших обширные
прерии, произошло в позднем миоцене или незадолго до этого. В таком случае,
судя по истории нотоунгулят, прериеподобная пампа в Южной Америке должна
была возникнуть на 20 или 25 млн. лет раньше, чем в Северной Америке. Это,
мягко говоря, маловероятно. Освоение нового адаптивного типа развивающейся
группой животных зависит не только от наличия соответствующих возможностей
во внешней среде, но и от наличия в популяции животных с соответствующей
генетической конституцией и способностью адаптироваться к данному фактору
среды. Наиболее вероятная в данном случае гипотеза состоит в том, что травянистая
растительность была широко распространена, хотя и не обязательно в виде
прерий или пампы, уже в эоцене или раньше и что некоторые нотоунгуляты
просто обладали способностью стать травоядными и необходимой для этого
генетической конституцией раньше, чем некоторые лошади.
Итак, довольно о десеадии, которому здесь, вероятно, уделено непропорционально
большое внимание по причинам, о которых было сказано выше. К сожалению,
за десеадием следует почти абсолютный пробел в наших знаниях по млекопитающим
Южной Америки — перерыв более длительный, чем любой другой во всей истории
фаун от риочикия до современности. Промежуток времени между десеадием и
колуапием радиометрически оценивается примерно в 9 млн. лет. Фауны также
свидетельствуют об эволюционных изменениях, соответствующих длительному
времени. Десеадские локальные фауны млекопитающих значительно более многочисленны
и имеют более широкое географическое распространение, чем колуапские, а
поэтому непосредственное сравнение в данном случае неправомерно. Однако
у них, по-видимому, нет ни одного общего вида и только несколько общих
родов. Наиболее существенное изменение — наличие в десеадии нескольких
семейств, которые не известны в более позднее время и, вероятно, вымирают
до колуапия. Среди них сумчатые Polydolopidae (последний их представитель
известен из боливийского, но не из патагонского десеадия), ксенартры Palaeopeltidae,
копытные Pyrotheriidae и Isotemnidae.
В связи с колуапием перед историками возникает вопрос, не слишком серьезный,
но достаточно поучительный, чтобы следовало уделить ему внимание. В Патагонии
имеется широко распространенная группа осадочных пород, которые отлагались
морем, периодически вторгавшимся из Атлантики в пределы нынешней суши.
Уровень моря менялся, причем это происходило в течение довольно длительного,
даже в геологическом смысле, времени, и возраст этих морских отложений
в разных местах различен, хотя обычно их объединяют под общим названием
патагонской формации или группы, используя для ее подразделений локальные
географические названия. Типовой разрез колуапия около Большого Уступа
перекрыт морскими отложениями, представляющими какую-то часть этой группы;
то же самое можно сказать и о примерном эквиваленте колуапия в долине реки
Чубут. Нижнее, в провинции Санта-Крус, типовой разрез сантакрусия подстилается
морскими отложениями, относящимися к патагонской группе. Поэтому можно
сказать (а некоторые так и говорят), что колуапий имеет допатагонский возраст,
а сантакрусий — постпатагонский; на этом основании колуапий и сантакрусий
были разделены и получили отдельные названия. Но подобное утверждение может
ввести в заблуждение. Геологического разреза, в котором колуапские и сантакрусские
слои встречались бы вместе, а часть патагонских морских слоев залегала
бы между ними, неизвестно. Поскольку патагонские слои не везде имеют один
и тот же возраст, возможно, например, что та их часть, которая перекрывает
колуапий, моложе той, которая подстилает сантакрусий. В таком случае в
свете известных нам стратиграфических взаимоотношений нельзя исключить
возможность того, что колуапий одного возраста с сантакрусием или даже
моложе последнего. Историк должен отличать объективно известные данные
(например, последовательность фаун и веков в Большом Уступе) от результатов
умозаключений, подобных только что описанным стратиграфическим соображениям.
Принципы эти важны, но в данном случае, как я уже сказал, сама проблема
не очень серьезна. Известные колуапские и сантакрусские фауны не совсем
идентичны, и при внимательном изучении развития эволюционных линий, представленных
и в той и в другой фауне, становится практически несомненным, что колуапий
действительно древнее, чем сантакрусий, как уже было установлено братьями
Амегино, хотя они и пользовались другими терминами. Таким образом, с последовательностью
все в порядке, но фауны этих двух веков тем не менее очень сходны, и даже
в большей степени, чем обычно требуется для формального выделения самостоятельных
веков развития фауны. Выделение колуапия в качестве самостоятельного века,
а не раннего сантакрусия — дело вкуса и традиции и не обусловлено реальной
необходимостью, Традиция — за выделение этих двух веков как самостоятельных,
а в моем вкусе — следовать в этом отношении обычаю.
Сантакрусские фауны (рис. 30 и 31) известны только из ограниченного района
в Южной Патагонии, но они особенно важны для излагаемой нами истории, поскольку
эти фауны чрезвычайно богаты и представлены в музеях большими коллекциями,
содержащими удивительно много почти полных скелетов отдельных млекопитающих,
о чем уже говорилось в предыдущих главах. Сантакрусские фауны представляют
собой продолжение второй фазы истории южноамериканских млекопитающих. Сообщества
млекопитающих в этой фазе, начало которой возвестил десеадий, представляют
собой комплекс потомков первых поселенцев, относящихся к отрядам Marsupialia,
Xenarthra, Condylarthra, Litopterna, Notoungulata и Astrapotheria, к которому
позже добавились Rodentia и Primates, переселявшиеся по цепочке островов.
(Кондиляртры в сантакрусии не найдены, однако известно, что в то время
они существовали в Южной Америке, поскольку они присутствуют как в более
древних, так и в более молодых фаунах.)
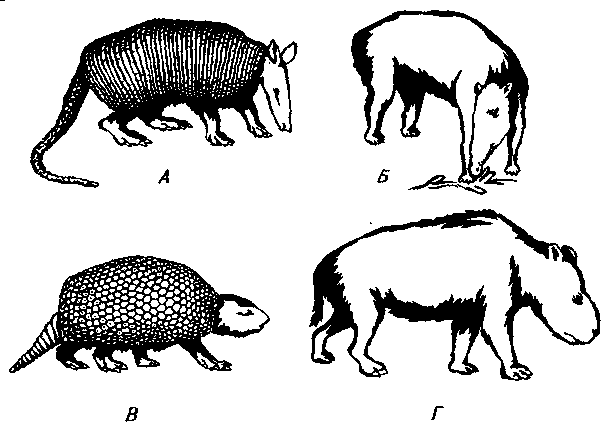
Рис. 30. Некоторые раннемиоценовые (сантакрусские) млекопитающие. А. Реконструкция Stegotherium — броненосца, питавшегося муравьями. Б. Листоядное копытное с бивнями Astrapotherium — типовой род отряда Astrapotheria. В. Propalaeohoplophorus — ксенартр из глиптодонтов. Г. Nesodon — нотоунгулят из токсодонтов.
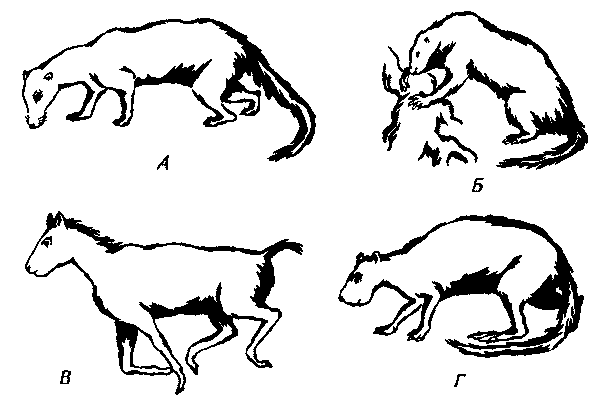
Рис. 31. Реконструкция некоторых раннемиоценовых (сантакрусских) млекопитающих. А. Prothylacinus — хищное сумчатое из боргиенид. Б. Hapalops — наземный ленивец из мегалонихид (отряд Xenarthra). В. Thoatherium — однопалый литоптерн. Г. Neoreomys — кавиоморфный грызун.
Существует несколько локальных сантакрусских фаун, но все они найдены
в одном и том же регионе и, за исключением деталей, сходны по составу.
Слои, в которых они встречаются, отложены ручьями и реками и образованы
в основном обломочными породами вулканического происхождения; они более
или менее сходны с более древними осадками, формировавшимися в других частях
Патагонии от касамайория до колуапия, но содержат также некоторое количество
галечных отложений, связанных с эрозией воздымавшихся на западе Анд. Вся
масса осадков отлагалась на низкой наклонной равнине, оставшейся после
отступания патагонского моря. В этом районе, вероятно, выпадало достаточное
количество дождей и было достаточно много деревьев, кустарников и трав,
чтобы прокормить большие стада листоядных и травоядных млекопитающих. Климат
также был, вероятно, значительно более ровным и теплым, чем сейчас в этой
области; возможно даже, что он был ровнее и теплее, чем в начале кайнозоя
или когда-либо впоследствии. Одним из указаний на такой климат служит,
вероятно, то, что в Патагонии ископаемые остатки обезьян найдены только
в колуапии и сантакрусии; это свидетельствует о существовании в те времена
в этом регионе тропических или субтропических лесов, которых, возможно,
там не было ни раньше, ни позже. Последний вывод, однако сомнителен, поскольку
остатки приматов вообще редки в большинстве южноамериканских ископаемых
фаун и, быть может, просто не попадались в сборах.
Огромным разнообразием отличались сумчатые, в частности ценолестиды и хищные
боргиениды; то же самое относится к броненосцам, глиптодонтам и особенно
к наземным ленивцам. Продолжалась экспансия грызунов, которые, в пределах
своего пока еще ограниченного размерного класса, стали близки к доминированию
в этих сообществах. Нотохиппиды — травоядные нотоунгуляты — не отличались
разнообразием, но они были очень многочисленны в более ранних локальных
фаунах. По-видимому, они вымерли перед концом сантакрусия. Остальные травоядные
нотоунгуляты по-прежнему оставались обычными.
Следует отметить, что в сантакрусских слоях обнаружено также довольно много
ископаемых остатков птиц. Среди них — древнейший из известных представителей
семейства нанду, или так называемых южноамериканских страусов. Там было
много также фороракид — нелетающих бегающих птиц; некоторые из них были
огромных размеров, с большими крючковатыми клювами. Иногда высказывают
мнение, что эти и другие нелетающие южноамериканские птицы, например нанду,
выжили благодаря тому, что на этом континенте долгое время не было плацентарных
хищников. Такое предположение далеко не убедительно. Нанду существуют и
по сей день, хотя плацентарные хищники входят в состав одних с ними сообществ
уже на протяжении по меньшей мере двух миллионов лет. Большинство фороракид
вымерло до того, как плацентарные хищники попали в Южную Америку, и только
одна или две последние уцелевшие формы — после этого. Многие из боргиенид,
живших среди этих птиц на протяжении многих миллионов лет, были в высшей
степени хищными, и вряд ли можно думать, что они не были способны убивать
их. Более того, сами фороракиды явно были хищниками, и, по всей вероятности,
скорее они убивали млекопитаюших, а не наоборот. Их основную пишу, по-видимому,
составляли мелкие нотоунгуляты и некоторые грызуны, после того как они
появились.
Хотя номинальное типовое местонахождение фриасия, так же как и остальных
веков развития крупных млекопитающих Южной Америки, находится в Аргентине,
здесь этот век охарактеризован недостаточно, и его фауна, или фауны, известны
плохо. Остатки нескольких млекопитающих, которых сейчас относят к этому
веку, были собраны Карлосом Амегино и описаны Флорентино Амегино, назвавшим
одну из местных фаун в своем французском тексте Friaseenne по реке Рио-Фриас;
при этом он конкретно указывал на ископаемые остатки, собранные там Сантьяго
Ротом. Рот сделал сходные сборы в различных местонахождениях вдоль восточно
го фасада Анд в Патагонии; значительно позднее, в 1930 г., Лукас Краглевич
(Lucas Kraglievich) повторно изучил экземпляры из Рио-Фриас, Рио-Феникс,
Лагуна-Бланка и других мест и показал, что именно лежит в основе так называемого
фриасского века и фауны, хотя едва ли можно сказать, что этот век имеет
какое-то одно типовое местонахождение или типовую фауну. Немногие сведения
о фриасских фаунах Аргентины, которыми мы располагаем, указывают на продолжение
того, что я назвал здесь второй фазой истории, причем по своей эволюционной
продвинутости известные отсюда млекопитающие занимают промежуточное положение
между лучше изученными формами из более древнего сантакрусия и более позднего
часикия. В геологическом отношении фриасские слои Патагонии свидетельствуют
о происходившей в это время следующей, более резко выраженной фазе поднятия
растущих Анд и о том, что эти горы, вероятно, начинали отбрасывать ту самую
тень, которая заслонила Патагонию от дождей и превратила ее в пустыню.
Значительно лучше известна относимая к фриасию фауна Ла-Вента в Колумбии,
собранная Р. А. Стёртоном (R. A. Stirton) из Калифорнийского университета
и другими исследователями в 40-х годах, но она все еще полностью не описана.
Хотя в Колумбии и Венесуэле было сделано несколько отдельных более древних
находок, фауна Ла-Вента — самая древняя из достаточно хорошо известных
фаун млекопитающих в северной части Южной Америки. В основном она отражает
постепенное развитие главным образом тех же групп, которые присутствовали
в сантакрусии Патагонии. Из этого следует, что в то время существовала
совершенно особая южноамериканская фауна млекопитающих и что она встречалась
лишь с локальными географическими различиями как на крайнем юге, так и
на крайнем севере этого континента. Из Ла-Венты происходит последний известный
нам кондиляртр Megadolodus, а также странный интератерий Miocochilius,
кратко рассмотренный в гл. 10.
По-видимому, наиболее интересная черта фауны Ла-Вента состоит в том, что
в нее входит по крайней мере три, а возможно, четыре вида обезьян, отнесенных
Стёртоном к трем родам, один из которых, Homunculus, впервые найден в сантакрусии
Патагонии. В то время, как и сейчас, Колумбия находилась в тропиках, и
этот район, вероятно, был частично покрыт лесами, хотя, судя по некоторым
другим млекопитающим, там были и саванны. Осадки формации ла-вента были
отложены водотоками бассейна реки Магдалены, уже тогда сформировавшегося
как основная система стока области, занятой современной Колумбией. Помимо
млекопитающих в составе фауны Ла-Вента много рыб, некоторые водяные змеи
и исключительно разнообразные крокодилы.
Лежавшая к востоку от Анд основная зона формирования осадков, в которых
были погребены остатки млекопитающих после сантакрусия сместилась к северу.
Во фриасское время эти осадки распространялись вдоль подножия Анд в северную
Патагонию. В следующий век, часикский, названный по местонахождению на
юге провинции Буэнос-Айрес, осадконакопление начало распространяться на
обширную равнину, которая сейчас представляет собой область аргентинской
пампы. Млекопитающие этого возраста обнаружены в нескольких местонахождениях,
часть которых находится еще дальше к северу, чем Часико. Типовая фауна
отражает еще одну стадию развития фаун второй фазы и не содержит существенных
дополнений. Степень изученности этого века и его фаун такова, что, по-видимому,
необходимо как вновь изучить имеющийся материал, так и провести дальнейшие
его сборы.
В следующем веке развития наземных млекопитающих, уайкерии, начинают появляться
первые провозвестники третьей фазы, или Великого американского обмена;
к его рассмотрению мы приступим в гл. 15.
ЛИТЕРАТУРА
Hirschfeld S. E., Marshall L. G. 1976. Revised faunal list of the La Venta
fauna (Friasian — Miocene) of Colombia, South America. Journal of Paleontology,
50, 433 — 436.
Hoffstetter R. 1974. El origen de los Caviomorpha у el problema
de los Hystricognathi (Rodentia). I. Congreso Argentine de Paleontologia
у Bioestratigrafia,
preprint, 27 p. Hoffstetter R. 1976. Introduction au Deseadien de Bolivie.
Paleovertebrata, 7 — III, 1 — 14.
Hoffstetter R. 1977. Phylogenie des Primates. Bulletin et Memoires, Societe
d’Anthropologie de Paris, 4, 327 — 346.
Lavocat R. 1974. What is an hystricomorph? Symposia of the Zoological Society
of London, 34, 7 — 20.
Pascual R., Odreman Rivas O. E. 1971. Evolucion de las comunidades
de los vertebrados del Terciario Argentine: los aspectos paleozoogeograficos
у paleoclimaticos relacionados. Ameghiniana, 7, 373 — 412.
Simpson G. G. 1974. Chairman’s introduction. Taxonomy. Symposia of the
Zoological Society of London, 34, 1 — 5 (Введение в дискуссию Лавока и
Вуда о происхождении и классификации «гистрикоморфных» грызунов.)
Szalay F. S. 1976. Systematics of the Omomyidae (Tarsiiformes, Primates),
taxonomy, phylogeny and adaptations. Bulletin of the American Museum of
Natural History, 156, 159 — 449. (Содержит дискуссию о происхождении южноамериканских
приматов.)
Wood А. Е. 1974. The evolution of the Old World and New World hystricomorphs.
Symposia of the Zoological Society of London, 34, 21 — 60. (См. также ссылку
на работу Скотта (1932), приведенную в конце гл. 2. К сожалению, после
Амегино не было сделано ни одного надежного обзора или ревизии аргентинского
десеадия.)
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ...............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
................................................ 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ .....................................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .................................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
.......................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ........................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252