| Главная |
или
Условные обозначения:
Чёрным цветом дан текст книги В. Тена «Человек изначальный. Из пены морской»
Зелёным цветом дан текст комментариев П. Волкова
Стр. 204:
Формирование морфологического типа зубатых и усатых китов началось около 60 млн лет назад. Этот процесс непосредственно связан с самой масштабной трансгрессией Мирового океана, когда наряду с четырьмя известными нам океанами образовался из группы морей пятый – Тэтис. Акваторией Тэтиса являлась вся Европа, за исключением приподнятой части Скандинавии. Сюда входят Русская равнина, Западная Сибирь и далее на юг, включая тот сектор земного шара, который ныне занят Индостаном. Не существовало не только Индостана, но также нагорий и гор Передней Азии, Средней Азии, Кавказа, Гималаев.
Не совсем правильно пишет Тен. Индостан
исправно существовал, и мало того – считается, что именно там наземные млекопитающие
сделали первые шаги по пути приспособления к водному образу жизни, породив в
дальнейшем китообразных. Просто Индостан был в это время в другом месте – он
дрейфовал огромным островом с юга на север, теряя за собой мелкие острова и
ещё только готовясь врезаться в Азию и поднять этим столкновением Гималаи.
В то время, конечно, никто не знал, что киты будут усатыми и зубатыми, и даже
не предполагалось их постоянноводного образа жизни, характерного для настоящего
времени. Формировались только самые основные признаки китов как отряда вообще.
Причём архаичнейшие китообразные в то время даже не были полностью водными животными.
Но, пожалуйста, продолжайте.
Стр. 204:
Все это занял разлившийся Тэтис, соединявшийся на юге с Индийским океаном, на
севере – с Ледовитым, на западе – с Атлантическим, на юго-востоке – с Тихим
(там, где ныне Зондский архипелаг, которого в меловом периоде тоже не существовало).
В широтном направлении от того места, где ныне высятся Пиренеи, до современной
Новой Гвинеи дно Тэтиса представляло собой геологически активную зону, которая
термически подогревала акваторию. Это была гигантская геосинклиналь, которая
около 17 млн лет назад взбухла Альпийско-Гималайской складчатостью, породив
крупнейшую в мире горную систему.
Здесь, на изрезанных и малопредсказуемых берегах Тэтиса, неизвестные науке сухопутные
животные 60 млн лет назад начали эволюционировать в малых и больших китообразных.
Ну, остатков предковых китообразных возрастом 60 млн лет (средний палеоцен) ещё неизвестно, поэтому данное утверждение Тена безосновательно. А вот Himalayacetus возрастом 53,5 миллионов лет уже известен, правда, лишь по немногим останкам, но зато уже несущим некоторые основные признаки китообразных, о которых, заметим, Тен не говорит ни слова на протяжении всей книги. Обычно же первым в ряду китообразных называют более известного пакицетуса (Pakicetus). И ближайшие родственники ранних китообразных известны – это парнокопытные индохиусы (Indohyus).
Стр. 205:
Побудило их к этому геологическое половодье. Вода подобралась и затопила места
их обитания не за один день и даже не за одно тысячелетие. У них было время
изменить свои привычки сухопутных существ на привычки амфибий, а потом эволюционировать
в чисто морские существа. На это у них было в запасе примерно 55 млн лет, ибо
эволюция дельфинов, сделавшая их совершенными морскими хищниками, завершилась
примерно 5 млн лет назад.
Вряд ли тогда готовился Всемирный Потоп – он до сих пор существует только в сказках. И вряд ли они перешли к жизни в воде, потому что она пришла к ним – это они пришли к воде, потому что им просто было выгоднее искать пищу в воде при очень небольшой конкуренции (крупные морские рептилии мезозоя вымерли). Времени на полный переход к морскому образу жизни потребовалось значительно меньше: уже в эоцене около 42 миллионов лет назад моря бороздили первые постоянноводные киты типа базилозавра и дорудона. Думаю, кто-нибудь даже вспомнит серию из научно-популярного сериала «Прогулки с чудовищами», где показана драматичная жизнь этих животных.
Стр. 205:
Но как сформировались эти два отряда, сходные по среде обитания, но столь непохожие
внешне, а также по способу питания (одни хищные, другие – мирные потребители
планктона)? Ясно, что предками и тех и других являлись сухопутные животные,
ушедшие в воду. Но почему это должны были быть одни и те же животные, если результат
столь разный?
Отвечаю на последний вопрос: потому что
на момент написания анализируемой книги Тена (напомню, 2019 год) было собрано
достаточно много ископаемых остатков архаичных китов, чтобы на их основании
можно было сделать именно такой вывод. И даже на момент первого издания его
работы о ранних китах уже было известно намного больше, чем, допустим, в 70-е
или 80-е годы прошлого века. Или это был риторический вопрос?
Кроме того, зубатых и усатых китов не всегда считают отдельными отрядами. В
традиционной систематике это подотряды внутри одного отряда, а сторонники кладистики
вообще считают их инфраотрядами внутри отряда китопарнокопытных (Cetartiodactyla).
Стр. 205:
Впервые идея о дифилетическом происхождении китообразных была высказана в конце
XIX в. Смысл ее заключается в том, что китообразные представляют собой не один,
а два различных отряда морских млекопитающих, происходящих от различных предков.
Исследователям конца XIX века простительно
выдвигать такие гипотезы. Палеонтология делала лишь свои первые шаги, ископаемые
находки были крайне скудны и не показывали всей картины эволюции – в данном
случае китообразных. Биохимический анализ белков, сравнение ДНК – этих технологий
и в помине не было. Собственно, о молекуле ДНК тогда даже не знали.
У нас же на дворе, напомню, начало XXI века, и наука сделала шаг вперёд по сравнению
с концом XIX века. Многие ошибки во взглядах исследователей прошлого уже стали
частью истории науки благодаря новейшим исследованиям.
Стр. 205:
В течение XX столетия данная версия постоянно находила новые и новые подтверждения.
Различия в строении мозга говорят о дифилетическом происхождении зубатых и усатых
китов. «Это относится к местам выхода черепных нервов из ствола головного мозга,
внутричерепным взаимоотношениям, местам и характеру выхода из полости черепа,
внечерепным разветвлениям, топографии концевых ветвей, структуре нервно-тканевых
отношений, рецепторных и моторных аппаратов» (Хоменко, 1984, с. 146).
К сожалению, эта версия не подтвердилась. Тен оперирует, мягко говоря, весьма несвежей информацией. 1984 год – это время, когда НЕ были известны многие из современных находок, проливших свет на раннюю эволюцию китообразных и дивергенцию усатых и зубатых китов. Даже полный скелет пакицетуса не был известен: его открыли только в 2001 году (напомню, до первого издания книги Тена). Данные палеонтологии говорят о том, что все современные киты имеют вполне монофилетическое происхождение, являясь потомками археоцетов. Но многие анатомические признаки формировались у них параллельно и независимо друг от друга – например, сдвигание ноздрей назад и телескопизация черепа.
Стр. 205-206:
Строение мозга при прочих равных обстоятельствах имеет при установлении таксономических
различий приоритет. Дело в том, что мозг – самая консервативная часть организма.
Если с другими органами эволюция смело экспериментирует (вот почему нельзя полагаться
на внешнее сходство, на котором основан симиализм), то от мозга она никогда
ничего не отнимает, ничего не редуцирует (как, например, хвост), не атрофирует
(как, например, неиспользуемые мышцы), только добавляет сверху в случае прогрессивного
развития.
Хотелось бы увидеть тот источник, в котором
говорится о приоритете строения мозга над строением других частей тела. Естественно,
ссылок на него Тен не приводит, хотя во время нападок на книги Маркова и Дробышевского
он любезно приводил указания на том и страницу, что очень облегчало мне работу.
Внешнее сходство – это критерий, которым пользовались зоологи и ботаники веке
эдак в XVIII-XIX. Естественно, применение этого метода при недостаточной изученности
той же анатомии неизбежно приводило к ошибкам, которые, однако, исправлялись
позднейшими более тщательными исследованиями. В настоящее время биологи уже
научились отличать конвергентное сходство от признака, унаследованного от общего
предка. Например, ластоногие были признаны полифилетической группой и включены
в отряд хищных на правах трёх отдельных семейств. Среди тех же хищников «порублено»
семейство виверровых, из которого выделены семейства нандиниевых, мангустовых
и линзанговых. Также двупалые и трёхпалые ленивцы разошлись по собственным отдельным
семействам. Всё это сделано на основании более тщательных анатомических исследований,
плюс на основании данных по генетике, полученных в последнее время.
Тем не менее, анатомический критерий работает, и довольно неплохо. Так, классификация
на уровне отряда по анатомическим данным вполне корректна. Посмотрим хотя бы
на названия тех же отрядов млекопитающих. Рукокрылые, хоботные, грызуны, хищные
– эти названия даны на основании внешнего облика представителей отряда.
Кроме того, важным подспорьем в работе систематиков является палеонтология.
Пути эволюции живых существ, восстановленные на основании ископаемых остатков,
помогли понять, что парно- и непарнокопытные – это два разных отряда, а не один,
как говорилось в старой литературе. «Толстокожие» (Pachydermata), объединявшие
слонов, носорогов и бегемотом, «разбрелись» по трём разным отрядам.
Далее, если допустить, что точка зрения Тена верна, и строение мозга имеет значение
для систематики, как мы можем устанавливать родственные связи у вымерших млекопитающих,
представителей полностью вымерших групп животных? От очень многих из них не
то, что мозг – даже череп неизвестен. Но палеонтологи успешно устанавливают
их родство на основе строения зубов. Уж в этом вопросе они изрядно поднаторели,
и легко отличат, например, зуб некрупной лошади от зуба осла, и даже различат
зубы летучей мыши, суслика и крота.
Естественно, что ни один признак, взятый в отдельности, не сможет служить основанием
для выводов по систематике, поэтому время от времени воззрения на систематику
и родственные отношения представителей тех или иных групп живых организмов меняются.
Но это происходит не по произволу, а строго на основании оценки и переоценки
имеющихся и вновь полученных данных.
Стр. 206:
Отсюда такие понятия, как «старый мозг» (палеокортекс) и «новый мозг» (неокортекс).
Новый мозг не сменяет старый, а вырастает сверху. Поэтому палеокортекс может
содержать тайны палеопсихики, когда сознание и неокортикальные структуры, являющиеся
его носителями, были только в проекте. Большинство зоологов-систематиков в настоящее
время поддерживают дифилетическую теорию.
Лингвист из господина Тена плохой: «кортекс»
– это не «мозг», а «кора». Кора – это всего лишь часть мозга. Неокортекс у млекопитающих
не «нарастает сверху»: чисто анатомически он расположен не выше и не ниже других
элементов коры полушарий мозга – архи- и палеокортекса.
«Первые слабые намеки на развитие коры по типу млекопитающих можно усмотреть
у ряда рептилий (…). У этих форм в полушарияхмежду древней и старой корой располагается
небольшой участок поверхностного серого веществ нового типа – область новой
коры (neopallium)» (А. Ромер, Т. Парсонс «Анатомия позвоночных», М.: Мир, 1992,
том 2, стр.308).
К сожалению для Тена, зоологи-систематики в настоящее время не поддерживают
идею дифилетического происхождения китообразных. Палеонтология и молекулярные
данные свидетельствуют строго против этого. В книге “Reproductive Biology and
Phylogeny of Cetacea: Whales, Porpoises and Dolphins” (редактор Debra Lee Miller,
Science Publishers, 2007 год, раздел 3.3.3, стр. 110-111 (источник
и для особо интересующихся)
однозначно указывается на монофилетическое происхождение усатых и зубатых китов.
Стр. 206:
Еще в первой книге по проблеме антропогенеза «Из пены морской...« (2005 г.) я усомнился в правильности версии «копытного» происхождения дельфинов, которая настолько общепринята, что попала даже в энциклопедии и детские издания. Мои сомнения основывались на законе Долло о необратимости эволюционных изменений.
Сомнение сомнению рознь. Если господин
Тен усомнился в чём-то из области биологии – это личные сомнения самого Тена,
не имеющие значения для науки, поскольку господин Тен не является специалистом
в области биологии. Такие сомнения обычно являются следствием личной неинформированности
сомневающегося и довольно легко излечимы интенсивным изучением материала по
теме. А вот если сомневается именно специалист, положивший жизнь на изучение
какой-то темы, причём сомневается именно в области своей специализации – такие
сомнения стоит принимать в расчёт хотя бы в силу значительно большей информированности
специалиста по сравнению с неспециалистом.
И за общепринятостью некоторой научной точки зрения тоже что-то стоит, верно?
Например, свидетельства в пользу этой точки зрения реальными фактами из разных
областей биологии.
Закон Долло я не раз вспоминал тёплым словом по ходу анализа книги, и не раз
ещё вспомню: очень уж часто приходится обращаться к нему. Но это лирика, а пока
мы продолжаем наше удивительное путешествие.
Стр. 206:
Копыта всех форм представляют собой не исходный, а конечный результат эволюции.
Универсальным нуклеусом эволюции является трех- или пятилучевая кисть, доставшаяся
млекопитающим от земноводных предков. Копыта образуются из отвердевших площадок
на разных уровнях сгиба кистей или на подушечках лучей.
Всякий конец – это начало чего-то нового.
Смотря какой отрезок времени брать, конечно. Если брать историю млекопитающих
вообще, то копыто не первично; первичен коготь. А если брать более узкий момент
истории млекопитающих – происхождение китообразных – то тут копыто является
несомненным началом, поскольку предки китов были копытными существами. Здесь
Тен лукавит, подменяя более узкое понятие (история китообразных как отряда)
более широким (история млекопитающих как класса), с заведомо отличной начальной
точкой.
По ходу чтения возникает вопрос: давно ли господин Тен перечитывал хотя бы школьные
учебники по зоологии? Откуда в его рассуждениях внезапно появляются какие-то
«сгибы»?
Строение конечности позвоночных (в частности, млекопитающих) довольно консервативно.
Элементы конечности могут срастаться, боковые элементы могут редуцироваться,
но количество тех же суставов остаётся неизменным. В передней конечности мы
находим локтевой сустав и запястный сустав (неграмотно называемый «коленом»,
и это в передней-то ноге!). В задней конечности наблюдаем колено и скакательный
сустав, соответствующий голеностопному суставу стопоходящего животного. Плюс
подвижные межфаланговые суставы. Никаких лишних «сгибов» в ноге нет. Анатомия
ноги той же лошади изучена досконально, учитывая огромную роль этого животного
в жизни человека и развитии цивилизации.
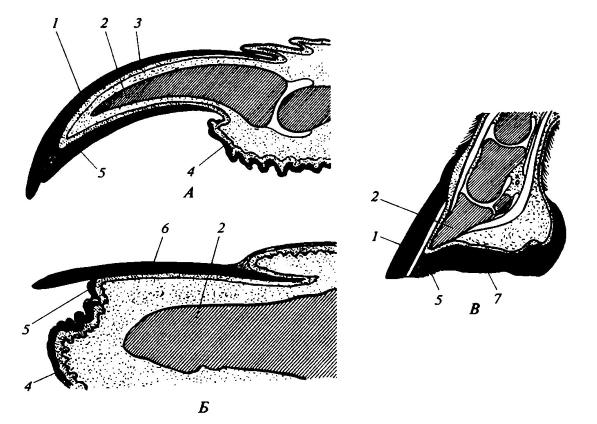
Повторю здесь ещё раз рисунок, на котором показано сравнение костей передней
конечности лошади (как представителя копытных) и человека. Гомологичные кости
закрашены одинаковым цветом.
Здесь наглядно видно, что план строения
конечности – общий.
Теперь немного о копытах.
«Когти, ногти и копыта — это кератинизированные эпидермальные структуры на кончиках
пальцев. Исходным типом является коготь, а ноготь и копыто — его модификации,
возникшие у млекопитающих» (источник: «Век
млекопитающих», процитирована книга А. Ромера и Т. Парсонса «Анатомия позвоночных»,
М., «Мир», 1992, том 1, стр. 168).
|
«…у наземных травоядных, у которых число
пальцев уменьшено и тело при передвижении опирается на кончики оставшихся пальцев,
усложнение когтей привело к формированию толстых роговых копыт. Их мощные когтевая
и подошвенная пластинки, а у лошадей и ороговевшая подушечка пальца (стрелка)
в совокупности образуют футляр, почти целиком охватывающий концевую фалангу.
При этом передняя стенка копыта соответствует когтевой пластинке, его подошва
— подошвенной пластинке, тогда как относительно твердая стрелка вдается в копыто
угловатым выступом сзади. Когтевой валик в копыте совершенно не выражен» (источник:
«Век млекопитающих», там
же).
«Копыто – ungula… . Если твёрдый кожный наконечник с формой когтя вместо простой
функции цепляния перестраивается на роль опоры о землю, присущую в примитиве
мякишу, то получается явление, обратное тому, что мы видели у когтя. В таких
случаях в наконечниках сильно развивается подошвенная часть и форма крючка изменяется
на усечённый конус. Запястный и подошвенный мякиши подвергаются сильной редукции
или совершенно исчезают, а пальцевый мякиш становится рядом с наконечником своеобразным
рессорным механизмом, как, например, стрелка у лошадей» (источник: «Всё
о животноводстве»).
И об «общепринятости» утверждения о происхождении китообразных от парнокопытных.
Ещё несколько десятилетий назад китообразных «выводили» от совершенно разных
млекопитающих – то от мезонихид, то от гипотетических меловых насекомоядных,
то ещё от кого-нибудь. Это было связано с недостаточностью палеонтологических
находок. Но находки последних лет вкупе с данными генетики дали нам доказательства
родства китообразных именно с парнокопытными. Этот вывод был сделан не просто
так, потому что кому-то этого захотелось, а основан на вполне материальных и
проверяемых доказательствах. Потому эта версия и общепринята, в отличие от выкладок
Тена, писаных вилами по воде.
Стр. 206-207:
В основе всех конечностей всех млекопитающих и птиц, населяющих Землю, та конечность,
которая досталась им от очень далеких земноводных предков.
[…]
Собака и кошка бегают «на цырлах». Их подушечки представляют собой то же самое,
что подушечки наших четырех пальцев, а большой палец редуцировался, и остаток
его можно видеть выше на лапе.
«На цырлах» пусть бегает господин Тен лично. В биологии способ передвижения перечисленных животных называется «пальцехождение».
Стр. 207:
Другая эволюционная модель воплощена в конечностях парнокопытных животных: быка,
оленя, козла, кабана и др. Они передвигаются на согнутых пальцах, на которых
от трения о землю образовались роговые наросты. Пальцы срослись, а в одном месте,
в центре, наоборот, разошлись.
Третий вариант ее реализации представлен у лошади и других непарнокопытных.
Это полностью согнутая кисть, на которой наросли копыта.
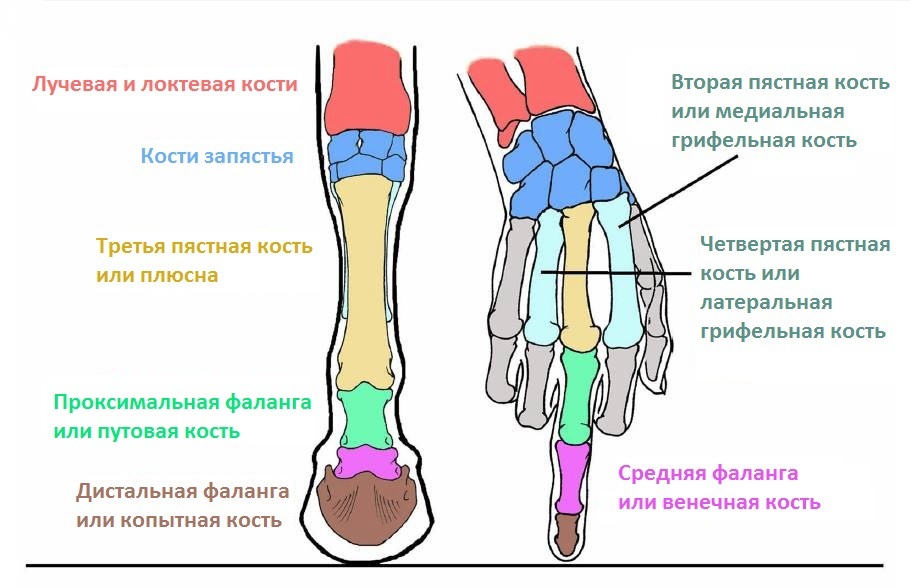
Я прочитал в разное время много книг по биологии. И 80-х годов, и новые, современные, и совсем старинные – вроде книг А. Э. Брема и В. Гааке, написанных в конце 19 века. Однако ни в одной из этих книг я не прочитал такого бреда, который сейчас читаю в книге Тена, изданной в 2019 году и повторяющей в данном абзаце его же книгу 2005 года. Я мог бы привести в качестве аргумента школьный учебник зоологии Козлова и Олигера, по которому учился, и в котором наглядно показано строение конечности парно- и непарнокопытного. Но я просто покажу здесь рентгеновские снимки дистальной части ног коровы (парнокопытного) и лошади (непарнокопытного).
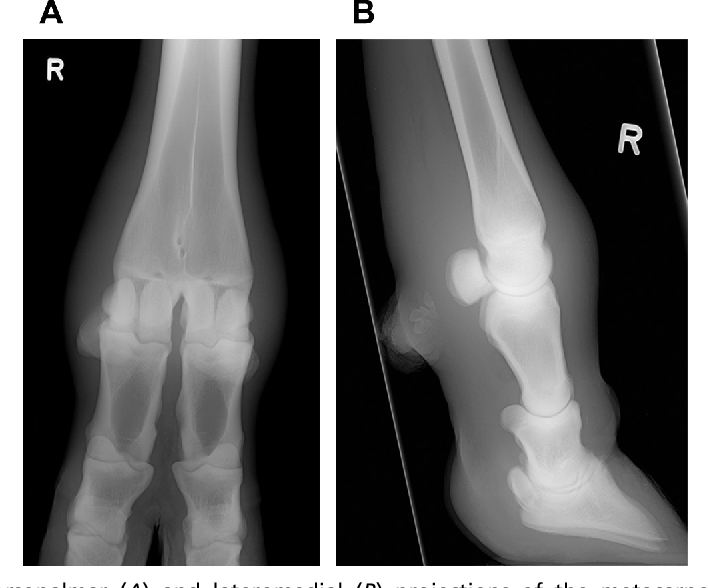
Рентгеновский снимок ноги коровы
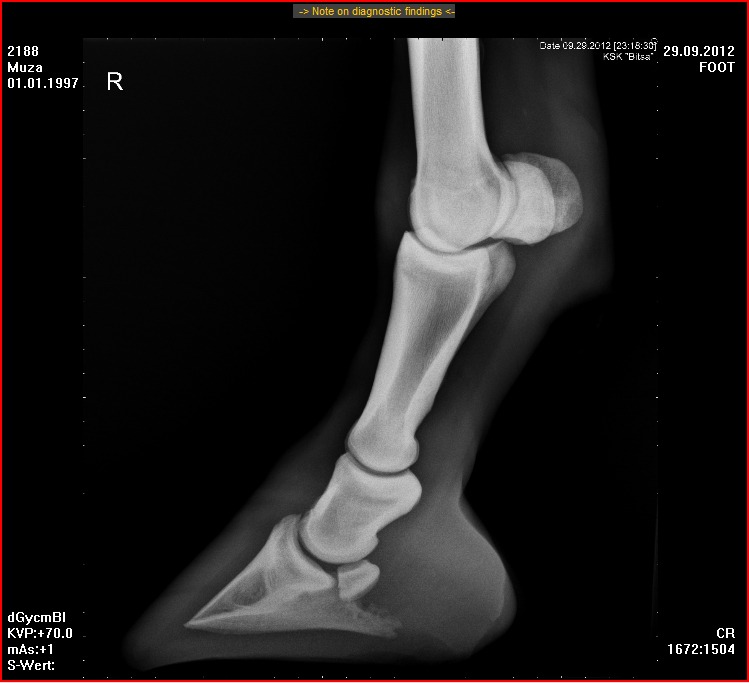
Рентгеновский снимок ноги лошади
Мне очень хотелось бы, чтобы Тен показал
своим прямым пальцем, в каком месте пальцы парнокопытного «согнуты»,
и где внутри конского копыта «полностью согнутая кисть».
Я также готов удовольствоваться видеосъёмкой анатомирования Теном варёной свиной
ножки из магазина с демонстрацией описанных в его книге особенностей.
От себя же скажу, что в конечностях копытных воплощена не «другая», а ровно
та же самая модель строения конечности, только кости дистальной (дальней от
тела) части конечностей частично срастаются между собой. Но пальцы выпрямлены,
и даже переразогнуты:
«Суставы пальцев фиксируются натяжением сухожилий сгибателей и разгибателей
пальца. Поверхностный и глубокий сгибатели пальцев, а также межкостный мускул
и связки сесамовидных костей в момент покоя животного, когда тяжесть тела давит
на верхний конец путовой кости вниз, натягиваются и удерживают суставы пальцев
в состоянии дорсальной флексии, т. е. переразгибания. Весь этот сухожильный
и связочный аппарат, расположенный ниже запястного сустава, называют «поддерживающим
аппаратом»» (источник:
«Ветеринарная хирургия, офтальмология и ортопедия» А. К. Кузнецов).
«Суставы пальцев тазовой конечности, как и суставы пальцев грудной конечности,
фиксируются натяжением трех сухожильных тяжей- поверхностного и глубокого сгибателей
пальцев и третьего межкостного мускула. Правда, на тазовой конечности поверхностный
сгибатель пальцев в отличие от такового грудной конечности не имеет точки опоры
на втором ее звене (голени), а прочно закрепляется по сторонам вершины пяточной
кости» (там же).
Стр. 207:
Все модели получились из одной универсальной формы. Из той, которой вы сейчас
переворачиваете страницы данной книги. Смотрите на нее как на самую древнюю
конечность млекопитающих, и забудьте бред симиалистов, будто ей всего полмиллиона
лет (иные уверяют, будто и того меньше).
Если уважаемый господин Тен имеет основание
полагать, что его конечности полностью идентичны конечностям древнейших млекопитающих
типа мегазостродона и морганукодона, вплоть до формы костей и суставов, то ему
следует обратиться с соответствующим заявлением в Академию наук и позволить
изучить его конечности во имя научного прогресса – возможно даже, что с посмертным
изъятием таковых в музейную коллекцию. Если же отличия всё же есть, то приведённый
абзац книги лучше изменить, чтобы он больше соответствовал истине.
Помнится, в книге Г. Уэллса «Остров доктора Моро» обезьяно-человек любил отмечать,
что у главного героя книги (человека) пять пальцев, как у него самого. Но тут
дело не только в пальцах, но и в степени их подвижности, о чём уже говорилось
при анализе предыдущих глав книги Тена. Пятипалость конечности приматов – примитивный
признак, но ногти вместо когтей, а у многих приматов ещё и противопоставляющийся
большой палец – это специализированные признаки, не характерные для предковых
млекопитающих.
Стр. 207:
Предки всех копытных – кондиляртры – имели пять лучей с коготками, которые у
некоторых животных стали сгибаться в «кулак» (кондилос). Благодаря этому и образовались
копыта разных форм, в зависимости от практики и точек сгибания. Кстати: «Среди
кондиляртр можно найти формы, весьма напоминающие приматов: их зубы тоже имеют
притупленные бугорки, более-менее прямоугольную форму коронок и прочие тонкости,
схожие с признаками нашего отряда. Неспроста статус некоторых животных долгое
время оставался или до сих пор остается под вопросом: кондиляртры это или приматы?»
(Дробышевский, 2018а, с. 281-282).
Книга “The Princeton Field Guide to Prehistoric
Mammals” (2017) под редакцией Дональда Р. Протеро любезно сообщает нам, что
в настоящее время кондиляртры не признаются самостоятельной группой (это было
установлено в 1988 году, задолго до выхода даже первого издания анализируемой
книги Тена) и фактически представляют собой «мусорный» (wastebasket) таксон,
куда ранее относили без разбора разнообразных архаичных копытных, которых нельзя
отнести к остальным отрядам копытных. Некоторые бывшие кондиляртры вроде фенакодуса
и гиопсодуса всё же нашли своё место среди непарнокопытных, тогда как хиракуса
сближают с парнокопытными.
Название происходит от греческих слов «кондилос» (кулак) и «артрос» (сустав),
и отражает форму таранной кости. Форма костей кисти и ступни говорит о том,
что конечности у них были пятипалыми с маленьким копытцем на каждом пальце,
и сами пальцы кондиляртров не были «согнуты в кулак». Поэтому утверждение Тена
о том, что «Благодаря этому и образовались копыта разных форм, в зависимости
от практики и точек сгибания» содержит в себе фактическую
ошибку. Разные формы копыт образуются вследствие редукции разного количества
пальцев и разрастания видоизменённых когтей на оставшихся пальцах. Да, «лучи»
господина Тена – это пальцы.
Стр. 207:
Копыто разогнуться уже не может, это движение в одном направлении, на «камбэк»
ставит категорический запрет биологический «закон Долло». У дельфинов под кожей
ласт удивительно длинные и изящные пять лучей.
А вот здесь уже становится интересно.
Раз за разом Тен показывает, что признаёт закон Долло, и использует его в своих
рассуждениях. Как говорил в мультсериале «Футурама» злой инопланетянин-телеведущий
Морбо, «это может оказаться полезным для моего вида!» И я тоже запомню это.
Мало ли – вдруг, да пригодится где-нибудь? Уже не раз пригождалось.
«Закон Долло», он же «закон необратимости эволюции», конечно, штука хорошая.
Но здесь он, увы, не к месту. Причина этого в том, что Тен изначально отталкивается
в своих рассуждениях от ошибочного мнения, что у копытных внутри копыта находится
согнутая в кулак кисть. Однако рентгеновские снимки ног копытных вкупе с данными
об эмбриогенезе этих животных полностью противоречат странному утверждению господина
Тена, которое он даже не потрудился обосновать.
Стр. 207-208:
Не все кондиляртры стали растительноядными копытными, некоторые стали хищниками.
В принципе от них могли получиться дельфины. Но нелепо называть этих древних
бескопытных животных копытными и говорить: «Дельфины произошли от копытных».
Их можно называть только предками копытных.
К сожалению (для Тена), кондиляртров
уже не считают предками китообразных. Одно время в качестве возможных предков
китов побывали родственные им мезонихиды – странные хищники с копытами, происходящие
от кондиляртров. Но потом была одна находка, о которой я скажу чуть ниже, и
пазл сложился.
И если Тен написал, что на ногах кондиляртров были «коготки», это ещё не означает,
что он был прав. В отличие от него, специалисты-палеонтологи считают образования
на пальцах кондиляртров копытами. Да, у них была пятипалая конечность с пятью
копытами.
Если же совсем придираться к словам, как это делает Тен, то говорить «дельфины
произошли от копытных» действительно не следует, но уже по другой причине. Семейство
дельфинов (Delphinidae) произошло от животных, которые уже были несомненными
полностью водными представителями отряда китообразных. Если мы говорим, что
китообразные произошли от копытных, то должны помнить, что эта фраза описывает
родственные отношения на уровне отрядов. Применительно к видам это верно только
для самого архаичнейшего кита. Как только появился он – первый кит, пусть даже
ещё четвероногий и сухопутный – все прочие (фактически, последующие) киты будут
происходить исключительно от китов, как бы эти киты ни выглядели.
Стр. 208:
Для включения в надотряд копытных существуют три критерия. Во-первых, маркерная
таранная кость – астрагал. Она хорошо известна даже неспециалистам, потому что
это та самая кость, которой играли в кости, в бабки, которую бросали при гадании.
При этом использовались бараньи или коровьи астрагалы. Во-вторых, наличие жевательных
зубов с площадками – моляров. В-третьих, копыта. Из всех признаков абсолютным
является только первый. Если будет найден крот, который ничем не отличается
от крота обыкновенного, живет под землей, питается червяками и имеет длинные
когти, но у него обнаружится миниатюрный астрагал, он будет записан в копытные.
Большое значение придается молярам, потому что абсолютное большинство копытных
– это не хищники, а растительноядные. Собственно говоря, необходимость в копытах
возникает, чтобы быстро убегать сломя голову, не рискуя сломать когти или ногти.
Недавно в Индии была найдена челюсть неизвестного ископаемого животного большой
древности. Оно тотчас было записано в копытные, потому что в челюсти были моляры.
И бинго! Потому что именно форма таранной
кости у Rodhocetus, одного из древнейших китов, ещё сохраняющих функциональные
задние конечности, позволила установить родство китов с парнокопытными. Об этом
я писал в своём старом обзоре «Первый кит, бегущий краем моря». Ещё стоит отметить,
что у любого крота есть астрагал. Да-да, у того самого, что роется в огороде,
отличаясь от «принципиального крота» Ф. Аквинского и А. Великого. Добро пожаловать
в стройные ряды копытных, милый пушистый землекоп? Кстати, отмечу, что у человека
эта кость тоже есть.
Идём дальше. Суём пальцы себе в рот, щупаем зубы, и с удивлением отмечаем, что
моляры у нас тоже есть! Внезапно! Видимо, неспроста в отношении интеллектуальной
деятельности издателей книги мне на ум приходят идиоматические выражения, представляющие
собой сравнения с некоторыми домашними животными – одно бородатое и рогатое,
второе похоже на лошадку с длинными ушами, львиным хвостом и голосом, как у
пингвина. А что? Право имею, поскольку у них, как у представителей вида «человек
разумный», тоже есть моляры – то есть, коренные зубы (после резцов, клыков и
предкоренных). Если следовать логическим конструкциям Тена, простое наличие
астрагала и моляров как таковых – это повод для их отнесения к копытным, причём
астрагал – это абсолютный признак. Значит, они – копытные, и я говорю чистую
правду. А на правду не обижаются – её принимают к сведению. Кстати, к Тену мои
слова тоже относятся. Если, конечно, у него от природы нет ни астрагала, ни
моляров, могу только пожалеть инвалида.
Если же оставить колкости в стороне, то видно, что тут господин Тен допускает
фактическую ошибку в рассуждениях – по незнанию (надеюсь), или намеренно (это
уже плохо). На самом деле имеет значение не простое наличие, а особенности строения
указанных элементов скелета.
Кроме того, в книге Тена 2019 года издания приводится откровенно устаревшая
информация. Молекулярная биология здорово перетряхнула родословное древо млекопитающих,
и копытные уже давно перестали быть надотрядом. Непарнокопытных объединили с
хищными и рукокрылыми в группу Pegasoferae (да, «пегасохищные»), парнокопытные
вместе с китообразными составляют группу Cetartiodactyla («китопарнокопытные»),
а хоботные, сирены и даманы вместе с тенреками и прыгунчиковыми стали Afrotheria
(«африканскими зверями»). То есть, бывшие «копытные» оказываются сборной группой.
Стр. 208:
С другой стороны, живут копытные, у которых копыт нет, например ламы и верблюды.
Копыта в качестве маркерного признака на последнем месте, потому что это даже
не экзоскелет, как, скажем, панцирь черепахи. Даже у черепах панцирь – второстепенный
маркерный признак: бывают черепахи кожистые.
Копыта просто появляются независимо друг
от друга в разных линиях млекопитающих, поэтому могут иметь разный облик при
одной функции. У халикотериев (непарнокопытных) вообще вместо копыт растут когтищи,
каким лев и тигр позавидуют – но по комплексу признаков они копытные.
Мне уже порядком надоело по ходу анализа книги Тена говорить себе «пардон»,
недоумённо протирать глаза и перечитывать текст, водя по нему пальцем. Но количество
странных биологических «открытий» сомнительного толка заставляет это делать
вновь и вновь. Вот и сейчас оказывается, что у черепахи (позвоночного, между
прочим) Тен находит… экзоскелет! Между прочим, это материал по зоологии из курса
средней школы: экзоскелет характерен для членистоногих. Напомню, что у черепахи
панцирь образован её рёбрами и позвоночником – внутренним скелетом. Если это
экзоскелет, то господин Тен вполне может происходить (персонально) хоть от дельфина,
хоть от крота, хоть от бешеного бандикута.
Укротив эмоции, возвращаемся к черепахам. Поскольку панцирь черепахи образован
в первую очередь внутренним скелетом (позвоночник и рёбра), именно форма грудной
клетки и расположение поясов конечностей относительно неё у кожистых и мягкотелых
черепах однозначно свидетельствуют о том, что они являются полноценными черепахами.
Наличие щитков – это лишь второстепенный признак строения панциря.
Да, не всегда бросающийся в глаза признак оказывается систематическим. Например,
не всякого жирафа узнаешь по шее.
Стр. 208:
Копыта образуются на базе ороговения и последующего перерождения клеток кожи.
Опять фактическая ошибка Тена, влияющая
на ход его рассуждений. Копыта фактически представляют собой видоизменённые
когти, о чём уже говорилось выше. То есть, они образовались не с нуля, а из
уже имеющихся когтей:
«Копыта – характерный признак различных копытных млекопитающих, у которых число
пальцев редуцировано и которые при передвижении опираются на кончики оставшихся
пальцев. Исходный коготь у них укоротился и расширился, превратившись в полуцилиндрическую
структуру, одевающую кончик пальца; на землю опирается дугообразный или V-образный
дистальный конец копыта, а подошвенная пластинка в виде подушечки располагается
внутри» (А. Ромер, Т. Парсонс «Анатомия позвоночных», М., «Мир», 1992, том 1,
стр. 169).
Стр. 208-209:
В связи с легкостью подобных образований эволюция предлагала их не только в
надотряде копытных. Палеонтологи находят собакоподобных с копытцами. Намек на
подобную адаптацию наблюдается у обезьян-брахиаторов, у которых лапы полностью
уже не разгибаются от ороговения кожи на внешней стороне. Обезьяны, слезшие
с деревьев на землю, скорее могли обзавестись копытами, чтобы быстрее ретироваться
от хищников, а не сводчатой стопой, которую они никак не могли заиметь. Это
гораздо более реальный вариант.
Эволюция млекопитающих, особенно в свете
молекулярных данных, показывает нам, что образование копыт легко происходит
у представителей разных групп зверей. Кроме перечисленных выше отрядов, они
появились, например, у архаичных травоядных диноцератов и пантодонтов.
И вновь господин Тен оперирует устаревшей информацией: копытные уже не представляют
собой надотряд, имеющий общее происхождение. Возможно, под «собакоподобными
с копытцами» Тен подразумевает мезонихид, но тут я не могу ничего утверждать,
хотя внешне именно они подходят под это описание.
Далее Тен вступает на крайне зыбкую почву. Сложно сказать, какой вид/род обезьян
подразумевает Тен, называя «обезьянами-брахиаторами»: конкретное название этих
приматов отсутствует, а домысливать за другого человека – дело неблагодарное.
Скажу лишь, что ороговение кожи, мешающее лапам разгибаться, неизвестно ни у
каких обезьян.
Утверждение господина Тена о том, что «обезьяны, слезшие с деревьев на
землю, скорее могли обзавестись копытами, чтобы быстрее ретироваться от хищников»,
я полностью оставляю на его совести. Скажу лишь, что копыто – это приспособление
для сокращения площади трения об землю при беге. Оно не возникает из воздуха,
копытами обзаводятся пальцеходящие бегающие животные. Чтобы у обезьяны появилась
потребность в копыте, она неизбежно должна вначале быть наземной стопоходящей,
затем стать бегающей пальцеходящей, и лишь после этого ногти
превратятся в какое-то подобие копыт. Такой путь повторяли все предки копытных
– как выяснилось, независимо друг от друга.
Стр. 209:
В конце XX в. в Пакистане, где некогда разливался Тэтис, американец Ф. Джинджериш
нашел останки собакоподобного существа, которое и было, как сейчас принято,
переходной формой к древним дельфинидам.
Даже не к дельфинидам, а ко всем прочим китообразным вообще. Этот зверь в равной степени был предком клюворылов, кашалотов, полосатиков, гладких китов и прочих современных китообразных, а также всех древних китов. Поэтому считать его исключительно «переходной формой к древним дельфинидам» логически неверно, даже если господин Тен питает некое особое отношение к этим существам.
Стр. 209:
Кстати, собакоподобных сближают с лемурами, потому что у них есть общая черта,
резко отделяющая от обезьян: у них влажные носы, а у обезьян сухие.
Странное утверждение. Кого имеет в виду
Тен под названием «собакоподобные»? Хочу поинтересоваться: означает ли отдалённое
сходство некоторого животного с собакой то, что его надо относить к лемурам
сугубо на основании внешнего вида? А если у нас есть собака, на 100% собакоподобная
– будет ли она лемуром, потому что в высшей степени собакоподобная (она же собака!),
и вдобавок с мокрым носом? Кстати, вспомнилось вот, что павианов называют собакоголовыми
обезьянами – стало быть, они тоже в чём-то собакоподобны. Если применять логику
Тена, то будут ли павианы мокроносыми (потому что они «собакоподобные»), или
сухоносыми (потому что обезьяны)?
Вот так – одна фраза, а сколько глупостей наговорил. Или не глупостей? На мой
взгляд, здесь имеет место ложная логическая связь – на основании отдельно взятого
признака поверхностного сходства объявляется родство, игнорируя огромные массивы
свидетельств против этого вывода. Ну, или Тен просто полез туда, где «ни в зуб
ногой», как говорится.
Стр. 209:
Основанием для того, чтобы видеть в пакицете предка дельфинов, стала «булла»
– слуховая кость, позволяющая чутко улавливать дрожание сред, более плотных,
чем воздух. Само название, слишком смелое, на мой взгляд, означает «пакистанский
кит». У него были крохотные копытца на каждом луче.
На каждом пальце! И
я рад, что Тен этой фразой признаёт, что у пакицета были копытца.
Строение буллы в данном случае – это именно «эксклюзив» китообразных, проходящий
«красной нитью» через всю историю отряда, от архаичных до современных представителей
китообразных. Помимо этого пакицетус был признан китообразным на основании строения
зубов, сходных с зубами более поздних, и уже несомненных китов.
Стр. 209:
Предки пакицета неизвестны. Вначале его возводили к мезонихидам – хищникам,
имевшим маленькие копытца на каждом луче и огромные клыки, почти как у саблезубого
тигра. Но сейчас эта дикая версия не имеет сторонников.
Почему-то Тен сказал, что было «вначале»,
но явно не хочет говорить о том, что было «потом». Потому что потом установили,
что предков пакицетуса нужно искать среди архаичных парнокопытных семейства
раоеллид (Raoellidae), и была найдена примитивная форма из этого семейства –
индохиус (Indohyus), напоминающий обликом современного водяного оленька, но
с длинным хвостом. Он не предок пакицетуса (жил позже него), но, судя по анатомии,
имел с ним общего предка. В частности, у него очень сходное с пакицетусом строение
ушной области.
Напомню, что китообразных отделили от мезонихид и сблизили с парнокопытными
именно особенности строения астрагала – таранной кости. Так что это решение
возникло не с бухты-барахты, а было вполне обоснованным.
Вновь и вновь обращает на себя привычка Тена обзывать пальцы «лучами». Причины
этой привычки лично мне неизвестны.
Наконец, среди мезонихид не было саблезубых форм – их клыки увеличены по сравнению
с прочими зубами не больше, чем у современных хищных млекопитающих.
Стр. 209:
Морские млекопитающие, происходящие от копытных, имеют короткие, толстые, тупые,
массивные ласты, не позволяющие эффективно маневрировать и развивать большую
скорость. Поэтому они не уходят от суши далеко. В настоящее время это сирены
– ламантины, дюгони. Рудименты копыт у них сохранились и хорошо видны. У их
предков были, как у пакицета, маленькие копытца на каждом луче. Разогнуться
они не смогли. У стеллеровых коров ласты оканчивались рудиментарными копытами,
похожими на лошадиные, которые тоже за миллионы лет эволюции так и не смогли
разогнуться.
Давайте говорить более корректно. Начать
надо так: «морские млекопитающие, происходящие от афротериев…» и далее по тексту.
Тогда у нас будет совершеннейшая правда. Сирены, о которых говорит Тен – это
другая линия морских животных, ведущая свой особый образ жизни и не имеющая
отношения к предкам китов. Стоит отметить, что они травоядны, а крупные растения
в океане растут лишь на хорошо освещённых мелководьях на небольшой глубине.
Поэтому сирены не стремятся уйти в океан не потому, что у них тупые ласты, а
потому что им там тупо жрать нечего. Нет им нужды покидать прибрежные мелководья,
где растёт их еда, потому и не приспосабливались сирены к пелагическому образу
жизни. А поскольку растения не умеют быстро плавать (вообще не умеют плавать)
и обладают низкой питательной ценностью по сравнению с мясом, эволюционировать
в сторону энергозатратного скоростного плавания также не имело смысла – гоняться
не за кем, и лишнюю энергию тратить не нужно.
Смущает то, что киты плотоядны, а копытные травоядны? Очень советую почитать
о гастрономических пристрастиях водяного оленька – будете (не)приятно удивлены.
Если предположить такие же наклонности у индохиуса, переход китообразных к поеданию
пищи исключительно животного происхождения уже не вызовет удивления и внутреннего
неприятия. Да и мезонихиды, хоть и разжалованы из предков китов, показывают
своим примером, что копытное вполне может эволюционировать в плотоядное существо.
Продолжим, однако, наш разговор. Вот скелет ласта ламантина:
Пальцы на нём видны отчётливо, все фаланги заметны, и я хотел бы увидеть столь же отчётливо, где же находятся те самые «сгибы», на которых, по мнению Тена, у этих зверей образовались копытца.
Стр. 210:
С линии копытных происходили схождения в водную среду, но не чаще, чем с других
линий, эволюция всегда конвергентна. Современное разнообразие амфибий-млекопитающих
огромно, а происхождение их различно. Это и грызуны (бобры, ондатры), и куньи
(выдры, каланы и т.д.), и дюгони, ламантины, которые на самом деле принадлежат
к копытным, и непонятно от кого произошедшие моржи.
Происхождение моржей как раз вполне понятно – это ластоногие, по сути своей водные представители отряда хищных. Причём предки настоящих тюленей ближе к куньим, а моржей и ушастых тюленей – к медвежьим.
Стр. 210:
Факт, что пакицет был переходной амфибийной формой, не доказывает, что он был
предком дельфинид, учитывая, сколько амфибий-млекопитающих вышло из других линий
в период разлива Тэтиса. Каждый отряд животных в период разлива Тэтиса давал
амфибий. Амфибийных форм было больше, чем наземных.
Я искренне надеюсь, что утверждение о
количестве амфибийных форм млекопитающих в Тетисе было сформулировано на основе
исследований, и потому хотел бы взглянуть на научную работу, в которой это сказано.
Раз уж «каждый отряд животных в период разлива Тэтиса давал амфибий»,
то я бы хотел взглянуть на подлинные ископаемые остатки полуводных рукокрылых,
многобугорчатых, сумчатых и приматов, а также почитать статьи с их описанием.
Ещё интересны статьи, где доказывается, что видов полуводных млекопитающих было
больше, чем наземных – с подсчётами, разумеется. Я ведь не слишком многого прошу?
Тену нужно всего лишь подтвердить свои слова, просто указав первоисточники,
которыми он (надеюсь) пользовался.
А вот утверждение «факт, что пакицет был переходной амфибийной формой,
не доказывает, что он был предком дельфинид» нуждается
в доказательствах. Ведь пакицетуса отнесли к китообразным именно на основании
того, что у него был китовый «эксклюзив» (воспользуюсь термином Тена, чтобы
и ему было понятно). Даже два. Сам Тен уже упоминал один из них – буллу. Второй
общий признак – звукопроводящий канал в нижней челюсти, который характерен для
всех современных китов.
Если Тен отделяет его от предков дельфинов, то получается, что он игнорирует
именно наличие «эксклюзивов» при установлении родства и тасует объективные научные
данные ещё гнуснее, чем осуждаемые им «симиалисты» в отношении обезьян и человека.
Так важны ли эти самые «эксклюзивы» в его рассуждениях? Если нет, зачем пугать
читателя списками этих признаков?
Стр. 210:
Когда открыли пакицета, можно было, действуя в рамках научной корректности,
сделать два осторожных вывода. Первый: это нормально адаптированный водно-земной
организм, скорее всего универсальный по питанию, которому в принципе незачем
было уходить дальше в воду и превращаться в китов.
Вывод о том, что пакицетус был предком
китообразных, сделан уже пост-фактум, на основании всего массива находок ископаемых
китообразных, а также общих признаков этих китообразных, которые и были обнаружены
у пакицетуса, позволив определить его как архаичное китообразное. Просто огромное
количество ископаемых находок китообразных – это свидетельство того, что пакицетус
по факту реализовал свой потенциал. Время было такое – в океане конкуренции
нет, а рыбы полно. Что бы не воспользоваться?
А вот как Тен умеет определять, какому животному «незачем», а какому «есть,
зачем» делать какой-то эволюционный шаг – это загадка.
Стр. 210:
Второй вывод: пакицет мог быть предком каких-то морских млекопитающих, например
морских леопардов, или вымерших, которых было множество. Но его сразу же, прямо
в день открытия, записали в предки дельфинов. Джинджериш взахлеб начал давать
интервью.
Господин Тен, ну какого чёрта? Доколе
вы будете забрасывать читателя сиюминутно сляпанными «на коленке» лжеаргументами?
Думаете, вас читают только скучающие домохозяйки, готовые проглотить любую вашу
чушь? Морские леопарды – это представители настоящих тюленей, и у них был свой
предок. Молекулярные и анатомические данные говорят, что он был из числа хищников,
близких к современным куньим. Кстати, он найден – это пуижила (Puijila darwini).
Далее, пакицетус известен учёным с 1981 года. Был обнаружен только неполный
череп, костей посткраниального скелета найдено не было – поэтому первая реконструкция
была неправильной, изображавшей пакицетуса как массивное тюленеобразное существо.
И родство животного с более поздними китами определили по строению черепа и
зубов – других частей скелета на тот момент в распоряжении учёных не было.
Пакицетус, как я уже сказал, был предком не только дельфинов, но и всех остальных
китообразных. Ставить особняком его родство именно с дельфинами – логически
некорректно. Однако, как я понимаю, Тен допускает возможность считать пакицеуса
предком кашалотов и, допустим, китов-полосатиков?
Стр. 210:
Сами по себе ископаемые амфибии-млекопитающие не являются сенсацией, их тьмы.
Но очень легко сделать сенсацию на предках дельфинов. Почему-то именно генеалогия
дельфинов является сенсационной темой наряду с антропологией. Когда находят
предков любого другого животного, это проходит тихо, по специальным изданиям,
и общественность особо не волнует. А как только появляется что-то новое о предках
дельфина или человека — поднимается звучный ор. Подключаются глянец, популярные
издания, блогеры... Загадочное явление само по себе. Не по Юнгу ли? Не архетип
ли – родовая память об истинных предках?
Попробую объяснить, тем более что когда-то
я изучал для себя этот вопрос. Дело в том, что предки китообразных долгое время
были неизвестны, и вопрос об их происхождении вкупе со специализированной и
уклоняющейся анатомией современных китов долгое время был предметом дискуссий.
Креационисты активно стебались над предположением Дарвина о медведе, плавающем
с открытой пастью, и приводили китов как пример несомненно сотворённой богом
группы существ, не имеющей переходных форм от наземных животных. Учёные спорили
о вероятных предках китов и о том, являются ли киты монофилетической, или же
полифилетической группой. Именно поэтому находка полного скелета пакицетуса
вызвала такой интерес – одним она сняла кучу вопросов и послужила превосходным
примером исполнения научных предсказаний, а другим даже не утёрла, а со всей
дури расквасила нос, превратив его в свиной пятачок. Потому и велик интерес
к находке пакицетуса.
Что же касается более узкого вопроса происхождения именно семейства дельфинов
внутри отряда китообразных, то вряд ли дяди во дворе за бутылкой пива и домино
когда-нибудь спорили, от какого рода семейства кентриодонтид могли произойти
первые истинные дельфины. Этот вопрос вряд ли волнует даже специалистов по трилобитам
или ископаемым рукокрылым, и обсуждается, скорее всего, исключительно в среде
специалистов по ископаемым китообразным.
Соответственно, интерес к предкам человека – это следствие желания понять, где
находятся наши корни, какова наша природа. Логично и объяснимо.
Стр. 210-211:
Так теперь, если каждый хочет стать автором сенсации, мы всех ископаемых амфибийных
гиен, копытных, собак и т.д. будем записывать в предки дельфинов? Джинджериш
сделал себе имя. Будем ждать очередной американской сенсации с какой-нибудь
другой амфибийной формой. Вон сколько имен зазвездилось в связи с ископаемыми
гомининами. Кости в африканских ущельях наружу торчат из обрывов. Даже антропологическое
сафари процветает: плати, выгребай, увози домой! Богачи получают удовольствие,
думая, что держат под стеклом кости предков человека. Ричард Лики, организовавший
сафари, зарабатывает: он знает истинную цену этим костям.
Дурацкое предположение, господин Тен.
Амфибийных гиен неизвестно, среди псовых тоже нет специализированных полуводных
форм. Более поздних полуводных копытных (например, носорогов аминодонтов или
целый отряд десмостилюсов) никто в предки ваших драгоценных дельфинов не записывает.
Успокойтесь, примите стаканчик обезжиренного кефира.
В патетическом описании поиска ископаемых остатков гоминид я явно ощущаю выплеснутую
в ругательные слова чёрную зависть, снедающую Тена изнутри. Просто явственно
видны слизистые и бородавчатые лапы той вселенской жабы, которая душит его –
ведь все находки гоминид лишь льют воду на мельницу «злых симиалистов», и ни
одной косточки в качестве прямого подтверждения его собственных выкладок так
и не обнаружено. Вот и приходится примазываться к чужим находкам, и пытаться
выворачивать их интерпретацию в свою пользу.
Я не исключаю, что какие-то находки ископаемых приматов осели в частных коллекциях,
как это было некоторое время с образцом знаменитой «Иды». Но вот придумка про
«палеонтологическое сафари» повеселила. Это где же находится то самое место,
сказочно богатое останками гоминин? Почему о нём знает дилетант Тен, но не знают
профессиональные антропологи? Это ж какую экспедицию можно забабахать! Десять
лет потом изучать и описывать результаты лишь одного полевого сезона!
Ископаемые кости, хоть гоминида, хоть антилопы или суслика, крайне хрупкие,
и вряд ли скучающий богач станет канителиться с их препарированием в поле или
в передвижной лаборатории – для этого надо иметь известный навык в раскопках
и фиксации образцов. Особенно прикольно, конечно, проводить раскопки, когда
в окрестностях шарятся банды голых негров или бородатых арабов с автоматами
в руке и именем одного из арабских божеств в сердце. Сафари, мать вашу, с преферансом
и падшими женщинами!
Стр. 211:
Наперсточники тоже не теряются. Поехал, прямо на поверхности нашел кривые кости,
поставил в начало цепи к человеку – и вот вам великий антрополог! Пока будут
разбираться, успеешь прозвучать. Хотите пример? Джохансон нашел безобразные
кости прямо на поверхности в первый вечер по приезде в экспедицию, даже лагерь
не успели развернуть. Пока другие ставили палатку, он якобы отошел «прогуляться»
и возвратился с кучкой косточек. Тут же был устроен праздник, по песне «Битлз»
находке дали имя Люси, и уже на другой день в США вышла статья друга Джохансона
журналиста Иди о том, что найдена «праматерь рода человеческого». Даже до камеральных
исследований, не говоря о лабораторных, как будто была подготовлена заранее.
Ага, один – напёрсточник, другой – трюкач,
третий – подтасовщик… И только один Тен – весь в белом, как в том анекдоте.
Только вот что-то я пока этого не наблюдаю, читая его произведение.
Теперь по существу. Приматы – большей частью обитатели лесов, где условия для
захоронения их ископаемых остатков очень плохие – кислая почва, обилие животных-падальщиков,
невозможность быстрого захоронения. Поэтому в большинстве своём ископаемые обезьяны
известны лишь по ископаемым зубам. От того же гигантопитека известны лишь зубы
и обломки челюстей, и ни одного фрагмента посткраниального скелета, хотя обезьянка
должна быть немаленькая. Поэтому находка скелета гоминида, у которого сохранилось
примерно 40% скелета – это само по себе уже незаурядное событие. И это была
первая находка австралопитеков в Восточной Африке – тоже масштабное событие.
Что же касается точности описания Теном обстоятельств находки скелета австралопитека,
получившего прозвище «Люси», то я просто процитирую книгу Д. Джохансона и М.
Иди «Люси. Истоки рода человеческого» (М., «Мир», 1984, текст взят отсюда;
указания страниц сделаны по бумажному экземпляру), приводя цитаты в порядке,
соответствующем их расположению в тексте и выделяя некоторые фрагменты текста
для ясности:
«Тридцатого ноября 1974 года я проснулся, как обычно просыпаюсь в экспедиции,
— на рассвете. Я находился в Эфиопии, в палаточном лагере на берегу илистой
речушки Аваш, в местности Хадар примерно в сотне миль к северу от Аддис-Абебы.
Здесь я работал уже несколько недель, будучи одним из руководителей
группы ученых, занимавшихся поисками ископаемых остатков» (стр. 13).
«Пошел всего лишь третий год полевых исследований в Хадаре,
а на моем счету было уже несколько находок. Я знаю, что мне везет, и не пытаюсь
этого скрывать. Вот почему я записал в дневнике: «Чувствую себя хорошо». Проснувшись
в то утро, я ощутил, что мне необходимо испытать свое счастье – что это один
из тех дней, когда может произойти нечто исключительное.
Но в первые утренние часы ничего не произошло. Грей и я сели в один из четырех
экспедиционных лендроверов и, трясясь на ухабах, медленно направились к участку
162. Это был один из нескольких сотен районов, которые мы изучали, чтобы нанести
на генеральную карту Хадара все полученные геологические и палеонтологические
данные. Хотя до места, к которому мы ехали, было всего лишь около четырех
миль пути, мы потратили на дорогу чуть ли не полчаса из-за неровностей
и ухабов. Когда мы прибыли туда, солнце уже начало припекать» (стр. 14).
«Сумасшедшая, невозможная мысль мелькнула в моей голове: а что если сложить
их вместе? Быть может, это части одного, чрезвычайно примитивного скелета? Такого
еще никогда и нигде не находили.
– Посмотри-ка сюда, – сказал Грей. – Ребра.
Неужели все это принадлежит одному индивиду?
– Я не могу поверить, – промолвил я. – Совершенно не могу поверить.
– Боже мой, почему бы нет? – закричал Грей. – Вот он. Здесь! – Его голос перешел
в радостный вопль» (стр. 16).
«После обеда все, кто был в лагере, отправились к оврагу, разделились по отдельным
участкам и стали готовиться к большой работе, которая в целом заняла
три недели. Когда она была закончена, у нас имелось несколько сотен
костей (из них многие – фрагменты), составлявших в совокупности около сорока
процентов скелета одной особи. Наше с Томом предчувствие не обмануло нас. Среди
костей не было дубликатов» (стр. 16).
Картина несколько отличается от той безобразной истории, которую состряпал для
нас Тен, верно? Вот и я говорю: в соответствии с его версией, все в дерьме,
и только он один – весь в белом. Однако мы видим, каково происхождение этого
«дерьма»: это довольно примитивная попытка Тена очернить специалистов. Но правда
– это такая упрямая штука, что она всегда вылезает наружу. Достаточно лишь обратиться
к первоисточникам.
По поводу информации о статье М. Иди пришлось спросить знающих людей. Сотрудник
библиотечного отдела Американского Музея естествознания Май Рейтмайер уточнила,
что статья Иди под названием «Люси возрастом три миллиона лет» (“Three-Million-Year-Old
Lucy”) была опубликована в ежегоднике “1976 Nature / Science Annual”. Надо же,
какое (не)совпадение в датах! Вполне ожидаемое, кстати. Вновь истории, которые
нам рассказывает Тен, оказываются «шиты белыми нитками».
Стр. 211:
Возвращающегося в США Джохансона при пересадке в Париже ждала рукоплещущая толпа.
Интернета не было, а люди откуда-то узнали, что великий антрополог будет делать
пересадку! Луис Лики, честный человек, перестал общаться с Джохансоном, вычеркнул
его из числа ученых, а Люси объявил подтасовкой.
Ей-богу, снова вижу одни эмоции и клокочущую
зависть. Интернета не было, но были столь же оперативные в плане передачи новостей
телевидение, телефон, телеграф и радио. Что, уже успели отвыкнуть от них в век
Интернета? Да, и вот, что пишет сам Джохансон в той же книге (ссылка,
стр. 138 бумажного экземпляра книги):
«На промежуточной посадке в Париже я вновь прошел таможню. Служащий потребовал
показать, что находится в маленьких пакетиках, лежащих в моем чемодане. Я объяснил,
что это ископаемые остатки из Эфиопии. «А, это, очевидно, Люси?» — спросил таможенник.
Он интересовался антропологией и прочел о Люси в газетах. Собралась порядочная
толпа, которая наблюдала, как кости Люси одна за другой выкладывались на таможенную
стойку. Я впервые почувствовал огромный интерес, который пробуждала Люси, где
бы она ни появлялась. И я понял, что превратился из никому не известного начинающего
антрополога в подающего надежды молодого исследователя, находки которого уже
могут соперничать даже с открытиями признанной палеоантропологической суперзвезды
– Ричарда Лики».
И напрашивается вопрос: а не из газет ли люди узнали о находке Люси? Капитан
Очевидность подсказывает мне, что, да, именно из газет.
История с отношением Луиса Лики к систематическому положению «Люси» на родословном
древе человека весьма занятна. Особенную пикантность ей придаёт один момент:
«Люси» была открыта в 1974 году, а Луис Лики умер в 1972-м.
Стр. 211-212:
Лет десять длились вокруг этой Люси танцы с бубнами, а сейчас она у большинства
антропологических шаманов в отставке, сейчас вокруг Арди камлают, теперь уже
«Ardipithecus ramidus – прекрасный кандидат на роль переходного звена» (Марков
А, 2011, с. 72). Без боязни, что уличат в очередной подтасовке. Чего бояться,
если контрольной линии для сверки нет? Была бы – пришлось бы оглядываться. Именно
потому, что нет ни одной «стройной линии», и не боятся позора разоблачения проходимцы
эти. На уродах классификация невозможна, во всяком случае, одна. Множество
– пожалуйста. А это означает, что ни одной, и это тоже объединяет генезис человека
и дельфина, для которого тоже нашли слишком много линий, чтобы признать хоть
одну.
Представим себе, что наш господин Тен
попал под следствие – как говорится, «от тюрьмы, да от сумы не зарекайся». И
обвинили его в чём-то гадком, бросающем тень на его репутацию. Следователи собрали
улики по делу, и на основании этих улик выходило, что Тен виновен. Он, конечно,
пытался говорить, что не виновен, но так все говорят на его месте. На основании
имевшейся на тот момент совокупности улик судья вынес приговор, Тену надели
наручники и отправили его по этапу куда-нибудь на солнечную Колыму. И вот господин
Тен наслаждается красотами колымской тайги, строго соблюдает режим дня, много
работает на свежем воздухе, а по вечерам тесно общается с интересными и духовно
развитыми личностями.
Внезапно следователи законным путём получают ранее неизвестную информацию, которая
проливает свет на один неясный момент. Так, ничего особенного – мелочёвка, которая
просто уточняет, что Тен находился в каком-то месте раньше (или позже) некоторого
промежутка времени, и потому просто физически не мог сделать то, в чём его обвиняют,
по причине своего отсутствия именно в момент преступления именно на месте преступления.
А судья на эту информацию в руках повертел, ленивым взглядом посмотрел, да и
сказал: «А зачем нам эту новую информацию учитывать? Приговор уже вынесен, нехрен
тут её в дело вшивать, решения менять. Скажут ещё в Следственном Комитете: подтасовками,
мол, всякими занимаетесь тут. Приговор вынесен – так что пусть трубит от звонка
до звонка!»
Теперь, собственно, сам вопрос. Будет ли в такой ситуации господин Тен сидеть
и дальше, считая годовые кольца на соснах при помощи топора, или сделает всё,
чтобы документ приобщили к делу, провели доследование по делу, его самого оправдали,
а судью за такое отношение к важной информации наказали?
Я специально утрирую ситуацию, переходя на личности, но принципиально она та
же самая: в случае с ардипитеком у учёных появляется новая, ранее неизвестная
информация, которая заставляет переоценивать массив уже имеющихся знаний. Поэтому
она не является подтасовкой – никакой фальсификации данных не происходит.
А сейчас вновь обратимся к словарям, чтобы определить, что же такое «подтасовка».
Вот определение слова «подтасовать» из словаря
Ушакова:
«1. Тасуя, подобрать в нужном для чего-нибудь (преим. для обмана), неслучайном
порядке. Подтасовать колоду карт.
2. перен. Исказить, намеренно приняв во внимание только выгодное для какого-нибудь
вывода и отбросив противоречащее. Подтасовать факты».
Старый «Большой
толково-фразеологический словарь Михельсона» проявляет редкостное единодушие:
«Подтасовка (иноск.) перемѣшиваніе фактовъ, для ложныхъ
выводовъ (намекъ на подтасовку картъ)».
Итак, для подтасовки нужно:
1) подобрать выгодные для себя факты;
2) отбросить невыгодные.
На протяжении примерно двухсот страниц книги Тена я убеждаюсь, что именно она
полностью подходит под указанный рецепт. Тен лишь делает видимость, что критикует
современные книги по антропологии: он цепляется к каким-то второстепенным вещам,
старательно обходя важные. Он избегает обсуждать важные для систематики признаки
анатомии, совершенно не обращается к хорошо проработанным в настоящее время
данным по биохимии и генетике, но зато выпячивает какие-то малозначительные
признаки вроде количества зубов и цвета склер. Много раз я замечал, что он избегает
сравнения человека и обезьян по каким-то обсуждаемым признакам – например, по
наличию и работе потовых желёз. Или же вовсе имеют место неоднократные замалчивания
и искажения «неудобных» фактов. Даже пресловутый закон Долло (закон необратимости
эволюции) Тен применяет лишь тогда, когда ему выгодно, «забывая» применить в
рассуждениях о более важных для систематики вещах.
Достаточно посмотреть книги «проклятых симиалистов», чтобы убедиться, что они
как раз ничего не скрывают. Излагая некие данные исследований, они делают ссылку
на эти исследования, и в конце книги приводится список использованных работ
– на несколько десятков страниц мелким шрифтом. Всяк желающий может отыскать
эти работы и посмотреть, на основании чего специалисты сделали те или иные выводы.
Зато в своей книге Тен не обременяет себя ссылками на источники, из которых
берёт данные для утверждений в свою пользу. Я уже отмечал это много раз по ходу
пьесы.
Но вернёмся к обсуждению биологических данных. Почему раньше боготворили Люси,
а теперь Арди? Тоже вполне объяснимо: потому что в ряду от «стандартных обезьян»
с деревьев до человека, шагающего по земле, Арди занимает место перед Люси.
Никто не сбрасывал со счетов образец
Относительно «множества линий», объединяющих генезис человека и дельфина, хотелось
бы узнать поподробнее. А то мы уже перевалили за середину книги, но пока Тен
не соизволил привести полное научное название (род + вид) хотя бы одного представителя
линии доисторических млекопитающих, связывающей происхождение человека с «дельфинидами».
Стр. 212:
Потрясающее сходство эмбрионов дельфинов и людей, согласно биогенетическому
закону, говорит об эволюционном родстве. И тут встает вопрос об общей исходной
форме, т.к. эмбрионы повторяют предковую взрослую форму. Кто похожий с круглой
головой и руками-ногами жил на Земле до начала эволюции дельфинид? Я сделал
предположение о лемуроподобных предках. Кстати сказать, я за него не держусь,
т.к. этот вопрос к антропогенезу прямого отношения не имеет.
И вновь Тен вступает на зыбкую почву,
пытаясь использовать в своих целях эмпирические законы, сформулированные естествоиспытателями
прошлого. С биогенетическим законом не всё так просто: он признаётся современной
биологией крайне ограниченно, поскольку в чистом виде никогда не выполняется.
Стоит напомнить формулировку этого закона, чтобы однозначно понимать, о чём
идёт речь.
Формулировка из «Википедии»
со ссылкой на «Большую Советскую Энциклопедию»:
«Каждое живое существо в своём индивидуальном развитии (онтогенезе) повторяет,
в известной степени, формы, пройденные его предками или его видом (филогенез)».
В этой же статье приводятся формулировки близких закономерностей, выявленных
различными учёными:
«Эмбрионы последовательно переходят в своём развитии от общих признаков типа
ко все более специальным признакам. Позднее всего развиваются признаки, указывающие
на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие
завершается появлением характерных особенностей данной особи» (К. М. Бэр, 1828
г.)
«Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше
более или менее затененный образ общего прародителя, во взрослом или личиночном
его состоянии, всех членов одного и того же большого класса» (Ч. Дарвин, 1859
г.)
«Историческое развитие вида будет отражаться в истории его индивидуального развития»
(Ф. Мюллер, 1864 г.)
«Онтогенез есть рекапитуляция филогенеза» (Э. Геккель, 1866 г.)
Самую прямую (и больше всех прочих критикуемую) формулировку биогенетического
закона выдвинул Эрнст Геккель. Прочие авторы высказываются более осторожно,
отмечая проявление этого закона в эмбриогенезе лишь в самых общих чертах, но
не буквально, как подразумевает геккелевская трактовка.
«Биологический энциклопедический словарь» (М., «Советская Энциклопедия», 1989,
стр. 60) в статье «Биогенетический закон» сообщает ещё один немаловажный факт:
«Б. з. – лишь частный случай соотношений онто- и филогенеза и соблюдается только
при эволюции онтогенеза данного вида путём надставки его последних стадий –
анаболии». То есть, у нас в руках далеко не волшебная палочка-выручалочка. На
разных стадиях онтогенеза возникают признаки, связанные с приспособлением к
новым условиям существования – достаточно вспомнить, например, причудливый облик
личинок морских рыб и ракообразных, а также гусеницу в сравнении с бабочкой.
Геккель знал о подобных признаках, нарушающих рекапитуляцию, и называл их ценогенезами.
А далее начинается кое-что интересное:
«Более поздние [чем геккелевские – П. В.] исследования онтогенеза организмов
мн. видов показали, что ценогенезы чрезвычайно обильны в любом конкретном онтогенезе
и рекапитуляция целых филогенетич. стадий невозможна. Рекапитулируют лишь отд.
признаки и процессы» (там же, стр. 60).
Это описание показывает, насколько ограниченное применение у биогенетического
закона.
Тен допускает ещё одну фактическую ошибку, когда говорит, что «эмбрионы
повторяют предковую взрослую форму». Сходство проявляется
не между эмбрионом и взрослой особью, а между эмбрионами предков и потомков.
У громадного количества млекопитающих даже молодые детёныши весьма короткоморды
по сравнению с взрослыми особями. И эмбрион дельфина на стадии «круглой головы»
похож на эмбриона человека не больше, чем на эмбрион свиньи или собаки. Единственное,
что можно сказать, исходя из особенностей эмбриона дельфина на данной стадии
– то, что его предки были четвероногими наземными млекопитающими, не более того.
Стр. 212:
Почему все-таки лемуры или догопяты, но не кондиляртры?
О том, что даже следы былой копытности у дельфинов отсутствуют, мы уже говорили.
Моляров у них тоже нет. И нет астрагалов. А вот у кондиляртр астрагалы есть,
поэтому их и считают предками всех копытных. Но предками дельфинов по данному
критерию они скорее всего быть не могли.
Так-так-так, помедленнее, я записываю…
«Былая копытность»? О, значит, она всё-таки была? Или это мне просто очень хочется,
чтобы Тен это признал?
Если у ныне вымерших птиц моа отсутствовали даже рудименты крыльев – это ещё
не означает, что крыльев у их предков вообще никогда не было. Если у змей сейчас
нет ног – это ещё не значит, что их никогда не было у их предков. Почему же
мы не можем утверждать, что у предков дельфинов (а равно и всех остальных китообразных)
не было копыт, если эти предки анатомически были родственны древним копытным
и имели концевые фаланги пальцев характерного для копытных строения? Плюс данные
биохимии гаденько так, исподтишка, но весьма уверенно указывают на родство китов
и парнокопытных.
И снова Тен демонстрирует чудеса познаний в области зоологии. По поводу лемуров
и долгопятов мы ещё поговорим, а вот почему не кондиляртры в предках у китов?
Да потому что это искусственная группа, фактически упразднённая современными
исследованиями.
Идём дальше. Моляры у дельфинов есть. Ибо это просто коренные зубы, независимо
от того, режущие они (как у кошек), или давящие (как у коровы). Никуда они не
делись, просто похожи на все остальные зубы. У примитивных речных дельфинов
иний их ещё можно отличить от остальных зубов, у настоящих дельфинов – уже нет.
Об астрагалах (таранных костях) я уже говорил – они есть даже у нас, людей.
Простое их наличие – не аргумент.

Череп инии показывает, что её зубная система гетеродонтна, и коренные зубы ещё можно отличить от остальных.
Но логика Тена вообще иррациональна:
у дельфинов нет астрагалов – значит, они не потомки кондиляртров. У дельфинов
вообще нет задних ног. Может, они – потомки змей? У них ведь тоже нет задних
ног…
Если уж искать предков дельфинов, то следует помнить, что они – не первые в
ряду потомков своих наземных четвероногих предков. Если построить цепочку от
наземных предков до нынешних дельфинов, то окажется, что между ними окажется
очень много промежуточных форм. И ещё окажется, что дельфины произошли от постоянноводных
форм, уже лишившихся задних конечностей. Это всё равно, что поставить рядом
пещерную живопись и полотно Рубенса, и доказывать, что человек не мог нарисовать
такую картину, а потому Рубенс был… допустим, марсианином. Или нибируанцем.
Или злокозненным рептилоидом с Сириуса.
Стр. 212:
Достаточно распространенная ошибка многих, читавших мои книги, заключается в
том, что мне приписывают гипотезу происхождения человека от дельфинов. Я этого
никогда не утверждал. Я считаю, что непосредственным животным предком
человека была неотеническая форма древних морских млекопитающих,
скорее всего дельфинид, а это совсем другое. Это другое существо, другой вид.
Подробней об этом поговорим позже.
Хорошо, учтём и посмотрим. Только окончание
«-иды» в биологии характерно для названий семейств. Поэтому слово «дельфиниды»
у человека, хотя бы немного знающего биологию, отождествляется с семейством
Delphinidae, и Капитан Очевидность подсказывает, что все его представители являются
дельфинами, даже косатки и гринды. Поэтому оправдание Тена о том, что «это
другое существо, другой вид» выглядит, мягко говоря, сильно
натянутым.
В биологии был такой прецедент – дронты на основании крайне специфических морфологических
особенностей были выделены в отдельное семейство. Биохимия вернула их в лоно
семейства голубиных – дронты фактически представляют собой голубей, вставших
на путь ювенилизации и сохранивших в облике взрослой особи множество особенностей
голубиного птенца.

Собственно, птенцы голубя с намёком на облик дронта.
Если бы версия Тена была реальностью, подкреплённой реальными зоологическими, палеонтологическими и биохимическими данными, то «неотеническую форму древних морских млекопитающих» и всех её потомков отнесли бы к тому же таксону, к которому относятся не-неотенические родственники данной формы, как в случае с дронтами и семейством голубиных.
Стр. 212-213:
До недавнего времени были известны только карликовые формы приматов древностью 60 млн лет. Трудно себе представить в качестве предка трехтонной косатки десятисантиметрового лемура. Вот это «трудно себе представить» и стало основанием для палеонтологов. Других оснований нет. Но сейчас известно, что среди древних лемуров были настоящие гиганты – ростом с человека и даже больше гориллы.
Так, у нас в рассуждениях появилась косатка.
Значит, речь всё же идёт о семействе дельфинов (Delphinidae), к которому она
относится.
Если уж брать больше, то кто бы мог представить себе в позднем триасе, глядя
на маленького мегазостродона, что его далёким потомком будет синий кит? Тем
не менее, будущее показало, что это так. У палеонтологов есть куча других оснований
признавать или не признавать тех или иных существ родственниками, помимо теновского
«трудно себе представить»: те же морфологические
особенности и распределение ископаемых остатков рассматриваемой группы живых
организмов в последовательности отложений. Какое-то специализированное состояние
признака у ранее жившей группы живых организмов может препятствовать его эволюции
в состояние, наблюдаемое у сходной с ней группы организмов, живших позже. Значит,
эти две группы – не предки и потомки, а в лучшем случае имели общего предка.
Плюс биохимия вносит свой вклад в установление родства изучаемых групп живых
организмов, если в распоряжении есть образцы их органических остатков. Поэтому
насчёт «других оснований нет» Тен просто врёт.
На кого рассчитана такая грубая и примитивная ложь – не пойму. Неспособность
что-либо представить – это не повод вносить изменения и исправления в родословное
древо той или иной систематической группы живых организмов, а всего лишь персональные
проблемы когнитивного плана. Кому «трудно себе представить»,
как работает наука, можно порекомендовать не соваться в науку и уж тем более
не поучать специалистов.
Стр. 213:
До катастрофы, произошедшей 65 млн лет назад, лемуры и долгопяты прятались в
прибрежных тростниках, не смея надолго выходить на сушу или пускаться в море.
Катастрофа уничтожила крупных холоднокровных хищников, потому что вслед за ней
наступила холодная пылевая ночь. Отсюда должно быть понятно, почему именно океан
Тэтис стал колыбелью новой жизни и очагом эволюции новых видов: он находился
на противоположной стороне планеты, где меж черных туч могло иногда появляться
солнце, согревая землю. Кроме того, весь океан пересекала обширная область вулканической
активности, через которую на поверхность проникало внутреннее тепло планеты.
Благодаря геотермальным источникам теплилась жизнь простейших и моллюсков, которые
питаются простейшими и которым свет не нужен вовсе. Что касается твердой поверхности,
то на ней возможности существования сократились до минимума.
А вот теперь становится интересно. На
момент написания этих строк единственным докайнозойским приматом является пургаториус
из Северной Америки. И то есть обоснованные предположения, что единственная
фрагментарная окаменелость, найденная в отложениях мелового возраста, была перезахоронена
в них из чуть более молодых палеоценовых отложений. А Тен уже говорит о лемурах
и долгопятах… Я бы хотел узнать названия их видов, места находок, имена авторов
описаний.
Единственные докайнозойские лемуры, которые мне известны – это герои давнишнего
диснеевского мультфильма «Динозавр» (2000). Хороший мультфильм, очень добрый.
Но… это вымысел, а не наука. И лемуры в том мультфильме были говорящими, впрочем,
как и динозавры.

Вот они, эти мезозойские лемуры. Из мультфильма.
Так что пока можно смело сказать, что
Тен пытается рассказывать читателям сказки – прямо с первых слов абзаца.
Нахождение Тетиса в ином полушарии не спасло бы гипотетических «лемуров Тена»:
пыль от столкновения астероида долго находилась во взвешенном состоянии в воздухе.
Земля имеет свойство вращаться вокруг своей оси, и ещё в атмосфере дуют ветры.
Поэтому спрятаться от пыли на другом полушарии было невозможно. Наличие иридиевой
аномалии фиксируется по всему земному шару.
Геотермальные источники? Вулканизм? Хмммм, суньте кипятильник в олимпийский
бассейн, и дождитесь, пока вода закипит. Вот, на Галапагосских островах активно
идёт вулканическая деятельность, в том числе под водой, а холодное Перуанское
течение хоть бы на градус теплее стало – на экваторе пингвины живут! Глубоководные
«чёрные курильщики» с перегретым кипятком по температуре воды на поверхности
океана вообще не угадываются – их открыли лишь с подводных аппаратов.
Стр. 213:
Лемуры и долгопяты устремились в теплую, темную воду. Они питались, нащупывая
в темноте чуткими длинными пальцами мелких моллюсков, и разделывали их при помощи
ногтей. Отсюда ногти (вместо когтей), чуткие, цепкие, супинаторно подвижные
кисти с отстоящим большим пальцем (для противоупора, когда надо разорвать двустворчатого
моллюска) и большие круглые глаза (впоследствии унаследованные человеком, почему
мы удивительно похожи на долгопятов). Лемуры и долгопяты лишились почти всех
средств самозащиты, потому что хищников не существовало несколько миллионов
лет. В то же время необходимость приспособления к новым тяжелым условиям вызвала
рост интеллекта. Лемуры настолько умны, что умеют лечиться травами: поранившись,
они останавливают кровь, прикладывая травы к ранам.
Тен всё-таки смотрел мультфильм «Динозавр»:
там говорящие лемуры как раз умели лечить травами – не только себя, но даже
динозавров. Тоже говорящих.
Мне нравится описание образа жизни этих существ: палеонтологи пока ни косточки
не нашли, чтобы подтвердить хотя бы само их существование, а Тен уже в красках
живописует их жизнь. Вот только при отсутствии доказательств его рассказ можно
со спокойной совестью поставить на одну полку со сказками народов мира.
«… хищников не существовало несколько миллионов лет»?
А змеи? А крокодилы? А птицы? Даже водяные черепахи – опасные хищники, охочие
до мяса и умеющие его добывать. Плюс отсутствие млекопитающих отряда хищных
(Carnivora) вовсе не означает, что в раннепалеоценовом мире не было млекопитающих,
поедавших мясо. Вот небольшая
заметка о них на сайте палеонтолога Martin Jehle.
Правильно ли утверждать, что «лемуры и долгопяты лишились почти всех
средств самозащиты»? Разве древесный образ жизни – не
защита от наземных хищников? Разве способность скакать по тонким веткам – не
защита от более крупных древесных хищников? Разве скорость и ловкость движения
по ветвям, быстрота реакции и умение следить за окружающей обстановкой – не
защита от пернатого воздушного хищника?
Наконец, водные животные вроде калана или выхухоли, питающиеся беспозвоночными,
не стремятся расставаться с когтями, меняя их на ногти – это исключительный
признак приматов. И обрели ногти приматы ещё до того, как разделились на лемуров
и долгопятов, до гипотетического «переселения в воду», которое пытается приписать
им господин Тен.
Круглых глаз мы от долгопятов не наследовали. Наши глаза обладают подвижностью
благодаря наличию глазодвигательных мышц. У крупных глаз долгопятов с этим проблемы
– эти мышцы не развиты, и долгопят вынужден сильно вертеть головой, чтобы что-то
разглядеть, поскольку скосить глаза он не в состоянии. Из более специализированного
состояния в менее специализированное признак не может перейти в силу закона
необратимости эволюции. Он же – закон Долло, хорошо известный господину Тену.
Стр. 214:
Эволюционно новый фенотип полуводных животных (ибо это были уже не лемуры и
долгопяты) формировался исключительно для добывания и разделывания моллюсков
в условиях пониженной освещенности в теплой воде – тот фенотип, который впоследствии
был унаследован людьми через несколько удивительных превращений.
Пылевые облака от столкновения Земли с тем злополучным астероидом висели в небе, самое большее, несколько лет. Неприятное событие, конечно, но это даже меньше времени жизни одной особи млекопитающего средних размеров – с кошку или опоссума. Возможно даже, что, когда небеса на Земле вновь прояснились, в разных местах планеты ещё продолжали жить те особи, которые помнили времена до падения астероида. Прошли даже не сотни, не десятки, а единицы поколений более мелких и быстро развивающихся животных. О каких эволюционных изменениях за такое короткое время может идти речь?
Стр. 214:
Здесь возникает вопрос: где они держали своих детей, когда земля была покрыта
льдом и снегом? Наиболее вероятный ответ: в теплых заводях. Детеныши росли в
воде и постепенно меняли внешний вид.
В теплой воде шерсть начала выпадать, редеть, пока не исчезла совершенно, а
для «сугреву» стал накапливаться подкожный жир по типу морских животных. Скорее
всего в воде начала увеличиваться и масса. Ледниковая фауна всегда крупнее межледниковой.
Чем больше масса, тем меньше вероятность замерзнуть. Постоянное пребывание в
воде также способствует накоплению массы тела в связи с изменением силы тяжести.
Таким образом, были сделаны первые полшага к превращению в археоцет.
Сказка продолжается… И это при том, что
за кратковременным похолоданием после падения астероида следовал относительно
тёплый палеоцен, а по окончании его случился палеоцен-эоценовый термический
максимум (ПЭТМ) – резкое потепление по всей Земле. Именно тогда в Гренландии
ненадолго начали расти пальмы. Это произошло около 55 млн. лет назад. Запомним
эту дату.
Теперь о драматичном описании перехода в воду. Мелкие водные животные, проводящие
часть жизни на суше, не стремятся расставаться с шерстью. Мало того – она у
них очень тёплая и густая: вспомним бобра, нутрию, выдру, калана, норку и выхухоль,
в разные эпохи бывших объектами промысла ради меха. У землеройки куторы, мелкого
полуводного зверька, шерсть даже водоотталкивающая. Тюлени (ушастые и настоящие)
также не спешат терять шерсть, хоть и имеют слой жира. Шерсть исчезла только
у постоянноводных млекопитающих, и ещё у крупных бегемотов.
Разгадка такой тенденции проста: даже тёплая вода в природном водоёме имеет
температуру ниже температуры тела млекопитающего, поэтому у теплокровного животного,
особенно мелкого, возникает неиллюзорная опасность переохлаждения. Из-за этого
обстоятельства выпадение шерсти у детёнышей, которые меньше взрослых особей,
в условиях жизни в воде при изначально небольшой массе тела будет антиадаптивным
признаком.
Если увеличивается масса тела (что есть адаптивный признак при переходе к водному
образу жизни), то закономерно увеличивается вероятность нахождения такого существа
в ископаемом состоянии – большие кости не затеряются и хотя бы частично сохранятся
и привлекут внимание палеонтолога. Из палеоцена известны фауны ископаемых млекопитающих,
но водных приматов Тена пока никто не обнаружил. Сказки, сказки…
Стр. 214-215:
Низшие (хвостатые) обезьяны произошли от полуобезьян сравнительно поздно. Их
эволюция началась позже эволюции археоцет, когда метеоритная зима окончилась.
Они представляют собой конечные продукты специализации отдельных видов полуобезьян.
Логично предположить, что от полуводного образа жизни на полностью морской фенотип
перешли не все. Когда на планете восстановился тропический климат, часть лемуров
и долгопятов продолжила наземную эволюцию.
Метеоритная зима окончилась за время
жизни 1-2 поколений млекопитающих среднего размера. Далее была стабилизация
природных условий, а в конце палеоцена начался тот самый термический максимум
– 55 млн. лет назад. Древнейшее возможное китообразное, плохо известный Himalayacetus,
имеет возраст 53,5 млн. лет. Если учесть, что вид получает больше шансов на
захоронение в ископаемом состоянии, когда его численность максимальна, можно
смело предположить, что и пакицетус, и гималайяцетус были на самом деле несколько
древнее, чем показывают их ископаемые остатки. То есть, термический максимум
как раз предшествовал появлению китообразных, и даже совпадал с этим событием,
и архаичные киты в любом случае появились в очень тёплом мире. То есть, в воду
предков китов загнал далеко не холод.
А когда же появились самые ранние обезьяны? И тут нас ждёт сюрприз. Archicebus
achilles и Teilhardina asiatica имеют возраст 55,8-48,6 млн. лет (информация
отсюда – очень полезный ресурс).
Стало быть, обезьяны оказываются одного возраста, и даже немножко древнее самого
древнего китообразного. То есть, снова имеет место фактическая ошибка в книге
Тена.
Стр. 215:
Превращение полуобезьян в низших обезьян представляло собой не прогресс, а продолжение
специализации для жизни в лесу или саванне. Интеллект низших обезьян не отличается
от интеллекта лемуров и в десятки раз, на целый порядок уступает интеллектам
дельфинов и человекообразных обезьян. Между антропоидами и низшими обезьянами
(вроде мартышки и павиана) такая пропасть, что о родстве говорить не приходится.
Их происхождение и генезис абсолютно различны. Эволюционная схема: полуобезьяны
– низшие хвостатые обезьяны – высшие бесхвостые обезьяны – человек – представляет
собой вульгаризацию и неоправданное спрямление эволюции.
Когда-то по всему Старому Свету и Северной
Америке жили лемуры и сходные с ними полуобезьяны (например, адапиды). Но после
появления обезьян они стремительно сдали позиции по всему миру, полностью сохранив
свои позиции лишь в изолированном мире Мадагаскара. В континентальной Африке
полуобезьяны – это всего лишь несколько видов галаго и потто, а в Азии – лори
и долгопяты. Их видовое разнообразие значительно ниже, чем у обитающих по соседству
обезьян, и они вытеснены ими в мелкоразмерный класс.
Относительно интеллекта низших обезьян и лемуров можно поспорить. Много ли лемуров
приспособилось к жизни в городах? Ни одного. А обезьяны в городах тропических
областей мира – самое обычное явление. Их умение приспосабливаться к быстро
меняющейся и изначально неестественной городской обстановке указывает на значительную
сообразительность. Плюс известны бабуины, которые справлялись с обязанностями
пастуха, знали всех коз в стаде и какой козлёнок какой козе принадлежит – это
не всякий человек запомнит. Павиан Джек-сигнальщик успешно трудился на железной
дороге, переводя стрелки по команде. В туристических районах макаки прекрасно
понимают ценность украденной у туристов вещи, и выкуп мобильного телефона за
пару орешков не получится. Так что не стоит недооценивать «низших» (мартышкообразных)
обезьян. Среди широконосых обезьян недюжинный интеллект демонстрируют капуцины.
Так что, говоря об интеллекте не-человекообразных обезьян, господин Тен либо
ошибается, либо сознательно врёт. Да, кстати, бабуины тоже способны к обману
– это говорит об их умении представить себе, о чём думает сородич, и манипулировать
этим.
Чтобы утверждать, что между мартышкообразными и человекообразными обезьянами
имеется «пропасть», Тену следует, как минимум, доказать, что древние обезьяны
семейств Proconsulidae и Dendropithecidae просто не существовали, или же были
типичными мартышкообразными обезьянами. Кстати, в 1-м томе книги Ст. Дробышевского
«Достающее звено» («Обезьяны и все-все-все») переход от низших узконосых обезьян
к человекообразным очень хорошо охарактеризован. Просто удивительно, почему
этот момент книги Дробышевского не подвергся сокрушительной и беспощадной критике
Тена. Могу лишь предположить, что Дробышевский говорит предметно, приводя названия
конкретных видов, а Тен слишком слабо представляет себе их признаки, чтобы делать
какие-либо возражения. Поэтому в данном случае мы видим, как Тен «бросается
слонами»: не было, мол, таких переходных форм, и всё тут. Можно, конечно, предположить,
что он чисто физически не дочитал книгу Дробышевского до этой главы, но пока
я склонен верить, что в людях ещё осталось что-то хорошее.
Стр. 215:
Лемуры (или долгопяты), бывшие всеядными обитателями прибрежных пространств,
полуостровов и островов Тэтиса, имевшие развитые, подвижные (более 100 степеней
свободы) конечности с длинными лучами, скорее всего и стали предками зубатых
археоцет. Для подобного вывода не имелось внешних оснований до тех пор, пока
не было известно о существовании в древности крупных лемуров. Во второй половине
XX в. поступили сведения о существовании гапалемура, который дожил до наших
дней в тропических болотах. Гапалемуров описывают как «довольно плотных животных
до 90 см роста с округлой головой, широкой мордой и короткими ушами, сходными
с человеческими» (Фридман, 1979, с. 78). В самом конце прошлого столетия на
Мадагаскаре было сделано открытие, вызвавшее сенсацию: гигантский ископаемый
лемур ростом более двух метров! Он жил в пещере, как древний человек. Когда
по ТВ продемонстрировали его скелет, мне показалось, что неспециалист мог бы
спутать его с человеческим. Подобное существо за 55 млн лет вполне могло превратиться
в дельфина.
Сложно анализировать текст Тена, когда
в нём отсутствуют конкретные названия описываемых видов. В таком случае есть
риск удариться в додумывание, а это крайне неблагодарное дело, поскольку чужая
душа – потёмки.
Лемуров нельзя назвать «или долгопятами»: это отдельное семейство Tarsiidae.
А лемуры – это инфраотряд в отряде приматов, включающий несколько современных
и вымерших семейств. То есть, эти понятия – не синонимы. Фактическая ошибка
Тена. Снова.
Гапалемур – крупный? Тен не очень точно цитирует Фридмана. У Фридмана в книге
написано следующее:
«Есть сведения, что гапалемуры сравнительно крупные животные: 70–90 см, но,
по данным Нейпир, их размеры (голова – туловище) 26–37 см с такой же примерно
длиной хвоста. Эти размеры соответствуют и тем, которые мог отметить автор этой
книги у экспоната в Ленинградском зоологическом музее.
Довольно плотные приматы с округлой головой, широкой мордой и короткими ушами,
сходными с человеческими…». (Э. П. Фридман «Приматы. Современные полуобезьяны,
обезьяны и человек» Академия наук СССР Издательство «Наука» Москва, 1979)
То есть, перед нами небольшие лемуры. В современной фауне есть и покрупнее –
тот же индри, например.
Действительно гигантских лемуров в фауне Мадагаскара не так уж и много, и они
все, к сожалению, вымерли в последние немногие тысячелетия. Мегаладапис до двух
метров не дорастал. А вот археоиндри, возможно, и дорастал: его вес оценивается
в 150-210 кг. И их остатки как раз обнаружены в пещере. Однако то, что они умерли
в пещере, ещё не означает, что они в пещере жили: пещера Ампасабазимба могла
сработать как естественная ловушка; такое часто встречается в разных частях
света. Далее, достаточно глянуть на морды таких лемуров (даже на череп), чтобы
понять, что с человеком их не спутаешь даже в состоянии сильного алкогольного
опьянения – они сильно вытянуты. Между прочим, цитируемый Теном «учёный-инволюционист»
А. Белов как раз отмечал внешнее свиноподобие морды мегаладаписа и аргументировал
этим свою мысль о том, что свиньи произошли от человека в процессе «инволюции»
через стадию вроде этого лемура.
И ещё одна мелочь: гигантские лемуры Мадагаскара жили в плейстоцене и голоцене,
когда моря уже кишели вполне современными по облику китообразными разных форм
и размеров. Эти гигантские лемуры немного опоздали появиться, чтоб от них произошли
киты: всего лишь на какие-то полста миллионов лет.
Стр. 216:
Судя по эмбриональному развитию дельфинов, предком этих морских млекопитающих
должно было быть наземное млекопитающее с круглой головой и пятипалыми конечностями.
Именно так выглядят дельфиньи зародыши на ранних стадиях развития.
Нет-нет, уважаемый господин Тен, биогенетический
закон касается эмбрионов – речь там идёт об онтогенезе. Мы уже говорили об этом,
комментируя страницу 212. У предка дельфинов был похожий эмбрион, но такие же
эмбриональные черты имеют очень многие животные. На стадии, когда у эмбриона
дельфина круглая голова, он мало чем отличается от эмбрионов множества других
млекопитающих. Так что из строения зародыша дельфина мы можем заключить лишь
то, что его предком было пятипалое четвероногое животное, у эмбриона которого
на ранних стадиях развития голова была круглой. А была ли она круглой у взрослых
особей этого предкового вида – это ещё бабушка надвое сказала. Но чем вести
разговоры, достаточно просто посмотреть на эмбрионы различных позвоночных.
Предком крота, судя по его эмбриону, тоже было явно очень умненькое круглоголовое
животное.
У волка – такой же круглоголовый умненький предок?
А летучие мыши – они, наверное, вообще создали крылатую цивилизацию, не дошедшую до наших дней?
А чем умненькие предки серых китов хуже? Это сколько ж архаичных неизвестных науке звероцивилизаций было на Земле?
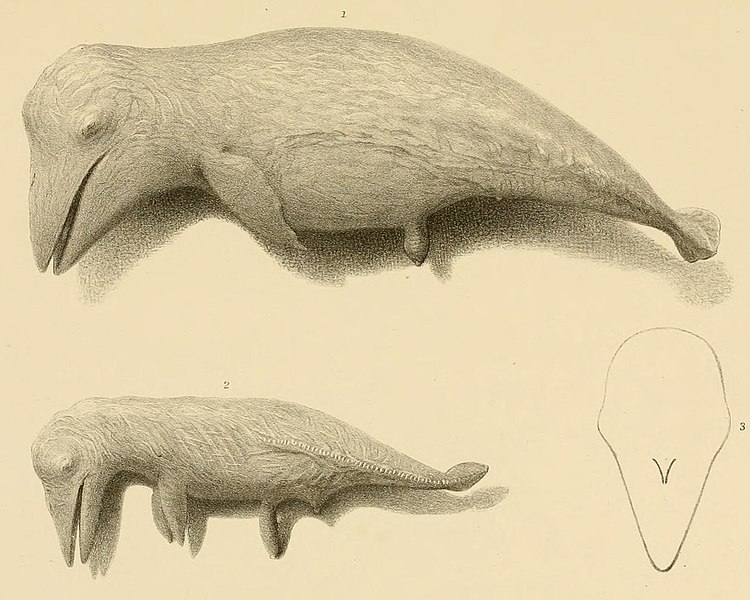
А вот для полноты картины эмбрион змеи:
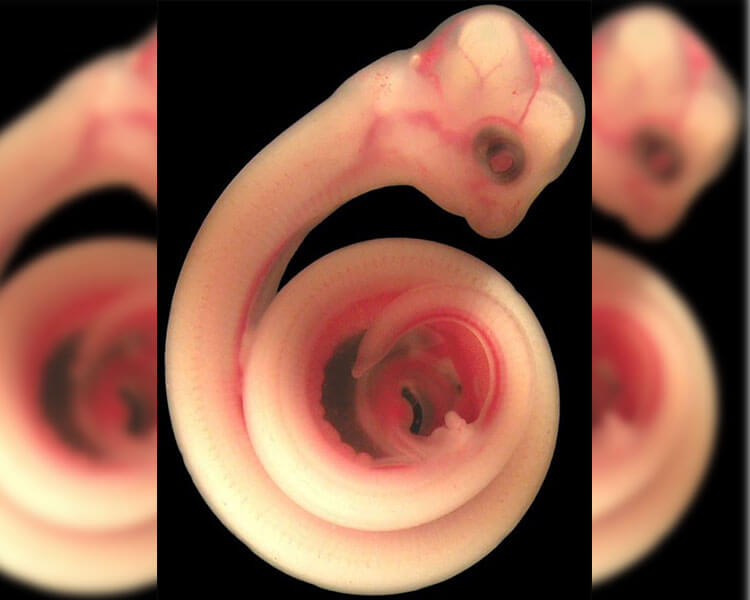
Исходя из особенностей облика змеиного эмбриона, предком змеи несомненно было безногое прямоползущее существо с круглой головой – скорее всего, рептилоид с Сириуса. Или с Проксимы Центавра. Или с Ахернара.
Стр. 216:
...В 1984 г. в южной Германии был найден целый скелет странного, весьма крупного
существа. У него была довольно круглая голова, плоские ноги и таранные кости,
как у высших обезьян, и почтенный возраст – 47 млн лет. В 2004 г. его объявили
искомым предком высших обезьян и, соответственно, самым древним предком человека.
Находке было придано великое значение. Карьер, где она была найдена, объявили
«памятником всемирного наследия», существу дали имя собственное Ида, а главное
– почетное название «дарвиний».
Какой-то странный «околопалеонтологический
экстаз» крутит Тена в этом месте его книги… Столько бреда намешано, что разбираться
в нём придётся долго.
Странно, что, другие животные, названные в честь иных деятелей науки – например,
жозефоартигасия, карлоамегиния, флорентиноамегиния, иностранцевия, суминия,
ивантозавр, томасхакслия – такого восторга, переходящего в экстатическое возбуждение,
у Тена не вызывают. Видимо, выработался рефлекс сугубо на имя Дарвина… И ещё
– её назвали просто в память о двухсотлетии со дня рождения Дарвина
(ссылка).
Что с того, что это «весьма крупное» (с домашнюю кошку: длина головы и тела
без хвоста 24 см) существо назвали Идой? Почему Тена не возбуждают аллозавр
Большой Ал (про него аж кино сняли!), тираннозавриха Сью, тесцелозавры Вилло
и Максимус, мамонтёнок Дима и многие другие? Ах, да, у нас есть ещё Люси с такой
же реакцией на неё. У Тена, что, совсем плохо с женщинами, что он на кости не
первой свежести в таком экстазе кидается? Да, это я в очередной раз злобно пошутил.
Настроение от чтения работы Тена весьма располагает, знаете ли. Но мы идём дальше.
Карьер в Месселе ЮНЕСКО объявило «памятником всемирного наследия» 9 декабря
1995 года вовсе не из-за находки Иды. Её нашли во время самодеятельных раскопок
в 1983 году, а описали лишь в 2007 году – образец долго хранился у торговца
окаменелостями, и на момент объявления карьера памятником ещё не был изучен.
Тен явно плохо осведомлён о фактах, которые с таким апломбом излагает, и вдобавок
немного путает причинно-следственные связи. Этот карьер даже без находки злополучной
Иды оказался исключительно богатым на находки разного рода доисторических животных
исключительной сохранности, в том числе редко встречающихся в иных условиях
– ёжиков, летучих мышей, мелких птиц и прочих. Потому и сделали Мессельский
карьер объектом Всемирного наследия и заповедником – чтобы всякие мутные личности
не занимались самодеятельными раскопками, попутно уничтожая что-то ценное.
Наконец, никто не ждал, что дарвиниус будет предком высших обезьян: этого примата
ставили на ступень значительно ниже: между лемурами и ранними обезьянами. Причём
дарвиний даже не является прямым предком низших обезьян, потому что жил позже
их появления: 47 млн. лет назад против 55,8-48,6 млн. лет.
Остаются таранные кости (астрагалы). В статье 2010 года “Astragalar morphology
of Afradapis, a large adapiform primate from the earliest Late Eocene of Egypt”
(в свободном доступе здесь)
говорится, что Darwinius вместе с анализируемым в статье родом Afradapis образуют
монофилетическую группу среди Strepsirrhini (то есть, полуобезьян, мокроносых
приматов), и тем самым поддерживается предположение о том, что антропоиды (обезьяны)
эволюционировали не из числа адапидных предков, а признаки, которые изначально
считались общими для антропоидов (их синапоморфиями), на самом деле могли возникать
независимо у разных приматов.
Стр. 216:
Существо называли «восьмым чудом света» и «Мона Лиза эволюции». Но каково же
было разочарование, когда выяснилось, что дарвиний – это не низшая обезьяна,
имеющая признаки высшей обезьяны, а лемур. Иду списали с корабля эволюции. Ибо
она нарушала Священную историю эволюции, написанную антропологами: низшие обезьяны
– высшие обезьяны – человек. Факты противоречат, тем хуже для фактов.
Всё было немного не так. Повторюсь, что
её вначале сочли переходным звеном между лемурами и низшими обезьянами. Никто
никогда не приписывал ей родства с высшими обезьянами – она даже по времени
существования не подходит под переходную форму, не говоря уже об особенностях
анатомии. Тен явно искажает информацию об этом примате и его месте на родословном
древе отряда. Делается это по глупости или злонамеренно – не знаю, но и то,
и другое – это очень плохо.
Пафосность выражений, употребляемых Теном, поистине зашкаливает: «… списали
с корабля эволюции». Поэзия, одно слово! Никто, правда, не списывал: место дарвиниуса
на родословном древе приматов просто было уточнено, и всё. Когда то же самое
происходит у непарнокопытных, рукокрылых или, допустим, в каком-нибудь из многочисленных
отрядов костных рыб, никто не рыдает, заламывая руки, и не хохочет зловещим
голосом, наблюдая за чужими страданиями. Но вот здесь вполне естественный процесс
уточнения систематического положения очередного ископаемого вида вызывает восторг
на уровне щенячьего.
А комплименты, упомянутые Теном… Что ж, не очень часто удаётся обнаружить такой
полный образец ископаемого примата, к тому же такого древнего. А наша Ида –
это самый полный образец ископаемого примата в мире – сохранилось 95% скелета
(у Люси – около 40%). Йорн Хюрум, специалист, описавший Иду, сказал об этом
образце так: «Чтобы привлечь посетителей в музей, вам нужен шедевр, или два.
Это наша Мона Лиза, и она будет нашей Моной Лизой на следующую сотню лет» (ссылка).
Вот так, просто и прозаично – этот образец просто красив и привлекателен, вне
зависимости от своего положения в родословной приматов. Да, просто «Мона Лиза»,
а не «… эволюции». И «восьмым чудом света» назвал Иду Йенс Франсен, один из
соавторов описания образца, именно за полноту скелета (ссылка).
О месте Иды в систематике приматов речь даже не шла.
Стр. 216:
Дарвиний обитал на территории, омываемой Тэтисом. Не исключено, что вел водно-земный
образ жизни. Он имел руки с длинными, изящными пятью пальцами, оканчивающимися
ногтями, которыми вполне мог разделывать моллюсков. У него были большие глаза,
большой палец отстоял на 90 градусов как на передней, так и на задней конечности.
Возможно, это и был предок дельфина? Тем более что наземная линия дарвиния неизвестна.
Возможно, перед нами одна из поздних особей существ, чья эволюция началась гораздо
раньше, 60 млн лет назад? Первым дельфинидам 48 млн лет. Дарвинию – 47 млн лет,
но это ведь единственный экземпляр.
А вот тут у меня начинается лёгкий когнитивный
диссонанс, потому что я потерял уверенность в том, каких животных Тен именует
«дельфинидами». 48 миллионов лет – это ещё эоцен, и в это время на Земле существовали
исключительно археоцеты, причём самые ранние – пакицетус и, возможно, уже жил
амбулоцетус (возраст – 47 млн. лет), или какой-то его непосредственный предок
примерно такого же уровня развития. Эти археоцеты были полуводными животными
с хорошо развитыми задними конечностями и тазом, прикреплённым к позвоночнику.
Семейство дельфиновых (Delphinidae) в том смысле, в каком его понимают учёные,
существует с позднего олигоцена по настоящее время. В каком объёме и смысле
понимает его состав Тен – из его книги неясно, и это обстоятельство определённо
создаёт трудности в ходе дальнейшего анализа его соображений, поскольку нельзя
понять, о какой группе животных он говорит в тот или иной момент времени. Впрочем,
нет худа без добра: чем больше точка зрения Тена в вопросах, отличных от «симиализма»,
не совпадает с мнением специалистов, тем быстрее даже несведущий читатель поймёт,
чего стоят соображения автора «инверсионной теории».
Вообще, отсутствие конкретики – это общая особенность рассуждений всех «альтернативных»
ниспровергателей теории эволюции вообще или в частностях вроде антропогенеза.
Дело в том, что отсутствие конкретики очень удобно для ведения нечестной полемики:
это как играть в футбол, имея незакреплённые ворота: захотел – развернул их
вправо, влево, или вообще задом наперёд, захотел – заменил на ворота меньшего
размера. Иными словами, честная игра превращается в плутовство. Креационисты
часто пользуются таким приёмом. Когда их просишь конкретизировать понятие для
целей дискуссии, они предпочитают эту дискуссию прекратить, либо пропускают
просьбу мимо ушей.
Но я отвлёкся…
Вернёмся к нашей маленькой Иде (почти Андерсен получается!). Не спорю, моллюсков
Ида вполне могла разделывать и кушать: на деревьях улитки тоже живут. Кто скажет,
что это не моллюски – пусть первый бросит в меня камень.
Стоит обратить внимание на такую особенность дарвиниуса, как отстоящие большие
пальцы. У современных водных млекопитающих палец не отстоит – ни у кого из них
нет ластов с большим пальцем, как у мистера Кобблпота (Пингвина) из «Бэтмена».
Если уж у них кисть или ступня превратилась в ласт, то в его образовании участвуют
сразу все пальцы этой части тела. Не нужно быть Шерлоком Холмсом, чтобы увидеть,
что пальцы Иды обладают очень длинными тонкими косточками, тогда как у водных
млекопитающих в ластах кости пальцев прочные, поскольку испытывают нагрузки
при гребле. Тонкие кости, как у Иды, могут сломаться поперёк во время гребка.
Хвост Иды не принимал участие в плавании – он для этого слишком тонкий, из небольших
позвонков, поэтому, если предположить водный образ жизни этого существа, приходится
признать, что главным органом плавания у него были конечности. Но отстоящие
пальцы и тонкие длинные фаланги пальцев строго против этого предположения.
Если же представить себе сочетание отстоящего большого пальца для лазания и
перепонки между четырьмя остальными пальцами для плавания, то лазать по деревьям
с такой конструкцией было бы крайне проблематично: любой сломанный сучок проколет
и распорет перепонку. Если же отстоящие пальцы есть и на передней, и на задней
конечности, это означает, что ни передняя, ни задняя конечность не была ластом.
Стр. 216-217:
Обращу внимание еще на один факт, на который до сих пор не обращали внимания:
пальцы дарвиния слишком длинные относительно тела. Гораздо длиннее, чем у человека
и всех известных обезьян. Они явно были неудобны при сухопутном образе жизни,
даже в качестве брахиатора. Не было ли между ними перепонок? Возможно, дарвиний
уже был адаптирован для жизни в воде с периодическими выходами на сушу? А ведь
предком дельфина было существо с чрезвычайно длинными пальцами.
Экземпляр дарвиниуса отличается превосходной
сохранностью – на нём сохранились отпечатки мягких тканей и шерсти. Отпечатки
передних и задних конечностей прекрасно различимы, и следов перепонок между
пальцами нет.
Длинные пальцы – очевидный признак лазающего существа. Ходить по ровной земле
на таких конечностях, конечно же, не очень удобно, но зато можно хвататься любой
из четырёх конечностей, отстоящие пальцы помогают в этом отлично.
Наличие хватательных хорошо развитых задних конечностей, длиной примерно в полтора
раза больше передних, говорит о том, что это существо не было брахиатором, подобно
гиббону, а с учётом маленького размера тела животного позволяет предположить,
что животное передвигалось по веткам на всех четырёх конечностях – как лемуры
или мартышкообразные обезьяны.
Наконец, стоит сказать и о длинных пальцах китов. Удлинение пальцев у китов
происходит иначе, чем у лазающих млекопитающих: не за счёт удлинения костей,
а за счёт увеличения количества фаланг (гиперфалангии). Это обычное явление
у вторичноводных животных, наблюдаемое также у ископаемых ихтиозавров и плезиозавров.
У наземного предка всех млекопитающих было заведомо меньше фаланг пальцев. Кстати,
у ранних китообразных количество фаланг в пальцах было не больше, чем у остальных
современных им млекопитающих: фаланговая формула 2/3/3/3/3 (источник:
“Evolution of hyperphalangy and digit reduction in the cetacean manus”).
Стр. 217:
Выводы об изменениях в строении тела и мозга дельфинов в процессе эволюции делаются на основе не только палеонтологических данных, но и на основе данных изучения эмбрионов современных дельфинов на разных стадиях их развития. Научным базисом последнего подхода является биогенетический закон Геккеля, который устанавливает воспроизводимость общей картины филогенеза в онтогенезе. Путь, пройденный видом, повторяется в стадиях развития эмбриона. Иначе говоря, эмбрионы животных в процессе развития в животе матери быстро «проходят» путь, пройденный за миллионы лет эволюции всем видом. Превращение зародыша дельфина в животе матери чрезвычайно интересны.
Сразу надо напомнить одну оговорку: этот путь повторяется в сокращённом и обобщённом виде, и эмбрион рассматриваемого нами вида не является абсолютным подобием эмбриона своего предка (с возможностью определения до уровня вида?), и уж тем более взрослой особи. По изменениям эмбриона в процессе его индивидуального развития можно приблизительно понять, как выглядели эмбрионы предков, и какие крупные эволюционные события претерпевала данная группа живых организмов в целом. Однако на процесс развития эмбриона накладываются более поздние эволюционные изменения, причём они могут не только добавляться к поздним стадиям развития предкового вида (так называемые анаболии), но и влиять на средние (девиации), и даже на ранние (архаллаксис) стадии развития эмбриона. Поэтому считать эмбриона дельфина полным подобием эмбриона его предков нельзя.
Стр. 217:
«Как известно, – пишет Г. Мчедлидзе, крупнейший специалист по палеобиологии
китообразных, – зачатки задних конечностей закладываются в эмбрионе всех современных
китообразных. Однако на определенных стадиях эмбриогенеза они исчезают. Так,
например, у эмбриона дельфина, уже достигшего 75-дневного возраста, зачатки
этого органа полностью утрачены. Совершенно очевидно, что сохранение на ранних
стадиях эмбриогенеза китообразных зачатков задних конечностей является не чем
иным, как рекапитуляцией признаков их наземных предков» (Мчедлидзе, 1976, с.
110). Передние конечности сохранились, преформировавшись в пятилучевые ласты.
Если даже задние конечности китообразных
в силу каких-то хитросплетений работы генов продолжат развиваться, то их степень
развития будет соответствовать всего лишь таковой у эмбриона – то есть, они
будут выглядеть всего лишь как зачатки конечностей, но не разовьются в полноценные
конечности. В эпоху интенсивного китобойного промысла достаточно часто фиксировались
случаи наличия у китов (в частности, у кашалотов) выступов по обе стороны от
генитальной складки, и внутри этих выступов обнаруживались рудиментарные мускулы
и кости. Были даже случаи, когда у кашалотов имелись лопастевидные образования,
отдалённо напоминающие ласты:
«В 1958 году мы исследовали самца кашалота длиной 11,6 метров, пойманного в
Беринговом море, с необычными выпячиваниями в тазовой области и по сторонам
генитальной складки (см. рис. №1) с общей длиной 28 и 34 сантиметров, пигментированных
подобно остальному телу. Часть левого выпячивания, которая выдавалась из тела,
имела вид округлой лопасти пропеллера, тогда как правое напоминало плавник с
пальцевидными отростками. Кости были заключены в плотную соединительную ткань.
Судя по рентгеновскому снимку (см. рис. №2) скелета выпячиваний на этом образце,
часть выпячиваний, которая находится внутри тела, соответствует бедру, а средняя
часть большой и малой берцовым костям. Часть выпячиваний, которая выдавалась
из тела, соответствует ступне задней конечности, а элементы, составляющие её
– вероятно, фаланги пальцев. Согласно числу фаланг, видимых на рентгеновском
снимке, это пятый и четвёртый пальцы. Скелет этих конечностей отличается от
типичного строения скелета пятипалой конечности млекопитающих отсутствием элементов
плюсны. Возможно, однако, что они просто не были найдены из-за их небольшого
размера, слабого окостенения и обилия соединительной ткани» (Зембский, В. А.,
и Берзин А. А. (1961) «О редком явлении атавизма у кашалота». Научные Доклады
Высшей Школы. Серия «Биологические науки.» (1972), источник).
На сайте Эда Бабински приводится ещё целый ряд примеров того, на что похожи
рекапитулированные задние конечности китов. В лучшем случае это отростки, лишь
отдалённо напоминающие плавники. Обычно – просто бугорки с отдельными бесформенными
костями внутри.
Стр. 218:
Переход в водную среду вызвал, выработку гидроприспособлений. «К гидростатистическим
приспособлениям относятся все изменения, способствовавшие улучшению плавучести
тела: увеличение общей поверхности животного, появление губчатой структуры во
всех костях скелета, изменение ориентации конечностей по отношению к телу (горизонтальное
взамен вертикального)» (там же, с. 111).
У господина Тена кривая не только логика, но даже руки – он не может нормально переписать слова первоисточника. В книге Мчедлидзе есть слово «гидростатическим». А словесный урод «гидростатистические» придуман лично господином Теном. Редактор книги, вычитывавший рукопись – редкий болван.
Стр. 218:
Выходит, что предки дельфинов, имевшие массу, сравнимую с массой человека, в
воде начали грузнеть. В то же время у них менялась структура костной ткани,
становилась губчатой. Вспомним приведенные выше данные о слабости костяка человека,
который не соответствует условиям наземного обитания.
Самый плохой (для рассуждений Тена) момент
здесь в том, что «предки дельфинов, имевшие массу, сравнимую с массой человека»,
не найдены в ископаемом состоянии, не описаны и не получили названия. Тен говорит
о животных, которых нет.
Напомню, что я говорил об отношении массы скелета к массе тела. Уменьшить относительный
вес скелета в весе тела можно, не только уменьшая скелет, но и увеличивая тело.
Если посмотреть хотя бы на положение скелета дельфина или тюленя в силуэте его
тела, можно увидеть, какое большое количество мягких тканей, в том числе жира,
окружает скелет у этих животных. Повторюсь с рисунком тела и скелета гринды:

Если сравнивать качественные характеристики скелета, то видно, что кости человека – это нормальные кости наземного животного, способные выдерживать значительные нагрузки. Думаю, многие помнят рисунки из учебников, иллюстрирующие способность бедренной кости выдержать вес автомобиля. А «недостаточность» массы скелета человека, как уже говорилось в ходе анализа предыдущих глав, является лишь кажущимся явлением, связанным с наличием у человека значительного количества жира.
Стр. 218:
Чрезвычайный интерес представляет изменение ориентации конечностей у дельфинов
в качестве гидростатистического приспособления. Задние конечности заняли положение,
параллельное туловищу, еще у археоцет и именно в таком виде закладываются у
эмбрионов современных дельфинов. Впоследствии данное обстоятельство сыграет
важную роль в процессе антропогенеза. Еще Э. Харди считал, что вытянутое вдоль
тела положение ног было у предков людей изначально.
Это говорит о том, что у предков дельфина
конечности закладывались в эмбриогенезе таким образом. Но не говорит о том,
что именно в таком положении они были у взрослой особи.
У первых археоцетов вроде пакицетуса, амбулоцетуса, даланистеса и родоцетуса
(именно он с астрагалом, как у парнокопытных, напомню) пояс задних конечностей
сохранял связь с позвоночником, и конечности принимали участие в передвижении
по суше. Разворот конечностей назад возможен лишь у видов, которые уже не выходили
на сушу – например, у базилозаврид, у которых элементы таза уже не соединены
с позвоночником, а задние конечности меньше передних и превращены в плавники.
Если говорить о положении конечностей относительно туловища у зародышей китообразных,
то приведу вот это фото:
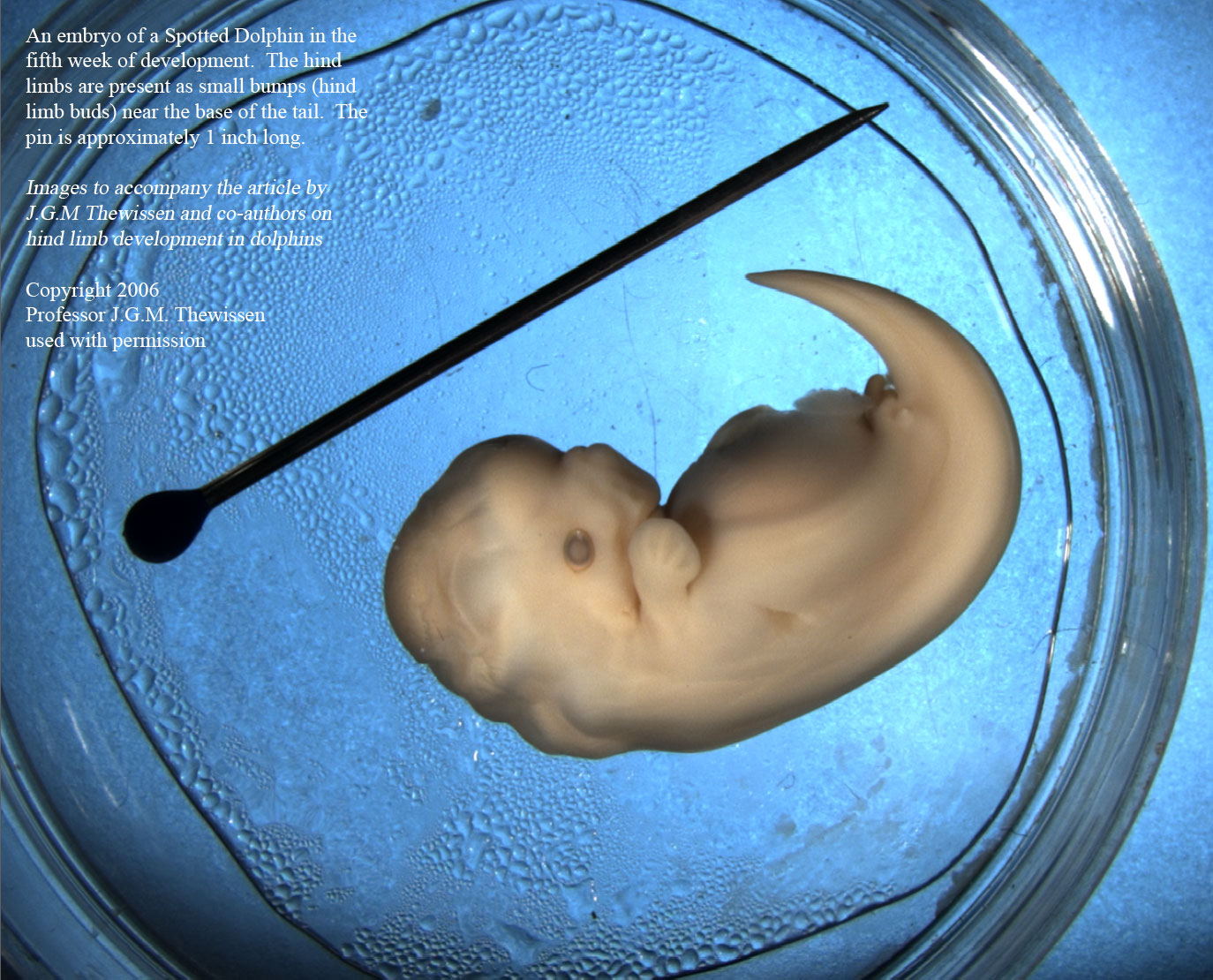
Зародыш дельфина
Думаю, это фото имеет достаточно хорошее
разрешение для того, чтобы каждый желающий смог оценить, каково положение конечностей
у эмбриона дельфина по отношению к позвоночнику. Ровно как у наземных млекопитающих,
не параллельно телу – не так, как утверждает Тен.
Для сравнения дадим фотографию зародыша человека:
Зародыш человека
Видно, что, несмотря на положение задних конечностей (ног) у взрослого человека параллельно телу, у зародыша человека они также находятся под углом по отношению к позвоночнику.
Стр. 218:
Еще одним важным гидростатистическим [sic – П. В.] приспособлением
является перемещение дыхательного отверстия в верхнюю часть головы. У предков
дельфинов имелось дыхало, расположенное, как у лемуров (и людей), спереди. При
этом, как уверяют палеонтологи, носовые кости нависали над дыхательным отверстием
(т.е. наземные предки дельфинов имели нос). У современных дельфинов одиночное
отверстие сверху. Перемещение дыхала вызвало изменение формы черепа: вытянутость
вперед оказалась заменена вытянутостью вверх. Сверху появился гребень. (Палеонтологи,
характеризуя верх черепа дельфинов, употребляют именно это слово, хорошо знакомое
также и антропологам.)
Перемещение ноздрей у китообразных –
это нечто большее, чем залепить дырку в одном месте черепа и провертеть её в
другом. Смещение ноздрей китообразных назад, на темя – сложный эволюционный
процесс, включающий перестройку всего черепа.
В очередной раз Тен неправильно использует термин «дыхало», который применяется
исключительно для описания ноздрей у современных китообразных – уже сдвинувшихся
на темя. У наземных предков китообразных дыхала не было, а были ноздри – просто
ноздри.
В процессе всё более глубокого приспособления к водному образу жизни носовые
отверстия китообразных смещались в сторону темени. Этот процесс начался ещё
у ранних археоцетов: даже у пакицетуса ноздри находятся уже не на самом кончике
морды, а у амбулоцетуса сдвинуты на границу передней трети черепа. Смещение
ноздрей на темя не завершилось даже в момент разделения усатых и зубатых китов,
и у каждой из групп после их разделения процесс смещения ноздрей на темя продолжался
независимо. В итоге у усатых китов дыхало образовано двумя сильно сближенными
ноздрями, а у зубатых осталась лишь одна ноздря (левая), а вторая заросла и
превратилась в хранилище для воздуха.
Достаточно взглянуть на череп современных зубатых китов, чтобы увидеть, что
он не очень-то и блещет «вытянутостью вверх». Голова дельфина может показаться
высокой, но эту иллюзию создают мягкие ткани.
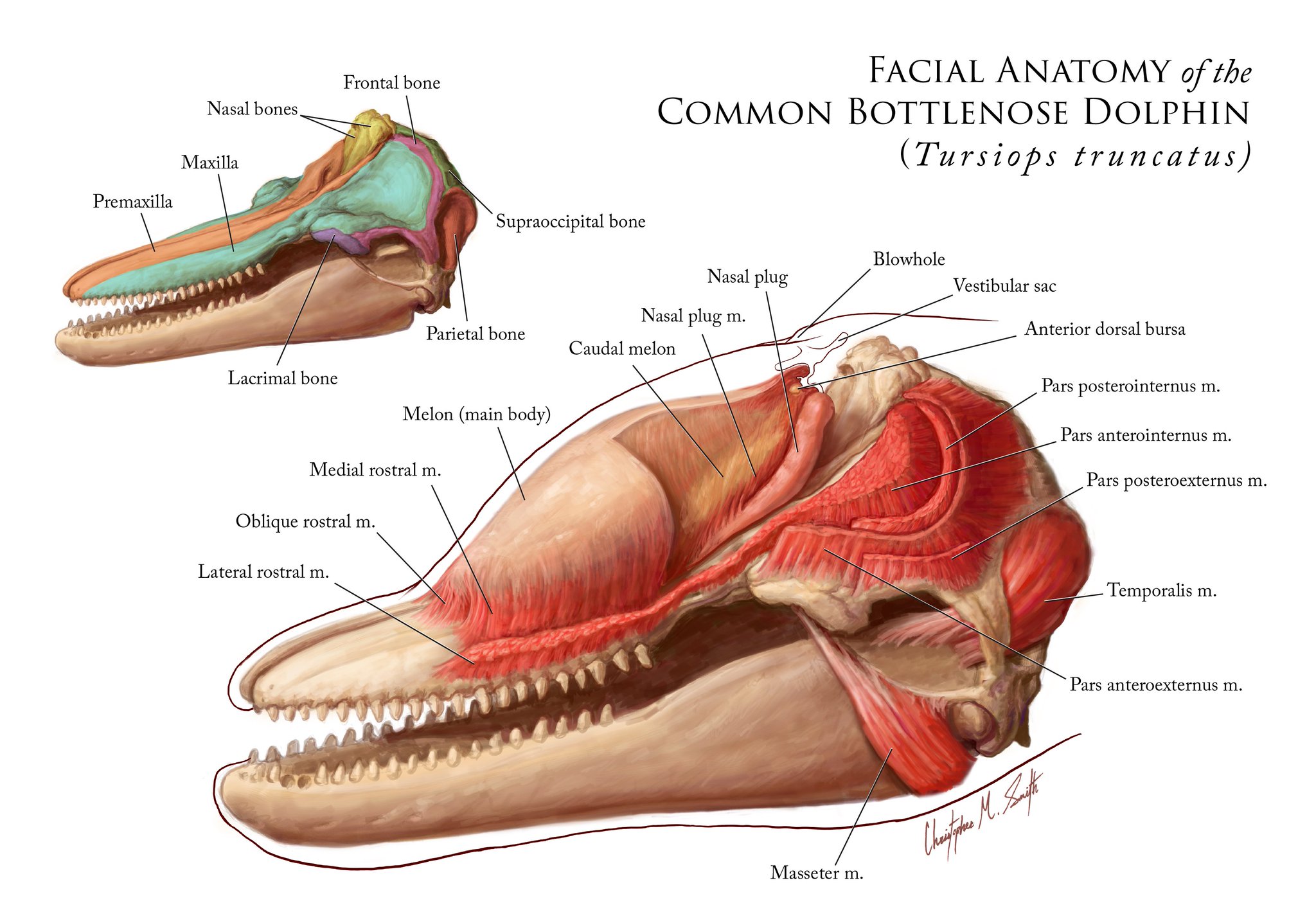
Схема строения головы дельфина афалины с мягкими тканями и очищенным от них черепом.
Стр. 218-219:
Отсюда своеобразная, никому более не свойственная в животном мире треугольная
форма черепа. Впрочем, аналогии существуют. Они известны среди тех существ,
кого принято считать предками или родственниками людей. Одним из самых архаичных
признаков в процессе гоминизации являются черепные гребни, которые отличают
массивных австралопитеков. Среди антропоидов гребни наличествуют у современных
горилл. Благодаря гребням черепа массивных австралопитеков и горилл имеют форму,
близкую к пирамидальной. Странно, что до сих пор никто не обращал внимания на
сходство внешних форм черепов древних гоминин, современных антропоидов и дельфинов.
Треугольная форма черепа – и никому более не свойственная? Да что вы говорите?

Череп кабана
Череп пекари

Череп жирафа

Череп тапира
Как видим, треугольная форма черепа довольно
распространена в природе. Однако показанные выше черепа треугольных очертаний
принадлежат существам, которых никогда не ставят в родословную человека. Кстати,
на некоторых из изображённых здесь черепов можно разглядеть хорошо выраженные
теменные гребни.
В связи с рассуждениями Тена возникает один интересный вопрос: какими костями
образован гребень у гоминид, и какими у дельфинов? У горилл и прочих приматов
сагиттальный череп образован теменными костями. У дельфинов теменные кости вообще
не соприкасаются друг с другом – их раздвинули в стороны соприкасающиеся друг
с другом верхнезатылочная и лобные кости. Поэтому сагиттальный гребень, гомологичный
таковому у приматов, у них не может образоваться в принципе. Какой-то другой
костный гребень вполне может появиться – и это имеет место, например, у кашалота,
череп которого зоолог Айвен Сандерсон сравнивал с ванной, у которой нет трёх
стенок. У него гребень – полукруглое поперечное поднятие по заднему краю черепа,
далёкое по форме и происхождению от сагиттального гребня на черепах приматов.
Это уже конвергенция – хорошо известное в биологии явление, когда у неродственных
видов возникают сходные внешне анатомические признаки. И, да, конвергентное
сходство не означает родства по определению.
Стр. 219:
Еще один интересный факт: древняя «подтреугольная» форма черепа с редким постоянством
рецидивирует у современных людей, страдающих микроцефалией (Домба, 1935). Это
говорит об эволюционной исходности данной формы в разрезе антропогенеза.
Это говорит о патологии, отличающейся от нормы для вида. Точно так же, как редкое заболевание «синдром Протея» не является доказательством происхождения человека от слона: такой диагноз имел знаменитый «человек-слон» XIX века Джозеф Меррик.
Стр. 219:
Известно, что у народов, относящихся к разным расам, встречается довольно обширный
набор генетических дефектов, проявляющихся внешне. При этом два признака встречаются
повсеместно, независимо от специфики генофонда: срощенные пальцы и триангуляционная
форма черепа. Вывод о рекапитуляции, о рудиментарности этих явлений просто напрашивается.
Или о том, что они могут быть обусловлены
редким сочетанием генов, которое время от времени всплывает. Или о том, что
различные нарушения в устоявшемся процессе онтогенеза приводят к внешне сходным
результатам разной природы.
Эволюция ласта у китообразных изучена, вот статья 2017 года об этом: “Review
and experimental evaluation of the embryonicdevelopment and evolutionary history
of flipper developmentand hyperphalangy in dolphins (Cetacea: Mammalia)” (ссылка
– статья в свободном доступе). Соединение пальцев китообразного в один плавник
– это результат не срастания, а неразделения эмбриональной конечности. У животных
с разделёнными пальцами после формирования в конечности эмбриона скелета пальцев
клетки в промежутках между пальцами подвергаются апоптозу (отмирают); ключевую
роль в этом процессе играют BMP – Bone Morphogenetic Proteins, костные морфогенетические
белки. Их антагонистами являются ингибиторы Gremlin и семейство белков, называемых
факторами роста фибробластов (FGF). Под действием этих антагонистов сохраняется
эмбриональная мягкая ткань между пальцами, превращая пятипалую кисть в ласт.
Схожим образом появляются перепонки в крыльях летучих мышей и в лапах утки.
Пальцы при этом формируются нормально, не срастаются. Увеличение количества
фаланг пальцев у китообразных связано с иными факторами.
Синдактилия у человека имеет много разных форм, включая срастание лишь некоторых
пальцев, либо всех пальцев, либо не только пальцев, но и костей конечности.
В статье “Webbed Fingers” (ссылка,
статья в свободном доступе) описаны эти несколько типов синдактилии у человека.
В статье 2012 года “The Epidemiology, Genetics and Future Management of Syndactyly”
(ссылка,
статья в свободном доступе) описаны механизмы формирования синдактилии у человека.
Это явление (наряду с полидактилией) связано с семейством межклеточных сигнальных
белков Hedgehog. В статье перечислены типы синдактилии и гены, с работой которых
связан тот или иной тип синдактилии. Среди «виновников» человеческой синдактилии
указаны гены Homeobox D 13, Fibulin 1, Gap Junction Protein Alpha 1, LMBR1,
LRP4, MF4. Одни из этих генов отвечают за формирование перемычки из мягких тканей
между пальцами, а другие – за слияние костей, и даже появление полидактилии,
а также исчезновение отдельных фаланг пальцев и нарушение порядка формирования
фаланг. То есть, синдактилия человека не имеет ничего общего с формированием
плавника китообразного. Рекапитуляцией здесь и не пахнет.
«Триангуляционная форма черепа»? Я не ослышался?
Триангуляция – это термин из архитектуры, геодезии, военного дела и радиолокации,
описывающий способы измерения геометрических величин и направлений. Есть триангуляция
в математике, психологии и социальных науках – также как метод измерений. Вполне
возможно, имелась в виду «триангулярная» (сиречь «треугольная») форма черепа.
Но слово написано, утверждено редактором и попало в печать. А я опять напомню
о склонности Тена поучать специалистов в области использования терминов, поэтому
с особым удовольствием плачу ему той же монетой.
Стр. 219:
Вообще, эмбрион дельфина удивительно похож на маленького человечка. Не последнюю
роль в поразительном сходстве играет наличие рук и круглой (!) головки. «Форма
черепа... в эмбриогенезе изменяется от округлой до вытянутой» – это научная
информация об эмбриогенезе дельфинов (Кузьмин, 1976, с. 23).
Опять же, это говорит о том, что у предков
китообразных у эмбрионов на определённой стадии развития тоже были круглые головы.
Облик взрослого животного может значительно меняться по сравнению с эмбрионом
и новорожденным детёнышем. В частности, очень характерным изменением является
удлинение головы из-за разрастания челюстей. А так-то даже эмбрион курицы отличается
очень крупной круглой головой.
Между прочим, В. Дольник в книге «Непослушное дитя биосферы» (есть в списке
использованной Теном литературы на стр. 365) отмечал одну общую особенность
облика неполовозрелых животных: он говорил, что у детёнышей животных морда более
укороченная, чем у взрослых, и что дети охотнее играют с игрушками животных,
несущими детские черты – крупную голову, укороченную и расширенную морду.
Стр. 220:
Предки дельфинов еще более походили на человека, чем эмбрионы современных видов
зубатых китов. У них, вне всякого сомнения, имелись руки. Как пишут палеобиологи,
«миоценовые дельфины обладали... длинными плечевыми костями» (Мчедлидзе, 1976,
с. 115). Это были довольно странные на вид существа. Представьте себе дельфина
с руками. Добавьте к этому ноги, обнаруженные у предков археоцет палеонтологами,
и способность дельфинов вертикально ходить по воде.
Не знаю, в каком объёме Тен использует
термин «дельфины» в данный момент. С учётом его невежественности и неразборчивости
логичнее вообще не делать оговорок на эту, хммммм…, «особенность» его стиля
изложения. Лично я при использовании термина «дельфины» подразумеваю, что данное
понятие включает представителей семейства Delphinidae, и в отдельных случаях
может относиться к речным китообразным семейств Iniidae, Platanistidae, Lipoditae,
Pontoporiidae, которых также именуют «речными дельфинами». Речные дельфины имеют
несколько иное происхождение, чем «истинные» дельфины семейства Delphinidae.
Но семейство Delphinidae чисто физически включает больше представителей, поэтому
здесь и далее под словом «дельфины» я буду подразумевать только виды семейства
Delphinidae.
Представители семейства дельфинов возникли в позднем олигоцене. Их предками
палеонтологи считают представителей семейства Kentriodontidae, которые уже являются
китообразными типичного облика, с полностью редуцированными задними конечностями,
не сочленённым с позвоночником тазовым поясом (также редуцированным), мощным
плавательным хвостом и обтекаемой формой тела. Если эти особенности их облика
кажутся Тену похожими на человеческие, то я не знаю, как это можно прокомментировать.
На фотографиях сам Тен, вроде, похож на человека.
Как бы ни выглядел автор анализируемой мной книги, в данный момент меня больше
интересует содержание его книги. Тену можно смело вручать «Шнобелевскую премию»
за открытие у дельфинов… рук! Что удивительно, специалист Мчедлидзе эти «руки»
не открыл, а профан Тен (о, счастливчик!) внезапно обнаружил, причём буквально
под носом у профи! Впрочем, вывод Тена хорошо подходит под выражение «В огороде
бузина, а в Киеве дядька». Из наличия у каких-то дельфинов длинных плечевых
костей ещё не следует автоматически, что эти кости находились внутри руки, а
не плавника. Не исключено, что эти кости были длинными только в сравнении с
ещё более укороченными плечевыми костями прочих китообразных, но не с руками
человека. Если развивать тему, то наличие у китообразных фаланг пальцев ещё
не означает, что у них руки с хватательными подвижными пальцами: эти фаланги
находятся внутри мягких тканей плавника, как рука в варежке.
Кстати, в миоцене у китообразных уже не было задних конечностей – лишь рудименты,
как у современных китообразных. Так что даже если бы у миоценовых дельфинов
внезапно были настоящие полноценные руки, из них получился бы не человек, а,
самое большее, русалка.
А пока перед нами в очередной раз ложная логическая связь.
Стр. 220:
Утрата рук стала для дельфинов своеобразной «трагедией», постольку поскольку
они до сих пор не могут отрешиться от того, что уже давно отсутствует. Так инвалид
чувствует отрезанную ногу.
«В конечностях китообразных – ластах – привлекают внимание две особенности:
1. стойкое сохранение ряда структур наземной многопалой конечности (включая
нервный аппарат) ... и 2. сохранение способности к использованию их в некоторых
поведенческих актах. Образно говоря, центральная нервная система современных
дельфинов «все еще помнит» о конечностях предков» (Василевская, 1984, с. 129).
Наверное, есть исследования учёных, которые
подтверждают «фантомные боли» дельфинов? Хотелось бы ознакомиться с ними. Пока
отсутствуют ссылки на данные исследования, слова Тена можно смело не принимать
во внимание, считая их домыслами.
Из приведённого отрывка работы Василевской можно узнать лишь об особенностях
строения передних конечностей китообразных, и о том, что они пока ещё могут
ограниченно использоваться этими животными. Всё. Описание эмоциональной составляющей
этого явления («до сих пор не могут отрешиться от того, что уже давно
отсутствует») полностью остаётся на совести Тена, поскольку
ссылок на какие-то научные работы, посвящённые эмоциональной стороне использования
конечностей китообразными, он не приводит.
Стр. 220:
Обратите внимание, речь идет не о копыте, а о многопалой конечности. Точнее
– о пятипалой, потому что все дельфины имеют пять лучей под кожей ласт.
Сочетание пяти пальцев и копыт демонстрируют нам архаичные копытные. Наличие копыта само по себе не диктует количество пальцев – у слона на ноге четыре-пять пальцев, у носорога и тапира по три, но у них у всех на кончиках пальцев есть копыта. Понимаю, что Тен в своих рассуждениях является пленником собственной ошибочной «кулачной» теории происхождения копыта. Просто лишний раз напомню, что копыто – это видоизменение всего лишь когтя, а не всей кисти или ступни, и потому наличие копыт и многопалая конечность – это не взаимоисключающие понятия.
Стр. 220:
Череп – та часть скелета дельфинов, которая пережила наибольшие метаморфозы
за время приспособления к жизни в воде через водно-земной этап. У самых ранних
предков дельфинов, обитавших на суше, имелись развитые лобные доли.
Подождите-подождите, мы о чём сейчас говорим – о черепе, или о мозге? Лобные доли – это элемент строения мозга (мягкого), а не черепа (твёрдого). Череп состоит из костей. В нём есть лобные кости.
Стр. 220:
Далее произошло следующее: «На начальных этапах приспособления к жизни в водной
среде лицевая часть черепа предковых форм значительно вытянулась в длину...»
(Мчедлидзе, 1976, с 30). (Еще одно эволюционное подтверждение того, что предки
дельфинов имели круглую голову.)
Из приведённой цитаты Мчедлидзе этого не следует. Это говорит лишь о том, что произошло относительное удлинение лицевой части черепа по сравнению с исходным состоянием, но не говорит о том, что это исходное состояние представляло собой именно круглую голову.
Стр. 220:
Этот гидродинамический сдвиг лицевой части повлек изменение краниальной части
черепа. Произошло сплющивание мозгового отдела, лобные доли оказались задвинуты
и сохранились только в виде небольших косточек.
Стоп! Лыко да мочало, начинай сначала.
У нас обсуждается мозг или череп? Лобные доли – это мозг. Части мозга сохранились
в виде косточек? Или речь идёт всё же о лобных костях?
Кстати, весьма существенным фактором, влияющим на форму черепа дельфинов, стала
эволюция эхолокационного аппарата. Именно он составляет значительную часть объёма
головы дельфинов, и его наличие заставляет череп дельфина уплощаться и становиться
вогнутым, а черепную коробку – ещё и сдвигаться назад внутри головы животного.
И эволюция эхолокационного аппарата делает череп зубатых китов асимметричным.
Стр. 221:
Но потом (!) дельфинам вдруг вновь понадобились лобные доли: произошло интенсивное,
в эволюционном смысле взрывное увеличение мозговой капсулы. Лобные доли вновь
появились в виде широкой полосы (там же, с. 115). Два этапа развития черепа
с их прямо противоположными тенденциями отражают этапы развития мозга, что крайне
интересно. Мы к этому еще вернемся.
На указанной странице ничего этого не
написано – я все глаза обсмотрел. Продолжив чтение книги Мчедлидзе в надежде
набрести на первоисточник вдохновения господина Тена, на стр. 124 у Мчедлидзе
я обнаружил вот что:
«Наличие полосы лобных костей на дорзальной поверхности черепа древних одонтоцет
связано с недостаточным еще увеличением поверхности контакта между лобными и
верхнечелюстными костями. У современных же форм наличие полосы лобных костей
обусловлено увеличением объема мозговой капсулы, вызванным, безусловно, прогрессивным
увеличением головного мозга».
Надеюсь, это именно тот отрывок у Мчедлидзе. В отличие от господина Тена, Мчедлидзе
отличает лобные кости черепа от лобных долей мозга.
Стр. 221:
Земноводные существа, с наступлением вод Тэтиса терявшие свои
лиманы, постепенно переходили на водно-земной образ жизни, а потом на водный
с периодическим возращением на землю для отдыха и деторождения.
Пришло время, когда им стало тяжело возвращаться на сушу, и тогда начался процесс,
который называется «эволюция археоцет». Начались гидростатистические и гидродинамические
приспособления, о которых говорилось выше. Однажды все эти замечательные адаптации
начали создавать проблемы для археоцет. Проблемы, связанные с неуклонной, происходившей
на протяжении всего кайнозоя, регрессией Тэтиса.
Эволюция археоцетов началась куда раньше
полного перехода к жизни в воде (у базилозаврид): с пакицетуса и плохо известного
гималайяцетуса. Это самые настоящие археоцеты, хотя внешне они сильно отличались
от китообразных в том виде, как мы привыкли их воспринимать в наши дни. Появление
приспособлений для жизни в воде началось буквально с первого шага в эту самую
воду: у пакицетуса кости были уже плотнее, чем у типичных наземных животных.
Дрейф континентов вызвал проблемы иного рода: Индостан и Африка своим движением
на север «закрыли» Тетис и вызвали поднятие горных массивов. Перераспределились
океанские течения, изменилось движение воздушных масс, и на Земле в целом поменялся
климат. Но это произошло уже тогда, когда археоцеты перешли к полностью водному
образу жизни, и на сушу уже не возвращались. Или жили на ней только после сильного
шторма – плохо и очень недолго.
Стр. 221:
Палеонтологам известны два семейства ископаемых археоцет: дельфиниды и акродельфиды
(еще недавно фигурировало третье – сквалодонтиды, но теперь они исключены из-за
акульих щечных зубов). В настоящее время акродельфиды известны только в качестве
ископаемых. От дельфинид произошли современные дельфины и косатки.
Бред. Эта характеристика процитированного
абзаца из книги господина Тена была дана мною не в порыве неконтролируемых эмоций,
а после внимательного, вдумчивого и многократного прочтения написанного
им текста. Данный абзац показывает, что господин Тен даже не обращался к справочной
литературе в процессе написания своей книги, а лепил, что бог на душу положит.
Напомню, я цитирую «расширенное и дополненное» издание 2019 года.
Начать следует с того, что среди одних только археоцетов семейств гораздо больше:
пакицетиды, амбулоцетиды, протоцетиды и ремингтоноцетиды, плюс базилозавриды
и кекенодонтиды. А сами археоцеты (Archaeoceti) – это самостоятельный подотряд
китообразных, хотя они считаются парафилетической группой. Если имеется в виду
семейство Delphinidae, то дельфиниды относятся к другому подотряду – зубатых
китов (Odontoceti). То есть, это заведомо не археоцеты. Если Тен подразумевает
под своими «дельфинидами» нечто иное, то пусть сам разбирается, определяется
и пишет конкретнее, не заставляя догадываться, что он имел в виду. К слову,
усатые киты – это тоже отдельный подотряд Mysticeti.
Не знаю, как самому Тену, а мне из приведённого отрывка неясно, куда же включили
сквалодонтид с их «акульими» зубами? Учёные относят их к подотряду зубатых китов,
в одно надсемейство с гангским дельфином. В подотряд археоцетов их никогда не
включали, потому и исключать не пришлось.
Далее у нас идёт самое интересное. Акродельфиды уже давно не признаются самостоятельным
семейством, а рассматриваются как сборная группа полифилетического происхождения
– то есть, как потомки не одного, а нескольких разных предков. Искусственность
выделения этой группы была доказана в 1988 году (C. Muizon. 1988. Le polyphyletisme
des Acrodelphidae, Odontocetes longirostres du Miocene europeen. Bulletin du
Muséum National d'Histore Naturelle, Section C Sciences de la Terre, Paleontologie,
Geologie, Minerologie 10:31-88, ссылка
(статья на французском языке)). После этого бывшие «акродельфиды» плавно «расползлись»
по другим семействам зубатых китов. Здесь и далее под «акродельфидами» я буду
понимать только представителей родов Acrodeiphis, Schizodelphis, Pomatodelphis,
Eoplatanista и Champsodelphis, которые включались в это семейство, пока его
выделение считалось обоснованным.
Данные по систематике излагаются по сборнику “Secondary Adaptation of Tetrapods
to Life in Water”, обзор “Evolutionary history of cetaceans: a review” (авторы
Ewan Fordyce и Christian de Muizon, ссылка).
В книге Мчедлидзе 1976 года издания, которой явно пользовался господин Тен,
акродельфид ещё признают самостоятельным семейством, но эти данные уже успели
устареть.
Наконец, современные дельфины и косатки не «произошли» от дельфинид – они являются
этими самыми дельфинидами, поскольку входят в семейство Delphinidae.
Стр. 221-222:
Между двумя семействами зубатых археоцет имеются солидные различия, касающиеся
прежде всего строения зубного аппарата. У дельфинид резко выросло количество
зубов в сравнении с наземными млекопитающими, но спецификация зубов была утрачена.
Не стало поверхностей, предназначенных для пережевывания пищи и дробления моллюсков,
и клыков. Все зубы у дельфинид (а также у современных зубатых китов) – резцы.
Дельфины не способны пережевывать пищу, они рвут добычу и тут же глотают. С
такими зубами можно быть только результативным морским хищником, потребление
растительной пищи и панцирных моллюсков исключено.
Сразу пошли «Сказки Венского леса» в
исполнении крыловского «Квартета»…
Кто же такие «дельфиниды» «по Тену»? Тен упорно не делает привязку этого термина
ни к одной научной работе, поэтому сложно понять, какой группе животных, признанной
специалистами, соответствует этот расплывчатый термин. Понять Тена несложно
– стоит ему перейти на конкретику, и его сразу же можно будет подловить на этом,
сравнивая признаки реального существа, указанного им в качестве предка, с человеком,
которого он объявляет потомком «дельфинид». Если имеются в виду дельфины семейства
Delphinidae, то к археоцетам они не относятся – это зубатые киты, другой подотряд
китообразных (если считать китообразных отдельным отрядом).
Какая, к чертям, «спецификация» зубов??? Спецификация
– это документ такой, устанавливающий некие требования (например, техническая
спецификация), или описывающий порядок выполнения какой-то работы. У нас что
– боженька внезапно потерял спецификацию, в соответствии с которой сотворял
китообразных? Если говорить об описании зубов китообразных, то здесь, думаю,
имеется в виду понятие «специализация». Зачем я заострил на этом внимание? Затем,
что Тен по ходу книги часто поучал своих оппонентов, как надо использовать тот
или иной термин. Это просто обратная связь, ничего личного.
У дельфинид не стало жевательных поверхностей? Берите выше, господин Тен! Их
не стало уже у археоцетов. У тех же базиозаврид зубы, хоть ещё делятся на резцы,
клыки и коренные, но уже приспособлены не для жевания, а для протыкания и разрезания
пищи. Дельфины унаследовали колющие зубы от своих предков.
Все зубы китообразных – резцы? Это неверно. У современных речных дельфинов семейства
иниид зубная система сохраняет гетеродонтность, и коренные зубы вполне различимы
среди прочих зубов, хотя близки по форме к зубам в передней части челюстей.
Это показывает, что зубы китообразных имеют разное происхождение, и однозначно
объявлять их зубами одного вида нельзя. Далее, зубатые киты не умеют рвать добычу
– их зубы не приспособлены для откусывания. По этой же причине их нельзя назвать
резцами – это колющие зубы, позволяющие лишь удерживать добычу, которую зубатые
киты глотают целиком.
Стр. 222:
Наибольший интерес для нас представляют акродельфиды, имевшие гетеродонтный
аппарат во рту. В отличие от своих гомодонтных родичей, акродельфиды имели не
только резцы, но и зубы с плоскими площадками. Общее количество зубов было не
увеличенным в сравнении с наземными животными.
Наконец-то, сподобилось! Пошла конкретика.
Вот только она какая-то ущербная. Акродельфиды с 1988 года не являются самостоятельным
семейством после упомянутой ревизии Christian de Muizon. Но у Тена они – живее
всех живых, как дедушка Ленин. Представители семейства известны по неполным
и плохо определяемым образцам, так что в целом о них можно сказать мало что
определённого. Систематическое положение известных родов акродельфид было пересмотрено,
и сейчас семейство сведено к одному неполному образцу с неясными диагностическими
признаками. Прочие роды в настоящее время относятся к иным таксонам. Вот, что
написано об этом в книге “Secondary adaptation to life in the water” (pp.169-233),
chapter: Evolutionary history of cetaceans: a review; Publisher: Pfeil Verlag,
Munich, editors: Mazin J. M & Buffrenil V. de, 2001:
Schizodelphis – переведён в семейство Eurhinodelphinidae;
Eoplatanista – выделен в собственное семейство Eoplatanistidae;
Pomatodelphis – теперь это представитель семейства Platanistidae, включающего
современных гангских дельфинов;
Champsodelphis – сомнительное название, часть образцов относят к роду Macrodelphinus
(семейство Eurhinodelphinidae), другие оставляют в этом роде, но его систематическое
положение неясное;
Acrodelphis – род неясного систематического положения, считается младшим синонимом
рода Champsodelphis. Только образец (частично сохранившийся череп) Acrodelphis
scheynensis может считаться относящимся к этому роду, но при этом сам род Acrodelphis
считается близким, или вовсе принадлежащим к роду Kentriodon (семейство Kentriodontidae).
Справедливости ради стоит отметить, что некоторые источники ещё используют название
семейства Acrodelphinidae, но считают его синонимом для семейства Eoplatanistidae
(например, здесь).
В общем, говорить как-то даже не о чем оказывается. Самостоятельного семейства
как такового нет, из-за чего проблема происхождения человека от китообразных
повисает в воздухе. Собственно, она и так там висела, ничем не подкреплённая.
Жевательных зубов у родов, ранее относившихся к акродельфидам, не обнаружено.
Стр. 222:
Интересно происхождение термина «акродельфид». «Акрон» – это конечность и одновременно
край. Последний смысл задействован в анатомическом термине «акромегалия», означающем
гиперразвитие рук у человека. Акродельфиды – это археоцеты с развитыми руками,
обитавшие на краю земли (у воды).
Термин акромегалия происходит от греческих
слов ἄκρος – конечность и μέγας – большой. Такая этимология приведена в «Википедии»,
и с ней согласна «Большая Медицинская Энциклопедия» (https://бмэ.орг/index.php/АКРОМЕГАЛИЯ).
Однако слово ἄκρος многозначное:
1) высший, верхний;
2) наружные, поверхностный;
3) крайний;
4) наилучший, превосходный, отличный (источник).
Поэтому название «акродельфиды» можно истолковать по-разному. Жаль, об этом
нельзя спросить описавшего род Acrodelphis палеонтолога Абеля – он уже умер.
Но в книге “Secondary adaptation to life in the water” упоминается, что представители
этого семейства отличаются длинным рылом (стр. 184). Вполне возможно, именно
эта особенность и послужила поводом для наименования рода и в дальнейшем семейства.
Тем более, в описании древнего кашалота акрофизетера авторы поясняют этимологию
названия, упоминая, что древнегреческое «акрос» означает «остроконечный». Это
очень хорошо соотносится с обликом длиннорылых существ, относившихся ранее к
акродельфидам.
Следует также помнить, что акродельфиды – не археоцеты, а представители подотряда
зубатых китов вне зависимости от того, являются они отдельным семейством, или
распределяются по иным семействам отряда.
Здесь я просто обязан напомнить утверждение Тена со стр. 84: «т.н. рабочая
рука была у наших предков изначально. Она – не результат, а предпосылка антропогенеза,
как и большой мозг, и прямохождение».
То есть, предсказание Тена говорит о том, что именно у этих китообразных должна
существовать «рабочая рука» гоминидного типа. Не забываем, однако, что передняя
конечность превратилась в плавник ещё у археоцетов эпохи эоцена. То есть, по
какому-то странному капризу эволюции, специализированный плавник предков «акродельфид»
должен переделаться обратно в руку с подвижными пальцами, и ещё на этих пальцах
должны появиться ногти! А как же закон Долло, который Тен использует в своих
рассуждениях (следовательно, признаёт его верность)? И, да, эта метаморфоза
должна была произойти ещё во время их жизни в воде.
А сейчас мы немного почитаем книгу Г. А. Мчедлидзе, но не «Основные черты палеобиологической
истории китообразных» 1976 года, которую использовал Тен, а чуть более раннюю,
«Ископаемые китообразные Кавказа» 1964 года. В ней приводится описание образцов
рода Champsodelphis, который по принятой на момент выхода книге систематике
относился к акродельфидам. На стр. 48 описаны образцы вида Champsodelphis sp.,
среди которых – передние конечности.
«Описанная кость [плечевая правой конечности – П. В.] обращает на себя внимание
и своими крупными размерами: длина ее равна 67,5 мм, ширина на дистальном конце
– 39 мм» (стр. 48).
«Плечевая кость левой конечности длинная, почти равная предплечью» (стр. 49).
Это к вопросу о «развитых руках» акродельфид. Вот, как выглядят эти образцы
в книге:

Таблица IX (Г. А. Мчедлидзе
«Ископаемые китообразные Кавказа», 1964)
1. Лопатка
2, 3. Скелеты передних конечностей.
Совсем как наши руки, верно? (да, это был сарказм). Именно так на самом деле выглядит передняя конечность акродельфида – та самая «рука», восхваляемая господином Теном. Сравним её со скелетом передней конечности современного дельфина:

Скелет кисти дельфина
Видно, что отличий не так уж и много, особенно в строении длинных костей (плечевая и локтевая + лучевая). Как выглядит плавник современного дельфина с мягкими тканями, думаю, объяснять не надо. Исходя из реальных особенностей строения скелета, «руками» у акродельфид, что называется, «не пахнет»: у них обычные плавники, мало отличающиеся от плавников дельфинов наших дней.
Стр. 222:
Гетеродонтность акродельфид говорит о том, что они пережевывали еду. Они занимали
экологическую нишу, которую оставили дельфиниды, устремившиеся в морские пучины.
Скорее всего нашими предками являлись акродельфиды, ибо они имели преимущественные
возможности для возвращения к наземному образу жизни, когда Тэтис начал усыхать,
превращаясь в цепь изолированных морских бассейнов. Один из последних останцов
некогда великого океана – Аральское море – исчез на наших глазах.
Способность выходить на сушу была утрачена
ещё у археоцетов – семейство базилозаврид в эоцене включает только полностью
водные формы. Они ещё сохраняли задние конечности в виде плавников, но их тазовые
кости начали редуцироваться и уже давно утратили связь с позвоночником, утеряв,
таким образом, пригодность для передвижения по суше. Все более поздние зубатые
киты, которые, по мнению специалистов, произошли от видов подсемейства дорудонтин,
также были полностью водными формами с редуцированными задними конечностями
и недоразвитыми тазовыми костями, не соединяющимся с позвоночником.
Соответственно, на сушу акродельфиды не возвращались – они чисто физически не
могли этого сделать.
Гетеродонтность как таковая – ещё не показатель способности жевать. У кошек
зубы тоже гетеродонтные, среди них чётко различаются резцы, клыки и коренные
зубы. Но коренные зубы у них режущие, приспособленные не для жевания, а для
разрывания плоти добычи. У видов, ранее относимых к семейству акродельфид, зубы
были гомодонтные, и жевать они не могли. Кроме того, длинный «клюв» видов, отнесённых
к акродельфидам, как-то не располагал к жеванию чисто физически, и указывает
на иной тип питания – это были хватальщики плавающей добычи в толще воды.
Стр. 223:
Особенно привлекательными были эти шхеры [вершины будущих гор Евразии – П. В.]
для акродельфид, которые, судя по гетеродонтному зубному аппарату, обожали именно
эту еду. Плоские зубы использовались не только для жевания, но и для раскалывания
мелких моллюсков. Этот прием известен с глубокой древности. […]
В отличие от дельфинид, акродельфиды предпочитали мелководье. Это были обитатели
шельфовой зоны. Для того чтобы поесть, им надо было плыть на мелководье, где
водились в огромном количестве моллюски и где была возможность время от времени
принимать вертикальное положение, чтобы прожевать пищу.
Сиреновые вынуждены жевать свою пищу
весь день, но для этого не всплывают на поверхность и движутся над дном, держа
тело горизонтально. Так что необходимость жевать не всегда связана с необходимостью
принимать вертикальное положение тела и всплывать к поверхности воды.
Я не буду уточнять, обладает ли господин Тен методикой, позволяющей реконструировать
с такой точностью поведение ископаемых животных (а он в своей книге неоднократно
делает заявления по поводу поведения доисторических животных). Вероятнее всего,
её попросту нет, и его слова о поведении акродельфид – чистейшая фантазия. Но
допустим, что они действительно принимали такое вертикальное положение тела
для еды. На будущем прямохождении гипотетических потомков этих существ такие
повадки не скажутся никак: задние ноги у всех китообразных к этому времени исчезли,
оставив лишь незначительные костные остатки в толще мягких тканей, не связанные
с позвоночником.
И повторюсь: у акродельфид были гомодонтные зубы и длинные челюсти, не приспособленные
к жеванию, и уж тем более – к питанию моллюсками с твёрдыми раковинами. Собственно,
посмотрим на картинку:

Челюсть Schizodelphis
А теперь смотрим на эту челюсть и показываем пальчиком: в какой стороне этой челюсти у нас находились жевательные зубы? Обращаем внимание на то, что справа у нас восходящая ветвь нижней челюсти – коренные зубы должны находиться ближе к ней. Итак, в каких же лунках сидели эти самые искомые жевательные зубы?
Стр. 223:
Черепа ископаемых акродельфид тоже отражают их какую-то сугубую историю на поздней
стадии развития.
Выше говорилось о том, что черепа всех археоцет в процессе адаптации к исключительно
водному существованию приобрели треугольную форму. И вдруг эта идеальная для
жизни в воде форма (идеальная тем, что позволяет иметь дыхало сверху) начала
преформироваться незадолго до исчезновения акродельфид.
«Среди акродельфид, – пишет Г. Мчедлидзе, – можно выделить ряд форм, отражающих
отдельные ступени сплющивания черепа...» (Мчедлидзе, 1976, с. 30). То есть стал
сглаживаться гребень.
Честно говоря, какая разница, что происходит
с черепом зубатых китов, если фактически он не играет практически никакой роли
в формировании внешнего вида животного? Он ведь буквально похоронен под толщей
мягких тканей головы, в том числе образующих дыхательные пути и эхолокационный
аппарат. Если начинает сглаживаться гребень, это означает эволюцию мягких тканей
над ним. Дыхало китообразного сформировано мягкими тканями, и у зубатых китов
положение дыхала и положение отверстий ноздрей мало связаны топографически –
их разделяет значительная толща мягких тканей, зачастую превышающая максимальную
высоту черепа.
Впрочем, усиление асимметрии черепа косвенно свидетельствует об усилении эхолокационных
способностей вида.
Стр. 224-225:
Наши предполагаемые предки акродельфиды имели много времени для того, чтобы
возвратиться на сушу из моря и освоиться на суше. Фактически это весь миоцен
длительностью 14 млн лет. Более чем достаточно, если учесть, что антропологи
в свое время за какие-то 0,7 млн лет из питекантропа Дюбуа «сделали» человека.
Последний ископаемый акродельфид имеет возраст 5 млн лет. Первый ископаемый
гоминин имеет возраст около 7 млн лет. Примерно два миллиона лет после появления
первых гоминин акродельфиды еще жили.
А вот теперь мы начнём удивляться. Сайт
Fossilworks
выдаёт нам время существования акродельфид (точнее – Acrodelphinidae) – от 23,03
до 7,246 млн. лет назад. Немного (на 2,24 млн лет) пораньше, чем их «хоронит»
Тен. С возрастом сахелантропа тоже получается неувязка: он существовал порядка
6-7 млн лет назад, чуть позже вымирания акродельфид, так что между ними обнаруживается
даже некий разрыв во времени существования.
Далее, «питекантроп Дюбуа» уже принадлежал к роду Homo. То есть, если бы он
был прямым предком человека, переход от него к человеку был бы гораздо меньшим
эволюционным шагом, чем путь от сахелантропа до человека. В случае гипотетического
перехода от китообразного к гоминиду изменения должны быть гораздо значительнее
– на уровне разных семейств, и даже более высоком. Хватит ли у них на это времени?
Терпение, скоро узнаем.
Стр. 225:
Толчком к очеловечению морских млекопитающих послужил процесс, обратный тому,
который привел предков акродельфид в море. Тех привела в море около 60 млн лет
назад трансгрессия Тэтиса, имевшая место в конце мелового периода. В течение
всего неогена Тэтис неуклонно сокращался из-за происходивших то тут, то там
поднятий земной коры.
Как говорится, не Тетисом единым… Тетис ещё существовал, а базилозавриды уже добрались до Северной Америки, где остатки тех же базилозавров довольно многочисленны. Так что киты просто ушли дальше в океан, не испытывая особых проблем. Пришлось, конечно, подстраиваться к изменениям океанских течений и искать новые источники пищи. Но никто из китов обратно на сушу не полез – ископаемых остатков, подтверждающих этот процесс, не найдено.
Стр. 226:
Эволюция акродельфид началась еще на пространствах мелеющего Тэтиса, от которого
ко времени начала, собственно, антропогенеза остались два больших водоема: собственно
Тэтис и Паратэтис, называемый также Сарматским морем.
[…]
Пропустим описание географии Тетиса, хотя могу предположить, что геологам нашлось бы, что сказать господину Тену.
Стр. 227:
В поисках места, где человечество начало быть, мы должны выбирать
между Средиземноморьем и Сарматским морем. Прошлое Средиземноморья удивительно
и трагично.
[…]
Восстановим картину произошедшего. Теряя связь с океаном через Гибралтар, Средиземное
море медленно превращалось в гигантскую теплую ванну с интенсивным испарением.
Вода буквально парила, в воздухе висел теплый морок. Дожди не компенсировали
ее расход на испарение. Чаша Средиземноморья была закрыта с севера высоким альпийским
поясом, из-за интенсивного испарения создавалась исключительно благоприятная
для жизни атмосфера умеренно теплой бани. Это та самая атмосфера, в которой,
учитывая особенности человеческого тела, оно только и могло сформироваться.
В период антропогенеза подобная атмосфера была только в Средиземноморье.
Очень интересно подумать о необходимости
появления потовых желёз у потомков китообразных в свете описания этой гипотетической
«бани». Тен правильно опасался, что его упрекнут в том, что у китообразных потовых
желёз нет, а у человека, внезапно, имеются. Здесь стоит уже в который раз вспомнить
закон Долло, который Тен неоднократно упоминает на страницах своей книги, тем
самым показывая, что знает его, и учитывает в рассуждениях. Нет, я понимаю,
что можно, конечно, знать, но не учитывать. Тем не менее, сейчас я хотел бы
почитать объяснение того, какие факторы в условиях описанной Теном парилки стимулировали
появление у человекообразных «потомков дельфинид» потовых желёз, и из каких
зачатков появились эти железы.
Ещё один интересный вопрос: сколько раз Средиземное море высыхало, и в какое
время это происходило? Ответ на этот вопрос легко ищется в Интернете, в
той же «Википедии». Это событие случилось 5,96-5,33 млн лет назад, геологами
признано 8 циклов пересыхания и возвращения моря. Промежуток существования акродельфи(ни)д
совершенно не совпадает со временем Мессинского кризиса: к этому времени они
успели вымереть. Хорошо, допустим, что акродельфиды существовали 5 миллионов
лет назад. Тогда на весь «цетоантропогенез Тена» у них было от 0,96 до 0,33
млн лет, ведь начался он не ранее начала этого кризиса и не позднее времени
вымирания последнего из акродельфид по версии Тена (не поддерживаемой наукой).
Стр. 229:
«Европа? Не может быть... Холодно». На самом деле в период антропогенеза нигде
в мире не было более благоприятных климатических условий, чем в Средиземноморье.
Условий, которые сочетали в себе эпохальную стабильность и поэтапную переменчивость,
которая – переменчивость – подстегивала и спасала от застоя.
Может, может, только успокойтесь, господин Тен. В кайнозое европейский климат был значительно теплее, чем сейчас. Люди более-менее сведущие в палеонтологии припомнят, что в Европе находят остатки обезьян и попугаев, не говоря уже о слонах и носорогах с бегемотами. Поэтому возражение воображаемого оппонента, приведённое Теном, явно дано тут «для красного словца».
Стр. 229:
Собственно, об антропогенезе надо говорить, на мой взгляд, только применительно
к мессинию. Обратите внимание: возраст самых древних гоминин полностью совпадает
с периодом мессинской регрессии. При этом надо учесть, что в течение по крайней
мере половины этого срока, т.е. более полумиллиона лет, в Средиземноморье существовали
не просто благоприятные, а сказочно благоприятные условия для предков людей
с их водно-земной организменной спецификой. Условия жизни для нашего предкового
вида были настолько «ясельными», настолько благоприятными, что на ум приходит
вмешательство Бога.
Возраст гоминин всё же несколько больше,
чем дата событий высыхания Средиземного моря. Воспользуемся английской
«Википедией» для того, чтобы увидеть, кого относят к подсемейству гоминин
– там список и систематика указаны полнее, чем в русской версии. Первый же упомянутый
в списке представитель, кениапитек (Kenyapithecus) имеет возраст 14 миллионов
лет. Мессинский кризис начался около 6 миллионов лет назад. Непорядок, несовпадение.
Может, мы слишком широко понимаем термин «гоминины»? Попробуем сузить его до
трибы Hominini, и даже до подтрибы Hominina. Но даже в этом случае грекопитек
(Graekopithecus) и сахелантроп (Sahelanthropus) оказываются древнее этого события:
их возраст 7,2 и 7 млн лет соответственно.
Есть ещё одно обстоятельство, которое сильно мешает принять умопостроения Тена
за истину. Дело в том, что на островах Средиземного моря в настоящее время находят
интереснейших представителей ископаемой кайнозойской фауны. Это карликовые слоны,
носороги и бегемоты. Это гигантские сони и зайцы. Это огромный лебедь и «холоднокровный»
козёл, впадавший в спячку. Пока Средиземное море высыхало, острова высились
среди рассола, как горы, и между ними появлялись участки сухой проходимой местности,
по которой расхаживали представители европейской и африканской фауны. Они забредали
на острова, а при подъёме уровня воды оставались в изоляции. И тогда на островах
происходили чудеса: гиганты становились карликами, а карлики, наоборот, гигантами.
Впрочем, эти чудеса закономерны и вполне объяснимы с позиций теории эволюции.
Думаем дальше. Если слоны, носороги и бегемоты задерживались на таких горах-островах,
это означает, что там была и питьевая вода, и возможность искупаться. То есть,
при повышении уровня моря такие острова сохраняли своеобразную «выборку» местной
фауны, и уже дальше в ход вступал отбор на выживание в специфичных островных
условиях.
А теперь вопрос. Если дельфино-антропоиды Тена, потомки его мифических «дельфинид»,
действительно существовали, почему они не попадали в эту «выборку»? Примерно
похожие фауны островных млекопитающих открыты на Крите, Кипре, Мальте, Сицилии
и других островах Средиземноморья. Островов достаточно много, и хотя бы на самых
крупных из них дельфино-антропоиды Тена должны были бы продержаться хоть какое-то
время, попав в отложения рек или прибрежных мелководий. Однако их не находят.
Стр. 229-230:
Геологи уверяют, что «период иссушения Средиземного моря и формирования здесь
мощной эвапоритовой толщи отвечает интервалу времени 0,5 млн лет» («Земная кора...»,
1982, с. 108). Общая продолжительность мессиния в два раза больше – 1 млн лет.
В первые полмиллиона лет и даже несколько далее соленость моря отнюдь не была
катастрофической, а иссушение создавало лиманы, шхеры, бухты, острова. Климат
скорее всего был почти неизменным благодаря «парилке» и высокой защите с севера.
Даже сейчас, если бы горная цепь, защищающая Средиземное море с севера, была
бы раза в два выше, там был бы другой климат – гораздо более теплый и ровный.
Еще одно важное обстоятельство: в Средиземноморье нет мангровых зарослей, затрудняющих
доступ к воде и делающих невозможной жизнь существ с длинными волосами.
Что удивительно, на стр. 64 Тен критикует
информацию о климате Африки, обрушиваясь с гневной критикой на «лживых» специалистов,
утверждая, что «пресловутая аридизация у антропологов находится всегда».
А здесь он же глотает данные по климату полным ртом, не задумываясь, верны ли
соображения геологов. Куда же делась такая яростная въедливость Тена на грани
параноидального недоверия? Или просто тогда речь шла о ненавистной обезьяне,
а сейчас – о «прельстивых и любовных» вымышленных «дельфинидах»?
Причём, что удивительно, и на стр. 64, и сейчас речь идёт об одном и том же
отрезке времени – чтобы в этом убедиться, можно посмотреть, о каких существах
идёт речь (сахелантроп, оррорин, ардипитеки – начало перехода от жизни в лесу
к жизни на открытых пространствах), и когда они жили. Время их существования
как раз совпадает со временем Мессинского кризиса Средиземноморья. Стало быть,
Мессинский кризис происходит на фоне того же самого по времени события аридизации
климата, да ещё и с похолоданием:
«Резкое похолодание и иссушение климата произошли в конце понтийского этапа.
Исчезли субтропические леса; их место заняли травянистые ландшафты, переходящие
на востоке в полупустыни. По оценкам палеоботаников, температуры января могли
опускаться до - 5…- 10 °C. Примерно 6 млн лет назад тектонические процессы привели
к тому, что Средиземноморский бассейн отделился от Атлантического океана. Образовавшийся
водоем получил название Мессинского. В этом замкнутом бассейне шло интенсивное
накопление солей. Мощные слои соли и гипса были обнаружены глубоководным бурением
и эхозондированием в различных частях бассейна, в основном в пределах глубоководных
впади и. Это событие известно в истории Средиземного моря как мессинский кризис
солености».
«На осушенной поверхности дна сформировался рельеф пустынного типа. Климатические
условия в то время отличались повышенной аридностью во всех частях Средиземноморского
бассейна. Вечнозеленые леса, существовавшие в раннепонтийское время, сократились.
Из животных широко распространились обитатели сухих саванн. Мессинский этап
продолжался всего около 500 тыс. лет» (источник:
Долуханов П. М. «История средиземных морей», М., «Наука», 1988).
Прошу прощения у господина Тена за то, что этой неосторожной цитатой разрушил
его утверждение о «парилке». И в свете этого примера видно избирательное отношение
Тена к научным данным – когда выгодно, они признаются и применяются, а когда
невыгодно – становятся объектом беспощадной (но пустой) критики.
Кстати, возникает вопрос: могут ли те самые изменения, которые столь красочно
расписывает господин Тен, произойти за примерно 500 тысяч лет этого кризиса?
Стр. 230-231:
[…]
Дно Средиземного моря превращалось в топкий солончак. Благодать обернулась катастрофой.
Земной рай превратился в гиблое место. Гоминины, потомки акродельфид, начали
уходить туда, где вода была менее соленой. Уходили на лиманы Сарматского моря
и далее, где находят останки гоминин возрастом примерно 2 млн лет (Дманиси на
Кавказе). Однако раньше и больше уходили на юг по течению Нила. Это вообще очень
показательно. Самые древние гоминины (сахелантроп и др.) найдены в районе озера
Чад. Менее древние – 4 и менее млн лет – в районе Великих африканских озер.
Нил 7 млн лет назад вытекал из озера Чад. Изменил течение, стал вытекать из
Великих африканских озер – и гоминины стали приходить сюда, и их находят здесь.
Это подтверждение излагаемой здесь гипотезы.
Тпрууууууу… В очередной раз придержите
коней, господин Тен! Где-то 5,33 млн лет назад случился Занклийский потоп, воды
Атлантики пробили путь в высохшую котловину Средиземного моря, и море очень
быстро вернулось в свои берега. Мессинский кризис закончился менее чем за полмиллиона
лет! Местность перестала быть «гиблой» задолго до времени, которое нам указывает
Тен.
Возраст дманисского человека насколько меньше, чем сказано у Тена: во 2-м томе
книги Ст. Дробышевского «Достающее звено» (стр. 174) указывается возраст 1,75-1,85
млн лет. Это ещё больше отдаляет его существование от времени Мессинского кризиса.
7 миллионов лет назад Нил мог вытекать откуда угодно: может быть, даже из озера
Чад. Есть другая проблема: на север он начал течь только во времена Мессинского
кризиса, который был, напомню, 5,96-5,33 млн лет назад. Эта фаза существования
реки называется «фаза Эонила» (источник: “Structural influence on the evolution
of the pre-Eonile drainage system of southern Egypt: Insights from magnetotelluric
and gravity data”, ссылка,
статья в открытом доступе). Поэтому добраться по нему из Средиземного моря до
озера Чад 7 миллионов лет назад никто не мог бы чисто физически. Стало быть,
сахелантроп, который ещё и жил до Мессинского кризиса, не был «беглецом» из
Средиземноморья.
Не забывает также, что сахелантроп сильно отличается от человека: если применять
логику господина Тена и признавать его «деградантом», то у него должно быть
дополнительное время на «деградацию» из исходного человекоподобного состояния.
В связи с описанным Теном расселением его гипотетических «потомков дельфинид»
следует упомянуть ещё одну тонкость. Тен говорит о расселении вдоль рек и жизни
у озёр. А ведь эти места очень благоприятны для захоронения ископаемых остатков
живых организмов, особенно окружающие болота. В водоёме больше вероятность попадания
тела погибшего животного в бескислородную среду, или хотя бы быть скрытым от
крупных хищников и в дальнейшем погрузиться в ил.
Однако, несмотря на такие благоприятные обстоятельства, странных дельфино-человеков
Тена в соответствующих по природе и возрасту отложениях не находят.
Стр. 231:
Но в Африке и на Кавказе, в полном отрыве от моря, без морской пищи, в условиях
дефицита йода и витамина D, началась деградация гоминин в обезьян. В людей превратились
те – самые сильные, изгонявшие остальных, – кто остался жить в устьях рек: европейских,
переднеазиатских, Нила. Кроме того, шансы были у тех, кто ушел берегом Красного
моря и дальше на юг Африки и в Азию – по побережьям.
Если посмотреть на карту распространения йододефицита, то мы увидим, что районы сильного дефицита йода далеко не всегда совпадают с областями находок ископаемых гоминид – предположительных «деградантов».
Карта распространения йододефицита
Внезапно обнаруживаются интереснейшие
вещи: значительное количество районов Африки относится к местам, где в организм
поступает достаточное, и даже избыточное количество йода!
Полезно помнить также, что в человеческом организме витамин D образуется в коже
под воздействием солнечного света. Поэтому представить себе гоминид, шагающих
целый день под африканским солнцем и испытывающих дефицит этого витамина, довольно
сложно. Понимаю, что есть Европа с пасмурными весной и осенью, и с холодными
короткими зимними днями – там опасность рахита вполне реальна. Но в Африке?
И на Кавказе, в горах, где недостатка в солнечном свете не наблюдается?
На Кавказе один из людей, найденных в Дманиси (образец D3444/D3900) – это старичок,
который дожил до весьма преклонных лет, лишился значительной части зубов, и
даже его нижняя челюсть начала истончаться. Такая продолжительность жизни –
это признак деградации?
Кроме того, признаки строения скелета древних людей не совпадают с симптомами
рахита – я уже говорил об этом, когда обсуждал аргументы Тена на стр. 187 его
книги.
В свете этих данных жёсткая «привязка» происхождения человека к прибрежным морским
районам представляется необоснованной.
Стр. 231:
И все же можно уверенно говорить, что в мессинский период было и место, и время
для протекания процессов антропогенеза в тепличных условиях. Полмиллиона лет
– временной минимум существования «условий наибольшего благоприятствования»
– это 25 тысяч (!) поколений современных людей. Со времени Христа прошло всего
100 поколений. Это достаточный резерв для эволюции.
Вспоминается фраза из еврейского анекдота:
«А что вам мешает говорить то же самое?» Уверенно говорить можно о
многих вещах. Можно даже выучить клингонский язык или наречие на'ви, и говорить
на этих вымышленных языках. Но пока технологии не дошли до того, чтобы менять
объективную реальность в соответствии с нашими словами. Поэтому Тен может писать
сколько угодно книг о своих «дельфинидах» и их разумных потомках. Пока не найдено
материального подтверждения его слов, все его книги не стоят и одной косточки
гоминида, найденной в Африке. Главное – писать книги красивым слогом, чтобы
раскупались, и избавляться от фактических ошибок, чтобы не попасться на ляпах
в элементарных вещах.
Итак, условия были не «тепличные», а суровые, аридные. Времени у гипотетических
«дельфинид» Тена было значительно меньше – всего лишь 630 тысяч лет. Но анатомические
преобразования, которые они должны были претерпеть, соответствуют уровню перехода
от отряда к отряду. В природе на это требуется несколько миллионов лет: формирование
предков современных отрядов млекопитающих заняло весь палеоцен и часть эоцена.
Так что «дельфиниды» Тена должны были эволюционировать в условиях жёсткого цейтнота.
Кроме того, не количество поколений определяет скорость эволюции, а скорость
изменения окружающих условий (как фактор отбора) и скорость появления и распространения
мутаций (как поставщик материала для отбора). В стабильных условиях среды хоть
сто двадцать пять тысяч поколений может пройти, а вид будет находиться в условиях
действия стабилизирующего отбора и не изменится ни на йоту. Облик рачка щитня
деликатно намекает нам, что в стабильных условиях и больше времени может пройти,
а внешний вид всё тот же едва ли не с самого триаса.
Стр. 231-232:
По мере усыхания моря гоминины были вынуждены концентрироваться в тех местах,
где еще сохранялись благоприятные условия для развития. Слабых убивали или изгоняли.
В свою очередь, изгнанные старались следовать по воде. В основном, конечно,
уходили в Африку: там было тепло и не приходилось перебираться через высокие
горы с их ледниками. Там их, этих изгоев формировавшегося человечества, и находят,
каждый раз делая сенсацию из отбросов антропогенеза.
Вот и я про то же: острова Средиземноморья – прекрасное изолированное и защищённое от бед с материка убежище для реликтовых «дельфино-гоминид» Тена, куда лучше, чем полная хищников континентальная Африка или Европа. Вместе с карликовыми слонами, оленями ростом с пуделя, а также с гигантскими сонями и зайцами, они вполне могли бы дожить хотя бы до эпохи Древнего Египта или античности, чтобы их привозили к царскому двору как редкую диковинку. На худой конец, их могли бы убивать и поедать первые люди-поселенцы, и их кости лежали бы в кухонных кучах людей со следами разделки, как кости карликовых слонов или козликов-миотрагусов. А нету.
Стр. 232:
Наиболее вероятный вариант эволюции видится в задержке развития археоцет на какой-то стадии онтогенеза, что позволило им сохранить универсальность.
Какие, к едреням, археоцеты? Мы о каком
отрезке времени говорим, вообще? Сначала Тен утверждает, что эволюция человека
из его мифических китообразных предков началась во времена Мессинского кризиса
– это было, напомню, 5,96-5,33 млн лет назад. Затем в рассуждение в качестве
предков человека вводятся археоцеты. А они (в понимании этого термина, принятом
наукой) существовали примерно 50-23 млн лет назад и вымерли в позднем олигоцене.
Мессиний – это конец следующей геологической эпохи – миоцена. Тогда археоцетами
на Земле уже не пахло.
Если Тен принимает для своих рассуждений термин «археоцеты» в каком-то собственном
оригинальном понимании, то это – его собственные проблемы. Опять же, это оригинальное
понимание у него ничем не обосновано.
Стр. 232:
Речь идет о том, что имело место развитие по типу педоморфоза. В таком случае
надо говорить о ретардации и неотении. Ретардацией называется эволюционная задержка
развития на определенной стадии. Неотенией называется способность к размножению,
к воспроизводству вида на стадии «детскости». В целом педоморфоз – это развитие
по детскому типу.
В таком случае возможны три варианта. Первый и второй: предки человека представляли
собой неотеническую форму дельфинид или акродельфид; третий вариант – акродельфиды
и есть способные к размножению дети дельфинид.
В целях однозначности понимания терминов
стоит дать их определения. Согласно «Биологическому энциклопедическому словарю»
(М., «Советская Энциклопедия», 1989, стр. 538), ретардация – это «замедление
темпов эмбрионального развития или запаздывание эмбриональной закладки органа
у потомков по сравнению с предками».
Неотения – это «задержка онтогенеза у нек-рых видов организмов с приобретением
способности к половому размножению на стадии, предшествующей взрослому состоянию.
У животных при полной Н. лишь половая система личинок достигает уровня развития,
характерного для взрослых организмов, а др. системы органов сохраняют личиночное
состояние. При неполной (частичной) Н. личинки длит. время растут, не приступая
к метаморфозу, но способности к размножению не получают» (там же, стр. 402).
Наконец, педоморфоз – это «способ эволюц. изменений организмов, характеризующийся
полной утратой взрослой стадии и соответствующим укорочением онтогенеза, в к-ром
последней становится стадия, бывшая прежде личиночной» (там же, стр. 455).
Для полноты картины мы должны определить также понятие «личинка»:
«… постэмбриональная стадия индивидуального развития мн. беспозвоночных и нек-рых
позвоночных животных (рыб и земноводных), у к-рых запасы питат. веществ в яйце
недостаточны для завершения морфогенеза» (там же, стр. 326)
Никого не насторожило это определение? В нашем случае речь идёт о позвоночных
животных – млекопитающих: если кто забыл, и китообразные, и приматы являются
млекопитающими. Но в определении они не перечислены. Из этого можно сделать
один простой вывод. А именно: явления неотении и педоморфоза не применимы в
данном случае, поскольку они относятся к животным, у которых в онтогенезе есть
личиночная стадия. В нашем случае у млекопитающих её нет. Остаётся ретардация.
Результатом данного процесса действительно может быть сохранение у организма
во взрослом состоянии некоторых зародышевых признаков, и в данном случае наличия
личиночной стадии не требуется. В случае позвоночных «неотенией» часто называют
частичное сохранение во взрослом состоянии ювенильных признаков.
Все «три варианта» эволюции китообразных в человека, предлагаемые Теном, отдают
неким сюрреализмом. Но понятно, в принципе, к чему клонит автор. Если выводить
человека от китообразных предков, необходимо каким-то образом объяснить появление
у людей, как минимум, ног – у китообразных с этим признаком как-то не сложилось
уже несколько десятков миллионов лет. А у зародышей китообразных задние конечности
начинают закладываться на одной из ранних стадий развития – поэтому, думаю,
Тен пытается «притянуть» упомянутые выше эволюционные явления к своим умопостроениям.
Но эти явления, как и явление рекапитуляции – не «палочка-выручалочка», поскольку
на саму возможность рекапитуляции признака во всей его полноте накладываются
некоторые другие обстоятельства, очень интересные в нашем случае.
Если Тен выводит человека от китообразных уровнем развития выше, чем примитивнейшие
археоцеты (а дельфиниды в научном смысле этого слова и акродельфиды в ранее
принимавшемся объёме группы к археоцетам не относятся), придётся задумываться
о том, как из плавника постоянноводного существа может обратно получиться конечность,
пригодная для передвижения по суше. Ну, и там всякая прочая мелочёвка типа обратного
деления позвоночника на грудной, поясничный и крестцовый отдел, разделения срощенных
шейных позвонков, и ещё прикрепления к позвоночнику тазовых костей, которые
успели в процессе эволюции от него отделиться, да ещё «до кучи» сильно редуцироваться.
Нужно придумать способ появления наружного уха, да и с гиперфалангией надо что-то
делать – у человека количество фаланг пальцев стандартно для наземных животных,
а у китообразных из различных систематических групп успело изрядно увеличиться.
Хорошо, представим себе, что имеет место ретардация, и зародыш китообразного
в ходе эмбрионального развития задерживает развитие задних конечностей вплоть
до родов. Попробуем предсказать результат этого мысленного эксперимента. Но
для этого нам полезно будет узнать, каким же именно способом киты лишились своих
задних конечностей. Благодаря современным технологиям изучения активности генов
в процессе эмбриогенеза этот процесс хорошо изучен.
«Учёные говорят, что постепенное сокращение размеров задних конечностей китов
на протяжении более чем 15 миллионов лет было результатом медленно накапливающихся
генетических изменений, которые оказали влияние на размер конечностей, и что
эти изменения произошли на какой-то поздней стадии развития, во время эмбрионального
периода.
Однако настоящая утрата задней конечности случилась на гораздо более поздней
стадии эволюционного процесса, когда произошло коренное изменение, инактивировавшее
ген, чрезвычайно важный для развития конечности. Этот ген, называемый Sonic
hedgehog, функционирует во время первой четверти беременности в ходе эмбриональной
стадии развития животных, перед стадией плода.
Всем позвоночным, имеющим конечности, ген Sonic hedgehog необходим для развития
нормальных конечностей ниже локтевого и коленного суставов. Поскольку задние
конечности древних китов оставались прекрасно сформированными до самых пальцев,
даже когда они уменьшились в размерах, позволяет предполагать, что Sonic hedgehog
по-прежнему работал над формированием скелета конечностей.
Новое исследование показывает, что ближе к концу этих 15 миллионов лет, когда
задние конечности древних китов уже были нефункциональными, но по-прежнему сохранялись,
явно сыграло свою роль отсутствие гена Sonic hedgehog. Хотя у животных по-прежнему
могли развиваться эмбриональные почки задних конечностей, как это бывает у современных
дельфинов стенелл, у них не было гена Sonic hedgehog, необходимого, чтобы отрастить
конечность полностью или даже частично, хотя он активен в других местах эмбриона.
Команда также показала, почему ген Sonic hedgehog стал неактивным, и все следы
задних конечностей исчезли в конце этой стадии развития кита … Было выяснено,
что ген под названием Hand2, который обычно функционирует как кнопка для запуска
гена Sonic hedgehog, в почках задних конечностей дельфинов, бездействовал. Без
него развитие конечности попросту останавливается» (источник:
статья “How ancient whales lost their legs, got sleek and conquered the oceans”).
В редукции тазового пояса китообразных были также задействованы иные гены, отвечающие
за формирование костей таза, их пропорции, лобковый симфиз, окостенение большой
и малой берцовых костей и пр. Изменения в их активности приводили к появлению
различных отклонений в развитии тазового пояса (источник:
Павел Гольдин «Naming an Innominate: Pelvis and Hindlimbs of Miocene Whales
Give an Insight into Evolution and Homology of Cetacean Pelvic Girdle»).
Бездействие гена обычно связано с мутациями, которые портят его, делая нефункциональным.
Но в данном случае «испорченный» ген привёл к получению эволюционного преимущества
– формы тела, более приспособленной для движения в воде.
Стр. 232:
В таком случае становится понятно, почему они исчезли, хотя были прекрасно приспособлены
к обитанию на мелких акваториях. При их пристрастии к отмелям сокращение глубоководной
акватории было «на руку». Они ж исчезли. Почему? Почему этот многообещающий
вид вдруг исчез именно тогда, когда условия стали наиболее благоприятны для
его развития? Данное предположение антиэволюционно. Скорее всего акродельфиды
не исчезли, а продолжились в ком-то. И почему бы не в нас?
У меня, конечно, есть ответ на вопрос
Тена – на тот, который повыше, конечно. Но он ему точно не понравится, потому
что этот ответ: «потому что они даже не появлялись!» Правда, с точки зрения
логики, то, что не появлялось, исчезнуть не может.
Исчезновение вида, приспособленного к обитанию в некоторых условиях, может быть
связано с множеством факторов – хотя бы с наличием конкурентов, которые могли
просто вытеснить рассматриваемую нами группу живых организмов из облюбованных
ею мест обитания.
Если же условия были благоприятны и не менялись, как же тогда будет меняться
вид – в стабильных-то условиях? Он просто попадёт под действие стабилизирующего
отбора, который будет сохранять его в сравнительно неизменном виде, отсекая
все отклонения от удачной хорошо приспособленной формы. Именно в стабильных
условиях обитают «живые ископаемые», удачно занимающие свою экологическую нишу,
неизменную уже миллионы лет. Если всё же представить себе этих самых «дельфинид»
Тена, плещущихся в своих гипотетических благоприятных для их жизни водоёмах,
то с каких щей они должны внезапно превращаться в кого-то другого? Их физиология
и образ жизни прекрасно приспособлены именно к данным условиям. Ни у одного
вида нет некоего заложенного богом/природой/инопланетянами «стремления» превращаться
в другой вид. Сподвигнуть на превращение «в нас» мифических «дельфиноидов» Тена
должно именно изменение среды. Тем самым оно запускает движущий отбор, который
способствует именно накоплению изменений, ведущих эволюцию вида в ту или иную
сторону.
Стр. 232-233:
Данные биологии говорят о том, что у дельфинид вполне могла быть неотеническая
форма. Это связано с особенностью деторождения у зубатых китов, которая называется
«обратный ход новорожденных».
[…]
Обратный ход новорожденных дельфинов представляет собой чрезвычайно сложное
гидростатистическое [sic – П. В.] приспособление.
Я специально провёл пальцем по странице
книги, перечитав это слово несколько раз. Нет, сканер не ошибся, и программа
распознавания текстов сработала без ошибок. Такое слово есть в книге, причём
появляется в тексте уже не первый раз. Не знаю, кто породил этого «словесного
урода»: сам господин Тен, или же господин ответственный редактор В. Ильин из
издательства «Эксмо». Но хрен редьки не слаще.
Не могу сказать, при чём тут статистика, и уж тем более – «гидростатистика».
Я вообще не понимаю, чем занимается «гидростатистика» как таковая. Естественно,
я понимаю, что это может быть описка, и автор имеет в виду слово «гидростатическое».
Гидростатика – это раздел механики жидкости и газов, изучающий свойства неподвижной
жидкости. Какое отношение имеет эта область знания к реальным и гипотетическим
китообразным и их анатомо-физиологическим приспособлениям – ума не приложу.
Стр. 233:
У всех наземных млекопитающих дети выходят из матки головой вперед. Это обеспечивает
значительную выживаемость при длительных родах. У большинства хищников, имеющих
возможность рожать в достаточно комфортной обстановке и при этом не истощенных,
смертность при родах практически отсутствует. У зубатых китов, хорошо обеспеченных
пищей (они – самые эффективные хищники моря, практически не имеющие конкурентов),
рожающих в защищенной среде (безопасность роженицы самоотверженно и эффективно
обеспечивает все стадо), высокий процент смертности новорожденных. При затяжных
родах дельфиненок задыхается в чреве, не успевая выйти. Это расплата за такое
эволюционное приспособление, как обратный ход.
Никому не приходило в голову, что китообразные просто живут в среде, изначально враждебной для дышащих воздухом животных? Я не спорю, у любого вида существует определённый процент смертности при родах от разных причин. Однако господин Тен не ссылается при этом ни на один источник, поэтому не представляется возможным ни подтвердить, ни опровергнуть его слова. Однако по этой же самой причине его слова не вызывают доверия – он сам не специалист в области биологии, явно не изучал причины смертности при родах у китообразных и у наземных животных, и не приводит никаких конкретных цифр и работ специалистов. Учитывая огромное количество ошибок в его работе, я не рекомендовал бы доверять его утверждениям, пока они не будут подкреплены научными работами.
Стр. 233:
Как считают палеобиологи, древние дельфиниды очень долго были вынуждены возвращаться
на какие-то постоянные нерестилища на берегу (Мчедлидзе, 1976).
Хотелось бы, чтобы господин Тен любезно сообщил номер страницы в книге Мчедлидзе, где есть слово «нерестилище». Обычно этот термин применяется к рыбам и земноводным, которые размножаются, вымётывая икру.
Стр. 233-234:
Такое эволюционное приспособление, как изменение положения младенца в матке,
позволяющее рожать в глубоких водах, не могло свершиться в один миг. Кого они
рожали – это тоже вопрос. Развитие эмбрионов дельфинов свидетельствует о том,
что их отпрыски долго сохраняли руки, ноги и круглые головы, что делало их похожими
на нас. Во всяком случае, дети дельфинид имели такой вид до тех пор, пока эволюция
не изменила ход новорожденных на обратный, сделав возможными роды в глубоких
водах. До тех пор скорее всего взрослые и детские особи дельфинид различались
между собой внешне, как различаются взрослые и детские особи лягушек, стрекоз,
саламандр. Выползая рожать на берег, археоцеты рожали существ, способных кормиться
на берегу и на мелководье. Их дети были похожи на их далеких предков, когда
предки дельфинид обитали в двух средах.
Остатки майяцетуса (Maiacetus inuus)
из семейства протоцетид (относившегося к археоцетам) позволяют нам увидеть,
какими были детёныши археоцетов. Образец самки (номер GSP-UM 3475a), погибшей
во время или незадолго до родов, был обнаружен в формации Хабиб Рахи в Пакистане
(средний эоцен). Он позволяет увидеть, что детёныш (образец GSP-UM 3475b) выходил
из родовых путей самки головой вперёд, уже при рождении обладал набором молочных
зубов и был вполне похож на родителя, за исключением неизбежного наличия признаков,
присущих всем детёнышам млекопитающих (источник:
“New Protocetid Whale from the Middle Eocene of Pakistan: Birth on Land, Precocial
Development, and Sexual Dimorphism” - статья 2009 года в свободном доступе).
Окаменелость самки майяцетуса показывает, что даже на ранних стадиях эволюции
китообразные развивались по выводковому (англ. precocial) способу – рождался
сразу хорошо развитый детёныш, способный обслуживать себя сам. Авторы упомянутой
статьи полагают, что новорождённый детёныш уже мог дополнять молочный рацион
другой пищей.
Круглые головы эмбрионов дельфинов говорят о том, что у эмбрионов их предков
тоже были круглые головы – не более того. И ещё – что признаки, типичные для
отряда, семейства, рода и вида начинают появляться на более поздних стадиях
эмбрионального развития. Круглая голова – типичный признак эмбриона у многих
видов млекопитающих; выше уже приводились фотографии весьма круглоголовых эмбрионов
представителей самых разных групп млекопитающих.
Ещё раз позволю себе заметить, что Тену всё же стоит конкретно определиться,
кем были его «дельфиниды». Если имеется в виду семейство Delphinidae, то это
заведомо не археоцеты, а зубатые киты. Если это археоцеты, то они заведомо не
могут принадлежать к семейству Delphinidae, и даже называться должны уже как-то
иначе, чтобы не возникало путаницы. Можно быть зубатым китом, но не быть дельфином.
Но невозможно быть дельфином, и в то же время не быть зубатым китом. Впрочем,
при образовательном уровне автора, который я вижу в его работе, это оказывается
одной из самых небольших ошибок.
Здесь также стоит обсудить утверждение Тена о различии ювенильных («детских»)
и взрослых особей у китообразных. В настоящее время неизвестно ни одной окаменелости,
подтверждающей это предположение Тена, поэтому его уже можно смело не принимать
во внимание. Для млекопитающих вообще, на уровне всего класса, неизвестно явление
глубоких различий между ювенильной и взрослой особями. Утверждать столь глобальные
вещи, не имея доказательств на руках, может либо гений, либо безумец. Но признаков
гениальности автора в анализируемой мною книге я пока не встречаю.
Стр. 234:
Потом, пережив метаморфоз, дети навсегда уходили в море. Когда началась регрессия
Тэтиса, многочисленные группы дельфинид, в силу изрезанности ландшафта, начали
попадать в ловушки: отдельные водоемы, не имеющие связи с океаном. Это были
крупные внутренние моря, сравнимые по размерам с Каспием, которые сокращались,
как шагреневая кожа, на протяжении нескольких миллионов лет. Запертым археоцетам
эволюция могла предложить только один выход: развитие по типу педоморфоза, через
неотеническую форму. Второй выход – вымирание.
Ещё бы сюда и ископаемые формы, подтверждающие
эту прекрасную сказку! Думаю, именно отсюда произрастает какая-то органическая
ненависть господина Тена к описаниям ископаемых гоминид, сделанным профессиональными
палеонтологами: у злых «симиалистов» есть подтверждения их взглядов, а у прогрессивного
приверженца «альтернативной науки» Тена – только его собственные сказки и легенды.
А задача заставить читателя поверить в эти байки оказывается в таких условиях
трудновыполнимой. Если «симиалист» предъявит ископаемые кости, укажет на их
возраст и отметит характерные особенности их отличия, допустим, от шимпанзе
или человека, то господин Тен в данный момент может предъявить в доказательство,
разве что, собственное голое седалище. Поэтому приходится уходить от конкретики,
напускать туману, допускать искажённое толкование терминов и пр. Понимаю глубину
трагедии «прогрессивного новатора», но ничего с собой поделать не могу – я должен
быть строгим и объективным. Я не учёный, но по мировоззрению «обезьяний» эволюционист,
и мне бросили вызов, если аннотация к книге не врёт. Так что терпите, господа
хорошие, я ещё не закончил дозволенные мне речи. У нас всего лишь 234-я страница
из 363, содержащих текст.
Напомню лишний раз, что Мессинский кризис случился 5,96-5,33 млн лет назад,
а археоцеты вымерли примерно 23 млн лет назад, поэтому тут получается ещё и
неувязочка по времени на какие-то 18 миллионов лет. И ещё, как мы выяснили выше,
обсуждая страницу 232 книги Тена, явления неотении и педоморфоза при обсуждении
млекопитающих побоку по причине отсутствия у млекопитающих личиночной стадии
развития. Но Тену это явно неведомо, поскольку новый раздел данной главы называется
именно так, как совсем не должен в данном случае.
Стр. 234:
У многих животных наличествуют неотенические формы, в которых взрослые особи совершенно неузнаваемы. Например, у стрекоз, у лягушек. У лягушек рождаются лишенные конечностей «головастики», похожие на рыбок.
Для начала нам нужно дать определение
понятия неотении. Согласно «Биологическому Энциклопедическому Словарю» (М.,
«Советская Энциклопедия», 1989, стр. 402), неотения – это «задержка онтогенеза
у нек-рых видов организмов с приобретением способности к половому размножению
на стадии, предшествующей взрослому состоянию». То есть, животное должно уметь
размножаться, и вместе с тем выглядеть, как личинка.
Тен даёт нам своё определение этого явления: «Неотения (от греческого
«неос» – юный, незрелый, и «тейньо» – растягиваю, удлиняю) – способность организмов
размножаться на ранних стадиях развития, до достижения состояния взрослости»
(стр. 374).
Биологический смысл явления неотении – утрата той стадии развития, которая в
онтогенезе является взрослой, и приобретение способности к размножению на предшествующей
ей личиночной стадии.
Оба примера, приводимые Теном, совершенно не подходят для иллюстрации данного
явления. Если бы у нас была, например, нелетающая взрослая стрекоза с «маской»
на морде, живущая в воде, с трахейными жабрами и недоразвитыми огрызками крыльев
на спине, но способная размножаться, или же взрослая головастикоподобная лягушка
с внутренними жабрами и хвостом – тогда да, это были бы примеры неотении. А
пока – нет. Опять фактическая ошибка. Очередная.
Стр. 234:
Закономерность во всех случаях неотении такова: в новорожденных
воспроизводится филогенетически более ранняя форма.
Нет, это неверно. В случае неотении во
взрослых особях некоторого вида в какой-то степени воспроизводится их же собственная
невзрослая стадия – перечитаем ещё раз приведённое выше определение, принятое
в науке. У Тена опять фактическая ошибка в рассуждениях.
Личинка не является «филогенетически более ранней формой»: это результат приспособления
ювенильной формы вида к иному образу жизни и среде обитания по сравнению с взрослыми
особями того же вида. «Биологический Энциклопедический Словарь» любезно сообщает
о причинах появления личинок:
«У мн. животных стадия Л. обусловлена разл. образом жизни на ранних этапах развития
и во взрослом состоянии; часто наличие стадии Л. связано со сменой условий обитания
во время развития» (там же, стр. 326).
Поэтому неотения – это, фактически, эволюционный шаг к переходу вида к образу
жизни личинки, если это оказывается выгоднее превращения во взрослую форму.
Применительно к млекопитающим идея о появлении у них личиночной формы, отличающейся
от взрослого животного строением и образом жизни, выглядит крайне сомнительной.
Различия в строении априори означают эволюцию ювенильной формы в сторону иного
образа жизни в сравнении с взрослой стадией, и в идеале образ жизни личинки
и взрослой особи не должен пересекаться – появление личинки призвано, помимо
прочего, снизить конкуренцию ювенильной и взрослой форм за пищу и места обитания.
Возможен ли такой трюк у млекопитающих? Нужно вспомнить, что у млекопитающих
есть одно важное приспособление: выкармливание детёнышей молоком. А это означает,
что детёныш и родитель должны заведомо жить в одной и той же среде, и на них
должны действовать одни и те же эволюционные факторы, следствием чего будут
одинаковые признаки строения у взрослой и ювенильной особей. Ну, или если Тену
хочется, чтобы у его «дельфинид/акродельфид» была личинка с ногами, эта личинка
должна жить в условиях, благоприятствующих образу жизни существа с ногами, но
неблагоприятных для жизни взрослых китообразных. А это означает, что ни глоточка
маминого молочка «личиночные формы» не получат – мама будет далеко. Не велика
ли цена появления «личинки» у млекопитающего?
Стр. 234:
У древних дельфинид (судя по онтогенезу) это должно было быть существо, похожее
на человека. Половозрелые зародыши археоцет должны были походить на человека
еще больше, чем зародыши современных дельфинов, потому что они находились ближе
к исходной форме.
Здесь мы видим, как ранее допущенная
в рассуждениях ошибка приводит к ещё большей ошибке. И на это накладывается
неправильное толкование Теном биогенетического закона – распространение его
на все случаи онтогенеза, хотя он является проявлением лишь частного случая
онтогенеза. Об этом уже упоминалось при обсуждении стр. 212 данной книги Тена.
«Половозрелые зародыши»? Это что-то новое в науке – напоминает гигантского гнома
и карликового великана. Нет, бывает, конечно, когда у тех же тлей личинка может
рождаться, уже содержа в себе собственных личинок. Но в этом случае мы просто
наблюдаем партеногенез – фактически, «клонирование». Да, я снова докопался до
формулировки. Раз Тен это начал, я имею право продолжить.
Стр. 234-235:
Неотении способствовала гиперсексуальность дельфинид, в том числе их детская
сексуальность.
При обсуждении стр. 171-172 книги Тена
уже говорилось о том, что сексуальное поведение у детёнышей не является каким-то
уникальным эволюционным фактором у зубатых китов: подобные формы поведения встречаются
у значительного числа видов млекопитающих. Это просто элемент игрового поведения.
К тому же Тен неправильно использует термин «гиперсексуальность», примеряя к
человеку особенности полового поведения других биологических видов. Об этом
также говорилось выше (см. обсуждение стр. 172-173 книги Тена).
Стр. 235:
Следует предполагать, что они не могли плавать с такой же скоростью, как сами
археоцеты, но превосходили их в искусстве ныряния. Кстати сказать, именно такой
способ «купания» является излюбленным для людей. […] В этом, как в любой мелочи
нашего поведения в природном формате, тоже должен быть заключен какой-то эволюционный
смысл, над которым стоит задуматься.
С такой же степенью уверенности я могу
предполагать, что у полуразумных пятиногих обитателей планетной системы звезды
Фомальгаут есть приспособления, позволяющие им пить концентрированную серную
кислоту, что очень способствует пищеварению. Ну, если не на Фомальгауте (эта
звезда молода), то на Глизе 867 точно. Таких умствований, не подкреплённых фактическими
объективными данными, я могу насочинять ворох. А другие люди на это жизнь кладут,
фантастические романы пишут, знаменитыми становятся даже.
Только вот какова она – эта степень уверенности? Учёный в чём-то уверен лишь
тогда, когда у него на руках имеются проверяемые материальные доказательства
– реальные ископаемые кости, результаты исследований с подробными описаниями
методики и оборудования. О поведении ископаемых животных можно судить по косвенным
данным – следовым дорожкам, особенностям строения костей (позволяющим реконструировать
мускулатуру), изотопному составу эмали зубов и пр. Методы исследований и полученные
с их помощью данные публикуются и обсуждаются. А предположения, сделанные «ни
на чём» – это чистой воды фантазии автора этих предположений. В данном случае,
например, господин Тен должен был:
1) описать особенности строения скелета своего «неотенического дельфиноида»
в сравнении с близкими видами археоцетов, отмечая в скелете черты приспособленности
к нырянию;
2) показать на форме зубов и особенностях строения челюстей, что его «неотенические
дельфиноиды» питались донными организмами в прибрежной зоне;
3) показать на изотопном составе эмали зубов, что он характерен именно для существа,
питающегося в прибрежной зоне, а не в открытом океане;
4) описать виды живых организмов (от одноклеточных до позвоночных), ассоциирующиеся
с «неотеническим дельфиноидом», и подтверждающие предположения о его образе
жизни, описанные выше.
И только после того, как будет проделана такая работа, господин Тен сможет с
полным правом утверждать, что его «неотенические дельфиноиды» вели именно такой
образ жизни, и не иначе. Однако вот уже 235-я страница его труда, а ни одного
конкретного названия ископаемых существ, подтверждающих выкладки Тена, у нас
нет. А те, что есть, оказываются неопределёнными (точнее, ошибочными) даже на
уровне отряда. Помните ту детскую песенку? «Далеко, далеко, на лугу пасутся
ко…». Вот и я думаю: то ли корифодоны, то ли котилозавры, то ли просто кошки.
Шутка. Но точность доказательств Тена примерно такая и есть.
Стр. 235:
Известны случаи, когда в силу изменения природных условий развитие по типу педоморфоза
начинает преобладать. Классический пример – аксолотль, обитающий на юге Северной
Америки. […] Аксолотли способны давать потомство, рожая таких же, как они сами.
Коэффициент цефализации у них выше, чем у амбистом – того вида саламандр, чьей
неотенической формой являются аксолотли.
Очень сожалею, что в очередной раз у
господина Тена отсутствует ссылка на научную работу, откуда взята данная информация,
и которая позволяет проверить приводимые автором данные о мозгах аксолотлей
и амбистом и корректность их интерпретации Теном. Кстати, амбистом существует
довольно много видов. Не может ли быть так, что различия в коэффициенте цефализации
отражают не особенности онтогенеза, а видовые различия?
И, да, аксолотли не живородящие, а мечут икру.
Стр. 235:
Более высокая степень цефализации в сравнении с родительской формой
является закономерностью педоморфоза. Даже у людей дети имеют
больше мозгов относительно массы тела, чем взрослые. Причем это не просто анатомический
показатель. Дети гораздо смышленее взрослых, они все – нераскрывшиеся гении.
Бывают бесталанные взрослые, но не бывает бесталанных детей, это аксиома педагогики.
Здесь не помешали бы ссылки на научные
работы, показывающие общую природную тенденцию к более высокой степени цефализации
у ювенильной формы. Пока это всего лишь умозрительное заключение Тена.
Больший размер мозга у детей также объясним: у них иные пропорции тела, можно
посмотреть даже в руководствах для художников. В процессе индивидуального развития
разные части тела растут с разной скоростью, и со временем тело чуть обгоняет
в скорости роста голову, и пропорции меняются на характерные для взрослых. Это
происходит потому, что индивидуальное развитие – процесс не только и не столько
линейного роста, но и качественных изменений. Те же половые железы, например,
у ювенильных организмов пока не работают.
Мозг ребёнка, в отличие от мозга взрослого, подготовлен для обучения – это приспособление
для выживания. У таких существ, как приматы, и в особенности человек, инстинктивное
поведение представлено «базовым минимумом», и несравнимо больший объём информации
усваивается в процессе взросления. Смышлёность детей – это проявление процесса
обучения в период активного роста мозга. У взрослого человека мозг сформирован,
поэтому усвоение новой информации идёт сложнее. Так что здесь на выводы Тена
накладываются иные биологические закономерности, что приводит к искажённым выводам.
Стр. 235-236:
Наверное, многие читатели будут удивлены, узнав, что неотения является чрезвычайно
распространенным явлением в мире животных, а педоморфоз – одним из главных инструментов
эволюции. Благодаря «детскому сексу» и задержке развития появились не только
отдельные виды животных, но и более крупные таксоны: семейства, роды, классы.
К примеру, все насекомые представляют собой итог развития неотенических форм
древних многоножек (Зотин, 1999, с. 201). «Считается, что не только насекомые
и некоторые другие группы беспозвоночных возникли путем неотении, но и некоторые
группы позвоночных», – пишут в книге «Направление, скорость и механизмы прогрессивной
эволюции» А.М. Зотин и АА Зотин (там же, с 242).
Относительно «чрезвычайной распространённости»
неотении можно поспорить. В той же самой книге написано следующее:
«В истории сравнительной анатомии беспозвоночных на протяжении последних 150
лет было высказано немало предположений о происхождении отдельных крупных таксонов
от преимагинальных стадий других групп животных. В случае насекомых, например,
их происхождение от ранних стадий развития многоножек сомнений не вызывает».
Обратим внимание на слова «отдельных крупных таксонов». Не всех и не многих,
стало быть. Также отметим в цитате из Зотиных слова «некоторые группы позвоночных».
В отношении земноводных и рыб – вполне можно согласиться, у них есть таксоны,
представители которых несут явные личиночные черты. Но даже в цитируемой книге
авторы не считают данное явление всеобщим.
«Детский секс» в случае указанных явлений является не причиной, а следствием:
личинки в своём исходном состоянии не проявляют элементов сексуального поведения.
Они начинают проявлять его, когда у них под действием иных эволюционных факторов
развиваются половые железы. Из-за примитивного поведения им недоступен такой
элемент поведения высокоразвитых позвоночных, как игры, в том числе с сексуальными
элементами.
Неотения подразумевает сохранение ювенильных признаков у взрослой формы. Кстати,
не задумывались, почему? Ведь не просто так эти признаки задержались у взрослых
особей, верно? Ответ один: в меняющихся условиях они приобретают адаптивное
значение, потому и сохраняются как можно дольше. Если образ жизни личиночной
стадии выгоднее, чем взрослой, эволюция с большой степенью вероятности может
пойти по пути «взросления» личиночной формы такого вида, отбрасывая ставшую
эволюционно невыгодной взрослую стадию онтогенеза.
Стр. 236:
Неотения – это эволюционная инверсия.
Хорошо, посмотрим для начала, что означает
слово «инверсия».
«Викисловарь»
даёт нам самое обобщённое определение этого понятия:
«Общее прототипическое значение — изменение обычного порядка с взаимной заменой
частей чего-либо».
Теперь посмотрим, насколько это понятие применимо к нашим явлениям. Напомню
понятия из «Биологического Энциклопедического Словаря»:
Ретардация – «замедление темпов эмбрионального развития или запаздывание эмбриональной
закладки органа у потомков по сравнению с предками» (стр. 538).
Неотения – «задержка онтогенеза у нек-рых видов организмов с приобретением способности
к половому размножению на стадии, предшествующей взрослому состоянию» (стр.
402).
Педоморфоз – «способ эволюц. изменений организмов, характеризующийся полной
утратой взрослой стадии и соответствующим укорочением онтогенеза, в к-ром последней
становится стадия, бывшая прежде личиночной» (стр. 455).
Замедление, задержка, утрата… Где же тут перестановка? Выходит, даже само название
новоявленной «теории» господина Тена – неправильное? Впрочем, тут я не удивлён.
А замедление темпов развития органов и систем органов, которое лежит в основе
этих явлений, носит название «отрицательная гетерохрония», поэтому теория Тена,
будь она реальностью, должна бы называться «нега-гетерохронной теорией».
Стр. 236:
В настоящее время она изучена до возможности выявления закономерностей. «Явление
неотении в случае измельчания вида или перехода его в неблагоприятные условия
существования связано с трудностями борьбы за существование взрослых форм животных»,
– говорится в цитируемой монографии. Далее приводится еще одна закономерность:
«Неотенические виды имеют преимущество при меньших массах и недостаток
– при больших по сравнению с предковыми формами» (там же, с. 242).
Простите, в цитируемой книге сказано: «Неотенические виды имеют преимущество в энергетическом обмене при малых массах и недостаток – при больших по сравнению с предковыми формами». На всякий случай выделю стыдливо пропущенный кусочек текста. Это не означает преимуществ в борьбе за существование вообще, верно?
Стр. 236:
Акродельфиды и люди имеют меньшую массу по сравнению с дельфинами, причем настолько,
чтобы можно было считать эти виды неотенической формой древних дельфинид в развитии.
Это тот случай, когда неотеническая форма имеет эволюционное преимущество в
сравнении с исходным видом.
Массы современных дельфинов – от 35 кг
(дельфин Коммерсона) до 10 тонн (косатка). Как видим, масса взрослого человека
находится в пределах этих значений. Бывшие акродельфиды известны по достаточно
фрагментарным останкам, однако размеры костей позволяют утверждать, что величина
взрослых животных укладывалась в пределы значений, характерных для дельфинов.
К тому же линейные размеры – это не признак неотенической формы. Отличия от
взрослой формы в данном случае не количественные, а качественные – наличие личиночных
особенностей в анатомии. У тех же земноводных аксолотль имеет примерно такие
же размеры, как взрослая амбистома, а обладающие наружными жабрами американский
протей и сирен значительно крупнее многих не-неотенических саламандр и тритонов.
Если же судить только по количественному показателю (масса тела, размеры), можно
договориться до идиотского вывода о том, что дельфин Коммерсона – это неотеническая
форма, допустим, косатки, гринды или серого дельфина.
То, что люди будут иметь меньшую массу, чем большинство дельфинов – вполне закономерно.
Китообразные обитают в среде, которая обладает значительной теплопроводностью
и даже в тёплых широтах постоянно создаёт опасность переохлаждения, поскольку
её температура заведомо ниже температуры тела млекопитающего. Поэтому от них
ожидаемо иметь достаточно большую массу тела, в том числе за счёт накопления
жира. И в связи с высокой теплопроводностью воды постоянноводное теплокровное
животное не может быть маленьким.
Так что меньший (в среднем) размер тела человека в данном случае не указывает
на неотению, а является следствием обитания в разных средах.
Стр. 236:
Отдельные ученые-антропологи, например Л. Больк и его сторонники, считают, что
без ретардации происхождение человека понять и объяснить невозможно. Разумеется,
пребывая в плену симиальной концепции, они исходят из того, что человек является
неотенической формой какой-то человекообразной обезьяны, скорее всего шимпанзе.
Между мозгами антропоидов и мозгом человека такая пропасть, что заполнить ее
представляется возможным только таким образом. Но выживание неотенической формы
древесной или саванной обезьяны было невозможно.
Судя по всему, господин Тен сам попал
в плен собственного мировоззрения. Иначе с чего бы более высокую компетентность
человека в каком-либо вопросе он безоговорочно записывает в последствия «плена
симиальной концепции»? А собственные ошибки и нестыковки он деликатно старается
не замечать, поэтому приходится разбирать его книгу едва ли не по абзацам.
Следует также помнить, что шимпанзе – не предок человека, а просто близкий родственник
в мире животных. Неоднократная ошибка в рассуждениях Тена – приписывание специалистам
заведомо неверного суждения.
Тен не приводит доказательства утверждения, что «выживание неотенической
формы древесной или саванной обезьяны было невозможно».
Не забываем, что даже в случае неотении как направления эволюции ювенильные
признаки остаются у взрослой формы не просто так, а из-за того, что приобретают
определённое адаптивное значение и именно способствуют выживанию. Иначе эволюция
просто не пошла бы в этом направлении. Следует помнить также, что неотенические
формы – это какие-то «вечные дети», ювенильные по всем параметрам. Ювенильные
признаки у неотенических форм сочетаются со вполне взрослыми – в частности,
со способностью к размножению. Поэтому в целом на жизнеспособности представителей
рассматриваемого вида неотения не отразится, как не отражается на тех формах,
которые демонстрируют это явление в наше время.
Стр. 237:
Дело в том, что ретардация, неотения и педоморфоз возможны только у определенной
группы животных. Это, по сути дела, еще одна закономерность неотении,
ландшафтная. Конкретно она формулируется так: вид, появившийся вследствие неотении,
может быть наземным, но исходная неотеническая форма может быть только водно-земной.
Тен представил нам всего лишь один пример,
который он сам считает примером неотении. И на основе этого примера, корректность
которого в целом крайне сомнительна, делает далеко идущие выводы и выстраивает
закономерности. Но по одной точке даже самый поднаторевший в статистике специалист
график не построит. Нужен некий массив примеров, который позволил бы вывести
закономерность.
Просто напомню, что наличие в онтогенезе личинки, отличающейся от взрослой особи,
связано с эволюцией в сторону различного образа жизни у личинки и взрослой особи.
Одним из частных случаев является онтогенез земноводного, у которого личинка
водная, а взрослая форма наземная. Видимо, этот частный случай Тен приводит
в качестве общей закономерности. В англоязычной «Википедии» в статье
о неотении говорится, что появлению неотении способствуют условия высокогорья
и низкие температуры. У аксолотля, классического примера этого явления, в наличии
оба фактора.
Стр. 237:
Наземные неотенические формы неизвестны, во всяком случае, специалисты не приводят
подобных примеров.
Хорошо. Приведу немного примеров, причём именно наземных. Например, вельвичия среди голосеменных растений и однолистные стрептокарпусы среди покрытосеменных – это, по сути, проростки, ставшие взрослыми и получившие способность размножаться. Ни голосеменные вообще, ни геснериевые среди цветковых растений в любви к водному образу жизни не замечены, но неотению нам демонстрируют наглядно.

Streptocarpus grandis - однолистный вид стрептокарпусов.
Страницей выше сам же Тен приводил пример
происхождения насекомых как результат неотении многоножек. И те, и другие –
наземные формы. Веерокрылы среди насекомых также демонстрируют неотению у самок.
Но они не водные животные, а наземные. Дронты происходят от «стандартных» голубей
путём сохранения во взрослом состоянии ювенильных признаков, но голуби ни в
привычном виде, ни в облике дронта в любви к водному образу жизни вообще не
замечены. Так что
Странный подход, кстати. Тен пишет, что «специалисты не приводят подобных примеров»,
а потом именно на этом основании объявляет их выводы несостоятельными. Почему
бы не применить эту же самую логику к его собственным умопостроениям? Ведь у
Тена в рассуждениях не встречается ни одного вида, подтверждающего его собственную
версию – только отдельные виды ископаемых китообразных (точнее, всего один),
или же открытые злыми «симиалистами» виды гоминид. Ни одного примера видов,
стоящих между несомненными китами и разумными людьми, Тен не приводит.
Стр. 237:
Из этого следует, что гипотеза ретардации Болька несостоятельна, если материнской
формой считать обезьян, живших в лесу или в саванне. Поэтому сторонники гипотезы,
согласно которой люди являются неотеническими потомками шимпанзе, вынуждены
в подтверждение ее приводить примеры с земноводными, которые в их устах звучат
как некорректные. Здесь можно вспомнить Дарвина, который, обосновывая происхождение
людей благодаря половому отбору, приводил примеры не с млекопитающими, а с птицами,
– они тоже, безусловно, некорректны, потому что человек – млекопитающее.
Если Тен не знает примеров наземных неотенических
форм, это ещё не означает, что их нет. К тому же для позвоночных, у которых
отсутствует личиночная стадия, неотения в чистом виде не характерна. Максимум,
на что они способны – сохранять отдельные ювенильные или фетальные признаки;
данное явление называется «фетализация» и достаточно широко распространено среди
наземных позвоночных.
Наконец, стоит прокомментировать рассуждения Тена о корректности тех или иных
примеров. Если некоторое явление признано общебиологическим, то это означает,
что оно изначально было выделено как устойчиво повторяющаяся закономерность
именно у разнообразных представителей живых организмов. Учёные сперва наблюдают
и собирают факты, а потом на основании фактов выделяют те или иные закономерности.
Стр. 237:
Наземные животные не могут позволить себе надолго задерживаться в детстве.
Весьма опрометчивое заявление, но ничем
не обоснованное. Всё зависит от избранной видом в процессе эволюции стратегии
размножения. Есть два вида стратегий размножения: r-стратегия и K-стратегия.
Одна из них – оставление немногочисленного потомства с высоким родительским
вкладом в выращивание каждого потомка (K-стратегия). Вторая – прямая противоположность:
многочисленное потомство при минимальном вкладе родительской особи в выращивание
(r-стратегия).
Примеры обеих стратегий у нас перед глазами. Детство орангутана длится до 4
лет, и первый раз молодая самка становится мамой в 8-12 лет. Мышь достигает
половой зрелости на 5-7 неделе жизни, в год даёт 5-10 выводков. Пока самка орангутана
родит одного детёныша, у мышей сменится от 59 до 120 поколений. Так что в детстве
задержаться иной раз можно – здесь есть свои недостатки, но и свои преимущества.
Достаточно сравнить интеллект мыши и орангутана, и количество усваиваемой ими
в течение жизни информации.
Стр. 237:
У водно-земных животных возможностей продлить период детства больше настолько,
насколько больше возможностей спастись от преследователей: в два раза. Обитание
на границе двух разных ландшафтов обеспечивает возможность при минимальной затрате
энергии в два раза эффективнее спасаться от естественных врагов.
А вот теперь убавим прыть бешеных кентавров
воображения. Разве сохранение ювенильных признаков в строении означает удлинение
детства? По-моему, здесь Тен путает два понятия, означающих совершенно разные
вещи: онтогенез (индивидуальное развитие отдельно взятого организма) и филогенез
(историческое развитие вида в целом). Так вот, явления вроде неотении и фетализации
относятся к филогенезу, а понятие «детство» – к онтогенезу.
Объяснение Тена относительно преимуществ «продления периода детства» выглядит
крайне наивным. Дело в том, что вид на границе двух ландшафтов оказывается в
положении барона Мюнхгаузена – между львом и крокодилом. Обитание в двух ландшафтах
означает наличие врагов в обоих этих ландшафтах. И не факт, что наш «дельфинидо-человек»,
спасаясь от льва, не прыгнет прямо в поджидающую его под водой пасть крокодила.
Или акулы. Или бригмофизетера. Плюс в каждой из этих сред найдутся паразиты,
способные жить в теле нашего гипотетического существа.
Стр. 237:
Интересно, что слово «дельфин» – древнегреческого происхождения и восходит к
слову «delphus», что означало «матка», «лоно», «утроба». Не говорит ли это о
том, что у древних сохранялись мифы еще более древних предков, в которых дельфины
воспринимались в качестве прародителей?
… Или же просто о том, что дельфин, в отличие от рыб, рождает живых детёнышей, что не могли не заметить греческие рыбаки.
Стр. 238:
[…]
Удивляет разорванность мышления некоторых ученых. С одной стороны, они уже совершенно
точно знают, что на базе обезьяньих мозгов не мог мало-помалу сформироваться
мыслительным аппарат человека. Это доказано уже десятки раз, включая математическое
моделирование.
Вот в этом самом месте, вместо того, чтобы читать заклинания, взывая к некой «общеизвестности» тех или иных фактов, нужно крепко, «с ноги», можно сказать, «припечатать» косных, с задеревеневшими мозгами «симиалистов» ссылками на научные работы, массово появляющиеся в рецензируемых журналах. Пока же этих ссылок в этом месте книги Тена днём с огнём не сыщешь. Прямо небо и земля по сравнению с теми частями книги, где он, стуча пяткой в грудь, набрасывается с разрушительной критикой на работы А. Маркова и Ст. Дробышевского, любезно приводя и том, и страницу. А тут, внезапно (тыдыщ!), работы общеизвестные, «доказанные уже десятки раз». Ну, покажите же хоть первый десяток-то?
Стр. 238-239:
С другой стороны, большинство ученых продолжают оставаться симиалистами, потому
что иначе их просто перестанут воспринимать всерьез, запишут в маргиналы и лжеученые.
Такое ощущение, что мировое академическое сообщество пребывает в коллективном
когнитивном диссонансе, причем осознанном. Дошли до тупика, но упорно продолжают
толкать тачку в стенку. Никакое другое существо вместо понгид нельзя даже предложить
на рассмотрение в качестве возможных предков человека, если хочешь, чтобы такие
люди, как уважаемый академик Иванов, восприняли тебя всерьез, а не в качестве
маргинала или юродствующего дурака, притом что сами уже давно знают, что человеческий
мозг мало-помалу никак не мог получиться из обезьяньих мозгов. Почему в таком
случае не ищут инверсию, других предков? Обезьяна держит лохматыми лапами?
Хорошо, мы спросим фразой из фильма «Красная
жара»: какие ваши доказательства?
Обезьяна не просто так расселась на почётном месте предка человека. В пользу
происхождения человека от обезьяны говорят многочисленные согласующиеся друг
с другом доказательства: сравнительно-анатомические, физиологические, биохимические,
генетические и палеонтологические. Вспомним, что в разделе критики книг «симиалистов»
даже сам Тен обильно сыплет конкретными названиями существ, включённых в родословную
человека, с точностью до рода и вида. А как начинаем говорить о его собственной
теории, у которой даже название, как выясняется, неправильное, так начинаются
пассы дланями в воздусях: предки человека дельфиниды, но не те дельфиниды, которые
Delphinidae, то есть, собственно дельфины, а те дельфиниды, а может даже акродельфиды,
которые археоцеты. Причём такие волшебные дельфиноидо-акродельфидо-археоцеты,
которые жили во времена Мессинского кризиса, хотя у учёных-палеонтологов все
археоцеты к этому времени уже почти пару десятков миллионов лет, как преставиться
изволили. Наверное, эти спецы по ископаемым китам в сговоре со злыми «симиалистами»,
не иначе. Естественно, Тен излагает основы своих воззрений без малейшей конкретики
в названиях даже на уровне рода, не говоря уже о том, что Тен совершенно не
знает современных взглядов на систематику тех же археоцетов. И всплывает вопрос
из анекдота про лекции Эйнштейна: «И с такими хохмами он едет в Одессу?» В смысле,
собирается опровергать ненавистный «симиализм»?
Возможно, я уже упоминал выше, что астроном и популяризатор науки Карл Саган
говорил: «Экстраординарные утверждения требуют экстраординарных доказательств».
Происхождение человека от обезьяны – достаточно ординарное утверждение, у которого
есть доказательства. Происхождение человека от китообразного предка требует
ответов на гораздо большее количество вопросов, поэтому по сравнению с позицией
«симиалистов» оно является экстраординарным. И ниспровергатель «обезьяньей»
версии антропогенеза должен безупречно с научной точки зрения:
1) опровергнуть доводы о близости обезьян к человеку;
2) объяснить все эволюционные процессы, сопровождавшие переход от китообразного
предка к человеку, с представлением материальных, проверяемых и однозначно толкуемых
доказательств.
Наука не держится на авторитете личностей. Многие из них ошибались. Даже Дарвин
в некоторых случаях был неправ.
«Опровержение устоявшихся представлений – неотъемлемая часть науки. Однако когда
настоящие ученые что-то пересматривают, они всегда делают это очень аккуратно
и последовательно. Данные должны быть представлены по возможности полно и со
ссылками на источники, если их добыл кто-то другой» (источник: А. Тимошенко
«Cмотри, какая дичь. Как опознать псевдонауку по ряду характерных признаков»,
ссылка).
У господина Тена собственных исследований в области анатомии, палеонтологии
и генетики нет. Поэтому на собственные работы он чисто физически не сможет сослаться.
А ссылок на работы признанных учёных, подтверждающие его измышления, он не приводит.
Стр. 239:
С целью избежать обвинений в домыслах, хотя ни один палеоантрополог без них
не обходится, я буду говорить только о фактах. Причем начнем не прямо с сапиентации,
а с ее предпосылок.
О фактах надо было говорить с самого
начала книги. Пока в подтверждение излагаемой «инверсионной теории антропогенеза»
я видел лишь домыслы и подтасовки фактов. Помните, при обсуждении стр. 211-212
книги Тена мы выясняли, что такое «подтасовка»? В данном случае это крайне избирательная
подача фактов, которые сами по себе ничего не доказывают, вроде цвета склеры.
По поводу домыслов у палеоантропологов я бы хотел увидеть хотя бы кучу этих
примеров, особенно в научных статьях. Прямо вот зайти на сайт Researchgate,
сделать выборку по теме, и давай хлестать меня по мордасам, да по сусалам цитатами
из первых же попавшихся в выборку научных статей из рецензируемых журналов:
вот, мол, тебе, злой серый Волков, подтасовка данных, а вот домысел, да вот
ещё один, другой, третий... Прямо вот чтобы я головы поднять не смел, увёртываясь
от града нелицеприятных фактов и виновато покусывая собственные локти. На всякий
случай уточню: не научно-популярных статей и книг, а строго научных, написанных
именно учёными и именно в рецензируемых журналах.
Стр. 239:
Факт, что из всех известных науке ископаемых животных наиболее ярко выраженную
тенденцию к увеличению объема мозга демонстрируют древние дельфиниды. В то время,
когда началась эволюция человека, они одни во всем животном царстве демонстрировали
показатели развития мозга, сравнимые с теми, что имеет Homo sapiens.
А у тоатерия из южноамериканских копытных
ноги стали однокопытными, как у современных лошадей, ещё раньше, чем у самой
лошади – тоже ярко демонстрируют тенденцию. И разве это делает его родственником
лошади? Характерный «парус» из остистых отростков спинных позвонков в разное
время имели платигистрикс (земноводное), диметродон и эдафозавр (зверообразные
рептилии), лотозавр (текодонт), спинозавр (ящеротазовый динозавр), оуранозавр
(птицетазовый динозавр). Они что – все родственники? А сколько раз на протяжении
эволюционной истории зверообразных рептилий и млекопитающих разных отрядов возникал
сабельный клык? И этот отдельный общий признак делает их родственниками? Заставляет
выстраивать между ними отношения «предок-потомок»? Нет. Примеры такого рода
лишь показывают, что сходные тенденции могут проявляться в процессе эволюции
у представителей разных, даже неродственных групп живых организмов, причём,
возможно, под действием разных факторов – как, например, сабельный клык у саблезубого
тигра и травоядного уинтатерия или кабарги.
В случае китообразных одним из факторов, способствующих развитию их мозга, может
быть сам факт их обитания в воде – среде, враждебной наземным по происхождению
животным. Социальность и взаимодействие между сородичами становятся факторами,
благоприятствующими выживанию, и отбор на усложнение поведения привёл к развитию
мозга.
Событие приобретения некоторого признака одной из групп живых организмов вовсе
не означает, что другие живые организмы, у которых такой признак появился позже,
являются потомками этого «первопроходца». Тен предлагает нам ложную логическую
связь: Post hoc ergo propter hoc («После этого – значит вследствие этого»).
Стр. 239-240:
Существует несколько критериев оценки мозга потенциальных кандидатов в люди.
Абсолютный критерий, т.н. мозговой Рубикон, предложенный в 1948 г. А. Кизсом,
в настоящее время не применяется в таком качестве. Ученые пришли к выводу, что
гораздо большее значение имеют такие показатели, как индекс цефализации («мозговитости»);
индекс кортиколизации [sic – П. В.] (соотношение
неокортекса и общей поверхности коры); количество ассоциативных областей в коре;
характер и число борозд; развитие мозолистого тела как основной комиссуры мозга
(предложено мной в 2005 г.), и еще несколько показателей, объединяемых в понятие
«архитектоника мозга».
Индекс цефализации изначально рассматривался как простая арифметическая дробь,
где в числителе масса мозга, а в знаменателе масса тела. Однако данная пропорция
имела слишком общий характер и не давала возможности надежно вычленить человека
из животного царства по такому важному для вида Homo sapiens показателю. В 1933
г. Я. Рогинский предложил т.н. квадратный коэффициент, который и стал базой
для расчета первых «индексов цефализации». После Рогинского появилось много
различных методик подсчета индекса цефализации, особенно на Западе. Там для
этой цели сформировалась разветвленная лженаука.
Никто из учёных не «вычленяет» человека
из царства животных, поскольку место нашего вида в природе уже определено, и
не подвергается сомнению никем, кроме больших оригиналов вроде господина Тена
и ему подобных.
Простые зависимости, выражаемые простыми формулами, не годятся в данном случае
хотя бы из-за того, что увеличение размеров тела в процессе эволюции нелинейно
– то есть, если некое живое существо стало в пять раз тяжелее, это не означает,
что вес его мозга также увеличился в пять раз.
Тен клеит к данной области биологии ярлык «лженаука». Стоит ли это делать? Для
этого нужно разобраться с тем, что такое лженаука. Русская «Википедия»
приводит такие признаки лженауки:
«Среди основных отличий псевдонауки от науки – некритичное использование новых
непроверенных методов, сомнительных и зачастую ошибочных данных и сведений,
а также отрицание возможности опровержения, тогда как наука основана на фактах
(проверенных сведениях), верифицируемых методах и постоянно развивается, расставаясь
с опровергнутыми теориями и предлагая новые».
Судя по отсутствию ссылок на первоисточники в данной части текста, именно построения
Тена грешат уклоном в лженауку, поскольку не дают возможности элементарно проверить
предлагаемые сведения. В любой научной публикации (хотя бы на сайте Researchgate)
можно увидеть большой список цитируемой литературы, а также описание использованного
в ходе исследований оборудования и методики измерений. Поэтому всякий желающий
может проверить изложенные в статье данные. Мало того, даже в научно-популярных
книгах А. Маркова и Ст. Дробышевского, на которые так яростно набрасывается
господин Тен, имеется по несколько десятков страниц ссылок на первоисточники.
Опровергаются ли публикации в научных журналах? Естественно, да. Информацию
об отозванных публикациях легко можно найти хотя бы в Интернете. И опровергаются
они именно благодаря подробному описанию оборудования и методики исследования,
что позволяет найти ошибку в процессе получения, фиксации и обработки данных.
Так что в данном случае ярлык «лженаука» от господина Тена беспочвен.
Стр. 240:
Существует расхожая истина: в каждой науке столько науки, сколько в ней математики.
Рядом бродит не менее расхожая: бывает ложь, наглая ложь и статистика. Так вот:
индекс цефализации – это статистика. Если б разум был у червяков, они придумали
бы такие коэффициенты, что выглядели бы самыми «цефализованными».
Вся закавыка заключается в том, что, если бы червяки стали разумными, у них бы этот факт неизбежно отразился бы на строении – появился бы некий мыслительный орган, материальный носитель сознания, имеющий более сложное строение по сравнению с их неразумными родственниками. Таким образом, между этим видом и его неразумными родственниками объективно и неизбежно существовали бы отличия, которые можно было бы подвергнуть количественной оценке.
Стр. 240:
Честный математический подход может быть только один: масса мозга, деленная
на массу тела. Так вот, человек по такому показателю существенно не вычленяется
из мира животных. Был бы Рубикон в мозгах согласно пропорциям по массе, он стал
бы главным критерием человечности.
Такой подход покажет чисто количественные различия, но не качественные. В английской «Википедии» в соответствующей статье для иллюстрации этого примитивного соотношения указаны такие цифры (приведу лишь некоторые):
Мелкие птицы: 1/12
Человек: 1/40
Мышь: 1/40
Дельфин: 1/50
Шимпанзе: 1/113
Лягушка: 1/172
Слон: 1/560
Как видим, если не учитывать дополнительных
параметров, то при «честном математическом подходе» воробей будет умнее человека,
мышь превзойдёт дельфина, а лягушка – слона. Соответствует ли «честность» этого
подхода реальной действительности?
Добавлю ещё немного цифр из другого источника:
«В самом деле, у дельфинов большой размер мозга с огромным числом нейронов и
множество извилин в нем не обязательно могли развиваться в связи с речью. Вес
мозга относительно веса тела сам по себе еще ни о чем не говорит. У человека
мозг составляет 1/34 веса тела. Но есть обезьяны в Южной Америке, относительный
вес мозга которых выше, чем у человека (у капуцина - 1/18, а у черной коалы
- до 1/15 веса тела). Однако никто не считает этих низкоорганизованных широконосых
обезьян "умнее" человека» (Томилин А.Г. «Снова в воду» М., «Знание»
1977, ссылка).
Имеется в виду, очевидно, не коала (сумчатое), а коата (как раз обезьяна). Слова
стоит иметь в виду и держать в уме – Томилин был специалистом и дельфинов знал
значительно лучше, чем Тен.
Стр. 240:
А в накрученные «индексы цефализации» не верьте: это очередное мошенничество
антропологов, которое, впрочем, не помогло им в доказывании обезьяньей версии
происхождения человека. Но занятие доходное: нелишний способ сделать имя и диссертацию,
хотя для науки абсолютно лишний.
Ладно-ладно, верю вам, господин Тен.
Лягушка действительно умнее слона. А мышь умнее дельфина. В свете этих выводов
данная мною вам оценочная характеристика «птичьи мозги» просто обязана восприниматься
вами как комплимент, если следовать «честному математическому подходу» и вышеприведённым
сведениям.
А теперь немного помыслим трезво. Что есть «индекс цефализации»? Берутся две
переменные – вес мозга и вес тела – и выводится некое математическое соотношение
данных величин. Чему тут не верить? Объективным проверяемым данным – весу мозга
и весу тела? Математической формуле? Хорошо, не поверим мошеннической формуле
«негодяев в белых халатах». Но она одна и для «теномерзкого» шимпанзе, и для
«теноугодного» дельфина. Отвергая данные, полученные при помощи некоторой формулы,
для шимпанзе и ископаемых гоминид, мы также автоматически отвергаем их для дельфина.
Тен в очередной раз критикует «накрученные «индексы цефализации»», хотя их появление
и изменение способов вычисления вполне объяснимы. Этому посвящён, например,
раздел 7.12.3 (стр. 316) книги У. Т. Фитча «Эволюция языка» (М., Языки славянской
культуры, 2013), которую господин Тен использовал в подготовке своей работы
и внёс в список литературы на стр. 368 своей книги. Но нам ведь надо ради красного
словца демонизировать злобных «симиалистов», верно?
Стр. 240:
Однако даже эти пресловутые коэффициенты – против обезьян и за дельфинов.
«По коэффициенту цефализации зубатые киты входят в одну группу с человеком,
ближайшие конкуренты – антропоиды – отстают в 3-4 раза» (Крушинская, Лисицына,
1983, с. 117,118).
Ну, что можно сказать? Киты молодцы! Не буду оспаривать данные, полученные специалистами – незачем. Но возникает вопрос: в процитированной книге что-нибудь говорится о родстве человека и китообразных на основании этих данных? И ещё вопрос: какие именно зубатые киты исследовались? А то к ним относится и кашалот с его 7,8 килограммами мозга на 40-50 тонн веса.
Стр. 240:
Интереса ради прикинем честную пропорцию, что называется, «по гамбургскому счету».
Средний вес мозга человека – 1,4 кг, тела – 75 кг. Дельфин афалина: 1,7 кг мозга
против 135 кг тела. Шимпанзе: 0,4 кг мозга против 50 кг тела (берем с осторожностью
в пользу шимпанзе, они весят до 70 кг). Честная пропорция получается следующая:
человек – 0,0186, дельфин – 0,0126, шимпанзе – 0,008. Получается, что от шимпанзе
мы выделяемся впечатляюще, а от дельфинов – нет. У горилл и орангутанов с мозгами
еще хуже.
Для того, чтобы счёт в данном случае
был «гамбургским», нужно привести хотя бы формулу, методику подсчёта этого самого
коэффициента, или первоисточник, из которого взяты данные цифры, чтобы можно
было уточнить там эту методику. Тен приводит нам лишь голые цифры неизвестного
происхождения.
Подозрительно, но дельфин у Тена получается какой-то худосочный. По данным той
же «Википедии», вес взрослой афалины – 150-300 кг (в англоязычной «Википедии»
указывается вес 150-500, максимальный – до 600 кг), вес мозга – 1800 г.
Гориллы и орангутаны своим примером подтверждают нелинейную зависимость между
размером мозга и тела, а в случае гориллы ещё относительно меньшую величину
мозга у растительноядных животных по сравнению с всеядными и хищниками.
Кроме того, здесь приводятся исключительно количественные показатели, которые
не отражают всей картины умственных способностей живых существ.
Стр. 241
Это не мешает С. Дробышевскому вопреки фактам писать:
«...Общеизвестной истиной является малая относительная величина мозга китов
и дельфинов, а преобразование лап в ласты – не лучший зачин для освоения трудовой
деятельности». Это он таким образом пытается объяснить, «почему китообразные
и ластоногие не стали разумными» (Дробышевский, 2018а, с. 288, 289).
«Вопреки фактам», значит… У Дробышевского
там же, на стр. 289, но чуть выше, о китообразных и их мозгах написано следующее:
«Еще в олигоцене они достигли церебральных показателей – по абсолютным размерам
и складчатости поверхности – уровня продвинутых гоминид (Lilly, 1977), а многие
современные виды вдвое превосходят по ним человека. Нынешние китообразные известны
своей повышенной социальностью и сложными способами общения, в том числе межвидового».
И про ласты всё понятно: это узкоспециализированная конечность с крайне ограниченными
функциями, которая сильно ограничивает возможности дальнейшей эволюции китообразных
– ласт уже не может превратиться в полноценный орган, способный выполнять иные
функции, кроме плавания.
Что же касается малой относительно массы мозга китообразных, она наглядно видна
хотя бы в «Википедии»:
у дельфиновых – от 1,40 (гринда) и 1,45 (косатка) до 5,3 (афалина), у морских
свиней – 3,54-4,9, у нарваловых – 1,76 (нарвал) и 2,24 (белуха), у кашалота
0,55-0,58 при самом крупном мозге среди позвоночных в абсолютном исчислении,
у усатых китов – 0,21-0,49.
Занятно, что у капуцинов коэффициент энцефализации оказался равен 3,4-4,8; у
шимпанзе – 2,2-3,1. Для австралопитеков приводятся величины 2,44-3,92, для парантропов
– 3,22-3,24, у ископаемых видов людей значение доходило до 4,31-5,44.
В той же статье сказано следующее: «Индекс энцефализации не даёт высокой точности
оценки интеллекта, поскольку большое значение имеет сама структура мозга. У
млекопитающих важна площадь коры головного мозга (неокортекса), которая увеличивается
за счёт мозговых извилин. Поэтому индекс энцефализации используют для выявления
тенденций развития и потенциальных возможностей различных видов».
Если продолжить исследовать довольно обширную таблицу, приведённую в статье,
то оказывается, что у утконоса коэффициент энцефализации – 0,94, у бурого медведя
– 0,91, у белого медведя – 0,53, у льва – 0,45-0,6, а у крысы – 0,4-0,79. Кто-нибудь
видел в цирке дрессированных утконосов? Вот и я о том же говорю: даже хитроумно
высчитанный коэффициент ещё не даёт объективной информации о возможностях мозга.
Крыса, отставая от утконоса по цифрам пресловутого коэффициента, в экспериментах
проявляет себя как умное и сообразительное существо.
Стр. 241:
А обезьяны стали – благодаря своей общеизвестной любви к труду и потому что
много мозгов на массу тела. За правду обидно, честное слово. А еще за Энгельса.
До сих пор на его трудовой теории паразитируют и при этом поносят повсюду и
как хотят! Ведь что у Дробышевского получается? Основной причинный фактор эволюции
– это «трудовая деятельность».
«Шимпанзе – наш ближайший родственник, самое умное существо на планете, не считая
человека», – уверяет он (там же, с. 449, цветная вкладка).
Умственные способности дельфинов явно
не коррелируют с размером их мозга. Специалист по китообразным А. Г. Томилин
в книге «В мире китов и дельфинов» (М., «Знание», 1974) даже главу 13 так и
назвал: «Мышление дельфинов конкретно, а действия трафаретны». Признавая высокую
степень развития мозга и способности дельфинов, он пишет о них следующее:
«Несмотря на свой гигантский мозг и зачатки рассудочной деятельности, дельфины
нередко совершают явно неразумные поступки, которые были бы исключены, если
бы они обладали отвлеченным мышлением, как человек. Находясь во власти инстинктов,
они зачастую не могут решать, казалось бы, весьма простые задачи. В океанариумах
дельфины часто гибнут от заглатывания посторонних предметов – кусочков стекла,
резины, веревок, тряпок и т. д. Когда дельфинов держат на привязи, они не могут
выбрать для себя наиболее безвредные формы поведения и запутываются так же,
как домашние животные, на длинных поводках. Особенно опасно привязывать дельфинов
за столбы в воде: наматывая бечевку вокруг столба и укорачивая ее, животные
погибают, лишив себя возможности выплывать для дыхания на поверхность. Чтобы
распутать веревку, им достаточно было бы сделать несколько кругов в обратном
направлении, но сообразить этого они не могут».
Если говорить о привязи, то даже обычная домашняя коза даст фору дельфину в
умении освобождаться от этой самой привязи – спросите любого козовода. В практике
наблюдений за поведением дельфинов есть и другие примеры «глупого» поведения
этих животных. Так, дельфины не догадывались перепрыгнуть через верхний край
сети, когда их ловили кошельковым неводом, выталкивали на поверхность откушенную
акулой голову сородича – таких случаев зарегистрировано немало.
Иными словами, измерить степень развития интеллекта живых существ неким единым
универсальным мерилом не удастся. Одни животные демонстрируют больше сообразительности
в решении одного типа задач, другие – иного. Поэтому дельфин вполне может превосходить
шимпанзе в решении каких-то задач, проигрывая в других.
Кстати, в запале критики Тен не заметил ляп в книге Дробышевского: на подписи
к фотографии указан вид Pan troglodytes, а на самом фото изображён бонобо Pan
paniscus.
Стр. 241:
А вот что специалисты по мозгу говорят об этом:
«Весь комплекс имеющихся морфологических данных позволяет перейти от осторожной
оценки уровня центральной нервной системы китообразных («где-то между собакой
и обезьяной») к определенному утверждению, что этот уровень выше уровня развития
мозга исследованных человекообразных обезьян и ближе к уровню человека» (Крушинская,
Лисицына, 1983, с. 125,126).
Хорошо, не спорю. Но тогда следует вопрос: свидетельствует ли это именно о том, что китообразные являются предками людей? Подтверждается ли родство данных животных и людей иными особенностями строения, генетикой, палеонтологией? Строение мозга – это лишь один признак. И определённое сходство в размерах и наличии извилин может быть результатом конвергенции – общебиологического явления. Независимо возникшая потребность в хорошо развитом органе мышления для жизни в разных, но похожих по степени сложности условиях у представителей разных групп млекопитающих привела примерно к одному результату. Выше я приводил примеры с копытом, спинным «парусом» и сабельным клыком: сходство есть, но родства оно не определяет.
Стр. 241:
«Профессор А. Портман из Зоологического института в Базеле (Швейцария) разработал
шкалу умственных способностей. По этой шкале места на своеобразном пьедестале
разума распределились для многих неожиданно. На первом месте – человек (214
баллов), на втором – дельфин (195 баллов), на третьем – слон (150 баллов). Почетное
четвертое место досталось обезьянам (63 балла)» (https://sokrovennik.ru/material/lichnost-delfina-dostojna-prav-chelovelka).
Стоп-стоп-стоп… Сам же Тен буквально на предыдущей странице говорит: «Честный математический подход может быть только один: масса мозга, деленная на массу тела». Какие баллы получаются при этом? Массу делим на массу, а в итоге получаем… некие баллы? Ведь это уже иная методика подсчёта, нечестный (по Тену) бог знает, чей ещё подход? Или, если у нас красиво получается поставить дельфина сразу после человека и низвергнуть эту противную волосатую обезьяну, шкала автоматически оказывается верной?
Стр. 241:
Интересно, что трое на пьедестале: человек, дельфин, слон – все имеют морское
происхождение. Предками слона были животные водно-земного ландшафта.
Если уж растекаться мыслию по древу, то водные предки были также у курицы и верблюда. Да, а как иначе? Какая-нибудь злополучная акантостега (или её родич) была вполне себе водной, и даже дышала жабрами. Лукавство Тена в рассуждениях состоит в том, что он не уточняет, насколько близкие предки того же слона были водными, насколько далеко по филогенетическому древу распространяется «влияние» водного образа жизни предка при смене образа жизни потомками. Если вспомнить меритерия – вполне возможно, что и да, он был полуводным. Но между меритерием и слоном укладывается целый ряд вполне наземных животных. И непосредственные предки современных слонов на уровне рода были вполне наземными животными. Возникает вопрос: на каком уровне ещё действует эффект наследования признаков от водного предка? На уровне рода, семейства, отряда? Когда кончилось влияние «водного образа жизни» меритерия в ряду от меритерия до слона? И кончилось ли оно?
Стр. 242:
К моменту начала эволюции человека на планете Земля жило только одно существо,
имевшее достаточно серого вещества для того, чтобы в нем образовался разум.
Это дельфиниды и, соответственно, их ближайшие родственники, акродельфиды. Даже
если б тогда жили шимпанзе, их 400 граммов было категорически недостаточно.
Ибо порог все же существует. Не для выстраивания «эволюционных схем», а для
понимания идиотии. «Ученые изучили более 60 случаев выживания микроцефалов.
Живут они 25-30 лет. Был только один случай, когда женщина с массой мозга в
277 граммов дожила до 74 лет, правда, в психиатрической клинике. Исследования
показали, что при массе мозга от 232 до 622 граммов микроцефалы могли произносить
отдельные слова и вести предельно упрощенную социальную жизнь» (С. Савельев,
КП, 22.07.2010; https://www.spb.kp.ra/daily/24527.3/673055/).
Помнится, на стр. 53-54 своей книги Тен
упоминал о родстве «доманов» (даманов) со слонами, и в качестве признака этого
родства упомянул особенности строения зубов. Однако родство даманов и слонов
подтверждается ещё и молекулярными данными, и анатомическими особенностями,
и палеонтологическими находками, которые взаимно дополняют и не противоречат
друг другу. Разве у нас есть какие-то сторонние данные, которые подкрепляют
идею родства человека и китообразных, выведенную исключительно на основании
похожей величины мозга? Есть у меня дурное чувство, что не дождаться мне обнародования
этих данных…
Эволюция мозга может быть настолько быстрой, что даже у близких родственников
в царстве животных мозг может иметь совершенно различное строение:
«У утконоса (Ornithorhynchus anatinus) извилины практически отсутствуют, тогда
как у ехидны (Tachyglossus aculeatus) степень их развития сравнима с высшими
плацентарными млекопитающими» (источник:
«Современные представления об эволюционном развитии и строении новой коры млекопитающих»,
Журнал анатомии и гистопатологии. 2019;8(3)).
«…закономерность, сформулированная Дарестом и Байярже (C.Dareste, J. G. F. Baillarger):
в пределах одного и того же отряда у крупных его представителей кора головного
мозга богата бороздами и извилинами (гирэнцефалы), а у мелких представителей
– бедна бороздами и извилинами (лиссэнцефалы). Так, даже в ряду наиболее высоко
стоящих в классе млекопитающих приматов есть лиссэнцефалы, напр, маленькая игрунка
(Hapale). И, наоборот, в отряде сумчатых, аплацентарных, мозг которых лишен
мозолистого тела, имеются гирэнцефалы, напр, кенгуру (Macropus)» (источник:
«Большая Медицинская энциклопедия»).
В свете этих любопытных фактов некоторое внешнее сходство в строении мозга человека
и дельфина представляется совсем не аргументом в пользу их родства.
Появление разума – это не самоцель эволюции как таковой. У эволюции нет цели,
это результат множества сиюминутных актов приспособления к изменениям условий
окружающей среды. И не факт, что эти изменения не ведут в тупик – пример аммонитов
и трилобитов тому свидетельство: сотни миллионов лет эволюции окончились тупиком
и вымиранием.
Если уж заговорили об идиотии, то мне почему-то очень хочется злобно пошутить
насчёт «эволюционных идей», описанных в рассматриваемой мной книге Тена. Но
я сдержусь.
Стр. 242:
В строении мозга следует обратить внимание на развитие коры. По абсолютному
объему коры дельфины среднего размера близки к человеку и далеко превосходят
всех прочих животных. По количеству извилин дельфины превосходят человека в
два раза.
Проверим? Пожалуй.
«Анализ микроструктуры коры показал, что общий уровень организации такого NC
[неокортекса – П. В.] довольно низок. Несмотря на большую площадь, NC китообразных
за счет небольшой толщины (1.5–2.2 мм) обладает очень малым относительным объемом
– меньшим, чем даже у насекомоядных и грызунов» («Современные представления
об эволюционном развитии и строении новой коры млекопитающих», ссылка).
Количество извилин, как показывает приведённый выше пример из этой же статьи
– не доказательство нашего родства с дельфинами:
«Конечный мозг китообразных по общему рисунку борозд и извилин отчасти напоминает
мозг копытных и хищных, но значительно отличается сильным развитием мелких борозд
и извилин (микрогирия), резким теменным изгибом (курватурой) и практическим
отсутствием височной доли» (там же).
Стало быть, по количеству коры китообразные – молодцы, а вот качество этой самой
коры – прихрамывает.
Стр. 242:
«В коре головного мозга китообразных находятся все шесть слоев, характерных
для коры мозга других млекопитающих... Строение коры на некоторых участках у
дельфинов оказывается более сложным, чем таковое у человека... Важной морфолого-физиологической
характеристикой коры головного мозга дельфинов и человека является преобладание
в ней так называемых непроекционных полей, т.е. тех областей, которые отражают
возможности абстрактного мышления, ассоциативной деятельности. Относительная
площадь областей коры, «молчащих» в ответ на раздражение органов чувств у неанестезированного
животного, у афалины сопоставима с таковой у человека» (Крушинская, Лисицына,
1973, с. 123).
А теперь в очередной раз подумаем. Шесть
слоёв клеток в неокортексе – это общий признак всех млекопитающих. То есть,
признак, не ставящий дельфинов в какое-то особое положение по отношению к человеку.
Нельзя исключать, что у китообразных в процессе эволюции какие-то зоны мозга
усложнились по сравнению с предками, а также с другими группами млекопитающих,
существующих одновременно с ними. Жизнь в сложной и фактически враждебной для
дышащего воздухом существа заставляет это сделать.
Относительно особенностей площади коры головного мозга у китообразных уже говорилось
выше. Большая площадь коры, не занятая обработкой сигналов от органов чувств
– это ещё не доказательство родства дельфина и человека. Это всего лишь количественный
показатель. Величина площади коры у китообразных и приматов могла увеличиться
независимо друг от друга – это изменчивый признак. Выше уже приводился пример,
когда у родственных друг другу утконоса и ехидны площадь коры головного мозга
резко отличалась.
То есть, данная цитата фактически не показывает никаких родственных связей между
китообразными и человеком.
Стр. 242-243:
При движении в глубь вопроса о мозге мы наталкиваемся на следующий важный критерий,
а именно: какое место в коре занимает неокортекс, ведь именно в нем сосредоточена
ответственность за самые сложные функции центральной нервной системы? У низших
плацентарных (от насекомоядных до зайцеобразных) неокортекс занимает 30-50%
площади всей коры. У хищных, копытных, ластоногих и верблюдов относительная
площадь неокортекса больше, но у большинства отсутствует такой важный признак
прогрессивного развития, как височная оперкуляризация (покрытие височных долей).
Височные доли не имеют новой коры, она отсутствует именно там, где больше всего
нужна существу с претензией стать когда-нибудь разумным. Височная оперкуляризация
в этой группе животных, выделяемых по уровню развития неокортекса, имеется только
у ластоногих. Очень интересный факт, подтверждающий, что обитание на границе
разных природных сред способствует развитию интеллекта.
И вновь никаких ссылок на научные работы,
подтверждающие данные утверждения, господин Тен не приводит, хотя ранее по книге
«зацепил» за то же самое Дробышевского.
Несмотря на большое количество информации о строении головного мозга, поисковые
сервисы Интернета не выдали ни единой ссылки на словосочетание «височная оперкуляризация».
Не исключаю, что господин Тен, будучи неспециалистом, опять что-то не понял.
Как говорится в низкопробной рекламе телемагазинов, «но и это ещё не всё!».
Вернёмся к статье «Современные представления об эволюционном развитии и строении
новой коры млекопитающих». Ведь там как раз приводится один интереснейший факт
о строении неокортекса китообразных, который уже упоминался при обсуждении написанного
господином Теном на стр. 242 – практическое отсутствие височной доли мозга.
А у приматов как раз отмечается хорошее развитие височных долей мозга.
«Неокортекс действительно возникает последним у сложных организмов, но это ни
в коем случае не означает, что он является высшей функцией организма. Неокортекс
– это всего лишь складирование разных частей функций, используемых в сознании
организмов. Полноценным сознанием обладают все самые сложные организмы, а это
млекопитающие. Сознание функционирует у млекопитающих с участием не только нейронов
неокортекса, но и целого комплекса ядер и образований нейронов, находящихся
ниже неокортекса, совместная работа которых приводит к появлению сознания у
этого организма. И поэтому работа нейронов неокортекса в осознании важна, но
она является одной из небольших частей для функционирования сознания.
Теперь по поводу височной доли и оперкуляризации (покрытие височных долей).
У разных организмов функции височной доли разные. Одинаковые функции височной
доли только у организмов одного вида, например, человека и обезьяны. И поэтому
неправильно рассуждать о покрытии височных долей корой у совершенно разных организмов,
потому что у человека это элемент умственной деятельности, инициируемой самим
человеком, а у других организмов это может быть или участие этой доли в каком-то
движении, или участие у других организмов в какого-то рода пищевой активности
(обычно неосознанной)» (комментарий Юрия Лучина).
Стр. 243:
У человекообразных обезьян неокортекс занимает от 85 до 96% поверхности коры.
У дельфинов — 97,8%, и этот показатель наиболее приближен к человеческому! (Данные
о развитии неокортекса даны по: Богословская, Поляков, 1981, с. 3—53.)
Жонглирование цифрами – это ход, который
может показаться убедительным для неспециалиста. Но специалист смотрит не только
на отдельные цифры: для него важна вся картина, где можно наблюдать те или иные
тенденции. Один из выводов в статье «Современные представления об эволюционном
развитии и строении новой коры млекопитающих» таков:
«в каждом отряде млекопитающих встречаются виды, уровень развития NC которых
не соответствует типу строения коры, наиболее характерного для данной группы;
это связано с параллельным, независимым характером эволюционного развития отрядов
млекопитающих и значительной адаптивной пластичностью мозговых структур».
То есть, в нашем случае площадь неокортекса у дельфина – это не показатель его
близости к человеку. Параллельная эволюция – только и всего. Хорошо известное
явление в природе, много раз наблюдаемое и много раз вводившее в заблуждение
специалистов по систематике, особенно в случае птиц.
Приведу ещё цитату из «Большой Медицинской энциклопедии» (ссылка):
«При исследовании мозга в восходящем ряду наземных млекопитающих обнаружено,
что увеличение поверхности полушарии, усложнение рисунка борозд и извилин соответствует
усложнению структурной организации корковой пластинки. Изучение водных млекопитающих
показало значительное развитие больших полушарий при весьма невысокой организации
неокортекса, что может свидетельствовать об определенной независимости эволюции
признаков архитектонической дифференцировки коры от величины поверхности полушарий.
Сопоставление данных площади поверхности полушарий (древняя, старая, промежуточная
и новая кора в сумме) и веса головного мозга различных представителей млекопитающих
(еж, кенгуру, муравьед, собака, свинья, тигр, тюлень, макак, павиан, орангутан,
шимпанзе, дельфин обыкновенный, дельфин афалина, кит финвал, человек) показало
прямую пропорциональную зависимость площади поверхности полушарий большого мозга
от веса головного мозга. Соотношения поверхности полушарий и веса мозга и у
ежа, и у дельфина, и у человека одни и те же, несмотря на отсутствие борозд
у ежа и их обилие у дельфина и человека. Процент площади коры, расположенной
внутри борозд (интрасулькальный компонент), казавшийся ранее эволюционно значимым,
оказался также зависящим от веса мозга (у кита — 88,1, у дельфина афалина —
85,0; у дельфина обыкновенного — 78,6 и у человека всего лишь 64,8)».
Стр. 243:
По количеству борозд и сложности их поверхностного рисунка дельфины превосходят
даже нас с вами, о чем можно судить по иллюстрациям, потому что это визуальный
показатель. Тем не менее многие специалисты зачислили мозг дельфинов в разряд
«примитивных». Почему?
Дело в том, что дельфины, превосходя всех, за исключением человека, по относительному
количеству нервных клеток, имеют чрезвычайно большое количество неспециализированных,
«первичных» нейронов.
Хороший пример, кстати. Он наглядно показывает,
что увеличение размера головного мозга может быть достигнуто либо качественными,
либо чисто количественными изменениями. То есть, налицо одинаковое внешнее проявление
признака, природа которого в сравниваемых нами случаях различна.
«То, что у дельфинов площадь борозд по соотношению со всей корой составляет
75-85%, а у человека меньше – это не говорит о том, что дельфины разумнее или
более продвинутые в эволюции по сравнению с человеком. Интеллектуальная сложность
организмов определяется толщиной неокортекса, а не площадью и количеством борозд.
У человека неокортекс образует до семи слоев, а у дельфинов и близко к этому
нет. Площадь некортекса человека закладывается по слоям 4 и 5, и соответственно,
толщина коры будет увеличиваться пропорционально усложнению функций сознания,
то есть увеличению количества слоёв неокортекса и нейронов в каждом из этих
слоёв. Такое высокое содержание борозд в некортексе дельфинов объясняется образом
жизни дельфинов: они вынуждены постоянно сознательно функционировать днём и
ночью, поэтому все их функции дублируются как левом, так и правом полушарии.
А у человека осознанная жизнь осуществляется днём, а ночью он спит, и тогда
отключаются принудительно все осознанные системы. И поэтому реально надо считать,
что у дельфина количество борозд в два раза меньше, чем есть на головном мозге.
Дельфин функционирует или правым полушарием, или левым полушарием» (комментарий
Юрия Лучина).
Стр. 243-244:
Специалисты отмечают «простоту организации дендритного аппарата нейронов» и
присутствие в слоях коры «ретикулоподобных элементов» (там же, с. 53). Речь
в последнем случае идет об элементах, выполняющих роль информационных сетевых
фильтров (от латинского rete – сеть) и активаторов. У человека ретикулярная
формация (особая структура мозга) тянется через весь мозг. Она получает информацию
от всех органов чувств, фильтрует ее и передает коре мозга. Она активирует кору
в нужные моменты, но не пропускает привычные или повторяющиеся сигналы и играет
огромную роль в механизме формирования целостных реакций организма.
А вот здесь господин Тен явно полез куда-то не туда. Как там он про Дробышевского-то говорил? Ах, да, «Слышавший звон Дробышевский» (стр. 121). Про себя господин Тен ничего подобного сказать не желает? Ну, если он лично стесняется, я скажу за него. Господин Тен, «слышавший звон», вцепился, «как кобель в падлу» (© М. Шолохов «Поднятая целина») в первое попавшееся похожее слово, и начал развивать свои умопостроения, невероятные по своей тупости. Мы о чём сейчас говорим? О неокортексе – самой эволюционно молодой части мозга, и об особенностях его строения у китообразных, в частности. А ретикулярная формация (в частности, у человека) – это филогенетически древняя структура в стволе головного мозга, а не в полушариях. Её-то к чему сюда примешивать?
Стр. 244:
Присутствие ретикулоподобных клеток непосредственно в новой коре является архаическим
признаком, свидетельствующим об архитектурной незавершенности, недостроенности
здания мозга. Но должны ли мы, исходя из этого признанного факта, «списывать»
дельфинов с нашего «Ноева ковчега», на котором мы собрали всех возможных предков
человека?
«Помимо пирамидных нейронов в NC китообразных
обнаружено большое число крупных радиальных звездчатых нейронов изодендритного
типа. Их длинные, слабоветвящиеся дендриты охватывают большую площадь, а аксонные
ветвления оканчиваются на телах и дендритах нейронов нескольких слоев. Клетки
такого типа относятся к разновидностям ретикулоподобных нейронов, являющихся
консервативно древними формами корковых нейронов, характерными для начальных
этапов эволюции NC млекопитающих. Высокодифференцированных звездчатых нейронов
различного типа в NC китообразных практически нет» («Современные представления
об эволюционном развитии и строении новой коры млекопитающих»).
Так что дело тут вовсе не в «незавершённости», а в том, что китообразные в процессе
эволюции достаточно рано свернули в удобную экологическую нишу, где качественное
усложнение мозга было ни к чему. Им хватило чисто количественных изменений,
хвала закону Архимеда, позволяющему тушке расти в воде без ограничений.
На последний вопрос отвечу: должны. У дельфиньих мозгов собственная эволюционная
история, мало похожая на таковую у приматов.
Стр. 244:
Возникает вопрос: что вы возьмете за основу, желая возвести большой, красивый
дом: большой недостроенный дом с прочным фундаментом, стенами, крышей, но без
специализации помещений или маленький, но хорошо устроенный внутри теремок,
подобный тому, в котором обитала мышка из сказки?
Здесь господин Тен допускает логическую
ошибку, которая носит название «апелляция к опыту». Вряд ли среди его читателей,
особенно оставляющих восторженные отзывы в интернет-магазинах, будет много нейробиологов,
знающих преимущества и недостатки каждого типа строения неокортекса у млекопитающих.
А простые читатели, судя по их отзывам, с удовольствием «хавают» те лженаучные
помои, которыми сочится анализируемая мной сейчас книга господина Тена, и повседневный
опыт вряд ли подскажет им правильный ответ.
Кстати, птицы выбрали второй вариант – в том числе такие умники, как попугаи
и врановые. См., например, книгу Дженнифер Акерман «Эти гениальные птицы» (М.,
«Альпина нон-фикшн», 2018), глава 2. Так что выбор далеко не очевиден.
Стр. 244:
Мозг дельфина подобен первому варианту, мозги других претендентов – второму.
Что с того, что у собаки или у обезьяны преобладают специализированные нейроны,
в мозгах нет ничего «лишнего», все «подогнано» «тютелька в тютельку»? Когда
каждая клеточка выполняет только одну свою узкую функцию и не способна действовать
по меркам других клеток, это плюс сиюминутный в рамках геологического времени.
(Мозг мал, но надежен и маловероятно, что он «сойдет с рельсов».)
А ведь эволюционный процесс как раз так
и действует – путём накопления множества именно сиюминутных выгод. У эволюции
нет цели в далёком будущем. Нет записанной в генах задачи: «через стопиццот
миллионов лет породить разумную форму». Для эволюционного процесса есть только
«здесь и сейчас»: разного рода приспособления возникают не загодя, а в ходе
последовательных многоэтапных перестроек, доводок и подгонок. Именно поэтому,
глотая пищу, мы рискуем поперхнуться – лёгкие возникли как вентральный вырост
пищеварительной трубки, и навсегда останутся таковым по своей природе. И при
прохождении по глотке пищевого комка придётся закрывать вход в лёгкие надгортанником.
Глаза позвоночного всегда будут иметь «вывернутую сетчатку» и слепое пятно там,
где аксоны нервных клеток покидают полость глаза. А возвратному гортанному нерву
даже у жирафа придётся два раза тянуться вдоль всей шеи и огибать дугу аорты,
потому что именно так обстояло дело у рыб, бывших предками наземных четвероногих.
Кто ж знал в девоне, что в кайнозое появятся жирафы? О маменьчизавре с его 10-метровой
шеей в этом плане даже подумать страшно.
Кстати, господин Тен почему-то умалчивает о том, по какому принципу построен
мозг у человека, и наблюдаются ли в его строении признаки, которые являются
производными от особенностей строения мозга китообразного.
Стр. 244-245:
Потенциально – это большой минус. В плане эволюции перспективна именно способность
сходить с ума, отвергая привычный образ действий. У дельфинид был выбор – вот
что важно. Его «простые» нейроны, подобно популярным ныне стволовым клеткам,
могли развиваться в любом направлении. Главное, что этих стандартных «кирпичиков»
хватало на дворец. У других выбора не было. Низший примат, как и собака, не
мог стать существом разумным благодаря специализированности нервных клеток,
а также общей недостаточности.
Но что удивительно: неокортекс всех современных
китообразных, несмотря на их бурную эволюцию и высокий интеллект, оказался почему-то
странным образом составлен из малодифференцированных клеток и имеет низкий уровень
организации в целом, что отмечено в вышеупомянутой статье. Выбор-то, может,
и был, но он оказался не реализованным в процессе эволюции. Строение неокортекса
показывает, что мозг китообразных нарастал главным образом количественно, а
не качественно.
Я предлагаю обдумать сравнение со стволовыми клетками, которое нам предлагает
господин Тен. Эта клетка реализует свой потенциал, когда превращается из неспециализированной
в специализированную. То есть, если применить эту логику к строению мозга, высокоразвитым
будет именно тот мозг, в котором примитивные исходные клетки также реализовали
свои возможности и стали специализированными. А у китообразных нейроны «простые»,
что отмечает сам Тен. Простое строение – простое функционирование. У человека
и остальных приматов строение неокортекса сложное и специализированное. И факт
наличия у человека разума говорит о том, что он достижим именно при специализированном
строении нейронов, что противоречит словам господина Тена.
Теперь подумаем, какова ценность для выживания у «способности сходить с ума»?
Для начала определимся со значением слова. Сборник
словарей Glosum довольно единодушно предлагает нам такие значения, как «психическое
расстройство, умопомешательство». Переносные значения рассматривать не будем
– на то они и переносные, чтобы не точно отражать явление.
И возникает вопрос: способствует ли выживанию особи в частности и вида в целом
такое поведение, которое не адекватно условиям среды? Естественно, нет. В природе
нет права на ошибку. Природа быстро выписывает таким «неадекватным» товарищам
«премию Дарвина».
Что такое «общая недостаточность», из книги Тена непонятно. Недостаточность
чего? Пока я наблюдаю у одного представителя приматов недостаточность биологических
знаний, изложенную в книге, и у целой кучи других приматов из издательства «Эксмо»
недостаточность критического мышления, которая позволила им подписать в печать
эту книгу, а также недостаточность совести, позволяющую им скармливать несведущему
читателю низкопробную дрянь под видом научно-популярной литературы.
Стр. 245:
Позволю себе еще одну цитату: «Эти и некоторые другие данные позволяют думать,
что, вероятно, именно у китообразных реализован тот способ построения нейронной
системы, теоретически предсказанный Лешли (в 1929 г. – В. Т.), при котором возможности
системы к обучению и памяти определяются количеством функционирующей коры, а
не ее анатомической специализацией. В недавнее время Ренш (в 1965 г. – В.
Т.), исходя из других посылок, также пришел к убеждению, что число взаимодействующих
элементов – самый важный показатель интеллектуальных способностей нервной системы»
(Богословская, Поляков, 1981, с. 53).
Вот и я о том же – в мозге китообразных наблюдаются количественные, а не качественные изменения в строении неокортекса. Единодушие различных авторов различных статей и книг, написанных в различное время, лишь подтверждает это.
Стр. 245:
Выходит, что в случае с зубатыми китами природа пошла по другому пути, чем в
случае со всеми другими животными. Она выделила их в какой-то свой эксперимент.
Уже эта исключительность должна наводить нас на размышления, ибо человек тоже
исключителен. Вместо небольшого количества узкоспециализированных нейронов природа
дала дельфинам много универсальных нервных клеток, которые призваны были «брать
количеством».
Я больше скажу: любой самый банальный
суслик на полях колхоза «40 лет без урожая» тоже исключителен. Любая изысканнейшая
орхидея тропического леса – исключительна. Любая глубоководная рыба самого страхолюдного
облика – тоже исключительна, единственная в своём роде.
Тен упрямо молчит об особенностях строения неокортекса у человека, хотя в данном
случае разговор об этом был бы крайне уместен. Именно сравнение, указание множества
параллелей, преемственности в строении – вот то, что должно однозначно и бесповоротно
убедить читателя в правоте господина Тена. Прежде, чем мы продолжим разговор,
я напомню его фразу, неосторожно и крайне неосмотрительно написанную на странице
205:
«Строение мозга при прочих равных обстоятельствах имеет при установлении
таксономических различий приоритет».
Это очень хорошая фраза в данном случае. Хорошая в том плане, что её написал
не кто иной, как сам господин Тен. Это означает, что данная фраза отражает именно
его собственные воззрения в данной области биологии. А теперь вернёмся к статье
(«Современные представления об эволюционном развитии и строении новой коры млекопитающих»).
В этой статье авторы описывают четыре типа неокортекса по строению. Я не стану
цитировать все особенности строения, отличающие один тип от другого: кому это
интересно, тот прочитает об этом в самой статье – для этого я привожу ссылку.
Приведу лишь распределение этих типов строения по таксонам млекопитающих:
«Первый тип неокортекса встречается у современных представителей таких отрядов
млекопитающих, как: однопроходные, сумчатые, насекомоядные, рукокрылые, грызуны,
зайцеобразные и ряда других».
«Второй тип неокортекса сформировался в ходе эволюции таких отрядов млекопитающих
как: хищные, ластоногие, копытные, хоботные».
«Третий тип неокортекса сформировался у приматов. Это один из древнейших отрядов
млекопитающих, и не удивительно, что именно в пределах этого отряда уровень
межвидовой и межсемейственной изменчивости очень велик».
«Четвертый тип неокортекса представлен у китообразных».
Замечу, что авторы не выделяют строение неокортекса у человека в какой-то особый
тип, рассматривая его особенности наравне с таковыми у прочих приматов.
А теперь возвращаемся к процитированной мною фразе господина Тена со стр. 205.
Итак, как же нам рассматривать родство китообразных и человека, отталкиваясь
от строения их мозга? Если строение мозга имеет приоритет, то особенности строения
неокортекса у приматов и китообразных говорят строго в пользу разделения тех
и других, и однозначного нахождения человека среди приматов, а не китообразных
и их гипотетических потомков. Если бы человек действительно происходил от неких
дельфиноидных предков (несомненных представителей отряда китообразных), то на
основании этого знания мы могли бы предсказать, что в мозге человека будут наблюдаться
признаки, характерные для китообразных, чётко отделяющие его от приматов.
Можно, конечно, предположить, что мозг «человека – потомка дельфиноидов» независимо
приобрёл в процессе эволюции особенности, характерные для мозга примата. Но
тогда возникает новый вопрос: какова вероятность независимого повторения в другой
эволюционной ветви такого огромного количества признаков, которые придают мозгу
«человека – потомка дельфиноидов» сходство с мозгами приматов «до степени смешения»?
Каким образом неокортекс китообразных должен утратить свои признаки специализации
в строении (а они есть), чтобы они вообще не проявлялись в строении неокортекса
существа, которое сватают ему в потомки? Какие движущие силы эволюции заставили
это сделать? Боюсь, если мы применим в наших рассуждениях ту самую хвалёную
«бритву Оккама» (стр. 84 книги Тена), то она в данном случае сработает не хуже
ножа для рубки капусты, разрубая в крошево умопостроения господина Тена.
Стр. 245:
Известно, что, когда природа хочет наказать, она дает слишком много. Не случайно,
говоря о дельфинах, мы оказались вынуждены использовать понятие универсальность
в качестве объяснительного принципа. Универсальность является основным отличительным
качеством также и человека как вида.
Здесь мы наблюдаем нечестный приём полемики,
называемый «апелляция к авторитету», подвид «анонимный авторитет»:
«Анонимный авторитет – утверждения с этой ошибкой содержат ссылку к определённому
весьма широкому кругу лиц, которые обычно пользуются авторитетом; конкретика
(например, фамилия, место работы и пр.) при этом не указывается. "Ученые
считают, что...", "есть мнение, что..." и т. д.» (источник).
Соответственно, хотелось бы увидеть, на чём основано подобное утверждение. Научный
первоисточник подойдёт.
В рассуждениях относительно качества универсальности применительно к дельфинам
и людям автор пытается подменить универсальность нейронов в мозге дельфина относительной
неспециализацией тела у человека как вида. Простое плутовство. «Лисячья правда»
– так, кажется, говорит сам господин Тен о таких вещах?
Стр. 245-246:
А вот что касается специализированности, то здесь благодаря науке об антропогенезе
(которая, занимаясь обезьяной, в целом пришла к отрицательному результату, но
отрицательный результат – тоже результат) все ясно. Именно: специализация является
самым страшным врагом эволюции. Там, где она выявляется, эволюция прекращает
свою работу. Ситуация выбора дважды не воспроизводится никогда. Сделав выбор
в пользу узкоспециализированного развития, вы попадаете в уютный тупик, который
всем хорош, кроме одного: в случае резкого изменения природных условий вы вымрете-с.
Что и произошло со многими видами.
Рассуждая о специализации, господин Тен
путает кислое с пресным. Специализация – не враг эволюции, между этими двумя
понятиями совершенно иные отношения. Специализация – это один из итогов эволюции,
поскольку она возникает и усиливается именно в процессе эволюции. Специализация
помогает избавляться от конкурентов, занимая какую-то крайне специфичную экологическую
нишу.
Если специализация заводит в тупик и вид вымирает – в этом нет ничего сверхъестественного
и противоестественного. А поскольку условия среды неизбежно меняются со временем,
любой вид рано или поздно сталкивается с опасностью вымирания. Это грозит даже
человеку. Но, пока условия среды относительно стабильны, узкая специализация,
напротив, благоприятствует выживанию благодаря отсутствию конкурентов.
Стр. 246:
Человека всегда спасала именно его универсальность, которой он обязан своим
предкам-археоцетам, имевшим большой мозг с универсальными нейронами.
А какие научные данные говорят о том, что у археоцетов был именно такой мозг, именно с такими нейронами? У господина Тена есть методика изучения особенностей нервной системы исчезнувших видов? Или это очередной домысел?
Стр. 246:
Видимо, с этим связано явление, которое специалисты называют «удивительной курватурой»
конечного мозга дельфинов (от «курва» – извилина). Такого количества извилистых
мелких борозд неокортекса нет более ни у кого, включая человека.
Слово «курватура» означает «кривизна,
изгиб линий» и относится к общим очертаниям органа (в нашем случае мозга). Возможно,
господин Тен не знает, но курватура есть даже у желудка, причём при очевидном
отсутствии в строении желудка такого элемента, как извилины. Мало того, в желудке
их целых две: большая и малая кривизна. Применительно к мозгу у китообразных
отмечается резкий изгиб (курватура) теменной части полушарий. Всего лишь. Это,
кстати, упоминалось в цитате при обсуждении содержимого стр. 242 анализируемой
книги Тена.
Напомню, что количество извилин – признак изменчивый, чисто количественный,
подверженный влиянию множества факторов, среди которых есть также размер тела
(с положительной корреляцией). То есть, строительство филогенетических связей
на основании одного лишь этого признака чревато ошибкой.
Стр. 246:
Борозды на поверхности коры мозга предназначены для разделения зон ответственности
отдельных популяций нейронов. В грубом приближении поверхность неокортекса можно
сравнить с современными электронными платами. На заре развития электроники отдельные
элементы схемы соединялись при помощи пайки проводами. Ныне схемы монтируются
на плате-проводнике со штамповкой непроводящих борозд, разделяющих функциональные
поля. Эти непроводящие борозды сравнимы с извилинами мозга, разделяющими популяции
клеток мозга. Дефицит специализации, то есть типологической различенности клеток,
требует скрупулезного распределения их по зонам ответственности, чтобы они не
пересекались и не мешали друг другу, составляя тем не менее одно целое, «единство
во многообразии». «Курвы» мозга дельфинов, т.е. крутые повороты извилин, составляют
сложный лабиринт, а не скопление тупиков.
«Конечно, не правомерно сопоставлять
борозды неокортекса и электронные платы. Это как сравнивать зелёное с круглым.
Там всё различается. Так как неокортекс является складированием разных элементов,
используемых в сознании, то их форма зависит от начала в эволюции у данных видов
появления разных сознательных функций. Это мы можем видеть в развитии неокортекса
в плоде человека во время беременности. Первыми будут появляться те борозды
или круги, которые раньше появились в эволюции у данных видов. Расположение
борозд, их величина и количество зависит от величины элементов, расположенных
в данной функции организма в неокортексе. Длинные, глубокие борозды и большое
их количество в неокортексе показывает, что осознанная одна функция представлена
большим количеством элементов в неокортексе. Под элементом понимается не отдельный
нейрон, а осознанная функция, допустим, осознанное движение руками или движение
глазным яблоком и так далее. Здесь необходимо заметить, что неокортекс формируется
сверху головного мозга, ранее образованного, в который входят разнообразные
ядра, проходящие пути. И поэтому неокортекс или большое количество разнообразных
элементов и их деталей, которые там находятся, не может иметь безбороздную форму,
то есть быть полностью гладким. Тогда бы получился просвет между неокортексом
и теми ядрами, проводящими путями, которые к этому моменту уже сформировались
в мозге.
Борозды, которые не похожи на участки борозд, рядом расположенные, показывают,
что здесь находятся элементы и их нейроны, которые у данных видов появились
в сознании в другое время. Поэтому в организме такая пёстрая картина борозд.
В первую очередь у сложных видов, в том числе у человека» (комментарий Юрия
Лучина).
Стр. 246-247:
В современной электронике наблюдается тенденция к унификации деталей, модульности.
Современная полупроводниковая микросхема, являясь в принципе чрезвычайно простым
устройством (гораздо более простым, чем былая ламповая схема), способна выполнять
множество различных функций. У ламп была узкая специализация, они не были взаимозаменяемыми.
Вопрос: не работают ли скопления простых нейронов мозга дельфинов по типу многофункциональных
микросхем, избежав узкой специализации, и это не недостаток, а, наоборот, прогрессивный
признак? Не примитивизм, а, наоборот, качественно более высокий этап, чем тот,
который явлен во взаимодействии узкоспециализированных нейронных систем у других
животных? Ламповый этап в развитии электроники тоже ведь не являлся необходимым.
Электроника в принципе могла бы и обойти его, развиваясь на полупроводниках.
Если продолжать сравнения с мозгами, то большая часть животных пошла по пути
дальнейшего развития по типу ламповой электроники: все больше специализированных
ламп, пока не наступает предел вместимости. А дельфины пошли по пути, похожем
на электронику полупроводников, избежав узкой специализации нейронных популяций.
Простите, я что-то немного недопонял
этот момент. Господин Тен пытается нам что-то доказать, не так ли? Пытается
– целую книгу об этом написал. Тогда он должен не сыпать вопросами типа «а может
быть… ?», «а если… ?», перекладывая на читателя обязанность отвечать на эти
вопросы. Так дела не делаются. Если уж господин Тен хочет что-то доказать, он
должен излагать материал не в виде глубокомысленных вопросов, а ставить читателя
перед массивом фактов: вот один факт, вот первоисточник; вот второй факт, вот
первоисточник… вот N-ый факт, и вот первоисточник. На основании фактов 1, 2,
… N можно сделать вывод о том, что положение дел обстоит именно так-то и так-то.
В биологии понятие «примитивный» толкуется вполне однозначно – как «близкий
по строению к особенностям предковых форм». Приматы, кстати, во многих особенностях
строения оказываются примитивными. Если возвращаться к статье «Современные представления
об эволюционном развитии и строении новой коры млекопитающих», то её авторы,
описывая строение неокортекса китообразных, дают ему такие характеристики:
«Подобные пирамидные нейроны встречаются в NC низших млекопитающих».
«Клетки такого типа относятся к разновидностям ретикулоподобных нейронов, являющихся
консервативно древними формами корковых нейронов, характерными для начальных
этапов эволюции NC млекопитающих».
Так что надежды господина Тена на то, что особенности строения неокортекса китообразных
– это (внезапно!) прогрессивный признак – откровенно напрасны. Специалисты высказали
своё мнение, сравнивая мозги китообразных и низших млекопитающих. Но, если переопределить
понятия и назвать низших млекопитающих высшими…
Возникает ещё один вопрос – означает ли неспециализация нейронов в неокортексе
дельфинов их многофункциональность? Господин Тен предлагает нам некую аналогию,
но он не доказал, что в живой природе всё обстоит именно так же, как в радиотехнике.
Чтобы это доказать, он должен сравнить функциональные возможности неспециализированного
нейрона из коры головного мозга дельфина и специализированного – допустим, из
мозга собаки или обезьяны, а лучше двух – человекообразной и мартышкообразной
либо широконосой. Пока же он предлагает читателю удовольствоваться аналогией,
и не факт, что корректной. Такая логическая ошибка носит название «ложная аналогия».
Стр. 247:
Изборожденность неокортекса мозга дельфинов вызвала, в свою очередь, явление,
которое тоже удивляет специалистов. Курватура конечного мозга вызвала округление
всего мозга по типу, прошу прощения за вольное сравнение, хлебного каравая с
бороздами на поверхности. Мозг дельфинов – самый круглый в природе. Он даже
более округлен, чем мозг человека. Круглый мозг, между прочим, – эволюционно
прогрессивный признак. У низших плацентарных и у большинства высших мозг имеет
вытянутую, продолговатую форму. Круглая форма мозга – это, согласно современным
представлениям, тоже важный показатель высочайшего уровня развития.
Здесь господин Тен ставит телегу вперёд
лошади.
«Относительно механизма возникновения борозд и извилин в процессе развития единого
мнения не существует. Полагают, что полушария растут неравномерно в различных
своих частях, вследствие чего и напряжение поверхности меняется в отдельных
участках; в свою очередь это должно вести к образованию складок или извилин.
Но, возможно, известную роль играет и первичный рост борозд, и, т.о., в какой-то
мере извилины возникают вторично» (источник:
«Большая Медицинская энциклопедия»).
«Ученые продемонстрировали, что в мозге с извилинами наличествует большое количество
клеток-предшественников, которые производят множество молодых нейронов, мигрирующих
к коре в процессе развития мозга. В результате этого движения клеточный слой
мозга заполняется нейронами. С помощью некоторых механизмов, ограничивающих
рост коры, формируются извилины, чтобы для собравшихся клеток было достаточно
места. Эксперименты на мышах с искусственно увеличенным количеством клеток-предшественников
показали, что одного только этого изменения недостаточно для появления извилин
и борозд, поскольку в некоторых случаях у мышей кора стала толще, но в целом
осталась гладкой» (источник).
То есть, механизм возникновения борозд и извилин на поверхности мозга не оказывает
влияния на очертания мозга в целом. На форму головного мозга влияет развитие
отдельных его частей. Например, развитые обонятельные луковицы придают мозгу
удлинённую форму. У приматов обонятельный мозг довольно сильно редуцирован (приматы
– микросматики) в связи с преобладанием зрения как источника получения информации
об окружающем мире. У китообразных также происходит редукция обонятельного мозга,
но она имеет иные причины – полую бесполезность обоняния в водной среде при
закрытых верхних дыхательных путях. И у тех, и у других произошло увеличение
размеров мозга в связи с жизнью в сложной среде обитания, хотя эта сложность
имеет различный характер. Так что сходство очертаний мозга человека и дельфина
сугубо внешнее. В целом же очертания мозга определяются очертаниями полости
для него в черепе. И при деформации черепа (искусственной или патологической)
форма мозга неизбежно меняется. А когда мозг не сохраняется (у ископаемых животных,
например), о его строении можно косвенно судить по полости черепа, в которой
он размещался. Например, слепки полости черепа часто упоминаются в книге Г.
Мчедлидзе «Основные черты палеобиологической истории китообразных», которой
пользовался господин Тен при подготовке своей работы.
Кстати, хотелось бы увидеть ссылки на первоисточники, где специалисты высказывают
это самое удивление очертаниями мозга. Но я уже привык к тому, что господин
Тен не очень щепетилен в вопросе предоставления доказательств для своих слов,
хотя от оппонентов требует это исправно.
Наконец, сам же господин Тен говорил, что внешнее сходство – это ость бывает
обманчива (стр. 129 его книги): «Внешность может быть обманчива, нельзя
начинать с подобия. В эволюционной биологии фактор похожести не работает слишком
часто, чтобы ему можно было верить». Поэтому, отталкиваясь,
в том числе, от слов самого господина Тена, можно со спокойной душой признать,
что внешнее сходство очертаний мозга человека и дельфина – это не показатель
их родства. Просто совпадение в общих геометрических очертаниях, не исключающее
огромной разницы в строении коры головного мозга.
Честно, мне приятно, когда слова господина Тена обращаются против него же самого.
Вот здесь как раз тот самый случай.
Посмотрев на форму головного мозга человека и дельфина, я могу сказать, что
мозг дельфина весьма далёк от хвалёной шаровидной формы – он сильно расширен,
его ширина превышает длину (у человека – чуть меньше длины). А очень крупный
мозжечок (крупнее, чем у человека) ещё сильнее искажает хвалёную шаровидную
форму мозга дельфина.
Стр. 247-248:
«К дельфинам нужно относиться как к «личностям», их права на жизнь и свободу
нужно уважать», — заявили ученые, обсуждавшие эту тему на форуме в Канаде. Специалисты
по философии, охране природы и поведению животных хотят добиться поддержки «Декларации
прав китообразных». Они уверены, что дельфины и киты обладают достаточным разумом,
чтобы оправдать применение к ним таких же этических принципов, как для людей.
Признание их прав означало бы конец китобойному промыслу и неволе, а также использованию
дельфинов и китов для развлечения. Решение было принято на ежегодном собрании
Американской ассоциации по продвижению науки, которое проходило в канадском
городе Ванкувере. Это крупнейшая научная конференция мира Вывод основан на многолетних
исследованиях, показавших, что дельфины и киты обладают крупным и сложно устроенным
мозгом, а также самосознанием, близким к человеческому уровню (https://sokrovennik.ru/
material/lichnost-delfina-dostojna-prav-cheloveka).
Источник, конечно, желтоватый, далёкий
от науки, но в данном случае не это имеет значение. Даже если поведение китообразных
будет признано достаточно сложным, чтобы законодательно закрепить отношение
к ним как к личностям, это не изменит их положения в системе природы. И вдобавок
этот факт станет лишним поводом признать, что человеческое сознание – это не
исключительный феномен нашего вида, и что уровня мыслительной деятельности,
сопоставимого с человеческим (хоть и не равного человеческому), могут достичь
различные биологические виды эволюционным путём. Это лишний раз покажет, что
разум человека суть порождение природы, лишь одно из многих приспособлений для
выживания.
Да, мозг дельфинов крупный, но вот объективные данные не позволяют считать его
сложно устроенным: выше уже говорилось об особенностях строения коры головного
мозга китообразных, ставящих их особняком от прочих млекопитающих и роднящих
с примитивными представителями класса. То есть, дельфины стали башковитыми благодаря
количественному увеличению, а не качественному усложнению мозга.
Стр. 248:
По двум очень значимым показателям, характеризующим возможности мозга, дельфины намного отличаются от человека. Настолько отличаются, что отдельные антропологи (например, М. Урысон), основываясь на этом, призывают прекратить все разговоры о дельфинах как родственниках людей. Речь идет о типе расположения борозд. У человека и человекообразных обезьян борозды расположены радиально, у дельфинов – циркулярно.
В этом месте не помешает рисунок – схема
внешнего строения и извилин на поверхности мозга человека и дельфина, которая
иллюстрирует слова господина Тена, потому что строить пальцами в воздусях разного
рода фигуры – дело нехитрое, но бесполезное. А рисунок сразу облекает мысли
в конкретную форму, не заставляя читателя домысливать за автора, что же на самом
деле имеется в виду, и позволяющую вести предметный разговор.
Конечно, здесь мои слова относятся больше не к автору, а к издателям, которые
не поместили в книгу ни одной иллюстрации. Впрочем, если такова была воля самого
автора, то моё замечание относится и к нему. Ведь давать как можно меньше конкретики
– это тоже защитный приём, позволяющий в нужный момент взвизгнуть «а я вовсе
не это имел в виду!» и раствориться в тумане.
Слова М. Урынсона полностью поддерживаю.
Стр. 248:
Для прогрессивной эволюции любое, самое маленькое различие в строении мозга
имеет большее значение, чем различие в строении тела, даже если последнее весьма
значительно, поэтому мимо данного возражения пройти нельзя.
Возникает закономерный вопрос: а почему
прогрессивная эволюция должна выражаться именно в изменении строения мозга?
Я напомню, что у нас есть такие понятия, как морфофизиологический прогресс/регресс
и эволюционный прогресс/регресс, и они не всегда идут рука об руку. Как всегда,
господин Тен не приводит мнение биологов (он сам – не биолог), не делает ссылок
на первоисточники, где изложена данная точка зрения. Это не есть хорошо – проверить
его слова нельзя. Однако в статье «Современные представления об эволюционном
развитии и строении новой коры млекопитающих» и в БМЭ (статья «Борозды и извилины
головного мозга») указывается, что даже родственные формы могут значительно
отличаться друг от друга строением мозга.
Если мы посмотрим на процесс усложнения мозга у тех же млекопитающих в целом,
то заметим одну закономерность. После мел/палеогенового вымирания млекопитающие
быстро эволюционировали, осваивая разнообразные жизненные формы – примерно до
конца эоцена. И тогда их мозги были весьма невелики, даже у крупных видов вроде
уинтатериев и пантодонтов. Но в олигоцене началось качественное изменение млекопитающих
– многие из них параллельно друг другу начали приобретать более крупный мозг,
одновременно вытесняя представителей архаичных отрядов и семейств, которые не
отличались особо развитым мозгом. Умнее стали и хищники, и их жертвы. То есть,
размер мозга – это не какой-то эксклюзивный признак немногих «избранных» групп
животных, а один из многих признаков, подвергающихся эволюции. Потому и рассматривать
его надо не «вместо», а «вместе», в контексте изменения других признаков. Потому
что у нас эволюционирует организм в целом, а не гигантский инопланетный мозг
без тела, который из мультсериала «Футурама».
Стр. 248:
Биологи-систематики давно перестали принимать во внимание такой фактор, как
внешнее различие видов. Они знают: за сравнительно короткий геологический отрезок
времени эволюция способна сотворить из слона норного грызуна, из крысы — «оленя»,
но строение мозга при этом останется однотипным.
Вот именно, что диагностическими признаками
являются другие, не бросающиеся в глаза. Поверхностное сходство общих очертаний
тела может быть результатом конвергенции, а может и не быть. Поэтому внешнее
различие видов вовсе не перестали принимать во внимание. Просто стали искать
действительно уникальные систематические признаки и смотреть внимательнее.
Кстати, это произошло не так уж и давно – в основном, как только появились технологии,
позволяющие считывать последовательность азотистых оснований в ДНК и строение
белков. Сходство и различие на молекулярном уровне не поддаются конвергенции,
как анатомические признаки, и очень многие вещи в систематике оказались повёрнутыми
с ног на голову. Эх, вы бы знали, господин Тен, что биохимия сотворила с родословным
древом птиц, построенным, например, к концу XX века… Достаточно сравнить данные
из «Большой Советской энциклопедии» 70-х годов с современным родословным древом
птиц.
Но, опять же, биохимические критерии прекрасно подтверждаются доказательствами,
предоставляемыми нам другими биологическими дисциплинами – например, палеонтологическими
находками. Так, палеонтология подтвердила предсказанное по биохимическим данным
родство китов с парнокопытными – у родоцетуса (Rodhocetus) из семейства протоцетид
(Protocetidae) нашли таранную кость (астрагал) типичного для парнокопытных строения.
Я вот всё думаю: когда же господин Тен начнёт припечатывать коварных «симиалистов»
конкретными названиями и признаками ископаемых организмов, подтверждающими его
умопостроения? Пока этого не было. Дождусь ли?
Стр. 248-249:
Этот подход биологов-систематиков позволяет (спасибо М. Урысону с его решительным
«нет», основанным исключительно на строении мозга) с ходу опровергнуть все возражения
против дельфинов как возможных предков людей, основанные на внешнем несходстве.
Внешность не имеет никакого значения. Человеческий облик за 5 миллионов лет
могла бы принять даже лягушка, если б имела просторные мозги и начала эволюционировать,
избегая узкой специализации.
Таких «опровергателей-с-ходу» в тех же
интернетах целые косяки встретить можно. Одни опровергают шарообразность Земли,
другие – всю теорию эволюции оптом, третьи – происхождение человека от обезьяны.
Только вот осведомлённость у этих «опровергателей» зачастую оставляет желать
лучшего – порой они не знают элементарных вещей, и не очень сложные уточняющие
вопросы по теме заставляют их извиваться, как уж на сковородке. Здесь у нас
тот же случай, неоднократно подтверждавшийся.
В частности, я думаю, читатели уже заметили одну вещь в рассуждениях Тена. Он
говорит нам, что «внешность не имеет никакого значения», но не говорит о том,
какие же критерии в данном случае всё же имеют это самое значение. Я думаю,
это неспроста. Ведь тогда проверка его же собственных выкладок с точки зрения
признаваемых им же самим критериев может привести к не очень хорошему (для него
же самого) результату. Думаю, именно поэтому Тен в своих рассуждениях избегает
конкретики – например, «размывает» тех же «дельфинид» в собственном понимании
этого термина чуть ли не до всех зубатых китов вкупе с археоцетами, что вряд
ли примут, поймут и простят профессионалы-цетологи.
А если начинается конкретика, начинаются страдания. Вот, стоило только господину
Тену заикнуться о том, что архиважное значение для систематики имеет строение
мозга, как оказалось, что и по строению мозга китообразные резко отличаются
от приматов, а человек, напротив, никак не выделяется из общего числа приматов.
Об этом мы говорили при обсуждении стр. 244-245 его труда (почему мне в голову
всё время лезет эпитет «мартышкиного»?).
Далее, если б у бабушки были усы, она б всенепременнейше была дедушкой. Мозги
– штука дорогая, ибо энергоёмкая, поэтому физиологии организма земноводного
попросту не хватит для обеспечения хорошо работающего мозга питанием и кислородом.
Это всё равно, что улитке пришпандорить крылья и заставить её взлететь. А уж
насколько человеческим был бы облик «батрахантропуса» – пусть это останется
на совести самого Тена. В течение большей части мезозоя и всего кайнозоя общий
облик лягушек оставался относительно узнаваемым.
Я не могу понять, как можно эволюционировать, «избегая» узкой специализации.
Это диктуется не желанием вида, а открывающимися перед ним возможностями, и
сознательный выбор тут не поможет.
Стр. 249:
Я не только не намерен проходить мимо трудности, на которую ссылается М. Урысон,
но приведу еще одну.
У дельфинов, в отличие от человека, слабо развито мозолистое тело – главная
комиссура мозга, которая, как написано мной выше, является главным критерием,
выделяющим мозг человека.
Возникает закономерный вопрос: верен
ли критерий, предложенный господином Теном? Он не ссылается ни на один специализированный
источник из области биологии или медицины, но утверждает это как нечто само
собой разумеющееся.
Специалисты выделяют несколько иной критерий, ставящий человека в особое положение
среди млекопитающих по строению мозга:
«Процесс эволюционных изменений корковых территорий больших полушарий головного
мозга от низших млекопитающих к высшим и человеку заключается в тенденции дивергентного
(«разнонаправленного») развития различных по генезу зон. Так, новая кора по
относительной величине ко всей коре значительно увеличивается, а древняя кора
явно уменьшается. И если по относительной величине новой коры человек почти
не выделяется из отряда приматов и даже уступает дельфину, то по соотношению
абсолютной площади новой и древней коры, как показателю максимальных девиаций
(ПМД), значительно превосходит и приматов, и китообразных» (источник: «Большая
Медицинская энциклопедия», ссылка).
Итак, как обстоят дела у человека с мозолистым телом? Площадь сечения мозолистого
тела у нашего вида – 700 квадратных миллиметров (ссылка).
А у дельфинов? Вернёмся к статье “Corpus Callosum Size in Deiphinid Cetaceans”
(ссылка),
которую мы уже упоминали, когда обсуждали страницу 198 книги Тена. Напомню,
у исследованных авторами видов дельфинов площадь сечения мозолистого тела составляет
от 104,1 до 828,6 квадратных миллиметров. Лишь синий кит с 850,1 кв. мм опередил
косатку в данном источнике. Отношение площади сечения мозолистого тела (кв.
мм) к весу мозга (г) у дельфинов из данной статьи колеблется в пределах от 0,079
до 0,310. У китов из разных семейств зубатых и усатых китов значения этого показателя
лежат внутри диапазона этих значений, кроме гангского дельфина (0,431). У калифорнийских
морских львов это отношение составляет от 0,348 до 0,529, у моржа 0,151, зато
у карибского ламантина – 0,829. У индийского слона значение – 0,181.
А если мы посчитаем это отношение для человека? Если условно взять вес мозга
равным 1200 г, а площадь сечения мозолистого тела – упомянутые 700 кв. мм, то
их отношение оказывается равным 0,5833. А теперь смотрим на морского льва и
ламантина, потом на себя, потом снова на морского льва и ламантина, потом опять
на себя… Так чем же наш мозг лучше-то? В абсолютном исчислении проигрываем одним,
в относительном – другим. Стало быть, уникальность мозга человека определяет
иной критерий, а Тен в очередной раз пытается выдать желаемое за действительное
и «протолкнуть» собственную идею «в массы», не удосужившись элементарно проверить
её.
Стр. 249:
Этот аргумент против близкого родства дельфинов и человека даже более значим,
чем внешний рисунок неокортекса, потому что касается глубинных вопросов обеспечения
работы мозга.
Но ради красоты своих умопостроений Тен,
видимо, готов отбросить этот аргумент? Иначе по какой причине была написана
его книга? Второе издание, между прочим! Если сам господин Тен признаёт наличие
аргументов (по его словам – значимых!) против вывода о родстве человека и дельфина,
но, однако же, пишет книгу именно об этом самом родстве – напрашивается вопрос:
как он поступил с этими самыми аргументами?
Внешний рисунок неокортекса не имеет особого значения для систематики млекопитающих.
Выше уже говорилось, что рисунок извилин возникал независимо в различных крупных
группах млекопитающих (на уровне надотряда и выше), и что внутри одной и той
же группы млекопитающих рисунок и глубина извилин мозгут значительно различаться.
Гораздо более значимым аргументом (с точки зрения специалистов, по крайней мере)
является строение неокортекса китообразных, ставящее их особняком среди класса
млекопитающих вообще и отделяющее от приматов в частности. Об этом также говорилось
выше, при обсуждении стр. 245 книги Тена.
Стр. 249:
Впервые на такую особенность рода Homo (включая древних гоминид) и человекообразных
обезьян, как радиальность (поперечность) борозд конечного мозга, обратила внимание
наш самый авторитетный палеоневролог В.И.Кочеткова, ученый с мировым именем
(Кочеткова, 1973, с. 40). Ее «Палеоневрология» до сих пор является «культовой»
книгой, во многом непревзойденной ни у нас, ни на Западе. Будем отталкиваться
от нее, тем более что М. Урысон в своем категоричном требовании «прекратить»
все разговоры о дельфинах опирается на нее же.
Если «радиальность» понимать как поперечность (именно так разъясняет смысл данного
термина Кочеткова), то как понимать «циркулярность»? Как продольность? С целью
избежать двусмысленности уточним понятия. Мы можем это сделать самостоятельно
(В.И. Кочеткова, к сожалению, уже скончалась), потому что имеем фото мозгов
дельфинов и человека.
К сожалению, очень многие другие понятия
Тен даже не пытался уточнить, считая их чем-то самим собой разумеющимся, а части
понятий (таким, как термину «брахиация») присвоил какие-то собственные, нигде
больше не встречающиеся значения.
«Поперечность или продольность борозд у человека определяется как количеством
органов, используемых в процессе осознания, так и сложностью самой первичной
функции в сознании. Граница между инициированными осознанными процессами и испытываемыми
осознанными процессами в неокортексе проходит по центральной борозде – спереди
инициированные, а сзади испытываемые. Извилина образуется по очередности расположения
органов у человека – например, двигательный гомункулус, а толщина извилины определяется
количеством и сложностью функции существ у человека, например, тактильным ощущением
у человека. Это поперечные извилины. Различного рода продольные или круговые
извилины – это результат появления новых функций в организме у данного вида
с развитием зрительной системы, слуховой системы и других, и сложностью этого
развития» (комментарий Юрия Лучина).
Стр. 249-250:
Думаю, что Кочеткова имела в виду под циркулярными бороздами те, которые очерчены
как бы широко расставленным циркулем, неподвижная нога которого стоит не в центре
мозга, а далеко за его пределами. Визуально они продольно-выгнутые. Главная
отличительная особенность их заключается в том, что они не продолжаются на другом
полушарии. Это у дельфинов. В отличие от них радиальные (поперечные) борозды
имеют тенденцию к продолжению, к переходу на другое полушарие. Это у человека.
Боюсь, что Тен вновь пытается выдавать
желаемое за действительное. Правильнее всего было бы узнать, что думает именно
Кочеткова. Однако её слов Тен не приводит, а жаль.
И даже если борозда визуально продолжается на другом полушарии, следует помнить,
что полушария мозга разделены продольной щелью, а медиальная поверхность полушарий
– гладкая.
Стр. 250:
Далее – о главном. Радиальные борозды свидетельствуют об интерактивности полушарий.
Это свидетельство нацеленности полушарий на совместную работу, ибо функция борозд
– вычленение популяций клеток мозга.
Функция борозд и извилин – увеличение поверхности мозга. А интерактивность полушарий определяется комиссурами, в том числе мозолистым телом.
Стр. 250:
Особенность, на которую указала В.И. Кочеткова, касается только конечного мозга,
неокортекса. Другой «недостаток» мозга дельфинов – отсутствие глубоких борозд,
просекающих оба полушария, и недоразвитость мозолистого тела – касается всего
мозга. При этом все три существенных различия сводимы к одному глобальному,
а именно: полушария мозга дельфина не являются интерактивными в такой степени,
как полушария мозга человека. Они гораздо менее зависимы друг от друга. Это
как бы два автономных мозга в одной черепной камере. Потрясающий
факт.
«Глубина борозды определяется, как уже
было сказано, площадью неокортекса, которая определяется по границе 4-го и 5-го
слоёв, а также толщиной неокортекса в данном месте. Мозолистое тело включает
проводящие пути с левого в правое полушарие и наоборот. У человека оно большое,
потому что одновременно функционирует и левое и правое полушарие, они оба функционируют
днём и перестают активно функционировать во время сна. Поэтому у человека мозолистое
тело очень большое, так как идет большая работа сложных функций головного мозга.
А у дельфина недоразвитость мозолистого тела вызвана раздельной работой левого
и правого полушария по времени, что не требует их постоянной совместной деятельности,
у них мозолистое тело нужно для переключения сознания с одного полушария на
другое» (комментарий Юрия Лучина).
У человека тоже нет борозд, «просекающих» оба полушария. А «недоразвитость»
мозолистого тела у дельфина можно объяснить скорее «переразвитостью» самих полушарий
мозга, из-за чего в соотношении площадь мозолистого тела (кв. мм)/вес мозга
(г) увеличивается знаменатель. В абсолютных значениях площадь сечения мозолистого
тела у них не так уж и мала – в вышеупомянутой (см. обсуждение стр. 249) статье
приводятся значения 300+, 400+ и 500+ квадратных миллиметров. Вполне возможно,
увеличение размеров мозолистого тела у человека связано с огромной ролью стереоскопического
зрения, и обмен сигналами между полушариями помогает строить трёхмерную картину
мира, что важно для примата – сам образ жизни настоятельно рекомендует уметь
оценивать расстояния, иначе однажды можно не допрыгнуть до ветвей соседнего
дерева, но зато быстро долететь до куда более далёкой земли.
Обратим внимание на оговорку «как бы», подразумевающую некую условность. Далее
она куда-то пропадает, и «как бы» плавно превращается в «так и есть на самом
деле».
Стр. 250:
...Одна загадка, связанная с дельфинами, волновала ученых много десятилетий.
Дело в том, что дельфины с момента рождения и до смерти ведут активный образ
жизни. Это уникальное явление в мире животных, где спят даже рыбы. Известно,
что мозг не может обходиться без отдыха. Чем более развит мозг, тем больше времени
ему требуется для восстановления работоспособности. Человек проводит во сне
треть своей жизни. Феномен дельфинов, очень мозговитых животных, которые при
этом «никогда не спят», пытались раскрыть многие.
В 1978 г. на базе Утришского дельфинария в Геленджике было выполнено исследование,
которое было внесено в реестр открытий под №28. Это означает, что в СССР к 1978
г. всего было сделано 28 открытий подобного масштаба. Это поразительное открытие
называлось следующим образом: «Явление межполушарной асимметрии состояний мозга
во время сна у дельфинов». Авторами его являются Л. М. Мухаметов и А. Л. Супин.
Мне захотелось проверить сказанное Теном,
и наиболее наглядным, хотя и не самым полным источником оказалась статья в «Википедии»
(ссылка).
Под № 28 там указано нечто иное:
«Явление адсорбционного понижения прочности металлов под действием металлических
расплавов. Е. Д. Щукин, Ю. В. Горюнов, Н. В. Перцов, Л. А. Кочанова, Л. С. Брюханова.
№ 28 с приоритетом от 26 января 1956 г» (ссылка).
От 1978 года там упоминается также нечто иное:
«Свойство нервных центров мозга человека и позвоночных животных содержать детерминированные
количества нейронов. С. Н. Оленев, В. Ю. Урбах, С. М. Блинков, В. А. Лившиц.
№ 399 с приоритетом от 19 сентября 1978 г».
Господин Тен сумел наврать даже в таком простом вопросе, где требуется просто
воспроизвести уже имеющуюся информацию. Есть ли упомянутое им открытие в Государственном
реестре открытий СССР? Есть. Вот оно (ссылка):
«Научное открытие "Явление однополушарного медленноволнового сна в мозге
позвоночного животного (дельфина)".
Формула открытия: "Установлено неизвестное ранее явление однополушарного
медленноволнового сна в мозге позвоночного животного (дельфина), заключающееся
в том, что медленноволновый сон возникает в одном (попеременно в правом и левом)
полушарии мозга, в то время как другое полушарие находится в состоянии активации,
что обусловлено односторонними влияниями регулирующих систем мозга на функциональные
состояния его полушарий".
Авторы: Л. М. Мухаметов, А. Я. Супин.
Номер и дата приоритета: № 266 от 5 марта 1974 г.»
Да уж, ни с номером не угадал, ни с годом. И достижений в науке в СССР на тот
момент гораздо больше было. За державу обидно, знаете ли.
Стр. 251:
Это открытие до сих пор не оценено по достоинству, ибо то, что оно дает для
понимания происхождения сознания человека, тянет на Нобелевскую премию. Искренно
надеюсь, что мой давний знакомый Лев Михайлович Мухаметов ее получит.
Я, в свою очередь, искренне надеюсь, что этот достойный учёный не станет притягивать к своему открытию за уши всякого рода наукообразные умствования, уподобляясь автору анализируемой мною книги.
Стр. 251:
Л. Мухаметов и А. Супин выяснили, что дельфины спят, причем, соответственно
уровню развития своего мозга, подолгу. При этом они не теряют активности, потому
что спят одним полушарием. Субъектом отдыха является не весь мозг, а только
одно из полушарий и никогда оба вместе, иначе дельфин погибнет. Интерактивность
полушарий заключается в том, что одно инактивирует другое, желая поспать. Один
глаз у дельфинов открыт всегда (Мухаметов, Супин, 1978).
Вот почему в каждом полушарии мозга дельфина присутствуют ретикулоподобные клетки,
которые у человека сконцентрированы в ретикулярной формации, расположенной между
полушариями мозга. Эти клетки – «часовые» и «фильтровщики информации». Они выполняют
задачу своевременной активации заторможенного полушария, как быстродействующие
дрожжи. Они в буквальном смысле слова «расталкивают» спящие нейроны. Наличие
ретикулоподобных клеток в коре мозга дельфинов не является признаком «неразвитости»,
это очень разумное приспособление, позволяющее просыпаться мгновенно.
Ай, да господин Тен! Ай, да… [здесь включился внутренний автоцензор]! Снова путает элементы неокортекса с ретикулярной формацией – два образования, выполняющие в мозге совершенно различные функции и появляющиеся в филогенезе в совершенно различное время. Но если есть похожие слова – «ретикулярная» и «ретикулоподобные» – между ними смело ставится знак равенства. Логика – как школьная линейка: прямая и простая. Только короткая.
Стр. 251:
«Два мозга» дельфинов поочередно находятся на круглосуточной вахте. Один будит
другого, какое-то время они обмениваются информацией, потом второй отправляется
отдыхать.
Так это что же получается – дельфин круглосуточно спит, даже днём?
Стр. 251:
Вот почему два полушария дельфинов не имеют поперечных борозд, соединяющих их
в одно функциональное целое. Вот почему у них не развита межполушарная комиссура.
Вот почему поверхностные борозды неокортекса у них продольные, а «перетекание»
борозд с одного полушария на другое только намечено, но не осуществлено, как
у человека с его поперечными бороздами.
Я сейчас вам напомню ещё одну вещь, господин Тен. В анализируемой сейчас книге на стр. 198 вы сами же написали, что мозолистое тело отсутствует у сумчатых и клоачных млекопитающих. А теперь я предлагаю подумать. Если неразвитость мозолистого тела у китообразных является следствием именно особенностей функционирования мозга, то это означает, что у сумчатых мы должны ожидать даже в большей степени наблюдения хотя бы части тех явлений, которые описываются для мозга дельфина. Или же мы должны признать, что степень развития мозолистого тела в различных группах млекопитающих определяется различными (не одинаковыми в каждом случае) факторами и не связана напрямую с описываемой особенностью мозга дельфина.
Стр. 251-252:
Межполушарная асимметрия, безусловно, является важнейшим гидростатистическим
приспособлением предков дельфинов. Она появилась у археоцет, когда те перешли
на исключительно морской образ жизни. В свою очередь, с ней связан ускоренный
рост всего мозга (о чем, основываясь на палеонтологических данных, мы говорили
выше), ибо та работа, которую у наземных предков дельфина выполнял весь мозг,
теперь оказалась возложена на одно полушарие. Рост мозга вызвал, в свою очередь,
рост мозговой капсулы, который отмечается палеонтологами.
Снова «гидростатистический»… Откуда взялся
этот «словесный урод», и что хотел этим сказать господин Тен? Да, я в очередной
раз придрался к словам – раз Тен позволяет себе это делать, значит, он не против
того же самого в отношении его собственного труда.
Можно предположить, что имеется в виду слово «гидростатический». Однако гидростатика
– это «раздел гидромеханики, в котором изучаются равновесие жидкости и воздействие
покоящейся жидкости на погруженные в неё тела» (определение
из «Большой Советской Энциклопедии»). И как влияет внешне ничем не проявляющаяся
функциональная асимметрия полушарий мозга на поведение тушки китообразного в
жидкости – ума не приложу. Поэтому применение данного эпитета в любой форме
к строению мозга, на мой взгляд, попахивает неким абсурдом.
Думаем дальше, и у нас становится, как говорила Алиса в Стране Чудес, «всё чудесатее
и чудесатее». Значит, полностью водный образ жизни начали вести археоцеты, и
это Тен признаёт. Хорошо, запомним это и посмотрим, что он скажет дальше.
Возникает ещё один важный вопрос: какие данные подтверждают «межполушарную асимметрию»
у археоцетов? У господина Тена есть электроэнцефалограмма мозга живого археоцета?
Да за одну только её расшифровку он уже получил бы звание не ниже доктора наук,
а сама по себе она была бы ценнейшим источником знаний об эволюции мозга китообразных.
В общем, мне действительно интересно, на каком основании Тен что-то утверждает
о работе мозга давно вымерших китов. Хочется припасть всеми фибрами души к этим
первоисточникам, что называется. Если что, черепа археоцетов были симметричными.
А по большому счёту межполушарная асимметрия довольно широко распространена
в природе: даже у кенгуру есть «рабочая» лапа, которой зверь пользуется чаще.
И уж конечно она присутствует у обезьян, даже у не относящихся к человекообразным.
Стр. 252:
Тогда и произошло вторичное появление широких лобных долей, которые на первом
этапе перехода с земного на водный образ жизни редуцировались во имя лучшей
гидродинамики. Здесь мы видим, как эволюция жертвует менее значимым во имя более
значимого. Рост мозга, вызванный асимметрией полушарий, оказался важнее фактора
гидродинамики; дельфины «отказались» от рыбьей морды, которую эволюция подарила
им при переходе к водному образу жизни, и таранят воду лбом. У некоторых появилось
такое гидродинамическое приспособление, как рострум, у некоторых (у гринд) –
нет. Независимо от наличия или отсутствия рострума все дельфины – лобастые животные,
что не слишком удобно при скоростном движении, учитывая высокую степень сопротивления
воды. Потеря скорости – плата за интеллект.
Прежде, чем так пафосно говорить, можно
было бы взглянуть хотя бы на схему внутреннего строения головы дельфина, чтобы
понять, что уплощённый острорылый череп дельфина как раз способствовал бы обтекаемости
очертаний его тела. Крутой лоб дельфина образован не выпуклым черепом с большим
мозгом, а приспособлениями для эхолокации – жировой «линзой» для звука («дыней»)
и носовыми ходами с воздушными мешками, образующими звуковоспроизводящий аппарат.
Далее, обсуждая страницу 220 творения господина Тена, я уже отмечал, что Тен
несколько путает лобные кости и лобные доли мозга: у него
«лобные доли оказались задвинуты и сохранились только в виде небольших
косточек». Поэтому в данном случае у меня отсутствует
уверенность в осознании Теном правильности того, о чём он пишет.
Асимметрия черепа дельфинов и других зубатых китов отражает эволюцию не столько
мозга, сколько эхолокационного аппарата: это следствие асимметрии мягких тканей
головы животного, наблюдаемой у современных представителей зубатых китов. Соответственно,
рост мозга животных был явно связан с прогрессивной эволюцией эхолокационного
аппарата.
Стр. 252:
Подтверждением высказанного является недавно выявленный факт: мозг дельфинов
рос скачками.
«Исследовательской группе из Калифорнийского университета под руководством Лори
Марино удалось установить, что интеллектуальная эволюция дельфинов началась
приблизительно 35 млн лет назад. Судя по размерам черепов древних дельфинов,
именно в это время у них резко увеличился объем головного мозга. Следующий скачок
произошел примерно 15 млн лет назад. В это время мозг дельфина еще раз увеличился
в размерах и по своим параметрам стал близок человеческому» (https://books.google.ru/books).
35 млн. лет назад – это ранний олигоцен.
Сложно время для истории Земли, надо сказать. Тетис закрылся, климат стал ухудшаться
– начало холодать, на суше изменилось движение воздушных масс, а в морях – течений.
Исчезали прежние места обитания, появлялись новые. Кстати, на суше глуповатые
эоценовые таксоны тоже начали стремительно исчезать, ровно по тем же самым причинам.
Соответственно, обострение борьбы за существование в условиях ухудшающегося
климата и изменяющихся мест обитания потребовало не только количественных, но
и качественных изменений.
Мне не удалось найти первоисточник этой новости на английском языке, а пересказу
этой новости в Рунете я не доверяю, поскольку при попытке отыскать первоисточник
постоянно наталкивался на сайты и книги, не имеющие отношения к науке. Есть
и ещё одна причина недоверия: дельфины как самостоятельное семейство появились
на Земле в позднем миоцене, 11 миллионов лет назад (иногда указывается время
15 млн лет назад). Впрочем, если под «дельфинами» господин Тен и неведомые пересказчики
научных новостей имеют в виду не тех дельфинов, которые Delphinidae, а тех,
которые кто-то там ещё, то здесь я умываю руки, потому что такие вещи логике
и научным фактам не подвластны.
Стр. 252-253:
Выше мы писали о том, что предки дельфинов долго были водно-земными, в частности,
для родов выходили на сушу. Возможность полностью уйти в море у них появилась
вместе с формированием функциональной асимметрии полушарий, что вызвало взрывной
рост мозга, потому что произошло, по сути дела, удвоение мозга. Теперь мы знаем,
когда это произошло: 15 млн лет назад. Но они миллионы лет возвращались на побережья
для родов в защищенных бухтах. Примерно в то время, около 10 млн лет назад,
началась эволюция их неотенических отпрысков в человека.
Нет, киты ушли в море гораздо раньше,
чем начался тот рост мозга, о котором упоминает господин Тен: это произошло
ещё в эоцене, когда появились первые постоянноводные китообразные – археоцеты
семейств протоцетид и базилозаврид. Собственно, выше, буквально на этой же странице,
Тен признаёт это. Значит, эти два события – переход к жизни в воде и увеличение
размеров мозга – не связаны друг с другом. Обратите внимание, я не тенденциозно
подбираю факты, а умница господин Тен сам выложил их на одной странице.
Археоцеты вымерли 23 миллиона лет назад, и после них в морях плавали киты других,
современных подотрядов (усатые и зубатые), представленные архаичными семействами
и родами. Соответственно, 15 млн лет назад в океанах плавали киты вполне современной
анатомии, полностью приспособленные к водному образу жизни.
Далее, откуда Тен взял цифру «около 10 млн лет назад»? Мессинский кризис начался
менее 6 млн лет назад. Сам Тен же утверждает, что его «новый антропогенез» начался
именно в связи с Мессинским кризисом, а он не совпадает с указанной датой. Странная
получается теория у Тена – куда ни ткни, вылезают какие-то нестыковки по датам,
отсутствие фактического материала и прочие неувязки.
Стр. 253:
Нобелевский лауреат Р. Сперри писал о мозге современного человека: «Каждое полушарие
имеет как бы отдельное собственное мышление» (Сперри; цит. по: Билич, Назарова,
2000, с. 111).
Контекст – великая сила! В этом я убеждаюсь
раз за разом, читая труд Тена. Потому что перед цитатой из книги Билич Г. Л.
и Назаровой Л. В. «Основы валеологии» (СПб, 2000), откуда они взяты, говорится
следующее:
«Оба полушария ответственны за сознание и самосознание человека, его социальные
функции».
Далее следует процитированный Теном кусочек, а после него авторы продолжают:
«При анатомическом изучении мозга были выявлены межполушарные различия. В то
же время следует подчеркнуть, что оба полушария здорового мозга работают вместе,
образуя единый мозг».
В контексте первоисточника фраза смотрится несколько иначе, чем хотел бы этого
Тен, не правда ли? Авторы первоисточника пишут о единстве мозга, а Тен теми
же словами иллюстрирует «разделённость» мозга.
Стр. 253:
Это получилось благодаря тому, что мозг человека не вырос мало-помалу из мозгов
обезьян, он достался нам от наших подлинных предков – акродельфидов, этой неотенической
формы древних дельфинид, уже избыточным и уже разделенным.
Соответственно, это высказывание Тена
не подкреплено ничем. Обратим внимание: здесь только авторский текст. Я вклиниваюсь
в него исключительно для удобства комментариев, но в данном случае не выбросил
ничего, чтобы не быть обвинённым в вырывании фраз из контекста. То есть, теперь
у нас акродельфиды стали неотенической формой дельфинид?
Если сравнивать время существования акродельфид и дельфинид, наблюдается такая
картина:
Акродельфиды: с 23,03 до 5,333 млн лет назад (ссылка);
Дельфиниды: с 20.44 млн лет назад до наших дней (ссылка).
Удивительная штука получается: в течение примерно 3 миллионов лет акродельфиды
представляли собой «неотеническую форму» животных, которые даже ещё не появились
в процессе эволюции как отдельное семейство! Что это: парадокс или невежество?
Итак, ссылок на уникальные работы по изучению работы мозга ископаемых китообразных
Тен не приводит, ископаемых переходных форм между «акродельфидами» и людьми
не показывает, их названий не даёт. Работ, описывающих неотению у «акродельфид»,
Тен также не указывает. Соответственно, сказанное им – это только слова, а написанное
– терзание клавиатуры компьютера, не более того.
Стр. 253:
Именно поэтому он избыточен даже для современных интеллектуалов. Именно поэтому
в мозге человека могут помещаться не одна, а две и более психик.
Избыточен ли мозг? Сложно сказать, откуда
Тен взял эту мысль. Можно лишь предположить, что в основе этого утверждения
лежит «городская легенда» (точнее – околонаучный миф) о том, что человек использует
свой мозг всего лишь на 10% его возможностей. Учёные давно разобрали этот миф
и показали, что подобное утверждение бездоказательно. В качестве одной из работ,
разбирающей этот миф, можно порекомендовать книгу “Understanding the Brain:
The Birth of a Learning Science”, доступную в Интернете. На страницах 113-114
подробно разбирается этот миф, и это можно совершенно свободно прочитать в Интернете,
вот
здесь (на английском языке). Подробно разобрав причины возникновения мифа,
авторы пишут в конце главы следующее:
«Миф также неправдоподобен по физиологическим причинам. Эволюция не позволяет
транжирства, а мозг, как и другие органы, но возможно, что и больше, чем любой
из них, подвергся формированию путём естественного отбора. Он представляет собой
всего лишь 2% от общего веса человеческого тела, но потребляет до 20% доступной
энергии. При таких высоких затратах энергии эволюция не допустила бы развития
органа, 90% которого бесполезно».
Думаю, авторы книги сказали достаточно, чтобы выкладку Тена можно было отправить
в утиль.
Помещается ли в мозге больше «одной психики»? Вопрос, конечно, занятный. Однако
господин Тен не приводит никаких первоисточников, которые позволяют проверить
это утверждение – как всегда, на самом интересном месте.
Стр. 253:
Как происходило становление мозга человека из разделенного мозга наших водно-земных
предков – это тема отдельной книги – о возникновении сознания.
Скажу честно, мне уже заранее становится страшно за людей соответствующих специальностей, если в этой книге аргументация будет такого же качества, как в «Человеке изначальном…». Это моё личное оценочное мнение.
Если у читателей этого обзора есть дополнения и исправления, большая просьба поделиться ими. Адрес почты автора обзора - на главной странице сайта.
Глава 1
Глава 2
Глава 3
Глава 4
Глава 5
Глава 6
Глава 7
| Главная |