Марсело Санчес
Эмбрионы в глубинах времени

Эмбрионы в глубинах времени
Биология развития
в летописи окаменелостей
Марсело Санчес
Embryos in Deep Time
The Rock Record
of Biological Development
Marcelo Sánchez

Издательство Калифорнийского Университета
Беркли Лос-Анджелес Лондон
Эмбрионы в глубинах времени
Пустая страница
Эмбрионы в глубинах времени
Биология развития
в летописи окаменелостей
Марсело Санчес
Embryos in Deep Time
The Rock Record
of Biological Development
Marcelo Sánchez
Издательство Калифорнийского Университета
Беркли Лос-Анджелес Лондон
Издательство Калифорнийского Университета, одно из
самых выдающихся университетских издательств в Соединённых Штатах, повышает
качество жизни во всём мире, пропагандируя образованность в гуманитарных,
общественных и естественных науках. Его деятельность поддерживают Фонд
Издательства Калифорнийского Университета и безвозмездные пожертвования
частных лиц и организаций.
Дополнительную информацию можно получить на сайте www.ucpress.edu.
Издательство Калифорнийского Университета
Беркли и Лос-Анджелес, Калифорния
University of California Press, Ltd.
Лондон, Англия
© 2012 The Regents of the University of California
Данные о публикации в каталоге Библиотека Конгресса
Санчес, Марсело Р.
Эмбрионы из глубин времени: биология развития в летописи окаменелостей/Marcelo
R. Sánchez.
p. cm.
Содержит библиографические ссылки и указатель.
ISBN 978-0-520-27193-7 (обл.: щелоч. бумага)
1. Палеобиология. 2. Эмбрионы. 3. Генетика развития.
I. Название.
QE719.8. S26 2012
560 - dc23
2011034435
Произведено в Соединённых Штатах Америки
21 20 19 18 17 16 15 14 13 12
10 9 8 7 6 5 4 3 2 1
Бумага, используемая в этом издании, соответствует минимальным требованиям
ansi/niso z39.48 - 1992 (r 1997) (Стойкость бумаги). |
Перевод П. Волкова
2013 г.
Содержание
Благодарности
vii
Пролог
xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания
197
Библиография
213
Предметный указатель
243
vii
Благодарности
Я хотел бы поблагодарить Цюрихский Университет и его Научный Факультет, а
также Палеонтологический Институт за создание вдохновляющей, побуждающей к
работе и благоприятной обстановки, в которой я смог написать эту книгу.
Я благодарю нынешних и бывших сотрудников моей лаборатории и близких коллег
за всё, что я узнал от них: Ингмара Вернебурга, Лауру Уилсон, Торстена Шейера,
Кристиана Митгатча, Массимо Дельфино, Яси Хьюджи, Кристиана Колба, Даи Койябу,
Джеймса Нинана, Фреди Карлини, Сандрин Ладевез, Коринн Уиммер, Яна Прочела,
Питера Менке, Мадлен Гейгер, Катю Полачовски, Фиону Стрэл, Патрисию Мейер,
Веру Вайсбекер, Фернандо Галлиари, Лайзу Рэйджер, Томаса Шмельцля и Анджали
Госвами. Я также благодарю некоторых коллег из Цюриха и из-за границы за обсуждение
идей и за любезно предоставленную информацию: Леннарта Олссона, Шигеру Куратани,
Майка Ричардсона, Хироши Нагашиму, Хьюго Бюше, Тома Кемпа, Джона Спайса, Норберто
Джанини, Йоханнеса Мюллера, Райнера Шоха, Шиге Кураку, Андреаса Вагнера, Инесс
Хоровиц, Орэнжел Агилеру, Лайонела Готье, Рено Лебрана,
viii
Анджали Госвами, Норму МакЛеод, Рика Мэддена, Мартина Сандера, Кэтлин Смит,
Пола Тейлора, Рафаэля Хименеса, Нико Гудмана, Клода Монне, Майкла Хаутманна,
Кристиана Клюга, Хайнца Феррера, Ричарда Хоффманна, Кеннета де Бэтса, Томаса
Мартина, Александра Нютцеля, Клаудию Хоффманн, Фреди Карлини, Карин Ниффелер
и Северина Урди. Я благодарю всех, кто внёс свой вклад в издание «Seminars
in Cell and Developmental Biology», которое я редактировал в 2010 году,
за помощь, вдохновившую на написание этой книги: Ричарда Клотье, Зерину Джохансон,
Мойю Смит и её коллег, Луизу Хэмфри, Кристофа Золликофера, Марсию Понс де
Леон, Мартина Сандера, Николь Клейн, Надю Фробиш, Флориана Витцманна, Райнера
Шоха, Дженнифер Олори, Торстена Шейера и Массимо Дельфино.
Роб Ашер в Кембридже, Англия, Лаура Уилсон и Торстен Шейер в Цюрихе, а также
Анна-Кристина Хоннен в Берлине любезно и критически перечитали ранние наброски
и сделали очень полезные замечания. Три анонимных рецензента, а особенно Мишель
Лорин, были очень щедры на критику и предложения, которые улучшили ясность
этой работы и помо¬гли мне избежать ошибок. Торстен Шейер, Кристиан Митгатч,
Джеймс Нинан, Лайза Рэйджер, Кевин де-Карли (Цюрих) и Найджел Хьюз (Риверсайд)
любезно просмотрели части текста. Торстен Шейер и Яси Хьюджи дали весьма нужные
советы, касающиеся главы о палеогистологии, а Кристиан Митгатч – по онтогенетическим
и историческим вопросам. Чже-Си Луо поделился информацией, касающейся «ранних
млекопитающих» со своей обычной
отзывчивостью и добрым юмором. Северин Урди (Цюрих) любезно представил развёрнутые
комментарии и догадки, касающиеся букашек и ракушек. Любые несоответствия
или огрехи, которые остались здесь – полностью на моей ответственности.
За помощь с графикой и вопросами форматирования я благодарю Морану Михальевич,
Кевина Де-Карли, Рози Ротт, Фиону Стрель и Торстена Шейера. Мадлен Гейгер,
Катя Полачовски и
ix
Клаудия Джоел прекрасно выполнили некоторые из рисунков. Джон А. Лонг (Лос-Анджелес),
Мэтт Фридман (Оксфорд), Ясмина Хьюджи (Цюрих), Йоахим Хауг (Нью-Хейвен), Панчо
Гоин (Ла-Плата), Чже-Си Луо (Питтсбург), Торстен Шейер (Цюрих), Кристиан Клюг
(Цюрих), Кристоф Золликофер и Марсия Понс де Леон (Цюрих), Луис Костер (Базель)
и Райнер Шох (Штутгарт) очень великодушно дали право воспользоваться принадлежащими
им изображениями.
Я благодарю за очень нужную и прекрасно оказанную помощь Хейке Готцманна –
за решение административных вопросов, и Хейни Уолтер – за помощь в вопросах,
связанных с информационными технологиями. Я также благодарен директору моего
института Хьюго Бюше за его поддержку.
Линн Мейнхардт, Кейт Уорн и Чак Крамли из издательства Калифорнийского Университета
быстро отвечали на мои многочисленные вопросы, возникавшие в процессе редактирования,
и любезно дали мне много рекомендаций разного рода, а Шейла Берг сделала в
отношении рукописи очень полезные и ёмкие замечания, касающиеся редактирования
и стилистики.
Вольфганг Майер в Тюбингене поддерживал мой интерес к онтогенезу на протяжении
многих лет и был очень ценным наставником.
Петер Хольцварт в Цюрихе, Роб Ашер в Кембридже, Энн-Кристин Хоннен в Киле
и Берлине, и моя мать Глория Виллагра в Пуэрто-ла-Крусе/Буэнос-Айресе оказали
мне моральную поддержку.
В последние несколько лет я пользовался поддержкой главным образом Швейцарского
Исследовательского Совета (SNF) и Цюрихского Университета.
x
Пустая страница
xi
Пролог
Разнообразие жизни обычно представляется в виде эволюционных древ: ветвящееся
изображение завершается рисунками животных и растений.
Это хорошо, поскольку очертания древа отображают общую историю, которую разделяют
друг с другом живые организмы, в том числе и мы сами. Но у этого способа отображения
информации есть одно ограничение. Изображённые живые организмы – это статичные
объекты, обычно во взрослом состоянии с узнаваемыми особенностями своего вида.
В реальной действительности организмы изменяются на протяжении своей жизни.
Если мы захотим изобразить биологию биоразнообразия, мы могли бы показать
для каждого организма фильм, сделанный в технике замедленной съёмки и демонстрирующий
различные стадии истории жизни – для многоклеточного организма от оплодотворения
до смерти. Варианты и количество изменений, которые происходят на протяжении
жизни индивида, настолько многочисленны и сложны, что каждой стадии этого
процесса посвящены различные научные дисциплины. Первые шаги деления клеток
и формирования эмбриона ассоциируются с эмбриологией. Многие исследования
в этой области относятся, например, к гаструляции – стадии, на которой образуются
зародышевые листки и устанавливается план строения тела взрослого организма.
Специалисты по биологии развития склонны изучать момент возникновения или
проис-
xii
хождение тканей и органов. Специалисты в области сравнительной анатомии и
зоологи обычно интересуются изменениями, возникающими после рождения, которые
некоторые биологи любят называть словом «рост», противопоставляя его развитию.
Для всех этих дисциплин молекулярная биология предложила методы и концепции,
позволяющие поставить совершенно новый набор вопросов, касающихся механистических
основ эволюции истории жизни, а также заново ответить на старые вопросы, поставленные
морфологами.
Индивидуальное развитие – это благодарный предмет для исследований в биологии.
А как вам тот факт, что большинство видов, представленных на древе жизни,
уже вымерло? Некоторые могут подумать что палеонтология нужна биологии развития,
как рыбе велосипед. Они также могут утверждать, что летопись окаменелостей
полностью умалчивает о многих аспектах эволюции индивидуального развития и
генетики.
Я не думаю, что дела обстоят именно так, и я написал эту книгу, чтобы объяснить,
почему. Конечно, палеонтология всё ещё в значительной степени умалчивает о
таких ключевых вопросах, как гаструляция. Потому и важно определить границы,
которых может достичь палеонтология. В этом контексте мне вспоминается вклад
известного специалиста по анатомии рыб в спорах 1980-х годов: роль окаменелостей
против молекул при реконструкции древа жизни. Колин Паттерсон (1933 – 98),
важная фигура в исследованиях окаменелостей, десятилетиями работавший в палеонтологическом
отделе Музея Естественной Истории в Лондоне, настоятельно выступал в пользу
главенства информации о ныне живущих видах над ископаемыми остатками при исследовании
эволюционных отношений. Большинство коллег-палеонтологов жёстко критиковало
его. Несколько лет спустя его идеи стали оцениваться намного выше, и в 1996
он был предоставлен к медали Ромера-Симпсона – это высшая степень одобрения
Общества Палеонтологии позвоночных. Роль палеонтологии в реконструкции древа
жизни в значительной степени признана большинством биологов, несмотря на ограничения,
возникающие при работе с фрагментарными данными, что резко контрастирует с
xiii
большим массивом информации (например, геномной), который известен сейчас
для большего и большего количества ныне живущих видов.
Интеграция информации из областей палеонтологии и эмбриологии имеет давнюю
историю. В викторианскую эпоху, например, Томас Гексли сделал важный вклад
в обеих областях знания: например, он был первым, кто предположил, что птицы
родственны динозаврам, и он же открыл главные аспекты ранних стадий жизни
книдарий, группы, к которой принадлежат кораллы и медузы. Фактически, многие
исследователи продолжают вносить большой вклад в эти очень различающиеся области
знания. Например, Фил Донахью из Бристоля изучает конодонтов, базальную группу
позвоночных животных, которая вымерла примерно 200 миллионов лет назад, но
у него также есть исследовательская программа, касающаяся микро-РНК и её роли
в морфологическом разнообразии. Взятые вместе, его исследования в области
палеонтологии, биологии развития и молекулярной биологии ведут к лучшему пониманию
истории жизни.
В этой книге я исследую то, что мы можем узнать о развитии организмов непосредственно
из летописи окаменелостей. Ископаемые остатки – это не просто статичные объекты,
части более не существующих животных. Глядя проницательным взглядом, обладая
соответствующей информацией и оставаясь в концептуальных рамках, мы можем
многое узнать о размножении и развитии вымерших животных.
Может показаться, что «моментальный снимок» какой-то стадии развития, который
даёт окаменелость, окружён таким количеством неизвестностей, что интерпретация
зачастую неполной анатомии остаётся, в лучшем случае, предположительной. Но
существуют методы изучения неполных окаменелостей, и выводы, сделанные большинством
палеонтологов, основаны на материальных свидетельствах.
Эта книга предназначена для людей, обладающих общим знанием биологии и интересом
к ископаемым остаткам и эволюции. Примечания и ссылки в конце книги могут
помочь разъяснить и уточнить разнообразные вопросы, к которым я здесь обращаюсь.
Я прошу, чтобы мне простили
xiv
несколько случайный характер упоминаемых ссылок, поскольку некоторые из затронутых
здесь тем очень обширны, и им сопутствует долгая история исследований и плодотворных
размышлений.
Многие примеры взяты из моей собственной работы и из работ моих самых близких
коллег, поскольку эти примеры я знаю лучше всего. В конце концов, каждая книга
представляет собой личный взгляд на проблему, и эта вовсе не является исключением.
1
Глава первая
Окаменелости, онтогенез и филогенез
| Человеческая история – это краткая
вспышка в космосе, и первый урок для него – скромность.
Уилл Дюран и Ариэль Дюран
«The Lessons of History»
|
Я помню, как на меня, тогда ещё ребёнка, произвело очень сильное впечатление
утверждение, ошибочно приписанное Томасу Гексли, где говорится, что, если
посадить обезьян перед пишущими машинками, а они будут случайно нажимать клавиши,
то при наличии достаточного времени они наверняка напечатали бы полную «Британскую
энциклопедию». У меня есть сокращённый вариант «Энциклопедии» на испанском
– пятнадцать толстых томов; поэтому я представляю себе, о каком массиве текста
идёт речь. Впервые я прочитал это утверждение в креационистском буклете, где
было указано на абсурдность этого утверждения.1
Но для меня оно имело смысл: маловероятно, но всё
же, при наличии бесконечного времени, это могло бы случиться. При написании
этой книги я решил немного исследовать этот вопрос, и выяснил, что этот мысленный
эксперимент с обезьянами и пишущими машинками всерьёз рассматривается с философской
и статистической точки зрения и используется в различных популярных работах.
Фактически, это один из
2
наиболее известных мысленных экспериментов, из датируемого 1913 годом эссе
французского математика Эмиля Бореля. С тех пор он стал популярной иллюстрацией
в теории вероятностей. Вполне очевидно, что вероятность того, что обезьяны
напечатают «Британскую энциклопедию» или произведения Шекспира, бесконечно
мала. Насколько это уместно при обсуждении эволюции и индивидуального развития?2
Здесь есть две точки обсуждения: временной отрезок и вероятность
эволюционного изменения, возникающего в течение этого времени.
Геологическое время не бесконечно, но оно долгое, очень долгое, или глубокое
– хорошее слово для описания, предполагающее, что в глубинах пород скрыто
то, благодаря чему мы можем многое узнать об этом отдалённом прошлом. Понятие
«глубокое время» возникло вместе с популярной работой по геологии американского
автора Джона МакФи под названием «Basin and Range» В ней он изложил, как геологи
интеллектуально и эмоционально развивают у себя ощущение необъятности времени.3
Осознание наряду с этим необъятности биологического разнообразия
ископаемых форм открывает исключительные перспективы, и это может быть наиболее
фундаментальным вкладом в человеческое понимание вселенной в сравнении с тем,
что могут дать геологи и палеонтологи.
Благодаря датированным ископаемым остаткам, размещённым на эволюционных древах,
и оценкам, основанным на молекулярных данных, мы знаем, что жизнь произошла,
по крайней мере, от 3,5 до 3,2 миллиардов лет назад, и что возраст многоклеточной
жизни составляет, как минимум, 700 миллионов лет.4
В двадцатом веке одним из главных достижений в геохимии была
разработка нескольких методов датирования горных пород, основанных на изотопах
различных химических элементов, что привело к надёжному установлению абсолютного
исчисления времени протяжённой истории Земли и жизни. Считалось, что эволюции
биологического разнообразия требовались длительные периоды времени. Дарвину
было важно собрать информацию о древности Земли и жизни. Он был озабочен тем,
чтобы
3
для эволюции действительно сложных структур, вроде глаза, было «достаточно»
времени. Дальнейшее открытие мутаций и их «беспорядочности», на первый взгляд,
сделало оправданным волнение Дарвина.
Эволюция далека от случайного процесса, и обезьяны за пишущими машинками –
это не лучшая аналогия для эволюционных процессов. Одно из главных замечаний,
сделанных в недавно вышедших книгах по эволюции – указание на нехаотичность
и предсказуемость эволюции. Это не теоретическое заключение, а скорее нечто,
проявляющееся эмпирически в характере явлений, заметных в фенотипах и генотипах
ныне живущих видов, а также в ископаемых остатках живых организмов. Факт того,
что мутации в некоторых генах имеют высокую вероятность быть отобранными неоднократно
в независимых родословных линиях, сталкивающихся со схожими условиями окружающей
среды, получил название «параллельная генотипическая адаптация». Это делает
генетические траектории адаптивной эволюции до некоторой степени предсказуемыми,
что даёт ключ к реконструкции молекулярных процессов, которые с большой степенью
вероятности работали у вымерших видов, несмотря на случайный характер эволюции.
Индивидуальное развитие у вымерших видов также можно реконструировать благодаря
открытию принципов, которые являются общими для огромных групп видов и даже
включают использование одних и тех же генов, отвечающих за индивидуальное
развитие. И всё же разнообразие, как в смысле количества видов, так и в смысле
многообразия форм, настолько же обширно, насколько протяжённо время, в течение
которого оно эволюционировало.
Вымирание большей части жизни на Земле
Теория эволюции даёт рациональное объяснение богатому биологическому разнообразию,
которое окружает нас. С каждым днём всё больше людей по всему миру живёт в
городах, но даже те, кто редко сталкивается с природой, имеют представление,
по крайней мере, о части этого разнообразия благодаря телевидению или посещению
зоопарка. Уже описано
4
приблизительно 1,5 миллиона видов, и, по разным оценкам, планету населяет
от 10 до 100 миллионов видов. Но это огромное количество видов – лишь малая
часть от общего количества видов, которые существовали на планете; консервативные
оценки предполагают, что не менее 99 процентов от общего разнообразия жизни,
которая существовала на Земле, вымерло.5
Большая часть этого минувшего разнообразия в значительной
степени не документирована, несмотря на работу палеонтологов, поскольку ископаемых
остатков тех видов больше не существует, или же они ещё ждут своего открытия
и изучения.
Часть этого огромного разнообразия, которая знакома нам самым непосредственным
образом – это позвоночные, группа животных, обладающих позвоночным столбом,
к которой принадлежим и мы сами. Существует примерно 59000 ныне живущих видов
позвоночных, но было описано ещё больше видов*. Некоторые группы позвоночных
с палеонтологической точки зрения известны лучше, чем другие. Для динозавров,
например, известно приблизительно 550 описанных родов, но по оценкам специалистов,
должно было существовать примерно 1850 родов. Не все новые роды и виды ископаемых
организмов, которые были описаны, являются валидными, потому что палеонтологи
непрерывно проводят ревизии своих выводов, касающихся таксономии, и новые
исследования в области видового разнообразия и анатомии помогают уточнить
критерии, на которых основываются выводы. Обзор, сделанный в 2003 году, выявил,
что валидные на тот момент 4399 родов ископаемых млекопитающих составляли
80 процентов от общего количества когда-либо описанных родов. Это число составляло
67 процентов в 1945 году, как сообщал в своей классической статье американский
палеонтолог Джордж Гейлорд Симпсон. Свои названия получают всё больше и больше
валидных ископаемых родов млекопитающих и других позвоночных.
Случай с эволюцией нашего собственного рода Homo иллюстрирует противоречия
и уклоняющиеся мнения, касающиеся таксономии ископаемых форм. Однако по многим
вопросам имеет место согласие во мнениях. Например, большинство антропологов
подтверждает, что около двух миллионов лет назад по меньшей мере пять различных
видов людей населяли
* Очевидно, автор имеет в виду ископаемые виды позвоночных,
сравнивая их с современными. – прим. перев.
5
планету. Мы – последняя ветвь когда-то гораздо более ветвистого эволюционного
древа, существующего с момента отделения наших предков от шимпанзе примерно
семь миллионов лет назад.
Цифры, которые я привожу здесь, служат просто для иллюстрации того факта,
что, если мы хотим понять эволюцию биологического разнообразия, взгляд в прошлое
является основополагающим. Роль палеонтологии в описании исчезнувшего биологического
разнообразия очевидна, но, помимо простого описания, существует ещё много
способов, которыми палеонтология вносит свой вклад в эволюционную биологию.
Летопись окаменелостей и что
она рассказала нам об индивидуальном развитии
Новые области для исследования, возникшие в ходе подъёма молекулярной биологии
в последние десятилетия, вызвали целую волну вполне обоснованного энтузиазма.
Это также привело к постановке в определённых кругах вопроса об оправданности
дальнейшей работы в таких старых дисциплинах, как палеонтология. В контексте
обсуждения этого вопроса многие из моих коллег выделили ряд специфичных аспектов,
когда только обращение к палеонтологии способно помочь другим биологическим
дисциплинам и создать целостную программу исследований. Они таковы:
9
Чтобы проиллюстрировать важность ископаемых остатков, я упомянул о происхождении
перьев и волос. Они рассматриваются как «эволюционные новшества»: особенности,
которые очень отличаются от прежнего состояния и инновационные, и потому они
так или иначе вели к новой главе в эволюционной истории, или же коррелировали
с ней. Другой пример – эволюция рук и ног, поскольку она документирована для
самых ранних представителей группы, к которой принадлежат наземные позвоночные.
С чем биологи согласны, и на чём они делают упор вновь и вновь, так это с
тем, что для того, чтобы понять эти новшества, вы должны знать, как они развивались
в ходе индивидуальной истории организма, который обладает ими.
Мы знаем человеческую руку как оформленное и гибкое сочетание скелета, мускулатуры,
кожи, нервов, кровеносных сосудов и других тканей. Чтобы рассмотреть происхождение
руки, вы должны вернуться назад во времени к нашему эмбриональному периоду,
в течение которого формируются различные её компоненты. Какова их связь с
другими частями тела? Какие гены задействованы в их развитии? Как объединяются
мускулы, сухожилия и кости? Как утверждал эволюционный биолог Гюнтер Вагнер,
«объяснение эволюционных новшеств равнозначно идентификации изменений в ходе
индивидуального развития, которые делают новый признак возможным». Нам следует
предпринять ещё одно путешествие назад во времени, чтобы понять происхождение
руки, и это будет путешествие по геологическому времени, а не по времени жизни
индивида. Первые ископаемые позвоночные с мускулистыми руками и ногами жили
почти 400 миллионов лет назад. Изучая те ископаемые остатки, мы можем узнать
тот экологический контекст, в котором эволюционировали руки, и комбинацию
анатомических особенностей, которыми обладали те животные. Мы знаем что у
самых ранних позвоночных с руками и ногами было больше пяти пальцев на каждой
конечности, и что они были водными животными. Теперь представьте себе, что
мы смогли вернуться назад в геологическом времени, а затем вернуться назад
во времени индивидуального развития
10
ныне вымерших животных, чтобы понять, как возникли их руки. Это гипотетическое
путешествие воплощает объединение двух дисциплин, которые в первом приближении
могут показаться весьма различными: палеонтологии и биологии развития. Но
их отношения чрезвычайно близки. Мы можем вернуться назад во времени в истории
этих дисциплин и их отношений, и увидеть, насколько давно признаны параллели
между индивидуальными изменениями и трансформациями, которые документируют
ископаемые остатки.
Отношения между палеонтологией и
биологическими дисциплинами,
изучающими индивидуальное развитие
Целые дисциплины или идеи возникают или утверждаются в канонах человеческого
знания благодаря усилиям и успехам отдельных индивидуумов, у которых есть
интерес, стимул и возможности заниматься тем или иным вопросом. Так, несомненно,
произошло в случае с Жоржем Кювье, которого часто называют «отцом палеонтологии».
Кювье получал образование главным образом в Штутгарте и жил в Париже в конце
1700-х – начале 1800-х годов.7 В
течение нескольких десятилетий он описал анатомию сотен ископаемых организмов,
установив реальность процессов вымирания и изменения по мере течения геологического
времени. Он жил до Дарвина и никогда не сочувствовал эволюционным идеям своего
прежнего преподавателя Жана-Батиста Ламарка или же Этьена Жоффруа Сент-Илера,
человека, очень интересовавшегося индивидуальным развитием организмов и столь
же выдающегося, как и сам Кювье. Статуями или бюстами представителей всего
этого ушедшего цвета французской науки может полюбоваться любой турист в Париже,
посетив замечательный Музей Естествознания и Ботанический сад (Jardin des
Plantes). Анатомические труды Кювье были широко известны, а его видный политический
статус во время и после правления Наполеона также способствовал его заметности
и распространению его энциклопедических работ.
11
Главный труд Кювье, «Leçons d’anatomie», был переведен на немецкий язык известным
анатомом, работавшим в Галле, Иоганном Фридрихом Меккелем-младшим. Собственные
труды Меккеля также касалась анатомических сравнений, но они относились к
эмбрионам, и его особенно интересовали уродства в процессе индивидуального
развития. То же самое, что Кювье делал с ископаемыми организмами в Париже,
Меккель делал с эмбрионами в Галле, когда собрал огромную коллекцию зародышей
многих видов позвоночных животных. Его наследие можно увидеть даже в базовой
анатомической терминологии: меккелев хрящ, значительная часть скелета нижней
челюсти у эмбриона человека, и меккелев дивертикул, часть подвздошной кишки
у некоторых людей. Меккель видел свою задачу как дополнение к работе Кювье
в области анатомии, но с позиций эмбриологии. С него началась длинная череда
биологов, изучающих эмбрионы. Меккель видел параллели между различиями среди
взрослых особей ныне живущих и ископаемых видов с одной стороны, и различиями
между эмбрионами одного и того же вида на разных временных отрезках в ходе
индивидуального развития с другой стороны. Выяснение пределов, верности и
значения этого потрясающего факта – это как раз то, с чем было связано многое
в биологии на протяжении пары веков.
Жаль, что Кювье и Меккель не дожили до наших дней, чтобы увидеть окаменелость,
которая дала бы им шанс поразмыслить и над вымиранием, и над уродствами одновременно.
Команда французских и китайских палеонтологов, работающая в северном Китае,
опубликовала в 2007 году сообщение об открытии весьма необычного экземпляра
рептилии, принадлежащей к хористодерам, которые представляют собой группу
полуводных рептилий с неясными родственными связями. Найденный в породах мелового
периода, знаменитых скрытыми в них замечательными окаменелостями крылатых
динозавров и ранних млекопитающих, скелет прекрасной сохранности имеет две
головы, каждая из которых связана со своим собственным набором шейных позвонков,
и остальная часть скелета также полностью сохранилась. Свёрнутая поза этого
и других
12
сходных, но обладающих одной головой экземпляров предполагает, что эта особь
представляет собой уродливый эмбрион. Принадлежность этих ископаемых остатков
к группе Choristodera была определена главным образом по длинным шейным позвонкам
и коротким конечностям – это две характерных особенности этой группы, известной
по достаточно полным и многочисленным экземплярам взрослых особей из этой
формации. Уродство, представляющее собой раздвоение головы/шеи, обычно для
ныне живущих рептилий и называется аксиальной бифуркацией. Это состояние не
обязательно фатально, поскольку многие черепахи и змеи по много лет выживают
с ним в неволе. Очевидно, хористодеры обладают общей с ныне живущими рептилиями
системой индивидуального развития, в которой мог возникнуть этот вид аномалии
развития организма.
Возможность изучать уродства в развитии организма по ископаемым остаткам в
большей степени возможна в царстве беспозвоночных, где палеонтолог собирает
и изучает сотни, если не тысячи образцов. Позвоночные встречаются реже и в
большинстве случаев требуют больше времени для препарирования. Параллели между
эмбриологией и палеонтологией чаще всего ищутся не среди необычных окаменелостей,
а скорее путём сравнения двух свидетельств или морфологических преобразований.
Так происходило даже в случае некоторых учёных с антиэволюционным мировоззрением,
как до, так и после Дарвина. Например, натуралист Луи Агассиц, родом из Швейцарии,
ученик Кювье, занимался поиском отношений между «градациями» таксонов, преобразованиями
в развитии организмов и геологической последовательностю окаменелостей. Под
«градацией» он понимал то, что между таксонами существуют различия в той или
иной степени, поэтому можно мысленно преобразовать один таксон в другой, если
исследовать природу этих различий. Агассиц понял, что исследование индивидуального
развития, а также изучение ископаемых остатков в контексте геологического
времени представляют собой параллели, проясняющие понимание этих преобразований,
даже если он не думал, что механизмом, в конечном счёте ответственным за это,
была эволюция. Пример можно прочитать
13
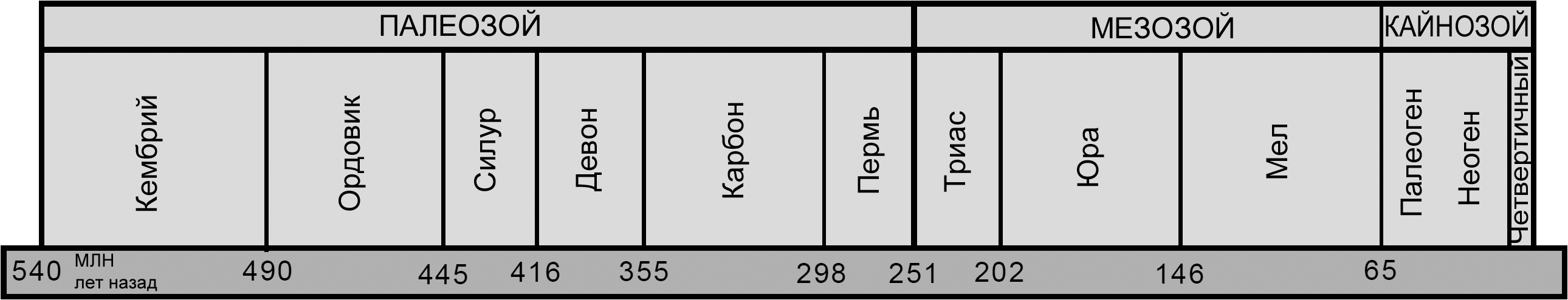
 Рисунок
2. Реконструкция позднего эмбриона или новорождённой рептилии-хористодеры
из меловых отложений Китая. Двухголовое состояние называется антериорной
аксиальной бифуркацией. Общая длина животного, не считая хвоста, была
приблизительно 40 мм. Рисунок Мадлен Гейгер, основанный на статье Buffetaut
и др. 2007. Рисунок
2. Реконструкция позднего эмбриона или новорождённой рептилии-хористодеры
из меловых отложений Китая. Двухголовое состояние называется антериорной
аксиальной бифуркацией. Общая длина животного, не считая хвоста, была
приблизительно 40 мм. Рисунок Мадлен Гейгер, основанный на статье Buffetaut
и др. 2007. |
в его крупном труде «Recherches sur les poissons fossils», изданном несколькими
томами в период между 1833 и 1843 гг. Агассиц утверждал, что хвост взрослых
ископаемых лучепёрых рыб, группы, к которой принадлежат почти все современные
виды рыб, кроме целакантов и двоякодышащих, демонстрирует параллели с развитием
хвоста современных видов этой группы от эмбрионов к взрослым особям. Молодые
особи имеют простой хвост, у более поздних стадий имеется гетероцеркальный
хвост, в котором верхняя часть крупнее, чем нижняя, а взрослая особь обладает
хвостом, у которого обе части сходного размера. Эти изменения подобны последовательности,
которую Агассиц описал для ископаемых форм.
То, что черты различия, а также и сходства между видами были результатом общего
происхождения с изменениями, произошедшими эволюционным путём, было признано
многими другими биологами после Дарвина, в особенности явно немецким интеллектуалом
Эрнстом Геккелем, которому было двадцать пять лет в 1859 году, когда книга
«Происхождение видов…»
14
была впервые издана. Как утверждает его самый новый и осведомлённый биограф,
в Европе, в том числе в Англии, больше людей прочитало об эволюции в книгах
Геккеля, чем в книгах самого Дарвина или любого другого автора. Геккель был
первым человеком после 1859 года, кто открыто опубликовал эволюционное древо
с существующими в природе группами организмов. Геккель был также склонен к
популяризации фактов и в конечном счёте главным образом его считают ответственным
за случившуюся позже историю с утверждением «онтогенез рекапитулирует филогенез».8
Этот принцип заключает в себе идею о том, что индивидуальное развитие так
или иначе отражает в себе различия между видами. Это утверждение имеет историю,
восходящую даже к главному учителю Кювье, Карлу Фридриху Кильмейеру из Тюбингена,
чьи лекции в университете, как говорили, привлекали целые толпы людей. Параллель
между развитием и эволюцией была классическим предметом исследований и обсуждения
в немецкой биологии, что отразилось и в немецком языке: одно и то же слово
«Entwicklung» может использоваться для описания как эволюции, так
и индивидуального развития.9
Параллели между трансформациями
в палеонтологии и в индивидуальном развитии
Рука раннего человеческого зародыша выглядит как весло, поверхностно напоминая
конечность наших водных предков. Существует много других примеров общего сходства
между палеонтологическими изменениями и изменениями, связанными с индивидуальным
развитием. Наиболее знаменитый из них – это тот, который относится к эволюции
и развитию наших нижней челюсти и среднего уха, особенностей, которые мы разделяем
со всеми остальными млекопитающими.
Этот случай часто приводится как выдающееся достижение сравнительной биологии,
и потому его стоит рассмотреть более подробно.
У млекопитающих имеется единственная кость, образующая нижнюю челюсть; она
называется
15
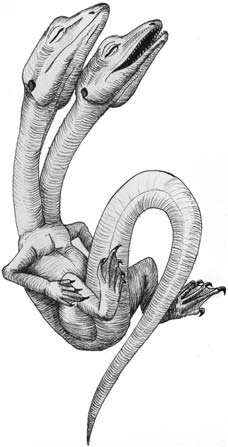
| 
|
Рисунок 3. Зародыши человека различного
возраста на протяжении первых трёх месяцев развития, какими они изображены
в классической работе Ф. Кейбеля (1906), частично по работе Вильгельма
Гиса. Не в одном масштабе. |
зубной костью – подходящее имя, поскольку это как раз то место, где расположены
зубы. У всех прочих позвоночных в челюстях имеется много костей. Млекопитающие
обладают тремя слуховыми косточками, тогда как у других позвоночных со сходным
типом уха есть только одна. Хотя у нас может даже мысли не возникнуть о том,
что наши челюсти и уши как-то связаны, фактически это именно так. Млекопитающие
– большие оригиналы с точки зрения анатомии, и мы, основываясь на дарвиновском
понимании эволюционной биологии, можем сделать вывод о том, что наши ранние
предки, должно быть, в какой-то точке нашей истории были подобны прочим позвоночным.
В действительности окаменелости показывают, что наш общий с рептилиями предок,
который жил, по крайней мере, около 315 миллионов лет назад, обладал
16
несколькими челюстными костями и простым средним ухом с единственной слуховой
косточкой. Первые окаменелости, документирующие преобразования, которые привели
к анатомии современных млекопитающих, были найдены в Южной Африке и в России
в 1840-е годы, но с тех пор ископаемые остатки самых ранних предков млекопитающих
были обнаружены на всех континентах.
На протяжении истории млекопитающих челюстные кости, кроме зубной кости, подверглись
редукции, и в процессе эволюции образовался сустав между зубной костью и черепом.
Прежний челюстной сустав становился всё меньше и меньшие и постепенно отделился
от нижней челюсти. Сочленение между костями сохранилось, но с другой функцией,
а именно для передачи звуковых волн, и со временем оно приобрело специализацию
к передаче высокочастотных звуков. Исходные сочленовная и квадратная челюстные
кости называются у млекопитающих malleus и incus как элементы уха, или, говоря
более простым языком, это молоточек и наковальня. Вместе со stapes (стремечком)
эти кости образуют три косточки среднего уха млекопитающих. Связи между косточками
образуют систему рычагов, с помощью которой высокочастотные звуковые волны
передаются к клеткам внутреннего уха, и в конце концов к мозгу для заключительной
обработки. Эти косточки уха млекопитающих, некогда бывшие частью жевательного
аппарата у наших вымерших предков и у наших современных кузенов рыб, по размеру
не намного больше, чем мелкие камешки. По крайней мере, у некоторых млекопитающих
было зарегистрировано существование двух челюстных суставов на ранних стадиях
индивидуального развития: исходный для челюстноротых позвоночных и всё ещё
характерный для рептилий, и новый, имеющийся лишь у млекопитающих – между
зубной костью, нашей единственной костью нижней челюсти, и чешуйчатой костью
черепа. Кости среднего уха прикреплены с помощью меккелева хряща к внутренней
стороне зубной кости. Некоторые виды из раннего триаса демонстрируют точно
такие же особенности строения: два одновременно существующих челюстных сустава.
Первая из найденных окаменелостей, показывающая эту замечательную особенность,
происходит из Южной
17
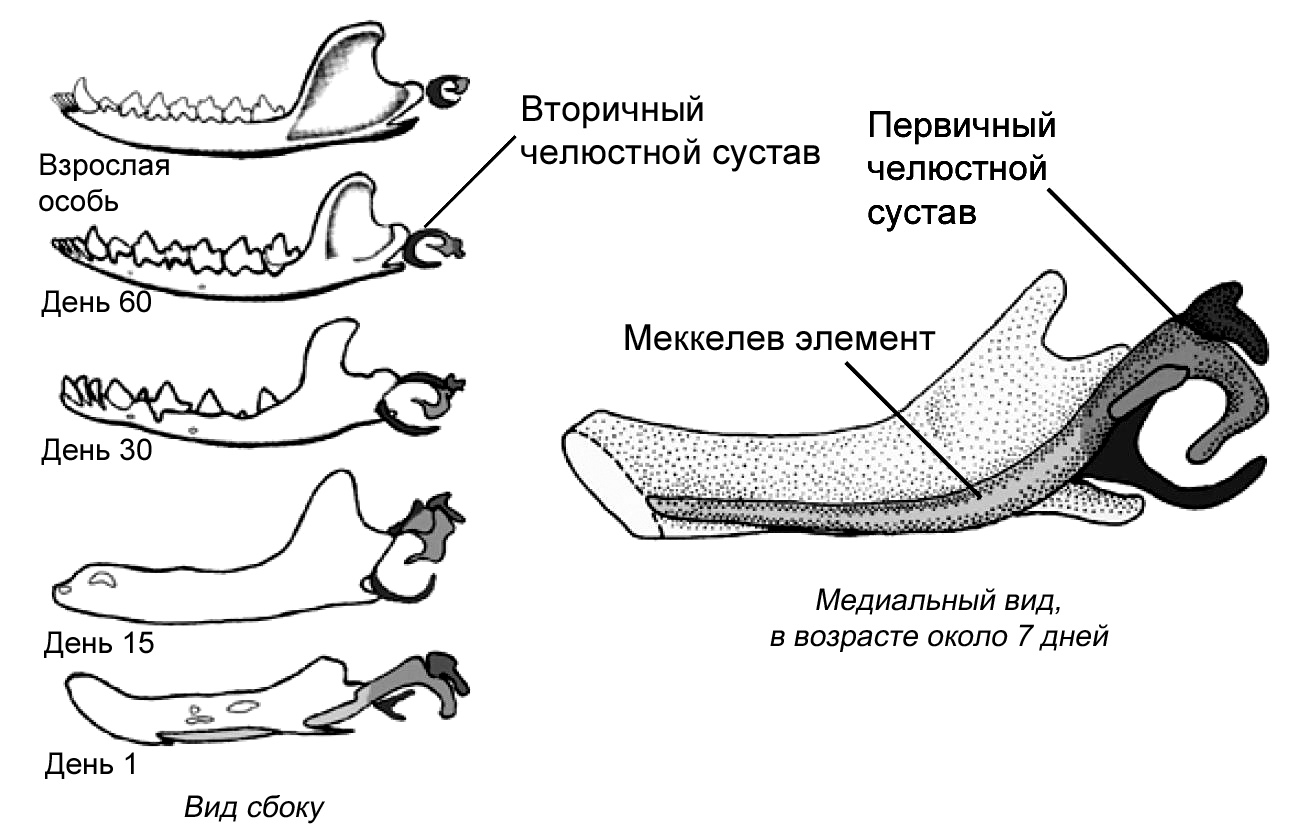
Рисунок 4. Развитие нижней челюсти и косточек среднего
уха у сумчатого млекопитающего Monodelphis domestica. Серия
образцов показывает постепенное уменьшение относительного размера
элементов, которые станут ушными косточками у взрослой особи. Медиальный
вид молодой особи из сумки демонстрирует длинный и заметный меккелев
хрящ, который становится относительно меньше, когда животное растёт.
Не в одинаковом масштабе. По Rowe 1996; и Luo 2007, 2011, с изменениями.
|
Африки и Альфред Кромптон из Гарвардского Университета10
в 1958 году дал ей название Diarthrognathus. Другое
«раннее млекопитающее», хорошо известное палеонтологам – Morganucodon
из позднего триаса/ранней юры, который обладал типичным для млекопитающих
челюстным суставом только между зубной и чешуйчатой костью, но имел видимые
остатки «старой челюстной анатомии». Morganucodon был мелким, размером
с землеройку, и, хотя теперь он известен из Евразии, Африки и Северной Америки,
первоначально он был описан из местонахождений в Англии и Уэльсе.
Этот замечательный пример параллелей между онтогенезом и филогенезом из летописи
окаменелостей не подразумевает того, что одно может использоваться для заполнения
пробелов в другом. Общее подобие обнаруживается в изобилии,
18
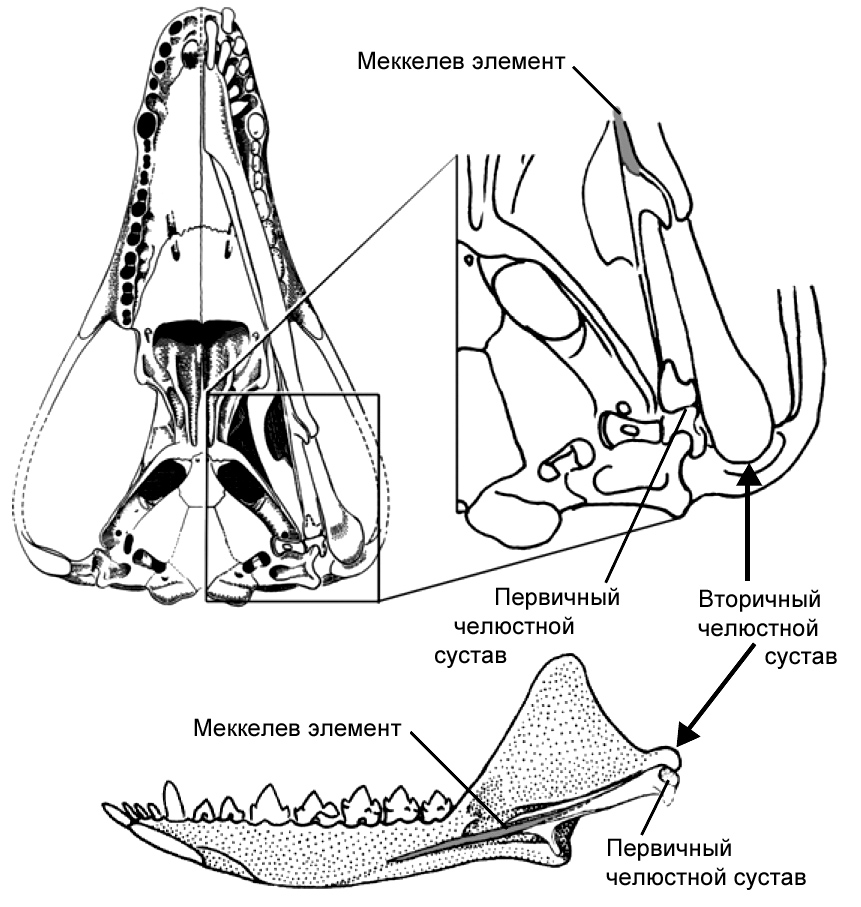
Рисунок 5. Челюстной сустав у «раннего млекопитающего»
Morganucodon. Череп показан с вентральной стороны, а нижняя
челюсть – с медиальной стороны. Длина черепа – около 2,5 см. Рисунок
любезно предоставлен Ч.-С. Луо, с изменениями (см. Luo 2007).
|
но отклонения также обычны. Когда черты сходства присутствуют, они иногда
кажутся поразительными, но при более пристальном рассмотрении оказываются
лишь поверхностными. Молодая особь сумчатого, всё ещё прикрепляющаяся к своей
матери, обладает двойным челюстным сочленением на стадии, когда череп ещё
не окостенел и расположение мускулатуры и мягких тканей ещё плохо различимо.
Это довольно сильно отличается от анатомии взрослого Diarthrognathus
(с двумя челюстными суставами), который
19
бродил примерно 200 миллионов лет назад по тем местам, которые сегодня являются
южноафриканской пустыней Карру.11
Онтогенез обычно не является
рекапитуляцией филогенеза
Тот факт, что онтогенез не является простой рекапитуляцией филогенеза, достаточно
общепризнан даже при том, что общие параллели находятся в изобилии. Чтобы
понять, что означает явление рекапитуляции в том смысле, в каком его определил
аналитическим путём Геккель, мы используем простую схему. Чтобы произошла
истинная рекапитуляция, дополнение должно было бы возникнуть в конце исходной,
или предковой последовательности появления признаков или на возможном продолжении
траектории. На рисунке 6 у нас изображены четыре вида, каждый из которых характеризуется
обычным путём индивидуального развития, состоящим из первого шага, M1 – M1.
У каждого вида новая стадия добавляется в конце последовательности. В этом
идеальном случае вид с наиболее специализированным состоянием, вид «D», сохраняет
в своём онтогенезе последовательность эволюционных преобразований, которые
совершились путём добавления новых стадий в конечные этапы развития. В ходе
эволюции случаются отклонения от гипотетического хода рекапитуляции, что признавал
и сам Геккель. Эти изменения означают, что последовательность появления признаков
в цепочке может сдвигаться, один или больше признаков могут исчезать, или
же может возникнуть совершенно новый признак. Эти изменения иногда настолько
драматичны или считаются настолько важными, что их рассматривают как эволюционные
новшества; так произошло в случаях происхождения волос у млекопитающих или
перьев у динозавров. Существуют различные виды отклонений от рекапитуляции,
среди них гетерохрония (изменения во времени появления) и гетеротопия (изменения
в пространственном расположении в структуре).
Некоторые палеонтологи исследовали эволюционные преобразования, документированные
ископаемыми остатками, и сравнили их непосредственно с онтогенетическими преобразованиями,
отыскивая параллели или
20
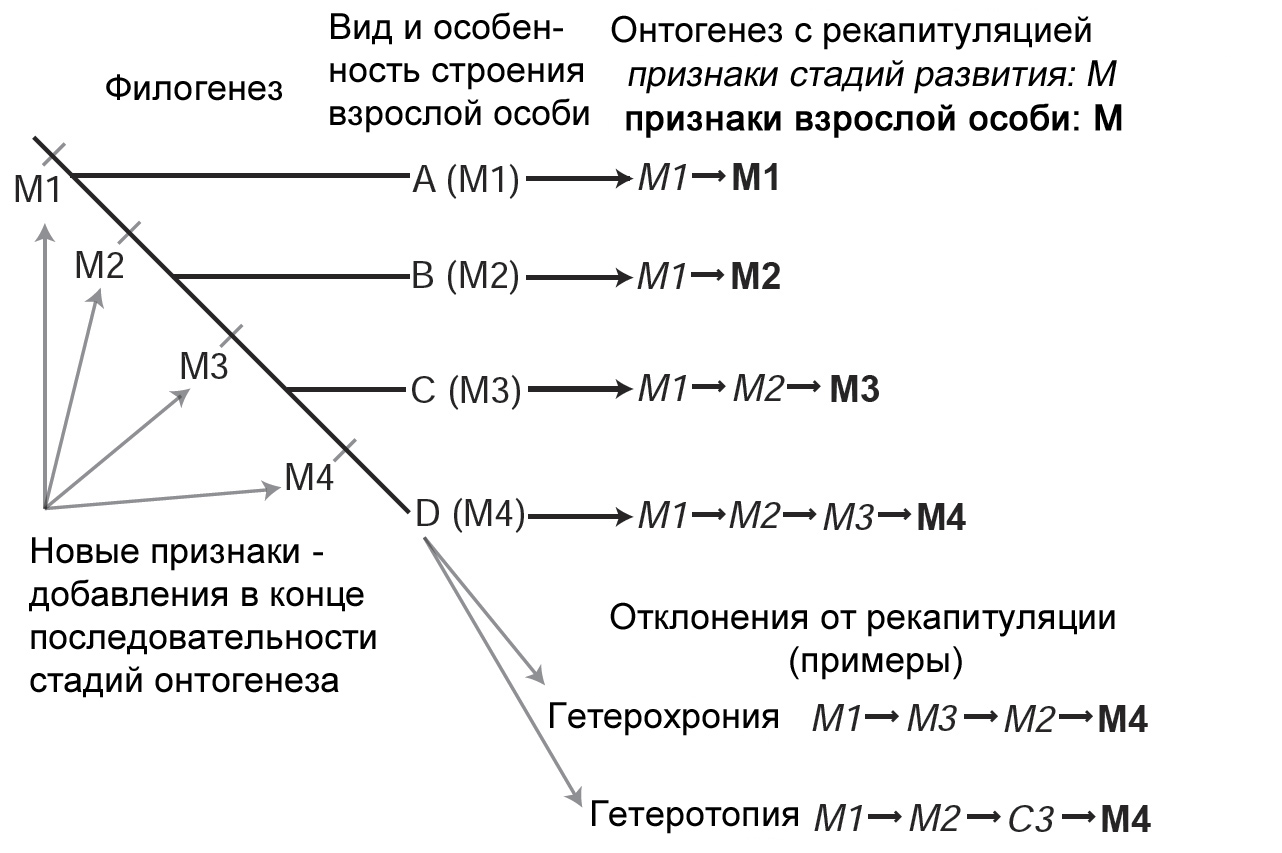
Рисунок 6. Последовательный характер
добавления новых признаков на конечных стадиях в эволюции онтогенетических
последовательностей приводит к онтогенетической рекапитуляции, но имеются
отклонения от этого способа преобразований. По Wägele 2005, с изменениями. |
их отсутствие. Например, изменения в развивающемся черепе эмбрионов аллигаторов
очень схожи с теми, которые наблюдаются в ряду ископаемых остатков ранних
крокодилов. Это сходство не просто поверхностное, но соотносится со статистически
существенной корреляцией в порядке, в котором в обеих последовательностях
происходят дискретные изменения тех же самых признаков. Итак, несомненно,
что существуют параллели между онтогенезом и летописью окаменелостей, показывающих
морфологические изменения в некоторых структурах у некоторых животных. Но
это исключения.
Эволюция онтогенеза
Каждый этап развития в жизни организма может подвергнуться изменениям в процессе
эволюции. Этого и следует ожидать, потому что живые организмы подвергаются
давлению со стороны окружающей среды на каждой стадии
21
и в каждый момент своей жизни. Головастик противостоит таким обстоятельствам,
как взаимоотношения с хищниками или поиск пищи, и на этой стадии в процессе
эволюции возможно появление специализаций. Фактически, при изучении лягушек
и других амфибий,12 таких,
как саламандры, было выявлено ошеломляющее разнообразие эмбриональной морфологии
и видоспецифичных особенностей индивидуального развития. Кристиан Митгатч,
ранее работавший в моей лаборатории в Цюрихе, и научный руководитель его работы
на соискание степени доктора философии Леннарт Олссон, в Йене (Германия),
в том же месте, где Геккель сделал свою долгую и знаменитую академическую
карьеру, продемонстрировали большое разнообразие у ранних эмбрионов лягушек,
проявляющееся в разном времени развития клеточных предшественников многих
черепных структур. Они осуществили это, прослеживая движение клеток из нервного
гребня, который у позвоночных является зародышевым листком, ответственным
за развитие многих тканей, специфичных для позвоночных. Эти ткани включают,
например, элементы зубов и много черепных костей. Работа Митгатча и Олссона
дала нам прекрасные изображения эмбрионов, показывающие потоки клеток, мигрирующих
в разное время и в различных пропорциях у разных видов (рисунок 7).
Самые ранние сухопутные позвоночные, которые населяли Землю, представляют
собой превосходный палеонтологический пример изменений на разнообразных стадиях
онтогенеза. Среди этих животных были некоторые из предков современных лягушек,
саламандр и группы безногих амфибий. Наиболее разнообразными среди этих первых
«амфибий» были темноспондилы (Temnospondyli), группа, существовавшая с нижнего
карбона, примерно 340 миллионов лет назад, до нижнего мела, около 120 миллионов
лет назад. Большая часть времени их существования перекрывается со временем
существования наземных динозавров. Самые мелкие из них были менее 30 сантиметров
в длину, но некоторые формы напоминали крокодилов и достигали длины около
7 метров. Ювенильные и личиночные стадии описаны для многих темноспондилов,
22
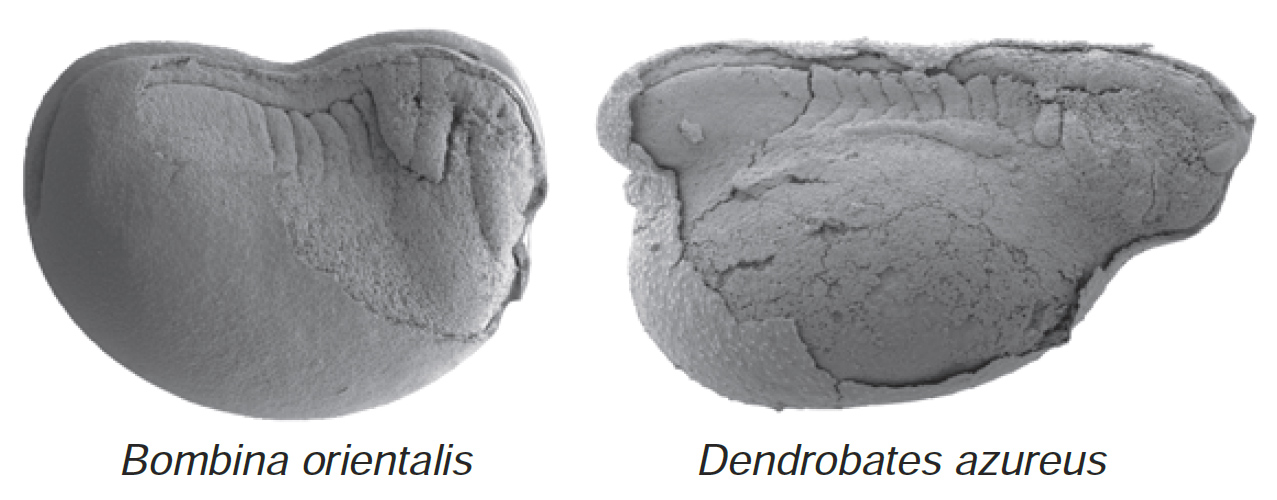
Рисунок 7. Различная степень развития сегментов,
или повторяющихся блоков эмбриональных тканей тела (сомитов) в начале
миграции клеток нервного гребня у эмбрионов лягушек Bombina orientalis
и Dendrobates azureus. Фото любезно предоставили Кристиан
Митгатч и Леннарт Олссон.
|
и они показывают большое разнообразие экологии личинок и характера роста.
Все темноспондилы выводили своё потомство в воде, но типы водных местообитаний
были различными. Некоторые группы предпочитали или выдерживали солоноватую,
или даже морскую воду; другие ограничили своё распространение озёрами.13
Эти личинки были исключительно плотоядными, на что указывают
характер их зубов и, в некоторых случаях, мощные челюсти. Другая ископаемая
фауна, зарегистрированная на участках, где были обнаружены эти окаменелости
прекрасной сохранности, а также подробное изучение анатомии жевательного аппарата
или случайно сохранившегося содержимого желудка темноспондилов показывает,
что некоторые из них охотились на рыбу, тогда как другие поедали различных
мелких «амфибий».
Замечательная сохранность разнообразных жизненных стадий у темноспондилов
отчасти объясняется их большим размером, что повышает вероятность их сохранения,
а также делает их легче заметными для палеонтологов во время работы в поле.14
A родственная группа, лепоспондилы, была, как и самые ранние
«амфибии», представлена мелкими формами: в размерном диапазоне от 5 до 20
сантиметров. Об онтогенезе лепоспондилов почти ничего известно, и это бросает
вызов пале-
23
онтологам, пытающимся точно установить их родственные связи и происхождение
современных земноводных, используя данные об индивидуальном развитии.
Значительное разнообразие путей онтогенеза демонстрирует группа одних из самых
ранних позвоночных с ногами, или четвероногих, похожих на тритонов – бранхиозавриды.
Почти все двадцать видов этой группы сохраняли личиночные особенности, такие,
как наружные жабры или зубчики на жаберных дугах во взрослом состоянии; это
неотенические признаки. Некоторые другие виды претерпевали метаморфоз и утрачивали
свои личиночные признаки во взрослом состоянии. Все представители группы были
мелкими, в противоположность прочим темноспондилам. Тысячи экземпляров некоторых
бранхиозаврид известны из местонахождений в Германии, включая одно в бассейне
Тюрингенского Леса. Там Ральф Вернебург много лет изучал ископаемые остатки
темноспондилов, таких, как бранхиозавры. Некоторые виды бранхиозаврид обладали
особенностями, которые позволяли считать их ручьевыми формами; другие, с видимыми
наружными жабрами, описаны как озёрные формы. Один из спорных вопросов для
экспертов – могут ли в некоторых случаях два типа возникать как различные
экоморфотипы одного и того же вида, продвигая онтогенетическую изменчивость
ещё на один шаг вперёд, демонстрируя случай «фенотипической пластичности»,
тему, которую я обсуждают в главе 2.
Как и в вышеприведённых примерах, любая стадия на протяжении развития индивидуума
может изменяться. Это демонстрировалось опытным путём для многих видов позвоночных
и стадий процесса. Эволюционируют все стадии онтогенеза, а не только взрослые,
объект приложения усилий многих исследователей. Многие авторы представляют
данные и идеи, призванные подвергнуть критике «соломенное чучело», защищающее
наивную рекапитуляционную идею, согласно которой онтогенетические и эволюционные
преобразования точно отражают друг друга. И важнее всего тут то, что желание
исследовать фундаментальный аспект того, как эволюционировало индивидуальное
развитие, привело к пристальному изучению организмов из многих групп, и итогом
этого стало несколько значительных открытий. Например, Майк Ричардсон и
24
| 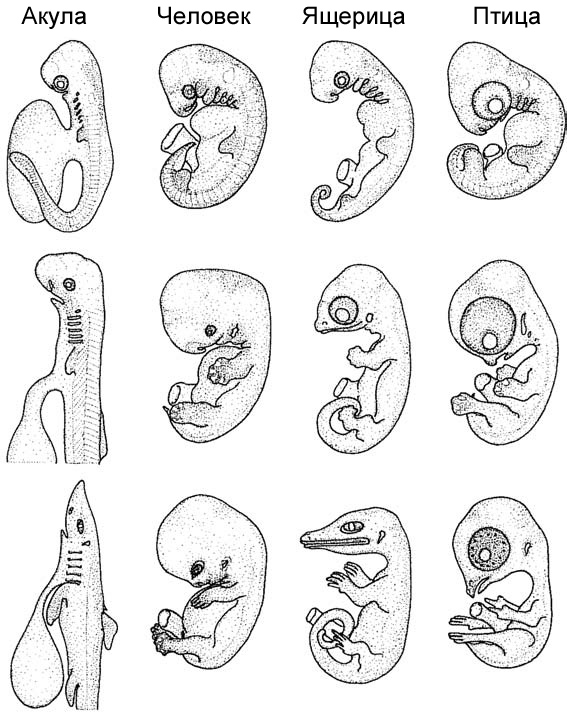
|
| Рисунок 8. Три стадии развития у акулы, человека, ящерицы
и птицы. По Westheide и Rieger 2009, с изменениями. |
25
немного позже Ингмар Вернебург документировали внешнюю морфологию эмбрионов,
которая уже в первом приближении показывает, что в более или менее сопоставимые
периоды индивидуального развития имеются существенные различия в пропорциях
и формах, а также в размере желтка у рыб, змей, лягушек, птиц, ящериц и млекопитающих,
в том числе людей. Эта особенность – лишь одна из многих, иллюстрирующих различные
физиологические и морфологические особенности на протяжении времени индивидуального
развития и у различных видов.15
Онтогенез и условия для
поддержания жизни
Если индивидуальное развитие и рассказывает
нам историю, то она похожа на те страшные истории, которые рассказывают
дети, сидящие вокруг походного костра, когда каждый мальчишка добавляет
свой отрывок к тому, что уже было до него. Нет ни одного единого рассказчика,
и нет внутреннего диалога. Есть только то, что уже произошло, что происходит
сейчас, и что произойдёт потом.
Марк С. Блумберг «Freaks of Nature»
|
Изменения в тканях, которые происходят в ходе индивидуального развития, протекают
для живого эмбриона в его настоящем времени. С этой точки зрения может показаться,
что взрослые особи – это почти случайность, или же один из нескольких моментальных
снимков жизни. Это не означает, что особенности какой-либо стадии развития
случайны; они являются результатом процессов, которые мы можем во многих случаях
количественно определить и предсказать – продуктом чётко скоординированных
и детерминированных механизмов, где пластичность и случайность играют главную
роль в формировании результата. Есть генетическая система, отвечающая за индивидуальное
развитие, которая работает в контексте окружающей среды. Действие этой системы
завершается там, где начинается фенотипическое проявление процессов индивидуального
развития, которое управляется физическими и химическими законами, которые
работают в природе.
Объяснение существования того или иного признака на любой из стадий
26
развития, отличающейся от взрослого состояния, обычно делается в рамках понятий
будущего состояния, к которому движется животное. Как утверждал Джон Реисс
в своей книге «Not by Design», объяснение существования признака должно делаться
в рамках его необходимости на момент его существования, но не в терминах будущего
состояния, движению к которому он способствует.
Чтобы понять этот основной принцип, полезнее всего обратиться к идее «условий
для поддержания жизни», выдвинутой Кювье. «Условия», о которых говорил Кювье
– это особенности живого существа, без которых оно не могло бы выжить. Организм
не может существовать, пока не будут удовлетворены его условия для поддержания
жизни. Это может показаться тавтологическим или тривиальным, но, подобно многим
великим истинам, идея становится фундаментальной лишь после того, как она
была полностью признана.
Эта идея имеет важные значения для концептуальных рамок, которыми мы пользуемся,
чтобы исследовать эволюцию индивидуального развития. Например, при рассмотрении
экологических аспектов эволюционного разнообразия понятие «ниша» часто используется
так, словно она должна была существовать в окружающем мире, и словно это цель,
к которой стремится группа организмов. Как обсуждалось ранее, ниша не существует
независимо от организмов, которые фактически создают её. Среди любых форм
жизни организм воздействует на окружающую среду, но также подвергается воздействию
с её стороны.16 Когда
образуется новый океанический остров, со временем некоторые организмы, случайно
оказавшиеся там, будут вести себя так же. Они не будут поступать так в соответствии
с неким планом или целью; они просто будут пытаться выжить. Большинство биологов
не настолько наивно, чтобы думать, что у живых организмов есть план, но изложение
часть строится на основе разработанного плана. Естественный отбор ограничивает
эволюционирующую совокупность, «выбирая» без плана среди множества экспериментов,
которые индивидуальное развитие проделывает в новом контексте окружающей среды.
Поскольку живые организмы – это мозаика признаков, которые все объединены,
но некоторые – теснее прочих, индивидуальное развитие под действием естественного
отбора будет эволюционировать сложным образом. Эволюционные изменения у островных
видов иногда объясняются
27
как результат остановки или замедления развития, при которых островная форма
просто напоминает ювенильную стадию своего предка. Но живые организмы мозаичны.
Каждый орган или часть тела, хотя и составляет единое целое с остальными,
управляется различными правилами относительного роста. Островной вид может
быть в общих чертах подобен ювенильной особи предкового вида, но это может
быть лишь поверхностным сходством, или же быть верным лишь для некоторых особенностей.
«Мозаика» образуется в каждый из моментов эволюционного времени из-за того,
что условия для подержания жизни должны удовлетворяться на каждой из стадий
индивидуального развития.
Я обсудил несколько параллелей между биологией развития и палеонтологией по
предмету и по подходу. Ещё одна – это телеологическая позиция, которая утверждает,
что в основе изменений лежит замысел, и он направлен в сторону улучшения.
Это та позиция, которая преобладала в этих двух дисциплинах. Идея наличия
замысла и цели связана с телеологией, термином, доставшимся нам от греческих
философов, которые первыми обсуждали в различных контекстах вопросы наличия
предназначения и цели у жизни. Если даже кто-то и принимает принципы множества
религий или иных форм веры в сверхъестественное, телеология совершенно неуместна
в любой научной дискуссии, касающейся эволюции. И всё же поиск объяснений
и незнание принципов самоорганизации жизни привели в прошлом к телеологической
концепции жизни.
Изучение ископаемых остатков не поддерживает телеологическое представление
о жизни. Летопись окаменелостей демонстрирует многочисленные образцы отличающихся
друг от друга и параллельных морфологических изменений. Классические примеры
– это ветвистое эволюционное дерево лошадей, берущее своё начало в раннем
эоцене, примерно 55 миллионов лет назад, или родословная людей, когда примерно
два миллиона лет назад одновременно жило пять видов гоминид, но лишь один
из них существует сегодня.
Формы телеологического мышления, которые по современным стандартам выглядят
весьма причудливыми, были весьма обычными в истории био-
28
логии. Во многих случаях утверждается о существовании некоторого рода жизненной
силы, которая осуществляет эволюционное изменение в предопределённом направлении,
будучи вызванной свойствами, присущими организмам. Примеры такого рода – принцип
врождённого стремления к совершенствованию Ламарка и идея «ортогенеза»; к
последней обращался на страницах своих книг влиятельный биолог Теодор Эймер.
Эти идеи витализма и внутренних движущих сил жизни очень повлияли на палеонтологию.
Идею эволюции, идущей посредством «аристогенеза», вида ортогенеза, подразумевающего
постоянное совершенствование, отстаивал, например, Генри Ф. Осборн, ведущая
фигура в американской палеонтологии, много лет занимавший должность президента
Американского Музея Естественной Истории в Нью-Йорке в первые десятилетия
двадцатого века.17
Онтогенез и филогенез
Некоторые иллюстративные материалы весьма поучительны и помогают глубже понять
отношения между онтогенезом и филогенезом. Я приведу здесь два своих любимых
примера. Первый принадлежит немецкому ботанику Вальтеру Циммерману (1892 –
1980), который, помимо своей работы в области морфологии растений, был оригинальным
и влиятельным писателем на темы теории эволюции и методов биологической классификации.
На варианте «схемы Циммермана», воспроизведённом здесь (рисунок 9), есть спирали
онтогенеза, включающие в себя различные стадии жизни, изменяющиеся в ходе
геологического времени. Филогенез является эволюцией онтогенеза, в данном
случае он проиллюстрирован происхождением птиц, с символичным археоптериксом,
«недостающим звеном между птицами и рептилиями» из Баварии, реконструкция
которого помещена на несколько ступеней ниже текущего состояния онтогенеза
птиц. Полный спектр изменений во времени, включающий и филогенез, и онтогенез,
Циммерман называет «гологенезом», термином, который никогда не был общепринятым
в стандартной терминологии на моём поле деятельности.
29
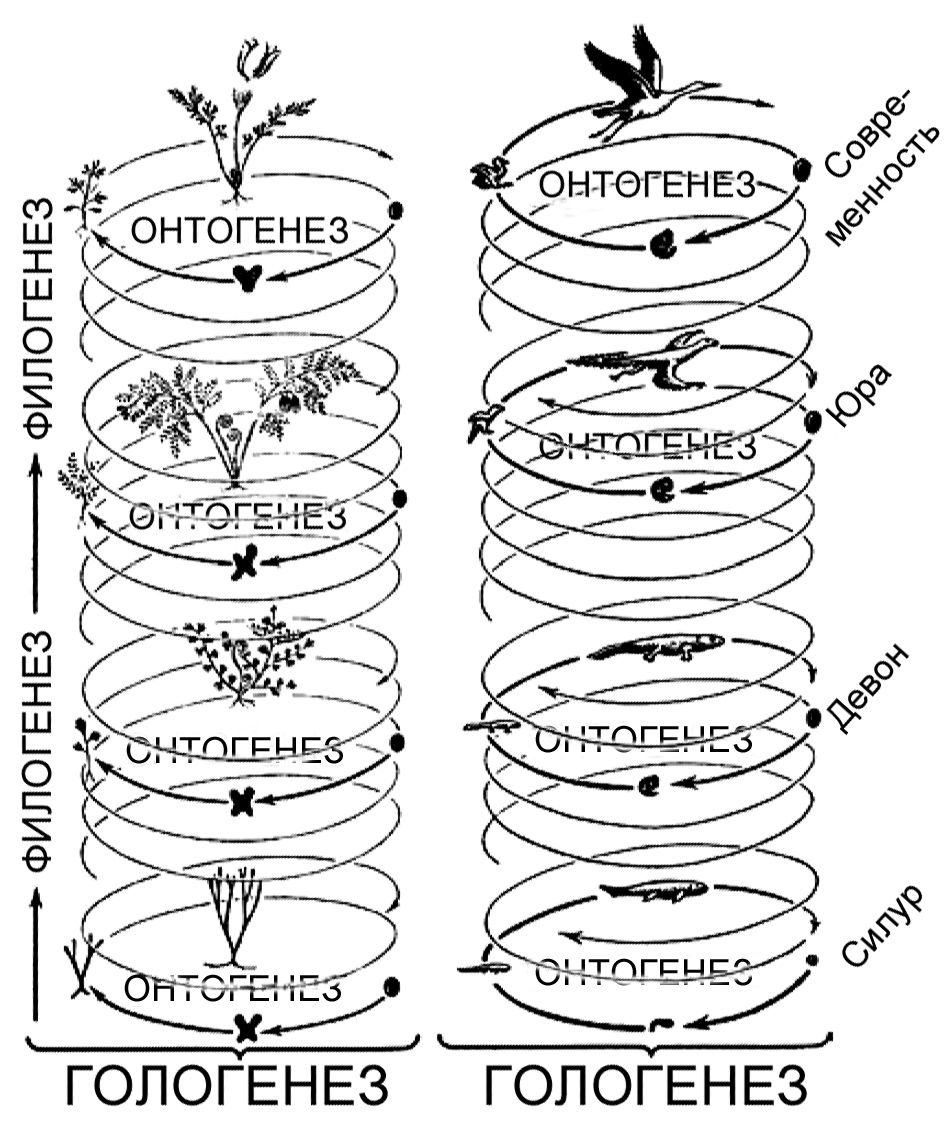
Рисунок 9. Отношения между онтогенезом
и филогенезом, как их изобразил Вальтер Циммерман. По Циммерману, 1968,
с изменениями. |
Другую схему, которая прекрасно иллюстрирует эволюцию, создал Вольфганг Майер,
мой бывший наставник в Тюбингенском Университете. Среди его самого значительного
вклада в науку – исследования анатомии млекопитающих ближе к моменту рождения,
когда новорождённые сталкиваются с новыми испытаниями со стороны окружающей
среды. Как любит замечать Майер, разные виды приобрели в процессе эволюции
различную анатомию на этой стадии развития, и это же справедливо для всех
стадий, от зиготы до смерти – на протяжении истории всей жизни. Естественный
отбор действует на всех стадиях, и в процессе эволюции меняются пакеты генов,
структуры и функции, работающие на разнообразных уровнях биологической организации.
В некоторых случаях это изменение происходит
30
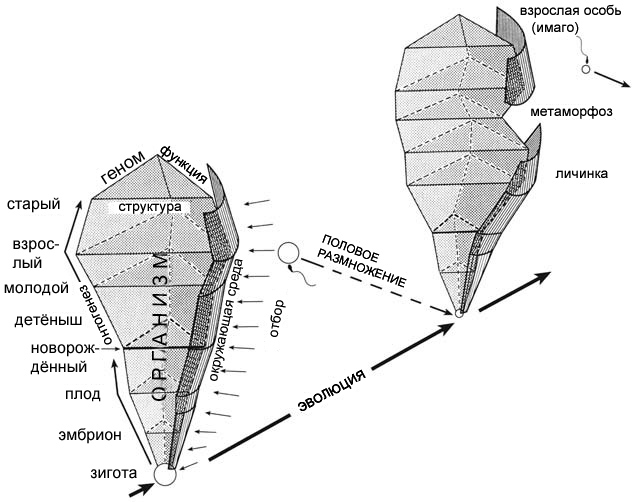
Рисунок 10. Отношения между онтогенезом и филогенезом,
изображённые Вольфгангом Майером. По Maier 1999, с изменениями.
|
постепенно, в сумме приводя к крупным эволюционным преобразованиям по ходу
течения геологического времени. Генетические изменения передаются следующим
поколениям посредством полового размножения.
Можно использовать много переменных величин, чтобы охарактеризовать процессы
онтогенеза, представленные в абстрактной форме на этих рисунках. Среди них
продолжительность эмбриональной и ювенильной стадий, возраст полового созревания,
состояние в момент выхода из яйца или при рождении, а также репродуктивная
долговечность. Все они входят в историю жизни животного, и каждая из них может
эволюционировать. Аналогичным образом эволюционирует также история жизни у
растений. В случае с ныне живущими видами способность связывать эти переменные
величинами с исследованиями в области популяционной динамики важна для вопросов
охраны природы, поскольку они могут
31
выделить стратегии управления для тех организмов, которые рассматриваются
как находящиеся в зоне риска. С эволюционной точки зрения знание этих переменных
величин может помочь нам понять, почему определённые группы вымирают, тогда
как другие становятся многочисленными и разнообразными, или же просто выжили.
Некоторые из этих переменных также связаны с крупными анатомическими новшествами,
как крупный мозг людей отчасти стал возможным благодаря долгому периоду беременности.
Ископаемые остатки являются ключом к нашему знанию о том, когда возникли эти
особенности, даже если они не могут предоставить прямую информацию, касающуюся
некоторых переменных величин, вроде продолжительности беременности. Это становится
возможным, потому что большое количество эмпирических данных показало тесную
ассоциацию с некоторыми анатомическими особенностями, которые сохранились
в ископаемом виде вместе с деталями истории жизни. Как в случае с ранними
наземными позвоночными, которые обсуждались выше, окаменелости, сохранившиеся
наилучшим образом, являются первыми и наиболее очевидными объектами изучения,
позволяющими документировать эволюцию истории жизни индивида в геологическом
прошлом.
Онтогенез и реконструкция
древа жизни
Со времён новаторской работы Эрнста Геккеля и с теорией эволюции в качестве
точки отсчёта основной задачей была реконструкция древа жизни. История подходов
и проблем длинна и сложна. Вопрос, который неоднократно оказывается на поверхности
в ходе дискуссий, состоит в том, как дополнить это древо информацией об онтогенезе.
При использовании морфологии вывод об отношениях между видами обычно делается
путём сравнения наличия и специфичности деталей различных особенностей строения.
В случае специалистов по позвоночным мы главным образом исследуем кости, мышцы
и нервы взрослого животного. Но наследуемые и отличающиеся от вида к виду
особенности, которые необходимы для анализа, позволяющего реконструировать
родословное древо, присутствуют на
32
всех стадиях жизненного цикла животного, поэтому нет никаких причин ограничиваться
только взрослыми особями в качестве источника информации об анатомии. Онтогенез,
таким образом, важен, но как же его правильно использовать? Большинство практикующих
систематиков, биологов, чья деятельность непосредственно связана с реконструкцией
родословного древа и классификацией, согласилось бы со следующим рядом утверждений,
который претендуют на обобщение вопросов, в отношении которых специалисты
большей частью достигли консенсуса.