| Главная | Библиотека | Форум | Гостевая книга |
176
Глава десятая
О трилобитах, ракушках и букашках
До этого момента я рассматривал главным образом развитие позвоночных животных.
В иных, нежели позвоночные, группах животных существует намного больше ныне
живущих видов, и конечно, то же самое справедливо для вымерших видов. В этой
главе я постараюсь показать некоторые из открытий в этой области и тот большой
потенциал, которым обладают исследования этих животных. Чтобы палеонтология
развития была применима к беспозвоночным, потребовалась бы полная переработка
объекта исследования, и здесь этого, разумеется, не пытались делать.
Скелет позвоночного, который как раз и сохраняется в ископаемом состоянии
чаще всего, находится внутри тела и, таким образом, представляет собой эндоскелет.
Для многих других животных характерен наружный экзоскелет, который покрывает
собой мягкие части тела. Так обстоят дела у членистоногих животных, таких,
как обладающие богатым видовым разнообразием насекомые, омары и крабы, а также
вымершие трилобиты.
Как я уже говорил ранее, касаясь палеогистологических исследований, эндоскелет
позвоночных подвергается растянутой во времени модификации по мере роста животного.
Модификация имеющихся в наличии частей
177
эндоскелета позвоночных – это один из четырёх основных типов роста, которые
мы можем наблюдать у животных. Добавление слоёв по мере роста – это другой
вид, это нарастание, типичное для минерализованных раковин. Колебания условий
окружающей среды и история жизни индивидуума отражены в скорости и характере
этого нарастания. Новаторское исследование этого явления привело к открытию
того, что в девоне в году было примерно 400 дней, а в пенсильвании* около
387, как позже подтвердили астрономы, которые выяснили, что скорость вращения
Земли замедляется. Джон Уэллс изучал кораллы в 1960-е годы и заметил у них
тонкие слои нарастания, отражающие ежедневные циклы, а также толстые слои,
отражающие ежегодный прирост. Собрав эту информацию воедино, он сделал это
удивительное открытие.
Другой вид роста происходит не посредством добавления ещё большего количества
слоёв, а добавлением вместо этого большего количества отдельных элементов:
когда морские ежи наращивают новую пластинку на своём панцире или трилобиты
добавляют новый сегмент тела. Случай с трилобитами особенно показателен в
плане эволюции индивидуального развития, о чём я буду говорить далее. Трилобиты,
подобно другим членистоногим, также претерпевали линьку. При таком способе
развития имеющийся экзоскелет неоднократно сбрасывается и образуется новый.
Поскольку, сформировавшись, экзоскелет не может растягиваться, данное животное
должно проходить через стадии «мягкого панциря» между сменами экзоскелета
меньшего размера на новый, больший. Абсорбция минеральных веществ из экзоскелета
для их дальнейшего использования выглядит энергозатратным процессом, но фактически
она имеет место у многих современных членистоногих, хотя явно отсутствовала
у трилобитов. Линечные шкурки могут сохраняться в виде окаменелостей, которые
способны ввести в заблуждение даже специалиста. Форма экзоскелета может сильно
варьировать от одной стадии к другой, и по этой причине нужно множество образцов,
чтобы понять, что серии ископаемых образцов, не связанных друг с другом плавным
переходом, фактически представляют собой стадии развития одного и того же
вида, разделённые промежутками, представляющими собой периоды роста.
* Пенсильваний – верхний карбон. – прим. перев.
178
Трилобиты!
Лишь немногие группы животных, возможно, кроме динозавров и аммонитов, могут
конкурировать с трилобитами за право быть самой узнаваемой группой ископаемых
животных. Восклицательный знак, проставленный выше, таким образом, вполне
оправдан, и он относится к книге Ричарда Форти с таким же названием*. Эта
разнообразная группа существовала с кембрия, примерно 526 миллионов лет назад,
до конца перми, около 252 миллионов лет назад, когда Земля столкнулась с крупнейшим
случаем массового вымирания. Кальцитный скелет трилобитов хорошо сохраняется
в виде окаменелости. Существует около 22000 описанных видов, и это большое
количество видов и их отдельных представителей содержит в себе потенциал для
исследования эволюции индивидуального развития в масштабах, колеблющихся от
уровня популяции до истории группы в целом.
План строения тела трилобитов включает деление на головной/туловищный отделы.
Голова несёт на себе парные антенны плюс три пары других придатков. А туловище
– это самая важная часть тела для исследований в области индивидуального развития,
поскольку мы можем изучать эволюцию количества сегментов тела, их распределение
по отделам тела и способы их роста. В эволюции туловищных сегментов трилобитов
существует важная тенденция. В ходе ранней эволюции трилобитов, в кембрии,
существовало значительное разнообразие количества этих сегментов во взрослом
состоянии на внутривидовом, межвидовом и родовом уровнях. Изменчивость на
этих уровнях снизилась на более поздней стадии эволюции трилобитов. Была выдвинута
гипотеза о том, что это изменение представляло собой «застывание» систем,
управляющих развитием, означавшее возросшее количество ограничений после установления
«каналов» в развитии. После времени эволюционных «экспериментов» процесс развития
более или менее установился, и в нём наверняка были задействованы Hox-гены.
Известно, что у членистоногих, как и у позвоночных, Hox-гены играют важную
роль в разделении тела на отдельные сегменты, и можно принять как факт
* В 2013 году эта книга выпущена на русском языке. – прим. перев.
179
то, что контроль сегментации и разбиения на отделы тела у трилобитов мог
достигаться подобным же образом. Добавление сегментов в тело происходит на
его конце, и, подобно тому, как множество молекул играет сходную роль в этом
процессе у насекомых и ракообразных, так же, вероятно, происходило и у трилобитов.
В главе 7 я указал, что некоторые группы позвоночных
обладают очень пластичными генетическими механизмами сегментации, результатом
чего становится изменчивое количество позвонков, тогда как другие группы очень
консервативны. Аналогичная ситуация имеет место у трилобитов, и Найджел Хьюз,
который провёл много исследований в области эволюции их индивидуального развития,
документировал примеры значительной пластичности в развитии сегментации у
геологически более молодых форм. Если ограничения и существуют, то они не
являются незыблемыми.
В ранний период эволюции трилобитов разбиение тела на отделы было неглубоким
– все сегменты выглядели более или менее похожими друг на друга. У более поздних
трилобитов изменение плана строения тела привело к более ярко выраженному
разделению его на отделы. Различные разновидности трилобитов в процессе эволюции
независимо приобрели отчётливо выраженную многосегментную хвостовую пластину,
состоящую из многочисленных слившихся воедино туловищных сегментов и называемую
пигидиумом. Это новшество имело экологическое значение: оно сделало возможным
сворачивание тела в капсулообразное защитное положение, когда все мягкие части
оказываются эффективно защищёнными. Защита от хищников, вероятнее всего, была
главным фактором отбора, стимулирующим эволюцию механизма сворачивания тела.
Сегменты в новом, увеличенном хвостовом отделе обычно чётко отличались от
предшествующих туловищных сегментов. У трилобитов с данной особенностью строения
количество сегментов стало менее изменчивым. У позвоночных существует аналогичная
ситуация. Животные, у которых разделение позвоночника на отделы нечётко выражено
или вовсе отсутствует, обладают более изменчивым количеством сегментов. Это
наблюдается у змей и китов: и у тех, и у других отсутствует тазовый пояс.
180
Различие между стабильным и изменчивым количеством сегментов может выглядеть
тривиальным фактом. Но оно имеет самое прямое отношение к исследованиям индивидуального
развития, поскольку изменчивость – это ключ к эволюционному потенциалу. Пластичность
в развитии может быть фактором, который следует рассматривать при исследовании
потенциального риска вымирания вида или группы организмов. Исследователи обнаружили,
например, что трилобиты, демонстрировали снижение пластичности с течением
геологического времени, и есть повод задуматься над тем, какое воздействие
это оказало на характер их эволюции – это предмет исследований в настоящее
время.
Эволюция трилобитов с течением времени также исследовалась с использованием
различных подходов. На протяжении десятилетий многие специалисты пробовали
реконструировать части эволюционного древа трилобитов, исследуя особенности
строения, сравнивая их у разных видов и далее проводя их числовой анализ.
Для этого они используют кладистическую методологию, в который распределение
особенностей строения оптимизируется с целью создания «лучшего» эволюционного
древа, с применением принципа парсимонии. В основном лучшее из возможных объяснений
для совокупности данных представляет собой та топология древа, отражающая
главенствующую гипотезу отношений, для которой должно быть сделано наименьшее
количество предположений (отсюда «парсимония»*). В процессе кодирования морфологических
особенностей в целях анализа можно заметить, что у некоторых видов они стабильны.
Но другие особенности могут изменяться в пределах вида – это случаи полиморфизма.
В 2007 году Марк Уэбстер опубликовал исследование, в котором он количественно
определил степень полиморфизма для 982 видов трилобитов с течением геологического
времени и обнаружил, что более древние виды, которые находятся ближе к основанию
эволюционного древа трилобитов, проявляли больший полиморфизм, чем поздние,
более продвинутые виды. Почему же имеет место эта чёткая закономерность? Она
может быть следствием изменений в окружающей среде, которые со временем приводили
к различным типам давления отбора. Или она могла быть связанной с «внутренним»
сужением системы, управляющей индивидуальным развитием. Но как бы то ни было,
* «parsimonious» (англ.) – буквально: «скупой» – прим. перев.
181
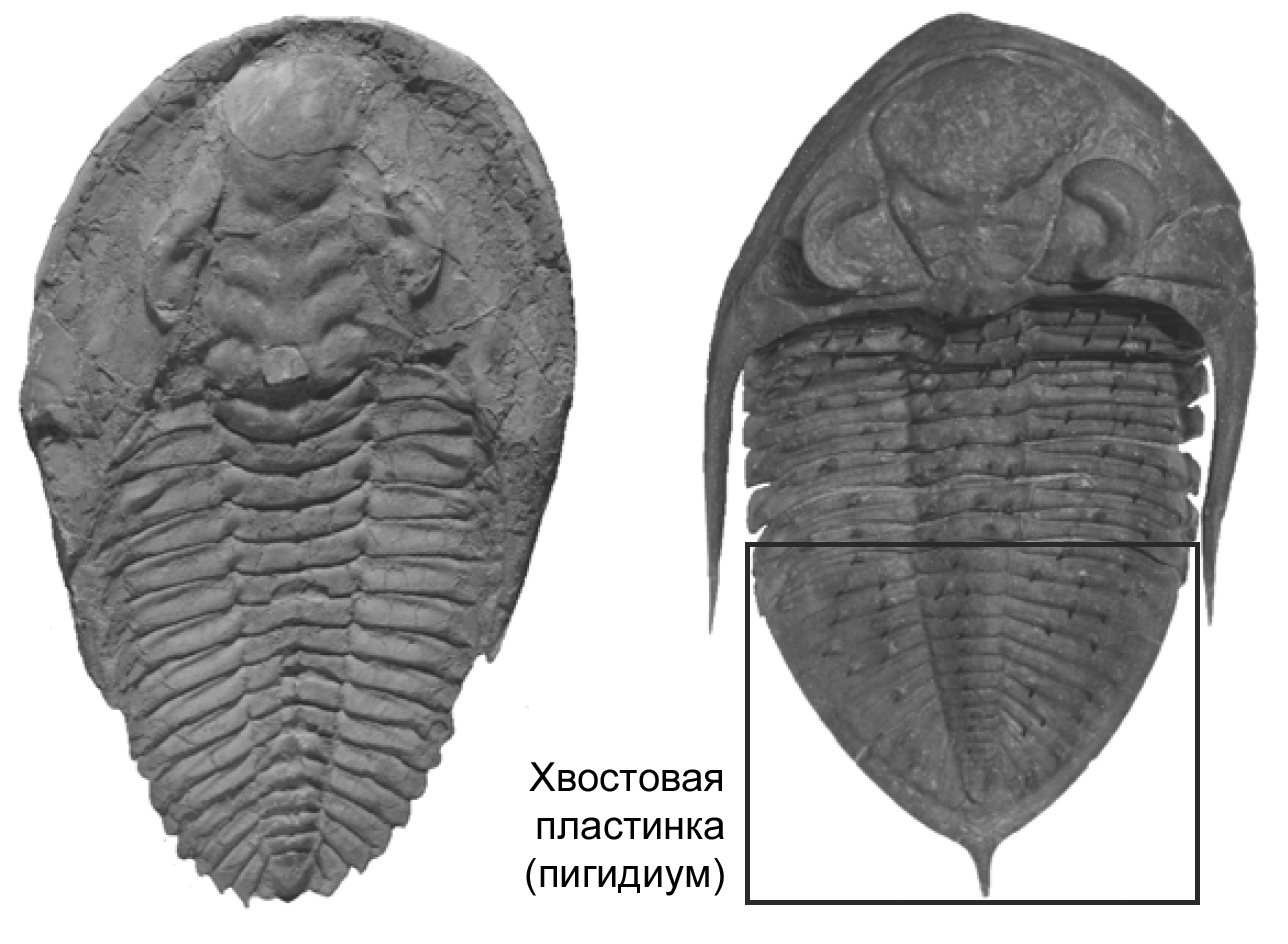
Рисунок 48. Два различных плана строения тела у трилобитов. Слева: в данном случае всё тело состоит из более или менее однородных сегментов до самого конца животного. Этот вид – Cambropallas telesto из среднего кембрия Марокко. Справа: самые задние сегменты достаточно сильно отличаются по строению и образуют хвостовую пластину; это пример разбиения тела на отделы. Данный вид – Zlichovaspis rugosa из раннего/среднего девона области Майдер, Марокко. Фотографии любезно предоставил Кристиан Клюг (Цюрих). |
важно то, что Уэбстер открыл закономерность, сходную с той, которая в общих чертах наблюдается для количества сегментов в масштабах геологического времени. Во всех этих исследованиях индивидуального развития сквозь призму геологического времени очень важно безошибочное определение видов. В случае с ныне живущими видами, если виды распознаны на основе репродуктивной изоляции, экспериментальными манипуляциями можно проверить эту изоляцию. С другой стороны, виды, определяемые таким образом, невозможно выявить в ископаемом виде. Но таксономия трилобитов имеет под собою твёрдые основания. Исследованы сотни и тысячи экземпляров, и степень изменчивости среди них измерена. Непротиворечивые и воспроизводимые критерии, которые работают применительно к ныне живу-
182
щим членистоногим, используются для определения морфологических границ видов. Другие группы беспозвоночных также известны по тысячам образцов, и исследования в области палеонтологии развития применительно к ним также являются возможными.
Биоминерализация
Прекрасная способность к биоминерализации у нескольких групп беспозвоночных
делает их пригодными для палеонтологических исследований, поскольку образующиеся
в итоге панцири обладают потенциалом для сохранения в ископаемом состоянии.1
В процессе эволюции биоминерализация много раз независимо возникала у многих
групп живых организмов, и палеонтологические знаки этого просто невозможно
не увидеть.2 Один
из классических примеров – «меловые утёсы» южной Англии. Этот белый известняк
мелового периода образует белые утёсы Дувра и обращённые к ним утёсы на противоположном
берегу Ла-Манша, во Франции. При большом увеличении можно увидеть, что меловые
утёсы состоят из миллионов кокколитов, каждый из которых размером лишь в несколько
тысячных долей миллиметра в поперечнике. Их произвели одноклеточные морские
водоросли, называемые кокколитофорами. Некоторые растения также могут подвергаться
биоминерализации.
Процесс, который приводит к образованию силикатов в морских водорослях, раковин
моллюсков, а также других карбонатов у беспозвоночных, по своей сути аналогичен
тому, который участвует в образовании карбонатного скелета у позвоночных.
Фосфат и карбонат кальция – это биоминералы, встречающиеся чаще всего. Органические
составляющие, такие, как коллаген и хитин, используются в комплексе с ними,
обеспечивая структурную поддержку костей и панцирей. Получающиеся в итоге
материалы имеют сложную архитектуру на микроскопическом, и даже на наноуровне.
Результатом этого может исключительная стойкость к износу или сопротивление
на излом. Учёные, исследующие материалы и структуры на наноуровне, очень заинтересовались
этими органическими структурами и используют
183
изощрённые методы получения изображений, чтобы обнаружить структуры, которые
развились в ходе эволюции у живых организмов, как у ископаемых, так и у современных.
Одной из целей этого является получение знаний, которые можно применить при
искусственном синтезе органо-неорганических материалов.
На более высоком макроуровне эволюция способности к биоминерализации также
привлекает всё больше и больше внимания по причине понимания того, что это
фундаментальная часть исследований эволюции истории жизни крупных групп организмов,
которые населяют океаны сейчас и населяли их в прошлом (см. главу
6). Кораллы, организмы, активно участвующие в процессах биоминерализации,
представляют особый интерес в настоящее время. Повышение содержания двуокиси
углерода в атмосфере увеличивает кислотность морской воды, что приводит к
декальцификации карбонатных скелетов. Смогут ли получившиеся в итоге «голые»
кораллы выжить и возобновить процесс кальцификации, если содержание двуокиси
углерода в атмосфере уменьшится? Что происходило с кораллами в геологическом
прошлом, когда они подвергались давлению отбора, действующему в ином направлении?
Примером такого рода являются склерактиниевые кораллы. Их предки были «голыми»
кораллами, похожими на актиний, которые пережили массовое вымирание пермского
периода. Первые склерактинии появились в среднем триасе; это были первые кораллы
после события массового вымирания в конце пермского периода. До недавнего
открытия, опубликованного Ярославом Столярски и его коллегами, считалось,
что склерактиниевые кораллы образовывали исключительно арагонитовые скелеты.
Но превосходно сохранившаяся окаменелость из верхнего мела обладала полностью
кальцитным скелетом. Это даёт основание полагать, что эти кораллы могли строить
скелеты из различных полиморфных модификаций карбонатов, как делали некоторые,
хотя и не все группы морских организмов, отлагающих карбонат кальция. Исследование
геохимических свойств ископаемых организмов открыло их способности изменять
систему биоминерализации, о чём даже не подозревали ранее. Что определяет
эту гибкость? Как и в случае с трилобитами, которые обсуждались выше, не все
группы были одинаково гибкими, или «пластичными» в отношении процесса индивидуального
развития. Какое воздействие оказывало это обстоятельство на картину эволюции
в прошлом,
184
и какое воздействие может оказать на будущее нынешних морских экосистем, можно узнать в ходе изучения организмов, обладающих раковинами и панцирями.
О раковинах моллюсков
Благодаря биоминерализации раковины моллюсков хорошо сохраняются в виде окаменелостей.
Тысячи их экземпляров собраны в палеонтологических коллекциях, отражая видовое
разнообразие и изобилие экземпляров и тем самым позволяя проводить всесторонние
исследования картины эволюции сквозь призму геологического времени. Чтобы
извлечь пользу из этого богатого источника информации, нам следует охарактеризовать
это разнообразие морфологически и таксономически, дав названия видам и разместив
их на правильно построенных филогенетических древах, по которым и будут исследоваться
закономерности эволюции. Проблема состоит в том, что каждая раковина обладает
лишь немногими дискретными морфологическими признаками, с помощью которых
её можно охарактеризовать, а явления конвергенции в форме и орнаментации представлены
в летописи окаменелостей в большом количестве. Сравнение отличающихся друг
от друга естественных групп моллюсков, таких, как аммониты и брюхоногие, например,
показало, что в основе морфогенеза и роста раковины лежат общие принципы генерации.
Поскольку в плане индивидуального развития раковина моллюска является высокоинтегрированным
объектом, многие особенности раковины часто изменяются в пределах вида совместно.
Этот высокий уровень интеграции делает трудным, и даже невозможным определение
свойств, которые полностью независимы от других, но это также означает, что,
работая с верно выбранной моделью, мы можем многое предсказать в эволюции
формы раковины. Это интенсивно исследовалось на примере наиболее известных
ископаемых моллюсков, аммонитов. Они образуют полностью вымершую группу головоногих
моллюсков, класса, к которому принадлежат осьминоги и кальмары. Их более отдалённым
родственником является Nautilus, также представитель головоногих моллюсков,
который фактически является единственным ныне живущим видом этой группы с
наружной раковиной.
185
Онтогенез раковины аммонитов был объектом интенсивных исследований, и было
выявлено три его основных аспекта: секреция раковины по мере того, как мягкое
тело увеличивается в размерах, секреция дополнительных слоёв на внутренней
поверхности камер тела и секреция септ в задней части тела, посредством чего
образуются камеры в раковине. Первым шагом в онтогенезе аммонитов была эмбриональная
стадия, которая обладала раковиной, называемой «аммонителла». Ископаемые остатки
её – это маленькие узелки, которые обнаруживаются в хорошо сохранившихся окаменелостях
в начале спирали, или же среди кучи неопределённых окаменелостей в углу музейного
кабинета. По мере роста раковины возрастает сложность её орнаментации – разумеется,
при наличии таковой. Орнаментация принадлежит к числу особенностей строения,
как и концентрические и продольные рёбра, шипы, бугорки и кили. Обширные отметины
на некоторых раковинах аммонитов позволяют предположить, что могла происходить
частичная резорбция раковины, поскольку они отмечают собой разрывы в линиях
роста. Орнаментация у аммонитов используется в целях таксономических исследований,
и, как было известно на протяжении долгого времени, её степень коррелирует
с другими особенностями раковины. Наиболее декорированные виды обладают наименее
плотно свёрнутой раковиной, с почти круговыми завитками. Эта корреляция стала
известна как закон Бакмана, в честь автора, который первым опубликовал сообщение
об этой идее (1892). Ойвинд Хаммер и Хьюго Бюше пришли к заключению, что картина
корреляции чётко проявляется внутри вида, но менее чётко на межвидовом уровне.
Используя компьютерное моделирование, они продемонстрировали геометрические
свойства системы, которые в итоге сводятся к связи развития рёбер и формы
устья раковины.
Достижение понимания геометрии раковины, не только у аммонитов, но и у моллюсков
вообще, заняло много времени у многих палеонтологов. В 1960-е годы, используя
в свое работе осциллограф, Дэвид Рауп стал пионером в области компьютерной
параметризации витков раковин, работы, которая заложила основу для возникновения
области теорети-
186
Рисунок 49. Компьютерные модели Хаммера и Бюше (2005) для гипотетических раковин аммонитов, иллюстрирующие корреляцию особенностей для пропорций, меняющихся на основе закона Бакмана. Вид сбоку (сверху) и со стороны устья (снизу). A: Расширенная в стороны (депрессная) раковина, у которой боковые рёбра становятся пропорционально мощнее. B: Раковина, сжатая с боков посредством уменьшения длины поперечной оси, с соответствующим образом слабее выраженными боковыми рёбрами. C: Аммонит с круглой в сечении полостью раковины и равной амплитудой боковой и вентральной ребристости. Поскольку амплитуда вентральных рёбер остаётся постоянной при изменении масштаба поперечной оси, возникает изменчивость в соотношения между амплитудами вентральных и боковых рёбер. По Hammer и Bucher, 2005, с изменениями. |
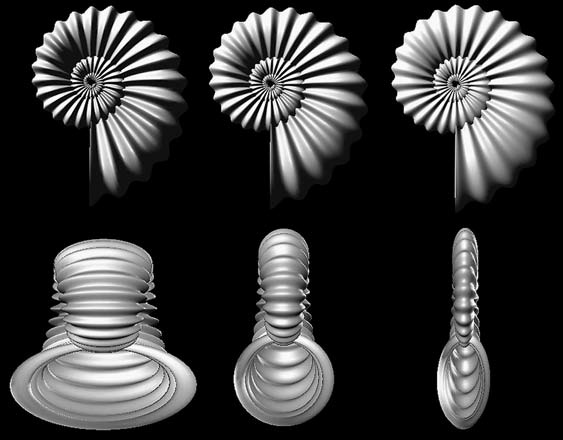
ческой морфологии. Его работа, а также другие ранние методы оперировали главным образом двумя измерениями и предполагали наличие оси вращения, которая не имеет реального биологического значения, поскольку возникает a posteriori как результат приращения раковины. Кроме того, многие раковины нельзя рассматривать как закручивающиеся вокруг единственной оси вращения. Традиционные модели не принимали во внимание онтогенетические изменения
187
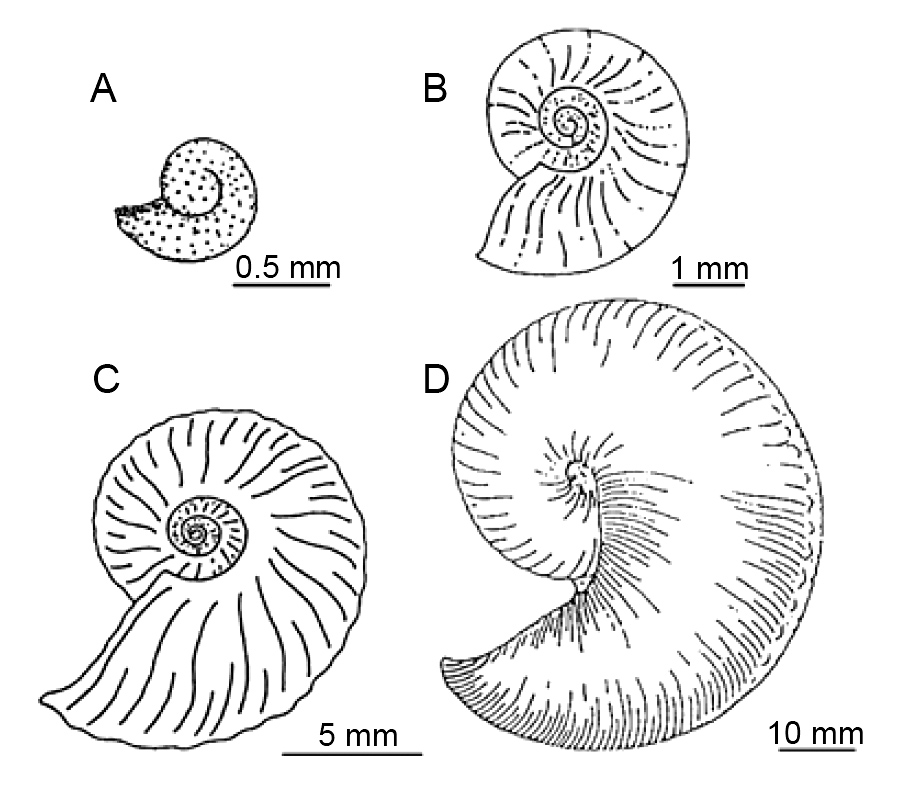
Рисунок 50. Схематические изображения
четырёх стадий онтогенеза позднемелового аммонита Hoploscpahites
nicoletti, вид сбоку. A) аммонителла; B) неаноконх, первое постэмбриональное
время; C) молодая особь; D) взрослая особь. Масштабная линейка = 5 мм.
Обратите внимание на увеличение степени орнаментации. Раковина ювенильной
особи сильнее сжата с боков и больше скручена, чем неаноконх. По Bucher
et al. 1996, с изменениями.
|