| Главная | Библиотека | Форум | Гостевая книга |
46
Глава третья
Онтогенез позвоночных
в ископаемом состоянии
Большинство ископаемых остатков позвоночных – это минерализованные части
скелета. Поскольку скелет в большинстве случаев лишь частично сформирован
у эмбрионов и на других ювенильных стадиях развития, неудивительно, что большинство
ископаемых остатков принадлежит взрослым или почти взрослым особям, которые
также крупнее, чем другие стадии жизненного цикла, и потому вероятность их
обнаружения выше. Но ископаемые остатки эмбрионов и молодых особей существуют.
В недавнем обзоре, посвящённом рептилиям, я и мой коллега Массимо Дельфино
нашли сотни научных статей, подтверждающих наличие таких образцов (www.developmental-palaeontology.net);
многие из них относятся к динозаврам. Некоторые окаменелости интерпретируются
как только что выклюнувшиеся из яйца или новорождённые особи, но, учитывая
неясности, окружающие эти интерпретации, их лучше считать особями «возраста,
близкого к времени рождения», или «перинатальными» особями.
Идентификация ископаемого образца в качестве эмбриона неоспорима, когда он
найден внутри ископаемого яйца. Более сложный случай – живородящие виды, то
есть такие, которые не откладывают яиц, а рождают живое потомство, как это
бывает у людей. Разграничить зародыша и последнюю трапезу может быть сложно,
когда имеешь дело с ископаемыми остатками
47
возрастом много миллионов лет. Зародыш – это маленький скелет «внутри» более крупного, и этот маленький скелет не несёт признаков повреждений, связанных с пищеварением. Анатомия и размер меньшего скелета, несомненно, имеют основополагающее значение для вывода о том, что он принадлежит к этому же самому виду. Противоречивые интерпретации не так уж и редки. Установление таксономической идентичности эмбрионов и ювенильных стадий – это также сложная задача. В некоторых случаях ассоциации настолько чёткие, что эмбрионы могут быть уверенно отнесены к конкретному виду. В других случаях отнесение образца к той или иной группе основано на некоторых уникальных особенностях, которые являются диагностическими для этой родословной ветви, но более точное таксономическое определение оказывается невозможным. Во многих случаях образцы, вначале описанные как представители различных видов, оказываются с большой степенью вероятности различными стадиями развития одного и того же вида. Редкие ископаемые остатки невзрослых особей – это словно моментальные снимки процесса индивидуального развития, и мы можем многое узнать о росте организма, если будем располагать их серией, или же если нам повезёт обнаружить критическую стадию, например, близкую ко времени рождения, которая сохранилась.
Ископаемые птенцы и новорождённые
При изучении истории индивидуальной жизни, относящейся ко времени рождения, перинатальный период и альтернативные стратегии развития – зрелое состояние (выводковое) против незрелого (птенцового) – имеют ключевое значение. Это состояние развития выклюнувшихся из яйца или новорождённых особей коррелирует со многими экологическими и поведенческими параметрами. В случае млекопитающих некоторые новорождённые детёныши весьма незрелые, у них нет шерсти и закрытые глаза – как у хомяков после периода беременности длиной примерно шестнадцать суток. Другие – гораздо более зрелые, рождаются с открытыми глазами, как у морской свинки после беременности, длящейся от шестидесяти до семидесяти суток. У новорождённого детёныша морской свинки и других близкородственных грызунов зубы уже частично сточены из-за движений челюстей, которые начались в
48
утробе. Аналогичные крайности в изменчивости стадий развития при появлении
на свет можно наблюдать у птиц.1
Птенцовые птицы включают пингвинов, чаек, аистов, хищных птиц, попугаев, колибри
и певчих птиц, и их птенцы в большой степени зависят от родительской заботы.
Выводковые птицы – утки, различные околоводные птицы, тетеревиные, гоацины
и индейки. Можно сравнивать разнообразные аспекты анатомии на момент их появления
на свет, например, степень дифференциации мозга, или же измерять проявления
физиологических процессов. Когда имеешь дело с ископаемыми остатками, степень
дифференциации скелета – вот ориентир для выбора. Среди ящериц только что
вылупившиеся гекконы обладают значительно менее окостеневшим скелетом, чем
большинство других видов, в том числе безногие формы и тегу.
Благодаря редким окаменелостям, сохранившим перинатальные стадии развития,
палеонтологи получили шанс узнать многое о репродуктивной стратегии и о поведении,
связанном с инкубацией яиц. В этом отношении динозавры изучались интенсивнее,
чем любая другая группа позвоночных. Один из примеров – это эмбрионы одного
из первых когда-либо описанных динозавров. Massospondylus, динозавр
из группы прозауропод из ранней юры, получил своё название в 1854 году от
Ричарда Оуэна, который ввёл в употребление термин «динозавры» (Dinosauria).
Примерно через 120 лет в Южной Африке были найдены яйца этого животного, а
через несколько десятилетий, в 2005 году, были описаны их эмбрионы, самые
старые когда-либо найденные эмбрионы динозавров. Massospondylus принадлежит
к группе, наиболее близкородственной появившимся позже гигантским динозаврам-зауроподам,
таким, как Diplodocus. Изучение характера их роста, этих самых крупных
животных среди когда-либо бродивших по земле, представляет большой интерес.
Крошечные скелеты эмбрионов Massospondylus превосходно сохранились.
Почти готовые к выклеву особи не имели зубов, что предполагает, что у них
не было возможности питаться самостоятельно. Четыре ноги этих поздних зародышей
имели равную длину, что указывает на то, они были четвероногими, хотя взрослые
особи, вероятно, были факультативно бипедальными. Череп и глаза были пропорционально
более крупными по сравнению с взрослыми особями, что является общим признаком
с другими очень
49
молодыми особями позвоночных, хотя черепа молодняка были более высокими и
узкими. Основываясь на отсутствии зубов и на других признаках анатомической
«незрелости», Роберт Рейсз и его сотрудники выдвинули предположение о том,
что потомству после появления на свет мог требоваться уход – это форма поведения,
которая также отмечена для других динозавров. Ещё больше об истории жизни
Massospondylus известно на основании многочисленных скелетов, представляющих
различные стадии роста. Оценивая изменения размеров тела и абсолютное изменение
возраста с использованием методов гистологии, мы знаем, что Massospondylus
рос со скоростью примерно 35 килограммов в год и всё ещё продолжал расти по
достижении примерно пятнадцатилетнего возраста.2
Также известны прекрасно сохранившиеся эмбрионы в яйцах нескольких других
видов динозавров, документирующие тем самым разнообразие репродуктивных стратегий.
Среди них – эмбрионы теропод-терезинозавров из верхнего мела Китая. В отличие
от прозауропода Massospondylus, молодые тероподы были явно выраженного
выводкового типа, или зрелыми на момент выклева из яйца, поскольку им было
присуще очень развитое окостенение позвоночного столба, рёбер, тазового пояса
и нижних конечностей, в которых две кости, большая и малая берцовые, были
уже явственно слитыми и окостеневшими.
Применение сканирования методом компьютерной томографии (КТ) сильно облегчает
изучение яиц, содержащих эмбрионы. Эта техника позволяет нам заглянуть внутрь
яйца, не вскрывая его, не трогая руками. Я и мои студенты используем КТ для
разного рода проектов, и это больше не редкость в палеонтологии. Окаменелость
аккуратно укладывается на платформу, которая скользит в КТ-сканер, напоминающий
гигантский пончик с узкой лежанкой. Обычно объектом исследования являются
люди, но окаменелости – более простые объекты: они не шевелятся, не страдают
клаустрофобией и не должны задерживать дыхание, пока их сканируют.
Прекрасный пример исследований с применением КТ относится к ископаемым остаткам
с острова Мадагаскар, принадлежащим недавно вымершей «слоновой
50
птице» Aepyornis, экстраординарному животному, которое достигало
высоты около 4 метров и массы тела приблизительно 400 килограммов. Её яйца
превышали 7 литров в объёме – они самые крупные среди всех ископаемых или
ныне живущих птиц, и крупнее, чем известные яйца зауропод и теропод. Остатки
этих животных, в том числе яичные скорлупки, известны из отложений возрастом
всего лишь несколько тысяч лет. «Слоновые птицы», возможно, были современниками
европейцев, поселившихся на Мадагаскаре в последние несколько веков.
У взрослых особей Aepyornis кости черепа срастаются вместе. У эмбрионов
на стадиях, которые обычно представлены в виде окаменелостей, можно гораздо
легче распознать кости, а также форму и степень их контакта. Это является
важным аспектом при сравнении черепов различных групп, ставящем своей целью
проследить морфологическую эволюцию. Наиболее заметной особенностью эмбриона
эпиорниса является толщина его костей; в сравнении с ним недавно вылупившиеся
птенцы страусов и нанду имеют грацильное телосложение.
В ходе исследований «слоновой птицы» мои коллеги сумели сложить мешанину костей
в реконструкцию эмбрионального черепа и вместе с этим решили вопросы идентификации
элементов черепа взрослой особи. Они оценили степень дифференциации скелета
у позднего эмбриона: В какой степени минерализуются костные элементы? Существовали
ли уже у эмбриона размерные пропорции и пространственные отношения костных
элементов, характерные для взрослой особи?
Живорождение у ископаемых позвоночных
У позвоночных существует широкое разнообразие репродуктивных стратегий, которые много раз появлялись параллельно в процессе эволюции. Один из примеров этого явления – плацентация, когда существует орган, соединяющий мать с развивающимся эмбрионом и облегчающий получение питательных веществ, газообмен и выведение отходов. Плацента не является исключительным признаком
51
млекопитающих; она достаточно широко распространена среди других основных
групп позвоночных, кроме птиц, черепах и крокодилов.
Независимая эволюция такой явно уникальной особенности строения, как образование
плаценты, также верна для живорождения, стратегии, при которой эмбрион развивается
внутри тела матери, а не в яйце. Яйцеживорождение означает, что яйца развиваются
и проклёвываются внутри тела матери, также получая защиту и позволяя избегать
излишних затрат на воспроизводство. Живорождение несколько раз независимо
появлялось в процессе эволюции у большинства групп позвоночных; у одних только
ящериц и змей около восьмидесяти раз.3
Мы и другие плацентарные млекопитающие – живородящие, конечно, как и все сумчатые,
группа, которая включает поссумов, кенгуру, коал и их родственников. Но утконос
и ехидна, эти ныне живущие представители когда-то гораздо более многочисленной
радиации млекопитающих, отдалённо родственных нам, откладывают яйца.
Предпосылка для развития живорождения – это механизм, который позволяет оплодотворению
произойти внутри тела, где в итоге и будет развиваться эмбрион. Было обнаружено,
что самцы некоторых плакодерм, самых первых челюстноротых рыб, живших в палеозойскую
эру, обладали придатками, похожими на совокупительные органы и напоминающими
подобные образования у современных акул; это даёт основание полагать, что
они практиковали внутреннее оплодотворение. Это не только не выглядит такой
уж большой неожиданностью, но также подкрепляет тот факт, что древнейшее свидетельство
живорождения, датированное поздним девоном, примерно 380 миллионов лет назад,
относилось к самке плакодермы, в которой сохранился плод размером в одну треть
длины взрослой особи. Важная окаменелость сохранилась настолько хорошо, что
демонстрировала то, что должно было быть пуповиной с кровеносными сосудами,
как показала компьютерная томография высокого разрешения. Эта и другие экстраординарные
окаменелости происходят из формации Гого в Западной Австралии.4
Одни из самых красивых окаменелостей, чётко документирующих наличие живорождения,
известны для разнообразной группы морских рептилий, все представители которой
были высоко специализированными к плаванию и населяли моря в
52
Рисунок 13. Рыба Incisoscutum ritchiei, принадлежащая к плакодермам, с придатками, похожими на совокупительные органы (указаны стрелкой) (Ahlberg и др. 2009). Рисунок любезно предоставил Джон А. Лонг (Лос-Анджелес). |
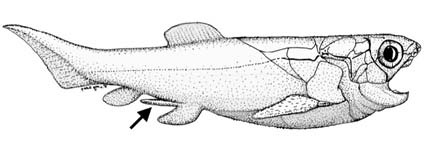
мезозойскую эру. Эволюция живорождения у этих водных рептилий была связана
с потерей способности откладывать яйца на суше и происходила параллельно значительным
морфологическим изменениям, направленным на вторичное приспособление к жизни
в воде, подобным тому, что мы наблюдаем у китов и ламантинов, чьи предки уже
были живородящими.
Ихтиозавры принадлежат к числу наиболее высокоспециализированных среди всех
групп мезозойских морских рептилий и внешне напоминали дельфинов. Мозазавриды
также были морскими рептилиями, и были, вероятно, родственны змеям.5
Эти группы вымерли в меловой период, в конце которого, примерно 65 миллионов
лет назад, также вымерли все динозавры, кроме птиц.
Живорождение, несомненно, имело место у ихтиозавров и мозазавров; у них это
непосредственно демонстрируется присутствием останков эмбрионов в пределах
брюшной полости матери. Ориентация эмбрионов у ихтиозавров и мозазавров показывает,
что они рождались хвостом вперёд, а голова появлялась самой последней, как
это происходит у ныне живущих полностью водных млекопитающих – китов и ламантинов.
Эмбрионы ихтиозавров зарегистрированы для видов, живших на протяжении всех
эпох мезозойской эры: триаса, юры и мела.
Другой разнообразной группой ископаемых рептилий, включающей преимущественно
водные формы, были зауроптеригии, к которым принадлежали наиболее замет-
53
Рисунок 14. Плиозавр Peloneustes philarchus из верхней юры Англии, демонстрирующий слабую связь между костями таза и позвоночником. Из Andrews, 1913 и Sander, 1994, с изменениями. |
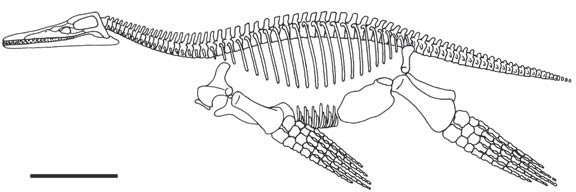
ные длинношеие и крупные плезиозавры. Свидетельства в пользу живорождения
в этой группе главным образом косвенные, основанные на морфологии их скелета.6
В частности, заслуживает подробного рассмотрения форма тазового пояса, поскольку
она даёт нам свидетельство ширины родовых путей. У зауроптеригий, так же,
как у ихтиозавров и мозазавров, отсутствует жёсткое соединение между костями
бёдер и позвоночным столбом. Наиболее вероятно, что это приспособление к плаванию,
поскольку конструкция такого плана позволяет делать быстрые остановки или
резкие повороты. Предполагается, что такого рода свободное соединение также
позволяет этим животным увеличить до максимальной степени пространство для
родовых путей.
Физиологические требования к яйцу, находящемуся внутри матки вместо того,
чтобы быть вне её, конечно, совершенно иные. Получение кислорода, необходимого
для роста, при нахождении в жидкости внутри матки и на воздухе вне её – вот
главное отличие. Может показаться, что эволюция живорождения – это трудный
или маловероятный эволюционный шаг. Но эта репродуктивная стратегия не только
много раз независимо появлялась в процессе эволюции: для неё известны также
случаи эволюционной реверсии. Молекулярная филогения удавов позволяет полагать,
что в ходе эволюции арабского песчаного удавчика Eryx jayakari происходила
эволюция от живорождения к яйцерождению.
54
У этого вида также отсутствует яйцевой зуб, который другие яйцеродящие змеи
используют, чтобы выбраться из яйца – дополнительное свидетельство того, что
способность откладывать яйца была утрачена и приобретена вновь. Ископаемые
свидетельства этого изменения ещё не известны.
Несмотря на повторяющуюся эволюцию живорождения среди ископаемых форм, большинство
из немногих групп современных морских рептилий представлено яйцекладущими
формами. Ныне живущие морские или частично морские рептилии – это морская
игуана Галапагосских островов, морские черепахи, шестьдесят два вида морских
змей и пингвины (птицы – это рептилии!). Галапагосские игуаны – это замечательные
ящерицы, которые щиплют морские водоросли на дне холодного моря, но в остальном
они преимущественно наземные и яйцекладущие. (Многие люди видели документальные
фильмы, где показаны самки морских черепах, использующие свои ласты, чтобы
выбраться на пляж, отложить там яйца, засыпать их песком и вернуться в море.)
С другой стороны, большинство морских змей – живородящие виды.
Ещё не было найдено ни одного свидетельства того, что живорождение появилось
у динозавров, и, хотя этот способ воспроизводства нельзя исключать для этой
группы, его появление у них выглядит маловероятным. Ни среди существующих
в наше время динозавров (птиц), ни в самой близкородственной динозаврам ныне
живущей группе рептилий (крокодилы) нет ни одного живородящего представителя;
нет их и в соответствующей летописи окаменелостей. Этот факт представляет
собой большую загадку для эволюционных биологов, но, похоже, это связано с
ролью яичной скорлупы в обеспечении развивающегося эмбриона важными минеральными
веществами.
Загадочное отсутствие явления живорождения среди приблизительно
9300 современных видов птиц и всех их ископаемых форм, известных к настоящему
времени, нельзя объяснить недостатком геологического времени, в течение которого
оно могло эволюционировать, поскольку долгая эволюционная история птиц восходит
к юрскому периоду, когда жили Archaeopteryx и другие специализированные
динозавры-тероподы. При объяснении этого отсутствия живорождения ссылаются
на морфологические или физиологические факторы, яв-
55
|
|
 Рисунок
15. Дышащий птичий эмбрион незадолго до выклева из яйца. Рисунок Клаудии
Джоэл.
Рисунок
15. Дышащий птичий эмбрион незадолго до выклева из яйца. Рисунок Клаудии
Джоэл.но несовместимые с воспроизводством путём живорождения. Например, птицы обладают
высокоспециализированными лёгкими с мелкими капиллярами, которым требуется
много времени, чтобы наполниться воздухом после того, как они были заполнены
амниотической жидкостью яйца в ходе индивидуального развития. По этой причине
большинство птичьих яиц имеет воздушную камеру, в которую поздний эмбрион
просовывает свой клюв, чтобы начать заполнять лёгкие воздухом. A стратегия,
основанная на живорождении, сразу поставила бы выклюнувшуюся из яйца птицу
перед внезапной необходимостью наличия воздуха в лёгких – физиологическая
проблема, явно не соответствующая анатомии выклюнувшегося птенца.
Сохранение всеми без исключения птицами яйцерождения должно отражать принципиальные
особенности птиц, препятствующие тому, что происходило так много раз у столь
многих других групп. Такие предположительные факторы включают полёт, яйцо
с изолирующей крепкой скорлупой, которая предотвращает потери воды, особенности
определения пола, иммунологические препятствия и особый характер развития
лёгких. У каждого из этих факторов есть свои «за» и «против».7
Фактор полёта в первом приближении выглядит наиболее значимым. Какой матери
хотелось бы таскать детёныша во время полёта? Но ни в одной из многочисленных
и разнообразных групп
56
нелетающих птиц никогда не эволюционировало живорождение, а беременные летучие
мыши могут летать.
Возможно, ни одна особенность птиц, взятая отдельно, не является несовместимой
с живорождением или с предшествующим ему состоянием в ходе его эволюции –
задержкой яйца в организме. Возможно, имеется целый коктейль из особенностей,
которые несовместимы с этим. Они должны включать эндотермию, инкубацию яиц
и родительскую заботу, выделение мочевой кислоты в качестве главного компонента
азотного обмена, изначально птенцовый тип потомства и известковую скорлупу
яйца. Биологи математически смоделировали затраты на задержку яйца в организме,
которые включают сниженную частоту появления потомства, повышенную материнскую
смертность и сниженный отцовский вклад в размножение, и сделали заключение
о том, что они перевешивают потенциальные выгоды для большинства птиц. Возможно,
некоторые из этих факторов также объясняют отсутствие живорождения у динозавров,
черепах и крокодилов.8
Вымершие предки птиц, как известно, откладывали яйца. Отдельные окаменелости
не только подтверждают наличие яиц, но также дают указания на наличие насиживания,
связанное с ними.
Яйца, родительская забота
и насиживание
Изучение ископаемой яичной скорлупы выявило различные стратегии в устройстве
скорлупы, призванные обеспечить безопасность, сохранение жидкости и газообмен
с окружающей средой.9
Отличительные особенности характеризуют несколько групп, и на основании гистологических
типов скорлупы сложилась номенклатурная система классификации, или паратаксономия,
яиц и яичных скорлупок. Наличие эмбрионов в ископаемых яйцах открыло путь
для хотя бы частичного соотнесения паратаксономии яиц с таксономией, основанной
на скелетных особенностях.
Соотнесение яиц с конкретным видом также можно осуществить
57
на основании ассоциации скелета и яиц, предоставляющей нам свидетельство
насиживания или разорения кладки. Классический пример путаницы между этими
двумя вариантами – случай с Oviraptor, динозавром из мелового периода
Монголии. Ископаемые остатки были впервые найдены во время знаменитой экспедиции
в пустыню Гоби, организованной Американским музеем естественной истории в
Нью-Йорке в 1920-х гг., и о которой можно узнать из книг Шона Кэрролла «Into
the Jungle» и Майка Новачека «Dinosaurs of the Flaming Cliffs». В 1923 году
был найден скелет беззубого динозавра поверх кладки разбитых яиц. В дальнейшем
Генри Фейерфилд Осборн, в то время бывший главой Музея в Нью-Йорке, присвоил
новому животному говорящее само за себя название Oviraptor philoceratops:
«ovi» означает «яйцо», «raptor» – «разбойник», а «philoceratops»
– «любящий рогатых динозавров». Находку черепа, лежащего прямо на гнезде,
интерпретировали как знак преступника, застигнутого на месте преступления
песчаной бурей и превратившегося в ископаемое как раз во время кражи яиц из
гнезда. Благодаря новым исследованиям, которые провели Марк Норрелл и его
коллеги из этого же самого учреждения в последние два десятилетия, мы теперь
знаем, что совместные находки скелетов взрослых особей Oviraptor
и кладок яиц не означают, что они грабили их; вместо этого они откладывали
их и исполняли родительские обязанности.
Один из видов родительской заботы – это высиживание яиц, при котором тело
родительской особи находится в прямом контакте с яйцами на протяжении долгих
периодов времени. Классический пример – это знакомая картина птицы-родительницы,
сидящей на яйцах: тело поверх гнезда, задние конечности подогнуты под него,
живот соприкасается с яйцами на большой площади, а передние конечности сложены
вдоль боков. Есть чёткие свидетельства того, что некоторые динозавры инкубировали
яйца подобно птицам. Когда родитель оставался при гнезде или яйцах, он, вероятнее
всего, активно защищал их от сородичей или от хищников.
Мы знаем, что Oviraptor, как и многие другие виды динозавров,
58
Рисунок 16. Сочленённый частичный посткраниальный
скелет динозавра-овирапторида из позднего мела Монголии, сохранившийся
поверх гнезда. Из Clark, Norell и Chiappe, 1999, с изменениями. |
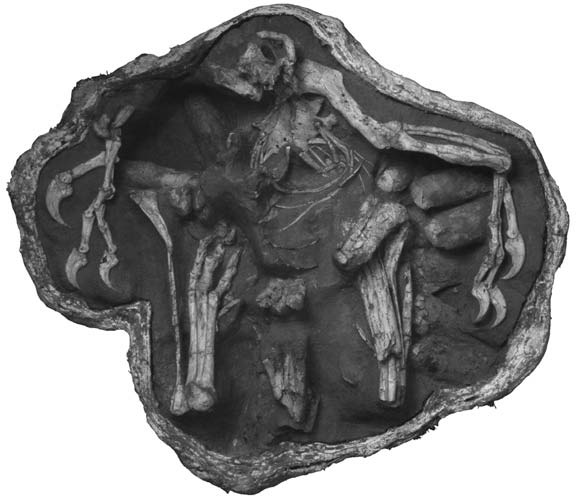
заботился о своих яйцах. Насиживание способствует инкубации, повышая температуру
яиц.
Среди ныне живущих рептилий насиживание яиц в гнезде известно только для змей
семейства ложноногих и широко распространено у питонов. Многие змеи, если
они находятся у гнезда, ищут пищу. Прямое свидетельство этого известно из
мела западной Индии, где останки почти полного скелета змеи Sanajeh indicus
были обнаружены обвившимися вокруг недавно проклюнувшегося яйца в гнезде динозавра-зауропода.
Другие особи змей, ассоциированные с кладками
59
яиц на этом же местонахождении ископаемых остатков, поддерживают догадку
о том, что происходило их кормление молодыми динозаврами. Такие ископаемые
остатки образовались при весьма особенных обстоятельствах. Очевидно, в данном
случае образование отложений и захоронение в мелкозернистом осадке были необычно
быстрыми, вероятно, ускорившись во время бури.
Существуют также другие виды инкубации, кроме яиц в гнезде. Среди позвоночных
множество разнообразных примеров демонстрируют рыбы и лягушки. Инкубация у
рыб и амфибий относится к таким случаям, когда эмбрионы удерживаются где-нибудь
на теле или внутри него, но не в яйцеводах. Существует великое разнообразие
местоположений яиц, в которых происходит их инкубация: в желудке, в выводковых
сумках на спине, в голосовых мешках, в сумках в паху, или же во рту.10
Темноспондил Trimerorhachis, обычный в пермских отложениях Техаса,
вероятнее всего, демонстрирует пример особого вида инкубации среди ископаемых
наземных позвоночных. В области глоточных мешков взрослого животного были
обнаружены остатки очень мелких особей, что даёт возможность предполагать,
что животное было занято инкубацией в тот момент, когда умерло и затем сохранилось
в ископаемом состоянии. Альтернативная интерпретация находки – это каннибализм
по отношению к собственной молоди. Но кажется более вероятным, что этот хищник
с острыми зубами и мощными челюстями питался рыбой и другими позвоночными
с помощью своей пасти, а не молодью своего собственного вида с помощью жабр.11
Ископаемые эмбрионы млекопитающих
В противоположность знаменитым случаям находок эмбрионов динозавров и других рептилий в летописи окаменелостей, ископаемые эмбрионы млекопитающих являются очень редкими, спорными, или представляют собой главным образом лишь курьёз из-за плохой сохранности. Эмбрионы редки даже в местонахождениях, известных благодаря
60
исключительной сохранности ископаемых взрослых млекопитающих. Большинство
млекопитающих обладает репродуктивной стратегией, при которой они рождают
слаборазвитых, незрелых детёнышей, что делает маловероятным обнаружение эмбрионов.
Чаще встречаются ископаемые остатки зародышей млекопитающих тех видов, которые
выработали «выводковый» тип появления потомства: более долгая беременность,
меньший размер выводка и больше масса тела взрослой особи. Вероятно, мы никогда
не узнаем продолжительность беременности у ископаемых видов, но во многих
случаях мы сможем узнать размер выводка. Лошадиные эмбрионы из эоценовых местонахождений
Германии показывают, что самки вынашивали только один плод, показывая, что
ранние лошади в процессе эволюции выработали стратегию рождения немногочисленного
потомства, вероятно, сочетающегося с интенсивной родительской заботой. Это
типичная стратегия для стадных копытных, и она соотносится с гипотезой о том,
что эоценовые лошади жили стадами, основанной на высокой частоте встречаемости
ископаемых остатков в некоторых местонахождениях. В одном случае образование
из мягких тканей, окружающее зародыш, было с уверенностью интерпретировано
как плацента.12
Небольшой размер выводка также был зарегистрирован для других хорошо сохранившихся
ископаемых млекопитающих. Скелет ореодонта Merycoidodon culbertsoni из
олигоцена Южной Дакоты, вымершего родственника парнокопытных (Cetartiodactyla),
демонстрирует наличие в тазовой области двух хрупких, но довольно полных черепов,
которые были описаны как «близнецы» в матке.
Более молодые ископаемые эмбрионы млекопитающих известны для некоторых вымерших
гигантских наземных ленивцев из плейстоцена Южной Америки, и для мамонтов
из Сибири. Их не полностью сформированные кости демонстрируют анатомические
детали, которые решают вопросы, связанные с идентификацией костей, которые
во взрослом состоянии слишком изменились по сравнению с костями других видов.
В некоторых случаях черепа взрослых особей из-за слияния отдельных элементов
или изменения пропорций несут неожиданные особенности, если их сравнивать
с другими родственными формами, или у них в анатомии отсутствует ожидаемый
элемент. Например,
61
на первый взгляд, на черепе взрослого человека отсутствует предчелюстная кость. Но, как обнаружил в девятнадцатом веке Гёте, исчезновение шва – вот причина, которая объясняет кажущееся отсутствие кости, которая фактически присутствует в нашем черепе.
Эмбрион или последняя трапеза?
Редкая возможность взглянуть на шаги эволюции истории жизни у весьма особенной
разновидности млекопитающих стала реальностью благодаря открытию Maiacetus
inuus, раннего кита из Пакистана возрастом 47,5 миллионов лет. Был найден
ископаемый образец с меньшей особью внутри его полости тела. Авторы исследования
интерпретировали меньший экземпляр как расположенный в положении in vivo внутри
матки взрослой беременной самки, собирающейся рожать. Положение головой вперёд
при родах у предполагаемого зародыша похоже на положение зародыша у наземных
млекопитающих и показывает, что эти животные, в отличие от современных китов,
рождали потомство на суше. Хорошо развитый набор зубов у зародыша был истолкован
как признак относительной зрелости при рождении, предполагающий, что новорождённые
особи у Maiacetus были, вероятно, активными сразу после рождения.
Как и у других предковых китообразных, у Maiacetus было четыре ноги,
видоизменённых для гребущего плавания, но способных, вероятно, выдерживать
его вес на суше, с похожими на плавники лапами.
Но некоторые коллеги с богатым опытом в области анатомии китообразных привели
доводы против истолкования находки как плода Maiacetus: положение
меньшей особи относительно позвоночного столба старшей особи слишком близко
голове предполагаемой матери, на уровне желудка, и отсутствие позвонков её
хвоста больше соответствует интерпретации меньшей особи в качестве добычи.
Но, каким бы ни был результат этой дискуссии, этот пример показывает важность
исследования судьбы трупов умерших животных, когда они разлагаются и начинают
процесс сохранения
62
в ископаемом состоянии. Наука, посвящённая такого рода исследованиям, тафономия, становится значимой при исследовании ископаемых эмбрионов и других ключевых окаменелостей, сохраняющих мягкие ткани.
Сгнившие и окаменевшие эмбрионы
Благодаря исследованиям в области тафономии мы достигли понимания того, как
лучше интерпретировать анатомические детали важных окаменелостей. Исследуя
характер посмертного разложения, мы знаем, как анатомические составляющие,
отличающиеся друг от друга по своим химическим свойствам, теряются на разных
стадиях разложения. Также могут возникать топографические изменения структур
организма, которые ещё более затрудняют интерпретацию на первый взгляд полных
окаменелостей.
Примечательный пример такого рода исследований касался темпа и последовательности
разложения отдельных деталей строения у видов базальных позвоночных. Задачей
Роберта Сансома, Марка Пурнелла и их коллег в Лестере была интерпретация анатомии
предположительных ранних ископаемых хордовых путём исследования их ныне живущих
аналогов после смерти. В процессе разложения детали анатомии, образованные
мягкими тканями, имеют тенденцию к разрушению, но не все особенности исчезают
в одинаковом темпе. Хотя эта работа не касалась эмбрионов или особенностей
онтогенеза, она упоминается здесь, поскольку иллюстрирует тафономический подход,
а также имеет отношение к приобретению первыми, вымершими представителями
родословной ветви особенностей строения, чем-то способствующих пониманию происхождения
систем развития, которые обеспечивают появление этих особенностей. Сансон
и его коллеги изучали разложение только что умерших особей ланцетника Branchiostoma
lanceolatum и миноги Lampetra fluviatilis. Среди деталей, важных
для изучения у ископаемых остатков, были глаза, хвост и предшественник позвоночника,
известный как нотохорд. Органы были помещены для гниения внутрь прозрачных
пластиковых коробок, заполненных солёной водой
63
и инкубируемых при 25 градусах Цельсия сроком до двухсот дней.
Исследователи зафиксировали процесс распада для каждой анатомической детали
и подсчитали изменения в соответствии с простой системой оценки. Деталями
строения, более стойкими к разложению были те, которые являлись общими особенностями
всех позвоночных, например, нотохорд. Напротив, детали строения, эволюционировавшие
позднее, сгнили первыми. Это прискорбно, поскольку позже возникшие детали
строения, которые различают близкородственных животных внутри родословной
ветви, являются наиболее информативными для целей реконструкции эволюционного
древа.
Исследование показало, что, будь окаменелость остатками примитивного хордового
или разложившегося позвоночного, по отношению к ней требуются тщательные анатомические
и химические исследования. Некоторые из кембрийских ископаемых форм, которых
касается это тафономическое исследование, при более тщательном изучении могут
оказаться более близкими родственниками позвоночным, чем считалось до этого.
Вероятно, мы этого никогда не сможем узнать, если принимать во внимание превратности
отклонений тафономического характера. Отложения во всём мире полны окаменелостей,
которые ещё ждут того момента, когда их обнаружат, равно как и сотни кабинетов
в музеях, и эти окаменелости не представляют собой такой сложности для интерпретации,
как эти кембрийские формы.
Другой пример тафономических исследований, на сей раз более подходящий к теме
этой книги, касается процесса разложения и тафономии крошечных эмбрионов.
Эти исследования были инициированы открытием в последние двадцать лет кембрийских
эмбрионов многоклеточных животных из Китая. Эти окаменелости по расположению
клеток и форме похожи на эмбрионов современных морских животных, но они очень
мелких размеров и сложны для изучения. Поэтому даже их органическая природа
может оказаться под вопросом и в некоторых случаях действительно подвергалась
серьёзным сомнениям.
Наиболее информативные исследования стадий развития кембрийских эмбрионов
от дробления до стадий, непосредственно предшествующих выклеву, включали получение
изображений с помощью синхротрона, крупное
64
технологический достижение, которое позволяет проводить исследование крохотных
окаменелостей на клеточном уровне.13
Порода подвергается химическому воздействию в целях растворения карбоната
кальция, окружающего крошечные окаменелости, состоящие из фосфата кальция.
Поскольку каждая из этих окаменелостей составляет менее полумиллиметра в длину,
это является трудоёмкой задачей. Если принимать во внимание эти ограничения
и тот факт, что для поиска таких крохотных, клеточных окаменелостей было сделано
ещё очень мало, таксономический и филогенетический охват ископаемых эмбрионов
очень ограничен. Фактически, в большинстве случаев вид животного, представленного
различными сохранившимися стадиями, остаётся нерешённым вопросом. Некоторые
тщательные исследования раскрыли характер дробления у некоторых эмбрионов
– это картина деления клеток, ведущего к образованию скопления клеток на первых
стадиях развития. Но некоторые из особенностей трудны для распознания с точки
зрения как биологии, так и геологии. Тафономические исследования предоставили
нам свидетельства того, какие эмбриологические стадии могут сохраниться и
как долго и в каких условиях жили эти, главным образом кембрийские, организмы.
Пионерными в этой области были исследования Элизабет Рафф и её коллег, которые
провели эксперименты с эмбрионами и личинками морских ежей, группы организмов,
предпочитаемых у специалистов в области биологии развития из-за того, что
о ранних стадиях их онтогенеза известно многое. Рафф и коллеги обнаружили,
что сохранение эмбрионов в основном не связано с размерами, и что минерализация
тонких особенностей строения, которые могли бы затем быть изученными, была
осуществима во многих обстоятельствах. Одни химические условия больше способствовали
точному сохранению клеточной анатомии, чем другие. Некоторые стадии обладали
незначительным потенциалом для сохранения. Подобные эксперименты в настоящее
время проведены с различными видами, но ещё больше должно быть сделано, прежде
чем будут разработаны точные или общие протоколы для интерпретации крошечных,
сложных органических структур из отдалённого прошлого.
При рассмотрении разнообразия в области индивидуального развития и таксономии,
65
которое должно было существовать, и превратностей геологического сохранения остатков применение универсального метода может быть очень сложным. Но, по крайней мере, экспериментальный подход с использованием ныне живущих видов скажет нам и тем самым позволит нам более критически относиться к тому, чего мы можем достигнуть, обладая новыми инструментами отображения и лучшим пониманием процесса фоссилизации.
Содержание
Благодарности vii
Пролог xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез
позвоночных в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания 197
Библиография 213