141
Глава восьмая
«Недостающие звенья» и
эволюция индивидуального развития
Многие люди привыкли думать об эволюции жизни как о прогрессии, похожей на
лестницу, где каждое животное стоит на своей ступени. В случае эволюции позвоночных
они могут вообразить рыбу на нижней ступени, саламандру на следующей, затем
ящерицу, мышь, и, наконец, человека на верхней ступени. Следуя этому средневековому
мифу, казалось вполне естественным предположить, что «низшие» животные эволюционируют
в «высших» животных. И, если это так и было, мы должны рассчитывать увидеть
«связующие звенья» между ними на всём пути вверх и вниз по «лестнице». Эта
идея Великой Цепи Бытия, или Scala naturae, представляет собой образ эволюции,
который всё ещё обычен в ненаучной среде, хотя учёные уже давно поняли, что
такая концепция является неточной. Миф о «лестнице» связан с неверным пониманием
эволюции и не соответствует реальному процессу. Вместо того, чтобы быть похожей
на лестницу, эволюция жизни больше напоминает ветвящийся кустарник. Каждая
ветка кустарника представляет собой самостоятельную родословную ветвь живых
организмов. Места, где из одной точки на кустарнике вырастают две или больше
ветвей, указывают на то, что родословные линии, представленные ветвями, в
какой-то момент своей истории должны были иметь общего предка. Каждый из ныне
живу-
142
щих видов является «недостающим звеном», поскольку каждый вид является частью
древа жизни и обладает мозаикой как «примитивных» пережитков прошлого, так
и продвинутых особенностей, которые своим наличием подтверждают изменения.
От каждого вида можно многое узнать об этих изменениях.
Каждый из миллионов видов животных имеет общих предков с любым другим видом.
«Недостающее звено» – это предковый вид, который дал начало двум современным
группам, например, людям с одной стороны и шимпанзе с другой. Но было бы не
разумно ожидать обнаружить именно тот критически важный вид; для его идентификации
потребовался бы полный ряд ископаемых предков-потомков. Конечно, мы можем
проследить эволюцию птиц из крылатых динозавров, но мы точно не уверены в
том, какой именно ископаемый вид был прямым предком современных птиц. Наши
усилия в деле реконструкции истории жизни не должны быть направлены на попытки
вообразить себе, как шимпанзе мог превратиться в человека, поскольку этого,
конечно, никогда не случалось. Мы скорее попытаемся установить, какие признаки
шимпанзе и люди унаследовали от их общего предка, и какие признаки появились
в их родословных линиях в процессе эволюции уже после того, как они разошлись.
Некоторые авторы правильно говорят, что не только концепция «недостающего
звена» архаична, но также его поиск – это устаревший подход к исследованию
макроэволюции.
На нынешней стадии развития эволюционной биологии фокус в исследованиях переместился
с обнаружения переходных таксонов на обнаружение переходных особенностей,
которые являются общими для близкородственных форм с общим происхождением.
В наше время хорошо известно много примеров такого рода, и они снабжают нас
информацией не только об анатомических преобразованиях, но также и о функциональном
и экологическом контекстах этих переходов. В этом контексте особенно важны
стволовые* виды, те, которые свидетельствуют о времени и способе приобретения
диагностических признаков ныне живущих видов в ходе эволюции.1
Пример такого рода даёт нам сама наша родословная ветвь млекопитающих. Согласно
молекулярным оцен-
* В оригинале сказано «stem species», то есть, виды, находящиеся
на «стволе» родословного древа группы. – прим. перев.
143
кам, подтверждаемым свидетельствами со стороны ископаемых остатков, последний
общий предок всех ныне живущих млекопитающих, то есть последний общий предок
человека (плацентарного млекопитающего), кенгуру (сумчатого) и утконоса (однопроходного)
жил, по крайней мере, 220 миллионов лет назад. Но история происхождения млекопитающих
от нашего последнего общего с рептилиями предка уходит в глубины времени ещё
дальше, до отметки, по меньшей мере, в 315 миллионов лет назад. Эти самые
ранние 100 миллионов лет эволюционной истории подтверждаются стволовыми видами
млекопитающих. Они стоят на эволюционном древе между последним общим предком
ныне живущих видов и последним общим предком с ближайшими ныне живущими родственниками,
в данном случае с рептилиями.
Для нескольких крупных таксонов ископаемые остатки предоставляют довольно
полную картину морфологических преобразований, связанных с их происхождением.
Летопись окаменелостей многих групп изменилась с почти отсутствующей на довольно
разнообразную в течение лишь немногих десятилетий, как в случае с мезозойскими
млекопитающими.
Непрерывные сообщения о новых открытиях «переходных форм» показывают большой
шаг вперёд, который был сделан и делается сейчас в области документации макроэволюционных
шагов позвоночных со времён Дарвина. Эти сообщения также показывают, что для
органической эволюции не характерны цель и план. Организмы выполняют свои
«условия для поддержания жизни» в каждый конкретный момент времени, в котором
они живут – и важно именно это, о чём я и буду говорить далее.
Глаза камбалы
Среди огромного множества современных рыб, примерно тридцати тысяч видов,
существует много форм с признаками анатомической специализации, происхождение
которых в большинстве случаев не зарегистрировано в ископаемых остатках. Классическим
примером была группа камбал, к которой принадлежат знакомые нам
144
с гастрономической (жаль, что в основном так) палтус и камбала, необычные
тем, что оба их глаза расположены на одной стороне головы. Мэтт Фридман из
Оксфордского Университета описал ископаемые остатки из эоценовых пород Италии
и Франции возрастом около 50 миллионов лет, которые показывают расположение
глаз, промежуточное между таковым у нормальных рыб и камбал: один глаз смещён
к противоположной стороне черепа, но сделал это не совсем в полной мере. Открытие
предковых камбал с неполным смещением глазниц демонстрирует нам то, что происхождение
плана строения тела камбалы возникло постепенным, пошаговым путём. Это эволюционное
происхождение напоминает индивидуальное развитие асимметрии у камбалы, когда
увеличение степени смещения глазницы преобразует симметричного эмбриона в
полностью асимметричную взрослую особь. «Промежуточные» окаменелости представляли
собой взрослых особей, а не молодь, поэтому, фактически, не было зарегистрировано
никакого «онтогенеза в ископаемом виде». Но отдельные окаменелости позволяют
сделать красивое сравнение с данными об индивидуальном развитии, полученными
для этих животных. Как эти промежуточные формы использовали наполовину смещённые
глаза, остаётся загадкой.
Случай с камбалами имеет отношение к одной из наиболее фундаментальных дискуссий
в эволюционной биологии, корни которой уходят в такие влиятельные и спорные
идеи, как «перспективный монстр». Этот термин был введён Ричардом Гольдшмидтом
(1878 – 1958), который наряду с Гансом Шпеманом и Виктором Гамбургером сделал
Германию выдающейся страной в области биологии развития. Он стал профессором
зоологии в Калифорнийском Университете в Беркли в 1936 году. Гольдшмидт особо
отметил отсутствие «промежуточных форм» в эволюции и предложил в качестве
примера асимметричный череп камбалы. Идеи Гольдшмидта были в значительной
степени упрощёнными и подвергались жёсткой критике со стороны людей, приводивших
доводы в пользу градуалистической эволюции. Он стал в некотором роде карикатурным
образом намертво застрявшего в ошибках учёного и остаётся изображённым так
в некоторых упрощённых обзорах дебатов в области эволюционной биологии.
Фактически, Гольдшмидт подчеркнул роль индивидуального развития
145
|
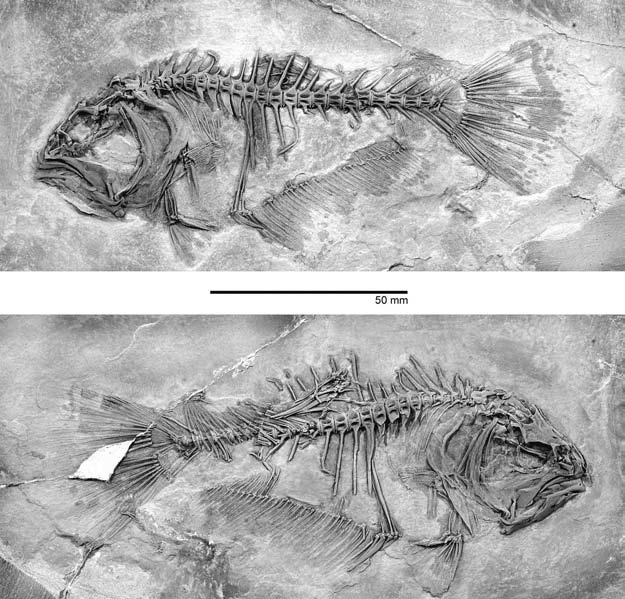
Рисунок 38. Heteronectes chaneti
из нижнего эоцена Монте-Болька, Италия. На одной стороне (внизу)
глазница находится в своём обычном положении, но на другой она расположена
намного дорсальнее (сверху), вблизи средней линии головы. Фотографию
любезно предоставил Мэтт Фридман (Оксфорд). |
в происхождении его «перспективных монстров», утверждая, что они не просто
появлялись из небытия как результат генетической мутации. Его акцент был сделан
не просто на генах, а на эпигенетических системах, возникающих в результате
комбинации генетических факторов и факторов окружающей среды. Из злодея, каким
он был, Гольдшмидт теперь стал чем-то вроде героя.2
Ископаемые остатки камбал, казалось бы, являются доводом против идеи Гольд-

Рисунок 39. Перемещение глаза в ходе индивидуального
развития камбалы.
147
шмидта. Они, конечно, противоречат его предсказанию, касающемуся этих рыб,
и относятся к обсуждаемой проблеме характера и скорости эволюции. Но идеи
Гольдшмидта, однако, важны, и в иных случаях эмпирические свидетельства поддерживают
их. Кроме того, хотя мы пытаемся вывести общие принципы, каждая отдельно взятая
эволюционная история уникальна, поэтому в обобщениях стоит проявлять осторожность.
Этот пример с камбалой, с переходными формами, о которых бы Гольдшмидт и не
подумал, что они могли существовать, не является залогом верности градуализма;
это лишь пример того, как летопись окаменелостей в некоторых случаях являются
единственным способом проверить эволюционную гипотезу непосредственно. Случай
с летучей мышью является примером противоположного рода – это случай, когда
именно молчание летописи окаменелостей может показать истинную картину эволюции.
Крылья летучей мыши
Многое в анатомии летучих мышей является специфичным – например, специализация
ушей у многих видов, которая позволяет им использовать эхолокацию, способность
слышать звуки очень высокой частоты, чтобы определять расположение добычи,
врагов и объектов в пространстве. Ископаемые остатки из эоцена Северной Америки
и Европы показывают, что эхолокация появилась на ранних этапах эволюции летучих
мышей, хотя уже после полёта. Специализации к слуху в области высокочастотных
звуков у летучих мышей, вероятно, возникали в процессе эволюции более одного
раза в пределах этой группы. Своего рода эхолокацию также используют землеройки,
а иным способом* и в другой среде – некоторые киты. Но действительно исключительный
аспект летучих мышей – это их крылья, которые позволяют им использовать активный
полёт – признак, который среди прочих позвоночных появился в ходе эволюции
только у птиц и (до позднего мела) у вымерших птеродактилей. Крыло летучих
мышей уникально тем, что оно состоит главным образом из очень удлинённых пальцев,
кроме большого пальца, и подвижной кожи между ними. Как могли бы выглядеть
передние
* Китообразные воспринимают звук с помощью нижней челюсти,
где имеется специальный звукопроводящий канал. – прим. перев.
148
конечности «недостающего звена»? Среди ныне живущих видов много разных млекопитающих
способно к планирующему полёту и для этой цели у них имеется кожа между конечностями.
Планирующий полёт выглядит первым шагом к полёту, включающим движение по воздуху
с использованием складки кожи. Но различия всё же огромны. Способность постоянно
махать передними конечностями с очень большими пальцами – это нечто совсем
другое.
Исчерпывающие молекулярные исследования ныне живущих млекопитающих ясно указывают
на то, что летучие мыши являются самыми близкими родственниками копытных,
вроде коров и лошадей, и вместе с ними образуют одну группу с хищными млекопитающими,
такими, как собаки и кошки*. Вопрос о родственных связях летучих мышей с современными
млекопитающими, таким образом, решён, но вопрос о происхождении уникальнейшего
морфологического новшества летучих мышей, активного полёта, всё ещё остаётся
без ответа.
Приблизительно через десять миллионов лет после того, как вымерли динозавры,
не относящиеся к птицам, адаптивная радиация существующих в наше время групп
млекопитающих становится хорошо документированной ископаемыми остатками. Многие
эоценовые окаменелости замечательно полны и вне всяких сомнений классифицируются
среди самых ранних летучих мышей. У них уже имеются крылья, и, несмотря на
крупномасштабные поиски в богатых отложениях хорошо сохранившихся окаменелостей,
неизвестно никаких промежуточных звеньев. Возможно, это эволюционное новшество
возникло очень быстро. Биология развития даёт подсказки в отношении механизмов,
которые, вероятно, сделали это возможным.
Передние конечности, на раннем этапе развития почки конечности относительно
более крупные по сравнению с задними – это довольно обычная картина для млекопитающих.
Так происходит в период индивидуального развития, который некоторые коллеги
называют филотипическим и который характеризуется некоторой морфологической
и, возможно, генетической общностью для различных таксонов. От этой в чём-то
сходной стартовой точки пути развития летучих мышей и других млекопитающих
начинают расходиться. Это было детально документировано для некоторых видов
летучих мышей и для обычных мышей. У летучих мышей увеличение размеров пальцев
на передней конечности происходит быстрее, чем пальцев на задней конечности
– это пример аллометрического роста для конечностей. У обычных мышей и
* Их объединяют в одну группу без определённого систематического
ранга под названием Pegasoferae – прим. перев.
149
передние, и задние лапы растут со сходной скоростью. Удлинение пальцев летучей
мыши в ходе развития достигается благодаря ускоренному темпу разрастания и
дифференциации клеток хряща. Карен Сирс и её коллеги обнаружили, что этот
процесс связан с более высоким содержанием костного морфогенетического белка
(bone morphogenetic protein) Bmp2. Другая работа сравнительного характера
показала также, какие молекулы задействованы в сохранении кожи между пальцами
в ходе индивидуального развития, что приводит к образованию крыла у взрослой
особи. На стадии раннего эмбриона у нас есть кожа между пальцами, как у всех
позвоночных с руками и ногами. В некоторый момент времени клетки, образующие
эту кожу, начинают отмирать, и поэтому у нас есть наши подвижные пальцы, а
не весло или крыло, как лапы уток или крылья летучих мышей. На лапах утки
и в передней конечности летучей мыши кожа не отмирает, а сохраняется и даже
разрастается. Механизмы, задействованные в этих двух случаях, аналогичны,
но, как открыли Скоттом Ветерби и его коллеги, участвующие в них молекулы
не одинаковы. Было известно, что у цыплят и мышей, модельных организмов для
исследований в области генетики развития, костные морфогенетические белки
(Bmp), которые способны стимулировать рост и дифференциацию клеток,
запускают процесс гибели клеток в недифференцированной, свободной соединительной
ткани (мезенхиме) между развивающимися пальцами «весла» передней и задней
конечностей, характерного для эмбрионального состояния. В ноге утки молекула
Gremlin служит ингибитором этого действия Bmp, и её работа
приводит к образованию перепончатой лапы. Летучая мышь также обладает этим
механизмом, но в нём дополнительно участвует другой белок, а именно, фактор
роста фибробластов Fgf8, также играющий роль в различных аспектах
морфогенеза.
Важность этих открытий состоит в том, что они предоставляют нам эмпирическое
свидетельство существования простого механизма, задействованного в процессе
образования крыльев – новшества, возникающего в ходе индивидуального развития.
Теперь «внезапное» возникновение летучих мышей выглядит менее маловероятным,
и свидетельства со стороны окаменелостей и их отсутствие могут указывать именно
на такую ситуацию. Существуют ли какие-то ископаемые эмбрионы летучих мышей,
которые предоставляют какое-либо дополнительное свидетельство? Единственный,
насколько я знаю, происходит
150
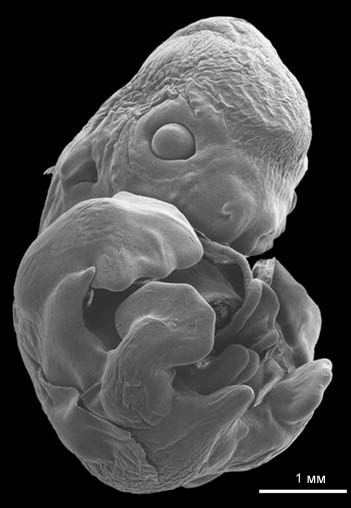
Рисунок 40. Эмбрион крылана Rousettus
amplexicaudatus, изображение получено с помощью сканирующего электронного
микроскопа. Фотография М. Санчеса, из Giannini, Goswami и Sánchez-Villagra,
2006, с изменениями. |
из эоценового местонахождения Мессель в Германии, известного, помимо всего
прочего, одними из самых ранних и наилучшим образом сохранившимися ископаемыми
остатками летучих мышей. Скелет ископаемого эмбриона уже сформирован, а крылья
более или менее видны.
Панцирь черепах
Другой специфичный план строения тела среди позвоночных, на сей раз среди
рептилий, – это план строения тела черепах. Главные признаки специализации
скелета –
151
это наличие панциря, отсутствие зубов, короткий хвост и анапсидный тип строения
черепа. Черепа у черепах лишены окон для прикрепления мускулов, типичных для
других наземных позвоночных. Это состояние может являться как вторичным, так
и первичным; мы не можем сказать об этом главным образом из-за того, что мы
не уверены в том, где размещать черепах на древе жизни рептилий.3
Если они находятся в основании древа, то их черепа, вероятнее всего, являются
просто результатом сохранения примитивного состояния, поскольку самые ранние
рептилии, хорошо представленные в летописи окаменелостей, являются анапсидами.
Если же черепахи являются сестринской группой по отношению или к ящерицам,
или к крокодилам, то тогда их черепа в значительной степени специализированы.
Пока известно недостаточно ископаемых остатков для решения этого вопроса,
даже несмотря на некоторые великолепные образцы, которые были описаны недавно.
Возможно, помогут будущие находки окаменелостей.
В 2008 году Ли и коллеги опубликовали описание нового рода и вида Odontochelys
semitestacea из отложений Китая возрастом около 220 миллионов лет. Это
животное имело анапсидный череп, но в остальном оно демонстрировало много
переходных особенностей, вроде наличия зубов и длинного хвоста.4
Но, что ещё важнее, пример Odontochelys касается того, что уникально
для черепах, их панциря.
Черепаший панцирь состоит из дорсальной, или спинной части, вентральной, или
брюшной части, и костных мостиков, соединяющих их*. Вентральная часть состоит
из расширенных брюшных рёбер, слившихся с некоторыми костями исходного для
позвоночных плечевого пояса, такими, как ключицы, которые теперь сильно видоизменены.
Спинная часть панциря намного сложнее по своему происхождению, поскольку она
является результатом слияния рёбер с кожными окостенениями, которые аналогичны
чешуям или остеодермам многих рептилий. Наиболее примечательно то, что у черепах
лопатка находится под спинным панцирем, или карапаксом; это уникальное состояние:
у всех прочих рептилий и у млекопитающих, например, как у людей, лопатка находится
снаружи грудной клетки. Наличие панциря и его анатомические связи у черепах
настолько уникальны, что для объяснения его происхождения традиционно обращались
к идее скачкообразных эволюционных изменений.
* Спинной щит черепахи в отечественной литературе называется
«карапакс», а брюшной – «пластрон». – прим. перев.
152
Желая добиться понимания развития такой странной анатомии, Хироши Нагашима
и его коллеги изучили, как формируется панцирь по отношению к лопатке и к
мускулатуре тела. Их анатомическое исследование показало, что новизна в строении
панциря черепахи – это то, что мои коллеги назвали осевой задержкой роста
ребра, и складывание мускулов для подгонки под необычное положение плечевого
пояса. Исследование индивидуального развития проводилось не с наивным ожиданием
проявления рекапитуляции, а скорее с целью понимания топографического положения
мускулов у современных черепах и их развития. Их работа дала ключи для формулировки
вероятной гипотезы морфологического преобразования, которую можно проверить
на окаменелостях.
Odontochelys, самая древняя ископаемая черепаха, является «недостающим
звеном» для понимания происхождения панциря, поскольку у неё имеется пол-панциря
– хорошо различимая нижняя половина. На спинной стороне широкие и короткие
рёбра Odontochelys напоминают рёбра позднего эмбриона ныне живущих
черепах. Фактически, анатомия Odontochelys очень хорошо соответствует
модели, которую представили Нагашима и его коллеги: рёбра укорочены настолько,
что без панциря лопатка расположена не выше и не в дорсальном положении по
отношению к рёбрам, но вместо этого находится перед первой парой рёбер.
Пример с черепахами иллюстрирует три фундаментальных положения:
-
Ископаемые остатки являются важнейшим свидетельством,
показывающим, в каком порядке возникали новшества процесса развития: брюшной
панцирь раньше спинного панциря.
-
Вместе с данными эмбрионального развития ископаемые
остатки представляют нам хорошо обоснованные сценарии процесса возникновения
эволюционных новшеств.
-
Ископаемые остатки могут быть очень информативными в
отношении некоторых особенностей и ничего не говорить о других. В случае
с черепахами мы многое узнали о панцире, но гораздо меньше о черепе.
153
|
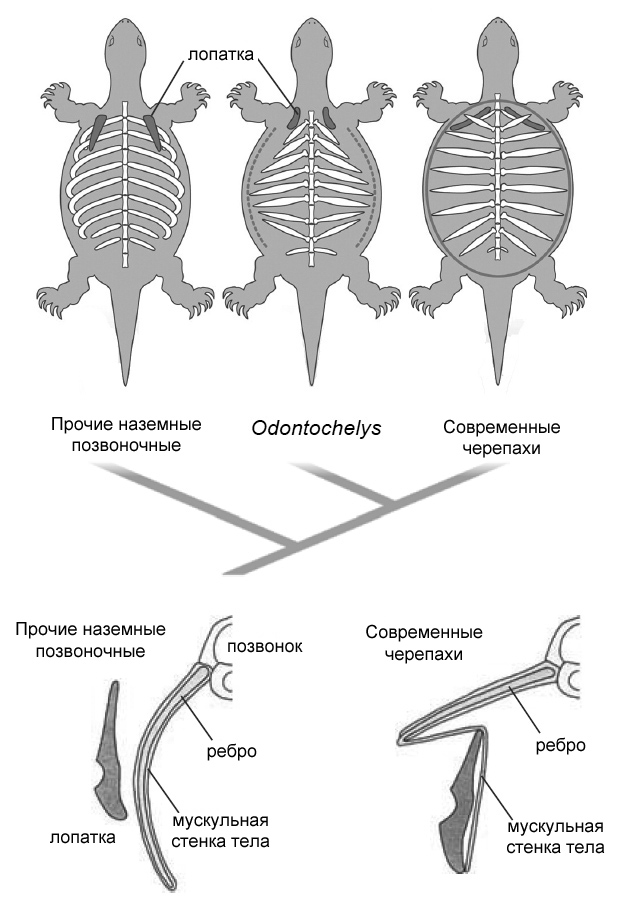
Рисунок 41. Карапакс черепах состоит
из расширенных рёбер и позвоночника, слившихся со скелетными тканями,
происходящими из кожи. План строения тела очень отличается от других
наземных позвоночных, поскольку у черепах лопатка расположена внутри
грудной клетки. Хироши Нагашима и его коллеги предполагают, что у Odontochelys
semitestacea из позднего триаса Гуйчжоу (Китай) эмбриональный «гребень
карапакса» (пунктирная линия), возможно, развивался у эмбриона лишь
временно и не полностью, стимулируя частичное веерообразное разрастание
рёбер. Уникальное расположение лопатки в ходе индивидуального развитии
черепахи приводит к образованию складки на мускульной части стенки тела
в области плеча. |
154
Другой аспект, который следует рассмотреть на примере черепахи и сальтационного
сценария происхождения черепашьего плана строения тела – это объяснение одновременного
«скачка», который приводит к появлению в значительной степени интегрированного
набора особенностей, с которыми связан панцирь; среди них мускулатура и её
управление с помощью нервов, анатомия конечностей и механизмы вентиляции лёгких.
Здесь более вероятным был бы такой сценарий, который делает возможным относительно
быстрое изменение, но также и учитывает потенциальную корреляцию между различными
особенностями строения черепахи.
Ноги китов
До 1980-х годов происхождение китов было изучено плохо. На основе лишь одних
анатомических исследований было трудно разобраться в том, какая группа наземных
млекопитающих была самой близкой к ним. Ископаемые киты были известны с девятнадцатого
века, но это не помогало разрешить эту загадку, поскольку они уже были животными,
очень специализированными в плане физиологических требований к жизни в воде,
и обладали плавниками и хвостовыми лопастями.
После этого времени серия замечательных открытий окаменелостей, сделанных
международной командой Фила Джинджерича и Ханса Тевиссена, дала нам довольно
полный ряд окаменелостей, которые демонстрируют преобразования и документируют
шаги, ведущие от обладающих нормально развитыми ногами наземных копытных к
млекопитающему без задних конечностей, имеющему лишь остатки тазового пояса.
Большинство этих окаменелостей происходит из эоцена Индийского субконтинента
и из Египта. Параллельно палеонтологическим открытиям молекулярные исследования
чётко определили место китов как самых близких родственников бегемота, и это
означает, что китов следует классифицировать в рамках группы парнокопытных
(Artiodactyla), к которой принадлежат коровы, жирафы, олени и свиньи. Летопись
окаменелостей показывает, что адаптация китов и бегемотов к водному образу
жизни возникла в процессе эволюции независимо.
155
Пакицетиды – это наиболее примитивная группа ископаемых китов. Анатомия их
уха занимает промежуточное положение между ушами наземных и полностью водных
млекопитающих, что означает, что оно, вероятно, могло функционировать в обеих
средах. Пакицетиды были способны бегать по суше, что отражено в анатомии скелета
их конечностей и позвоночника.5
Данные о химических анализах соотношения изотопов кислорода и исследования
отложений, в которых были обнаружены пакицетиды, дают основание полагать,
что они жили в реках или вблизи них, а не в морских местообитаниях. Следующее
важное ископаемое животное в цепочке – это Ambulocetus, который вёл
земноводный образ жизни. У него были длинные шея и хвост, а крупные ступни
задних лап и другие особенности подсказывают, что это животное плавало, как
выдра. Ремингтоноцетиды были экологически схожими с ныне живущими крокодилами
гавиалами из Юго-Восточной Азии; они обладали длинными и узкими черепами и
были способны совершать быстрые боковые движения для схватывания рыбы. Морфология
ушной области указывает на их способность к подводному слуху. Ещё несколько
ископаемых форм, таких, как Rhodocetus и Dorudon, документируют
дальнейшие морфологические преобразования вроде редукции задних ног.
Что же ископаемые остатки китов рассказывают нам об эволюции индивидуального
развития? Крупные эволюционные преобразования, такие, как совершенно новый
план строения тела, приспособленный к жизни в новой среде обитания, являются
особенно подходящим объектом исследований в области индивидуального развития,
потому что новшества, которые мы видим у полностью сформированных взрослых
особей – это результат процесса онтогенеза, который должен был в значительной
степени измениться с течением геологического времени. Ископаемые остатки показывают
темп и путь образования новых особенностей. Последовательность их возникновения
особенно важна.
Ханс Тевиссен и его коллеги исследовали некоторые аспекты специализированной
генетической системы развития конечностей у видов дельфинов, результатом действия
которой является почти полная редукция задних конечностей. У большинства Odontoceti,
или зубатых китов, к которым принадлежат
156
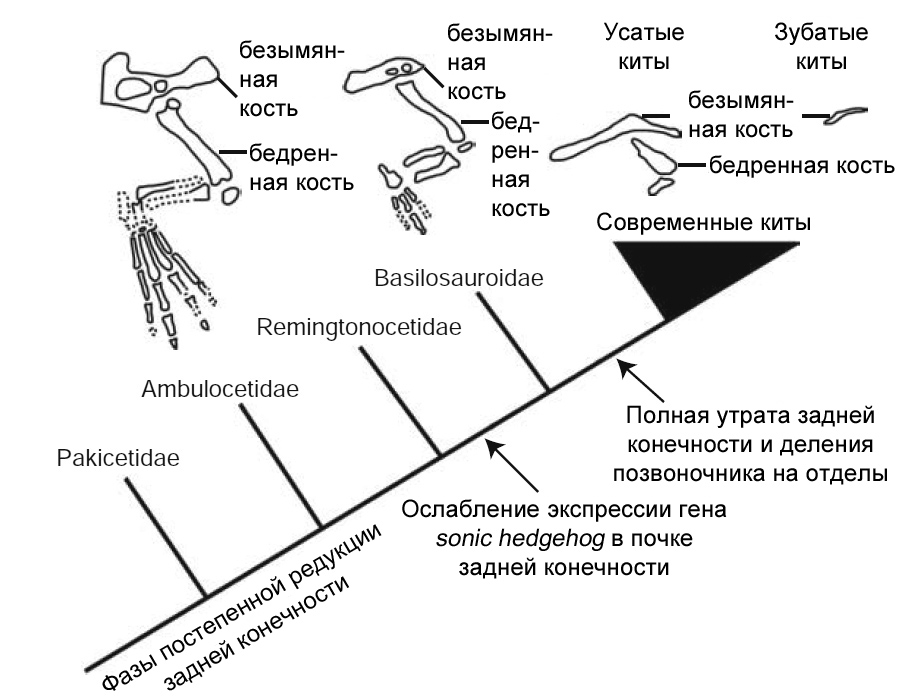
Рисунок 42. Постепенная редукция костей
задней конечности в ходе эволюции китообразных, с примерами из их основных
групп. Из Thewissen и др. 2006, с изменениями. |
дельфины, единственный элемент задней конечности – безымянная кость, тогда
как у усатых китов (Mysticeti) в целом сохраняются некоторые из самых проксимальных
костей задней конечности.
Исследование эмбрионов дельфинов показало, что многие из молекул, задействованных
в развитии конечностей у других млекопитающих, также экспрессируются у дельфинов,
но в иное время и в другом местоположении. Например, апикальный эктодермальный
гребень, дистальный край почки конечности, где происходит экспрессия гена
fgf8, у дельфина является лишь временной особенностью строения. Другая
важная молекула, sonic hedgehog, которая является посредником в развитии
основной части почки конечности у других наземных позвоночных, здесь отсутствует.
Тевиссен и его сотрудники выдвинули гипотезу о том, что это отсутствие
157
связано с утратой дистальных элементов конечности, таких, как ступня. Зафиксированная
последовательность реально происходившего эволюционного изменения, которую
предоставляют нам ископаемые остатки, и знание генетических основ индивидуального
развития, задействованных в процессе редукции задней конечности, предлагают
подробный сценарий изменений. Не имея ископаемых остатков, мы могли бы лишь
предполагать, на что были похожи эти изменения. Но благодаря ископаемым остаткам
могут быть реконструированы потенциальные генетические механизмы, задействованные
в этом процессе.
Содержание
Благодарности
vii
Пролог
xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие
звенья» и эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания
197
Библиография
213
Предметный указатель
243