| Главная | Библиотека | Форум | Гостевая книга |
34
Глава вторая
Эво-дево, пластичность
и модули
Дисциплину, которая объединяет области ведения генетики развития и эволюции, окрестили «эво-дево»*. Очень немногим новым аспектам эволюционной биологии было уделено столько же внимания от практиков и философов от биологии, если такое внимание вообще оказывалось. Предполагалось, что «эво-дево» создаст новый вид синтеза знания, который поможет понять происхождение биологического многообразия. По этой причине важно подумать над тем, как палеонтология вносит свой вклад в эту область знания. Главный объект внимания «эво-дево» ещё является объектом споров. Некоторые видят его в объяснении способности эволюционировать, или эволюабельности. Многие видят его в объяснении эволюционных новшеств или инноваций в смысле действительно новых, крупных шагов в морфологических изменениях – например, эволюцию глаз, зубов, конечностей или черепашьего панциря.1 Другие видят в «эво-дево» лишь «мимолётную комету», дисциплину, которая будет вытеснена более новым слиянием областей знания, или новым порядком групп исследователей и тем, возможно, продиктованным подъёмом геномики.2 Но, как бы то ни было, никто не отрицает того, что эти дискуссии стали источником многих размышлений и импульса к изучению индивидуального развития с эволюционных позиций.
* В оригинале «evo-devo», от слов «эволюция» (англ. «evolution») и «развитие» (англ. «development») – прим. перев.
35
Начала «эво-дево» были во многом связаны с выяснением того, что общего имеют
друг с другом многочисленные отдалённо родственные группы, как, например,
мухи и люди, или несопоставимые элементы анатомии, как зубы и конечности.
То, что глаз мухи и глаз позвоночного образуются благодаря действию сходных
молекулярных механизмов – это крупное открытие. Выяснение того факта, что
некоторые молекулы принимают участие в развитии и зубов, и конечностей – это
тоже важное достижение. Но эти образования, начиная с момента своего появления,
имеют различные формы и цвета, и именно это является захватывающим моментом.
«Эво-дево» изменила акценты с попыток понять, почему организмы столь различны,
на вопрос о том, как могли возникнуть эти несходство и разнообразие, если
помнить о широко распространённом явлении генетического консерватизма, характеризующего
эти организмы. Понимание отношения между генотипом и фенотипом – основное
в этом предприятии.
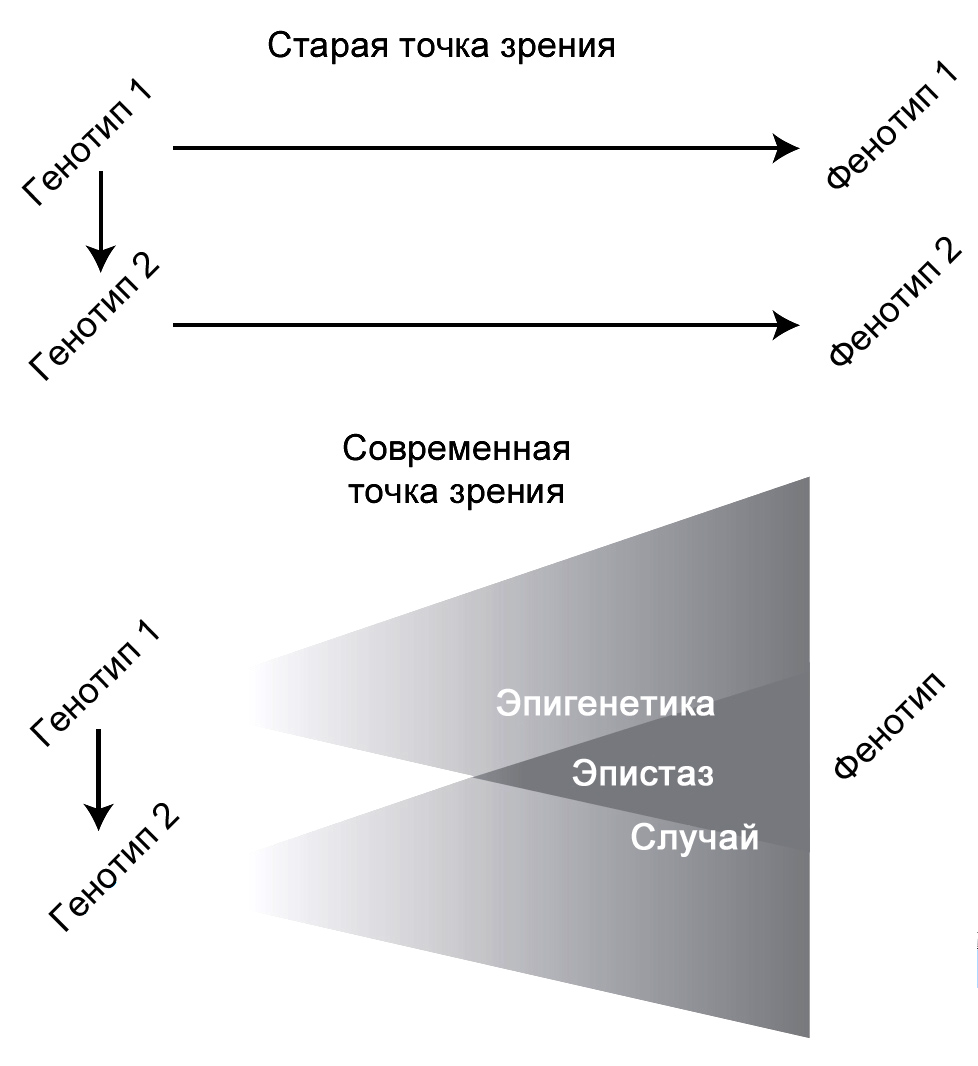
В прошлом многие биологи предполагали, что отношения между генотипом и фенотипом
прямолинейны и просты, когда жизнеспособная мутация непосредственно приводит
к новому фенотипу. На всех уровнях организации, даже на молекулярном, это
оказалось неверным. Уже сто лет назад некоторые биологи писали о норме реакции,
явлении, при котором один и тот же генотип способен образовывать различные
фенотипы в зависимости от внешнего воздействия со стороны окружающей среды.3
Эта идея была во многом забыта ведущими эволюционными биологами, но теперь
она является центральной для исследований индивидуального развития с эволюционных
позиций. Область ведения недавно окрещённой экологической биологии развития,
прекрасно описанная в учебнике Скотта Гилберта и Дэвида Эпела, суммирует множество
данных, касающихся взаимодействия между окружающей средой и индивидуальным
развитием.
Два аспекта разнообразия, которые стали центральными темами исследований «эво-дево»
– это фенотипическая пластичность и модульность. О том, что означают эти важные
концепции и какое отношение имеет к ним палеонтология, говорится далее.
36
Фенотипическая пластичность и изменчивость
Развитие организма – это не река генетической информации, неуклонно текущая по своему руслу в сторону создания биологической формы, а скорее множество рек, притоков и водоворотов – бурный, циклический процесс, задействующий генную регуляцию и синтез белка.
Ничто в изменчивости не имеет смысла вне контекста развития организма.
|
У изменчивости есть негенетический компонент, и фенотипическая пластичность
относится к явлениям изменчивости, вызванным воздействием окружающей среды.
Такое воздействие могут оказывать физические факторы, такие, как температура,
или биотические факторы вроде взаимодействия с другими видами, или стимулы
типа рациона. Конечно, это очень важно, поскольку изменчивость – это как раз
то, на что действует естественный отбор. Чем больше изменчивость, тем активнее
происходит эволюция и тем больше разнообразие.4
Фенотипическая пластичность в характере роста, связанная с факторами окружающей
среды, была отмечена для нескольких видов динозавров, в том числе для прозауропода
Plateosaurus. В периоды изобилия пищи или благоприятного климата
животные росли быстрее. Такой вид пластичности характерен не для всех прозауропод;
он отсутствует, например, у близкородственного Massospondylus. Этот
вид исследований можно предпринять только для того вида, для которого известно
большое количество ископаемых остатков, достаточное для построения графиков
роста с использованием палеогистологии для оценки возраста отдельных экземпляров.
Фенотипическая пластичность предполагалась для нескольких групп ранних наземных
позвоночных палеозоя; это мнение основано на тщательных анатомических и палеоэкологических
исследованиях сотен экземпляров. Райнер Шох в Штутгарте продемонстрировал
на прекрасных по своей сохранности ископаемых остатках темноспондилов, что
один и тот же вид был способен изменять
37
Рисунок 11. Старое и новое представление отношений между генотипом и фенотипом. На основе сходного рисунка из Sholtis и Weiss (2005). |
свою онтогенетическую стратегию в зависимости от условий окружающей среды – это же известно для многих современных саламандр. Данное животное, Sclerocephalus, достигало максимального размера в 2 метра и на первый взгляд напоминало странного крокодила. В больших мелководных озёрах, которые были бедны пищей, Sclerocephalus затормаживался в развитии, что приводило к появлению мелких взрослых особей, поедавших мелкую добычу. В озере с более сложной экосистемой, где и численность, и количество видов пищи были больше, Sclerocephalus имел более продолжительную стадию роста, становился крупнее и мог бы даже рисковать выходить на сушу, хотя содержимое желудка показывает, что это было преимущественно водное существо. В другом доисторическом озере, отложения которого сохранили очень крупных и прожорливых акул, были найдены только личинки, и это заставляет предполагать, что взрослые особи жили где-то
38
Рисунок 12. Скелет Sclerocephalus haeuseri из бассейна Саар-Нахе, юго-западная Германия, примерно 297 миллионов лет. Изображение любезно предоставил Райнер Шох (Штутгарт). |
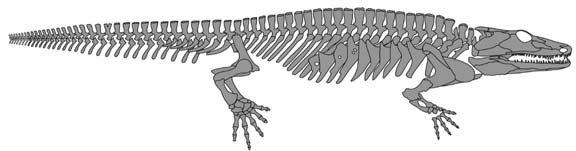
ещё, возможно, чтобы избежать хищников. Связь между экологическими условиями
и изменениями в онтогенезе была установлена для этих палеозойских позвоночных
на основании надёжных свидетельств.
Sclerocephalus явно проявлял пластичность в своей стратегии развития;
у других ранних наземных позвоночных, также известных по сотням экземпляров
из различных местностей, она проявляется гораздо слабее. К сохранению таких
ископаемых остатков и их открытию привели исключительные обстоятельства. Болота
и озёра зачастую являются хорошими местами для захоронения ископаемых остатков,
и в некоторых случаях бескислородные условия илистого дна этих водоёмов, со
слабой биологической переработкой органики привели к превосходному сохранению
в ископаемом виде костей и даже мягких тканей мёртвых тел этих животных. Поскольку
онтогенез отдельно взятой особи у этих ископаемых животных нельзя проследить
непосредственно, как это можно сделать сегодня у лягушки или саламандры, палеонтологам
приходится опираться на тщательное анатомическое сравнение многочисленных
экземпляров, и, хотя определение различных форм с точностью до вида может
быть проблематичным, обычно мы можем идентифицировать этих животных с высокой
степенью достоверности.
Палеонтологи традиционно интересовались изменчивостью из-за потребностей таксономического
характера. Чтобы определить границы вида по ископаемым остаткам, требуется
установить максимальную степень морфологической изменчивости, которую может
проявлять вид.
39
Для этой цели подходит метод оценки или измерения некоторых черт строения
у как можно большего числа индивидуумов, а сравнение с соответствующими ныне
живущими видами и рассмотрение онтогенетической изменчивости являются основополагающими.
Другой необходимый аспект – это знание эволюционного древа, в которое включался
бы новый вид, и вместе с этим установление продвинутых признаков, которые
характеризуют этот вид.
Форма и размеры костей изменяются в пределах вида, и даже в пределах популяции.
Но не все элементы скелета изменяются в одинаковой степени. Один из факторов,
который может оказывать влияние на степень изменчивости – это происхождение
кости в процессе индивидуального развития. Некоторые кости образуются из хрящевых
предшественников и являются частью основы скелета, во многих случаях глубоко
интегрированными в составе скелетного модуля, или группы костей, которая развивается
в большей или меньшей степени в унисон.
У других костей нет никаких хрящевых предшественников. Для тех костей, которые
образуются из соединительной ткани, как, например, коленная чашечка в человеческом
колене, присутствующая также у многих других видов млекопитающих, ожидается
проявление большей изменчивости по размерам и по форме. Коленная чашечка и
другие кости, известные как сесамовидные, неплотно сочленяются с другими костями,
и подвергаются большему воздействию со стороны мускулов и сухожилий, работающих
вокруг области, в которой они формируются. Потому для сесамовидных костей
ожидалось, что они будут более изменчивыми, чем другие кости. Эти предсказания
были проверены и подтверждены в ходе изучения плейстоценовых млекопитающих,
проведённого Кристиной Раймонд и Доном Протеро, которые измерили почти две
тысячи костей из двух местонахождений ископаемых остатков: Ранчо Ла Бреа в
Лос-Анджелесе и пещеры Сан Хосито в Нуэво Леон, Мексика. Кости принадлежали
шести видам млекопитающих: саблезубой кошке Smilodon fatalis, льву
ледникового периода Panthera atrox, бизону Bison antiquus,
лошади Equus occidentalis, верблюду Camelops hesternus и
наземным ленивцам Paramylodon harlani и Nothrotheriops shastensis.
Раймонд и Протеро сравнили в рамках статистического подхода изменчивость
40
для нескольких промеров 989 мембранных костей, включая коленную чашечку и другие сесамовидные кости, с промерами для 811 энхондральных костей, прежде всего для таранных костей сопоставимого размера. Для двадцати одного из двадцати семи промеров у мембранных костей изменчивость оказалась значительно выше, чем у энхондральных.
Модульность и интеграция
Концепции модульности и интеграции находятся в центре внимания при разговоре
об эволюции и индивидуальном развитии. Модули – это подмножество признаков
данного организма, которые больше объединены друг с другом, чем с остальными
признаками этого организма.
Например, бугры коренного зуба гораздо больше влияют друг на друга, чем любой
из них влияет на форму клыка, или, естественно, на какие-то другие аспекты
анатомии скелета. Сам Дарвин признавал интеграцию между частями организма,
и он привёл много примеров корреляций роста в первой главе «Происхождения
видов…». Все особенности строения организма интегрированы, но некоторые –
в большей степени, чем другие; поэтому интеграция не гомогенна. То, как мы
подразделяем анатомию организма, несомненно, связано с этим. Коренной зуб
можно было бы осмыслять как единое образование, без его разделения на бугры
и другие части, такие, как корень зуба. Если отмежеваться от этого субъективного
аспекта, то интуитивно очевидно, что мускулы, части сустава и кости в области
колена будет объединены друг с другом более тесно, чем любая из этих частей
с любой частью шеи. Эта интеграция не наблюдается непосредственно и должна
выводиться из статистического характера изменчивости исследуемых особенностей.
Цель состоит в том, чтобы разграничить модули путём количественной оценки
характера взаимодействия между чертами строения – так называемого характера
ковариации.
41
Подробности того, как это делается, разнообразны и являются предметом споров,
поскольку это расширяющаяся область для исследований. Морфологические модули
существуют в различных контекстах: развития организма, генетическом, функциональном
и эволюционном. Методы исследований, изучающие их, различны в каждом из случаев.
Первое всестороннее исследование модульности с использованием количественного
подхода было представлено Эвереттом Олсоном и Робертом Миллером в их книге
«Morphological Integration», вышедшей в свет в 1958 году. Они определили программу
исследований и представили первые попытки статистического подхода к количественному
определению корреляции признаков. Олсон и Миллер весьма часто обращались к
ископаемым видам, например, упоминая о явно модульной эволюции зубных бугров
в родословной линии эоценового млекопитающего Hyopsodus. Они обратили
внимание на «прогрессивную утрату у последовательно сменяющих друг друга видов»
параконида, одного из небольших бугров в нижних зубах млекопитающих, которая
выглядит независимой от других изменений в форме зубов. Более современная
работа, проведённая в лаборатории Юкки Йернвалла в Хельсинки, показала, что
несколько признаков бугров на коренных зубах млекопитающих в значительной
степени интегрировано из-за связи, возникающей между ними в процессе развития.
Эти наблюдения имеют большое значение для реконструкции эволюционных древ,
которая в случае ископаемых млекопитающих основывается в значительной степени
на признаках строения зубов, которые считаются независимыми друг от друга.
Некоторые люди активно реализуют исследовательские программы по изучению модульности,
и среди них Крис Клингенберг, работающий с очень разнообразными группами позвоночных
и беспозвоночных, и Габриэль Марро – с млекопитающими. Марро и его коллеги
изучили тысячи черепов некоторых групп млекопитающих, чтобы провести сложные
анализы, которые требуют больших выборок материала, чтобы дать статистически
существенные результаты. Объединяя свои данные с известными генеалогиями индивидов,
которых они изучали, и эволюционными древами в случае целых групп, Марро получил
возможность исследовать отношение между
42
генетической структурой популяций и морфологической изменчивостью черепа
в них. Главнейшее открытие, сделанное в ходе этого исследования – то, что
важнейшим определяющим фактором способности группы к эволюции является её
диапазон изменчивости в размерах.
Характер интеграции и модулей, обнаруженный у взрослых особей, отражает ассоциации,
которые возникают в ходе развития, имеет генетические основы и наследуется.
Это означает, что, изучая взрослых особей, мы можем косвенно изучить индивидуальное
развитие. Это открывает область исследования для палеонтологии. Анджали Госвами
также изучала модульность в черепе млекопитающих, но старалась также охватить
исследованием ископаемые формы, чтобы изучить значение модульности для крупномасштабных
картин эволюционных процессов. В своём исследовании, на которое часто ссылаются
специалисты, она изучила 105 видов, восемь из которых – ископаемые. Она зафиксировала
сведения об опознавательных точках многих участков черепа, сопоставимых (гомологичных)
для разных видов, таких, как границы между двумя костями. Далее этот трёхмерный
массив данных был статистически проанализирован с проведением тестов, которые
показали, какие опознавательные точки в значительной степени коррелируют друг
с другом и изменяются в унисон. Анджали идентифицировала в черепе шесть модулей:
в передней части лица, вокруг коренных зубов, вокруг глазницы, вокруг боковой
части черепа, в своде черепа и в основании черепа. Почти все ископаемые формы,
которые исследовала Анджали, соответствуют этому стандартному набору модулей,
в том числе один саблезубый вид с очень специализированным черепом. Ископаемая
саблезубая кошка Smilodon fatalis демонстрирует характер интеграции
черепа, отличающийся от такового у любого из остальных исследованных видов:
у него лицевой скелет и увеличенные клыки образуют два отдельных модуля вместо
одной передне-ротовой–назальной группы. В чём причина такого отклонения? Некоторые
аспекты картины роста Smilodon, вероятно, связаны с новым распределением
модульности. Для Smilodon характерно уникальное распределение времени
смены зубов. Молочные
43
сабельные зубы прорезаются в молодом возрасте, вероятнее всего, позволяя
охотиться в то время – и влияя на характер корреляции между частями черепа
на ранних этапах его формирования. У других ископаемых саблезубых форм молочные
сабельные зубы появлялись поздно, что в итоге вело к более длительному сохранению
предкового характера интеграции черепа в эволюционной линии. Эмпирические
свидетельства показывают, что модульность сама по себе, как и изменение пропорций
между частями черепа в процессе роста (аллометрия), подвержена процессу эволюции.
Ископаемые остатки значительно расширяют диапазон морфологически возможных
изменений, или же подтверждают закономерности, выявленные в ходе изучения
исключительно ныне живущих видов.
Поскольку специалисты по палеонтологии позвоночных вынуждены сосредотачивать
своё внимание только на скелете, многие другие аспекты анатомии, а вместе
с ней и индивидуального развития, которые могут проявлять себя иным образом,
выпадают из поля зрения. Из-за явления модульности анатомические области могут
вести себя по-разному и независимо друг от друга. Например, у обезьян, группы
млекопитающих, к которой принадлежим и мы, скелет в общих чертах сходен у
всех видов, тогда как пищеварительная система чрезвычайно изменчива и включает,
в том числе, адаптации к ферментации пищи в передней и задней кишке.
Выживание здесь и сейчас:
утраченные и замещённые модули
Всё в жизни является вопросом альтернатив. Это не наличие универсального оптимума, а локальное достижение лучшего результата. Хороший пример этого «принципа» – самоампутация. Это действие, посредством которого животное отделяет от себя один или больше своих собственных придатков, когда лучше потерять хвост или конечность, модуль, нежели потерять свою жизнь – особенно, если вы сможете затем регенерировать этот придаток, как это известно для многих групп позвоночных, за исключением млекопитающих. Саламандры могут даже регенерировать конечности. Ящерицы, находящиеся ближе к нам на древе
44
жизни, могут регенерировать хвост, но уже не способны регенерировать свои
конечности.
Хвост – это модуль организма, обычно характеризующийся большей степенью своей
изменчивости по сравнению с другими частями осевой области тела у позвоночных,
как уже отмечал в 1913 году Э. С. Гудрич. В зависимости от рассматриваемого
вида живых организмов, хвосты могут играть важную роль в передвижении, запасании
жира или защите от хищников среди прочего. Чтобы вырастить хвост, нужно затратить
энергию, и потому, если он есть в наличии, его роль, вне всяких сомнений,
принадлежит к числу условий для поддержания жизни. Но много разных животных
может выживать и без него. В случае с автотомией хвоста некоторые ящерицы
и саламандры, которые схвачены за хвост, будут отбрасывать его часть и таким
образом получат возможность спастись бегством. Отброшенный хвост отвлекает
внимание хищника от убегающего животного. У сцинков одного из видов (Scincella
lateralis) особи, которые отбросили хвост, возвращаются на место, где
они его потеряли, и, если они находят его, то заглатывают, чтобы возместить
значительную часть затраченной энергии. Ящерицы могут частично регенерировать
свой хвост, но при этом исходная структура с позвонками замещается хрящевым
тяжом. Внешне хвост также может выглядеть иначе: тоньше или толще, или же
иметь окраску, отличную от исходной. Медицинское сообщество, конечно, очень
интересуется этим явлением, сознавая важность возможности регенерации тканей
для людей, которым была сделана ампутация. В отличие от полной способности
к регенерации у амфибий, частичная способность к ней у ящериц – это лучшая
модель, демонстрирующая регенерацию ткани в хвосте в противоположность неспособности
к регенерации у конечности.
Чтобы понять, как действует автотомия хвоста, полезно понять, как она появилась
в процессе эволюции. Эволюционировала ли эта способность только один раз,
или же много раз и независимо? У каких групп она есть, и что у них есть общего?
Эти вопросы всё ещё остаются без ответа. Если рассматривать вымерших животных,
то очень трудно, если вообще воз-
45
можно сказать, какой вид не обладал способностью к регенерации придатков
тела, поскольку в данном случае отрицательное свидетельство – это отсутствие
свидетельств вообще. Но летопись окаменелостей представляет свидетельства
наличия автотомии хвоста. Об этом можно сделать вывод по ископаемым остаткам,
на которых сохранилась особого рода пластинка, служащая для разрыва хвоста.
На ранних стадиях развития исходные эмбриональные сегменты, называемые сомитами,
подвергаются ресегментации, при которой задняя половина сомита соединяется
с передней половиной следующего за ним сегмента, и так далее. Пластинка для
разрыва хвоста возникает на границе структур, образующихся из двух эмбриональных
предшественников, которые объединяются на ранней стадии развития. Когда эту
особенность смогли идентифицировать в окаменелых хвостовых позвонках, она
была отмечена в разнообразных группах, в том числе у пермских капторинид и
других базальных групп рептилий, возможно, у морских рептилий, называемых
мезозаврами, возможно, у представителя пролацертиформ Tanystropheus5,
у клювоголовых, ящериц и морских крокодилов из юрского периода, и у ящерицы
из среднего эоцена Германии.
Поскольку в ходе регенерации позвонки хвоста замещаются хрящевым тяжом, до
недавнего времени мы могли только гадать, происходила ли регенерация у ископаемых
таксонов. Хельмут Тишлигер из Баварии, который на протяжении ряда лет специализировался
на использовании длинноволнового ультрафиолетового излучения для распознавания
образований в окаменелостях, сумел открыть новые детали у окаменелостей, известных
на протяжении долгого времени, например, отпечатки мягких тканей у берлинского
экземпляра Archaeopteryx. Недавно, используя метод фотографирования
в длинноволновом ультрафиолетовом свете, который показал ореол, соответствующий
былому наличию хвоста, состоящего исключительно из мягких тканей, он обнаружил,
что регенерация происходила у некоторых верхнеюрских рептилий отряда чешуйчатых
из Германии.
Содержание
Благодарности vii
Пролог xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево,
пластичность и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания 197
Библиография 213