| Главная | Библиотека | Форум | Гостевая книга |
66
Глава четвёртая
Кости и зубы под
микроскопом
Может показаться весьма невероятным, но самым важным рабочим инструментом для большинства палеонтологов, помимо молотка, является микроскоп. Значительная часть людей, изучающих доисторическое биологическое разнообразие, работает в интересах нефтедобывающей промышленности, исследуя мельчайшую пыльцу ископаемых растений или ископаемых фораминифер; последние являются представителями группы одноклеточных организмов, важной для установления стратиграфической корреляции между геологическими подразделениями. Палеонтологам микроскоп позволяет также изучать микроструктуру тканей ископаемых организмов, в особенности кость, которая стала важным предметом исследований, имеющих отношение к изучению индивидуального развития у вымерших таксонов. Изучение живых тканей, или гистология, является обширным полем деятельности, и многое в ней касается идентификации патологий, например, важная диагностическая процедура при исследовании рака. Сравнительные исследования тканей различных организмов, бывших предметом изучения на протяжении столетий, стали возможными благодаря технологическому прогрессу в изготовлении тонких срезов нежных и зачастую мелких комплексных структур различной консистенции и формы. Самые первые работы с тканями на микроскопическом уровне
67
осуществил швейцарец Вильгельм Гис (1831–1904), уроженец Базеля. Он изобрёл
микротом, механическое устройство, состоящее из хитроумно расположенного лезвия,
которое использовалось для того, чтобы делать тонкие срезы тканей для микроскопического
исследования. Благодаря этой новой технологической разработке ему удалось
проследить происхождение различных типов тканей животных на эмбриональной
стадии. Насколько я знаю, Гис не изучал микроструктуру ископаемых костей;
он скорее интересовался эмбрионами и мягкими тканями, например, делая открытия,
которые дали ему повод использовать термин «дендриты» для проводящих отростков
нервных клеток. Гистологические срезы с использованием преимущественно того
же самого метода, который применял Гис примерно 150 лет назад, в настоящее
время используются при изучении эмбрионов ныне живущих видов, которые, учитывая
их размер, представляют собой очень сложный объект для изучения методом анатомирования.1
С достаточно давних пор палеонтологи поняли, что тонкие срезы могут показать
важные анатомические детали ископаемых остатков. Британский специалист по
ископаемым рыбам Уильям Джонсон Соллас (1849 – 1936), профессор геологии из
Оксфорда, и его дочь Игерна стали пионерами в этих исследованиях, когда в
1903 году издали работу о девонской рыбе с неясными родственными связями.
Совершённый ими технический прорыв состоял в последовательном стачивании образца,
создании рисунков появившихся при этом сечений и переводе их в восковую модель,
увеличенную по сравнению с оригиналом. Соллас вначале создал модели ископаемых
змеехвосток (офиурид) и граптолитов, группы вымерших беспозвоночных, родственных
баланоглоссу, которые были выставлены в Британской научной ассоциации в 1901
году.2 Соллас сделал
модели многих окаменелостей, но похоже, что сохранились лишь немногие из них.
Позже метод Солласа был мастерски использован школой шведских специалистов
в области сравнительной анатомии. Но изготовление восковых моделей отнимает
чрезвычайно много времени. Например, известная восковая модель головы Eusthenopteron,
животного, важного звена в переходе между рыбами и наземными позвоночными,
отняла пятнадцать лет времени у двух техников на её
68
изготовление. В наше время цифровая камера и правильно подобранные оборудование
и программное обеспечение могут сделать ту же самую работу в течение нескольких
дней. Это технический прогресс огромного масштаба.
Следующий крупный технологический шаг вперёд был представлен в работе 1970
года французскими исследователями Сесиль Поплен и Арманом Ж. де Рикле, которые
изобрели метод получения срезов ископаемых образцов с помощью микротома. Они
подбирали технику разделения окаменелостей на срезы, которые можно было бы
разместить на стекле, как при стандартных гистологических методах. Это были
вовсе не тривиальные сложности, поскольку у окаменелостей твёрдая ткань пористая,
хрупкая, ломкая, более гетерогенная по составу и менее эластичная при нагрузках,
чем кость современных животных. При пропитке ископаемых костей смолами в вакууме
и под давлением каждый срез становится более устойчивым. Химический состав
ископаемой кости также отличается от её состава у ныне живущих таксонов. Получившиеся
в результате срезы сохраняют очертания различных структур, имеющихся в окаменелости.
Следовательно, даже если химический состав изменился или исходные компоненты
были замещены, границы между ними сохранились. Эта техника использования микротома
была первым шагом к замене метода последовательного стачивания, которому приносится
в жертву исходная окаменелость. Новейшая техника визуализации внутренней структуры
окаменелости не требует получения срезов. С помощью мощной компьютерной томографии
с высоким разрешением теперь возможно изучить детали микроанатомии, даже не
разрезая окаменелость.
Информативные срезы ископаемых костей
Причина того, что гистология ископаемой кости является богатым источником информации, состоит в том, что кость – это живая ткань, которая претерпевает много последовательных процессов резорбции и реконструкции. Этот факт приносит нам пользу после того, как с нами приключается несчастный случай, при котором мы ломаем кость; мы страдаем от этого в пожилом возрасте из-за остеопороза. Для палеонтологов изменение кости
69
в течение жизни означает, что многое из этой записи жизни позвоночного животного
можно увидеть. Ключевой момент здесь – умение различать следы первичного роста
кости и процессов вторичной перестройки среди отметок, образовавшихся в процессе
роста.
Самую ненарушенную информацию о росте скелета позвоночного обычно получают
из района средней части длинных костей, таких, как в руках (плечевая кость)
или в ногах (бедренная кость). Перед изготовлением срезов кости погружаются
в синтетическую смолу, затем они шлифуются и полируются, пока не достигают
соответствующей толщины, обычно 0,6 – 0,8 см. Далее эти срезы изучаются под
микроскопом при различном освещении и с разными светофильтрами. Обычно предметом
исследований палеонтологов являются отдельные обломки костей, поскольку музейные
кураторы обычно не любят агрессивного способа исследований, который включает
разрезание ископаемых образцов на тонкие кусочки. Это приводит к проблемам,
поскольку зачастую нет уверенности в том, что известно точное с позиции анатомии
или таксономии место взятия образца. Несмотря на эту трудность, количество
новых открытий, сделанных путём исследований тонких срезов окаменелостей,
увеличивается экспоненциально.
Моё первое знакомство с палеогистологией произошло благодаря моему более молодому
коллеге Торстену Шейеру, который работал в моей лаборатории вскоре после моего
прибытия в Цюрихский Университет, и благодаря его бывшему научному руководителю
работы на соискание степени доктора философии Мартину Сандеру из Бонна, мировому
лидеру в этой области. Я посетил лекцию Сандера в Тюбингене, посвящённую зауроподам,
крупнейшим среди динозавров, на примере которых он показал, как много может
дать палеогистология в понимании характера роста и устройства костей у этих
животных. Мне пришло в голову, что этот подход можно было бы использовать
для исследования Stupendemys geographicus, самой крупной водной черепахи
из когда-либо существовавших, представителя гондванской группы, остатки которой
мы собрали в ходе моего полевого проекта в Венесуэле. Сандер направил меня
к Шейеру, и мы исследовали Stupendemys и других животных, главным
образом млекопитающих и морских рептилий, о чём я и расскажу далее.
70
Панцирь Stupendemys принадлежит к числу самых толстых и самых крупных
среди известных панцирей черепах; у одного экземпляра он достигает 3,3 м в
длину. С точки зрения гистологии он демонстрирует хорошо выраженную слоистую,
словно бутерброд, структуру, где внешние слои, включающие плотную кость, устойчивую
к напряжению, окружают ячеистую внутреннюю губчатую кость. Такой тип структуры
был важен для уменьшения веса панциря при сохранении прочности в процессе
роста. Совокупность отметок, характеризующих рост, была неполной из-за эрозии,
которой, несомненно, подверглись образцы, изучавшиеся нами, и которая характеризует
местонахождение окаменелостей Урумако в Венесуэле.3
Однако, вид ткани и просветы между отметками, образовавшимися в процессе роста,
сопоставимы с таковыми у ныне живущих черепах с «нормальным» медленным темпом
роста. Если предположить темп роста подобным таковому у большинства современных
морских черепах с сопоставимыми гистологическими характеристиками, то экземпляр,
который мы изучили, должно быть, рос примерно от 60 до 110 лет, прежде чем
достиг гигантского размера панциря.
Организация ткани, наблюдаемая у современных животных, служит ключом для понимания
особенностей гистологии, которые можно наблюдать в ископаемых костях. Остеобласты
– это клетки, создающие кость. Также существуют пустоты, называемые лакунами,
которые связаны друг с другом посредством канальцев, ответственных за распределение
питательных веществ и кислорода. Эти и другие основные особенности строения
кости могут наблюдаться в окаменелостях. Остеобласты и остеоциты, кровеносные
сосуды или васкулярные каналы и коллагеновые волокна в большинстве случаев
разрушаются в ходе фоссилизации, но их положение и форма могут быть зафиксированы
в полной мере.4 Некоторые
исследования ныне живущих позвоночных показали, что костные ткани у животных,
которые растут быстро, обладают иными особенностями в сравнении с тканями
тех, кто растё медленно. Быстрый рост связан с костной тканью, богатой кровеносными
сосудами, означающей усиленное снабжение кости кислородом и питательными веществами.
Медленный рост коррелирует с менее васкуляризированной костью, часто прерываемой
концентрическими
71
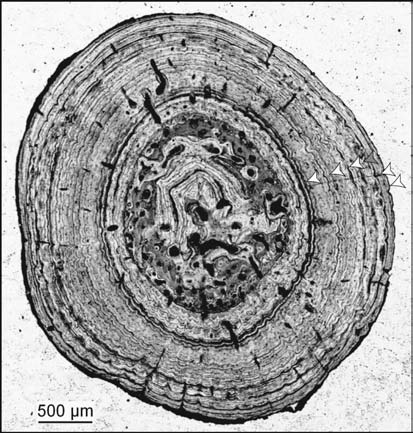
Рисунок 17. Тонкий срез фрагмента щитка Stupendemys geographicus из позднемиоценовой формации Урумако, Венесуэла. Образец находится в коллекции Университета Франсиско де Миранда в Коро (UNEFM-CIAPP-2002-01). |

линиями, сходными с годичными кольцами дерева. Это открытие – то, что скорость
роста влияет на тип откладываемой костной ткани – было опубликовано Родолфо
Амприно из Туринского Университета в 1947 году и сейчас известно как правило
Амприно. Изучение гистологии костей было продолжено после выхода статьи Амприно,
и палеонтологи следовали ходу его мыслей, пытаясь реконструировать аспекты
физиологии и истории жизни ископаемых видов.
В микроструктуре кости также существуют следы, которые предоставляют нам информацию
о продолжительности жизни, возрасте и зрелости; это предмет изучения для скелетохронологии.
Оценка возраста основывается на идентификации отметок, возникающих в процессе
роста, таких, как кольца, образующиеся при замедлении роста, и линии задержек
роста (ЛЗР) между зонами более быстрого роста кости. В большинстве случаев
отметка, свидетельствующая об одном замедлении роста (коль-
72
Рисунок 18. Схематическое представление двух основных видов гистологии костей. Они обычно отмечаются для эктотермных «рептилий» (слева) и эндотермных птиц и млекопитающих (справа). По Chiappe 2007, с изменениями. |
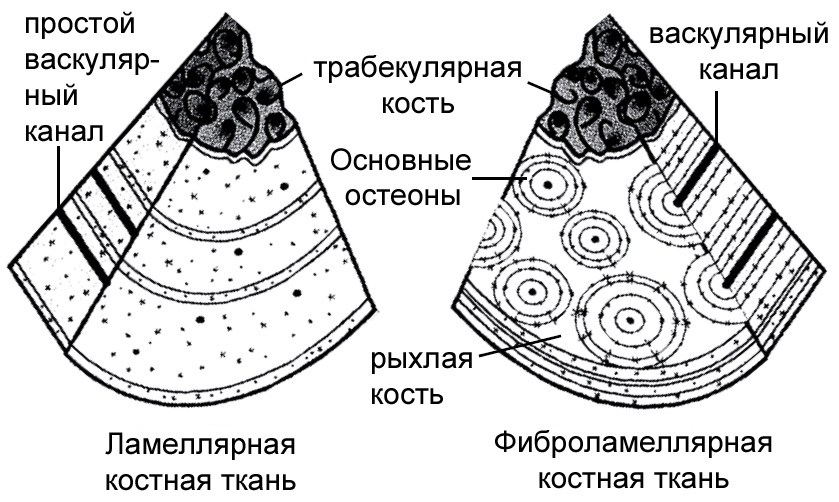
цо) или его остановке, откладывается каждый год, во время более холодного,
или же более сухого сезона, и это вызвано характером питания и физиологией
животного наряду с переменными составляющими вроде болезни и репродуктивной
цикличности. Из-за этих событий, происходящих в жизни отдельной особи, мы
знаем, что образование маркеров роста может быть нерегулярным. Типичная картина
отклонения – наличие короткого периода роста между годичным кольцом и ЛЗР,
или когда за один цикл роста откладываются два кольца (плюс одна ЛЗР). Кроме
минимального возраста, по микроструктуре костей можно дать оценку половой
зрелости.
Уменьшение ширины зон быстрого роста и обычно изменение вида откладывающейся
костной ткани может подразумевать достижение половой зрелости. В наилучших
случаях может быть изучена ростовая серия образцов, и данные, полученные из
различных элементов скелета, в противоположность единственному образцу, являются
намного лучшим материалом для того, чтобы делать достоверные выводы о характере
роста вымерших позвоночных. Для оценки минимального возраста животного в каждой
кости
73
подсчитываются циклы роста; для заключения о возрасте половой зрелости анализируется
размер годичных слоёв. Когда в разных костях одной и той же особи наблюдается
резорбция костной ткани, для оценки минимального возраста индивида используется
кость с минимальным количеством резорбции и максимальным количеством циклов
роста.
Разрушение и перестройка кости осуществляется клетками, которые носят название
остеокласты.5 Это интенсивный
и энергоёмкий процесс; поэтому развитие кости тесно связано с метаболизмом.
Это представляет непосредственный интерес для людей: самые последние исследования
в области молекулярной медицины показали, что ожирение, диабет и остеопороз
– это взаимосвязанные состояния.6
В этом контексте были важны эксперименты с лабораторными мышами. Многое из
того, что мы знаем о том, как растут и изменяются кости, получено прежде всего
в ходе контрольных исследований, когда изучаются кости отдельно взятых животных,
для которых исследователи знают пол, историю жизни, условия жизни и рацион.
Например, у многих видов позвоночных беременность может привести к значительной
перестройке костных тканей.7
Это было зарегистрировано для многих видов рептилий, таких, как черепахи и
крокодилы. В зависимости от группы источником кальция являются строго определённые
кости, но не какие-то другие. Одной из них является малая берцовая кость,
более тонкая из двух длинных костей, которая также присутствует в задней конечности
и у нас, у людей. У некоторых видов крокодилов остеодермы у откладывающих
яйца самок функционируют как хранилища кальция в ходе оогенеза. Эта стратегия,
вероятно, характеризовала, по крайней мере, некоторых ископаемых рептилий,
поэтому, как правило, использование остеодерм для целей скелетохронологии
не является оптимальным решением, потому что в них происходит перестройка
костного вещества. В случае с черепахами можно было бы подумать, что панцирь
представляет собой превосходный источник материала, но пока изученные виды
не демонстрируют использование самками этого источника. Возможно, панцирь
просто слишком важен для обеспечения биомеханической целостности животного.
Вместо него самки используют длинные
74
кости вроде бедренной. Это явление в ходе оогенеза аналогично тому, что происходит
у людей во время беременности и лактации, в периоды, характеризующиеся уменьшением
плотности минеральной составляющей костей у женщин.
Птицы производят твёрдую скорлупу яиц, состоящую больше чем на 90 процентов
из кристаллов карбоната кальция. У птиц перед овуляцией в полости костного
мозга образуется особый вид эфемерной костной ткани, служащий источником кальция,
который позволяет матери образовывать скорлупу яиц. Этот тип кости называется
медуллярной костью, и теперь он описан для Tyrannosaurus rex и для
других видов, представляющих основные группы динозавров. Это открытие стало
важным не только для того, чтобы вывести происхождение птиц из динозавров,
но также (что даже ещё важнее) для более точной реконструкции кривых роста
для многих видов динозавров с использованием надёжного маркера состояния репродуктивной
зрелости. У динозавров этот момент совпал с переходом от стадии ускорения
роста к стадии замедления.
Характер роста динозавров
и происхождение птиц
Мы очень много знаем об онтогенезе костных тканей у динозавров, и гораздо меньше о нём у видов млекопитающих. Главной проблемой, связанной с палеогистологическими исследованиями, был ответ на вопрос, на кого больше были похожи динозавры в физиологическом плане: на эктотермных рептилий, или на эндотермных птиц и млекопитающих. В 1960-х и 1970-х годах Арман де Рикле и его коллеги в Париже выяснили, что кость динозавра не является типичной рептильной костью, но вместо этого она сильно васкуляризирована, подобно костям птиц и млекопитающих. В целом это дало основание полагать, что динозавры быстро росли, и что рост поддерживался высоким уровнем метаболизма. Эти
75
открытия внесли свой вклад в смену парадигмы динозавров – с медлительных,
тупых и холоднокровных гигантов на красочных, динамичных и теплокровных существ.
Однако, некоторые исследования показали, что быстрый рост не может просто
коррелировать с эндотермией, или теплокровностью, поэтому простого ответа
на вопрос относительно эндотермии и эктотермии не существует. Вместо этого
у динозавров обнаружена богатая и разнообразная картина уникальных типов роста
и физиологии.
Существует много видов динозавров, они жили на протяжении большого отрезка
времени, приблизительно 160 миллионов лет, и достигали максимального для взрослой
особи размера в диапазоне от примерно 1 килограмма до 75 тонн. Некоторые динозавры
выжили и в процессе эволюции превратились в птиц*, и это ещё один крупный
переход. Де Рикле, Жак Кастане, Джек Хорнер и их коллеги по всему миру улучшили
качество выборки по различным костям и их сравнению с ныне живущими аналогами,
и также разработали скелетохронологический метод, который дал информацию о
темпах роста.
Кевин Падиан и его коллеги сообщили, что у многих видов динозавров в компактной
костной ткани имеется кость различных типов, отражающая изменения в темпе
роста; похоже, что в конце жизни рост замедляется или даже прекращается. Мы
знаем, что, даже если стратегии роста у динозавров сильно различались, эти
животные росли в темпе, существенно опережающем таковой у ныне живущих рептилий,
далёких от птиц с позиций систематики. Даже крупнейшие из динозавров достигали
взрослого размера меньше, чем за три десятка лет. Длинные кости крупных динозавров
и птерозавров в большинстве случаев состоят из хорошо васкуляризированной
фиброламеллярной кости, что отражает быстрый темп роста. Этот вид костной
ткани обнаруживается у коров, лошадей, лосей и других крупных млекопитающих,
а также у крупных птиц. Но ранние птицы и многие динозавры не всегда обладают
костью такого типа, и если он у них есть, это не обязательно вся компактная
костная ткань.
Большой объём палеогистологических исследований был проведён в отношении динозавров-
* Это не совсем точное утверждение. Вначале всё же появились птицы, и лишь потом вымерли динозавры. Какое-то время они существовали бок о бок. – прим. перев.
76
зауропод.8
Особый интерес к этим животным вполне оправдан, поскольку они были самыми
крупными наземными животными. Особенности гистологии костей передней и задней
конечности были использованы Мартином Сандером и его студентами в Бонне для
изучения темпа метаболизма и поиска ответа на вопрос, достигались ли большие
размеры за счёт увеличения темпов роста, или же продления времени роста. Крупные
юрские формы, потенциально эндотермные, явно росли быстро – несомненно, быстрее,
чем самые ранние виды зауропод. Mamenchisaurus, позднеюрский зауропод
из Китая, рос с максимальной средней скоростью 2 тонны в год, и это замечательное
достижение для метаболизма. Некоторые виды демонстрируют пластичность в развитии,
и действительно, различные онтогенетические «стадии» можно распознать, основываясь
на различных видах гистологии костной ткани в пределах одного вида.
Если использовать для сравнения только современных рептилий с их замедленным
темпом метаболизма, то можно прийти к выводу, что огромному юрскому зауроподу
Apatosaurus должно было требоваться до 120 лет, чтобы дорасти до
своего полного огромного размера. Но, изучая гистологию костей, мы знаем,
что Apatosaurus должен был достигнуть своего полного размера в возрасте
всего лишь от десяти до двенадцати лет; эта картина больше сходна с наблюдаемой
у эндотермных птиц и млекопитающих.
Другие виды динозавров также стали объектом палеогистологических исследований,
включая, разумеется, харизматичных теропод – группу, к которой принадлежит
не только Tyrannosaurus rex, но также и птицы. Возможность определить,
когда и как появился эндотермный тип роста у птиц, представляет большой интерес.
У Tyrannosaurus масса тела взрослой особи достигала приблизительно
5 тонн, а максимальная скорость роста – 2,1 килограмма в день.9
По массе и скорости роста T. rex был сходен с современным африканским
слоном. T. rex доживал до двадцати восьми лет и достигал скелетной
зрелости в возрасте примерно двух десятков лет. Как и у других наземных теропод,
но в отличие от современных птиц, половая зрелость у T. rex
77
наступала задолго до того, как животное вырастало до своего полного размера.
Как и в случае с гигантскими зауроподами, T. rex достигал своего
огромного размера главным образом за счёт ускорения темпа роста по сравнению
со своими предками.
Уменьшение размера тела при эволюционном переходе от не-птичьих динозавров-теропод
к птицам документировано многочисленными и довольно полными окаменелостями.
В ходе изучения палеогистологии Кевин Падиан и его коллеги достигли понимания
изменений в процессе роста, лежащих в основе этого изменения размеров. Они
пришли к заключению, что уменьшение размера, связанное с происхождением птиц,
включает сокращение продолжительности фазы быстрого роста предков.
Кости моа
Объектами палеогистологических исследований также стали другие рептилии и птицы, особенно те, которые демонстрируют крайне выраженные признаки адаптации. Опираясь на них, мы знаем, что продление времени роста было механизмом, задействованным в эволюции двух очень крупных и впечатляющих форм: гигантского североамериканского крокодиломорфа Deinosuchus из позднего мела Северной Америки и Varanus (Megalania) priscus из плейстоцена Австралии, вдвое превосходившего по размеру своего близкого родственника, комодосского варана. Многие животные, обитающие на островах, проявляют на протяжении истории своей жизни своеобразные адаптации, которые могут быть объяснены при помощи палеогистологических исследований, и среди них моа. Моа были нелетающими травоядными птицами, эндемичными для Новой Зеландии. Среди одиннадцати их видов из шести родов два самых крупных вида, Dinornis robustus и D. novaezealandiae, достигали почти 4 метров в высоту с вытянутой шеей и весили примерно 230 килограммов. Моа принадлежат к бескилевым птицам, которые включают африканского страуса, австралийского эму, новозеландского киви и южноамериканского нанду. Вероятнее всего, в результате охоты маори моа, как полагают, вымерли к 1500 году
78
н. э. Моа демонстрируют явно выраженную «K-стратегию», которая означает долгую
беременность и немногочисленное потомство – что сопровождается длительным
периодом роста и поздним наступлением половой зрелости – в противоположность
другим группам птиц, включая других бескилевых.
В ходе изучения палеогистологии нескольких длинных костей, Сэмюэль Тёрвей
с коллегами зарегистрировали у четырёх из шести родов моа несколько ежегодных
задержек роста, или линий задержек роста, обнаружив, что у этих особей типичный
размер взрослой особи достигался лишь по истечении многих лет прерывистого
роста. Но у гигантских форм моа, у которых самки весили более 200 килограммов,
а самцы до 85 килограммов, наблюдаются особенности гистологии костей, отличные
от характерных для других, более мелких видов. У гигантских форм рост был
ускоренным: наружные слои кости богаты кровеносными сосудами и демонстрируют
лишь немногие ЛЗР, если они вообще есть. Виды Dinornis вырастали
за промежуток времени около трёх лет, при том, что самые мелкие виды моа,
вроде видов рода Euryapteryx, не достигали взрослого размера до девяти
лет. Медленный рост и позднее наступление половозрелости появились в процессе
эволюции в среде, свободной от хищников и, возможно, с ограниченными ресурсами
и сниженной конкуренцией, хотя это лишь предположение. Прибытие людей, как
и в других похожих и печальных случаях, вскоре привело к исчезновению этих
животных, жизненная история которых была неподходящей для сопротивления интенсивному
хищничеству. В настоящее время в Новой Зеландии какапо, единственный в мире
нелетающий попугай, возможно, тоже обречён – в результате разрушения среды
обитания и прочих форм давления со стороны человека.10
Развитие костей и локомоция
у ископаемых видов и у галапагосских игуан
Каждое утро по пути к своему офису я имею удовольствие проходить мимо наших музейных экспозиций, в которых выставлены серии образцов, демонстрирующих рост, или полные скелеты
79
взрослых особей ихтиозавров, плакодонтов, пахиплеврозавров и плезиозавров.
Эти окаменелости прекрасны, и, хотя с таксономической точки зрения они были
далеки от моей исходной специализации, у меня появился интерес к ним. У нас
есть превосходные коллекции триасовых морских рептилий из местонахождения
Монте-Сан-Джорджио в Тичино, италоговорящей области в южной Швейцарии.11
Среди ихтиозавров представлены самые древние свидетельства наличия живорождения
в этой группе. Другие морские рептилии принадлежат к родословной ветви, которая
показывает различные степени приспособленности к жизни в воде. Плезиозавры,
вероятно, были живородящими и полностью ограниченными жизнью в воде, тогда
как некоторые плакодонты были, самое большее, полуводными. Пахиплеврозавры
из Монте-Сан-Джорджио включают различные виды, которые отличаются друг от
друга по своему водному образу жизни. «Пахи» (как их неофициально именуют)
выглядят очень изящными. Голова и всё тело у них удлинены, а конечности обычно
сохраняются в положении, в котором они, вероятнее всего, находились во время
движения, а именно, прижатыми к телу. Вероятнее всего, они питались рыбой
и кальмарами. Если основываться на форме и сочленении их костей, то очевидно,
что они двигались вперёд с помощью волнообразных движений, и при этом хвост,
должно быть, играл важную роль. Микроструктура костей также важна для понимания
локомоции у этих животных; вес и балласт должны играть основную роль. Кости
у пахиплеврозавров были тяжёлыми, потому что они были плотными – это служило
для противодействия подъёмной силе со стороны лёгких и таким образом облегчало
ныряние и поиск корма под водой. Фактически, обиходное немецкое название группы
«Dickrippensaurier» означает «толсторёберная ящерица», как и название
«пахиплеврозавр», которое также может быть переведено как «толстый плеврозавр»
или «толстобокая ящерица». Вопрос, который представляет большой интерес –
как развивались кости этих животных.
Ясмина Хьюджи из моей лаборатории исследовала эту тему, изучая гистологию
ростовой серии четырёх видов пахиплевро-
80
завров. Ясмина обнаружила, что после выклева из яйца происходит непрерывная минерализация костей, которой управляют три процесса:
1. Обызвествлённая хрящевая сердцевина никогда не рассасывается полностью, поэтому на любом этапе обнаруживается лишь небольшая медуллярная полость, или же её нет совсем.
2. Отложение кости начинается одновременно в различных местах в пределах хрящевой сердцевины.
3. Наружный слой кости увеличивается в толщину благодаря непрерывному нарастанию при минимальной или совсем не протекающей резорбции внутреннего слоя кости, формирующейся вокруг основного стержня, надкостницы.
Как сравнить «пахи» с ныне живущими аналогами? Изучение современных видов всегда важно для интерпретации данных, полученных от окаменелостей. Ныне живущих представителей пахиплеврозавров не существует, но их самые близкие родственники – это группа, к которой принадлежат ящерицы и змеи. Постепенное удлинение тела и редукция конечностей у ископаемых форм много раз происходили в ходе эволюции ящериц. Многочисленные примеры этого можно обнаружить у сцинков (Scincidae), наиболее разнообразной группы ящериц, насчитывающей примерно тысячу двести видов, половина из которых живородящие. Сцинки – наземные животные, но многие из форм, у которых частично или сильно редуцированы конечности – роющие и обитающие в песке животные, где они показывают особый род локомоции, напоминающий плавание. Вот, почему в моей лаборатории мы исследовали развитие скелета различных видов сцинков и сравнивали его с таковым у пахиплеврозавров. Но мы также хотели увидеть, как влияет жизнь в воде на рост костей. Единственная ныне живущая ящерица, которая регулярно плавает и кормится в воде – это галапагосская морская игуана Amblyrhynchus cristatus. Среди близких родственников Amblyrhynchus есть несколько видов наземных игуан, с которыми можно сделать сравнение, в том числе обыкновенный конолоф Conolophus subcristatus.
81
Ясмина Хьюджи исследовала галапагосских и других игуан для своей докторской
диссертации в Цюрихе после того, как мы смогли убедить нескольких кураторов
музеев по всему миру предоставить нам кости конечностей этих животных, чтобы
сделать срезы. Поскольку игуаны, подобно большинству многоклеточных животных,
обладают двусторонней симметрией, исследование, сопряжённое с разрушением
образца, затрагивает одну сторону, но оставляет другую неповреждённой. Я всегда
напоминаю об этом факте музейным кураторам, когда прошу у них разрешение.
Для своих исследований мы берём лишь небольшую пробу в диафизах костей конечностей.
Кости этих игуан многое рассказали нам об адаптации и эволюции процесса роста.
Морская игуана уникальна среди современных ящериц из-за плотности её костей
и истории жизни. Полностью наземные ящерицы демонстрируют равновесие между
процессами нарастания кости и её резорбции, которое позволяет кости расти,
не теряя прочности. У морской игуаны мы зафиксировали замедленный процесс
роста, при котором кости приобретают более высокую плотность. Дальнейшее увеличение
плотности кости происходит благодаря процессам перестройки, которые заменяют
рыхлую волокнистую кость первых лет жизни на более плотную ламеллярную кость.
Гистология костей морской игуаны показывает высокие и постоянные темпы нарастания,
что отражает относительно высокую скорость метаболизма по сравнению с другими
игуанами.
Скелетохронология, основанная на подсчете маркеров роста и циклов роста между
ними, позволяет сделать оценку возраста достижения половой зрелости и минимального
возраста на момент смерти. Скелетохронологические данные игуан были соотносимы
с информацией об истории жизни, полученной в ходе полевых исследований. Подобно
другим крупным игуанам, морская игуана достигает половой зрелости достаточно
поздно; самки достигают половой зрелости в возрасте от трёх до пяти лет, тогда
как самцы половозрелы в возрасте от шести до восьми лет. Кости и их линии
задержек роста показали ожидаемую картину. Соответствие между данными, полученными
при изучении от костей, и полевыми данными от экологов, ещё раз убедило нас
в верности выводов.
82
Рисунок 19. Микроструктура длинных костей взрослого самца морской игуаны Amblyrhynchus cristatus (экземпляр из Музея естествознания в Берлине, NKMB 30260). A: Поперечные срезы диафиза лучевой кости, показывающие толстый и неваскуляризированный наружный слой. B: Группа морских игуан, греющихся на берегу (фотография Урсины Коллер). C: Срез лучевой кости при большом увеличении в нормальном проходящем свете. Ежегодно образующиеся линии задержки роста (ЛЗР) отмечены белыми стрелками, а ЛЗР, появлявшиеся на протяжении года, отмечены серыми стрелками. D: Срез при большом увеличении в поляризованном свете. Заметен чётко выраженный узор из светлых и тускнеющих полос, соответствующих циклам роста (тёмные и светло-серые зоны). Изображения любезно предоставила Ясмина Хьюджи. |

Мы обнаружили коренное различие между морской игуаной и пахиплеврозаврами. У игуаны образуется медуллярная полость, а эффект балласта достигается за счёт дополнительных (образованных надкостницей) слоёв окостенения. У пахиплеврозавров, напротив, медуллярная полость отсутствует, а высокая плотность кости дополнительно возникает за счёт минерализованного
83
Рисунок 20. Поперечный срез плечевой кости, показывающий плотность кости и линии роста у взрослого пахиплеврозавра Neusticosaurus pusillus из триаса Монте-Сан-Джорджио, Швейцария (Палентологический институт и музей, Цюрихский Университет, T4178). Фото K. Waskow, в нормальном проходящем свете. Образец показывает шесть линий задержки роста (белые и серые стрелки). Возраст наступления половой зрелости оценивается в три-четыре года из-за резкого уменьшения ширины четвёртого цикла роста (единственная серая стрелка). Две самых наружных ЛЗР расположены близко друг к другу. Медуллярная полость полностью заполнена обызвествлённым хрящом, который подвергся частичной перестройке. Изображение любезно предоставила Ясмина Хьюджи. |
хрящевого центра. Это сравнение современных животных с ископаемыми – ещё один пример изменчивости механизмов развития, когда они могут быть результатом различной степени адаптации к водным местообитаниям и различных «отправных точек» (различного состояния соответствующих предков) в процессе эволюции.
84
Кости, клетки и гены
Это может показаться невероятным, но данные палеогистологии могут давать
косвенную информацию о геноме вымерших позвоночных. Это возможно потому, что
у ныне живущих видов существует строгая корреляция между размером клетки и
размером генома. Размер клетки может быть количественно определён в тонких
палеогистологических срезах кости путём измерения полостей остеоцитов, мест,
которые при жизни организма занимали клетки, ответственные за формирование
кости. Такие оценки были сделаны для десятков видов динозавров, для птерозавров
и для некоторых других вымерших пресмыкающихся и палеозойских наземных позвоночных.
Достоверность этих оценок ограничена, поскольку просто размер сам по себе
– это очень упрощённый аспект характеристики генома, но он, однако, представляет
важность при проведении общего сравнения групп живых организмов.
Сообщается, что размер генома коррелирует с темпом метаболизма у четвероногих
позвоночных, обладателей мускулистых конечностей, обитателей прибрежной полосы
или суши. Клетки меньшего размера более эффективны энергетически и позволяют
поддерживать более высокую скорость метаболизма, которая, в свою очередь,
связана с гомеотермией. В качестве рекомендации стоит более тщательно исследовать
эти отношения у ныне живущих родственников изучаемых групп, и следует достичь
гораздо большего понимания значения размера генома, прежде чем будет раскрыт
полный потенциал палеогистологии в этой области. Соотношение между эволюцией
размера генома и морфологическим разнообразием далеко от простой связи, и
даже одна только дупликация гена, которая, как считали когда-то, должна тесно
коррелировать с морфологическим разнообразием, не имеет прямого отношения
к сложности в целом. Генные дупликации – это удвоение части генома, создание
добавочных копий уже существующих генов, в результате чего у новых копий возникает
потенциальная возможность получения новых функций.12
85
Природа зубов
Жорж Кювье похвалился, что он может реконструировать целое млекопитающее
только по одному зубу. Это, конечно, преувеличение, но, вероятно, любой палеонтолог-специалист
по млекопитающим при наличии выбора предпочел бы взять в качестве источника
информации о вымерших видах зуб, а не какую-то другую часть тела. Зубы – это
деталь строения позвоночных животных, которая благодаря высокому содержанию
минеральных веществ сохраняется в ископаемом состоянии гораздо чаще, чем остальная
часть скелета. Эмаль действительно является самым твёрдым веществом в теле
млекопитающего. Это происходит потому, что эмаль содержит гидроксиапатит в
большем количестве в единицу объёма, чем кость.
Ряд серия окаменелостей документирует изменения скелетных тканей в самый ранний
момент эволюционной истории позвоночных. Происхождение зубов вызывает споры
по нескольким причинам. Одна из них – то, что взгляды на эволюционное древо
включённых в этот процесс животных, одних из самых примитивных позвоночных,
всё ещё не согласованы, и в результате этого возможны альтернативные реконструкции
последовательности событий. Фактически, некоторые авторы, основываясь на ревизии
анатомии и родственных связей плакодерм, панцирных челюстноротых рыб, которые
были очень разнообразны в девонский период, утверждают, что зубы, вероятно,
появлялись в процессе эволюции больше, чем один раз.13
Но перед решением этого вопроса следует обратиться к ещё более фундаментальному
вопросу: чем в действительности является зуб? Не удивительно, что, чем глубже
мы проникаем в тайну происхождения позвоночных, тем менее чёткими становятся
определения. Так происходит из-за того, что не все особенности, например,
местоположение зубов в черепе и связанных тканях, возникли одновременно или
в форме, существующей у уже изученных ныне живущих видов. У ранних позвоночных
не существует чёткого характера распределения зубных тканей. Конодонты, вероятно,
обладали эмалью (этот вопрос ещё не решён окончательно), но у других бесчелюстных
поз-
86
воночных была ткань, называемая энамелоидом.14
Энамелоид должен был существовать у последнего общего предка челюстноротых
позвоночных, и эта и другие зубные ткани появились и стали более сложными
в ходе их эволюции.
Другой вопрос заключается в том, связаны ли критерии определения зубов с индивидуальным
развитием, или же являются генетическими, или возможны оба варианта; выяснить
это можно только по ископаемым остаткам. Мойя Смит и её коллеги сделали важное
открытие, касающееся времени и места экспрессии гена sonic hedgehog,
известного, помимо всего прочего, как ключевой регулятор индуцирования развития
зубов. Исследования ими работы этого гена у кошачьей акулы показали, что,
как и у мышей, места экспрессии соотносятся по времени и месту со временем
и местом образования зубов и зубных бугров. Факт наличия общего механизма
этого процесса у этих отдалённо родственных видов даёт основание полагать,
что такой тип гена, отвечающего за развитие, был у последнего общего предка
современных челюстноротых позвоночных. Но положение дел тут гораздо сложнее.
Как показали Юкка Йернвалл и его сотрудники, зубы являются полигенным образованием,
то есть, на форму зуба оказывают воздействие мутации многих генов. Это особенно
справедливо для млекопитающих, что иллюстрируют собою мыши, являющиеся в этом
отношении классической моделью для исследований и обладающие мутациями более
чем тридцати генов, включённых в этот процесс. Генетические механизмы не просты,
и между видами должны существовать определённые различия. Эти факторы невозможно
изучить непосредственно по окаменелостям.
Типичный зуб позвоночного состоит из тканей, которые происходят из различных
зародышевых листков или клеток раннего эмбриона.15
Две ткани являются самыми основными: эмаль, которая происходит из эктодермы,
самого наружного слоя клеток эмбриона, и дентин, который образуется из нервного
гребня. Клетки нервного гребня образуются у раннего эмбриона, сосредотачиваясь
вдоль дорсальной средней линии нервной трубки в виде цепочки, тянущейся в
передне-заднем направлении, и оттуда они мигрируют для образования множества
видов тканей, в том числе дентина зубов. Развитие зуба начинается с взаимодействия,
образования складок и дифферен-
87
циации двух слоёв в коже, так же, как в случае чешуй, волос и перьев. Некоторые аспекты развития этих структур гомологичны с точки зрения анатомии, поскольку в их основе лежат общие механизмы развития и генетические основы. Эти механизмы должны присутствовать у животного, которое было последним общим предком видов, обладающих зубами и специализированными особенностями покровов тела, в том числе чешуёй, перьями и волосами.
Зубы и история жизни
Ископаемые зубы могут дать некоторые подсказки к распределению во времени
событий из истории жизни индивидуума. И не случайно зубы являются предпочитаемым
объектом при исследовании онтогенеза. Морфология зубных бугров развивается
в направлении от кончика к корню, путём своего рода приращения, напоминающего
характер роста раковин моллюсков или рогов жвачных (группа парнокопытных,
которая включает коров, жираф и оленей). Последовательность прорезывания зубов
также коррелирует с событиями в истории жизни, о чём я говорю в главе
8, касаясь наших человеческих предков.
Зубы млекопитающих были благодатным объектом для исследований истории жизни.
В конце 1960-х годов объектом внимания стало использование слоёв в зубном
цементе и дентине для оценки возраста. Эти слои откладываются с регулярной
цикличностью, и те из них, которые связаны с долговременными, повторяющимися
из года в год сезонами, можно подсчитать, чтобы получить значение абсолютного
возраста индивидуумов. Слои в дентине и цементе различимы при стандартном
микроскопическом исследовании с использованием простых гистологических методов.
Различия в скорости метаболизма, отчасти определяющиеся сезонными изменениями
в окружающей среде, вызывают периодичность в отложении слоёв. Зубные маркеры
обычно представлены в виде толстых слоёв, отложившихся в течение сезона сравнительно
быстрого роста («лето») и сезона ограниченного роста и, следовательно, более
тонких полос. Цемент и дентин ред-
88
Рисунок 21. Смена зубов (указано стрелкой) у ископаемого носорога Brachypotherium brachypus из среднего миоцена Эрликона, кантон Цюрих, Швейцария. Фотография любезно предоставлена Палеонтологическим музеем Цюрихского Университета. |

ко становятся объектом перестроек или резорбции, и по этой причине в силу
многих обстоятельств могут быть лучшим скелетохронологическим инструментом,
нежели кости, которые подвергаются значительным изменениям и разрушению в
процессе роста.16
Существуют многочисленные примеры исследований истории жизни ископаемых млекопитающих;
некоторые из этих вымерших видов сосуществовали с людьми в не столь далёком
прошлом, как мамонты и большерогие олени. Состоящие из дентина бивни мастодонтов
и мамонтов построены из слоистых образований, которые, подобно другим зубам,
нарастали путём приращения. Таким образом, их анализ может дать информацию,
касающуюся возраста, скорости роста, возраста наступления половой зрелости
отдельных животных и характера смертности в популяции. Такого рода работу
провёл Дэн Фишер, который также изучал контекст доисторической окружающей
среды,
89
используя анализ стабильных изотопов в бивнях. С помощью таких методов можно
получить информацию о температуре и рационе в разное время жизни животного.
В недавнем исследовании изучение гистологии зубов большерогого оленя Megaloceros
(к сожалению, вымершего) установило, что посмертный возраст для нескольких
особей из ирландской популяции составлял от восьми до четырнадцати лет. Это
похоже на средний посмертный возраст большинства других видов оленей. Крупный
размер Megaloceros, особенно его большие рога, а также необычная
плотность его черепа дали бы основание предположить для этих животных долгую
жизнь, соотносимую с ожидаемым продлением времени роста по сравнению с его
предками. Оценки возраста, которые были получены в ходе изучения зубов, противоречат
этому предсказанию. И вновь мозаика из скорости и характера эволюции процессов
развития разных частей тела животного делает сравнительные исследования истории
жизни более сложными и интересными.17
Износ зубов
Морфология зубов может радикально изменяться на протяжении времени жизни
индивида, в некоторых случаях вплоть до исчезновения бугров и обнажения тканей
зубов, изначально покрытых эмалью, когда индивид был моложе. Степень износа
можно использовать для определения относительного возраста ископаемых организмов,
но этот подход имеет множество неудобств, в том числе изменчивость в скорости
и характере износа зуба даже в пределах популяции, а также в зависимости от
местности.18 Существует
две причины износа зубов: абразивное воздействие, производимое пищей или частицами,
которые её сопровождают, и истирание, вызванное контактом между зубами.
В случае позвоночных с многократной сменой зубов простая замена – это очевидная
стратегия для борьбы с износом.
90
Рисунок 22. Нижний зубной ряд трёх
особей Dremotherium feignouxi, безрогого жвачного из местонахождения
Монтегю-ле-Блин (Бассейн Алье, центральная Франция) раннемиоценового
возраста. Обратите внимание на значительные изменения формы зубов из-за
износа в течение жизни. |

Но млекопитающие, для которых характерна максимум двукратная смена зубов, в процессе эволюции приобрели альтернативные стратегии. Травоядные особенно часто имеют дело с абразивным воздействием пищи, и долговечность зубов является критически важным моментом для борьбы с их износом. Для этого в процессе эволюции много раз и в различных группах возникали стратегии, суть которых состоит в появлении более высоких и даже постоянно растущих зубов. Изучение характера износа зубов имеет давнюю традицию в палеонтологии млекопитающих, и в ходе исследований затрагиваются также аспекты истории жизни и вопросы функционального плана. В этом отношении были исследованы даже некоторые из самых ранних млекопитающих, такие, как Haldanodon exspectatus из поздней юры Португалии, и было показано, что их рацион проявлял абразивные свойства. Исключительный пример адаптации, призванной компенсировать износ зубов, был зарегистрирован у Hyaenodon, млекопитающего из отряда креодонтов, которое жило на протяжении длительного времени в кайнозое и имело широкое распространение на нескольких континентах. Hyaenodon был плото-
91
ядным и обладал режущими зубами, среди которых верхний и нижний хищные зубы должны были тесно контактировать друг с другом, чтобы осуществлять функцию резания. Чтобы сохранить эту тесную подгонку зубов вопреки их износу, происходило поворачивание верхних зубов в медиальном направлении. Это явление было выражено очень явно: верхние зубы у самых старых особей повёрнуты под углом почти в девяносто градусов, а эмалевая коронка полностью стёрта. Эта специализация, очевидно, помогала увеличить среднюю продолжительность жизни Hyaenodon.19 Некоторые виды ископаемых плотоядных плацентарных и сумчатых млекопитающих также обладают такой специализацией.
Содержание
Благодарности vii
Пролог xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы
под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания 197
Библиография 213