| Главная | Библиотека | Форум | Гостевая книга |
126
Глава седьмая
Ископаемые остатки и
генетика развития
Онтогенез в ископаемом виде не сохраняется. Но сохраняются структурные образования, которые некогда были результатом процесса развития. В этом случае ископаемые остатки взрослых особей могут дать информацию о развитии благодаря сохранению фенотипов, обладающих непосредственной и чёткой корреляцией с определённым процессом, протекающим в ходе индивидуального развития. Чтобы реконструировать такой процесс, важно изучить положение ископаемой формы на эволюционном древе жизни, чтобы быть уверенным в том, что исследования основаны на правильном предположении. Этот подход называется заключением в современные филогенетические скобки*. Он был введён специально для того, чтобы можно было делать вывод об анатомии мягких тканей и создавать реконструкции поведения для ископаемых форм, но его также можно использовать для реконструкции онтогенеза вымерших форм. Вымершее животное сравнивается с его ближайшими ныне живущими родственниками. Исследуется распределение некоторой особенности развития среди существующих в наше время таксонов, которые «окружают» вымерший таксон на данном древе. Если особенность существует у ныне живущих видов, входящих в группу, внутри которой находится изучаемая ископаемая форма, следовательно, можно сделать вывод о том, что она существовала у общего предка, и вследствие этого также у ископаемой формы.
* В оригинале это звучит как «extant phylogenetic bracketing» – прим. перев.
127
Рисунок 33. Динозавры заключены в филогенетические скобки между крокодилами и птицами. |
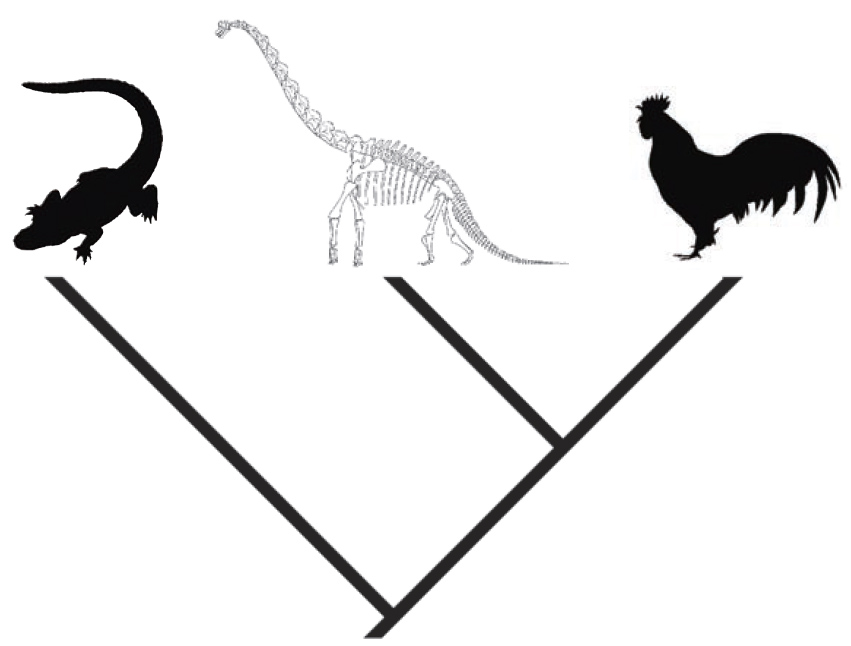
Например, динозавры заключены в «скобки» между птицами и крокодилами. Особенность, имеющаяся и у птиц, и у крокодилов, например, яйцерождение, вероятно, присутствовала бы и у динозавров. С другой стороны, особенность, отсутствующая и у птиц, и у крокодилов, вроде метаморфоза, вероятнее всего, также отсутствовала бы у динозавров. Но дело усложняется, когда у одних из ныне живущих родственников особенность имеется, а у других её нет. Например, крокодилы обладают костями преимущественно ламеллярного типа, которые коррелируют с медленным ростом, тогда как птицы обладают фиброламеллярными костями и способны к быстрому скачкообразному росту. А как обстояли дела у динозавров? В таких неоднозначных случаях, а также для проверки выводов, основанных исключительно на использовании метода филогенетических скобок, особенно важным является рассмотрение случаев сохранения деталей, которые представляют прямые свидетельства существования той или иной особенности. В случае с процессом роста у динозавров исследования гистологии костей дали прямую и определённую информацию о
128
наличии фиброламеллярной кости и своеобразных кривых роста со стадией быстрого
роста у некоторых групп этих животных.
Можно поставить под вопрос научную ценность выводов, сделанных в отношении
организмов, отделённых от нас несколькими сотнями миллионов лет независимой
эволюции. Но такие выводы подтверждены тем, что они основаны на эмпирических
свидетельствах, собранных на материале ныне живущих организмов. Крупное открытие
нескольких последних десятилетий состоит в том, что многие механизмы индивидуального
развития и молекулы остаются одинаковыми для таксонов, не сравнимых друг с
другом напрямую и отдалённо родственных филогенетически. Известно, что многие
молекулярные процессы, которые контролируют изменение фенотипа, сами по себе
намного старше, чем группа, у которой эти фенотипы проявляются; Шон Кэрролл
назвал это явление «предковой сложностью». Этот консерватизм имеет место у
очень непохожих друг на друга и весьма отдалённо родственных видов. Это ставит
нас перед фактом наличия глубоких, или фундаментальных уровней, находящихся
на ранних отрезках геологического времени и времени индивидуального развития,
до которых может быть отслежена история структурных образований, имеющихся
у взрослых особей таких отличающихся друг от друга видов, как мухи и люди.
В этом случае мы говорим о «глубинной гомологии» этих образований, подразумевая
«тождество», или гомологию, на некотором уровне организации (молекулярном,
клеточном) и на некотором уровне иерархии на эволюционном древе жизни.
Может показаться, что использование настоящего для того, чтобы делать выводы
в отношении прошлого, как в случае с «филогенетическими скобками», накладывает
на него свои ограничения. По сути, эволюционный биолог Марк Пэйджел пишет,
что этот подход «обрекает прошлое на сходство с настоящим. И, возможно, ещё
хуже то, что прошлое, картину которого мы получаем, оглядываясь назад – это
очень обычное прошлое, усреднённое прошлое». Но у нас нет выбора, и это не
означает того, что мы не обнаруживаем важных, и даже неожиданных примеров,
когда рассматриваем окаменелости таким образом. И вообще, что такое «обычное»
или «усреднённое»? Не существует ни одного современного трилобита, но есть
все основания, чтобы полагать, что разделение тела на сегменты, которое они
претерпевают в ходе индивидуального разви-
129
тия, было связано с теми же самыми процессами, какие выявлены у ныне живущих
членистоногих.
В приведённых ниже примерах я постарался продемонстрировать, как мы можем
изучать эволюцию индивидуального развития, рассматривая фенотипы взрослых
особей вымерших видов.
Количество позвонков
Сложные животные состоят в значительной степени из повторяющихся частей,
и позвоночные здесь не являются исключением. Это не столь очевидно на первый
взгляд, но, если мы исследуем наш собственный скелет или скелет любого другого
позвоночного, то мы увидим последовательное повторение позвонков, лишь плавно
отклоняющихся друг от друга по размеру и по форме. Ткани, окружающие эти позвонки,
также образуют повторяющиеся структуры, в том числе мышцы, нервы и сосуды.
Эти «пакеты» анатомических структур, эти сегменты целого тела, возникают в
ходе индивидуального развития из строительных блоков, или сомитов. Сомиты
образуются на протяжении особого отрезка времени раннего эмбрионального развития.
Скорость сегментации определяет в этом случае, сколько позвонков будет иметь
взрослый организм. У некоторых змей развивается около трёхсот сегментов, тогда
как черепахи останавливаются примерно на двадцати пяти, со строго определённым
количеством, восемнадцатью, перед крестцом (часть позвоночного столба, находящаяся
прямо перед костями хвоста). Есть своего рода часы сегментации, и в зависимости
от вида они тикают по-разному. Иногда они тикают дольше или быстрее в пределах
одного вида, особенно в области хвоста.
Наш собственный позвоночный столб легко разделяется на отделы, обладающие
своими отличительными особенностями. У нас нет никаких рёбер в шее, в шейном
отделе. У нас есть грудной отдел с большими рёбрами, которые сходятся
друг к другу, чтобы прикрепиться к грудине, и которые образуют клетку, где
наши сердце, желудок и другие органы надёжно защищены. У нас нет
130
грудной кости или рёбер на животе, поэтому мы должны удерживать органы на
своих местах, приобретая сильную брюшную мускулатуру в нашем поясничном
отделе. Наконец, некоторые позвонки слились, чтобы образовать крестцовый
отдел, к которому прикрепляется таз, или пояс задних конечностей, а за ним
следует каудальный отдел, или хвост. Границы между отделами соответствуют
границам экспрессии специальных генов, которые регулируют процесс развития
и устанавливают признаки отличий отделов, образующихся вдоль оси тела. Эти
гены – члены семейства регуляторных генов Hox.
В этом контексте процесса индивидуального развития вещи, кажущиеся обыденными,
вроде количества позвонков в различных отделах скелета, выходят в новое измерение.
Количественные значения отражают два фундаментальных аспекта раннего этапа
развития: образование сомитов и границы экспрессии генов Hox. Тогда
изменчивость в количестве позвонков должна отражать эволюционную пластичность
этих механизмов. Млекопитающие консервативны в плане количества предкрестцовых
позвонков. И у мыши, и у жирафа имеется семь шейных позвонков. И коала, и
опоссум, которые оба фактически являются сумчатыми, обладают двенадцатью позвонками
в грудо-поясничном отделе, и изменчивость в этой области мала у этой группы
в целом. Рептилии, которые включают птиц, гораздо более разнообразны в этом
отношении. У лебедя есть около двадцати пяти шейных позвонков, а у утки шестнадцать
или меньше. У всех черепах восемнадцать предкрестцовых позвонков – восемь
шейных, десять спинных и два крестцовых – хотя примитивная черепаха Odontochelys
из триаса Китая, как сообщают, обладает лишь девятью спинными позвонками.
Я впервые узнал о существовании этих жёстких шаблонов от Шигеру Куратани.1
Куратани-сэнсей, как я называл его, будучи в Кобэ, в своей жалкой попытке
сработаться с ним, догадался исследовать проблему количества позвонков среди
наземных позвоночных, отобразив в виде карты данные, изложенные в классическом
труде покойного сэра Ричарда Оуэна, современника Чарльза Дарвина. Совместно
с Юичи Наритой
131
он выяснил, какие группы консервативны в плане индивидуального развития,
а какие пластичны. Посетив его лабораторию в 2006 году и обсуждая их анализ,
я заметил в их работе другую своеобразную картину. У большинства млекопитающих
из всех групп сумма числа позвонков в грудной клетке и в области живота, в
грудо-поясничной области, равна девятнадцати. Это количество постоянно и у
сумчатых, и у яйцекладущих однопроходных, и у многих групп плацентарных млекопитающих,
и, вероятно, закрепилось на ранней стадии эволюции млекопитающих. Но, кроме
некоторых других беспорядочно распределяющихся исключений, крупная группа
плацентарных млекопитающих Afrotheria обладает общим признаком – увеличенным
суммарным количеством грудных и поясничных позвонков. Эта картина сохраняется
даже тогда, когда исследуются самые древние и наименее специализированные
ископаемые виды каждой из этих групп. Afrotheria, группа, изначально признанная
лишь на основании молекулярных свидетельств, включает таких разнообразных
существ, как даманы, слоны, дюгони, тенреки и златокроты. После того, как
мы заметили эту картину, Роб Ашер и Томас Лехманн открыли, что задержка в
развитии зубов также является диагностическим признаком для афротериев, как
ископаемых, так и ныне живущих.2
Формирование количества позвонков связано с механизмами, которые у вымерших
видов невозможно изучить непосредственно. Но у количества позвонков существует
однозначное соответствие сегментации и границам экспрессии Hox-генов.
Существующие в наше время филогенетические скобки чётко поддерживают предположение
о том, что эти процессы работают у всех наземных позвоночных, ископаемых и
современных. Тогда богатая летопись окаменелостей наземных позвоночных даёт
нам шанс исследовать индивидуальное развитие у вымерших форм. Таким способом
мы смогли исследовать раннюю историю млекопитающих и рептилий, чтобы увидеть
картину эволюции и ответить на ряд вопросов: Когда в процессе эволюции возник
консерватизм млекопитающих в количестве позвонков? Какой из механизмов был
эволюционно более пластичным – сомитогенез или границы экспрессии Hox-генов?
Какие ограничения
132
накладывают на пластичность механизмов индивидуального развития экологические
адаптации, такие, как вторичноводный образ жизни, наблюдаемый у китов и морских
рептилий?
Чтобы ответить на эти вопросы, мы вместе с Йоханнесом Мюллером и Торстеном
Шейером собрали группу экспертов, которые предоставили актуальную и исчерпывающую
информацию по филогенезу и количеству позвонков для групп, на которых они
специализировались. Обладая этой информацией, мы провели анализ, результатом
которого было выявление различной степени пластичности процессов развития
в эволюции вымерших групп и в ранней истории всех основных ныне живущих групп.
Экспертный анализ ископаемых остатков показал, что увеличение и уменьшение
общего количества позвонков или в границах отделов позвоночника, при сохранении
одинакового общего количества, возникало независимо в нескольких эволюционных
линиях. Мы отследили назад во времени появление канальности и консерватизма
в количестве предкрестцовых позвонков у млекопитающих до момента возникновения
этой группы, по крайней мере, около 315 миллионов лет назад, когда количество
шейных позвонков колебалось между пятью и шестью. Установление почти постоянного
количества шейных позвонков, равного семи, произошло приблизительно на 100
миллионов лет позже, в юрский период. Это открытие подвергает сомнению выдвинутую
ранее гипотезу, пытающуюся объяснить консерватизм, впервые отмеченный только
у ныне живущих млекопитающих. На основании факта высокой частоты смертности
зародышей и молодых особей человека с ненормальным количеством шейных позвонков
для объяснения консерватизма в количестве шейных позвонков у млекопитающих
предполагалось наличие летального плейотропного эффекта, относящегося к мутациям
в области регуляции Hox-генов, которые оказывают воздействие не только
на осевой скелет, но также и на появление клеточных новообразований. Лишь
млекопитающие с низким темпом обмена веществ демонстрируют отклонения от стандартного
количества шейных позвонков, и эта особенность также связана с более низкой
подверженностью раковым заболеваниям. Однако наш результат подвергает сомнению
эту гипотезу. Самые ранние представители эволюцион-
133
Рисунок 34. Tanystropheus longobardicus. Другие реконструкции этого животного изображают его с шеей, которая насчитывает тринадцать позвонков и вытянута перпендикулярно главной оси тела. Из Wild 1974, с изменениями, сделанными Кристианом Клюгом на основе работы Tschanz 1988. |

ной линии, ведущей к млекопитающим, уже были весьма консервативными в плане
количества шейных позвонков, а их физиология, как видно из палеогистологических
данных и других характерных анатомических особенностей, была рептильной. Возможно,
это пример того, что Томас Гексли назвал «убийством прекрасной гипотезы ужасным
фактом».
Другую крупную закономерность в эволюции количества позвонков мы отмечаем
в группах, которые населяют водные местообитания. В этих группах имеет место
отклонение от стандартной картины развития: в процессе эволюции появляется
большее количество предкрестцовых позвонков. Это справедливо для всех рептилий
и млекопитающих. Ослабление регионализации – то есть, когда различия между
группами позвонков исчезают и они становятся похожими друг на друга – похоже,
способствует проявлению большей пластичности в развитии.3
Но в случае водных млекопитающих количество шейных позвонков остаётся постоянным.
Это отличает их от рептилий, у которых общее количество позвонков (посредством
сомитогенеза) также увеличивается, но наряду с ним увеличивается и пластичность
в количестве шейных позвонков, что показывают различные вымершие группы вроде
плезиозавров, у которых много шейных позвонков, или пахиплеврозавры и Tanystropheus,
у которых количество шейных позвонков намного меньше. Картина, наблюдаемая
у динозавров, ясно показывает, что количество позвонков и их распределение
по отделам позвоночника не определяется общим размером, которого достигает
животное во взрослом состоянии: эмбрионы зауропод не показывают высокой скорости
сомитогенеза ради достижения гигантского размера, характерного для этой группы.
134
О Saurichthys и рыбах-мутантах
Я обсуждал главным образом то, как палеонтологические данные могут дополнить
наше понимание эволюции истории жизни и механизмов индивидуального развития.
Но могут данные из области эволюции индивидуального развития сообщить что-либо
палеонтологам? Могут ли открытия из области молекулярной эволюционной биологии
развития внести свой вклад в понимание разнообразия вымерших таксонов? Поскольку
нуклеиновые кислоты и белки очень редко сохраняются с течением геологического
времени, реконструкция молекулярной основы морфологических изменений у ископаемых
форм должна основываться на умозаключениях.
Здесь я исследую основания для таких выводов, рассматривая их на примере ископаемой
рыбы.
Среди богатых коллекций среднетриасовых позвоночных в Цюрихе есть замечательная
рыба. Внешне она выглядит похожей на современную щуку, с удлинённой и обтекаемой
формой тела, которая выдаёт в ней хищника. Её название – Saurichthys,
«рыба-ящерица». Этот род располагается довольно близко к основанию родословного
древа костных рыб, близко к осетрам, веслоносам и ильной рыбе, своим ныне
живущим родственникам. Существуют четыре вида, известных из местонахождения
в Монте-Сан-Джорджио в Швейцарии, и они выставлены в Цюрихе, но род имеет
всесветное распространение и включает более тридцати видов. Те виды, которые
имеются в нашем музее, наиболее очевидно характеризуются различным рисунком
и количеством чешуй, как было установлено в 1980-х годах Оливье Риппелом,
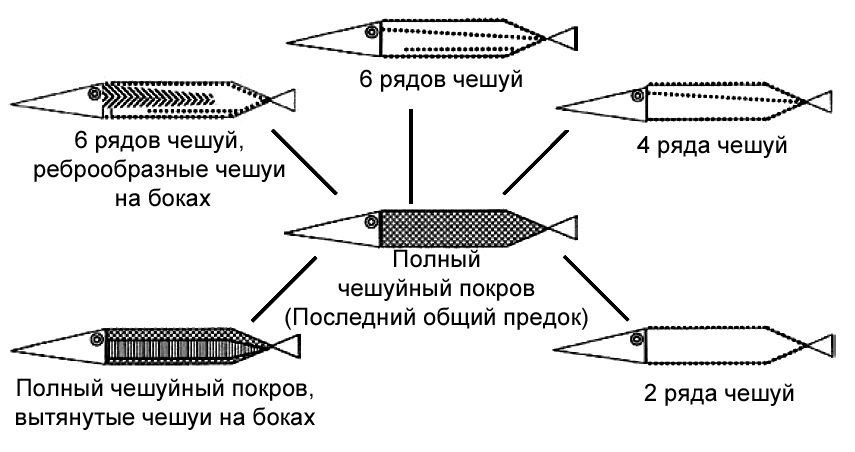
когда он работал в Цюрихе. Гипотетический последний общий предок, давший начало
радиации видов Saurichthys, может быть уверенно реконструирован на
основании нескольких других окаменелостей как полностью покрытый однородной
ромбовидной чешуёй, с многочисленными, сильно сегментированными плавниковыми
лучами и с большим количеством дермальных костей в черепе. Виды рода Saurichthys,
происхождение которых можно отследить назад к этому гипотетическому предку,
характеризуются различной степенью редукции чешуйного покрова (ряды
135
и вытянутая форма чешуй), редукцией количества плавниковых лучей и их сегментов,
сокращением количества дермальных костей в черепе и их формой, отражающейся
в форме жаберной крышки, а также снижением степени массивности зубов и челюстей.
Фактически, новизна в эволюции во многих случаях ассоциируется с утратой структур.
Хорошие примеры этого явления – общая редукция экзоскелета во многих родословных
линиях и утрата тяжёлой брони у силурийских и девонских рыб.4
Но нам пока нет необходимости возвращаться во времени так далеко. Утрата или
редукция чешуй у других рыб не представляет собой ничего необычного. Например,
у осетров и веслоносов (отряд Осетрообразные) полный чешуйный покров, зарегистрированный
у образцов из позднего мела, редуцировался до пяти рядов щитков у ныне живущих
видов*. Утрата чешуй независимо происходила в процессе эволюции, по крайней
мере, тринадцать раз у гольянов и карпов (карпообразные) и несколько раз у
галаксиевых, саланксовых (корюшкообразные) и икромечущих карпозубых (карпозубообразные).
В пределах некоторых видов возможна значительная изменчивость. Одомашненный
карп Cyprinus carpio существует в виде различных разновидностей:
кожистый карп с фактически отсутствующей чешуёй; зеркальный карп с немногочисленными
увеличенными чешуями и зеркальный линейный карп с единственным рядом крупных
чешуй, тянущимся вдоль боков. Эти примеры изменчивости и эволюционных изменений
вызвали интерес к расшифровке генетических механизмов, лежащих в основе явления
редукции чешуи.5
Известны сотни образцов Saurichthys, и изучение некоторых из них
привело к открытию живорождения у этого животного. Я всегда думал, что когда-нибудь
я бы хотел изучить эту рыбу и поработать с ней, но это случилось довольно
неожиданным образом. Генетика развития и любезный и вежливый отставной доктор
медицины из Базеля Лиени Шмид являются ключевыми фигурами в этой истории.
В последние годы Лиени Шмид присоединился к Рудольфу Стокару и его коллегам
в Лугано, собирая Saurichthys в триасовых породах в южной Швейцарии.
Он также посещал курсы эво-дево, на
* У веслоноса чешуя отсутствует или имеются лишь отдельные чешуйки. – прим. перев.
136
Рисунок 35. Схемы, показывающие разнообразие картины чешуйного покрова у Saurichthys. Последний общий предок должен представлять собой рыбу, полностью покрытую однородной ромбической чешуёй. По Schmid и Sánchez-Villagra 2010, с изменениями. |
которых обсуждались самые новые работы, связывающие морфологическую эволюцию
с генетикой развития. Среди них были исследования некоторых видов рыб, которые
хорошо известны с точки зрения генетики и служат моделями для понимания эволюционных
изменений. Шмид заметил сходство в характере морфологических изменений, представленных
в исследованиях генетики развития видов рыб, с изменчивостью у Saurichthys.
Он рассказал мне о своих идеях, и мы исследовали вопрос глубже и написали
об этом работу, которую я вкратце излагаю здесь.
Серия изящных исследований, которые включены в учебники, иллюстрирует генетические
и фенотипические изменения на популяционном уровне в экологическом контексте.
Лучше всего известный пример – это поразительная изменчивость в строении чешуйчатого
покрова у колюшек.6
У современных видов был зарегистрирован быстрый рост морфологического разнообразия,
порождённый изменением регуляторных функций и взаимодействием между генами,
экспрессия которых происходила посредством изменяющихся экологических условий.
Трёхиглая колюшка (Gasteros-
137
Рисунок 36. Образец Saurichthys curionii из триаса Монте-Сан-Джорджио, Швейцария (Палеонтологический институт и музей, Цюрихский Университет). |
teus aculeatus) несёт на своих боках ряд из примерно 36 пластинок
– видоизменённых чешуй. В пресной воде последовательно отбиралась мутация
эктодисплазинового сигнального пути (совокупности взаимодействий на молекулярном
уровне), которая уменьшала количество этих видоизменённых чешуй с 36 до 0
– 7 в гомозиготном состоянии и до промежуточного количества в гетерозиготном.
Молекулярные генетические факторы, ведущие к утрате чешуйчатого покрова, были
изучены в лаборатории и в полевых условиях на колюшках, а также на мутантах
нескольких видов рыб.
Существуют другие мутанты по эктодисплазиновому сигнальному пути. На примере
японской медаки (Oryzias latipes) было продемонстрировано, что мутант
с утратой чешуи возник из-за мутации в рецепторе этого пути. У данио-рерио
(Danio rerio) было идентифицировано несколько мутаций, как в сигнальной
молекуле самого эктодисплазина, так и в рецепторе, которые приводили к появлению
фенотипов со сходными типами утраты чешуи. Другой молекулярный механизм, приводящий
к утрате чешуи, был обнаружен у мутанта данио-рерио и у домашних карпов. Причиной
этого является мутация утраты функционирования в рецепторе-1 фактора
138
роста фибробластов. Известна важность факторов роста фибробластов на всех
стадиях развития. Жизнеспособный мутант с мутацией этого пути возможен только
благодаря дублированию гена, поскольку летальные побочные эффекты мутации
в одном гене компенсируются немутированной версией этого же гена, которая
в терминологии генетиков носит название «паралог».
Мы с Шмидом постулировали, что подобные генетические процессы лежат в основе
фенотипических изменений у Saurichthys и у мутантов этих рыб. Молекулярные
основы морфологического разнообразия Saurichthys включали мутацию
или регуляторные изменения сигнального пути в качестве ключевого механизма.
Существуют серьёзные основания полагать, что эти механизмы действовали у триасовых
рыб так же, как в наши дни.
Анализ последовательности генов, кодирующих компоненты эктодисплазинового
сигнального пути, показывает, что эти гены у позвоночных очень консервативны.
Известно, что эктодисплазин контролирует формирование зубов и дермальных костей
у челюстноротых позвоночных, чешуи у рыб, перьев у птиц и волос и кожных желёз
у млекопитающих. Что же касается факторов роста фибробластов, то их работа
существенна на очень ранних стадиях развития и они явно унаследованы по меньшей
мере всеми позвоночными животными. Эти сигнальные пути представляют собой
пример того, что названо «инструментальными белками»* и хорошо сохраняется
в ходе эволюции.
В дополнение к редукции количества чешуй, вышеупомянутые виды рыб благодаря
мутации в эктодисплазиновом пути или в пути фактора роста фибробластов демонстрируют
тенденцию к выстраиванию оставшихся чешуй в виде рисунка, схожего с тем, который
наблюдается у отдельных рядов чешуй у Saurichthys. Кроме того, сохраняющиеся
чешуи на боках мутантов вытянуты в дорсовентральном направлении так, что их
можно сравнить с удлинёнными боковыми чешуями, как у видов Saurichthys
с реброобразными боковыми чешуями. Мутации в компонентах
* В оригинале названы «tool kit proteins» – прим. перев.
139
Рисунок 37. Схематичные изображения рыб дикого типа и мутантов различных модельных видов, используемых в исследованиях в области генетики развития. Эктодисплазиновый путь: данио-рерио и японская медака, по Harris и др. 2008; трёхиглая колюшка, по Colosimo и др. 2005. Путь фактора роста фибробластов, по Rohner и др. 2009. Рисунки сделаны Л. Шмидом (Базель), из Schmid и Sánchez-Villagra 2010, с изменениями. |

140
эктодисплазинового пути также ведут, по крайней мере, у некоторых мутантов
данио-рерио, к сокращению количества и снижению сложности строения плавниковых
лучей, как было отмечено для некоторых морфологических типов Saurichthys.
Кроме того, были сообщения об утрате зубов и изменениях размеров дермальных
костей у мутантов данио-рерио, опять-таки изменения органов, которые менялись
в процессе роста видового многообразия у Saurichthys.
Особенности строения, контролируемые инструментальными белками, в том числе
чешуйчатый покров, строение плавниковых лучей и особенности зубной системы,
являются особенно важными для целей самозащиты, хищничества и питания, и таким
образом для адаптации в целом. Изменение в важном гене, определяющем ход развития,
даёт хороший шанс для симпатрического видообразования. Чем больше будет фенотипический
эффект единственной мутации, тем больше шанс немедленной репродуктивной изоляции
мутантов, если они окажутся жизнеспособными и успешными. Изменения в сигнальных
путях, наличие которых мы предполагаем в случае увеличения видового многообразия
у Saurichthys, могут, таким образом, дать объяснение явлению очень
быстрого видообразования. В противоположность градуалистической модели эволюции,
изменение в важном гене, контролирующем развитие, представляет собой реалистичное
объяснение существенных различий между видами рода Saurichthys в
палеоэкологическом контексте, в котором они возникли. Тот факт, что не были
обнаружены промежуточные, плавно отличающиеся друг от друга ископаемые формы
Saurichthys и большинства других видов, хорошо вписывается в общую
картину.
Содержание
Благодарности vii
Пролог xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер роста и процессы
увеличения многообразия
105
7. Ископаемые
остатки и генетика развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания 197
Библиография 213