Bestiarium (or, to be more exact, Herbarium) of the future - Plants


Bestiarium (or, to be more exact, Herbarium) of the future - Plants |
|
| In this section descriptions of various animals and plants, which could live on the Earth in Neocene epoch. The section will be supplemented as new ideas about possible ways of evolution of life will appear. If readers will not find here any species placed here earlier, it means that it is a reason to search for a new chapter in English version of "The Neocene Project". | |
|
Plants |
Dicotyledon plants |
Carpet
water lily (Nataronymphaea pustulosa)
Order: Nymphaeales (Nymphaeales)
Family: Nymphaeaceae (Nymphaeaceae)
Habitat: rivers of Central Africa.

Picture by Carlos Pizcueta (Electreel)
Climatic changes of boundary of Holocene and Neocene have affected
tropical flora. The area of woods was reduced, and thereof some rivers have
dried up completely. Later, when conditions in biosphere were normalized, the
climate became more damp and steady, and plants have started to evolve actively.
Among them there were species mastering new ecological niches.
Nymphaeas family to which water lilies, candocks, and some other plants belong,
from the moment of occurrence at the end of Mesozoic took a convenient place
of shallow water plant. But in Neocene among them one unique plant, which has
completely lost connection with bottom, has appeared. It is carpet water lily,
filled up numbers of plants freely floating on water surface. Thickets of this
plant species cover surface of lakes and shallow rivers of Africa from Sahara
Nile up to Okavango Lake.
Carpet nymphaea is floating plant of the small size: diameter of crown is at
all about half meter (at some of its relatives only one leaf may reach such
size!). Leaves are rounded, growing on short petioles. Tissue of leaves is thick
and spongy; intervals between nerves are strongly thickened and also filled
by lacunose parenchyma, forming air bubbles (leaves look as if “quilted”). It
permits a plant to keep swimming. The basic stalk is short, covered with thick
spongy crust. At reservoir drying it falls to ground, and sustains drought.
Roots are long; they hang down up to meter depth. They serve to the plant for
maintenance of balance, and also for nutrients absorbing. Roots are covered
with long hairs and look fluffy. At plants floating near of coast, roots grow
into silt, and plant starts to develop more intensively.
Similarly to all floating plants, the carpet water lily breeds mainly by sarments,
on which ends young plants develop. For one year from one plant theoretically
it can turn out up to three hundred new ones.
Vegetative multiplying is not a unique way of breeding of this plant. The carpet
water lily easily and quickly breeds by seeds. It flowers frequently and plentifully.
Flowers are small (diameter of flower is up to 5 – 6 cm) and numerous – up to
10 ones and more at the plant. They rise above water surface a little on short
peduncles. Flowers are brightly colored and well appreciable from afar: petals
are pink, and stamens are yellow. They are pollinated by bees and small beetles.
Seeds are covered with layer of spongy tissue, therefore they do not sink. Under
layer of spongy tissue there is a dense environment due to which seeds can endure
a long drought. They quickly sprout only in fresh rain water. If in water there
is a plenty of products of decay, and its reaction is acidic – it is a signal
of drought approaching, and seeds stay in condition of rest. Even if adult plants
will be lost, the part of seeds remains, and the population of carpet water
lily will be restored from them. Sprouts quickly start vegetative breeding –
having formed four leaves, the young plant already forms sarment.
Lace-leaved
buttercup (Ranunculus lacinifolius)
Order: Ranunculales (Ranunculales)
Family: Buttercups (Ranunculaceae)
Habitat: mountain streams and rivers of the east of North America.
Being older mountains in comparison with Rocky Mountains at the west, Appalachian
Mountains have rather non-uniform geological structure – they are destroyed
by time in greater degree, and in northern part by ice sheet of Holocene-Neocene
congelation. In these mountains there is a lot of shallow streams and rivers
with slower and less destructive current. Sometimes, if the district has a small
slope, rivers can spill, forming wetland areas with slow current. In such places,
amphibious plants grow plentifully, entrapping silt and sand by their roots,
and in due course nutritious soil is formed. In such places special plant communities
establish and one of typical plants of these areas is lace-leaved buttercup.
Lace-leaved buttercup is a typical representative of its genus – grassy plant
with crawling stalk, which easily taking roots in nodes, and upright flowerstalks
about 15-20 cm tall. Leaves have oval general outlines, deeply palmatisect leaf
blade of bright green colors, are covered with numerous rigid brownish trichomes
– it is a way of protection against caterpillars, snails and other herbivorous
invertebrates. Leaf is divided into 5-7 lobes almost up to a place of attachment
of leafstalk to leaf blade, and each lobe is multifidi with less deep incisions.
Leafstalks grow vertically; because of it a particular leaf resembles a tiny
palm tree. Crawling stalks and flowerstalks are also covered with hairs. Shoots
formed under water have thinner hairs. For protection against herbivorous vertebrates
this buttercup secretes various substances giving to its greenery a sharp smell
and making it poisonous, but local ungulates graze it without any significant
harm for themselves, but in very small amounts only. By means of this plant
animals kill a significant part of intestinal parasites.
Lace-leaved buttercup is a plentifully blossoming plant, but its flowers are
not too bright, of chartreuse color, dimmer than at other species of local flowers.
But they smell intensively and pleasantly, and many insects search for these
flowers, being attracted by smell of this buttercup instead of colouring of
its flowers. Near to this buttercup, other plant species grow, including insectivorous
ones, which also have yellow flowers; therefore the plant has made its flowers
clearly recognizeable, having chosen another way of attraction of insects. Flowers
of this species have 7-12 petals, in particular plants their number is even
more. Petals have rounded shape. One flowerstalk bears one flower on its top
and 2-3 additional ones on short lateral flowerstalks are formed in leaf axils.
Lace-leaved buttercup breeds by seeds, which frequently fall in water and float
in current along the coast. Seeds can also be dispersed by herbivores – they
endure action of their digestive juice. Young plant blossoms at the second year
of life. The basic way of settling is vegetative propagation. This plant has
long catching roots capable to fix in ground of coastal shoaliness. Plant gives
a significant number of crawling runners, settling both above and under water,
and is capable to form even small thickets. For winter lace-leaved buttercup
dies off and in the spring it gives new shoots – at first leaves, and then flowers.
Life expectancy of this plant is about 10-15 years.
This species of plants was discovered by Bhut, the forum member.
Pacific
splendid bleeding-heart (Dicentra pacifica)
Order: Ranunculales (Ranunculales)
Family: Poppies (Papaveraceae), subfamily: Fumeworts (Fumarioideae)
Habitat: North America, Pacific humid forests.
In Neocene epoch, to the west from Rocky Mountains range in North America very
unusual ecosystem has established: very humid forests of temperate climate and
the expressed change of seasons. In this area rains fall regularly (rather cold,
sometimes with snow in the winter), and the level of humidity is very high.
There is a special fauna there: high vertebrates, birds and especially mammals,
are present mainly by large species, whereas among small vertebrates reptiles
and especially amphibians dominate. Invertebrates are present in an unusual
proportion also: lepidopterans, dipterans and hymenopterans are poorly present,
but a variety of beetles and terrestrial molluscs – slugs and snails – is very
great. Such invertebrates are bad pollinators; therefore plants prospering at
northwest coast of North America widely use different kinds of pollinators.
Pacific splendid bleeding-heart, one of characteristic grassy florwering plants
of these forests, uses local birds for this purpose, namely, hummingbirds.
Hummingbirds are rather characteristic inhabitants of these forests, but they
live in such forests only seasonally, since spring up to the end of summer.
In return for life rather safe from predators, local hummingbird species pay
with a significant monotony of their diet: in these forests, there are not many
species of fodder plants, therefore only unspecialized species of these birds
here can live.
Pacific splendid bleeding-heart prospers in humid forests. It is rather large
rhizomatous plant up to half meter tall; rosette of its leaves can reach 60-70
cm in diameter. This bleeding-heart plant resembles externally a kind of fern
– it has the similarly looking wide and pinnatisect leaves of green color with
a glaucescent shade, which favourably contrasts with flowers of this species.
At various species of bleeding-heart plants, flowers have different colors;
at a Pacific species they are very extended, white or whitish-yellow with a
red eye inside. Flowers are formed on large flowerstalks – more than one meter
tall, arched and swinging from a slight breeze. Flowerstalk has a prominent
coral red color attracting pollinators, and it bears some leaflets of the same
red color in the basis of inflorescence. Hummingbird perfectly distinguishes
red color, flies to this plant from apart and thrusts its beak into brightly
colored fauces of flower. Ancestral species blossomed primarily from spring
up to the beginning of summer, but at Pacific splendid bleeding-heart, it is
possible to find flowers during the most part of year, except for winter months.
In winter, the plant stays in a relatively dormant condition, keeping only a
small number of leaves.
Seeds of this species ripen in silicular capsules, which superficially resemble
pods of impatiens and dehisce from any touch in similar way, scattering seeds
far enough to give them a chance to sprout. This Dicentra species does not form
extended rich thickets.
Pacific splendid bleeding-heart is a perennial plant and is able to live 10-20
years. Grubs and snails frequently damage rhizome of this species.
This species of plants was discovered by Bhut, the forum member.
Antarctic
barberry (Berberis notalissimus)
Order: Ranunculales (Ranunculales)
Family: Barberries (Berberidaceae)
Habitat: bushes of Antarctica.

Picture by Lyagushka
In Neocene epoch, Antarctica still remains a very cold continent
with rather poor flora even in comparison with a southern extremity of South
America. The main reason of it is the isolation of continent: very few plants
managed to settle hundreds kilometers far across the sea to occupy this inhospitable
ground. In this respect some berry-bearing grasses and bushes were more lucky
than anothers: their seeds were brought there by birds with their dung, which
became at the same time an original fertilizer given seeds an opportunity to
sprout and to establish in a new place. But more often seeds got on the continent
on legs of birds, being dried on crushed pulp – digestion at small birds passed
too quickly, and seeds appeared in the sea more often. But large birds frequently
brought seeds in their intestines.
One settler berry plant was a descendant of box-leaved barberry (Berberis buxifolia)
prospered on Tierra del Fuego in human epoch. This bush was rather late immigrant
on continent – it has settled down there only after relative warming of the
climate. During many thousand years from its descendants some subspecies of
a new species – Antarctic barberry – had formed. They differ from each other
a little in size, leaf shape and length of thorns, but share one general property
– an improved frost resistance. This low-growing plant is capable to winter
under snow even in fifty-degree frost.
Antarctic barberry is a crawling bush having a trunk from 1 up to 1.5 meters
long with brown pubescent shoots. Trunk woodens; dark grey bark is thick and
friable, serving for a trunk as a kind of “fur coat”. Under bark on the surface
of trunk, a significant amount of dormant buds forms – in case of damage of
a plant, they quickly germinate and a plant restores itself. In addition, the
lower branches frequently take roots and at destruction or damage of a parent
plant continue growth as independent plants. Sometimes at the place of destruction
of an old bush, ring-like thickets of its clones are formed from its branches.
Because of shortening of growing season in a new habitat, this species is an
evergreen plant keeping its foliage till the spring. Leaves are elliptic, at
some subspecies smooth-edged, at others with denticles on the edge; leaves are
cuneate at the basis and caudate on top, entire, from 1 up to 5 cm long at different
subspecies. Closer to the coast relatively large-leaved forms live, and in mountains
and far from the ocean there are mainly small-leaved and very branchy forms
with dense crones not blown through by winds. Thorns are ternate, from 0.5 up
to 2 cm long at different subspecies. Their presence is a vital necessity: meadow
swans – local herbivorous birds – like to treat themselves on foliage of
this species.
Flowering begins at an establishment of stable warm weather and continues up
to the end of summer. Flowers are yellowish-orange, reddish at some subspecies,
growing by 1-2 on thin flowerstalks up to 2.5 cm long. Flowers are pollinated
by beetles, flies, mosquitoes, local butterflies and bees flying to them and
by mousebirds, which peck
off insects from flowers.
Fruits ripen during all second half of summer. They are almost spherical, bright
red, 5-9 mm in diameter. Berries of barberry represent a fine food for numerous
local ornitho- and enthomofauna. In years of especially blessed harvest, berries
fallen on the ground in the autumn are kept rather edible during all winter
and help local birds to survive in winter under snow layer.
The young plant develops rather slowly and begins to fructify only at the fifth
year of life. Slow development is compensated by significant longevity of this
plant, up to 150 years and more. Ring-like clones are capable to live up to
500 years.
This species of plants was discovered by Wovoka, the forum member.
Snake-stem
oak (Quercus serpenticaulis)
Order: Fagales (Fagales)
Family: Fagaceae (Fagaceae)
Habitat: Mediterranean lowlands, slopes of the Alpes.
Mediterranean lowlands represent the area rather adverse for life of plants.
After drying of Mediterranean Sea the climate of Southern Europe became very
dry. Only in spring the snow, thawing in the Alpes, forms the rivers which dry
soon in sand and among the saline grounds remained from Mediterranean Sea. In
this place plants had adapted to endure adverse conditions of environment in
various ways. Some of them stocke water in trunks and leaves, others go through
dry season as tubers or bulbs, and the third ones turned to ephemers and grow
up and produce seeds within short spring time. In this area trees turn the most
freakish shapes.
One tree of Mediterranean lowlands does not rise higher than to one meter above
the ground surface. But its trunk reaches the length of 30 – 40 meters and even
more. Characteristic acorn fruits indicate the belonging of this plant to oaks,
but its trunk resembles more any liana lacking a support. It is a snake-stem
oak, one of characteristic plants of Southern Europe.
The trunk of this tree reaches half meter thickness, but sometimes there are
specimens having trunks of almost one meter thickness. Plants growing in places
with sufficient humidifying – close to channels of drying up rivers or in places
where subsoil waters rise higher – reach such size. Trunk is covered with friable
dark grey bark, wrinkled with numerous small cracks. Wood of this species is
very firm, with a plenty of thin annual growth rings: the plant with a trunk
of half-meter thickness can have age of about 300 years. Trunk and branches
of snake-stem oak take roots very easily. The main trunk grows, rising slightly,
but later lays on the ground and takes roots from the bottom side. Young trunk
of this plant is flexible – it bends around stones and thickens, as if filling
in emptiness between them. Contours of the plant resemble typical tree crone,
but flattened out, uniplanar and expanding in sides. Frequently two or three
large branches at the basis of trunk reach about the same size, as the main
trunk. Taking roots and keeping stones, snake-stemed oak strengthens mountain
slopes. “Love” of this plant to stones has obvious practical orientation – stones
are condensers of moisture from air, and the plant receives enough moisture
for growth, simply growing roots right under stone.
Leaves of snake-stem oak don’t correspond too close to stereotyped image of
oak tree. They are narrow and long (their length is about 10 cm at width of
up to 7 mm), with undulate edges. Such leaf shape helps to distribute light
and shadow in regular order both on the surface of the leaf, and in depth of
crone. Buds and young leaves are covered with rich white downiness; completely
grown up leaf has leathery dark green surface, but keeps white downiness on
the bottom side. In conditions of sufficient humidifying the plant keeps leaves
within the summer, but specimens from droughty habitats can drop leaves in second
half of summer and stay leafless up to the next spring.
The tree begins to blossom in early spring, at the appearing of first young
leaves; pollen of this plant is transferred with the wind. Small acorns of snake-stem
oak (their diameter is about 6 mm), half hidden in ligneous cupule, ripen in
the beginning of summer. They are carried by birds and rodents. These animals
frequently make hidden stocks of acorns, but then not always find them. From
such hidden acorns new plants grow.
Eastern
mountain oak (Quercus appalchiensis)
Order: Fagales (Fagales)
Family: Beeches (Fagaceae)
Habitat: east of North America, slopes of Appalachian mountains.

Picture by Biolog
Prior to Neocene epoch, various species of oaks have managed
to adapt to anthropogenous pressure and to exist rather successfully in conditions
of human-induced changes of nature. Early Neocene, marked by human extinction,
has caused certain harm in oaks, having pushed these thermophilic trees to the
south because of the beginning of the ice age. Then the glacier has retreated,
and thermophilic plants, and trees among them, have returned to the north and
have formed the new species adapted to somewhat changed conditions of life.
For example, Eastern mountain oak, the descendant of a scarlet oak (Quercus
coccinea), one of North American oaks species of Holocene epoch, is a such species.
Like many oaks, it is rather tall tree reaching 10-20 m in height and forming
rich groves everywhere in Appalachian Mountains, but in the north of the range,
large thickets of this species are less often. Leaves of this species have typical
“oak” shape, but have deep cuts of semicircular form and seven (less often five
or nine) lobes slightly expanded on their tips. Leaf blade is of dark green
color, which is replaced in the autumn by equally intense red color, like at
its ancestral species. Bark is rather smooth; it has only shallow cracks and
exfoliates with small scales. Tree is single-stemmed more often; its thickness
at the basis reaches 150-180 cm. Root system is deep and well-developed; it
enables a tree to fix even in cracks of rocks. The plant prefers well-drained
substratum and poor stony soil.
Tiny wind-pollinating flowers have the shape typical for oaks; acorns ripen
within two years and have small knobby acorncaps. Ripen acorns keep on branches
rather loosely; sometimes they may be torn off by wind and roll down the slopes.
They frequently get in places suitable for germination and rooting – into deep
cracks among stones. Acorns have rounded shape and are slightly extended, with
yellowish-brown shell. These acorns represent one of the important fodder resources
for rodents and some birds.
Acorns of eastern mountain oak have rather bitter taste, and the majority of
animals does not eat them, but one jay species, parrot
jay (Cyanocitta pecularia), eats mainly acorns and nuts, and eastern
mountain oak is one of its basic fodder trees. In a diet of parrot jay, a part
of acorns of this oak species is proportionally greater, than at many other
animals. It hides these acorns in the ground as a food stock for winter much
more often, or stores them up in crack between stones and frequently forgets
about them to the spring. If these stocks would not be found out during the
winter by other animals, they would sprout one year later, and if they would
not die in the first 10 winters, they can live a very long life. An average
life expectancy of eastern mountain oak is about 200-300 years.
This species of plants was discovered by Bhut, the forum member.
Prairie
birch (Betula endurabilis)
Order: Fagales (Fagales)
Family: Birches (Betulaceae)
Habitat: prairies of North America.
During Holocene epoch, birches managed to go through the mass extinction becoming
a result of human activity. They have endured climatic changes of early Neocene,
and among them new species have appeared, many of which, however, are rather
conservative, especially non-tropical forms inhabiting zones of temperate climate
in Eurasia and North America. Range of birches had undergone a significant influence
from the part of glaciers, which have shifted their range to the south. At retreating
of glaciers, some species has also settled to the north, to more habitual conditions,
but some species have remained in southern latitudes and have adapted to climatic
changes.
One of such trees is North American prairie birch. Its appearance is typical
for birches of Northern hemisphere: bark of very light color (white or light
grey) with black spots. Due to it some sites of its bark are heated up in the
sunlight much faster, than others, and prairie birch receives an amount of sun
heat necessary for it and does not overheat at the same time. This is a very
low-growing tree of open areas – the height of an adult tree does not exceed
2-3 meters. In a typical case the young tree intensively grows, being single-stemmed;
then growth in height slows down, and active growth of branches begins. On roots
the significant amount of buds is formed; they wake up at damage of trunk, and
very soon form a clone thicket up to 10-15 meters in diameter.
Leaves of a steppe birch have easily recognizeable shape, rounded or oval with
serrated edges, on long and thin leafstalks. For protection against drying up
the bottom side of leaf is covered with white downiness, and on the top side
of leaf numerous hairs grow. In the autumn leaves get bright yellow color and
fall down. In some cases a part of foliage sheds during especially hot and droughty
summer and is partly restored after autumn rains.
It is a wind-pollinating plant blossoming early in the spring, and even in the
end of winter. Its inflorescences represent long drooping aments. Because of
features of pollination, like many other trees of the American steppes and prairies,
this birch quite often forms one-specific groves of several large old clones
of the same species.
Seeds of this species are sispersed also by wind. They are small and are edible
not for many animals, but some prairie songbird species eat them, especially
before winter, when other kinds of food are not plentiful in North American
prairies. But much numerous bird search in thickets of prairie birch for various
invertebrates eating foliage, wood and other parts of this tree. Elevated parts
of this plant represent forage for larger herbivores – for vertebrates. Prairie
birch resists to them due to intensive growth, but peccasons
have strong enough jaws to browse even large branches and to pick bark on tree
trunk. In places, where large herbivores frequently feed, this tree grows less
often, than other tree species of North American prairies.
A particular plant of prairie birch lives no more than 12-15 years, but the
clone successfully exists in the average 40-70 years and more, especially in
conditions of browsing by large herbivores and the subsequent rejuvenation of
thickets.
This species of plants was discovered by Bhut, the forum member.
Patagonian
false hazel (Nothavellana antipodorum)
Order: Fagales (Fagales)
Family: Southern beeches (Nothofagaceae)
Habitat: the south of Patagonia, Tierra del Fuego, forests.
Southern beeches are characteristic plants of ancient Antarctic flora widespread
on continents and islands of Southern hemisphere. One center of their specific
variety is a far south of South America, an area of temperate climate with the
expressed seasonal changes. Various species of southern beeches occupy various
ecological niches in temperate forests of the south. In due course of evolution
among them original “twin” of hazel (Coryllus) of Northern hemisphere – Patagonian
false hazel – had appeared.
This species is an undersized treelike plant with large nut-like seeds. The
height of this plant in favorable conditions does not exceed 3-4 meters, more
often only a little taller than 2 meters. Usually it is a monocormic plant,
but at damage of a top it easily branches and grows in several trunks, keeping
spherical general outlines of a crone. It is a shadow-resistant species growing
in underbrush. Its bottom branches frequently lay down on the ground, take roots
and continue their growth, forming new vertical trunks from lateral buds. The
fallen trunk also can take roots – in this case lateral branches start to grow
upwards, forming rich thickets in underbrush. Leaves are wide and smooth, of
rounded outlines, with obtuse tip and dentate edge, up to 15-20 cm long. This
species is a deciduous plant; in the autumn its foliage turns yellow.
Flowers are very small and wind-pollinated. The plant blossoms in the spring
in the beginning of leaf growth.
Seeds ripen in the middle of summer; the size of seed is up to 15 mm and it
is covered with hard ligneous shell. It is one of the basic food sources for
local birds and mammals, which gather significant stocks of seeds of this kind
and are their basic dispersers. Seeds keep germination capacity for a long time
– up to 3 years. Young plants begin to fructify at the age of 5 years, life
expectancy reaches 200 years; due to rejuvenescence via young growth the age
of clones may reach 400 years.
True
cabbage tree (Brassixylon crassus)
Order: Capparales (Capparales)
Family: Crucifers (Brassicaceae)
Habitat: dry districts of Southwest Asia, Northern Africa and Southern Europe.

Picture by Biolog
The lot of species of plants of Holocene epoch had been named
“cabbage trees”. But this name was given to them exclusively for external similarity
of some parts of plant to cabbage cultivated by people. But the tree has appeared
in Neocene in dry areas of Southwest Asia (in foothills to the east from the
Mediterranean lowland), and later settled on coast of Mediterranean swamps,
is named more deservedly: it is one of close relatives of cabbage, belonging
to the same botanical family.
The true cabbage tree will not make a competition to giant trees of tropical
forest: its maximal height does not exceed 13 – 15 meters. But it can survive
in places, where dry and hot wind blows and settles on ground thin coating of
salts from salt Mediterranean swamps.
In connection with very dry climate in places of inhabiting of this tree, at
it the number of adaptations was developed, permitting to survive in extreme
conditions. True cabbage tree is stem succulent: very thick trunk similar to
a bottle serves to it for accumulating of water stock. It is strongly expanded
in the bottom third, and sometimes lays on the ground: it seems, as if tree
has not sustained weight of the “beer stomach”. Wood of this plant is fragile
and friable, plentifully sated with water. But only few ones venture to drink
it: tree reliably protects itself with the help of bitter substances dissolved
in sap. Leaves of this tree also have bitter taste. They are juicy and rather
wide: the tree leads active photosynthesis the most part of year, running in
stagnation (“catalepsy”) only in hottest summer months. But also then the most
part of leaves is kept, only oldest ones cast. Young leaves form original crowns
on tips of tree branches. Petioles of leaves can make movements, and depending
on environmental conditions tree folds leaves in similarity of cabbage heads,
or spreads them. Usually leaves are folded together at night and in hottest
time of day, being opened in the morning and in the evening. But in cloudy days
crowns of leaves may open all the day. Besides for reduction of moisture loss
leaves are covered with thin wax cover.
True cabbage tree extracts water from deep layers of ground, therefore it prospers
in areas where underground waters approach close to a surface. In rare oases
of Northern Africa this tree forms original thin woods both with palms and some
other plants.
This plant, as against to cabbage, is perennial: the age limit of it may exceed
200 years. It also blooms more than once in life, but annually approximately
from 5-years age. Large pink flowers with four petals in big corymb-like inflorescences
open on uppermost branches in middle of short rain season, involving butterflies
and flies. Each flower blooms not for long time – only two days and one night.
Therefore many flowers of this plant remain not pollinated. Fruits of true cabbage
tree look like long and braided, as if a drill, pods. At a touch (for example
if any bird sat on inflorescence) ripe fruit “blows up”: folds instantly braid
and scatter seeds far. An environment of seeds is naked, but very strong and
bright: red with white speckles. Birds involved with such colouring, peck these
seeds. But in their stomachs the environment is not digested, but becomes only
a little thinner and more permeable. Having dropped out from intestines of bird
with portion of dung, seeds have good chances of growing.
The first year young plant has underground tuber-like stalk (“root crop” formed
by hypocotyl of sprout, and the crown of small leaves on the ground surface.
At the second year growth of trunk begins though the underground thick stalk
does not disappear still long time. If the trunk will be damaged (more often
simply had eaten), the plant has chance to survive due to underground part.
False-leaved
prickly pear tree (Equadopuntia pseudophyllata)
Order: Caryophyllales (Caryophyllales)
Family: Cactuses (Cactaceae)
Habitat: Galapagos Islands, coastal areas.

Picture by Lyagushka, improvement by Biolog
Galapagos Islands in human epoch had differed in droughty
climate: at them there were no constant reservoirs, and fresh water could be
found only in rain pools. Therefore the vegetation of three types prevailed
at this archipelago: xerophytes (enduring significant dryness of ground), succulents
(stocking water in trunks) and mangrove vegetation (able to live in sea water).
Succulent vegetation of Galapagos Islands had been presented by several species
of cactuses among which prickly pears (Opuntia) were most typical ones. These
cactuses with wide flat stalks had reached the size of tree at Galapagos Isles.
Huge local tortoises had been unique animals ate these cactuses. Introducing
of goats to these islands even turned to cactus’s advantage: number of tortoises
had reduced, and goats were eaten away other plants, had lost cactuses of natural
competitors. Therefore prickly pears had remained dominant plants at Galapagoses.
Neocene descendant of Galapagos prickly pears has remained a treelike plant.
But changes of climate had forced it to change shape though some basic features
of cactuses nevertheless had kept at it. Galapagos false-leaved prickly pear
tree grows up to 20 meters in height, and trunk of old tree may grow up to 6
meters in diameter. At this species the differentiation of sprouts to two sharply
distinguishing types is precisely shown. Trunk and main branches of this tree
are covered with thick, friable and easily crumbled bark. Such bark reliably
protects trunk of plant from overheat, thus reducing losses of moisture. Wood
of prickly pear tree is friable, watery and rather fragile. During hurricanes
some trees fall, but continue to grow, taking roots in ground. Also separate
branches of prickly pear tree easily take roots.
Sprouts older than three years keep spikes gathered in bunches by some ones
on the surface. On young branches sprouts of other type grow: green and soft
oval annual “leaves” almost without spikes. Only on edges of such “leaves” short
spikes stick up. Most part of “leaves” of this plant lives only one year, drying
and falling down in dry season. Only youngest “leaves” may go through it and
keep two seasons in succession.
Prickly pear tree flowers almost the year round, only in summer drought it may
stop flowering, simultaneously shed “leaves”. Flowers of this plant are bright
red with long petals, up to 10 cm in diameter. They produce a lot of nectar,
but are smellless: this plant is pollinated by birds. Fruits of prickly pear
tree are sweet and juicy, pear-shaped and covered with spikes. Local birds peck
them, eating pulp and carrying seeds, and fallen fruits are eaten by large local
tortoises.
Prickly pear tree grows rather quickly, giving for one year apical growth about
a meter tall. But this tree lives not for long: no more than 200 years.
The idea about existence of this plant had been supposed by Simon, the forum member.
Coral
opuntia (Rhodoopuntia corallina)
Order: Caryophyllales (Caryophyllales)
Family: Cacti (Cactaceae)
Habitat: southern slopes of the Alpes.

Picture by Biolog
Millions years after human disappearance have passed, but
traces of human activity in the field of introduction of various species of
animals and plants will be kept for a very long time as descendants of these
species living in places where they could not get in the natural way. The Mediterranean
hollow is surrounded with the Alpes from the north and with deserts from the
south, the east and partly from the west. Lands, not poisoned with salt of dried
up Mediterranean Sea, differ in dryness, and only few plants can grow there
successfully. Among the ones managed to get accustomed in these places, there
were cactuses, descendants of prickly pears (Opuntia spp.) introduced by people.
Some species of these plants have got freakish shape because of influence of
rigorous conditions of habitats.
Plants of Cacti family produce betalains – red pigments close to beet betanin.
They give characteristic red color to flowers of cactuses. These pigments absorb
ultra-violet radiation successfully and protect the plant from burning rays
of sun. In Neocene in Mediterranean lowlands unusually looking species of Opuntia
evolved, at which the ability to synthesis of these pigments is amplified in
great degree. Therefore its stalks have bordeaux-red color (the chlorophyll
is present, but it is masked by red pigment) and are similar to thickets of
precious (red) coral, as if it was kept on the place from the moment of sea
drying. Hence the name of this plant – coral opuntia.
Because of high content of the pigments synthesized by this plant, petals of
its flowers have very dark purple, almost black color, as flowers of the black
tulip bred once by people. Occasionally in a population there are carriers of
rare gene alleles having citreous, black and green colouring of stalk.
Coral opuntia grows not in marshy hollow, but on southern slopes of the Alpes
adjoining to it. Red leaf color is characteristic for mountain plants as in
mountains solar ultra-violet rays are especially dangerous. At rather small
heights (less than 3000 – 3500 m above sea level) this prickly pear can take
the treelike shape and grows almost as big as saguaro (Carnegia gigantea) of
human epoch, but branches much more plentifully, forming a crone of flat branches.
At such height in its population there are individuals having atypical colouring
of stalks as here the intensity of ultra-violet radiation is not so great, and
mutants do not receive burns. Higher the coral opuntia strongly decreases in
size, loses the basic trunk and at greatest heights, a little below the snow
line, it forms an elfin wood of creeping perennial stalks from which shoots
of the current and last years rise upwards. Highly in mountains red colouring
turns more intensive. Flat stalks of this species are similar to shoots of human
epoch prickly pears. They keep the pointed spikes, growing in bunches of 5 –
8 ones, and young sprouts have the underdeveloped leaves similar to little horns
and rather rich downiness of thin hair protecting growing tissues against solar
burn and frosts. The completely developed shoots become hardier, and flowers
and fruits form on them.
Roots are adapted to fastening on rocky ledges. Elfin wood form in high mountains
easily takes roots at the bottom part of perennial shoots and plays the important
role in prevention of landslides.
This species of plants does not show strict requirements to pollinators; various
animals – from beetles and butterflies up to small birds and rodents can carry
the pollen of these cactuses. Flower buds are formed at the edge of last year’s
shoots in winter. In spring and in the beginning of summer unusual black flowers
of this plant blossom. They are strongly heated up in sunlight and involve carrion-eating
flies and beetles. And the insects gathering on flowers, in turn involve insectivorous
birds. Fruits of coral opuntia have very dark colouring and are covered with
glaucescent layer of wax. This cover reflects ultra-violet rays, making fruits
more appreciable for birds. Also rodents and other mammals eat fruits of this
cactus. Carnivores willingly eat fruits fallen from treelike cactuses of this
species, being involved with a moisture contained in them. Pulp of fruits is
sapful, dark red and very sweet; tiny seeds are dispersed in pulp. In dry years
the pulp of fruits can not reach the complete ripeness, but seeds normally develop
and can even sprout right inside the fruit. This feature gives an additional
guarantee of survival of sprouts. In typical case seeds are distributed by birds
and mammals.
This species of plants was discovered by Anton, the forum member.
Naked
halopuntia (Halopuntia nuda)
Order: Caryophyllales (Caryophyllales)
Family: Cacti (Cactaceae)
Habitat: Mediterranean lowlands, saline soils.

Picture by Biolog
In Neocene there is no more adverse place for life of plants,
rather than areas of the former Mediterranean. Collision of Africa and Europe,
continued for millions years, had resulted in closing of strait of Gibraltar
and drying of Mediterranean Sea. Giant amounts of the salts formerly dissolved
in water, appeared on land after water had receded and evaporated. Salt had
impregnated the ground, having made it unsuitable for growth of plants. Only
Mediterranean “oases” formed on heights support vegetative life which feeds
the poor animal population. And in salt swamps the creatures, able to resist
salt, prosper. Wast space of the hollow is covered with the crust of salts and
gypsum, and only in few places in cracks of firm minerals a stunted vegetation
can exist.
In such conditions there is no place to competition – it is replaced quite with
difficulty of life in such environment. And ones had found a way of survival
prosper even in such rigorous conditions – on the heated waterless ground under
hot rays of sun.
On Mediterranean saline soils one species of plants grows, which is a descendant
of the species introduced by people. It is naked halopuntia, the representative
of cactuses, a halicole spikeless opuntia.
This plant represents a branching bush about 1 meter high, forming dense cushion-like
thickets. This species resembles externally a prickly pear of human epoch, because
the external shape of the plant has changed only a little, and it turned smaller
in comparison with taller ancestors. Halopuntia has the same segmented stalks
made of thick oval-shaped segments. But at close look it is appreciable, that
the cactus does not have spikes, very characteristic for ancestral prickly pears.
One more difference from prickly pears of human epoch is a color of plant: it
is covered with plentiful wax film and has silver-gray colouring.
Reduction of the size of this plant is connected to adverse conditions of existence;
at life on the salted ground a lot of energy is spent for pumping out of ions
of sodium and chlorine through cell membranes against a gradient of concentration.
The plant accumulates in the tissues water with rather low concentration of
salts, and surplus of salt is secreted by special glands on the surface of cuticle.
Salt is initially secreting as very rich brine and stiffens on the surface of
a plant as drops. Due to layer of wax it does not dry on the thin skin of shoots;
as accumulation proceeds, crystals of salt break off and fall, or they are blown
off with a wind. Salt is secreted mainly on old shoots, and young sprouts are
reached with the sap already deeply filtered from salt.
The root system of halopuntia is very advanced, as at the majority of desert
plants: it exceeds stalk many times in the size, and it is fair to assert, that
the most part of cactus is hidden under the ground. The bottom parts of stalks
form a caudex covered with thick layer of dry skin which protects this plant
from penetration of salt. The root system of halopuntia is located in top layer
of the ground, at the depth up to 10 – 15 cm under surface level. During rare
rains it quickly absorbs water, while salts in ground are not dissolved yet.
In dry season the tips of roots wither, and roots become covered with dense
peel interfering the drying out.
Usually naked halopuntia is imperceptible from apart – colouring of this plant
merges with a landscape spotted with white areas of salts. Therefore the rare
herbivores, venturing to go far into salt desert, do not notice the plant. Growth
of this plant occurs in two waves – in spring and in an autumn when air is rather
moist. In summer, in peak of heat, halopuntia runs into a condition of catalepsy,
and stays in rest in winter. Flowers of this species are bright red, large (their
diameter is over 10 cm) and multipetaled, with a characteristic wax cover on
petals which reflects ultra-violet rays and involves pollinator insects. During
the flowering thickets of halopuntia become seen from apart, and there is a
probability of their damage by herbivores. Fruit of this plant is a berry characteristic
for cactuses. The pulp of fruits has sweet taste with slight saltish taste.
It is eaten willingly by birds distributing tiny seeds of halopuntia.
Halopuntia grows very slowly, and the survival rate of sprouts is lowest. The
plant easily breeds by cloning: the wind can break off a part of branch which
takes roots successfully in the ground and can live for a long time due to internal
stocks of water. Life expectancy of this plant is more than 100 years.
This species of plants was discovered by Anton, the forum member.
Compass
cactus (Platyopuntia compassa)
Order: Caryophyllales (Caryophyllales)
Family: Cacti (Cactaceae)
Habitat: Mediterranean lowland, Northern Africa, deserted areas.
Extreme conditions of existence cause occurrence at live organisms living in
such conditions of various adaptations for the survival which frequently give
freakish shape to their owners.
In arid areas at the north of Africa and along the edge of Mediterranean hollow
the complex of vegetation is formed, including various species hardy to extreme
conditions of existence. Some of them grow and develop in more favorable season
while the sun heats not so intensively, and survive in summer drought as tubers
or bulbs. Others have time to live the whole life within several weeks and exist
in difficult time as seeds. And the third ones do not hide anywhere and resist
to drying up heat successfully.
People introduced American prickly pear cactuses to the Mediterranean region.
They successfully developed in these places even without people; therefore they
had kept after human extinction and their descendants began the expansion in
arid areas of Mediterranean hollow. One of Neocene species of Old World opuntias
is a compass cactus. This plant is named for prominent feature of growth: its
flat branches are always oriented along the north – south line by an edge; therefore
hot beams of midday sun slide along their surface and do not cause burns.
Compass cactus keeps characteristic shape of its ancestors: it has flat stalks
made of flattened segments. Segments have rounded outlines, are rigid, gristly,
strongly squeezed from sides and fibrous. Outlines of the plant are fan-shaped:
all branches are located almost in the same plane and do not overlap each other
at the side view. At this plant some perennial stalks are kept - its “trunk”
and “branches”, and the significant part of shoots does not live more than three
years. In due course of growth sprouts change shape and functions. One year
old shoot is covered with thin layer of wax protecting tender growing tissues
from solar burn and giving glaucescent color to the shoot. For the second year
shoot grows ripe and loses wax cover. Its surface gets bright green color, and
shoot begins active photosynthesis. Very large spikes of compass cactus grow
not so densely, but are ligneous and strong. Herbivores live in deserts seldom,
but protection against them is necessary all the same. The height of plant does
not exceed one meter.
Some shoots of the second year form only vegetative buds, but at the majority
of them flower buds are formed. At the third year of shoot existence in the
beginning of spring flowers blossom. They are white and small, with numerous
dulled petals, and are located along edge of shoot segment in groups of 4 –
7 ones. Petals of these flowers are covered with a layer of wax due to what
they reflect sunlight and are appreciable for rare pollinator insects.
After pollination on the plant small edible berry-like fruits of reddish-pink
color ripe; they have sourish taste and numerous tiny seeds. At the ripening
of fruits the segment, on which they ripen, gradually dries out, giving to fruits
the nutrients stored up in the previous year. Simultaneously on such shoot spikes
fall down. Dried up segments of stalk with fruits remain on the plant where
they are found by the birds, which are the main distributors of seeds of compass
cactus. When all fruits appear eaten, shoots gradually wither and fall down.
This way natural thining out of the plant crone takes place. The broken off
one-year-old shoots perish, as a rule. The two-year-old shoot stocked up enough
nutrients, can live, not taking roots, till three years in extremely droughty
conditions. Due to such survivability it easily strikes roots in favorable conditions
– a rare rain is enough for it to take roots in the ground. Gradually during
the next three – five years it forms a normal-shaped plant and starts blossoming.
Baobab
opuntia (Adansopuntia obesa)
Order: Caryophyllales (Caryophyllales)
Family: Cacti (Cactaceae)
Habitat: savannas and semideserts of northern Africa.
In human era cacti of prickly pear genus (Opuntia) were introduced to the Mediterranean
region. After the disappearance of mankind they survived climate change and
became widespread in arid regions of the Old World, giving rise to various life
forms. The largest of the descendants of prickly pears in the Old World is the
baobab opuntia. It is a large tree, similar to a baobab in appearance, size
and shape, but upon closer inspection it turns out to be a cactus. This species
preserved and enhanced the characteristic-to-cacti ability to store water in
its trunk, so that it can withstand prolonged drought and grow in an environment
where other woody plants die.
The trunk is about 20 m around, but the height of the tree is not too great
– also about 20 m. The wood of this species is friable, with plenty of water
storing parenchyma, and an outer layer is strengthened with a network of fibers.
The lower part of the trunk is unbranched, the first large branches grow at
a height of about 10 meters. Perennial branches are rounded in cross-section,
the young growth until the age of three years is similar to the typical flattened
stems of prickly pears. Young branches are thorny; spines up to 5 cm long, with
jagged edges, are gathered in groups of 4-7 ones. Trunks of young plants are
also spiny, but the spines disappear as they grow. The bark of the trunk of
the adult plant is smooth and spineless, but very dense and thick – up to half
a meter at the base of the trunk. Thus the plant protects itself against rare
desert herbivores who want to get primarily to the succulent wood. This plant
endures damage of the bark very easily, quickly restoring the lost parts of
the bark. Also, the thick layer of bark serves as protection against overheating.
The baobab opuntia’s flowers are very large – about the size of a dinner plate,
bright red and having many petals. They appear on biennial shoots; buds are
formed in autumn and winter. The tree in bloom is an unforgettable spectacle.
Pollination is generated by insects and birds, which it tempts with watery nectar.
The fruits are the size of a grapefruit, with red-purple flesh, sweet and juicy.
They are eaten by monkeys, who are among the main seed distributers. In dry
areas harelopes and birds feed on its fruits.
Seedlings successfully develop only in rare rainy years, when they have time
to store enough water to survive the dry season. Usually the first two years
of the plant’s life are critical for survival. Baobab oputia blooms for the
first time at the age of about 15 years, having reached 4 meters in height.
Life expectancy is more than 1000 years.
The idea about existence of this species of plants was proposed by Anton, the forum member.
Translated by FanboyPhilosopher.
Mole
cactus (Cryptocactus subterraneus)
Order: Caryophyllales (Caryophyllales)
Family: Cacti (Cactaceae)
Habitat: Mexican desert, arid areas with a low precipitation level.
Cactuses belong to the plants most adapted to arid conditions. Almost all cactuses,
except for the most primitive ones, are stem succulents, and their green stalks
incur functions of both leaves and water-stocking organs. Stalks of cactuses
take various shapes – from small spherical ones up to huge barrel-like and even
treelike plants. The exceptional
variety of cacti species indicates their success in struggle for existence.
As a rule, the stalk of cactus is perennial though in Holocene epoch there were
separate exceptions of this rule. In Neocene among cactuses the species had
evolved, which had departed the common rule: its each stalk actually exists
less than two years. This cactus is very original “invisible” plant: it can
be seen only within several months per year. Every other time this plant is
hidden under the ground from hot sun rays; and this plant is named because of
this feature – it is a mole cactus.
The occurrence of mole cactus at the ground surface begins from its flowers.
Shortly before a season of spring rains, when the winter cool gradually leaves,
bright red flowers, appreciable from apart, about 4 – 5 cm in diameter blossom
right at the ground surface. Usually there are some of them. Insects notice
them due to feature of a structure of petals – they reflect ultra-violet light
intensively because of special structure of petals. This species is pollinated
by various insects – even ants and beetles feed in its flowers. After pollination
petals fade, and under the ground ripening of the fruit, a small rounded berry
with numerous tiny seeds and sapful pulp begins. Usually one or two berries
develop successfully on the same plant, and only in very favorable conditions
their number may increase. Berries ripen to the beginning of summer, and the
pedicel, growing roughly, takes them out to the ground surface, where they are
eaten by rodents and birds.
When the flower is pollinated or has simply faded, active growth of stalks of
this plant begins. Shoots of mole cactus are spherical and have numerous not
deep plicas on the surface. Colouring of plant skin is bright green with glaucescent
wax film; there is white downiness at tops of stalks and in areolae. Spikes
of this plant are strong, but not numerous. In every areola there are one long
hook-like spike and some smaller straight ones. With the help of hook-like spikes
shoots of this cactus are hooked on wool and feathers of large inhabitants of
desert. They easily break off the basis and thus are settled to long distances.
Shoots of the current year grow from a last year’s stalk which is located under
ground and gradually gives all saved up nutrients for growth of new stalks and
fruit ripening. In due course of stalk growth and seed ripening last year’s
shoot gradually corrugates and then dies off, and to the middle of summer new
stalks already begin to grow independently. In such way during some years in
succession the initial plant forms some kind of a clone expanding as almost
right ring. Usually flowers and fruits appear at the internal edge of ring,
and new shoots grow on external side. Diameter of stalks of mole cactus reaches
6 – 8 cm.
In the middle of summer, when last year’s stalk dies off finally, every young
plant forms some long thick roots which stretch downwards and branch at small
depth. They represent a kind of “anchors” for the plant, but their role in its
life is much more important. When weather becomes too hot, roots begin drying
up and convolving. Thus they form significant effort which draws the plant under
the ground. Till one or two weeks cactus hides under ground, and these stalks
will not appear at the surface any more. Thus spikes gradually weaken and fall
off, breaking at the retraction of the stalk under ground. In darkness the stalk
gradually loses chlorophyll and becomes similar to tuber. During the winter
buds form on it, and from them the next year flowers and new stalks will appear.
Octopus
sundew (Arbopendera nocturna)
Order: Caryophyllales (Caryophyllales)
Family: Sundews (Droseraceae)
Habitat: tree groups in steppes of Southern Europe.
In Neocene, the south of Europe has changed only a little: from temperate zone
up to the Balkan extensive steppes extend, where grassy plants of various families
grow. Only in places, where there are more-less constant rivers and other large
reservoirs, larger plants – bushes and trees – grow also.
But plants adapt for an inhabitancy not worse, than animals and fungi do; even
there, where trees and bushes grow, smaller representatives of flora could find
the place for life.
Acacia-leaved maple is one of the most successful
and tall trees in the European steppes; it frequently forms rich thickets at
the banks of the steppe rivers. But on its branches, in addition to its own
pinnate leaves, there are also other ones not similar to them – elongated, green,
shining, hanging down from tree branches like tentacles of a predatory octopus.
These leaves belong to octopus sundew, one of local kinds of insect-eating plants.
Octopus sundew is a rosette plant: its stalk is very short, topped with the
rosette of long and wide leaves, which hang down from branches of acacia-leaved
maples (or other steppe trees) and rock in air freely. Root system of this plant
represents a set of long and thin roots of white color, which form a kind of
grid under bark and in cracks of bark of the host tree. However, ocnopus sundew
is not a parasite, but an epiphyte: it extracts the most part of necessary nutrients
independently, like its relatives do.
But octopus sundew has a unique feature, which, perhaps, is observed in related
species: it is strictly nocturnal plant. In the daytime leaves of this plant
are curled in hard ball and are actually not visible for various herbivorous
animals coming into a grove to browse tree foliage. In such condition plant
receives enough light for photosynthesis. During the sunset, when the ambient
temperature decreases a little, and humidity of air increases, leaves unwrap
and hang down from branches of a tree, where this sundew grows. Various mosquitoes,
moths and other nocturnal insects fly up to it leaves, being attracted by drops
of sweetish liquid, and stick to them. Having felt, that something has stuck
to its leaves, sundew rolls them and begins to digest caught insects.
In addition to catching insects, this plant gets necessary moisture with the
help of its foliage. In the morning on sundew’s leaves dew begins to condense,
and the plant reacts to it right in the same way, absorbing it with the whole
surface of leaves. In steppe rain represents a rare event, therefore actually
all water accessible for sundew is dew. Leaves curled up in the daytime help
to save up moisture in the hot sun.
Octopus sundew beging to blossom in second half of spring. Its flowers similar
to tiny bellflowers open at night and grow on long flowerstalks lifted upwards
(leaves hang down, therefore pollinator insects are in relative safety). The
same insects, that represent a prey of octopus sundew, pollinate it and carry
its pollen from one flower to another. Fruits of octopus sundew represent tiny
undehiscent capsules with fleshy walls, looking like little whitish berries.
They are eaten willingly by local songbirds; seeds pass through their digestive
systems without any serious harm and successfully sprout, when get into the
environment suitable for their growth (branches of a maple or other steppe tree).
This species of plants was discovered by Bhut, the forum member.
False-flowering
sundew (Drosera pseudofloribunda)
Order: Caryophyllales (Caryophyllales)
Family: Sundews (Droseraceae)
Habitat: humid subtropical forests at the coast of Fourseas, Caucasian peninsula.
In human epoch sundews growed in Europe were the small plants lived at the wetlands.
Fitness to existence in conditions of deficiency of nutrients has enabled these
plants to “lose touch” with the ground and to develop new habitats: the stones
and tree branches covered with a moss. In Neocene Europe even epiphytic sundew
species have evolved. One of such species is false-flowering sundew widespread
in forests along the Fourseas coast.
It is a small rosette plant with very short thick stalk; a general diameter
of the rosette is about 10 cm. Plant has very original leaves, which are the
basic suppliers of nutrients for it. Like at all sundews, leaves of this species
are covered with brightly colored tentacles, on which tips, drops of sticky
liquid are secreted. But the shape of leaves determines the name of this species:
at false-flowering sundew, leaf mimics a flower of a certain plant. Leafstalk
is attached to a leaf not from the edge, but in the center of the bottom side
of a leaf blade. Leaf blade is very much concave and in general, it resembles
a wine-glass on a long stem. Diameter of such leaf does not exceed 12-15 mm.
Trapping tentacles are arranged only along the edge of such leaf, imitating
petals of a flower. They are colored pinkish-red, increasing the similarity
to a flower. Some thin tentacles grow in the middle of a leaf, imitating stamens
of a flower. In their basis there is large gland secreting a drop of nectar.
Large moths with long proboscises are quite able to suck this nectar, not landing
on this plant, but smaller insects, when creeping on such “flower”, appear inevitably
pasted to it and perish. Having felt a touch of prey, the “flower” leaf closes
like a real blossom and digests it. An adult plant can keep simultaneously up
to seven such leaves. In the similar way tropical plants of flowerleaf
genus imitate flowers of plants, being thus an example of convergent evolution.
False-flowering sundew can grow as an epiphytic plant in tree crones, frequently
in shadow of other plants. This species has not completely lost connection with
the ground and frequently grows among moss cushions on rocks.
Stalk of this species is tuber-like, similar to a small nut and is covered with
rough skin. It forms some the tenacious attaching roots, penetrating into a
bark or cracks in stone. Atop of them the network of thin soaking roots develops,
providing plant with water. In the autumn all leaves and soaking roots die off,
but attaching roots keep viability within several years in succession. Life
expectancy of this plant makes about 10-15 years.
Vegetation of false-flowering sundew begins in early spring. The first leaves
have tongue-like shape like at its ancestral forms. Approximately in one month
after the beginning of vegetation leaves of the typical shape develop.
In the middle of the summer, false-flowering sundew begins to blossom. Its true
flowers are small, blue ones having five petals, composed in lax raceme on long
elastic flowerstalk rising upwards to the height of about 30 cm. They are pollinated
by small dipterans and wasps. Fruit represents dry capsule; seeds are wind-dispersing.
Sprouting
sundew (Drosera prolifera)
Order: Caryophyllales (Caryophyllales)
Family: Sundews (Droseraceae)
Habitat: tropical rainforests of Florida, coast of gulf of Mexico, Cuba.
In Neocene in territory of Florida and North American coast of gulf of Mexico
favorable conditions for development of tropical forests had established. It
is a world of extensive wetlands and lowlands, a kingdom of moistureloving plants.
Among grassy plants of this area insectivorous plants of various families are
frequently found. On land and in coastal zones of bogs various species of sundews
frequently grow. One of their representatives is sprouting sundew, a rather
large species of this family.
Sprouting sundew represents a perennial rosette plant with short tuber-like
stalk. Leaves grow on strong long leafstalks; diameter of the rosette of an
adult plant reaches 30 cm. This species can grow not only on the ground: sprouting
sundew is a facultative epiphyte. If birds or wind will bring seeds of this
plant on trees, it normally grows, using wood dust and moss as a substratum.
Usually it is an inhabitant of moss cushions in the underbrush.
Leaves of sprouting sundew are covered with characteristic trapping tentacles
secreting drops of a sticky liquid to which the caught animals paste. Diameter
of leaves at this species is about 5 cm. Leaf is spoon-shaped, and its top side
is slightly concave. Leaf colouring is reddish-brown, and tentacles are bright
red with white points formed by groups of cells reflecting ultra-violet rays.
In this way the plant attracts insects, but they are not its only prey. Due
to rather large size sprouting sundew is capable to catch and to digest small
frogs and lizards.
Due to a tropical climate in its habitat, this plant blossoms almost the year
round. Flowerstalk is about one meter tall; it is thin and elastic, of purple
color. Sprouting sundew has small white or yellow flowers of five petals.
For this species ease of vegetative propagation is characteristic, due to what
it can settle quickly. Leaf of this plant easily breaks off in the place of
an attachment of leaf blade to leafstalk. A leaf broken off from a plant keeps
viability and motor reactions in relation to an animal got on it. Sometimes
a tiny frog can break a leaf off, having casually got on it and trying to escape.
But the broken off leaf continues to secret digestive enzymes and can kill and
digest such prey independently. On the break place of a leaf a bud appears very
soon, from which later a young plant develops. An adventives bud can also form
on the leaf tip. If a tip of normally growing leaf touches the ground, the bud
begins to develop and forms a young plant soon. It gradually takes roots and
starts to grow independently near to a parental plant. An adventive bud develops
also on the tip of broken off leaf, but only in conditions of an additional
feeding.
Young plants begin to blossom already at the second year of life.
Pitcherplant-leaved
sundew (Neodrosera nepenthifolia)
Order: Caryophyllales (Caryophyllales)
Family: Sundews (Droseraceae)
Habitat: edges of the Mediterranean lowland.
In Holocene the sundew genus Drosera was one of the most widespread among insectivorous
plants. Sundews could grow on various soils – from wetlands to mountains and
sandy grounds. Their hunting device has rather simple arrangement: long glandulous
hairs with drops of sticky liquid on their tips grow plentifully on leaves of
various forms and sizes. The irritation of a leaf by substances of protein nature
forces it to roll, and secreting juice splits these substances. In Neocene sundews
grow at all continents, even in Antarctica.
One descendant of European sundews has settled in the Mediterranean lowland
and acquired some almost constant microscopic cohabitants enabling the plant
to survive in such harsh conditions. Pitcherplant-leaved sundew lives on dry
soils of edges of this lowland, avoiding saline soils. It has kept numerous
features of its ancestors: a radical rosette 20-30 cm in diameter of 15-20 leaves
and thin tall (up to 50 cm) flowerstalk, on which tip an inflorescence develops.
Root system of pitcherplant-leaved sundew is remarkable in its impressive length
– up to several meters in ground depth that is connected to necessity of extraction
of fresh water from deep soil layers. The main root is napiform right under
rosette, and on the collet buds form, from which new shoots grow. Leaves resemble
a smoking pipe in their shape: on the end of leafstalk, there is a transformed
leaf blade – small (about 3 cm deep) pitcher with a lid. Leaf is deeply concave
and short glandulous hairs grow on its internal surface, covering two thirds
of its area. One side of leaf is extended upwards and bent, forming a lid. Edges
of pitchers are smooth and sloping inside, promoting falling of insects inside
of the trap. Hairs inside pitcher secret water, sugar and digestive enzymes
for digestion of prey. Pitchers are always filled with a liquid up to a level
of top hairs on their surface, and the whole pitcher is covered with dense cuticle
from within. Evaporation is compensated by plant’s secretions and by rains and
dew, when available. This liquid emits the sweet smell attracting insects.
All aerial parts of a plant (stalk, leafstalks and outer sides of trapping leaves)
are bright green; edges of pitchers are dark red.
Pitcherplant-leaved sundew blossoms in the summer, during the most active swarming
of pollinator insects. Flowers about 1.5 cm in diameter are composed to spike-like
inflorescence, which numbers up to 10-12 blossoms. Flower structure is the same,
as at its ancestors: pentamerous green calyx, pentapetalous bright red corolla,
five stamens and one pistil with superior multilocular ovary, in which basis
there are nectaries producing fragrant nectar. The plant is not specialized
to pollinators: its flowers are visited by solitary bees and wasps, flies, beetles
and butterflies. Fruit is a lobular dehiscent capsule with a lid, full of tiny
seeds like poppy ones, which are dispersed by wind. Germinating capacity of
seeds is kept for about one year; they usually sprout during rare spring rains.
But the most unusual side of life of this plant is its microscopic symbiotes.
In tissues of pitcherplant-leaved sundew leguminobacteria
live, forming tubercles on roots and leaf stalks, when the plant needs nitrogen
because of prey shortage. This plant survives on poor soils due to ability to
extract nitrogen compounds with the help of symbiotes. Above a liquid and on
prey floating there rhizopodes Drosarcella
and Droseroflugia –
descendants of Holocene genera Arcella and Difflugia respectively – are frequently
found. And the liquid inside the pitcher is inhabited by Endobodo
flagellates – descendants of Holocene Bodo genus. Rhizopodes and flagellates
are commensals near sundew’s “table”, finding in it shelter, livelihood and
moisture, so hard to in salty desert.
This species of plants was discovered by Biolog, the forum member.
Mauritian
false-berry tree (Pseudobaccodendron mauritianus)
Order: Caryophyllales (Caryophyllales)
Family: Goosefoots (Chenopodiaceae)
Habitat: forests of Mauritius Island.
In human epoch flora and fauna of Mauritius Island had put heavy on trail –
till some centuries as much species of plants and animals had been introduced
to this island, as, probable, had evolved her during millions years of natural
evolution. The degradation of primal flora and fauna of island and its full
extinction up to Neocene had been quite natural result of it.
In conditions when the primal Mauritian flora appeared in depression, the introduced
species had started to evolve actively. In favorable tropical conditions even
grassy plants became giants and had turned to trees. One of such trees, Mauritian
false-berry tree, is widespreaded in humid valleys and coasts of island. Its
ancestor is one species of orach (Atriplex) introduced to Mauritius by people.
At the process of evolution orach had turned from grassy plant at first to woody
half-shrub, then to bush, and farther to tree. At the continents the goosefoots
family is represented mainly by grassy plants, but occasionally among them treelike
forms appear. Saxaul (Haloxylon) is one of such species. It is adapted to extreme
desert conditions and shows an opportunity of occurrence of treelike plants
in family represented mostly by grasses. As against saxaul, false-berry tree
has not got used to such Spartan life: this is a plant of tropical forest preferring
moisture and an abundance of sunlight.
The height of false-berry tree may be up to 20 – 25 meters at thickness of ridge
trunk up to 3 meters at the basement. This is the tree with powerful deep root
system (as the answer to presence of hoofed mammals in Neocenic fauna of Mauritius)
and firm white wood. Bark of false-berry tree is thick, covered with deep longitudinal
cracks and colored light grey. The crone of this tree is similar to cypress
one – it is high and pointed to top.
Leaves of false-berry tree are externally similar to willow ones – they are
narrow and oblong, richly covering young sprouts. From above each leaf is covered
with microscopic bubbled hairs protecting from surplus of sunlight and giving
to tree crone the silvery shade appreciable from afar. There is especially large
amount of hairs on young leaves having pinkish shade. Hairs protect from solar
burns, disseminating light reaching the surface of leaves, but prevent the water
evaporation. However, the plant has the adaptation solving such problem: at
the edge of leaves hydatodes (water excreting glands) are located. From time
to time from them the drop of water emits and is quickly got rolled from the
leaf due to hair cover. Therefore, animal wandering in thickets of Mauritian
false-berry tree may get wet quickly even in dry day. This tree grows in river
valleys and near bogs.
False-berry tree blossoms extremely ordinary: tiny flowers pollinated by wind
do not have petals, and it is possible to detect externally the flowering of
tree only having examined its flowers specially. Tiny flower without petals
is the common feature of family representatives. But the fructifying tree is
appreciable from apart. This plant has received the name because of the appearance
of fruits. At the most part of plants belong to goosefoots family fruits are
usually dry, densely covered with wrapper of sepals and falling off in common
with it. The juicy fruit looks the exception from the rule. And false-berry
tree is just one of such exceptions. At it to the moment of seed ripening the
calyx does not dry out, but expands, overgrowing fruit from all sides, and becomes
juicy and even sweetish. Ripen fruit of false-berry tree in size is like a large
cherry. Its color also changes: the calyx becomes bright red and is appreciable
from afar on a background of silvery foliage, involving birds and monkeys eating
fruits. Ripen fruits are easily showered from wind, and they are eaten willingly
by local land animals: very small forest pigs and dryocaprellas – dwarf descendants
of goats.
When the seed of tree passes through the stomach of animal, its environment
becomes more permeable for water and it sprouts easier. Numerous germs of false-berry
tree perish or appear oppressed in shady underbrush. In shadow their growth
is strongly slowed down, and plants as though stop in growth at height no more
than one meter. The majority of such young plants may be eaten by local herbivores.
But when any old tree falls, young sprouts of false-berry tree start to grow
further quickly. In first year they grow up two meters, having “slipped by a
mouth” of local dwarf herbivores having the minimal damage. Further rate of
growth decreases, but the tree trunk starts grow thick and at its basis powerful
plank-buttress roots grow.
False-berry tree lives till 300 years and more, and starts to fructify from
10-years age, having reached about half of maximal height.
Five-winged
tree (Pentalaminodendron fragilis)
Order: Caryophyllales (Caryophyllales)
Family: Goosefoots (Chenopodiaceae)
Habitat: tropical forests of South-Eastern Asia.
Restoration of forests of South-Eastern Asia, suffered from human activity in
greatest degree, was accompanied by numerous attempts of grassy plants to colonize
these woods. A part of grassy plant species still plays a diminished role, being
among pioneer vegetation of the damaged habitats. But some of former grasses
have made successful attempt to turn to trees. One of such plants was orache
(Atriplex). On Mauritius island one descendant of this plant is Mauritian
false-berry tree (Pseudobaccodendron mauritianus). In the Asian tropics
an original five-winged tree grows – a plant of easily recognizeable appearance.
Among numerous trunks of various colors and bark textures, this tree species
may be distinguished without any mistake. This species is a tall (up to 30 meters
in height) tree with a pentahedral trunk. Already in young plant the trunk shows
the characteristic pentahedral shape with edges extended to slightly wavy crests,
with large alternated leaves, which arrangement corresponds to these edges.
It is a fast-growing and very photophilous tree – appearing in the “window”
of forest canopy, it gives an increment up to 5 meters per one year, and already
at the second year of life it beging to branch. During its whole life, this
species keeps five longitudinal crests along the whole trunk, and the cross-section
of tree trunk looks like a quinqueradiate star with thin beams. The width of
these edges can reach 2-3 meters. In the bottom part of a trunk they gradually
pass to plank-buttress roots expanding in various sides to the distance approximately
equal the width of crone of this tree. Crone of five-winged tree is wide and
umberella-like. Along the most part of a trunk, branches dry up and fall down
from lack of light. Branches are oriented strictly in directions of growth of
edges, and every edge continues along the bottom side of a branch, reinforcing
it. When the tree reaches the top level of forest, its growth slows down considerably.
Wood of this species is firm, but fragile, therefore wide edges on trunk serve
to give a tree more durability and stability, but simultaneously enable not
to increase huge mass of wood, occupying the top level of the forest. Leaves
of this species are wide, rhomboid, with tips extended to droppers. Young leaves
have a white farinose cover from both sides; on adult leaves, bottom side of
a leaf is covered with a layer of silvery trichomes, and top side is smooth
and bright green.
Flowering of this species is very inconspicuous – its flowers lack petals and
have five leathery sepals. They smell unpleasantly and are pollinated by beetles,
which eat a part of plentiful pollen. Flowers are composed to small racemes
growing on trunk and the bottom part of large branches, on narrow parts of edges.
Fruits of five-winged tree represent hard dry capsules full of firm seeds. They
may be opened only by birds and rodents dispersing its seeds. Seeds of this
species have inherited from an ancestor – an orache – an ability to keep germination
capacity for a long time and non-simultaneous germination. Sometimes seeds can
lay in ground up to 5 years before sprouts will appear from them. At five years’
age five-winged tree begins to blossom, and its life expectancy may be about
120 years.
Ivy-leaved
grape (Vitis hеderomorphus)
Order: Vitales (Vitales)
Family: Grapes (Vitaceae)
Habitat: humid forests of Caucasian Peninsula and the Balkan.
After climate warming in Neocene zones of tropical and subtropical climate have
considerably extended, and in Caucasus and Balkan regions the climate similar
to tropical one, with rather mild winter, was established. It resulted in appearance
of plants composing forests of these regions. In shapes of many endemic species
there are features more characteristic for plants of tropic areas.
A characteristic component of flora of these places is wild grape of various
species – descendants of cultivated grapes (Vitis vinifera), occupying various
ecological niches in plant community.
In humid shady thickets of forests, on mossy trunks of giant forest trees, numerous
species of epiphytic plants grow, including small descendant of cultural grape
– ivy-leaved grape, a dwarf plant with thin long stalks and small leaves. Like
all epiphytes, it sprouts on trunks of trees, taking roots in cracks of tree
bark and moss thickets. When plant’s stalk contacts a suitable substratum, on
it additional roots develop. On firm substrata plant gives numerous short roots,
which penetrate into the slightest cracks of substratum.
Vine of this plant is thin, bright green, poorly wooden and plentifully branching.
Leaves are small, pentagonal, with serrulate edge and thin “dropper” tip. With
age, on older parts of plant friable shelled bark grows. Length of stalks reaches
4-5 meters; they densely cover bottom parts of trunks of large trees.
Flowers are tiny, yellowish, composed in multifloral panicle. Fruits look like
small (not exceeding five millimeters) berries of violet color, gathered in
very dense cluster, more similar in structure and shape to large raspberry.
Plant fructifies twice, and in warmer regions – three times per one year. Seeds
are dispersed by birds eating berries. Young plant begins to blossom already
at the second year of life. In the winter, the plant loses a part of leaves
and runs into state of dormancy. Life expectancy is about 15 years.
This species of plants was discovered by Morgot, the forum member.
Mexican
clawleaf (Phyllonyx mexicanum)
Order: Fabales (Fabales)
Family: Peas (Fabaceae)
Habitat: deserts of Mexican plateau.
In deserts plants grow, able to resist to severe environment and to win in rigid
and uncompromising struggle for existence. At them there are two main enemies
– dryness and herbivores. And plants during the evolution develop various strategies.
In desert stretched on Mexican plateau, one of plants had adapted to repulsing
herbivores.
When in desert spring begins, from the ground accurately closed feather-like
leaves appear. They quickly grow, forming compact crowns. Each complex feather-like
leaves about 30 – 40 cm long consists of approximately ten pairs of oval simple
leaflets up to 3 cm long. On the tip of each leaf the middle nerve forms very
acute spike. This feature has determined the name of this plant – Mexican clawleaf.
Spikes are very widespread weapon of desert plants. But the clawleaf has added
to this feature of structure one more skill, characteristic for peas (however,
not only for them only). In the basis of each leaflet there is the layer of
tissue consisting of cells able to pump up water fast and to release it. It
gives remarkable properties to leaves.
Usually crowns of clawleaf freely lay on the ground, and leaves are stretched
towards to the sunlight. But if the herbivore will touch to them, the surprising
transformation comes: leaves start to close, folding along the middle nerve,
and simultaneously rise upwards. The plant literally before the eyes “bristles
up” with spikes. If it is not disturbed any more, in some minutes leaves take
former position again.
This plant maximally fully uses time favorable for growth. In spring clawleaf
vegetates actively, growing foliage. Not less roughly it grows under the ground.
This plant reserves water in underground tubers covered with thick peel. On
the peel there are large warts – it is not the display of disease, but the feature
absolutely necessary for life of plant. Colonies of nitrogen-fixing bacteria
live in these warts. These symbionts help the plant to receive nitrogen necessary
for growth right from air, and due to them clawleaf can settle on poorest soils,
and prospers in places where other plants decay. Growing up, clawleaf forms
affiliated tubers on long fragile stolons. When any large herbivore (for example,
bear porcupine, the characteristic
inhabitant of Mexican desert) digs ground and eats main tuber, stolons break
off, and affiliated tubers, not noticed by this animal, survive.
Soon after development of young leaves the plant begins to blossom. Flowers
of clawleaf are zygomorphic, with well advanced standard petal. They are brightly
colored – lilac with silvery strips converging in fauces of flower. Hairs have
property to reflect ultra-violet light, due to what insects notice them from
apart. This plant is pollinated mainly by bees and beetles. The nectar of flower
is hidden deeply in fauces, and to get it, the insect must creep in depth of
flower. At this moment stamens touch its back, and the insect appears strewn
with pollen. One of desert insects, cactus
hawkmoth, can simply “steal” nectar. This moth hangs in air in front of
flower, pushes long thin proboscis between petals, and exhausts nectar, not
making pollination.
Each flower blooms not for long – about four days. But this plant forms long
inflorescence on which flowers are placed in clusters, therefore as a whole
the plant blooms till two months and more. If the pollination has passed successfully,
fruits are formed approximately of third of flowers. To the moment of fruit
ripening the peduncle lies down on grass. Fruits at clawleaf are special – each
fruit contains only one large (up to 2 cm long) flat seed. Usually fruits of
leguminous plants dehiscent because of non-uniform drying of fibers penetrating
their parts. But at clawleaf the fruit is not dehiscent, and on its tip the
double bent spike covered with bristles develops. Such fruit resembles spoon-bait
with fish-hook, and on one peduncle it is total up to two tens of such fruits.
To the moment of ripening the fruit keeps on pedicel very poorly – it is ready
to meeting with animals which distribute them. Large praire
groundowl, marsupial hyena
or bear porcupine are main distributors of fruits of this plant. Having clung
to their covers, fruit of clawleaf is carried by them on large distance. Sooner
or later the animal combs it out from body covers, and fruit falls on the ground.
Near the shelter of owls or dust baths of bear porcupine congestions of this
plant form often. The seed keeps germinating capability for a long time, and
because of dense cover germination is usually delayed. It is the expedient tactics
– the plant will not sprout during the casual rain, after which it may be some
more weeks of hot dry weather, or during short-term winter warming. At the first
year of life clawleaf forms only the crown of short leaves; for the second year
leaves reach the typical size, and since the third year of life the plant blossoms.
Trickster tree (Pseudonecrodendron mimicus)
Order: Fabales (Fabales)
Family: Peas (Fabaceae)
Habitat: Japan Islands, humid mountain forests.
In the nature the phenomenon of a mimicry is very usual, at which the unprotected
species survives due to similarity to the protected neighbour. Usually animals
protect itself in such way, and insects, spiders and snakes in particular.
In Neocene there are even birds which imitate the protected neighbours. Among
plants examples of mimicry are much rare, but they also take place. At the
Japan Islands one of the largest species of toxic plants of the Earth grows
– “death tree” (Necrodendron mortalis). Poison literally impregnates it, protecting
reliably against the majority of herbivores. Only few insects can harmlessly
eat its leaves, and even use poison of this tree for self-defense.
One neighbour of “death tree”, the trickster tree, in due course of evolution
had got significant similarity to the deadly neighbour. This plant very precisely
imitates young growth of “death tree” – leaves of these species are very similar
at first sight, and also emit similar bitterish smell which considerably amplifies
at casual injuring of the leaf. But, when “death tree” belongs to the number
of giants of Japan forests, its imitator does not grow above five meters even
in its better condition. Moreover, in due course of growth this plant develops
numerous lateral shoots and forms multi-stalked bush.
Leaves of trickster tree are similar to a leaf of “death tree”, but they are
not compound, but deeply dissected simple ones. However, this feature has not
an effect for external similarity of two plants, and herbivores equally diligently
avoid eating their foliage. But trickster tree with all evidence “betrays itself”
during the flowering – the young growth of “death tree” simply does not begin
blossoming at such size. Flowers of trickster tree also differ in great degree
from ordinary-looking flowers of “death tree”, bearing characteristic features
of flowers of pea family plants. They develop in leaf axils and are white,
growing one by one, large (length is up to 3 – 4 cm) and well appreciable from
apart. Flowers of trickster tree are labiate and have dense petals; banner
is densely pressed against keel and the pollinator animal must make some efforts
in order to reach nectar. Flowers of trickster tree produce nectar at night
especially intensively because they are pollinated mainly by moths and small
species of bats. In the evening and in the early morning flowers of this plant
are visited by solitary bees. Occasionally beetles can reach nectar of these
flowers, simply biting through their petals at the basis.
Fruits of trickster tree represent long spirally twirled legumes containing
numerous large seeds with dense environment. At drying the ripened legume becomes
very sensitive to touches. The casual touch causes sharp twisting of sides;
thus seeds are scattered to great distance. Seeds of trickster tree can keep
viability during very long time. They are found frequently by rodents willingly
eating seeds of trees. Usually they eat seeds at once, but sometimes make stocks,
burying some of found seeds in secluded place. Frequently such stocks remain
unclaimed, and from them young trees grow.
Trickster tree is a photophilous plant. It grows plentifully in places where
any large tree has fallen and in forest canopy the gleam is formed. At sufficient
illumination trickster tree roughly develops and forms rich thickets which
constrain development of sprouts of other species of trees for many years.
When surrounding trees gradually close by their branches a gleam in forest
canopy, trickster tree gradually degrades. Its growth becomes less; the significant
part of branches dries out and crone becomes thiner. In shaded place this plant
grows and blossoms, but feels like oppressed and does not reach the maximal
size.
Hunting tree (Prolesrobinia venator)
Order: Fabales (Fabales)
Family: Peas (Fabaceae)
Habitat: steppe areas of Eurasia.
In XVII century various acacia species had been introduced to Eurasia as ornamental
plants – in human epoch trees of several genera had been known under this name.
Among them there was also clammy locust (Robinia viscosa). A distinctive attribute
of this 12-meter plant was a presence of sticky hair of glandulous downiness
richly covering sprouts, petioles and flowerstalks. After human disappearance,
during the period of early Neocene congelation this species appeared at the
edge of extinction, but extreme conditions had broken a unite area of species
to some isolated sites and this circumstance sped up evolution of this plant.
And in Neocene steppes of Eurasia the descendant of Robinia grows – the hunting
tree. Like fly-catching tomato, hunting tree displays an early stage of specialization
to feeding on insects and other small invertebrates though this specialization
is more expressed, rather than at fly-catching tomato. Hunting tree is the
largest predatory plant of Neocene, reaching up to one and a half meters in
height. In appearance this plant is similar to Holocene ancestor, and also
to other acacias of Holocene epoch. It has long odd-pinnate leaves consisting
of 8 – 11 pairs of small rounded leaflets.
As well as at ancestral species, sprouts, petioles and flowerstalks of hunting
tree are richly covered with sticky hairs. But, as against the ancestor, this
is an adaptation for hunting. A lot of thick and sticky liquid with the smell
attractive to insects is secreted on hairs. If the insect lands on this liquid,
it sticks and then hairs grow into prey’s body. Each hair represents huge cell
with strong cytomembrane. When the insect lands on leaf and sticks to hairs,
the substances secreting by its body stimulate the penetrating of hair top
into the tissues of insect. The top of hair starts to divide actively and sprouts
into the body of prey as a chain of thin-walled cells which absorb its liquid
contents. After prey is exhausted, hairs penetrated into it die off. Old leaves
stop the liquid secretion and participate in photosynthesis for certain time.
Hunting tree is dangerous even to small vertebrates like small tree frogs.
Growth of foliage is slowed down by an autumn. Young leaves stop growth and
secretion of sticky liquid, and old ones fall down. In spring growth of this
plant recommences.
Flowers of hunting tree are large and beautiful, having pinkish shade. They
are gathered in narrow double-row bunch and rise above the crone on long flower-bearing
stems, in order to prevent casual landing of pollinator insects on trapping
hairs. Near flowers lack of trapping hairs not to interfere with process of
pollination. Flowering of hunting tree takes place in the middle of summer
during two or three weeks. Fruits of this species represent long thin legumes.
Their sides at drying curl like hard spiral and scatter seeds far from parent
plant.
Hunting tree is photophilous, but it also loves damp ground. It grows at the
riverbanks and at the coasts of lakes, frequently on marshy ground. Within
the first years of life this plant develops due to symbiosis with bacteria
in root nodules, but in adult condition it receives a significant part of nitrogen
from the caught prey, and nodules on roots decrease. At the third – fifth year
of life this plant starts blossoming. Life expectancy does not exceed 20 years.
This species of plants was discovered by Odin, the forum member.
Antarctic
strawberry (Xerofragaria antarctica)
Order: Rosales (Rosales)
Family: Rosaceae (Rosaceae)
Habitat: tundra and meadows of Antarctica.

Picture by Pavel Volkov
After global warming and receding of glaciers, flora of Antarctica
was formed actually from zero, exclusively due to settling of plants from other
continents with the help of wind and birds. South America became the main source
of newly settling species: the main route of seasonal migrations of birds crossed
the Drake Passage. Getting in new habitats with rigorous conditions of life,
some species survived and gave rise to new Antarctic flora. Among plants of
Neocene Antarctica there are plants adapted to dispersing of their seeds by
numerous local birds. Antarctic strawberry belongs to their number; it is a
small grassy plant, which is widely settled across the continent and grows along
all coasts of Antarctica – from sandy beaches up to mountain areas, reaching
the altitude of one kilometer above sea level. Wetland subspecies of Antarctic
strawberry lives in boggy areas, including the strip along the edge of ice sheet.
As against many other plants of Neocene Antarctica, Antarctic strawberry was
accustomed at the continent for a long time, during some millions years. During
first millions years of Neocene, wild strawberry was a rare species in Antarctica,
dependent completely on migrations of birds. Before the beginning of their migration,
birds ate berries of wild strawberry of Tierra del Fuego and transported seeds
of this plant in their dung or on toes on the Antarctic ground. Here plants
sprouted, got accustomed temporarily and began to fructify, but in the cold
winter plants completely froze, and their population was supported only by migrating
birds. Only after some millions years, with gradual warming of global climate,
a stable population had established, and on the continent a species of local
Antarctic strawberry, which appeared capable to endure cold winter, has gradually
evolved.
Antarctic strawberry is a perennial dioecious grassy plant characteristic in
drought and cold resistance of its flowers and sets. It is a rosette plant with
short perennial stalk and fibrous root system growing not deep in the ground.
The whole plant is richly pubescent with numerous white trichomes – stalk, leafstalks
and flowerstalks, and the bottom side of leaves. Leaves are trifoliate, aeruginous
from the top side; leaflets are rounded with slightly serrated edges. Foliage
of this strawberry species does not die off and does not decay in the autumn,
but dries at its root and simultaneously curls in ball-like shape, protecting
rosette of young leaves and apical point in conditions of cold winter. In the
spring at snow thawing, last year’s leaves become wet, and leaf ball openes,
exposing apical point to sunlight. At this time active growth of plant begins.
The second type of plant shoots represents crawling sarments, on which young
plants are formed. During a vegetation season, adult plant gives 2-4 sarments
and forms up to 5 daughter plants.
Flowering begins soon after formation of the rosette of 3-4 fully developed
leaves. Flowers are white, on short flowerstalks – they do not rise above leaves.
Fruits represent characteristic achenes on juicy expanded receptacles of red
color, very large (from 2 up to 4 cm in diameter), with torulose surface. Dense
“berries” easily detach from calyx and vary in colouring from dim red up to
yellow and white, have juicy pulp and sweet taste.
Flowers are pollinated by beetles, flies, mosquitoes, local butterflies and
solitary wasps, and even by mousebirds
pecking insects on them. Many species of local beetles and birds eat these berries.
Flies mosquitoes, butterflies and bees can drink juice from a berry pecked by
bird.
Plant begins blossoming and fructifying at the third or fourth year of life.
Life expectancy is about 10 years. Old plant fructifies less actively and, as
a rule, dies off in the autumn, having given rise to daughter plants.
This species of plants was discovered by Wovoka, the forum member.
Spikeberry,
or Haides’ thorns (Rubus thornus-aidii)
Order: Rosales (Rosales)
Family: Rosaceae (Rosaceae)
Habitat: subtropical woods of foothills of the Caucasian ridge, Asia Minor,
the Balkan.
Rough expansion of heat-loving broadleaf forests at the coast of Fourseas has
resulted in occurrence of unusual shape in descendants of plants that had been
usual satellites of people in Holocene. Among such successful species there
was European blackberry (Rubus fruticosus), which prospered in cultural landscapes.
Tens millions years of evolution in the wild have generated a really demonic
shape at one of its descendants. This descendant of blackberry is spikeberry,
or Haides’ thorns, representing a giant perennial bush. Usually it forms extensive
thickets occupying a large area. Shoots of this species look like whips, reaching
the length of up to 10 meters at thickness of up to 5 centimeters. They grow
from a repent underground rhizome and during their growth bend their top parts
to the ground, forming something like an arc. Frequently old stalks lay their
tops on the ground and take roots, giving rise to new shoots. During their growth
shoots branch, and on tips of branches clusters of very sweet berries of dark
violet color grow ripe, attracting numerous animals. Birds are the main seed
dispersers of this species.
Old shoots wooden, but wood of this plant is fragile and friable, and trunks
are hollow. Colouring of bark is light brown; it exfoliates in thin paper-like
plates. Shoots of spikeberry are covered with the numerous bent thorns up to
3 centimeters long. On the oldest part of stalk thorns in due course fall off
together with a shelled bark. The bottom side of leaves along main nerves is
also covered with little spikes. Leaves are dense, dark green from above and
silvery because of hairiness from below, consist of three leaflets of about
equal size and have triangular outlines. Their edges are dentate and surface
is rugose. Spikeberry forms so rich thickets that some animals, having penetrated
on imprudence in its thickets in searches of berries, get in a deadly trap.
Young non-lignified shoots are armed with hard thorns, being also flexible and
rupture-resistant. They are easily capable to hook and to keep a careless animal.
As a rule, a panicing animal, trying to free, rushes in various sides and causes
to itself wounds by thorns of large old stalks and can die from loss of blood.
In depth of bushes it is frequently possible to see many skeletons and semidecomposed
corpses of animals, among which there is a lot of remains of cubs.
Haides’ thorns fructify during almost whole vegetative season – since May till
November. During almost all this time the plant blossoms, being the important
fodder resource for nectar-eating insects. Flowers of this plant are white,
pentapetalous, gathered in racemes of some tens ones. In winter at the north
of range and in mountain areas the plant sheds foliage completely, in warm regions
at the south of range only a part of the oldest leaves falls. At this time it
is in a condition of relative dormancy, and on shoots new flower buds are formed.
The age of many large bushes reaches a half of century, though old trunks exist
no more than 6-8 years.
This species of plants was discovered by Morgot, the forum member.
Malikaat
(Aromadryas malikaat)
Order: Rosales (Rosales)
Family: Rosaceae (Rosaceae)
Habitat: North American forest-tundra and tundra.
In Neocene, during short polar summer the North American tundra literally blossoms.
Despite of poverty of specific variety, tundra vegetation amazes with its quantity.
Snow is just thawn, and first flowers begin blossoming, and to the moment of
establishment of stable warm weather, tundra is already in bloom. Thicket of
Nearctic pine spread their pollen, pinkish
flowers decorate green thickets of qijuktaat,
and between them smaller plants grow – for example, polar aupillantinguat
saxifrage and crawling kinnikinnik
bush. One of the most colourful flowering plants of tundra is low-growing malikaat
opening its large white flowers.
Malikaat is a descendant of white dryad (Dryas octopetala), low-growing grassy
plant of Holocene epoch. Relationship of these species is shown by its flowers
– like flowers of its ancestor, they are large (diameter up to 8 centimeters),
white with yellow center, which is formed of numerous stamens and pistils. In
their colouring, they resemble a blossom of camomile, only of a little bit larger
size: malikaat plant is 15-20 cm tall on the average. However, it is not a capitulum
like at a camomile, but a single flower: malikaat belongs to different family
of plants. On stalks of plant, the rests of leafstalks are kept for a long time.
They dry up and serve as a kind of spikes, protecting a plant in some degree
against large herbivorous mammals. But on malikaat leaves caterpillars of various
local butterflies feed. In winter the greenery of this plant is eaten under
snow by rodents.
Malikaat is an evergreen cold-resistant plant, which does not suffer from polar
colds, like its neighbours like kinnikinnik. At the same time this species breeds
only by seeds. As against the ancestor, which seeds were dispersed by wind,
malikaat has evolved to adapt to zoochory. Seeds at their ripening quickly lose
fluffy outgrowths, but are buried into the developing receptacle having pleasant
smell attracting herbivorous mammals (hence the genus name meaning “fragrant
dryad”), which nibble the plant, gathering seeds from it. Having gone through
a digestive path of an animal, seeds drop out with its manure and due to processing
by gastric juice sprout faster in the next spring. Because of this feature of
settling malikaat usually grows in well defined places – along migration ways
of skewhorns and similar mammals. During the flowering
of this species, tracks of animals become especially visible on the background
of tundra landscape. As the plant does not form long crawling shoots and is
dispersed exclusively at seed stage, its population is less numerous, than populations
of other polar plants. However, it is compensated by endurance of this plant
to trampling and its significant longevity – a particular bush can be 40-60
years old.
This species of plants was discovered by Bhut, the forum member.
Vigorous
lichenomorpha (Lichenomorpha vivax)
Order: Malpighiales (Malpighiales)
Family: Riverweeds (Podostemaceae)
Habitat: western Madagascar, seasonal flowing reservoirs.
Riverweeds – original water grasses, at which a significant part of a body is
presented by thallus – belong to the most specialized aquatic plants. Thallus
at these plants is a flattened formation resembling thallus of macroalgae, making
a significant part of mass of a plant and tightly attaching to substratum. Riverweeds
are thermophilic plants widespread mainly in tropics of both hemispheres.
In the western part of Madagascar, freshwater reservoirs are inconstant and
dry up for the term of from several weeks to about several months. Aquatic plants
living in them have developed during evolution ability to endure a drought.
Some representatives of riverweeds developed reservoirs of such type also have
such features. One common species in the western part of island is vigorous
lichenomorpha, a plant capable to endure significant drying up. It is rather
large species: length of thallus of adult specimens reaches 30-40 cm. Thallus
is tightly attached to substratum (stones) and is covered with thick skin, and
at drying of a reservoir on covers of a plant wax layer is formed, due to what
this species is capable to endure exposing to the air within several months.
In a dry season, vigorous lichenomorpha exists as a dry crust on the stones,
resembling lichens – hence the name of this species. Thallus has irregular leaf-like
shape and is usually divided into some blades having wavy edges and indistinctly
separated from each other. In a dry season edges dry up and crumble, but under
water they grow intensively and restore the form. Colouring of thallus in growth
condition is bright green, in brightly lit place its edges redden.
In tissues of thallus a plenty of buds – both vegetative and generative – is
formed. In rain season thallus quickly comes to life; within several days buds
sprout and the vegetative shoots looking like thalluses of macroalgae – flat,
transparent, very wide, leaf-shaped with pointed tips – develop. First shoots
are simple, later ones are furcated one or twice. Plant forms some tens vegetative
shoots. During the vegetation time, in thallus a significant amount of starch
and proteins is stored.
At the end of rain season, when the water level is declined, vegetative shoots
appear on the surface of water and begin to dry up. It is a signal for the beginning
of flowering, and the plant produces numerous tiny flowers with reduced perianth.
The height of flowerstalks does not exceed 3-4 cm. The plant is wind-pollinated,
less often self-pollinated. Seed is covered with a sticky slimy tegument. Seeds
stick to legs of waterfowl and are dispersed by them to other reservoirs. In
the first 2-3 years of life, young plant develops only vegetative shoots and
grows mass of thallus; later in begins to blossom annually. Life expectancy
reaches 20 years and more.
Najad’s
hair (Hydrocapillatura najadalis)
Order: Malpighiales (Malpighiales)
Family: Riverweeds (Podostemaceae)
Habitat: mountain rivers of Southern Europe, southern slopes of Alps.
Changes of climate in Neocene epoch have changed ranges of various groups of
plants, having enabled warm-loving species to settle into higher latitudes.
Riverweeds, a specialized family of aquatic plants, in Neocene has considerably
expanded its range, and its representatives became characteristic aquatic plants
of flowing waters of Europe. Some species of these plants occupied even high-mountainous
areas – apparently, following routes of migrations of waterfowl dispersing seeds
on their legs and plumage. In new habitats after millions years new species
of these plants had evolved. One of such species is an Alpine plant named naiad’s
hair.
This species is a typical representative of riverweeds; the significant part
of a body of this plant is transformed into crawling thallus tightly attaching
to substratum. Branches of thallus resemble flat twisting tapes with slightly
wavy edges. On the top side of thallus numerous vegetative and flower buds appear.
Plant has clearly expressed seasonal prevalence of the vegetation connected
to a water level in rivers. This species lives lives in coastal shallow waters
at the depth not exceeding 20-30 cm.
The main period of vegetation includes winter and spring, when rivers are deeper
due to thawing of glaciers. Vegetative buds develop at this time, and plant
produces long ribbon-like vegetative shoots spreading in current and fluttering
in a stream of water, which resemble hair of green color (hence the name). The
length of such shoots reaches 20-50 cm at width not exceeding 3 mm. At this
time at this plant active photosynthesis takes place and it accumulates in thallus
nutrients necessary for successful flowering.
In the summer, when the water level in rivers is declining and current is slowed
down, the part of thalluses appears above water. At this time on them flower
buds begin to develop and plant blossoms plentifully. Flowers are tiny, have
extremely reduced perianth, thin flowerstalk and light pink color. Numerous
single flowers are gathered in bunches sticking out on small knobbles above
thallus surface. Pollination is performed by wind or by casual insects. Some
flowers are eaten by snails and water scavenger beetles.
Tiny seeds are covered with sticky slime and easily paste to plumage, legs and
beaks of shorebirds, which execute the settling of this plant. During first
two years of life sprout forms thallus and occupies vital space; at the third
year of life it forms some vegetative shoots. Flowering of a young plant takes
place at 4-th year of life.
Saharan
potamomatrix (Potamomatrix saharensis)
Order: Malpighiales (Malpighiales)
Family: Riverweeds (Podostemaceae)
Habitat: Africa, Saharan Nile basin, sesonal water bodies and drying parts of
riverbed.
Riverweeds are deeply specialized underwater grasses, and during evolution they
have undergone radical changes in anatomy in comparison with terrestrial plants.
For the moost part they occupy not drying up reservoirs, however some species
managed to develop the habitats not too suitable to life of specialized aquatic
plants. One of such plants is Saharan potamomatrix inhabiting Saharan Nile basin.
It is an atypical species of riverweeds adapted to life in slowly flowing water
and in drying up rivers. During evolution this plant has adapted to survive
in drying up reservoir, despite of specialization to life under water. Thallus
of this species is corticated; it develops on firm substratum – usually on a
stone or a sunken tree – and is tightly attached to it. From it numerous thin
photosynthesizing shoots grow – they are ribbon-like, transparent and lack leaves.
They reach length of 30-80 cm and spread on water surface. In shadow they have
green color, but get pinkish color in well-lit places. In rain season among
them elastic flowerstalks grow quickly, towering above waters surface and bearing
modest-looking spike-like inflorescences. Seeds are very small and sticky; they
are dispersed on legs and plumage of shorebirds and waterfowl.
At drying of reservoir flowerstalks and photosynthesizing shoots die off, and
thallus forms brood buds covered with a multilayered tegument and capable to
endure drying. Slime covering them stiffens as a crust protecting a plant from
drying and death. Brood buds easily detach from thallus and are dispersed by
wind and birds. At wetting slime quickly swells and pastes bud to substratum.
The growth of bud is delayed for some days, and it absorbs water at the same
time. If the reservoir does not dry up, and water supply does not interrupt,
it becomes a signal to continuation of growth. During river floods, potamomatryx
forms new shoots quickly and expands actively. A young plant usually begins
to blossom the next year, and seed breeding it happens at the third year of
life. Life expectancy of a plant makes up to 5-6 years.
Dendrofuchsia,
fuchsia tree (Dendrofuchsia penduliflora)
Order: Myrtales (Myrtales)
Family: Willowherbs (Onagraceae)
Habitat: temperate areas of South America, northern part of Tierra del Fuego.

Picture by FanboyPhilosopher (flowers are pollinating by tajin hummingbird)
Plants of fuchsia genus (Fuchsia) in human epoch had been
a characteristic component of flora of temperate and subtropical areas of South
America. They represented bushes with characteristic hanging flowers of tubular
shape pollinated by butterflies, moths and birds. In Neocene one species of
this genus had made a successful attempt to develop the new vital form, and
had turned to small tree. Nevertheless, this vital form is rather changeable:
at the north of an area this plant represents semi-deciduous tree up to 8 meters
in height, but at the south of area it sharply decreases in growth and turns
first to multi-stalked bush, and then to deciduous elfin wood form with creeping
trunk about 10 meters long which winters under snow. It is dendrofuchsia, or
fuchsia tree.
Leaves of this plant are leathery, dark green and ovate with the dulled tip.
Surface of leaf between large nerves is slightly inflated. At the north of an
area in the beginning of winter only old leaves fall, and at the south of area
the plant loses foliage for winter completely. If young sprouts freeze slightly,
the plant is quickly restored from resting buds. Because of it at the south
of an area this plant usually has richly ramified sprouts. At the north of an
area sprouts of dendrofuchsia are long and slightly hanging. Wood of this plant
is friable and rather fragile. At the north of an area where plants keep foliage
for winter, during sudden snowfalls the trunk of dendrofuchsia can not sustain
loading and breaks. But the plant does not perish – its sprouts take roots easily,
and the plant continues development as a lying bush. At the first years of life
the annual growth of dendrofuchsia reaches 1 – 2 meters.
During the flowering dendrofuchsia represents a true gem of forests of the utmost
south of South America poor in bright colors. In spring, when the temperature
of air raises and days become longer, on tree numerous flowers blossom. Hanging
flowers of tubular shape with four peaked sepals and four rounded petals are
very similar to flowers of common fuchsia. They are formed on sprouts of the
last year. One flower-bearing stem bears a cluster of three or four flowers.
Flowers are changeable in colouring, but are always colored in shades of red
color which is distinguished well by birds, their pollinators. In the most typical
case calyx of flower is dark red, and petals are carmine pink. During the flowering
the tree is entirely covered with flowers, and the foliage is hardly distinguished
among the mass of flowers. Blossoming dendrofuchsia is easily seen from apart
– the red mass of flowers is notable on the background of surrounding bushes
and foliage of Nothofagus – dominant kind of plants of the utmost south of South
America. This plant is pollinated by local butterflies, moths and migrating
tayin hummingbird. Dendrofuchsia
is exclusively cross-pollinating plant.
Fruits of this plant are berry-like and sapful, with thin skin and tiny seeds.
They are distributed by fruit-eating mammals and birds. Life expectancy of dendrofuchsia
does not exceed 60-80 years.
Undersnow
fuchsia (Fuchsia subnivalis)
Order: Myrtales (Myrtales)
Family: Willowherbs (Onagraceae)
Habitat: far south of South America, Antarctica (Antarctic Peninsula), plains,
stony barren lands.
In human epoch various species and hybrids of fuchsias were popular indoor plants
– they were fast-growing, plentifully blossoming and evergreen. In some areas
of the Earth they survived for any time after human disappearance, more or less
successfully winning for them places in ecosystems of early Neocene. In their
homeland – in South America – fuchsias have evolved actively, adapting to changing
conditions at the boundary of Holocene and Neocene. Some of their species have
adapted to extreme conditions of high mountains and high latitudes with the
expressed seasonal climate and snowy winter. One of such extremal species is
undersnow fuchsia, which managed to adapt to cold climate, to cross the Drake
Passage and to occupy Antarctica.
This plant is semifrutescent crawling fuchsia with wooden perennial shoots,
which take roots in ground between stones and give a plentiful young growth.
The length of shoots can reach 150-200 cm, but the height of plant does not
exceed 6-10 cm. This species is an evergreen plant with very small rounded leaves
with short leafstalks. Leaf surface is leathery and smooth; edges are serrulated.
Foliage accumulates great amount of glycosides, therefore it has very bitter
taste and is inedible for local herbivorous birds – meadow
swans. Plant successfully winters under snow, frequently keeping its flowers.
The plant blossoms during almost the whole
season of vegetation, but especially plentifully in the spring, after an advent
of warm weather and snow thawing. Its flowers are very bright – petals are red
with white nerves, sepals are green with red spots and red from within. Nerves
reflect ultra-violet rays and it reliably attracts rare local butterflies, which
are pollinators of the plant. Flowers are pendant, having vertical flowerstalks
rising to 9-12 cm above the ground. Such arrangement of flowers attracts also
the Antarctic mousebirds,
which lick its flowers, standing on the ground. After pollination bright red
berries appear, being a delicacy for local birds. Their pulp contains numerous
tiny seeds, which frequently stick to legs and plumage of birds, due to what
the plant is settled even to isolated islands, where at absence of pollinators
it uses vegetative propagation. Young plant begins to blossom at the age of
5-6 years, life expectancy is about 80 years.
Siberian
fireweed (Chamerion sibiricus)
Order: Myrtales (Myrtales)
Family: Willowherbs (Onagraceae)
Habitat: Siberia, areas of broken forest, “obda roads”.
In human epoch rosebay willowherb (Chamerion angustifolium) was one of the most
usual plant species in human-destructed habitats, where it prospered, reaching
great numbers. Due to ability to settle and to grow on poor soils this plant
has kept high number of populations in epoch of biological crisis, and after
stabilization of natural conditions in early Neocene it became an ancestor of
several species living in Northern hemisphere. One descendant of this species
is Siberian fireweed living in biotopes establishing as a result of activity
of local megafauna breaking continuous thickets of
trees. This is rather tall grassy plant reaching 50-150 cm in height: on wet
ground it expands more plentifully, forming a bush of ten and more stalks. It
usually grows in communities of grassy plants along “obda roads”, but sometimes
appears also far from them, in sites of the broken forest stand. Plant’s stalk
is strong, tetrahedral in cross-section, of reddish color, especially at the
basis. Leaves are narrow, especially ones growing in the end and in the beginning
of growing season. Colouring of leaves is bright green from above and lighter
from below. This plant represents a food source for obda,
local harelope species,
shurga, various rodents and
other herbivores, and also for insects. After being eaten up by mammals, plant
restores its biomass very soon, and has time to finish blossoming and to produce
seeds, but in this case it blossoms less plentifully.
Flowers grow in rich cylindrical racemes on tops of stalks and have red or pink
color; during the peak of vegetation shade of petals turns brighter. Flowers
have four rounded petals. This species is pollinated by various insects, mainly
dipterans and hymenopters, less often by butterflies. At the end of summer fruits
ripen – dry capsules containing small seeds with the long silky “parachutes”
enabling them to fly away with wind. Seed teguments are rather hard and can
easily endure the way through digestive system of large herbivorous animals
and rigorous Siberian winters. Sometimes the ripened seeds are dispersed by
local herbivores – having eaten a plant with ripen seeds, the animal disperses
its seeds with manure. In the spring seeds germinate and rather soon reach height
of half meter and more. Flowering usually begins at the second year of life.
The plant prefers good lighting with shading in hot midday hours. For winter
Siberian fireweed dries up, but its dry stalks are kept for a long time and
represent a part of winter diet of herbivores. In Eurasia related species (probably,
only local forms) grow, differing in colouration of their flowers: white, yellow,
pastel etc.
This species of plants was discovered by Bhut, the forum member.
Hardy
fireweed (Chamerion endurabilis)
Order: Myrtales (Myrtales)
Family: Willowherbs (Onagraceae)
Habitat: North America, open areas – North American prairies, tundra.
One of the main features of various species of fireweed (Chamerion) is this
plant’s rate of colonization of open areas, and also indiscriminateness to climatic
conditions and inhabitancy. It helped rosebay willowherb (Chamerion angustifolius)
to survive both in epoch of anthropogenous pressure in Holocene, and during
the ice age of early Neocene. In warm postglacial time the plant has continued
to prosper, and its descendants have occupied various ecological niches in the
nature of Northern hemisphere.
Hardy fireweed is a typical representative of flora of North American open areas.
This is a tall grassy plant, usually 100-150 cm tall, with thin graceful stalks,
narrow leaves and large racemes of pink or dark pink blossoms on tops of stalks.
At the north of the range hardy fireweed blossoms in second half of polar summer,
adding more bright colors to short polar summer, and at the south, in warmer
climate it takes place in second half of summer and in early autumn. It makes
this plant a very important food source for many North American insects, which
actively pollinate its flowers before winter colds. Frequently even migrating
hummingbirds moving to wintering areas are engaged in it. Autumn flowering is
connected to droughty summer, when plant stops its growth and saves moisture.
At the end of summer, the plant literally “flashes” with bright colors and fades
completely for two weeks. Large herbivores like donkeyhorses
and even peccasons willingly
eat this species – this plant grows at open areas in extensive thickets and
represents a plentiful source of forage.
Hardy fireweed propagates by wind-carried seeds; representatives of this species
growing together in small territory, blossom and produce seeds very synchronously.
Seeds are very hardy, successfully winter in new place under snow, and start
to sprout roughly in the spring. In second half of summer they frequently are
already adult and can blossom. In tundra, the young plant comes into full force
and blossoms only at the third year of life, and flowering begins earlier, than
in the south of range.
This species is a perennial plant; underground part with fleshy roots enables
it to be restored after being eaten by herbivores. Duration of existence of
thickets exceeds twenty years and in tundra up to 60 years and is more.
This species of plants was discovered by Bhut, the forum member.
Riverine
almond (Neoterminalia riparia)
Order: Myrtales (Myrtales)
Family: Leadwood (Combretaceae)
Habitat: gallery forests near the river courses of central and northeastern
South America.
During the Holocene, the man introduced many species, animal and vegetal, for
ornamental purposes. The tropical almond (Terminalia catappa) was one of these
introduced species. During the mid-20th century of the human epoch, this tree
was used extensively in Brazilian urban landscaping, since being a rare case
tropical deciduous, their fallen leaves would give a “European” flair to the
street. This species adapted very well and in the Neocene they give rise to
the Genus Neoterminalia, with the riverine almond being the type species. This
tree forms, together with the violet
trumpet tree (Tabebuia violacea) and the dendém
oil palm (Elaeis dendem), the gallery forests that exist near the river
courses of Central and Northeastern South America, being only fond in those
riparian areas.
The riverine almond grows to 30 m tall, with an upright, symmetrical crown and
horizontal branches. As the tree gets older, its crown becomes more flattened
to form a spreading, vase shape. Its branches are distinctively arranged in
tiers. The leaves are large, 23 cm long and 12 cm broad, ovoid, glossy dark
green, and leathery. They are dry-season deciduous; before falling, they turn
yellow-brown.
The trees are monoecious, with distinct male and female flowers on the same
tree. Both are 3 cm in diameter, whitish pink, fragrant with no petals; they
are produced on axillary or terminal spikes. The fruit is a drupe 6 cm long,
green at first and red when ripe, containing a single seed. The fruit is dispersed
by water and animals. In comparison with that of their ancestors, the fruits
are meatier; this way the animal will consume the pericarp leaving the seed.
The flowers are pollinized by insects.
Other species in this genus is coastal
almond (Neoterminalia atlantica). Larger than the riverine almond,
growing to 40 m tall, this species are components of the Atlantic forests of
Southeastern South America. Their 5 cm long fruits are yellow when ripe and
the flowers are white with no petals. This species lost less leaves in dry seasons
than the riverine species.
This plant species was discovered by João Vitor Coutinho, Brazil.
Armourless
paranepenthes (Paranepenthes dearmatus)
Order: Nepenthales (Nepenthales)
Family: Nepenthaceae (Nepenthaceae)
Habitat: Southeast Asia, Jakarta Coast, Sunda Land; rainforest canopy.
Among branches of trees and sprouts of various lianas of tropical rainforest
of Southeast Asia there are various species of carnivorous plants of family
Nepenthaceae - true Old World pitcher plants, and related to them flowerleaf
having more specialized trapping leaves imitating flowers of some plants. But
in the neighbourhood with these plants their relative grows, which, as opposed
to these plants, had developed completely other form of relations with small
inhabitants of forest canopy. This epiphytic plant is named armourless paranepenthes.
It has long creeping stalks covered with air roots. It attaches to thickets
of moss and creeps on bark, having densely attached to layer of moss covering
it by very short sucker-like roots. The stalk of this plant reaches length of
3 – 5 meters and more.
Leaves of paranepenthes have the shape characteristic for representatives of
family: they are large, pitcher-looking, and their volume reaches two liters.
Edges of leaves are colored dimly, and cap is wide and has long outgrowths on
edges. Leafstalks, as at true Nepenthes, had undertaken a role of photosynthesizing
organs – they are strong, expanded leaf-likely with rounded outlines.
“Standard” representatives of family use pitcher-like leaves as traps, but paranepenthes
has no nectaries and digestive glands on the internal side of pitcher leaf (hence
the name). In leaves rain water simply accumulates, and it strongly involves
various climbing animals. The plant has entered symbiosis with them, and uses
their dung and excretions as fertilizer, receiving from them mineral and organic
substances necessary for growth. Climbing frogs and crabs living in forest canopy
use these leaves as constant shelter. Besides various mosquitoes lay eggs in
leaves of paranepenthes. To use substances emitted by its “lodgers”, in the
bottom part of pitcher leaves of paranepenthes fibers develop, with which help
the plant absorbs excretions of animals. Sometimes air roots from overlying
sprouts grow gown in leaves.
The adult plant of armourless paranepenthes develops up to 20 – 25 normally
advanced leaves and turns to “apartment house” for various tree-climbing frogs
and other moistureloving animals.
Because the plant is not “interested” in insects as food source, it has adapted
to pollination by butterflies and hymenopters. Its 4-petaled flowers about 3
cm in diameter are gathered to big spike-like inflorescences. They are colored
bright pink and emit sweetish smell.
Aupillatinguat
(Saxifraga aupillatinguat)
Order: Saxifrages (Saxifragales)
Family: Saxifragaceae (Saxifragaceae)
Habitat: tundra of North America, stony areas.
Glaciers rendered significant influence on the lay of the land in the Arctic
latitudes of Eurasia and North America. Moving to the south, they raked up layers
of sediments, and after their thawing at the district there were the areas actually
lack of humus layer. Only among stones, a small amount of ground accumulated,
and it enabled to exist fairly well to plant species, which have got used to
poor conditions of life in tundra.
In the beginning of a polar summer, stony districts created by glaciers turn
bright and multicolored. Light pink bell-like blossoms rock on shoot tips of
qijuktaat, the descendant of Arctic bell-heather
(Cassiope tetragona) of human epoch, and nearby from cracks between stones as
if lilac-pink foam splashes: it is a blossoming of another plant, aupillatinguat
a descendant of purple mountain saxifrage (Saxifraga oppositifolia). Aupillatinguat
is the Eskimo name of saxifrage. Like many saxifrages of Holocene and Neocene,
aupillatinguat does not form separate bushes like qijuktaat does, but crawls
on the ground, branching plentifully and forming rather dense carpet 5-10 cm
tall. Its monotonous greens is broken, and during the beginning of flowering
is completely hidden by star-shaped flowers of violet color with 6-8 large petals.
They are large (up to 30 mm in diameter) and well-visible, as against scales
which replace leaves at aupillatinguat like at many other polar plants. These
flowers are pollinated by various polar insects, and aupillatinguat is the important
fodder plant for insects, drawing their attention with “honey marks” – strips
and spots. Shoots of this plant are truncated and thickened, resemble little
tetrahedral green strobiles. Root system is shallow and is made of numerous
short roots.
Aupillatinguat does not synthesize resinous “antifreeze” substances in tissues,
but the size of a plant enables it to hide completely under snow and to spend
rigorous polar winter in relative heat. Large herbivores like skewhorns
are capable to dig it out from under snow, but its mean of protection is its
growth in “inconvenient” places among stones, where a large animal not always
succeed to nibble it. Various small rodents easily find and eat this plant under
snow, but aupillatinguat is very hardy and is capable to propagate not only
by seeds, but also vegetatively – by radical shoots and branches easily taking
roots. Large animals eat blossoms of this plant more often – it may be too difficult
to bite off stalks because of their specific shape. Seed capsules at this species
are elevated on long stalks above a level of flowers and easily dehisce, and
the surface of seeds is supplied with numerous fibers. Therefore, skewhorns
inevitably touch them when trying to eat flowers. At a touch seed capsules dehisce
and clinging seeds stick to a wool of animals then to fall somewhere in other
part of tundra.
Life expectancy of aupillatinguat makes about 20 years.
This species of plants was discovered by Bhut, the forum member.
Egg-leaved
wood crassula, “egg tree” (Dendrocrassulopsis ovifolium)
Order: Saxifrages (Saxifragales)
Family: Stonecrops (Crassulaceae)
Habitat: Southwest Asia, Atlas mountains.
Ice ages are characterized by climate drying: huge amounts of water were accumulated
in glacier, the level of ocean lowers, and at the place of shallow seas land
areas appear. Evaporation of water is reduced, and as a consequence, smaller
amount of rains falls. Expansion of area of continents results in sharper temperature
contrast between coasts and land areas far from sea. In such conditions number
of moisture-loving plants is reduced, but hardy species, able to reserve water
in organs and endure cool winter climate, prosper. In Old World among such hardy
plants it is possible to note crassulas family, distinguished by significant
variety. When in ice age instead of Mediterranean sea the salt desert had appeared,
at its edges, rather free from salt, various crassulas prospered. Later, when
climate began damper, some of their representatives had got absolutely fantastical
appearance.
Wood crassula is heavy-build succulent tree with creeping trunk which is easily
taking roots in ground. Its height is not so big: about 3 - 4 meters, but the
trunk frequently reaches 1 meter thickness. Roots of this plant grow near the
ground surface and do not grow deeply in ground, but they are spread to big
area, permitting to gather rain water very fast even after the lightest rain
which had hardly moistened dust. At drought the part of roots may die off, but
in rain season they restore very quickly. Superficial root system is a bad support
for trunk of plant, therefore wind frequently fells massive plant down. However
it does not cause inconveniences to the plant: tumbled down sprouts quickly
grow roots, and tree continues life.
The bark of wood crassula is very thick and friable – it serves for thermo isolation,
protecting tree trunk from solar burns and overheat.
Leaves of this plant are thick, prolonged-spherical form, covered with thin
wax film, gathered at tops of sprouts. Their length is about 5 cm at thickness
up to 4 cm. In leaves lifted above the ground and inaccessible to the majority
herbivores, the significant part of stock of moisture, which has been saved
up by this plant, is concentrated. Besides each leaf is a pledge of original
“immortality” of plant: as well as all crassulas, this plant may multiply by
rooting of separate leaves.
This original plant lives not for long: no more than 10 years, giving for this
time a significant biomass due to leaves have taken roots on the ground and
branches grown in different sides.
Wood crassula concerns to monocarpic flowering plants, and it means, each plant
of this species blooms only once per life, and after flowering quickly dies
off. Flowering begins at the end of dry season. On each sprout ordinary-looking
white flowers open in a plenty, emitting out strong aroma. To them flies and
beetles living in desert fly basically. Flowering proceeds a long time: about
one month. For this time the irreversible exhaustion of plant begins: leaves
collapse, becoming similar on wrinkled sacks, and eventually cast. The trunk
of plant appreciably “grows thin”, on it longitudinal furrows appear, and bark
falls off by large pieces. To time of seed ripening tree is completely exhausted,
and during damp season completely decays. But seeds have fine chances to survive:
during rain season they have time to sprout and young plant may save up enough
moisture for surviving in drought.
Wooly
cabbage (Platyaeonium lanatum)
Order: Saxifrages (Saxifragales)
Family: Stonecrops (Crassulaceae)
Habitat: Zinj Land, Usambara Mountains.
In Neocene epoch split of Africa along the line of Great Rift walley has resulted
to that all highest mountain tops of continent appeared at the East-African
subcontinent, Zinj Land. In some senses the highest mountain tops are similar
to islands in the sea: these are the islets, where the special environmental
conditions establish, surrounded by extensive territories, where conditions
of life are strictly different. Isolation of these territories results in formation
of very special flora and fauna at mountaintops of tropical latitudes.
In Usambara Mountains, in the top part of cloud forest zone, an unique species
of perennial grassy plants of original shape – wooly cabbage – grows. This plant
really resembles externally a heap of cabbage heads with leaves covered with
numerous white trichomes laying on the ground. On closer examination, it appears
that this plant is not related to cabbage and has only the remote external similarity
to this species. Wooly cabbage is a descendant of one species of Aeonium genus,
widespread in Africa in human epoch.
The adaptation to life in mountain areas has caused significant changes in shape
of a plant. Ancestors of this species were bushy plants with flat rosettes of
leaves on tips of branches. Life in mountains has transformed wooly cabbage
into a creeping plant, which stalk trails on the ground and branches plentifully,
taking roots along its whole length. Length of stalk can reach 2 meters. Leaf
rosettes at this species are large, up to 30 cm in diameter. They grow on ground
surface rather densely, forming cushion-like thickets, and between them, only
few plants of other species can grow. Leaves have spoon-like shape with expanded
tops and bend to the centre of rosette, covering apical point. Bottom side and
edge of leaf are plentifully pubescent with white trichomes; top side is covered
with wax – it is a protection against intensive ultra-violet radiation. Leaf
blade is succulent, is up to 10 cm wide and about 1 cm thick, of gristly consistence.
Stalk of plant during the growth period bears small tongue-shaped leaves, which
die off very soon. Having got out of a shadow of already formed rosettes, tip
of stalk takes roots and begins to form a new rosette. In the first year, it
reaches diameter of about 10 cm, for the third year it gets maximal diameter
and begins accumulation of the nutrients necessary for flowering. At the fifth
or sixth year of existence the rosette begins to blossom.
Flowering of wooly cabbage occurs very intensively. At the basis of each advanced
rosette a stalk reaching two-meter height is quickly formed. Flowers of this
species have star-like shape and narrow white petals, are composed to a large
compound raceme of conic shape. Bunches of flowers produce a honey smell and
plentifully produce nectar attracting bats and butterflies – the basic pollinators
of this plant. Seeds are tiny, supplied with hairs and dispersed by wind. After
flowering rosettes of a plant die off, and from the whole plant only youngest,
no more than 2 years old rosettes remain. They continue to grow and in due course
form a clone of a parental plant.
Below on mountain slopes the closely related species – waxy
parasol (Platyaeonium umbellatum) living on rockslides – grows. It
also has a trailing stalk, and flat umberella-like rosettes up to 50 sm in diameter
grow on rising stalks. Leaves have tongue-like shape, a layer of plant wax from
above, especially plentiful near the obtuse top of leaf. Plant is very photophilous
and due to dense rosettes of leaves, it simply shades its competitors. Flowers
of this species have carmine-red color and slight smell, being pollinated mainly
by birds and wasps scraping wax from leaves of plant for building of their nests.
Flower stalk is formed at the basis of rosette and sprouts upwards between leaves.
Rock
fig (Ficus platycaulis)
Order: Urticales (Urticales)
Family: Moraceae (Moraceae)
Habitat: mountains of Southern and South-Eastern Asia, steep slopes.
In human epoch figs differed in big variety of vital forms: among them it was
possible to meet trees, bushes and lianas of any size. Figs may form the original
vital form banyan: one tree, expanding in sides and giving numerous prop roots,
may form the true wood. Among figs “suppressors” squeezing by numerous roots
tree trunk in which forked crown they begin growing, and gradually suppress
and replace it in forest canopy, are also known. In Neocene among various figs
in tropics of Old World the climbing species have mastered hillsides have appeared.
Rock fig grows in conditions of plentiful rains and high temperature the year
round. This plant begins its life as an epiphyte when its small seed gets in
crack of rock where moss and vegetative dust accumulate. The sprout quickly
starts growing, clinging by rich brush of adventitious roots for the slightest
cracks. Gradually thin stalk expands and reaches next cracks where it fixes
and continues reaching upwards. Gradually stalk becomes thicker, lignifies,
and roots start to penetrate into cracks of rocks, filling and fastening them
more deeply, as if cement. The stalk of adult plant accrues more in width,
than in thickness: it is flat and band-shaped. The plant plentifully branches,
covering with itself huge areas of hillsides. The length of stalk can reach
100 m and more, and the area occupied with plant may number up to several thousand
square meters.
Certainly, such heavy green mass of a plant can not exist only due to poor
mineral feed, founded by roots in cracks of rocks. Having grown up to certain
size, rock fig lowers long crampons from rocks downwards to ground. In the
beginning it is only one thin shy root reaching to the ground. And sometimes
it must grow down to some tens meters. When it reaches ground, additional inflow
of mineral feed stimulates the further growth of plant, and some roots begin
growing to ground. Being bound and branching, they grow into ground and grow
thick, becoming similar to tangle of giant snakes.
Rock fig forms on hillsides and stony sites original “oases”, involving numerous
birds and insects. Besides the plant strengthens slopes and interferes with
landslides. Certainly, sometimes especially strong landslides tear down thickets
of this plant, but all the same the fig rather quickly restores from survived
branches.
Leaves of rock fig are small, heart-shaped and skinny. They form continuous
cover, under which various insects and spiders find shelter, and also mosses
and lichens plentifully expand. The tip of each leaf is strongly extended to
original “dropper”, characteristic for tropical plants of damp climate. In
damp hot weather it seems, that under thickets of rock fig there is a constant
rain – drops continuously fall from leaves of plant, and flow down in streams
from stones on which it grows.
Inflorescences of this species are ordinary-looking, as at all figs. These
are small green “berries” - siconiums on stalk and large branches. Rock fig
is pollinated by microscopic wasps living only in its inflorescences. Larvae
of wasps destroy a part of seeds, but staying ones will suffice for reproduction
of this plant. Ripen fruits of this species are small, red - brown, very sweet
and soft. Birds are the basic carriers of seeds of this plant. They swallow
fruits entirely, disseminating seeds of rock fig both with dung.
Mushroom-leaved begonia (Begonia fungifolia)
Order: Begoniales (Begoniales)
Family: Begoniaceae (Begoniaceae)
Habitat: Southeast Asia, rocks; epilithic plant.
Mountain slopes in Southeast Asia represent a favorable place for growth of
grassy plants had got used to be content with a small amount of ground and
moisture. Rains plentifully spill on rocks, but water quickly flows down on
them, and plants had developed the adaptations, allowing to keep and to accumulate
moisture.
Among thickets forming on rocks a species of plants, which may be easily confused
with fruit bodies of any mushrooms, frequently grows. It seems, as if among
moss the set of uniform mushrooms with green leathery pileus and the thicken
stem covered with hairs has grown. But among thickets of these “mushrooms”
thin flower-bearing stems bearing clusters of tiny pink flowers rise. These
flowers are very characteristic – thy have four petals and three-edged “winged”
ovary. They indicate to the belonging of their owner to extensive Begoniaceae
family. In Neocene some representatives of this family turned to trees, but
the majority of them remained grassy plants. The present species of begonias
is named mushroom-leaved begonia.
It is semi-succulent plant, at which aquferous tissue is advanced. Begonias
of tropical forest have large and thin asymmetric leaves. At mushroom-leaved
begonia leaf, on the contrary, is small, rounded and leathery. Its diameter
does not exceed 8 – 9 cm. Leaf of this plant has almost regular rounded outlines,
gristle-like consistence and smooth top side with slightly pressed nerves.
As opposed to the majority of begonias with bright leaves, leaves of mushroom-leaved
begonia are dim: they have dark green color with lighter border, and along
large nerves from the centre of leaf the stripes of bright green color stretch,
forming star-like figure.
The leafstalk is attached in the centre of buckler-like leaf and is the main
water-reserving organ of this plant. Leafstalks at this begonia are inflated
and similar to stalk of mushroom. They are bright green and semi-translucent;
the most part of their volume is made of water-reserving parenchyma. The surface
of leafstalk is covered with hairs that reduce evaporation.
The root system of this begonia is fibrous and consists of set of thin roots.
On stalk of plant numerous additional roots develop. During a rain this plant
soaks up water, which is quickly reserved in leafstalks. The stalk of this
plant represents thick creeping rhizome; leaves are kept only at the top part
of sprouts. Leafstalks are slacky attached to rhizome, and break off at small
effort. The broken off leaf keeps viability due to a stock of moisture in leafstalk
for a long time. At the place of fracture additional buds develop, from which
young sprouts grow. While the root system is poorly advanced, they grow due
to stocks of the moisture kept in a leafstalk.
Mushroom-leaved begonia blossoms almost the year round, except for several
weeks, when the amount of rains is minimal. It is pollinated by insects of
various species – mainly by bees and butterflies. Tiny seeds of this plant
simply spill out from the opening pods, and they are carried with wind.
Silver brachybegonia (Brachybegonia argentimaculata)
Order: Begoniales (Begoniales)
Family: Begonias (Begoniaceae)
Habitat: Southeast Asia, rainforest canopy.
Begonia family including numerous species in human epoch showed relative poverty
at the genus level – at that time there were only four genera of which three
ones included only one species, and only Begonia genus was very variative and
widely settled in tropics. In Neocene among begonias original species of begonian
trees (Begoniodendron) had evolved, representing an original attempt of begonias
to enter large size class and to form new vital forms. At the level of grassy
plants evolution of begonias had also proceeded, and in Neocene at the territory
of Southeast Asia the genus of epiphytic plants brachybegonia (Brachybegonia)
related to begonias had evolved. These plants had developed life in forest
canopy and represent exclusively epiphytic grassy forms. They keep the characteristic
asymmetrical leaves growing in two rows. The stalk of brachybegonias is very
short, and leaves lay on each other. Leafstalks are also so short, that leaves
seem growing directly from the stalk, forming the symmetric crown. Roots of
these plants are very thin and tender; root system is diffuse. Roots of brachybegonias
branch plentifully and penetrate into cracks on bark of tree, strongly fixing
even large bushes on its branches.
The bottom leaves in the crown, the oldest ones, differ in shape and color
from top ones. They are more rounded, shorter and wider. Colouring of such
leaves is dim. When the plant develops completely, these leaves appear in shadow.
But they do not die off, and serve as original traps for organic substances.
In them dust, rests of bark and dung of birds and insects is gathered. From
stalk, and even from leaves growing higher the additional roots sprout here,
and the plant receives an additional feeding. A similar role is played by sterile
fronds of “staghorn fern” (Platycerium) which also is kept in tropical forests
of Neocene. Thus, brachybegonias and “staghorn fern” represent an example of
convergence.
The top leaves of brachybegonias differ strikingly from bottom ones. They start
to develop when the plant will grow stronger enough. These leaves are larger;
they have long tip “dropper”, and leaf surface may have a pattern of strips
and spots, similar to pattern of begonia leaves.
Brachybegonias had inherited from begonias ease of vegetative breeding. Formation
in leaf axils of long sprouts on which tips crowns of leaves develop is typical
for them. Such “offshoots” easily come off sprouts and take roots in suitable
places. On leaves of some species the sprouting buds develop, which give rise
to young plants.
Breeding by means of seeds plays an important role in settling of plants to
longer distances. Adult plants grow from top of the main sprout a flower-bearing
stem on which the inflorescence develops – a cluster of several tens, and at
large species even of hundreds small flowers. Fruit of brachybegonia is a capsule
filled with dry smallest seeds. They are easily carried by wind to several
kilometers away from parental plant. After flowering the stalk dies off, but
in its basis from buds one or several lateral shoots develop, replacing parental
plant. One shoot lives up to four years at large species, and no more than
two years at small ones.
Various species of brachybegonias differ from each other in shape and colouring
of leaves, in size of adult plant and amount of flowers. Silver brachybegonia
lives in the top level of tropical forest. Its crowns reach 50 – 60 cm in width
and total up to 10 – 15 leaves. The top side of leaves is almost entirely covered
with small hairs, giving silvery shine to the leaf surface. Only in the middle
of leaf closer to the place of leafstalk attachment hairs are absent and leaf
has typical green color and smooth surface; the bottom part of leaf is always
covered with white downiness which in due course thins. Young leaves look almost
white because of rich downiness. Also flower-bearing stems of plant are hairy.
The inflorescence of this species is corymb-like. Flowers have a colouring
sharply contrasting with colouring of leaves – they are dark red. They are
pollinated mainly by solitary bees and flies of various species. This species
lives in tropical gallery forests of Indochina.
Live-bearing brachybegonia (Brachybegonia vivipara) differs from other species
of this genus in higher ability to vegetative breeding. It has large leaves
with serrated edge, dark green with reddish middle, covered with rough rigid
hairs along the edge. On denses along the edge of leaf adventive buds are formed,
and from them small daughter plants develop. The inflorescence represents sprawling
cluster of several tens bright pink flowers. This species is common in humid
mountain forests of Jakarta Coast.
Climbing brachybegonia (Brachybegonia scandens) had choosed another tactics
of vegetative breeding. This species forms rather short runners having ability
to branch and take roots in nodes. From the crown of parental plant in various
sides runners grow. The part from them takes roots in bark and in thickets
of moss, and continues growth upwards or downwards. The shoots having no abilities
to take roots stop growth and die off soon. Successfully growing runner branches
and continues growth in length. Having reached up to the place where humus
accumulates, such runner takes roots and forms a young plant. Leaves of this
species form the crown of about 50 cm wide of rounded leaves of bright green
color with small denticles along the edge. The surface of leaf is covered with
short thin hair of red color. This species blossoms, forming high peaked cluster
of several tens small white flowers. Climbing brachybegonia grows in warm rainforests
of lowland areas of Jakarta Coast and nearby islands, preferring the top part
of forest canopy.
Giant brachybegonia (Brachybegonia gigantea) from tropical forests of Hindustan
and Southeast Asia represents the largest species of the genus. It forms huge
leaves – up to one meter long at width of about 70 cm. Leaves of this species
are entire-kind and have denticles along the edge. Their colouring is bright
green; along the edge narrow pinkish-red border is stretched. On smooth surface
of leaf separate spots of white color are scattered, located more often in
the place of attachments of leafstalk to leaf plate. The flower-bearing stem
reaches the length of two meters and bears up to one thousand tiny red flowers.
This species produces a small number of runners, but they easily take roots
in nodes and stretch along tree trunk and branches to the distance up to five
meters.
Graceful brachybegonia (Brachybegonia elegans) is the smallest species of the
genus – the width of its crown does not exceed 10 – 12 cm. Its rounded leaves
have bright red color with green irregular-shaped border varying from separate
“leakages” and “splashes” up to wide border reaching half of leaf radius. It
grows quickly and gives plentiful offsprings. The adult plant starts to form
lateral shoots even before flowering. The flower-bearing stem of this species
is very long in comparison with the size of the plant – it reaches almost half-meter
length. The inflorescence of this species is a cluster of large pink flowers
hanging on long pedicels. This species is widespread in humid forests from
eastern part of Hindustan up to the south of Jakarta Coast.
Dissected brachybegonia (Brachybegonia dissecta) has leaves of unusual shape.
First leaves of this plant are ordinarly looking rounded ones. But in due course
of growth and maturity they change the shape – between nerves cuts appear,
and the largest leaves have cuts as deep as one third of general width of leaf.
This adaptation helps to avoid the shading of bottom leaves by top ones. Leaf
surface is smooth, dark green with separate dark red spots. Width of crowns
of this plant is about 40 cm. This species inhabits the bottom part of forest
canopy where an effect of lack of sunlight takes place. Dissected leaves represent
the adaptation permitting to compensate it. This species is widespread in lowland
forests of Jakarta Coast and Indochina.
Tree-climbing tectobegonia (Tectobegonia repens)
Order: Begoniales (Begoniales)
Family: Begonias (Begoniaceae)
Habitat: Southeast Asia, tropical rainforest.
In human epoch begonias were exclusively grassy plants or suffrutices. In epoch
of ecological crisis caused by human activity many species of these plants
became rare, and even had died out. But after disappearance of mankind and
restoration of tropical forests begonias could develop other vital forms. Among
them begonian trees (Begoniodendron) and epiphytic brachybegonias
(Brachybegonia) had evolved. And on trunks of trees of tropical rainforest of Southeast Asia
their relatives climb upwards. It is easy to find out begonia descendants in
them: leaves of these plants have characteristic unequal-sided shape. But appearance
of this plant is atypical for begonias – leaves grow on stalk in two lines,
overlapping each other, as if a tile. Hence the name of this plant – tectobegonia
(from Latun “tectus” – covered).
The stalk of tectobegonia is very long – at old specimens it reaches the length
of 6 – 8 meters. The stalk nestles densely against the bark of trees, and the
plant creeps on the surface of tree trunk. It is root-climbing liana similar
ti ivy (Hedera) - in stalk nodes a plenty of white strong roots develops, penetrating
into the bark and moss pillows. Stalk is flattened and ribbon-like. It is fragile
and easily breaks off; besides the plant branches and total length of stalks
of one individual can reach 20 – 30 meters. Usually this plant begins its life,
taking roots in vegetative dust. The first weeks of life sprout grows vertically,
but then lays and begins creeping. Gradually shoots get the shape typical for
the present species: they begin creeping on a substratum, stalks turn ribbon-like,
internodes turn shorter, and then leaves grow, overlapping each other like
tile. Tectobegonia grows in rainforests and such phyllotaxy helps to deflect
streams of rain water. The plant grows quickly and its shoots creep to light
forest canopy soon. At old specimens initial roots and the bases of stalks
die off, but the top part of stalk does not perish. It continues growth as
an epiphyte, nourishing due to humus which accumulates in hollows and among
branches. This plant is well adapted to vegetative breeding: the broken leaf
easily takes roots, and forms a normal plant shortly.
Leaves of tree-climbing tectobegonia are dark green and rounded, with the lengthened
tip extended as a “dropper”. Leaf length does not exceed 5 cm; width is up
to 3 cm. In shadow leaves look ordinary, but at the sunny place, when the plant
appears in the top part of tree crone, silvery spots appear on leaves.
During the flowering tectobegonia forms in leaf axils short flower-bearing
stems on which rich spherical clusters of tiny white flowers grow. On young
leaves in the basis of flower-bearing stem large silvery spots appear, involving
insects from apart by shine. Usually tectobegonias are pollinated by butterflies
and flies, less often by solitary bees.
Fruit of tectobegonia is a winged capsule. Tiny seeds of these plants (weighting
some milligrams only) are carried by wind.
Sour-leaved
begonian tree (Begoniodendron oxyphyllus)
Order: Begoniales
Family: Begoniaceae
Habitat: tropical forests of Southern Asia. Close species are expanded from
Zinj Land up to South-Eastern Asia.
In Holocene epoch exclusively perennial grassy plants had belonged to begonian
family; at some species annual sprouts died off, but large perennial tuber
was kept. Among these plants there were ground grasses (frequently they were
too sizable), grassy lianas and epiphytes. But in Neocene epoch, when the structure
of wood vegetation in tropics had essentially changed, among begonias treelike
forms, representatives of begonian trees (Begoniodendron) genus, had appeared.
Plants belonging to this genus differ in attributes characteristic earlier
for grassy begonias. They grow very quickly - the annual height increase reaches
five meters and even more. Wood of begonian trees is friable, very sappy and
fragile. Sometimes during strong winds the tree trunk breaks, but this event
does not bring a big harm to tree: one more feature inherited from grassy begonias,
ability to vegetative multiplying, has an effect. The tumbled down tree laying
on the ground easily takes roots. From trunk vertical sprouts start to grow,
and branches bend upwards. This way one tree turns to rich thicket. Through
any time the part of trunks will be choked and will be lost, but some trunks
will develop normally and gradually thickening will grow up to height 25 -
30 meters - usual height of begonian trees.
Except for it begonian trees have kept ability to multiply by rooting of branches
and even separate leaves.
Due to growth rate begonian trees are pioneer vegetation of tropical woods.
They are among first plants settling on free ground where the old tree has
fallen, or on river deposits (begonian trees are too water-resistant plants).
They grow quickly, but live not for long time: no more than 50 - 70 years if
they will not be fallen down with any storm.
Wood of begonian trees, rather soft and sappy, would be fine food for every
possible larvae of insects. But trees of Begoniodendron genus have developed
the special strategy of protection against them, having developed one of features
of their ancestors. Sap of begonian trees is true chemical “cocktail” of every
possible organic acids – malic, citric, and even formic and acetic ones. At
various species the structure of sap varies, but the principle of its action
stays constant: larvae of insects practically do not live in very sour environment,
and development of mushrooms and bacteria is essentially suppressed.
As well as grassy begonias, begonian trees differ by original oblique leaves.
They are colored not as brightly as at small grassy begonias, but at some species
on leaves there are silvery spots and downiness, and some species have graceful
dissected leaves. These are evergreen trees.
Having reached about ten years' age, begonian trees start to blossom plentifully.
Small pink four-petal flowers of these trees are gathered to large corymb-like
inflorescences. They are surrounded with two brightly colored bract leaves/
Pistillate flowers grow on ends of branches in medium and bottom part of crone,
and staminate ones blossom out basically at top of tree. If insects visit flowers
not so actively, that is chance of pollination by means of wind. Begonian trees
bloom practically all year round. Only in places with seasonal droughts flowering
interrupts at two - three months. Seeds of begonian trees are very tiny. They
are carried by wind to far distance; therefore begonian trees frequently grow
on islands of Indian Ocean.
Sour-leaved begonian tree is the typical representative of genus. Its height
is up to 20 meters. This species meets in river valleys, sometimes forming
there dense thickets. This plant endures flooding of roots during high waters,
keeping stability with the help of well advanced plank-buttress roots. The
trunk of this tree receives additional durability because it is not cylindrical,
but ridge-shaped - as five- or six-sided prism. It is connected by that this
species of trees grows in places where strong winds frequently blow. The bark
is colored grey: it protects tree trunk from overheat.
Leaves of this tree are dark green with numerous silvery spots; edges are denticulate.
Length of leaves is about 25 cm, width is up to 7 – 10 cm. Bracts are very
large (length is up to 30 cm), rounded, dark red with scarlet spots. The surface
of bracts and leaves differently reflects ultra-violet beams, therefore insects
easily notice flowers of plant. Flowers are small, light pink, gathered in
clusters of 30 – 40 ones. Fruit is three-edged pod.
Other species also belong to begonian trees genus:
“Silver
hand” begonian tree (Begoniodendron chira-argentea) is settled on
islands of Indian Ocean. Its height is up to 15 - 20 meters. Trunk is thick,
“bottle-like”, adapted to storage of stock of fresh water. The rootage is not
deep (tree avoids salted subsoil waters of islands), but is submitted by powerful
plank-buttress roots growing far in sides – they keep tree from falling during
gales sweeping over islands. Leaves are about 20 cm long, rounded, dissected
to five – six lobes, and plentifully covered with silvery hairs. Hairs serve
for protection against superfluous sunlight, and absorb water during rains.
Bark is dark grey and thick.
Flowers are bright red; bracts are pink. It flowers in season of monsoons,
at lack of moisture casts a part of foliage.
Sprouting
begonian tree (Begoniodendron proliferus) lives at the
East-African subcontinent, and inhabits tropical woods. Its height
is about 25
meters. It grows in forest wood, avoiding swampy soil. Leaves are
oblique-heart-shaped, with large jags on edge, light green with red
spots between main nerves. In place of attachment of petiole on blade
the additional bud develops. On branches especially strongly lighted
by the sun, it starts to develop in young plant. This time 1the attachment
of petiole to branch weakens, and during a storm such leaves with
young plants take off from parent tree and are carried away by wind
to many tens meters aside. Leaves of parent plant serve as sails,
increasing range of flight of young plants. Having fallen on the
ground, such offspring quickly takes roots and starts active growth.
Pink flowers are small. Bracts are very large and red-colored.
Creeping
begonian tree (Begoniodendron repens) grows in rainforests of South-Eastern
Asia, most frequently meeting in mountain woods and on steep riverbanks. At
this
tree it is impossible to tell unequivocally, what is possible to think “height”
– the main trunk of this tree in the beginning grows upwards, then lies down
and continues growth in horizontal direction. From it numerous thick vertical
trunks branch off. Such kind of growth permits a plant “to wedge” between large
trees, considerably outstripping in growth sprouts of competitor trees. To
“fix” achievements, the horizontal trunk actively grows thick. The length of
basic trunk may reach 50 meters and more, vertical trunks of 2-nd order reach
15 - 20-meter height. On river breakages the basic trunk can overhang downwards,
growing downhill up to the water, forming sometimes original natural “bridges”
across small rivers.
Leaves are large (length is up to 40 cm, width is about 30 cm), oblique-heart-shaped,
edge of leaves is wavy. The tip of leaf is strongly extended to “dropper”:
it is a common feature of plants of damp rainforest. Leaves are colored dark
green with faltering silvery strips along large nerves.
Bracts are silvery with several green spots; flowers are large and pink.
Desert cyclamen (Aridocyclamen macrotuber)
Order: Primulales (Primulales)
Family: Primroses (Primulaceae)
Habitat: deserts of Mediterranean lowland.
During millions years Africa moved to the north, aside Europe. And millions
years after human disappearance it has resulted in significant changes of environment
in Mediterranean area. When strait of Gibraltar was closed, Mediterranean Sea
had dried up very quickly, and instead of it the severe salt desert was formed,
edged by swamps with lifeless supersalt water. Drying of the Mediterranean
had considerably changed a climate of Southern Europe – it became dry and severe,
with significant seasonal and daily differences of temperatures. The desert
is crossed with some small rivers. Some of them run into swamps, and others
dry in sand. All of them have head waters in Alpine glaciers and give life
to not numerous inhabitants of desert.
Dry deserts stretch as a ribbon along edges of areas of former sea-bottom covered
with a crust of salt. These areas differ in severe climate, but nevertheless
they are populated with plants capable to reserve or to keep moisture. In desert
there is much more plants, than it seems at first sight. For desert the perennial
plants are very characteristic – these ones are ephemers, blossoming in spring,
when the ground is humidified enough with streams of water flowing from mountains.
One of the most beautiful plants of the Mediterranean desert is desert cyclamen,
the representative of ephemeral vegetation. This species is the native from
the Alpes, the descendant of European cyclamen (Cyclamen europaea). It lives
at the coast of temporary reservoirs, where the moisture in ground is kept
longer.
This plant develops within two months in spring, while ground is moistured
enough. The desert cyclamen survives in summer, autumn and winter in condition
of big spindle-like tuber. At the depth of about 40 cm, where there is a tuber,
the ground softens differences of temperatures and protects it from the majority
of herbivores. But during the vegetation the tuber literally creeps out upward,
closer to the sun. This property of a plant is connected to features of its
structure. Roots of desert cyclamen are very long. They reliably keep this
plant in ground, and have ability to twist at drying. In spring roots impregnate
moisture and push a tuber closer to ground surface, and in the beginning of
drought they contract and involve tuber deeper under ground. In foothills of
the Alpes there are populations of this species able to vegetate much longer,
rather than in desert. But this species differs in lowered competitiveness
- desert cyclamen is very photophilous plant and does not suffer shadowing.
In spring, when the ground becomes enough humid, the plant starts to grow.
Within several days it promptly develops 4 – 5 large rounded leaves (their
diameter is up to 15 cm) on long leafstalks. Leaves have a grayish shade because
of layer of wax developing on them. After that desert cyclamen begins to blossom.
Its flower-bearing stem quickly expands and reaches the height of 70 cm. No
more than 2 – 3 large flowers with petals curved back by characteristic way
keep on plant simultaneously. Flowers emit intense aroma, which becomes especially
intensive to the evening. This plant is pollinated by butterflies and solitary
wasps. Wasps especially frequently visit flowers of desert cyclamen. These
insects search on the plant not only nectar, but also gather from leaves wax
necessary for construction of nests. Flowers of desert cyclamen are pink with
silvery edges of petals reflecting ultra-violet light that helps to involve
insects.
To the beginning of summer at desert cyclamen seeds ripen. As against an ancestral
species, which seeds very quickly lost germinating capacity, seeds of this
species can keep germinating capacity till 3 years – in the changeable world
of desert conditions are not always favorable for sprouting, and the part of
shoots simply can be lost. But in ground always there is a reserve of seeds
of desert cyclamen. Seeds have strong environment on which the edible oily
appendage grows. Due to it they are pecked out willingly by birds, with which
help this species is settled. Desert cyclamen frequently grows in “oases” of
Mediterranean swamps.
Digitate
wing-seeded maple (Pteracer palmatus)
Order: Sapindales (Sapindales)
Family: Maples (Aceraceae)
Habitat: warm and humid forests of Southern Europe (Balkan), Caucasus and Asia
Minor (at the southern coast of Fourseas).
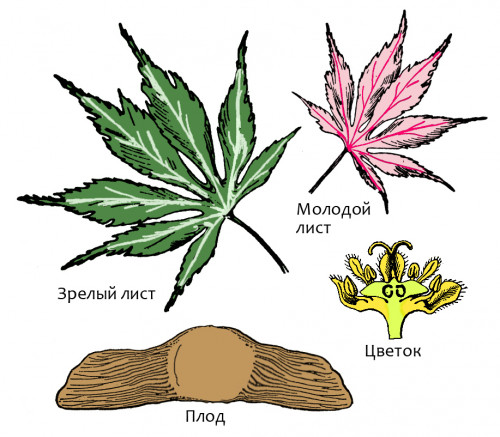
Picture by Biolog
In Holocene the wood flora of Europe had benn tested for survival
rate. People had cut down earlier almost continuous and impassable woods and
after that test the ice age had came. When Mediterranean Sea had dried up, and
instead of it the hot desert had formed, woods of Europe had kept only at the
narrow strip between northern tundra and rigorous salt Mediterranean desert.
Woods had remained at the south of Western Europe irrigated by clouds from Atlantic,
and also along mountain ridges of Alpes and Carpathian Mountains. Rather large
forest areas had formed in the Balkan.
When the warming began in Europe, woods had started to settle actively farther
to the north. At that time people had already disappeared and the wildlife evolved
according its own laws. In Neocene the edge of warm climate area had moved closer
to poles, but it occured not in all parts of Eurasia. Eurasia is a huge land
and the area of sharply expressed continental climate. Significant territories
of this continent are removed far from large reservoirs making climate steady
and humid, therefore at the northeast of Eurasia winter is still cold and snowy.
At the south and east of Europe the influence of Fourseas is felt, and attributes
characteristic for tropical plants have appeared at descendants of species of
temperate flora. These features are wide and often compound or dissected leaves
with “droppers” – pointed tips. Nut-tree, chestnut, maples and sycamores are
the typical species making the European subtropical woods of Neocene epoch.
The strict competition has caused occurrence of effective adaptations for breeding
and settling.
Wind carries above crones large flat seed having two wide transparent wings.
From each capful it flies up higher ana higher, and flies to long distance.
It had fallen down from the high tree reaching 30 – 35 meters height. This is
tree of average size in European subtropical forests – it belongs to species
makinf forest canopy and does not “stick up” above it. At this tree there is
umbrella-like crone, and on flexible young branches clusters of the same winged
seeds swing.
The seed of this tree with two big wings indicates the origin of this plant:
this tree is the descendant of maple (Acer) which various species were characteristic
for Northern Hemisphere. For the characteristic form of seeds this plant is
named wing-seeded maple. At the maple the fruit consists of two seeds having
one wing is characteristic. At ripening they break up and keep in air due to
rotation of falling seed around the axis. At at fruits of wing-seeded maple
one fruit of the pair is reduced and from it only wing remained. The rest of
this seed is small scar on one side of flat disc-shaped seed having two wide
wings. Blasts carry such seeds far from parental tree. The seed is easy also
and can stay in air for a long time, reacting even slight wind whiffing.
If it will have luck to appear in place favorable for life and to sprout, eighty
– hundred years a huge tree of wing-seeded maple will develop later from it.
Trunk of adult tree of this species is straight; till the growth its lower branches
fall down. At adult tree first large branches remain at height of 20 meters
from the ground. The trunk of wing-seeded maple is covered with smooth grey
bark covered whith constantly shelling small scales – such feature does not
permit this tree to overgrow by epiphytes forming in favorable conditions “hanging
gardens” weighting up to tens kilograms. Some thick plank-buttress roots growing
in the bottom part of trunk provide to it additional stability. Such roots start
to develop since the tree reaches approximately half of normal height.
Leaves of digitate wing-seeded maple are typical for plants of humid wood: they
are dissected to 7 – 9 pointed lobes separated to 9/10 of their length and arranged
on petiole like fan. Tissues of leaves are rather soft. Leaves grow on young
sprouts in pairs, and reach diameter of about 50 – 60 cm. Young leaves of this
tree are pink with bright crimson strips along main nerves: it is so-called
anthocyan colouring protecting against sunlight. The mature leaves of digitate
wing-seeded maple are light green, but along nerves darker zone stretches. Nerves
of mature leaves are white.
This plant is semi-deciduous – in the autumn, at first cold snaps wing-seeded
maple casts approximately one third of old leaves and falls the relative rest
till 2 months. On branches only the youngest foliage remains. At the north of
area this tree can lose almost all leaves. The rest period is very important
feature of wing-seeded maple: at this time at tree flower buds are formed.
The foliage prevents pollinator insects to find flowers, therefore tropical
trees produce flowers more appreciable with the help of strong smell or bright
colouring of corone. At wing-seeded maple the inflorescence is long and carries
set of small five-petaled flowers. This tree is pollinated by bees and wasps.
Flowers are located by clusters on long peduncle hanging from branch down. Flower
of this tree is about 2 cm in diameter. It is yellow, fragrant, with short petals
and wide receptacle in which middle two carpellas grow forming the structure
similar to head of ancient key. After pollination one ovary is reduced, and
the second one develops. The wing of reduced ovary reaches the same size as
at normal ovary, and as a result one large seed with two wide wings keeping
in air for a long time is formed. It sprouts in spring and starts to grow quickly.
Usually in underbrush many sprouts of this plant grow, but the most part of
them perishes not having found favorable conditions for life. Till the first
year of life the young plant reaches height of about two meters. First years
of life wing-seeded maple can grow in shadow of underbrush competing with other
plants, but do not receiving the necassary amount of light it starts to wither.
If the old tree closing access to light to underbrush falls, young wing-seeded
maple starts to grow quickly, aspiring to pass competitors. This tree begins
to blossom first time at the age of about forty years, having reached height
of approximately 20 meters. The maximal life expectancy of this plant reaches
400 years.
Umbrella-leaf
horse chestnut (Aesculus umbelliferus)
Order: Sapindales (Sapindales)
Family: Horse chestnuts (Hippocastanaceae)
Habitat: the Balkan, Carpathian mountains, Caucasian peninsula, southern and
western coast of Fourseas.

Picture by Biolog
The forming of Fourseas rendered significant influence to
the climate of surrounding territories. At the Balkan the narrow strip of land
between mountains and coast of Fourseas has turned to area of widespreading
of original kind of subtropical forests formed by descendants of northern species
of trees. Tree species growing here are remarkable with their broad leaves on
which tips characteristic “droppers” develop for removal of water surplus. In
winter trees cast a part of foliage and stop growth, but this period is rather
short, and annual growth rings in their wood are expressed poorly.
One of characteristic inhabitants of subtropical forests at the coast of Fourseas
is umbrella-leaf horse chestnut. This tree is easily distinguished from apart
because of its original leaves which differ in great degree from leaves of other
trees of these forests. At umbrella-leaved horse chestnut leaves resemble palmately
compound leaf of its ancestor, horse chestnut (Aesculus hippocastanum), but
differ from it in several features: size and number of leaflets. Umbrella-leaf
horse chestnut has giant leaves similar simultaneously both to umbrellas and
to fan-like leaves of palm trees. The leafstalk of each such leaf is about one
meter long and up to four centimeters thick; it carries leaf plate of about
one and a half meters in diameter, which is made of 15 – 19 approximately equal
leaflets expanded and rounded on the terminal part. The midrib in each leaflet
forms thin extended dropper.
Having huge leaves, umbrella-leaf horse chestnut is very exacting to presence
of water. This tree grows along riverbanks and occasionally develops even at
marshes where grows on drier areas. Most frequently this species of trees grows
in forests along the coast of Fourseas.
Bark of this tree smooth and dark grey. Umbrella-leaf horse chestnut can reach
height of 25 – 30 meters. Usually it is one-trinked tree, but frequently it
produces radical offsprings and grows several trunks, forming dense crone. However,
more often such trees break on separate trunks and fall, because each trunk
forms asymmetrical one-sided crone, and the trunk can not sustain weight of
foliage and branches. Trees with normally advanced crone can grow till 300 years
and more. As they grow, tree trunk becomes ridged – a kind of original “edges
of rigidity” appear on it, like the resemblance of plank-buttress roots of tropical
trees. Due to a friable bark respiration of roots is provided and this plant
successfully grows on overmoistened ground.
In winter umbrella-leaved horse chestnut loses a significant part of foliage.
On branches only youngest leaves remain, which do not reach the complete development
and grow only up to 30 – 40 cm in diameter. And the strong frost can kill them
at all and then the huge tree stands leafless. But in spring, when warmth returns,
the tree quickly puts on new foliage. The leaf of this plant develops very quickly
– for one week it reaches the usual size. At the same time tree begins blossoming,
becoming a true ornament of subtropical forests.
Inflorescence of umbrella-leaf horse chestnut umbellate is thyrsoid one up to
one meter high. Flower-bearing stems rise vertically, as at its ancestor. Flowering
begins from lower part of inflorescences. Flowers of this tree are rather large
– their diameter reaches 4 – 5 cm. They are zygomorphous and have white color
with yellow middle; in fauces of flower black specks are scattered. This tree
is not specialized to the certain pollinators. In its flowers butterflies, beetles
and solitary bees search for food. Very frequently birds visit flowers of this
tree, playing an important role in pollination of this plant. In due course
of deflowering each flower of this species changes colouring from white to yellow
and pink. Right before falling off petals get crimson-red color for one or two
days.
Fruits of umbrella-leaf horse chestnut are similar to fruit of its ancestor
– they are large and leathery; the environment is covered with short spikes.
However, in each fruit not one, but two seeds ripen, as it was, obviously, at
ancient species of this genus. However, the second ovule has secondarily appeared
at this species. In due course of evolution the umbrella-leaf horse chestnut
had adapted to distribution of its seeds by small animals, and its seeds are
rather small – their diameter is only about 2 cm. They are distributed by forest
rodents and birds. In autumn woodpeckers eat seeds of this plant.
Many seeds of this tree appear eaten in autumn and in winter, but some ones
succeed to escape, and they sprout. This tree begins to blossom first time at
the age of about 30 years, at height of about 10 – 15 meters.
Crimson-leaved balsam (Impatiens cinnabarifolia)
Order: Ericales (Ericales)
Family: Touch-me-nots (Balsaminaceae)
Habitat: rainforests at the north of Meganesia, sites of broken forest stand
and riverbanks.
Symbiosis between plants and animals is more often forming relatively to pollination
of plants to which the certain species of pollinators adapts strictly, and
its presence guarantees reproduction of plant. Less often the plant forms the
shelter to insects like ants, which protect it from pest insects.
One species of grassy plants, crimson-leaved balsam, has established symbiotic
relations with the representative of vertebrate animals that is a rarity in
flora in itself. This plant entered symbiosis with one local bird, the gardener
bowerbird (Hortulornis plantarum-cultor). Two circumstances have allowed to
symbiotic relations to be established: requirement of bird for bright objects
on the one hand, and red anthocyan colouring of young leaves of plant, on the
other hand. Probably, ancestors of the present species of bowerbird originally
simply broke off such bright tops of plants and dropped them around of their
bowers for attraction of females. Tops of balsam plant easily take roots, and
this plant in many cases started to grow near constructions of bowerbirds.
Gradually evolution of bird had gone in direction of maintenance of life of
such “ornamental plants”: birds broke off sprouts of other grasses in order
to make raising bush even more appreciable and by that reduced a competition
of balsam to other plants. Besides birds tried to choose for cultivation plants
with brightest leaves. Similar relations had formed characteristic shape of
this plant.
This species of plants expands well at enough of light – in sites of broken
forest stand, and also at riverbanks. Leaves of crimson-leaved balsam have
bright anthocyan colouring – they are green with numerous red spots, and the
youngest leaflets are completely ruby-red. This species differs in fast growth
and development: from germination of seed up to the flowering less than four
months pass. If the plant develops without care on the part of bird, for this
time it grows up to one meter height and branches a little, and its leaves
in the bottom part of stalk gradually fall down. But at care on the part of
gardener
bowerbird the shape of plant is completely other: bird pinches out by beak
top of plant, and it starts to branch plentifully, turning to spherical bush
about 40 – 50 cm high. Due to branching it has plenty of points of growth,
and bright red color of young leaves it well visible on the background of environmental
green.
But symbiosis with the bowerbird has the return side: balsams belong to number
of insect-pollinating plants and presence of symbiote bird frightens off pollinator
insects, or bird simply catches and eats them. Therefore flowering of crimson-leaved
balsam proceeds exclusively at night. In axils of top leaves flower buds are
forming. They develop for a long time before will reveal only for one night.
Their pedicle is turned to spiral, having wrapped up a bud by itself. At night
the pedicle quickly starts to untwist, and in one hour is straightened completely,
taking out a flower bud above leaves. The flower with long spur and dazzling-white
petals is opened, exhaling strong sweet aroma with light putrefactive shade,
involving moths. The plant secrets a plenty of liquid nectar, and moths willingly
visit its flowers. If the flower is not pollinated, within day it literally
turns to slime and the pedicle falls off. Frequently the bird, cleaning dust,
pulls and throws out such pedicles. If pollination has taken place, the pedicle
turns to spiral again, and under covering of leaves a fruit starts to grow
ripe – dehiscent pod. To the moment of seeds ripening the pedicle is straightened
and extended horizontally. At casual touch of bird it bursts with crashing
sound and scatters seeds to the distance of several meters.
Leafless pounchstem (Gastrocaudex aphyllus)
Order: Apiales (Apiales)
Family: Carrots (Apiaceae)
Habitat: South-Eastern Europe, Balkan.
In Neocene Africa had closely come nearer to Europe. It had caused a lot of
changes in nature of Mediterranean. First of all, the result of this slow,
but steady movement was disappearance of Mediterranean Sea – strait of Gibraltar
was closed for ever, and till some thousand years the sea had completely dried
up. On the western part of saline lowland, once being a bottom of Mediterranean
Sea, severe deserts are stretched. East part of Mediterranean is the world
of saline swamps, where only creatures steady against salt can live. These
swamps are fed by streams flowing down in spring from the Alps. The second
consequence of movement of Africa to the north is a volcanic activity in Southern
Europe, and growth of Alps in height. They are still not as high, as Himalayas
are, but the first step is already made, and during the following millions
years Alps can become the highest mountains of the planet.
Narrow strip between southern slopes of Alps and Mediterranean swamps is the
area of droughty plain. Salt of the receding sea has remained in swamps, and
Alps block the way to damp northern winds. Water gets in these places in winter
as fogs, in spring as thawed snow from mountain tops. At this time for some
weeks the rich ephemeric vegetation, which spends the most part of year as
seeds, tubers and bulbs, develops here. But some plants are not afraid of cold
fogs and burning summer heat.
On clay ground the grotesque plant, partly similar to fossil plants of Paleozoic
and early Mesozoic, grows. It is not going to hide from sun and differs in
high specialization to extreme conditions of inhabiting. Thick branchless stalks
of this plant stick up from the rhizome hidden under ground and reach the height
of 3 – 4 m at thickness at the basis about 20 cm. Trunks are slightly bent
and completely lack of foliage, and internodes are inflated. This plant is
named leafless pounchstem. The plant is similar to Ottoa oenanthoides, the
plant dwelt in Andes (Venezuela) at height more of 4 kilometers above sea level,
and belongs to family common with it. Conditions of inhabiting of both species
are similar in one feature – they differ in dryness. In mountains dryness is
physiological – because of cold plants soak up water worse. And in desert it
is hot, and actual shortage of water is felt.
To save the precious moisture, leaves of pounchstem are reduced. They are present
by wide petioles without leaf plate, with advanced stipules, and exist rather
short time. Petioles and stipules are thin, turned to the kind of cap. They
serve for protection of apex, covering it from scorching beams of sun. In process
of stalk growth these caps quickly die off, but under them new ones are constantly
formed. Trunks of pounchstem have strong bark and are filled with friable water-bearing
tissue. In desert there are some species of herbivores, for example, whistlehorns.
But the plant is reliably protected from them by the weapon popular among Apiaceae
family – liquid accumulated in its stalk is poisonous and very bitter. Pounchstem
has not armed with spikes as cactuses or devil’s-milk plants (Euphorbia), but
chemical protection provides its survival.
Trunks of this plant grow from short turnip-like rhizome. As a signal for growth
ground humidifying serves. In spring from Alps streams flow down, and in desert
small temporary reservoirs are formed. The plant as if shakes from itself winter
catalepsy, and begins rough growth. Each stalk grows till some years, gradually
thickening. In nods stalk does not grow thick, therefore its trunks seem drawn
across. The young stalk has thin bark and friable tussue inside. Concentration
of poisonous substances in it is especially great: from bark of young stalk
the bitterish smell, warning herbivores that the plant is protected by poison,
is emitted. Active growth proceeds during three – four years of life of stalk.
When it grows ripe, bark hardens and turns drier, but cells of core are poured
by water and in them poisonous substances accumulate. Till first years of life
each stalk accumulates nutrients, and at the 6 – 8-th year of existence begins
to blossom.
The inflorescence of pounchstem is typically for its family – it is large (about
80 cm in diameter) umbel consisting of set of tiny white floscules. It rises
above stalks on low thick peduncle. Floscules are pollinated by insects. They
are not poisonous, and from them pleasant aroma is emitted. Seeds of pounchstem
are prickly; their environment is covered with numerous hook-like bristles.
They are carried by birds, clinging to their feathering. After ripening of
seeds trunk dries up and breaks off at the basis.
Leafless pounchstem lives up to 30 – 40 years, and starts to blossom at the
tenth year of life and blossoms annually up to the death.
Despite of poison, this plant gives food and home to some species of desert
insects. On young trunks of pounchstem water-bearing
plant louse parasitizes.
It is not afraid of poisons containing in plant, and can neutralize them. This
insect accumulates poison in body and plentifully secrets almost clean water.
This insect serves as though as natural “filter” of poison and the source of
water for some insects. Populations of this plant louse are “pasturing” by
shepherd beetle. It licks off water secreted by plant lice, and survives in
desert due to this. This beetle preserves plant lice against other insects,
feeds on inflorescences of pounchstem and pollinates its flowers.
Mediterranean
ant root (Myrmiradix mediterraneus)
Order: Apiales (Apiales)
Family: Carrots (Apiaceae)
Habitat: deserts of Mediterranean lowland, maquis of Southern and Western Europe.
In due course of evolution plants very frequently entered the union with insects.
The most widespread form of relations between plants and insects is the adaptation
of certain species of plants exclusively to one species of pollinator insects.
Phenomenon of myrmecophilia, the formation of symbiosis with ants, is less widespread.
In exchange for shelter and food, given to its six-legged lodgers, plants receive
reliable protection.
Frequently symbiosis of plants and ants arises in tropical forests where the
competition and danger on the part of herbivores are especially great. But occasionally
such unity arises in less favorable conditions. One case of such symbiosis had
arisen in arid areas of Mediterranean lowlands where herbivores are rare, but
damage of shoots in very adverse conditions of habitat can weaken the plant
considerably. This plant is Mediterranean ant root, the representative of umbellifers.
Appearance of this species of plants differs from typical shape of its relatives
– high piping stalks, alternate wide leaves and very characteristic inflorescences
– umbel of tiny modest flowers. Leaves of Mediterranean ant root have thick
midrib and leaf plate dissected into three rounded lobes with dulled tips. The
upper surface of this leaf is covered with thin layer of wax reflecting a part
of ultra-violet rays. The bottom side of leaf is richly covered with tiny white
hairs that create a favorable microclimate in thin layer of air adjoining the
surface of leaf. Stalks of this plant reach one meter height at thickness of
about 5 – 6 cm at the basis. At dying off of stalks their tissues quickly dry
up and are kept in vertical position almost all next year. For one year this
plant is capable to form about three fully developed blossoming stalks. The
inflorescence up to 20 cm in diameter is composed of 150 – 200 tiny flowers.
Flowering begins in early spring and proceeds during about one month. When seeds
ripen, pedicels of not pollinated flowers straighten and twist as prickles,
protecting seeds from an eating by birds. Seeds of Mediterranean ant root are
distributed by wind. For this purpose each seed has wide leathery sail surrounding
it along the edge.
The main feature of this plant is its thick underground rhizome. It reaches
length over 1 meter at diameter up to 7 – 9 cm, branches to some shoots and
lays in ground at the depth of up to half meter. It is hollow and separated
to chambers by internal cross partitions. The plant in itself is not poisonous,
but it has other protection against herbivorous mammals and insects: ants. Mediterranean
ant root gives a place for life to several species of ants – it is inexpedient
to become too strictly specialized in conditions where this plant lives. Actually,
its root and last year’s stalks represent a ready dwelling for ants. They only
bore entrances in partitions and surround a rhizome of this plant with tunnels,
improving air circulation. The fact that Mediterranean ant root is true myrmecophilous
plant, is proved to be true by one more feature – this plant constantly supplies
ants with some food. On internal walls of rhizome drops of liquid are secreted
from time to time, and ants drink it, and on edges of leaves amylaceous secretions
gathered by ants are forming. The plant grows successfully without ants, but
in this case its leaves are strongly damaged by insects.
The young plant begins to blossom for the first time only at the fifth year
of life. Life expectancy can reach 20 years and more.
Desert
carrot (Daucus aridophilus)
Order: Apiales (Apiales)
Family: Carrots (Apiaceae)
Habitat: deserts of the Near East.
As a rule, cultivated plants left without care from the side of people, appeared
inviable. Centuries-long selection has resulted in too heavy dependence of the
majority of species of cultivated plants on people, and after human extinction
only few of them were able to compete to wild species, which had been only undesirable
weeds in cultural plantings of human epoch. But on the other hand, due to the
human activity, cultivated plants were settled across the whole globe and could
receive advantages in a survival in some habitats. One of such species is carrot
(Daucus carota). One descendant of this species living in deserts at the edges
of Mediterranean lowland is desert carrot.
Life in harsh conditions of desert has considerably affected shape of plants
of this species. Desert carrot has large root tuber (about 80 cm long at thickness
of up to 10 cm), deeply penetrating into the ground. Due to long roots, plant
reaches a level of underground water, anchors firmly in the ground and gradually
buries itself in it each year until the time of flowering comes. In the beginning
of a dry season, roots dry up, curl up slightly and drag the plant deeper into
the ground. In first two years of life root is very thin, but to the moment
of flowering it grows very thick and accumulates lots of nutrients. Sugar gives
to pulp distinct sweetish taste. At this plant, pulp of root is paler, than
at cultivated ancestral species, and has pale yellow color. In a dry season
the bud is protected by a leaf having deeply reduced blade and expanded leafstalk.
Up to the moment of flowering the plant forms every year a rosette of deeply
divided leaves on rigid vertical leafstalks. In the year before flowering, the
size of leaves is maximal – up to one meter long on half-meter long leafstalk.
Desert carrot, like its ancestor, is a monocarpic plant and begins to blossom
at the age of 4-6 years. Flowerstalk at this plant reaches height of 3-4 meters,
an inflorescence from tiny yellow blossoms is an umbel about 60 cm in diameter.
Seeds of this species are covered with tiny hooks and prickles, being dispersed
by birds and bats having a rest on this plant. Seeds keep germinating capacity
for a long time and sprout in the spring during rains. At a young plant the
main root grows intensively; at this time its damage brings plant to death.
“Toad tree” (Bufodendron toxicus)
Order: Apiales (Apiales)
Family: Ginsengs (Araliaceae)
Habitat: Southern Europe, coast of former Mediterranean Sea.
In Neocene Europe and North Africa make a whole landmass – till the Cenozoic
Africa slowly drifted to the north, and had gradually joined Europe. Mediterranean
Sea had dried up till the previous glacial epoch, and its hollow occupied by
saline swamps, is gradually narrowed in Neocene. Millions years later it can
disappear completely, having turned to highland, but in Neocene this district
is rather extensive.
Above saline swamps the zone of dry stony semidesert with separate sites of
saline lands is stretched. The desert differs in hot and dry climate, but plants
had developed even such inhospitable district.
Here and there in dried up channels of streams flew down from Alps in spring,
the unusual plant grows. This is thick tree similar to representatives of genus
Adenium, lived in human epoch. The trunk of this plant is short – the plant
does not exceed two meters in height. But it is thick and stumpy, reaching
almost one meter thickness in the basis. In top part from trunk some the same
thick branches curved upwards and in sides grow. Roots of this plant are also
very thick, tuber-like inflated in basis. The bases of roots reaching about
one meter width, surround tree trunk; therefore it seems, as if the tree has
“thawn” from drying up heat on stony ground, as if any huge candle. This plant
is named “toad tree”.
Relations of this species with plants of Holocene epoch are unusual: “toad
tree” is the succulent descendant of typical European species – the ivy (Hedera
helix). So sharp transition of one vital form to another was shown by representatives
of genus Cissus of grape family. The majority of species of these plants is
submitted by lianas, but Cissus macropus is strongly expressed succulent plant
from South Africa with tuber-like stalk.
“Toad tree” accumulates a plenty of water in friable wood and the bases of
roots. But the tree is not intended to share it with anybody at all: sap of
“toad tree” is poisonous, and it protects this plant from an encroachment of
herbivores. Bark of this plant is very friable and soft, light grey with dark
knobs, outwardly resembling the toad skin. Such structure of bark is typical
for a lot of plants living in similar places – the bark badly transfers heat
and rescues the trunk from overheat, and also rescues the plant from rare light
frosts in winter. The plant forms sprouts of two types: long-term skeletal
branches and thin annual sprouts dying off and falling with approach of dry
season. Skeletal branches accrue slowly – one meter high plant exceeds the
age of 50 years, and two-meter giants have lived for one and a half centuries.
This tree is very durable – its age may reach 400 years and more.
The relationship of “toad tree” with the ivy is more obviously shown by leaves
and flowers. Leaves of this plant grow at the time of first spring rains and
keep on plant till only about three months a year. There are two types of leaves
of “toad tree”: they differ on peduncles and vegetative sprouts. On vegetative
sprouts leaves are palmately parted with thin lobes. The shape of leaves is
the parameter of development and state of health of plant. On healthy and strong
plant leaves have five and even seven lobes. And young or very old plant usually
forms three-lobed leaves.
At the age of about 20 years the plant begins to blossom the first time in
life. “Toad tree” blossoms in winter, before developing of leaves. At this
time from Alps cold air masses move, and in mornings on places of growth of
this plant a rich fog is rising. The cold period stimulates development of
flower buds, and fog serves as stimulus for flowering. The plant flowers and
fructifies early, because to dry season its fruits must have time to ripen
and sprouts could be developed enough go through a droughty season.
Bracteal leaves differ from usual ones: they are oval-shaped with dulled tip.
They richly grow on short floriferous sprouts, forming the cover for inflorescence,
similar to heads of asters. Mean-looking floscules with strong smell are gathered
in dense cyme-like inflorescence. Their spicy smell involves small beetles
keeping activity even in winter cool. At this time food sources are very rare,
therefore insects for a long time keep on inflorescences of “toad tree” and
spend night on them, having hided in depth of cluster. Fruits of this plant
are the round grey berries covered with wax. They ripen in spring, to the moment
of migration of birds from Africa. This plant is specialized to carrying of
seeds by birds: wax reflects ultra-violet light, and ripe fruits are well appreciable
for birds. Besides berries are poisonous for mammals occasionally coming to
places of inhabiting of this plant. Birds willingly eat berries and carry seeds
in vicinities. While in ground there is a lot of moisture, seeds sprout. The
young plant forms the crown of peaked leaves and grows a long rod root. It
grows in height very slowly: in first year the “toad tree” forms small turnip-like
tuber and the crown of 4 – 5 leaves. At the fifth year of life growth of stalk
in height begins.
Palmated aepyhedera (“over-ivy”) (Aepyhedera dissecta)
Order: Apiales (Apiales)
Family: Ginsengs (Araliaceae)
Habitat: Southeast Europe, Balkan.
Warm subtropical forests are obliged by their occurrence to influence of Fourseas.
Rains from this reservoir give enough moisture for growth of forests, and Fourseas
softens climate. Forests at the coast of this reservoir have some features
of humid tropical forest. They are made of deciduous flora, and in such forests
the abundance of lianas and epiphytes is observed. At the same time, these
forests are adapted to survival in conditions of expressed seasonal climate.
The huge mountain circuit complicates communication of subtropical forests
with tropical forests of Asia and Africa. Therefore in subtropics of Fourseas
at local plants features, characteristic for true subtropical plants of a southern
origin had evolved – large size, wide or dissected leaves, long leaf tips –
dripping tips, the adaptation for removal of surplus of water.
Among lianas of European forests descendants of an ivy of several species dominate.
Some of them had remained hardy, as their ancestor, evergreen lianas with rigid
leathery leaves. They had settled far to the north and twine round tree trunks
practically in whole Europe, except for Scandinavia and northeast. Northern
species are perennial small-leaved lianas with thin lignificated stalk.
The huge descendant of an ivy (Hedera helix), small liana of historical time,
is one of dominant lianas in southern subtropical forests of Europe. The southern
ivy descendant, palmated aepyhedera, differs from small northerners – it is
a magnificent evergreen ligneous liana with thick trunk, rough bark and holdfast
roots growing at all extent of trunk. With their help thick stalk of this plant
(“Aepyhedera” means literally “over-ivy”) strongly keeps on tree trunks. It
is a perennial plant reaching the age of 200 years and more. The age of separate
rods of this species can be only a little less, than the age of environmental
trees. Aepyhedera is the giant among lianas: at adult plant up to ten stalks
more than 70 meters long can develop, entangling some near trees. The bases
of trunks lay on the ground, being bound with roots of trees and making almost
impassable sites of forest where they grow. They are covered with thick bark,
which slivers as separate plates. On trunk not only short holdfast roots, but
also long crampons develop; these ones freely hang down and reach the great
length. Reaching the ground, they grow thick, lignificate, and also deliver
an additional nutrients to the plant.
Leaves of aepyhedera are digitate and have rounded shape. As the plant develops,
leaf shape changes. At young plant leaves are small and oval. Then they become
pentagonal, resembling leaves of ivy. As the plant grows, leaves gradually
become lobed, and spaces between leaf lobes increase. The size of leaf is increased
further, and additional lobes develop. At completely developed plant the leaf
reaches diameter of 70 cm and has 7 – 9 lobes. The tip of each lobe is extended
to dripping tip, helping to excrete the surplus of water.
Having reached the stage of full physiological maturity, aepyhedera begins
to blossom. Before flowering it changes the appearance of leaves again. On
trunk the flower-bearing shoots covered with integral leaves, having the lengthened
shape and dripping tip start to develop. Flowers of aepyhedera are small, white
with yellowish shade, composed to wide cyme-like inflorescences. Their smell
is rich and heavy; it can cause a headache. Because of this feature climbing
and flying mammals do not come nearer to flowers of this plant during its flowering.
But aroma of aepyhedera flowers is very attractive to large beetles and solitary
bees which pollinate this plant. From the end of summer aepyhedera fructifies.
Its fruits are bluish-violet berries up to 5 cm in size covered with thin layer
of wax. They are poisonous for mammals, but birds eat them without any harm
and carry seeds. One of basic seed-carriers of aepyhedera is cavedove
(Cavernicolumba
strigops) nesting in Balkan caves.
Ginseng tree (Megalopanax flabelliphyllum)
Order: Apiales (Apiales)
Family: Ginsengs (Araliaceae)
Habitat: South-East Asia, Jakarta Coast; rainforests.
In human epoch South-East Asia was one center of variety of ginseng family
plants. These plants form various life forms – lianas, perennial grasses, and
also small trees. When the area of tropical forests had extended, some representatives
of ginseng family appeared in conditions of strict competition to plants of
other groups. Among ginseng plants of Neocene epoch large species of trees
reaching the age of several hundreds of years had not evolved. But they had
rather succeeded as pioneer vegetation in the damaged areas of forest.
In places, where the wood patriarch undermined by time has fallen, or where
the river cuts forest, in forest canopy extensive “window” is formed and the
sunlight reaches the ground surface. Growth of large forest grasses and also
seedlings of trees of various species here begins; they compete to each other
for the right to develop and to occupy this site of forest. Among these species
prospering within few years, there is one representative of treelike ginsengs
– the ginseng tree.
This species of plants has rather characteristic appearance. Ginseng tree is
fast-growing tree about 15 meters high, having long prickles on trunk. The
tree reaches such height within approximately six years. The trunk of this
plant usually does not branch; only at damage of top 2 – 3 lateral branches
start to develop. More often one strongest branch continues to grow, and the
others dry up and break off. But in the basis of trunk there is a plenty of
buds, from which shoots develop. Bark of this plant is lacquered brown and
shining; on young sprouts it has reddish shade. In the bottom part of trunk
the old bark exfoliates and shells by long strips between thorns. Thorns, serving
for protection against herbivores, are black with white tips. On the bottom
part of trunk a plenty of thick additional roots, serving for support, develops.
Leaves of ginseng tree also have very characteristic outlines. They are alternate,
digitate, deeply dissected to 15 – 19 long lobes with rounded tips sharply
passing to thin “dropper”. Leaves are huge: they reach two meters in diameter.
Leaf has long elastic leafstalk (up to 5 meters long); foliage forms umbellate
crone characteristic for treelike ginsengs. The leaf surface is durable and
leathery; therefore dying off leaf keeps the shape for a long time and is decomposed
rather slowly. Completely advanced leaf has dark green color with white nerves;
young leaf is reddish: it is the anthocyan colouring protecting from surplus
of sunlight.
Ginseng tree belongs to the number of moistureloving plants. It grows on riverbanks,
where the forest canopy is broken by natural way. This species especially frequently
grows on river shallows and sandy spits. This species belongs to pioneer vegetation:
it is photophilous and has low competitive ability. Till the process of settlement
of other species of plants ginseng tree degrades and is quickly superseded
by other trees. The main advantage of this species in struggle for existence
is its indiscriminateness to nutrients. In fact, seeds of ginseng tree sprout
and further the plant normally develops on sand, being content with minimum
quantity of organic substances.
In tissues of ginseng tree a plenty of biologically active substances is synthesized.
They have tonic influence and raise sexual activity at males of many species
of mammals. Therefore thickets of ginseng tree constantly suffer damage from
various animals, and even strong thorns on tree trunk do not stop them. Climbing
animals eat foliage of tree, and massive ground animals gnaw bark or feel young
trees in order to eat foliage. Even predators in courtship season do not pass
this species of trees by.
Ginseng tree blossoms annually since the third year of life. In leaf axils
hanging umbellate inflorescences consisting of ordinary-looking pale flowers,
pollinated by flies and beetles, develop. Fruits of this plant are small, but
numerous berries of dark blue color with grey wax bloom. Birds (especially
pigeons and parrots) eat them willingly, and mammals find the fallen berries.
Berries also have tonic action. Seeds of this plant are carried by animals.
“Death
tree” (Necrodendron omnimortalis)
Order: Rutales (Rutales)
Family: Anacardiaceae (Anacardiaceae)
Habitat: Japan Islands, humid forests at eastern slopes of islands.

Picture by fanboyphilosopher
In human epoch Japan Islands represented one of places where
animals and plants of tropical origin penetrate to the north, using the influence
of warm sea currents softened climate of this place. The glacial epoch marked
change of Holocene by Neocene had damaged flora and fauna of islands, having
thrown heat-loving species away to the south, and having exterminated ones could
not make it. In Neocene the new wave of expansion of southern species to the
north began. However, some relicts of Holocene managed to survive in glacial
epoch, and in Neocene they had reached prosperity in ecosystems formed anew.
Among such species there was a descendant of one Far Eastern plant – poison
sumach, or poison dogwood (Toxicodendron verniciflua). In warm and humid climate
of Neocene when literally each blade of grass has chance to turn a tree, this
plant completely realized its opportunities having turned to large tree. It
has kept and increased greatly poisonous properties of the ancestor, and deservedly
has frightening name “death tree”. If it should grew in human epoch, it would
cause set of legends, inspiring mystical horror by deadly properties. But it
grows in epoch when the Earth is humanless.
The height of “death tree” reaches 25 meters: it is rather high tree of Japan
woods. In its appearance there is a set of features of former being a bush of
its ancestor. Trunk of “death tree” is strongly waved and rough; this tree often
grows as two trunks and more which fantastically bound by branches and grow
together. Branches of this tree grow bending upwards, thus they often overwound
and weave. In common they form high egg-shaped crone, and the top of tree looks
pointed. On contrary to the name, “death tree” differs in unique vitality: fallen
on the ground, but not dead, trunk and branches easily take roots, and the plant
continues growth. Bark of “death tree” is dense and grey with darker spots,
in the basement of trunk it is dot with deep cracks.
“Death tree” grows in places where the ground layer may be rather thin. Therefore
in the basement of trunk this tree huge plank-buttress roots develop giving
to the plant additional support. On these roots dormant buds develop often,
and from them new trunks sprout at damage of root.
Leaves of “death tree” are compound and odd-pinnate, about one meter long. Each
leavf consist from 7 – 11 pairs of small peaked leaflets. Leaves are light green
with skinny shining top side and soft wrong side covered with thin hair. “Death
tree” belongs to number of plants able to move by leaves: for night its leaves
hang. They also behave the same way before the rain.
This plant has kept poisonous properties of its ancestor, and due to them is
reliably protected from herbivores at any stage of development. Transparent
viscous sap of tree is extremely poisonous: it is an adaptation against the
wood-boring vermins. On this plant only some species of herbivorous insects
live, and it is reliably protected against the majority of species of insects
damaging surrounding trees. Due to alcaloids the plant has ability to suppress
fungi growth, therefore on its trunk sponks do not settle. Wood of “death tree”
as if emphasizes the impression from plant as a whole: it is colored meat red.
Wood is so rich in alcaloids that even the dead tree is decomposed for very
long time – about three years pass before on its trunk first wood-decomposing
fungi settle.
The touch to young sprouts of “death tree” may cause a chemical sting: in young
bark of this plant urushiol alcaloid accumulates, causing skin inflammation.
Therefore herbivorous mammals avoid eating young growth of this plant. Also
for young shoots and sprouts of “death tree” have a bitterish smell is characteristic,
warning of inedibility.
“Death tree” blossoms till almost all year having a break in winter months.
Flowers of this tree are ordinary-looking and tiny, have five white petals.
They are gathered to cluster inflorescences hanging down from leaf axils down
on long pedicels. They are pollinated by small moths involved with aroma amplifying
at night. Fruits of this plant are small dry drupes surrounded by strongly grown
juicy cup-shaped pedicels of red - orange color. They are eaten by unique species
of birds – the sumach finch.
In stomach of this bird under the action of gastric juice the environment of
seeds becomes thinner and permeable. Germinating ability of seeds passed through
intestines of bird increases. But finches in every case exterminate a part of
seeds: when pedicels dry up, birds begin simply peck seeds out from them and
crush by beak. However at high competitiveness of “death tree” the damage caused
by finches is much less than benefit brought by them to tree in seeds carrying.
In winter “death tree” falls into condition of relative rest: the plant stops
growth and casts a part of foliage. Also at it separate small branches thickened
the crone fall down. It brings to the plant considerable benefit – secretions
of decomposed foliage sterilize soil and kill sprouts of other plants.
“Death tree” is very moistureloving plant; in dry years its growth turns slow,
and the tree casts part of leaves. Till the long drought tree grown tens years
may even perish. In some degree it restrains the expansion of “death tree” in
woods of islands. At the same time this plant is sensitive to soil inundating
and perishes during strong floods from roots inundating. Therefore “death tree”
grows only in places where soil is well drained and simultaneously is constantly
damp – more often on hillsides turned to ocean. In these places “death tree”
forms the continuous thickets lasting to hundreds meters.
In continuous thickets of “death tree” only few animals live – birds avoid this
plant, and only sumach finches feel like at home among branches of this plant.
They can eat seeds and young sprouts of this tree without harm for themselves.
Strangler
sandal tree (Odoroxylon scandens)
Order: Santalales (Santalales)
Family: Sandalwoods (Santalaceae)
Habitat: tropical woods of Southern Asia.
The sandal trees family is most richly submitted in tropical woods. Plants leading
partly or completely parasitic way of life belong to it. Some species of these
plants have suffered from cutting down by people (at least one species was completely
destroyed) because of valuable aromatic wood. After human disappearance on Earth
and global warming, when the area of tropical woods has increased, plants of
this family have returned lost positions, having formed new original species.
Strangler sandal tree is very original semi-parasitic liana. This plant reaches
very impressive size: length of its horizontally growing trunk may reach up
to 90 - 100 meters. The plant gives set of lateral branches and expands in width
to tens meters, adhering to the majority of trees suitable to growth in vicinities.
The plant begins life as an epiphyte: birds or insects wear out its seeds in
crown of any large tree. During the first some days sprout develops due to spare
nutrients received from parent plant. But its roots gradually start to bore
bark of tree, on which the plant has lodged. If the tree appears not suitable
for being of the host plant, the strangler sandal tree can develop long time
as an epiphytic liana. It forms sprouts reaching for nearest trees. Having reached
the next tree, sprout forms some additional roots penetrating through bark of
the tree.
If the young plant was lucky, or if sprout has to found a tree satisfying needs
of strangler sandal tree, in places of contact to bark of tree sucking additional
roots boring bark of tree and taking root in its wood are formed. Due to emitting
phytogormones the place of accretion forms original “unit” in which wood of
the host tree and this liana are “bound”. On cuts of such “units” interesting
pattern is formed; it is especially effectively looking, when wood of parasite
and host plant differs by color. At the strangler sandal tree wood has very
dark, almost beet-red colouring. It is rich in essential oils and smells pleasantly.
Oils protect plant from wood-boring insects. The trunk of the strangler sandal
tree, which has received generous inflow of additional feeding, starts to grow
in length and thicken strongly. Lateral branches reach to new trees, and soon
the big area of forest appears braided by one large plant. It is easy for noticing
among branches: bark of the strangler sandal tree is very light, almost white.
As against to some relatives, completely passed to parasitic existence, the
strangler sandal tree depends on host tree only partly. From host tree it receives
only water and mineral salts which usually does not suffice to epiphytic plants.
At this species leaves are not lost; the plant is able to independent photosynthesis.
From horizontal trunks the strangler sandal tree gives numerous vertical photosynthesizing
sprouts with characteristic oval leaves. In three - five years' age the strangler
sandal tree begins blossoming. At the tips of photosynthesizing sprouts vertical
inflorescences develop – truss of set of small pinkish flowers with strong pleasant
smell. They are pollinated by insects – basically by beetles and flies though
sometimes they are visited by wasps and single bees. This plant is monecious,
but staminate flowers blossom only after decaying of pistillate ones. Fruits
are white sweetish berries with thin skin and one large seed inside. They are
carrying by birds, sometimes fallen berries are eaten by rodents.
Creeping
aromoxylon (Aromoxylon scandens)
Order: Santalales (Santalales)
Family: Sandalwoods (Santalaceae)
Habitat: Southeast Asia, tropical forests.
In human epoch sandalwoods had an important value as a source of valuable wood,
that has resulted in significant decrease of their number and to extinction
of some species, especially of local endemics. Reduction of areas of tropical
forests resulted in extinction of highly specialized semiparasitic species of
these plants and in survival of mainly less specialized forms capable to parasitize
on numerous plant species. In Neocene, at stabilization of environmental conditions
and expansion of the area of forests, among plants of sandalwood family two
strategies expressed: narrowing of specialization at increase of a degree of
fitness to parasitism and, on the contrary, expansion of spectrum of host species
at keeping of rather low specialization to parasitic habit of life.
In tropics of Southeast Asia a semiparazitic species of sandalwoods lives –
creeping aromoxylon, a very large lignifying liane growing in forest canopy.
Aromoxylon begins its life as small liane-looking plant of underbrush. It does
not lose green leaves, and is capable to develop for many years in underbrush,
like sprouts of numerous tropical trees. Creeping stalk chooses a direction
of growth, being guided on the chemical substances emitted into the ground by
roots and fallen leaves of suitable species of host plants. Having reached up
an attractive tree, liane begins to climb up on its trunk, attaching to its
trunk and branches by adventive roots. In places of contact roots start to pierce
wood of support tree, and in these places in due course woodknobs of mix of
tissues of tree and liane develop. Plant receives water and mineral substances
from host tree and due to it grows to huge size. An adult plant can die off
in the lower part and grow in tree crones exclusively as a hemiparasite, depending
from them entirely in terms of water and mineral substances supply. Despite
of it, plant keeps foliage and normal ability to photosynthesis. Merging with
conducting system of host tree, plant supplies it with sugars and other products
of photosynthesis.
Stalk of adult plant of this species reaches thickness of 15-20 cm; it is covered
with thin and smooth grey bark. Wood has very expressed characteristic pleasant
aroma. Leaves are simple, peaked, with leathery surface and smooth edges.
Flowers are tiny, modestly-looking, gathered in racemae developing on young
branches. Fruit is a large stony fruit having bright yellow peel and juicy pulp,
containing one large seed. Seeds are dispersed mainly by large ground-dwelling
mammals, which are attracted by strong smell of the ripened and fallen fruits.
Though birds willingly eat fruits of this plant, they almost do not play a role
as dispersers of fruits: in bird stomaches, seed covers dissolve.
Young plant begins to blossom approximately at the age of 10-12 years. Life
expectancy frequently makes up to 200 years and more. Frequently the plant outlives
host trees, and exists due to the nearby trees. Thus parts of the plant merged
to the old host, die off with it, and the tree breaks up to a number of clones
living in crones of new host trees.
Flaming poisonous tree (Poinsettia arborea)
Order: Spurges (Euphorbiales)
Family: Spurges (Euphorbiaceae)
Habitat: mountain forests of Hawaii.
In human epoch the flora of Hawaiian islands had suffered great damage. A number
of species of plants had died out because of extinction of pollinator animals
connected to them. Some species of trees had become extinct because of cutting
down, and others had suffered because of introduced herbivorous mammals. Not
the smaller damage to endemic flora was put with introduction to the islands
of exotic plants from various continents. “Tempered” in more inclement conditions
of life at the continent, they had quickly superseded a significant amount
of species of native flora. In Neocene flora of Hawaii descendants of species
of plants introduced by people take a significant place.
Slopes of the volcanos forming Hawaii are covered with magnificent vegetation.
The monotony of green color of foliage is broken by bright colors of blossoming
plants. Among them crones of one species of Neocene Hawaiian flora are especially
brightly allocated – they are red with yellow, as if covered with fire. This
plant bringing bright colors in palette of tropical forest, is the descendant
of poinsettia, or “Christmas star” (Euphorbia (Poinsettia) pulcherrima), introduced
by people. This species is named flaming poisonous tree, and its features justify
this name.
Flaming poisonous tree is a moistureloving plant which very well feels like
in damp air of gorges of the Hawaiian mountains. It is especially plentiful
in humid forests of eastern slopes of mountains. Usually it is suffrutex or
low tree growing by some trunks. The height of this plant reaches 6 – 8 meters.
Bark of flaming poisonous tree is greyish-brown, thick and friable. In the
bottom part of trunk it exfoliates as long strips. Leaves of this plant are
simple, up to 20 cm long, wide, with large angular lateral lobes. The tip of
leaf extends as a characteristic “dropper” which helps to remove surplus of
water from tissue. Sap of this tree is white, as at the majority of representatives
of spurges family. It contains a plenty of rubber which at drying up forms
drips and stops up the wounded places on trunk and branches. Sap is not as
poisonous, as at some continental species of spurges, but has very burning
taste and strong irritating action. This tree is protected from wood-boring
insects, filling with plentiful lacteal sap apertures, which they drill in
trunk. The foliage of this tree is also inedible, but some animals of Hawaii
specially eat it in small amounts in order to remove internal parasites.
This species of trees belongs to the pioneer vegetation of the archipelago
inhabiting fields of stiffened volcanic lava after eruption. When volcanic
lava hardens and among it grassy plants expand, flaming poisonous tree appears
one of the first settlers among wood plants. It grows very intensively – young
plant within one year forms three-meter trunk and starts to form a crone. The
trunk of this tree in youth is four-costal, but later friable core is destructing
and in trunk apertures appear, frequently through ones. Thus secondary growth
proceeds, edges of cracks become covered by bark and the trunk of elder specimens
resembles some trunks growing in parallel and partly accreting with each other.
At old trees trunk can break on some parts in case of hurricanes or simply
under weight of wet foliage after rain. However thus tree stays alive and the
parts of trunk, which has kept connection with roots, continue their growth
– branches bend upwards and form additional roots. Frequently trunk lays on
mountain slope, and tree continues to live, giving new vertical shoots. Life
expectancy of this species does not exceed 100 years.
Inflorescences of flaming poisonous tree are very small, as at all typical
representatives of family. An inflorescence represents cyathium typical for
spurges: the pistillate flower with reduced perianth is surrounded with several
small staminate flowers. But flowering of this tree will not pass imperceptibly
for pollinators: inflorescences are surrounded with bright bracts. This plant
is pollinated by various birds which are descendants of species introduced
to Hawaii by people, as well as ancestors of this species of plants. Bracts
of flaming poisonous tree have defined the name of species: they involve birds
with contrast colouring, crimson-pink with yellow border. When the wind blows,
bracts wave, making an impression of fire. When the plant sheds its blossom,
its bracts lose brightness – they turn brown and gradually get usual green
colouring. Flaming poisonous tree blossoms in several “waves”, simultaneously
developing a plenty of inflorescences on tops of young shoots.
Small seeds of this plant are supplied with cop of white hairs. They are easily
carried by wind, and get in cracks of stiffened lava at lava fields of islands
among the first. This tree quickly develops, requiring a minimum quantity of
humus, and its roots deeply penetrate in lava along cracks. But in adult condition
it is very photophilous, and in due course of overgrowing of lava fields this
species is gradually superseded by other plants. Also flaming poisonous tree
frequently settles on slopes of mountains where the competition to other plants
is less, rather than in valleys.
Mint
tree (Menthoxylon leucophyllum)
Order: Lamiales (Lamiales)
Family: Lamiaceae (Lamiaceae)
Habitat: southern slopes of Himalayas, the north of Hindustan, tropical forests.
Plants develop the most various ways for protection against insects: some of
them form a plenty of fibres, others are protected with the help of poisons,
and third ones enter the favorable unions with enemies of their enemies. One
of representatives of Neocene flora, the mint tree, had chosen way of perfection
of chemical protection. This plant accumulates a lot of menthol and various
alkaloids in leaves, reliably having protected itself from many pest insects.
Certainly, there are some species
of beetles, adapted to eat leaves of this species, and even using substances
received from tree for protection against their own enemies, but nevertheless
their number is much less, than could be, if the mint tree should not have means
of chemical protection.
Mint tree grows on mountain slopes, opened to wind and sun. During evolution
it has adapted to life on well lighted places, forming at times continuous thickets
in favorable conditions. From height of bird's flight thickets of mint tree
are well appreciable on the background of environmental wood: at this tree the
foliage has silvery shade. For protection against the sunlight leaves and young
sprouts of plant have rich silvery downiness, reflecting surplus of sunlight.
Leaves of mint tree are rather large (their length is about 30 cm), oval-shaped
with denticulate edges.
Mint tree is one of trees of the first size in Asian woods. Its height is up
to 50 meters. This tree grows very long time: duration of its life can reach
300 years and more. Wood of mint tree is very firm, pleasantly smelling on break:
it is literally impregnated with menthol for protection against wood-boring
insects.
But the mint tree has concluded the favorable union with pollinating insects:
its flowers are simply irresistible for every possible butterflies and bees.
Tubular flowers of this tree are gathered to long inflorescences rising vertically
on tips of branches. Each inflorescence numbers up to fifty such flowers. Colouring
of them is rather ordinary-looking: perianth is pale lilac. The fauces of flower
is opened, and the lower lip represents fine “airfield” for insects. All secret
of appeal of flower is inaccessible to sight of vertebrates, but involves insects
from apart: internal parts of flower reflect ultra-violet light, and on lip
in ultra-violet light the strip is appreciable, indicating to insect a way to
nectar.
Except for ultra-violet pattern, flowers involve insects with strong pleasant
smell with a shade of mint. Fruits of mint tree are dry pods with firm seeds.
On tips of seeds hooks, assisting them to cling to wool of animals, wandering
under tree, grow.
Except for seeds, the mint tree can breed by root offsprings: on the surface
of thin lateral roots there are buds sprouting at damage of root. So at some
distance from adult tree young trees of the same species appear. When the parental
plant falls because of old age or hurricane, young growth grow competing in
speed, and soon in a wood the whole group of high trees with silvery foliage
is formed.
Splendid
aepymona (Aepymona superba)
Order: Lamiales (Lamiales)
Family: Lamiaceae (Lamiaceae)
Habitat: eastern part of North America in the area of temperate and warm-temperate
climate.
Human epoch is marked by extensive selection work with particular species of
animals and plants, which have got new features in domesticated condition. Usually
these features appeared harmful to survival in the wild, because their consequences
represented changes in physiology and acquiring of other non-adaptive features.
After crash of an industrial agriculture and human extinction, numerous domesticated
plants and animals have quickly died out. However, in a case of a decorative
oswego tea (Monarda) grassy plant, selection work has presented a plant with
bright red colouring of blossoms, which has enabled descendants of a garden
plant to get new pollinators, hummingbirds. Restoration of natural habitats
has enabled the feral descendants of oswego tea to settle widely, supplying
migrating hummingbirds with food and evolving to new species.
A descendant of oswego tea in North America is splendid aepymona – perennial
rhizomatous grassy plant. Stalks are robust, tetrahedral, covered with short
white trichomes, slightly lignifying in bottom part. In southern part of range,
stalk can continue its growth till 2 years in succession, reaching height of
250-300 cm. Hence the name of a plant: the prefix aepy-means a superlative degree,
and -mona is from the name of an ancestor – Monarda. In the north of range,
stalks are annual, but up to 200 cm tall. Leaves are opposite, up to 20 cm long,
bright green, elongated with much extended tips. Nerves are foveate, protrudent
from the bottom side of leaf.
This species is a moistureloving plant, which prefers to grow at damp meadows
and slightly waterlogged ground. During flooding the plant grows and blossoms
normally, enduring flooding within several weeks.
Flowering begins at the end of spring and continues up to the middle of summer;
this species is an important fodder plant for migrating hummingbirds. Flowers
are large (up to 8 cm long), tubular, narrow, slightly curved, ornithophilic.
They have bright red color from outside, grow in the top part of stalks in verticils
in leaf axils. In flower fauces there is a pattern of dark oculate irregular-shaped
spots with white edges; on a lower lip of a flower longitudinal white strokes
stretch, being directed deep into the fauces of a flower. White marks are formed
of dense transparent trichomes, reflecting ultra-violet rays – these are indicator
marks of a way to nectar. Reserve pollinators of plants are bumblebees and large
solitary bees capable to distinguish these marks and having long enough proboscii
to get nectar from depth of a flower.
Fruit is a dry capsule; seeds are easy, capable to float of water surface. Usually
they are distributed by water – during the summer flooding or autumn rains.
Young plant begins to blossom at the third year and reaches a peak of development
at the age of 8-9 years. The rhizome can normally develop up to 15-20 years.
Polar
mint (Menthula densa)
Order: Lamiales (Lamiales)
Family: Lamiaceae (Lamiaceae)
Habitat: tundra areas of Eurasia, Northern Europe, Polar Ural, Taimyr Peninsula,
islands of Arctic Ocean; stony wastelands.
In human epoch among cultivated plants a plenty of hybrid forms has appeared.
Many of them have died out after human disappearance, not having endureed a
competition to wild plant species, but some hybrid plants appeared viable enough
to survive in the wild for rather long time and to evolve, becoming ancestors
of new species of plants of Neocene epoch. In Neocene flora there are many species,
which ancestors were the hybrids cultivated by people in historical epoch. One
of such species is polar mint, the descendant of hybrid peppermint (Mentha piperita).
This species has mastered life in inhospitable areas of tundra and forest-tundra
in the northern Eurasia, where it rather successfully competes to other plant
species.
Polar mint is an evergreen grassy plant with short shoots forming a dense cushion.
Plant is a rhizomatous perennial grass; underground rhizomes spread in all sides
from parental plant. Shoots are up to 10 cm tall; they grow very densely next
to each other, branching from the basis. Leaves are opposite, rounded, up to
2 cm in diameter, rugose, with denticulate edges. Surface of leaf is covered
with short trichomes, bottom side is plentifully pubescent. Epigeal portion
of plant contains a lot of menthol giving to greenery a characteristic mint
smell.
Flowers are composed in short spherical inflorescences on tips of shoots. Flower
of tubular shape is light lilac, covered with trichomes reflecting ultra-violet
light. The present plant species is pollinated by solitary bees, bumblebees,
flies and small butterflies.
Seeds are very small, covered with hook-like bristles enabling to hooked against
plumage of birds. They grow ripe rather seldom, and the plant usually propagates
by means of repent rhizomes. It frequently happens, that all plants of this
species from a certain island represent only few large old clones.
Plant is the important fodder resource for herbivores – from insects up to mammals.
In some areas polar mint is a major melliferous plant supporting existence of
a population of insects.
Violet
trumpet tree (Tabebuia violacea)
Order: Mint (Lamiales)
Family: Trumpetvines (Bignoniaceae)
Habitat: gallery forests of central and northeastern South America.
The Genus Tabebuia of the family Bignoniaceae survived the Holocene, and in
the Neocene a new species appeared, the violet trumpet tree. This species is
native to savannas, plains and semi-open areas of South America, but being found
in larger numbers, together with the riverine
almond (Neoterminalia riparia) and the dendém
oil palm (Elaeis dendem), in the gallery forests of South America to the
south from selva zone.
The violet trumpet tree is a dry season-deciduous tree growing to 20 m tall.
The leaves are palmately compound, with six leaflets, each leaflet 15 cm long,
green with silvery scales both above and below. The tree crown is wide, with
irregular, stratified ramification and only few thick branches. The bark is
dark grey and may be vertically fissured.
Flowering occurs in dry periods, when the tree has none, or very few, leaves.
The flowers are bright violet, up to 5 cm diameter and are produced several
together in a loose panicle. The polinization is realized by birds and insects.
The long and slender fruit capsules can measure up to 30 cm. After the drying
fruit dehisces the anemochorous, winged seeds are released. They can be transported
by the wind for long distances.
This plant species was discovered by João Vitor Coutinho, Brazil
Australian
Bushy Olive (Austrolea edibilis)
Order: Lamiales (Lamiales)
Family: Olives (Oleaceae)
Habitat: Southern Meganesia, zone of Jarrah bushland, bushy savannah.
Australia during the age of man, as with many countries on earth, played host
to many plants and trees which were invasive “weeds”. A common tree which was
a weed in the southern part of Australia was the European Olive (Olea europaea)
due to commonly being grown in this area for oil due to favourable climate.
It has left a few descendants in Neocene Meganesia, though none are tree-shaped
as its ancestor sometimes was.
This plant is a mid-sized shrub reaching as much as 1.7 meters tall, though
its spread may be as much as 3 meters in diameter. It most often has a single
trunk that branches many times into a brush of fine branches, shape of this
bush is generally round or squat shaped but browsing by herbivores may cause
it to appear different. The open nature of its habitat means that it is only
partially able to deal with shade. Branches extend all the way down to the ground,
no trunk is visible outwardly, and small animals frequently shelter and nest
within or under this bush. Leaves are thin and spindle shaped, resembling their
ancestor, but are somewhat smaller and more dense. This tree is evergreen, but
prone to burning during fires due to oil within leaves and fruit. Bark is a
very dark greyish brown and the surface of the central trunk has a gnarled texture.
Root system is very extensive and this tree tolerates drought very well.
Flowers are small, white and feathery, producing large amounts of very light
pollen; tree is pollinated by the wind. Blooming begins midway through the wet
season. Fruits ripen in the beginning of the dry season; fruit is a small drupe
which is considerably smaller than that of its domestic ancestor. Small seed
5 millimetres long is encased in a sheath of oily flesh which tastes both bitter
and sweet. Whole fruit is round, up to 1.7 cm in diameter and dark purple when
ripe. Many birds and mammals consume the fruit of this tree, and distribute
its seeds in their dung. Seeds are thick shelled and may resist a fire if it
passes over quickly enough, in this way discarded seeds may survive even if
many adult trees are killed off by burning. Young plants begin to bear seed
in as little as 5 years. Individual bushes may grow in girth and spread over
their lifetime, and live as long as a hundred years.
This plant species was discovered by Timothy Donald Morris, Adelaide, Australia
Alien
rootflower (Rhizoflosculus xenos)
Order: Scrophulariales (Scrophulariales)
Family: Figworts (Scrophulariaceae)
Habitat: forests of South-Eastern Europe (southern and western coast of Fourseas)
In Holocene among representatives of figworts family it was a plenty of species
which leaded semi-parasitic habit of life – they were attached by underdeveloped
roots to root system of other plants and received through them mineral substances
from ground. But they kept green leaves, and looked externally like other most
usual grassy plants. Also in family true parasitic species were represented
in small amount.
In Neocene the tendency of passing to parasitic habit of life had amplified
at them. It was favoured with changes of climate resulted in formation of new
productive ecosystems. Humid subtropical woods at the coast of Fourseas became
a habitat of new species of parasitic plants.
On roots of high trees of European subtropical woods it is possible to see flowers
of this plant, hence it has received the name rootflower. This parasitic plant
has no own green leaves and lives practically completely due to the host tree.
Its relations with host tree develop originally: it is parasitism inclined to
symbiosis. The rootflower exists the most part of year as tissue bundles under
bark and in thickness of wood of host tree. Its presence causes fantastical
knotty inflows on roots and bottom part of trunk (not higher than at one meter
above the ground). But the most interesting fact is that at rootflower its own
root system is very well advanced. Evolution had made an original step back:
first the plant had “attached” to host tree and became dependent on it, and
then relations of parasite and host began to develop to absolutely another direction.
Under the ground rootflower develops rich circuit of roots connected to mycelium
of various species of local fungi. It brings huge benefit both to parasite,
and its host: the host tree receives from rootflower mineral substances from
ground, but in exchange the parasitic plant takes a part of organic substance
synthesized by the host. Vascular system of host and parasite plants interlace
inside knot-like inflows on trunk which in due course strongly expand. The rootflower
can parasitize at the plenty of species of deciduous trees of European woods.
It avoids only plants on which lianas of family Araliaceae, descendants of ivy,
grow. Obviously, the substances emitting by these plants oppress sprouts of
rootflower at early stages of development.
In summer on inflows formed of tissues of parasite and also on roots of the
host tree flowers of this plant form. Two strong gristle-like bracts punch bark
of the host plant and open, letting outside a single flower. Flowers of this
parasitic plant are large – the diameter of flower reaches 4 – 5 cm; they also
are numerous – at one square meter up to several tens flowers blossom. Flower
of rootflower is bilabiate with perianth extended to tube, bright yellow with
lilac “ocula” in the middle. The top petal is covered with silvery hair reflecting
ultra-violet light, which develop in longitudinal strips converging in fauces
of the flower. It is the original “sign” for pollinating insects. They are involved
also with strong aroma with slight putrefactive shade. Flowers of this plant
are pollinated mainly by single hymenopters though occasionally they are visited
by butterflies. Petals of rootflower are densely closed and only the strong
insect can move them apart, therefore small casual pollinators like flies can
not reach nectar. Butterflies with long proboscis, large species of diurnal
hawkmoths, may simply penetrate thin proboscis between densely closed petals
and drink nectar not making pollination.
The fruit of rootflower is very original – in it the “constructive decision”
of plants of Cucurbitaceae family known as squirting cucumber (Ecballium elaterium)
was independently developed and repeated. Ripen fruit of rootflower is rounded
pod about 3 cm long, filled with liquid in which ripen seeds float. The liquid
is under high pressure. The fruit is opened with the help of small cover at
the tip. When large animal casually touches fruit, it “blows up” and throws
to the animal sticky liquid with seeds. Seeds of rootflower are tiny; they easily
stick to wool and are carrying by animal to other places. When animal rubs against
trees, seeds remain on bark and sprout. Sprout of rootflower is very delicate
and sensitive: phytoncides emitted by other plants easily suppress it. Therefore
rootflower meets in woods of Europe rather seldom. If the species of host tree
is suitable for rootflower and nothing interferes with development of parasite,
the sprout takes root under bark of tree and expands. Till first years of life
it develops in ground thin roots, searching for mycelium and forming mycorhiza.
In this time at the bark of host tree characteristic outgrowths start to form.
Having reached about ten years' age, rootflower begins to blossom.
Veey's root (Sublathraea wiyii)
Order: Scrophulariales (Scrophulariales)
Family: Figworts (Scrophulariaceae)
Habitat: forests of South-Eastern Europe (southern and western coast of Fourseas)
In Holocene at the territory of Australia people had discovered two species
of orchids distinguished by original way of flowering – cryptanthemys (Cryptanthemys
slateri) and rhizantella (Rhizanthella gardneri). These both plants were lack
of chlorophyll. They grew and blossomed under the ground. Even at the moment
of discovery these both plants were extremely rare. The Cryptanthemys orchid
had become extinct in historical epoch, and Rhizanthella had vanished after
it at the epoch of anthropogenous pressure despite of protection efforts.
In Neocene epoch at Earth one more plant leading underground life and even
blossoming under the ground had appeared. It is the descendant of toothwort
(Lathraea squamaria), the parasitic plant of figworts family. In Holocene epoch
this plant had lost chlorophyll, and in Neocene specialization of its descendant
had reached the top. The descendant of toothwort is completely underground
flowering plant, Veey's root.
Veey is the underground monster at Slavic myths, which appears on the ground
only once in four years, February, 29, and was able to kill by sight*. According
tales Veey spent all rest time under ground. The Neocenic plant growing in
forests at coast of Fourseas leads equally secretive underground habit of life.
The Veey's root is a parasitic plant which grows under the ground the most
part of year. This species parasitizes at bush plants of birches family (Betulaceae)
living in underbrush. Veey's root prefers mellow soft ground, and grows near
riverbanks or in valleys, where the ground is mellow and humid enough. Obviously
it is connected to optimum conditions of growth of its hosts.
At Veey’s root there is lack of chlorophyll, and its leaves had turned to thick
scales. The body of plant is creeping rhizome about one meter long, growing
at the depth of 20 – 30 cm. The root system of plant is reduced up to several
sucker roots, with which help the Veey's root attaches to roots of host plants.
This parasitic plant completely depends on the host.
In summer when ground is warm and damp, and the host plant vegetates actively,
Veey's root begins to blossom. This event cannot be seen without any special
efforts, because Veey's root even blossoms under ground. But inflorescences
of this plant may be found out guiding at the characteristic smell involving
pollinators.
Veey's root forms inflorescences in axils of scale-like leaves densely covering
the rhizome. The inflorescence develops in wrapper of several strong juicy
opposite leaves. They protect flowers against damages. The wrapper “bores”
the ground thickness and slightly opens at the tip, having reached the ground
level. At ripening flowers the wrapper of inflorescence expands in width, forming
the chamber in which plant pollinators get.
Flowers of Veey’s root do not differ in refined form - they involve pollinators
not with appearance, but with smell. At them there are reduced petals and pale
grey colouring with pinkish shade. Each flower is surrounded with two big bracts.
Flowers densely sit in longitudinal lines on thick rachis of inflorescence.
Aroma of flowers of this species is strong and sweet with putrefactive nuance.
Veey's root is pollinated by ground beetles involving very much with such smell.
On inflorescences of Veey’s root it is possible to see often small ground snails
and slugs also involved with smell of blossoming plant. They do not play an
essential role in pollination, but can eat a significant part of ovaries. The
plant is escaped from damages by its basic pollinators – beetles: many beetles
feeding on inflorescences of Veey’s root are carnivorous and hunt molluscs.
After pollination, shortly before seed ripening, the plant of Veey's root is
shown at the ground surface. The rachis of inflorescence is strongly extended
and lifts ripening fruits to the height about one meter above the ground. Fruits
of Veey’s root are dry pods opened by small operculums on tips. When the wind
shakes the plant, through apertures tiny seeds carrying by wind run out. In
one pod it may be some thousands of small seeds.
The seed of Veey’s root sprouts at once having got on the ground. The sprout
searches for chemical substances – secretions of roots of the host plant, and
grows directly to these roots. Usually significant part of sprouts perishes
not having found suitable place for life. The Veey's root develops slowly:
from germination of seed to the first flowering over 10 years may pass.
---------------
* Note for foreign readers about Veey: this monster is described like very
stumpy-built human-like creature with long and heavy eyelids. It is not able
to lift them, and demons do it using pitchforks. The glance of Veey is harmful
and deadly for all live beings. Veey spends almost all life in caves or in
other holes.
Uzambarian snowleaf (Niveophyllum usambaricus)
Order: Scrophulariales (Scrophulariales)
Family: Gesneriaceae (Gesneriaceae)
Habitat: Uzambara Mountains (Zinj Land).
Gesneriaceae is one of characteristic families of plants of tropical zone.
In Holocene epoch it was presented by extremely grassy plants of different
habitus - from tuberous perennial plants and evergreen epiphytes up to annuals
and monocarpic grasses with unique leaf. In glacial epoch the specific variety
of this family had considerably suffered because of reduction of the area of
suitable for inhabiting places of tropical zone. But in Neocene when evergreen
rainforests had widely distributed at Earth, these plants had received
a new opportunity for evolution, and used it maximum full.
Neocenic Gesneriaceae had occupied even such habitats which were earlier inaccessible
to them. One of such atypical representatives of family lives in mountains
of Zinj Land. When the African continent had splitted along the line of Great
Rift Walley, and its eastern part had broken away, the highest mountains of
Africa had ceased to belong to the continent. Live creatures had stayed on
Zinj Land, began to evolve irrespectively to continental species.
It is rather low plant – up to one meter height. Its vital form is uncharacteristic
for family: at it there is thick lignificating trunk. At top of trunk the crown
of wide leaves covered with white downiness is stretched. Young leaves are
haired especially rich: it seems, that snow lies in the middle of plant. For
this feature the plant has received the name “snowleaf”. However this species
do not grow where the true snow falls: snowleaf has kept relative heat-loving
characteristic for family. The ancestor of this plant is the species widely
known in human epoch as “African violet” (Saintpaulia ionantha).
The shape of this plant is somewhat reminiscent some huge species of lobelias
(Lobelia) dwelt in mountains of Africa in human epoch. The trunk of snowleaf
is covered with leaves only in the top part where they form the wide crown.
The basis of trunk is environmentproofed by friable spongy bark which softens
difference between day time and night temperatures: at night in habitats of
snowleaf it happens rather cool, and occasionally even there are light frosts.
Therefore such protection is very useful for this plant. If light frosts are
too delayed, old leaves die off, but do not fall off, and remain on trunk as
additional thermal isolation. But some of young leaves most richly covered
with silvery hairs, always survive, and the plant is quickly restored after
colds.
If light frosts appeared too strong, the plant does not perish all the same:
at the basis of trunk of snowleaf from dormant buds young crowns of leaves
develop. In due course they expand, forming additional trunks, and form new
crowns. So for some tens years the single plant can turn to thicket with several
tens trunks.
Almost the year round snowleaf blossoms, making a break only in coldest time.
Flowers of snowleaf are gathered in big inflorescence numbering up to 20 flowers.
Flower is long, tubular, white with strong smell. In fauces of flower there
is a pattern of small red spots merging in depth of flower to one spot. Flowers
of snowleaf are opened constantly, but their specific spicy aroma is not almost
felt in the afternoon, but amplifies by night. At this time tiny
bats eating
exclusively nectar of plants fly to the feeding. They should not fly far for
the forage: they live under leaves of snowleaf where they have the convenient
warm dwelling protected from wind. The same animals pollinate this plant. Peduncle
is very strong, and bats licking nectar can simply jump from one plant to another.
Fruit of this plant is small dehiscent pod with very small seeds which are
carried by wind. Sprouts of snowleaf are very weak and delicate, and the most
part of them simply perishes among grass. But some sprouts succeed to survive
and grow up. One more way of breeding of snowleaf is vegetative breeding with
the help of leaves. Leaf of plant broken off by wind or animals can easily
take roots in ground and give some young plants.
Right-sided asymmetrocalyx (Asymmetrocalyx dexter)
Order: Scrophulariales (Scrophulariales)
Family: Gesneriaceae (Gesneriaceae)
Habitat: humid tropical forests of Amazonia, forest canopy.
Tropical forests of Amazonia in congelation epoch had gone through an extreme
degree of decline – their area at some stages of congelation broke up to a
number of isolated islets where only small part of former variety of their
inhabitants was kept. After deviation of glaciers, when the climate became
warmer again, rainforests had formed a continuous cover in northern half of
South America. A variety of rainforest inhabitants of Neocene has caused a
competition, and strict specialization as its consequence. Flowering plants
began to develop various ways of attraction of pollinators, leading up them
up to various degrees of refinement. Some flowers are hospitably open to all
pollinators, and others give nectar only to those ones, which are adapted to
extract it. Representatives of Gesneriaceae family belong to the plants pollinated
by birds and insects. Representatives of one genus of these plants evolved
long time in common with the special genus of hummingbirds, and indestructible
connection between bird and plant species became a result of this evolution.
Flowers of this species of Gesneriaceae have long tubular nimbus with bright
limb, hiding nectar from strangers by simple way: it is bent sideways. For
this feature plant has received name Asymmertocalyx (“an asymmetrical calyx
of flower”). The bend can be both to the right, and to the left, and two species
of this genus of plants realized both variants of a structure successfully.
These species differ from each other in habitats, and their pollinators also
do not meet in common with each other.
Both species of these plants are epiphytes living in forest canopy. In tropical
forests of Amazonia right-sided asymmetrocalyx lives; it is the species, at
which tubular nimbus bends to the right. The inflorescence at this plant is
a long ear on strong vertical flower-bearing stem, and the axis of inflorescence
is spirally twirled. Flowers grow in pairs on short stems in axils of wide
rounded leaflets. In one node of flower-bearing stem, thus, four flowers grow.
Flower-bearing stems of this species are covered with rich white hairs – it
is the “family feature” of Gesneriaceae.
The length of flower tube is about 5 centimeters that approximately corresponds
to the length of beak of the humming-bird pollinating this species. For attraction
of the pollinator bird the limb of nimbus has bright red coloring with strips
of silvery hairs stretching from edges to the pharynx of flower. Diameter of
the flower of this species is about 2 centimeters. A fruit of asymmetrocalyx
is rounded dry pod dehiscent in casual direction. Smallest seeds are supplied
with cops need for flight.
Asymmetrocalyxes are perennial plants with strong creeping stalks. They attach
to bark of host tree - by set of thin roots, and the beard of white roots gathering
dew and rain water hangs down in air. Leaves of right-sided asymmetrocalyx
are dark green, relief and impressed. They are not so large, leathery and having
a small amount of hairs on top side. Leaves are gathered in dense crowns, and
during the rain they direct rain water to roots.
This plant is pollinated by bacororo hummingbird, or left-sided
crookbill hummingbird (Asymmetrochilus bakororo), which beak is adapted to extracting of nectar from
such flowers. Fitness of asymmetrocalyx to pollination by hummingbird is not
absolute – some large butterflies also can extract nectar of this plant, having
turned sideways. And bees simply gnaw through tube flower, and also extract
nectar, passing a bend of flower nimbus.
The related species – left-sided asymmetrocalyx (Asymmetrocalyx sinister) –
lives in cool foggy forests on mountain slopes. This species has adapted to
life in conditions of high humidity and cool climate. The whole plant is covered
with intensive white downiness, which detains drops of dew, protecting leaves
from rotting. Numerous short air roots gather moisture from the surface of
plant. They develop even on flower-bearing stems, from the same node, as leaves.
Flowers of this species are carmine-pink and larger, than at rainforest species
– their diameter is up to 5 cm. Left-sided asymmetrocalyx is pollinated by
itubory, right-sided crookbill
hummingbird (Asymmetrochilus itubory). But also
this plant can settle effectively with the help of vegetative breeding – on
tips of its large leaves brooding buds with short leaves and hairs modified
to hooks are forming. The brooding bud clings to plumage of birds and easily
breaks off parental plant, and birds carry such buds in forest, leaving them
in places which may be suitable for growth.
Tube-flowered
kohleriopsis (Kohleriopsis tubiflorus)
Order: Scrophulariales (Scrophulariales)
Family: Gesneriads (Gesneriaceae)
Habitat: South America, tropical rainforests of Amazon and Hyppolythe river
basin.
Destruction of tropical forests in human epoch was accompanied by mass extinction
of animal and plant species, for which forest was an only inhabitancy. Local
species with the low number, strictly dependent on locally established conditions
of their inhabitancies have especially strongly suffered. After human disappearance
and restoration of acceptable life conditions, active evolution of epiphytic
plants mastering canopy of new tropical forests had begun.
Tube-flowered kohleriopsis is a representative of epiphytic gesneriads. Due
to tiny seeds these plants managed to master easily a forest canopy, where various
species of this family live. Kohleriopsis grows in tree crones on large horizontal
branches and trunks of lianes. It is a perennial grassy plant; the basic attachment
organ is its rhizome covered with scale-like rudiments of leaves. It grows horizontally,
and imbeds in cracks of tree bark, filling them in due course of growth. Sometimes
plant can “creep” on tree trunk and expand upwards and in sides. Plant attaches
to substratum by means of numerous small roots, among which the wood dust accumulates
– plant takes mineral substances from it. From rhizome short vertical shoots
grow, on which flowers are formed. Stalks and leaf petioles leaves are covered
with reddish-brown trichomes, in nodes air roots develop. Leaves are simple,
opposite, rounded, dark green with pointed tips and denticulate edges, up to
10 cm long. Bottom side of leaf is greyish-white. Leaf surface is covered with
numerous trichomes giving to it a velvety look. On the shoot up to 15 cm tall
4-6 pairs of leaves usually develop; after that growth of stalk slows down,
and on its top the development of flowers begins.
Flower stalks are short and strong, rising for 5-8 cm above leaves. Flowers
of tube-flowered kohleriopsis are remarkable in their very characteristic shape
– they are tubular, about 15 cm long at tube width not exceeding 8 mm. Because
of great length of tube end of flower sticks out over the edges of leaves and
pollinator is compelled to drink nectar exclusively in flight. Diameter of pentapetalous
flower does not exceed 3 cm. Corolla tube is straight; flowers are ornithophilous,
but can be visited by large bees incidentally. Flower tube is colored in various
shades of red – from dark pink up to wine-red; one ecological race of this species
has white flowers and is pollinated by hawk moths. Petals are spotty – on background
color dark red spots are scattered (except for white-flowered race), and on
them lines of trichomes grow, reflecting ultra-violet light. Trichomes serve
as nectar indicators, and are especially well-advanced at white-flowered race.
On borders of ranges of various races, forms having intermediate characteristics
of flowers exist.
Seeds of this species are very small and anemochoric. From germination of seed
to the first flowering of young plant about one year passes.
Chimaeric
bladderwort (Utricularia amphibiotica)
Order: Scrophulariales (Scrophulariales)
Family: Bladderworts (Lentibulariaceae)
Habitat: temporary reservoirs in pampas of South America.
Many groups of plants had rather safely overcome the boundary of Holocene and
Neocene, and evolved to new species in restored natural communities. Flesh-eating
bladderworts (Utricularia) represented one of widely settled groups of carnivorous
plants. In human epoch they lived in all continents, except for Antarctica,
and this state of affairs had not changed in Neocene.
The majority of new species of bladderworts differs only a little from their
ancestors, but there are also exceptions; for example, in swamps of Siberia
and China giant bladderwort grows; it is a true
giant among its relatives, and in opposite side of world, in prairies of South
America, its distant relative named after antique monster grows.
In ancient myths the chimera was three-headed monster symbolically uniting three
elements – fire, air and ground. Its South American namesake is capable to live
at once in three elements: roots in the ground, leaves and stalks in air and
in water, if such opportunity will be presented. This plant lives at the banks
of shallow rivers and lakes in pampean zone of South America, which tend to
dry up in dry season. The majority of bladderworts, certainly, represents more
overland, rather than underwater plants, and prefers humid habitats of tropical
forests. Other bladderworts, on the contrary, lead strictly aquatic way of life
and can not survive on land. Chimeric bladderwort manages to use both worlds
in any measure. It grows on the coasts of reservoirs and grows stalks of two
kinds. The plant lives on damp ground and forms an upright stalk with short
internodes and rounded leaves with slightly cut edges. At the top of stalk during
rain season high (up to 50 cm) not branching inflorescence, cluster with alternate
zygomorphic labiate flowers is formed. Each flower is up to 2 cm in diameter,
colored white with large reddish-brown spot on lower labium. The flower is densely
closed, and only heavy insect like beetle, fly or bee can creep into it. Having
opened fauces of flower, it pours out pollen on its back; then insect will transfer
it to another flower. Also mosquitoes and butterflies feed on flowers, but usually
they simply push proboscis between lips of flower and do not make pollination.
Seed breeding at this species occurs successfully: about a quarter of flowers
produces seeds.
While the main plant lives on land, it grows long liana-looking shoots from
axils of the bottom leaves. If they grow aside water, they begin changing, reacting
to humidity of air. Leaves on them still have rounded outlines, but turn dissected
stronger. Because of its own weight shoot falls in water soon and starts to
form thin stalk with fluffy leaves dissected on threadlike lobes on which trapping
bladders grow. With the help of these bladders plant catches tiny aquatic invertebrates
while shoots of terrestrial type form new plants on land in vegetative way:
the tip of long shoot takes roots and gives rise to new plant. If the shoot
lays on the ground, it also takes roots in nodes; from these nodes new vertical
shoots grow.
Underwater shoots can be transported to other reservoirs on legs of birds. In
dry season they perish, but form in leaf axils pinecone-looking buds in firm
environment, which keep viability in wet dirt. In the beginning of rain season
from these buds both terrestrial and aquatic shoots can grow, depending on conditions
in which the bud sprouts. Seeds easily endure drying and sprout soon after ground
humidifying. Each ground shoot of plant lives till three years in favorable
conditions. The plant is actively breeding in asexual way and forms dense thickets
at the coast and in shallow water.
This species of plants was discovered by Bhut, the forum member.
Fly-catching tomato (Neolycopersicon
muscicarpus)
Order: Polemoniales (Polemoniales)
Family: Solanaceae (Solanaceae)
Habitat: South America, light forests in tropical and subtropical areas.
In human epoch South America was known as the centre of variety of tomatoes
(genus Lycopersicon). Among these plants there were the species adapted to
various habitats: for example, at Galapagos Islands one halicole species lived.
The species had not been entered by people to agriculture appeared hardy enough
in order to go through epoch of anthropogenous pressure and global ecological
crisis at the boundary of Holocene and Neocene. In Neocene some descendants
of tomatoes had got surprising adaptations for survival. People managed to
select cultivars of tomato, which protected against small insects independently,
keeping them with the help of sticky hairs. The descendant of one South American
species of tomatoes developed the same ability independently of such tomato
cultivars, but it had promoted in this feature even further. From the simple
protective adaptation at this plant sticky hairs had turned to hunting instrument
– the plant specially involves and catches tiny insects and mites. This species
of plants is fly-catching tomato.
This plant catches insects not only to protect itself from harm. This species
had adapted to life on poor ground, and its well-being depends on presence
of insects. The fly-catching tomato exhausts the caught animals, receiving
in such way necessary amount of mineral and organic substances. Hairs on its
leaves have a special structure. Each hair represents a huge cell with strong
covers. On the tip of hair the drop of sticky substance is secreted. When the
insect lands on leaf and sticks to hairs, substances emitted by its body stimulate
growth of tip of hair into the tissues an insect. The hair sprouts into the
body of prey as a chain of thin-walled cells, which absorb its liquid contents.
The plant consumes tiny soft-bodied insects – plant lice, thripses, small caterpillars
and maggots – especially successfully. After nutrients will be absorbed, the
hair dies off and withers, and the wind carries away devastated exoskeleton
of insect. The young leaves richly covered with hairs catch insects more successfully.
They are reddish because of anthocyan pigment protecting them from solar burn.
For attraction of insects on some hairs drops of sweet liquid are secreted.
This plant is positioned at early stage of specialization to feeding on insects
and yet has no adaptations permitting to keep prey actively.
The first leaves of young plant are rounded, but on next leaves denticles at
edges appear, passing later to deep cuts. Completely generated leaves are pinnatisected,
dark green, slightly sticky and velvety to the touch. On adult leaves trapping
hairs remain only on lateral lobes and on the edge of leaf. All parts of plant
are protected from herbivores by alcaloids and have a specific smell.
Fly-catching tomato is a perennial plant. At the first year of life its stalk
is vertical, but later it becomes ascending and takes roots in nodes. Lateral
sprouts and top rise vertically, and the plant forms the dense thicket. This
plant is pollinated by insects, and, in order not to catch casually its own
pollinators, it moves flowers highly upwards on long flower-bearing stem. Flowers
of fly-catching tomato are small, five-petaled and similar to flowers of the
majority of Solanaceae. Petals grow together to short tube, and white peaked
tips of petals are turned back. Fruit of this plant is small red berry. In
ripen condition the fruit of plant is edible, and it is eaten willingly by
large ground rodents.
Some species of bugs and beetles are adapted to “robbing” of this plant: they
seize stuck insects from its leaves.
Pipe-flowered tobacco tree (Nicotiana longissimoflora)
Order: Polemoniales (Polemoniales)
Family: Solanaceae (Solanaceae)
Habitat: Great Antigua, tropical forest areas.
The inhabitancy of any species of live organisms includes not only abiotic
factors (climate, precipitation amount, features of relief and ground), but
also surrounding species. Every species inevitably interact with other species
which play roles of predators, prey or parasites, pollinators or seed and spore
distributors, commensals or symbiotes. And the pattern of interaction with
these species renders great influence to evolution of any separately taken
species.
At flowering plants interaction with animals is most brightly (in literal sense)
shown at the example of evolution of adaptations to pollination. Flower corollas
have various shapes and structure, and flowers produce the certain smell in
order correspond to maximum to features of anatomy or behaviour of one or another
pollinator and to exclude the appearing of any casual pollinators.
One example of strict specialization to the pollinator is displaying by the
plant living in tropical forests of Great Antigua island, in tropical America.
This species, pipe-fllowered tobacco tree, descends from feral domestic tobacco
(Nicotiana tabacum), which had been cultivated widely in New World by people.
Its descendant represents a large perennial plant with treelike stalk up to
6 m high. Pipe-flowered tobacco tree belongs to photophilous and fast-growing
species. In underbrush of tropical forest plants of this species grow pale
and thin and quickly perish because of lack of sunlight. This species grows
as a pioneer plant at the sites of forest where the forest stand is damaged.
Usually thickets of this species are formed in places where hurricanes had
tumbled down an old tree, and it had crushed under itself some more trees,
falling on the ground. Continuous thickets of this species meet also on the
riverbanks.
In anatomy of pipe-flowered tobacco tree everything specifies that the plant
uses to the full short-term advantages of growth on well light sites of the
damaged forest. This is very fast growing plant which is capable to grow up
to about three meters for one year. Its stalk initially is grassy and has bunches
of rigid longitudinal fibres inside. But in due course of growth it lignifies
in the bottom part, grows thick fast and becomes much stronger. The surface
of stalk becomes covered with grey bark which gradually exfoliates as narrow
strips. Leaves of this species are very large; in the bottom part of stalk
their length may be about 1 meter. Leaves of young plant are simple and oval,
with feather-veined reticulate venation. Surface of leaf between veins is slightly
inflated and folded. Edges and the top side of leaf are covered with silvery
hairs. Young leaves have richer downiness, and tops of shoots seem silvery
because of it. Having reached the height of about five meters, the plant loses
leaves on the bottom part of stalk and starts branching. Leaves on branches
are smaller, than initial ones were – their length does not exceed 70 cm. But
in shape they differ from ones developed in the first year of life of the plant:
in the basis of leaf two appreciable lobes develop, which gradually increase
to branch tips. On tips of branches leaves are three-lobed; lateral lobes make
about half of the length of central blade of leaf in length. On tips of leaves
long narrow “droppers” develop, helping to excrete the surplus of water.
Flowers of this plant begin appearing already at the second year of life. They
appear as clusters on tips of branches. Flowering of this species represents
a magnificent show. Flowers of pipe-flowered tobacco tree are tubular and hanging,
with five rounded petals. Diameter of corolla is about 10 cm, but its length
is unique for flora: about 50 cm. Flowers are bright red, covered with hairs
reflecting ultra-violet radiation. They are pollinated only by one moth species
– Antiguan giant
hawk moth. Though insects do not distinguish red color, hairs
on petals make flower well appreciable in ultra-violet light, and help the
pollinator to find it. Moreover, flowers of this plant are very fragrant, and
pollinator insect can feel their smell at the distance of about one kilometer.
The biggest inflorescences develop on the top of plant: pollinator moth does
not like to fly down too close to the ground. Occasionally insects of other
species can get nectar of this plant. Solitary wasps and bees simply gnaw through
the tube of flower at the basis and lick nectar, not making pollination.
Fruit of this plant represents dry capsule. After flowering growth of branch
in length stops and it starts forking. Counting the number of branchings it
is possible to define the age of this plant approximately. But pipe-flowered
tobacco tree does not live for a long – the maximal age of this plant makes
no more than ten years. In due course of growth this species is superseded
by young growth of the trees making tropical forest, and gradually decays.
For protection against herbivores this plant produces a mix of poisonous alkaloids
of similar chemical structure – nicotine, anabasine, coniine, hygrine etc.
Only few herbivores can eat it. However animals frequently eat the old and
fallen leaves containing residual amount of alcaloids in order to remove internal
parasites, or wallow in foliage to expel parasites from wool or feathers. Only
caterpillars of the pollinator moth can eat leaves of this plant unpunishedly.
The idea about existence of this species of plants was proposed by Anton, the forum member.
Bromeliad
dodder (Baccuscuta bromeliophila)
Order: Solanales (Solanales)
Family: Bindweeds (Convolvulaceae)
Habitat: South America, tropical rainforest canopy.
Picture by Biolog
One of favourable strategies of a survival of plants in tropical
rainforest is parasitism. Parasitic plants are capable to develop life in any
level of the forest, and the high degree of adaptation to parasitism enables
them to grow at any level of the forest, even in the shadiest underbrush, due
to more successful host plants. One species of such plants lives in tropical
rainforests of South America of Neocene epoch – a large dodder parasitizing
on trees in forest canopy. Like all related species, it is a nonchlorophyllic
plant lacking leaves and having long grassy threadlike stalks. Perennial stalk
reaches length of 3-4 meters; it branches and plentifully forms haustoria at
contact with a host plant of suitable species. Colouring of shoots is light
orange, light pink on tips of shoots.
Inflorescences represent clusters of numerous tiny florets on a thin long stalk.
They rise from masses of shoots vertically and rock in light wind, attracting
pollinator insects. “Flower stalk” actually represents short lateral shoot –
its tip is seen from a top of an inflorescence, stopping its growth at this
stage. Tiny florets are pentapetalous and actinomorphic. Plant is not specialized
to certain pollinators, and its pollen may be carried by small flies, beetles,
wasps and moths. As against related species of human epoch, fruits of this species
represent indehiscent capsules. Their walls are juicy and fleshy, in process
of ripening they grow together. They get bright red color and sweetish taste,
imitating berries – hence the name of genus meaning “berry dodder”. Seeds are
buried in pulp of the infructescence. It is an adaptation for life in conditions
of strict competition in tropical forest. Seeds of plant are dispersed by birds
and mammal, and also are eaten by small iguanid lizards. Due to zoochory the
probability of delivery of seeds to the potential host plant increases. They
keep germination capacity for very long time and sprout in different time.
This plant sprouts more often in epiphytic “gardens”, and for the first time
develops on the epiphyte – usually it is a plant of bromeliad or orchid family.
Shoots stretch out of borders of epiphytic thickets, reach bark of tree and
penetrate it with their haustoria. Bromeliad dodder usually parasitizes on trees,
using epiphytic plants as intermediate hosts, but in case of growth on an improper
tree species it can pass the whole life cycle on epiphyte.
Tree-strangling
dendrovolvulus (Dendrovolvulus arbrosphinga)
Order: Solanales (Solanales)
Family: Bindweeds (Convolvulaceae)
Habitat: islands of Caribbean Sea, tropics of Central and North America, forests.
Restoration of biological diversity of tropical forests in early Neocene was
accompanied by numerous attempts of plants from various families to occupy new
ecological niches and to form new life forms. In New World, presence of hummingbirds
promoted active evolution of ornithophilic plants, which have got an advantage
due to use reliable pollinators from among these birds. One example of successful
evolution of ornithophilic plants in tropics of New World is a large liane,
tree-strangling dendrovolvulus.
This plant is a large perennial liane with strongly ramified lignifying stalk.
Length of stalk of adult plant can reach 20 meters at thickness of up to 5 cm
in the basis. This species is a descendant of a grassy plant Calystegia (false
bindweed, morning glory); the name occurs from the invalid name of the ancestral
genus – Volvulus. Stem is climbing, at early stages of growth it creeps on wood
litter and does not lignify. Having found a support, plant quickly twists it
and begins intensive growth in height. The bottom part of stalk lignifies with
age; wood is elastic and durable, grey bark peels off in small scales. Lignifying
stalk prevents a thickening of a trunk of support plant and can literally strangle
twisted trunks of trees at their bases, thus enabling a plant to compete to
them for light. Dendrovolvulus especially frequently strangles young and intensively
growing trees. This plant has inherited from its ancestor a repent underground
rhizome, which grows to the next trees and grows new runners from the ground.
On rhizome there are numerous dormant buds sprouting at its damage.
Leaves of this liane are petiolar and have spear-shaped outlines. Lateral blades
in the basis of a leaf are much smaller than middle one and are deflexed in
sides. At a mature plant on large leaves they are separated from the main lobe
of a leaf by narrowings and can be strongly curved in sides, because of what
leaf looks like trilobate one.
Flower of this species is characteristic for bindweed family: it has narrow
tube and wide limb of five merged petals and almost lacks smell. Perianth has
red color, sometimes with white spots in fauces of flower. Flowers develop on
young shoots one by one in leaf axils, grow on flowerstalks horizontally or
are slightly pendent. Blossoming of each separate flower lasts for 3-4 days;
they close for night and before a rain. In the daytime flowers are widely open
and produce liquid nectar plentifully; they are pollinated by hummingbirds and
occasionally by large hawkmoths. Some wasps gnaw through a tube of a flower
from the side and lick nectar through an aperture, not making pollination.
The adult plant of this species is very photophilous, at shading it blossoms
bad or does not begin to blossom at all.
Fruits of dendrovolvulus represent ligneous capsules dehisceing with a longitudinal
crack and scattering seeds forcefully. Due to hard teguments seeds easily endure
stay in sea water within several days. This feature has enabled the present
species to settle widely at the islands of Caribbean Sea and in tropics of Central
and North America. Sprouts of this species may be found at the coast of North
America in zone of temperate climate, but young plants perish in the winter.
Seeds keep germinating capacity for 2-3 years; young plant begins to blossom
for the first time at the third year of life. Life expectancy of plant is about
30-40 years.
New
Azora banana plantain, “Azorean banana” (Neomusa plantagoidea)
Order: Plantaginales (Plantaginales)
Family: Plantains (Plantaginaceae)
Habitat: New Azora, mountain areas of island.
The flora of islands in human epoch differed in presence of set of endemic species
of grass plants which had turned from small species to giants of plant kingdom.
At Hawaii treelike geraniums and lobelias lived, and at Saint Helena’s Island
huge species of plantain Plantago robusta dwelt earlier. This plant had even
received the name “St. Helena’s banana”. But the destiny of these species was
not surviving after human extinction. Many species, including giant plantains,
had died out, despite of human efforts of their preservation. In Neocene flora
of islands new species of giant plantains had appeared, but they are not related
to the similar species known in human epoch.
In mountain areas of New Azora one of such species of plants grows. This plant
is a little similar to banana, but differs from it in rounded leaves with arc
nervation. Because of similarity to banana the plant has received the name “Azorean
banana”, and is officially named New Azorean banana plantain. However at the
closer look at this plant the set of differences from the prototype of human
epoch is appreciable. While the “trunk” of a banana was formed by petioles of
leaves, at New Azora banana plantain there is true trunk – thick rhizome. It
grows vertically, and can thicken and branch till the growth. It is covered
with fibrous rests of leaves and rough bark protecting from damages. Under the
ground surface thin rhizomes of other type stretch: they can bend upwards at
any distance from parental plant and form new vertical stalks. Trunks of this
plant are fibrous, elastic and rigid. The height of New Azorean banana plantain
usually does not exceed 1 meter, but in remote mountain areas giants grow –
long-livers about 2 meters high.
Tops of rhizome stalks of this plant are topped with crowns of wide oval leaves
of dark green color on thick petioles. Leaves are downed with short rich hairs
of grey color. Young leaves have especially rich downiness, that’s why the middle
area of tuft looks silvery. The length of completely formed leaves of adult
plant reaches one meter.
Approximately since the fifth year of life the stalk starts to form inflorescences.
The inflorescence is formed in apex. It is the spadix characteristic for plantains
covered with numerous small floscules. The height of inflorescence may reach
one meter at the same length of peduncle. This plant is pollinated by insects;
especially frequently on flowers of New Azorean banana plantain it is possible
to meet beetles and various wasps. Small birds, and occasionally even bats fly
to these inflorescences to feed on nectar, but they only incidentally participate
in pollination of “Azorean banana”.
Fructification of New Azorean banana plantain is very interesting: when flowers
are pollinated, the axis of inflorescence starts to grow thick, and the peduncle
bends to the ground. Ovaries are immersed in expanding axis of inflorescence
and become covered by dense environment. The pulp of inflorescence axis becomes
friable and sweetish by taste, and thin skin gets bright orange color. Such
inflorescence forms infructescence similar a little bit to pineapple one by
the principle of formation. Seeds of this plant local gallinaceous birds – rock
Azorean partridges – distribute. Birds peck infructescenses falling to the
ground. Seeds, passing through the stomach of bird, not only do not perish,
but even raise germinating ability: because of action of gastric juice their
environment grows thin, that helps the germ to receive water and the mineral
salts necessary for germination and development. The dung of birds gives the
additional top dressing necessary for growth. But reproduction of this species
is braked by small birds, which frequently peck off unripe ovaries from growing
infructescenses.
After fructification growth of stalk stops for some time. When infructescence
starts to ripen, near the basis of peduncle the new apex is formed, and strong
stalks occasionally start to branch.
“Azorean banana” grows slowly: large plant can reach the age of 120 – 130 years,
and the general life expectancy reaches 200 years and more.
Common
boatflower (Hydroplantago vulgaris)
Order: Plantaginales (Plantaginales)
Family: Plantains (Plantaginaceae)
Habitat: rivers, lakes and brackish waters of Europe and Asia Minor.

Picture by Nem
In human epoch various plants of plantain family began to master aquatic habitats.
After the ending of human epoch, evolution of Plantaginaceae in this direction
has continued, and in Neocene freshwater and brackish reservoirs of Europe are
occupied by various descendants of hydrophilic plantains belonging to Hydroplantago
genus. These plants have a number of shared characteristic features: all of
them are rooting or floating forms having a surface inflorescence representing
unilateral spirally twirled spica. Flowers are tiny, lack petals and have sticky
stigmas. One leaf in the basis of an inflorescence is boat-shaped – hence the
common name of genus. It functions as a wrapper of an inflorescence, helping
it to keep on water. Surface of bract leaf is covered rich thin trichomes and
is water-repellent. Other leaves, as a rule, are underwater, narrow and growing
on bipalmate shoots.
In reservoirs of Central and Southern Europe, and also in nearby areas of Asia
Minor common boatflower – one of the most widespread species of the genus –
lives. This species has two types of shoots: rooting and sarmentous ones. Rooting
shoots are rosette-like and have reduced scale-like leaves and numerous buds,
in which axils elongated sarment-like shoots with long internodes and narrow
lanceolar leaves of brigh green color develop. This species inhabits shallow
sites of rivers and lakes – well lightened and warmed up by sun. Growth takes
place at once both at the bottom, and in thickness of water, therefore significant
areas of coastal waters appear filled with thickets of this species. Propagation
is mainly vegetative.
Pollination of boatflower takes place by means of wind. In the summer on the
surface of water the inflorescences appear, being closed by rolled bracts. During
the maturing of flowers, bract turns away and down from an inflorescence, laying
on the surface of water like a boat, and inflorescence rises above the surface,
where its pollination takes place. Seeds are small and have firm porous tegument;
they sprout not simultaneously, frequently having a delay of up to 2-3 years.
The least specialized kind living in the rivers of the south of Europe (to the
south from the Alpes) is simple
boatflower (Hydroplantago primitiva). It is species rooting at the
bottom, considerably different in its form of growth: it has rosettes with short
vegetative shoots growing from leaf axils and underground runners giving rise
to new rosettes. Only flower stalks with bract leaves reach water surface. Populations
of this species are almost fully isolated from each other.
In brackish reservoirs along the coast of Atlantic Ocean small floating species
lives – pondweed-like
boatflower (Hydroplantago potamogetonis), an inhabitant of well illuminated
sites of reservoir. It lacks rosette shoots, and leaves are very small, covered
with water-repellent trichomes and densely covering its stalks. Due to intense
growth this species forms on the surface of waveless reservoirs floating islets
capable to hold weight of small bird or rat. Inflorescences are short and have
small bracts, which do not lay on the surface of water.
Floating
boatflower (Hydroplantago natans) from the rivers of a southeast of
Europe also has completely passed to aquatic way of life and has lost connection
to substratum. This plant is remarkable in having a short stalk with air-bearing
cavities, and its leaves are not numerous, but large, with peaked tips. Nerves
are thickened, in them air-bearing parenchyma serving as a float is well-advanced.
In nodes numerous roots develop. This plant forms a continuous cover of leaves
on water surface. Bract is very large (3-4 times exceeding a usual leaf), boat-shaped,
having strongly advanced nerves and serving as a support for horizontal inflorescences.
This species of plants was discovered by Nem, the forum member.
Rain-loving
dogwood (Cornus pluviphilus)
Order: Cornales (Cornales)
Family: Dogwoods (Cornaceae)
Habitat: humid Pacific forests of North America.
In Neocene epoch at Pacific coast of North America the special type of plant
community was formed – a rainforest of a temperate climate. From ocean rain
clouds constantly come, spilling rains over rather narrow strip of land between
ocean and Rocky Mountains. Flora of this place is presented by moistureloving
plants having adaptations for evaporation of surplus of moisture. One prominent
feature of local plants is width of leaves. Even local
thuja species has very wide needles.
In shady underbrush of such forests mosses and ferns expand, and one of characteristic
species of bushes is rain-loving dogwood. This is rather large plant with a
sprawling crone, 3-4 m tall and up to 5 m in diameter. This plant usually forms
one main trunk, though rather often from its basis the lateral branches grow,
which are almost equal to it in size. Despite of the size, rain-loving dogwood
externally resembles a bush more, and forms extensive thickets. This plant very
rarely grows as monocormic tree. Its bark has very characteristic colouring.
Young shoots are covered with red smooth bark, which in due course darkens;
on trunk bark is dark brown with a pattern of small cracks.
Leaves of this plant are large, dark green, elliptic with extended tip forming
a “dropper” more characteristic for plants of tropical rainforests. In this
place at plant hydatodes are located, removing a surplus of moisture as drops
of clean water. Young growing leaves are pinkish; with age their colouring changes
to green, and pinkish color is kept on nerves, but disappears later. Autumn
foliage is dark red; it is a deciduous plant. In winter colouring of young branches
becomes especially well-appreciable.
Dogwood blossoms in the beginning of summer and it is pollinated by animals;
hummingbirds and hawk moths frequently visit it. At its Holocene ancestor (most
likely, Cornus nuttallii) flowers were rather small whereas at rain-loving dogwood
they look like large white stars, which are well-visible against the background
of dark green leaves of this tree. Capitate inflorescence of small flowers is
surrounded by 5-6 large bracts of white or slightly pinkish color. Wide and
flat inflorescences of dogwood are convenient for beetles, which land on them,
feed on nectar and a part of pollen, and transport its pollen from one flower
to another.
Fruits of rain-loving dogwood represent large white berries flattened at top
and at pedicel. At ripening they are very well visible on the background of
autumn foliage of plant. They are not so nutritious, and their pulp is rather
watery. But a local jay species feeds on them; the pulp is digested, and seeds
are dispersed with bird dung. Also herbivorous mammals eat these berries. The
young plant begins to blossom for the first time at the third-fifth year of
life. Rain-loving dogwood can live for some tens years.
This species of plants was discovered by Bhut, the forum member.
North
American oak ghost (Monotropa querciphila)
Order: Ericales (Ericales)
Family: Heathers (Ericaceae), subfamily: Monotropes (Monotropoidae)
Habitat: oak forests of the eastern part of North America.
An ancestor of this plant was false beech-drops (Monotropa hypopitys) – very
unusual plant, the parasite of mycelium of various fungi, lost its chlorophyll
and looking as if it is chemically decoloured. Despite lacking chlorophyll,
false beech-drops appeared rather hardy plant – shade-loving and capable to
survive until the mycelium feeding it and trees, with which mycelium penetrating
this plant entered symbiosis, were alive.
In Neocene the union with mushrooms was useful to a plant: this ancestor has
evolved to some new species, in part due to restoration of habitats after human
disappearance. Evolution of these plants has gone the way of specialization
to parasitism. One species of this genus – North American oak ghost – parasitizes
on various oak species growing in eastern forests of North American continent.
This is a medium-sized plant, about 10 cm tall. Like its ancestral species,
it lacks chlorophyll and its tissues have pale yellow color. This species depends
on fungi only in the early period of life, and in due course of development
it penetrates into roots of trees of suitable species, more often oaks (hence
the name) and continues to develop due to these plants. Its roots penetrate
deep into wood of various oak species and if the oak is too young or old, oak
ghost can sometimes kill the whole tree.
An only exception is Eastern mountain oak (Quercus appalchiensis). In due course
of evolution this tree species has developed an original immunity to parasitism
of oak ghost: it produces substances inhibiting growth and development of oak
ghost, though not killing it. Thus, this plant can live on the aforementioned
oak species in a kind of “suppressed” condition, not threatening life of host
tree.
Oak ghost breeds by means of tiny seeds, which are dispersed even by slight
whiffs of wind. Seeds get on surface of ground and sprout only at penetrating
of mycelium of soil fungi inside them. Getting stronger, plant forms a short
underground rhizome and grows in all sides thin roots, which find roots of host
trees and sprout into them; if it is done successfully, plant quickly develops
and forms numerous adventives buds in the place of penetration into the host’s
root. After a certain time, flowers are formed – usually at the end of spring
and in the beginning of summer. Their smell is sourish and spicy, and they are
pollinated by various insects of underbrush, more often by beetles or solitary
bees. To winter fruits have time to ripen and to disperse seeds and then epigeal
portion of plant dies off, and plant winters as rhizomes or chords of tissues
under bark of host trees. This plant is a food of various species of hares and
rabbits, and also foe forest-dwelling peccary species.
North American oak ghost is remarkable in its longevity and frequently lives
as long, as its host tree.
This species of plants was discovered by Bhut, the forum member.
Mountain
mayflower (Alloepigaea montana)
Order: Ericales (Ericales)
Family: Heathers (Ericaceae)
Habitat: mountain forests in the eastern part of North America.
Mountain forests in the east, and partly in northeast of North America differ
a little from mountain forests of the west: they are drier than forests of Pacific
coast because of features of windrose, and slopes of mountains are flatter,
therefore fauna and flora are more homogeneous, without strict distinctions
seen on eastern and western slopes, as it takes place in Rocky Mountains. In
general, flora of Appalachian Mountains is present with broadleaf trees (conifers
are rarely found), mainly various species of oaks (conifers are more diverse)
and shade-loving grassy plants prospering in these woods. Some
of them have turned to parasites living without chlorophyll, but only few
species are those. Among plants of more traditional way of life, the typical
inhabitant of forests of Appalachian Mountains is mountain mayflower, the descendant
of North American mayflower, or trailing arbutus (Epigaea repens). However,
this species has a hybrid origin – a certain part of genes is inherited from
the Asian forms and their garden hybrids cultivated in human epoch.
Mountain mayflower sprawls on the ground, giving numerous lignifying shoots,
which easily take roots in wood litter. Ancestral mayflower species was an evergreen
plant, and mountain mayflower has kept its features: it is resistant to cold
and successfully winters under snow. However, the way of life of this species
has appreciably changed in comparison with ancestors – this plant is in the
way to adaptation to parasitic way of life. In winter it sheds old leaves, keeping
them only on tips of shoots, and exists as a semiparasite, absorbing nutrients
from roots of trees to which it has stuck by its roots – usually these are various
oak species. One species of trees, Eastern
mountain oak, contains in its sap substances, that kill parasitic plants
and inhibit their growth, and other tree species can die, especially young specimens.
In the spring, as soon as snow thaws, mayflower grows new leaves and passes
to an independent feeding by means of photosynthesis. Its leaves are disproportionately
wide, of rounded shape and dark. They absorb a lot of sunlight, especially when
on trees the foliage only begins to appear. During this time there is a peak
of parasitism from the part of a plant, which actively produces greenery and
supersedes its competitors in underbrush. In early spring mayflower already
grows its own flowers – small, pale whitish-pink star-shaped ones having five
petals and darker middle, which will blossom until the beginning of summer,
when other mountain flowers will appear and grassy plants will grow up. By then
various insects already pollinate mountain mayflower, and instead of flowers,
berries appear on it; they are vaguely similar to wild strawberry in their shape.
These berries are eaten by various herbivorous and omnivorous mammals and birds,
but seeds of mountain mayflower are resistant to their gastric juice. Having
passed through digestive system of an animal, they only sprout better.
Mountain mayflower cap propagate successfully by repent shoots and roots, and
even by shoots of roots, if the part of root system will be lost, being dug
out from the ground by local herbivores. On the roots torn off from a plant
the adventive buds appear, forming functional shoots. The plant is capable to
exist independently for a long time, not parasitizing on another plant’s roots,
but thus it blossoms less abundantly. At the presence of suitable species of
host trees, mountain mayflower easily passes to parasitic way of life.
This species is a perennial bush capable to live up to 40-50 years.
This species of plants was discovered by Bhut, the forum member.
Awl-leaved,
or Antarctic crowberry (Empetrum subulatum)
Order: Ericales (Ericales)
Family: Heathers (Ericaceae)
Habitat: Antarctica, wetlands and thickets of mosses at the basis of glaciers,
mountain areas near the snow line.
During one of Pleistocene congelations crowberry from North America managed
to cross equatorial areas of the Earth and to settle in the Southern hemisphere.
In the far south of South America the plant has formed the viable populations,
which have turned to valid separated species in early Neocene, and after regression
of glaciers in Antarctica crowberry from Tierra del Fuego appeared one of colonists
of free from glaciers coast of this continent. Due to its unpretentiousness
it had quickly settled across the grounds free of a glacial cover, and in Neocene
it is a widespread species in Antarctica.
Awl-leaved crowberry is a perennial plant with creeping lignifying stalks and
red berries, which at ripening attract birds with their colouring. Berries have
a thin layer of wax reflecting ultra-violet rays – it helps berries to be visible
for birds from apart. This species is an evergreen plant, which has very narrow
leaves with edges turned inward, that resemble conifer needles, up to 20 mm
long. A specific appearance of leaves is connected to necessity of saving moisture
at life in cold bogs. Thickets of this plant resemble an undersized grass and
differ from it in modestly-looking flowers, appearing at the ground, which are
replaced in summer by bright berries. Pulp of berries is sweet, but very watery.
Shoots of this plant reach length of up to 100 cm. One plant forms some large
branchy shoots and occupies an area of up to 3 square meters. Shoots laying
on a substratum easily take roots, anchoring in the smallest roughnesses of
substratum. Short roots are covered with tubercles – inside them nitrogen-fixating
bacteria settle, helping this plant to get nitrogen necessary for growth. Also
on roots of a plant soil fungi settle, supplying it with nutrients. Due to such
allies this plant grows successfully even on the most nutrient-poor and barren
ground and is an important species of pioneer vegetation, promoting soil formation.
At the same time for awl-leawed crowberry low competitiveness is characteristic
and in mixed thickets it is easily superseded by other kinds of plants.
Slow growth of this plant is compensated by significant longevity – plant is
capable to live up to 50 years and more. It frequently happens, that at dying
off of the basic root and the death of the central shoot lateral branches continue
their growt, forming a ring-like clone.
Qijuktaat
(Cassiope qijuktaat)
Order: Ericales (Ericales)
Family: Heathers (Ericaceae)
Habitat: North American tundra.
In the summer North American Arctic regions of Neocene epoch represents rather
noisy place. Various species of geese and ducks, and also auks form unnumerable
rookeries, which attract various predatory mammals and birds trying to hunt
them. Numerous birds hasten to brood posterity, getting all advantages of polar
summer and long polar day. Life of inhabitants of tundra is humming on the background
of local vegetation, which has evolved, reacting to glaciers advance and thawing,
and to changes of borders of natural zones accompanying it. Like in human epoch,
in Neocene in tundras undersized grassy and suffruticose vegetation dominates.
Because of small density of human populations in tundra, the plants lived in
this natural zone have suffered the least damage from human activity, and in
Neocene in tundras of Northern hemisphere their descendants grow. One of such
species is qijuktaat, a descendant of Arctic bell-heather (Cassiope tetragona)
of Holocene epoch: “qijuktaat” is a name of bell-heather at Eskimos. This is
rather tall plant on a scale of tundra vegetation – about 25-30 cm in height.
It is a bush with creeping trunks and large branches. Spreaded on the ground,
the bush forms set of vertical photosynthesizing shoots. Young branches are
covered with thick scale-like leaves of dark green color. On the old branches
laying on the ground leaves dry up, but do not disappear and form a protective
cover. Tips of shoots with a young increment are reddish, and tops of shoots
are coloured crimson-red, therefore thickets of these plants in a phase of active
growth have an original look.
Among leaves vertical flower stalks sprout, rising above tips of shoots flowers
of bell-like shape (like at ancestral species) colored light pink; the inner
side of petals, pistil and stamens have brighter shade. White-flowered morph
of this species is rather common. Qijuktaat is an important nectar-bearer and
a fodder plant for numerous insects of North American tundra; even males of
blood-sucking insects willingly feed in these flowers. Vertebrate animals like
wild geese willingly eat branches with young increment and scale leaves of this
heather plant in particular. Grazing birds literally “shave off” leaves, dragging
shoots through slightly opened beaks. Herbivorous mammals bite off photosynthesizing
shoots almost near the ground.
Qijuktaat is hardy plant in conditions of polar winter and cold. Its sap contains
resinous substances acting like antifreeze; their concentration increases to
winter, making this plant less edible, than in summer. However it does not stop
skewhorns, which dig qijuktaat by the root and
eat the bottom parts of a plant, where concentration of these substances is
lowered. Thus for winter they cause great damage to a plant, but this plant
is very hardy: the shoots not eaten by them can take roots in the spring, and
the rests of stalks become overgrown with new green shoots very soon. By the
end of a polar summer, a plant frequently restores former abundance, though
during the recovery it blossoms less actively. Qijuktaat successfully breeds
by seeds, and repent branches easily take roots, that helps it to survive at
the presence of large herbivores.
Young plant begins to blossom for the first time at the third year of life,
and the maximal age of qijuktaat may be up to 20-25 years and more.
This species of plants was discovered by Bhut, the forum member.
Kinnikinnik
(Arctostaphylos kinnikinnick)
Order: Ericales (Ericales)
Family: Heathers (Ericaceae)
Habitat: North American forest-tundra and tundra.
North American tundra in Neocene is a place rich in life, especially in polar
summer, when from wintering places numerous birds return, and plants try to
take a benefit from each day of short and cool northern summer. On dry watersheds,
there are thickets of Nearctic pine, in other
places green waves of qijuktaat thickets
decorated with pinkish foam of tiny flowers swing in the wind. Among stones
there are tiny plants – polar aupillatinguat
rockfoil grows or berries of kinnikinnik redden.
Kinnikinnik is a descendant of a bearberry (Arctostaphylos uva-ursi) of Holocene
epoch. This is rather tall plant – creeping spreading evergreen bush about 30
cm tall, frequently much lower. At the northern border of the range this plant
only slightly rises above the ground. Shoots easily take roots; root system
is shallow and filaceous. Leaves of this plant are rounded, leathery and dark
green from above, bright green and rough from below. Along the leaf edge dense
fringe of white hairs stretches. Flowers of this plant are vaguely similar to
qijuktaat flowers: they are also bellflower-like, but smaller and narrower and
have an appreciable rose-red shade inside: pollinator insects precisely differ
these two species of flowers and do not mix they – some of them pollinate qijuktaat,
others prefer kinnikinnik more, however any strict specialization of pollinators
is absent. Flowering begins early in the spring, soon after snow thawing. Flowers
easily endure freezings.
Like its ancestral species, kinnikinnik is a berry-bearing plant: during fructification,
plants may be entirely covered with bright red berries having dryish starchy,
but sweet pulp. This is one of few berry plants in North American tundra, therefore
at the end of the summer many mammals, and even some birds like traveller
goose eat its berries willingly and in great amounts – and not only such
herbivores like skewhorns do it, but also geopossums
– omnivorous ones having a bias to predatoriness. And these marsupials eat not
only berries, but also greenery of this plant: it contains the substances strengthening
health of these marsupials. As a result, animals of various species quite often
eat away thickets of kinnikinnik completely, but seeds of this plant past through
digestive system of mammal sprout faster, than without treatment by its gastric
juice. In addition, seeds and young plants of kinnikinnik endure a winter cold
easier, than adult specimens freezing in severe winters with a small amount
of snow. Compared to other polar plants, kinnikinnik grows faster and endures
easier the damages put by herbivorous animals. A young plant fructifies already
at the third year of life, and a peak of fructification takes place at the tenth
year. Life expectancy frequently reaches 50-60 years.
This species of plants was discovered by Bhut, the forum member.
North
American bellflower (Campanula popularis)
Order: Asterales (Asterales)
Family: Bellflowers (Campanulaceae)
Habitat: North American prairies, meadows, woods and forest-tundra in the zone
of temperate and cold climate.
Many species of bellflowers look fragile and vulnerable plants, but this impression
is deceptive: bellflowers are actually very hardy plants easily forming various
life forms. Evolutionary flexibility and human horticulture have helped them
to survive during the time of anthropogenous pressure of Holocene and an ice
age of the beginning of Neocene and to continue to prosper and evolve.
North American bellflower is one of the most widespread species of this family
in North America. It grows everywhere to the east from Rocky Mountains – both
at open spaces, and in rather shady, but dry broadleaf and mixed forests of
the east of the continent, and in colder forests of the north, and even in forest-tundra.
This species forms shoots of two types. In the summer during flowering, this
plant forms tall upright shoots typical for bellflowers, with elastic thin stalks,
numerous blue flowers of bell-like shape and long and narrow leaves. In the
spring the plant forms a rosette of wide rounded leaves absorbing sunlight better
and enabling North American bellflower to grow faster than its competitors.
To the middle of summer this plant grows long stalks with leaves of “summer”
shape and flowers, which are visited willingly by various insects – this plant
is not specialized to any certain kinds of pollinators. Large animals eat these
flowers, sometimes pulling out from the ground and damaging their roots. Underground
part of plant represents short perennial rhizome, from which vertical shoots
grow. This species does not form underground sarments and grows as compact clumps.
North American bellflower settles to large distances by means of seeds. Seeds
are hidden in dehiscent capsules, which “blow up” when ripen, scattering seeds
some meters far in sides. As a result of it, the present plant species grows
locally in numerous groups at rather small area. In an autumn stalks die off
up to a ground level and under snow only rhizomes with roots winter to sprout
in the spring after snow thawing. In northern part of range plants winter with
the radical rosette of leaves.
Rodents and grubs frequently eat rhizomes of this plant. If the main rhizome
will avoid being eaten, North American bellflower can live up to 8-10 years
and even more.
This species of plants was discovered by Bhut, the forum member.
Cobold
bellflower (Campanula coboldiana)
Order: Asterales (Asterales)
Family: Bellflowers (Campanulaceae)
Habitat: the Alpes, stony areas of meadows and a mountain-tundra zone at snow
line.
In human epoch the Alpes were one of the centres of a species diversity of bellflowers
in Europe – here a plenty of rare local and endemic species lived. In epoch
of human domination the significant number of species of these plants has died
out because of destruction and pollution of habitats, climate change and destruction
of plants. Local species sensitive to changes of an inhabitancy appeared especially
vulnerable. In Neocene epoch processes of mountain building in the Alpes continued
because of slow collision of Europe and Africa, and it has resulted in appearing
of new habitats, which had been occupied by specialized endemic species evolved
already after people.
In the zone of mountain meadows and tundras a little bit below the snow line
a specialized high-mountainous bellflower species lives – cobold bellflower.
It is an undersized grassy plant forming a rich cushion of short shoots. Height
of shoots does not exceed 5 cm, but they branch intensively at the basis and
adjoin to each other densely. Leaves are rounded, alternate, growing on stalk
densely, forming rich rosette shoots. Young leaves and apical points are plentifully
pubescent for protection of growing tissues against ultra-violet radiation;
on mature leaves downiness is kept only on their bottom sides, preventing losses
of moisture. This species is a perennial evergreen plant growing on stony soil.
Short underground rhizome is hidden in the ground at the depth of 1-2 cm and
frequently sprouts between stones.
During the flowering this plant becomes only a little bit taller: small solitary
flowers of deep mazarine colouring opened one by one on thin shoots 6-8 cm tall.
They are not specialized to any specific type of pollinator; on them flies,
small beetles and butterflies feed. This plant is the important fodder resource
for insects: cobold bellflower begins to blossom right after snow thawing and
its flowering goes on up to the middle of summer.
Seeds ripen the next year early in the spring and are dispersed by streams of
thawed snow water. Fruit is an indehiscent capsule serving as a float and containing
3-4 seeds. Fruit shell gradually absorbs water, due to which seeds swell and
sprout. After germination the strongest sprout suffocates others and develops
in an adult plant. The first flowering takes place at the third year of life;
life expectancy reaches 20 years.
Tree-climbing
stenocampana (Stenocampana scandens)
Order: Asterales (Asterales)
Family: Bellflowers (Campanulaceae)
Habitat: Southern Asia, tropical forests.
Epoch of restoration of forests in early Neocene is a time of active speciation
and an intensive competition between the plants forming natural community. At
this time plants of various families tried to occupy new ecological niches and
as a result of it among them vital forms unknown in human epoch appeared among
them. Among such plants there were, for example, begonias,
grapes and daisies
turned to treelike forms. Bellflower family, being the most diverse in flora
of Old World, was presented mainly by grassy plants. But in epoch of restoration
of forests they also managed to enter large-sized class – but they became not
trees, but lianes. One of the largest species of lianes in this family is tree-climbing
stenocampana, which represents partly lignifying fast-growing liane. It inhabits
areas of the damaged forest stand and quickly expands, giving an amount of growth
up to 10 meters per one year. The stalk of an adult plant reaches thickness
of 10-15 cm; length of stalks of an adult plant is up to 40 meters. At this
plant, bottom part of stalk lignifies, but wood is soft and fragile. Bark is
smooth, exfoliating longitudinal cracks and peels from stalk in strips.
Stenocampana twines trees and bushes, attaching to them with short adventitious
roots. Stalk branches mainly in its top part, near to a crone of tree being
a support for a plant. Leaves are large, simple, alternate, having long elastic
petioles. Leaf blade grows to 20-25 cm long and 15 cm wide and has oval outlines.
Leaf tip it extended to a narrow dropper 2-3 cm long. Surface of leaf is smooth,
bright green with thin plant wax cover. In dry season a part of foliage from
the bottom part of stalk falls off; leaves remain only on young shoots.
At the end of dry season plant begins to blossom. On old increment axillary
buds sprout, from which short shoots develop, forming single flowers in leaf
axils. This species has original long and narrow tubular flowers with small
limb – hence the name meaning “narrow bell”. Length of a flower reaches 12-14
cm, its elastic pedicel grows up to 10 cm. Calyx is blue-colored with marks
in fauces reflecting ultra-violet part of spectrum and showing a way to nectar
to pollinator insects. Flowers are pendulous or directed almost horizontally,
rocking on their pedicels. This plant species is pollinated by hawk moths and
other large lepidopterans having long proboscii.
Fruit is an indehiscent berry with dryish sweet pulp and thin skin of red color.
Usual distributors of fruits are arboreal and ground-dwelling birds. Germination
of seeds occurs much more successfully after their passage through a digestive
path of a bird. Young plant begins to blossom at the second year of life and
reaches the maximal size at the age of 10 years. Life expectancy is over 50
years.
Spiniest
astreca (Astreca spinosissima)
Order: Asterales
Family: Asteraceae
Habitat: Northern Africa, border of Mediterranean swamps.
Drying in late Holocene Mediterranean sea has essentially affected climate
of districts surrounded it earlier. The territory of former Mediterranean began
more droughty, and wind from Mediterranean hollow carry clouds of salt dust
sedimenting far in depth of continents. In such conditions not all plants can
normally exist, but some species adapted to such conditions prosper.
At the former African coast of Mediterranean sea among sickly grass groups
of interesting plants about 2 meters height grow here and there. At first sight
they are similar to small palm with prickly trunk, but, at more close look,
it becomes evident, that it is not a relative of palm at all
This fantastical plant is spiniest astreca, one of inhabitants of droughty
districts surrounding Mediterranean lowland. The name “astreca” is made of
words “aster”, a plant showing belonging to family Asteraceae, and “areca”
that means “palm”.
The thick succulent stalk of astreca has five longitudinal sharp edges, and
its cross cut is similar to star. Each edge is plentifully bristled up by sharp
strong spikes reliably preserving plant against large herbivores. Stalk poorly
branches, giving from the basis two - three large lateral sprouts. Trunks of
astreca are covered with dense thin skin partly lignifying in the bottom part.
But spikes on trunk are not the main feature of astreca giving to it “palm-like”
shape. Stalks of this plant are topped with crown of dark green pinnate leaves
about one meter long. Along the middle nerve of leaves light silvery strip
passes. Under alive leaves the rests of dried up last year's leaves hang down
like fringe – though they are dead, they still play the vital role, protecting
stalk from sun burns. Each leaf from above and from below along main and large
lateral nerves has lines of sharp thin spikes – it is the reliable weapon against
teeth of gluttonous herbivores. Died out leaves do not fall down also because
their spikes so regularly take part in protection of plant.
On the bottom surface of leaves silver-white hairs richly grow, giving to wrong
side characteristic felt-like structure. Such adaptation helps to evaporate
water in less quantity: stomas are on the bottom side of leaves, and hairs
soften contrast between medium inside leaves and drying up heat outside.
Plants of family Asteraceae are champions of vegetative multiplying. At meter
depth under ground astreca forms an extensive circuit of rhizomes on which
offsprings are formed. Therefore one plant in suitable conditions quickly expands
to original thin grooves.
During short rain season astreca grows quickly. For some months the plant increases
up to 20 cm in height, and forms about two tens large leaves. In conditions
of plentiful humidifying on old 7 - 10-years trunks peduncles develop. Inflorescences
of astreca are small yellow heads of hermaphrodite flowers, surrounded with
dry reddish-brown leaves of inflorescence covers. Heads are gathered in large
common corymb-like inflorescence on long peduncle. It develops on top of old
trunk of plant and is visible from apart. Solitary wasps and butterflies pollinate
astreca basically, though occasionally it is possible to see beetles and flies
on inflorescences of plant.
To the beginning of dry season seeds of astreca ripen. After their ripening
the stalk of plant given a peduncle, dies off. But near it the young growth,
which soon will get stronger, already grows up.
In desert there is not so much animals may become seed carriers, therefore
astreca had selected simple vital strategy, characteristic for set of plant
species: its seeds are carrying by wind. Tiny seeds of astreca have white parachutes
with which help they are carried at the big distance. In such way astreca was
settled on all coast of Mediterranean swamps, and even grows on some mountains
of Mediterranean lowland, having formed some well differing species.
Seeds of astreca keep germinating ability for a long time, waiting for favorable
conditions till some years. After rain they sprout quickly, and young sprout
starts to form the crown of leaves on ground surface, and its long root reaches
to damp deep layers of ground. For the second year of life the elevated trunk
starts to form, reaching full development to 10-years age. At the third year
of life plant starts to form horizontal rhizomes from which new offsprings
grow in various sides.
In Northern Africa, on islands of Mediterranean swamps and in Southern Europe
some related species of this plant live:
Astrophytum-like
astreca (Astreca astrophytoides) is the inhabitant
of dry mountain areas of Mediterranean lowland. It differs
by short thick trunk (height of plant is about half meter)
with large fleshy edges covered with short spikes. The form
of trunk of this species of astreca resembles cactus of genus
Astrophytum. Leaves are narrow, densely downed from below,
covered from above with numerous sharp hairs.
The peduncle is very short, inflorescence consists at all from 3 - 4 heads
of yellow flowers with pink dry leaves of cover.
More often this species meets in cracks of rocks by small groups, avoiding
salt soils.
Palm-like
astreca (A. palmetto) meets in Northwest Africa, at
the Gibraltar isthmus and in Southern Europe. Living in rather
damper conditions, this species is characterized by high trunks
(up to 5 meters height), a plenty of long narrow leaves (length
about 1 meter at width about 10 cm). Leaves are pinnatilobate,
on the tip of each lobe sharp spike sticks up. Trunk is 5 -
6-edged; edges are narrow, slightly wavy, covered with single
long spikes.
The peduncle is long (up to 1 meter); it carries a plenty of small heads with
white cover leaves.
This species meets in separate groups lengthways of riverbeds and near temporary
reservoirs, preferring relatively humidified ground.
Giant desert daisy (Taraxacaulis crassiradix)
Order: Asterales (Asterales)
Family Asteraceae (Asteraceae)
Habitat: semi-arid areas of Mediterranean lowland.
Rigorous conditions of life result in occurrence in animals and plants of the
various adaptations permitting them to survive. In the epoch previous to Neocene,
climate of Earth was rigorous and contrast, having caused to life set of
species of live organisms adapted to extreme conditions. In Neocene the climate
began much softer, and species adapted to rigorous conditions of inhabitancy
have started to die out. But even in Neocene on Earth places with very
rigorous conditions of life were kept. They became original “reserves” and
“ranges” for the plants living in extreme conditions, and in these places evolution
has continued “to perfect” their adaptation. One of such areas is the district
around of former Mediterranean sea which has turned to congestion of salt swamps
and lakes in huge hollow had being earlier its bottom.
In epoch of large geological or climatic cataclysms the most undemanding and
widely widespread species survive with the greater probability. Therefore it
is no wonder, that an ancestor of one of interesting plants of semi-desert
at the edge of Mediterranean bogs was the usual dandelion (Taraxacum officinalis).
However, in human epoch classification of dandelions to species was the extremely
uncertain because of wide variability of these plants. Obviously, the pledge
of success of these plants in Neocene also had been hidden in it.
The descendant of dandelion, the giant desert daisy, lives in droughty areas
of Asia and Northern Africa, preferring stony and clay ground. It represents
stalk-less grass, being somewhat reminiscent of the ancestor. At this plant
leaves are much larger, than at dandelion: about 30 - 40 cm long with thick
main nerve and pinnated edges. On angular lobes of leaves along the edge thin
sharp spikes stick up. The bottom surface of leaves is covered with grayish
felt film of hairs; top is glossy. Young leaflets are richly covered with hairs
even from above, but in process of development of leaves downiness remains
only on the bottom surface. Leaves lay on the ground as wide crown, and last
year's leaves can remain on plant one - two years more, protecting plant from
herbivores, and ground under it from overheat.
On depth of approximately one meter under ground surface the most important
part of plant, the thick tap root forming some kind of “root crop” grows. At
the largest plants of this species it may reach length one and half meters
at thickness more than half meter and weight about 100 kg. It is covered with
thin elastic bark, and from it some lateral roots grow asides. Here there is
a stock of water carefully gathered by plant during rare rains. At this time
on ground surface numerous soaking roots grow, absorbing rainwater and dew.
In drought they wither, but the plant does not endure lack of water: its main
root grow deep into ground down to 20 - 30 meters and more, reaching water-bearing
layers. Even if the animal like an insect or a rodent will manage to burrow
to the “root crop”, it will have not enough benefit from it: sap of plant contains
a plenty of latex, and thickens on air. At top of “root crop” there is sanctuary
of plant – its apex. If it appears damaged, the plant stops growth, but the
next year in the beginning of rain season it will develop two, or even three
apexes developing from sleeping buds.
In the beginning of rain season wrinkled “root crop” gradually absorbs water,
and the plant slightly “rises”, growing up from under the ground a crown of
young leaves. And in dry season roots retract and involve “root crop” deeply
in ground, leaving on surfaces only dying off leaves.
In short rain season giant desert daisy begins to blossom. Peduncles of these
species are short, and yellow inflorescences (heads typical for all family)
almost lay on the ground in middle of crown formed by leaves: so the plant
is less appreciable from afar for herbivorous
mammals. But it involves to itself
not numerous pollinators (mainly beetles and flies) with a specific smell:
a mix of pleasant flower and putrefactive aromas. Flowering lasts not for long:
no more than one week. After pollination seeds quickly ripen, and the peduncle
starts to grow. It bears ripening seeds on meter tall above ground. Seeds of
giant desert daisy are similar to seeds of dandelion: they are small, have
thick environment and characteristic pappus as they are carried by wind. After
fructification the plant keeps leaves not for long, and by the end of summer
at it the dormant period comes: the elevated part dies off, and the plant spends
in rest an autumn, winter and the beginning of spring, remaining green less
than half-year.
This species develops for a long time: the plant begins to blossom for the
first time only at five years' age when its “root crop” reaches weight of 10
- 15 kg.
Giant desert daisy easily distributes by seeds, therefore it frequently grows
in foothills on island “oases” of the Mediterranean swamps. But here large
local herbivores, Antheos tortoises, can eat it, causing the big damage to
slowly growing plant.
Silver
oromatrica (Oromatrica argenteofolia)
Order: Asterales
Family: Asteraceae
Habitat: Himalayas, alpine meadows at southern slopes.
In Neocene Himalayas still keep a title of highest mountains of Earth.
Here, at huge height, still the most part of year the cold and snow reign,
though in lower valleys and on southern slopes it happens very hot in summer.
Vegetation of Himalayas is variable, though severe conditions of life have
made many plants very similar to each other. Sometimes, it is difficult to
distinguish poppy from carnation or bluebell while plants will blossom – their
leaves and stalks are so similar. The majority of grasses of Himalayas has
creeping or strongly short branchy stalks, and leaves forming rich polsters
between stones. Such form of growth protects them from cold winter winds: snow
quickly closes these undersized plants, and they successfully winter, despite
of winter colds.
But among mountain flora there are not only undersized species pressing closely
to the ground. Here and there on slopes inaccessible to herbivorous animals,
the unusual plant, oromatrica (literally “mountain camomile”) grows. It strongly
differs by shape from the majority of Himalayan grasses. At this plant there
is thick upright trunk about half meter tall, covered with dry rests of leaves.
At top of trunk live leaves grow – they are strongly dissected, having silvery
downiness on the bottom part. It is main protection of plant against colds:
to winter leaves turn upwards, closing apex and protecting it from frost. Covered
by snow, this plant is practically indistinguishable from stones. Leaves have
unpleasant smell and bitter taste protecting them from different herbivores.
But, despite of it, large herbivorous animals nevertheless eat these plants
- in winter even such forage is better, than starvation is.
In spring when snow thaws, growth of oromatrica begins. Live leaves on top
of plant open, and between them young leaves grow. Silvery hairs covering leaves
protect them from superfluous sunlight and reflect pernicious ultraviolet rays.
In stalks there is a stock of nutrients, therefore this plant can recover even
after all its top part was eaten by animals or has been broken by snow avalanche.
Then on the rest of stalk lateral sprouts grow, and the plant forms sprawling
bush. Young leaves can endure rather strong night frosts – spring comes in
high mountains not at once, and sometimes winter may return for some days,
and even weeks. But sooner or later the rests of snow thaw, and there comes
warm weather. The plant actively forms new sprouts and restores roots. The
root system of oromatrica is basically superficial and also forms the ramified
circuit covering large stones: it helps to gather dew which is flowing down
on them in the morning. Landslides and avalanches frequently damage roots of
oromatrica, but they are quickly restored - slow growth in mountains obviously
does not promote survival.
When warm weather is finally established, and danger of night light frosts
leaves, oromatrica begins blossoming. Inflorescences of this plant are similar
to camomiles with short white petals (more precisely – edge semiflosculous
flowers in inflorescences). They tower above a plant on long ramified peduncle.
Flowers smell pleasantly, involving various insects: this plant is undemanding
to pollinators. After the pollination on inflorescence small seeds with fluffy
pappus ripen. When they will completely ripen, their pappuses extend and open.
The first rush carries seeds, but only few of them will get in conditions favorable
for growth. Having fallen on the ground, seeds do not sprout at once: winter
should pass before they will go to growth, and some seeds sprout only after
two winters at all.
Within the first summer of life the young plant forms the crown of leaves on
short stalk, and starts blossoming only since the second year. Life expectancy
of this plant is about 13 – 16 years.
At highest mountains the closely related species grows:
Cabbage
oromatrica (Oromatrica pseudobrassica) – it is more undersized plant:
it forms “cabbage head” of leaves on short stalk. In severe conditions when
even in heat of summer the temperature does not increase above +15°С, and light
frosts at night are possible, this plant manages to grow and blossom. The apex
of this plant is constantly closed by fluffy young leaves, and light frosts
do not harm to it. Peduncles of this plant are short, and petals are covered
with thin layer of wax and reflect ultra-violet light, involving rare insects
of high mountains.
Leafless sea aster (Aphyllaster littoralis)
Order: Asterales (Asterales)
Family: Asteraceae (Asteraceae)
Habitat: littoral zone from Northwest Europe up to North Africa, Iceland, New
Azora.
In Neocene the shore line of Northern Europe had undergone significant changes.
When weight of thousand cubic kilometers of glaciers holding down the significant
areas of continent, had disappeared, the north of Euroasian lithospheric plate
began to rise. The sea receded in significant territories, and the former sea-bottom
turned to plains on which there were the saline lakes drying up later. The
territory, occupied by Northern Sea in human epoch, had turned to extensive
marshy plains with the separate heights overgrown with forest. Baltic Sea had
turned to extensive brackish Venedian Lake isolated from Atlantic Ocean basin.
Cold current Anti-Gulf Stream determines the climate of these places: cool
foggy summer similar to late spring, smoothly passing to early autumn. In summer
in the mornings there are thick mists leaving plentiful dew on leaves of coastal
plants.
The coast of Northern Europe to the south of Scandinavia represents wide strip
of saline marshlands, where action of inflow is strongly felt. Ground and water
are saline here, and sea representatives of fauna meet in shallow reservoirs
at the great distance from ocean. Flowering plants actively develop this area,
forming set of endemic species adapted to existence in conditions of ground
salinity.
At small distance from ocean in littoral zone there is the interesting plant
maintaining twice per day the pressure of tidal waves – leafless sea aster.
This is low plant with thick edged branches divided to segments. Edges are
extended in flat wide “wings” with spikes on edge, and serve for photosynthesis.
The true leaves of plant are reduced up to filmy covers protecting young sprouts.
When sprout starts to develop, cover leaves quickly dry up and fall down. Such
development of leaves is the necessity: during the inflow there is a threat
of damage of plant, therefore leafless sea aster has lost soft and delicate
leaves, having turned in rigid plant similar to coral.
On young sprout edges are small – the probability of the damaging of young
sprouts by tidal wave is less so. Till the process of development of mechanical
tissues they expand and strengthened. Leafless sea aster lives in conditions
of surplus of salt, therefore along edges of stalk glands secreting brine as
rich drops are located. In outflow brine dries up on the sun as white flakes
on edges, and then is washed off by inflow.
For additional protection against salt sea water the surface of plant is covered
with thin wax film especially strongly advanced on young sprouts. Because of
it they have strongly expressed glaucescent shade. The sense of this adaptation
is, that at outflow water does not stay on the plant and does not form the
crust of salt, complicating photosynthesis, on it.
In spring, when it becomes warm enough, leafless sea aster begins to blossom.
Inflorescences of this plant look beautiful – they are white with pinkish middle,
a little bit similar by the shape to chamomile flower with “fluffy” middle
and wide petals; their diameter is about 4 – 6 cm. Semiflosculous flowers are
located in one line at the edge of inflorescence. Medial flowers are supplied
with long pinkish outgrowths of perianth. Inflorescences form on very high
strong stems growing above the level of highest inflow. On one plant it may
be over 20 inflorescences blossoming simultaneously. This plant blossoms after
cool wintering: during the period of relative rest in axils of last year's
stalks flower buds are formed. This feature limits the ability of leafless
sea aster to breeding by seeds – the plant exists normally only in conditions
of expressed seasonal changes of climate. This plant is pollinated by various
insects – more often by beetles and bees, but occasionally on its inflorescences
butterflies feed. Flowering lasts till more than two months. At this time places
of growth of leafless sea aster look very beautifully: flowers bring bright
accent in dim green-and-brown palette of interdial zone.
Seeds with fluffy crest ripen at the end of summer. When they develop completely,
the base of inflorescence is curved outside, and they are carried by wind.
Due to this feature the plant is widely settled along the coast of Europe,
having reached even the coast of Iceland. At New Azora and African coast this
plant blossoms seldom – only after especially cool winter, when flower buds
form.
Except for seeds, leafless sea aster is settled with the help of sprouts. Tidal
waves frequently break off branches of this plant and carry them away in ocean.
Parts of plant are able to survive for a long time, floating in sea water.
Due to such ability this plant was settled even further to the south, but southern
populations of this species do not blossom.
“Flora’s
fan” (Flabellisenecio monstrosus)
Order: Asterales (Asterales)
Family: Asteraceae (Asteraceae)
Habitat: South Africa, hillsides.
At plants of various botanical families and orders one interesting feature of
growth is known – it is the producing of cristate forms. In this case the apex
turns to a line of growth, and the stalk of plant instead of the cylindrical
or ridge form becomes flat and crest-like, or has twisting edges. Such forms
were frequently observed in human epoch at plants of cacti family: among them
a lot of species of various genera formed such forms. Less often the producing
of cristate form was observed at plants of other families: at succulent spurges
(Euphorbiaceae), crassulas (Crassulaceae) and Celosia (Amaranthaceae). Usually
cristate forms of plants could exist for a long time only in conditions made
by people, and in nature they did not maintain a competition to usual form of
the same species. But in droughty deserts of South Africa a rare case had taken
place, when this kind of deformity has turned to an attribute favourable to
survival.
Among the rocks heated by the midday sun, strongly keeping by roots for poor
ground, one of the freakish plants of Neocene, “Flora’s fan” grows. It is stem
succulent, which has got water-stocking stalk due to formation of the cristate
form. “Flora’s fan” has two types of sprouts – cristate and typical ones. Perennial
caudex of this species has cristate form and represents fan-shaped structure
(hence the name of this plant) about one meter high. As it grows, it becomes
thicker, the bottom part covers by suberificate bark, and from the basis to
edges of caudex sharpen edges stretch. Edges of caudex, where the growth zone
is located, are slightly wavy. The edge of fan-shaped caudex has green border:
the growth of last two years keeps a function of photosynthesis. But older parts
of caudex become covered by dense skin, turning later to dense grey-brown peel.
“Flora’s fan” is original “compass plant” of deserts of the south of Africa:
fan-shaped caudex grows by the north – south line in order to avoid hot beams
of midday sun (in southern hemisphere in midday it stays at the north). In the
basis of caudex additional buds form, and from them young stalks of fanlike
shape develop.
On edge of fan-shaped caudex the short normal sprouts grow. They have thick
cylindrical pentahedral stalks – possible, the ancestral form of this plant
initially had similar stalks. On sprouts of usual type prickles develop in the
beginning, and then narrow leathery leaves with small prickles on edge. These
sprouts are short and do not branch. Having reached the length of about 20 cm,
they stop growth, and on them small inflorescences develop – heads of yellow
color gathered to common corymb-like inflorescence at the tip of stalk. For
protection against herbivores the surface of caudexes and stalks is covered
with small prickles. Also prickles develop on edges of caudex and annual sprouts.
Prickles on caudex are the modified leaves had kept only thick central nerve,
and on sprouts of usual type prickles are formed by lignificate outgrowths of
edges.
After deflowering leaflets of inflorescence covers supplied with tiny prickles,
protect developing seeds. In dry season when seeds with fluffy cops ripen, the
cover opens, unbends, releasing seeds, and dries up. Seeds of “Flora’s fan”
are carried with the help of wind. Soon after seed ripening sprouts of usual
type die off. After them at the edge of caudex the lines of stamps of characteristic
appearance stay; they are especially large at plants of the age of 8 – 15 years.
By their quantity it is possible to count the age of plant which begins to blossom
for the first time at the fourth year of life. During the drought on caudex
wrinkles appear, and the last year’s growth dries up a little. At the beginning
of new rain season the caudex quickly restores stocks of water. On its edge
the growth of new tissue begins, and then the next line of new sprouts is formed.
Life expectancy of this plant seldom exceeds 30 years.
Silver-shining
bitter tree (Dendrartemisia amarissima)
Order: Asterales (Asterales)
Family: Daisies (Asteraceae)
Habitat: steppes of Three-Rivers-Land, northern and eastern coasts of Fourseas,
to the east up to Mongolia.
Plants of daisy and sunflower family of human epoch were presented mainly by
grassy plants; treelike forms represented an exception among them and lived
mainly in tropics. Climatic changes of the boundary of Holocene and Neocene,
and in addition, changes in biosphere of anthropogenous origin prior to them
have enabled daisies to take the place among treelike plants. This transition
had taken place not only in tropics and in isolated habitats, but also in temperate
latitudes of Eurasia, in conditions of a continental climate.
Representatives of mugwort (Artemisia) genus successfully realized an opportunity
of transformation into the treelike form. In human epoch within the mugwort
genus there was a transition from grassy plants to bush-like forms (like Gmelin’s
wormwood (Artemisia gmelini)). In Neocene in territory of Eurasia thickets of
treelike wormwoods of several species belonging to Dendrartemisia genus have
appeared. The typical representative of this genus is silver-shining bitter
tree.
Silver-shining bitter tree is a treelike plant having a crone of various shapes,
reaching height of 5-6 meters. Being protected from wind, this plant expands
in height and then its crone has the outlines close to oval-shaped. In steppes
the plant does not grow taller than 2-3 meters, and its crone turns low, but
very stretched in sides. Sometimes long branches can touch the ground and take
roots, and then secondary trunks appear. Wood of this species is very hard,
thin-layered, having expressed “sagebrush” smell.
Trunk of silver-shining bitter tree is covered with friable grey bark shelled
as small plates; young branches are covered with dense brownish bark. Shoots
of the current year are covered with grayish short hair vanishing to an autumn.
Leaves of this species have very characteristic shape and colouring. Leaves
of an adult plant are dense and leathery, about 10 cm long, cleft into 5 peaked
narrow lobes. Middle lobe is little bit larger, two shorter lobes adjoin it,
and marginal lobes are the shortest, measuring less than one third of length
of the middle lobe of a leaf. The bottom side of leaf is richly covered with
white felt downiness. Top side of a leaf is dark green, covered with thin layer
of plant wax protecting it from loss of moisture. From apart and in the wind
the plant looks silvery. On the most part of its range, this plant is deciduous;
only on eastern coast of Fourseas it keeps a part of leaves in winter.
Foliage and annual shoots of silver-shining bitter tree contain a significant
amount of glycosides and alcaloids, therefore the plant is actually not eaten
by local herbivorous mammals. Nevertheless, some species of insects eat foliage
of this tree, and wood is bored by a specific species of longhorned beetles.
Mammals eat foliage of this tree in small amounts, ony as an anti-helminthic
agent. For protection against ectoparasites various animals wallow on fallen
foliage of silver-shining bitter tree.
Inflorescences of this species represent small capitula containing 5-9 tiny
florets with reduced calyx, which role is partly taken by leaflets of covers
of inflorescences. The external surface of an inflorescence is covered with
rich white hair reflecting ultra-violet light, due to what insects find out
its flowers. Capitula are composed in long raceme-like inflorescences growing
on wintered shoots. As seeds ripen, capitula break away entirely and are carried
by mammals, being clinged to their wool.
Term from germination of a seed to the first flowering takes about 10 years
at this species, and life expectancy can make over 100 years.
Some more species of treelike wormwoods exist in Eurasia:
Hairy
bitter tree (Dendrartemisia pubescens) grows in mountains of Near East
and at the Persian Ridge. It is rather haet-loving species, deciduous only in
part. This tree growing as several trunks is up to 5 meters tall, very photophilous
and inhabits sunny mountain slopes. Its young shoots and pinnatipartite leaves
with narrow lobes are plentifully pubescent with white hair. Top side of leaf
is covered with a mealy powder.
Creeping
bitter tree (Dendrartemisia repens) grows in mountains of Altai and
represents a typical high-mountain plant. This species shows a characteristic
form of growth: repent trunk and main branches easily taking roots in the ground.
The length of trunk can reach 10 meters, but usually it is shorter. Young branches
rise above a surface of the ground. This is an evergreen plant with leaves cleft
to 7-9 narrow lobes and plentifully covered with downiness from both sides.
The present species winters under, keeping its foliage.
Spiced
bitter tree (Dendrartemisia tropicalis) grows in the south of the range
of genus and is common in Himalayan foothills. This is an evergreen tree up
to 10 meters tall having dark green leaves cleft to narrow lobes. The bottom
side of leaf is covered with a grey tomentose layer; top side of leaf is naked,
dark green; middle nerve is white. Foliage of this plant is rich in ether oils
and emits a pleasant smell especially well appreciable in dry hot weather. Its
greenery has very bitter taste and is poisonous for small herbivorous mammals.
The idea about existence of this group of plants was proposed by Nem, the forum member.
Scarlet
thistle (Pachycirsium coccineus)
Order: Asterales (Asterales)
Family: Daisies (Asteraceae)
Habitat: Pacific coast of North America, humid deciduous forests.
Humid forests of a temperate climate formed along the Pacific coast of North
America represent a favorable place for life of moistureloving grassy plants.
Rains and fogs from ocean favour to growth of rich underbrush in forests formed
of thuja and moistureloving deciduous trees.
Local plants get freakish shapes, and sometimes it is difficult to recognize
in them modestly looking ancestral forms of human epoch.
One of characteristic plants of underbrush of humid Pacific forests is scarlet
thistle, perennial rhizomatous grassy plant with spiked stalks and prickly leaves.
This species descended from perennial creeping thistle (Cirsium arvense), very
common and widespread weed plant. In conditions of abundance of moisture this
plant reaches height of 2-3 meters, and its leaves grow up to half-meter length.
They are pinnatipartite, have large pointed prickles on edges and on the top
side of leaf closer to edges. Nerves are very thick, boldly bulging from the
bottom side of leaf; along the primary vein large prickles grow. Bottom side
of leaf is bright green. Stalks are pentahedral in cross-section, tubular and
annual; along edges rows of thin pointed prickles stretch. After fructification
stalks die off.
In the beginning of vegetation on the surface of the ground a rosette of small
prickly leaves is formed. By the end of spring large adult leaves grow, and
from the middle of the rosette blossoming shoots grows.
Plant develops a massive root system accumulating a significant amount of nutrients.
This species breeds actively by repent rhizomes, which grow to 2-3 cm in diameter.
They have dense cover and friable pulp rich in starch. On the surface of a rhizome
dormant buds are formed, which at damage of rhizomes begin growth and form vertical
shoots. Vegetative reproduction emables a plant to form the extensive clones
stretching to tens meters.
It is a dioecious plant; male individuals are indistinguishable from female
ones externally. Capitula are large – top one reaches diameter of 6-8 cm, lateral
ones grow to 4 cm. In capitula numerous narrow tubular florets of bright red
color develop, because of what capitulum gets hemispherical form. This species
blossoms actually during all summer. Producing nectar plentifully, the ancestor
of this species has attracted new pollinator, a hummingbird. The union with
this pollinator has determined an appearance of a plant and colouring of its
florets, having directed evolution of a plant in the new side. Red color of
inflorescences is attractive to birds, however on florets there are also spots
reflecting ultra-violet light and making them appreciable for reserve pollinators
– butterflies and moths.
Seeds grow ripe by the end of summer. They are small, anemochoric, supplied
with a pappus of long strings enabling to soar in air. Seeds quickly lose germinating
capacity; therefore, this species uses vegetative reproduction more often, forming
large clone thickets.
Prairie
oxeye daisy (Leucanthemum americanum)
Order: Asterales (Asterales)
Family: Daisies (Asteraceae)
Habitat: North American prairies.
Sapient humans as a biological species have existed not for too long time –
for some hundreds millenia only. But during this rather short term they has
proved as the force equal to the other forces of nature: they have destroyed
some species, have created artificially other ones, and have introduced the
third ones to other continents, where they would never get in natural way. Among
such immigrants there was also oxeye daisy (Leucanthemum vulgare), or dog daisy,
which, having got in North America, has easily got accustomed there, having
endured people and glaciers without any problems, and has evolved to new species
for this time. One of these species is prairie oxeye daisy, a perennial grassy
plant of the American prairies.
This species is a perennial grass producing shoots of two types. In the top
part the plant looks as easily recognizeable “camomile” about 10-20 sm tall,
having large capitulum inflorescences with white ray flowers, and on poorly
branching stalks lengthened leaves grow – simple ones with slightly dentated
edge. In the bottom part of a plant leaves are wider and rounded, that is better
to absorb sunlight. The bottom surface of leaf is covered with silvery felt
downiness, on the top side numerous transparent trichomes develop, slightly
diffusing sunlight above leaf surface. Prairie oxeye daisy is photophilous,
avoids dense thickets of bushes and grows only on open spaces or in a light
forest. This is an evergreen plant – in autumn flower-bearing shoots die off,
and only dense radical rosettes of medium-sized leaves keep to give new flower
stalks the next year.
Prairie oxeye daisy breeds by seeds and is able to vegetative propagation. Its
white inflorescences represent one of the most usual and habitual flowers of
North American prairies. It is especially favourite plant for butterflies –
both as source of nectar for them, and as a fodder plant for their caterpillars.
Herbivorous vertebrates – from various deer
and donkeyhorses to
peccasons – also willingly
eat these flowers, but prairie oxeye daisies do not suffer from it: radical
part of a plant usually remains untouched. This plant breeds actively by underground
rhizomes – at prairie oxeye daisy they are “creeping” and are capable to give
new shoots if old ones will be eaten. It forms new bushes at the distance of
up to several meters from a parental plant. Seeds of prairie oxeye daisy have
tiny hook-like bristles, due to which they easily stick to muzzles and legs
of various herbivorous animals, which then carry them across prairies, where
they sprout later. As a result of it prairie oxeye daisy grows more often along
migrational ways of various herbivorous animals of prairies. The young plant
begins to blossom in favorable conditions the next year, but forms a radical
rosette of leaves more often and begins vegetative reproduction, especially
at the western edge of its range, in more arid conditions, where it begins to
blossom only in wet years. In nonfavourable conditions the plant reproduces
mostly vegetatively, producing the large clones stretching a network of shoots
across hundreds meters. Duration of existence of such clone of prairie oxeye
daisy reaches 60-80 years.
This species of plants was discovered by Bhut, the forum member.
Monocotyledon
plants
|
Pelagic
pondweed (Potamogeton pelagicus)
Order: Najadales (Najadales)
Family: Pondweeds (Potamogetonaceae)
Habitat: Fourseas, top layers of water far from coast.
Sea algae grew in Black Sea, have died out long before the sea has dried up.
During the ice age of the boundary of Holocene and Neocene, immense masses of
water were accumulated in glaciers, and the river flow into Black Sea has drastically
decreased. Salinity of its water has increased gradually up to oceanic level,
and then has exceeded it and Black Sea has turned to some hypersaline reservoirs
with water poisoned with hydrogen sulphide. Such conditions were completely
unsuitable for life of algae.
In Neocene at the territory of Eurasia Fourseas has appeared – large midland
reservoir with brackish water. Its flora is presented exclusively by descendants
of freshwater and marsh plants, which have found conditions favorable for life
here. The majority of plants of Fourseas clusters in a narrow zone of coastal
shoalinesses, and between them a severe competition for habitats and sunlight
takes place. But far from the coast in open water nothing shades sun rays. Here
microscopic algae plentifully develop, and from among the higher aquatic plants
only one fast-growing species – pelagic pondweed – exists there.
The majority of pondweeds represents grasses ecizing in the bottom. Pelagic
pondweed is free of connection with a bottom, and it has enabled it to enter
new habitats not occupied by its competitors. It forms extensive floating thickets
on the surface of water stretching to tens meters and representing real rafts
of bound stalks.
Pelagic pondweed is a long-stalked a plant. The length of its stalk reaches
4-5 meters, and the daily amount of growth makes about 5 sm at each stalk. Stalk
of this species is thin, reddish, plentifully branching in leaf axils and very
rupture-resistant. In nodes of stalk long roots hanging down and covered with
thin fibrils develop.
Floating leaves are its main floats. They are oval with obtused tip and long
petiole, their length is about 10 cm. Tissues of leaf have rich air-bearing
parenchyma, and the top surface is smooth and glaucous, with thin wax cover.
Due to this covering leaves are water-repellent and protected from surplus of
sun light. Leaves are alternate, and in their axils new shoots develop. When
during storm the stalk appears broken off, it does not harm to a plant – it
quickly grows new shoots. Thickets of pelagic pondweed are so dense, that small
birds can walk on them, almost not slumping. Frequently flocks of birds taken
by wind in the sea, save their lives in thickets of pelagic pondweed, where
they can have a rest and find food – small invertebrates gathering in thickets.
Underwater leaves of this species are small, transparent and emerald-green.
They develop on the youngest shoots and do not provide meaningful buoyancy.
Having only formed a first floating leaf, shoot begins keeping on water steadily.
Blossoms of this species are small and modestly looking; they have four petals
and are arranged in spike-like inflorescence. Inflorescences develop in leaf
axils and rise from under water vertically. Seed breeding seldom happens successful,
because insects only casually appear in place where this species grows. Pollination
by means of wind is inefficient, as the large thicket of this plant may represent
a clone, and self-pollination actually takes place. Nevertheless, this disadvantage
is quite compensated by active vegetative reproduction.
In an autumn floating stalks and leaves quickly die off and decompose. At this
time leaf axils short shoots with scale-like leaves – turions – develop. They
float in the top layer of water due to presence of cavities in a core rich in
starch. Turions easily endure deep freezing and frequently winter in thickness
of the ice covering coastal water areas. At rise of temperature of water they
quickly sprout to new shoots.
Common
dendronajas, sea tree (Dendronajas vulgaris)
Order: Najadales (Najadales)
Family: Dendronajadaceae, sea trees (Dendronajadaceae)
Habitat: Atlantic ocean, coastal waters and underwater shallows up to depth
of 20 meters.

Picture by Nem, colorization by Biolog
Initial image by Nem |
In Neocene some angiosperm plants have taken advantage of consequences
of global ecological crisis and have made successful attempt to force macroalgae
to retreat from their place in plant communities of sea shoalinesses. In early
Neocene from Cymodoceaceae family an extensive Dendronajadaceae family descended,
evolved roughly during the next 15 million years. 25 million years after human
epoch it includes 10 genera and more than 100 species settled in subtropical,
tropical and boreal zones of all oceans.
The shape of plants of Dendronajadaceae family is very unusual. They have developed
vertically standing rhizomes (“trunks”) – stout, ligneous, up to 5-7 m tall
and up to 70 cm in diameter, capable to secondary growth due to cambium-like
tissue. Inside trunks there are vessel elements; walls of old vessels in trunk
core can accumulate calx. Reproduction of these plants is mainly vegetative,
but in favorable conditions seeds ripen successfully.
These are dioecious plants; pistillate flowers reduced in great degree develop
inside the spatha (frequently long and narrow) spirally wrapped up along edges;
stile and stigma are long (sometimes branchy) and threadlike. Male flowers develop
on special short shoots, which die off after flowering. During the ripening
bracts around of seeds become fleshy, and the external spatha opens, exposing
two lines of fruits, which are actively eaten by some fishes. Seeds have dense
envelope, therefore they are not digested and with faecal masses fall on sea
bottom where sprout. At some genera an envelopeof fruits is bright and contains
air cells; such fruits emerge on water surface, where sea birds peck them.
Being widely spread along the Atlantic coast of Old World, common dendronajas,
or sea tree, forms extensive thickets in coastal sea waters. The height of its
“trunks” reaches 3-4 meters, up to 5 meters in subtropical waters. Old parts
of “trunks” of kinds of this sort are covered with specific crust-forming lime-depositing
algae of Lithophaneroderma genus, which during their growth form a thick bark
up to 3-4 cm thick. During the secondary growth of trunk this bark bursts, but
during their growth algae close cracks formed this way. Common dendronajas lives
strictly in sublitoral zone, because apical points of this plant do not endure
drying in air. If tops have dried, growth stops for a certain time, and under
the bark new apical points form. New plants grow also from the bases of broken
“trunks”. A piece of “trunk” rolled by sea waves easily takes roots at the bottom
in a silent place and gives vertical shoots.
Leaves of sea tree are simple, ribbon-like, of dark green colors, up to 3 meters
long and about 3-4 cm wide. They keep alive for about 2 years, constantly growing
up at the basis. Tips of leaves are obtused, but usually the top of leaf blade
is damaged by sea animals and large whole leaves are almost not observed.
This plant species blossoms seldom. On the northernmost edge of its range common
dendronajas begins blossoming only in the warmest years though it can grow even
in cold water of Antigulf Stream. In a southern part of its range flowering
of this species takes place almost every year. Male plants differ from female
ones in smaller height of “trunks” and shorter leaves. They prefer to grow on
shallow sites of sea bottom and use vegetative propagation actively, due to
what resist to occurrence of female plants of this species in their thickets.
Male flowers have three large anthers and are covered by leathery calyx. They
produce a plenty of fine pollen, which is washed away by water in due course
of its ripening. Female inflorescences develop in the centre of leaf rosette.
Long inflorescence contains up to 10 pairs of tiny blossoms with long stigmas,
wrapped up in edges of spatha. Pistils jut outside like long brushes, on which
pollen settles. Fruits are fleshy and berry-like. Each fruit contains up to
20 small seeds sprouting approximately in two weeks after passage through intestines
of a fish.
Dendronajadid reef forests became the most highly productive coastal sea biocenoses
of Neocene. Due to multilayered structure of these thickets various fishes and
invertebrates find shelter there.
This species of plants was discovered by Nem, the forum member.
Poseidon’s
onion (Thalassoalium bulbosum)
Order: Najadales (Najadales)
Family: Dendronajadaceae, sea trees (Dendronajadaceae)
Habitat: the south of the Atlantic and Indian oceans, coastal waters and underwater
shallows up to depth of 10 meters.

Thalassoallium bulbosus and Th. pyramidalis (picture by Nem, colorization by Biolog)
Initial image by Nem |
Representatives of Dendronajadaceae family represent an unique phenomenon in
flora: these are perennial lignifying plants capable to grow in water of oceanic
salinity. The majority of species of these plants has a tall trunk and inhabits
depths up to 20 meters, frequently reaching surface of water by their leaves.
But representatives of one genus of these plants have evolved in the other direction,
having developed stormy coastal shoalinesses. These are representatives of Thalassoallium
genus, original “bulb plants” of seas of southern hemisphere.
These plants differ in very short and thick non-branching “trunks”. Due to massive
trunks, these plants easily endurethe dash of the waves and long roots hold
them in place even during a storm.
The typical representative of genus is Poseidon’s onion. Trunk of this plant
reaches a half meter height at almost same diameter. At young plants, trunk
is more extended in height, but with the age, it grows thicker a lot. Surface
of trunk is covered with a dense bark keeping in the top part scars from fallen
leaves. At top of a trunk narrow linear leaves with obtused tops grow in a bunch.
They are short – no longer than one meter long even at large specimens; tip
of leaf decays in due course, but it grows up from the basis for a long time.
Core of trunk is rich in starch. At young plant trunk is spongier, in the core
gases (usually carbonic gas and oxygen) gather. Due to this feature the young
plants uprooted during storm can drift in waves and settle this way.
In the basis of trunk of adult plant buds appear. At damage of apex new vertical
shoots form of them instead of repent horizontal rhizomes in normal conditions.
At the some distance from an adult plant daughter plants appear. At Thalassoallium
genus vegetative reproduction prevails above seed breeding: sometimes a population
present by thousands plants appears a clone of one plant expanded during several
centuries.
Poseidon’s onion belongs to dioecious plants; male and female specimens are
externally similar, and in the same conditions, male plant is only a little
bit smaller than female one, but in numerous populations this difference is
poorly appreciable. Seed breeding takes place rather seldom: usually the population
of plants begins to blossom synchronously once in some years. On female plants
long flower stalks grow, bearing up to 10 pairs of small flowers closed by spatha
turned in a tube. From the top part of spatha long thin stigmas divided on their
tips to the set of strings jut out. Due to resistant and flexible flower stalk,
the inflorescence floats on the surface of water and stigmas appear on the film
of a surface tension. Male plants form raceme-like inflorescences of numerous
modesly looking blossoms lacking petals and producing pollen with a waterproof
layer on the surface. Pollen emerges on the surface of water, looking like spots
of yellowish color, and the pollination takes place. The main condition of successful
flowering is a quiet surface of the sea. After pollination flower stalk twists
like a spiral and seeds ripen under water.
In waters of South Africa another species of the genus grows – pyramidal
sea onion (Thalassoallium pyramidalis). This species is better adapted
to life in conditions of storm weather. Its stalk grows extremely thick, and
old roots gradually twist, drawing it in thickness of sea bottom. Therefore
the part of stalk seen above the ground surface has conic shape. This species
also uses mainly vegetative reproduction, but is capable to blossom and to be
pollinated even in conditions of often storms. Its inflorescences do not reach
surface of water, and pollen floats in thickness of water.
This species of plants was discovered by Nem, the forum member.
Sea
banyan (Thallasoficus ramosus)
Order: Najadales (Najadales)
Family: Dendronajadaceae, sea trees (Dendronajadaceae)
Habitat: tropical waters of Indian ocean, depth up to 10 meters.

Picture by Nem
Initial image by Nem |
In Neocene angiosperm plants master shallow waters of oceans
in new quality – as perennial lignificating plants, frequently of large size.
Due to the large size and ability to vegetative breeding they successfully compete
to macroalgae and their thickets represent a basis of productive ecosystems
of sea shallows. These plants have various forms of growth that enables them
to exist successfully in places having various conditions of life: from quiet
coves up to the coasts exposed to dashes of the waves during storm.
In tropical areas of Indian Ocean at coasts of Africa and Asia the representative
of dendronaiad family remarkable in its original form of growth lives: its “trunk”
is made of a network of plentifully branching and bound rhizomes which are frequently
merged with each other in places of their contact. Thickets of this species
resemble in their appearance the banyan – ficus species; because of it the plant
is named as sea banyan. Rhizome of this plant begins its growth in vertical
position, but further it lays on sea bottom, takes roots and begins branching.
Runners creep on a site of sea bottom and on them numerous vertical shoots develop,
branching, bound and merging with each other. As a result, a three-dimensional
cellular structure up to 2 meters high, consisting of lignifying and gradually
growing thick rhizomes covered with a glaucous elastic bark, is formed. Among
stalks of this plant fishes and crustaceans willingly settle. Diameter of thickets
can reach 10 meters and more. Underground rhizomes do not appear at this species.
Due to ability to form elastic and strong thickets sea banyan frequently settles
along the external edges of reeves, where shoaliness breaks and is replaced
by a slope descending in depth. In such places sea banyan forms “ledges” up
to 3 meters long, which hang above a deep-water site. During a storm they frequently
crack, but keep viability, and the plant increases a new layer of rhizomes from
above. Thickets falling down from a slope represent an important food source
for the animals inhabiting slopes of underwater mountains.
Leaves of sea banyan are simple, lengthened, with obtused tip. They grow on
tops of shoots and cover with a continuous carpet an interlacing of rhizomes
of the plant. At damage of thickets, when light penetrates deep into interlacings
of rhizomes, on them sleeping buds quickly develop, from which new shoots grow.
The broken rhizome quickly takes roots and forms thickets soon.
Sea banyan plentifully blossoms annually. Pistillate flowers are small and inconspicuous,
lack petals and grow inside the involuted spatha in 3-4 pairs. Long ramified
stigmas look like brushes jutting out from the spatha. Inflorescences on short
stalks are formed in leaf axils. Seeds of this plant are easily dispersed by
water to great distances. As against many related species, sea banyan is capable
to produce seeds without fertilisation. Male individuals of this species may
be found much less often than female ones. They have smaller size and prefer
to grow in places with constant slow current. Manle flowers are tiny, having
three leathery sepals and growing in leaf axils in groups of three ones. Pollen
is washed away by water and float in thickness of water, keeping viability for
a long time. Seeds are small and heavier than water. Due to a fleshy nutritious
envelope they are willingly eaten by fishes which disperse them to great distances
along the coast. Near the islands separated from the mainland by open ocean,
this species is presented mainly by clones of few individuals and breeds exclusively
due to vegetative reproduction.
This species of plants was discovered by Nem, the forum member.
Avian
pneumospermium (Pneumospermium ornithochorus)
Order: Najadales (Najadales)
Family: Dendronajadaceae, sea trees (Dendronajadaceae)
Habitat: Pacific Ocean, everywhere in tropical and subtropical zones.

Picture by Nem, colorization by Biolog
Initial image by Nem |
During the rough evolution in early Neocene representatives
of dendronaiad family have developed various strategies of survival in marine
habitats. They were connected usually to changes in growth mode and helped to
resist sea waves and currents. But at the majority of dendronaiads seeds were
dispersed exclusively by fishes – they were enclosed in nutritious shell attractive
to them. Pneumospermium genus from dendronaiads family during evolution has
developed a different way of seed dispersing – with the help of sea birds.
Thickets of avian pneumospermium represent a very characteristic feature of
tropical sea shallows: they as if continue mangrove thickets under water, keeping
a ground by roots and preparing the ground for the incursion of mangrove forests
to the sea. Avian pneumospermium forms extensive thickets stretching to tens
meters. They may be submitted both by genetically diverse plants, and by clones
of one plant. The height of stalks of this plant does not exceed one meter.
As against the majority of dendronaiads, this species forms repent ground sarments
only seldom. Usually the clone is formed, when the old bush of this plant begins
to die off, and the bottom branches lay on the ground. “Trunk” of this plant
has the treelike shape, having irregular biparous branching. The surface of
rhizomes is covered with an elastic brownish bark; it is yellowish-brown on
young parts of rhizomes. On the tips of rhizomes bunches of elongated leathery
leaves with a spongy interior develop, among which modestly looking, but rather
large inflorescences grow. Pneumospermium usually grows up to a level of the
lowest outflow. The plant rather harmlessly endures drying of its tops, because
the top of shoot and young leaflets are covered with a dense layer of jelly-like
protective substance.
The root system is deep, enabling this plant to endure impacts of storm waves.
If the plant is damaged by storm, it quickly restores itself, and the broken
off parts of a rhizome are carried away by waves and take roots in a new place.
Due to their spongy structure, leaves serve as a fine float, not giving a heavy
rhizome to sink, and stocks of starch in a rhizome enable this plant to keep
viability for a long time and to be carried by currents to the great distances.
Male and female plants in a non-blossoming condition almost do not differ from
each other: male plants only have smaller leaves. Male blossoms lack petals,
have only three leathery sepals, develop in leaf axils in rich clusters. They
form a plenty of powderlike floating pollen forming yellowish spot on the surface
of water. Flower stalk is short, spatha resembles a plant leaf involuted lengthways,
but on it a double line of swellings marks female flowers. Stigmas stick out
in pairs from a longitudinal silt in the involuted spatha. Pollination takes
place during the outflow, when stigmas reach water surface. Fruits are berrylike,
having thick and brightly colored skin, which contains air-filled cells. At
ripening of fruits, spatha withers away and the fruits broke away from pedicels
emerge on a surface, where sea birds peck them. In a stomach and an intestines
of a bird seed covers turn thinner and after that they sprout much faster. Due
to sea currents and birds this species is widely settled at the islands of Pacific
Ocean, not forming any clearly defined subspecies.
This species of plants was discovered by Nem, the forum member.
Fiery
lily (Lilium igneum)
Order: Liliales (Liliales)
Family: Lilies (Liliaceae)
Habitat: prairies of North America.
One of the basic problems in survival at plants from prairies of North America
in Neocene is not a winter cold, but activity of hungry animals. Numerous herbivorous
mammal of this natural zone eat vegetative forage in plenties, and this circumstance
appeared an important factor determining structure of vegetation of prairies
and their adaptation for a survival. Many plants of this ecosystem breed in
mass amounts and also grow very fast; but if in case of some kinds of trees
and bushes it promotes a survival, other plants have chosen a little bit different
way – for example, they became poisonous, and herbivorous mammals avoid them.
One of such poisonous plants is fiery lily from North American prairies. This
is a tall plant, up to one meter high, with bright greens, hard stalks and long
leaves. On tops of stalks bright red flowers having six long petals with obtused
tops blossom. Inside a flower there are spots and strips covered with hair reflecting
ultra-violet light. They are signs of a way to nectar for pollinator insects.
Seeds develop infrequently: on the average, only 1-2 capsules per plant for
one season of vegetation. Flowering lasts since late spring up to the middle
of summer, and some flowers have time to blossom on young plants at the end
of the summer.
Colouring of flowers makes fiery lily rather noticeable in prairies, and it
is necessary for its survival: greenery of a plant has very unpleasant taste,
and only few eaten stalks of this species can even poison such big animal, as
peccason or large deer.
Numerous phytophagous invertebrates also do not eat this plant – there are only
few beetle species damaging greens and gnawing bulbs of this species.
Fiery lily reproduces mainly by bulbs and underground runners, which form on
the bottom, underground part of a stalk. Runners grow in all sides, forming
there new bulbs, from which the new plants sprout, which will blossom already
the next year. Many herbivorous animals avoid eating them, but peccasons sometimes
wallow on these plants, using their poisonous sap for relief from ectoparasites.
Fiery lily is a perennial plant, and a separate individual can exist till 20
years and more, reproducing by vegetative propagation at this time.
This species of plants was discovered by Bhut, the forum member.
Live-bearing
papyrus (Cyperus viviparus)
Order: Poales (Poales)
Family: Sedges (Cyperaceae)
Habitat: Central Africa, swamps and riverbanks.
In human epoch the significant part of forests of Central Africa had been cut
down, and plantations and other artificial plantings were stretched instead
of them. But along riverbanks, where extensive bogs had stretched, forests had
kept in more ar less complete condition. Difficulties of lumbering and niduses
of infections dangerous for people have helped to islets of natural communities
to be kept, and then they became a pledge of the future revival of forests after
extinction of mankind. In Neocene tropical rainforests occupy a significant
part of Central Africa. Many forests are swampy, and here in forest canopy there
are gleams, and trees are replaced by giant grassy plants. One of dominant species
of swamp plants is live-bearing papyrus.
This species is not a descendant of true papyrus (Cyperus papyrus) – that species
became a rarity in human epoch and had gradually died out. Live-bearing papyrus
descended from any small species of Cyperus which stretch their umbellate shoots
in marshes of tropical countries.
In comparison with its ancestors this species had considerably changed. The
height of stalks of this species reaches 5 – 6 meters at the diameter at the
basis of about 40 cm. Stalks of live-bearing papyrus are smooth and strong,
filled inside with friable parenchyma. They grow from the thick rhizome reaching
10-meter length at thickness from above half meter. Rhizome grows under ground
and creeps on the surface of ground if thickets grow in water. An internal part
of rhizome is amylaceous, and external layers of tissues are very strong and
suberificate. On the surface of rhizome additional buds develop, from which
lateral branches grow, and also buds giving rise to stalks are formed. On underwater
parts of rhizome scale-like rudimentary leaves, in which axils buds develop,
are well appreciable.
The top of overground shoot at this species is crowned with an umbrella of long
(up to 1 meter) narrow dark green leaves rigid and rough to the touch. The young
stalks, which have not finished the growth, differ in bright green color and
sticky surface of leaves. Young shoot grows actively during approximately two
weeks, and then is kept during two years if it will not be eaten by herbivorous
animals. At the second year of life shoot begins to blossom. Flowers of this
plant are ordinary-looking, gathered in spikelets at tops of long thin flower-bearing
stems. This plant is pollinated by wind, but the significant role in its pollination
is also played by small beetles which gather on the inflorescences, involved
with an abundance of pollen.
Not each flower of this plant turns to fruit. Nevertheless, this plant has developed
a special way of breeding due to which it can occupy new territories quickly.
In crone of shoot, in leaf axils, buds sprout. From them instead of inflorescences
young and tiny plants with roots and shoots develop. Heavy roots of daughter
plant are rich in starch – it is a stock of nutrients and means of maintenance
of balance. Shoots, on the contrary, are lightweight and parenchymatous. The
horizontal rhizome at daughter plant does not develop yet, and some shoots grow
from buds at the basis of first of them. The young plant receives water and
mineral substances from parental shoot, and grows actively. When it becomes
too heavy, it breaks off a crone of adult plant, falls in water and floats in
current, keeping vertical position. Cast ashore, such plant quickly takes roots
and starts development. In such way live-bearing papyrus easily occupies new
habitats. Sometimes young plants of this species float to the ocean, but in
sea water they perish quickly.
Hamster's
wheat (Syntriticum dimorphosperma)
Order: Poales (Poales)
Family: Graminoids (Poaceae)
Habitat: steppes of Three-Rivers-Land, Middle and Central Asia.
Picture by Biolog
In the nature very often there were unions of plants and animals,
in which animals played a role of “owners” of plants, creating by the activity
favorable conditions for plants development. Such unions arose in parallel in
different areas of Earth at species of different systematic groups. For example,
within human memory different insects, ants and termites, cultivated microscopic
fungi on rotting vegetative material. Millions years of co-evolution had resulted
in occurrence of separate species of fungi normally developing only in symbiosis
with insects.
In Eurasia of Neocene epoch the situation had repeated: in steppes the new symbiosis
of plant and animal had appeared. The background of this symbiosis is extremely
remarkable: it originates in human epoch. Human agricultural activity had generated
the special forms of graminoids, wheat and rye, with large seeds ideally adapted
to settling with the human help. Cultural cereals appeared so changed by artificial
selection that the majority of their forms appeared unable to exist without
people and had died out. But some forms of cultural wheat had survived in steppes
of Eurasia due to activity of animals against which people waged ruthless war
till thousand years – rodents. When the people had disappeared, rodents began
to carry seeds of graminoids cultivated before by people. Running wild graminoids
lost the attributes acquired till thousand years of selection and adapted to
new seed carriers. This way in Eurasian steppes new species of these plants
had appeared.
The separate species of graminoids was generated in conditions of symbiosis
with farmer hamster, one
of steppe rodents. This animal had passed from simple gathering of seeds of
wild graminoids to true cultivating of fodder plants at its territory. One species
of graminoids grows especially good in conditions of similar care and has adapted
to coexistence with farmer hamster. Its possible ancestor is feral wheat (Triticum)
widely cultivated by people, and its descendant is named hamster's wheat. Main
feature of hamster's wheat permitting it to receive advantages from the union
with gluttonous rodent is features of seeds. At hamster's wheat there is the
special structure of flowers: they grow together in clusters of three ones,
and the ear of this plant is made of such “triplets”. The middle flower in “triplet”
is not pollinated, but from it large parthenocarpic seed with normally advanced
embryo is formed. It is rather large and also is able to sprout in favorable
conditions. From sides two another flowers forming seeds only at pollination
grow to the middle flower. For this feature the plant had received the genus
name “Syntriticum” – “joint wheat”. At lateral flowers of hamster's wheat seeds
are small and covered with dense environment. They have the concave shape and
surround the middle parthenocarpic seed like a peel. Their covers are firm,
and it is difficult to gnaw them; the rodent, as a rule, disregards these seeds,
clearing from them and cracking only larger middle seed. Seeds of the second
type thrown out in common with dust sprout the next year.
At northern and western borders of area this plant is annual – the underground
part of plant does not maintain a frost in conditions of continental climate.
Only under thick layer of snow the part of roots hibernates more or less successfully.
At the coast of Fourseas and in western part of Three-Rivers-Land steppes climate
is rather warm, and the plant becomes perennial – it grows at the same place
till three years in succession and even longer at the extreme south of area.
The shape of hamster's wheat is typical for graminoids: at it there are high,
up to one meter, strong culm stalks and long narrow leaves. Hamster's wheat
gives new buds in the basis of stalk and forms friable bushes. One sprout forms
till the growth season a bush on which it is formed about ten ears weighting
20 grams each other to an autumn.
In mixed herbages competitiveness of this species is too low. The plant survives
much more successfully in the union with farmer hamster. This animal raises
chances of hamster's wheat to survival, carries its seeds and looks after “allotments”
cutting plants competing to this species.
First weeks of life the plant develops rather slowly – at this stage of development
other grasses, especially wide-leaved ones not belong to graminoids easily muffle
it. Hamster's wheat expands, giving from the basis of main stalk lateral sprouts.
To the beginning of summer all sprouts start to grow quickly and the plant overtakes
former competitors. The plant quickly reaches one meter height and forms ears
with long stigmas and plentiful dry pollen. Pollen of hamster's wheat falls
from anthers approximately in midday time during one hour. In same time stigmas
of pestles are susceptible to pollen. The plant is self-sterile and is only
cross-pollinated. The wind carries its pollen: it is the common feature of all
graminoids. Seeds ripen to the middle of an autumn. In southern areas where
the vegetative season is long, flowering and fructification pass by two waves
– the plant flowers second time on lateral sprouts approximately one month after
the basic flowering. Thus seeds also ripen in two portions.
Mountain
rice (Oryza montana)
Order: Poales (Poales)
Family: Grasses (Poaceae)
Habitat: Himalayas, tropical high mountain areas.
Cultural species and cultivars of rice occur mainly from the Far East and from
Africa. This plant genus includes the moistureloving species growing in marshes
and in other wetlands. In Neocene epoch cultural rice was introduced to many
countries of tropical zone and has naturalized in new places as a crop plant.
After collapse of a civilization range of rice decreased drastically, the majority
of cultivars has died out without any posterity, but somewhere in new places
it has settled down – for example, in a southern part of Himalayan mountain
range. In due course of evolution here a new kind evolved – mountain rice adapted
to life in local reservoirs.
Mountain rice is not less moistureloving, than its ancestors, but conditions
of its growth differ strictly from warm marshes with stagnat water, where its
ancestors lived – these are wild mountain rivers with fast current and much
cooler water. Life in such conditions has considerably changed a plant. The
adult plant of this species resembles most of all a bush with numerous branches,
which are almost leafless. Branches of different plants frequently entwine with
each other, fixing thickets of this grass even firmer. These shoots with strong
stalks are generative one: above water, spikes develop on them, and shoots are
able to resist to water flow in certain degree. Leaves of mountain rice look
like thin tapes: above water, only flowers and spikes stick out.
This plant forms one more kind of shoots, vegetative ones. They differ in weaker
and flexible rupture-resistant stalks, and long ribbon-like leaves trailing
in water flow. They easily form roots in nodes, and internodes in them are long,
due to what shoots reach the length more than 150 cm. They enable a plant to
occupy new territories, and on the established shoots growth of generative stalks
begins soon. Roots of vegetative shoots serve a plant as anchors; they do not
give current to carry it away, or help it to fix at the new place. Due to them
mountain rice is able to vegetative reproduction – its shoots torn off from
a parental bush can produce new roots and sprout at the new place. Roots have
ability to “stick” hard to a rough surface of stones, fixing a plant on the
substratum.
Modestly looking membranaceous flowers of mountain rice are gathered in hanging
lax panicles and are wind-pollinated. Its spikelets are rather dark and ordinary-looking
in comparison with its domesticated ancestor. Seeds of this plant remain on
a parent plant for a long time – it is a heritage got from its cultural ancestor.
They look rather unpresentably and keep bracts surrounded a flower. Waterfowl
and shorebirds, and even some fishes willingly eat them in great amounts: these
seeds are rich in starch and nutritious.
Mountain rice is adapted to life in the flowing water; in reservoirs with slow
current it loses fight for territory to other plants, therefore its populations
are most numerous in mountain areas. The maximal life expectancy of a plant
of mountain rice is up to 6 years. Usually the plant dies off after mass flowering,
and the significant part of a population may be present as sprouts and young
plants of an underwater phase.
This species of plants was discovered by Bhut, the forum member.
Drying
cryobromelia (Cryobromelia arescens)
Order: Poales (Poales)
Family: Bromeliads (Bromeliaceae)
Habitat: Andes, high mountain areas near to snow line.
In human epoch the greatest variety of representatives of bromeliad family was
observed in tropical rainforests. Nevertheless, in various ecosystems of South
America there were the representatives of this family adapted to different,
more severe conditions of life. This pattern is actual also in Neocene: some
representatives of family live in extremely severe conditions, having developed
original adaptations for survival.
At the high-mountainous meadows of Andes one bromeliad species lives, growing
actually up to snow line – it id drying cryobromelia. This plant is adapted
to life in conditions of physiological dryness and the increased level of ultraviolet
radiation, and in the south of range it is capable to endure winter cold. This
species is a rosette plant with thick spherical stalk and numerous narrow leaves
pointed on tips to hard wooden prickles. The bottom side of leaf is covered
with downiness of silvery hair and numerous bristle-like prickles. Top side
of leaf is smooth, dark green, covered with thin layer of wax reflecting sun
rays. In winter external leaves at this plant dry up and serve as an insulating
wrapper for the rosette of live leaves, protecting them from cold and drying
up wind. At this time plant is similar to ball or cactus – it is entirely covered
with a hair layer, in which prickles stick out, which also protect it from not
numerous local herbivores. Root system of plant represents several strong fibrous
roots and serves only for attaching of this plant to substratum. This species
grows among stones, on sites with thinned soil layer, and even simply in cracks
of rocks with a minimum amount of soil. This species gets a significant part
of nutrients from air with a dust which is kept by its leaves. The plant is
extremely photophilous and does not grow in places where bushes appear.
In the spring plant passes to a phase of active growth. Due to increase of humidity
of ground and air, wrapper of dry leaves opens and active growth of new foliage
involved in active photosynthesis begins. Leaves of this plant in active growth
phase form widely spreaded rosette: height of an adult plant reaches 20 cm,
and diameter of its rosette is up to 50 cm.
This is a monocarpic plant, blossoming at the age of 3-5 years, having stored
enough of nutrients in its stalk. To this moment at the basis of a parental
plant 2-4 young daughter plants are already formed. Inflorescence of this species
represents a tall cluster of small ears with bright red bracts. Flowers have
three petals and are tubular, having a cover of plant wax on edges of their
petals. Plant is pollinated by high-mountainous species of hummingbirds and
by hawk moths. Seeds are tiny, have fluffy cops and are capable to be dispersed
by wind across great distances.
Wax-bearing
cryptobromelia (Cryptobromelia cerosa)
Order: Poales (Poales)
Family: Bromeliads (Bromeliaceae)
Habitat: southern areas of South America, Tierra del Fuego, open areas and hillsides.
In human epoch, bromeliads were known mainly as tropical plants loving humidity,
though rather considerable number of species successfully surviving in arid
habitats was also known. In time of Holocene-Neocene congelation, among these
plants cold-resistant forms began to appear, which in process of thawing of
glaciers receded to high mountain areas or to the far south of the continent.
During evolution they have got the adaptations enabling survival in conditions
of cold climate.
In the far south of South America one of such cold-resistant forms of bromeliads
grows – wax-bearing cryptobromelia. This species has kept rosette form of growth
typical for representatives of this plant family. Rosette about half meter in
diameter is formed of approximately ten leathery tongue-shaped leaves of dark
green color covered on edges with a thin crust of plant wax. The bottom side
of leaf is also covered with the wax giving a glaucescent shade to a plant.
The bottom part of leaf blade is thick and fleshy, staying alive for 2 years
and serving for storage of nutrients. The top part of a leaf blade dies off
to winter; the pulp of a leaf in this part decays, and the leaf turns to “brush”
of fibres. In addition, to winter some young leaves rise vertically and curl
up, closing an apex. On edges and on the bottom sides of leaves the wax is plentifully
secreted, “sealing” leaves and protecting an apex from freezing. In such condition
plant winters, surviving under snow.
Flowering of wax-bearing cryptobromelia begins in the spring and is one of the
most beautiful sights among plants of the far south of South America. When snow
thaws, the leaves protecting apex open, and soon from the middle of the crown
a flower stalk up to one meter tall grows. On its tip a truss of the spikes
formed of bright red bracts with white speckles on edges is formed. Every bract
covers some small white tubular flowers, each of which blossoms for approximately
2-3 days. Wax-bearing cryptobromelia is pollinated by local tayin
hummingbird (Tayin migratorius), less often by butterflies and moths. White
specks on bracts are formed of fuzz covered with a thin layer of wax and reflecting
ultra-violet rays – it is a signal for pollinator insects. Wax from leaves of
this kind is gathered by some solitary bees, which also pollinate this plant.
Flowering of this species lasts for about one month; at the end of blossoming
bracts gradually fade and turn brown. After pollination, the fruits ripen, containing
tiny flying seeds with a cop, which are dispersed by wind.
This plant is a monocarpic grass: after blossoming the rosette dies off. In
its basis buds appear, from which young plants develop later. Also some tiny
young rosettes appear on flower-bearing stem – they are carried by large animals
in their wool. The young plant begins to blossom at the third year of life.
Diving
eichhornia, swamp watermelon (Eichhornia submersa)
Order: Commelinales (Commelinales)
Family: Pontederiaceae (Pontederiaceae)
Habitat: North America, rivers and lakes in the central and southern parts of
the continent.
In human epoch, water hyacinth (Eichhornia crassipes) began one of the most
noxious invasive species. It caused great damage to a nature, outcompeting native
species, destroying ecosystems and changing hydrological regime of reservoirs.
In human epoch its number had been controlled rather successfully, but it was
not possible to have an ultimate victory over this species, and it had entrenched
itself in new territories. The ice age has forced it to recede to the areas
of warmer climate, but after the end of the ice age descendants of this species
began to settle to the north again. During the ice age, the range of the ancestral
species appeared disrupted to some isolated sites that has resulted further
in formation of various species. One of them is diving eichhornia living in
pbasins of large rivers in conditions of subtropical climate. This plant, as
against its heat-loving ancestor, has adapted to survival in conditions of cool
winters, and even can survive for some time under thin layer of ice. In this
connection it developed the certain features of life cycle.
In a warm season this species is a floating rosette plant similar to its ancestral
species. The basic difference in shape from an ancestor is large floats formed
from leafstalks. They are pressed against each other so densely, that form a
spherical structure up to 20 cm in diameter. Leaf blade is separated from petiole
by thin layer of cells capable to suberificate. Leaf blades are of oval shape,
leathery and dark green. In the summer plant breeds actively by sarments, giving
in total up to 30-40 young plants per a season. Actively breeding plants form
on the surface of a reservoir a dense cover capable to bear a weight of large
bird. Flowers are pinkish-white with yellow “eye” on the top petal, resembling
flowers of an ancestral species. The plant seldom gives seeds, actively breeding
in vegetative way. Its greenery is eaten by various herbivorous mammals, insects
and fishes.
Roots are long, covered with numerous fibrils actively absorbing nutrients dissolved
in water.
To winter leaf blades die off, but petioles remain to winter. In the lowermost
petioles, during the vegetation starch grains are stored in parenchyma; therefore
to an autumn they grow heavy considerably and plant crowns immerse in water.
Leaf blades remain only on the youngest leaflets, forming a sort of “cop”. Instead
of air, in petioles water is gradually accumulated and a plant “dives” under
water. The wintering plant has the spherical shape, hence the second name of
it. At this time water-dwelling mammals eat them. Broken off petiole is capable
to form on the breaking point a brood bud, which develops later into a normal
young plant due to stocks of starch. To spring stocks of starch are exhausted
and broken off petioles gradually emerge on the surface, where developing daughter
plants begin independent life. In the north of a range young plants begin blossoming
since the second year of life, after wintering; in the south flowering is almost
all-the-year-round with a break for some weeks for wintering. Life expectancy
of a separate plant is no more than 3-4 years.
Gorgeous
urushiphila (Urushiphila spectabilis)
Order: Orchids (Orchidales)
Family: Orchids (Orchidaceae)
Habitat: forests of Japan.

Picture by Lyagushka
Orchids are most variative family of flowering plants in Holocene
epoch. They had reached the success in struggle for existence with the help
of two important adaptations. First of them is high specialization to entomophily.
Each species of orchids is pollinated by few species of insects, and sometimes
only by one of them. The plant develops the special adaptations in structure
of flower, excluding the feeding of “strangers”, casual species of insects,
on flowers. The second component of their success is the close symbiosis with
fungi. Due to it orchids had solved two problems. They can survive on poor substrata
due to the ability of fungi to transform mineral substances to easily consuming
by plant form. It has permitted to orchids to succeed in ecological niche of
epiphyte. And the second problem, tiny seeds assisting to epiphyte to settle,
also was solved with the help of fungi. Penetrating to seed of orchid, fungus
delivers to microscopic germ nutrients, providing its development at early stages.
Symbiosis has permitted to some species of orchids of Holocene epoch to pass
to other way of life, rather than the majority of plants – to become saprophytes
having lost chlorophyll and leaves. Such orchids live, eating substances delivered
to them by fungi. And some orchids of Neocene epoch had replaced the character
of relations with surrounding species of plants and fungi, having turned to
specialized parasites. One species of parasitic orchids grows in Neocene in
Japan Islands. It grows attaching to roots of amazing local plant – “death
tree”, one of the most poisonous plants of the world. The orchid is named
urushiphila: “urushi” is the Japanese name of poisonous sumach.
The union of the parasitic plant and the poisonous host plant had taken place
in Holocene. One species of sandal tree (Santalum) parasitized at the poisonous
upas tree (Antiaris), and the poison of the host plant had penetrated into it
up to leaves. Urushiphila is also literally impregnated with poison of “death
tree”, using it for protection against pest insects. Only its flowers are lack
of poison.
Urushiphila represents a plant lack of chlorophyll and deeply specialized to
parasitic habit of life. Leaves of this orchid are reduced up to small scales
surrounding the basis of peduncle. Stalk of urushiphila is long ramified rhizome
growing in ground at the depth about 20 cm. It reaches length of several meters
and forms numerous knotty inflows. On them flower buds from which flowers are
serially formed develop.
Besides of “death tree” urushiphila adheres to roots of some sumach species.
Flowers of urushiphila large and single, reaching the diameter of about 5 cm.
at the peduncle it may be two flowers occasionally – at the ancestor of urushiphila
the inflorescence represented a rich truss. Flowers rise from ground to the
height of about 30 – 40 cm and have strong pleasant aroma. Their petals are
white and pointed; labium is cup-like and pinkish. Due to white colouring flowers
of urushiphila are well appreciable in twilight of underbrush. At the labium
of the plant the long spur directed downwards is formed; honeycups are located
in its deep. It makes the pollination of urushiphila the privilege of insects
with long proboscises. This plant is pollinated by butterflies, and on its flowers
the urushi swallowtail,
the large species of local butterflies, feeds often. It is the basic pollinator
of urushiphila.
Urushiphila exists literally “at the verge of nonexistence”, balancing between
life and death. On the one hand, microscopic soil fungi are necessary for germination
of its seeds, but on the other hand secretions of leaves of the host plant kill
the mycelium. The plant had solved this problem rather simply: it patiently
waits while the host plant will find it itself.
Seeds of urushiphila have the microscopic size: it is the common feature of
all orchids. They sprout with the help of mycelium which penetrates into seeds.
First years of life the orchid exists as saprophyte, accumulating forces for
“attack” to the host tree. It grows rather slowly and does not blossom, representing
a shapeless underground tuber with thin skin. The situation changes when the
sprout of host plant appears near it. When young “death tree” starts to develop,
under influence of substances emitting by it the mycelium starts to degrade.
It is one of signals for the tuber of urushiphila modifying the physiology of
the plant: at orchid the feeding root grows actively searching for roots of
host tree under the ground. The orchid feels root secretions of the host plant
and directs feeding root to its side. For the formation of this root stocks
of nutrients had been accumulated till many years of underground existence are
expended. The orchid even starts to “digest” actively hyphae of the fungus permitted
it to sprout. Urushiphila “inserts” to the development of root all nutrients.
If it could not reach the host plant, the tuber starts to degrade and the plant
perishes. Many tubers of this plant may be eaten by various underground animals
or perish when the necessary plant had not appeared near the tuber.
But at successful coincidence of circumstances the plant wins “main prize” in
struggle for existence – after the attachment to tree the orchids start to develop
much more actively. It quickly increases the weight of rhizome and actively
generates flower buds. From germination of seed to the first flowering sometimes
about 15 years pass. Expanding, the rhizome of urushiphila can adhere to near
trees of the host plant species.
Urushiphila grows in woods of Japan Islands seldom enough. But in places favorable
for life it forms mass congestions – among trees tens flowers of this plant
rise.
Bladder-bearing
neocalanthe (Neocalanthe pustulifera)
Order: Orchidales (Orchidales)
Family: Orchids (Orchidaceae)
Habitat: South-Eastern Asia, forests of various types with constant or seasonal
humidifying.
Among the animal-pollinated plants orchids show the highest degree of specialization
to such symbiosis. In human epoch there was a plenty of orchid species showing
various ways of attraction of insects and their use for pollination. On the
border between Holocene and Neocene numerous specialized species have died out
because of destruction of habitats and extinction of pollinators. In Neocene
a plenty of new species has evolved, developing highly specialized ways of pollination
again.
On branches of trees in evergreen and seasonal tropical forests of South-Eastern
Asia, one orchid species lives, which had formed in due course of evolution
the union with nectarivorous bats – it is bladder-bearing neocalanthe, a descendant
of one species of Calanthe genus existed in human epoch. This is a typical epiphytic
plant, which stalk has turned to large onion-like pseudobulb. It grows in forked
branches, among stalks of perennial lianes with lignifying stalks, or in “epiphytic
gardens” formed by ferns. At top of this stalk every year a pair of large (up
to 50 cm long) leathery leaves with obtuse tips develops, dying off before the
beginning of a rest period. Roots of this plant grow hard into the bark of tree
where it develops. Also during the growth rich “beard” of air roots of white
color covered with a spongy tissue develops.
During 1-2 months (in seasonal forests – for about 4 months) per year the plant
stays in relatively dormant condition. At this time in the basis of stalk flower
buds develop. In seasonal forests dormant period coincides with a dry season.
At this time air roots of a plant die off in full or in part.
After the end of a dormant period at the basis of plant’s pseudobulb thick strong
flowerstalk begins to develop, being directed sideways from a tree trunk. On
it 2-3 large flowers blossoming one by one develop. Top sepal and lateral petals
of a flower are colored white; in fauces there are some red strokes showing
to the pollinator the way to nectar. Flower emits very strong vanilla scent.
Bottom sepals are very extended and leathery. Their ends are twisted in spirals
and rock on each side of a flower. Bottom petal (labellum) is pinkish-white
and has small limb, but on its edges rich and strong threadlike outgrowths develop,
and it is extended to long spur, on which tip there is a bladder-like expansion.
Internal walls of this bladder are rich in nectar-secreting glands.
A bat attracted with a smell of flower, perches on it, seizing by hinder legs
in “beard” of a flower, and clings by thumbs of its wings on hard gristly sepals.
Then bat pushes its long tongue into the spur and licks nectar off from walls
of that expansion. At this moment animal touches by head the middle of a flower.
Pollinia paste on bat’s forehead and it carries them to the next flower.
Fruit represents a small capsule filled with microscopic seeds. After the finishing
of blossoming plant has a new wave of growth of new leaves and a period of active
vegetation. At this time large old plants also form lateral shoots.
Development of a young plant takes place rather slowly. Within the first year
of life the sprout is fed exclusively due to fungus it enters symbiotic relations,
and only during the second year of life it develops the first leaf about 2-3
cm long. At six-year age the plant can produce its first flower.
Protected myrmiorchis, ant orchid (Myrmiorchis tutus)
Order: Orchids (Orchidales)
Family: Orchids (Orchidaceae)
Habitat: rainforest of South America; epiphytic plant.
In human epoch many species of plants entered symbiosis with ants were known.
Plants formed in the body special cavities in which ants settled, and in exchange
not only used excretions and waste products of ants, taking nutrients from
them, but also received reliable protection. Among “green friends” of ants
there were acacias, some other trees, and also numerous epiphytes. Among numerous
species of orchids plants of genera Schomburgkia, Coryanthes and Gongora had
hollow stalks served as a refuge of ants. In Neocene independently of them
similar relations with these bellicose insects one more orchid had formed –
myrmiorchis, the epiphytic plant of South American rainforests.
This plant has sprouts of two types. Dense oval leaves of orchid grow on short
vertical sprouts. The main mass of plant is made of thick creeping rhizome,
which creeps along a branch on which the orchid has settled. The rhizome is
especially unusual, because on it zones of secondary growth develop. This phenomenon
extremely seldom meets not only at orchids, but also at monocotyledon plants,
to which orchids belong, in general.
Young apical part of rhizome is smooth. It partly rises above substratum and
is covered with leaves. When leaves fall down, from buds secondary sprouts
start to develop. The old part of rhizome begins changing: occurrence of zones
of secondary growth has casual character, and the rhizome gets irregular shape.
It becomes knobby, in it apertures and cavities, and also additional apexes
form. Old rhizome reaches length of about one meter and half-meter width.
Ants of various species protecting orchid against pests live in cavities of
myrmiorchis rhizome. Besides dung and rests of prey of ants are sources of
organic substances necessary for growth of this epiphytic plant.
But the union of orchid and ants has the back side. Ants perfectly protect
orchid from various herbivorous insects damaging leaves and rhizome of plant.
But they equally successfully can attack pollinator insects, vital for this
orchid. Therefore myrmiorchis had developed the adaptation allowing to pollinating
insects to visit its flowers, not risking getting to ant’s mandibles. The plant
simply interferes with access of ants to flower-bearing stems. At the medium
part of each flower-bearing stem, lower than the lowermost flower is located,
there is original “cuff” some centimeters long consisting of long rigid bristles
of different length. Ant may hardly overcome it, and these insects seldom appear
on flowers of myrmiorchis.
Flowers of myrmiorchis are adapted to pollination by butterflies. The inflorescence
of this plant represents lax truss of 15 – 20 flowers on high (about 60 cm
long) vertical flower-bearing stem. Flower of myrmiorchis has rounded petals,
and also is small and white. It has small labium passing to long spur turned
down and forward. In labium there are some thin stripes of purple hairs, indicating
to pollinator insect the way to nectar. Some dots of the same color may be
in the basis of petals.
Myrmiorchis develops quickly enough – this plant begins to blossom already
at the fifth year of life. This orchid differs in fast growth, forming a bush
up to three kgs in weight to the moment of flowering.
Bromelia-leaved
lady’s slipper (Coronopedium bromeliaephyllum)
Order: Orchidales
Family: Orchidaceae
Habitat: islands of Indian Ocean.
Orchid family is the biggest and rich in species one among all flowering plants.
Its representatives are grass plants have mastered various places of inhabiting.
Among them there are most of all epiphytes, but there are ground species, and
also orchids living in marshes, and even aquatic plants are known. In Australia
in human epoch some species of orchids constantly growing, and even blossoming
under ground were known. One of features, which have provided success of orchids
in struggle for existence, is their symbiosis with fungi. Some orchids have
developed it excessively: they have lost chlorophyll, and live only due to organic
substances of ground which acquire with the help of fungi.
Seeds of orchids are very tiny, looking similar to dust. They sprout and develop
only then when the mycelium of the certain species of fungus will sprout in
them.
The significant part of orchids lives in tropics, but in temperate climate they
also are rather various. These plants may be met both in damp tropical woods,
and in rather dry savannas. In Neocene epoch some representatives of orchids
due to tiny seeds carrying by winds have got from Indonesia to islands of Indian
Ocean and have occupied their ecological niche in island woods.
Orchid living on these islands, is rather original even then, when it does not
bloom. It is an epiphyte: strangely enough, on trees it is easier to find water
necessary for life, than on the ground: subsoil waters on islands are salt because
of affinity of ocean. Therefore rain is a unique reliable source of fresh water
on islands. But rain water needs to be gathered and kept, and orchid from islands
successfully makes it. At it there is short stalk, and curved leaves form the
dense crown. During rains water gathers in crown, forming special mini-pools.
The same feature is characteristic for South-American plants of bromeliad family
(Bromeliaceae), therefore because of this analogy the orchid has received the
name bromeliad-leaved lady’s slipper. Leaves of thislady’s slipper are covered
with spots merging to cross strips – it also amplifies similarity to bromeliads.
For bromeliads such similar striped figure is also characteristic. Diameter
of such crown reaches half meter, and depth is up to 20 cm. In one crown of
adult plant about two litres of rain water may gather. In this water larvae
of mosquitoes and midges frequently find a shelter, and also larger animals
may settle: snails and ground cancers which even hatch posterity, sitting in
these plants.
To save water, at this orchid other adaptation was also developed: leaves are
covered with thin layer of wax. It prevents, on the one hand, superfluous evaporation
of water, and on the other hand - leaves are not moistened with water, and there
is no danger of their rotting. Due to wax coating this plant can normally grow
in dry air.
Peduncles of bromeliad-leaved lady’s slipper are not protected by wax film,
but it is not necessary for them: the peduncle sprouts at the basis of the old
crown. In the beginning from under the crown thick stolon sprouts – it is the
sprout with reduced leaves. Some time it grows aside, and then bends upwards
and forms peduncle carrying one large flower.
The flower at bromeliad-leaved lady’s slipper is pollinated by beetles. For
these insects the plant prepares the special “entertainment” in the bottom petal
(at orchids it is named “labellum”) curved like bag. It is the fragrant sweetish
liquid formed from nectar plentifully flowing down in petal, diluted with rain
water. Trying to get it, insects fall in petal, and then start to search for
an exit. Edges of lip are bent inside, forming “visor” above puddle of nectar,
and it is possible to leave from this petal, only crawling near stigma and having
received after that a lump of sticky pollen. Having sated and having taken “sweet
bathing” in nectar, the beetle leaves flower and flies … to the following one,
where all events repeat. The plant involves insects with contrast colouring
of flower: on vine-red background of petals white spots reflecting ultra-violet
light are scattered. The top petal (“sail”) is entirely red, but its edge is
bordered by white strip. The lip is red - brown, but for insects there are special
“signs” – edges of lip are covered with thin white strokes.
The adult plant forms up to 3-5 flowers at one rosette. The flower keeps on
peduncle rather long time: about 2 months, but not always on its place the fruit
– narrow pod with microscopic seeds – is fastened.
After flowering on sarment in place of peduncle forming the new rosette grows.
But frequently sarments simply dry up after flowering.
The plant actively propagates by offsprings, but extremely slowly renews from
seeds. The first year sprout exists due to grown in it mycelium and represents
a tiny lump of cells. Only for the second year of life the sprout gives the
first leaf, and begins blossoming only at eight-year age, and even later. But
with the help of seeds the plant was settled at the most distant islands of
Indian Ocean.
Oak
lady’s slipper (Cypripedium quirinus)
Order: Orchidales (Orchidales)
Family: Orchids (Orchidaceae)
Habitat: oak woods in the east of North America.
Orchids represent an original family of floral plants; the significant part
of their species grows in tropics, and such species, as a rule, belong to the
number of the most colourful and/or unusual species of orchids. In human epoch
many of their species appeared threatened or extinct completely. But there are
also kinds of orchids growing in the zone of temperate climate and even settle
into subpolar latitudes. Orchids from areas of temperate climate are ground-dwelling
flowering plants, having, nevertheless, rather bright and beautiful flowers.
In their size, these flowers all the same are inferior to flowers of tropical
forms, but are not without a sort of grace and have attractive colouring.
One of these orchids is oak lady’s slipper, a descendant of yellow lady’s slipper
(Cypripedium parviflorum), rather numerous orchid species of Holocene epoch.
This is rather tall grassy plant – about 40 cm in height, with elongated peaked
leaves arranged on stalk alternately. Leaves in the base of stalk grow 15-20
cm long, having numerous relief nerves protruding from the bottom side of a
leaf; to top of stalk the size of leaves decreases. Stalk bears some slipper-like
flowers (usually 2-3) of pale yellow color, pollinated by various insects. Lateral
petals have numerous brown speckles in the basis. Specialization to pollinators
is low – this plant is pollinated by many species of solitary and social hymenopters,
and also by small beetles.
Fruiting rate is low – fruit develops of one flower from ten on the average.
Fruit of this plant represents a narrow capsule containing dust-like seeds.
After fructification the plant “has a rest” for two years, not forming blossoms.
Plants not produced seeds blossom annually.
This is a perennial plant, but it does not endure cold, therefore its area is
limited by eastern broadleaf forests and mountain oak groves. In mountain areas
this orchid finds especially favorable conditions for life – it frequently “shares”
its symbiotic fungi with oaks, receiving through them some amount of nutrients.
This orchid is more symbiote or commensal and does not harm to trees. Under
the ground, the plant forms a repent rhizome, which forms lateral shoots once
in some years and in due course forms large beds numbering tens large stalks.
Rate of growth of oak lady’s slipper is very low: in first three years of life
the plant feeds exclusively due to mycelium and stays under the ground. At the
fourth year of life of a plant a first overground leaf appears, and at ten years’
age the plant begins to blossom for the first time. This orchid occasionally
eaten by various mammals and insects. The neighbourhood with oaks brings a certain
advantage to a plant: orchid accumulates in its body tannins from their fallen
foliage and even from roots of live tree by means of ground-dwelling fungi,
and it makes a plant less edible for herbivores.
Oak lady’s slipper can reach age of several tens years, quite often living up
to one-century age. Vegetatively breeding clones of a parental plant can live
till 200 years and more.
This species of plants was discovered by Bhut, the forum member.
Sea
coconut (Cocos maritima)
Order: Arecales (Arecales)
Family: Palms (Arecaceae)
Habitat: Hawaii, other islands of Pacific Ocean, east of Indonesia, sea coasts.

Picture by Biolog
In the antiquity, in human epoch “sea coconut” (“coco de of
mar”) was the name of giant nuts of Seychelles coco palm (Lodoicea), which were
rarely found on the coasts of Indian Ocean. Since no one saw a living palm of
that species for a long time, a legend grew about it that it grows on the bottom
of the ocean. Then the palm of this species was discovered, and the legend remained
a legend.
But in Neocene this legend as if revived: one species of plants has practically
“realized” it into life.
In Holocene coco palm was the most characteristic inhabitant of the coasts of
tropical seas. Where its true native land was located is possible only to guess,
since mankind had substantially enlarged its habitat, and it is difficult to
say unambiguously, whether the coco palm grew on some of its territory initially,
or was it the descendant of run wild plants initially planted by people. Seacoasts
proved to be very convenient places to live: because of the salt contained in
the oceanic water few plants capable of competing with the coco palm grew there.
Because of its endurance this species successfully survived in ice age, although
the area of its habitat was substantially reduced, and even broke into some
isolated regions. Possible, on the atolls of Pacific Ocean, one of the populations
of coco palm was forced to make a step towards the ocean. The island-building
corals began to die out everywhere because of catastrophic change in the conditions
of their habitat, and some islands were washed away by the ocean. In order to
survive, the coco palms began to adapt to new conditions: they began to conquer
the oceanic shallows. The plants at first simply “laid down” stem-first into
water, and endured only short-term flooding. Then some of them began to develop
a new adaptation: thick stilt roots upon which the trunk could remain on surface
of water, and even above it. Gradually, the type of growth changed: the lower
part of the stem began to die off, and the plant remained standing only on its
supplementary stilt roots.
Gradually such palms, after “detaching” from the place where they grew, began
“to crawl” farther into the sea, forming unique associations in the shallows.
Thus the new species of the palms, the sea coconut evolved.
Sea coconut is characterized by one other special feature, untypical for the
palms: this plant forms additional buds on the stem, from which new shoots germinate.
This way one palm can form rather wast thickets, which successfully oppose the
sea surf. The older the plant gets, the greater is the number of its crowns,
and farther it penetrates into the sea.
Sea coconut preserved the recognizable appearance of the representatives of
its genus. This palm has long (up to 4 - 5 meters and more) featherlike leaves.
On the edges of leaves special salt-removing glands appeared, which release
the surplus of salts, which penetrate into the plant via its roots. Thick brine
excretes on tips of leaflets and dries out like semi-transparent clusters. When
the wind swings the tree, the clusters of salt fall off from the leaves and
simply fall into the water.
The flowers of sea coconut are small and are unprepossessing as in the other
palms. The fruits have the characteristic “nut-like” appearance. They ripen
in large clusters - by ten and more. The “nuts” of sea coconut are similar to
ones of true coconuts, but are smaller (they are not larger than an apple in
size), covered with durable shell with felt fibrous surface, which makes it
possible for them to float longer over the surface of water. Sometimes the fruit
even germinates, while floating in the sea and during the storm an already finished
“seedling” of the palm is cast ashore. The young plant is very hardy: it can
endure flooding by tidal waves. Some nuts, after being tangled in stilt roots
of adult palms, germinate there, and develop normally, taking root on the sea
floor, enduring the tides and strengthening by its growth the palm “groves”
in the sea.
Sea coconut begins to blossom and to bear fruits approximately at the age of
ten years, and it can live up to 200 years. Usually, the old palms perish during
the storms and the tsunamis.
Translated by Bhut.
Rapa
Nui coconut palm (Cocos paschalis)
Order: Arecales (Arecales)
Family: Palms (Arecaceae)
Habitat: Easter Island, coastal zone.
A descendant of the coconut palm (Cocos nucifera), which was introduced in Easter
Island during the Holocene. With the disappearance of the humans, these plants
were capable of not only surviving in the Neocene, but also of giving rise to
descendants.
The Rapa Nui coconut palm is a large palm, growing up to 35 metres tall, with
pinnate leaves 4 to 6 m long, and pinnae 60 to 90 centimetres long; old leaves
break away cleanly, leaving the trunk smooth. These plants produce their first
fruit in six to ten years, taking 15 to 20 years to reach peak production.
The palm produces both the female and male flowers on the same inflorescence;
thus, the palm is monoecious. The female flower is much larger than the male
flower. Flowering occurs continuously; the plant is not specialized to pollinators.
Like their ancestors, the fruit of the Rapa Nui coconut palm is a drupe, not
a true nut. Like other fruits, it has three layers: the exocarp, mesocarp, and
endocarp. The exocarp is the glossy outer skin, usually yellow-green to yellow-brown
in color. Both the exocarp and the mesocarp make up the “husk” of the coconut,
while the endocarp makes up the hard coconut “shell”. The endocarp is around
4 millimetres thick and has three distinctive germination pores on the distal
end. Two of the pores are plugged, while one is functional.
The interior of the endocarp is hollow and is lined with a thin brown seed coat
around 0.2 mm thick. The endocarp is initially filled with a multinucleate liquid
endosperm. As development continues, cellular layers of endosperm deposit along
the walls of the endocarp up to 11 mm thick, starting at the distal end. They
eventually form the edible solid endosperm which hardens over time. The small
cylindrical embryo is embedded in the solid endosperm directly below the functional
pore of the endocarp. During germination, the embryo pushes out of the functional
pore and forms a haustorium inside the central cavity. The haustorium absorbs
the solid endosperm to nourish the seedling.
The fruit is elongated and triangular in cross section. It is also smaller than
that of their ancestors, its length reaches 35 cm.
This plant species was discovered by João Vitor Coutinho, Brazil.
Dendém
oil palm (Elaeis dendem)
Order: Arecales (Arecales)
Family: Palms (Arecaceae)
Habitat: tropics of South America, river valleys.
During the Holocene, the man introduced many species, animal and vegetal, for
different purposes. The African oil palm (Elaeis guineensis) was introduced
in many regions to the production of vegetal oil. The species introduced on
South America survived on the Neocene, giving raise to the dendém oil palm.
The word dendém was used as a popular name for its ancestor in Brazil. This
plant is native to the tropical regions of South America, being commonly found
near rivers of Central and Northeastern regions of the continent, together with
the riverine almond (Neoterminalia
riparia) and the violet trumpet
tree (Tabebuia violacea).
Mature palms are single-stemmed and grow to 20 m tall. The leaves are pinnate
and are 5 m long. A young palm produces about 30 leaves a year. Established
palms over 10 years produce about 20 leaves a year. The flowers are produced
in dense clusters; each individual flower is small, with three sepals and three
petals.
The palm fruit takes 6 months to mature from pollination to maturity. It is
reddish-yellow, measuring 2.5 cm, and grows in large bunches. Each fruit is
made up of an oily, fleshy outer layer (the pericarp), with a single seed. Animals,
such as the psittacids and monkeys eat these fruits. Some monkeys use tools
to crack the seeds.
This plant species was discovered by João Vitor Coutinho, Brazil
Mole
flower (Talpanthus subterraneus)
Order: Arales (Arales)
Family: Arums (Araceae)
Habitat: forest areas of Europe.
Deciduous forests in areas of temperate and subtropical climate are a place
where change of seasons causes significant distinctions in conditions of inhabiting
of plants and animals. In summer tree leaves almost completely trap sunlight,
and underbrush is very shady – only shade-enduring plants can grow there. An
autumn the significant part of foliage falls down, and within approximately
half-year light exposure of underbrush becomes much better. But winter colds
interfere with growth of grassy plants, and therefore time of active vegetation
of photophilous grasses lasts till some spring weeks. Representatives of this
ecological group of plants are known as ephemers (annuals) and ephemeroids (perennial
plants).
In spring there are few flowers in forest, and after wintering insects search
for them actively. Any flower had appeared in the beginning of spring will be
necessarily visited by insects. But frosts, which may kill flowers, prevent
to plants to blossom too early. Nevertheless, one of plants of European forests
has adapted to blossom early in spring.
When snow thaws and the top layer of ground gets warm a little, under the ground
thick knobby rhizome awakes. From the last autumn on it large flower buds developed,
and they waited for spring warming. When the ground gets warm enough, flower
buds sprout. Their cover representing a modified leaf (spatha) punches a layer
of ground and last year’s foliage, and grows on height of about 30 centimeters.
Spatha is densely curtailed in tube which has yellowish-white color from outside.
At top spatha opens, and its inner side – meaty-red with grayish spots – becomes
visible. Spots are covered with hairs reflecting ultra-violet light, therefore,
insects, not distinguishing red color at all, notice inner side of spatha, and
move right to it. When spatha opens, the putrefactive smell is emitted in air,
and beetles – the basic pollinators of this plant – willingly gather to it.
The most interesting feature of this plant is the position of its flowers below
the ground level – hence this plant has got the name “mole flower”. Tiny and
homely flowers of this species are gathered in spadix inflorescence, which has
no flower-bearing stem and develops directly on rhizome. The spatha forms a
tube through which insects reach the underground inflorescence, and hairs directed
downwards keep insects near the inflorescence while pistillate flowers will
lose a susceptibility to pollen. After fertilization the top part of spatha
dies off, and the bottom part serves as protection of developing infructescence.
The infructescence of mole flower ripens an autumn. It represents strongly grown
up spadix with sweetish amylaceous pulp, in which seeds are immersed. The bottom
part of spadix starts to grow actively, and the infructescence becomes taken
out to the ground surface, where it is eaten by birds and mammals – seed carriers
of mole flower.
In middle of spring, when flowering is finished, the mole flower takes out to
the ground surface the large bud, in which some leaves curtailed together are
contained. The leaf of mole flower has rounded outlines; it is large (its diameter
is about 40 – 50 cm), deeply dissected to three, five or seven lobes. At young
plants did not blossom yet, leaves are integral or triple, and at mature ones
with good advanced rhizome they reach complete development. The surface of mole
flower leaf is smooth and dark green with lighter nerves. Leaves die off in
first half of summer. During the vegetation this plant has time to stock enough
nutrients for formation of new flower buds and for ripening of infructescence.
The young plant of mole flower begins to blossom for the first time at the third
year of life, and in northern parts of an area at the fourth, and even at the
fifth year of life.
At the north of Europe and in the Alpes the close species of plants lives –
serotinous
mole flower (Talpanthus tardus). At this species flowering and fructification
are separated by winter colds, and vegetation lasts during the whole summer.
Feature of this species is the blossoming of serotinous mole flower at the beginning
of an autumn. In winter this plant stays in rest condition, and the last year’s
infructescence appears from under the ground in late spring. This species has
large leaves of dark green color, only slightly cut on edges.
Sea
taro (Paracolocasia transpacifica)
Order: Arales (Arales)
Family: Arums (Araceae)
Habitat: tropical zone of Pacific Ocean - islands, soasts of Asia, Meganesia
and New World.
Taro (some species and numerous varieties of Colocasia genus) was one of the
most ancient food plants cultivated by people. Range of this culture in historical
epoch included tropics of both hemispheres – continents and many ocean islands.
After human disappearance, many cultural plants have become extinct completely;
however, in some places taro has successfully grown wild and has shown significant
competitiveness in formed ecosystems of early Neocene. In tropical areas of
the Earth there are various descendants of cultivated taro. One of their species
is widely settled at the islands of tropical zone of Pacific Ocean – it is sea
taro.
The present species is a large grassy plant with large underground tuber weighting
up to 4-5 kg. An adult plant forms a crown of 8-10 leaves on petioles up to
2 meters long. Leaf blade is heart-shaped with extended tip, of dark green color.
Leaf surface is naked and leathery. Leaf petioles are green with greyish-white
spots – large in the basis and small closer to leaf blade.
At present plant species there is an interesting way of settling enabling it
to colonize new habitats. Tubers of sea taro easily endure travel in salt water
due to a dense tegument. The core of old tuber is cave, at small tubers it is
filled with friable starchy pulp. Plants from coastal populations frequently
appear in the sea after severe storms. At floating tuber leaves quickly die
off and the skin becomes denser. Due to currents and storms, such tubers are
capable to cross hundreds kilometers in the sea, due to what they settle. Growth
rate at such tubers is rather active. Having got on the coast, they sprout in
some hours, and in one week have the advanced root system and a crown of 2-4
medium-sized leaves. Sea taro prefers for growth sandy coasts at absence of
shading by other plants, therefore the present species frequently appears among
pioneer plants. Having established on the coast, the plant breeds actively by
stolons and quickly forms thickets at the coast. Tubers frequently get with
currents far to the north and the south, to temperate latitudes, where they
develop successfully in the summer, but usually perish to winter.
This species in an entomophilic plant. Spatha of an inflorescence is greenish-white
from outside and meaty-red from within. An inflorescence is a thin spadix, like
at ancestral species, emitting an unpleasant smell. The plant is pollinated
by flies and small beetles – the most widespread pollinators present actually
everywhere. Seed breeding usually takes place on large islands, where the races
differing in size and colouring of leaves appear. Plants from coastal populations
of various islands are more similar to each other.
Madagascar
xenocolocasia (Xenocolocasia madagascarensis)
Order: Arales (Arales)
Family: Arums (Araceae)
Habitat: Madagascar, seasonally drying marshlands of western part of the island.
In human epoch taro (Colocasia spp.) was one of the most ancient and widespread
cultural plants, and it was widely cultivated in tropics of Old World, including
the remote oceanic islands. Significant variability and wide range have enabled
a plant to sustain in Neocene flora of tropics and to form a number of high-grade
species. Despite of domestication and changes accompanying it, taro appeared
competitive enough in relation to island flora of Madagascar, and in early Neocene,
it has occupied the ecological niche in local flora.
The descendant of taro in wetlands of the island is Madagascar xenocolocasia,
a large grassy moistureloving plant. The stalk of this species represents a
perennial tuber weighting up to 10 kg, developing under the ground and covered
with thin rough skin. From parental tuber stolons up to 2 meters long grow,
on which tips daughter tubers develop. Due to active vegetative breeding, the
plant forms extensive thickets in favorable conditions.
On the surface of the ground the plant forms thick pseudostem, which consists
of leaf petioles strongly expanded in the basis and nested one into anothers
as at a banana. Thickets of this species look similar a little to banana plantation
because of large leaves up to 3 meters long having narrow sagittate shape. The
length of a leaf exceeds 5-6 times its maximal width. Leaf blade is leathery
and smooth, having two ear-like appendages in place of attachment of leaf blade
to petiole and thin long dropper on the tip. Colouring of an adult leaf is bright
green with a brownish zone along midrib; young leaves are pink, in process of
development, they become brownish, and further turn green. Leaves and tubers
of the plant have burning taste; however local herbivorous mammals eat them.
In tubers of plant grubs frequently develop.
The plant has well defined seasonal vegetation with a rest season during the
drought. At drying of the marshland, leaves of a plant quickly die off, and
tubers is retracted deep into the ground due to drying of roots. In a new vegetation
season the young part of tuber grows on its top.
Flowering takes place in the middle of rain season, when the plant has formed
all leaves of the current season. Inflorescence is a thin spadix with a meaty
red spatha. This plant is pollinated mainly by beetles. Fruits are sweet red
berries; seeds are dispersed by birds and mammals.
Life expectancy of a plant is up to 10 years.
“River
daisy” (Spongiolemna gigantissima)
Order: Arales (Arales)
Family: Duckweeds (Lemnaceae)
Habitat: subtropical and tropical areas of Far East and East Asia, Japan Islands.
Duckweeds are the most specialized and most strongly changed family of flowering
plants. Their body has turned to frond – the original alloy of leaf and stalk.
Some duckweed species may lack of roots. Flowers of duckweeds are reduced up
to pestles and stamens covered with thin wrapper.
Despite of such deep degeneration of body, duckweeds are prospering group of
plants. Due to high growth rate and ease of vegetative breeding duckweeds form
continuous cover at the surface of reservoirs, and also are the important food
resource for water animals.
In Holocene duckweeds were submitted by tiny plants, and to this family the
smallest plant of the Earth belonged – Wolfiella of only about 0.5 mm long.
In Neocene family is also submitted mainly by tiny plants poorly changed in
comparison with human epoch. But among them some specialized forms had evolved,
including one original giant. It is “river daisy”, the aquatic plant floating
on a surface of the rivers and lakes of East Asia. At first sight “river daisy”
can be confused with other plants – floating leaves of water lilies. But at
close examination the difference becomes obvious. Rounded fronds of this species
remotely similar to leaves of water lilies have diameter about 15 – 20 cm. Their
edges are slightly cut up and wavy, blades are inflexed a little, and frond
of plant resembles the stylized image of flower. Body of plant in the bottom
part is spongy and has advanced aerenchyma. Because of numerous air bubbles
it has silvery colouring. The top side of frond is dense and leathery, and has
bright green color. One adult plant is able to grow on its edges some new fronds
simultaneously. Buds, from which they form, develop on edges of frond blades.
At the bright sunlight young fronds redden. At northern edge of an area in autumn
the plant produces small turions similar to fronds of common duckweed species.
They float at the surface of water and gather at the coast. In winter adult
plants die off, and turions immerse to the bottom where winter successfully.
Spring warming and amplification of light exposure stimulates their development.
Growth rate of this plant is very great. The development of one frond from occurrence
up to reaching of full size takes about one week, and on one adult frond simultaneously
up to seven fronds of new generation may develop. On the bottom side of frond
“beard” of thin not branching roots reaching length of 10 cm grows. They actively
absorb organic substances dissolved of water due to what this plant effectively
clears water.
This plant is dioecious. Male plants differ in smaller size, but breed vegetative
more intensively, and the biomass of male and female individuals in population
is approximately identical. Flowers of “river daisy” are reduced, as at all
duckweeds, but differ in special structure. Pestles and stamens at them are
very long and strong; they are covered at the basis with small spatha leaflet,
which dries up soon. Flowers develop between blades of frond. “River daisy”
is an entomophilous plant, but its relations with insects are considerably simpler,
than at orchids, for example. Damselflies like to sit on pestles and stamens
sticking up above thickets of “river daisy” for rest. They carry sticky pollen
of this plant on legs. Seeds of “river daisy” are tiny, covered with sticky
slime. They float on water surface for a long time, and stick to feathers or
wool of water tetrapods, which settle this plant.
The idea about the opportunity of evolving of such plants is proposed by Nem, the forum member.
Module
duckweed (Heterophyllemna integra)
Order: Arales (Arales)
Family: Duckweeds (Lemnaceae)
Habitat: Eurasia, marshes and lakes in temperate and subtropical zones.
Duckweed family shows a special direction of evolution – the tendency to extreme
simplification of structure of a body, which at them has turned to frond, formed
of stalk and leaf. Simplicity of body structure, rapid rate of growth and breeding,
and also simplicity of settling and ability to adaptation – these qualities
appeared essential for a successful survival of duckweeds in Holocene epoch.
In Neocene these plants have gone through a new coil of evolution – complication
of body structure via specialization of fronds and formation of a modular design
of a uniform body of a plant. Modular duckweeds are descendants of Spirodela
duckweed widespread in human epoch. A new group of modular duckweeds is characteristic
for reservoirs of Eurasia in various climatic zones, except for the coldest
areas. Like their ancestors, these are the plants floating on water surface.
Module duckweed from freshwater reservoirs of Eurasia shows a characteristic
structure of a body of plants of this group. This is a small plant not exceeding
15-20 mm in diameter, with a compound body of fronds of different structure
and purpose. The basic frond is the main thing in structure of a plant. It takes
an active part in vegetative breeding, forming buds, from which new fronds and
wintering turions capable to survive in winter under ice develop. In it, a stock
of starch spent during the wintering is also stored. This frond lives for two
years, developing from a bud. Fronds of all other types form from it. It takes
part in vegetative breeding in growth season.
Some fronds on edges of the basal one are the specialized bodies of photosynthesis.
They are thin and transparent, lack roots and are raised above water, working
as sails and bodies of photosynthesis. Due to insignificant thickness, photosynthesis
takes place in the whole volume of these fronds.
The specialized type of frond is a generative or flower-bearing frond – a bladder-like
frond having the transparent top side, inside which groups of flowers develop.
When reduced flowers develop completely, they break through a transparent membrane
and are pollinated by tiny insects. This frond is supplied with a stock of starch
in its bottom part; after pollination it breaks off a parent plant and spends
all stocked resources for ripening of seeds, then dies off and seeds get in
water. They are spreaded by streams of water or waterfowl, sticking to their
plumage.
To winter photosynthesizing fronds die off and resolve, and on the basic frond
of the second season of growth wintering turions are formed; they disengage
it and sink. Resources of this frond are completely exhausted for their formation,
and it dies off. Fronds of the first year of life winter under water, forming
only separate flower buds.
Greenhouse
schizolemna (Schizolemna anabaenophila)
Order: Arales (Arales)
Family: Duckweeds (Lemnaceae)
Habitat: Eurasia and North America, lakes in a zone of cold climate.

Picture by Biolog
Plants of duckweed family have successfully gone through human
epoch due to their unpretentiousness in a choice of habitats, fast breeding
rate and ease of settling. Simplification of body structure has enabled them
to cut down expenses for maintenance of ability to live up to a minimum, channeling
all the power to breeding. In due course of evolution in early Neocene among
duckweeds the species using different life strategies have appeared. One successful
evolutionary course is symbiosis of one duckweed species with cyanobacterium
of Anabaena genus. This step has enabled duckweed to grow in oligotrophic
lakes due to ability of cyanobacterium to fix nitrogen from an atmosphere and
to use it for its own needs. Greenhouse schizolemna duckweed has given symbiotic
bacterium favorable conditions for life in exchange for a part of the nitrogen
received by it. Specialization to symbiosis has resulted in change of appearance
of this species.
An adult individual of greenhouse schizolemna represents disc-like or oval frond
about 10 mm in diameter. The middle part of this frond is translucent, and under
a layer of integumentary tissues of a plant friable parenchyma lays, in which
cyanobacterium cells live and breed. In summertime the plant becomes as if a
“greenhouse” for cyanobacterium (hence a specific epithet), and it breeds plentifully
inside a plant body. Chlorophyll-bearing tissues of the duckweed itself are
kept only as a margin along the edge of the frond. In the summer schizolemna
breeds asexually by formation of fronds from sides. The process of formation
of new frond is original: it separates from the edge of an adult plant, taking
a part of “greenhouse” with symbiotic cyanobacteria. Also frequently a simple
dividing of the adult frond via the formation of a cross partition takes place:
it simply falls apart in two halves along it – hence the name: “schizo-” means
“to split”.
Due to symbiotic cyanobacteria greenhouse schizolemna is capable to grow in
oligotrophic lakes above the Arctic Circle, using advantages of polar day to
the full. This plant is an important fodder resource for local waterfowl.
In an autumn plant forms wintering turions inside old frond, which dies off.
Turions keep inside spores of cyanobacterium, from which the new population
of symbiotes develops. In the spring after ice thawing, turions emerge; on legs
of waterfowl they settle to new reservoirs, where their growth begins.
Gymnosperms |
Nearctic
pine (Pinus nearctica)
Order: Pinales (Pinales)
Family: Pines (Pinaceae)
Habitat: North American forest-tundra.
Relations between forest and tundra had never beed unequivocal. When the world
was enveloped by global warming, tundra receded to higher latitudes, and forest
advanced into its former possessions; when an ice age began, the situation was
opposite. The beginning of Neocene was marked by sharp receding of forests to
lower latitudes and by extensive congelation. In that time the zone of forest-tundra
was absent almost completely.
But the ice age was over, the climate has become warmer, and the tundra has
receded again. Between it and taiga forest the transitive zone of forest-tundra
populated with plants of both natural zones has appeared again. In North America,
especially at the northwest, in Beringia, one of characteristic trees of forest-tundra
is Nearctic pine.
Nearctic pine is a descendant of jack pine (Pinus banksiana) of Holocene epoch,
and this is a rather small tree – about 4-5 m tall. It grows as light forests
on drier places protected from polar winds, avoiding wetlands and lowlands.
At the windswept places this tree forms creeping elfine wood form, which height
does not exceed half meter. Trunk of Nearctic pine may frequently of irregular
shape, curved hard and looking almost as a huge bush. Branches at this pine
are sprawling, covered with long paired needles and frequently grow from a trunk
at different angles. Bottom branches of plant can frequently raise the main
trunk in growth, forming very wide crone; therefore crones of these plants are
frequently closing and groves of Nearctic pine are always rather shady and gloomy.
The stony ground in such places is covered with a layer of fallen needles, and
because of lack of light the most part of grassy plants can not grow there.
Growth of this tree is very slow, frequently not exceeding several centimeters
per one year.
Despite of small height of Nearctic pine and absence of grassy plants in its
thickets, large polar mammals frequently visit these strange forests. There
they always have an opportunity to hide from a polar wind and there is much
less amount of snow there. In the summer thickets of this pine help animals
to hide from midges.
As well as the majority of coniferous plants, Nearctic pines are pollinated
by wind. Breeding of these plants occurs in the beginning of a polar summer,
and plants produce plentiful dusty pollen. Ripening of seeds takes a lot of
time: cones can develop till five years before seeds – small, lightweight ones
supplied with wings – will develop completely. They are easily carried by wind
to great distances. During the first years of life they grow very slowly and
perish from various herbivores, but are capable to resist colds of polar winter.
As against many conifers, this plant easily survives at damage of an apex by
animals or frosts, and in due course forms branchy tree with a sprawling crone.
The first seeds are formed on this plant at the age of about twenty years, and
life expectancy can make up to three hundred years and more.
This species of plants was discovered by Bhut, the forum member.
Swamp
larch (Larix paludiphila)
Order: Pinales (Pinales)
Family: Pines (Pinaceae)
Habitat: Western Siberia, wetlands.
In human epoch larches (Larix spp.) were the most widespread trees on the Earth
and formed extensive large forests in territory of Eurasia. At the end of human
epoch, their range decreased because of mass cutting down of forests, and further
these trees receded to the south, being pressed by glaciers in time of Holocene-Neocenic
congelation. At the end of an ice age they were among the first plants to occupy
territories, from which ice sheet retreated, and have formed extensive forests
again. In territory of Western Siberia they had to face conditions of life new
to them: plains became boggy because of slow, but a continuous raising of northern
edge of Eurasian lithospheric plate released from mass of glacial sheet. As
a result, a part of population of ancestral species appeared in extensive boggy
territories, where in due course of evolution a new species – swamp larch –
had formed.
This tree species is an original cold-resistant analogue of a North American
swamp cypress (Taxodium), forming extended thickets in boggy sites. Populations
of this species are widespread across the whole West Siberian Plain, except
for its northernmost areas. Swamp larch endures flooding of the bottom part
of trunk and normally grows in reservoirs, which depth may be up to 2 meters.
This species is remarkable with its extremely firm and rotting-resistant wood:
even trunks of dead plants continue standing vertically for a long time. In
the wood bored by insects numerous hollows appear, where forest and marsh birds
nest.
Swamp larch grows up to height of 25-30 meters; trunk diameter of an adult tree
is up to 2 meters. Crone of this tree is rather lax, passing a significant part
of the sunlight down, to the surface of a bog. Outlines of crone are wide conical;
its diameter in the bottom part may reach 15 meters. Needles are gentle and
soft, falling for winter; the length of needles reaches 80-100 mm; they grow
in bunches of 5-8 ones. Young female cones have bright pink color, reddening
and turning brown later. Seeds of this plant ripen within one year and fall
out from cones in the spring, during a high water. Due to a significant amount
of fat oil, seeds float on the surface of water. They are dispersed by streams
of water or stick to legs and beaks of waterfowl. Sprouts of this species easily
endure suppression and grow very slowly in the first years of life. Survival
rate and germinating capacity of seeds are rather low: no more than 10% of the
formed seeds give sprouts.
Root system of this species stretches to great distance from a plant – to 10
meters and more in separate roots. In the basis of trunk 4-5 powerful plank-buttress
roots are present, establishing a plant in viscous ground. As against the majority
of coniferous trees, this species successfully breeds in vegetative way, actively
giving root sprouts at the distance of up to 5 meters from a parental plant.
More often, these root sprouts grow along the external edge of thickets of swamp
larch, and grow much faster, rather than plants from seeds. New trees can rise
to the surface of water from two-meter depth, spending under water up to 5 years
and more, until their tops will appear from water. Shoots formed under water
have a long annual accretion, thin branches and expanded needles. On the surface
of water such plants begin forming shoots typical for this species, and the
trunk begins growing thick quickly.
Life expectancy of swamp larch reaches 500 years. Tree gives its first seeds
at the age of 20 years, and further forms them almost annually. Seasons of the
greatest crop of seeds repeat every 4-6 years.
Eastern
mountain spruce (Picea appalachiensis)
Order: Pinales (Pinales)
Family: Pines (Pinaceae)
Habitat: slopes of Appalachian mountains near mountain tops.
Appalachian Mountains of North America differ in many respects from Rocky Mountains
on the west of the continent. They are much lower, and their slopes are flatter,
because they are much older. Tops of these mountains are deeply destroyed by
aeration, on them plateaus frequently form. The trees growing on slopes of Rocky
Mountains are more often coniferous, than deciduous ones: above on slopes continuous
coniferous woods are formed. Mountain forests of Appalachian Mountains aremixed
and much more diverse in number of species. Typical trees of these forests are
oaks, but conifers also grow here woods. Eastern mountain spruce is a species
of coniferous trees from the east of the continent.
Its appearance is typical for representatives of the genus – it is a tall, slender
tree about 20 m high, growing in one trunk, having a conic crone and wide boughs
reaching the ground. As against spruces of human epoch, it looks disproportionately
thin because of “pendent” branches – width of its crone in the middle part is
about 100 cm; top of tree is peaked.
Branches of mountain spruces are long and narrow, but have numerous lateral
branches, with wide and long needles 20-40 mm long. Needles have bright green
color, being pointed and densely growing. Branches form rather dense crone,
which almost does not pass sunlight, therefore roots of mountain spruces have
no contenders in extraction of nutrients and water from ground, but small animals
often arrange shelters under this tree – under such spruce there is always rather
warmer and less snowy even in windy mountain winters. Due to a narrow crone
the tree easily dumps from its crone snow accumulating on it.
Eastern mountain spruce is pollinated by means of wind, as well as others conifers,
and its seeds are dispersed by animals. This tree has short thick cones – 5-10
cm long – and proportionally large and heavy seeds lacking wings. They are large,
rich in nutrients, mainly in fats, therefore serve as food to various rodents
and small birds. Due to them a part of seeds appears dug in the ground as a
stock for winter. If the owner has forgot about the stocks or had been caught
by a predator, in the spring the hidden seeds sprout, providing reproduction
of species.
Enemies of this kind are mainly insects and fungi causing diseases and death
of plants of this species across wide areas. Herbivorous mammals only occasionally
eat tops of these plants during winter food shortage, but nutritional value
of them is very small. After damage of top the tree is sick and is restoring
for a long time. If mountain spruce will be not not damaged by herbivores at
young age, it quite can live for some hundreds years. Between this spruce and
Eastern mountain oak a constant competition
takes place – oak is more photophilous, and its sprouts can not develop among
shady crones of mountain spruces, but its young growth supersedes spruce from
more fertile ground areas.
This species of plants was discovered by Bhut, the forum member.
Pacific
thuja (Thuja pacifica)
Order: Pinales (Pinales)
Family: Cypresses (Cupressaceae)
Habitat: humid Pacific forests of North America.
In Neocene to the west from Rocky mountains of North America a very unusual
ecosystem has developed: rainforests with a temperate climate. From ocean the
rain clouds come regularly, providing a high level of humidity of air and ground.
In these forests a natural community of moistureloving kinds of plants and animals
is formed. One of typical species of trees determining the appearance of these
forests, is Pacific thuja – hardy and moistureloving coniferous plant.
Pacific thuja is a descendant of Pacific red cedar (Thuja plicata) of Holocene
epoch. Like its ancestor, it is a very tall tree growing up to 60-65 meters
in height, with numerous roots growing deep into the ground and helping this
plant to resist impacts of elements – wind, flooding, and mudflows on hillsides.
Additionally, roots of these trees play the important role in stabilizing of
hillsides, and mighty trunks stop the large stones rolled from slopes. Trunk
at this tree is smooth and straight, covered with reddish small-grooved bark;
branching at an adult plant begins at significant height – in the top third
of trunk. Branches form wide conic crone on the top of tree. Branches are long
and rather direct; photosynthesizing shoots are covered with wide scale-like
leaflets, on which surface the thinnest layer of wax is secreted, helping rainwater
to fall down. Leaflets are much wider, than at any other thuja species in human
epoch. Due to the contents of resinous substances all parts of a tree smell
pleasantly.
Pacific thuja plentifully forms pollen in the beginning of summer, when spring
rains are over, and weather becomes rather dry. This tree not only is pollinated
by wind, but also settles with its help: its cones are proportionally small,
up to 10 cm long, made of 4-5 pairs of large ligneous scales. Seeds are small
and easy, having a wide wing and are dispersed by wind. Western coast of North
America is a windy place and though in underbrush of Pacific forest the wind
is felt slightly, at height of crones of Pacific thuja presence of a wind is
felt clearly, and both pollen and seeds of this tree travel very long distances
– enough for an exchange of a genetic material between the remote populations.
This species is a very durable tree, but a payment for longevity is low survival
rate of sprouts. Its sprouts are initially very small, and they grow slowly
– until the plant becomes strong enough, it has a lot of enemies from among
fungi and insects. Young plants in a underbrush frequently grow in a depressed
condition, being in a shadow of adult trees. If the young Pacific thuja manages
to survive in first ten to twenty years of life and to reach even 5-10 meters
in height, there is a high probability, that it can enter the forest canopy
woods and live for some centuries – age of adult specimens reaches 700-800 years,
and even more.
This plant species was discovered by Bhut, the forum member.
Sporiferous
plants - ferns
|
Two-shaped
tongue fern (Epiphyllitis heteromorphus)
Order: Cyatheales (Cyatheales)
Family: Aspleniaceae (Aspleniaceae)
Habitat: warm and humid forests along the western and southern coasts of Fourseas.
Climate warming has resulted in formation at the south of Europe of deciduous
forests having separate features characteristic for tropical forest. But the
area of these forests is very limited – the Mediterranean clutched between Europe
and Africa had turned to salt desert, and the water comes to these forests only
from Fourseas – huge brackish sea-like lake at the southeast of Europe. Warm-
and moistureloving forests cover its southern and western coasts. The plants
making these forests are descendants of local European plants – mountains and
Mediterranean lowlands reliably isolated them from “true” tropical forests.
Besides the features of the trees forming these forests, the presence of epiphytes,
characteristic life forms, gives them tropical appearance. In European forests
of tropical type mosses and ferns are epiphytes.
One of epiphytic ferns of these places is two-shaped tongue fern, the descendant
of common European hart’s tongue fern (Phyllitis scolopendrium) of Holocene
epoch. This species grows in forks of branches, where the needed amount of vegetative
material for germination of its spores accumulates.
Similarly to the ancestor, this species is crown plant forming dense funnel
of fronds. At this species two types of fronds develop – sterile and fertile
(hence the specific epithet). Sterile bottom fronds are rounded, wide and leathery.
In them vegetative dust, bird dung and other substances accumulate, and the
plant receives nutrients from them. The additional roots developing on short
stalk of plant penetrate there. Being young, sterile fronds take part in photosynthesis
for some time, but as they develop, the amount of chlorophyll in them reduces
and suber tissues develop. These fronds turn rigid, and only at their edges
the area of growing tissue stays. The similar type of fronds develops at staghorn
fern (Platycerium), which has successfully survived up to Neocene epoch and
lives in South-East Asia and Meganesia.
Top fronds of two-shaped tongue fern are softer. They photosynthesize and on
their bottom side along lateral nerves, at an angle to central nerve, sporangiums
gathered to linear soruses develop. This species has lengthened, non-divided
and dense fronds – this feature is inherited by it from the ancestor. Fertile
fronds of this fern have slightly wavy edges and rather soft tissue. At short-term
winter cold snaps they may die off. But up to this time the plant forms in the
center of the crown some young fronds, which do not unwrap in winter and cover
a vulnerable apex. They produce slime rich in polysaccarides, which protects
apical tissues of damage by cold. In spring, when it becomes warm enough, they
open, and new fertile fronds grow very quickly. Completely advanced plant forms
the crown about 50 – 60 cm in diameter.
Two-shaped tongue fern strongly attaches against bark of trees by roots easily
growing into roughnesses of bark and penetrating deeper while its grows thick.
At the old part of rhizome new sprouts grow, and one plant can form thickets
of several large crowns.
This plant breeds by spores easily carrying by wind and by insects. But two-shaped
tongue fern has one more way of breeding. Fertile fronds easily form brooding
buds being damaged. Usually the bud is quickly forming at the place of break
of nerves, and the broken off piece of fertile frond also may produce at least
one bud, from which the young plant successfully develops.
Sporiferous
plants - horsetails |
Epiphytic
horsetail (Equisetum dendrophilus)
Order: Equisetales (Equisetales)
Family: Horsetails (Equisetaceae)
Habitat: forests of Balkan and South Europe.
The climate of Neocene differs in heat and humidity. It has resulted in expansion
of tropical and subtropical zones and shifting of borders of heat-loving vegetative
communities areas to higher latitudes. In Europe heat-loving forests had appeared
in southern part, in Alps foothills, at the Balkan and in Asia Minor. To the
south, behind the Alps, the hollow is spread being earlier the bottom of Mediterranean
Sea. This is a place where salt swamps and rigorous saline deserts dominate.
To the south of Mediterranean Lowland savannas of North Africa cut by huge river
named Sahara Nile spread. Extensive plains appeared an absolute obstacle for
heat-loving vegetation of Africa, and mountains have prevented penetration into
Europe of Asian heat-loving plants. Therefore subtropical woods of Europe are
formed by descendants of local plants evolved in favorable conditions: the huge
sea-sized lake Fourseas provides these forests with moisture.
In multilevel forests of south-east of Europe the set of opportunities for evolution
had appeared. Huge trees are twined with lianas and on branches epiphytes live
as in true tropical forest. Basically they are various ferns – from tiny and
delicate up to huge ones with skinny leaves. But among their leaves (fronds)
of various shape long and similar to brushes sprouts of other sporous plant,
epiphytic horsetail, the endemic of the south of Europe, are visible.
It is easier to sporiparous plant to occupy a niche of epiphyte, rather than
to flowering one – spores are easily carrying by wind (orchids growing often
as epiphytes have microscopic seeds with reduced germ). But sporous plants have
one vulnerable feature: from the spore not a sprout of adult plant, but prothallus,
the sexual generation develops. On it feminine and masculine genitals, archegones
and antheridiums develop. And drops of water are necessary for fertilization
and development of adult plant (sporophyte). Therefore sporous plants have reached
especial success in woods with high air humidity.
In early Neocene seas of south-east of Europe had returned to their coasts after
drying up ice age, and later had merged to Fourseas. It had favorably resulted
in climate of Balkan – the climate became more humid and rainy. The ancestor
of epiphytic horsetail, obviously, settled in these conditions in moss cover,
and then became climbing plant. Gradually descendants of this species began
to pass a complete cycle of development in tree crones. The Neocenic epiphytic
horsetail had appeared so.
The epiphytic horsetail represents rhizomatous grassy plant. Its rhizomes creep
on trunk upwards and downwards, hiding from sunlght in cracks of bark. Covers
of rhizomes are thick and spongy – it is a reliable protection against excessive
evaporation of water.
Stalks of epiphytic horsetail are thin and low – not higher than 20 cm. But
this plant forms rich cushion thickets of bright green color. As all species
of horsetails, this plant is lack of leaves, but lateral branches located in
nods of stalk by verticils of ten ones and more. The bottom nods of stalks are
located closer to each other and lateral branches are denser here, rather than
on tops of sprouts – this way inside thickets in growth zone of rhizome and
young sprouts the optimum microclimate is kept.
Rhizomes of epiphytic horsetail have two types. The basic rhizome is rather
short, with internodes pulled together. From it vegetative photosynthesizing
sprouts grow. From the basic rhizome keeping the certain distance lateral rhizomes
of the second type grow. These rhizomes are very thin and have long internodes,
reduced roots and vegetative sprouts. They creep on trunk, and sometimes freely
hang down from branches. Wind shakes such rhizome and throws it to the next
tree. There it is fixed, quickly forming roots clinging to the tree bark, and
starts to grow, forming characteristic green “bottle brushes”. This plant is
settled so.
Sporiferous sprouts of this plant strongly differ from vegetative ones – they
are long (reaching one-and-a-half-meter length), without lateral branches, with
narrow and long strobil on the end. Such sprouts develop in the beginning of
summer. The stalk of sporiferous sprout is elastic; therefore sprout is arcuated
and hangs, being shaken in the wind. The plant simply uses one behavioural feature
of flies – these insects like to perch on hanging down thin objects. Not only
wind, but also insects carry spores of epiphytic horsetail. Spores with outgrowths
(elaters) cling to legs of flies and are delivered by insects to places where
the wind could not bring spores. This habit of spores carrying gives appreciable
advantage in settling to the epiphytic horsetail.
In the best way the spores had got in damp places (for example, on the friable
bark of tree holding moisture) develop. From spore small prothallus similar
to hat with cut up edges grows. On its convex middle part the masculine genitals
(antheridiums) are located, and edge lobes female archegones are. Fertilization
is promoted by the form of prothallus: the dew drop gathers on antheridiums,
spermatozoids get in it, and then the drop simply rolls down to archegones.
Sometimes from connected spores some prothalluses growing near each other are
formed. Even the cross fertilization giving more viable plant is possible in
this case.
Young plant at the first years of life forms only sterile sprouts and does not
give long rhizomes with which help it is settled. Approximately at the fourth
year of life at it first sporiferous sprouts are formed and got stronger plant
at the age of about eight years starts to settle with the help of rhizomes.
It forms set of clone thickets stretching to tens meters in forest canopy.
The close species, hanging
wood horsetail (Equisetum pendulus) as if keeps features of the previous
stage of development of epiphytic horsetails. This species grows on bark and
keeps against it by roots sprouting deeper in process of its thickness increase.
Vegetative sprouts of this plant freely hang down, reaching the length of meter
and more. Their lateral branches are thin and long, therefore this horsetail
looks fluffier. Strobils are formed on ends of sprouts which are not distinguished
externally from vegetative ones. Strobil of hanging horsetail is long, and spores
of this species are also carried not only by wind, but also by flies.
Siberian
sweet horsetail (Equisetum dulcis)
Order: Equisetales (Equisetales)
Family: Horsetails (Equisetidae)
Habitat: wetlands of Western Siberia, river valleys.

Picture by Biolog
In human epoch horsetails represented rather conservative
and poor in species diversity group of sporous plants. Nevertheless, many species
had rather extensive range, and some species coexisted with people successfully,
becoming weeds in agricultural areas. Human activity and climatic changes of
the boundary between Holocene and Neocene have resulted in large-scale changes
in ecosystems, which resulted in active speciation of survived species, among
which there were horsetails also.
At the territory of wetlands of Siberia one new horsetail species of Neocene
epoch grows – Siberian sweet horsetail. This is a grassy perennial plant seldom
reaching a height of 1 meter; usual height of vegetative shoots is about 50-60
cm. These shoots bear whorls of short lateral branches – up to 15 ones in one
whorl. Stalk is brigh green, brownish in nodes.
This species has repent underground rhizome storing starch and other polysaccharides
to winter. It is knotty and easily breaks up, when large mammal digs it out
from the ground, and thus some parts of rhizome avoid being eaten. In a season
of vegetation from a rhizome vertical shoots of two types grow in turns. After
snow thawing from the ground fleshy nonchlorophyllic shoots of light pink color
appear, bearing on their tips starchy spore-bearing strobili containing large
spores with dense environments. Height of such shoots is up ti 10-15 cm. Spore-bearing
strobili have juicy starchy pulp with the expressed sourish-sweet taste; in
the spring they are willingly eaten by rodents and gallinaceous birds, which
disseminate spores in their feces. After a high water the period of active growth
of vegetative shoots begins. They are annual, dying off for winter. Tissues
are rich in silica; therefore herbivorous mammals reluctantly eat green parts
of this plant.
This species is a perennial plant breeding actively in vegetative way. Due to
carring of spores by birds and mammals in populations of this plant a probability
of joint growth of genetically diverse plants and occurrences of durable posterity
in them are high.
Brown algae |
Leafy
sea duckweed (Algolemna prolifera)
Order: Fucales (Fucales)
Family: Algolemnaceae (Algolemnaceae)
Habitat: Pacific ocean to the north of equator, surface of water.
Picture by Biolog
In human epoch large brown macroalgae were presented by bottom-dwelling
forms attached to a firm substratum with the help of rootlike outgrowths named
as rhizoids. Thalli of these species frequently reached huge size, extending
to many meters. In Neocene one group of brown algae has made a real breakout:
they separated completely from sea bottom and have turned of plants floating
on the waters surface and forming a separate algal family. For the approximate
similarity to floating plants of fresh waters they have got the name “sea duckweed”,
though appreciably differ from them in large body size.
Thallus of sea duckweed is clearly divided into two parts: large rounded float
with leathery environment filled with spongy air-bearing parenchyma, and a photosynthesizing
part – leaf-like outgrowths of thallus growing fast on sides of a float, for
which the plant is named as leafy sea duckweed. Diameter of a float of this
species is up to 40-70 mm, and length of leaf-like outgrowths is up to 20 mm.
Colouring of plant tissues is reddish-brown, and outgrowths are transparent.
At the edges of a float “apical points”, where a formation of new daughter float
takes place, form from time to time. Their arrangement is not fixed, and growth
of daughter individual can begin in any place. The adult plant can form up to
three floats simultaneously. The old plant can break half-and-half: each half
completes missing parts within several days. Usually plants of this species
may be met one by one or with not completely generated shoots. During storm
due to small size and weight plants actually are not damaged by waves – only
lateral sprouts break off. In silent weather sea duckweed grows intensively,
forming extensive thickets. Breding of these plants is mainly vegetative – by
separating of daughter plants. Growth very rough: plant doubles its biomass
every three-four days.
Breeding via spores at sea duckweed occurs in regular way. Life cycle is diplophasic:
the haploid generation is absent. Antheridii and oogonii develop on edges of
a float between shoots of thallus and free sexual cells into the water, where
fertilisation and development of young plants take place.
Sea duckweed is an important component of oceanic biocenoses, giving shelter
and food to small sea animals. Plants represent a substratum for polyps and
sedentary crustaceans. Pelagic fishes and small cephalopods also lay eggs on
them. Presence of these plants settling with sea currents facilitates transoceanic
settling of sea animals.
A close species lives in Southern hemisphere: fringed
sea duckweed (Algolemna marginalis), at which lateral “leaflets” of
thallus are short, forming only a fringe along edges of floats. Diameter of
a float at this species reaches 30-40 mm. It is more cold-resistant, in equatorial
waters grows quickly, but degrades in due course. Populations of this kind live
in waters of the currents washing coast of Antarctica, but currents frequently
carry them to equatorial waters.
In Indian Ocean mermaid’s
griddle-cake, or flattened sea duckweed (Algolemna platycystis)
lives – a species having wide floats flattened both from above, and from below,
and filled with a spongy tissue. Diameter of such float may be about 130-150
mm at thickness of up to 12-20 mm. Its surface is leathery from above and lumpy
from below, of yellowish-brown color. Daughter plants at this species keep very
hard on parental thallus, therefore present species forms sometimes “carpets”
of one parental plant, several daughter and numerous granddaughter ones, which
break up only during a violent storm.
Green algae |
Mediterranean
filodunaliella (Filodunaliella mediterranea)
Order: Chlamydomonadales (Chlamydomonadales)
Family: Dunaliellaceae (Dunaliellaceae)
Habitat: hypersaline waters of the Mediterranean basin.

Picture by Biolog
In human era, very few algae could live in hypersaline water.
The most known genus of such organisms was Dunaliella – the unicellular motile
green algae, very similar to Chlamydomonas, and often causing water bloom. In
Neocene, their descendants have undergone significant changes to master new
ecological niches. Filodunaliella mediterranea has mastered the hypersaline
water bodies of Mediterranean basin, where it thrives, being almost rid of enemies
or competitors.
Filodunaliellas became filamentous algae from unicellular – they form short
chains of 4-5 oval cells enclosed in a collective slimy-oily capsule. The cells
are about 10 μm in diameter, the filaments are around 50 μm long, and one of
two terminal cells bears a polar tuft of flagella, their rotation pulls the
chain of cells forward.
Each cell, like the ancestors, contains a large chloroplast with a spherical
pyrenoid and starch grains, and also a large amount of glycerol which protects
the cell from the osmotic action of the brine. In the ancestral forms, the chlorophyll
in the chloroplast was usually masked by a large amount of carotenoids for protection
from UV, giving the cells a red-orange color. In filodunaliellas, their content
is lowered: in its habitat it is covered by the plates of “blood-of-the-salt”
halobacterium (Halobacterium sanguinisalis) and often clings with them.
For this reason, its cells are golden-yellow in color.
Filodunaliellas reproduce the same way as its ancestors: by mitotic division.
In adverse conditions (dehydration, desalination), the cells within a chain
form zygospores by pairs, and unpaired cells die. Zygospores have a thick resistant
envelope, which allows to wait a long time for favorable growth conditions to
be restored, and are easily carried by wind and animals. In new places, they
germinate with many new cells that divide to form new chains. Large numbers
of zygospores of this species are found along the shores of saline lakes and
at the bottom of dried water bodies.
Production of large amounts of glycerol by filodunaliellas provides the life
conditions for halobacteria (a carbon source). During a rapid development of
filodunaliellas jointly with halobacteria, a dark blood-red color of the brine
is observed, and without the halobacteria it is golden-yellow. In both cases,
the water surface can become oily due to the glycerol.
This species of plants was discovered by Biolog, the forum
member.
Translated by Biolog.



