Animalcularium of the future - Protists and other microorganisms


Animalcularium of the future - Protists and other microorganisms |
|
| In this section descriptions of various animals and plants, which could live on the Earth in Neocene epoch. The section will be supplemented as new ideas about possible ways of evolution of life will appear. If readers will not find here any species placed here earlier, it means that it is a reason to search for a new chapter in English version of "The Neocene Project". | |
|
Microorganisms |
Ciliates |
Sousaphone
(Tubafon suzafon)
Order: Heterotrichida (Heterotrichida)
Family: Stentoridae (Stentoridae)
Habitat: fresh waters of the Northern Hemisphere, in temperate and warm climate
zones.
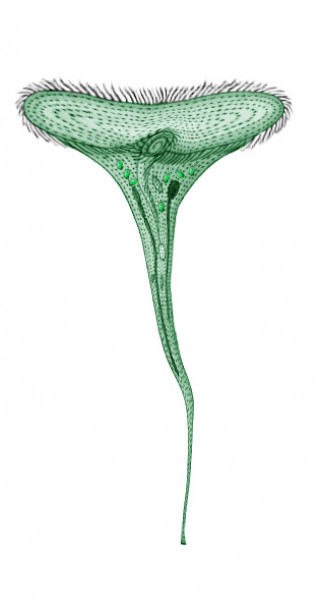
Picture by Biolog
Small living funnels with fans of cilia, sitting on underwater
objects (rocks, plants etc.), can be found in still fresh waters of small water
bodies and river backwaters of Neocene. These are sousaphones, the descendants
of Stentor species of the human era.
Sousaphones are very large (up to 15 mm high) ciliates, resembling their ancestors.
The stalk is hair-thin at the base, but gradually thickens, and at a height
of about 10 mm merges into a funnel, which can stretch up to 13-14 mm in diameter.
The funnel with the cytostome in its bottom is surrounded with rows of cilia,
which can reach 10 mm long. Sousaphones effectively yield their organic food
(bacteria, algae etc.) from the surrounding water by beating of such fans.
Sousaphones have inherited the contractile ability of the cells from their ancestors
and have brought it to perfection. A disturbed ciliate first fulminantly coils
its stalk in one plane (now resembling the sousaphone instrument, after which
it is named), and then shrinks into a ball about 5 mm in diameter. When necessary,
sousaphone can leave its substrate to swim by means of the cilia, like its ancestors
stentors did. The ciliate’s body elongates considerably during swimming and
loses its typical shape.
Symbiotic zoochlorellae often occur inside the cells of sousaphones, they give
the ciliate a greenish-yellowish tint, while the cell itself is hyaline. A macronucleus,
several micronuclei and a contractile vacuole are visible under the cell envelope.
The ciliate receives a part of required organic substances from the symbiotic
zoochlorellae. This species often becomes a prey of predators like worms, rotifers
and fish fry. Only its growth and multiplication rates save it from them.
Sousaphone reproduces by mitosis around once per day. This process can be preceded
by conjugation (but not necessarily). In adverse conditions, the animal produces
thick-walled cysts and loses moisture significantly, keeping viable with this.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Kraken
ciliate (Teuthosoma horribilis)
Order: Evaginogenida (Evaginogenida)
Family: Dendrosomidae (Dendrosomidae)
Habitat: still fresh waters of temperate climate of the Northern Hemisphere.
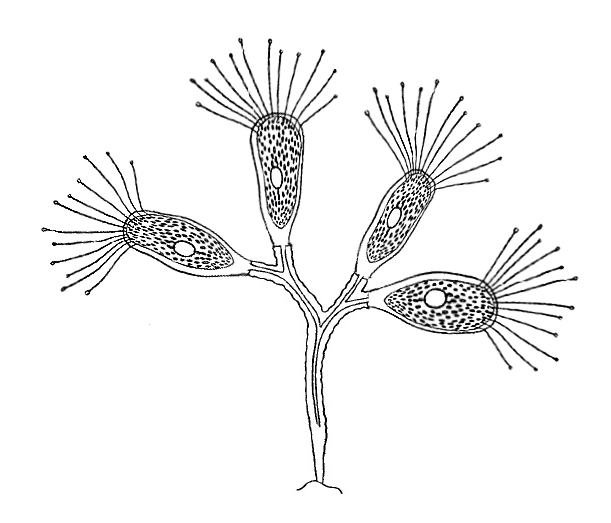
Picture by Biolog
Some sucking ciliates (subclass Suctoria) of holocene possessed
a special way of nutrition, atypical for this group of protozoans. They became
totally sessile and lost the cytostome and cilia. Instead of the cilia, they
got tentacles and vesicles to discharge enzymes for prey digestment into the
medium, then the prey sucked directly into a digestive vacuole.
In Neocene, one of their descendants developed further this way of nutrition.
The kraken ciliate, inhabiting fresh waters of lakes and river backwaters, became
a colonial form: its dendriform branching colonies attached to rocks, plants
and other underwater objects consist of 15-20 individuals and are about 15 mm
tall. Separate zooids, 3-4 mm large, sit on the tips of stalks. They have an
elongated shape and bear tufts of 8-10 long flexible tentacles on the front
end of the cell, which makes them distantly resemble squids (hence the species
name). The zooid cells and their stalks are colorless and hyaline.
Kraken ciliate is a voracious predator. When a zooid catches a prey, other also
lean towards it to help digest the prey, and have their shares of food. Apart
from other protozoans, due to its size, kraken ciliate consumes small crustaceans
(both larvae and adult), eggs of small invertebrates, larvae of mollusks and
other small animals. Potent enzymes allow it to soften even the shells of adult
crustaceans. The same enzymes serve for protection of the colony by repelling
small planarians and mollusks.
Kraken ciliate reproduces the same way as its ancestors: its zooids bud. Buds
have cilia and swim away to settle in a suitable place, shed the cilia and give
rise to a new colony. Sometimes this happens on the body of an animal (e. g.
on a mollusk shell), and the ciliate can travel with it.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Ichthyocinete
(Ichthyocineta symbiotica)
Order: Endogenida (Endogenida)
Family: Acinetidae (Acinetidae)
Habitat: all oceans of the Earth, bathyal zone.
Marine sucking ciliates (subclass Suctoria) in holocene used to inhabit shallow
depths, living on many substrates, including animal bodies (fish, cetaceans),
and feeding on various food. One of their descendants mastered great depths
and entered into a symbiosis with fish.
Ichthyocinetes live at depths of hundreds of meters, almost exclusively on the
bodies of fish, usually on the sides, less often on fins. It is a quite large
(1-1.2 mm tall) ciliate. An oval or almost spherical cell sits on a thin stalk
and is covered with a goblet-shaped lorica (additional proteinaceous coat).
The front end of the body bears a tuft of 10-15 tubular tentacles, each having
an extrusome (vesicle) with digestive liquor.
Inhabiting the fish body, ichthyocinetes get special food at their disposal:
unicellular fish parasites, particularly glowing swarmers of photopiscidium
(Photopiscidium bathichthybolis).
Moreover, ichthyocinetes capture them both before they are swallowed by the
fish, and when a new generation exits from a cyst in the fish’s skin. When a
captured photopiscidium swarmer is digested extracellularly, its symbionts –
infusorial photobacteria (Photobacterium
endoprotistum) – fall out into the water and partly are also captured and
digested by the ichthyocinete (as a result, their luminescence stops). One ichthyocinete
individual can capture simultaneously several photopiscidium swarmers and/or
bacteria. Thus, ichthyocinetes at least partially protect the fish from photopiscidium
and other parasites in exchange for treat. At the same time they interfere with
unmasking of the host fish by the colonies of glowing parasites, and save it
from predators.
Ichthyocinete reproduces the same way its ancestors did: by budding. The swarmer
buds(around 0.2-0.3 mm in diameter) have cilia and swim away to settle in new
places. In the deep, where these protozoans live, such places usually are represented
by fish bodies. Less often, they settle on organic particles which come from
upper layers of water – in this case they relocate onto the bodies of pelagic
worms or crustaceans and passively wait for relocation onto a fish body, when
a fish eats those animals. The swarmers of ichthyocinete have large trichocysts
that launch long acerate anchoring filaments to attach to a fish body. Hundreds
of swarmers (and, accordingly, new ichthyocinete individuals) can accumulate
on one fish. They exchange their hereditary material by conjugation.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Sheltering
funnel (Infundibulum refugium)
Order: Sessilida (Sessilida)
Family: Zoothamnidae (Zoothamnidae)
Habitat: all tropical seas of the Earth.
Colonial marine and freshwater ciliates (Vorticella, Carchesium, Zoothamnium
and other) existed in the human era. They formed dendriform or feather-shaped
colonies, consisting of various numbers of goblet-shaped cells (only several
to hundreds), on solid substrates. One of their descendants became a common
inhabitant of warm marine shallow waters of Neocene, it lives on rocks, seaweeds
and other underwater objects.
The sheltering funnel, just like its ancestors, forms branching dendriform colonies
up to 5 cm tall and about 4 cm in diameter on a single stalk, which is about
2 cm long. The zooids (2-3 mm in diameter) of the colony face cytostome upward
and slightly to the center. The peripheral branches of the colony are longer
than the inner ones, which gives the colony a funnel-like shape. The top zooids
on peripheral elongated branches are larger than others (4-5 mm) and bear long
(up to 12-13 mm) slightly thickened cilia at the outer side of the colony. The
beating of these cilia effectively drives food particles into the cytostomes
of colony members (both own and other zooids), which allows to surpass smaller
competitors in getting food.
The colonies of sheltering funnel seem white under natural illumination because
of light refraction in small hyaline zooid cells.
A disturbed colony of sheltering funnel instantly contracts the stalk and branches,
shrinking into a ball about 1.5 cm in diameter. With this, the outer cilia of
the peripheral zooids are not retracted, but pile up in a roof shape (hence
the species name), covering the colony from the top.
The reproduction cycle of sheltering funnel is the same that in its ancestors:
differentiated macrozooids transform into rather large (up to 4-5 mm) free-swimming
“swarmers”, which secede from the mother colony, settle on a substrate and give
rise to a new colony. The “swarmer” can settle and give a new colony on bodies
of various animals (e. g. on mollusk shells), and if the animal is motile, the
colony travels together with it.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Bathytrichodine
(Bathytrichodina parasitivora)
Order: Mobilida (Mobilida)
Family: Trichodinidae (Trichodinidae)
Habitat: all oceans of the Earth, bathyal zone.
Trichodines were specialized ciliates of holocene, inhabiting bodies and gills
of fish. Their body was flattened-convex, usually in the shape of a thick one-sidely
convex lens. The bottom surface of such lens, adjoining to the fish’s body,
had an attachment sucker disk with a concave center and a ring of of radial
cytoskeletal denticles. The cytostome (cellular mouth) was on the top, surrounded
by a ring of cilia.
In Neocene, the bathytrichodine, inhabiting the bodies of deepsea fish, has
kept these features. Its cell, about 0.7-0.8 mm in diameter, has the same lens-shaped
appearance. The attachment disk has 30 denticles, diverging from the center,
and is surrounded with a ring of cilia. The upper ring around the cytostome
consists of long, slightly thickened cilia.
The ciliate creeps freely across the fish’s body by means of movement of the
cilia of the lower ring, but it also can detach to swim freely – usually this
happens when the host dies. But the fish’s body gives it a plentiful treat –
it can catch swarmers of photopiscidium (Photopiscidium
bathichthybolis) and digest them. Moreover, the ciliate does this including
during rupture of a mature cyst of the parasite on the fish’s skin. Bathytrichodines
can sense the presence of the cysts by their luminescence (infusorial
photobacteria often survive in new swarmers) and creep closer to them, thus
demonstrating positive phototaxis. Unlike its ancestors, which were partial
parasites of fish, bathytrichodine helps the fish get rid of parasites in exchange
for food.
Bathytrichodine reproduces by simple division, like its ancestors: the fission
occurs parallel to the sucker disk, and the new cell grows cilia and a new sucker
disk to swim away and settle in a new place, more precisely, on a fish body.
Usually, such swarming ciliates soar motionless in the water column and are
activated only at the emergence and subsequent enhancement of odor of a possible
host.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Photopiscidium
(Photopiscidium bathichthybolis)
Order: insertae sedis
Family: insertae sedis
Habitat: all oceans of the Earth, bathyal zone.
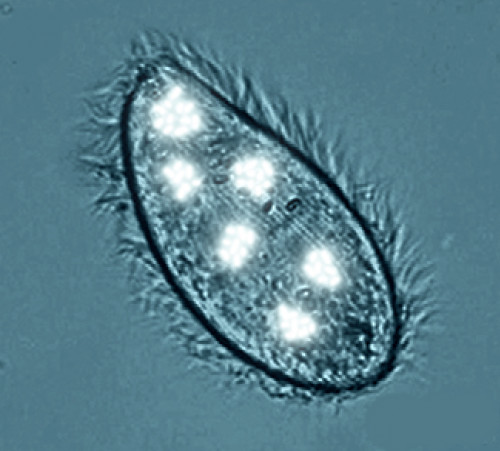
Picture by Biolog
One of marine holocenic ciliates – Cryptocaryon from
Prostomatea class – had adapted to parasitize on fish, causing a disease named
cryptocaryonosis (“marine ichthyophthiriosis”). This parasite formed cysts in
the fish’s skin, with new swarmers forming inside them. Some descendants of
this species changed their life style in Neocene.
Photopiscidium lives in deep waters of oceans and parasitizes on deepwater fish.
A mature swarmer appears as an oval or ovoid cell about 0.5 mm in diameter,
covered with cilia, with a bean-shaped macronucleus and a small micronucleus.
It glows from inside with a bluish light – its cytoplasm contains symbiotic
infusorial photobateria (Photobacterium
endoprotistum) as several aggregations. Bacteria receive food (which is
hard to find in the deep of the ocean) from the ciliate in exchange for help
in infecting fish.
Glowing ciliates soar in the water column and attract young and small fish,
which eat them. Inside the fish’s intestine, the ciliate produces potent enzymes
to pierce through the intestinal wall, get into the bloodstream and settle in
the host’s skin, along the way consuming the fish’s body substances and feeding
them to their symbiotic bacteria. At this stage of invasion, part of the bacteria
sometimes dies from the ciliate’s enzymes. In the fish’s skin, photopiscidium
forms cysts, the cell repeatedly divides inside it. The cyst grows up to 1.5-2
mm with this, and glows with intensity depending on the number of ciliate rods
that have survived in the new cells. In favorable conitions, the bacterial population
restores rapidly.
Over time, the cyst ruptures, and a new portion (several dozens) of new swarmers
comes out. In the water, they feed freely for some time, swallowing new living
ciliate rods, and then, once they acquire sufficiently bright glowing, they
are swallowed by a fish.
Dozens of such cysts can appear on one fish, and it gleams like the night starry
sky. Such gleaming attracts other fish that feed by predation, and photopiscidium
gets a chance to enter another victim’s body even without leaving the previous
one. But this rarely happens at great depths.
Fish have their own protection from this parasite – dangers await the swarmers
before they infect and just before they exit from the cysts: sucking infusoria
ichthyocinetes (Ichthyocineta symbiotica)
and creeping ciliates bathytrichodines (Bathytrichodina
parasitivora) inhabit the fish skin, and they often feed on the photopiscidium’s
swarmers.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Flagellates |
Iridescent
noctiluca (Multiluca versicolor)
Order: Noctilucales (Noctilucales)
Family: Noctilucaceae (Noctilucaceae)
Habitat: tropical and moderately warm seas worldwide.

Picture by Biolog
In the human era, some marine flagellates had adapted to repel
enemies by bright light flashes of bioluminescence. The organisms of this kind
in human era were the noctiluci (Noctiluca), which used to flare brightly
when disturbed. Forming large aggregations, they caused sea mareel (or milky
sea) on large areas. Their descendants in warm and temperate seas of Neocene
have kept many of their features, but the evolution of some of them took new
directions. The iridescent noctiluca became one of such species.
This species became colonial from solitary: dividing cells, 3-4 mm in diameter,
do not come apart and form laminar coenobia resembling those in a Holocenic
alga Gonium. The coenobia in the iridescent noctiluca are 15-16 mm
wide and consist of 8-32 identical cells enclosed in a collective slimy capsule.
The iridescent noctiluca has inherited the cellular structure from its ancestors:
it has a cellular mouth and a modified contractile flagellum tentacle, and also
fat inclusions in the cytoplasm, which allow the cells to soar in the water
column.
When a coenobium is disturbed, all of its cells flare up brightly, simultaneously
then separately, and the bioluminescence color can be different in different
cells within one coenobium, and with this the colony shimmers with reddish,
yellowish, bluish and greenish light. Such illumination will repel almost any
dinoflagellate hunter.
The ways of reproduction in the iridescent noctiluca are the same that in its
ancestors, but adjusted for colonial life style: either the coenobium breaks
into separate cells and they give rise to new colonies, or the cells bud within
the coenobium. Both ways occur equally often and alternate with each other.
In favorable conditions, iridescent noctiluci multiply in great numbers and
cause a bright shimmering mareel which gleams with all colors of the spectrum.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Sundew
bodonid (Endobodo droserae)
Order: Bodonida (Bodonida)
Family: Bodonidae (Bodonidae)
Habitat: outskirts of the Mediterranean basin.

Picture by Biolog
Bodonids were one of the most widespread groups of flagellates
in small or even temporary, polluted freshwater ponds of holocene. They were
abundant in puddles, ponds and wetlands, but preferred places where bacteria
– their main food – multiply.
One of its descendants – the sundew bodonid – inhabits the pitcher-like traps
of the pitcher-leaved sundew
(Neodrosera nepenthifolia), which grows in arid conditions of outskirts
of Mediterranean basin. These are very small (no more than 50 μm) flagellates
– their habitats are too small in volume. The cells are ovoid, bearing two flagella
(anterior and posterior) on the wider end. The flagella have large basal bodies,
and a cytostome (cell mouth) is located in the bottom of the flagellar pocket.
The entire surface of the cell is strengthened with microtubules that underlay
the cellular membrane – this allows to maintain a constant cell shape and to
move in a viscous medium. The posterior flagellum is shortened (three times
shorter than the anterior one) and bears the function of driving bacteria and
other food into the cytostome. The cell often has several (up to 5-6) digestive
vacuoles filled with food and appearing dark, almost black.
In arid conditions of Mediterranean basin, this species has found a habitat
and a food source in the trap leaves of the sundew, where some moisture remains.
Prey caught by the sundew and partly digested always can be found there, and
this prey becomes a substrate for bacteria, and the bacteria, in turn, are food
for the sundew bodonid. This species is a commensal that eats from the sundew’s
table, although it partially performs a cleaning function: it prevents spoilage
of the sundew’s digestive fluid by eating bacteria. The cell wall of this species
is more durable than in its ancestor, so that it can withstand the digestive
action of the fluid in the sundew’s leaves.
In adverse conditions, the flagellate can form cysts, in which it keeps viable
for a long time (up to several weeks). Usually, this happens when the aerial
part of the plant dries due to lack of moisture. The cysts are easily carried
by the wind and driven into the traps of other sundews. Also, the are easily
transmitted on the feet of insects that managed to escape from the plant’s traps.
This species of protists was discovered by Biolog, the forum
member.
Translated by Biolog.
Amoebozoa |
Host
amoeba (Gregariamoeba structurata)
Order: Tubulinids (Tubulinida)
Family: Amoebas (Amoebidae)
Habitat: almost everywhere, where there are temporary or long term reservoirs
with the polluted water and low contents of oxygen.
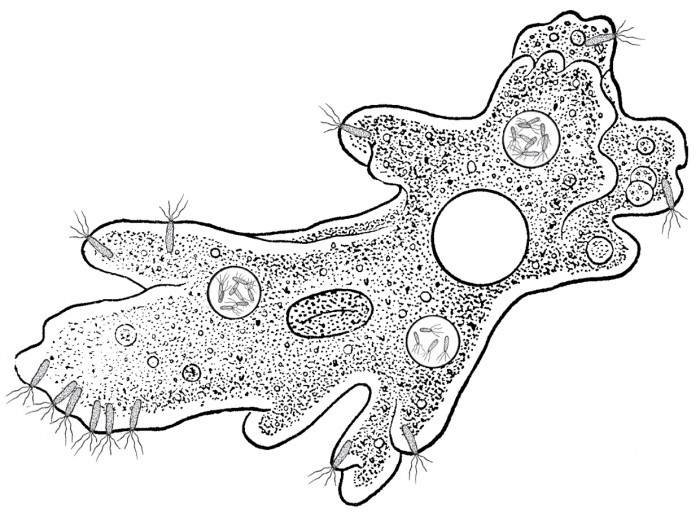
Picture by Biologist
Sponges hardly endure the polluted water, therefore in human
epoch the variety of freshwater sponges had sharply decreased, and organisms
of other systematic groups had started to substitute them in their niche.
Possible, as a consequence of this redistribution of roles in Neocene ecosystems
colonial protozoans had evolved, that eat plankton and the weighed particles
of 1 – 5 microns in size, living in eutrophic reservoirs. One of their representatives
is host amoeba named because of some features of colony life.
The colony of host amoebas looks like a shapeless slimy crust of 0.01 – 0.05
mm thick on underwater objects – plants, stones and driftwood. The cell has
the shape of flat cake; its diameter is about 70 microns, and height is approximately
10 microns. As against related forms, host amoeba has no its own shell.
In cytoplasm of these protozoans 2 species of symbiotes live: lophotrichous
bacteria attached in deepenings of plasmalemma (at the encystment of the host
cell they are located in vacuoles) and parasitic bacteria of the unknown origin,
synthesizing folic acid for the host and having the important role in chemotaxis
(they are able “to remember” a direction and to adjust the formation of pseudopods
at this place). These symbiotes constantly live in vacuoles of host amoebas.
At division of the cell they are distributed between daughter cells.
At the end of cell most removed from the substratum there is a large bunch
of lophotrichous bacterial symbiotes, moving food objects closer. Near to
this bunch the hyperactivity of cytoplasm is observed: pseudopods grow more
often and phagocytosis proceeds more actively. Sated cells creep away deep
into the colony. Amoebas use the bacterial symbiotes to create a current of
water to vertically extended site of body.
These protozoans live in eutrophic reservoirs and consequently are periodically
compelled to struggle against burying of colonies with the deposit. Host amoebas
struggle against this adverse phenomenon in two ways. They mainly migrate
through a layer of deposit, forming thus of a chain of cells following the
“leader” cell (at this time the colony is frequently separated to some daughter
ones). Otherwise they migrate, forming “guides” (one more species of the symbiotic
bacteria producing a number of amino acids and determining behaviour of amoeba
in many respects) that allow “to remember” the direction of movement, and
the whole colony creep in this direction till 5 – 70 minutes following each
other like caterpillars of pine processionary moth. If the layer of deposit
is thin, cells creep through it upward in order to remain everytime at the
border of two environments. In more serious cases the colony is had to migrate.
Migration begins from occurrence of “leaders” to which cells at the edge of
colony turn. Then the formation of chains of cells follows, and they search
for more favorable conditions independently from each other. In this case
amoebas alternate “a choice of direction” at which “leaders” replace each
other, and the periods of straight crawling. At this time amoebas eat bacteria
living in silt. During the migration colony frequently breaks up to some parts.
The second problem to which these protozoans face is a shortage of oxygen
frequently happening in polluted reservoirs. Host amoebas struggle against
it by the formation of planktonic settling forms. They have numerous bacterial
symbiotes with flagella, facilitating the swimming (the direction of flagellae
rotation can change to opposite one), and long “thorns” of cast away bacterial
flagella, facilitating soaring in thickness of water. After the period of
plankton life these protozoans can settle again on the ground or floating
objects and form colonies.
The colony keeps the certain orderliness due to “skeleton”. The common “skeleton”
of the colony represents friable enough construction of flagellae having polysaccharide
chemical structure cast away by lophotrichian symbiotes with the insignificant
addition of proteins. This “skeleton” also serves also as food stock for the
whole colony.
Settling of host amoebas proceeds in two ways. In standard case this species
is settled with the help of cysts which are not so steady, especially to drying,
and are only temporary condition of the cell. Less often some amoebas “stick
together” to the chain, grow long (about 20 – 40 microns) cross “hairs” representing
modified bacterial flagella, and due to them get much better hydrodynamical
characteristics. Such planktonic settling stages are formed at shortage of
oxygen. The whole colony breaks up to set of such chains and “flies up”, leaving
the substratum.
Sexual breeding in life cycle of host amoebas is absent. A free-floating stage
can easily become a commensal of various aquatic animals, more often Daphnia
species on which shell small temporary colonies of filterers are formed. Sometimes
the floating stage gets on gills of fishes, and due to them can be transported
to rather long distance. Cysts at this species are formed at approach of adverse
conditions (food shortage and/or drying of reservoir), and do not endure complete
drying, therefore they are kept only in places where silt keeps sufficient
dampness. These protozoans are rather numerous and sometimes form in their
habitats a kind of original tiny stromatolites.
This species of protists was discovered by Mutant, the forum member.
Glass-shelled
drosarcella (Drosarcella vitrea)
Order: Arcellinida (Arcellinida)
Family: Arcellidae (Arcellidae)
Habitat: outskirts of Mediterranean basin.
Some amoebas known to humans were different from their relatives by the presence
of a shell that protected them from external impacts. The shell could be formed
by different ways: either by producing structural material (chitin, silica)
by the cell itself or by deposition of phagocytized and undigested particles
(usually mineral) on the cell surface.
In Neocene, the descendants of testate amoebas have mastered various ecological
niches, but one species, the glass-shelled drosarcella, became a commensal
of the insectivorous plant pitcher-leaved
sundew.
This species is a descendant of Arcella genus, which had inhabited
fresh water, including puddles and wetlands. It is a small (about 0.2 mm)
amoeba that produces a bilayer test in the shape of a one-sidedly convex lens
with an aperture underneath. The inner layer of the test is brown chitin (like
in the ancestors), and it is covered with a layer of silica (silicon dioxide)
from outside, which gives the amoeba’s test a glass shine and a reflective
ability. There is an aperture in the bottom side of the test, with several
pseudopodia protruding from it, which gives the amoeba a mushroom-like shape.
The reflective “glazed” test protects the amoeba from the sun of the Mediterranean
basin, where this species finds food and moisture in the pitchers of the pitcher-leaved
sundew. Climate drying, caused by closing of the Strait of Gibraltar and drying
of the Mediterranean Sea, made this species to back off to such an unusual
habitat. There, it eats from the sundew’s table, feedind on its prey. It creeps
across the internal walls of a pitcher and down into the liquid – along the
sundew’s hair and on floating captured prey. In order to avoid being digested
by the plant’s digestive enzymes, the animal keeps in the topmost layer of
the liquid, where the concentration of enzymes is the least. In the conditions
of the Mediterranean basin, this amoeba lives almost exclusively in the trap
leaves of the sundew.
Drosarcella reproduces by division. The cell partly “flows out” of the aperture,
its nucleus and other organelles divide, and formation of a new test starts.
In adverse conditions, the amoeba can form thick-walled cysts, which is preceded
by full exiting of the cell from its test. The cysts are transmitted by insects.
This species of protists was discovered by Biolog, the
forum member.
Translated by Biolog.
Purple
megalosarcodina (Megalosarcodina purpurea)
Order: Arcellinida (Arcellinida)
Family: Megalosarcodines (Megalosarcodinidae)
Habitat: the Mediterranean hollow, hyperhaline swamps.
Hyperhaline swamps and lakes, formed in a hollow of former Mediterranean Sea,
represent an extreme inhabitancy for live beings. Here, among white salt plains,
only few species survive, which can adapt to presence of great amount of salt
in water. But the ones managed to survive in such conditions, prosper and
are virtually out of competition. Frequently the evolution of such organisms
derivates freakish lifeforms.
Very large testate amoeba externally similar to fossil nummulites of Tertiary
lives in hyperhaline water of Mediterranean swamps. It is purple megalosarcodina,
a species of shell-bearing protozoans, making the separate family. Its size
is measured in centimeters – it is a giant among protozoans. The size of flat
disc-shaped shell of megalosarcodina is about 5 – 6 cm. The cell of this amoeba
represents multinuclear syncytium (the number of nuclei is measured by thousands)
having flat leaf-like shape. The shell is also flat and disc-like, made of
carbonates. The surface of this shell has numerous small transparent crystal
fenestellae. The shell grows in concentric circles along the edge. It is capable
to regeneration, and as it grows, it also turns thicker in the middle due
to accumulation of new layers of material. On the surface of the shell borders
of the previous layers of increase are seen. After the dying off of megalosarcodinae
their shells fall on the bottom of reservoir, forming deposits of limestone
in ultrahaline swamps.
Numerous bacteria living in cytoplasm of purple megalosarcodina are extreme
halophiles performing the photosynthesis by means of bacteriorhodopsine pigment.
Due to their presence the amoeba has beautiful purple color. Symbiotic bacterium
Rhodolampropedia hexagonalis is the direct descendant of bacterium
Lampropedia. In human epoch purple bacteria of unusual cell shape
– triangular or square (such cell shape Lampropedia had) lived in
Dead Sea. Their descendant has cells of rectilinear hexagon shape. At very
large width reaching two millimeters (in human epoch such bacteria had not
been known) thickness of cell does not exceed one tenth of hair thickness.
Bacteria form piles of cells like thylacoids in plant chloroplasts, and piles
are stacked in cytoplasm of amoeba as honeycombs: the hexagonal shape of bacterial
cells enables it. The shell of megalosarcodina has the cellular structure
adapted to the best focussing of light penetrating through fenestrellae at
the top side of shell.
Like all amoebas megalosarcodina has no strictly determined body shape. The
cell forms pseudopods able to creep from shell in searches of food – mainly
organic particles and bacteria. Pseudopods are supplied with the protective
weapon – poisonous harpoons like infusorian trichocysts. Therefore water turtlebeetles,
crustaceans and salt-loving
skinkfishes do not eat megalosarcodina. Also megalosarcodina can stretch
pseudopods in brightly lighted sites of the bottom in order to allow symbiotic
bacteria to perform photosynthesis more actively – megalosarcodina receives
from them an essential part of feeding, from time to time “attacking” very
much multiplied symbiotes with digestive enzymes.
This species of protozoans breeds by means of cell division, and one half
of former cell stays on shell, and another half forms its shell anew within
several days. Megalosarcodina also has sexual process – merge of nuclei in
cell with the subsequent division. Sexual process appears in adverse conditions,
for example, at drying of the reservoir. This protozoan endures drought, closing
shell apertures with secretions of glycoproteid nature and running into anabiosis.
Bacteria thus turn to cysts. If adverse conditions are delayed, megalosarcodina
gradually eats a part of symbiote cysts.
In favorable conditions, and also at some freshening of water (for example,
during the rainy time) megalosarcodina breeds more intensively, separating
pseudopods and other parts of the body. If division has taken place inside
the shell, the daughter individual abandons it through one of apertures for
pseudopods.
Accumulating fat drops in cytoplasm, purple megalosarcodina gets a side benefit,
the buoyancy. Hyperhaline water helps to float, pushing out this animal to
the surface. This way of movement is characteristic for younger individuals
with rather lightweight shell. Larger animals with thick massive shell creep
on the bottom.
This species of protists was discovered by Anton, the forum member.
Insect-eating
droseroflugia (Droseroflugia insectivora)
Order: Arcellinida (Arcellinida)
Family: Difflugiidae (Difflugiidae)
Habitat: outskirts of Mediterranean basin.
Some testate amoebas known in the human era did not synthesize their tests
themselves, but deposited undigested particles (usually mineral) captured
by phagocytosis on the cell surface, where they were agglutinated by an organic
matrix.
In Neocene, one of their descendants, the insect-eating droseroflugia, settled
in the traps of pitcher-leaved
sundew on outskirts of the Mediterranean basin, and adapted to make shelter
from particles of the plant’s prey. This is almost the sole habitable place
for relics of freshwater microbiota after aridization of this terrain.
This amoeba is the same size as drosarcella
– about 0.2 mm. Inhabiting the trap leaves of the sundew and ingesting microparticles
of its prey, droseroflugia excretes solid undigested particles onto its surface
and agglutinates with an organic matrix. Therefore, the outer appearance of
its test is variable and depends on the sundew’s prey. But generally the test
consists of solid particles obtained from insects captured by the sundew:
microscopic parts of their shells, antennae, legs, wings etc. All this creates
a variegated spotted shell on the amoeba, and it protects the protozoan from
the sun. The test is bell-shaped with a wide aperture in the bottom, from
which the amoeba’s pseudopodia protrude.
The life style of this species is the same as in drosarcella, but it is commonly
found on the surface of the liquid in the traps, or creeping on partly submerged
prey.
Droseroflugia multiplies by division with the same mechanism as in drosarcella.
In adverse conditions, this species forms cysts that are spread by insects.
This species of protists was discovered by Biolog, the
forum member.
Translated by Biolog.
“Spelean
sulfur” (Sulfophysarum spelaeus)
Order: Fuscisporida (Fuscisporida)
Family: Physaridae (Physaridae)
Habitat: caves of North America (the Appalachians).
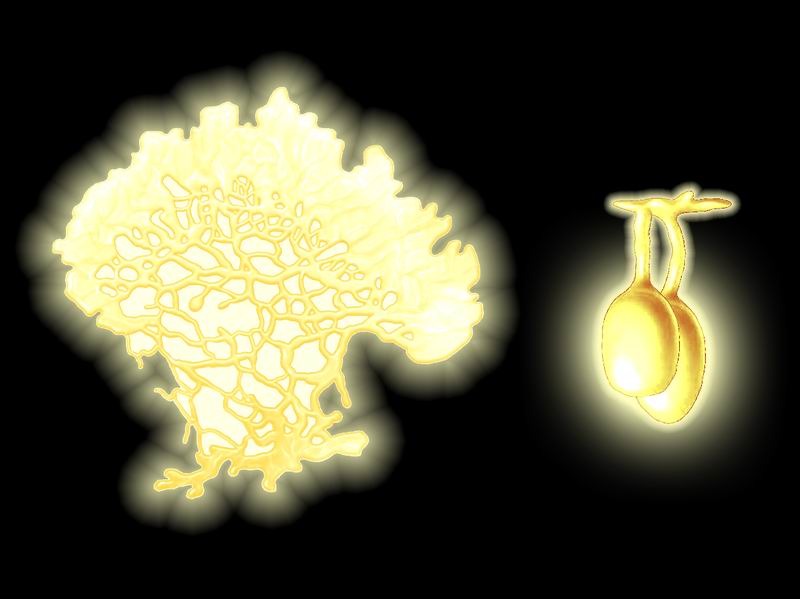
Picture by Biolog
The slime molds (myxomycetes) are a quite widespread group
of holocenic fungoid organisms inhabiting moist places in temperate forests
of all continents. They play an important role as decomposers of organic remains
and regulators of bacterial populations. Some of their descendants in Neocene
mastered new ecological niches, and one species has relocated to caves, having
adapted to specific life conditions.
“Spelean sulfur” is a descendant of Physarum genus which had lived mainly
on dead wood. The plasmodium of this species appears as a thin and soft pale
yellow moist crust permeated with a dendriform network of growth tubules.
It is a single large multinucleated cell that is the vegetative stage of the
organism. The plasmodium can move slowly (about 5 cm per hour), and simultaneously
grow from the edges. During sporulation, the plasmodium stops moving and covers
with a tough coat. Sporangia of the same color, about 1 cm in diameter on
small stipes, appear on it. They are covered with cellulose coat and open
from the base. Colorless spores also have cellulose coats, and are spread
(and partly eaten) by cave insects and mites. Myxamoebas – amoeboid gametes
– emerge from spores that found favorable conditions with proper moisture
and temperature. Fusing in pairs, they give rise to new plasmodia.
This slime mold feeds on guano and organic remains of animals (birds and mammals)
inhabiting the cave, and also on bacteria, which multiply in large numbers
on these substrates. Feeding on bacteria, the plasmodium can creep on stone
walls and even the arch of the cave; in both cases, the sporangia will hang
down. Spores also can spawn new myxamoebas and new plasmodia on the walls
and ceiling of the cave.
Insects that spread the spores of “spelean sulfur” are guided by light: the
myxomycete gives off a yellowish glow, dim but clearly seen in the dark, and
the cave brightens up with yellow stains, resembling deposits of native sulfur
(hence the species’ name).
This species of slime molds was discovered by Biolog, the
forum member.
Translated by Biolog.
Retaria |
Corallogerina
(Corallogerina natans)
Order: Globigerinida (Globigerinida)
Family: Globigerinidae (Globigerinidae)
Habitat: seas of warm and temperate climate around the globe.
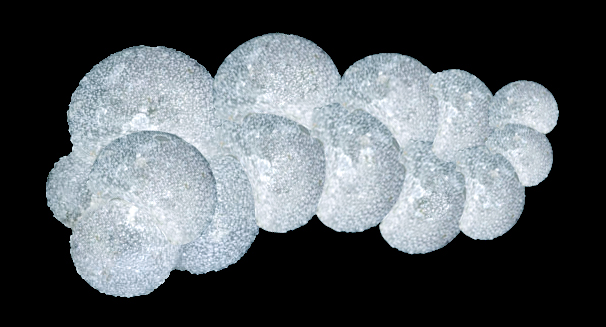
Picture by Biolog
In prehistoric and human eras, foraminifera held a special
place among the marine protozoans, playing an immense role in geological processes.
Their calcareous shells formed bottom sediments – the limestone and chalk.
The thickness of these deposits reached dozens and hundreds of meters, and
sometimes even kilometers. In Neocene, this geological process continues,
although the species composition of its participants has changed significantly.
Corallogerina is one of typical foraminifera of Neocene, a descendant of Globigerina
genus of human era. It inhabits warm and temperate seas at shallow depths
(up to several meters) as a part of marine plankton. Their ancestors, globigerinas,
were also planktonic foraminifera, their tests were thin and translucent,
and extremely thin (like a spiderweb) pseudopodia protruded through the pores.
Corallogerina has inherited all of its ancestor’s traits, but the nuclei of
agamont (diploid generation) in it divide repeatedly and give rise to an aggregate
of cells, which do not secede and stay together. These cells cover with tests,
and the colony resembles pieces of coral branches (hence the species name)
soaring in the water column. The diameter of one cell in the test is small
(2-3 mm), but the branching colony reaches 2-3 cm in length and consists of
10-15 haploid cells.
Each of these cells can then spawn gametes, and the gametes, by fusing, produce
a zygote, which gives rise to a new agamont generation, thus continuing the
life cycle.
The tests in corallogerina are thin, spherical, almost transparent, bearing
small spikes on the outer surface, and pores through which extremely thin
pseudopodia protrude and surround the cell with an arachnoid halo.
The tests of corallogerina compose a significant part (half and more) of bottom
calcareous sediments of neocenic seas, forming very thick deposits of lime
ooze, up to 4-5 meters thick.
This species of protists was discovered by Biolog, the
forum member.
Translated by Biolog.
Golden
heliochromococcus (Heliochromococcus aureus)
Order: Arthracanthida (Arthracanthida)
Family: Heliochromococcidae (Heliochromococcidae)
Habitat: tropical seas around the globe.
Starting from early Paleozoic, one group of marine planktonic protozoans had
mastered the construction of skeletons mainly from silica (SiO2)
and entered into symbiosis with algae to obtain energy of sunlight. These
are radiolaria – very peculiar and beautiful marine microorganisms. In Neocene,
one of the groups of these organisms began to evolve towards total autotrophy.
Heliochromococci are spherical radiolaria of Acantharia group inhabiting surface
(up to several meters) layers of water in warm tropical seas of Neocene. Just
like their ancestors, they synthesize skeletons from strontium sulfate (SrSO4,
celestine). These are small pelagic organisms about 0.5-1 mm in diameter.
Their skeleton appears as two cribriform spheres nested in each other and
connected by beams. Both spheres are transparent like glass, and shine in
the sun. The radius of the inner sphere is ½ of that of the outer sphere,
and it is entirely occupied with a large central capsule filled with golden-yellow
zooxanthellae. Ectoplasm is between two spheres, and also covers the outer
sphere from outside. Numerous pseudopodia, extremely thin like spiderweb,
stretch out of it. The outer surface of the outer sphere is smooth, but bears
20 thin straight spikes arranged in a regular icosahedral pattern. The sphere’s
walls between the spikes bear thickenings arranged checkerwise and performing
the role of optical lenses to provide zooxanthellae with light.
Zooxanthellae have undergone assimilation in the cells of heliochromococci
and lost the ability to live independently. And the radiolarian, due to this,
switched to full autotrophy – it provides the zooxanthellae with mineral substances
from the water, and they provide it with organic substances and energy by
photosynthesis.
The life cycle in heliochromococci is the same as in their ancestors: a fully
formed organism produces flagellated zoospores, which lose flagella and produce
their own skeletons to give rise to a new organism. During zoospore formation,
a part of zooxanthellae passes to the spore and then to the new organism.
In favorable conditions, heliochromococci can multiply in large numbers, causing
a golden sea bloom.
Blue-green
heliochromococcus (Heliochromococcus cyaneus) is a species with green
zoochlorellae, its spikes are thickened at the ends and have a bluish tint
(the color of strontium sulfate). This species causes a bluish-green sea bloom.
Red
heliochromococcus (Heliochromococcus ruber) contains reddish-orange
zooxanthellae and has thin spikes branched at the ends, and causes orange-red
sea bloom.
This species of protists was discovered by Biolog, the
forum member.
Translated by Biolog.
Azure
vanadella (Vanadella azurea)
Order: Vanadellata (Vanadellata)
Family: Vanadelloideae (Vanadelloideae)
Habitat: tropical seas around the globe, near-surface and middle layers of
water (up to dozens of meters deep).
Radiolaria (phylum or subphylum Radiozoa) in holocene were well-known for
their ability to form cytoskeletons from silica (SiO2), and one
group from strontium sulfate (SrSO4). However, the sea water contains
many compounds of other elements, and one of the main places is held by vanadium
which is present there as the vanadyl ion (VO2+). Vanadium is toxic
as pure oxides, but as vanadyl it was used by some marine organisms of holocene.
These organisms learned to include vanadium in cofactors of enzymes (some
algae and bacteria) and in protein molecules with further accumulation in
special cells (tunicates, particularly sea squirts – as a way of protection
against predators). By the Neocene time, some radiolarians also switched to
using vanadium compounds to produce their cytoskeletons.
Vanadellas (Vanadellae) are a new class of neocenic radiolarians. Their main
distinctive feature is the cytoskeletons made of complex silicates with vanadium
and calcium (such composition is inherent for vanadium minerals cavansite
and pentagonite). These skeletons consist of intersecting ring structures
with common intersection points on the skeleton’s “poles”. Tufts of rather
long needles go from these points, and both rings and needles can be smooth
or spiked. The entire skeleton is stained a color with various tints depending
on the vanadium content: azure, sapphire, saturated blue of various tints.
The central capsule in vanadellas is not separated from the ectoplasm: their
cells do not contain zooxanthellae, they are totally heterotrophic. This allows
them to inhabit greater depths than their relatives, who depend on the symbionts’
photosynthesis. The life cycle in vanadellas does not differ from that of
their ancestors and relatives. Vanadellas feed on bacteria and organic particles
suspended in the water.
In the azure vanadella, the cells are 0.2-0.4 mm large, have 5 rings and 5
needles in the skeleton, which is covered with small spikes and entirely stained
bright-blue.
By converting vanadium, vanadellas take part in its mineralization, enriching
the bottom sediments with it. Multiplying in large numbers, vanadellas cause
water bloom, giving it a color corresponding the color of the vanadella’s
skeleton.
Turquoise
vanadella (Vanadella caerulea) has 7 smooth rings and needles stained
bright blue with a turquoise tint. It inhabits greater depths of water than
the azure vanadella (30-40 m).
Purple
vanadella (Vanadella purpurea) has 6 rings and needles covered with
spikes and stained purple-blue. It inhabits subsurface layers of the water
(first few meters).
This species of protists was discovered by Biolog, the
forum member.
Translated by Biolog.
Bacteria |
Philanthomycete
(Philanthomyces antibioticus)
Order: Actinomycetales (Actinomycetales)
Family: Streptomycetaceae (Streptomycetaceae)
Habitat: Fennoscandian forests, a symbiont in burrows of hymenopterans
Streptomycetes (Streptomyces genus) were one of main tools for humans to control
infectious diseases – humans obtained dozens of antibiotics of various action
spectrum. These filamentous bacteria produced erythromycin, tetracyclines,
levomycetin, nystatin, amphotericins, neomycin, vancomycin, and many other
antibiotics, and also other important substances (tacrolimus, allosamidin
and other). Representatives of streptomycetes entered into symbiosis with
insects – they lived in pockets of antennae of female beewolf (Philanthus
triangulum) and protected its eggs and larvae from soil mycelial fungi. In
Neocene, this symbiosis got further development, and the relationships between
the actinomycete and the insect became even stronger. A new actinomycete –
philanthomycete – emerged in the process of their joint evolution.
Philanthomycete is a typical representative of actinomycetes. Its cells appear
as thin branching aseptate filaments which divide into thinner substrate filaments
and thicker aerial filaments. The aerial filaments form dense tufts with chains
of thick-walled spores formed on their ends by successive fragmentation. The
filaments and spores are hyaline, filaments up to 1.3 μm thick.
This species is a symbiont of a crabronid wasp named beelynx
(Neophilanthus apilynx). The actinomycete is present as spores in the
antennal pockets of female wasps. The spores germinate quickly on the walls
of burrow chambers and on the prey – paralyzed bees. The actinomycete forms
a thin white powdery coating there. A wasp hatching from the pupa touches
the coating with antennae to gather spores into the mycangia.
The main help for the wasp from the bacterium is protection: the bacterium
has inherited the ability to produce antibiotics from its ancestors. Philanthomycete
produces large amounts of philanthomycin – a nystatin derivative which is
a potent antifungal antibiotic with a broad action spectrum. Philanthomycin
prevents the growth of any fungi inside the insect’s burrow, thereby protecting
the larva and its forage from being destroyed. After eating a prey with the
bacterial coating, the wasp’s larvae become resistant to fungal infections,
including Entomophthorales and Cordyceps that are specialized insect parasites.
In exchange, the bacterium receives nutrition – parts of the wasp’s prey and
excrements and secretions of the larva.
The ancestors of this species were less specialized and inhabited a broader
spectrum of niches – soil, surface of plants, organic-polluted water etc.
But philanthomycete, due to the close symbiotic relationships, is no longer
found outside the wasp’s burrows.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Infectiofly
asporoclostridium (Asporoclostridium dolichomusci)
Order: Clostridiales (Clostridiales)
Family: Clostridiaceae (Clostridiaceae)
Habitat: tropics and subtropics of the Old World – Africa, Zinj Land, Asia
and southern Europe.
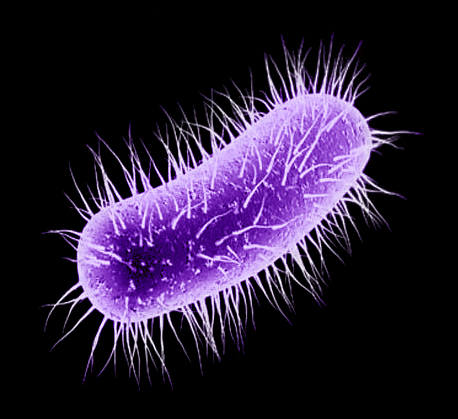
Picture by Biolog
In human era, clostridia were very abundant group of anaerobic
soil bacteria, although some species inhabited (permanently or temporarily)
human and animal intestine. Some species were deadly dangerous for humans
due to very potent exotoxins, e. g. agents of botulism (Clostridium botulinum),
tetanus (C. tetani) and gas gangrene (C. perfringens and
similar species). In Neocene, some clostridia have taken a new evolutionary
step: they have lost the ability to produce spores because of favorable conditions
they found after entering into symbiosis with dipteran insects.
Asporoclostridium dolichomusci is a descendant of C. perfringens.
It is a large (3-4 μm long and 2 μm thick) gram-positive, obligately anaerobic
rod, inhabiting pockets of digestive tract of some carnivorous flies (particularly,
infectioflies and sambios).
This bacterium is a chemoorganotroph, and inside the insect’s organism it
uses the host’s food for nutrition, without harming the fly. But when it enters
the body of a living vertebrate animal, the picture changes dramatically.
The bacterium produces a potent exotoxin (neurotoxin), which acts like the
botulism toxin of C. botulinum and tetanospasmin of C. tetani,
and also some hemolytic enzymes. This complex quickly (without “aid” of other
bacteria in 2-3 days) kills the animal since the fly injects this “biological
weapon” with its proboscis directly into the tissues and bloodstream of the
victim, causing an instant sepsis.
When the fly shows up to “have lunch”, it sucks in a new portion of bacteria
(which have multiplied in the victim’s body) with food. In fact, the symbiotic
bacteria circulate between the fly and its victims, almost totally avoiding
getting into the environment, which allowed them to do without endospores.
Besides clostridia, this deadly microbiome of flies includes fly
escherichia (a descendant of E. coli – a former inhabitant of
animal intestines, including insects) and sambio
fly staphylococcus (a descendant of S. aureus – a former member
of human and animal normal microbiota).
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Sambio
fly staphylococcus (Staphylococcus sambiorum)
Order: Bacillales (Bacillales)
Family: Staphylococcaceae (Staphylococcaceae)
Habitat: tropics and subtropics of the Old World – Africa, Zinj Land, Asia
and southern Europe.

Picture by Biolog
Staphylococci – gram-positive cocci in bunches – were common
inhabitants of human and animal organism in holocene. They also included very
dangerous forms, e. g. Staphylococcus aureus. A descendant of this species
entered into symbiosis with carnivorous dipteran insects and began to play
an important role in their life.
Staphylococcus sambiorum inhabits the pockets of digestive tract of sambio
and infectioflies, feeding
on the insect’s food without harming it. When the fly bites a victim (a vertebrate
animal), the bacterium enters the animal’s tissues and bloodstream and multiplies
quickly, and simultaneously produces its main “weapon” – coagulase. This enzyme
in this species possesses a very high activity and causes a very quick clotting
of the animal’s blood directly in the blood vessels. This is a fatal process,
resulting in a rather quick death of the victim. When the fly starts feeding
on the carcass, the bacteria are sucked in through the proboscis, and the
fly’s “biological weapon” is “reloaded”. The fly’s larvae receive the bacteria
with food.
Morphologically, this species does not differ from its ancestors: it is a
gram-positive coccus forming bunches. It is a facultative anaerobe, resistant
to high concentrations of salt. A chemoorganotroph, a mesophile, resistant
to immune responses of animals. Besides coagulase, S. sambiorum produces potent
leukocidins (toxins that kill leukocytes) to protect itself from the immune
system of the insect’s victim, and at the same time to help its neighbours
– fly escherichia and clostridia.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Pitcherplant-leaved
sundew leguminobacter (Leguminobacter neodroserae)
Order: Rhizobiales (Rhizobiales)
Family: Rhizobiaceae (Rhizobiaceae)
Habitat: outskirts of the Mediterranean basin.
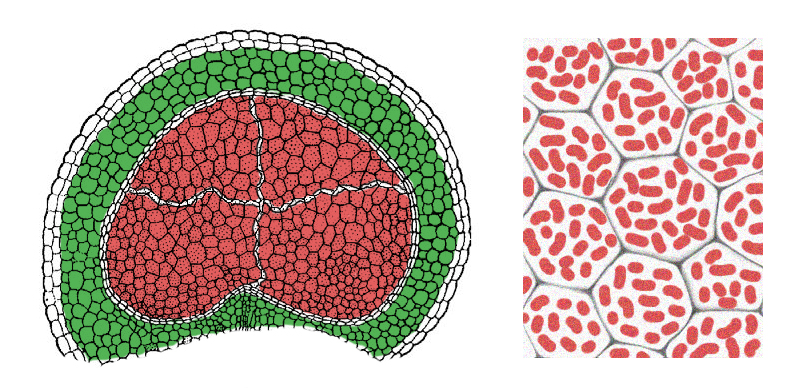
(
Picture by Biolog
In Holocene, some soil bacteria capable of fixing atmospheric
nitrogen did this in a peculiar and interesting way. The nitrogen fixers of
Rhizobium genus formed nodules on plant roots (mainly of leguminous plants).
In the nodules they, being provided with required nutrition and protection
from oxygen (since nitrogen fixation is an anaerobic process) by the plant,
in exchange provided the plant with fixed nitrogen in a metabolizable form.
But rhizobia could also live freely in the soil, contributing to the soil
part of nitrogen cycle, and underwent deep changes (formation of bacteroids)
when entering into a plant root. In Neocene, their descendants have kept and
modified this ability, and have continued the symbiotic relationships with
plants.
Leguminobacters differ from their ancestors in that they live only inside
the tissues of plants, leguminous and some other
families. And each plant species has its own species of bacteria, isolated
from others. Morphologically, it is all the same gram-negative bacteroids,
and they are still capable of fixing atmospheric nitrogen anaerobically in
the nodules. Their distinctive feature is that they mastered colonization
of the plant’s vascular bundles and get to the ovules by them, and from there
– to the embryos in seeds. During vegetative reproduction, they just pass
into a new rooting plant. These are the ways they are transmitted to new host
plants. This close union allowed such plants to settle in unsuitable, nitrogen-poor
soils, but in some cases the bacteria form nodules only when the plant is
short of nitrogen, the plant in this case sends a chemical signal to its symbionts.
The system of nitrogen fixation in leguminobacteria remained the same, but
they began to form larger nodules (the size of a pea and larger) not only
on roots, but also on other parts of the plant: on the stem and even on the
leaves. The tissues of stems and leaves with this do not lose their photosynthetic
function (the outer layers of the tissues of such nodule remain green). The
nodules have internal partitions (compartments) that contribute to more uniform
distribution of the bacterial mass and enlarge the area of absorbtion of the
fixed nitrogen by the plant. The pulp of the nodules is stained pink and even
red by leghemoglobin. Leguminobacteria have a defective metabolic system,
and therefore they cannot live outside the host plant: if the plant itself
is destroyed or a nodule is damaged, the bacteria quickly stop fixing the
nitrogen and die.
The bacteroids of leguminobacteria are very small (about 0.1-0.2 μm) polymorphic
cells with a single, partly incomplete murein layer in the cell wall. They
are organoheterotrophs by the life style, totally dependent on the host plant.
They are aerotolerant: they can live in the presence of oxygen, but the nitrogen
fixation occurs only in anaerobic conditions. They do not form capsules and
spores.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Fly
escherichia (Escherichia muscina)
Order: Enterobacteriales (Enterobacteriales)
Family: Enterobacteriaceae (Enterobacteriaceae)
Habitat: tropics and subtropics of the Old World – Africa, Zinj Land, Asia
and southern Europe.
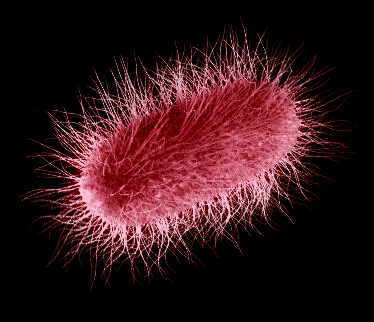
Picture by Biolog
In human era, enterobacteria were one of the most numerous
and abundant groups of bacteria. Their members inhabited all available niches
– water, soil, and animal (including invertebrates) and human organisms. Escherichia
genus was the most common among all intestinal bacteria, partly very important
intestinal symbionts and partly dangerous pathogens causing infections (toxigenic
strains). Their descendants in Neocene only weakly changed in general, but
some have switched to another life style, taking advantage of the rapid evolution
of fauna in the period of ecosystem recovery.
Escherichia muscina is the same gram-negative facultatively anaerobic rod,
motile by means of peritrichous flagella, a chemoorganotroph. But it inhabits
a new medium – the pockets of digestive tract of robberflies (infectioflies
and sambio). Having entered
into an animal’s body with the fly’s sting, it multiplies quickly and produces
a set of potent toxins, the main role belonging to hemolysins. Being produced
in large amounts, they cause a fulminant lysis of red blood cells and hemoglobin
in the animal blood, which is fatal to the animal. Along with them, the animal
is affected by the clostridial
neurotoxins and staphylococcal
coagulase. This species hardly produces endotoxins, the cells just do not
have time to be disrupted to discharge the exotoxins – the affected animal
dies too quickly.
E. muscina, like clostridia and staphylococci, multiplies quickly in the doomed
animal’s body, and the fly gets a chance to “reload” its “biological weapon”.
And the fly’s larvae, feeding on the carcass, acquire their own microbiome
of deadly symbionts.
Infusorial
photobacterium (Photobacterium endoprotistum)
Order: “Vibrionales” (“Vibrionales”)
Family: Vibrionaceae (Vibrionaceae)
Habitat: all oceans of the globe, bathyal zone.
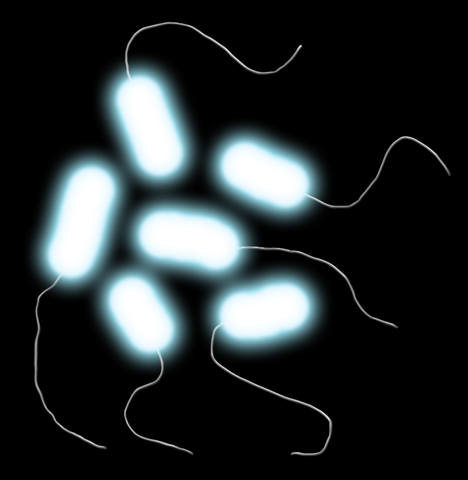
Picture by Biolog
Photobacteria, known in human era, had a peculiar feature
that allowed them to develop a specific “quorum sensing” – they were able
to glow. The luciferin-luciferase system allowed them to emit bluish light
upon forming cell aggregates. Their descendants in Neocene have kept this
ability, but switched to another life style.
Photobacterium endoprotistum is a medium-sized (1-1.5 μm) rod-shaped bacterium
with rounded ends, one of which bears a single polar flagellum. This species
is still gram-negative by the structure of the cell wall, but the cell is
covered with a slimy capsule from outside. The species is a chemoorganotroph,
and does not form spores. It is a facultative anaerobe, a psychrophile, and
a moderate barophile by the requirements to the medium it lives in. It finds
favorable conditions in the deep of the oceans.
These bacteria can live both in the water and inside the cells of a parasitic
ciliate photopiscidium (Photopiscidium
bathichthybolis), which swallows them during the stage of a young swarmer.
Inside the cell, the bacteria are protected against the ciliate’s digestive
enzymes by capsules. They lose flagella and feed from the ciliate while it
parasitizes on a fish body.
Accumulating in the ciliate’s cytoplasm, they start glowing (quorum reactions
turns on). As the ciliate passes along the fish organism to the sites of final
settling, the bacteria partly die and the luminescence weakens, but newly
released swarmers replenish the losses. Also, the bacterial number increases
at the expense of the nutrients from the ciliate’s cell.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Volvox-like
nostoc (Nostoc volvoxoides)
Order: Nostocales (Nostocales)
Family: Nostocaceae (Nostocaceae)
Habitat: still freshwaters of temperate climate in the Northern Hemisphere.
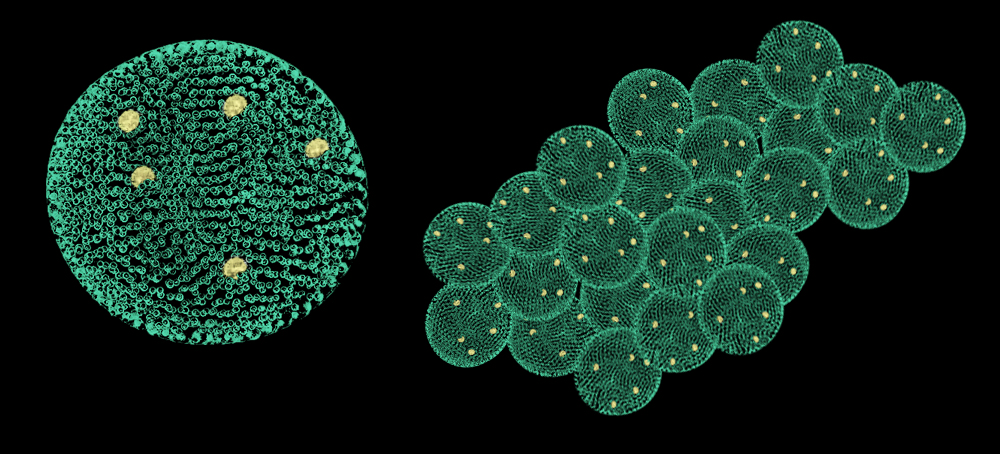
Picture by Biolog
Cyanobacteria have been the largest and the most important
group of photosynthesizing autotrophic prokaryotes during the whole history
of the Earth. They inhabited fresh and saline water bodies and the soil. Some
species entered into symbiosis with fungi as part of lichens, and even with
flowering plants, in the latter case the plant gets required nitrogen since
cyanobacteria can fix atmospheric nitrogen.
Members of Nostoc genus were common in fresh water bodies and in moist soils
where they formed spherical jelly-like colonies. Some of their descendants
in Neocene have switched to another life style.
Nostoc volvoxoides has kept the cellular morphology of its ancestors. Its
cells, with a membrane system (thylakoids) bearing photosynthetic pigments,
instead of chains form dense single-layer mats closed into hollow balls the
size of a pea, covered with a gelatinous capsule. Large heterocysts (specialized
cells that fix atmospheric nitrogen) inherited from ancestors stand out among
the ball’s cells. Also, the species has large floater cells with gas bubbles
in the cytoplasm, which allow it to soar in the subsurface water layers (and
even to surface up and protrude from the water) where the colony gets maximum
light and nitrogen.
The balls are stained usually blue-green; pale yellowish spherical heterocysts
and almost hyaline “gas” cells stand out on them; both heterocysts and “gas”
cells are 1.5-2 times larger than usual cells.
The cells of N. volvoxoides reproduce by fission, and daughter cells are deposited
inside the ball, while mother cells die gradually. Thus, 2-3 daughter colonies
form inside the ball, they are released via rupture of the mother colony as
it dies.
In favorable conditions, N. volvoxoides multiplies in large numbers and causes
blue-green water bloom. Living and dead colonies in this can stick together
to form carpets (mats) on the water surface. The carpets reach 1-2 meters
large and are so solid that insects and other small animals walk or crawl
freely on them. Small numbers of this “alga” are consumed by herbivorous fish
and also by land herbivorous mammals.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
Symbiotic
anabaena (Anabaena schizolemnae)
Order: Nostocales (Nostocales)
Family: Nostocaceae (Nostocaceae)
Habitat: fresh waters of the North Hemisphere in cold climate zones, a symbiont
of Schizolemna anabaenophila
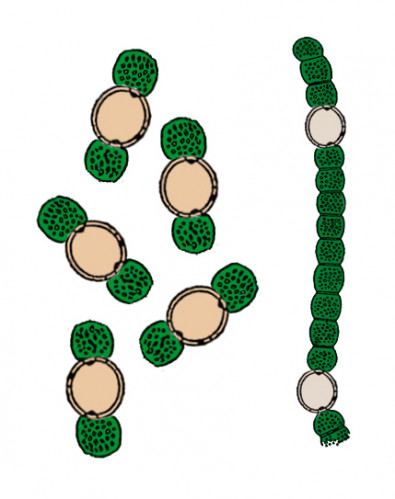
Picture by Biolog
During the whole history of life on the Earth, cyanobacteria
repeatedly entered into symbiosis with various plants. Receiving “shelter”
from the plant, they provided it with nitrogen which they fixed from the environment.
Some species of such symbionts, particularly of Anabaena genus, inhabited
tissues of aquatic plants, e. g. Azolla fern and angiosperms duckweeds (Lemna).
Their descendants in Neocene have evolved and changed according to the changes
in the structure and life style of their hosts – the duckweeds.
Anabaena schizolemnae has undergone significant morphological changes. It
has two morphologically different stages: a free stage living in water and
a symbiotic stage living in the tissues of Schizolemna,
a duckweed.
The free-living stage appears as long filaments composed of green barrel-shaped
cells, alternating with large spherical yellowish heterocysts – this appearance
is characteristic of ancestral and related forms. And the symbiotic stage
appears as a heterocyst about 10 μm in diameter with two blue-green photosynthetic
vegetative cells on the sides. These cells are 1.5 times smaller than the
heterocyst, and provide its development. Such structure allows A. schizolemnae
to provide the plant with nitrogen (fixed by heterocysts) at a maximum efficiency
with keeping its own photosynthesis. A large number of such three-cell symbionts
accumulates in the duckweed’s tissues, and the compactness allows to distribute
more uniformly, giving the Schizolemna’s symbiotic tissues a brownish-green
color. During vegetative reproduction of the host plant by fission of the
leaflet, the symbiont’s cells distribute in it equally enough. In the Schizolemna’s
growth process, the vegetative cells of A. schizolemnae divide to form new
undifferentiated cells outside of the heterocyst. When their number in a chain
reaches three, they secede and differentiate into a heterocyst and vegetative
cells. Undifferentiated cells of A. schizolemnae penetrate even the ovules
of the duckweed, and in rare cases of seed reproduction of the host plant,
the seedlings already bear the symbiont in their tissues. In the fall, the
duckweed plants stop the photosynthesis, accumulate starch and submerge into
the water to winter there. In these conditions, A. schizolemnae forms spores
that can winter in the duckweed’s tissues.
The free stage forms only when the symbionts for any reason fall out of the
duckweed’s leaflet into the water. They cannot enter the organism of the same
or another duckweed again, but instead they divide to restore filamentous
structure, and live independently. Since such situation is quite rare, A.
schizolemnae is almost absent in water blooms.
This species of bacteria was discovered by Biolog, the
forum member.
Translated by Biolog.
“Blood-of-the-salt”
halobacterium (Halobacterium sanguinisalis)
Order: Halobacteriales (Halobacteriales)
Family: Halobacteriaceae (Halobacteriaceae)
Habitat: hypersaline wetlands of Mediterranean basin, in water and on salt
deposits.

Picture by Biolog
In human era, Archaea inhabited various niches with extreme
conditions, mainly elevated temperature, very high acidity, or salt concentrations
at a level of strong brine. They possess such abilities due to biochemical
differences from bacteria and eukaryotes: cell walls in most of them lack
murein and consist of highly resistant proteins, and the membrane lipids have
a unique structure, also with high resistance. Their descendants in Neocene
have kept most of the ancestral features, and this helped them to master new
niches.
Halobacterium sanguinisalis inhabits the world of salt in the Mediterranean
basin. It has inherited the ability of bacteriorhodopsin-mediated photosynthesis
from its ancestors, whose cells were stained red by bacteriorhodopsin. With
this, it does not assimilate carbon, but obtains it from other sources. Like
its ancestors, H. sanguinisalis accumulates sodium and potassium salts inside
the cell (crystals in the cytoplasm) to resist the osmotic drain of water
from the cell to the environment. But the life under scorching sun has resulted
in emergence of a new adaptation – accumulation of additional pigments that
protect the cell from excess solar UV. These pigments give the rhomboid cells
of this species a dark red-purple color resembling human venous blood – hence
its name. The rhomboid cells are 8-10 μm large, flat and very thin, they adjoin
each other like an argyle pattern and stick together, forming plates of 10-20
cells (up to 100 μm large). The edge cells bear archaella – transformed archaeal
flagella. The whole plate can swim freely by using them.
Breeding in large numbers, these organisms cause a dark-red color with a purple
tint of brine, and also of salt formations and crystals where enough moisture
is present on them for the microorganisms to develop.
H. sanguinisalis participates in the phosphorus cycle, converting its compounds,
and also enters into symbiosis with a filamentous
alga Filodunaliella mediterranea, from which the archaeon gets carbon
as glycerol. The filaments of F. mediterranea often cling to the plates of
the halobacterium and float in the brine together with them.
This species of archaea was discovered by Biolog, the forum
member.
Translated by Biolog.