
| Главная | Библиотека | Форум | Гостевая книга |
Живые драгоценности влажных тропических лесов
Неотропической области: группа пчёл-эуглоссин (Euglossa spp.) вьётся и
садится на цветы орхидеи (Clowesia thylaciochila). Увеличенная вставка
показывает пчелу, которая только что посетила цветок и несёт на своей
груди похожие на нагрудный значок жёлтые поллинии орхидеи – мешочки, содержащие
пыльцевые зёрна. |
ГЛАВА 3
ВСПОМИНАЕТ СТИВ:
Это было в самом начале дождливого сезона, когда трястись на
ухабах вечно грязной Трубопроводной дороге в Панаме было ещё достаточно легко
– мы смогли бы даже приехать вовремя, чтобы проделать работу, как запланировали.
Мой друг за рулём привык работать и в гораздо худших условиях, чем сейчас. Я
также был хорошо знаком с этой узкой извилистой дорогой, много раз приземляясь
на ружьё, когда массивные ребристые шины зелёной «Тойоты» катили по колее, прорезающей
«Национальный парк Соберания» – последний равнинный первичный тропический лес,
оставшийся в зоне Панамского канала. Мы дали водителю Дейву Рубику прозвище
«Бешеный пёс», потому что он был одним из самых отчаянных тропических биологов.
В академических кругах доктор Рубик был известен своими исследованиями так называемых
безжалых пчёл, которые опыляют многие из растений в этом лесу. Разносторонние
интересы Рубика, однако, выходят далеко за рамки изучения безжалых пчёл, и простираются
от печально знаменитых (жалящих) африканизированных пчёл до современного джаза.
В этот день я собирался помочь Дейву Рубику разложить пахучие приманки, чтобы
привлечь самцов великолепной группы пчёл, известной как эуглоссины – этих летающих
драгоценностей тропического леса.
К тому времени, когда мы сели в «Тойоту» на рассвете, дождь уже шёл. Его стук,
усиленный шумом цикад и кобылок, почти заглушал ритмы джаза из стерео Рубика,
пока мы петляли по дороге к нашей самодельной рабочей станции на дорожной отметке
«Десятый километр». Внезапно Дейв яростно затормозил, и меня отбросило на отделение
для перчаток, в котором он хранил свои вонючие приманки. Я взглянул вперёд как
раз вовремя, чтобы заметить огромную пятнистую кошку, которую мы едва не сбили.
Моё сердце бешено колотилось, потому что я осознал, что кошка, скрывающаяся
в подлеске, была ягуаром – первым, которого и Дейв, и я увидели в дикой природе.
Это был ещё один знак того, что Национальный парк Соберания сохранил участки
девственного тропического леса – редкость в Панаме в наше время.
Наконец мы припарковались и продолжили давно начатую работу – составление ежемесячной
переписи пчёл-эуглоссин, чем Рубик и его коллеги непрерывно занимались на протяжении
более чем 100 месяцев. В этом густо заросшем лесом местообитании мы не должны
были упорно искать пчёл на цветах – они сами искали бы нас. Мы хотели привлечь
их тремя синтетическими химическими приманками: цинеолом – с лекарственным запахом
мази «Vicks Vapo-Rub», метилсалицилатом – с сильным конфетным ароматом, похожим
на леденцы с «зимней свежестью», и печально известным скатолом – с сильнейшим
запахом свежего, тёплого помёта млекопитающих.
Рубик хранил все три пахучих вещества в контейнерах фирмы «Тапперуэр», но проявлял
особую осторожность при обращении со скатолом – его контейнер буквально вопил
любопытному: «ЗЛОВОННО – ХРАНИТЬ ПЛОТНО ЗАКРЫТЫМ!» Достаточно небольшого зазора
– и эта острая вонь держалась бы на нашей одежде и коже несколько дней. Но Рубик
проходил через эту энтомологическую рутину так много раз, что теперь она была
его второй натурой. Мы предлагали насекомым эти три приманки на кусочках промокательной
бумаги, прикрепляемых к деревьям у обочины на высоте груди. А потом мы с Дейвом
раскладывали алюминиевые садовые стулья посреди дороги, наблюдали за полётом
искристо-голубых бабочек-морфо, слушали трубный гул цикад и ждали, пока разноцветные
с металлическим блеском самцы пчёл-эуглоссин последуют по воздушным запаховым
дорожками к нашим смоченным приманкой бумажкам.
Нам не пришлось ждать слишком долго. Уже в первые пять минут я услышал высокий
гул любопытного самца целиком зелёной Euglossa imperialis со свисающим вниз
хоботком такой же длины, как и его тело. Вскоре до бумажек добрались другие
самцы пчёл-эуглоссин, скрыв их под блеском своих металлических зелёных, пурпурных,
медных и золотых красок, сверкая, словно драгоценные камни, в пылающем солнечном
свете тропиков. Многие из них ненадолго взлетали, хотя некоторые бродили вокруг
бумажек в течение нескольких минут, если их не пугали.
Иногда Дейв вставал со стула, подходил к одной из бумажек и быстро захватывал
пчелу кончиками пальцев, чтобы определить её. Похватав руками и отпустив за
эти годы уже десятки тысяч таких пчёл, он мог быстро определить любой из 57
видов, которые жили по соседству в этом тропическом лесу. Все они пользуются
специализированными, уплощёнными в скребки волосками на передних ногах, чтобы
выскрести запах из рваных волокон промокательной бумаги. Они нетерпеливо собирали
летучие запахи с промокательных бумажек, поскольку распознавали их как те же
самые химические вещества, которые имеются у большинства орхидей в тропическом
лесу Соберании.
Послание было очевидным: эти запахи служили указателями для пчёл – так же, как
отметка «Десятый километр» была ориентиром для нас с Дейвом. Однако остаётся
неясным, как пчёлы на самом деле используют химические вещества, которые собирают.
Какие преимущества, возможно, могли получать пчёлы, добавляя некие запахи к
своему репертуару полового поведения?
Пчёлы подсемейства эуглоссин были достаточно плохо известными до конца 1960-х
годов, когда Каллоуэй Додсон и Роберт Дресслер воспользовались методом газовой
хроматографии для того, чтобы проанализировать химию некоторых пряных ароматов
неотропических орхидей – ключ, как они считали, к экологии эуглоссин. К тому
времени в музейных коллекциях насекомых находилось мало экземпляров самцов пчёл-эуглоссин,
потому что никто не нашёл надёжного способа их отлова. (Это ещё остаётся особенно
справедливым для самок и в наши дни.) После выяснения состава некоторых из этих
остро пахнущих химических зелий из запахов, напоминающих мяту, корицу и ваниль,
учёные заметили, что самцы эуглоссин в больших количествах собираются на любой
материал, смоченный этими имитациями цветочных ароматов. Они начали прикреплять
промокательные бумажки, пропитанные каждая своим типом химических веществ, к
коре тропических деревьев, как сделал Рубик в Панаме, и как мы сделали тогда
на северных границах тропических лесов, где они соседствуют с мексиканской пустыней
Сонора. И – вуаля! Малоизвестные пчёлы, кажется, появляются из ниоткуда, пополняя
собой разнообразие эуглоссин, которое и представить себе нельзя было до тех
пор, пока любопытными натуралистами не стали повсеместно использоваться запахи.
В начале 1960-х годов учёные сперва полагали, что самцы соблазнялись орхидеями,
не дающими им вознаграждения в виде пищи, потому что цветки имитировали смолистые
гнёзда, построенные самками пчёл. Позже предположили, что цветочные ароматы
использовались как аттрактанты, так как выяснилось, что самцы пчёл используют
запахи как точку сбора, к которой они могли бы сходиться, устраивая место для
свиданий, похожее на токовища, используемые в качестве мест для брачных демонстраций
птицами вроде полынного тетерева. Однако, по мере того, как накапливалось всё
больше информации, стало ясно, что эти пчёлы находятся в зависимости от острых
запахов цветков орхидей, которые являются прекурсорами для их собственных половых
феромонов, нужных для привлечения к себе самок для спаривания. По странному
капризу природы пчёлы могут красть сексуальное послание орхидеи и превращать
его в свой собственный афродизиак.
Чтобы использовать запахи, счищенные с лепестков орхидеи (или с промокательной
бумаги), пчёлы каким-то образом «перерабатывают» летучие соединения в увеличенных
губчатых железах в своих задних ногах. Очевидно, пчёлы смешивают их со своими
собственными химическими выделениями или иначе преобразуют их биохимическим
путём. В любом случае такие афродизиаки, представленные в виде аэрозоля, выделяемого
группой самцов пчёл, собирающихся и устраивающих демонстрации на «токовище»,
оказывают непреодолимое действие на их потенциальных брачных партнёрш.
Некоторые детали этой истории остаются лишь предположительными. Это не такая
уж и лёгкая задача – подтвердить в условиях тропического леса, что самки эуглоссин
выбирают своих брачных партнёров по принципу «у кого лучше пахнет лосьон после
бритья». Однако такой выбор брачного партнёра по запаху не вызвал бы особого
удивления, так как пчёлы являются превосходными химиками. На самом деле среди
пчёл есть и другие известные примеры выбора брачного партнёра при помощи обоняния.
Самки пчелы-плотника (Xylocopa varipuncta) на американском Юго-западе выбирают
себе брачных партнёров, будучи привлечёнными их сладким ароматом, далеко распространяющимся
по ветру. Это продемонстрировали, используя синтетический феромон самца пчелы-плотника
для привлечения самок этого вида, Стив и его коллеги по работе в поле – Уильям
Всисло, в настоящее время в Смитсоновском институте тропических исследований
в Республике Панама, и Роберт Минкли из Обернского Университета – на нескольких
участках в пустыне Сонора близ Финикса и Тусона.
Сама плотность скопления пчёл в каждом листе приманки из промокательной бумаги
позволяет легко узнать, активно ли самцы эуглоссин опыляют растения в ближайших
окрестностях. Конечно, эти временные скопления служат лишь косвенным признаком
их местного изобилия на самих цветах. Чтобы узнать об этом поточнее, вам нужно
будет подняться высоко в полог тропического леса и поискать цветущие орхидеи,
на которых видно, что их пыльцу уже унесли. Есть несколько простых способов
получить прямое свидетельство посещения пчёлами этих эпифитных цветков – цветков,
которые укореняются или прицепляются высоко на стволах деревьев.
Как только пчела поймана, вы легко сможете разглядеть большие жёлтые сдвоенные
выпуклости, торчащие на её голове. Каждая из них представляет собой свежий поллиний
– ярко-жёлтый мешочек пыльцы, который прикрепился к этому самцу, пока тот добывал
на орхидее её восхитительный сексуальный парфюм. Поллинии орхидей – это изумительные
изобретения, особый способ упаковки зёрен пыльцы в контейнеры с плотной оболочкой
для переноски их животными. Поллинии орхидей напоминают два ярко-жёлтых яичных
желтка, прикреплённых к кожистому, но гибкому коричневому стебельку. Когда пчела-эуглоссина
протискивается сквозь ранее не посещавшийся цветок орхидеи, поллинии приклеиваются
к её панцирю, часто на труднодоступных «островках безопасности», откуда пчёлы
не могут удалить их даже самыми энергичными движениями передних ног – аналогично
тому, как вы с трудом сможете почесать зудящую поясницу. В течение нескольких
дней пчела-эуглоссина может таскать поллинии, прикрепившиеся в таких местах,
которые могут показаться людям смешными – между глаз, на макушке, торчащими
на брюшке – пока не посетит другой цветок орхидеи того же самого вида, который
может отцепить свою награду с злополучного посредника. Когда самец эуглоссины
заползает в другой цветок орхидеи на спине, животе или боку, в зависимости от
формы цветка, поллинии в буквальном смысле «отстают». Давление цветка заставляет
два мешочка пыльцы оторваться – их сдирает восприимчивое клейкое рыльце, успешно
гарантируя перекрёстное опыление.
За те годы, что Рубик и его коллеги месяц за месяцем осматривали пчёл, по их
оценкам значительно меньше 5 процентов из числа пчёл, собирающихся на промокательных
бумажках, недавно посещали орхидею – у них отсутствуют «значки» из мешочков
пыльцы и клейкие отметины. Учёные заинтересовались тем, почему так мало особей
в этих местах носит поллинии орхидей. Это представление в целом соотносится
с другими исследованиями опыления орхидей: не все посещения пчёл заканчиваются
успешным взятием пыльцы или переносом её с одного цветка на другой. Даже при
том, что исследования Рубика говорят, что пчёлы-эуглоссины Соберании имеют более
стабильную по годам численность популяций, чем почти все остальные насекомые,
когда-либо изучавшиеся, нет никаких признаков того, что посещения пчёл и доступность
поллиниев орхидей находятся в состоянии некоего «совершенного равновесия», которое
подразумевали ранние трактовки мутуализма между растениями и животными из учебников.
Хотя пчёлы-эуглоссины зависят от пыльцы и нектара, собранных с растений других
семейств, в плане питания, они обладают многочисленными специализированными
адаптациями, которые помогают им извлекать выгоду из ароматов цветков орхидей.
В свою очередь формы цветков тропических орхидей, размер пыльцевого мешочка
и его размещение на пчёлах, а также «клей» поллиниев – всё это указывает на
некоторый уровень адаптации цветка к эуглоссинам в качестве агентов распространения
пыльцы. Однако сообщения о «непоколебимой» приверженности каждого вида пчёл-эуглоссин
к партнёрству исключительно с одним особым видом орхидеи-хозяина не получили
подтверждения в недавнем исследовании специалистов по экологии растений.
Теперь мы знаем, что каждый отдельно взятый вид орхидей часто посещается «гильдией»
пчёл-эуглоссин, часто из нескольких родов, и каждого из них привлекает свой
особый аромат. Чтобы получить свою собственную специфическую смесь ароматов,
пчела может посещать несколько видов орхидей, грибов, или даже сочащиеся раны
на дереве. Орхидеи, в свою очередь, выработали способы, гарантирующие то, что
их поллинии будут перенесены только на другой цветок их собственного вида и
не будут «разбазарены», оставленные на цветке совершенно иного вида. Каждый
род орхидей – а иногда также и некоторые виды – обладает заранее определённой
«картой» мест на теле посещающей его пчелы, куда приклеивается его драгоценный
груз пыльцы. Во время каждой последующей встречи с цветком этого же вида специфическая
форма цветка вынуждает пчелу вертеться и разворачиваться так, что эти движения
увеличивают вероятность успешной передачи нужным образом размещённого пакета
пыльцы. Этот не просто гарантирует опыление и последующее оплодотворение: это
также гарантирует перекрёстное опыление между растениями одного и того же вида.
Независимо от того, сколько видов пчёл посещает одну и ту же орхидею, все они
носят её наклейки с мешочками пыльцы на одном и том же месте, прежде чем сдадут
их орхидее того же вида.
Для цветов выгоды, полученные от животных, очевидны: перенос и правильное размещение
пыльцы – именно поэтому мы называем этих животных «опылителями», а не просто
«посетителями цветов». Но какую пользу получают от цветов животные? Это гораздо
более скользкий вопрос. Ответы на него так же различны, как различны виды пользы,
предоставляемой растениями: защита от непогоды, укрытие от хищников, безопасные
убежища для спаривания, ключевые точки для защиты территории, места засады для
охоты, а также притягательные источники питательного нектара и пыльцы, смол,
масел, снадобий, духов и других дополнительных химических соединений. И как
мы уже видели, некоторые пчёлы подсемейства эуглоссин могут преобразовать запахи
орхидей для изготовления своих собственных сексуальных приманок.
Если это польза, то какова её цена? Иногда биологи определяют «цену услуг по
опылению» только в рамках цены калорий, сожжённых опылителями, перемещающимися
от одного цветка к другому в границах своей территории – по своему кормовому
маршруту. Например, затраты на перемещение колибри-отшельника, движущегося по
кормовому маршруту между цветками геликонии металлической могут быть высокими,
поскольку эти растения растут зачастую более чем в сотне ярдов одно от другого,
заставляя колибри-отшельника преодолевать почти милю по лесным воздушным коридорам,
чтобы получить достаточно нектара для питания во время своего утреннего путешествия.
Но разнообразные затраты на услуги по опылению могут также рассматриваться с
позиции эволюционных представлений, то есть как вклад в специализированные морфологические
и химические адаптации, а также ограничения, которые специализация накладывает
на использование других ресурсов. Например, чтобы добыть нектар одного особого
вида Heliconia, колибри сопутствующего ему вида должны поворачивать свою голову
под углом более чем на 90 градусов, почти вверх тормашками, чтобы позволить
своим клювам добраться до скрытых нектарников. Конечно, их клювы также должны
иметь соответствующую длину и кривизну. Геликония в изобилии предлагает богатый
энергией нектар своим жужжащим друзьям, которые могут совершить этот акробатический
трюк. Как отметил орнитолог Стив Хилти, «при этом пыльца помещается на подбородок
и в основание клюва, тем самым снижая вероятность того, что пыльца [этой геликонии]
позже окажется на цветках неправильного вида растений». Иными словами, когда
переносящий пыльцу колибри погружает клюв в трубчатый цветок нужного вида, форма
цветка определяет точность переноса пыльцы с подбородка или клюва; в цветке
неподходящего вида пыльца может быть стёрта, но попадёт в неправильное место,
вдалеке от восприимчивого к ней рыльца.
Поэтому применительно к опылителям затраты на посещение цветов можно оценивать
множеством различных способов. В дополнение к непосредственным затратам энергии
на полёт от одного цветка к другому существуют также последующие поведенческие
и метаболические затраты на сбор, перенос и обработку вознаграждения, полученного
от цветов. Пчёлы-эуглоссины должны не только переносить поллинии; они должны
также обрабатывать цветочные запахи, прежде чем те приобретут адаптивную ценность.
Их визиты на цветы орхидеи, отнимающие много времени, также делают их лёгкой
мишенью для хищников – клопов-крупноглавов и пауков-бокоходов – или для таких
паразитов, как клещи. Все эти затраты достаточно заметны: биолог-счетовод может
приблизительно подсчитать время, которое пчела-эуглоссина тратит на сбор корма,
или вероятность того, что она будет атакована хищником или паразитом.
Но эволюционную цену сосредоточения на вознаграждении орхидеи трудно спрогнозировать
даже самому способному счетоводу. Пчёлы-эуглоссины – это очень энергичные летуны,
способные преодолевать длинные кормовые маршруты, и вдобавок снабжённые сенсорным
аппаратом для обнаружения орхидей на пути следования. У них есть специализированные
органы – хоботки, которые напоминают рыболовные удочки – позволяющие им добираться
до пищи, скрытой глубоко внутри цветков. Также вероятно, что значительная часть
процесса обработки пчелой аромата орхидеи и использования его в качестве афродизиака
запрограммирована генетически. Возможно, следуя на ранних стадиях становления
своей эволюционной линии по пути специализации на сборе половых аттрактантов
с орхидей и из других источников, эти сильно специализированные настоящие пчёлы
оказались запертыми в своей роли сборщиков запахов. И как же самцам пчёл-эуглоссин
приходится делить своё время между цветами, которые богаты запахом, и цветами,
которые предлагают кормовой нектар? (Самцы не заинтересованы в сборе пыльцы
с цветов.) Это возможность выбора затрат на время кормления, с которой мало
кому из других пчёл приходится сталкиваться в жизни.
Однако для нескольких других сочетаний растений и опылителей эволюционные затраты
такого обязательного мутуализма были приблизительно рассчитаны. Легче всего
это оказалось сделать для насекомых, которые вознаграждаются за услуги по опылению
питательными семенами для своего потомства, как в случае с купальницей Trollius
и мухами, которые размножаются в её цветах. Количество яиц мухи, найденных в
цветке, почти точно соответствует расчётному балансу между затратами и выгодами,
предполагаемыми при поддержании этого мутуализма на протяжении долгого времени.
Однако классический пример такого мутуализма «семя за семя» наблюдается у пустынной
юкки и её опылителя-моли – облигатная зависимость, впервые открытая в 1876 г.
Чарльзом Валентайном Райли, всего лишь за два года до того, как он стал национальным
энтомологом в правительстве Соединённых Штатов. Основываясь на кропотливых исходных
наблюдениях Райли, такие учёные, как Джон Аддикотт, Джудит Бронштейн и Олле
Пеллмир, продемонстрировали, как у некоторых видов самка моли обменивает размещение
пыльцы на рыльцах юкки на небольшую часть жизнеспособных семян для прокорма
своего потомства. Эти самки юкковых молей обладают специализированными ротовыми
органами, которые позволяют им собирать пыльцу с бледных бархатистых цветков
юкки – пыльцу, которую они затем носят с собой большую часть своей недолгой
жизни. Также существуют моли-«подражатели» или «разбойники», которые пользуются
преимуществами этого мутуализма, но их нашли ещё не у всех видов юкки.
Пример мутуализма, наблюдаемого в пустыне Чиуауа
в Аризоне, Нью-Мексико и Мексике, где растения юкки вроде этой Yucca elata
опыляются специализированной молью рода Tegiticula. Моль активно собирает
пыльцевые зёрна, сгребает их в шаровидную массу своими удлинёнными ротовыми
органами и заталкивает пыльцу в восприимчивое к ней рыльце пестика. Мы
не знаем ни одного другого опылителя, который преднамеренно опыляет цветок.
Эта моль поступает таким образом, чтобы гарантировать запас семян для
своих прожорливых личинок, развивающихся внутри незрелых плодов. |
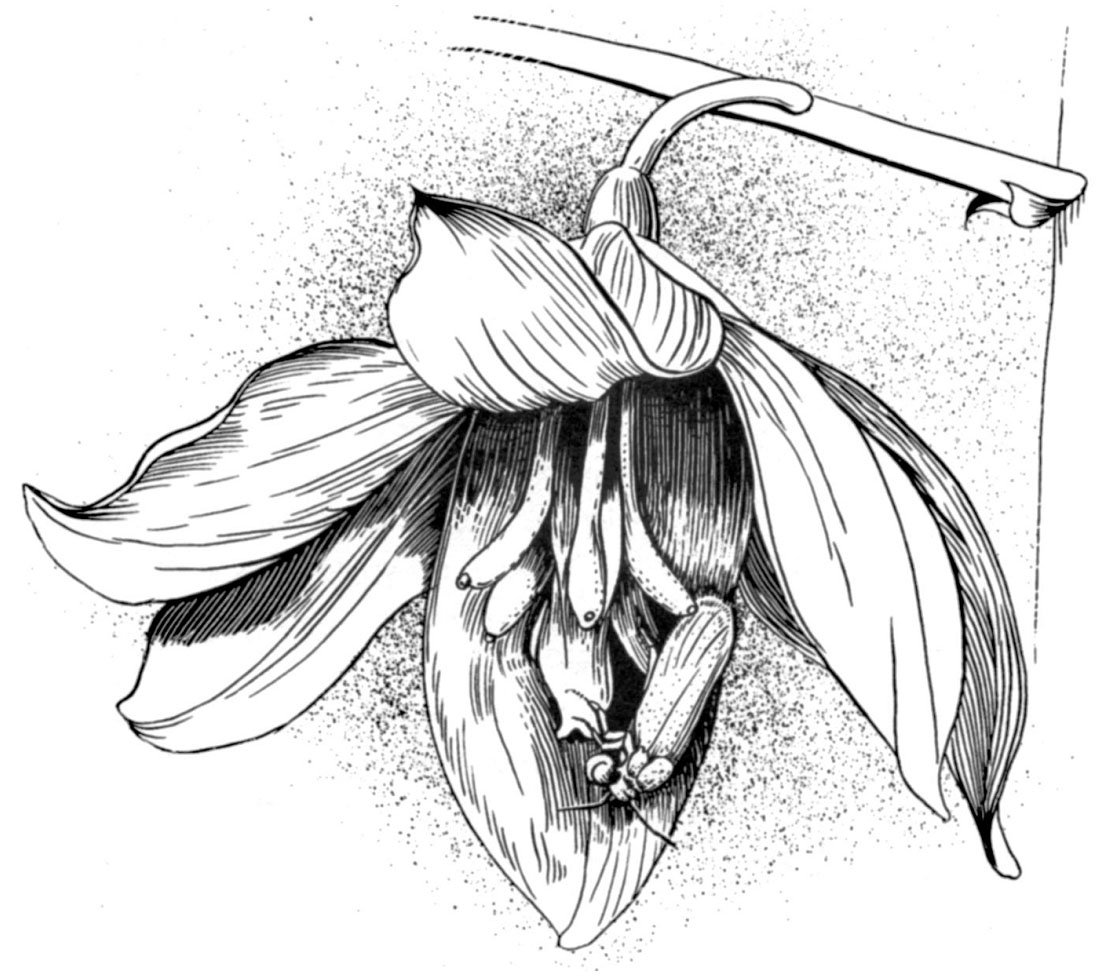
Олле Пеллмир из Университета Вандербильта описал, что происходит, когда моль прорезает завязь цветка юкки и откладывает внутри неё свои яйца:
Затем она приближается к рыльцу и активно закладывает небольшое количество пыльцы. Поступая таким образом, она гарантирует, что доступность развивающихся семян не будет ограничена отсутствием пыльцы, ведь это единственная пища её потомства... После появления на свет личинка потребляет лишь малую часть развивающихся семян, оставляя многие из них нетронутыми. Возвратная специализация растения путём исключения иных опылителей привела к появлению облигатной взаимной зависимости между юкками и их опылителями, поскольку ни один из этих видов не может успешно размножаться без другого.
Согласно Пеллмиру, это тот случай, когда исчисление энергетических
затрат становится очень интересным. Если бы моль могла увеличить число яиц,
откладываемых в плод, она получила бы выгоду, но воспроизводство растения, вероятно
пострадало бы из-за меньшего количества способных к прорастанию семян, что стало
бы закономерным итогом большего успеха моли. Вместо этого Пеллмир и его коллега
Чед Хат обнаружили, что юкки склонны сбрасывать цветы, содержащие большое количество
отложенных яиц, особенно если цветок был плохо опылён молью. Иными словами,
естественный отбор благоприятствует моли, которая откладывает как раз нужное
количество яиц, чтобы обеспечить размножение, но многократно и надёжно опыляет
этот самый цветок. С эволюционной точки зрения моли, которые перегружают развивающийся
плод слишком большим количеством яиц, а также бездельники, когда речь идёт о
предоставлении услуг по опылению, не имеют преимуществ: цветок сбрасывается,
семена перестают развиваться, личинки моли остаются без пищи и умирают. Это
избирательное сбрасывание плодов поддерживает симбиоз между молями и юкками
устойчивым на протяжении эволюционного времени, так, чтобы выгода от приспособленности
одного вида не ложилась бременем на другой.
Конечно, окончательная цена этого мутуализма совершенно очевидна: каждый из
мутуалистов терпит неудачу в размножении, когда не может найти другого в нужном
месте в нужное время. Джудит Бронштейн из Аризонского университета наблюдала
такое явление на краю травянистых равнин в горах Санта-Рита, всего лишь в 30
милях к югу от своей лаборатории, где она постоянно исследует биологию юкки
высокой (Yucca elata). Бронштейн пишет: «юкки в Санта-Рита цвели на месяц позже,
чем в прошлом году, и закончили цветение не синхронно с появлением большинства
особей юкковой моли. В результате плоды юкки, в которых завязались семена, были
самыми ранними; растения, которые цвели позже, цвели совершенно впустую».
Мы наблюдали более долговременное нарушение мутуализма между юккой и молью,
работая в парке Папаго в Аризоне – сейчас это деградировавшая природная область
в центре Финикса – столицы штата. Вплоть до 1930-х гг. фотографии парка Папаго
показывают, что юкки ягодоносные (Yucca baccata) представляли собой заметную
отличительную особенность естественной растительности тех мест. Возможно, из-за
взрыва численности грызунов и кроликов, последовавшего за сокращением численности
хищников в этой местности юкки, похоже, оказались просто выгрызенными начисто.
Единственные юкки, остающиеся в парке Папаго – те, что были высажены там в пустынном
ботаническом саду в 1940-х и 1950-х гг., и они цветут лишь местами, и то не
каждый год. Культивируемые в саду юкки никогда не давали семян – или моль исчезла
в этих местах, когда снизилась численность местной популяции, или количества
цветущих в настоящее время растений недостаточно для поддержания существования
жизнеспособной популяции личинок моли.
Одной из самых парадоксальных эволюционных затрат, сопровождающих партнёрство
по опылению в мутуалистических отношениях, оказывается уязвимость для «мошенников»
и «разбойников» вроде дополнительных видов молей, которые в настоящее время
ассоциированы с партнёрством моли и юкки. Эти оппортунисты присваивают преимущества,
даваемые растением-хозяином, но не обеспечивают взаимной выгоды. Также существуют
разбойники и мошенники, пользующиеся преимуществами, которые дают пчёлам, молям
и колибри мутуалистические связи с различными цветами. Например, некоторые особи
колибри пилоклювого рамфодона прокалывают венчики тропических цветов, чтобы
получить нектар в обход пыльцы, истощая тем самым запас пищи, доступной эуглоссинам
или колибри, которые проникают в цветок со стороны его зева.
Конкуренция с мошенниками и разбойниками может не только ограничить ресурсы,
доступные надёжному опылителю-мутуалисту, но и снизить успешность плодоношения
того цветка которому он тысячелетиями предоставлял услуги опылителя. К счастью
для растений, численность мошенников и разбойников редко превосходит численность
мутуалистов, которых привлекает та же самая совокупность цветков, иначе коэволюционирующее
сообщество могло бы прийти в упадок. Несомненно, существует пороговая плотность
растений-хозяев – точка, за которой такие «паразиты» не получат поддержки в
своих обманных играх. Мексиканский эколог Франсиско Орнелас определил, что численность
многих ворующих нектар колибри значительно ниже, чем у тех видов, которые предоставляют
услуги опылителей:
У такого рода предприимчивости – получения нектара в обход пути, используемого опылителями – есть одно следствие: редкость данного явления. Представители третьего вида используют преимущества мутуалистических содружеств и попадают в зависимость от плотности мутуалистических взаимодействий. Редкость расхитителей нектара стала бы эволюционным результатом специализации растения к определённому набору истинных опылителей... Ворующие нектар колибри выглядят редкостью в природе и/или отклонением от нормы в местных масштабах.
Любой разрыв связей между растением и его истинными опылителями,
который снижает обилие цветов, также ставит в опасное положение разбойников
и мошенников. Поэтому естественная редкость расхитителей нектара может усугубиться
либо сведением леса, которое напрямую уничтожает имеющиеся источники нектара,
либо снижением численности «законных» опылителей, которое в конечном счёте ведёт
к снижению репродуктивного успеха в популяции кормового растения.
Случаи облигатного мутуализма – в частности, симбиоз между единственным видом
опылителя и единственным видом растения – составляют лишь небольшой процент
от общего числа взаимоотношений между растением и опылителем: риск сверхзависимости,
должно быть, слишком велик, чтобы такие отношения сложились у многих видов.
Тем не менее, даже при том, что случаи облигатного партнёрства редки по сравнению
с взаимодействиями, складывающимися между неспециализированными видами, они
зачастую оказывают непропорционально большое влияние на структурирование сообществ
растений и животных. Лучший пример такого рода можно найти во влажных тропических
низменностях по всему миру, где деревья фикуса-душителя играют ключевую роль
в прореживании леса и поддержании существования плодоядных видов.
В целом ряде стран, от Перу до Новой Гвинеи, фикусы представляют собой критически
важный ресурс для самых разнообразных животных в сообществах тропического леса
– от летучих мышей и приматов (в том числе людей) до попугаев и райских птиц.
В некоторых лесах фиги составляют до 70 процентов рациона позвоночных. В мире
существует более 750 разных видов фикусов, и большинство из них полагается на
различные виды крохотных ос как на единственных опылителей. А эти осы, в свою
очередь, зависят от части развивающихся семян фикуса, используемых в пищу на
критически важной стадии их жизни. Вот, что пишет Джудит Бронштейн:
Фикусы были названы замковым камнем среди мутуалистов тропических лесов – замковым камнем называется тот, который скрепляет всю арку. Основная мысль здесь состоит в том, что, если вы начнёте убирать замковые камни, это станет бедствием для многих животных, которые полагаются на них... Это могло бы случиться, например, при выборочной заготовке деревьев, на которых селятся фикусы-душители. Или же, распыляя инсектициды, вы истребляете ос, что в дальнейшем приведёт к тому, что их деревья-хозяева станут более редкими, или, возможно, вымрут, поскольку деревья не смогут рассчитывать ни на каких других местных опылителей. Они должны полагаться на самых высокоспециализированных фиговых ос для достижения успеха в размножении. Оба мутуалиста исполняют причудливый эволюционный танец и не могу сменить партнёров... Считается, что популяции могли бы погибнуть, если бы исчезли фикусы или их облигатные мутуалисты-опылители. Если бы это случилось, можно было бы ожидать целый каскад событий вымирания. Можно предположить, что облигатные паразиты мутуалистов вымрут, а обезьянам придётся коренным образом изменить свой рацион, или же они начнут голодать. Вместе с более мелкими животными, питающимися плодами и семенами фикусов, возможно, уменьшилось бы количество корма для таких хищников, как хищные птицы и ягуары. И это не тривиальный пример, а весьма реальная угроза из-за сведения тропических лесов.
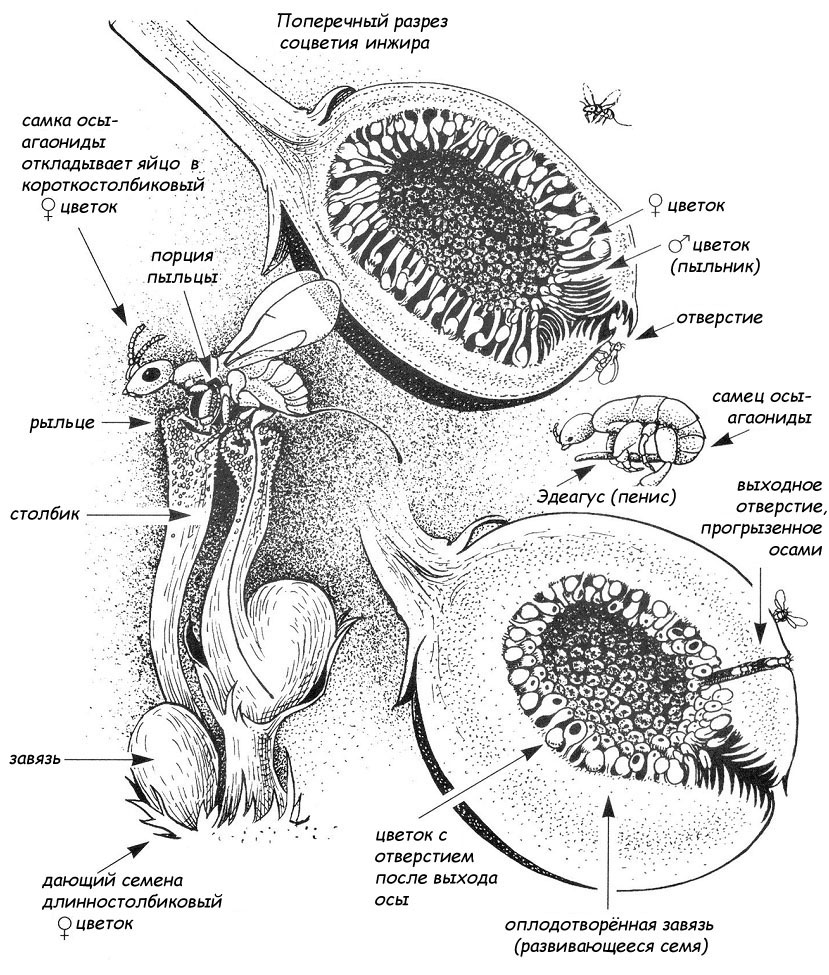
Один из лучших примеров взаимной коэволюции представляют
собой соцветие инжира (Ficus carica) и крохотные осы-бластофаги (Blastophaga
spp.), которые опыляют эти деревья, растущие главным образом во влажном
тропическом лесу. Здесь мы видим сложную анатомию фиги с сотнями её цветков,
осу, откладывающую яйцо внутри цветка, бескрылого самца осы и новое поколения
бластофаг, появляющихся на свет сквозь стенку зрелого плода. |
Пытаясь предсказать воздействие фрагментации тропического леса
на фикусы и их партнёров, Бронштейн и её коллеги задались вопросом: «Какое количество
деревьев фикуса необходимо для того, чтобы позволить сохраниться популяции осы?»
По их изначальным оценкам нужно было бы от 95 до 294 деревьев какого-то одного
конкретного вида фикуса, чтобы поддержать существование популяции опылителя
на протяжении минимум четырёх лет. Позднее Дойл МакКей, коллега Бронштейн из
Монпелье, подсчитал, что потребовалось бы минимум 300 взрослых деревьев фикуса,
чтобы гарантировать 99%-ную вероятность выживания ос-бластофаг. Затем МакКей
задался вопросом: «Какую площадь леса следует сохранять, чтобы гарантировать
поддержание существования минимальных жизнеспособных популяций фикусов и ос?»
Его ответ обескураживает. В зависимости от вида, могло бы потребоваться от 800
акров до 800 квадратных миль, чтобы гарантировать долгосрочное выживание фикусов,
осы и всех прочих зависящих от них птиц и млекопитающих.
Чаще риски, связанные с такой жёсткой зависимостью, оказываются слишком высокими,
чтобы способствовать столь строгому облигатному мутуализму. Вместо этого мы
обнаруживаем, что большинство мутуалистов может считаться продуктом распределённой
коэволюции – то есть, «гильдия» опылителей совместно специализируется на работе
с набором определённых растений, которые обладают схожими формами цветков и
способами подачи себя, но могут цвести либо последовательно, либо перекрываясь
в пространстве и времени. Если не заметно, что цветок либо опылитель участвует
в мутуалистической игре, то у них обоих найдётся другой выбор.
Конечно, не все опылители обязательно специализируются на определённом наборе
цветов со схожим обликом: некоторые, подобно медоносной пчеле, представляют
собой неспециализированных «мастеров на все руки». Чтобы увидеть, насколько
широкими могут быть пристрастия опылителя, давайте вернёмся из тропического
леса обратно в пустыню Сонора, где мы живём и работаем, и рассмотрим исследование
реакции медоносных пчёл на разнообразные вознаграждения из цветочной пыльцы,
сделанное в прошлом десятилетии.
ВСПОМИНАЕТ СТИВ:
Ай! Я остановился, чтобы отцепить рукав моей рубашки и кусочек
живой плоти от ветки сенегалии Грегга, преодолевая скалистый склон бахады над
каньоном Пима. Здесь, в горах Санта-Каталина, не далее чем в 15 милях от Национального
парка Сагуаро и центра Тусона, я еженедельно в течение последних десяти лет
собирал образцы в пяти домашних колониях европейских медоносных пчёл, чтобы
проверить богатый урожай пыльцы, собранный местными пчёлами. Хотя дело было
прохладным утром в середине ноября, медоносные пчёлы летали в окрестностях,
словно большие звенья миниатюрных самолётов. Отыскав ульи, я услышал гудение
машущих крыльев тысяч рабочих пчёл, находящихся в воздухе и активных в этих
местах, как минимум, 11 месяцев в году. Всё говорило о том, что я наверняка
найду новую пыльцу, если соберу образцы из этих ульев.
В отличие от сбора мёда, сбор образцов пыльцы из ульев легко сделать – это не
беспокоит пчёл, находящихся внутри, как бывает, когда их дом разламывают на
куски, чтобы стащить 50 фунтов мёда. Не надевая костюма пчеловода или защитной
сетки, я просто медленно и целенаправленно прошёл мимо струящихся потоков пчёл
и присел за ульями. Там я одновременно вытянул выдвижные ящички из дерева и
плотной ткани, называемые пыльцевыми ловушками. В каждой из них находились низкие
округлые холмики из кучек шариков пыльцы из пыльцевых корзиночек пчёл, окрашенные
во все цвета радуги. Пчёлы действительно работали на совесть.
Там лежали тысячи этих комочков – все примерно одинакового размера, но пестреющие
разными оттенками жёлтого, оранжевого, зелёного, коричневого и белого цвета
– в каждом из них преобладал определённый вид пыльцы диких цветов, произрастающих
в окрестностях. Это было любопытно, поскольку во время своей экскурсии в каньон
я видел мало цветов. Однако пчёлы способны увидеть дикие цветы, которые ускользают
от нашего взгляда – и, кроме того, они могли недавно собирать корм на площади
в 60 квадратных миль. Как однажды выразился Э. О. Уилсон, если бы пчёлы были
размером с людей, а расстояние их вылета увеличилось бы в той же пропорции,
то единственная колония пчёл, размещённая ровно в центре Техаса, собирала бы
пыльцу и нектар с диких цветов на половине этого штата. Эти пчёлы обладали глубокими
знаниями того, что находилось в цветке в любой час дня или время года на удивительно
большой площади пустыни Сонора близ Тусона.
Цвет каждого шарика указывал на то, что каждая отдельная пчела, собирающая корм,
обнаружила и успешно доставила в своё гнездо два маленьких груза пыльцы. Каждый
из этих шариков содержал до нескольких десятков тысяч пыльцевых зёрен, склеенных
вместе слюной пчелы или отрыгнутым мёдом. И за целый год в этих ловушках я в
итоге собрал комочки пыльцы, принадлежащей 55 видам цветковых растений из 40
родов и 25 семейств – около четверти всех видов растений, цветущих в пределах
их досягаемости.
Короче говоря, медоносные пчёлы обладают самым широким спектром пыльцевой диеты
– полилектией – среди всех известных опылителей. Известно, что они также собирают
мельчайшие частицы самых неожиданных материалов, отличающихся от пыльцы, в том
числе споры плесени, сырных клещей, муку, угольную пыль и опилки. Они даже собирают
пыльцу с обычно ветроопыляемых и лишённых нектара цветков вроде злаков и амброзии.
Фактически, до 30 процентов пыльцевого рациона в пустыне Сонора может поступать
от аборигенных и завезённых растений, которые считаются анемофильными – по сути,
ориентированные на опыление ветром через строение их цветка, летучесть пыльцы
и общее строение растения.
Используя пыльцевые ловушки в каньоне Пима, я определил не только количество
пыльцы, ежегодно собираемой медоносными пчёлами – часто от 50 до 85 фунтов на
колонию в год – но также и то, какие типы цветков они посещали. В прошлом десятилетии
маргаритки, мескитовое дерево, креозотовый куст, лилии, хохоба, амброзия дельтовидная,
омела и горчица давали до трёх четвертей пыльцевого рациона медоносных пчёл
в предгорьях Санта-Каталины, но эта пропорция вскоре может измениться. Ниже
по склону бахады, на котором я стоял, было видно землеройное оборудование, готовое
содрать аборигенную растительность, живущую на изучаемом мною участке.
Это был знак предстоящих перемен массового характера в окрестностях этого места
и во многих других местах. Кактусовый лес вскоре должны были заменить 500 новых
роскошных домов, и пейзаж вокруг них украсят лишь немногие эффектно выглядящие
аборигены вроде кактусов сагуаро и опунций, а вдобавок к ним – множество разнообразных
новичков. Когда мигранты с востока, из Калифорнии или Мексики приезжают в Тусон,
они привозят с собой множество экзотических растений для украшения ландшафта
– австралийский эвкалипт, африканский сумах, оливковое и перечное деревья. Хотя
медоносные пчёлы, несомненно, приспособятся к ним, юкковая моль и одиночные
пчёлы, коренные жители местообитаний вроде этого, вряд ли заживут лучше. И что
касается личной жизни местных аборигенных растений: какое воздействие окажут
на них очередная фрагментация среды обитания и длительное опыление экзотическими
для них пчёлами? Масштабные изменения в местной растительности могут означать
истощение их пищевых ресурсов и могут неизбежно привести к их локальному вымиранию
– временной или постоянной утрате популяции в данном месте. Локальное вымирание
одной популяции не ведёт к неизбежному вымиранию вида. Но в этом случае встаёт
один большой вопрос: сколько популяций может исчезнуть до того, как целый вид
будет признан находящимся под угрозой исчезновения? Как мы увидим далее, единого
ответа на этот вопрос нет, но есть очень чётко выраженные и тревожные тенденции.
В горном облачном лесу близ Сан-Вито в Коста-Рике
крошечный солнечный колибри (Phaethornis longuemareus) собирается посетить
необычное растение (Columnea florida) из семейства Gesneriaceae. В течение
года близ кончиков его листьев ненадолго появляются парные красноватые
пятна – визуальные указатели, которые птицы учатся ассоциировать со сладким
нектаром, скрытым в трубчатых цветках на стеблях под ними. |

ГЛАВА 4
ВСПОМИНАЕТ СТИВ:
Я оглядывал предгорья над долиной Напа, пока мы бродили по
горным хребтам – мы шли группой, растянувшись хвостом за моим профессором энтомологии
Роббином Торпом из университетского городка Дэвис Калифорнийского университета.
По сравнению с нижней частью склонов и дном долины, где пышные виноградники
заглушали ещё остающиеся островки естественной растительности, растительность
на возвышенностях была очень скудной. Высоко на горных хребтах, где на поверхность
выходила поблёскивающая зелёная серпентиновая скала, карликовый чапараль был
ещё более разреженным из-за того, что на серпентиновых почвах на него действовало
адское варево странных химикатов – некоторые из них ядовиты для растений.
Хотя многие из моих друзей-студентов отпускали комментарии по поводу того, насколько
бедным был растительный покров, каждый куст или дерево, растущие в отдалении
от остальных, можно было разглядеть целиком по отдельности и почувствовать их
запах: сосна серая, кипарис Сарджента, местная форма толокнянки. Это преимущество
не осталось незамеченным для меня. Я занимался изучением флоры и фауны Внутреннего
Берегового хребта, протянувшегося вдоль края континента. Я попросил Роббина
показывать мне какие-либо растения, уникальные для этой области, какие он заметит.
Примерно в этот момент сильный и необычный аромат заставил меня замереть на
тропе. Хотя мы находились на несколько миль выше любого из виноградников, я
распознал запах как практически идентичный букету вина «Каберне-Совиньон», разлитого
в долине Напа, которое я недавно с удовольствием попробовал. Оглядевшись, я
понял, что аромат мог исходить лишь из одного места – от тёмно-каштановых цветков
широколистного кустарника, растущего вдоль тропы. Это был не просто какой-то
кустарник, а каликантус западный, известный ботаникам как Calycanthus occidentalis.
Даже сейчас, наслаждаясь стаканчиком богатого танином вина, я вспоминаю ту вспышку
обонятельного распознания, которая пронеслась у меня в голове в тот момент,
когда я ощутил запах каликантуса: «Жуки. Он должен опыляться изнывающими от
страсти жуками-блестянками!» Не видя опылителей, я предсказал – просто по подсказкам
окраски и аромата цветков – как минимум, одну из групп животных, которые могли
бы переносить пыльцу с растения на растение у каликантуса западного в этих предгорьях
Напа.
Таблица I: Синдромы опыления
| Синдром |
Опылитель | Период цветения (a) | Окраска | Запах | Форма цветка (b) | Глубина цветка | Указатели нектара |
Вознаграждение |
| Кантарофилия | Жуки | Днём и ночью |
Разнообразная, обычно неяркая |
Сильный, фруктовый или аминный |
Актиноморфный | От плоского до чашевидного | Нет | Пыльца или пищевые тельца |
| Сапромиофилия | Падальные и навозные мухи | Днём и ночью |
Лилово-коричневая или зеленоватая |
Сильный, часто как разлагающийся белок | Обычно актиноморфный | Нет, или глубокий, если имеются ловушки | Нет | Нет |
| Миофилия | Журчалки и мухи-жужжала | Днём и ночью | Разнообразная | Разнообразный | Актиноморфный или зигоморфный | Нет, или до умеренной | Нет | Нет, или пыльца и нектар |
| Мелиттофилия | Пчёлы | Днём и ночью или только днём | Разнообразная, но не ярко-красная | Присутствует, обычно сладкий |
Актиноморфный, цветок горизонтальный или висячий | Нет, или до умеренной | Присутствуют | Нет (41,6%) и пыльца; открыто расположенный или скрытый |
| Сфингофилия | Бражники | Ночью или в сумерках | Белая или от бледной до зелёной | Сильный, обычно сладкий | Обычно актиноморфный, цветок прямостоячий | Глубокая узкая трубка или шпорец | Нет | Обильный нектар (22,1%); скрытый |
| Фаленофилия | Мелкие ночные бабочки | Ночью или в сумерках |
Белая или от бледной до зелёной | Умеренной интенсивности, сладкий | Актиноморфный или зигоморфный | Умеренно длинная трубка | Нет | Нектар; скрытый |
| Психофилия | Дневные бабочки |
Днём и ночью или только днём | Ярко-красная, жёлтая или синяя |
Умеренной интенсивности, сладкий | Актиноморфный или зигоморфный | Глубокая узкая трубка или шпорец | Присутствуют | Нектар (22,8%); скрытый |
| Орнитофилия | Птицы | Днём | Ярко-красная | Отсутствует | Актиноморфный или зигоморфный | Глубокая широкая трубка или шпорец |
Нет | Обильный нектар (25,4%); скрытый |
| Хироптерофилия | Летучие мыши | Ночью | Грязно-белая или зелёная | Сильный, ферментированный | Щёткообразный или чашевидный | Нет | Обильный нектар (18,9%) и обильная пыльца; открытый |
Источник: С изменениями из Wyatt (1983). Концентрации нектара
даны по Pyke and Waser (1981).
a. «Период цветения» означает время раскрытия цветков
b. «Актиноморфный» означает радиально симметричный; «зигоморфный» означает двусторонне-симметричный.
Эта игра в вопросы и ответы интриговала и заставляла страдать
от головной боли биологов на протяжении большей части этого века: насколько
возможно логическим путём выявить животных-опылителей какого-либо вида растения,
просто заранее зная ключевые характеристики цветка и животного? Взаимодействующий
набор признаков растения и животного, который имеет последовательный характер,
известен как синдром опыления. (Таблица I). Как мы увидим далее, эта рутина
распознавания характера взаимодействия, как и любая игра в вопросы-ответы, можем
быть очень интересной, но в ней зачастую недооценивается сложность и изменчивость
отношений, сложившихся в природе. Некоторые биологи наших дней откладывают в
сторону эту игру по выявлению «классических отношений» и берутся за другую,
более сложную. Из-за того, что эта новая игра требует больше эмпирических наблюдений
над сбором корма опылителем и над характером цветения всех растений того или
иного ландшафта, она лучше отражает разнообразие типов взаимодействия растения
и опылителя.
Эта новая парадигма, недавно предложенная экологом Джудит Бронштейн, позволяет
нам выделить ландшафты растения и опылителя. Эти образцы ландшафта рассказывают
нам о взаимодействии жука не только с каликантусом, но и обо всех других цветах,
которые жуки последовательно посещают, а также обо всех других животных, которые
посещают кусты каликантуса и его соседей. Как и основное правило в игре синдромов
опыления, это также предполагает, что существуют ключевые признаки цветковых
растений и опылителей, которые не изменяются независимо, а сгруппированы вместе
в пространстве и времени. Но, в противоположность другому предположению игры
синдромов опыления, мы в настоящее время признаём, что такие признаки изменчивы
как внутри вида, так и от вида к виду. Не все популяции каликантуса могут цвести
в одно и т же время или привлекать одних и тех же жуков. Аналогичным образом
одни жуки могут активно опылять каликантус, тогда как другие могут лишь изредка
посещать его, предпочитая обращать свою активность на другие растения по соседству.
Чтобы играть в какую-то игру, мы должны узнать, какие особенности цветков имеют
значение для опылителей, и какие особенности животных формировали морфологию
и фенологию цветка с течением эволюционного времени. Чтобы более чётко показать
некоторые из этих особенностей, давайте вернёмся к каликантусу.
Внимательнее разглядывая цветы каликантуса каштановой окраски, мы начинаем замечать
подсказки, указывающие на то, что это действительно могли быть цветки для жуков.
Цветки длиной полтора дюйма образованы мясистыми, почти суккулентными тепалиями
– это нечто среднее между лепестками венчика и чашелистиками чашечки – которые
защищают вознаграждение, находящееся внутри них. Они не производят никакого
нектара, но наполнены ярко-жёлтыми, довольно маслянистыми пыльцевыми зёрнами.
А на кончике каждой из красноватых долей околоцветника, находящихся в самой
глубине цветка, мы встречаем шероховатые белые «пищевые тельца», содержимое
которых известно как трубчатый белок. Такие пищевые тельца встречаются внутри
многих примитивных цветов и, возможно, поддерживают пищевой интерес жуков, поэтому
они меньше повреждают нежные части растения, когда ищут богатую белком пыльцу
себе на обед. Держась внутри защищающего их цветка, вне досягаемости для большинства
хищников, жуки также часто спариваются с другими особями своего вида.
Эти особенности цветка напоминают о примитивных цветковых растениях Новой Гвинеи
и Новой Каледонии, которые изучались как «живые ископаемые» с целью реконструкции
условий, при которых в процессе эволюции впервые возникло опыление животными.
Такие цветы обладают либо сладким и пряным, либо сильным и неприятным ароматом
– пахнут, словно гниющий тропический фрукт, или, что ещё хуже, как пара заношенных
грязных и потных носков, привезённых домой из летнего лагеря. В провинции Гуанакасте
в Коста-Рике есть один вид дерева, Sapranthus palanga, которому студенты дали
прозвище «дерево тысячи грязных носков». В обоих случаях эти цветки с широко
раскрытым венчиком предлагают множество аттрактантов (вроде их ароматов) и видов
вознаграждения (маслянистая жёлтая пыльца и беловатые пищевые тельца), которые
могут чувствовать и переносить как жуки, так и мухи. Когда жуки кормятся пыльцой
каликантуса, часть её прилипает к их твёрдой кутикуле. Позже, когда жуки перемещаются
для кормления на другое растение, она пассивно переносится на органы размножения
(в том числе на восприимчивые к пыльце рыльца) других цветков.
На холмах долины Напа в Калифорнии произрастает
каликантус западный (Calycanthus occidentalis). Крупные цветки обладают
многими примитивными особенностями, в том числе мясистыми тепалиями и
белыми пищевыми тельцами. Цветочный аромат напоминает смесь перезрелых
фруктов и вина «Каберне Савиньон». Этими цветами привлекается в качестве
опылителей множество жуков-блестянок и стафилинид. |

Однако жуки не всегда оказываются самыми успешными опылителями.
Фактически же они безмятежно кормятся на разных цветах, многие из которых никогда
не опыляют. У некоторых видов вроде каликантуса они переносят пыльцу, стараясь
отыскать побольше белых пищевых телец и партнёров по размножению. Но чаще они
сбрасывают больше пыльцы, чем доставляют на другой цветок, пока бродят между
растениями. Кроме того, если неподалёку окажется другой пряно пахнущий привлекательный
вид, они могут сразу же броситься к нему для кормления и вообще не доставят
пыльцу на совместимый цветок первого вида.
Этот способ случайного переноса пыльцы жуками специалисты в области биологии
цветения назвали «беспорядочным»* опылением, потому что эти насекомые склонны
поедать и загрязнять экскрементами такие цветы во время путешествия по ним.
Редко когда один вид цветов зависит исключительно от одного вида жуков (если
так вообще бывает). Бродящих повсюду жуков, в свою очередь, едва ли когда-либо
привлекал аромат исключительно одного вида из их окружения. Мутуализм в данном
случае свободный, рассеянный, если, конечно, эти отношения можно назвать мутуализмом.
И всё же среди экологов, опытных в игре синдромов опыления, стало общепринятым
относиться к цветкам каликантуса как к «цветкам жуков». Вытянутые цветки фукьерии
известны как «цветки колибри», а белые трубы цветущих по ночам цереусов – «цветки
бражников», хотя учёные зафиксировали и других животных, опыляющих или, как
минимум, посещающих эти цветы и кормящихся на них.
|
* В литературе на английском языке такой способ опыления носит название mess-and-soil pollination – буквально «грязно-земляное опыление», что больше соотносится с описанием автора. – прим. перев. |
Это соотнесение типов цветка с «гильдиями» сходных животных,
кормящихся на цветках, похоже, стало повсеместным способом времяпрепровождение
для людей. Словно участники игры синдромов опыления, представители многих аборигенных
культур предполагают, что близкородственные друг другу животные посещают цветы
определённой формы, чтобы питаться их похожим на мёд нектаром. Экологи просто
отсеяли из этого фольклора всё лишнее, выявив множество цветочных ресурсов,
которые привлекают животных, а также множество приспособлений животных к форме
цветов. Например, дневные и ночные бабочки пользуются скручивающимся в спираль
хоботком, похожим на соломинку, чтобы сосать нектар, находящийся на дне глубоких
цветочных трубок.
Любопытным натуралистам не потребовалось слишком много времени, чтобы начать
ассоциировать кормовое поведение чешуекрылых с теми или иными формами, окрасками
и размерами цветков. Во многих языках существуют названия для растений, которые
означают, по сути, «бабочкина трава» – как в случае с ластовнем (Asclepias tuberosa)
в умеренных областях Северной Америки. Монарх и данаида гилиппа – одни из многих
видов, которые часто посещают этот многолетник с оранжевыми цветками на всём
пространстве его ареала. В испанском лексиконе народов Латинской Америки есть
несколько замечательных ассоциаций между цветком и опылителем, в которых оба
они известны под одним и тем же народным названием. Например, название chuparosa
(«сосальщик роз») относится не только к колибри, но и к красному трубчатому
цветку такого типа, какой есть у Justicia californica. Это правда, что колибри
в обеих Америках активно разыскивают трубчатые цветки, которые зачастую обладают
красной или оранжевой расцветкой и богаты нектаром. Колибри понимают, что красный
цвет сигнализирует о возможном присутствии нектара неподалёку. В действительности
этот цветовой стимул для колибри настолько силён, что, если накрасить губы помадой,
набрать в приоткрытый рот вина и встать там же, где установлены кормушки красного
цвета с сахаром или на улицу выставлены пурпурно-красные цветы в горшках, колибри
будут часто кормиться прямо у вас изо рта.
Исходное предположение о том, что некоторые «гильдии» животных обладали сильной
привязанностью к конкретным классам цветков, впервые были формально выведено
натуралистом из Mилана Фредерико Дельпино в серии статей, изданных в период
между 1868 и 1875 гг. Пауль Кнут, ещё один гигант в области биологии цветения,
часто считается автором классификации синдромов опыления, созданной около 1910
г. Эти синдромы позже были выверены и дополнены великим немецким специалистом
по биологии опыления Стефаном Фогелем в новаторском очерке, опубликованном в
1954 году. В этом очерке Фогель предположил, что, выявив некоторые характерные
особенности, которые он назвал “Stil” («стиль»), специалисты по биологии цветения
могли бы предсказать, кто из посетителей цветков может являться истинным опылителем.
Неважно, идёт ли исследование в поле или по гербарию: Фогель утверждал, что
кто угодно смог бы увидеть, что формы цветов указывают на их гармоничные отношения
с определёнными животными. А его очерк иллюстрировал эти синдромы опыления такими
красочными гравюрами, что каждый из видов отношений быстро превратился в прописную
истину в биологической литературе. К его чести, Фогель предупреждал своих читателей
о том, что такие синдромы должны рассматриваться как рабочие гипотезы, которые
следует проверять в полевых условиях. Однако другие учёные приняли цветные гравюры,
иллюстрирующие классификацию Фогеля, за истину в последней инстанции. На два
десятилетия, прошедшие после его публикации в 1954 году, биологи превратили
синдромы опыления в стереотипы, как будто отношения тет-а-тет между растениями
и их опылителями были нормой в мире природы.
Влияние Фогеля на исправление этих упрощений было несколько ослаблено, поскольку
его публикации появились только в Германии. Другие передовые учёные вроде Христиана
Шпренгеля, Пауля Кнута и Германа Мюллера помогли превратить использование синдромов
в обучающий инструмент в современных исследованиях в области биологии цветения,
но в более популярных работах их предостережения не были учтены. К тому моменту,
когда в 1979 г. был издан классический учебник Кнута Фегри и Леендерта ван дер
Пейла «Основы экологии опыления», синдром опыления надёжно закрепился в качестве
преобладающей парадигмы для изучения взаимодействия между цветами и их опылителями.
Хотя ранние исследователи подразумевали, что это лишь концепция, которую нужно
использовать экспериментально, гибко, в современной ботанической литературе
она так или иначе превратилась в догму. Слишком многие биологи-путешественники
рассматривали классификационный подход, связанный с синдромами опыления, как
указатель, дающий ответы на все вопросы, и забывали о поиске других признаков
и того, на что они указывали. Однако мы полагаем, что эта методология всё равно
может использоваться в качестве неоценимого эвристического инструмента студентами,
натуралистами и биологами, которые не являются специалистами в области биологии
опыления per se.
Некоторые формы, ароматы, питательные награды и время раскрытия цветков оказались
прочно связанными с эволюционным влиянием единственного вида жука, пчелы или
летучей мыши. Словарь названий для этих «цветов, любящих животных», модный в
настоящее время среди некоторых биологов – кантарофилия, мелиттофилия, хироптерофилия
– это побочный продукт более ранних систем классификации. Герберт Бейкер с коллегами
разработал эту классификацию, чтобы принимать во внимание не только морфологию
цветка, но также и химический состав нектара и пыльцы.
В настоящее время мы знаем, что многие из этих взаимоотношений представляют
собой вовсе не простые истории близких отношений тет-а-тет. Эколог Калифорнийского
университета Николас Вейзер впервые столкнулся с трудностями при использовании
такого рода проецирования, когда пытался объяснить время цветения и завязывания
семян «цветками колибри» у фукьерии временем миграции колибри по засушливым
районам юго-запада США. Вейзер и другие были удивлены, обнаружив, что во многих
зарослях фукьерии пчёлы-плотники воровали нектар или опыляли красные трубчатые
цветки чаще, чем птицы. Позже летом, пока пустынные пчёлы опыляют «цветки колибри»,
те же самые колибри, которые движутся через пустыню, опоздав к цветению фукьерии
на две недели, могут опылять 13 различных видов растений с горных лугов, представляющих
восемь различных ботанических семейств и несколько синдромов.
Исследования Лиз Слоусон из Пустынного ботанического сада в Финиксе, штат Аризона,
в области биологии цветения и опыления агавы Пальмера (Agave palmeri) также
показали сложную историю адаптации цветков и «гильдий» первичных и вторичных
опылителей. Эти массивные агавы используют репродуктивную стратегию «большого
взрыва», то есть, цветут лишь один раз после того, как в течение нескольких
десятилетий накопят запасы питательных веществ; затем каждое отдельное растение
погибает, а его гены передаются следующему поколению через семена, высыпающиеся
из плодов. Их цветоносы выглядят, словно декоративные канделябры на фоне пламенеющих
летних небес Аризоны во время заката.
Хотя устройство цветоноса явно видоизменено (открытые и тесные группы цветков
находятся на коротких горизонтальных ответвлениях), когда ночная тьма сменяется
рассветной прохладой, на сцене появляются другие нарушители. В эти ранние утренние
часы можно увидеть и услышать тучи голодных шмелей, медоносных пчёл и пчёл-плотников,
а также бумажных ос и гигантских оранжево-чёрных ос-помпилид. Они жадно хлебают
обильный нектар, а самки пчёл уносят в гнёзда пыльцу. Так происходит потому,
что даже на участке, плотно заселённом местными летучими мышами, цветочных богатств
с избытком хватает для всех едоков. Но, как уже говорилось ранее, на многих
участках нет мигрирующих летучих мышей, следующих по «нектарному коридору»,
либо их численность сильно снизилась из-за разрушения укрытий летучих мышей
внутри пещер или из-за отравления пестицидами. Лиз решила, что, если летучих
мышей действительно не хватает, пчёлы, прилетающие рано утром, получают большую
часть пыльцы и нектара агавы. К счастью для агав Аризоны, пчёлы весьма успешно
осуществляют опыление цветов агавы, даже если эволюция их цветка, вероятнее
всего, определялась их танцем в паре с летучими мышами.
В другом месте, в низинных тропических лесах, единичный вид пчёл-эуглоссин может
посещать до восьми различных видов орхидей, цветущих один за другим в своих
панамских местообитаниях. Вейзер и его коллега Мэри Прайс сделали вывод о том,
что «отсутствие узкой специализации, похоже, является правилом среди опылителей»,
добавляя, что среди различных растений, которые посещали колибри на определённом
участке, «большинство цветов не соответствует «синдрому опыления птицами»».
Почему же тогда некоторые экологи продолжают использовать синдромы опыления
как инструмент для обучения теме взаимоотношений растения и животного? Ответ
прост. Игра синдромов остаётся полезной в качестве отправной точки, когда о
посетителях цветков растения мало что известно. Она может не предложить нам
полного диапазона животных, среди которых следует искать потенциальных опылителей,
но поможет определить нескольких из них. Подход с использованием синдромов опыления
по-прежнему может с успехом применяться студентами и другими новичками в игре
опылителей.
В некоторых случаях мы можем лишь догадываться о том, какими могли бы быть исторические
отношения между растением и его основными опылителями, поскольку эти отношения
мутуализма были в значительной степени разрушены изменениями в современном пейзаже.
Такой дедуктивный подход обычно используют биологи, посещающие фрагменты местообитаний,
в которых сохраняются лишь неспециализированные опылители типа медоносных пчёл.
Морфология цветка и химия нектара, например, могут использоваться для того,
чтобы предсказать присутствие опылителя в недавнем прошлом – если этот опылитель
был особенно важным для определения характера эволюции признаков цветка растения.
И даже если растение могло вымереть ещё до того, как была соответствующим образом
исследована его экология опыления, животные, которые когда-то его посещали,
всё ещё могут демонстрировать признаки адаптации к виду его цветков. Кроме того,
некоторые выводы, сделанные на основе изучения современной фауны и образцов
недавнего исторического прошлого, могут быть использованы для того, чтобы подтвердить
то, каким образом происходило изначальное взаимодействие между растением и опылителем.
Известно несколько замечательные примеров детективной работы в области экологии,
результатом которых была идентификация опылителей иного рода, чем те, которых
наблюдают за посещениями этих растений в наши дни. В одном из случаев специалист
по экологии растений Пол Кокс скептически воспринял прочитанное сообщение из
вторых рук о том, что крысы опыляют туземную гавайскую лиану Freycinetia arborea,
известную как иеие. Она обладала весьма немногими признаками растения, опыляемого
нелетающими млекопитающими. Поэтому он решил провести четыре дня в укрытии в
лесах над Кеалакекуа, на Гавайях, где росло некоторое количество лиан. За всё
это время он не видел на растениях никаких крыс и выявил лишь одного регулярного
посетителя цветов иеие – птицу, известную как японская белоглазка. Но белоглазка
была впервые завезена на Гавайи из Японии в 1929 году, поэтому её недавний вклад
в опыление цветков иеие не мог объяснить некоторых особенностей соцветий растения.
Женские цветоносы иеие богаты сахарами-гексозами и некоторыми аминокислотами,
которые независимо эволюционировали у цветков многих растений, привлекательных
и для древесных птиц, и для летучих мышей. Кокс чувствовал, что история лианы
иеие должна была скрывать других птиц.
Просматривая тысячи страниц журналов гавайских натуралистов девятнадцатого века,
Кокс встретил записи, свидетельствующие о том, что острова были некогда населены
несколькими птицами, которые в настоящее время находятся под угрозой исчезновения
или вымерли: гавайская ворона, оуэ и похожая на клеста птица, последний раз
наблюдавшаяся в 1894 году и жившая когда-то в районе Кона Кост*. Кокс взял старые
музейные образцы этих птиц и начал изучать перья на их головах с использованием
сканирующего электронного микроскопа, разыскивая пыльцевые зёрна. К удивлению
своих коллег Кокс обнаружил значительные количества пыльцы характерного облика
на оперении головы экземпляров оуэ и «клеста» Кона, и некоторое количество такой
же пыльцы на перьях головы гавайской вороны. Эти пыльцевые зёрна выглядели идентичными
тем, которые он получил с 57-летнего экземпляра лианы иеие. Очевидно, эти три
птицы регулярно совали свои головы в прицветники мужских растений лианы иеие,
а также, несомненно, посещали женские цветоносы ради награды иного рода. Сомнения
Кокса имели под собой солидное основание: в цветках иеие не было ничего такого,
что предполагало бы приспособление к опылению крысами; фактически же то старое
утверждение основывалось на ошибке, которая усилилась за счёт многократного
обращения к ней за все эти годы. Вместо этого, предположил Кокс, цветонос иеие
эволюционировал для того, чтобы позволить доступ к цветкам самым разнообразным
птицам, варьирующим по размеру от завезённой белоглазки (птица длиной 4 дюйма)
до редкой гавайской вороны (почти 20 дюймов в длину). Оуэ, гавайская ворона
и «клёст» Кона – все эти птицы почти вымерли незадолго до конца девятнадцатого
века, поэтому иеие, должно быть, некоторое время страдала от недостаточного
опыления и плодоношения. Однако эти многолетние лианы смогли сохраниться, потому
что они могут энергично размножаться, ветвясь и повторно укореняясь. К концу
1930-х годов завезённые японские белоглазки, вероятно, начали брать на себя
опыление лианы. Интенсивность плодоношения, несомненно, вновь возросла.
* Вероятно, речь идёт о цветочнице Chloridops (Psittirostra) kona. – прим. перев. |
В другом широко освещённом случае с Гавайскими островами большое
количество «кардинальских цветов» семейства лобелиевых (Lobeliaceae) пострадало
от вымирания в прошлом веке, вынуждая ко-адаптировавшихся к ним опылителей перейти
к использованию других цветочных ресурсов. Из примерно 273 видов, подвидов и
разновидностей лобелиевых, описанных в историческое время с Гавайских островов,
лишь у 27 процентов есть достаточно значительные популяции, способные уберечь
их от скорого вымирания. Четверть видов вымерла в течение прошлого века; ещё
19 процентов видов, подвидов и разновидностей могут считаться редкими или находящимися
под угрозой исчезновения. Это катастрофическое снижение разнообразия в крупнейшем
семействе растений Гавайских островов ставит перед нами вопрос: что же случилось
с их коэволюционировавшими опылителями? В течение многих лет биологи полагали,
что множество гавайских птиц, известных как гавайские цветочницы, могли коэволюционировать
совместно с лобелиевыми, поскольку некоторые из них обладают длинными, изогнутыми
вниз клювами, которые соответствуют форме цветочных трубок некоторых «кардинальских
цветов».
Но когда биолог Колин Кори отправилась искать гавайских цветочниц, посещающих
цветки двух редких лобелиевых в области Коолау на острове Оаху, в течение 136
часов наблюдения она не смогла зафиксировать на этих растениях ни одной птицы.
Несколько бражников, мелкие чёрные пчёлы (местный род пчёлы Hylaeus, семейство
коллетид) и завозные медоносные пчёлы были единственными посетителями цветов,
которых она зарегистрировала на обоих видах лобелиевых. Она также обнаружила,
что эти два редких растения в настоящее время способны к самооплодотворению:
им может быть не нужна птица или любое другое животное для переноса пыльцы с
растения на растение, чтобы завязать семена. Кори пришла к выводу, что взаимозависимость
между гавайскими цветочницами и лобелиевыми не поддерживается современными наблюдениями
обоих организмов на Оаху.
Тем не менее, недавно команда зоологов пришла к выводу иного рода, утверждая,
что признаки ныне живущих организмов невозможно понять исключительно в рамках
их современного состояния. Когда Томас Смит, Леонард Фрид и их коллеги изучили
собранные в недавнем прошлом музейные образцы одной из гавайских цветочниц,
ииви, они обнаружили, что её клюв в девятнадцатом веке был длиннее, чем в наши
дни. Затем они обнаружили немного записей из недавнего прошлого – девятнадцатого
века. Эти записи документировали кормление цветочниц ииви на ныне редких или
вымерших представителях лобелиевых с длинными трубчатыми цветками, которые производили
обильный, богатый сахарами-гексозами нектар, но не имели никакого запаха – это
особенности цветков, которые соответствуют скорее птицам, чем насекомым, которых
зарегистрировала Кори. Длинный клюв хорошо служил бы цветочнице ииви, когда
эти цветы встречались значительно чаще.
Большинство особей ииви наблюдалось кормящимися на открытых цветках дерева охайя
лехуа (Metrosideros), которые лишены трубчатых венчиков, определённо приспособленных
к птицам. Смит, Фрид и их коллеги выдвинули гипотезу о том, что, когда в девятнадцатом
веке численность лобелиевых снизилась, некогда обычные цветочницы ииви начали
переходить на другие цветочные ресурсы ради выживания. Цветочницам не требуются
никакие специальные адаптации клюва, чтобы добывать нектар из цветков охайя.
Зоологи предполагают, что изменение в рационе с трубчатых цветков на открытые
привело к возникновению движущего отбора в сторону появления более коротких
клювов. Иными словами, особи ииви с самыми длинными, сильнее изогнутыми вниз
клювами пропали из размножающихся популяций в течение последней сотни поколений.
Надклювья у ииви нашего времени на 2-3 процента короче, чем у ииви, поступивших
в коллекции до 1902 г., когда лобелиевые были ещё довольно обычны в лесах.
Почти вымершая цветочница ииви (Vestaria coccinea)
с Гавайского архипелага является единственным известным опылителем нескольких
местных цветов вроде этого вида Clermontia. Её сильно изогнутый клюв позволяет
птице успешно добывать нектар из такого рода изогнутых цветков. |

Такие примеры указывают на то, что действительно существуют
долгосрочные сделки, заключённые между совокупностью опылителей и определенным
растением, но эти сделки редко бывают настолько эксклюзивными или жёсткими,
что другие организмы постоянно остаются «за рамками», будь эти «другие» японскими
белоглазками или гавайскими деревьями охайя. К сожалению, многие экологические
исследования недавнего прошлого демонстрируют тенденцию к исключению этих других
организмов из той картины, которую они создают в соответствии с тем, как определены
их цели. Чтобы преодолеть эту тенденцию к рассмотрению только пары из растения
и животного – а не взаимодействия цветочных ресурсов и опылителей в местообитании
в целом – Джудит Бронштейн убедила нас отступить на шаг назад и взглянуть издалека
на весь пейзаж экологических взаимодействий, вращающихся вокруг доступности
нектара и пыльцы.
Правда, Бронштейн, как и многие другие до неё, признаёт, что «мы удивительно
мало знаем о том, как опылители отвечают на пространственные и временные изменения
своих цветочных ресурсов». Мы знаем, что опылители в процессе кормления обычно
перемещаются между участками цветочных ресурсов. Однако мы редко знаем, насколько
далеко они путешествуют каждый отдельно взятый день, чтобы свести концы с концами,
не говоря уже о тех расстояниях, которые они преодолевают на протяжении всей
своей жизни, путешествуя за кормом. Ботаники на острове Барро Колорадо, в Ла-Сельва
и Монтеверде в Коста-Рике – это одни из тех немногих, кто точно зафиксировал,
сколько видов растений конкурирует за одного и того же опылителя в данном ландшафте
– или, напротив, как растения подверглись действию отбора, чтобы цвести в разное
время и с разной интенсивностью, чтобы минимизировать конкуренцию друг с другом.
Самый полный обзор, который нам известен – это 50-месячное исследование насекомых,
которые обслуживают 133 вида растений, обитающие на 80 акрах греческой фриганы
– пострадавшей от перевыпаса и пожаров средиземноморской экосистемы трав и кустарников
близ Афин. В этом нарушенном сообществе, которое мало кто из экологов счёл бы
особенно богатым, Петаниду и Эллис документировали самое высокое разнообразие
опылителей из когда-либо зарегистрированных: ошеломляющие 666 видов насекомых,
в том числе более 225 видов одиночных пчёл. Как ни странно, климат на этом участке
настолько переменчив, что лишь 20 процентов от всей фауны опылителей обнаруживались
на протяжении всех пяти лет исследований. В этом месте не было зафиксировано
никаких летучих мышей или птиц. Но на каждый цветок во фригане приходится в
среднем по пять видов насекомых-посетителей, будь то осы, пчёлы, дневные или
ночные бабочки, жуки или мухи.
Постепенно Бронштейн и другие учёные занялись интеграцией различных исследований
вроде работы по греческой фригане, чтобы дать нам расширенное представление
о ландшафте растения и опылителя. Хотя дополнительные исследования, в настоящий
момент находящиеся в процессе работы, ещё добавят красок и глубины резкости
в эти картины, здесь уже можно предварительно обрисовать некоторые из данных
закономерностей. В первом типе ландшафта господствуют неспециализированные опылители,
связанные с цветами, которые цветут последовательно один за другим или не перекрываясь
по времени друг с другом. Этот вид ландшафта очень обычен в тропиках, но может
также встречаться в местообитаниях с резко выраженными сезонными изменениями,
вплоть до арктической тундры. В тундре и во влажных тропиках периоды цветения
множества растений в одном местообитании дополняют друг друга, и они не обязательно
конкурируют за одних и тех же опылителей. Кроме того, одно растение на протяжении
долгого сезона цветения могут последовательно посещать разные опылители. Например,
лаванда в южной Испании посещается 70 различными видами пчёл, дневных и ночных
бабочек – у каждого вида свой пиковый период активности на протяжении трёхмесячного
сезона цветения.
На полевой станции в Скалистых горах в Колорадо Лесли Риэл и Ник Вейзер обнаружили
свидетельство наличия последовательных случаев мутуализма: цветущие дельфиниумы
снабжали колибри пищей сразу по их прилёту на горные луга. В дальнейшем колибри
опыляли поздно цветущие трубчатые цветки растений рода Ipomopsis. Где бы эти
два цветка ни росли вместе, они посещались одним и тем же набором видов колибри,
но демонстрировали лишь небольшое перекрывание по времени цветения. Однако на
заливных лугах западного Колорадо ниже полевой станции в Скалистых горах орхидея
Spiranthes diluvialis демонстрирует опасности последовательного мутуализма.
Седония Сайпс и Винс Тепедино показали, что из-за того, что шмели полагаются
на эту редкую орхидею только как на источник нектара, для обеспечения их пыльцой
на протяжении всего сезона им должен быть доступен ряд последовательно цветущих
растений. При отсутствии этих кормовых ресурсов в данной местности для перекрёстного
опыления этой небольшой орхидеи останется слишком мало шмелей. Следовательно,
присутствие других видов, обеспечивающих дополнительные источники пыльцы для
шмелей, может быть столь же критичным, как и сами опылители для орхидеи.
Другой пример этого первого вида ландшафта был недавно описан на участках леса
в Японии, где шмели на протяжении всего сезона последовательно посещают множество
облигатно перекрёстноопыляемых цветов. Самая ранняя трава зацветала в апреле,
но плохо давала семена, особенно там, где фрагменты леса находились в обжитых
районах. Похоже, что шмелиные колонии этого сезона ещё не стали достаточно большими,
чтобы обслуживать области, где цветов было недостаточно для их привлечения.
Когда наступал июнь и зацветала следующая трава, на островках леса среди сельскохозяйственных
и жилых местностей было вполне достаточно вознаграждения для шмелей; вторая
трава лучше обсеменялась в обеих местностях. Третья многолетняя трава цвела
в августе, и она также не была ограничена в опылителях.
Во втором типе ландшафта растения и опылителя господствуют неспециализированные
опылители растений, которые цветут все одновременно. Такая ситуация часто наблюдается
в пустынях и субтропических местообитаниях, где растения дружно реагируют на
начало короткого сезона дождей, но может быть обычной также в умеренных и высокогорных
областях. Растения в этих ландшафтах явно конкурируют за опылителей. И каждое
из них может пострадать от низкого репродуктивного успеха, потому что там пчёлы
или бабочки могут двигаться без разбора от одного вида к другому, по пути растрачивая
пыльцу впустую. Некоторые тропические травы вроде Calathea occidentalis устраивают
необычайно роскошные цветочные представления, конкурируя за самых успешных опылителей,
которыми в данном случае являются пчёлы, редко появляющиеся в изобилии. Бабочки,
более обычные посетители цветков, гораздо менее надёжны в плане переноса пыльцы
от одной калатеи к другой, хотя они извлекают очевидную выгоду из обилия нектара,
связанного с этими многовидовыми представлениями. Весенние дикие цветы на сицилийском
побережье демонстрируют схожую тенденцию бурного одновременного цветения, которое
привлекает самых разнообразных насекомых-опылителей.
Третий тип ландшафта отличается господством специализированных опылителей, которые
посещают растения, обладающие продолжительным периодом цветения в среде обитания,
где не выражена сезонность климата. Недолго живущие осы, например, должны искать
фикусы нужного вида, последовательно цветущие в их ограниченном местообитании
на протяжении всего года, иначе они вымрут. К счастью для ос, очень мало фикусов
из одной популяции цветёт одновременно.
В четвёртом ландшафте растения и опылителя неспециализированные мигрирующие
опылители вроде нектароядных летучих мышей последовательно переходят на разнообразные
растения, снабжающие их нектаром по мере их миграции из тропических в аридные
местообитания умеренного климата. Даже там, где есть только один источник нектара,
доступный определённому виду летучих мышей, он обычно цветёт в одной последовательности
с другими растениями, опыляемыми летучими мышами, растущими на севере и на юге
вдоль пути миграции летучей мыши. Таким образом летучая мышь может специализироваться
только на одном виде цветов в каждом отдельно взятом местообитании, но они связаны
в «нектарный коридор» последовательных периодов цветения вдоль миграционного
маршрута летучей мыши.
Эта концепция «нектарного коридора» для мигрирующих видов, в общих чертах определённая
в середине 1970-х гг. Донной Хауэлл из журнала «Исследования Национального Географического
общества», недавно была окончательно сформулирована и разработана Тэдом Флемингом.
Его работа уделяет основное внимание географической последовательности растений,
используемых летучими мышами малыми листоносами – одним из нескольких опылителей
гигантских колонновидных кактусов, агав и манфред. Эти летучие мыши склонны
использовать нектар агав поздней осенью и ранней весной, и нектар ипомеи древовидной
зимой. Летом, перелетая каждый месяц на большие расстояния, они явно разыскивают
обильно цветущие заросли различных агав и колонновидных кактусов, собирая корм
на расстоянии целых 60 миль от своих дневных укрытий. Миграция колибри из северной
Мексики на юго-запад США в течение месяца каждую весну явно совпадает со сходной
последовательностью цветения с юга на север различных видов и популяций фукьерий.
Мы сопровождали Тэда в некоторых из его ночных визитов в земли кактусов пахицереуса
и сагуаро рядом с морем Кортеса близ Байя Кино в поисках вылизывающих нектар
мигрирующих летучих мышей. Хотя летучие мыши – это не исключительные опылители
ни для гигантских колонновидных кактусов, ни для растущих рядом с ними агав
– вспомните, что Лиз Слоусон показала, что они могут вполне приемлемо опыляться
пчёлами и осами по утрам, когда раскроются цветки – фактически на некоторых
территориях растения могут сталкиваться с нехваткой пыльцы, особенно в те годы,
когда летучие мыши редки и нечасто посещают цветы.
И последний из типов ландшафта, описанных Дж. Бронштейн, довольно редок: здесь
господствуют специализированные опылители, каждый из которых связан с ограниченным
числом видов растений, цветущих одновременно. Этот ландшафт можно обнаружить
в пустынях, сезонных субтропических кустарниковых зарослях или в высокогорной
тундре. Там мы обнаруживаем господство олиголектичных одиночных пчёл, устраивающих
гнездо в земле и связанных с единственным широко распространённым доминирующим
растением типа креозотового куста или мескитового дерева, или ночных бабочек,
которые специализируются только на одной местной юкке. Такие специализированные
насекомые сталкиваются с серьёзными проблемами, если появляются до или после
цветения своего единственного типа цветочных ресурсов – например, если запоздалый
снег убьёт самые ранние цветы или даже задержит весь сезон цветения. Это может
также происходить в пустынях после тёплых безморозных зим, которые способствуют
раннему развитию цветочных почек – за целые недели до того, как прилетят мигрирующие
опылители. Это ещё один тип ландшафта, где все опасности беспорядочных связей
проявляются слишком очевидно. Юкки и юкковые моли составляют один из примеров
облигатного партнёрства, которое складывается между специализированными опылителями
и синхронно цветущими растениями – и потому они сильно подвержены влиянию множества
дестабилизирующих сил. Неудивительно, что такие отношения тет-а-тет составляют
менее одного процента от всех наблюдаемых случаев взаимодействия растения и
опылителя в местообитаниях, которые были интенсивно исследованы.
Но другие ландшафты растения и опылителя могут оказаться столь же уязвимыми,
как те, в которых опылители специализируются на синхронно цветущих растениях.
Если специализированный или неспециализированный вид, зависящий от последовательного
цветения нескольких видов, обнаруживает, что одно звено в этой цепи разорвано,
скажем, из-за разрушения среды обитания, или из-за избирательного удаления растения,
представляющего собой критически важное звено, он может оказаться неспособным
выждать без корма, пока станут доступными другие ресурсы. Иными словами, даже
опылители, которые не демонстрируют жёсткой зависимости от единственного вида
цветов, могут стать уязвимыми из-за своей зависимости от целого списка цветочных
ресурсов.
Более 20 лет назад выдающийся специалист по экологии тропиков Дэн Дженсен обеспокоился
уязвимостью таких взаимодействий, когда сухие субтропики Коста-Рики фрагментировались
всё сильнее, превращаясь в мешанину островков вдоль Панамериканского шоссе.
Дженсен пророчески писал об опасностях, с которыми придётся столкнуться облетающим
кормовой маршрут колибри, которые каждый день должны искать пищу на маршруте
длиной от 10 до 12 миль, пролегающем через сухой лес, чтобы собрать достаточно
корма для себя и для своего молодняка. Когда значительная часть леса вдоль их
маршрутов вырублена или превращена в пастбище, весь маршрут превращается в ничто.
Дженсен писал: «То, что скрыто от наших глаз… представляет собой гораздо более
коварный вид вымирания: это исчезновение экологических взаимодействий».
Хотя немногочисленные зарегистрированные случаи истинно облигатных опылителей
и синхронно цветущих растений остаются особенно уязвимыми, другие три типа ландшафтов
растения и опылителя также могут пострадать из-за исчезновения мутуалистических
отношений. Однако каждое из них произойдёт по своему собственному сценарию и
в своём собственном темпе. Легче предсказать взаимосвязанные вымирания для строгих
мутуалистов, у которых не окажется японских белоглазок или деревьев охайя лехуа,
способных остановить их падение в пучину вымирания. Однако и другие типы взаимоотношений
также могут подвергнуться глубокому разрушению. И едва мы начинаем уделять внимание
ландшафтам растения и опылителя во всей их полноте, как понимаем, что большая
картина быстро ломается, трескается или разлетается в клочья.