AFTER MAN
A ZOOLOGY OF THE FUTURE
BY DOUGAL DIXON
INTRODUCTION BY DESMOND MORRIS
ST. MARTIN'S PRESS
NEW YORK
Published in the United States of America in 1981
by
St. Martin's Press
175
Fifth Avenue
New York
NY 10010
Library of Congress Number 81-56345
ISBN 0-312-01163-6
© Harrow House Editions Limited 1981
For information, write: St. Martin's Press
All rights reserved. No part of this work covered by the copyright hereon may be reproduced or used in any form by any means - graphic, electronic, or mechanical, including photocopying, recording, taping or information storage and retrieval systems - without written permission of the publisher.
Edited designed and produced by
Harrow House Editions Limited
7a Langley Street, Covent Garden, London, WC2H 9JA
Edited by James Somerville
Designed by David Fordham
Phototypeset by Tradespools Ltd., Frome, England
Illustrations originated by Gilchrist Bros, Ltd., Leeds, England
Printed and bound in the United States of America.
ПРЕДИСЛОВИЕ Десмонда Морриса 9
ПРЕДИСЛОВИЕ АВТОРА 10
ЭВОЛЮЦИЯ 11
Генетика клетки : Естественный отбор : Поведение животных : Форма и Развитие : Цепи питания
Происхождение Жизни : Ранние формы жизни : Эра рептилий : Эра млекопитающих : Эпоха человека
Мир после Человека
ЛЕСА И СТЕПИ УМЕРЕННОГО ПОЯСА 36
Кролопы : Хищники : Жители подлеска : Древесные жители : Ночные животные : Заболоченные земли
Травоядные млекопитающие : Охотники и их добыча : Жизнь на деревьях
Мигранты : Мичинг и его враги : Северный Ледовитый Океан : Южный Океан : Горы
Обитатели песков : Крупные животные пустынь : Североамериканские пустыни
Поедающие траву : Гиганты равнин : Мясоеды
Полог леса : Живущие на деревьях : Подлесок : Жизнь в воде
Австралийские леса : Подлесок австралийских лесов
ОСТРОВА И ОСТРОВНЫЕ КОНТИНЕНТЫ 100
Южноамериканские Леса : Южноамериканские пампасы : Остров Лемурия
Острова Батавия : Острова Пакаус
Участь жизни
Словарь : Древо Жизни : Указатель:
Благодарности
ПОСВЯЩАЕТСЯ ГЭВИНУ
Автор и издатели хотели бы выразить благодарность иллюстраторам этой книги. Это Диз Уоллис (представлен Folio); Джон Батлер и Брайан МакИнтайр (представлены Ian Fleming and Associates Ltd); Филип Худ; Рой Вудард (представлен John Martin and Artists Ltd); и Гарри Марш
Как только я увидел эту книгу, я пожалел, что не написал
её собственноручно. Это изумительная, красиво поданная идея. Много лет назад,
будучи молодым зоологом, я начал придумывать несуществующих животных, рисуя
их карандашом и красками, в качестве приносящего удовольствие отступления
от требований моих научных исследований. Свободный от ограничений эволюции
такой,
какая
она есть, я был волен следовать моим собственным, личным эволюционным капризам.
Я мог придумывать чудовищ и странных существ, растущих как растения, и невероятных
животных любого цвета, формы и размера, как мне нравилось, позволяя им изменяться
и развиваться согласно моим собственным правилам, давая полный простор своему
воображению. Я называл их своими биоморфами, и они стали для меня столь же
реальными, как животные и растения реального мира.
Мышление Дугала Диксона, очевидно, работало похожим способом, хотя существа,
которых он вызвал к жизни, сильно отличаются от моих. Вместо изобретения параллельной
эволюции, как если бы это имело место в другом мире, он поставил перед собой
интригующую задачу рассмотреть эволюцию в будущем на нашей собственной планете,
основанную на видах, которые существуют в настоящее время. Взмахивая волшебной
палочкой времени и устраняя сегодняшние доминирующие виды, в том числе человека,
он получил возможность наблюдать мысленным взором, как небольшие животные постепенно
занимают место главных обитателей поверхности земли.
Распространив свой сценарий на отдаленное будущее, приблизительно 50 миллионов
лет после нашего времени, он предоставил членам его нового животного царства
время, чтобы пройти через драматические изменения в строении и поведении. Но,
делая это, он никогда не позволял себе становиться слишком экстравагантным
в своём изобретательстве. Он создал свою фауну будущего настолько кропотливо,
что каждый
вид животных преподает нам важный урок, повествующий об известных процессах
эволюции в прошлом – об адаптации и специализации, конвергенции и адаптивной
радиации. Представляя нам вымышленные примеры этих реально существующих процессов,
его книга не только приносит большое удовольствие от чтения, но также имеет
реальную научную ценность. Животные на этих страницах могут быть воображаемыми,
но они наглядно иллюстрируют целую цепь важных биологических принципов. Вот
он, способ, которым он превосходно уравновесил своё яркое воображение со строгой
научной дисциплиной, который делает его книгу настолько успешной, и его животных
настолько убедительными и, кстати, настолько же превосходящими монстров, зачастую
откровенно смешных, изобретенных второсортными представителями жанра научной
фантастики.
Единственная опасность от чтения этой восхитительной книги состоит в том, что
некоторые из вас могут достигнуть такой точки, когда внезапно чувствуешь печаль
от мысли, что животные, скрупулёзно описанные в ней, не существуют сейчас.
Было бы очень здорово, если бы можно было отправиться в экспедицию и наблюдать
их всех, бродящих по поверхности сегодняшней земли, с помощью бинокля. Если
говорить о себе лично, я ощущаю это очень остро, когда листаю эти страницы,
и, вероятно, нет на свете большей похвалы, которую я могу предложить автору,
чем эта....


 Эволюция
– это процесс усовершенствования. Следовательно, смотреть на животных и растения
наших дней, и на их взаимодействие – хрупкое равновесие между флорой, травоядными
и хищниками; точную конструкцию несущих нагрузку структур в позвоночнике жирафы;
изящную форму ступни обезьяны, которая позволяет ей как схватывать предметы,
так и лазать по деревьям, изысканную окраску кожи шумящей гадюки, полностью
скрывающую её среди мертвых листьев лесной подстилки – и пробовать спроецировать
всё это на будущее – это занятие на грани невозможного. А как вы смогли бы
улучшить что-то сверх совершенства?
Эволюция
– это процесс усовершенствования. Следовательно, смотреть на животных и растения
наших дней, и на их взаимодействие – хрупкое равновесие между флорой, травоядными
и хищниками; точную конструкцию несущих нагрузку структур в позвоночнике жирафы;
изящную форму ступни обезьяны, которая позволяет ей как схватывать предметы,
так и лазать по деревьям, изысканную окраску кожи шумящей гадюки, полностью
скрывающую её среди мертвых листьев лесной подстилки – и пробовать спроецировать
всё это на будущее – это занятие на грани невозможного. А как вы смогли бы
улучшить что-то сверх совершенства?
Одна из тенденций, которую, однако, можно легко заметить – это разрушительный
эффект, который человек оказывает на выверенное природное равновесие. Я не
напрасно взял его крайнее проявление, когда человек уже уничтожил виды, численность
которых снижается в наши дни, и позволил себе чудовищное разрушение их естественной
среды обитания перед тем, как исчез сам, и тем самым позволил эволюции вновь
работать, восполняя нанесённый им ущерб и заполняя оставленные опустошёнными
места обитания. Исходным материалом для этого восполнения послужили виды животных,
которые преуспевают вопреки или благодаря присутствию человека, и которые переживут
его – те, которых человек расценивает как вредителей и нахлебников. Они выживут
с наибольшей вероятностью, нежели сильно изменённые и ослабленные близкородственным
скрещиванием животные, которых он выводит и искусственно сохраняет ради удовлетворения
своих собственных потребностей. Результат этого – зоология мировой фауны, рассмотренная
мною по своему желанию через 50 миллионов лет в будущем, и которую я использовал,
чтобы объяснить некоторых основные принципы эволюции и экологии. Результат
этого – предположение, основанное на фактах. То, что я предлагаю, не является
точным предсказанием – это скорее зондирование возможностей.
Мир будущего описан как бы путешественником во времени из наших дней, который
совершил путешествие в мир того времени и изучил его фауну. Такому путешественнику
будет известно кое-что о жизни современных животных, так что он сможет описывать
происходящее со ссылкой на разновидности животных, знакомые читателю. Его отчёт
написан в настоящем времени, как будто адресованный коллегам- путешественникам
во времени, которые отправятся в тот же самый период и пожелают сами исследовать
мир.
Усаживайтесь поудобнее, дорогие путешественники во времени, и наслаждайтесь
драматическим зрелищем эволюции жизни на вашей планете.

Дугал Диксон,
Варехам,1981
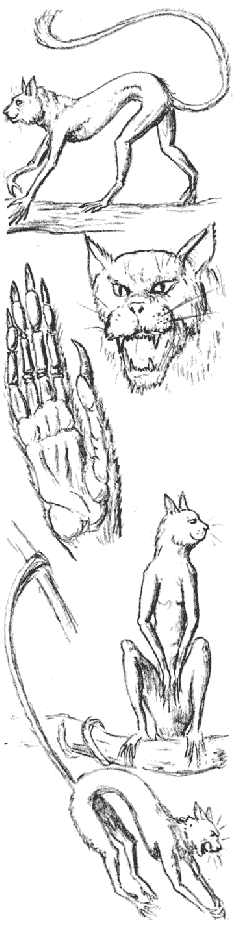
Эскизы на этой странице подобраны из собственных рабочих набросков автора, и были использованы художниками, чтобы создать таблицы и иллюстрации для «После Человека…».
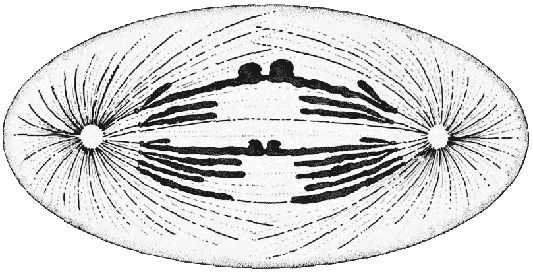
Живая клетка, показанная здесь в процессе самовоспроизводства, является основным строительным элементом, из которых состоят все живые существа. Способность клетки к неопределённой изменчивости, которая имеет место при половом размножении, лежит в основе эволюционного развития.
Форма и место живых существ на Земле могут быть отнесены на
счёт двух вещей –
эволюции и среды обитания.
Изучение эволюции выясняет, как возникла жизнь, как она стала разнообразной
в том виде, как сейчас, и как разные живые существа происходят от иных видов.
Изучение среды обитания (экологии) живого существа показывает, как разные
формы жизни взаимодействуют друг с другом и со средой, которую они населяют.
Иными словами, эволюцию можно представить себе как продольный срез жизни
на нашей планете,
тогда как экология показывает эту же ситуацию в поперечном
срезе.
Каждый из этих компонентов неотделимо связан с другим,
и они оба не могут изучаться
абсолютно раздельно.
Хотя оба этих аспекта способствуют выживанию, не следует забывать,
что вымирание
является очень важным фактором.
Без него не было бы пространства для эволюции.
В природе не было бы новых
экологических ситуаций, требующих заполнения путём эволюции новых животных
и растений из
«старых запасов». Эволюция, имеющая место сейчас, прослеживается как по палеонтологической
летописи, так и по свидетельствам, которые предоставляют
ныне живущие растения
и животные.
Исследование ископаемых остатков показывает общую тенденцию развития
от простого
к более сложному, и ту роль, которую играет среда в подгонке организма
к
преобладающим условиям. У ныне живущих организмов сходство в строении,
эмбриональном
развитии и биохимии является чётким указанием сходной эволюционной истории
или общего
происхождения. Поэтому эволюция – это не процесс, который происходил только
в прошлом, для того, чтобы приспособить растения и животных к сегодняшней
экологии; но она является процессом, который происходит постоянно,
который мы можем изучать
как по его результатам,
так и по ископаемым свидетельствам прошлого.
Она
происходила, происходит сейчас, и будет продолжаться настолько долго,
насколько
долго жизнь
будет существовать на нашей планете.
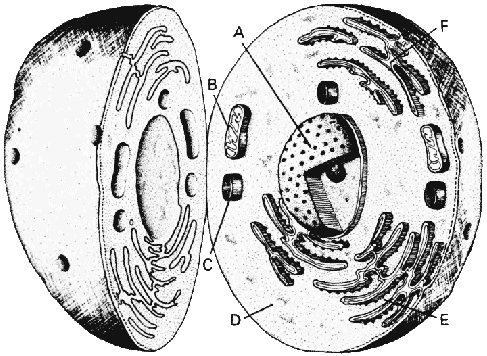
Большинство клеток животных содержит одни и те же главные компоненты. В центре лежит ядро (А), которое соджержит генетический материал клетки. Митохондрии (В), отвечающие за выработку энергии, и лизосомы (С), которые секретируют химические соединения, лежат ближе к поверхности цитоплазмы (D). Рибосомы (Е), где происходит биосинтез белков, лежат вдоль складчатых производных клеточной мембраны, известной как эндоплазматическая сеть (F). |
Животные, а также, разумеется, растения, сложены из микроскопических
кирпичиков, называемых клетками. Клетки, находящиеся в различных органах и
тканях одного и того же существа, обладают весьма различными размерами и формой
– кости сложены из угловатых клеток, почки – из округлых клеток, нервы из длинных
и тонких клеток – но все сложены из сходных компонентов. Снаружи каждую клетку
окружает клеточная оболочка – кожица, мембрана клетки, заключающая в себе студенистую
цитоплазму, которая несёт в себе множество маленьких структурных образований,
называемых органеллами. Наиболее важное из них – клеточное ядро, которое находится
в центре клетки и содержит информацию, на основе которой строится весь организм.
Информация хранится в виде кода, представляющего собой последовательность
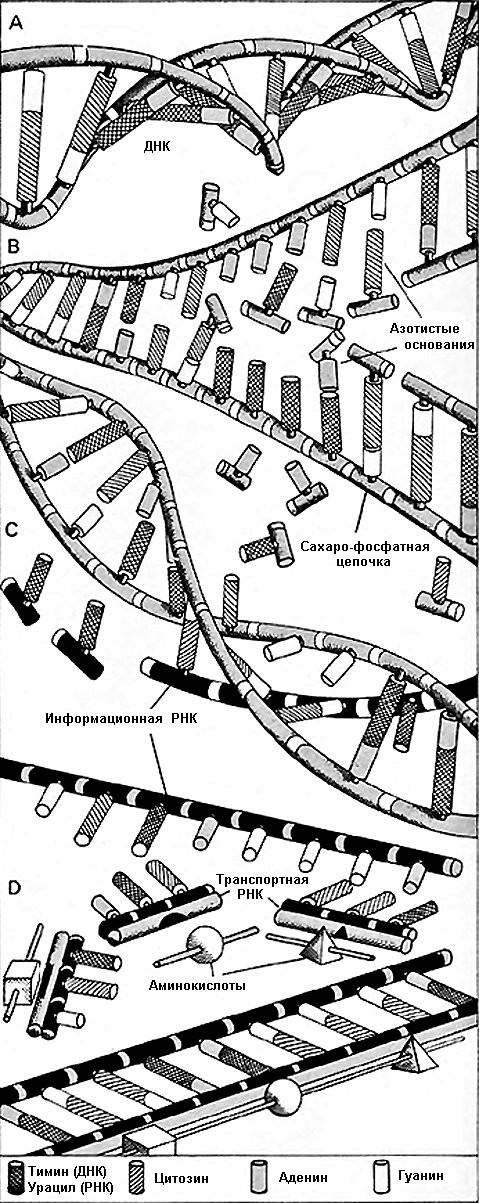
компонентов, содержащихся в длинной молекуле сложного соединения, известного
как дезоксирибонуклеиновая кислота (ДНК). Молекула ДНК немного похожа на лестницу,
скрученную вдоль. Пролёты лестницы состоят из сахаро-фосфатных молекул, а каждая
ступенька состоит из пары молекул, известных как азотистые основания. Существует
только четыре таких основания, и последовательность, в которой они располагаются
вдоль скрученной «лестницы», даёт кодированные инструкции, согласно которым
формируется организм. Хотя код полностью повторяется в ядре любой клетки организма,
лишь немногие участки кода используются для формирования каждого отдельного
органа.
Замечательной способностью молекулы ДНК является её способность к самовоспроизводству.
Молекула расщепляется вдоль и раскручивается таким образом, что каждая половина
«лестницы» состоит из «перил» и половинок «ступенек». Недостающие половинки
«лестниц» достраиваются из набора сахаро-фосфатных основ, который пополняется
за счёт пищи живого существа и находится в ядре любой клетки. Поскольку каждый
из четырёх типов азотистых оснований в цепочке притягивает к себе строго специфичный
тип азотистого основания, то, когда полностью формируются две новых цепочки
ДНК, они абсолютно одинаковы между собой по последовательности компонентов.
Это наиболее важный процесс, который лежит в основе размножения клеток и роста
всех организмов.
   |
|
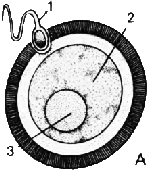
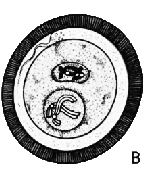
| ОПЛОДОТВОРЕНИЕ: 1. Сперматозоид 2. Яйцеклетка 3. Ядро яйцеклетки 4. Хроматида |
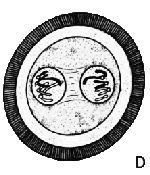 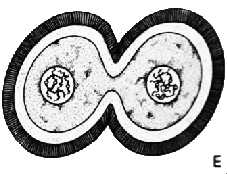 |
Сперматозоид
проникает в яйцеклетку (А) и движется к находящемуся неподалёку ядру
яйцеклетки (В). Хромосомы сперматозоида и яйцеклетки разделяются
на отдельные цепочки, известные как хроматиды. Соответствующие друг
другу хроматиды движутся к противоположным полюсам яйцеклетки (С),
где их окружает ядерная оболочка (D). Затем это образование разделяется
на две самостоятельных клетки (Е).
|
|
Тем не менее, чтобы расти, организму необходимы белки в форме тех или иных структурных элементов, таких, как коллаген, в случае соединительной ткани между органами, или фермент, который поддерживает специфические биологические процессы. Хотя биосинтез белков происходит вне клеточного ядра, он контролируется ДНК и происходит путём, аналогичным репликации ДНК. Посредником, который переносит инструкции ДНК к центрам биосинтеза белка, рибосомам, является молекула, известная как РНК. Она формируется вдоль частично «расстёгнутых» участков ДНК и лишь незначительно отличается от них. Информационная РНК движется к рибосоме, где к ней присоединяется другая форма РНК, транспортная РНК, которая переносит аминокислоты. Именно из этих аминокислот формируются белки. Молекулы РНК являются единственными переносчиками кода, и гарантируют, что аминокислоты соединяются в правильной последовательности, формируя белок. Таким образом ДНК контролирует работу всей клетки и всего организма..
| Во время деления клетки, когда образуются две самостоятельных клетки, ДНК (А), содержащаяся в делящейся клетке, разделяется вдоль и формирует новые молекулы ДНК вдоль своих свободных концов (В) из азотистых оснований и сахаро-фосфатов, содержащихся в клеточном ядре. Чтобы создать информационную РНК, цепочки ДНК расплетаются частично (С) и достраиваются очень похожим материалом; сахаро-фосфатный скелет незначительно отличается химически, и одно из азотистых оснований замещено (D). Информационная РНК содержит код, который даёт гарантию, что транспортная РНК присоединяется к верной последовательности, чтобы создать цепочку аминокислот, которая образует нужный белок. |
 |
Молекулы ДНК в клеточном ядре собраны в структуры, называемые
хромосомами, и определенные группировки последовательностей оснований нуклеиновых
кислот на ДНК дают начало определённым признакам организма. Эти группировки
называются генами. Половина хромосом в клетках живого существа, и, следовательно,
половина его генов, получена от его матери, и половина от его отца. Это отражается
в выравнивании хромосом в процессе деления клетки. В этот момент хромосомы
выстраиваются парами, полученные от матери выравниваются относительно идентичных,
полученных от отца, таким образом, чтобы соответствующие гены находились рядом.
Даже при том, что каждый ген в паре вносит вклад в определение той или иной
особенности, один ген часто маскирует эффект воздействия другого.
Будучи частью процесса размножения, в половых органах формируются
особые клетки, известные как гаметы – сперматозоиды и яйцеклетки – и содержащие
только половину набора хромосом, содержащегося в обычных клетках. Хотя в каждой
гамете представлена одна хромосома из каждой пары, ни одна из них не идентична
никакой из хромосом, полученных от отца или матери, поскольку содержит смесь
генетического материала от обоих родителей. Эта особенность хромосом в гаметах
в первую очередь отвечает за различия между разными индивидуумами одного вида,
которая наблюдается в природе. Во время оплодотворения гаметы объединяются
с гаметами другой особи, чтобы образовать полноценную клетку с полным набором
хромосом, который, в свою очередь, делится и формирует совершенно новый организм
с генетическими характеристиками обоих родителей.
Таков, в общих чертах, замысловатый процесс, который позволяет растениям и
животным самовоспроизводиться и передавать свои отличительные особенности от
поколения к поколению. В генах, вовлечённых в этот процесс, происходят небольшие
изменения, или мутации, которые позволяют происходить эволюции. Последствиями
мутации являются вариации признаков взрослого организма, выросшего из клетки,
содержащей ген. В большинстве случаев произошедшее изменение губительно, и
ведёт к проигрышу организма в окружающем мире конкуренции. Организм гибнет,
и вместе с ним гибнет мутантный ген. Тем не менее, иногда мутантный ген даёт
тенденцию, которая даёт организму заметное преимущество в борьбе за существование.
Разнообразие генетического материала, которое возможно благодаря половому размножению,
даёт определённый спектр характеристик, которые встречаются у представителей
отдельно взятого вида. Естественный отбор, который считается основной движущей
силой эволюции, оказывает воздействие на это разнообразие, благоприятствуя
отдельным характеристикам, и отметая остальные соответственно их пользе для
выживания.Молекулы ДНК в клеточном ядре сосредоточены в виде структур, называемых
хромосомами, и специфические группировки последовательностей азотистых оснований
в ДНК дают начало специфическим чертам самого организма. Эти группировки называются
генами. Половина хромосом в клетке живого существа, и, следовательно, половина
его генов, достаётся ему от матери, и половина от отца. Это отражается в расположении
хромосом во время деления клетки. При этом хромосомы выстраиваются парами,
и полученные от матери хромосомы располагаются параллельно полученным от отца
так, что соответствующие гены находятся друг напротив друга. Даже при том,
что каждый ген в паре вносит свой вклад в формирование какой-либо особенности,
один ген часто маскирует действие другого.
Естественный отбор, являющийся результатом условий, в которых живёт организм, может оказывать один из трёх видов воздействия на популяцию. Он может быть стабилизирующим, движущим, или разрывающим (дизруптивным). Стабилизирующее воздействие наблюдается, когда условия среды обитания остаются неизменными в течение долгого времени. Поэтому такая среда поддерживает хорошо сбалансированную популяцию животных и растений, в которой эволюционные изменения невыгодны. В силу таких обстоятельств любое изменение, происходящее у растения или животного, выведет его из чётко очерченной, эффективной, проверенной временем совокупности факторов, обеспечивающих выживание, и поставит его в невыгодное положение, которое в конце концов приведёт к его вымиранию. С другой стороны, его более консервативные сородичи выживут. Животные, длительное время подвергавшиеся действию стабилизирующего отбора, могут выглядеть сравнительно неспециализированными и примитивными по сравнению с животными, которые пережили более насыщенную событиями эволюционную историю. Часто они характеризуются пассивными механизмами выживания, такими, как тяжёлый панцирь или высокая плодовитость, компенсирующая потери от хищничества.
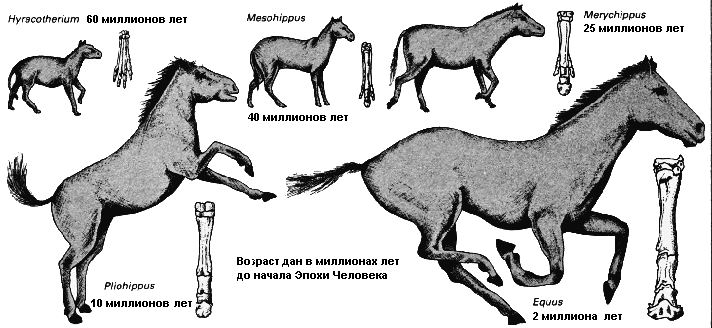
Самый ранний предок лошади, гиракотерий, маленькое длиннопалое существо не крупнее собаки, населял обширные лесные массивы, произраставшие на Земле между 60 и 50 миллионами лет до начала Эры Человека. Поскольку к концу третичного периода условия окружающей среды становились более сухими и лесные массивы отступали, это существо начало прогрессивно улучшать адаптации к жизни на равнине. Коренным образом изменились его ступни; наружные пальцы исчезли, оставив единственное роговое копыто. По мере того, как он превращался в полностью сформированное бегающее животное, его ноги становились длиннее, зубная система и пищеварительный тракт изменились от листоядного типа к травоядному, когда рацион менялся от листвы к траве. Самые важные структурные изменения произошли примерно в эпоху мерикгиппуса, который появился 25 миллионов лет назад. |
Действие движущего естественного отбора более ясно видно, когда меняется сама среда. В силу этих обстоятельств происходят изменения, которые производят впечатление, словно организм эволюционирует по определённому пути с некоей конечной целью. Это крайне ошибочное впечатление, которое возникает из-за того, что на фоне своей среды обитания наиболее поздний член эволюционного ряда всегда оказывается намного лучше приспособленным, чем более ранние промежуточные стадии, которые, там, где они известны, выглядят по сравнению с ним наполовину сформированными и неполноценными, даже хотя при этом они были столь же хорошо приспособленными к более ранним промежуточным состояниям самой среды обитания. Примером этого служит эволюция лошади, которая развивалась от мелкой листоядной формы, жившей в лесу, до крупной, длинноногой бегающей формы, питающейся травой, тогда, как её среда обитания менялась от леса к открытой травяной равнине. Небольшие изменения, которые позволяли ей более эффективно взаимодействовать с меняющимся окружением, непрерывно отбирались на протяжении её истории, и таким путём эволюционировала лошадь.
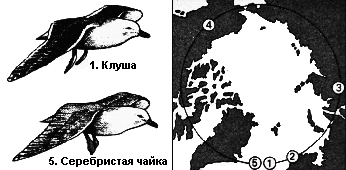
(1) Британская клуша, Larus fuscus graellsii, (2) клуша Larus fuscus fuscus, (3) восточносибирская чайка, Larus argentatus vegae, (4) американская серебристая чайка, Larus argentatus smithsonianus, (5) серебристая чайка, Larus argentatus argentatus*. * Иногда некоторые подвиды из числа указанных здесь считаются самостоятельными видами. – В. П. |
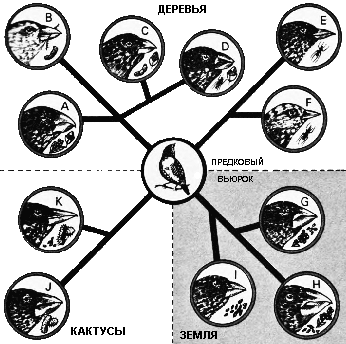
От исходного вьюрка, который прилетел
на Галапагосские острова из Южной Америки, произошло около пятнадцати
различных видов, чтобы заполнить свободные экологические ниши островов
– каждый вид с особыми характеристиками приспособлен к своему собственному
рациону. Вьюрки распадаются на три обширных группы по местообитанию
– кактусные, древесные и наземные – и различаются главным образом по
форме клюва. Полагают, что это началось со скудности птичьего населения
островов, что позволило вьюркам эволюционировать в формы, подходящие
ко всем доступным местообитаниям. |
Птицы с их способностью к полёту обычно являются первыми позвоночными, достигающими нового острова, и потому далёкие острова обычно могут быть местом обитания интересной авифауны. Типичны тяжёлые нелетающие птицы, такие, как моа, Dinornis, с Новой Зеландии, дронт, Raphus, с Маврикия, и «слоновая птица» эпиорнис, Aepyornis, с Мадагаскара, которые все эволюционировали в отсутствии наземных хищников*. Море, отделяющее остров, служило эффективным барьером, предотвращающим скрещивание между далеко улетевшими особями, достигшими острова, и исходной популяцией, оставшейся дома. Подобные барьеры для скрещивания обязательны для эволюции новых видов.
| * Для эпиорниса это сомнительно. Со времён написания книги Диксона открыты остатки нелетающих эпиорнисообразных птиц (Stromeria, Eremopezus) из континентальной Африки, и в свете этого эпиорнис на Мадагаскаре представляет собой скорее сохранившегося реликта более ранних эпох. – В. П. |
Действие разрывающего (дизруптивного) отбора
имеет место, когда новое место обитания представляет широкий выбор пищевых
ресурсов и мест обитания. Вид животных, попав в такие условия окружающей среды,
может успешно дать начало различным формам, которые специфическим образом приспособлены
к каждому из этих жизненных пространств, или экологических ниш. При отсутствии
животных-конкурентов эти различные формы со временем разовьются в совершенно
новые виды. Это событие, которое случается, когда остров или группа островов
образуется из-за вулканической активности в открытом океане. Необитаемый остров
медленно колонизируется животными, которые постепенно образуют новые виды,
чтобы эффективно использовать всю его территорию. Классический пример нарастания
эволюционного разнообразия наблюдается на Галапагосских острова в Тихом океане.
В ранний период их истории их заселил мелкий вьюрок, который со временем эволюционировал
в древесные насекомоядные формы, потребителей семян с массивными клювами и
форму, которая поедает личинок жуков, обитающих в древесине, вытаскивая их
кактусовыми иглами. Большое количество получающихся в итоге видов указывает
на большое количество экологических ниш, доступных на островах.
Расы или подвиды часто сосуществуют в пределах одного и того же ареала, осваивая
немного различающиеся среды обитания или источники пищи, но сохраняя способность
к скрещиванию. Они могут даже существовать в виде цепи подвидов, протягивающейся
из одной области в другую, и каждый подвид способен скрещиваться с соседними.
Когда виды на концах цепи достаточно различны, цепь носит название клина. Иногда
клин может формировать кольцо, например, окружающее горный хребет, где два
концевых члена, хотя и соседствующие, и родственные, настолько различаются,
что скрещивание между ними невозможно, и являются, с формальной точки зрения,
разными видами. Это ставит проблемы в таксономии, поскольку, если скрещивание
возможно везде на протяжении кольца, его члены должны быть чётко определены
как подвиды одного и того же вида.
Когда группа оказывается изолированной от исходной популяции, она может развиваться
самостоятельно до такой стадии, что позже, когда изолирующий барьер исчезает,
и две популяции встречаются снова, скрещивание между ними более невозможно.
Они являются, по определению, двумя различными видами. Различия ещё более обостряются,
когда новое местообитание, которое нашла для себя изолированная группа, является
большей частью неблагоприятным для жизни. Группа быстро исчезнет, за исключением,
может быть, нескольких особей, обладающих крайней степенью выраженности видовых
признаков, которые проявляют некоторую приспособленность к новой среде. Вид,
который позже разовьётся, будет происходить от тех нескольких особей, которые
изначально были генетически отличными от основной популяции, и случайно были
носителями генетических задатков, которые делали их от рождения более перспективными
для выживания.
Поскольку живые организмы способны к неограниченной изменчивости и имеют наследственную
тенденцию изменяться, поселяясь в нестабильной среде, новые виды возникают
гораздо быстрее, когда среда быстро меняется. Эволюция настолько результативна,
что ни одна экологическая ниша не остаётся пустой надолго. Кто-то всегда развивается,
чтобы заполнить её.
Эволюция не использует осознанное стремление организма. Она
или протекает посредством какой-то адаптации, которая вызвана воздействием
на него среды обитания, или некой жизненной стратегии, которую организм выработал
в течение жизни и передал своему потомству. Это происходит, потому что некоторые
особенности генетической структуры живого организма отбираются ради, или вопреки
некоторым особенностям среды, в которой он обитает. В этом контексте среда
понимается как физическое окружение организма, такое, как топография, температура,
или количество осадков, а также другие организмы, которые сосуществуют с данным,
кого ест он, или кто ест его.
Скорость эволюции имеет мало общего со скоростью, с которой происходят генетические
мутации – более важным фактором является скорость изменения окружающей среды,
скорость, с которой открываются новые пути, по которым могут эволюционировать
и развиваться новые формы жизни.
Будучи ответственным за структурные и морфологические особенности животного,
набор генов в клетке также лежит в основе поведенческих особенностей, которые
позволяют животному взаимодействовать с сородичами и средой обитания таким
способом, который гарантирует его выживание.
Это может быть объяснено тем, что особенности организма передаются следующим
поколениям исключительно через гены. В поддержку этой точки зрения могут быть
приведены доказательства из моделей поведения, наблюдаемых у животных. Поведение,
проще говоря, является активным ответом животного на его окружение, и наряду
с ростом и размножением является одним из факторов, которые являются определяющими
для живого.
 |
 |
 |
 |
 |
 |
Токовые игры птиц
являются важной частью поведения, которое также включает песню; песня
самцов птиц предназначена для того, чтобы в равной степени привлекать
самок и отпугивать пришлых самцов. Визуальные демонстрации могут
иметь место независимо от намерения привлечь брачного партнёра. Особь,
обычно самец, принимает определённые позы и подаёт сигналы, как только
заметит внимание возможного брачного партнёра. Затем пара токует
совместно; каждый из партнёров отвечает на жесты другого партнёра
с целью выяснить желание или готовность другого партнёра к образованию
пары. Многие виды используют для токования брачное оперение. В большинстве
случаев самцы нарочито ярко окрашены, тогда как самки выглядят скромно
по сравнению с ними. Движения и жесты в брачном поведении обычно
связаны с агрессией или умиротворением. У некоторых видов частью
токования являются чистка оперения или имитация сна. (A) Олуша, Morus bassanus, (B) Полынный тетерев, Centrocercus urophasianus, (C) Баклан, Phalacrocorax carbo, (D) Бролга, австралийский журавль, Grus rubicunda, (E) Чомга, Podiceps cristatus, (F) Пингвины Адели, Pygoscelis adeliae |
||
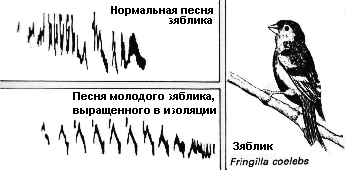
Исследования песни зяблика Fringilla coelebs привели к интереснейшей догадке о роли обучения в поведении. Было выяснено, как показано на сонограмме напротив, что молодые зяблики, выращенные в изоляции, способны только на примитивную песню, а чтобы воспроизвести её в полноценном виде, они должны вначале услышать песни других особей в природе. |
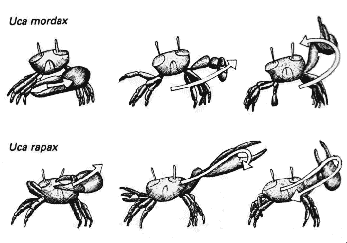
Самцы манящих крабов, Uca spp., привлекают брачных партнёров, взмахивая своими крупными «манящими» клешнями. Жесты, которые они производят, а также траектория и скорость движения варьируют у видов, обитающих в одной и той же местности и гарантируют, что будут привлечены самки только нужного вида. Поскольку только спаривание особей одного вида приводит к появлению плодовитого потомства, самцы, не обладающие генами, контролирующими правильный тип движений, будут выбраковываться. |
Птицы сбиваются в стаю, когда появляется ястреб, тем самым
делая сложнее для ястреба задачу схватить отдельную особь. Бегающие травоядные
меняют направление движения, чтобы спастись от более быстроногого хищника,
и потому он выбивается из сил, прежде чем добывает кого-то из них. Молодые
птицы остаются рядом с матерями, пока не станут достаточно зрелыми, чтобы защищать
себя самостоятельно. Эти явления, как и все аспекты поведения, эволюционировали,
чтобы способствовать выживанию. Ген, который контролирует способ поведения,
который не вносит вклада в выживание вида, вскоре устраняется.
Брачные ритуалы являются очень сложным аспектом поведения. Точное движение
птицы в демонстрационном танце или движении головы ящерицы, когда она приближается
к предполагаемому брачному партнёру, указывает её будущему партнёру, что они
находятся в состоянии готовности к размножению, и что они являются представителями
нужного вида. Последнее обстоятельство важно, поскольку, хотя спаривание между
двумя родственными, но различными видами может приводить к появлению потомства,
оно почти наверняка будет бесплодным. Такое спаривание – совершеннейшая трата
времени и сил с эволюционной точки зрения, поскольку оно не распространяет
успешно гены существа, и поэтому его следует избегать.
Все эти действия – инстинктивные наследственные типы поведения. Другие типы
поведения приобретаются путём научения, но в конечном счёте также проистекают
из общей совокупности генов животного. Способность вырабатывать соответствующее
действие методом проб и ошибок, или по примеру других особей вокруг себя является
способностью, которую дают животному его гены.
Агрессия – это элемент поведения, который, возможно, сложнее, чем это кажется
на первый взгляд. Можно было бы задать вопрос: почему, если цель агрессии состоит
в том, чтобы убрать конкурентов, животные не сражаются насмерть каждый раз,
когда возникает конфликт? Кроме того, что здесь возникает очевидный риск, ответ,
вероятно, состоит в том, что у животного нет никакой возможности убить всех
своих потенциальных конкурентов; убивая отдельных особей, оно с равной вероятностью
и помогает своим конкурентам, и приносит пользу себе. В большинстве случаев
схватка в животном мире принимает форму ритуализованных сражений и демонстрации
агрессии, которые наносят незначительные физические повреждения вовлечённым
в них существам, но устанавливают господство того или иного из участников.
Таким образом животное, которое выигрывает состязание, достигает того, что
оно намеревалось сделать – получить или отстоять в состязании ресурс без того,
чтобы самому пострадать от ранений. Проигравший также получает выгоду: он избегает
серьёзных ранений и сохраняет возможность участвовать в дальнейших состязаниях,
где он может в конечном счёте достичь успеха. Сложно увидеть, как эта стратегия
могла быть выработана путём научения, и наиболее вероятно, что она является
итогом эволюционного развития; животные, применяющие эту стратегию, с большей
вероятностью будут участвовать в воспроизводстве, и поэтому гены, ответственные
за поведение, получают преимущество при передаче перед другими, которые проявляются
в менее успешных способах поведения.
Во всём животном царстве различные формы поведения предназначены для того,
чтобы гарантировать скорее выживание генов индивидуума, нежели выживание самого
индивидуума. Исключение делается в отношении самых близких родственников, поскольку,
чем ближе родственник, тем большее количество сходных генов в его генотипе.
Оборонительный
инстинкт, который заставляет птицу-мать идти навстречу опасности, и даже жертвовать
собственной жизнью, чтобы спасти выводок
– это особенность поведения, рассчитанная на то, чтобы обеспечить выживание
собственных генов. Поскольку гены птицы-матери представлены в её потомстве,
и несколько особей из выводка имеют лучшие шансы для размножения и распространения
своих генов, нежели одиночная птица-родитель, сохранить жизни её птенцов, даже
ценой её собственной жизни, выгоднее для её генов. Менее очевидно поведение,
способствующее выживанию генов, у общественных насекомых, таких, как пчёлы
и муравьи. Член группы будет сражаться до смерти безотносительно выживания
колонии. В этом случае члены колонии гораздо ближе друг к другу генетически,
нежели другие животные в пределах одной размножающейся популяции. Потому выживание
колонии гарантирует выживание генов особи, несмотря на гибель особи.
Может показаться, что многие брачные приспособления, особенно такие, как у
птиц, скорее снижают шансы на выживание особи, нежели повышают их. Брачное
оперение самцов многих птиц, будучи заметным для брачного партнёра, столь же
заметно для хищника. Птицы, обладающие особенно длинными и расцвеченными перьями
хвоста, должны испытывать большое неудобство, спасаясь от врага. Возможно,
такие помехи для выживания могут быть способом показать, насколько успешен
данный самец – если он выжил, когда всё работает против него, стало быть, он
должен быть хорошим! Потому самку инстинктивно привлекает самец, устраивающий
наиболее экстравагантное токование.
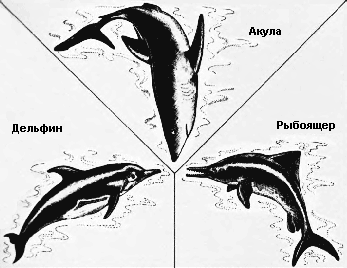
Естественный отбор устанавливает правило точности, согласно которому форма жизни наиболее пригодна для заселения определённого места обитания. Эта эволюционная тенденция может дать начало большому количеству различных, но внешне сходных животных. Когда животные произошли от общего предка и независимо развивались в сходном эволюционном направлении, говорят, что они эволюционировали параллельно. Когда предки различны, а животные эволюционировали в разных направлениях, которые в итоге привели к образованию сходной внешности, их эволюция называется конвергентной. Пример параллельной эволюции можно увидеть в развитии Equus, лошади, которая появилась в конце третичного периода в Северной Америке, и Toatherium, тоатерия, замечательно сходного с ней копытного, которое в это же время эволюционировало на изолированном тогда континенте Южной Америке. Эти две формы развивались независимо в сходном направлении от похожих копытных предков как ответ на сходный набор условий внешней среды.* Пример конвергентной эволюции можно найти в развитии акулы Carcharodon, рыбоящера Ichthyosaurus, и дельфина Delphinus – трёх животных из совершенно различных классов, выработавших сходную обтекаемую форму, плавники и хвост, чтобы занять одну и ту же нишу в одной и той же среде обитания – нишу активного морского рыбоядного хищника.
Среди акулы, рыбоящера и дельфина только акула произошла от морских предков. Рыбоящер и дельфин произошли от живших на суше рептилии и млекопитающего соответственно. Несмотря на их коренным образом различающееся происхождение, они все выработали сходную обтекаемую форму тела, соответствующую их водному образу жизни, и все вместе образуют яркий пример конвергентной эволюции. |
| * Сходство тоатерия и лошади не совсем полное, и параллелизм в развитии ограничивается опорно-двигательным аппаратом: несмотря на то, что тоатерий значительно опередил лошадь в развитии однопалых конечностей, приспособленных к бегу, его зубы остались приспособленными к пережёвыванию мягкой листвы, а не жёсткой травы. Другие примеры параллелизма более знакомы. Бывший отряд ластоногих (Pinnipedia) сейчас разделяют на семейства в составе отряда хищных (Carnivora). Так, по новейшим данным, настоящие тюлени произошли от куньих, а ушастые тюлени и моржи близки по происхождению к семейству медвежьих. Другой пример – южноамериканские ленивцы, которых делят на два семейства – трёхпалые ленивцы (Bradypodidae), произошедшие от древних ленивцев мегатериев, и двупалоленивцевые (Choloepidae), происходящие от мегалониксов. Иногда двупалых ленивцев напрямую относят к семейству Megalonychidae. – В. П. |
Одним из следствий того, что определённые формы облика животных
соответствуют определённым экологическим нишам, является то, что сильно удалённые
друг от друга местности с одинаковыми условиями климата и окружающей среды
могут поддерживать существование фаун, очень сходных внешне, даже если они
эволюционировали от разных корней. Тропические травянистые равнины Южной Америки,
Африки и Австралии, все одновременно, были населены животными со сходными физическими
характеристиками – длинноногими бегающими травоядными, быстрыми хищниками,
подземными насекомоядными существами и медлительными тяжеловесными пожирателями
листвы. В Австралии они были сумчатыми, в Африке плацентарными, а в Южной Америке
представителями обеих групп. Несмотря на разное происхождение, многие из этих
существ были сходными внешне. Подобные ситуации возникали не только в разных
местах одновременно, но также в разное время в разных местах.
Влияние географической широты оказывает два причудливым образом контрастирующих
друг с другом эффекта на облик и форму животных. Один, известный как правило
Бергмана, гласит, что в пределах группы родственных форм животные, обитающие
ближе к полюсам, будут крупнее. Другой, правило Алена, утверждает, что, опять-таки
в пределах группы родственных форм, те, кто живёт ближе к полюсам, будут иметь
более короткие конечности. Оба эффекта являются по сути мерами по сохранению
тепла, которые, с одной стороны, сохраняют температуру тела, а с другой, предохраняют
от обморожений.
Генетические изменения могут быть небольшими и почти незаметными, или могут
давать результат, коренным образом меняющий облик вида. Наземная улитка Cepaea
nemoralis живёт в разнообразных местообитаниях в лесном поясе умеренного климата,
и может иметь один из нескольких типов окраски раковины. Если местность открытая
и безлесная, однородная жёлтая окраска раковины маскирует улитку лучше всего,
и улитки с иными отметинами легче замечаются хищниками и быстрее съедаются.
Когда земля покрыта листовой подстилкой, коричневые полосатые формы являются
лучше замаскированными, и вместо них уничтожаются другие формы. Это даёт начало
популяциям преимущественно жёлтых улиток на открытой местности, поросшей травами,
и коричневым полосатым улиткам в лесной местности. Сходный эффект наблюдался
у берёзовой пяденицы Biston betularia в раннюю эпоху человеческой промышленной
революции. До той поры вид состоял преимущественно из крапчатых серо-белых
особей, которые отлично маскировались на покрытых лишайниками стволах деревьев,
где они жили. Чёрная форма, также встречавшаяся в популяции, была легко заметна
и чаще поедалась птицами, из-за чего была редкостью. С развитием тяжёлой промышленности
деревья покрывались копотью и чернели, обеспечивая превосходный фон для маскировки
чёрной формы. Белая форма, напротив, отсеивалась хищниками, и популяция бабочек
стала состоять преимущественно из чёрных особей. Позже, когда были приняты
законы о чистоте воздуха, в атмосферу и на стволы деревьев стало попадать меньше
копоти, и популяция пяденицы вернулась назад, к преобладанию серо-белых особей.
Эти изменения задействовали только разновидности в пределах одной и той же
размножающейся популяции, и всё время, пока они имели место, существовал постоянный
обмен генетическим материалом. Тем не менее, если изменения условий окружающей
среды были постоянными, и разные разновидности были изолированы одна от другой,
со временем они станут различными видами.
 Изменения
окружающей среды, связанные с промышленной революцией, дали чёрным
мутантным формам (B и C) берёзовой пяденицы Biston betularia преимущество
в промышленных районах, где они почти полностью заместили ранее доминировавшую
серо-белую крапчатую форму (A). В связи с низким уровнем загрязнения
атмосферы это почти не сказалось на популяциях из сельскохозяйственных
районов. Изменения
окружающей среды, связанные с промышленной революцией, дали чёрным
мутантным формам (B и C) берёзовой пяденицы Biston betularia преимущество
в промышленных районах, где они почти полностью заместили ранее доминировавшую
серо-белую крапчатую форму (A). В связи с низким уровнем загрязнения
атмосферы это почти не сказалось на популяциях из сельскохозяйственных
районов. |
ЮЖНАЯ АМЕРИКА |
АФРИКА |
АВСТРАЛИЯ |
|
КРУПНЫЕ ЛИСТОЯДНЫЕ
ФОРМЫ |
 |
 |
 |
Astrapotherium |
Носорог |
Diprotodon |
|
| ФОРМЫ, ПИТАЮЩИЕСЯ ТРАВОЙ |  |
 |
 |
Thoatherium |
Зебра |
Кенгуру |
|
| ХИЩНИКИ |  |
 |
 |
Prothylacynus |
Гиеновая собака |
Сумчатый волк |
|
| МЕЛКИЕ ВСЕЯДНЫЕ ФОРМЫ |  |
 |
 |
Neoreomys |
Соня |
Бандикут |
|
| ВИДЫ, ПИТАЮЩИЕСЯ МУРАВЬЯМИ |  |
 |
 |
Stegotherium |
Трубкозуб |
Сумчатый муравьед |
|
| РОЮЩИЕ НАСЕКОМОЯДНЫЕ ФОРМЫ |  |
 |
 |
Necrolestes |
Златокрот |
Сумчатый крот |
|
| НЕЛЕТАЮЩИЕ ПТИЦЫ |  |
 |
 |
Phororacos |
Страус |
Эму |
Рассматривая жизнь травянистых равнин Африки и Австралии в эпоху, близкую к эпохе человека, и сравнивая её с жизнью, существовавшей на равнинах Южной Америки немного раньше, в середине третичного периода, мы можем видеть, что животные со сходным образом жизни имели склонность эволюционировать в существ сходной внешности и размеров в соответствующих местообитаниях. Не имеет значения, что эти местообитания разделены во времени, в пространстве, или и тем, и другим образом одновременно; они являются единственным наиболее значимым эволюционным фактором, определяющим внешность живых существ. Крупные растительноядные звери, очень похожие внешне на носорогов, и длинноногие стремительные травоядные появлялись во всех трёх местообитаниях. Эволюционировали хищники, насекомоядные и всеядные животные, внешне похожие друг на друга. Наиболее явно сходными группами были роющие насекомоядные существа и нелетающие птицы, которые, в связи с их высоко специализированным образом жизни, явно развивались сходными путями.*
* Данное утверждение Диксона сомнительно
в отношении американских фороракосов, образ жизни которых сильно отличался
от образа жизни африканских
страусов и
австралийских эму. Если страусы и эму – крупные всеядные птицы, употребляющие
много растительных кормов, то фороракосы, судя по строению их скелета и клюва,
были активными хищниками и падальщиками, зачастую огромных размеров. Относительно
тоатерия см. примечание выше. – В. П.
Мимикрия – это особый случай явления имитации, при котором существо, обычно с целью защиты, принимает облик иного животного или растения, или даже полностью неодушевлённого объекта вроде птичьего помёта. Если животное подражает другому животному, возможны две важных формы этого явления. Первая, известная как мимикрия Мюллера, имеет место в случае, когда некоторое число опасных или несъедобных видов вырабатывает сходную окраску или рисунок, чтобы получать защиту совместными усилиями. Животные, демонстрирующие данную форму мимикрии, имеют тенденцию приобретать броские цвета, которые делают их видимыми издалека на окружающем фоне и служат предупреждением. Вторая форма, мимикрия Бейтса, распространяется на совершенно безвредных животных, которые выработали окраску или внешнее сходство с несъедобными или опасными видами, чтобы получить преимущество от их предупреждающей окраски и таким способом избегать хищников. Существуют другие формы мимикрии, которые позволяют хищнику приблизиться к жертве, которой он сам подражает. Насекомые, а особенно бабочки с их броскими узорами на крыльях, являются мастерами мимикрии, но она также встречается у позвоночных, и среди растений.*
| * Диксон смешивает два явления: мимикрию и покровительственное сходство. Мимикрия включает случаи подражания животных животным с целью защиты, а имитация растений и объектов неживой природы относится к покровительственному сходству. Иногда, впрочем, покровительственное сходство включается в понятие мимикрии под названием «мимезия». – В. П. |
Как мы видим, скорость эволюции в большей степени зависит
от скорости изменения окружающей среды, нежели от каких-то особенностей животных
как таковых. Но даже
в этом случае заметно, что, чем выше ступень эволюционной лестницы, на которой
стоит животное, тем быстрее оно эволюционирует. Например, роды двустворчатых
моллюсков существуют, в среднем, 80 миллионов лет, роды рыб – 30 миллионов
лет, а роды копытных и хищных млекопитающих от шести до восьми миллионов лет.
Чем
меньше продолжительность жизни рода, тем быстрее другой род эволюционирует,
чтобы занять его место. Результатом этого является более активная смена родов
в наземных
местообитаниях, где жизнь в целом гораздо более эволюционно продвинутая, чем
в море.
Растения имеют тенденцию эволюционировать гораздо медленнее, чем животные,
и флора, существующая в Эпоху Человека, состоит преимущественно из растений,
которые
эволюционировали в начале мелового периода, когда динозавры ещё были доминирующими
формами наземных животных.
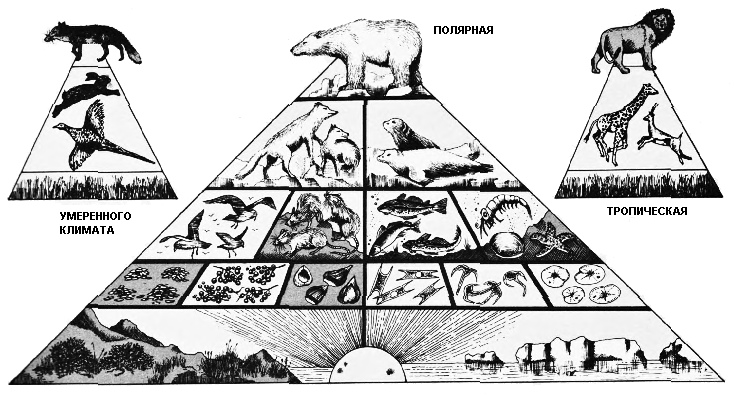
Пищевая цепь – это фундаментальное понятие экологии; это
последовательность, в соответствии с которой организмы едят друг друга. Вернее
всего будет представить этот процесс как пирамиду, чем как цепь, поскольку
в любом местообитании гораздо больше животных является низшими звеньями цепи,
нежели высшим звеном.
В основании пищевой пирамиды находятся растения, первичные продуценты, которые
используют энергию солнца, чтобы синтезировать первичную пищу из углекислого
газа воздуха и минеральных веществ почвы. От миллиардов членов этого широкого
основания пирамиды все цепи питания протягиваются наверх, к хищным животным,
стоящим на её вершине. Например, на далёком Севере в эпоху человека растения,
которые развивались в течение короткого полярного лета, служили пищей насекомым,
которых поедали мелкие птицы, которых, в свою очередь, ели мелкие хищники вроде
лисиц, которых, в конце концов, поедали крупные хищники типа белого медведя.
Сходным образом микроскопический фитопланктон, существующий в море, в то же
время находится в основании пищевой цепи, которая протягивается через рыб и
тюленей снова к белому медведю. Никто не охотится на живых белых медведей,
хотя, когда он умирает, падальщики и микроорганизмы из числа находящихся в
основании пищевой цепи кормятся на его трупе, разлагая его вплоть до неорганических
веществ, которые служат пищей растениям в основании пирамиды. Если исключить
мир паразитов, где количество организмов, питающихся на каждом уровне, скорее
возрастает, нежели уменьшается, пищевые пирамиды, подобные показанным выше,
могут быть воссозданы для любого типа местообитаний на Земле, в каждом случае
с одним хищником или небольшой группой хищников, находящихся на вершине.
На вершине пищевой пирамиды находятся хищники, последнее
звено в цепи передачи энергии, которая началась от растений – изначальных
продуцентов пищи. Эта пища в форме листвы и плодов переходит к травоядным,
находящимся на пирамиде выше, и исключительно через них к хищникам.
Подобные пирамиды существуют по всему миру, от тропиков до полюсов. |
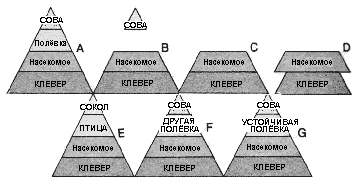
Общими уровнями пирамиды являются уже упоминавшиеся выше первичные продуценты, травоядные и хищники. На всех уровнях пирамиды действуют падальщики и микроскопические разрушители органики. Если один из ключевых членов уровня пищевой пирамиды будет удалён из-за болезни или изменений внешней среды, её структура будет нестабильной. Виды, находящиеся ниже освободившейся ячейки, могли бы увеличивать свою численность бесконтрольно до тех пор, пока позволяет пищевая база, и их численность контролировалась бы голодом. В действительности это происходит редко, и вскоре появляется другой хищник, способный занять свободную нишу.
На поле клевера (A) интересно предположить, что может случиться, если один уровень пищевой пирамиды будет удалён. Если полёвки будут большей частью уничтожены болезнью (B), совы будут лишены пищи, и вскоре покинут это место (C), предоставив популяции насекомых возможность бесконтрольно размножаться (D). Эта ситуация вряд ли продолжится, и свободная ниша будет быстро занята одним из трёх путей: совершенно новое насекомоядное существо, например, птица, поселится здесь, принеся с собой своего хищника (E), или здесь поселится иной вид полёвок, и за ним вернётся сова (F), или остатки былой популяции полёвок, устойчивые к болезни, возродят её (G). |
Согласно приблизительному «правилу большого пальца» хищник, требующий для существования одну единицу энергии, должен взять десять эквивалентных единиц энергии от травоядных, на которых он охотится. Сходным образом каждое травоядное должно получить десять единиц от растительности. Энергия для растений исходит исключительно от солнца, и вновь из десяти единиц, поглощённых растением, не более одной используется эффективно. |
Растения могут использовать только столько энергии солнца,
сколько могут уловить. Её трудно измерить, но только не более, чем примерно
одна восьмая процента энергии солнечного света, падающего на растение, может
быть запасена путём превращения в сахар. Химическая энергия из сахара используется
растением, чтобы строить всевозможные органические составляющие, которые идут
на построение его тела. Это тот сахар, и та энергия, заключённая в нём, которые
получает травоядное животное, когда ест траву. Тем не менее, и оно не может
использовать всю запасённую растением энергию для своих нужд – максимальная
эффективность животных составляет около десяти процентов. Этот фактор десяти
процентов присутствует на всех стадиях пищевой цепи, и означает, что в любом
местообитании сто травоядных могут обеспечить пищей только десять хищников,
которые, в свою очередь, обеспечивают существование только одного хищника «второго
уровня». Эти построения упрощены и относятся только к животным одного размера.
Вес животных является более важным фактором, чем число особей. Фактор десяти
процентов справедлив для любого уровня в сложном переплетении пищевых цепей,
и является важным фактором, поддерживающим стабильные очертания пищевой пирамиды.
Зависимость достатка в снабжении пищей от солнечного света – это причина, по
которой разные части Земли обеспечивают существование весьма различного количества
живых организмов. В тропиках, где солнечное освещение интенсивно, растениям
для усвоения доступно гораздо больше солнечной энергии. Следовательно, там,
где позволяют другие факторы, такие, как количество осадков, на единицу площади
приходится больше растительности, чем в областях с умеренным и полярным климатом.
Это большое количество растительного материала может обеспечить пищей большее
количество травоядных, которые, в свою очередь, являются пищей для большего
количества хищников. С другой стороны, в Арктике меньшее количество солнечной
энергии поддерживает гораздо более редкую растительность, и следовательно,
там меньше травоядных и гораздо меньше хищников.
Разнообразие видов на каждом уровне пищевой пирамиды зависит от разнообразия
растений в её основании. В тропической саванне, например, где есть низкие злаки,
травы, не относящиеся к злакам, высокие злаки, кустарники и деревья, каждый
из большого числа видов туземных животных поедает разный набор растений.* Потому
животное, которое ест корни, не конкурирует с животным, которое ест низкорослые
травянистые растения, или с животным, поедающим высокие злаки. Даже те виды,
которые имеют во многом похожий рацион, достаточно заметно различаются в иных
аспектах таким образом, что не конкурируют непосредственно – например, один
вид питается днём, а другой ночью.** Таким способом увеличивается число экологических
ниш, а процесс эволюции гарантирует, что все они будут заполнены.
| * Противоположным примером может служить примитивная экосистема сверхсолёных
водоёмов, где продуцентом является микроскопическая водоросль дюналиелла
(Dunaliella salina), которой питается только жаброногий рачок артемия (Artemia
salina). – В. П. ** Такое явление известно для осьминогов, живущих на Гавайях – один вид активен днём, второй ночью, а третий в сумерках – утром и вечером. – В. П. |
Принцип, согласно которому «природа не терпит пустоты», верен в биологии
так же, как и в физике. Экологическая ниша никогда не остаётся пустой надолго,
и кто-то эволюционирует, чтобы её заполнить, как только она образуется. Внутри
каждого вида, однако, конкуренция сильна, и каждая отдельно взятая ниша прокормит
только определённое количество особей. Столкновения между особями одного вида
обычно преобразованы в стилизованные демонстрации, в которых участникам наносится
минимальный ущерб. Территория сохраняется и брачные партнёры выбираются без
помощи какой-либо реальной схватки. Это является стратегией, которая обеспечивает
больший успех в поддержании позиции живого существа в экосистеме.
Влияние хищничества плотоядных животных слишком мало, чтобы нарушить баланс
пищевой пирамиды. Добывая только слабых, больных и стареющих особей – вынужденная
практика в свете того факта, что здоровая взрослая особь обычно может спастись
бегством или отразить атаку – они гарантируют, что выживет только наиболее
приспособленный. Если приспособленные и здоровые взрослые особи вида не могут
спастись бегством или отразить нападение, этот вид быстро вымирает, а нишу
занимает другой вид. В этом смысле хищники могут считаться не более, чем нетерпеливыми
падальщиками.
 |
 |
 |
 |
 |
 |
|
ВЕЛИКОЛЕПИЕ ЭВОЛЮЦИИ
|
||||||
ПРЕДИСЛОВИЕ Десмонда Морриса 9
ПРЕДИСЛОВИЕ АВТОРА 10
ЭВОЛЮЦИЯ 11
Генетика клетки : Естественный отбор : Поведение животных : Форма и Развитие : Цепи питания
Происхождение Жизни : Ранние формы жизни : Эра рептилий : Эра млекопитающих : Эпоха человека
Мир после Человека
ЛЕСА И СТЕПИ УМЕРЕННОГО ПОЯСА 36
Кролопы : Хищники : Жители подлеска : Древесные жители : Ночные животные : Заболоченные земли
Травоядные млекопитающие : Охотники и их добыча : Жизнь на деревьях
Мигранты : Мичинг и его враги : Северный Ледовитый Океан : Южный Океан : Горы
Обитатели песков : Крупные животные пустынь : Североамериканские пустыни
Поедающие траву : Гиганты равнин : Мясоеды
Полог леса : Живущие на деревьях : Подлесок : Жизнь в воде
Австралийские леса : Подлесок австралийских лесов
ОСТРОВА И ОСТРОВНЫЕ КОНТИНЕНТЫ 100
Южноамериканские Леса : Южноамериканские пампасы : Остров Лемурия
Острова Батавия : Острова Пакаус
Участь жизни
Словарь : Древо Жизни : Указатель: Благодарности
 В
эпоху человека цепочка подвидов, или клин, существовала вокруг Северного
Полюса между британской клушей, Larus fuscus graellsii, и серебристой
чайкой, Larus argentatus argentatus, её концевыми членами. Все соседствующие
виды клина могли скрещиваться между собой, за исключением концевых
членов, которые, к тому времени, как цепочка замкнулась, стали слишком
далёкими друг от друга, чтобы успешно скрещиваться.
В
эпоху человека цепочка подвидов, или клин, существовала вокруг Северного
Полюса между британской клушей, Larus fuscus graellsii, и серебристой
чайкой, Larus argentatus argentatus, её концевыми членами. Все соседствующие
виды клина могли скрещиваться между собой, за исключением концевых
членов, которые, к тому времени, как цепочка замкнулась, стали слишком
далёкими друг от друга, чтобы успешно скрещиваться.