Глава вторая
Вторая ступень: древние люди
(палеоантропы)
1. Ледниковая эпоха
Известные нам остатки древнейших людей — питекантропа и синантропа — найдены
в слоях второй половины четвертичного периода, который длился около 2 млн.
лет. Его можно подразделить на три эпохи, неравные по длительности и характеру:
доледниковую, ледниковую и послеледниковую, или современную (рис. 117). Первые
две ступени — это эоплейстоцен и плейстоцен, а третья соответствует голоцену
по терминологии, аналогичной той, которая принята и для третичного периода
кайнозойской
эры.
Для плейстоцена характерны наступание и отступание ледникового покрова на
значительных территориях северных областей Европы, Азии и Северной Америки.
По мнению П. П. Лазарева (1929), причиной похолодания были, по-видимому,
некоторые изменения в расположении суши и воды в северном полушарии, в околополярной
области. Это повлекло за собой нарушения в ходе теплых и холодных течений.
В конце третичного и начале четвертичного периодов «имелась полная изоляция
части будущего европейского материка от теплых океанских течений, и полюс
не получал совершенно теплой воды от
экваториального течения»,— пишет Лазарев.
Напротив, Г. Симпсон (Simpson, 1930) полагает, что причиной развития ледника
в плейстоцене послужило усиление солнечной радиации и испарения воды в океанах.
Однако этот взгляд не получил признания. Вернее полагать, что увеличение
влажности в атмосфере и некоторое понижение температуры оказались достаточными,
чтобы на севере начали накопляться массы льда наподобие тех, какие еще сохранились
в современной Гренландии, но по ее краям понемногу тают.
Распространяясь в южных направлениях, в частности по Европе, ледниковый покров
в период наибольшего своего развития покрывал не только Скандинавский полуостров
с островами Великобритании и Ирландии, но и части территорий, ныне занимаемых
Францией, Германией и Польшей. Он распространился также на
309

значительную часть территории современной Европейской части Советского
Союза, доходя на юге и юго-востоке до районов нынешних городов — Днепропетровска,
Тулы и Пензы.
Если сюда присоединить также области, бывшие подо льдом в северной Азии и
Северной Америке, то общую площадь, некогда покрывавшуюся льдом местами толщиной
в сотни метров, можно исчислить примерно в 8 000 000 км2.
Оставшиеся после всех ледниковых и межледниковых эпох наносы из песка, глины,
валунов, лёсса покрывают в настоящее время значительные площади, например,
в Европейской части Советского Союза. По ним, как и по соответствующим напластованиям
остальной Европы, устанавливаются
основные черты истории четвертичного периода, во время которого и происходило
формирование человечества начиная с питекантропа и синантропа. Неоднократные
колебания климата, надвигание и уход масс льда — все это имело огромное влияние
на ход развития животного и растительного миров в местах, близких к леднику,
тем более что в то же время происходило
Рис. 117. Ландшафты эпох плейстоцена
1 — доледниковая; 2 — ледниковая; 3 — начало послеледниковой. Из архива
Музея антропологии, Москва

понижение или повышение уровня некоторых областей этой части света и прилегающих
к ней участков других материков. Вследствие этого возможно было переселение
некоторых животных из Азии и Африки в Европу и обратно. Должен был кочевать
и древний человек, жизнь которого была тесно связана с наличием съедобных
растений и дичи.
Многие ученые считают, что было четыре ледниковые эпохи: 1 — гюнцская, 2
— миндельская, 3 — рисская и 4 — вюрмская. Они названы так ученым Альбрехтом
Пёнком по местностям в Швейцарских Альпах, где он подробно изучил следы этих
эпох. По его мнению, каждая эпоха длилась около 25 тыс. лет. Три несколько
более теплые «межледниковые» эпохи длились соответственно 175, 200 и 100
тыс. лет.
Для исчисления длительности четвертичного периода Пёнк предложил взять за
единицу времени промежуток, прошедший после вюрмского оледенения до настоящего
момента, т. е. 25 тыс. лет. При удвоении этой цифры, согласно предложениям,
сделанным советскими геологами, получаются в два раза большие длительности
эпох, абсолютные величины, которые установлены по совокупности данных, полученных
при изучении отложений четвертичного периода в Западной Европе и Европейской
части Советского Союза. Межледниковые эпохи называются: 1 — гюнц-миндельская,
2 — миндель-рисская, 3 — рисс-вюрмская.
В последнее время теория множественности ледниковых эпох, или полиглациализма,
начинает
колебаться в своих основах в связи с работами советских геологов и палеонтологов.
В этом смысле весьма интересны работы В. И. Громова (1948). Он считает,
что полиглациализм основывается преимущественно на данных литологии, с помощью
которой пытаются решать, например, вопрос о принадлежности валунных камней
и разных геологических образований к формациям ледникового происхождения;
что на данных только геоморфологии исчисляются, например, колебания снеговой
линии в прежние эпохи, причем исходят из наличия рядов речных трасс с нахождением
моренных образований на различной высоте. Между тем, говорит В. И. Громов,
данные флоры, фауны, ископаемых остатков человека для доказательства повторности
ледниковых и межледниковых эпох либо совершенно не принимаются во внимание,
либо учитываются не в полном объеме. По его мнению, учение о множественности
ледниковых эпох, в течение четвертичного периода чередовавшихся с межледниковыми,
характеризовав шимися даже более мягким климатом, чем современный, а также
более высоким положением снеговой линии, не имеет ни палеонтологического,
ни археологического обоснования.
По В. И. Громову (1948), различаются три фазы четвертичного периода, если
основываться на изучении особенностей истории фауны на территории Советского
Союза до 60° северной широты, преимущественно в Европейской части СССР:
1) доледниковая (миндель и миндель-рисс); 2) ледниковая (рисская стадия,
рисс-
312
вюрмский интерстадиал и вюрмская стадия) и 3) послеледниковая фаза.
Таким образом, В. И. Громов считает, что на территории Советского Союза холодоустойчивая
фауна арктического типа развилась и была характерной на протяжении длительного
времени лишь во второй половине четвертичного периода. До этого фауна не
была однообразной, так как обнаруживает смену трех генетически связанных
между собой фаунистических комплексов. Но арктической фауны ни один из
них не содержит.
На основании резкого изменения характера фауны непосредственно перед минделем,
когда окончательно исчезла субтропическая фауна и появилась новая, характеризующаяся
эласмотерием, южным слоном и этрусским носорогом, В. И. Громов проводит
границу между третичным и четвертичным периодами. В. И. Громов резкую смену
фауны
ставит в связь с ухудшением климата, которое в доминдельское и миндельское
время сперва выражалось усилением континентальности, а позже похолоданием
климата и оледенением Евразии. (Позднее В. И. Громов изменил свою точку
зрения и перенес границу третичного и четвертичного периодов на более раннее
время.)
В свою очередь резкое потепление после вюрмской стадии во время раннего
послеледникового времени и соответствующее изменение других природных условий
способствовали
резкому перелому в истории фауны, более быстрому вымиранию арктической
фауны и постепенному вымиранию или
обеднению фауны из-за исчезновения таких животных, как лошадь, верблюд,
росомаха.
Самое сильное испытание для очень многих видов животных наступило, несомненно,
при великом оледенении во времена рисской стадии. Оно привело к вымиранию
одних видов, миграции других и приспособительной эволюции третьих видов.
Характерными животными для этой эпохи являются трогонтериев слон, длиннорогий
бизон, гигантский олень
По мнению В. И. Громова, рисcкая ледниковая эпоха была критической эпохой
в развитии животного и растительного миров в соответствующих областях Евразии.
Нелегко пришлось в это время и древним людям, поскольку начало и развитие
максимального похолодания захватило мустьерскую культурную эпоху, т.е.
время неандертальцев, а вторая его половина — время кроманьонцев.
В. И. Громов считает, что на основании изучения остатков животных и орудий
можно относить неандертальцев к первой половине и более поздним эпохам
максимального оледенения, а кроманьонцев — ко второй его половине, с тем
что ориньяк падает
на вторую половину максимального оледенения, а солютре и мадлен на рисс-вюрм
и вюрм. Люди позднего палеолита охотились на мамонтов и северных оленей,
которые вместе с шерстистым носорогом были характерными представителями
этого времени: охотились они также и на лошадей.
Неандертальцы и гейдельбергский человек жили, по схеме
313
В. И. Громова, в мустьерскую, шелльскую и ашёльскую культурные эпохи,
в то время как синантропы жили в дошелльские времена, соответствующие гюнц-миндельской
эпохе, переходной от третичного периода к четвертичному периоду. Наконец,
питекантроп отодвигается В. И. Громовым в еще более раннюю эпоху, соответствующую
прежней гюнцской.
В общем из миллиона лет четвертичного периода первые 500 тыс. лет приходятся
на доледниковую эпоху, начало ледниковой и первую половину наибольшего
похолодания. На это время приходится, первая фаза культурного развития,
или так называемая
эпоха раннего палеолита, с очень низким уровнем техники выделки каменных
орудий (Ефименко, 1953).
Это стадия аморфных орудий, не имевших определенной формы и не классифицирующихся
на ясные типы. Между тем типы ручных рубил, скрёбел и наконечников сравнительно
легко определяются в культурных остатках на местах поселения людей эпохи
среднего палеолита, длившейся 300— 400 тыс. лет. Эпоха позднего палеолита
с ее богатством новых типов орудий относится к последним 100—150 тыс. лет
четвертичного периода.
Творцами аморфных орудий были преимущественно древнейшие люди, среднепалеолитических
— неандертальцы, позднепалеолитических, гораздо более сложных и своеобразных,
в том числе резцов и костяных орудий,— кроманьонцы.
Во время последовательных похолоданий, или ледниковых эпох,
животный и растительный миры, а вместе с ними и человек отходили перед
наступающими льдами на юг, юго-запад и юго-восток. Во время межледниковых
эпох люди вновь
распространялись по освобождавшимся территориям, которые заселялись в первую
очередь растениями, а за ними и животными.
Под влиянием непрерывной борьбы за существование при таких резких изменениях
природных условий постоянно изменялись и формы существования человека.
Вырабатывая орудия труда и сообща охотясь на небольших, а позже и на крупных
млекопитающих,
например слонов, человек превращался в их опасного соперника. Острые камни
и дубина в руках древнего человека нередко с лихвой возмещали недостаток
в собственных природных органах нападения.
Неандертальцы
и их физический тип
Неандертальская стадия эволюции гоминид (Бунак, 1966в) представлена большим
количеством костных остатков, обнаруженных в разных местностях Европы,
Азии и Африки. Неандертальцы жили преимущественно в среднюю эпоху древнего
каменного
века, в среднем палеолите. Это время относится к первой половине максимального
оледенения, соответствующего рисской эпохе. Иначе это отдаленное время
называют эпохой мустье.
Многие западные авторы приурочивают мустье к рисс-вюрму и даже вюрму. Весь
средний палео-
314
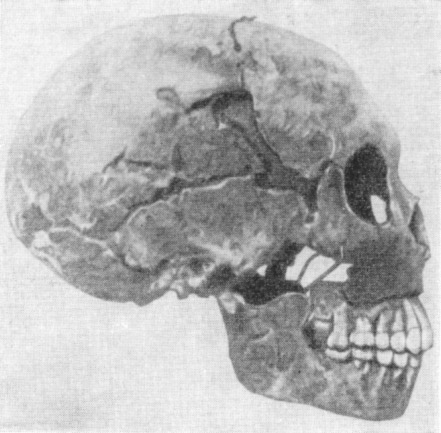
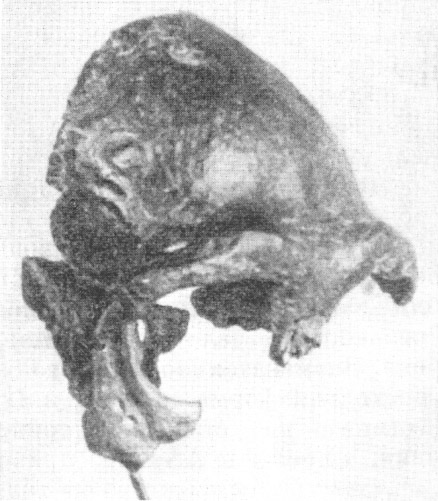
Рис. 118. Череп юноши неандертальца (Homo neanderthalensis King) из
нижнего грота Ле Мустье
По А. Грдличке, 1930, 1/3 нат. вел. (ширина изображения
56 мм)
|
лит, как мы уже говорили, занимает несколько сот тысяч лет. Более подробно
его подразделяют по характеру выделки каменных орудий. Костяные орудия
появляются лишь в самом конце мустье. Характерные для неандертальцев каменные
орудия
были найдены при скелете в нижнем гроте Ле Мустье, Франция, в 1908 г. (рис.118)
и в других местах.
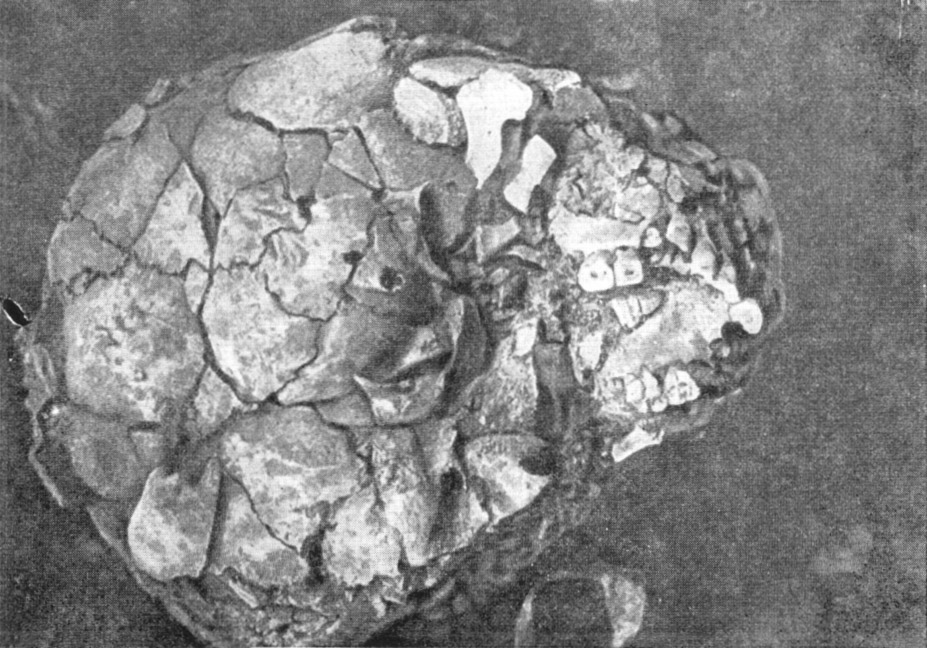
Первый череп взрослого неандертальца был найден в 1848 г. в каменоломне
на скале Гибралтар (Испания). Это был весьма неполный женский череп со
слабо
развитым надглазничным валиком (рис. 119). Объем мозговой коробки составляет
у него лишь 1080 см3. В 1926 г. там же были найдены
каменные орудия и остатки черепа ребенка неандертальца в возрасте пяти
лет.
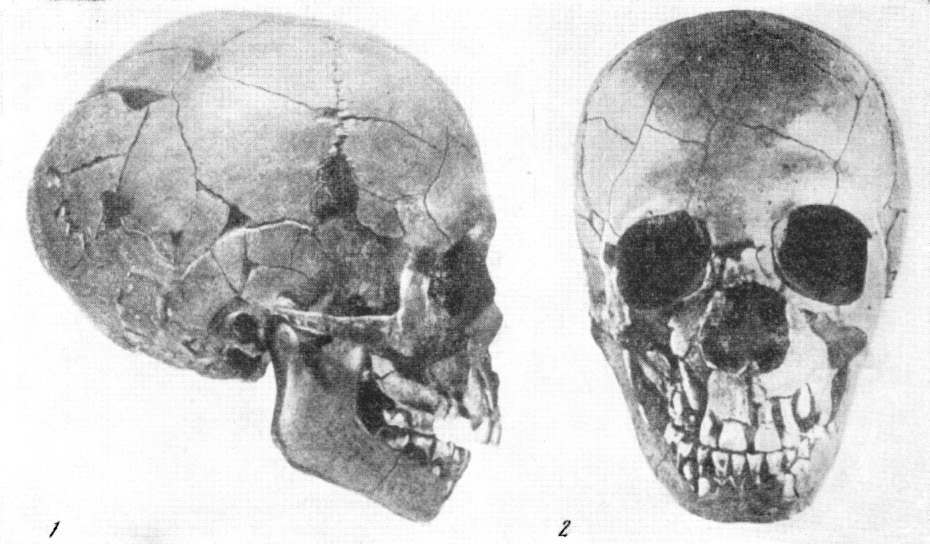
В 1856 г. в долине Неандерталь, в устье р. Дюссель, близ г. Дюссельдорфа
(Германия), в гроте Фельдгофер, также были найдены черепная крышка (рис.
120) и части скелета неандертальца. Для древних людей подобного типа вместимость
мозговой коробки значительна — около 1400 см3, но лоб очень покатый, а
выше глазниц располагается сплошной надглазничный валик, напоминающий подобное
же образование у шимпанзе.
Вокруг черепа из Неандерталя разгорелись большие споры. По мнению немецкого
ученого Рудольфа Вирхова, череп принадлежал современному человеку, но под
влиянием патологических процессов принял такую своеобразную форму. Он считал,
что позже на форме черепа отразилось также давление пластов земли.
Дарвин, не придавая черепу из Неандерталя должного значения, отметил лишь
его древность и значительные размеры. Эта находка долгое время стояла изолированно,
как и гибралтарский череп. Лишь гораздо позже начали находить все новые
черепа и кости подобного типа вместе с аналогичными грубыми каменными орудиями.
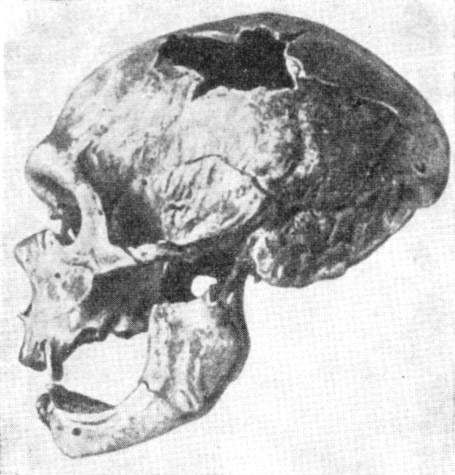
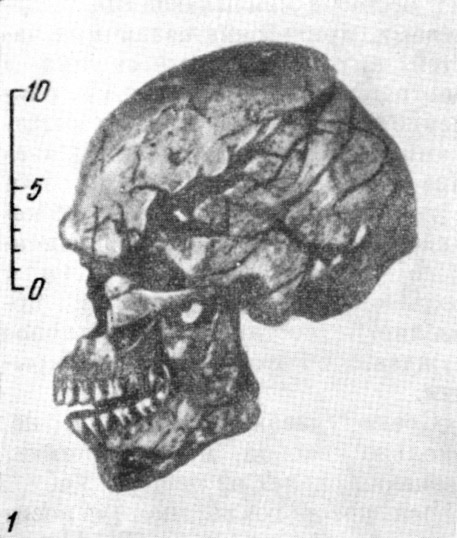
Особое значение имела находка в 1908 г. остатков скелета мужчины неандертальца
в возрасте 50—55 лет. Скелет был найден близ с. Ла Шапелль-о-Сен (Франция)
в пещере Буффия, в самом глубоком слое.
Его череп (рис. 121) имел совокупность особенностей, характерных для большинства
неандертальцев:
1) мощный надглазничный валик и сильно покатый лоб:
2) как бы сплющенный сверху вниз затылочный отдел;
315
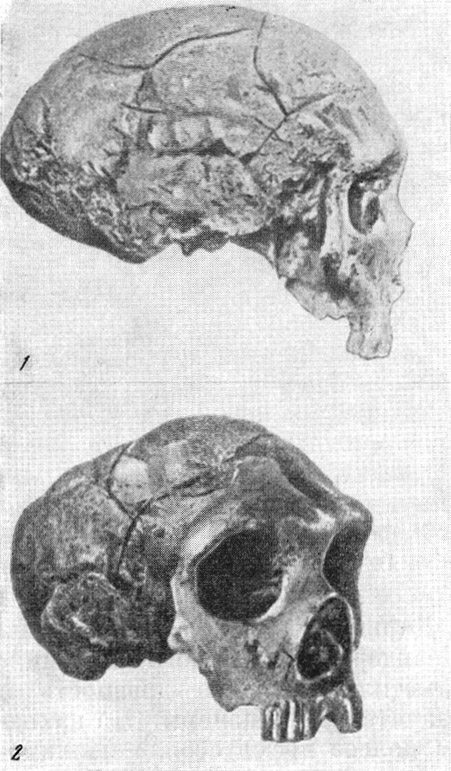
Рис. 119. Череп женщины неандертальца из Гибралтара
1 — вид в профиль; 2 — в три четверти (по слепку). По А. Грдличке, 1930
(1) и Т. Моллисону, 1932 (2), 3/8 нат. вел. (ширина
изображения 57 мм) |
3) горизонтально расположенный верхний край чешуи височной кости;
4) несколько притупленный сосцевидный отросток;
5) плоские и скошенные назад скуловые кости;
6) верхние челюсти без клыковых ямок, характерных для людей современного
типа;
7) массивную нижнюю челюсть без подбородочного выступа.
Но вместимость мозговой коробки этого черепа очень велика, приблизительно
1600 см3, что значительно превышает среднюю вместимость
неандертальских черепов.
По размерам головного мозга неандертальцы, таким образом, не уступали современному
человеку (рис. 122). Но лобные доли у них были невелики, и мозг имел ряд
черт сходства с мозгом человекообразных обезьян.
Форма черепа неандертальцев удлиненная, в частности из-за сильного развития
надглазничного валика. Черепной свод очень низкий. Черепа некоторых неандертальцев
по своей форме более близки к черепам современного типа, как, например,
череп из Эрингсдорфа (Германия, 1914 г.). Многочисленные находки остатков
костей
неандертальцев позволяют составить общее понятие о предшественниках типа
современного человека. Рост их был невелик, в среднем около 160 см.
Длина рук по отношению к длине ног была относительно немного короче, чем
у современного
европейца (Gieseler, 1959; Heberer, 1959, 1968).
Строение длинных костей нижней конечности у неандертальцев показывает,
что их ноги еще не были вполне хорошо выпрямленными в коленном суставе.
У них,
вероятно, была довольно неуклюжая походка, что характеризуется некоторыми
другими особенностями тела. Так, позвоночник у неандертальцев имеет малую
выраженность изгибов, столь характерных для людей современного типа (рис.
123).
Кроме европейских неандертальцев, за последние десятки лет открыто много
их представителей в
316
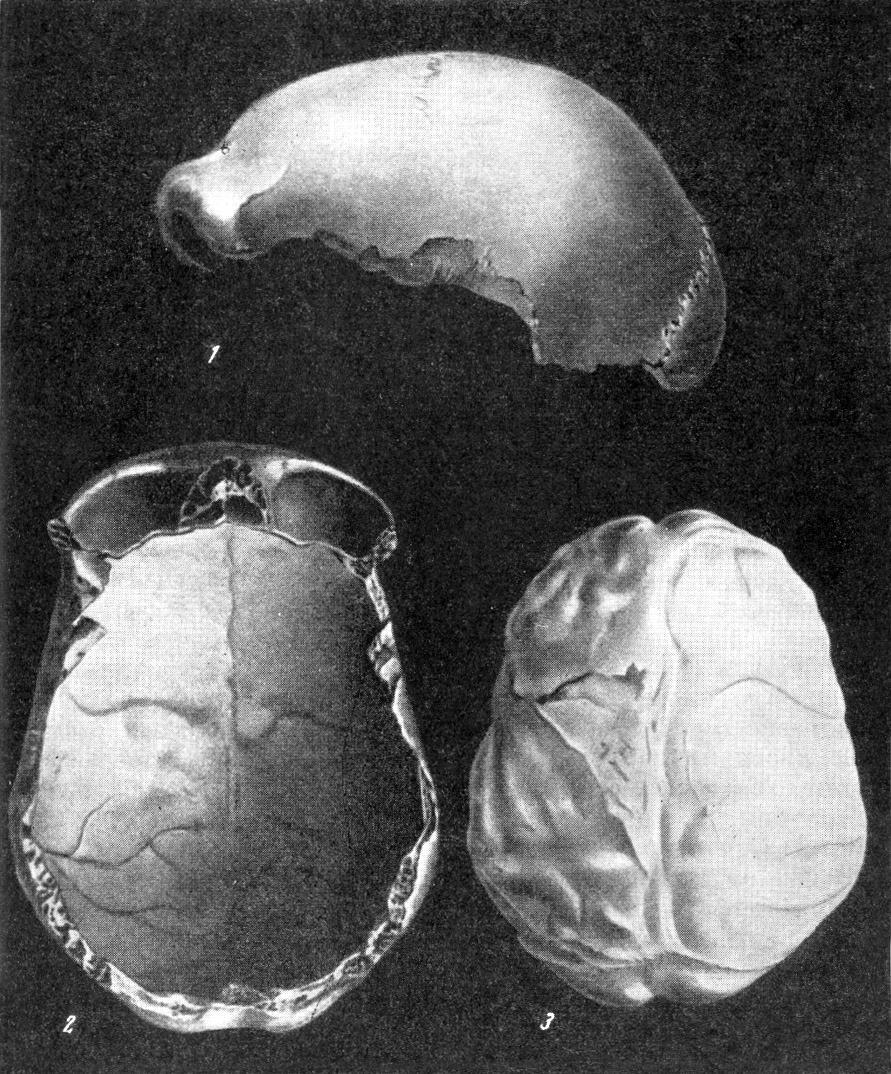
Рис. 120. Череп древнего человека из Неандерталя
1 — вид слева; 2 — снизу; 3 — слепок его мозговой полости. По Г. Шааффгаузену,
1858, 1/3 нат. вел. (ширина изображения 114 мм)
Рис. 121. Череп старика неандертальца из Ла Шапелль-о-Сен
По М. Булю, 1952, 1/4 нат. вел. (ширина изображения 58 мм)
|
Африке и Азии (рис. 124), благодаря чему выявилось еще большее разнообразие
типа палеоантропов и укрепилось мнение о преемственности генетической
связи с неоантропами, с видом «человек разумный» (Рогинский, 1966а).
В 1921 г. в Северной Родезии (на территории современной Замбии), в пещере
холма Брокен-Хилл, вместе с костями разнообразных, большей частью мелких
животных были обнаружены череп, крестец, фрагмент левой безымянной кости
таза, бедренная и большая берцовая кости левой нижней конечности древнего
человека. Эта находка имеет большое значение, хотя геологический возраст
ее неизвестен. Кроме того, принадлежность черепа и костей скелета одному
индивидууму не доказана.
Для черепа родезийского неандертальца характерен чрезвычай-
но мощный надглазничный костный валик с развитыми боковыми отделами,
очень покатый лоб, причем вдоль лобной кости заметен невысокий костный
валик;
на затылочной кости есть поперечный гребнеобразный валик; затылочное
отверстие располагается ближе к середине основания черепа, чем у большинства
неандертальцев:
это напоминает его положение у современного человека (рис. 124). Нижняя
челюсть
не найдена, но, вероятно, была крупнее гейдельбергской.
Объем мозговой коробки равен 1325 см3. По мнению некоторых
ученых, очертания мозга родезийца указывают на сходство с питекантропом (Abel,
1931).
Тип родезийца представляет форму, в которой совмещены очень древние признаки
(длинный череп, продольный валик на лобной кости, сильно покатый лоб,
очень мощный надглазничный валик) с прогрессивными — более переднее положение
затылочного отверстия, форма зубов, как у современного человека, высокий
рост (около 180 см). В общем же родезиец является примитивной
формой, и его положение в группе неандертальцев представляется до сих
пор неясным,
равно как
и
его значение для эволюции позднейших гоминид.
Некоторые авторы считают родезийца предковой формой для негроидной расы,
но для этого нет достаточных оснований. Правильнее считать его одним
из древних африканских представителей неандертальской стадии эволюции
человека,
уже
обнаруживающих некоторые черты перехода к типу строения современного
человека.
318
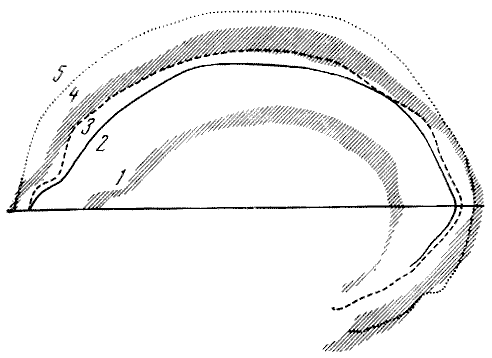
Рис. 122. Сравнение очертаний черепов
1 — шимпанзе; 2 — питекантроп; 3 — синантроп; 4 — неандерталец; б — современный
человек (кроманьонец из Комб-Капелль). По Г. Вейнерту, 1936
|
Если полагать, что череп принадлежит другому индивидууму, чем прочие
кости, то его архаичность делается нагляднее.Некоторые авторы даже полагают,
что
родезиец по грубости и массивности черепа напоминает антропоида вроде
гориллы (Герасимов,1955, стр. 196). Там же в Африке, но более далеко
на север, в
1932 г. были обнаружены фрагментарные остатки трех черепов древних неандертальцев.
Находка была сделана у озера Ньяраса, иначе Эяси, к юго-востоку от озера
Виктория.
Наиболее сохранившийся череп был реконструирован Вейнертом, который
считает, что у него имеется очень много примитивных особенностей,
как, например, мощный надглазничный и сильно выраженный затылочный валики,
наибольшая
ширина черепа в области сосцевидных отростков, наклон большого затылочного
отверстия
назад, как у антропоидов. Вместимость мозговой коробки невелика, всего
около 1200 см3.
Вейнерт называет эту форму ископаемых людей обезьяночеловеком, а ньярасского
человека — африкантропом. Однако другие ученые полагают, что реконструкция
сделана недостаточно точно. Да ее и трудно осуществить с полной точностью
из-за фрагментарности находок. Многие антропологи считают, что африкантроп
относится скорее к неандертальцам или к формам, переходным от обезьянолюдей
к первобытным людям (Washburn, 1963).
О широком распространении неандертальцев по территории Старого Света
свидетельствуют находки их остатков в Азии. Для уяснения процесса развития:
типа человека
в юго-восточной Азии большое значение приобретает находка остатков неандертальцев
на острове Ява. В 1922 г. геолог Ч. Тен Хаар производил изыскания на
террасе реки Соло в 30 км от Триниля — места находки питекантропа. Он
обнаружил
много костей ископаемых животных, например оленя, свиньи, бегемота, буйвола,
носорога,
крокодила, стегодона. Там же были найдены остатки пяти черепов древних
людей (рис. 125). Среди них была и очень хорошо сохранившаяся лобная
кость детского
черепа. В раскопках позже принял участие В. Ф. Ф. Оппенурс, который и
дал первоначальное описание черепов (Oppenoorth, 1937).
Величина черепов значительная. Длина одного из них достигает 221 мм,
т. е. он гораздо длиннее, чем череп средних размеров современного человека;
стенки
мозговой коробки толстые; вместимость ее составляет лишь 1000 см3.
По
319
названию селения Нгандонг, близ которого был найден этот череп, его обозначают,
как «Нгандонг I». Оппенурс предложил называть обладателей этих черепов
явантропами, вследствие того, что их обнаружили на Яве.
По Оппенурсу, череп I обнаруживает сходство с родезийским. Лобный отдел
сильно покатый, но сам черепной свод приблизительно на 2 см выше,
чем у питекантропа.
Значительно развит затылочный поперечный валик. Характерен мощный надглазничный
валик, по чрезвычайному развитию краев сходный с аналогичным валиком
на родезийском черепе.
Многие признаки сближают яванского неандертальца с европейскими. Но объем
мозговой полости черепа у него гораздо меньше, хотя череп I принадлежит
старому индивидууму.
Явантропа сближает с питекантропом большая длина лобной кости и возвышение
в области брегмы, т. е. пункта ее соединения
Рис. 123. Скелеты
1 — горилла; 2 — неандерталец; 3 — современный человек. По Г. Рэйвену,
1951 (1), по Г. Вейнерту, 1936 (2) и по М. А. Гремяцкому, 1945 (3)
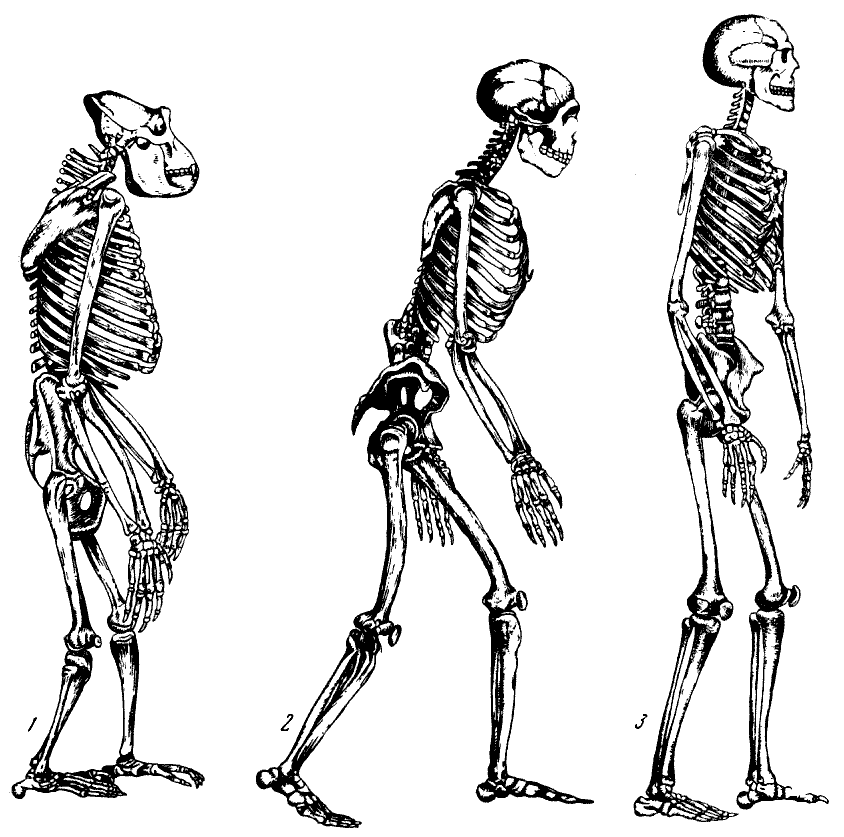
320
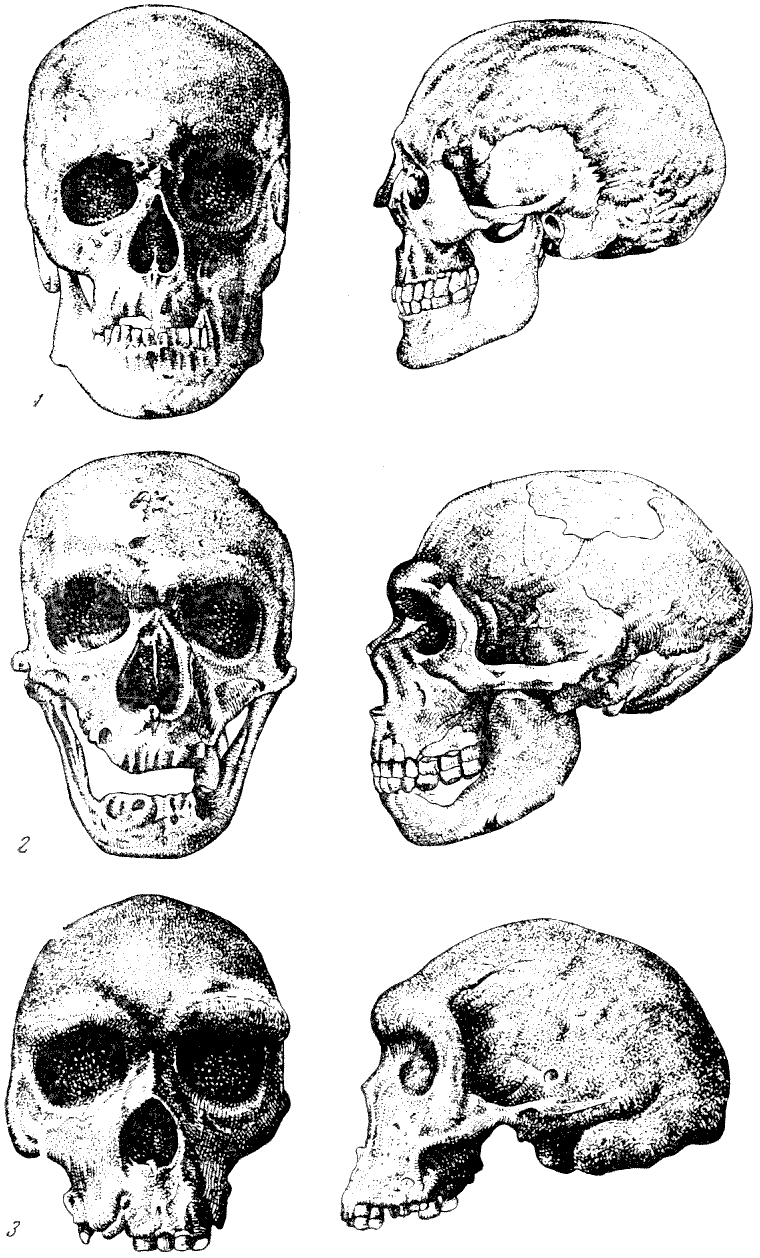
Рис. 124. Черепа гоминид
1 — современный человек; 2 — неандерталец из Ла Шапелль-о-Сен; 3 —
неандерталец родезийский из Брокен-Хилла. Фас и профиль (профиль 2 по
Дж. Г. Мак-Грегору), 2/9 нат. вел. (ширина изображений, соответственно,
33,5 мм и 45 мм; 36,5 мм и 49 мм; 37 мм и 49,5 мм)
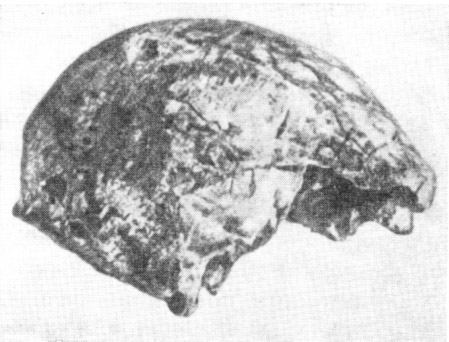
Рис. 125. Череп неандертальца (явантропа) из Нгандонга
По В. Оппенурсу, 1937, 1/3 нат. вел. (ширина изображения 57 мм)
|
с теменными при пересечении лобного шва со стреловидным. Зато с современным
человеком его сближает хорошая выраженность сосцевидного отростка
и значительная глубина сочленовной ямки, куда входит соответствующий отросток
нижней
челюсти.
Древность яванских черепов довольно значительна. По Оппенурсу и Кизсу,
явантроп жил немногим позднее питекантропа, для которого эти авторы
принимают позднеплиоценовый
возраст. Но ближе к истине мнение Осборна, по которому яванский человек
жил около 150 тыс. лет назад, в третью межледниковую (рисс-вюрмскую)
эпоху.
На,основании изучения найденных черепов Оппенурс рисует такую картину
эволюции древних людей: яванский человек был предком для родезийцев
и австралийцев,
синантропы же дали начало гейдельбергскому человеку и через него европейским
неандертальцам.
В этих построениях Оппенурса неубедительно выведение территориально столь
отдаленной формы, как родезиец от явантропа, а также гейдельбергского человека
от синантропа.
Путешествие синантропов в Европу не могло иметь места. Скорее можно
предполагать, что в близкой к Европе области, недалеко от границы между
Азией и Африкой,
одновременно с синантропами развивались сходные с ними группы древних
людей, из которых произошли некоторые более поздние формы, например
гейдельбергский человек, европейские неандертальцы, палестинские неандертальцы
и родезиец.
Вейнерт, как и Оппенурс, считает, что нгандонгский человек — это прямой
потомок яванского питекантропа и предок австралийцев. Это мнение
неверно, так как
австралийцы прибыли на свой материк из юго-восточной Азии и притом
сравнительно недавно, несколько десятков тысяч лет назад. В советской
антропологии
после исследования Я. Я. Рогинского (1949) гипотезе полицентризма
в проблеме
происхождения типа современного человека противопоставляется гипотеза
широкого моноцентризма,
основанная на теснейшей генетической близости строения тела в разных
группах современных людей, в частности их черепа.
Находки неандертальцев
в СССР
Летом 1938 г. археолог А. П. Окладников, производя раскопки грота Тешик-Таш
близ г. Байсуна в южном Узбекистане (рис. 126),
322

Рис. 126. Ущелье Заутолош-сай: участок вблизи грота Тешик-Таш
По
А. П. Окладникову, 1949
обнаружил каменные орудия мустьерской эпохи и неполный скелет ребенка
8—9 лет (рис. 127— 130). Там же он нашел остатки кострищ, а также
многочисленные кости и рога горных козлов теке, на которых, очевидно,
охотились тешикташские
неандертальцы (Окладников, 1940).
А. П. Окладников передал костные остатки ребенка неандертальца в
Институт антропологии (Москва), где их подвергли изучению. Череп
реконструирован
из ста пятидесяти обломков. Эту трудную работу выполнил археолог-скульптор
М.
М. Герасимов,
который реконструировал и внешний облик ребенка неандертальца (рис.
131—133).
Г. Ф. Дебец (1940) первоначально исследовал череп. Вместимость мозговой
коробки очень большая — 1490 сма. Если бы этот неандерталец достиг
взрослого состояния,
то его мозг по величине вряд ли уступал бы мозгу неандертальца
из Ла Шапелль-о-Сен (1600 см3). По описанию
эндокрана тешикташского мальчика
(Бунак, 1951),
в нем отмечаются некоторые черты прогрессивности, перехода к типу
строения мозга современного человека. На лбу
Рис. 127. Череп ребенка неандертальца и рога горного козла на дне
грота Тешик-Таш в начале расчистки
По А. П. Окладникову, 1949
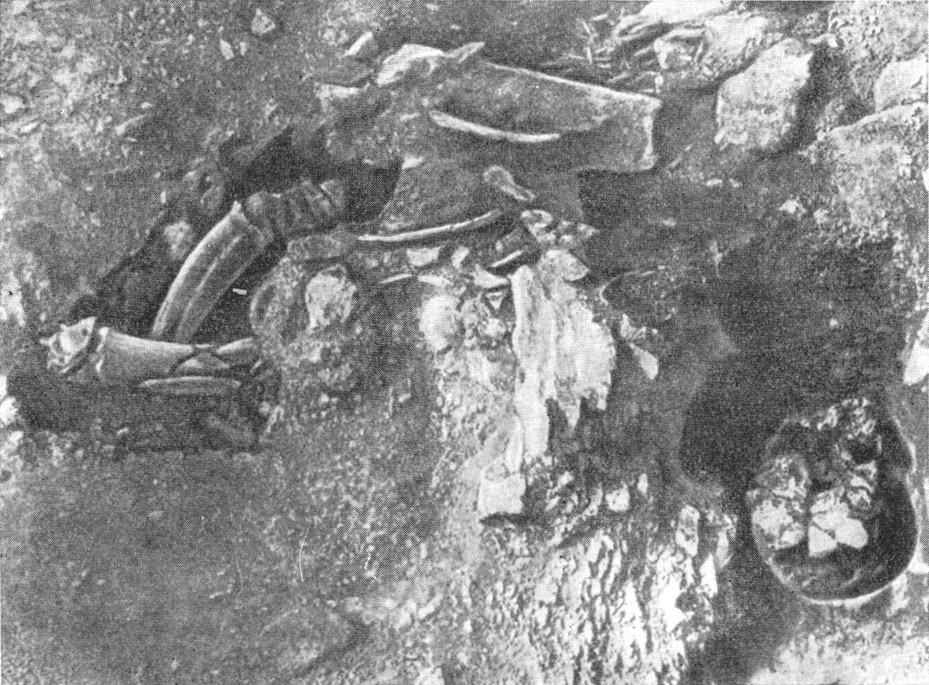
324

Рис. 128. Череп ребенка неандертальца в непотревоженном состоянии
на возвышенном участке своего местонахождения — останце; впереди
кости
скелета
По А. П. Окладникову, 1949
Рис. 129. Череп ребенка неандертальца
на месте находки в гроте Тешик-Таш; вокруг черепа порода расчищена
По А. П. Окладникову, 1949
заметен сплошной, но еще относительно слабо развитый надглазничный
валик.
Подбородочный выступ отсутствует. Все это характерно для неандертальца,
но зубы тешикташского ребенка обладают малой полостью, т. е.
относятся к цинодонтному
типу, как у большинства современных людей (Гремяцкий, 1948).
Находка тешикташского ребенка имеет мировое значение. Она свидетельствует
о том, что неандертальцы жили около ста тысяч лет назад в глубине
материка Азии в условиях среды, близких к современным. Там
не было заметно влияния
ледникового покрова, столь изменившего области Северной Азии.
Этому влиянию зарубежные ученые, как Ганс Вейнерт, приписывают решающую
роль в формировании
современного человека из неандертальца. Но тешикташская находка
противоречит такому
мнению, которое несостоятельно прежде всего потому, что противоречит
основному тезису
о решающей роли общественного труда в антропогенезе.
Костные остатки другого очень древнего человека были обнаружены
в Крыму раньше. В 1924 г. в гроте Киик-Коба, в 25 км к
востоку от Симферополя,
Г. А. Бонч-Осмоловский
(1940, 1941, 1954) обнаружил остатки неандерталь-
326
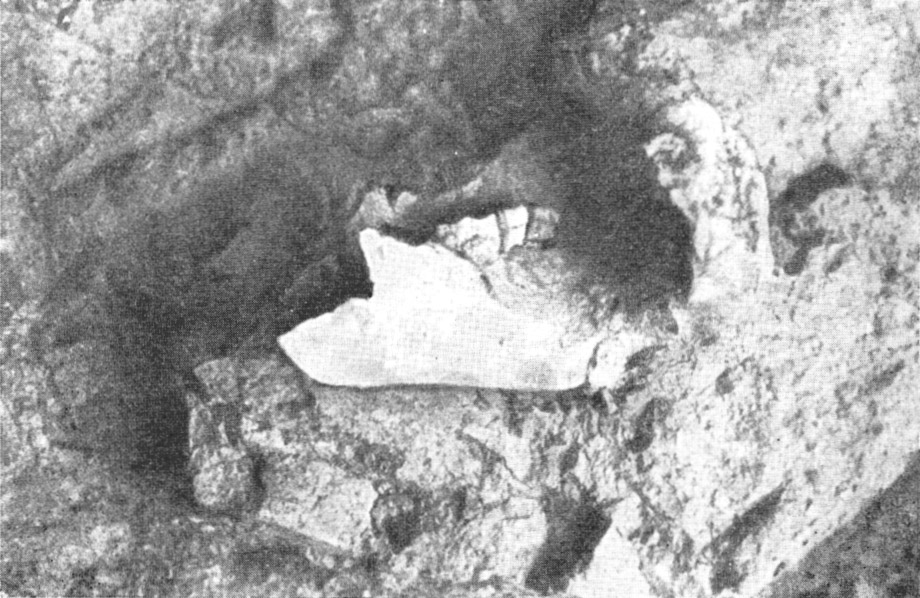
Рис. 130. Нижняя челюсть от черепа ребенка неандертальца в
гроте Тешик-Таш (на месте нахождения, в процессе расчистки)
По А. П. Окладникову, 1949
Рис. 131. Череп ребенка неандертальца из грота Тешик-Таш,
реставрированный М. М. Герасимовым
Вид справа (1) и спереди (2). По А. М. Гремяцкому, 1949, 2/7
нат. вел. (ширина изображения 119 мм)
Рис. 132. Слепки мозговой полости черепов
1 — ребенок неандертальца из грота Тешик-Таш; 2 — ребенок современного
человека 8 лет
По В. В. Бунаку, 1954, 2/7 нат. вел. (ширина изображения 119 мм)
Рис. 133. Ребенок неандертальца из грота Тешик-Таш (реконструкция
М. М. Герасимова)
По М. М. Герасимову, 1949
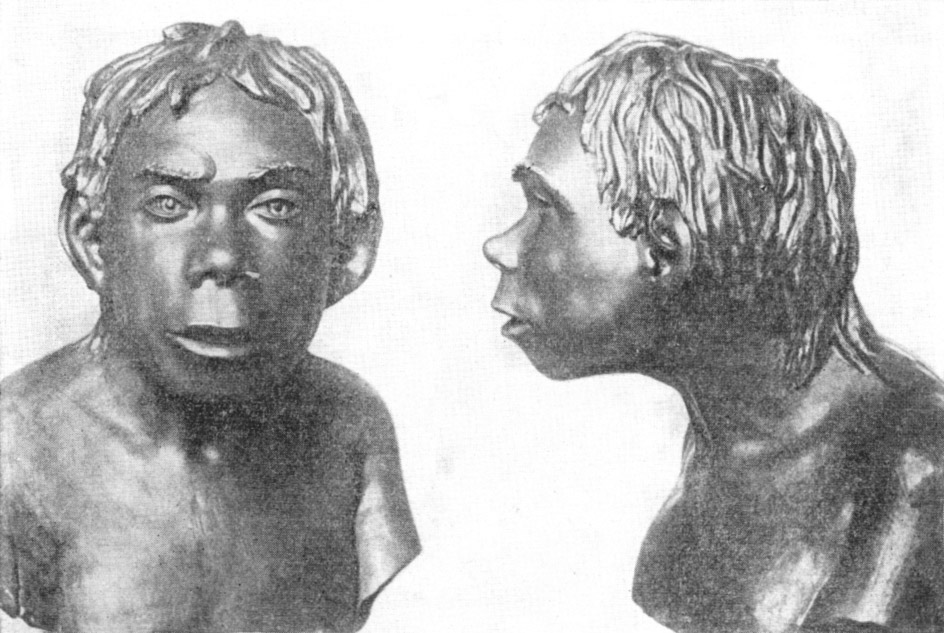
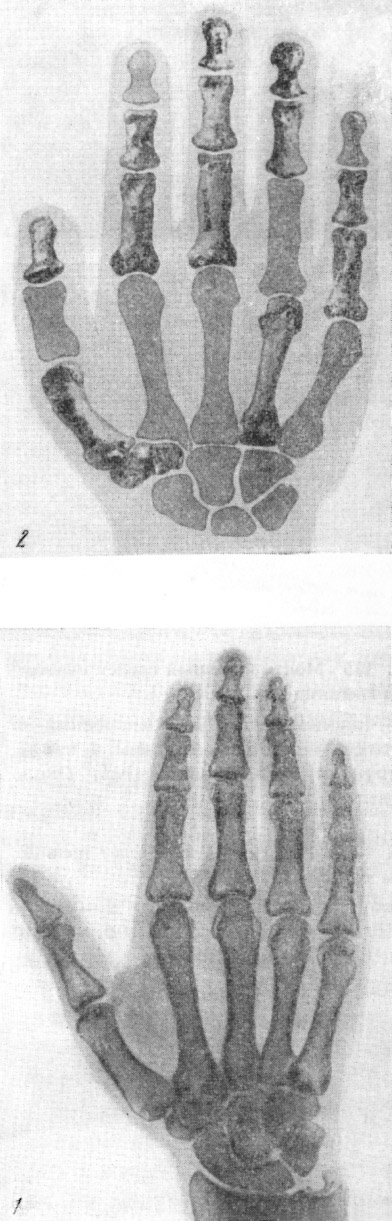
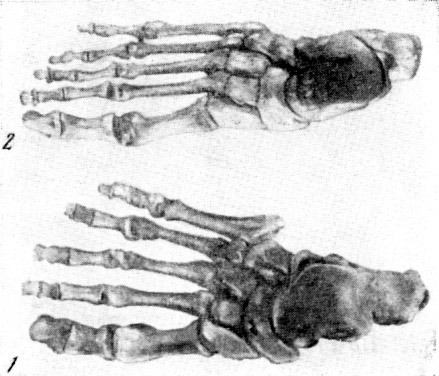
Рис. 134. Скелет правой кисти гоминид
1 — кииккобинский человек (реконструкция
2 — современный человек, тыльная поверхность. По Г. А. Бонч-Осмоловскому,
1941, 1/3 нат. вел. (ширина изображения 50 мм)
|
цев: кости стопы, голени и кисти взрослого (рис. 134—135)
и неполный скелет годовалого младенца. Там же, в разных
слоях дна пещеры,
были найдены тысячи
каменных орудий, которые, по мнению Бонч-Осмоловского,
относятся к аморфной и позднеашёльской стадиям культуры
раннего палеолита.
Обнаружены
и костяные
орудия, например ретушеры (наковаленки). Открытие и
описание находок в гроте Киик-Коба составляют существенный вклад
в палеантропологию.
Большой интерес представляют кости конечностей кииккобинского
человека. Г. А. Бонч-Осмоловский, судя по особенностям
скелета конечностей,
полагает, что кииккобинцы обладали еще далеко не столь
высоко развитыми кистью
(рис. 136) и стопой, как современный человек. Вместе
с тем он, как и В. В. Бунак,
считает, что строение конечностей кииккобинца противоречит
мнению о происхождени[и] человека от предков, живших
некогда на деревьях.
В пользу
такого мнения
приводятся
разные факты, в том числе черты сходства кисти кииккобинца
с кистью человеческого плода. Однако эти утверждения
Г. А. Бонч-Осмоловского
недостаточно обоснованы.
Так, уже самое определение геологического возраста нижнего
и верхнего слоев грота с их культурными остатками,
как шелльского и ашёльского,
встречает
возражения со стороны некоторых археогов и палеонтологов
(Бадер, 1940). Что же касается отрицания Г. А. Бонч-Осмоловским
и В.
В. Бунаком (1954)
древесной стадии в эволюции наших обезьяноподобных
предков, то
в пользу призна-
329
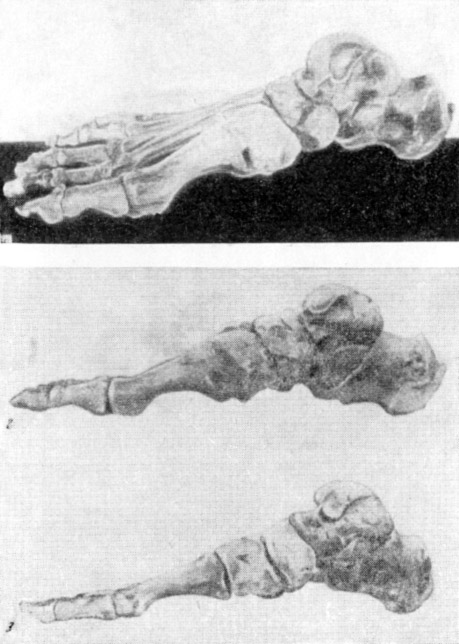
Рис. 135. Монтированный скелет правой стопы гоминид
1, 2 — кииккобинец, 3 — современный человек; вид с медиальной стороны. 1
и 2 — реконструкция В. В. Бунака, 1954. Около 2/3 нат. вел. (ширина изображения
58 мм)
Рис. 136. Монтированный скелет правой стопы гоминид
1 — кииккобинский человек; 2 — современный, тыльная поверхность. По В. В. Бунаку,
1954, 1/3 нат. вел. (ширина изображения 56 мм)
|
ния этой стадии, установленной еще Дарвином, есть слишком
много доказательств, чтобы можно было бы ее отвергнуть
на основании
далеко не достаточных
фактов.
Здесь стоит напомнить о наличии у человека поперечной
головки мышцы, приводящей большой палец стопы. Она
характерна для
обезьян в отличие
от прочих млекопитающих.
Эта особенность убедительно свидетельствует, что в
нашей родословной были древесные формы с обезьяньим
типом строения
тела. Фактов,
подтверждающих это мнение, можно привести множество.
Но, вероятно, надо считать,
что прямохождение
у предков человека начало развиваться в связи с круриацией
раньше, чем это полагали до сих пор.
Палестинские неандертальцы
К ископаемым людям, в строении тела которых можно усматривать
переходные черты к типу современного человека, принадлежат
палестинские неандертальцы.
С 1931 по 1936 г. в Палестине, близ городов Хайфы и
Атлита, в пещерах Схул и Табун на склонах горы Кармел,
были найдены
костные
остатки
почти от двух
десятков особей неандертальцев, в том числе скелет
ребенка (Рогинский, 1966б; Коробков, 1963).
В пещере Схул было обнаружено несколько скелетов, что
указывает, по-видимому, на коллективное погребение.
Большинство скелетов
найдено в 1932 г.
американским ученым Теодором Мак-Коу-ном, который в
августе того же
330
Рис. 137. Череп неандертальца
1 — череп из пещеры Схул V с горы Кармел;
2 — его реконструкция. По Т. Мак-Коуну и А. Кизсу, 1939. Около 1/5 (1)
и 1/3 (2) нат. вел. (ширина изображений, соответственно, 58
мм и 62 мм)
|
года демонстрировал некоторые находки на Всемирном
конгрессе археологов в Лондоне.
Огромный интерес к этим находкам вполне оправдывается
их количеством. Кроме того, исследования показали,
что кармельские
скелеты
обнаруживают также
черты, дающие понятие о переходе неандертальцев к современному
типу человека.
Признаки кармельских людей ярко выражены на скелетах
взрослых из пещеры Схул. Рост их велик, бедренные кости,
как правило,
прямые.
Череп (рис.
137) отличается
выраженным надглазничным валиком, который Мак-Коун
даже называет зонтикообразным. Нижняя челюсть очень
тяжела,
мощна и широка.
Однако в отличие от типичных неандертальцев на некоторых
нижних челюстях отмечается подбородочный выступ, что
сближает кармельцев
с современным
человеком. За правомерность такого сближения свидетельствуют
более высокий и округлый
свод черепа, а также более развитые теменные и лобные
части, чем у типичного неандертальца. Зато зубы сходны
с неандертальскими.
Однако коренные зубы
обладают небольшой полостью, следовательно, они не
тавродонтного типа,
если не считать некоторой сближенности корней. Судя
по черепу, к нему прикреплялась мощная мускулатура.
Поперек
затылка
идет костный
гребень
— место прикрепления
шейных мышц.
Большинство скелетов кармельцев, в том числе и скелет
ребенка, оказались в скорченном положении. Были также
найдены каменные
орудия, судя
по кото-
331
рым скелеты должны быть отнесены к эпохе мустье.
Мак-Коун вместе с Кизсом в специальной монографии (McCown,
Keith, 1939) дали весьма обстоятельное описание скелетов,
отдельных костей и черепов.
Археологическую
часть находок описали Д. Гаррод и Д. Бейт (1937).
Интересующие нас остатки скелетов происходят преимущественно
из небольшой пещеры Схул, но часть их обнаружена в
крупной пещере Табун. Они найдены
и извлечены начальником экспедиции Дороти Гаррод.
В пещере Схул, в твердой, как камень, брекчии, были
найдены: почти полный скелет ребенка, по-видимому девочки,
около
4 1/2 лет; части
четырех скелетов
двух мужчин 30—35 и 50 лет, женщины 35—50 лет и ребенка,
вероятно мальчика, 8—10 лет; фрагменты черепа, зубы
и отдельные кости
взрослого мужчины,
женщины 30—40 лет, ребенка, вероятно мальчика, 5—5
1/2 лет, а также 16 отдельных
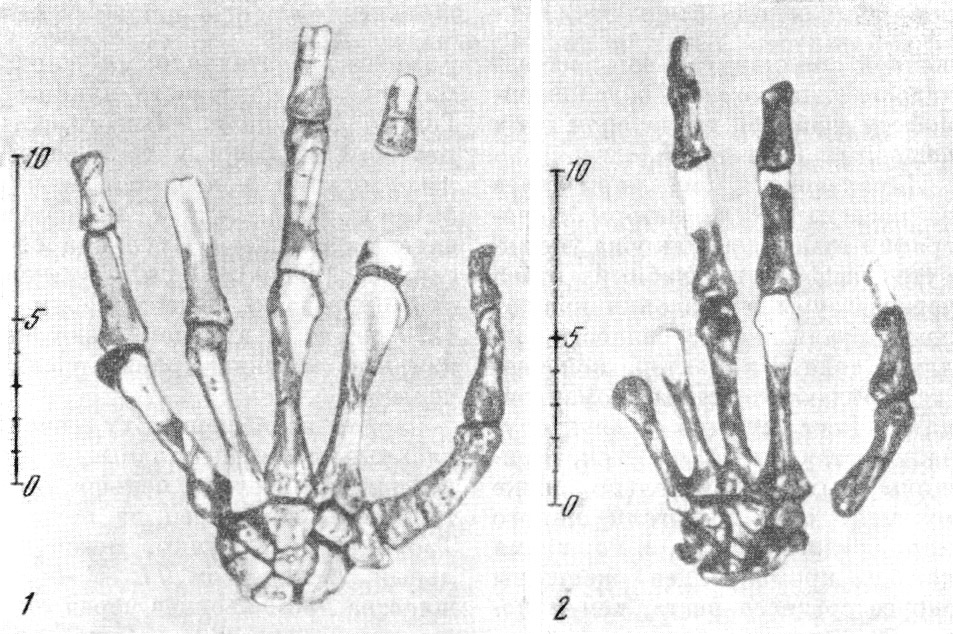
костей (рис. 138—139).
В пещере Табун из породы были извлечены: почти полный
скелет женщины в возрасте около 30 лет, почти целая
нижняя челюсть
мужчины около
30—35 лет,
с зубами
в ячейках, три серии костей (среди них диафиз правого
бедра) и зубов (среди них первый и второй нижние моляры)
из верхнего
слоя
шелльской
эпохи. Таким
образом, в обеих пещерах сохранились кости 23 особей
разного возраста и пола. Черепа хорошо сохранились
на трех скелетах
из пещеры Схул,
равно как и от
женского скелета из пещеры Табун (рис. 140). Извлечение
костей из породы, закрепление
их особыми связывающими веществами, приведение различных
частей того или иного скелета в соответствие одной
с другой, сличение
с другими
костными остатками, измерения, описание, анализ и сравнение
их между собой, а
также
с другими находками неандертальцев и людей современного
типа, — все произведено с большой тщательностью и позволяет
составить
отчетливое
суждение об
этом крупном открытии.
Среди указанных находок несколько особое место занимает
женский скелет из пещеры Табун, череп которого во многом
носит явно
неандертальские черты и
обладает значительным общим сходством с женским черепом
Гибралтар I (находка 1848 г.). В 1925 г. примерно в
85 км к северо-западу
от
горы
Кармел,
в пещере эль-Зуттие, в Галилее, найден фрагмент человеческого
черепа (рис.
141), который,
по мнению Мак-Коуна и Кизса, относится к той же группе,
что и кармельцы.
Остатки скелетов из пещеры, Схул по форме и строению
представляют такое смешение черт неандертальцев и кроманьонцев,
какое
до сих пор еще не
удавалось подметить.
Однако оба типа, Схул и Табун, относятся почти к одной
и той же фазе, леваллуа-мустьерской культурной эпохи
(сопровождающая их
фауна в
общем сходна, хотя в пещере
Схул найдено больше костей быков, а в пещере Табун
чаще встречаются
кости газелей).
Надо думать, что палестинцы, обнаруженные в пещерах
горы Кармел,
представляют все же единую группу с сильно выра-
332
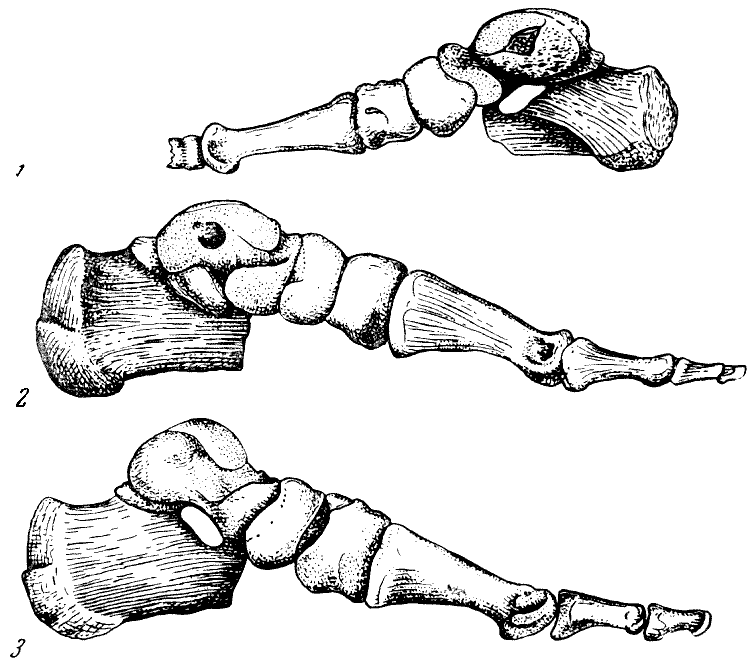
Рис. 138. Скелет стопы неандертальцев
1 — Табун I; 2 — Схул IV; 3 — современный человек.
По Т. Мак-Коуну и А. Кизсу, 1939. Около 1/3 нат. вел. (ширина изображений
66 мм, 91 мм, 88 мм)
женной индивидуальной изменчивостью, возможно, обусловленной
смешанным характером этих пещерных обитателей.
Характеризуя тип кармельцев из пещеры Схул, авторы
монографии отмечают, как уже упомянуто выше, что
лобный отдел черепа,
зубы и позвонки
являют неандертальские
особенности, но длина тела и характер конечностей
придают сходство с кроманьонцами. Рост мужчин в двух случаях
составлял 170 и
178 см. Женщины
были значительно
ниже мужчин: они достигали малого или среднего роста,
в то время как у кроманьонцев женщины скорее среднего
роста,
чем
малого.
Позвоночник обладал
некоторыми неандерталоидными чертами, нижние конечности
длинные. Голова крупная. Вместимость мозговой коробки
у трех
взрослых мужчин
колеблется между
1518 и 1587 см3. У женского
черепа из пещеры Схул она составляет 1300—1350 см3,
а у женского черепа из пещеры Табун — 1270 см3,
т. е. в общем такая же, как и у женских кроманьонских
черепов.
Черепа из пещеры Схул очень длинные, долихоцефальные,
с индексом значительно меньше 75, в то время как
череп из
пещеры Табун
средней длины, мезоцефальный,
с индексом 77. У большинства кроманьонцев череп имеет
удлиненную, долихоцефальную
333
форму. Черепа из пещеры Схул имеют свод средней высоты
и в этом отношении занимают промежуточное положение
(о новой
реконструкции
черепа Схул
V см. в работе: Snow, 1953).
Судя по слепку мозговой полости, головной мозг вполне
сходен с мозгом кроманьонцев по форме и близок по
общим размерам,
но рисунок
извилин
и борозд несколько
проще.
Лоб умеренно выпуклый, глазницы широкие, но не высокие
(у большинства кроманьонцев они очень низкие). Имеется
надглазничный
костный
валик, на котором намечается
разделение на среднюю и боковые части. У кроманьонцев
это разделение уже завершилось.
Лицевой отдел черепа у большинства кармельцев из
пещеры Схул сравнительно мало выступает, лицо ортогнатное,
прямое, умеренно
длинное, как
у кроманьонцев. Однако у черепа Схул V прогнатизм
выражен
весьма резко.
Клыковые ямки
на верхней челюсти отсутствуют, верхний край скуловой
кости утолщен, носовой
отдел варьирует как по степени выступания носовых
косточек, так и по ширине грушевидного отверстия.
Обратимся теперь к нижней челюсти. Она варьирует
по величине и по массивности: найдены и крупные и
малые
челюсти, притом
малые относительно
массивные,
а крупные относительно тонкие, не массивные. Ширина
восходящей
Рис. 139. Скелет левой кисти неандертальцев
1 — Табун I; 2 — Схул V; тыльная сторона. По Т. Мак-Коуну
и А. Кизсу, 1939, 3/7 нат. вел. (ширина изображения 122 мм)
334
Рис. 140. Женский череп из пещеры Табун (реконструкция)
По Т. Мак-Коуну и А. Кизсу, 1939, 2/5 нат. вел. (ширина изображения 59
мм)
Рис. 141. Фрагмент черепа неандертальца из пещеры
эль-Зуттие, на северо-западном берегу Геннисаретского озера, Галилея
По Т. Моллисону, 1932. Около 1/2 нат. вел. (ширина изображения 56 мм)
|
ветви очень большая, как у неандертальцев и некоторых
кроманьонцев. Угол нижней челюсти у одних умеренно
развит, как и у некоторых
кроманьонцев, но у других развит хорошо. Подбородочный
выступ отсутствует или умеренно
развит,
а у кроманьонцев он развит умеренно или хорошо. Наконец,
зубы у кармельцев умеренно крупные либо, как и у
неандертальцев, крупные (у кроманьонцев
они умеренно крупные). Узор на жевательной поверхности
коренных зубов удерживает больше примитивных особенностей,
чем у кроманьонцев.
В итоге произведенного анализа по главным признакам
сравнения Мак-Коун и Кизс нашли, что обитатели
из пещеры Схул обладают
лишь тремя
существенными признаками, общими с неандертальцами:
надглазничным валиком, плоской
формой скуловой кости и узором зубов. В то же время
есть 8 признаков, общих с
кроманьонскими,
12 носят промежуточный характер. Когда же авторы
добавили еще 86 признаков более мелкого значения,
то в итоге
из 113 общих
с неандертальцами
оказалось
16 признаков, с кроманьонцами (и даже шире с неантропами)
— 32, промежуточного характера — 48, неопределенных
— 13 и имеющих
характер
черт специализации
— 4.
Таким образом, налицо известный и довольно значительный
уклон в сторону типа современного человека. Все же,
придавая особый
вес
надглазничным
. костным
разрастаниям и другим особенностям строения черепа,
зубов и скелета, Мак-Коун и Кизс классифицируют кармельцев
как
335
входящих в группу неандертальцев, но образующих связующее
звено между последними и людьми современного типа.
Однако Мак-Коун и Кизс понимают эту связь не в
филогенетическом смысле, так как полагают, что
неандертальцы и современные
люди суть потомки
одного общего
предка — питекантропа. С нашей точки зрения, кармельцы
— это, несомненно, эволюционная ступень в развитии
неандертальского типа в кроманьонский
(в широком смысле этого понятия).
Серьезное анатомическое отличие неандертальцев
выражается в отсутствии у них подбородочного выступа.
Но именно
у кармельцев мы видим
переход от отсутствия
выступа до его наличия: при таком смешении признаков
современного человека с неандертальскими этих людей
надо считать за
переходную форму. Так
как черты современного человека у них даже преобладают,
то некоторыми исследователями
было высказано мнение, по которому кармельцы —
первые истинные люди, самые древнейшие представители
вида
человека разумного.
Во всяком случае находка кармельцев — одно из
наиболее выдающихся событий в области антропогенеза
за последние
десятилетия.
Она равноценна находке
неандертальцев на острове Ява или даже открытию
китайского древнейшего человека — синантропа. Знаменательно
и то, что эта переходная
форма найдена в Палестине
— на стыке материков Азии, Европы и Африки, причем
она имеет черты сходства не только с неандертальцами,
но и с кроманьонцами Европы.
Другие замечательные находки древних людей с некоторыми
переходными чертами сделаны в 1951—1960гг. в Загросских
горах иракского
Курдистана в пещере
Шанидар американским антропологом Р. Солецким (Solecki,
1955, 1961). Наибольшее внимание из остатков семи
скелетов привлекают
мужские
черепа I и II (рис.
142), реставрированные Т. Д. Стюартом (Stewart,
1959,1961). На них (Коробков, 1963) заметна смесь
неандертальских
черт (вроде
выраженного
надглазничного
валика) со свойственными современному человеку
(например, сильноразвитые сосцевидные отростки
и намечающийся
подбородочный выступ),
а также со специфическими для неандертальцев особенностями
нижней
челюсти
и носовой
области.
Обнаружение костных остатков неандертальцев и близких
к ним форм на столь широкой территории Старого
Света дает
основание
полагать,
что
неандертальцы
были предками современного человека. Трудно предположить,
чтобы могло бесследно вымереть многочисленное население
неандертальцев, орды
которых с успехом
боролись за свое существование в тяжелых условиях
похолодания, охотились на животных, поддерживали
огонь, жили прочными
первобытными
стадами.
По своему физическому типу неандертальцы в большинстве
вполне могли быть предшественниками и предками
современного человечества.
Да
и трудно предположить
иной путь происхождения типа современного челове-
336
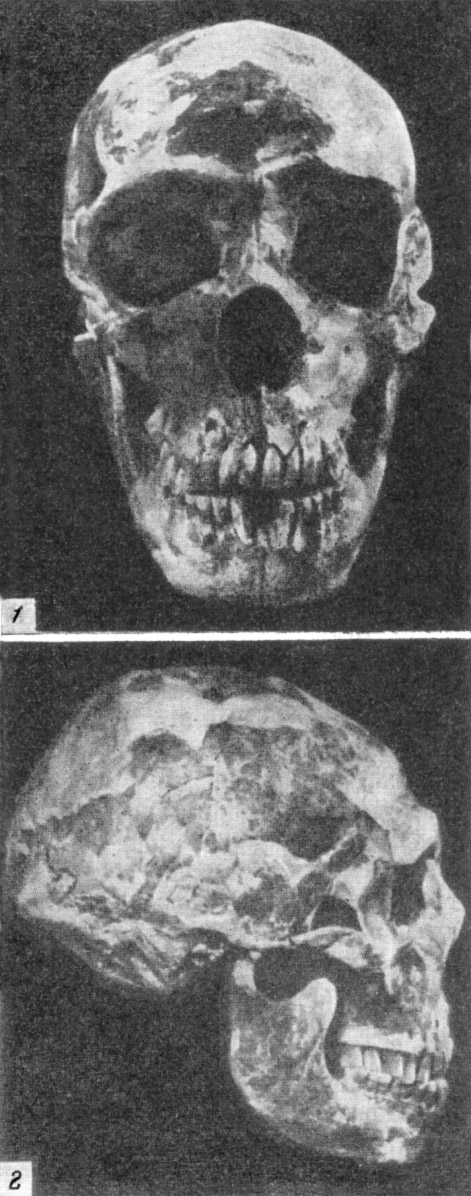
Рис. 142. Ископаемый человек
Неандерталец, Шанидар I. Череп, вид спереди и справа
1 — около 2/5 нат. вел.; 2 — 1/3 нат. вел. По Р. Стюарту,
1959 (ширина изображения 60 мм)
|
ка. Надо было бы тогда думать, что он зародился,
например, в небольшой области Азии, откуда стремительно
распространился
по
всей территории
Старого Света,
но никто не поддерживает этой теории миграций древнего
человека.
Новейшие работы все более и более ясно подтверждают
обоснованную в 1927 г. прогрессивным американским
антропологом Алешом
Грдличкой гипотезу
о развитии
кроманьонцев из неандертальцев.
Эту гипотезу подтверждают и некоторые другие последние
находки неандертальцев в Европе и Азии. Так, в
1933 г. у Штейнгей-ма,
близ г. Штутгарта
(Германия), был найден неандертальский женский
череп с малым объемом мозговой коробки
и сильно развитым надглазничным валиком. Затылочное
отверстие располагается почти так же, как у современного
человека.
Верхний край чешуи височной
кости не идет горизонтально, как обычно у неандертальцев,
обезьянолюдей и обезьян,
а округлый, как у современного человека. (О находках
неандертальцев в Чехословакии см. монографию Э.
Влчека — Vlcek, 1969.)
М. А. Гремяцкий (1948) предложил делить неандертальцев
на более генерализованные формы, как исходные для
современного человека,
и более специализированные,
«классические» западноевропейские формы (рис. 143).
На этом
основании, а также исходя из других неандертальских
находок, более ярко
свидетельствующих о возникновении типа современного
человека из типа человека первобытного,
в последнее вре-
337

Рис. 143. Лицо неандертальца в фас и в профиль
Реконструкция М. М. Герасимова, 1948
мя появляются новые соображения о необходимости
дифференцированного анализа обширной и разнообразной
группы ближайших
предшественников человека разумного.
Обратимся к новейшему разбору групп неандертальцев,
или палеоантропов (Алексеев, 1966). К более генерализованному
типу принадлежат
черепа из находок в Штейнгейме,
Эрингсдорфе, Саккопасторе и Крапине, а также
в Сванскомбе и Фонтешеваде. Набор их морфологических
особенностей
носит
более
прогрессивный
характер, чем в составе черепов неандертальцев
из Ла-Феррасси, Ла-Кина, Ла-Шапелль,
Спи и Мон-те-Чирчео. Эта вторая группа носит
явные черты значительной специализации, и мнение о ее
возможности превращения
в тип
современного человека встречает
гораздо больше препятствий, чем для более древней
прогрессивной группы, которая имела достаточно
времени, чтобы превратиться
в Homo sapiens,
дать начало
неоантропам. Подобные лашапелльскому черепа
относятся к вюрмской эпохе, а близкие к эрингсдорфскому
— к рисс-вюрмской.
Такое подразделение неандертальцев на две группы
предлагалось и другими советскими антропологами
(Гремяцкий, 1948;
Якимов, 1949),
однако
в отличие от них В.
П. Алексеев допускает возможность преобразования
и более специализированной группы палеоантропов
в вид
современного
человека.
В общем если учитывать характер каменной индустрии
ашёльской и мустьерской эпох и широту географического
распространения
неандертальцев по стра-
338
нам Европы, Африки и Азии (Рогинский, 1966), то
палеоантропов, или древних людей неандертальского
облика, вполне можно
считать группой
в целом промежуточной
между древнейшими людьми и современными (Гохман,
1966). Другими словами, оправдывается мнение
Алеша Грдлички
о неандертальской
стадии в ходе
развития собственно гоминид, в то время как гипотеза
«пресапиенса» А. Валлуа (Vallois,
1954) лишается подтверждений (Войно, 1959). Особой
точки зрения держится Б. Ф. Поршнев (1969). По
его мнению, некоторые
палеоантропы,
лишившись
прежней культуры, все-таки смогли сохраниться
на разных материках в качестве реликтов.
Правда, до сих пор реальные остатки от них или
их представители в руки специалистов еще не поступали,
так что вопрос
остается открытым.
Морфологические особенности ископаемых представителей
человечества говорят в пользу того, что оно прошло
три ступени формирования
физического типа.
По нашему мнению, в семействе гоминид (Hominidae)
следует вообще различать только один род людей
(Homo), который
представлен тремя подродами,
соответственно ступеням становления человека
(Нестурх, 1941; см., однако, статью Г.
Ф. Дебеца, 1948а, б).
Первый подрод обезьянолюдей, или питекантропов
(Pithecanthropus), включает в свой состав две
четко выраженные и достаточно
полно представленные формы древнейших людей.
К подроду обезьянолюдей относятся два вида:
1) яванский,или тринильский, обезьяночеловек,
т. е. питекантроп прямоходящий — Homo (Pithecanthropus)
erectus;
2) китайский питекантроп, или пекинский синантроп
— Homo (Pithecanthropus) pekinensis.
Второй подрод образуют древние люди, или палеоантропы
(Раlaeoanthropus), группирующиеся вокруг типа
неандертальца. Сюда относятся,
например, западноевропейские неандертальцы, родезиец,
палестинские неандертальцы,
древний человек
явантроп, всего несколько видов (или подвидов).
Третий подрод — новые люди, или неоантропы (Neoanthropus),
представленные ископаемыми людьми современного
типа, в том числе кроманьонцами,
и современными. Все они составляют один вид «разумный
человек» — Homo
(Neoanthropus)
sapiens.
Образ жизни древних людей
Многие неандертальцы обитали в пещерах, которые
защищали их от холода во время ледниковой эпохи.
Вместе с
остатками неандертальцев
ученые
нередко находили кости пещерных медведей, тигров,
львов, гиен. Это свидетельствует
о том, что человек вел с хищными животными борьбу
из-за жилища. Остатки других зверей, в том числе
и таких
крупных, как мамонт
и носорог,
свидетельствуют об интенсивности охоты, которая
особенно развивается во времена мустье.
Более
древние люди добывали пищу еще в значительной
мере собиранием плодов и
корней, поимкой мелких животных.
Неандертальцы мустьерской эпохи охотились не
только на
339
открытых пространствах, но и в лесах, преследуя
преимущественно животных средних размеров. Нередко
они нападали сообща
и на более крупных
млекопитающих, иногда на беззащитных, больных,
попавших в яму или болото, поедали
и их трупы.
Убив животное, неандертальцы разрезали кожу каменными
орудиями, снимали мясо с костей, разбивали длинные
кости и доставали
из них питательный
костный мозг, а из черепа извлекали головной
мозг. Мясо ели сырым или предварительно
жарили его на кострах. Вероятно, неандертальцы
использовали шкуры животных для укрытия тела
и на подстилку (Ефименко,
1953; Осборн,
1924).
В эпоху мустье значительно усложнялась техника
и ведение хозяйства, вызванные охотой. Происходило
дальнейшее
разделение труда и
выделение наиболее
опытных охотников как руководящих сочленов первобытного
стада. Хотя европейские неандертальцы были удовлетворительно
приспособлены
к
жизни, даже в тяжелых
условиях мустье,
но весьма вероятно, что разные болезни и трудности
борьбы за жизнь сильно понижали их долговечность
(Рохлин, 1965).
Вообще жизнь первобытных людей была полна лишений
и опасностей. Оценку той отдаленной эпохи
формирования человечества
мы находим в словах
В. И. Ленина:
«Что первобытный человек получал необходимое
как свободный
подарок природы,— это глупая побасёнка... Никакого
золотого века позади
нас не было, и
первобытный человек был совершенно подавлен трудностью
существования, трудностью борьбы с природой» (Полное собрание сочинений,
т. 5, стр. 103).
Каменные орудия неандертальцев (рис. 144) были
уже более разнообразны, кроме того, они лучше
обрабатывались, чем
в предшествующие
культурные эпохи. Так,
начало нижнего палеолита характеризуется дошелльской
культурой с ее более простыми орудиями, чем ручные
ударники, или
рубила, характерные
для более
поздней шелльской культуры.
Ручные рубила шелльской культуры получались путем
обивки каменного ядрища с таким расчетом, чтобы
один конец
его мог служить ударным,
колющим и
режущим инструментом, а другой конец было удобно
держать рукой, сжатой в кулак
(рис. 145). Кроме ручного рубила, из эпохи шелль
известны и другие формы орудий.
Культуре ашёль свойственны более симметричные
ручные рубила с обивкой по всей поверхности,
следовательно,
тогда
возникают приемы
новой техники.
Тут встречаются и орудия, сделанные из осколков,
сбитых с ядрища; они характерны также для еще
более поздней
— мустьерской
культуры
палеолита.
В эпоху мустье наиболее типичны скребла и остроконечники,
вырабатывавшиеся из отщепов, а не из самого кремневого
ядрища. Техника изготовления
орудий в эпоху мустье, судя по их выделке в европейских
местонахождениях, заметно
изменилась по сравнению с ангельской (С. А. Семенов,
1968).
340

Рис. 144. Каменные орудия среднего палеолита
1 — мустьерское скребло, Франция; 2, 3, 4 —
два орудия типа остроконечников из верхнего сло[я] грота
Киик-Коба, Крым;
5 — мустьерский остроконечник,
Франция; 6 — мустьерский дисковидный
нуклеус из кварцита со стоянки на р. Деркул,
Донбасс, вверху показан способ откалывания
пластин от этого ядрища; 7 — кремневое
рубило ашёльского
типа (Англия);
8 — шелльское
кремневое рубило
раннего типа
(Бельгия). По Г. Мортилье, 1910 1, 5)
и П. П. Ефименко, 1953 (2—4, 6—8).
Размеры орудий: 1 — 70 мм; 2, 3, 4 — нат. вел.;
5 — 74 мм; 6 — 1/3 нат. вел., 7 — 1/3, 8 —
около 1/5
нат.
вел. (высота изображений: 2 - 30 мм; 3 - 30,5 мм; 4 - 32 мм; 6 - 32 мм; 7
- 45 мм; 8 - 40 мм)
Форма и величина древних орудий нередко дают
возможность со значительной степенью точности
судить о способе
их употребления. Орудия находят
иногда в больших количествах вместе с разбитыми
вдоль и поперек костями животных,
с остатками кострищ, с человеческими костями.
Орудия древних людей и другие признаки их деятельности
позволяют сделать
важные
заключения
об
образе
жизни этих людей, об уровне их общественно-экономического
развития.
Маркс говорит: «Такую же важность, какую строение
останков
костей имеет для изучения организации исчезнувших
животных видов, останки средств труда имеют
для изучения исчезнувших
общественно-экономических
формаций. Экономические эпохи различаются не
тем, что производится, а тем, как производится,
какими средствами труда» (Капитал, т. I. Маркс
и Энгельс. Сочинения, т. 23, стр. 191).
В общественном труде у древних людей принимали
участие, конечно, не только мужчины, но и женщины.
Но форма
их участия, очевидно,
была иной,
так
как в силу анатомо-физиологических
341

Рис. 145. Неандерталец
Реконструкция Н. А. Синельникова
и М. Ф. Нестурха, рисунок художника С. Г.
Оболенского, 1941
|
особенностей, свойственных женщинам, им не всегда
представлялось возможным с той же легкостью,
как мужчинам, участвовать
в охоте на более крупных
животных, требовавших долгой и быстрой погони.
Женщинам труднее было метать камни и
одолевать опасных зверей.
Не только охота, но и другие особенности жизни
наших предков вызывали необходимость разделения
труда между мужчиной и женщиной. Это вполне согласуется
со
следующим положением
Маркса:
«В пределах
семьи — а
с дальнейшим развитием в пределах рода — естественное
разделение труда возникает вследствие половых
и возрастных различий,
т. е. на чисто
физиологической почве, и оно расширяет свою
сферу с расширением общественной жизни, с
ростом населения, особенно же с появлением
конфликтов между различными родами
и
подчинением одного рода другим» (Капитал, т.
I. — Маркс и Энгельс. Сочинения, т. 23, стр.
364).
Усложнение коллективных действий и общественных
взаимоотношений уже в первобытном стаде неандертальцев,
несомненно,
повлияло положительным образом на прогрессивное
развитие головного мозга и возникновение новых
средств общения вместо
первоначального звукового языка из нечленораздельных
звуков, характерного для первых людей,
или обезьянолюдей.
Таким новым средством общения явилась членораздельная
речь, которая, вероятно, начала развиваться
еще у неандертальцев (если не у
синантропов), но окончательно
сложилась лишь позже — у кроманьонцев.
К кроманьонской стадии эволюции человечества
многие относят зарождение некоторых, например
связанных
с охотой на животных,
с колдовством,
или магией, фантастических
представлений о природных силах, а затем и
о силах общественных, из чего складываются
зачатки
религии.
О зарождении религии
Энгельс
342
писал: «...всякая религия является не чем иным,
как фантастическим отражением в головах людей
тех внешних
сил, которые господствуют
над ними в их повседневной
жизни,— отражением, в котором земные силы принимают
форму неземных. В начале истории объектами
этого отражения являются
прежде
всего силы природы,
которые
при дальнейшей эволюции проходят у различных
народов через самые разнообразные и пестрые
олицетворения... Но вскоре,
наряду с
силами природы, выступают
также и общественные силы...» (Маркс и Энгельс.
Сочинения, т. 20, стр. 328). Нахождение скелетов
первобытных
людей в пещерах приводило
ряд
ученых к идее
о культовых погребениях у поздних неандертальцев.
Развитие мозга
у ископаемых людей
Мозг современного человека является одним из
наиважнейших результатов развития гоминид как
общественных существ,
производящих орудия
труда. В человеческом
обществе начиная уже с первых стадий его развития
умственные способности индивида, полезные для
жизни в обществе,
начали выдвигаться все
более и более на первый план.
В ходе эволюции человека имели значение не
только сам процесс изготовления орудий, но
и их применение.
Труд,
членораздельная
речь, совместная
охота способствовали выживанию индивидов с
более сильным развитием головного
мозга.
Уже мозг питекантропа был в 1 1/2 раза крупнее,
чем мозг гориллы, а по относительному весу
должен был
превышать его примерно
в 3—4 раза.
На
протяжении 500 тыс.
лет мозг обезьянолюдей развился в мозг неандертальцев,
которые по его размерам и весу близки к современным
людям
(рис. 146).
Однако у неандертальцев мозг по строению был
значительно примитивнее, чем у современного
человека, например
по малому развитию передних
участков лобной доли.
Труд оказывал могущественное влияние на прогрессивное
развитие головного мозга. В стадной жизни наших
предков—обезьян — такой могучий стимул,
как труд, еще отсутствовал. Но у обезьянолюдей
он появился, и мозг под влиянием
общественной трудовой деятельности начал интенсивно
развиваться
и увеличиваться в размерах.
Средняя вместимость мозговой коробки яванских
питекантропов была 900 см3,
у китайских древнейших людей, или
синантропов,— около
1050 см3,
у неандертальцев
1300—1400 см3, т. е. вместимость была такая
же, как и у людей современного типа. Следовательно,
в течение
примерно
двух
третей плейстоцена
прирост объема мозга в 400 см3 равняется такому
же увеличению мозга, которое,
вероятно, произошло у предков гоминид на протяжении
во много раз большего промежутка
между временем расцвета дриопитеков до момента
появления первых людей.
Таким образом, темп развития головного мозга
за период от пи-
343
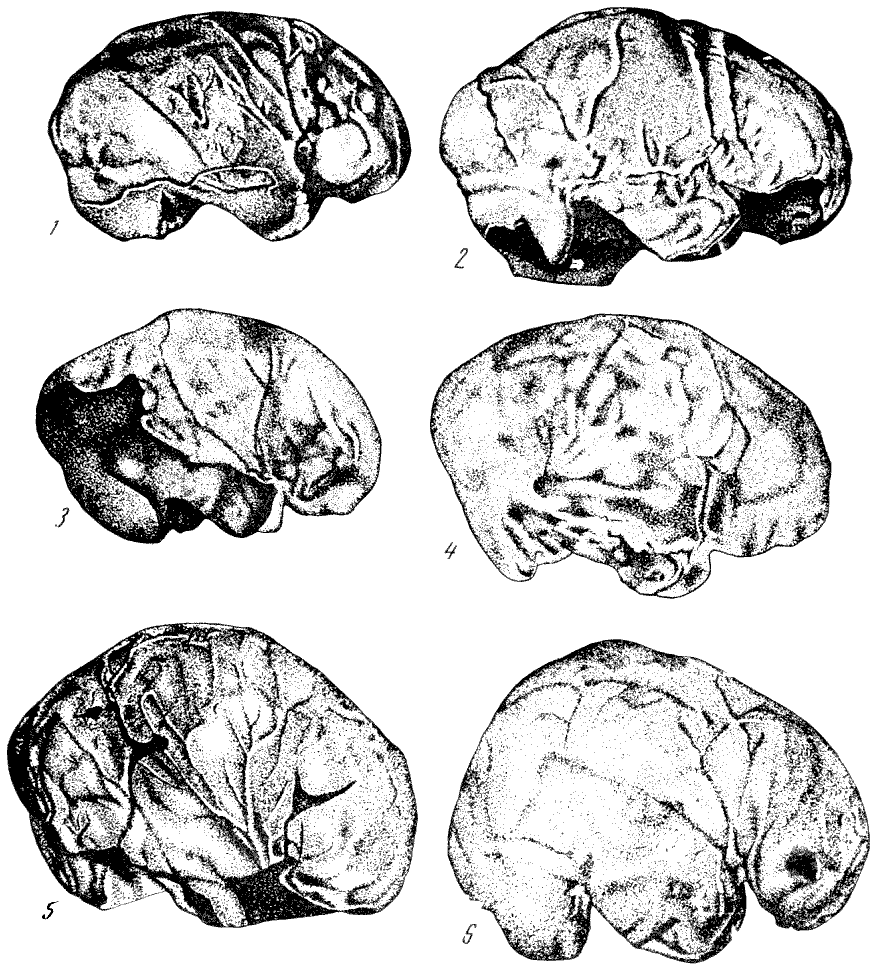
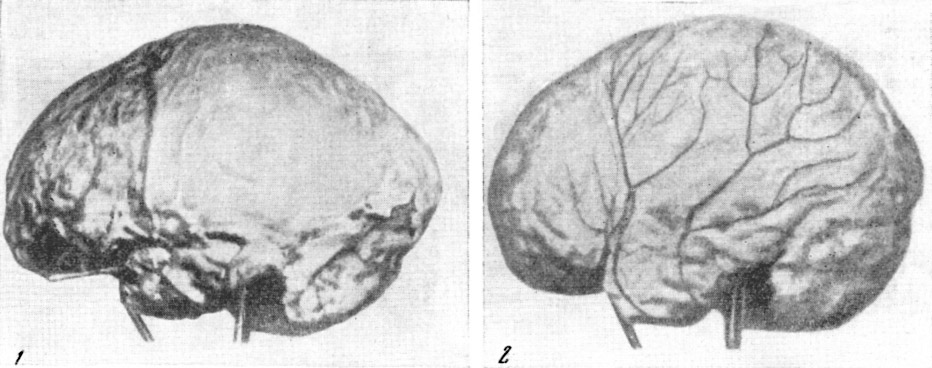
Рис. 146. Слепки полости мозговой коробки (эндокраны)
черепов:
1 — питекантроп; 2 — синантроп; 3 —
неандертальцы из Брокен Хиллаи; (4)
из Ла Шапелль; 5 —
ископаемый человек
из
Пржедмости; 6 —
современный человек. По Ф. Тильнею, 1928
(1, 3, 4, 5, 6)
и Э. Дюбуа, 1933 (2), 1/5—1/4
нат. вел. (ширина изображений, соответственно, 47 мм, 53 мм, 44 мм, 51 мм,
53 мм, 54 мм)
текантропа до неандертальцев является относительно
и абсолютно весьма интенсивным, хотя в то
время приемы первоначальной
техники и примитивные
формы человеческого
общества в течение сотен тысяч лет сравнительно
мало изменялись.
Благодаря, однако, новизне и силе воздействия
труда на человеческий организм головной
мозг первых людей
испытал
такие темпы развития,
каких никогда
не было да и не могло быть ни у одного
животного. Если у наших миоценовых предков
— дриопи-
344
теков — головной мозг имел объем, вероятно,
400—500 см3, а
у питекантропов увеличился почти вдвое,
сохранив еще
многие примитивные
черты,
то у людей современного типа его размеры
возросли уже втрое, причем форма
мозга
и сложность его строения сильно изменились
(Кочеткова, 1967). Очень сильное развитие,
большие размеры и вес головного мозга
человека составляют для
идеалистов, для религиозно настроенных
людей препятствие к допущению правильности
научного
объяснения естественного хода процесса
антропогенеза. Однако именно совершенно новый, непривычный
для обезьяны фактор
труда в обществе
себе подобных
с выделыванием и употреблением искусственных
орудий при жизненно необходимейших
потребностях в пище и в защите от врагов очень интенсивно
побуждал
творческие функции мозга к быстрому
и могущественному уникальному
прогрессу в процессе
руппового отбора (Нестурх, 1962а).
В течение плейстоцена шла прогрессивная
эволюция абсолютных размеров, формы
и строения головного
мозга гоминид
параллельно с редукцией
некоторых его
участков. Определенные сведения об
изменении формы и размеров мозга ископаемых
гоминид
были получены
при изучении слепков
внутренней полости мозгового
отдела черепа.
На внутренней стенке черепа ископаемого
человека хорошо видны следы от кровеносных
сосудов,
некогда шедших
по поверхности мозга, но
извилины мозга проецируются
слабо. Даже подразделение мозга на
части не всегда возможно установить с достаточной ясностью.
Те же трудности испытывают
и при изучении слепков
мозговой полости
черепов современных людей. Все это
осложняет и подчас делает невозможным
изучение
более мелких, но важных участков, как,
например, двигательной, речевой и нижнетеменной
областей,
имеющих большое значение
с эволюционной точки зрения.
Мозг человека заключен в оболочки,
которые прилегают к стенке мозговой
полости гораздо
ближе у ребенка,
чем у
взрослого,
поэтому и слепки
мозговой полости
черепа ребенка лучше выражают строение
поверхности мозга. Тилли Эдингер (Edinger,
1929) указывает,
что у человека,
а также
антропоидов, слонов,
китов и прочих
животных с крупным мозгом, покрытым
извилинами, поверхность слепка мозговой
полости представляется
почти гладкой,
Эдингер пишет,
что, если «кто хочет
исследовать мозг по слепку полости
черепа, как это и вынужден делать палеоневролог,
тот блуждает
в потемках».
В этом отношении Эдингер скорее согласна
с Симингтоном (1915), который, считает,
что:
1) по слепку полости человеческого
черепа нельзя судить о простоте или
сложности
рельефа мозга;
2) по слепкам мозговой полости черепа
неандертальца из Ла Шапелль-о-Сен нельзя
даже приблизительно
судить об относительном
развитии
чувствительных и ассоциативных
зон коры;
3) различные заключения Буля, Антони,
Эллиота-Смита и других
345
исследователей в отношении примитивных
и обезьяньих черт мозга некоторых
доисторических людей,
полученные путем
изучения шлепков
полости мозгового
отдела черепа, в высшей степени
умозрительны и ошибочны.
Но всё же эти слепки дают возможность,
как соглашается и Эдингер, сделать
некоторые заключения о форме
и главнейших особенностях
мозга, например
о степени развития лобной и затылочной
долей.
Так, Э. Дюбуа (Dubois, 1924) при
описании слепка мозговой
полости питекантропа
подчеркивает,
что на
отпечатке
видны важные, хотя и не прямые
указания на характерные особенности
первоначальной формы мозга человека.
Мозг питекантропа, судя по муляжу,
обладал очень
узкими лобными долями при сильном
развитии нижней лобной извилины.
Дюбуа полагает,
что последнее доказывает возможность
развития членораздельной речи.
По описанию Дюбуа, весьма характерна
уплощенность слепка мозга питекантропа
в теменной области.
С мозгом прочих
гоминид сходство
состоит в том,
что наибольшая его ширина лежит
на 3/5 длины от переднего края
лобного
отдела.
В общем, мозг питекантропа, по
мнению Дюбуа,
является как бы увеличенной копией
мозга человекообразных обезьян.
Некоторые особенности сближают
его больше с мозгом гиббона: об
этом, по Дюбуа, свидетельствуют
положение
верхней
прецентральной извилины и другие
признаки.
Для суждения о типе неандертальца
обычно пользуются слепками со следующих
черепов:
Неандерталь, Ла Шапелль-о-Сен,
Гибралтар, Ла Кипа. Эдингер дает (с оговорками)
следующую
характеристику
мозга
неандертальца: по типу
строения это
человеческий мозг, но с явственно
выраженными обезьяньими чертами.
Он длинен и низок,
впереди более узок, сзади более
широк; возвышение в теменной области
ниже,
чем у современного
человека, но выше, чем
у человекообразных обезьян. По
меньшему числу борозд и их расположению
в известной степени напоминает
мозг человекообразных обезьян.
О
том же свидетельствует
угол
отхождения
продолговатого
мозга и заостренность лобной доли
в виде клюва, а также большее развитие
затылочных
долей,
заключающих в себе
зрительную зону. Червячок в
мозжечке относительно более развит,
чем у
современного человека,
и это более
примитивный признак.
Больше доверия, по Эдингер, можно
оказать сведениям об основных размерах
мозга
ископаемых гоминид
(табл. 5).
Из табл. 5 видно, что у некоторых
неандертальцев были сравнительно
крупные головы и
крупный мозг.
Точно так же можно было, хотя и
не всегда, получить достаточно
точные
цифры, характеризующие
объем
мозговой полости черепа
других гоминид.
Из всех формировавшихся
(древнейших и древних) людей неандерталец
из Ла-Шапелль-о-Сен обладал, по-видимому,
максимальным
объемом мозговой
коробки (1600 см3),
а питекантроп II — минимальным
(750 см3).
У неандертальцев размах вариаций
ее объема был срав-
346
Таблица 5
Размеры черепа и слепка мозговой
полости (эндокрана) у гоминид
(по Т. Edinger,
1929)
Гоминиды |
Длина, см |
Ширина, см |
Череп |
| Современный человек (баварцы; указан размах вариаций) |
14,3—22,5 |
10,1—17,3 |
| Ла Шапелль-о-Сен |
20,8 |
15,6 |
| Ла Кина |
20,3 |
13,8 |
| Неандерталь |
19,9 |
14,7 |
Слепок мозговой полости |
| Ла Шапелль-о-Сен |
18,5 |
14,5 |
| Ла Кина |
17,75 |
13,1 |
| Неандерталь |
17,5 |
13,8 |
нительно еще невелик, составляя
примерно 500 см3 против
900 — у современного
человека. Однако
не
следует забывать,
что
минимум
и максимум (размах
вариаций) зависит также
от числа изученных индивидов.
Длина эндокрана
современного
человека составляет примерно
166 мм, а ширина — 134
(Бунак,
1953).
Для мозга ископаемых гоминид
характерно развитие асимметричности
в его
форме. Более сильно
развито обычно левое полушарие,
что может указывать
на преимущественное
употребление правой руки. Праворукость
или леворукость составляет
характерный признак
человека в
отличие от млекопитающих
животных. Значительная
асимметрия верхних конечностей
могла появиться
лишь после того, как у
наших предков развилось
прямохождение
и
появился труд.
Асимметрия в величине полушарий
замечается уже у питекантропа.
По Э. Смиту (Smith,
1934), он
должен был быть левшой.
Напротив, Ф. Тильней
(Tilney,
1928) обращает
внимание на то, что левая лобная
доля у питекантропа была
крупнее, и
полагает, что это указывает
скорее на его
праворукость.
Вообще, о более
сильном
развитии левого полушария у
питекантропа можно судить
по тому, что
на его черепе
более
заметное углубление видно на
внутренней поверхности
левой затылочной кости.
Отмечены
асимметрии и на слепке
мозговой полости
черепа синантропа.
Ясно обнаруживается асимметрия
мозга у неандертальцев,
у которых она видна
в форме, типичной для
современного человека.
На слепке
мозговой
полости
черепа из Ла Шапелль-о-Сен
левое полушарие короче
правого
на
347
3 мм, но зато шире его на
7 мм и выше, причем
теменно-височный участок
выступает
на нем
сильнее. К этому присоединяется
еще то, что в
скелете правой руки
плечевая кость из Ла Шапелль-о-Сен
имеет более крупные
размеры, чем левая.
На слепке мозговой
полости гибралтарского
черепа
затылочная доля левого
полушария явственно
сильнее выдается назад.
На слепке
полости черепа
из Ла Кина левое
полушарие длиннее, правое
же
более развито. Наконец,
на слепке
полости черепа
из Неандерталя
правое полушарие крупнее
левого.
Из этого описания видно,
что у древнейших и древних
гоминид
праворукость
встречалась,
как кажется,
чаще или же наравне
с леворукостью.
Форма и способ выделки
каменных орудий, а
также стенные рисунки
древних людей иногда тоже
позволяют судить
о преимущественном пользовании
левой или
правой рукой.
По Р. Коблеру
(КоЫег, 1932),
у людей развилась сперва
левору-кость; позже,
в связи с употреблением
более
сложных форм оружия (например,
в сочетании
с таким оборонительным
приспособлением,
как
щит), стала использоваться
преимущественно правая
рука. Коблер ссылается
на
то, что на большей
части
древнейших
орудий видны
следы их обработки
левой рукой.
Но Эдингер сообщает, что
у первобытных людей верхнего
палеолита 2/з
всех кремневых орудий
были сделаны
правшами, равно
как и стенная живопись
в
пещерах.
Слепки мозговой полости
черепов ископаемых
форм людей современного
типа и их
потомков во всем существенном
сходны.
В результате скорее можно
согласиться с Д. Ж. Кеннингемом
(1902),
который еще до
того,
как
стали известны
слепки мозга ископаемых
людей, писал,
что праворукость
развилась как характерная
особенность человека уже
в очень ранний
период его эволюции,
по всей вероятности,
до того,
как развилась
способность
к членораздельной речи.
Он отмечает, что левое
полушарие
у большинства
современных людей развито
сильнее правого.
Итак, в итоге длительного
развития от обезьяны до
человека на
протяжении нескольких
последних
миллионов
лет мозг
наших, предков
— миоценовых
и затем плиоценовых
антропоидов — увеличился
и видоизменился, а в плейстоцене
испытал особый
подъем развития у
ископаемых гоминид
и достиг высокого
развития к стадии
людей современного
типа (Koenigswald, 1959).
Эволюция человеческого
мозга становится понятной
в свете
учения Дарвина
о развитии органического
мира и учения
Энгельса о роли
труда в процессе
формирования
человека.
Мозг достиг высокого развития
уже у ближайших предшественников
гоминид,
т. е. у австралопитеков,
но особый, могучий
толчок это развитие получило
лишь
при возникновении
трудовых действий у питекантропов.
Переход от обезьяны к человеку
был бы немыслим без наличия
высокоразвитого мозга у
его ближайшего предка.
Это сильно
способствовало тому,
что произошли резкие
изменения в поведении наших
предков,
появились новые
348
формы жизнедеятельности,
т. е. способы добывания
пищи и
защиты от врагов,
особые приемы при
осуществлении других
необходимых действий с помощью
искусственных органов
в виде изготовленных
орудий.
Дарвин ставил на видное
место высокое умственное
развитие
наших предков.
По его словам,
ум должен был иметь
для человека первостепенное
значение
даже в
очень
древнюю эпоху, так
как
позволил изобрести
и применять членораздельную
речь,
выделывать оружие,
орудия,
ловушки и т. д. Вследствие
этого человек при
содействии своих
общественных привычек
уже
с давних пор стал господствующим
из всех
живых существ.
Далее Дарвин пишет:
«Развитие ума должно
было сделать
значительный шаг вперед,
когда благодаря
прежним
успехам у человека
вошла в употребление
речь, как полуискусство
и полуинстинкт.
Действительно,
продолжительное употребление
речи
должно было отразиться
на мозге и обусловить
наследственные изменения,
а эти,
в свою
очередь, должны были
повлиять на усовершенствование
языка.
Большой
объем
мозга у человека
сравнительно с низшими
животными,
по отношению к величине
их тела, может
быть главным
образом отнесен,
как справедливо
заметил
м-р
Чонси Райт,
на счет раннего
употребления какой-либо
простой формы речи
— этого дивного
механизма, который
обозначает
различного
рода предметы
и свойства определенными
знаками и
вызывает ряд мыслей,
которые никогда не
могли бы
родиться из одних чувственных
впечатлений, или
если бы даже и родились,
не
могли
бы развиваться»
(Соч.,
т. 5, стр.
648).
Для эволюции человеческого
мозга исключительное
значение имело
возникновение и развитие
членораздельной речи,
которая является,
вероятно, очень
древним приобретением
человека. По мысли
Энгельса, она
зародилась уже
во время
переходного периода
от обезьяны к человеку,
т. е. у формировавшихся
людей.
Характеризуя
исторические
ступени культуры, Энгельс
предположительно говорит
о низшем отделе первого
из них, т. е. эпохе
дикости, следующее:
«Детство человеческого
рода. Люди находились
еще в
местах своего первоначального
пребывания, в
тропических или субтропических
лесах. Они жили, частью
по крайней мере, на
деревьях;
только этим
и можно объяснить
их существование
среди крупных хищных
зверей.
Пищей служили
им
плоды, орехи,
коренья; главное достижение
этого периода — возникновение
членораздельной
речи.
Из всех народов, ставших
известными в исторический
период, уже
ни один не находился
в этом первобытном
состоянии. И хотя оно
длилось, вероятно,
много
тысячелетий,
однако
доказать
его на основании
прямых свидетельств
мы не можем; но,
признав происхождение
человека из царства
животных, необходимо
допустить
такое
переходное состояние»
(Маркс и Энгельс. Сочинения,
т.
21, стр.
23—178).
Зарождение звуковой
речи некоторые относят
довольно
далеко, ко
временам нижнего
или среднего палеолита.
Синантроп,
может
349
быть, ею уже обладал
в зачатке. У неандертальцев,
вероятно,
уже должна
была быть начальная
ее стадия.
Блэк считает, что
способность к членораздельной
речи
была уже у
синантропа. Надо
полагать, что яванские
питекантропы были
еще действительно
неговорящими
людьми;
они, как и животные,
имели ряд жизненно
важных нечленораздельных
звуков,
обозначавших то
или иное внутреннее
состояние,
но имевших сигнальное,
трудовое значение
и бывших более
разнообразными,
чем у современного
шимпанзе. Вероятно,
у древнейших
людей,
как и у антропоидов
тина шимпанзе,
были в
ходу и неэффективные,
сравнительно
тихие вокальные
звуки, или «жизненные
шумы», имевшие,
по
В. В. Бунаку, особо
важное значение
для возникновения
речи (Бунак,
1951, 1966,
Yerkes, Learned,
1925).
Американские
ученые Роберт Йеркс
и Бланш
Лёрнед специально
изучили
звуки, издаваемые
шимпанзе.
Они пришли к убеждению,
что у шимпанзе
есть около
тридцати своеобразных
звуков и что каждый
из этих звуков
имеет свое специфическое
сигнальное
значение, обозначая
какое-нибудь внутреннее
состояние или же
отношение к происходящим
вокруг явлениям.
Возможно, однако,
что этих звуков
у шимпанзе не так
много, десятка
два
— два с
половиной.
О звуках, издаваемых
гориллами, мало
известно. Описывают
обычно рев самца,
идущего на
врага. Один ученый
наблюдал самца
горной
гориллы,
который
сидел на лежавшем
дереве вместе
с двумя самками:
ученый слышал негромкие
звуки,
которыми
они мирно обменивались
друг с другом.
Число основных
звуков у горилл
невелико
(Шаллер, 1968).
У орангутанов мало
звуков: они молчаливы
и издают
рычание, рев
или визг только
при каких-нибудь
особых обстоятельствах
— при испуге, в
гневе, от боли.
Громкие
звуки,
издаваемые гиббонами,
слышны
за километры.
Все попытки Роберта
Йеркса научить
своих шимпанзе
говорить окончились
неудачей,
хотя он и применял
различные методы
обучения. Йеркс
предполагал применить
к шимпанзе также
и методы, с помощью
которых
специалисты-педагоги
учат
говорить глухонемых
детей. Если подобные
попытки и могли
бы увенчаться
известным
успехом,
то только при условии
применения подходящих
приемов
обучения к самым
маленьким детенышам,
так
как онтогенетическое
развитие
головного мозга
у шимпанзе заканчивается
раньше, чем у человека.
Но надо иметь в
виду, что основной
причиной, почему
обезьян
очень трудно
обучить хотя
бы немногим словам,
является прежде
всего зачаточное
состояние
их речевых зон.
Кроме того,
нельзя не учитывать
и заметных отличий
в строении голосового
аппарата у обезьян
по сравнению с
человеком (см.
упомянутые
выше статьи
В. В. Бунака,
1951 и 1966б).
Людвиг Эдингер
(Edinger, 1911),
отмечая высокое
развитие коры
головного мозга
у шимпанзе,
допускает, что
терпеливый
дрессировщик
мог бы
обучить человекообразную
обезьяну нескольким
словам, но обезьяна
всегда оста-
350
валась бы на неизмеримо
далеком расстоянии
от человека,
так как основы для
отчетливого
понимания, т. е.
соответствущие
участки
мозга, у
нее не развиты.
Многие авторы
считают, что
анатомической
предпосылкой
для
развития речи
у человека
является наличие
подбородочного
выступа.
Этот
выступ есть
лишь у
современного
человека. Он
отсутствовал,
как правило,
у неандертальцев,
его
не было у обезьянолюдей,
а также (кроме
сростнопалого
гиббона
—
сиаманга) его
нет
у современных
и ископаемых
обезьян и полуобезьян.
Возникновение
звуковой речи
нет необходимости
непременно
связывать
с наличием
подбородочного
выступа, так
как для производства
членораздельных
звуков требуется
прежде
всего четкая
согласованная
работа всего
речевого аппарата,
включая сенсорную
и
мнестическую
зоны
головного мозга,
располагающиеся
в филогенетически
новых областях
теменной и
височной
долей.
Образование
подбородочного
выступа у человека
произошло,
по Л. Больку,
главным
образом
в силу редукции
той части нижней
челюсти,
которая
несет зубы.
Нижняя же половина,
составляющая
само тело челюсти,
подверглась
процессу
редукции
в меньшей
степени, вследствие
чего и обозначился
подбородочный
выступ.
Среди млекопитающих
некоторую аналогию
можно было
бы усмотреть
в выдающейся
вперед подбородочной
части нижней
челюсти
слона, так
как
зубная система
его подверглась
еще более сильной
редукции, вследствие
чего
состоит лишь
из четырех
коренных
зубов и
двух верхних
резцов, или
бивней,
т. е. всего
из шести
зубов.
Речевая функция
могла лишь
второстепенным
образом влиять
на
основной процесс
формирования
подбородочного
выступа
(Гремяцкий,
1922).
Для развития
речи у людей
не меньшее
положительное
значение имели
преобразование
формы челюсти
из вытянутой
в
подковообразную,
увеличение
объема полости
рта,
в которой движется
язык,
а также более
свободное
движение
челюсти
в новых направлениях
в связи с
уменьшением
размеров клыков.
Несравненно
важнее для
развития
членораздельной
речи анатомо-физиологические
особенности
соответствующих
участков коры
лобного отдела
больших
полушарий
головного
мозга (наряду
с височной
и теменной).
Были
сделаны попытки
установить
на слепках
мозговой
полости
ископаемых
людей степень
развития
этого
столь
важного отдела
коры. К сожалению,
по слепку мозговой
полости
черепа, или
эндокрану,
даже обладая
слепком
мозговой полости
черепа современного
человека,
трудно сделать
вывод о пользовании
членораздельной
речью
(Эдингер,
1929). Это
весьма затруднительно
и при изучении
самого
мозга.
Муляж
полости черепной
коробки дает
понятие лишь
о том, какова
была
форма
мозга, одетого
в свои оболочки,
которые
образуют настолько
плотный покров,
что очень скрывают
извилины
и борозды
мозга, обнаруживая
ярко лишь
картину расположения
более крупных
кровеносных
сосудов.
Но-
351
вая, успешная
попытка
изучения эндокранов
гоминид
предпринята на большом
материале
в лаборатории
мозга Института
антропологии
(Кочеткова,
1966).
Членораздельная
речь не
является
врожденным
свойством.
Это
следует,
в частности,
из описания
редких
случаев,
когда
дети вырастали
в полной
изоляции
либо среди
животных,
вдали
от человеческого
общества,
и, будучи
найдены,
не умели
говорить.
Из связей
и взаимоотношений
индивидуального
и группового
характера
среди
древних
гоминид
для
возникновения
речи наибольшее
значение
имели
те,
которые
развивались
на основе
трудовых
процессов.
Во
время коллективной
охоты
на животных
и последующего
распределения
мяса между
членами
общества,
во время
совместного
изготовления
орудий,
при деятельности
в течение
трудового
дня, наполненного
борьбой
за существование,
люди постоянно
испытывали
необходимость
в
такой звуковой
сигнализации,
которая
регулировала
бы и направляла
их
действия.
Тем самым
различные
звуки,
а
также связанные
с ними
мимика
и жесты
становились
для них
жизненно
важными,
показывая
в общепонятной
форме необходимость
тех,
а не иных
поступков,
полезность
актов,
так
или иначе
согласованных
между
членами
первобытного
стада.
Особое
значение
звуки
голоса
имели
в
условиях
темноты.
С другой
стороны,
объединение
наших предков
вокруг
костра
в пещере
должно
было
тоже способствовать
развитию
звукового
языка.
Пользование
огнем
и
изобретение
способов
его добывания
дали, надо
полагать,
мощный
толчок
развитию
членораздельной
речи уже
у неандертальцев.
Марксистское
объяснение
того, как
возникла
и развивалась
членораздельная
речь, дано
Энгельсом.
Он пришел
к выводу,
что
речь
как средство
общения
между
людьми
по необходимости
возникла
из звуков
голоса,
сопровождавших
и предварявших
трудовые
операции,
а также
другие
совместные
действия
членов
коллектива
формировавшихся
людей.
Энгельс
пишет:
«Начинавшееся
вместе
с развитием
руки,
вместе
с
трудом
господство
над природой
расширяло
с каждым
новым
шагом вперед
кругозор
человека.
В
предметах
природы
он постоянно
открывал
новые,
до того
неизвестные
свойства.
С
другой
стороны,
развитие
труда по
необходимости
способствовало
более тесному
сплочению
членов
общества,
так как
благодаря
ему стали
более
часты случаи
взаимной
поддержки,
совместной
деятельности,
и стало
ясней
сознание
пользы
этой совместной
деятельности
для каждого
отдельного
члена.
Коротко
говоря,
формировавшиеся
люди пришли
к тому,
что
у них появилась
потребность
что-то
сказать друг другу.
Потребность
создала
себе
свой
орган:
неразвитая
гортань
обезьяны
медленно,
но неуклонно
преобразовывалась
путем модуляции
для все
более развитой
модуляции,
а органы
рта постепенно
научались
произносить
один членораздельный
звук за
другим»
(Маркс
и
Энгельс.
Сочинения,
т. 20,
стр. 489).
Если высокое
развитие
головного
мозга
наряду
с прямохож-
352
дением
и рукой
послужило
важнейшей
предпосылкой
появления
речи,
то
не менее важно и
обратное
влияние
речи
на
мозг.
Энгельс писал:
«Сначала
труд,
а затем и
вместе
с ним
членораздельная речь явились
двумя
самыми главными
стимулами,
под
влиянием которых
мозг
обезьяны постепенно
превратился
в человеческий
мозг»
(там же,
стр.
490).
Будучи
чрезвычайно
выгодным,
общественно
полезным
явлением,
речь
неизбежно
развивалась
дальше
и дальше.
В подтверждение
своей
теории
развития
языка
в процессе
труда
Энгельс
привлекает
примеры
из
жизни
животных.
В то
время
как
для
диких
животных
звук
человеческой
речи
может
обозначать,
вообще
говоря,
лишь
знак
возможной
опасности,
для
домашних
животных,
например
для
собак,
человеческая
речь
делается
в ряде
отношений
понятной,
на
каком
бы
языке
человек
ни
говорил,
но,
конечно,
лишь
в «пределах
свойственного
им
круга
представлений».
Для
домашних
животных
слова,
произносимые
человеком,
становятся
сигналами
определенных
действий,
которые
должны
последовать
со
стороны
человека
либо
быть
выполнены
ими
самими.
Животные,
более
способные
к
быстрому
и прочному
образованию
условных
рефлексов,
к дрессировке,
оказываются
и наиболее
понятливыми
в прирученном
или
домашнем
состоянии,
когда
соблюдение
нужных
действий,
согласно
этим
сигналам,
может
повлечь
за
собой
одобрение,
а несоблюдение
вызывает
наказание.
Звуки
членораздельной
речи,
первоначально
служившие,
наиболее
вероятно,
сигналами
действий,
начали
затем
обозначать
также
предметы
и явления;
возрастало
количество
звуковых
сигналов;
все
большее
значение
приобретали
их
сила,
высота,
тембр
(обертоны),
интонация,
последовательность.
В связи
с развитием
звукового
языка
эволюционировал
и производивший
их
речевой
аппарат.
Совершенствовался
и слуховой
анализатор,
который
является
у человека
по
сравнению
с некоторыми
млекопитающими
не
всегда
столь
утонченным
в отношении
улавливания
мельчайших
различий
в высоте
тона
и в
тембре
звуков
членораздельной
речи.
Но
человек
резко
превосходит
в понимании
их
внутреннего
смысла,
в частности,
когда
дело
касается
тех
или
иных
сочетаний
звуков:
в этом
отношении
его
слуховой
анализатор
является
высокоспециализированным,
позволяя
различить
гораздо
большее
число
и значение
звуков,
чем
это
доступно
любому
животному.
В
то
же
время
периферический
отдел
слухового
анализатора
у человека
подвергся,
как
и у
некоторых
обезьян,
редукции,
на
что
указывает,
в частности,
почти
полная
неподвижность
ушной
раковины
человека
с ее
рудиментарными
мышцами.
Корковый
отдел
слухового
анализатора
человека,
согласно
исследованию
С.
М.
Блинкова
(1955),
качественно
отличается
и сложностью
строения
резко
превосходит
соответствующий
участок
даже
у антропоидов;
то
же
относится
и ко
всей
височной
доле.
Однако
в формировании
речи
принимают
участие
не
только
лобная,
височная и теменная
доли,
но
и вся
кора
в
целом.
353
Словесное
мышление встречается
только у
человека: вторая
сигнальная система,
по термину
И. П.
Павлова, является
важнейшей основой
развития сознания.
Будучи неразрывно
связана с
первой сигнальной
системой, охватывающей
условные рефлексы
обычного типа,
вторая сигнальная
система объединяет
свойственные лишь
человеку сознательные
условные рефлексы
на слова,
знаменующие действия,
предметы, отношения
между ними,
понятия и
т. п.
Тезис И.
П. Павлова
о второй сигнальной
системе —
одно из
величайших завоеваний
советской науки.
Он позволяет
углубить разработку
идеи Энгельса
о происхождении речи
в трудовых процессах.
Эта проблема
привлекала внимание
крупнейших отечественных
мыслителей. Очень
интересные строки
относительно возникновения
речи мы
читаем у
А. М.
Горького: «Известно,
что все
способности, отличающие
человека от
животного, развились
и продолжают развиваться
в процессах труда,
способность членораздельной
речи зародилась
тоже на
этой почве».
(Полное собрание
соч., 1953,
т. 27,
стр. 164).
Сперва, говорит
он, развились
формы глагольные
и измерительные (тяжело,
далеко), далее
наименования орудий
труда. По
мысли А.
М. Горького,
бессмысленных слов
в начальной речи
не было
(стр. 138).
Как речь,
так и
ум человека
ставятся A.M.
Горьким в
теснейшую, органическую
связь с
трудовой деятельностью:
«Разум человеческий
возгорелся в
работе по
реорганизации грубо
организованной материи
и сам по
себе является
не чем
иным, как
тонко организованной
и все
больше тончайше
организуемой энергией,
извлеченной из
этой же
энергии путем
работы с
нею и
над ней,
путем исследования
и освоения
ее сил
и качеств»
(там же,
стр. 164—165).
Вероятно,
членораздельная речь
способствовала прогрессивному
развитию
человечества уже
на неандертальской
фазе его
формирования: интенсивное
развитие речи
в это
время, наверное,
в значительной
мере способствовало
превращению древних
людей в
более высокий тип
кроманьонцев. Позднейшие
неандертальцы с
их умением
добывать огонь,
возникающим обычаем
закапывать умерших
в пещерах,
гротах, служивших
жилищем, с
их приемами
обработки кости
стояли выше
своих предшественников,
т. е.
более ранних
неандертальцев (Семенов,
1959).
В
еще большей
мере членораздельная
речь развивалась
и усложнялась
у ископаемых
людей современного
типа, т.
е. у
«новых», или
«готовых» —
«разумных», людей,
все более
быстрыми темпами
проходивших дальнейшие
эпохи истории
материальной культуры,
стадии общественно-экономического
развития
(Войно, 1964).
Как
видно из
предыдущего изложения,
современное человечество
представляет собой
результат длительной
эволюции, которая
в первом,
наиболее продолжительном
отрезке филогенетической
родословной человека
была составной
частью общего
хода развития
животного мира
с характерными
для него
биологическими закономерностями.
Но
самое появление
первых людей
с их
трудом, общественностью,
354
языком
было скачком,
особым перерывом
постепенности в
ходе эволюции
их ближайших
предков. Путем
резкого перехода,
крутого, решающего
поворота в
ходе эволюции
началась новая
стадия развития
живой материи,
когда возникало
древнейшее человечество.
Это и
было начало
совершенно нового
процесса формирования
человека —
гоминизации. Формировавшиеся
древнейшие
и древние
люди не
были животными,
как это
полагает Б.
Ф. Поршнев
(1955а), считающий
за людей
только представителей
вида Homo
sapiens.
Труд
древнейших и
древних людей,
изготовлявших орудия,
принципиально, качественно,
отличается от
«труда» бобров,
муравьев, пчел,
птиц, вьющих
гнезда. В
эволюции животных
действуют лишь
природные, биологические
факторы.
Под
влиянием совокупности
социальных и
биологических факторов
шло превращение
обезьяны в
человека: этот
качественно отличный
от эволюции
животного мира
процесс формирования
может быть
правильно понят
лишь в
свете диалектико-материалистического
учения
Энгельса о
решающей роли
труда.
По
мнению Я.
Я. Рогинского
(1967), появление
трудовых действий
обозначило начало
диалектического скачка
от животного
к человеку
— первый
поворотный пункт
в эволюции
гоминид, а
второй —
с появлением
современного человека
и открытием
эры господства
законов социальных
означает конец
скачка. Развитие
культуры человека
современного не
связано с
прогрессивной эволюцией,
как это
было у
палеоантропа или
архантропа. Весь
ход формирования
гоминид под
влиянием труда
закономерно привел
к появлению
нового качества
в неоантропе.
У любого
современного народа
независимо от
его расового
состава переход
к более
высокой социально-исторической
формации
совершается вне
зависимости от
эволюционного процесса,
под влиянием
лишь исторических
закономерностей.
Диалектико-материалистическое
представление о
процессе формирования
человека, его
мозга, речи,
мышления служит
в советской
антропологии прочнейшей
базой для
углубленного исследования
антропогенеза, для
борьбы против
всех и
всяческих идеалистических
гипотез в
этой области
науки о
человеке, а
также в
области расоведения
для разоблачения
расизма на
основе антропологических
данных.
Оглавление
| Предисловие |
5
|
Часть I. Дарвиновская и другие гипотезы антропогенеза
|
7
|
| Глава первая |
Дарвин о происхождении человека |
8
|
| |
Идея антропогенеза до Дарвина
|
8
|
| |
Дарвин об эволюции животного мира
|
13
|
| |
Человеческая родословная по Дарвину
|
17
|
| |
Очерк развития знаний о приматах
|
29
|
| |
Развитие приматологии в СССР |
40
|
| Глава вторая |
Человекообразные обезьяны и их происхождение |
56
|
| |
Современные антропоиды
|
56
|
| |
Ископаемые антропоиды |
70
|
| Глава третья |
Позднейшие гипотезы происхождения человека
и их критика
|
95
|
| |
Религиозные трактовки антропогенеза
|
95
|
| |
Тарзиальная гипотеза
|
99
|
| |
Симиальные гипотезы
|
104
|
| |
Гипотеза антропогенеза Осборна
|
111
|
| |
Гипотеза антропогенеза Вейденрейха
|
119
|
| |
Некоторые факторы гоминизации и вымирания ископаемых антропоидов плиоцена
и плейстоцена |
124
|
| Часть II.Особенности строения человеческого тела
и возникновение древнейших людей |
133
|
| Глава первая |
Человек как примат |
134
|
| |
Черты приспособленности человеческого тела к прямохождению
|
134
|
| |
Характерные отличительные особенности человеческого тела, не стоящие
в непосредственной связи с прямохождением
|
140
|
| |
Специальные черты сходства между человеком и антропоидами
|
145
|
| |
Рудименты и атавизмы у человека |
148
|
| Глава вторая |
Роль труда и прямохождения в антропогенезе
|
154
|
| |
Роль труда
|
154
|
| |
Способы передвижения у человекообразных обезьян
|
157
|
| |
Вес тела в центр тяжести у человека и обезьян
|
165
|
| |
Нижние конечности
|
168
|
| |
Костный таз, позвоночник и грудная клетка
|
173
|
| |
Верхние конечности
|
180
|
| |
Пропорции тела и асимметрии
|
185
|
| |
Череп |
189
|
| Глава третья |
Головной мозг и высшая нервная деятельность
человека и обезьян
|
199
|
| |
Головной мозг и анализаторы человека и обезьян
|
199
|
| |
Развитие периферических частей анализаторов
|
210
|
| |
Высшая нервная деятельность обезьян
|
226
|
| |
Вторая сигнальная система — характерное отличие мышления человека |
246
|
| Глава четвертая |
Стадность у обезьян и зачаточные формы труда
|
249
|
| |
Стадность у обезьян
|
249
|
| |
Зачаточные формы труда
|
263
|
| |
Антропогенез и его факторы |
272
|
Часть III. Формирование человека по данным палеоантропологии
|
233
|
| Глава первая |
Первая ступень: древнейшие люди (питекантропы)
|
284
|
| |
Питекантроп острова Явы
|
284
|
| |
Синантроп
|
292
|
| |
Гейдельбергский человек |
305
|
| Глава вторая |
Вторая ступень: древние люди (палеоантропы)
|
309
|
| |
Ледниковая эпоха
|
309
|
| |
Неандертальцы и их физический тип
|
314
|
| |
Находки неандертальцев в СССР
|
322
|
| |
Палестинские неандертальцы
|
330
|
| |
Образ жизни древних людей
|
339
|
| |
Развитие мозга у ископаемых людей |
343
|
| Глава третья |
Современные люди (неоантропы)
|
356
|
| |
Человек верхнего палеолита
|
356
|
| |
Ошибочные гипотезы происхождения современного человека и их критика
|
370
|
| |
Человеческие расы
|
380
|
| |
Факторы формирования рас
|
387
|
| |
Наука против расизма |
394
|
| Summary |
|
403
|
| Приложение 1. |
Предложения по биологическим аспектам
расовой проблемы
|
406
|
| Приложение 2. |
Декларация о расе и расовых предрассудках
|
410
|
| Литература |
|
414
|