Глава третья
Головной мозг и высшая нервная деятельность
человека и обезьян
Головной мозг и анализаторы человека и обезьян
В ходе эволюции животного мира появилась и развивалась нервная система,
наибольшего уровня достигшая у позвоночных животных с их сложным головным
мозгом и в
особенности у высших млекопитающих.
Теория Дарвина и в этот вопрос внесла большую ясность. О важности эволюционной
трактовки строения и жизнедеятельности человеческого мозга в свете дарвиновского
учения Энгельс писал:
«В основных чертах установлен ряд развития организмов от немногих простых
форм до все более многообразных и сложных, какие мы наблюдаем в наше время,
кончая человеком. Благодаря этому не только стало возможным объяснение существующих
представителей органической жизни, но и дана основа для предыстории человеческого
духа, для прослеживания различных ступеней его развития, начиная от простой,
бесструктурной, но ощущающей раздражения протоплазмы низших организмов и
кончая мыслящим мозгом человека. А без этой предыстории существование мыслящего
человеческого мозга остается чудом» (Маркс и Энгельс. Сочинения, т. 20, стр.
512).
Для понимания процесса развития головного мозга и анализаторов у предков
человека в ходе филогенетической эволюции первостепенное значение имеет рефлекторная
теория И. П. Павлова, одной из основ которой является учение Дарвина. Отметим
в связи с этим, что и для современного дарвинизма продолжают быть важнейшими
следующие вопросы: об объеме влияния и воздействия условий внешней среды
на организм, в особенности на ранних стадиях его индивидуального развития;
о передаче приобретенных признаков по наследству; о механизме возникновения
новых видов в результате качественного преобразования на основе количественного
накопления анатомо-физиологических изменений в организме; о наличии и объеме
межвидовой и внутривидовой борьбы в мире растений и животных.
Что касается животных, то необходимо специально учитывать
199
развитие нервной системы, которая осуществляет особые связи организма с
окружающей средой и играет все более возрастающую роль в ходе эволюции, в
процессе беспрерывного
приспособления к постоянно изменяющимся условиям жизни. Физиологическое учение
И. П. Павлова, его рефлекторная теория заставляют нас по-новому понять связи
организма со средой.
«Окружающий животное внешний мир, вызывая, с одной стороны, беспрерывно условные
рефлексы, с другой стороны, так же постоянно подавляет их, заслоняет другими
жизненными явлениями, в каждый данный момент более отвечающими требованию
основного закона жизни — уравновешиванию окружающей природы»,— пишет И. П.
Павлов (Полн. собр. соч., т. III, кн. 1, стр. 129), подразумевая под словом
«уравновешивание» беспрерывное приспособление организма к среде. И далее:
«Таким образом, большие полушария являются органом анализа раздражений и
органом образования новых рефлексов, новых связей. Они — орган животного
организма, который специализирован на то, чтобы постоянно осуществлять все
более и более совершенное уравновешивание организма с внешней средой,— орган
для соответственного и непосредственного реагирования на различнейшие комбинации
и колебания явлений внешнего мира, в известной степени специальный орган
для беспрерывного дальнейшего развития животного организма» (там же, стр.
273).
И. П. Павлов установил, что в больших полушариях находятся
центральные концы анализаторов, с одной стороны связанных с восприятием раздражений,
которые поступают извне: будучи представлены на периферии глазами, ушами,
кожным покровом, носом, ртом, анализаторы, с другой стороны, связаны с импульсами,
поступающими из скелетной мускулатуры и внутренних органов.
Развитие анализаторов в эволюции приматов было теснейшим образом связано
с преобразованием коры большого мозга, где находятся их центральные зоны,
и перестройкой самого головного мозга в целом. Сведения об общих чертах развития
анализаторов обезьян, шедшего под влиянием природных факторов, позволяют
яснее представить себе значение отличий человеческого мозга (рис. 55). Можно
полагать, что они сформировались под главнейшим воздействием труда и звукового
языка.
Энгельс пишет: «Сначала труд, а затем и вместе с ним членораздельная речь
явились двумя самыми главными стимулами, под влиянием которых мозг обезьяны
постепенно превратился в человеческий мозг, который, при всем своем сходстве
с обезьяньим, далеко превосходит его по величине и совершенству. А параллельно
с дальнейшим развитием мозга шло дальнейшее развитие его ближайших орудий
— органов чувств» (Диалектика природы. Маркс и Энгельс, Сочинения, т. 20,
стр. 490).
Сильное развитие головного мозга человека было первоначально обусловлено
анатомо-физиологическими прогрессивными особен-
200
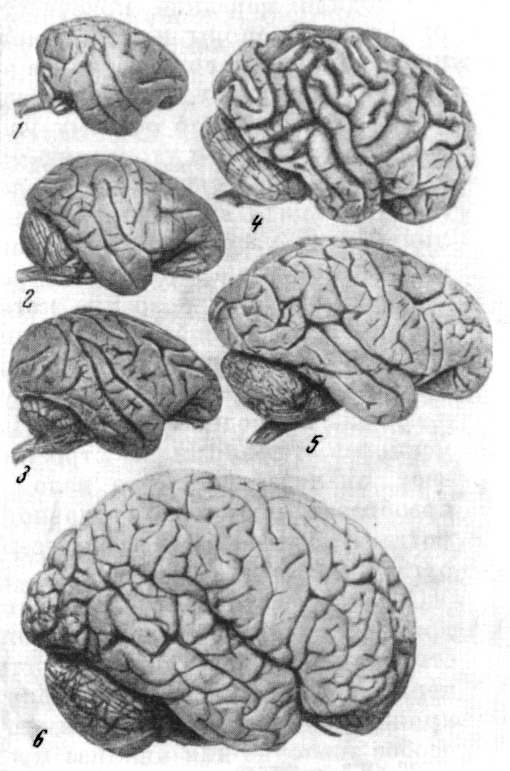
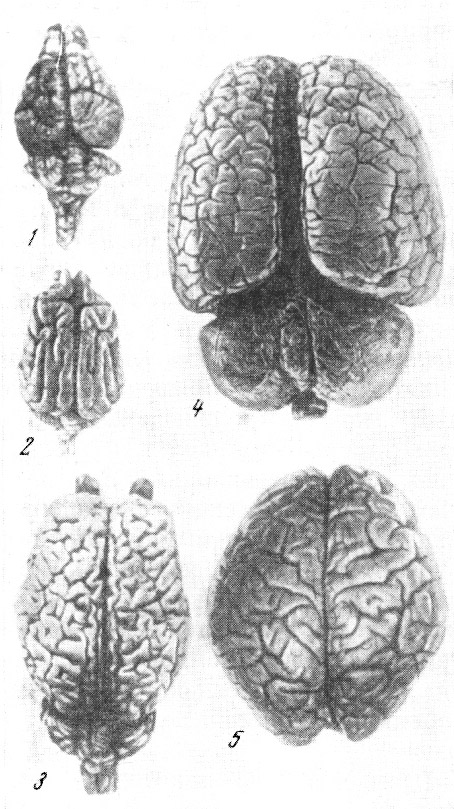
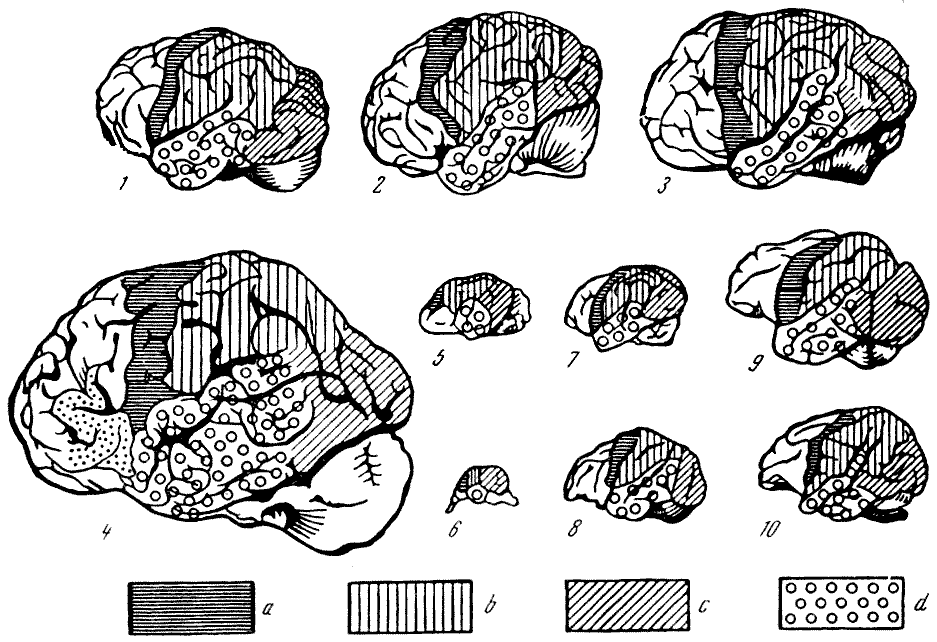
Рис. 55. Головной мозг
обезьян и человека 1 — цепкохвостая обезьяна коата
Жоффруа (Ateles geoffroyi Hasselt und Kuhl); 2 — павиан гамадрил
(Cynocephalus hamadryas Fr. Cuvier); 3 — быстрый гиббон (Hylobates
agilis Fr. Cuvier); 4 — орангутан (Simla satyrus Linnaeus);
5 — обыкновенный шимпанзе (Pan troglodytes Blumenbach); 6 —
современный человек (Homo sapiens Linnaeus). Вид справа. По
Т. Моллисону, 1932
|
ностями мозга его ближайших предков — древних обезьян. Большое значение
при этом имели влияние нового для них наземного образа жизни, переход к прямохождению
и животной пище, усиление стадного образа жизни, наконец, освоение орудий
в виде простого употребления камней и палок при добывании пищи и обороне
от хищников. Как уже отмечалось выше, при вертикальном положении тела очень
уменьшилась нагрузка шейных мышц, которая была максимальной у более древних
четвероногих предков человека, когда голова поддерживалась в горизонтальном
положении. Затылочный гребень, служащий одним из важных пунктов прикрепления
шейных мышц, ослаблялся в своем развитии, становился ниже и глаже. Уменьшился
и весь наружный рельеф черепной коробки, что явилось важным условием прогрессивного
развития, в частности увеличения размеров головного мозга и его отдельных
долей. Но, конечно, главное здесь не только и не столько в количественном
разрастании, сколько во внутренней перестройке и усложнении строения мозга
вплоть до тончайших его структур.
Усовершенствование прямохождения и усложнение функций руки как органа добывания
пищи с помощью орудия способствовали прогрессивному развитию головного мозга
и соответственному преобразованию черепа. Но настоящий толчок был дан лишь
с того знаменательного, поворотного момента в человеческой родословной, когда
наши предки перешли к изготовлению орудий, когда зародились подлинные трудовые
действия, когда появились один за другим совершенно новые и могущественные
факторы развития.
Таким образом, прямохождение способствовало, хотя и косвенным образом, развитию
головного мозга у предков человека, в частности, еще и потому, что
201
освободило передние конечности от функции опоры и локомоции, направило
по-новому развитие их хватательных и ударных движений, способствовало особому
усовершенствованию
и утончению органов кожного осязания на ладонях и пальцах в виде папиллярных
линий и их узоров, не только на кистях, но и на стопах.
Изощрялось также зрение, которое помогало лучше заметить пищу или врага,
расширяло горизонт видимости предметов окружающей природной среды. Изменялись
функции и строение всех анализаторов, что вело к преобразованию коры головного
мозга и в известной мере всего организма предка человека.
Следует отметить, что И. П. Павлов подчеркивал важность свойства высочайшей
пластичности нервной системы. Павловские принципы общей биологии имеют особое
значение для анализа процесса филогенетического развития головного мозга
и периферических частей анализаторов, которые сильно изменялись у предков
человека — ископаемых обезьян и еще более древних полуобезьян, испытавших
за 60 млн. лет третичного периода крупные видоизменения.
Изучение анатомо-физиологических особенностей головного мозга требует применения
самых разнообразных методов. Среди них видное место занимают макро- и микроскопические
исследования,в особенности коры больших полушарий, являющихся важнейшей частью
центральной нервной системы (Саркисов, 1964).
В основе отечественной науки о цитоархитектонике коры больших
полушарий мозга человека и животных лежат классические исследования крупного
отечественного анатома прошлого столетия В. А. Беца, открывшего гигантские
пирамидные клетки в том слое коры, который теперь обозначается как пятый
(V), из основных шести слоев, считая от поверхности коры в ее глубину (рис.
56). Бец заложил фундамент учения о функционально-структурной архитектонике
мозга, установил основы для деления коры в сравнительно морфологическом аспекте.
Кардинальная идея взаимосвязи формы и функции плодотворно развивалась далее
в трудах В. А. Бехтерева, Л. Б. Блуменау и позднейших прогрессивных церебрологов.
В настоящее время в СССР головной мозг изучают многие биологи, медики и другие
специалисты на основе физиологического сеченовско-павловского учения. Большая
работа, осуществляемая в Институте мозга, Институте антропологии Московского
университета и других научных учреждениях, позволяет считать, что кора больших
полушарий человека и обезьян плодотворно изучается в историческом, эволюционном
аспекте. В частности, обнаружены важные качественные отличия звездчатых клеток
в третьем слое коры у человека (Поляков, 1964).
В области антропологического исследования мозга немалое значение имеют также
его объем, вес и внешний рельеф. У человека объем головного мозга колеблется
индивидуально в пределах 1000 — 2000 см3. Если же говорить
о труп-
202
новых вариациях, которые, например, касались бы рас или территориальных
антропологических типов, то варианты лежат в пределах 1200—1600 см3. Объем
мозга ниже 1000
см3 у человека встречается редко, но описаны случаи, когда у отдельных индивидуумов
независимо от их расовой принадлежности, ничем особенным не отличавшихся
от окружающих людей, после их смерти оказывалось, что объем их мозга составлял
только 900 или 800 см3, а то и еще меньше.
Такая сильная вариабельность заметно выделяет человека по сравнению с дикими
животными, у которых уклонения в весе или в объеме мозга не выходят из пределов
нескольких процентов. Что же касается домашних животных, то хотя изменчивость
у них и больше, чем у диких, но зато несравнима с человеческой еще и вследствие
уменьшения размеров мозга в неволе у некоторых видов
и пород.
Объем головного мозга и вместимость черепной коробки не равны между собой,
так как между самим мозгом и черепом располагаются не только твердая, мягкая
и паутинная оболочки, но также сосуды и спинно-мозговая жидкость, вследствие
чего получается довольно ощутимая разница. У взрослого человека разница составляет
5—10%, а у старого даже 15%: иначе го-
Рис. 56. Схема послойного строения коры больших полушарий человека
Слои: 1 — молекулярный; 2 — наружный зернистый; 3 — с
малыми и средними пирамидальными клетками; 4 — внутренний зернистый; 5 —
с большими пирамидальными клетками; 6 — с полиморфными клетками; 7 — белое вещество. По Э. Виллигеру, 1930

203
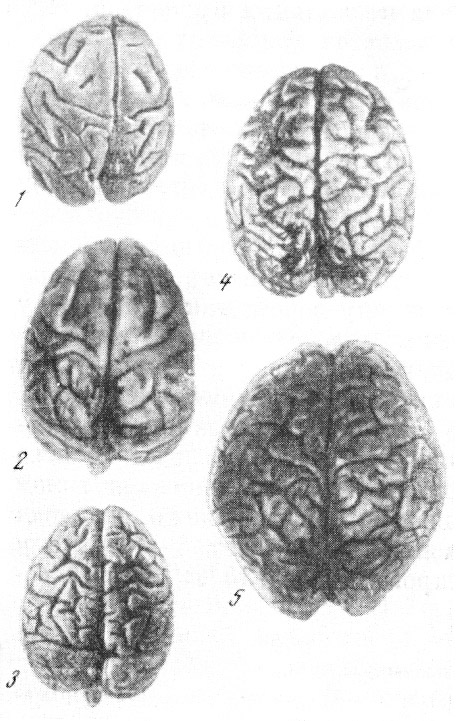
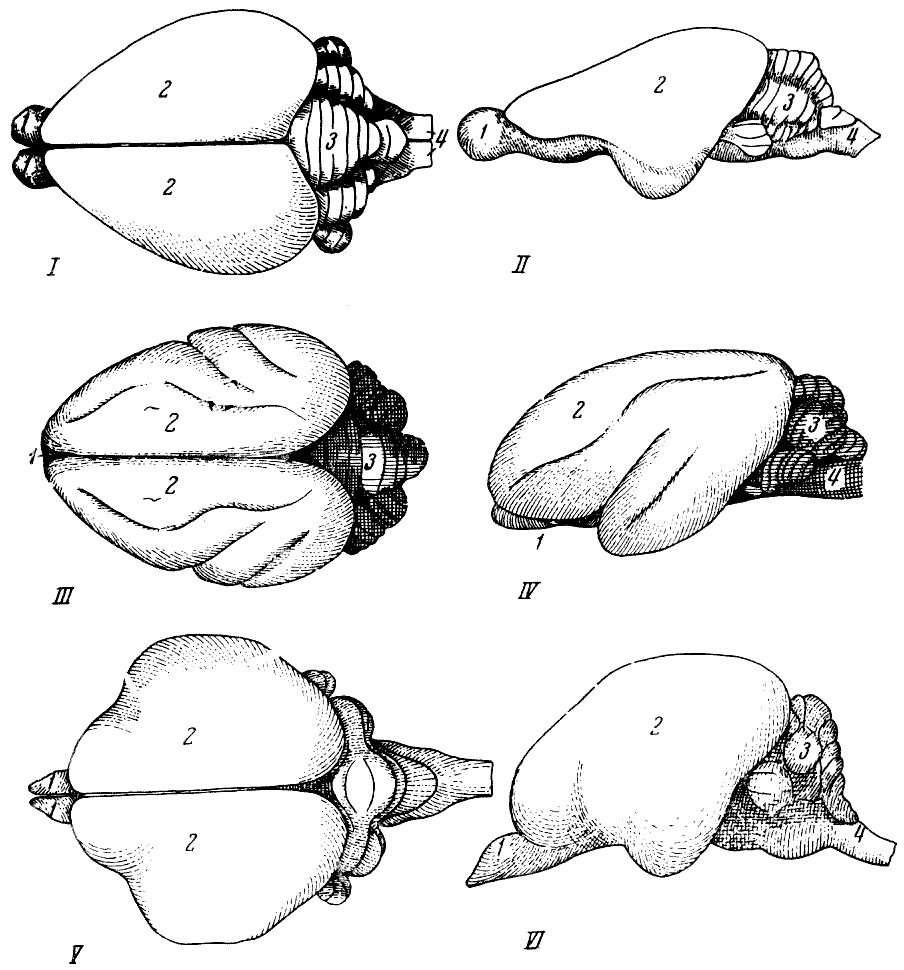
Рис. 57. Головной мозг приматов
1 — павиан; 2 — гиббон; 3 — шимпанзе; 4 — горилла; 5 — человек. По Ф. Тильнею,
1928
|
воря, объемы разнятся соответственно на 100 и даже 200 см3.
По объему мозга (в см3) можно судить о весе, учитывая,
что удельный вес его немного выше, чем у воды. Главная часть массы мозга
приходится на полушария
(87%) и мозжечок (11%). Что касается не абсолютного, а относительного веса
мозга, то и здесь человек занимает видное место по сравнению с человекообразными
обезьянами (рис. 57), превышая в десять раз гориллу, шимпанзе в четыре,
орангутана в шесть и гиббона в два раза.
Правда, относительный вес мозга, т. е. отношение его веса к весу всего тела,
у некоторых американских обезьян больше, чем у человека, у которого он составляет
примерно 1 : 35. Именно, у капуцина обыкновенного отношение составляет 1
: 18, у паукообразной обезьяны черной коаты 1 : 15. Но нужно иметь в виду,
что чем мельче млекопитающее животное, тем обычно относительный, вес его
мозга больше. В свете этой закономерности понятнее становятся и приведенные
выше отношения для человека и антропоидов.
В добавление отметим, что хотя мозг такого огромного животного, как индийский
слон, по абсолютному весу и превышает мозг человека приблизительно в четыре
раза (рис. 58), но по относительному весу (1 : 560) сильно уступает (в
пятнадцать раз).
Таким образом, человек по совокупности абсолютного и относительного весов
занимает столь видное место среди животного мира, что уже ставит людей
выше, любого из названных животных.
Специфические черты человеческого головного мозга заметно обнаруживаются
и в его онтогенетическом развитии (Развитие мозга ребенка, 1965), которое,
с другой стороны, ясно свидетельствует о ближайшем родстве человека с высшими
обезьянами. Сперва развитие коры наступает в средней части полушарий и
идет по принципу радиальности от центра к периферии по поверхности; происходит
дифференциация на слои вплоть до шести. В дальнейшем процесс развития коры
большого мозга плода испыты-
204
Рис. 58. Головной мозг человека и млекопитающих
1 — кенгуру; 2 — собака; 3 — лошадь; 4 — слон; 5 — человек. По Ф. Тильнею, 1928
|
вает коренной перелом: дифференциация формы и строения происходит по
принципам региональности и ареальности, т. е. по пути оформления в составе
коры
областей, полей, подполей вместе с извилинами и бороздами.
Возникновение борозд и извилин следует за глубокими цитоархитектоническими
преобразованиями в новой коре, или неокортексе, составляющем около 95%
объема всей коры у человека.
В начале четвертого (лунного) месяца утробного развития на бывшей до того
гладкой поверхности больших полушарий появляется первая борозда, а именно
ринальная, или обонятельная. Вслед за обонятельной начинает формироваться
сильвиева борозда и появляются некоторые борозды височной, лобной и теменной
долей в средней части латеральной стороны полушария, в районе той же сильвиевой
борозды (рис. 59).
После сильвиевой борозды, как и у антропоидов, на поверхности будущей теменной
доли появляется центральная, или роландова, борозда, между тем как у низших
обезьян позже сильвиевой возникает так называемая параллельная борозда,
соответствующая верхней височной по терминологии анатомии человека. Затем
в течение пятого
(лунного) месяца на больших полушариях человеческого плода появляются и
некоторые другие борозды. В следующем месяце края сильвиевой ямы начинают
смыкаться,
центральная доля постепенно погружается на ее дно. В ходе зародышевого
развития на поверхности полушарий последовательно формируются борозды,
характерные
для животных, у которых поверхность полушарий более или менее покрыта извилинами,
т. е. животных гирэнцефалических, в отличие от животных с гладким мозгом,
т. е. лиссэнцефалических.
Особенности процесса онтогенетического развития мозга зародыша человека
свидетельствуют о ранее пройденных нашими предками этапах филогенетического
развития начиная
с низших представителей типа хордовых животных.
205
Ко времени рождения большой мозг плода уже характеризуется извилинами и
бороздами в такой мере, что по характеру их расположения и развития вполне
может служить
прототипом для мозга взрослого: ему предстоит почти двадцатилетний путь
по линии увеличения размеров и усложнения узора на поверхности больших
полушарий
за счет появления дополнительных борозд второго и третьего порядка (рис.
60— 62). У плацентарных млекопитающих животных детеныши появляются на свет
с достаточно сформированным мозгом, и вскоре после рождения он перестает
у них увеличиваться. Таким образом, у животных рано в целом заканчивается
процесс онтогенетического развития головного мозга, или церебрализация,
а человек составляет, следовательно, своеобразное исключение из закона
ранней
церебрализации. Это обнаруживается даже при сравнении головного мозга человека
и обезьяны по весу и объему,
так как у ребенка они равны примерно лишь четверти, а у детеныша обезьяны
— половине или даже двум третям по сравнению со взрослыми формами.
Мозг крупной человекообразной, обезьяны вроде орангутана достигает значительного
веса в течение первых двух-трех лет послеутробного развития, но дальше
увеличивается немного. Возрастные изменения веса мозга у человека были
изучены В. В. Бунаком
(1936), который, применив сложный метод и произведя специальные расчеты,
пришел к заключению, что представления о наибольшей церебрализации младенца
оказались ошибочными, так как, относительная «масса мозга достигает максимальной
величины в, возрасте 3—5 лет, после чего постепенно и очень немного снижается»
(стр. 298) в связи с более интенсивным нарастанием массы тела.
Приведем еще некоторые данные по соотношению веса мозга и веса всего тела
в ходе развития
Рис. 59. Развитие головного мозга человека
1 — плод трех месяцев; 2 — пяти месяцев; 3 — семи месяцев; 4 — новорожденный. Вид слева
По слепкам
коллекции Музея антропологии (Москва)
206
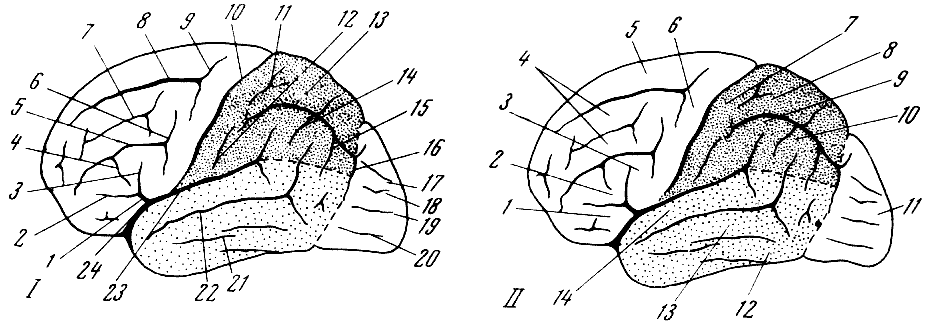
Рис. 60. Боковая поверхность большого полушария мозга человека
I — борозды, II — извилины. Слева не заштрихована лобная доля; густыми
точками заштрихована теменная, редкими точками височная; справа не
заштрихована затылочная
доля. 1, 2 — передняя горизонтальная ветвь начальной части боковой
мозговой фиссуры (сильвиевой); 3 — передняя восходящая ветвь 1 и 2;
между линиями
к 3 и 4 — лобный полюс; 4 — радиальная борозда; 5 — нижняя лобная
борозда; 6 — прецентральная нижняя борозда; 7 — средняя лобная борозда;
8 —
верхняя лобная борозда; 9 — прецентральная верхняя борозда; 10 — центральная,
или
роландова, борозда; 11, 12 — постцентральная борозда, 13 — среднетеменная,
или интерпариетальная, борозда; 14 — первая промежуточная борозда от
13 (с другой стороны 13 напротив отходит теменная борозда), между 14 и 13 на краю
(между теменной и затылочной долей) видна теменно-затылочная фиссура;
15 — вторая промежуточная борозда; 16 — поперечная затылочная; 17, 18,
19,
20 — верхние и боковые затылочные борозды; справа вниз от 20 — затылочный
полюс;
между затылочной и височной долями на нижнем краю видна предзатылочная
вырезка, на височной доле видна нижняя височная борозда; 21 — средняя
височная борозда;
22 — верхняя височная борозда (ее передний конец, будучи продолжен
вперед налево, упирается в височный полюс); 23 — задняя ветвь боковой
фиссуры
мозга (сильвиевой); 24 — ствол сильвиевой фиссуры.
II — извилины: 1, 2, 3 — нижняя лобная: глазничная треугольная, оперкулярная
части; 4, 5 — средняя и верхняя лобные; 6, 7 — передняя и задняя центральные; 8 — верхняя теменная; 9, 10 — верхнекраевая и угловатая; 11 — нисходящая;
12, 13, 14 — нижняя, средняя и. верхняя височные. По Э. Виллигеру, 1930
человека,
сообщаемые Дж. Таннером (Харрисон, Вейнер, Таннер и Барникот, 1968).
Во-первых, головной мозг по весу обнаруживает уже с раннего этапа
зародышевого развития наибольшую близость к окончательной его массе
(в этом отношении ему вряд ли сильно уступает лишь глаз). Из этого проистекает
более
быстрое развитие мозга, чем остальной части тела. У новорожденного
вес
мозга равен 25% от взрослого состояния, в полгода — 50, в два с половиной
— 75,
в пять лет — 90
и в десять — 95 %, между тем как вес остального тела при рождении равен
5% по сравнению с состоянием половозрелой юности, а в десять лет —
50%.
В ходе возрастного развития все ярче обнаруживаются половые различия
в весе мозга; у взрослого, мужчины он достигает в среднем: 1400 г,
а у взрослой женщины — 1270, т. е. меньше на 130 г.
Следует добавить, что относительный вес женского мозга в среднем несколько
больше, так как мужчины на 8—9 кг тяжелее жен-
207
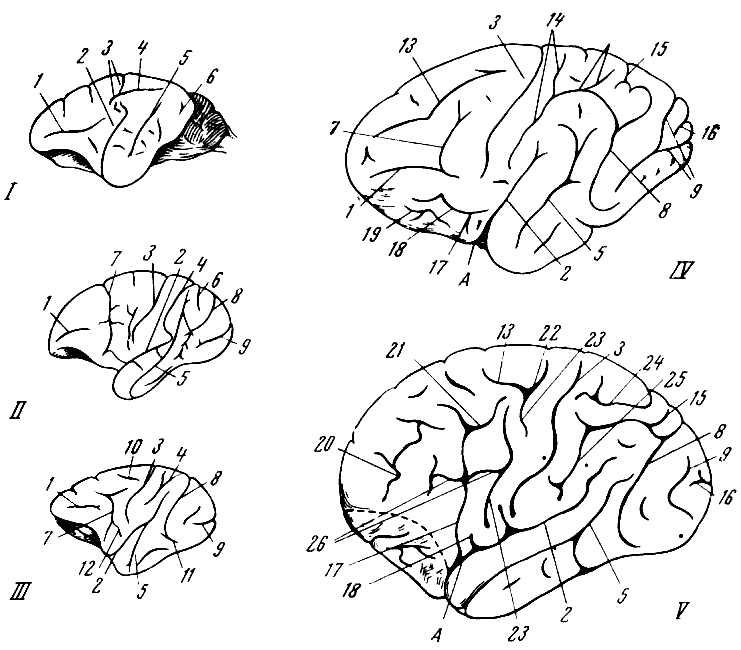
Рис. 61. Расположение борозд на больших полушариях мозга приматов
I — черный лемур (Lemur macaco Linnaeus); II — бурый
капуцин (Cebus fatuellus Linnaeus); III — свинохвостый макак (Macacus
nemestrinus Linnaeus);
IV — гиббон хулок (Hylobates hoolock Harlan); V — шимпанзе
(Antropopithecus troglodytes
Blumenbach). A — наружный участок центральной доли; 1 —
борозда прямая; 2 — сильвиева; 3 — центральная; 4 —
интерпариетальная, или интрапариетальная;
5 — параллельная; 6 — поперечная затылочная; 7 —
нижняя прецентральная, или
дуговидная; 8 — обезьянья; 9 — боковая затылочная; 10 —
верхняя прецентральная;
11 — нижняя затылочная; 12 — нижняя поперечная; 13 —
верхняя лобная;
14 — постцентральная; 15 — парието-окципитальная;
16 — шпорная; 17 — фронтоорбитальная; 18 —
верхняя ограничивающая;
19 — орбитальная; 20 — фронто-маргинальная
Вернике;
21 — средняя лобная; 22 — верхняя прецентральная;
23 — нижняя прецентральная; 24 — добавочная и
интрапариетальная;
25 — нижняя постцентральная;
26 — нижняя лобная борозда. По Ч. Зоннтагу, 1924
щин в соответствии с тем, что по длине тело мужчины в среднем превышает
тело женщины сантиметров на десять. То же справедливо и для таких антропоидов,
как горилла или же орангутан, у которых самец гораздо тяжелое самки,
и относительный
вес мозга последней соответственно выше, чем у самца.
К числу специфических особенностей человеческого мозга относится еще
заметная асимметричность формы и строения больших полушарий. Так,
у правшей более
развито и крупнее левое полушарие и там находится двигательная речевая
зона. Мозг обезьян далеко не так асимметричен, хотя по отношению
к че-
208
Рис. 62. Мозг низших приматов
I, II — малая тупайя (Tupaia minor Gunther); III, IV — черный лемур (Lemur
macaco Linnaeus); V, VI — долгопят привидение (Tarsius spectrum Storr).
1 — обонятельные лопасти; 2 — лобные доли; 3 — мозжечок; 4 — продолговатый
мозг.
По Ф. Вуду Джонсу, 1929
ловекообразным обезьянам отмечались случаи преимущественного развития
левого полушария, которое оказалось, например, заметно длиннее
правого на мозге
молодого самца орангутана Морица (Московский зоопарк, 1933 г.).
Из подобных фактов, устанавливаемых при изучении размеров мозговой
полости человека и
обезьян, можно сделать вывод, что человек по признаку асимметричности
головного мозга стоит не совсем одиноко. Очевидно, такая особенность
в более слабой
степени была свойственна и ближайшим предкам человека — ископаемым
антропоидам конца третичного периода вроде австралопитеков. Возможно,
что более резкая
асиммет-
209
рия мозга у людей развилась в коррелятивной связи с усовершенствованием
прямохождения, освобождением рук, преимущественным применением
правой руки в трудовой деятельности
гоминид.
Развитие периферических частей
анализаторов
Полагают с достаточной уверенностью, что ближайшими предками приматов
были наземные мелкие насекомоядные млекопитающие конца мелового
периода мезозойской
эры, жившие более чем 60 млн. лет назад. У предков приматов орган
обоняния был развит сильно, о чем свидетельствуют удлиненная мордочка
и обилие
носовых обонятельных раковин, с другой же стороны, крупные обонятельные
участки
головного мозга, иначе говоря периферический и центральный отделы
обонятельного анализатора,
как у современных насекомоядных, очень хорошо были сформированы
и прекрасно функционировали.
Обонятельный анализатор у обезьян развит сравнительно слабо, и
его периферический отдел — наружный нос — редко бывает сильно
выражен; у человека слабое обоняние
сочетается с крупным носом, имеющим значительную опору из костей
и хрящей.
Развитие наружного носа человека, являющегося по своей форме и
строению особым, специфическим образованием, в значительной мере
зависит от
разрастания в
вышину верхнего отдела верхнечелюстных костей, перпендикулярной
пластинки решетчатой кости и сошника, а также от отступания альвеолярной
части верхней челюсти. Эти особенности отсутствуют у обезьян, даже у антропоморфных.
Нос гориллы или гиббона, характеризующийся
хорошо
развитым хрящевым остовом, обладает едва ли не наибольшим сходством
с человеческим.
По Веберу (1936) носовой скелет состоит из тех же костей и хрящей,
как и у человека; отсутствие выдающегося носа у обезьян обусловливается
сильным
развитием верхней челюсти, малой шириной наружного костного носового
отверстия, формой носовых хрящей, небольшой величиной и малым выступанием
носовых
костей.
Сказанное имеет значение также для живущего на острове Борнео носача
обыкновенного, принадлежащего к подсемейству тонкотелых обезьян.
Его крупный, мягкий,
подвижный, длинный нос относится больше к категории хоботных образований.
На кончике
он как бы раздвоен продольной глубокой бороздой и способен раздуваться
при гневе и половом возбуждении. У старых экземпляров нос отвисает
ниже подбородка.
К числу носатых обезьян относятся еще и некоторые другие тонкотелые
обезьяны. Таковы, например, немейский тонкотел с острова Хайнаня
и из северной (верхней)
Кохинхины, а также одноцветный симиас с южного острова Пагай недалеко
от Суматры с их заметно выраженными носами. К роду носатых обезьян
ринопитеков,
обладающих коротким носом со вздернутым кончиком, относятся виды:
рокселланов ринопитек из северо-западного Китая; биэтовский из
западнокитайской провинции Юннань и юго-восточ-
210
ного Тибета; бреличевский из центрального Китая и юго-восточного
Тибета; авункулярный из Тонкина, северный Вьетнам.
У очень многих обезьян наружный нос весьма невелик, что относится,
в частности, и к таким человекообразным обезьянам, как орангутан
и шимпанзе. По отношению
к обоим последним добавим, что их череп в области носового (грушевидного)
отверстия обнаруживает заметную вогнутость — явление симогнатизма.
Хрящевой остов человеческого носа состоит из срединно-продольного
хряща носовой перегородки, хрящей боковых стенок носа (лежащих
с каждой стороны
между носовой косточкой и большим крыльным хрящом), а также ряда
маленьких хрящиков.
В глубине носовых полостей на их боковых стенках располагаются
три носовые раковины (нижняя, средняя и верхняя). У новорожденных
имеется
еще четвертая
носовая раковина, или санториновая, которая позже в 50—75% случаев
исчезает.
Наличие 5-й раковины у человека отмечается очень редко. Случаи
сохранения 4-й и даже 5-й раковин свидетельствуют о том, что его
предки имели
более активно функционировавший орган обоняния. Ныне же у человека
к обонятельной
области относится лишь верхняя носовая раковина. Редукцию обонятельного
органа по линии носовых раковин человек разделяет с обезьянами
и долгопятом как
микросматическими приматами, в отличие от лемуров и гупаий, имеющих
более развитые органы обоняния, т. е. макросматических приматов.
Тонкое различение запахов как признаков вещей теснейшим образом
связано со способностью образовывать условные рефлексы, сильно выраженной
у птиц и еще больше
у млекопитающих.
Параллельно с регрессом обонятельного анализатора у обезьян и человека
шла также редукция специальных осязательных органов на лице: оба
эти процесса были связаны между собой и с общим укорочением лицевого
отдела,
особенно
ринариума. Среди низших приматов у полуобезьян на мордочке развивается
до
четырех парных и одного непарного пучков жестких и длинных волос,
которые называются вибриссами и представляют собой специализированные
осязательные
волосы (рис. 63—64), У их основания в коже располагаются особые
нервные окончания и небольшие скопления крови в виде лакун, или
синусов.
Малейшее прикосновение
кончика вибриссы к предмету тотчас вызывает колебания в кровяном
синусе, и по нерву в мозг животного передается сигнал, дающий представление
о характере поверхности данного предмета.
От первоначальных пучков вибрисс у низших обезьян остались только
три парных: в области бровей, на верхней губе и подбородке, как
и у антропоидов,
а
у человека нет ни одного. Даже в ходе эмбрионального развития человека
отдельные
зачатки подобных волос с синусами на лице зародыша появляются лишь
изредка.
С древесным образом жизни в органах осязания у приматов коррелятивно
связаны не только регрессивные, но и прогрессивные изменения. Так,
на ладонях и
по-
211
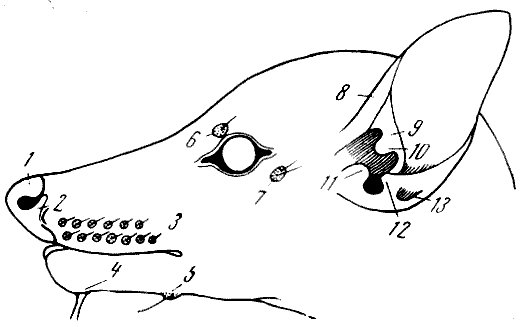
Рис. 63. Пучки вибрисс и ушная раковина мадагаскарского
лемура хирогале — Chirogale E. Geoffrey (схема)
1 — срединный носовой выступ; 2 — боковой носовой выступ; 3 — верхнечелюстной
пучок вибрисс (vibrissae mysticiales); 4 — подбородочный пучок (vv. submentales);
5 — пучок между ветвями нижней челюсти (vv. interramales); 6 — надглазничный
пучок (vv. supraobitales); 7 — заглазничный пучок (vv. genales); 8 — завиток
ушной
ркковины (helix); 9 — противозавиток (antihelix); 10 — выступ противозавитка
(processus antihelicis); 11 — козелок (tragus); 12 — противокозелок (antitragus);
13 —
углубление (bursa). По Ф. Вуду Джонсу, 1929
Рис. 64. Руконожка мадагаскарская (Chiromys madagascariensis
E. Geoffrey). На мордочке видны пучки вибрисс
По Г. Грандидие и Г. Пети, 1932
|
дошвах у обезьян вместо отдельных подушечек, свойственных многим
низшим млекопитающим, вся поверхность занята папиллярными линиями,
которые
сочетаются с флексорными
(рис. 65).
Папиллярные линии снабжены массой нервных окончаний, позволяющих
кисти или стопе получать тончайшим образом суммированные осязательные
раздражения
и
реагировать на соприкосновение с ветвями, плодами и другими предметами
окружающего мира. Токи папиллярных линий образуют в местах схождения
определенные узоры
в виде трирадиу-сов на самой ладони или подошве, кругов, петель,
дельт — на последних фалангах пальцев. Направление флексорных линий
в продольном
или поперечном направлении зависит от привычного способа складывания
кистей или стоп у особей данного вида обезьян и у каждого отдельного
индивидуума.
Сочетание узоров папиллярных и флексорных линий на кистях у любого
человека очень сложно и неповторимо. Оно представляет его антропологическую
подпись,
дактилоскопическое факсимиле. Изучение подобных узоров представляет
особый интерес для антропологии и судебной медицины (Гладкова,
1966). Развилась
даже ссо-бая отрасль науки — дактилоскопия: по отпечаткам пальцев
на предметах в помещении, в котором было совершено преступление,
зачастую
можно без
особого
труда установить личность преступника.
Отметим, что дерматоглифический узор линий представляет собой
результат филогенетического развития вида и показывает ход
212
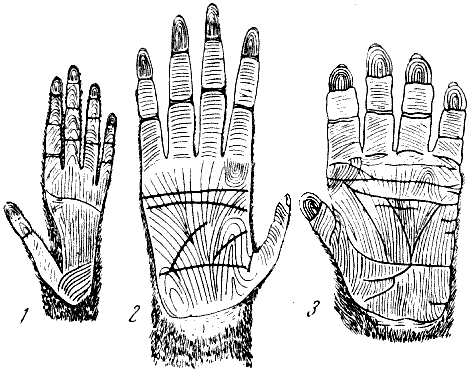
Рис. 65. Сгибательные (флексорные) и сосочковые (папиллярные) линии
на ладонной поверхности кистей антропоидов (схема)
1 — гиббон; 2 — орангутан; 3 — горилла. По Ч. Зоннтагу, 1924
|
видовой эволюции, путь приспособления приматов к древесному образу
жизни, а у человека — процесс преобразования его организма под
влиянием трудовой
деятельности. Изменения формы узоров при увеличении тонкости восприятия
характеризуют особый путь эволюции осязательного анализатора человека
по сравнению с обезьянами.
Следует особенно подчеркнуть, что тонкость осязания на кистях предков
человека — обезьян — оказалась несомненно одной из очень важных
предпосылок к правильному
использованию орудий и их изготовлению. Но справедливо и обратное:
трудовая деятельность привела на протяжении сотен тысяч лет к дальнейшему
усовершенствованию
человеческой руки. Происходило ее качественно особое развитие как
комплексного органа, теснейшим образом связанного в своей деятельности
с рядом анализаторов
—
кинестезическим, кожным в сочетании со зрительным и другими. Размеры,
форма, вес каменных орудий и других предметов оценивались человеком
по-новому
в сравнении с теми тактильными восприятиями, которые развивались
в ходе эволюции
его предков — обезьян, манипулировавших на деревьях плодами, ветвями.
И если изменения обонятельного анализатора происходили под воздействиями
внешней среды, то в неменьшей степени то же надо сказать и об осязательном,
потому что лазанье по деревьям с помощью обхватывания ветвей привело
к образованию не только хватательных стоп с ногтями, а также и
хватательных кистей тоже
с ногтями. Но это было органически связано и с мощным развитием
осязательных нервных телец в составе кожи подошв и ладоней. Все
преобразования
конечностей обеспечивали необходимую координацию и прочность сложного
и нередко
весьма
рискованного передвижения по ветвям деревьев.
Сохранение равновесия при лазаньи по деревьям и перепрыгивание
с ветки на ветку является для обезьян первейшей задачей, выполнение
которой
обеспечивается также благодаря особому развитию органов равновесия,
прежде всего вестибулярного
аппарата внутреннего уха и мозжечка. С этими органами координируются
действия кистей и стоп.
Участие конечностей в локомоции было теснейшим образом связано
со зрительным анализатором, испытавшим сильное прогрессивное развитие.
Укорочение мордочки
у древнейших приматов ти-
213
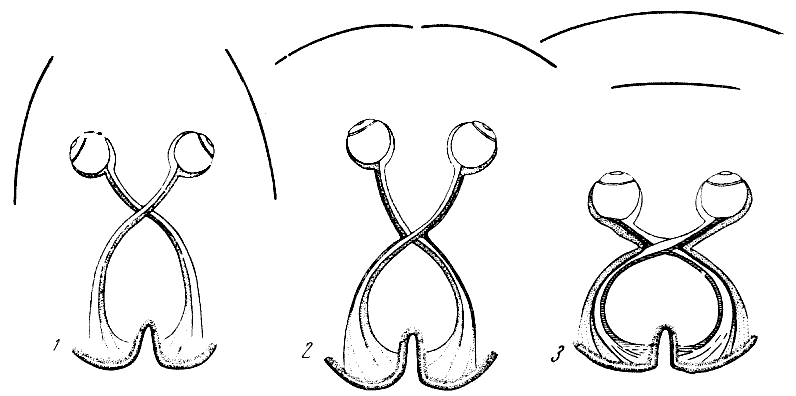
Pис. 66. Схема зрительного перекреста у разных представителей
млекопитающих животных
1 — с независимым движением глаз и раздельными монокулярными полями
зрения (лошадь и т. п.), полный перекрест зрительных нервов;
2 — с согласованными
движениями глаз и комбинированными монокулярными полями зрения
(заяц и др.), почти полный перекрест; 3 — с полностью согласованными
движениями
глаз и
реакциями зрачков, с бинокулярным ясно различимым полем зрения
и бимакулярным полем фиксации зрения (половинный перекрест
ретинных и макулярных волокон
зрительных нервов, т. е. от правых половин сетчаток волокна
идут в правое
полушарие, а из левых — в левое у обезьян и человека). По Ф.
Вуду Джонсу и О. Д. Портеусу, 1929
па полуобезьян позволило внешнему участку зрительного анализатора
переконструироваться. Зрительные поля, ранее раздельные, теперь
вследствие перехода глаз
из бокового положения вперед стали все более перекрывать друг друга
(рис. 66). Другими
словами, у приматов возникло и прогрессивно развивалось бинокулярное,
или стереоскопическое, зрение, которое позволило им видеть предметы
рельефными:
объемное зрение приматов обогатилось еще и за счет все более четкого
различения красочности тропического леса — их среды обитания.
Изменения анализаторов — обонятельного, осязательного и зрительного
— у приматов зависели от сложной совокупности факторов и влияний
внешнего мира,
но одним
из важнейших несомненно
было то, что стал другим способ питания. Вместо преследования различных
насекомых и их личинок в мире обильно стелющихся по земле разнообразных
запахов, как
это свойственно и современным насекомоядным млекопитающим, древнейшие
приматы, еще во многом сходные с насекомоядными и, вероятно,
похожие на современных
тупаий (Вебер, 1936), начали все больше и больше употреблять
в пищу ягоды, орехи и другие плоды, почки, листья, цветы, соки растений.
Цвет растительной
пищи, надо думать, приобретал важное значение хотя бы вследствие
того, что между окраской плода и степенью его съедобности уже
для полуобезьян
должна
была устанавливаться необходимая связь, имеющая первостепенное
жизненное
значение.
214
Современные обезьяны, во всяком случае по сравнению с их отдаленными
предками — полуобезьянами, обладают сильно развитым зрительным
анализатором. Глазницы
обезьян обращены более или менее вперед, и на глубине сетчатки
имеется даже участок особо острого зрения в виде так называемого
желтого
пятна с центральной
ямкой: сила зрения обезьян очень велика, они видят предметы окружающего
мира объемными и цветными.
Острота зрения у обезьян по сравнению с прочими млекопитающими
превосходна в том смысле, что позволяет им наиболее полно усматривать
объемность,
или трехмерность, предметов: у них развилось стереоскопическое
зрение. У наших
сородичей — обезьян — глаза обращены почти прямо вперед, а у
большинства других млекопитающих они обращены под большим или
меньшим углом
в стороны; у лемуров и долгопятов глаза обращены вперед по углом
около
45°, у тупаий
они обращены в стороны еще сильнее. В ходе эволюционного развития
у предков современных обезьян глаза сдвигались из бокового положения
вперед, и обезьяны
способны видеть находящиеся перед ними предметы объемными, рельефными.
Развитие объемного зрения у человека было обусловлено всем предыдущим
ходом эволюции его предков — обезьян — во времена третичного
периода. У обезьян
и у человека развитие объемного зрения является одной из самых
характерных черт их эволюции. У более отдаленных предков, как
и у современных
низших
млекопитающих, зрительные нервы испытывали полный перекрест:
правый глаз снабжался левым зрительным нервом, а левый глаз —
правым.
У обезьян и у
человека левый нерв снабжает лишь левую половину левого глаза,
другая же половина
волокон этого нерва идет в левую половину правого глаза, а правый
нерв снабжает правые половины обоих глаз: получается половинный
перекрест зрительных нервов.
Как сказано, близ места вхождения зрительного нерва на сетчатке,
на дне глазного яблока, у обезьян и у человека есть так называемое
желтое
пятно
с центральной
ямкой — это место наиболее острого зрения. Благодаря комбинированному
восприятию оптических впечатлений сразу двумя глазами, обращенными
вперед, и формированию
единого зрительного поля получается рельефное, точное изображение
предмета. Подобных желтых пятен с центральными ямками у других
млекопитающих не встречается. У человека желтое пятно имеет более
простое строение,
так как из числа
нервных специализированных окончаний здесь имеются лишь немногочисленные
колбочки,
а у обезьян есть и палочки. Кроме того, обе формы окончаний у
обезьян более тонки и многочисленны.
Итак, человек обязан своим бинокулярным зрением прежде всего
своим предкам — обезьянам. Им же он обязан и тем, что видит мир
красочным.
Человек, обладая
трехцветным зрением, близок именно к обезьянам Старого Света.
Шимпанзе хорошо различают синий и зеленый цвета, но значительно
хуже, чем
человек, воспринимают
215
желтый и красный, уподобляясь в этом отношении мартышкам. Между
тем капуцины, по-видимому, не замечают красного цвета и обладают
двуцветным
зрением вроде
человека, больного протанопией.
Сильная острота зрения у обезьян в соединении с бинокулярностью
и цветностью является биологически весьма выгодной при их жизни
в условиях
тропического
леса с его множеством хищных и ядовитых животных, разнообразных
паразитов и колючек. Когда они чувствуют себя в безопасности,
не заняты едой,
отдыхают после путешествия по лесу, начинается искание друг у
друга в шерсти колючек,
паразитов и тому подобных включений. Обычно одна обезьяна ложится
или становится в удобную для искания позу и остается внешне пассивной,
в то время как
другая, близко наклонившись, ищет в шерсти и на открытых местах
кожного покрова.
Тщательно перебирая прядь за прядью, обезьяна время от времени
схватывает пальцами и ногтями насекомое или иной посторонний
предмет, находящийся
в шерсти или на коже.
Насекомые бывают весьма мелки и назойливы, избавление от их присутствия
на теле составляет благодеяние для тропических животных и для
людей. Достаточно вспомнить приводимые разными путешественниками
описания
их длительных и
нередко
мучительных странствований по Африке, где одним из главнейших
препятствий были мириады всевозможных мелких жалящих и колющих
насекомых, в
том числе муравьев. Увидеть и поймать мелкое насекомое,
спасающееся быстрым бегством или ловкими прыжками, не так просто.
Это требует острого зрения, большой быстроты и координации движений,
хорошо
налаженной
связи в мозгу между двигательной зоной передней конечности и
зрительной зоной.
Нужно отметить, что термин «искание блох» мало отражает действительное
содержание поисков обезьян в шерсти друг у друга. Возможно предполагать,
что здесь.
большую роль играет удаление слабых или уже выпавших волос,
от которых тотчас же поедаются обезьяной луковица и основание волоса
(вместе
с находящимися здесь мельчайшими кристалликами веществ, входящих
в состав пота).
Бинокулярное зрение прогрессировало у предков человека — обезьян
— параллельно с развитием более тонких хватательных движений
передней конечности
и с усовершенствованием способности большого пальца кисти противопоставляться
остальным пальцам.
Манипулирование различными съедобными предметами, поворачивание
их перед глазами и рассматривание с разных сторон, откусывание
частей
плода и рассматривание
надкусанных частей, поимка и рассматривание мелких паразитов
— все подобные действия должны были содействовать в процессе
естественного
отбора комбинированному
прогрессивному развитию двигательного анализатора, передней конечности
и сочетанию движений с контролем зрения еще у обезьян третичного
периода. Трудовые
действия древнейших людей преобразовывали
216
этот процесс и усилили его уже в первые сотни тысяч лет четвертичного
периода, о чем в известной мере можно судить по слепкам мозговой
полости черепа,
или эндокранам. Приготовление орудий немыслимо без особо развитой
хватательной передней конечности — руки, без острого бинокулярного
зрения. А такое сочетание
анатомофизиологических особенностей как раз и имелось у наших
предков.
Следовательно, для возникновения человека и всей его дальнейшей
эволюции было необходимо непрерывное прогрессивное развитие общего
строения
нервно-мышечного аппарата руки, тонкой архитектоники соответствующих
частей центральной
нервной системы и зрительного аппарата. Взаимно влияя друг на
друга, рука и мозг
развивались в процессе общественного труда.
Осязательный и зрительный анализаторы в соединении с двигательным
развивались у древних приматов в сравнении с их наземными предками
по-новому, в
ином качественном состоянии, позволяя приспособляться не только
к смене типа
локомоции и способа питания, но и к условиям межвидовой борьбы,
в частности с такими
опасными животными, как средние и крупные кошачьи хищники, разные
змеи или налетающие с воздуха хищные птицы.
Немалое значение приобретает слуховой анализатор, специализирующийся
в различении разнообразных лесных звуков, имеющих жизненное значение.
Чуткость
органов
слуха очень важна для обезьян не только днем, но и ночью: она
нередко спасает их от подкрадывающихся ночных хищников. Обезьяны
постоянно прислушиваются к звукам голоса, издаваемым другими особями стада.
Некоторые звуки
обезьян
имеют особое
значение сигналов, причем довольно безразлично, какая именно
из них издаст звук, предупреждающий
об опасности.
Большое исследование средств общения у обезьян в стаде осуществила
Н. А. Тих на Сухумской медико-биологической станции Академии
медицинских наук
СССР (Тих, 1950). Работа была произведена на павианах гамадрилах,
у которых оказалось около пятнадцати разнообразных звуков голоса,
выражающих
эмоциональное состояние
обезьян. Тих установила, что звуки, издаваемые обезьянами, тесно
связаны с эмоциональной основой и соответствующими телодвижениями.
Их искусственное
вызывание даже по методу условных, рефлексов натолкнулось на
большие трудности, в частности вследствие стереотипности звукового
выражения
эмоций, связанности,
врожденных (безусловных) звуков с определенной биологической
значимостью. Так, звук оборонительного значения гораздо труднее
сделать пищевым
сигналом, чем звук на пищу. В опытах М. А. Панкратова при этом
наблюдался даже срыв
высшей нервной деятельности у обезьян (Воронин, 1952).
Сюда присоединяется то обстоятельство, что периферический и корковый
отделы слухового анализатора у обезьян, в том числе и человекообразных,
частью
выше дифференцированы, чем у человека. Например, обезьяны слышат
такие высокие тона (около 30 тыс.
колебаний в секунду), какие недо-
217
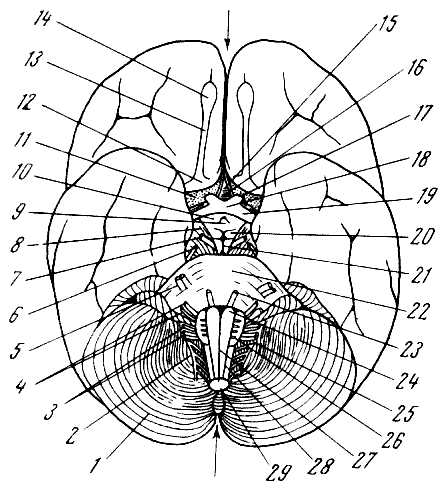
Рис. 67. Головной мозг человека (вид снизу)
1 — мозжечок; 2 — добавочный нерв; 3 —
языкоглоточный и блуждающий нервы; 4 — лицевой и слуховой
нервы; 5 — тройничный нерв; 6 — блоковый
нерв; 7 — глазодвигательный нерв; 8 — ножки
мозга; 9 — воронка; 10 —
серый бугор; 11 — серое переднее продырявленное поле; 12 — обонятельный
треугольник; 13 — обонятельный тракт; 14 — обонятельная луковица
(стрелка между 14 и 15 указывает на продольную фиссуру головного
мозга); 15 — конечная пластинка; 16 — диагональная связка; 17 — глазной
нерв; 18 — зрительный перекрест; 19 — зрительный тракт; 20 —
мозговые шарики;
21 — ямка Тарини между ножками моста; 22 — мост и основная борозда
моста; 23 — отводящий нерв; 24 — пирамида продолговатого мозга;
25 — олива; 26 — подъязычный нерв; 27 — срединная передняя фиссура
продолговатого
мозга; 28 — передняя боковая борозда продолговатого мозга; 29 — продолговатый мозг (в разрезе: стрелка между 29 и 1 — указывает
на заднюю вырезку
мозжечка). По Э. Виллигеру, 1930
|
ступны человеческому невооруженному уху и обнаруживаются только
с помощью специальных акустических аппаратов. Обезьяны обладают
развитыми
ушными
мышцами и свободно двигают сводами ушными раковинами, в то время
как наружное ухо человека при его больших размерах лишено или почти лишено
подвижности, как у гориллы и орангутана, обладающих
маленькими
ушами.
Точно так же в некоторых отношениях у обезьян сильнее развиты
такие анализаторы, как, например, глазной, уже не говоря об обонятельном.
Другими словами,
в области первой сигнальной системы обезьяны даже обладают известным
преимуществом. Зато человек несравненно превосходит их ближайших
сородичей в деятельности
головного мозга, свойственной его анализаторам в связи с работой
второй сигнальной
системы и обеспечивающей познание признаков вещей, недоступное
никакому животному.
Периферические отделы слухового, осязательного, зрительного и
других анализаторов в ходе эволюции приматов под влиянием внешней
среды
находились в неразрывной
связи с корковыми отделами больших полушарий мозга и развивались
вместе с ними. Перейдем к изложению некоторых необходимых данных,
касающихся
соответствующих областей коры.
Тип мозга обезьяны развился из типа мозга более примитивного
примата вроде полуобезьяны. Так, у лемура мозг невелик и весит
не более
нескольких десятков
граммов; его обонятельные части развиты довольно значительно,
а лобные доли слабо; височная, характерная для приматов вообще,
явно
очерчена;
поверхность полушарий у большинства приматов с небольшим количеством
извилин; на затылочной
доле заметна шпорная борозда, имеющаяся и у всех других приматов;
малый мозг, или
218
мозжечок, почти не прикрыт большим мозгом.
В стволовой части головного мозга всех приматов, в том числе
и у человека, подмечаются черты сегментированности, более заметно
обнаруживающиеся
в спинном мозгу. У последнего это ярко проглядывает в попарном
равномерном отхождении
пучков спинных чувствительных и брюшных двигательных нервов от
спинного мозга, а у первого — в симметричном порядке расположения
двенадцати
пар
черепных
нервов. Подобная сегментированность свидетельствует о том, что
не очень
далекими предками человека были четвероногие животные и что
более
отдаленными звеньями
человеческой родословной служили низшие позвоночные. Вместе с
полуобезьянами и обезьянами человек уходит своими корнями в глубину
животного
мира. Его головной мозг имеет в основе исходный обезьяний тип
строения.
Мозг низшей обезьяны вроде макака или павиана крупнее и тяжелее,
чем у лемура, достигая веса 50—100 г; обонятельные луковицы невелики,
а
в большом
мозгу
извилины и другие участки древнего обонятельного отдела коры
мало развиты, хотя и сильнее, чем у человека (рис. 67).
В коре человеческого мозга, по данным исследования И. Н. Филимонова
(Институт мозга, Москва), основная зона, связанная с обонятельной
функцией и обозначаемая
в качестве древней коры, или палеокортекса, составляет лишь 0,61%
площади поверхности коры (Филимонов, 1949). Если говорить о количественном
уменьшении обонятельной области в ряду
приматов, то оказывается, что, по данным того же автора, «величина
отношения древней коры ко всей коре уменьшается у человека в
сравнении с игрункой
очень резко (в 4,7 раза)».
В противоположность обонятельной области, или древней коре, большого
мозга, его новая кора, или неокортекс, представляет собой в ряду
приматов прогрессивную
часть мозга и достигает у человека огромного развития (рис. 68—69).
Это происходит, в частности, благодаря разрастанию некоторых
долей коры полушария
специфическим
для человека образом. В качестве примера остановим внимание на
нижнетеменной области неокортекса обезьян и человека.
Ю. Г. Шевченко, изучив в Институте мозга нижнетеменную (нижнепариетальную)
область коры у игрунки, мартышки, гиббона, шимпанзе, орангутана
и человека, установила сходство в чертах основной структуры и
обнаружила у антропоидов
поля 39 и 40, характерные для человека (Brodmann, 1925). Одновременно
она обнаружила и заметные количественные и качественные отличия.
С одной стороны,
границы между соседними полями и подполями данной области у
антропоидов более резкие, чем между соответствующими формациями
у человека,
с
другой стороны, у антропоидов они слабее развиты, не захватывая
всей нижнепариетальной
дольки, а лишь ее верхние отделы. У игрунки нижнетеменная область
равна 0,17% от всей поверхности коры, у орангутана и шимпанзе
составляет лишь 3,3%, в
то время как у человека она очень
219
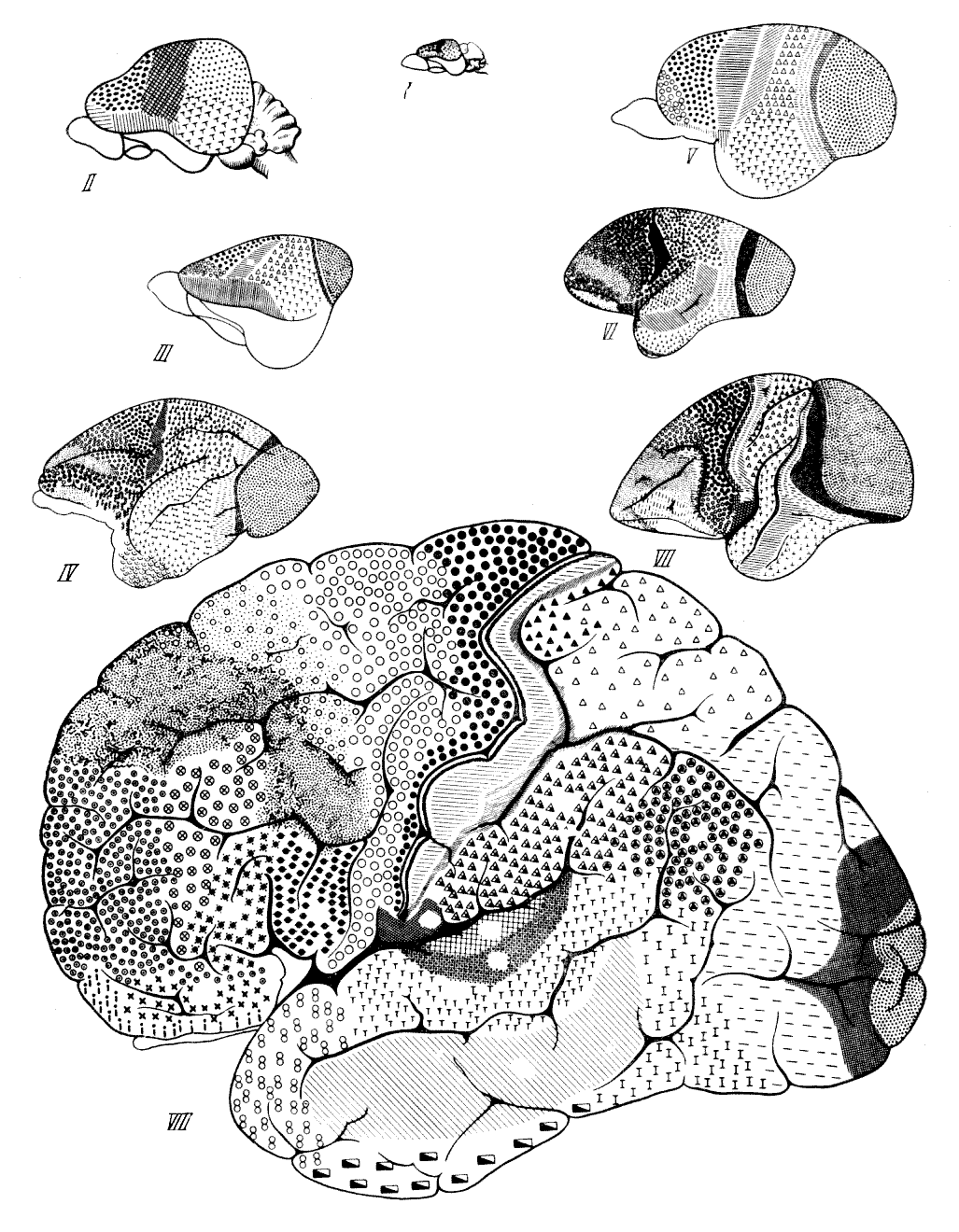
Рис. 68. Цитоархитектонические поля коры больших полушарий
мозга приматов
I — насекомоядное млекопитающее хоботный прыгунчик
(Macroscelides
elephantulus);
II — тупайя собственно (Tupaia sp.); III — тупайя перохвостая
(Ptilocercus lowii); IV — лемур (Lemur sp.); V — долгопят (Tarsius
spectrum);
VI — игрунка (Hapale jacchus); VII — макак (Macacus sp.); VIII
— человек (Homo sapiens);
I, II, II, V — по Ф. Вуду Джонсу, 1929; IV, V, VI и VII — по
К. Бродманну, 1925
Рис. 69. Области коры больших полушарий головного мозга
1 — оранг; 2 — шимпанзе; 3 — горилла, 4 —
человек; 5 — лемур;
6 — долгопят; 7 — капуцин; 8 —
макак; 9 — павиан; 10 — гиббон; а —
двигательная область, б — чувствительная (сенсорная); в —
зрительная;
г — слуховая. Вид слева.
Лобная доля не заштрихована (на рисунках видно ее прогрессивное
развитие от лемура и долгопята до антропоидов и человека).
Точками на лобной
доле у человека обозначена двигательная зона речи. 2/7
нат. вел. По
Ф. Тильнею, 1928 (ширина изображения 117 мм)
разрослась и охватывает 7,7% (Шевченко, 1936—1938).
Приведенные соотношения тем более представляются наглядными,
что, хотя по размерам площади поверхность коры полушарий
большого мозга
человека
в три
раза превосходит ее у шимпанзе, вся нижнепариетальная область
у человека в десять раз больше, чем у человекообразных обезьян.
Такое
сильное
развитие филогенетически новой нижнетеменной области у человека,
очевидно, зависело
от ее более усиленного и своеобразного функционирования под
влиянием трудовых действий, звукового языка, второй сигнальной системы.
Нарушения деятельности этой области обнаруживаются в сложных нервных расстройствах.
Таким образом, нижнетеменная область тесно связана с функциями
по линии второй сигнальной системы — со звуковой речью, чтением,
письмом:
филогенетически
молодая область коры у человека качественно отличает его мозг
от обезьяньего.
Лобная область также включает весьма существенные филогенетические
новые образования, высоко ставящие мозг человека над животным.
В Институте мозга
(Москва) ее подробно исследовала
221
Е. П. Кононова, которая сделала очень важное открытие, когда
обнаружила в коре нижней лобной доли мозга шимпанзе участки,
соответствующие
44 и 45 полям
полушарий человеческого мозга, между тем как зарубежные ученые
отрицали наличие их гомологов даже у антропоидов. Кроме того,
ей удалось найти
следы или зачатки
подобных участков даже у некоторых низших обезьян. Почему
так важны поля 44 и 45 для человека? Уже давно ученые подметили связь
этой
структуры с
функцией членораздельной речи. Соответствующий участок даже
называли по имени крупного
французского антрополога «центром речи Брока». Но позже было
установлено, что к речевой функции имеет ближайшее отношение
не только лобная
область, но также нижнетеменная и височная. Исследования
показали, что в нижней
лобной извилине помещается двигательная зона речевой функции.
При ее патологических состояниях в левом полушарии у правшей
возникает
двигательная
афазия и
иные
расстройства речи (Пенфилд, Роберте, 1964).
О том, что развитие лобной доли человека носило прогрессивный
характер, свидетельствуют и другие ее анатомо-физиологические
особенности,
например очень многочисленные
ассоциативные связи с помощью пучков волокон с другими отделами
мозга. Это подтверждают факты поздней миэлинизации волокон
(как и в нижней
теменной доле), сравнительного увеличения лобной доли в
ходе эволюции, а также заметного
усложнения узора борозд всех размеров в связи с обогащением
количества извилин на поверхности полушарий большого мозга.
Следующей областью, связанной с речевой функцией, является
височная: в ней находится зона восприятия звуков, в частности
членораздельной
речи
или, шире,
звукового языка. Работа С. М. Блинкова (1955) показывает,
что в поле 41, которое здесь имеет особое значение в качестве
так
называемой
«слуховой
зоны»,
кора более сложно дифференцирована, чем, у обезьян. То же
следует сказать и о филогенетически новой территории заднего
участка
височной доли.
Еще более резкие различия от мозга обезьяны находятся в тончайшей
структуре поля 21
средней, подобласти височной доли человеческого мозга.
О важности определенных территорий височной доли, в частности,:
участка, носящего название «зоны Вернике» (по имени немецкого
невролога), свидетельствует их патологическое состояние,
приводящее к непониманию
речи, т. е. к
сензорной афазии. Некоторые заболевания в соседней доле,
затылочной. ведут к
словесной слепоте типа оптической алексии, когда человек
перестает узнавать те или
иные буквы и целые слова. Таким образом, и эта доля,
вероятно, принимает значительное участие в осуществлении
функций специфической
только для
человека второй сигнальной системы, так или иначе связанной
со всей корой.
Затылочная область коры, в которой располагается мозговой
конец зрительного анализатора, как раз и служит у человека
главнейшим
органом восприятия
зрительных образов в связи с первой и второй сигнальными
системами.
222
Неудивительно,что и тут исследователи устанавливают заметные
отличия по сравнению с мозгом других приматов, можно сказать
даже обнаруживают
качественную
перестройку
в связи с общим процессом преобразования мозга обезьяны в
мозг человека.
Воспользуемся в данном случае материалами исследования, сделанного
в Институте мозга на полушариях большого мозга человека,
высших и низших обезьян (Преображенская,
Филимонов, 1949).
Установлено, что у человека из трех полей затылочной области
наибольшего развития и сложности строения достигло поле 19,
в то время как
поле 17 невелико. И если у человека поле 19 составляет 4,5%
всей поверхности
коры,
у орангутана
— 6,4% и у низшей, мартышкообразной обезьяны — 6,0%, то
поле 17 соответственно занимает 3,0, 8,5 и 10,0%.
Отсюда видно, что и в затылочной доле большого мозга человека
имеются такие черты, которые придают ему значительное своеобразие.
Другими
словами, развитие
полей 17, 19 и всей затылочной области у человека пошло совсем
в другом направлении, чем у его ближайших сородичей — обезьян.
Иной представляется и картина расположения полей на полушарии:
так, поле 17 у человека располагается не на боковой, или
латеральной, его стороне,
а почти целиком на серединной, или медиальной, вследствие
разрастания теменно-височных частей коры. Туда же переместилась
и шпорная
борозда, которая у обезьян
находится частично на латеральной стороне полушария у самого
затылочного полюса. У низших обезьян зрительно-окципитальная
кора надвигалась на теменную и частично перекрыла ее заднюю
грань.
В связи с этим
здесь и возникла обезьянья борозда, ярко обозначающаяся на
латеральной поверхности
больших полушарий в пограничном районе теменной и затылочной
долей.
У человека затылочная доля испытала сильное потеснение и
некоторое уменьшение относительных размеров, что зависит
от разрастания
нижнетеменной, височной
и лобной долей. Следовательно, в этом случае прогрессивное
развитие одних отделов коры большого мозга вызывало относительный
регресс
других ее частей
или элементов, что справедливо и для всего мозга в целом:
специфически человеческие черты в процессе антропогенеза
появились на базе
прежних и совсем заново,
обезьяньи же ослабевали и исчезали. Проиллюстрируем сказанное
еще одним примером.
На границе лобной и височной долей, в глубине сильвиевой
борозды, располагается центральная (стволовая) доля, или
так называемый
рейлиев островок (рис.
70). Он обнаруживается при раздвигании краев борозды, образованных
покрышками, или оперкулярными частями, соседних долей, в
том числе теменной. Иная
картина
у обезьян, так как у них центральная, или инсулярная, доля:
частично выдается на поверхность, располагаясь между названными
выше и
окружающими долями
и занимая срединное положение (Дешин, 1934).
Говоря о мозге предшественников древнейшего человека,
223
Рис. 70. Центральная доля большого полушария мозга человека (видна
на дне сильвиевой ямы, обнажившемся вследствие удаления покрышек, которые
образуются сдвинувшимися здесь частями лобной, теменной и височной
долей)
По Э. Виллигеру, 1930
|
Ю. Г. Шевченко (1967) считает, что он развивался под влиянием
прогресса передних конечностей в сторону прогрессивного конструктивного
манипулирования
предметами
при одновременном совершенствовании двуногого хождения, о
чем может свидетельствовать пример австралопитеков. И качественные
особенности
развития больших полушарий
их коры, равно как и системогенеза мозга человека в целом,
можно
познать лишь в свете онто- и филогенетического развития при
сопоставлении в
первую очередь с характерными чертами структуры, функций
и развития мозга других
групп приматов, в частности узконосых низших и высших обезьян
(Шевченко).
С другой стороны, важнейшее значение в процессе развития
мозга австралопитеков и формировавшихся древнейших и древних
гоминид
имело появление и
все более активное применение посредника между организмом
и средой
в виде орудий, у гоминид — уже искусственных. В последнем
случае опосредованные действия по-новому, качественно характеризовали
постоянный, еще более
активный и могучий трудовой фактор преобразования головного
мозга
ископаемых людей.
При этом, как показывает его эволюция по слепкам эндокранов,
сперва усиливается развитие их задних отделов с сенсорными
областями, а затем передние, которые,
в особенности префронтальные, проецируют самую сложную часть
двигательной
зоны коры (Кочеткова, 1967) и где у современного человека
реализуется программирование на высшем уровне сознания (Лурия,
1969).
Частные примеры разрастания извилин, например вокруг инсулярной
дольки, превращающейся в островок на дне сильвиевой щели,
свидетельствуют об
интенсивном процессе развития извилин, или гирификации, и
увеличения количества борозд
второго и особенно третьего порядка, или сулькации, в мозгу
человека.
Процесс разрастания коры больших полушарий человеческого
мозга есть выражение огромного преобразования центрального
органа высшей
нервной
деятельности
в ходе развития организма человека под влиянием труда и речи
в резкое отличие от эволюции мозга обезьяны под влиянием
одних лишь
биологических
или, шире,
природных факторов.
Если усиленная тарификация и сулькация коры человеческого
мозга частично и связаны с тем, что вызывались самим процессом
увеличения
его размеров,
то основная причина лежит в иной плос-
224
кости, в качественном преобразовании внутреннего клеточного
состава больших полушарий. Оказывается, что поразительная
сложность тонкой
архитектоники
коры очень сильно обусловлена и непропорционально богатым
составом нервных клеток, а именно невронов со всеми их отростками.
Если в мозгу орангутана или шимпанзе кора содержит лишь несколько
миллиардов невронов, то в большом мозгу человека кора их
вмещает около десяти
и даже больше миллиардов. Отсюда, как это и выясняется при
исследовании, огромное
усложнение связей между частями мозга, что опять-таки подтверждает
мнение о качественном отличии человеческого мозга от обезьян.
Итак, в свете данных современной науки полностью подтверждается
вывод о том, что головной мозг современного человека является
конечным продуктом чрезвычайно
долгого хода развития животного мира. Во время предыстории
и в последнее
время, когда из обезьян возникли первые люди, началось под
влиянием труда
и звукового языка более интенсивное развитие нервной системы,
формировалось человеческое сознание, позволяющее понять законы
природы, расшифровать
свое собственное происхождение (Леонтьев, 1959).
Весь ход изучения строения, развития и функций головного
мозга человека является сплошным опровержением религиозных
представлений
о душе
как о частице божественного
духа, управляющей человеческим телом, как о бессмертном двойнике
материального тела.
Учение о локализации функций в головном мозгу наглядно показывает
полную зависимость всех психических процессов от нормального
функционирования неповрежденных частей мозга и всего этого
наиважнейшего органа
в целом
(Поляков, 1964).
Уяснению нормальной деятельности головного мозга чрезвычайно
способствует глубоко материалистическое учение И. П. Павлова
об условных рефлексах.
Созданное величайшим физиологом мира, это учение неопровержимо
доказывает наличие общих
закономерностей деятельности нервной системы млекопитающих
животных и человека, общность у них первой сигнальной системы,
помогает
уяснить особенности
онтогении, филогении, строения и жизнедеятельности человеческого
тела, являющегося продуктом
также и более древних стадий развития животного мира.
Вместе с тем на основе своей физиологической концепции И.
П. Павлов пришел к созданию представления о второй сигнальной
системе
как
специфической для одного человека в отличие от животных,
и трудно переоценить важность
этой
идеи для исследования самых глубоких научных, философских
проблем, связанных
с речью, мышлением.
Рефлекторная теория И. П.Павлова служит разительным естественнонаучным
подтверждением ленинской теории отражения. Тем самым она
бросает яркий свет на проблему
антропогенеза в части, касающейся развития мозга как органа
мышления.
225
Высшая нервная деятельность
обезьян
Физиологи изучают высшую нервную деятельность животных строго
объективным методом условных рефлексов. Физиологическое учение,
созданное русскими
учеными И. М. Сеченовым и И. П. Павловым, рефлекторная теория,
учение о нервизме
— все это краеугольные камни единой концепции, позволяющей
биологам, в том числе антропологам, правильно и глубоко подойти
к решению
даже труднейших
вопросов эволюции на основе материализма.
Физиологическое учение И. П. Павлова — сильное, действенное
оружие в борьбе против реакционных течений в зоопсихологии,
особенно
в США, Англии, ФРГ.
Субъективный метод исследований высшей нервной деятельности
должен в конце концов уступить место объективному. Как известно,
И.
П. Павлов никогда
не позволял трактовать физиологические опыты по изучению
высшей нервной деятельности
в психологических терминах. В то же время он, борясь против
дуалистической, идеалистической психологии, утверждал правомерность
существования
и развития психологии материалистической. Основы последней
лежат в учении
марксизма-ленинизма,
методика связана с рефлекторной теорией, сама психология
теперь неразрывна с павловским учением. В сфере внимания
И. П. Павлова
находились такие
пункты первостепенной важности для учения об антропогенезе,
как высшая нервная
деятельность, членораздельная речь (вторая сигнальная система),
мышление, труд. Обращенная всем своим существом против идеализма, физиология
И. П. Павлова служит прочной
научной основой
для борьбы советских антропологов против всевозможных антидарвинистических
гипотез антропогенеза.
Поведение обезьяны, включающее моменты, которые более сильно,
чем у других животных, напоминают поведение отдельного человека,
составляло
предмет
интереса ученых с давних времен. Еще ученые XVIII в., например
Бюффон, наблюдали действия
обезьян, в том числе человекообразных. Особенный сдвиг в
изучении поведения,
движений, инстинктов этих животных произошел в тот период,
когда теория эволюции Дарвина победоносно проникала в самые
разнообразные
отрасли
биологии, в том
числе в зоопсихологию (Дембовский, 1963).
Как известно, Дарвин написал капитальное сочинение «Выражение
эмоций у человека и у животных», вышедшее в свет в 1872 г.
Он рассматривает
это
сочинение как
часть своего основного труда «Происхождение человека и половой
отбор». Дарвин доказал, что в выражениях основных эмоций
человек обладает
очень близким
сходством с обезьянами, которое можно истолковать только
как генетическое родство между первым и последними.
Забвение или непонимание качественных различий между человеком
и животными, отчасти свойственное самому Дарвину, ярко проявилось
в
работах многих
зоопсихологов, что несомненно мешало развитию правильных
представлений о психике живот-
226
ных и вело не раз к антропоморфизму в ее трактовке.
Немецкий зоопсихолог Вольфганг Кёлер посвятил большое исследование
поведению антропоидов. В его распоряжении было несколько
экземпляров молодых шимпанзе,
над которыми он поставил ряд опытов по выяснению уровня их
«умственных способностей». В этих опытах некоторые шимпанзе
обнаружили умение
лучше других находить
те или иные пути к пище, положенной в малодоступном месте
или высоко подвешенной: они подтаскивали ящики, взбирались
на них,
пользовались
палками. В одном
опыте шимпанзе научился даже составлять две короткие палочки
в одну длинную. Кёлер пришел к ошибочному выводу, что тип
интеллекта у шимпанзе
такой же,
как и у человека.
Подобное приравнивание человека к животному можно часто встретить
у американских зоопсихологов, среди которых самое видное
место занимает Роберт Йеркс.
Он изучал поведение крупных антропоидов и пришел к заключению,
что по уровню умственного развития горилла (рис. 71) стоит
на первом месте,
шимпанзе
на
втором и орангутан на третьем. Самый ошибочный вывод Йеркса
— признание
им у обезьян мышления, проявляющегося во внезапном «озарении»,
«догадке», «идеации». Другими словами, Йеркс и Кёлер идеалистически
трактуют
зоопсихологические проблемы.
Иную позицию в отношении проблемы интеллекта и поведения
животных занимают отечественные зоопсихологи. Мы остановим
внимание на
трудах тех исследователей, которые специально изучали обезьян
в связи с проблемой антропогенеза,— Н. Н. Ладыгиной-Котс (Москва), Н. Ю.
Войтониса
(Сухуми), Г. З. Рогинского
(Ленинград).
Н. Н. Ладыгина-Коте изучала умственные способности и особенности
зрительного восприятия у детеныша шимпанзе Иони по оригинальному
методу выбора
на образец (рис. 72). Опыты производились обычно по следующему
плану: разложив перед
шимпанзе предметы одинаковой формы, но разного цвета, Н.
Н. Ладыгина-Коте брала из запасного ящика какой-нибудь подобный
предмет и предъявляла
ему. Шимпанзе обнаруживал способность выбрать предмет такого
же цвета из лежащего
перед ним набора и подать экспериментатору.
Проделав многочисленные опыты по указанной методике, Н. Н.
Ладыгина-Котс пришла к следующему общему выводу: «Поведение
шимпанзе не
столько, быть
может, «предусмотрительно», сколько «послеосмотрительно».
Если можно так выразиться,
шимпанзе крепок задним умом: лишь на практике испытав пути
решения, он начинает применять их правильно». По мнению Н.
Н. Ладыгиной-Котс
(1923),
в этом случае
между шимпанзе и человеком разница очень велика, так как
разумный человек «провидит последствия и, не нуждаясь в конкретном
опыте,
делает правильный
вывод».
Кроме опытов над шимпанзе Н. Н. Ладыгина-Котс провела ряд
экспериментов над макаком резусом, ставя на его пути различные
препятствия
к пище или к
227

Рис. 71. Молодая самка горной гориллы (Gorilla gorilla beringei
Matschie), no кличке «Мисс Конго», выталкивает палкой пищу
(плоды) из трубы
Опыт
Р. Йеркса в Лаборатории биологии человекообразных обезьян
в «Орендж-Парке», полуостров
Флорида. По Р. и А. Йерксам, 1934
Рис. 72. Опыт с шимпанзе детенышем «Иони», который подает
экспериментатору кружочек того же цвета, какой ему показывается
По Н. Н. Ладыгиной-Котс, 1923
Рис. 73. Мимика лица шимпанзе — детеныша шимпанзе «Иони»
1 — волнение; 2 — внимание; 3 — удивление; 4 — отвращение;
5 — злоба; 6 — страх; 7 — печаль (плач); 8 — радость
(смех). По
Н.
Н. Ладыгиной-Котс,
1936
свободе. В отношении резуса она пришла к сходному выводу,
а именно, что характер элементарной познавательной
способности низшей обезьяны
резко
несходен, если
не прямо противоположен таковому у человека (Ладыгина-Котс,
1920).
Сравнивая же особенности поведения детеныша шимпанзе
и ребенка соответствующего возраста, Н. Н. Ладыгина-Котс
(1935, 1960,
1965) приходит к выводу,
что, хотя шимпанзе и обнаруживает большие качественные
отличия от человека, все же у него возможно предполагать
наличие
таких черт,
подобие которых
должен
был иметь и древнейший представитель гоминид, впоследствии
ставший человеком
разумным.
Различные выражения лица шимпанзе были изучены Н. Н.
Ладыгиной-Котс (1935) во время длительного наблюдения
поведения, привычек,
инстинктов и выразительных
движений того же детеныша шимпанзе Иони в возрасте
от 1 1/2 до 4 лет. Н. Н. Ладыгина-Котс изучала мимику его
лица во
время таких
основных
состояний
животного,
как общая возбудимость, «печаль, радость, страх, злоба,
удивление, внимание, отвращение». Движения рта и губ
занимают большое
место в общей мимике лица
шимпанзе (рис. 73). При сильном возбуждении у него
могла распушиться шерсть на голове, туловище, руках и ногах.
Наступали также различные
другие изменения
во внешнем виде и поведении животного. Даже по сравнению
с гориллой и орангутаном шимпанзе поражает богатством
своей мимики и ее
большим
сходством с человеческой.
Мимика крупных человекообразных обезьян гораздо богаче,
чем у гиббонов и низших обезьян, но весьма значительно
уступает
человеческой.
Превосходство последней обусловливается выдающимся
многообразием функциональной
деятельности лицевого нервно-мышечного аппарата, характеризующегося
более высоким
развитием двигательного анализатора, сложностью иннервации
мышц лица, очень сильной
дифференцированностью его мимической мускулатуры (рис.
74), комплексностью
связей с мозгом в целом.
Эволюция лицевой мускулатуры приматов была подробно изучена
Э. Хьюбером (Huber, 1931). Он придавал особое значение
центральной нервной системе:
ее высокий
уровень развития является важным предварительным фактором
усложнения
мимической мускулатуры и богатства выражения эмоций.
Их разнообразие увеличивается
с умножением связей между разными отделами коры мозга.
«Между тем как у низших
приматов выражение лица представлено очень простой
схемой с немногими стереотипными чертами, по мере поднятия по
ряду приматов оно
постепенно становится богаче
и более совершенным, до тех пор пока не достигает совершенно
поразительного разнообразия и сложности у крупных человекообразных
обезьян, среди
которых наибольшую степень сходства с человеком обнаруживают
африканские антропоиды
— горилла и шимпанзе, а меньшую — орангутан»,— пишет
Хьюбер.
Однако даже горилле и шимпанзе не приходится состязаться
по богатству мимики с современным
230

Рис. 74. Лицевая мускулатура у различных представителей
приматов
1, 2 — схема основного строения
поверхностной лицевой мускулатуры приматов по ее
изучению у лемуров, долгопятов
и примитивных
американских обезьян;
3 — банканский долгопят (Tarsius saltator
Elliot);
4 — львиная игрунка (Leontocebus=Oedipomidas
rosalia Linnaeus); 5 —
черная коата (Ateles
ater F. Cuvier); 6 — макак резус
(Pithecus=Macacus rhesus Audebert); 7 —
сиамский гиббон (Hylobates pileatus Gray); 8 —
орангутан (Pongo pygmaeus
Linnaeus),
полувзрослый самец; 9 —
обыкновенный шимпанзе (Pan troglodytes Blumenbach);
10 — береговая горилла (Gorilla gorilla
Savage and Wyman), молодая самка; 11 — новорожденный
европеец, 12 — взрослый европеец; а —
платизма (поверхностный слой);
б — платизма
(глубокий слой);
в — группа задних ушных и затылочных мышц; г —
вентральный отдел переднего
ушного отдела глубокого шейного сфинктера; д —
дорсальный отдел того же сфинктера, или мышца, опускающая
ушную
раковину; е —
козелково-завитковая мышца; ж —
группа передних ушных, височных и лобных мышц; з —
круговая мышца глаза; и — мышца, опускающая
надбровный участок;
к — носогубная
мышца;
л — ухогубная,
или скуловая, мышца. По Е. Хьюберу, 1931
человеком, у предков которого в дополнение к указанным
выше природным биологическим условиям развития (включая
сюда прогресс
соответствующей
области в моторной
зоне коры головного мозга в связи с прямохождением и употреблением орудий)
присоединились еще социальные моменты, оказавшие
могущественное действие на
усовершенствование головного мозга и психики, на
231
развитие лицевой мускулатуры и выражения лица, надо
полагать, еще у древних людей.
«Приобретение и постепенное совершенствование членораздельной
речи должно было оказывать дальнейшее побуждающее
воздействие на развитие
мимической
мускулатуры и выражение лица»,— говорит Хьюбер. Благотворное
влияние оказывала речь на лицевые мышцы древних людей,
и развитие мимической
мускулатуры
у современных людей, по Хьюберу, еще продолжается
в мышцах глабеллярной области
(выше корня носа) и надглазничных областей, а также
в мускулатуре вокруг рта.
Новорожденный имеет малодифференцированное выражение
лица в связи с недостаточным развитием органов чувств
и коры
полушарий, хотя
к моменту
рождения ребенка
мимическая мускулатура в главных чертах сформирована.
У маленьких детей ее функциональная деятельность
проявляется не сразу,
но вырабатывается довольно
быстро, особенно на протяжении первого года, а затем
в
течение следующих двух лет. Лицевая мускулатура у
взрослого человека
очень высоко дифференцирована.
Артисты способны с помощью тщательной и длительной
тренировки лицевых мышц осуществлять чрезвычайно
тонкую мимику.
Массу вариаций смеха,
гнева и других
эмоций можно видеть и в жизни. У человека имеется
хорошо выраженная мышца смеха, растягивающая углы
рта и обнажающая
зубы. У обезьян
эта
мышца не
является столь дифференцированной: оскаливание рта
представляет собой большей частью выражение угрозы,
а иногда, как, например, у шимпанзе, также и выражение смеха (Schrier, Harlow,
Stollnitz,
1965).
Очень характерен
для
обезьян их большой интерес к самым разнообразным
предметам окружающего мира, независимо от того, съедобны
они
или нет. Н. Ю. Войтонис
(1949) специально
изучал у низших обезьян «ориентировочно-исследовательскую
деятельность». Он установил, что обезьяну привлекает
не только новизна предмета,
но и сложность его строения вместе с податливостью
для воздействия. В
отличие
от человекообразной
обезьяны у павиана или макака интерес к предмету
не повышается, если его сделать звучащим.
Стремление обезьяны к новому, интересному зачастую
настолько велико, что превышает голод и заставляет
обследовать
данный ей предмет
со всех сторон.
Такой повышенный интерес к предметам окружающего
мира выработался у обезьян, очевидно, вследствие
условий
их жизни: обладая
довольно высокоразвитым
мозгом, прекрасным зрением и хватательными передними
конечностями, обезьяны пользуются в природе богатым
ассортиментом пищи
и разными путями реагируют
на явления и предметы окружающей природы.
Вывод Н. Ю. Войтониса о том, что эта ориентировочно-исследовательская
деятельность у обезьян, являющаяся, по И. П. Павлову,
их специфической особенностью,
далеко перерастает за пределы чисто пищевой потребности
и у низших обезьян, весьма
важен. Здесь можно видеть ценное под-
232
Рис. 75. Опыт А. И. Каца
с макакой лапундером «Патом» на Сухумской медико-биологической
станции: обезьяна использует палку (с вилкой на
конце) для добывания яблок из положенного на бок глубокого
деревянного ящика — «экспериментального колодца»
По фото из архива Института антропологии,
Москва
|
тверждение той мысли Дарвина, что нашими предками
были обезьяны, сумевшие обратиться к использованию
предметов
в качестве
средств добывания пищи.
С другой стороны, вывод, сделанный Войтонисом, заставляет
искать корни некоторых
отличительных умственных особенностей человека в
далеком прошлом у ископаемых обезьян.
Но одного только повышенного интереса к предметам
было, наверное, недостаточно для того, чтобы наши
предки
обратились к пользованию
орудиями и труду.
Необходимо было еще обладать способностью подметить
соотношения между предметами и изменить
их с помощью своих природных органов либо с помощью
какого-нибудь предмета, могущего служить орудием
в руках обезьяны.
Н. Ю. Войтонис установил,
что такая способность не только имеется у низших
обезьян, но и проявляется весьма своеобразно в том
смысле, что
вместе с повышенным
интересом
к цели
и упорством
в ее достижении низшие обезьяны пользуются весьма
разнообразными предметами, нередко легко отказываясь
от неудачного
приема и обращаясь
в результате
нескольких или многих попыток к более удачному.
Очень интересной была задуманная Н. Ю. Войтонисом
серия опытов, в которых один макак лапундер научился
доставать
из небольшого
колодца
воду или
песок с помощью ведерка, а яблоки с помощью длинной
вилки (рис. 75). Другой макак
лапундер самостоятельно научился применять, смотря
по обстоятельствам, короткую или длинную палку с
крючком на конце для захвата пищи. (Эта серия опытов
произведена А. И. Кацем. Фотография взята из его
кандидатской диссертации,
1949.)
На основе таких исследований удалось установить новые
факты, свидетельствующие о том, что обезьяны способны
формировать
устойчивые и достаточно
сложные навыки при достижении цели даже при отсутствии
пищевой приманки. Такая
способность, несомненно, должна была быть у человеческих
предков в еще более высокоразвитой
форме. Обладали они и умением воспользоваться орудием
как средством изменения соотношений между предметами,
например
при добывании
из земли луковиц,
кореньев, червей, насекомых и их личинок.
Согласно исследованиям Н. Ю. Войтониса, низшие обезьяны
удивительно замечают детали предметов. Такое свойство
должно бы-
233
ло иметь большое значение для наших предков при обработке,
например, каменных орудий, сперва еще очень грубой,
но впоследствии достигшей
тончайшего мастерства.
Таким образом, работа Н. Ю. Войтониса, выяснившая
некоторые характерные особенности поведения низших
обезьян, подтверждает
основное положение
дарвинизма о том,
что корни умственных способностей человека надо искать
в психике человеческих предков (насколько о них можно
судить по современным
обезьянам). С
другой же стороны, работа Н. Ю. Войтониса является
удачной иллюстрацией идеи Энгельса о важной роли
труда в процессе очеловечения обезьяны. Склонность
манипулировать орудиями, большую любознательность
и упорный интерес к новым предметам
обнаруживали в условиях опыта также и широконосые
обезьяны из группы капуцинообразных, или цебид.
О высоком развитии
капуцинов
свидетельствуют
опыты Г. З. Рогинского
(1946), в которых выяснилась важнейшая роль зрительных
рецепторов для правильного решения задач и обнаружилась
быстрая ориентировка
капуцина
при изменении
условий опыта. Очень высокую сообразительность обнаружили
капуцины также в опытах И. А. Биренса де-Хаана (Haan,
1931) и Г. Клювера
(Kluver, 1933).
Еще более интересны опыты Г. З. Рогинского (1948)
по исследованию навыков у шимпанзе в связи с вопросом
о зачатках интеллектуальных
действий
(рис. 76). В своих работах Г. З. Рогинский
в большой мере учитывал биологию обезьян, имеющих
сильно развитые органы зрения и осязания в соединении
с особой
моторикой и заметно
отличающихся
от своих сородичей в классе млекопитающих также другими
особенностями.
Учет особенностей жизни обезьян на воле важен потому,
что поведение биологически очень комплексно именно
у них по
сравнению с прочими
млекопитающими, хотя
количество прирожденных, безусловных рефлексов вряд
ли намного больше. Поведение не есть простая сумма
разных
рефлексов
или их ассоциаций.
По исследованиям Г. З. Рогинского, поведение шимпанзе
отличается большой пластичностью, приспособляемостью
к обстановке,
большой организованностью действий в трудных положениях,
причем индивидуальный
опыт имеет очень
важное значение. В противовес Кёлеру советский ученый
установил, что
шимпанзе
вовсе не «рабы зрительного поля», а способны легче
переходить из одной ситуации
в другую, быстрее и более ловко, чем низшие обезьяны,
применять палки взамен орудий. Для антрополога, занимающегося
исследованием
процесса
антропогенеза,
выдающийся интерес представляют те преобразования
высшей нервной деятельности, которые должны были
происходить
в процессе эволюции
у предков человека,
в частности у ископаемых приматов.
Энгельс характеризует ближайших предков человека
как необычайно высокоразвитых человекоподобных обезьян,
которые далеко
превосходили всех прочих
234
Рис. 76. Опыты с шимпанзе
1 — обезьяна выбрала нужную тесемку, за которую привязано
яблоко; 2 — обезьяна
манипулирует согнутой палкой, доставая яблоко. По Г. З. Рогинскому,
1948
|
смышленостью и приспособляемостью. Если понятие «приспособляемости»
больше увязывается через кору мозга с общими биологическими
и анатомо-физиологическими качествами организма,
развивающегося в зависимости от условий изменяющейся
среды, то «смышленость» следует сопоставить собственно
с деятельностью
коры больших полушарий головного мозга, включающих
центральные отделы анализаторов.
Советская антропология теснейшим образом соприкасается
с учением И. П. Павлова о закономерностях высшей
нервной деятельности,
которое она
органически
использует
в разработке своих проблем. Физиологическое учение
И. П. Павлова
в целом имеет выдающееся значение для антропогенеза,
оно позволяет лучше
понять,
как развилось на Земле «существо с мыслящим мозгом»,
как появился тот наисложнейший организм, в котором,
по словам
Энгельса, природа
дошла
до познания самой
себя.
Материалистическое учение И. П. Павлова о высшей
нервной деятельности человека и животных, резко противостоящее
идеалистической физиологии
и психологии
реакционных буржуазных ученых, помогает бороться
против
разнообразных, подчас весьма утонченных форм идеализма
в науке о человеке.
Для мировоззрения И.
П. Павлова характерна материалистическая концепция
эволюционного развития человека.
Проблему антропогенеза И. П. Павлов ставил в связь
с особенностями высшей нервной деятельности животных,
которые
он ис-
235
следовал в плане теории эволюции. Так, И. П. Павлов
писал в 1913 г.: «Возбудителем и вдохновителем современного
сравнительного изучения высших проявлений
жизни животных по всей справедливости надо считать
Чарлза
Дарвина,
который, как
это известно всякому образованному человеку, во второй
половине прошлого столетия своею гениальной иллюстрацией
идеи развития
оплодотворил всю
умственную работу человечества и в особенности биологический
отдел естествознания.
Гипотеза происхождения человека от животных придала
захватывающий интерес изучению
высших проявлений жизни животных.
Ответ на вопрос, как наиболее полезно вести это изучение,
и само изучение стали задачей последарвиновского
периода» (Павлов, 1951,
стр. 237).
Исследования И. П. Павлова открыли путь для углубленного
изучения проблемы эволюции интеллекта. Стремясь установить
филогенетическое
родство между
человеком и животным миром, Дарвин сосредоточил свое
внимание на общности в их организации.
И. П. Павлов, установив реальное сходство в физиологии
головного мозга человека и животных, подчеркнул в
то же время и качественные
различия,
выдвинул идею
о второй сигнальной системе, свойственной человеку.
В согласии со своей материалистической концепцией
И. П. Павлов уже давно боролся против дуалистического
взгляда на человеческую
природу,
столь характерного
для английских ученых типа Шеррингтона, американских,
например
Лешли и Йеркса, или немецких, как Кёлера. Бывший
«весьма одиозной персоной» для зарубежных и отечественных идеалистов, И.
П.
Павлов всегда
боролся против
идеализма, против уподобления
животным, или бестиализации, человека. Он возражал
и против антропоморфизации высшей нервной деятельности
животных,
приписывания им идей или
обобщений, возникающих у человека лишь на основе
второй сигнальной системы,
возражал против признания высшей нервной деятельности
животных разумной, а не
практически
рассудочной. Между тем подобные заблуждения очень
характерны для упомянутой категории исследователей,
охотно идущих
на отождествление поведения
человека с поведением животных. В отличие от их концепций
теория и практика физиологии
И. П. Павлова пропитаны духом диалектического, воинствующего
материализма.
Материалистически мыслящие психологи, биологи, антропологи
должны особенно ценить принцип детерминизма И. П.
Павлова, полностью
применимый и для
объяснения процесса антропогенеза в противовес всевозможным
телеологическим толкованиям
последнего, которые основаны на антинаучном постулате
непознаваемости сущности природы и человека, на идеалистическом
допущении
сверхъестественных сил.
Для характеристики ложных построений некоторых реакционных
американских ученых приведем «теорию телефинализма»
в эволюции органического
мира, предложенную Леконтом Дю-Нуи (из Рокфеллеровского
института, Нью-Йорк)
в книге «Человечес-
236
кая судьба». Здесь утверждается, что в основе эволюции
лежит сверхъестественное целенаправляющее начало
и что человек
создан богом с помощью чуда,
буквально так, как описано в библии. Теория телефинализма
есть лишь один из вариантов
этого наиболее заметного направления в умах тех американских
реакционных биологов, которые защищают в США во многих
книгах и статьях идеалистический
лозунг «Творение путем эволюции».
И. П. Павлов изучал проблему взаимоотношения различных
рефлексов и их изменений, учитывая преобладающее
влияние условий внешней
среды на организм.
Подходя к рефлексам с общебиологической точки зрения,
с позиций дарвинизма, И. П. Павлов утверждал, что
индивидуальное приспособление
существует
во всем животном мире, что безусловные рефлексы и
их комплексы
в форме инстинктов
представляют собой видовую форму приспособления,
что временные связи — условные
рефлексы и ассоциации — служат индивидуальной формой
«уравновешивания» организма со средой, т. е. приспособления.
В 1935 г. он писал:
«Образ поведения человека и животного обусловлен
не только прирожденными свойствами нервной системы,
но
и теми влияниями,
которые падали
и постоянно падают на организм во время его индивидуального
существования, т. е.
зависит от постоянного воспитания или обучения в
самом широком смысле
этих слов.
И это потому, что с указанными выше свойствами нервной
системы непрерывно выступает и важнейшее
ее свойство — высочайшая пластичность» (Поли. собр.
соч., т. III, 1949, стр. 517).
Идеи и фактические завоевания И. П. Павлова, послужившие
блистательным развитием научных достижений основоположника
материалистической
физиологии человека
и животных — великого русского ученого И. М. Сеченова,
имеют важное значение и для современной советской
физиологии. На
базе конкретных
исследований
воочию обнаруживается и правота утверждения И. П.
Павлова об изменчивости условных
и безусловных рефлексов, об их генетической теснейшей
взаимосвязи.
Основная мысль о переходе простых рефлексов в сложные
была высказана И. П. Павловым еще в 1922 г. в связи
с его идеей
о замы-кательной
функции нервной
системы. Если приложить данный основной тезис физиологического
учения И.
П. Павлова к той стадии эволюции обезьян, когда они
должны были сменить древесный образ жизни на наземный,
то для
антрополога становится возможным более точно
уяснить себе тот нервный механизм, на основе которого
происходила глубокая смена в осуществлении многих
существенно важных
рефлекторных
актов
в процессе приспособления наших предков к новым условиям
среды в
верхнетретичные времена.
Изучение высшей нервной деятельности обезьян было
начато по указаниям И. П. Павлова в его лаборатории
в 1923
г. А. Г. Ивановым-Смоленским,
Д. С.
Фурсиковым и продолжено исследователями не только
на низших, но и человекообразных обезьянах.
237
Высшая нервная деятельность обезьян чрезвычайно интересовала
И. П. Павлова, стремившегося установить ее качественные
особенности. Сравнивая
приматов
с прочими млекопитающими и с человеком, он говорил:
«Если обсудить еще раз, если сказать, в чем успех
обезьяны сравнительно с другими животными, почему
она ближе
к человеку, то именно потому,
что у нее
имеются руки, даже четыре «руки», т. е. больше, чем
у нас с вами. Благодаря этому она имеет возможность
вступать
в очень сложные
отношения с окружающими
предметами. Вот почему у нее образуется масса ассоциаций,
которых не имеется у остальных животных» (Павловские
среды,
1949, стр.
431—432).
Иначе говоря, для обезьян характерно очень высокое
развитие аналитико-синтетических процессов в коре
больших полушарий
их головного мозга. Последнее
связано, в частности, с высоким уровнем эволюционного
развития анализаторов у обезьян, вследствие чего
им стал доступен
более сложный анализ
окружающей
среды,
связанный с синтезом ее определенных элементов и
сторон. Особенное значение имеет разнообразие
двигательных сигнализаторов, высокое развитие моторики
и кинестетики.
«Соответственно этому,— добавляет И. П. Павлов,—
так как эти двигательные ассоциации должны иметь
свой материальный
субстрат
в нервной системе,
в мозгу, то и большие полушария у обезьяны развились
больше,
чем у других,
причем
развились именно в связи с разнообразием двигательных
функций. У нас же, кроме разнообразия движения рук,
есть сложность движений речи» (там же).
Физиологические воззрения И. П. Павлова позволяют
на новой фактической материалистической основе и
в свете
новых теоретических
положений
трактовать проблему возникновения
трудовой деятельности у ближайших предков человека.
И. П. Павлов очень близко подходит к проблемам трудовой
теории антропогенеза
Энгельса. Возникает возможность
подступа и к решению вопроса о начальных ступенях
развития человеческого мышления. Наблюдая за постройкой
шимпанзе
Рафаэлем
пирамиды из
деревянных предметов, посредством чего тот мог доставать
высоко подвешенную
приманку,
И. П. Павлов так оценивал его действия: «Я говорю,
что
это и есть разум, вся эта деятельность, когда обезьяна
пробует
и то
и другое,
это и есть
мышление в действии». И далее: «Вы точно воочию отчетливо
присутствуете при образовании
нашего мышления, видите все его подводные камни,
все его приемы. В этом разумность и есть, а господин
Кёлер
от этого
отмахивается:
это,
мол, метод
проб и ошибок»
(там же, стр. 431).
Многие реакционные физиологи и психологи глубоко
ошибочно трактуют высшую нервную деятельность обезьян
— столь
близких к человеку
животных, приписывая
им человеческие психические свойства. Йеркс считает
обезьян «почти людьми». Кёлер находит у них интеллект
того же
типа, что и у
человека. Качественные
различия между человеком и обезьянами, таким образом,
зату-
238
шевываются. Другие, наоборот, проводят слишком резкую
грань между человеком и животным, приписывая первому
обладание
частицей божества,
т. е. «бессмертной
душой».
Когда И. П. Павлов говорит о «примитивном мышлении»
у обезьян, то он вкладывает в этот термин чисто физиологическое
содержание
с учетом
качественных отличий
от человеческого разума. Так, он пишет, что, стремясь
«уйти от истины, психологи типа Йеркса или Кёлера
пользуются такими пустыми
представлениями,
как, например:
обезьяна отошла, «подумала на свободе» по-человечески
и
«решила это дело». Конечно, это дребедень, ребяческий
выход, недостойный
выход»,
у Рафаэля
же на деле «все поведение сложилось из анализа и
ассоциаций» (там же,
стр. 388).
Преуменьшая различия между человеком и обезьянами,
идеалистические ученые всячески поддерживают виталистическую
концепцию
происхождения человека,
соединяя ее с представлением о независимости духа
от тела, о преобладании первого
над последним. Получается характерное для таких ученых
противоречие, вызываемое культивированием в капиталистических
странах «религиозной
науки». По представлениям
некоторых реакционных биологов США божественное сотворение
мира обнаруживается в его развитии. В сочинениях
подобных ученых утверждается
истинность
библейского мифа о чудесном творении первых людей.
Такое же искажение истины в науке
о жизни и происхождении человека, высшей нервной
деятельности, допускается и некоторыми английскими
учеными-идеалистами.
Ознакомившись с «трудом» одного из подобных английских
реакционеров в науке, И. П. Павлов с глубоким возмущением
писал, что хотя
сей натуралист и «готов
признать интеллигентность у животных, как это делают
многие, особенно психологи, но тем не менее он резко
отличает этих
животных от человека
и возражает
против происхождения человека от животных, против
того, что мы представляем собой
продолжение животного мира». И по поводу того же
натуралиста И. П. Павлов добавляет: «Я давно уже
был поражен, каким
манером человек
ухитрился вырыть
такую яму между собой и животным» (Павловские среды,
1949, стр. 165).
Идеи И. П. Павлова углубленно разрабатываются в исследованиях
его учеников и последователей. Некоторые основные
работы по изучению высшей нервной
деятельности обезьян были сделаны еще под руководством
самого И.
П. Павлова над самцом
шимпанзе Рафаэлем (рис. 77) и самкой Розой в физиологической
лаборатории в селе Павлове (бывш. Колтуши) под Ленинградом.
Здесь на протяжении
ряда лет изучалось поведение шимпанзе. Так, Э. Г.
Вацуро вместе с другими сотрудниками
лаборатории высшей нервной деятельности произвел
многочисленные эксперименты с шимпанзе (Вацуро, 1948,
1955). И. П.
Павлов проводил очень много
времени у клеток, где помещались Рафаэль и Роза;
поведение антропоидов, их высшая
нервная деятельность представляли для него огромный
интерес.
239

Рис. 77. Шимпанзе «Рафаэль»
По Э. Г. Вацуро, 1948
|
Для нас в данном случае особенно важны опыты с добыванием
пищи из закрытых ящиков с помощью разных деревянных
втулок с круглым,
треугольным,
квадратным
и другими сечениями соответственно форме отверстия
на крышке. Рафаэль умудрялся применять разнообразные
способы
добывания
высоко подвешенной
пищи в виде
плодов, ягод; он подставлял шест и быстро влезал
по нему, громоздил друг на друга деревянные ящики
или
предметы
иной формы. Не менее
интересны и
примененные им способы прокладывания шестов с плота
на плот, чтобы добраться до завтрака
1.
Любопытны и опыты с огнем: на стол ставили деревянную
подставку, посередине которой лежало яблоко, вокруг
же него были наставлены горящие свечи. Не будучи в состоянии достать яблоко
из боязни
обжечь
руки, Рафаэль
начал
изобретать разные способы
тушения, задувал огоньки либо пристукивал их молоточком.
В других ситуациях подобного рода он заливал огонь
водой из кружки (рис.
78) или набирал
воды
в рот из водопроводного крана и извергал ее на огонь.
Весьма поучительны, наконец, и опыты, показывающие
способность обезьяны к составлению двух палок в одну.
В результате
своей работы Э. Г.
Вацуро приходит
к заключению, что главным фактором поведения антропоидов
является кинестетический импульс, т. е. что на первый
план надо поставить их двигательно-осязательные,
кинестетические восприятия, а не зрительные.
Э. Г. Вацуро выступает против тезиса Кёлера о том,
что обезьяны — рабы зрительного поля. Советский исследователь
опровергает
также другой
неверный вывод Кёлера,
заключающийся в приписывании шимпанзе интеллекта
того же рода, что
и у человека. Кёлер стирает качественное различие
между человеком и обезьяной,
антропоморфизирует
последнюю, одновременно как бы «озверяя» первого,
приписывает обезьяне сознательное изготовление орудий.
При употреблении и даже случайном составлении простейших
«орудий» обезьянами их поведение лишь отчасти сходно
с предполагаемым образом действий
наших предков в те отдаленные времена, когда они
начинали переходить
к употреблению
природных предме-
1 По указанию И. П. Павлова были
сняты кинофильмы «Роза и Рафаэль» (3 части) и «Опыты
с шимпанзе на плотах» (5 частей).
240

Рис. 78. Опыты с шимпанзе «Рафаэлем»
1 — тушение огня; 2 — на плотах. По Э. Г. Вацуро,
1948 (1) и по Г. P. Рогинскому, 1948 (2)
тов в качестве первоорудий и жили стадами.
Несомненно, однако, что изучение поведения обезьян
является в высшей степени ценным для уяснения
процесса антропогенеза. Сходство в некоторых основных чертах поведения,
высшей нервной
деятельности, выражения
эмоций указывает
на общие пути раз-
241
вития головного мозга и органов чувств у предков
человека и обезьян.
Наблюдения и опыты, проведенные на обезьянах
в лабораториях И. П. Павлова и на Сухумской
медико-биологической станции его учениками
и последователями
на протяжении десятков лет, подтверждают
учение
Дарвина
о происхождении человека из мира животных,
выявляют на новой
основе материалистической
физиологии
более близкое филогенетическое родство
с обезьянами.
Высшая нервная деятельность и поведение
обезьян обусловлены исторически сложившимися
взаимоотношениями
организма
и среды, составляют
их видовую характеристику.
Сообщим, что по словам Л. Г. Воронина
(1952, 1954), осуществившего крупные
исследования
на основе учения
И. П. Павлова,
обезьяны обладают наиболее
высокоразвитыми подражательными способностями,
ярким ориентировочно-исследовательским
рефлексом, средствами сигнализации в
виде звуков и мимико-жестикуляторных
реакций, сильными нервными процессами
—
возбуждением и торможением (Devore,
1965).
О биологической значимости условнорефлекторной
деятельности животных свидетельствуют
и эксперименты (рис. 79),
поставленные в лаборатории
сравнительной физиологии
высшей нервной деятельности позвоночных
животных Института физиологии имени академика
И. П. Павлова
в Колтушах
Л. Г. Ворониным и его
сотрудниками (Рокотова,
1953; Воронин, Фирсов, 1967). Последующие
важные исследования там же осуществили
Л. А. Фирсов (1969) и А. И. Счастный (1966).
Среди животных с более высоко развитым
интеллектом в последнее время особый
интерес вызвали дельфины
афалины. Действительно,
обладая большим
и сложным
головным мозгом (около 1700 г),
эти дельфины отличаются высокоразвитой
высшей нервной
деятельностью. Среди
ее
характеристик Л. Г.
Воронин и Л. Б. Козаровицкий
выделяют (1969) подражание, ориентировочно-исследовательскую
и игровую деятельность, по которой дельфины
очень напоминают других
высших
животных, в частности
обезьян. Однако исследователи подчеркивают,
что все же дельфины остаются просто умными
животными,
как
бы ни
хотели видеть
в них и «братьев
во разуме» некоторые любители сенсаций
(стр. 138).
Совокупность условных рефлексов, накопленная
животным в течение жизни, составляет
его индивидуально приобретенный
опыт, характер
и объем
которого обусловлен
прирожденными способноcтями, коренящимися
преимущественно в особенностях его нервной
системы. Кроме того,
этот опыт
не остается
без влияния
на изменение характера высшей нервной
деятельности в последующих поколениях
вследствие
унаследования признаков.
Характерное для высшей нервной деятельности
обезьян динамическое соотношение лабильности
и концентрации,
или возбуждения
и торможения (Воронин,
1951), развивалось в процессе их эволюции
в условиях стадного образа при жизни
на деревьях. У высших приматов особый
характер и слож-
242
Рис. 79. Опыты с шимпанзе-детенышей «Ладой». Световой сигнал показывает,
что пища положена в ящик, в который ведет дверца в стенке за решеткой
1 — до опыта; 2 — сигнал включен (круглое отверстие открыто); 3 — еда
в руках обезьяны. По Н. А. Рокотовой, 1953
|
ность носит осуществление рефлекса цели
— достижения разнообразных съедобных
предметов, в первую очередь
при использовании
некоторыми видами обезьян
опосредованных манипуляторных действий.
И. П. Павлов считал обезьян самыми
умными из животных вследствие обладания
четырьмя
«руками»,
четырьмя
кинестетическими волярными поверхностями
их цеплятельных и анализирующих акроподиев
при корреляции с бинокулярным и
цветным зрением:
налицо здесь большая гармония между
потенциальными способностями и кинетическими
возможностями
для появления большего
богатства
манипуляций по сравнению
с другими млекопитающими.
Наибольший интерес при определении
уровня интеллекта животных и его
прогрессивного развития представляет
количество и
качество кинетических
и потенциальных
вариантов в способах достижения жизненно
необходимой цели либо избегания врагов.
Мы позволяем себе
в связи с этим
высказать мысль
о важности
явления «вариантности» при подготовке
и осуществлении жизненно важных действий
и особой способности к ней в поведении
обезьян. А такое выгодное качество
обнаруживается
в наблюдениях и экспериментах над
обезьянами.
В нашем понятии оно коррелирует с
особенностями экологии
обезьян и с отсутствием
узкой
специализации
строения и функций, которое достаточно
характерно
для самого человека.
Среди вариантов способов добывания
пищи приматами с помощью опосредованных
(опосредствован-
243
ных) действий внимание антрополога
и психолога может больше привлечь
употребление камня,
чем дерева. Примеры
манипулирования
камнями
при разбивании орехов
капуцинами и шимпанзе были известны
еще
Дарвину. В 1951 г. было опубликовано
сообщение из Либерии
о том,
как
дикий шимпанзе,
сломав с масличной
пальмы ветвь с орехами, затем разбивал
их камнем на каменной плите и поедал
ядра. Советский антрополог А. И.
Кац (Тбилиси)
в 1966
г. впервые сумел
в экспериментах наблюдать, как
подопытный экземпляр шимпанзе спонтанно ударил
камнем о
другой камень, и разбил его, чего
с этими антропоидами пока не удавалось
достигнуть
другим исследователям (Хрустов,
1965).
Вариантность в поведении шимпанзе
Султана по сравнению с другими
особями в опытах
В. Кёлера
весьма наглядна.
Если
и допустить,
что служитель
показал Султану, как надо сложить
две бамбуковые палочки вместе,
вставив одну в
другую, то ведь и в дальнейшем
обезьяна добывала приманку самостоятельно,
варьируя
сочетание палки со щепкой, с пучком
соломы и
т. п. Подобная творческая иррадиация
целевого манипулирования,
несомненно,
высоко превознесла
Султана над его
соплеменницами по богатству вариантов
решения одной и той же задачи —
достать
пищу. Этот
самец проявил
вариантность
в искусственном
составлении «орудия»
из разных частей и в нужном удлинении
средства добывания
пищи.
Вспомним пример из жизнедеятельности
людей: талантливый математик сумеет
найти большее количество и притом
более удачных вариантов решения задачи. Тот же
критерий прилагается
и при определении
победителя в математических и других
научных
конкурсах, и в жизни.
Критерий степени вариантности выбора
путей достижения цели может быть
применен для
проблем, связанных
с онто- и филогенетическим
развитием человека. В
этом направлении антрополог должен
применить данные сравнительной
психологии к
освещению процесса перехода грани
между животным и человеком. В исторический
момент возникновения
нового
вида приматов
— древнейшего
человека
(Homo erectus) — трудовые манипуляции
с камнями означают новый, качественно
особый
вариант
поведения на основе прежней длительной
миллионолетней жизнедеятельности
прегоминид,
т. е. предков
первых людей, или архантропов.
По-видимому, предковый вид, давший
начало
человеку, сформировался в нижнеплейстоценовой
группе австралопитеков.
В свое время Энгельс (1895— 1896)
правильно говорил о подвидовой
группе ископаемых
крупных наземных
антропоидов (ближайших
предков древнейших
гоминид, или архантропов) как
о той «породе обезьян, которая
далеко
превосходила
все прочие смышленостью и приспособляемостью»
(Маркс и Энгельс. Сочинения, т.
20, стр. 491). Нам представляется,
что самым главным элементом в энгельсовском
термине «смышленость»
надо было бы считать
особо повышенную
способность
к видовой богатой вариантности
в поведении непосредственных пред-
244
ков человека. Она должна была развиться
при необходимости добывания
пищи с помощью природных предметов
как орудий в условиях жесточайшей
борьбы
за выживание на открытой местности
с недостаточностью пищи, но
со множеством хищников (Нестурх,
1964а). Подобные природные
условия
и привели предков
человека
— наиболее высокоразвитых двуногих
наземных
крупных антропоидов в
подсемействе австралопитековых
(семейство понгид) — к совершенно
новому, более
прогрессивному качественно
особому — трудовому — варианту
выживания
и преобразования предкового
вида.
Говоря о первых творцах орудий,
К. П. Оукли (Oakley, 1963)
пишет: «По
аналогии
с лошадиными
(Equidae)
мы должны ожидать
появления
огромной вариабильности
внутри гоминид во время взрывной
фазы их эволюции. Генетическая
изменчивость олицетворяет
эволюционную
потенцию» (стр.
167).
Качественно новый прогрессивный
сдвиг в процессе эволюции высших
приматов,
а тем
самым и всего
животного мира,
органически породил
и новое, активное
приспособление человеческих
существ к окружающей природной
среде
с внесением в нее все более
и более существенных
изменений. А
это требовало
беспрерывного
усиления вариантности тела
и ума в новых
жизненных действиях, в поведении.
Эта вариантность была
качественно отличной
от свойственной антропоидам.
Однако не следует упускать
из виду, что новооткрытая первыми
людьми
примитивнейшая трудовая функция
была еще глубоко
непривычна для них, возникших
из животного мира. Ведь прочие
двуногие
антропоиды,
пользовавшиеся
предметами
как орудиями,
не преодолели
барьера труда, вымерли,
были истреблены. Рубикон труда
для этих неудачных «попыток»
гоминизации по причине
меньшей вариантности
оказался
непреодолим. Путь к
истинному очеловечению
для них оказался закрыт навсегда
(Якимов, 1967).
Пусть процесс гоминизации закономерно
испытывали в условиях наземного
образа жизни десятки:
двуногих антропоидов
плиоцена и плейстоцена,
однако «марафон»
был завершен лишь одним из
них. И поэтому появление древнейших
людей на Земле носит характер
исключительного, единичного
события
на фоне
более
широкой
закономерности — гоминизации
родственных антропоидов.
Богатство условных рефлексов
стало особенно велико у людей
из-за сложности
общественной
среды, процессов
производственной
трудовой
деятельности
и постоянного общения с себе
подобными, благодаря высшему
развитию головного
мозга. Для
человека характерны высокоразвитые
и качественно отличные от животных
психика
и поведение,
что обусловлено развитием
у него
второй сигнальной
системы,
кроме первой, общей с животными.
По И. П. Павлову, у людей
наряду с первой сигнальной
системой условных рефлексов
в процессе
исторического развития возникла
и сформировалась новая
система сигналов в виде устных
и графических
обозначений того, что люди
непосредственно воспринимают
из внешнего мира
и из своего внутреннего.
245
Вторая сигнальная система —
характерное отличие
мышления
человека
Задумываясь в итоге своей
свыше чем полувековой
богатейшей научной деятельности
над отличиями
человека от мира
животных, И. П. Павлов
полагал найти качественные
особенности прежде всего
в
строении и функциях головного
мозга, в
высшей нервной
деятельности.
В поисках
того особого свойства,
которое
характеризует
человека и способствует
формированию только что
указанных его особенностей,
ой приходит
к развитию
понятия о
второй сигнальной
системе.
И. П. Павлов писал: «В
развивающемся животном
мире на фазе человека
произошла чрезвычайная
прибавка
к механизмам
нервной
деятельности.
Первая сигнальная
система действительности
есть и у человека. Но
слово составило
вторую,
специально
нашу, сигнальную
систему
действительности,
будучи сигналом
первых сигналов.
Многочисленные раздражения
словом, с одной стороны,
удалили нас
от
действительности, и поэтому
мы постоянно должны
помнить это,
чтобы
не исказить наши отношения
к действительности. С
другой стороны, именно
труд и связанное с ним
слово сделали нас людьми,
о чем,
конечно,
здесь
подробнее говорить
не приходится.
Однако не подлежит сомнению,
что основные законы,
установленные
в работе первой сигнальной
системы, должны также
управлять и второй,
потому
что эта работа все той
же нервной ткани» {Поли.
собр.
трудов,
1949, т. III,
стр.
568—569).
Говоря об общности закономерностей
деятельности клеток и
проводящих путей нервной
системы
при осуществлении функций
первой и второй сигнальной
систем
в области звукового языка,
И. П. Павлов отдавал
себе
ясный отчет в том, что
оба способа общения качественно
далеко
не
равноценны,
причем
не только
потому,
что один из них развился
на основе другого, а
и вследствие
того,
что слово, членораздельная
речь,
действительно
принадлежат к
числу самых
разительных
отличий человека от животного
мира.
В своих сочинениях И.
П. Павлов вновь и вновь
отмечает
принципиальное
различие
между первой
и второй сигнальными
системами:
«Конечно, слово для человека
есть такой же реальный
условный раздражитель,
как и все остальные общие
у него с животными,
но вместе с тем и
такой многообъемлющий,
как никакие другие, не
идущий
в этом отношении
ни
в какое количественное
и качественное сравнение
с условными раздражителями
животных. Слово,
благодаря всей
предшествующей
жизни взрослого человека,
связано со
всеми внешними и
внутренними раздражениями,
приходящими в большие
полушария, все их сигнализирует,
все их заменяет
и потому может
вызвать все
те действия,
реакции организма,
которые обусловливают
те
раздражения»
(Полн. собр. соч., 1951,
т. IV, стр. 428—429).
Учение И. П. Павлова
помогает глубже осознать
утверждение
Энгельса о
важнейшей роли,
которую наряду с трудом
играла членораздельная
речь в жизни
246
человека, одновременно
влияя на его организм.
Головной
мозг и органы
чувств сильно
преобразовались под
совокупным воздействием
труда
и речи, приобрели
новые качества и
усовершенствовались в прогрессирующем объединении
жизненных отправлений.
Специалисты по мозгу
уточняют локализацию
речевой функции,
в частности, путем
микроструктурного
изучения коры полушарий
большого
мозга.
В Институте мозга
Академии медицинских
наук СССР осуществлены
большие
исследования лобной
доли (Е.
П. Кононова), нижнетеменной
(Ю. Г.
Шевченко),
височной (С.
М. Блинков),
по онтогении коры
полушарий (Г. И.
Поляков), по
затылочной области
(И.
Н. Филимонов),
зрительным буграм
(М. М. Курепина).
Важны
также работы
по кровоснабжению
коры полушарий большого
мозга, произведенные
Б. Н. Клосовским
в Институте педиатрии
Академии медицинских
наук СССР.
Помимо изучения особенностей
коры головного мозга
человека, стоящих
в теснейшей
связи с речевой
функцией (Войно,
1964), интерес представляют
исследования
звукопроизводящего
аппарата. В последние
годы появился
ряд работ
анатомического
и антропологического
характера,
посвященных
гортани.
Обращает на себя
внимание исследование
В. В. Бунака (1951,
1966а), изучившего
строение
гортани человека
и обезьян в связи
с развитием речи
(см.
также:
Хрисанфова, 1956).
Все перечисленные
выше исследования
имеют
важное значение для
понимания формирования,
функций и строения
человеческого мозга,
развития речи,
мышления. По И. П.
Павлову, речь и мыслительная
способность
человека
неразрывно связаны
между
собой. В 1932 г.
он писал
«относительно той
прибавки, которую
нужно
принять, чтобы в
общем виде представить
себе и человеческую
высшую
нервную деятельность.
Эта прибавка касается
речевой
функции, внесшей
новый
принцип в деятельность
больших полушарий.
Если наши
ощущения и
представления, относящиеся
к окружающему миру,
есть для нас первые
сигналы
действительности,
конкретные сигналы,
то речь, специально
прежде всего кинэстези-ческие
раздражения,
идущие
в кору
от речевых органов,
есть
вторые
сигналы, сигналы
сигналов.
Они
представляют
собою отвлечение
от действительности
и
допускают обобщение,
что и
составляет наше лишнее,
специально человеческое
высшее мышление...»
(Полн. собр.
трудов, т. III, 1951,
стр. 232).
Трудно переоценить
то значение, которое
имеют
эти положения
И. П. Павлова
для науки о человеке,
его
разуме и сознании.
Советские антропологи
должны усилить планомерные
исследования
важнейших
в эволюционном отношении
анатомо-физиологических
особенностей человека
в сравнении с обезьянами
и другими высшими
млекопитающими. На
первой очереди здесь
стоят головной
мозг, ведущие рецепторы,
голосовой
аппарат,
рука, антропологическое
изучение которых
должно идти в плане
приспособленности
человеческого тела
к прямохождению,
труду,
речевой
247
функции, в разрезе
онто- и филогенетического
развития.
Большая близость
между человеком
и обезьянами
в анатомо-физиологических
особенностях
головного мозга
и органов чувств,
основанная
на близком филогенетическом
родстве, позволяет
искать и заметное
сходство
в чертах их высшей
нервной
деятельности,
несмотря на
то, что именно
здесь различия
особенно
резкие в силу
совершенно особого
пути
формирования
человеческого
сознания.
О психике И.
С. Беритов пишет:
«Сознательная
психонервная
деятельность
человека
есть особая,
новая форма психической
деятельности,
развившаяся
на основе
последней путем
исторического
развития трудовых
процессов. Короче
говоря,
сознание с самого
начала является
общественным
продуктом
и останется
таковым, пока
существует человек»
(Беритов,
1969,
стр. 225).
Чтобы объяснить,
как могла зародиться
специфически
человеческая
трудовая деятельность,
необходимо
обратиться, в
частности, к
тем ее истокам,
которые кроются
в палеобиологии
верхнетретичных
антропоидов.
Среди последних
были
и предки человека,
у которых
должны были развиться
зачаточные формы
трудовой деятельности.
О переходе наших
предков к пользованию
орудиями
можно до некоторой
степени судить,
изучая особенности
поведения
обезьян в искусственных
условиях опыта.
Многочисленные
наблюдения
и
опыты отечественных
ученых показали,
что обезьяны,
будучи поставлены
в такую
ситуацию,
когда они не
могли доставать
пищу
руками, оказывались
способными целесообразно
применять палки
и
другие
предметы и добывать
с их помощью
приманку.
Проводя параллели
между типами
поведения обезьян
и человека,
указывая на черты
сходства и различия
между
ними, необходимо
подчеркнуть,
что поведение
современного
человека
является в первую
очередь обусловленным
влияниями
общественной
среды. Необходимо
учитывать
глубокие качественные
отличия человеческого
поведения.
Следует указать
также на коренное
различие
между поведением
современных
людей и поведением
прочих животных,
проистекающее
из различия
в
характере мышления.
Поведение обезьян
нельзя оценивать
только как
индивидуальное.
Эти животные
живут, как правило,
стадами, и их
поведение в очень
сильной степени
отражает влияние
условий стадного
сожительства,
что
имеет немалое
значение для
понимания антропогенеза.
Действительно,
нельзя представить
себе, чтобы труд
в форме, свойственной
только человеку,
т.
е. в форме общественной
деятельности,
мог бы возникнуть
у
обезьян,
которые не вели
стадного образа
жизни.
Большую роль
в развитии сознания
необходимо
приписать общественной
среде с ее мощным
влиянием на процессы
взаимодействия
между развивавшимися
трудом,
мозгом и речью.
Для того
чтобы уяснить
возникновение
общественности
у древнейших
людей, следует
остановиться
на
явлениях стадности
у приматов.
248
Оглавление
| Предисловие |
5
|
Часть I. Дарвиновская и другие гипотезы антропогенеза
|
7
|
| Глава первая |
Дарвин о происхождении человека |
8
|
| |
Идея антропогенеза до Дарвина
|
8
|
| |
Дарвин об эволюции животного мира
|
13
|
| |
Человеческая родословная по Дарвину
|
17
|
| |
Очерк развития знаний о приматах
|
29
|
| |
Развитие приматологии в СССР |
40
|
| Глава вторая |
Человекообразные обезьяны и их происхождение |
56
|
| |
Современные антропоиды
|
56
|
| |
Ископаемые антропоиды |
70
|
| Глава третья |
Позднейшие гипотезы происхождения человека
и их критика
|
95
|
| |
Религиозные трактовки антропогенеза
|
95
|
| |
Тарзиальная гипотеза
|
99
|
| |
Симиальные гипотезы
|
104
|
| |
Гипотеза антропогенеза Осборна
|
111
|
| |
Гипотеза антропогенеза Вейденрейха
|
119
|
| |
Некоторые факторы гоминизации и вымирания ископаемых антропоидов плиоцена
и плейстоцена |
124
|
| Часть II.Особенности строения человеческого тела
и возникновение древнейших людей |
133
|
| Глава первая |
Человек как примат |
134
|
| |
Черты приспособленности человеческого тела к прямохождению
|
134
|
| |
Характерные отличительные особенности человеческого тела, не стоящие
в непосредственной связи с прямохождением
|
140
|
| |
Специальные черты сходства между человеком и антропоидами
|
145
|
| |
Рудименты и атавизмы у человека |
148
|
| Глава вторая |
Роль труда и прямохождения в антропогенезе
|
154
|
| |
Роль труда
|
154
|
| |
Способы передвижения у человекообразных обезьян
|
157
|
| |
Вес тела в центр тяжести у человека и обезьян
|
165
|
| |
Нижние конечности
|
168
|
| |
Костный таз, позвоночник и грудная клетка
|
173
|
| |
Верхние конечности
|
180
|
| |
Пропорции тела и асимметрии
|
185
|
| |
Череп |
189
|
| Глава третья |
Головной мозг и высшая нервная деятельность
человека и обезьян
|
199
|
| |
Головной мозг и анализаторы человека и обезьян
|
199
|
| |
Развитие периферических частей анализаторов
|
210
|
| |
Высшая нервная деятельность обезьян
|
226
|
| |
Вторая сигнальная система — характерное отличие мышления человека |
246
|
| Глава четвертая |
Стадность у обезьян и зачаточные формы труда
|
249
|
| |
Стадность у обезьян
|
249
|
| |
Зачаточные формы труда
|
263
|
| |
Антропогенез и его факторы |
272
|
Часть III. Формирование человека по данным палеоантропологии
|
233
|
| Глава первая |
Первая ступень: древнейшие люди (питекантропы)
|
284
|
| |
Питекантроп острова Явы
|
284
|
| |
Синантроп
|
292
|
| |
Гейдельбергский человек |
305
|
| Глава вторая |
Вторая ступень: древние люди (палеоантропы)
|
309
|
| |
Ледниковая эпоха
|
309
|
| |
Неандертальцы и их физический тип
|
314
|
| |
Находки неандертальцев в СССР
|
322
|
| |
Палестинские неандертальцы
|
330
|
| |
Образ жизни древних людей
|
339
|
| |
Развитие мозга у ископаемых людей |
343
|
| Глава третья |
Современные люди (неоантропы)
|
356
|
| |
Человек верхнего палеолита
|
356
|
| |
Ошибочные гипотезы происхождения современного человека и их критика
|
370
|
| |
Человеческие расы
|
380
|
| |
Факторы формирования рас
|
387
|
| |
Наука против расизма |
394
|
| Summary |
|
403
|
| Приложение 1. |
Предложения по биологическим аспектам
расовой проблемы
|
406
|
| Приложение 2. |
Декларация о расе и расовых предрассудках
|
410
|
| Литература |
|
414
|