Глава вторая
Человекообразные обезьяны
и их происхождение
Современные антропоиды
По строению тела человек должен быть отнесен к млекопитающим животным,
которых насчитывается свыше 4 тыс. видов (4237 — по Д. Моррису — Desmond
Morris,
1965). Человеку свойственны такие основные признаки млекопитающих, как млечные
железы, волосяной покров и постоянная температура тела.
В классе млекопитающих различают три подкласса: 1) яйцеродящие, или однопроходные
(примеры: утконос, ехидна); 2) сумчатые, или двуутробные (кенгуру, опоссум);
3) плацентарные, или одноутробные (собака, лошадь, мышь, крот, слон, обезьяна).
Третий подкласс, самый обширный, состоит из таких млекопитающих, у которых
во время родов выходит послед: детское место, или плацента, пупочный канатик
и плодные оболочки. Человек как примат относится к подклассу плацентарных
млекопитающих. Отряд приматов установлен Линнеем еще в 1758 г. и в настоящее
время включает человека, обезьян Старого и Нового Света, долгопятов
и лемуров (полуобезьян). К числу приматов, по мнению некоторых зоологов и
антропологов, следует присоединить тупаий, выключив их из насекомоядных (рис.
5). Характерные признаки у большинства приматов следующие: относительно крупный
головной мозг с выраженной височной долей, с менее развитым обонятельным
отделом и более развитым зрительным, с некоторыми особыми бороздами (шпорная
борозда на затылочной доле); пятипалые хватательные конечности с ногтями
на пальцах, большой (первый) палец противопоставляется другим; есть ключицы;
сбоку глазной орбиты костная стенка или полукольцо; зубы разнородные; желудок
простой формы; пара млечных желез на груди. Приматы рождают одного, реже
двух (или трех) детенышей. Живут на деревьях. Способы передвижения: лазание,
беганье и прыганье по ветвям. Питаются преимущественно растениями, у многих
в состав пищи входят также насекомые и другие мелкие животные. Приспособленность
приматов к древесному образу жизни обусловила мно-
56
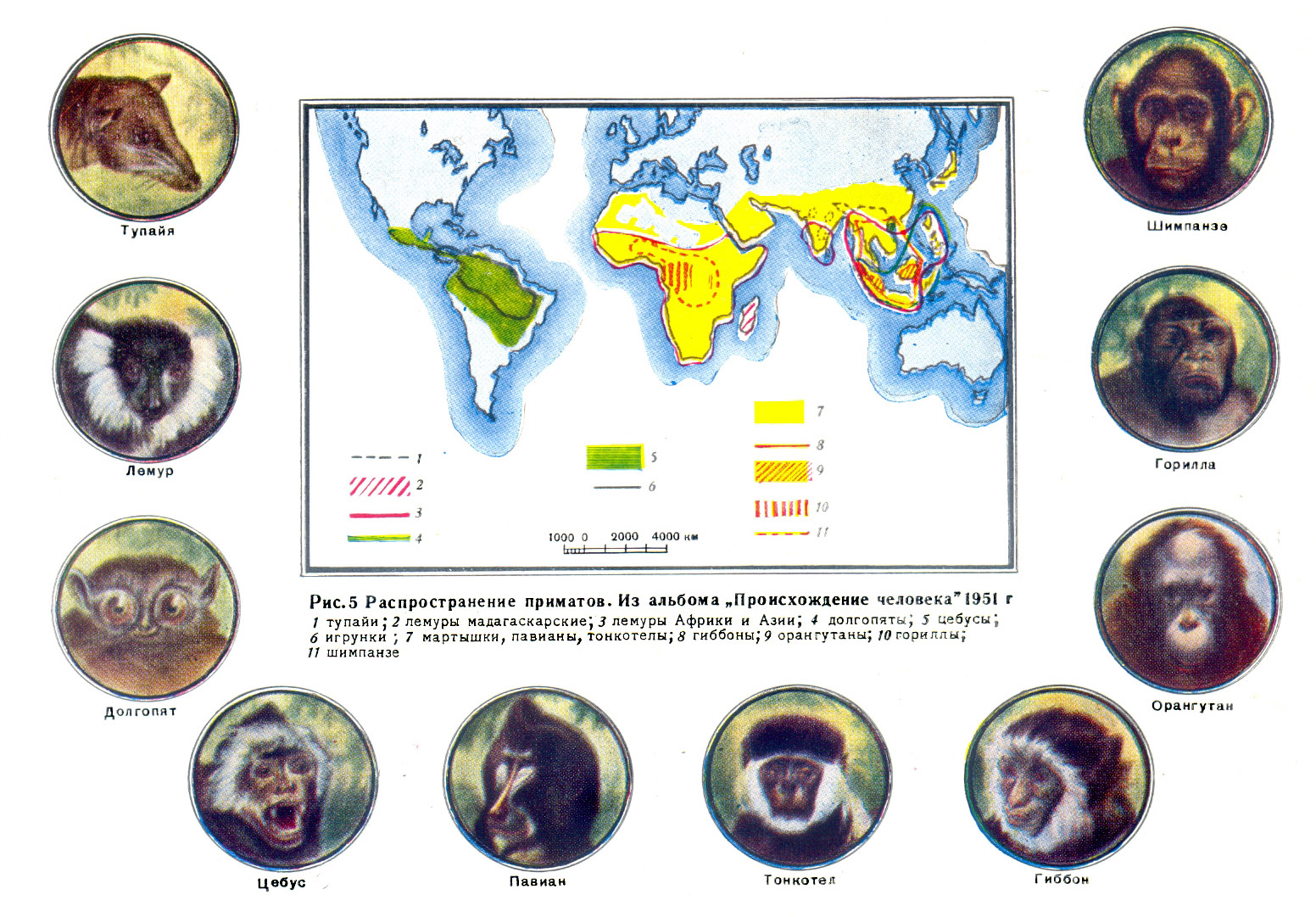
Рис. 5 Распространение приматов. Из альбома "Происхождение человека" 1951
г.
1 тупайи; 2 лемуры мадагаскарские; 3 лемуры Африки и Азии; 4 долгопяты;
5 цебусы; 6 игрунки; 7 мартышки, павианы, тонкотелы; 8 гиббоны; 9 орангутаны;
10 гориллы; 11 шимпанзе
Рис. 6. Низшие обезьяны Азии и Африки
1 — яванский макак моньет (Macacus cynomolgus
Anderson); родина — острова Малайского архипелага; 2 — белохвостая
гвереца мбега (Colobus
caudatus Thomas), Эфиопия Из архива Института антропологии,
Москва (1), и по Л. Геку из О. Абеля, 1931 (2)
|
гие характерные для них анатомо-физиологические особенности (Жедепов, 1962;
Нестурх, 1962).
Из приматов по степени близости к человеку первое место занимают человекообразные
обезьяны, второе — мартышковые (рис. 6), третье — американские обезьяны (рис.
7), четвертое —долгопяты, пятое — лемуры, последнее место — тупайи. Одной
из черт сходства между человеком и обезьянами Старого Света служит значительная
узость носовой перегородки по сравнению с обезьянами Нового Света, которых
даже называют широконосыми (или плосконосыми) в отличие от узконосых обезьян
Афревразии. Для человека характерна совокупность особенностей, из которых
должны быть в первую очередь названы: очень крупный и высокоразвитый головной
мозг; кисть с сильноразвитым и хорошо противопоставляющимся большим пальцем;
стояние и передвижение на двух ногах; стопа опорного типа с упругим продольным
сводом, причем большой палец развит лучше остальных, но неспособен им противопоставляться
(Нестурх, 1960).
В семейство человекообразных обезьян, или антропоидов, входят горилла, шимпанзе,
орангутан и гиббоны (рис. 8—10). Из них к человеку ближе всего обитающие
в Африке шимпанзе и горилла. Азиатские антропоиды отстоят несколько дальше,
особенно гиббоны — «малые антропоиды». Считают, что гиббоны составляют своего
рода переходную ступень от низших мартышкообразных обезьян к высшим, крупным
антропоидам (Вебер, 1936; Нестурх, 1941).
Все человекообразные обезьяны
отличаются от низших обезьян Старого Света — мартышек, макаков, павианов
и тонкотелов — отсутствием хвоста, седалищных мозолей (кроме гиббонов)
и защечных мешков. Кроме того, у разных низших обезьян есть подшерсток, сама
же шерсть
нередко гораздо гуще, чем у высших. У гиббонов хвоста и защечных мешков
нет, но имеются очень густой волосяной покров и седалищные мозоли.
По размерам и весу тела гиббоны также сильно уступают дру-
57
Рис. 7. Обезьяны Америки
1 — капуцин белолобый (Cebus albifrons Humboldt, 1911); родина — Никарагуа, Коста-Рика,
Колумбия, Гвиана, Бразилия; 2 — игрунка обыкновенная (Hapale jacchus Linnaeus);
родина — Восточная Бразилия. По Д. Ж. Эллиоту, 1912
|
гим антропоидам, равно как и по развитию головного мозга. Действительно,
длина тела даже у самых крупных гиббонов не превышает одного метра, вес
не достигает более 18 кг. Мозг весит всего лишь 100—115 г,
в то время как у
гориллы вес мозга доходит до 500—600 г, а в исключительных
случаях и до 650—752 г, т. е. больше, чем у шимпанзе или орангутана. Горилла
(рис.
11) превосходит этих антропоидов и длиной тела. Самцы гориллы достигают
высоты
180 см, а отдельные экземпляры и 2 м, вес же доходит до 200—300 кг.
Самки горилл гораздо мельче и легче, как и у орангутанов, самцы которых
достигают высоты 150 см, а веса 100—150 и даже свыше 200 кг. Самцы шимпанзе
легче,
но и они бывают ростом 150 см, однако вес их не больше 60 — 70 кг (самки
на 5 — 10 кг легче).
В свете приведенных данных можно называть гиббонов малыми антропоидами,
в отличие от крупных антропоидов, с которыми по росту и весу человек имеет
гораздо больше общего. Так, если принять, что средний рост мужчины колеблется
большей частью между 165— 170 см, а вес между 65—70 кг,
то ясно, что человек ближе к антропоидам типа шимпанзе (рис. 12—13) или
гориллы, чем к гиббонам.
Кроме того, у гиббонов в размерах тела и весе половые различия ничтожны,
в то время как у женщин длина тела в среднем меньше на 10 см,
а вес на 10
кг, чем у мужчин.
Из особенностей внутреннего строения антропоидов упомянем парные гортанные
мешки, дости-
58

|
гающие у них очень сильного развития (кроме некоторых гиббонов). Эти мешки
служат им резонаторами или усилителями производимых звуков. Что касается
силы голоса антропоидов, то путешественники не раз сообщали о том,
какие оглушительные крики издают в тропических лесах, например, стада шимпанзе
или гиббонов. Громкий рев производят также гориллы и орангутаны.
Гортанным мешкам антропоидов соответствуют у человека так называемые
морганьевы желудочки, располагающиеся в гортани тотчас выше истинных
голосовых связок:
эти крошечные выпячивания являются рудиментами гортанных мешков, некогда
функционировавших у ископаемых антропоидов, и свидетельствуют о человеческой
родословной. Что же касается низших обезьян, то у них морганьевы желудочки
отсутствуют, а имеющийся гортанный мешок непарный и совершенно иного
происхождения.
Другой важной особенностью строения может служить червеобразный отросток
слепой кишки, или аппендикс: он характерен для всех антропоидов, причем
у них он длиннее, чем у человека. У низших обезьян аппендикс отсутствует.
Следовательно,
и здесь человек тесно смыкается с группой человекообразных обезьян.
Рис. 8.
Гиббоны
1 — самка сиамского гиббона (Hylobates concolor Schlegel) с детенышем
«Примо», родившимся в 1938 г. в зоологическом саду г. Сан-Диего (США); 2 —
сростнопалый гиббон, или сиаманг (Symphalangus syndactylus F. Cuvier);
родина — остров Суматра.
По Б. Дж. Бенчли, 1944 (1), и по Р. и А. Йерксам, 1934 (2)
59

Рис. 9 Орангутаны (Pongo pygmaeus )
1 — самец; 2 — самка с детенышем; родина — острова Суматра и Борнео. По Цукерману,
1933 (1), и по Р. и А. Йерксам, 1934 (2)
|
Наряду с бросающимися в глаза чертами подчас удивительного сходства
во внешнем облике и внутреннем строении между человеком и антропоидами
существуют
и
очень заметные различия, которые в наибольшей мере зависят от различий
в способе передвижения, характере питания и образе жизни.
Коротко говоря, антропоиды— это высокоразвитые обезьяны, приспособленные
к жизни в глухих тропических лесах (Buettner-Janusch, 1962—1964;
Reynolds, 1967; J. R. and P. H. Napier, 1967). Там они перелезают
с ветки на
ветку, с дерева на дерево, очень часто перебрасываются с помощью
только одних
рук, причем поджимают ноги.
Однако на земле эти обезьяны (антропоиды) большей частью передвигаются
на четырех конечностях, держа туловище в полувыпрямленном положении:
их передние
конечности — руки — длиннее задних, или ног, вследствие более энергичного
употребления первых при передвижении по деревьям (рис. 14, 15).
Из антропоидов преимущественно наземный образ жизни ведет горилла.
Но и она не утратила некоторых привычек древесного образа жизни:
в случае
опасности
горилла может спрятаться на дереве, а каждый вечер делает в развилке
ветвей новое крупное гнездо, которое выстилает мягкими листьями,
травой, мхом
для
полноты ночного отдыха, или на земле (Шаллер, 1968).
Строят гнезда также шимпанзе и орангутаны, причем первые делают
их и на земле для дневного отдыха (Ладыгина-Котс, 1960).
60
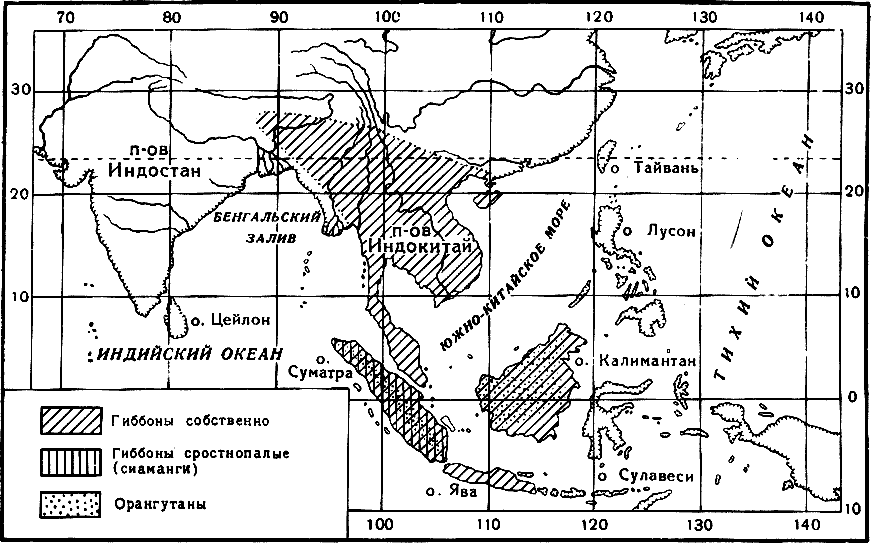
Рис. 10. Распространение гиббонов и орангутанов
По А. Брему, 1920 (несколько изменено автором)
Что же касается гиббонов, то они гнезд не строят, как и все остальные
обезьяны. Если у предков человека и был в свое время инстинкт
постройки гнезда на
деревьях или, как это делают шимпанзе, также дневного ложа
на земле, то в настоящее
время от такой привычки у современного человека, даже в раннем
детском волрасте, не осталось и следов.
Пищей для крупных антропоидов служат плоды, листья, цветы,
побеги и другие части растений. Гиббоны, тоже предпочитающие
растительную
пищу,
успевают
во время полета захватить бабочку, птичку, любят также
пошарить в птичьих гнездах в поисках яиц или птенцов. Крупные антропоиды
разгрызают
своими
могучими зубами самые твердые плоды: их челюсти очень сильны,
а у гориллы и орангутана они имеют огромные размеры и гораздо
более мощны, чем у менее крупного шимпанзе.
Сильные различия анатомо-физиологического характера между
человеком и антропоидами в весьма значительной степени
ослабляются многими
чертами сходства между
ними, которое, без сомнения, свидетельствует о родстве.
Чтобы убедиться в этом, достаточно познакомиться, например, со
свойствами их крови,
с
особенностями размножения и хода индивидуального развития.
Белые кровяные шарики, или
лейкоциты,
шимпанзе и гориллы наиболее близки к человеческим по типу
сегментации ядра нейтрофилов и эозинофилов, так как в нем
сегментов мало,
а у макаков и
других обезьян их заметно больше.
61
Но еще важнее биохимические черты близости крови человека
и антропоидов. Об этом ярко свидетельствуют, например,
осаждение почти одинаковых
объемов белков при реакции преципитации и общность
групп крови.
Кроме большого
количества других, в большинстве второстепенных, изо-серологических
факторов у людей
встречаются следующие четыре главные группы крови:
I или 0 (ноль); II или А; III или Б; IV или АБ, образующие единую
систему.
Оказывается,
что такие
же группы крови среди приматов встречаются лишь у антропоидов,
хотя у
низших обезьян Старого Света и были обнаружены группы
крови, похожие на человеческие
(табл. 1).
 Сделанные
Ж. Труазье (1931) неоднократные переливания крови от
разных шимпанзе группы II (А) людям той же
группы прошли
вполне
успешно,
без каких-либо
отрицательных явлений. Гораздо раньше, полвека назад,
не менее успешными были произведенные Г. Фриденталем
(Friedenthal,
1900) опыты переливания
человеческой крови в кровеносную систему шимпанзе.
В то же время при опытах на низших обезьянах оказалось,
что, судя
по
красноватому
цвету
мочи, человеческие
эритроциты испытали в крови этих обезьян гемолиз: следовательно,
кровь человека оказалась для низших обезьян чужеродной.
Такое мнение подтвердилось
при помещении
цельной крови павиана в сыворотку человеческой крови,
так как эритроциты обезьяны разрушались. При подобном
же опыте
с цельной
кровью шимпанзе
ее эритроциты оставались в сыворотке человеческой крови
без изменений.
Сделанные
Ж. Труазье (1931) неоднократные переливания крови от
разных шимпанзе группы II (А) людям той же
группы прошли
вполне
успешно,
без каких-либо
отрицательных явлений. Гораздо раньше, полвека назад,
не менее успешными были произведенные Г. Фриденталем
(Friedenthal,
1900) опыты переливания
человеческой крови в кровеносную систему шимпанзе.
В то же время при опытах на низших обезьянах оказалось,
что, судя
по
красноватому
цвету
мочи, человеческие
эритроциты испытали в крови этих обезьян гемолиз: следовательно,
кровь человека оказалась для низших обезьян чужеродной.
Такое мнение подтвердилось
при помещении
цельной крови павиана в сыворотку человеческой крови,
так как эритроциты обезьяны разрушались. При подобном
же опыте
с цельной
кровью шимпанзе
ее эритроциты оставались в сыворотке человеческой крови
без изменений.
Много общего между человеком и антропоидами можно подметить
и в области сравнительной патологии приматов, как это
видно из
монографии
Т.
Ру, посвященной болезням
низших и высших обезьян, изученным в лабораторных условиях
(Ruch, 1959).
Близко и число хромосом: у человека — 46, а у крупных
антропоидов — 48, в то время как у низших узконосых
— от 54 до 78.
По мнению итальянского биолога Б. Киарелли, высказанному
на VII Международном конгрессе антропологов и этнографов
1964
г. в Москве,
сокращение
на одну пару хромосом в процессе антропогенеза произошло
путем слияния между
их сходными экземплярами.
62
Таблица 1
Группы крови у человека и обезьян (по данным
А. Э. Муранта, 1954)
Приматы* |
Наличие группы крови (+) или отсутствие
(—) |
0 |
А |
Б |
АБ |
| Человек |
+ |
+ |
+ |
+ |
| Шимпанзе |
+ (13) |
+ (110) |
— |
— |
| Горилла береговая |
— |
+ (13) |
— |
— |
| Горилла горная |
— |
— |
+ (2) |
— |
| Орангутан |
— |
+ (7) |
+ (8) |
+ (4) |
| Гиббоны |
— |
+ (1) |
+ (6) |
+ (1) |
| Макак резус (группы найдены в слюне) |
+ |
+ |
+ |
+ |
* В скобках обозначено количество исследованных особей
Вероятно, подобный генетико-эволюционный процесс имел место и в ходе развития
высших обезьян из низших. Во всяком случае у шимпанзе есть 12 хромосом,
подобных; человеческим, у гориллы — 10, у орангутана — 8, как установил Б. Киарелли.
А это — одно из
Рис. 11. Гориллы
1 — самец горной гориллы (Gorilla gorilla beringei Matsehie), no
кличке «Мбонго», зоологический сад в г. Сан-Диего, США; 2 — молодая
самка горной гориллы
«Мисс Конго» (Лаборатория биологии приматов во Флориде). По Б. Дж. Бенчли,
1944
(1), по Р. и А. Йерксам, 1934 (2)

63
Рис. 12. Шимпанзе обыкновенные (Pan chimpanse Meyer=Anthropopithecus
troglodytes Flower and Lydekker)
1 — самец (Нью-Йоркский зоологический сад), по Д. Ж. Эллиоту, 1912;
2 — взрослая самка «Белла» (Московский зоопарк). Родина — Экваториальная
Африка
лучших доказательств глубокого родства крупных антропоидов с человеком.
У гиббонов 44 хромосомы, у сиамангов — 50 (Reynolds, 1967).
Удивительные черты сходства между человеком и антропоидами отмечаются
также в области размножения. Сперматозоиды и яйцеклетки шимпанзе или
гориллы по
форме и размерам почти неотличимы от человеческих.
Беременность длится у шимпанзе от 210 до 252 суток, в среднем около
235 суток, у орангутана — около 275 суток, у человека — примерно 265—280
суток
(у гориллы,
вероятно, столько же). В этом отношении гиббоны ближе к низшим обезьянам,
так как у
них срок беременности не превышает 210 суток. Более сходны с человеческими
и плоды антропоидов (рис. 16) по сравнению с прочими обезьянами (Штарк,
1956; Harms, 1956).
На Сухумской медико-биологической станции Академии медицинских наук
СССР за 30 лет родилось свыше тысячи детенышей низших обезьян, в связи
с чем
были сделаны многочисленные наблюдения (Воронин, Канфор, Лакин, Тих,
1948; «Труды
Сухумской биологической станции Академии медицинских наук СССР», т.
I, 1949; Елигулашвили, 1955; Лапин, 1963; Фридман, 1968). За это время
здесь
родились
три двойни: у гамадрилов (две) и резусов (од-
64

Рис. 14. Стадо белоруких гиббонов — ларов (Hylobates lar)
Панно Е. Д. Самойленко-Машковцевой. Музей Института антропологии Московского
университета
Рис. 15. Орангутаны (Simla satyrus), самец и самка
Панно Е. Д. Самойленко-Машковцевой. Музей Института антропологии Московского
университета
на). Гораздо реже рождаются в неволе человекообразные обезьяны. Гориллы
впервые дали потомство в США 22 декабря 1956 г.
Двойня у горилл в неволе впервые родилась 3 мая 1967 г. в Зоологическом
саду во Франкфурте-на-Майне (ФРГ). Это были, очевидно, разнояйцевые
близнецы Алиса
и Эллен: первая отличается дружелюбным выражением лица, в то время
как вторая глядит с неприязнью, нетерпелива и агрессивна. Отмечены
и некоторые
различия
во внешности. Близнецов пришлось выкармливать искусственно, так
как мать отказывалась заботиться о них. Течение беременности и рождение
детенышей
у шимпанзе наблюдались начиная с 1915 г. Всего было около 75 случаев
рождения шимпанзе в неволе,
в том числе были две двойни (рис. 17). Врачи и научные работники установили
большое сходство между человеком и шимпанзе в ходе беременности
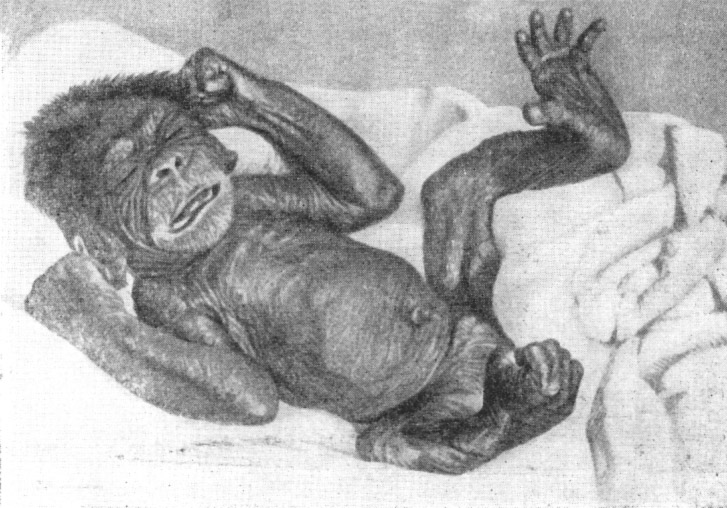
и акте родов. Да и сам новорожденный шимпанзе с густыми волосами на голове,
при их отсутствии
на теле (рис. 18), больше похож на новорожденного человека, чем
взрослый
шимпанзе на взрослого человека. Уровень развития скелета в утробный
период у антропоидов, судя по вторичным очагам окостенения, запаздывает
по сравнению
с низшими обезьянами и ближе к тому, какой наблюдается у человеческого
новорожденного (Воккен, 1949). Этот факт можно поставить в связь
с тем, что детеныш, например,
мартышки беспомощен лишь около двух-трех ме-
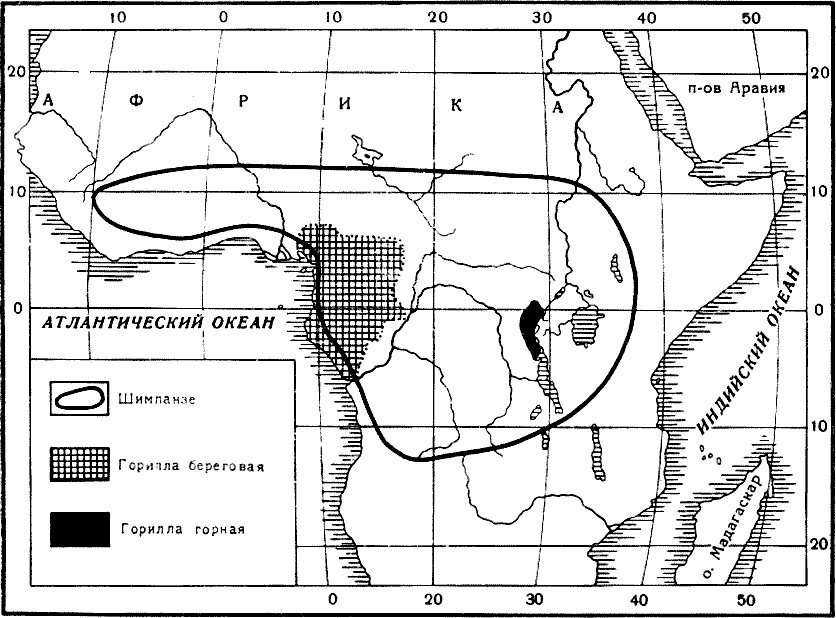
Рис. 13. Карта распространения африканских антропоидов: шимпанзе (по
А. Брему, 1920, дополнено автором) и гориллы (по Г. Дж. Кулиджу,
1929)
65
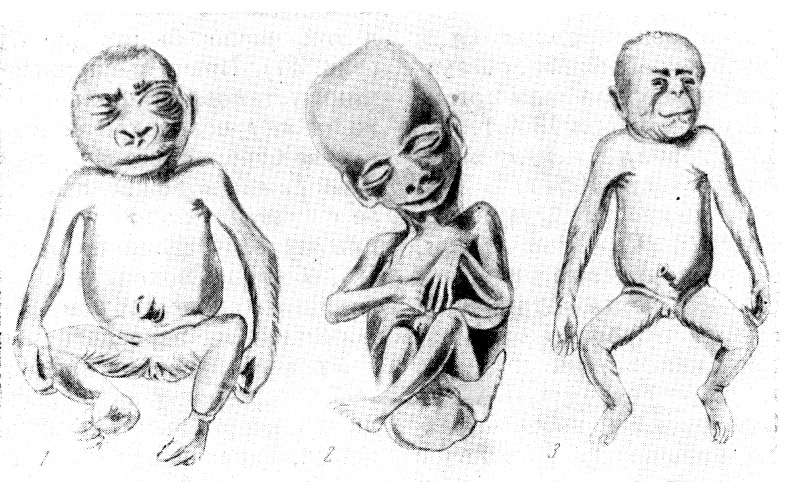
Рис. 16. Плоды человека и антропоидов
1 — горилла; 2 — человек; 3 — шимпанзе. По Г. Клячу, 1915 (1), Э. Зеленке,
1905 (2) и А. Шульцу, 1929 (3)
сяцев, а детеныш шимпанзе гораздо дольше, месяцев пять-шесть. Действительно,
детеныш шимпанзе начинает передвигаться самостоятельно только
после полугода жизни (рис. 19—21). Но еще лет до двух он держится на
теле матери или
в самой тесной близости к ней, сосет ее грудь, спит с ней в одном
гнезде.
В возрасте около года у детенышей шимпанзе прорезываются последние
из двадцати молочных зубов, которые после заметного, но не такого
длительного, почти
пятилетнего промежутка, как у ребенка, заменяются на постоянные:
на протяжении
десятка лет с небольшим сформировываются все тридцать два зуба.
В нарастании веса у детеныша шимпанзе были подмечены (Schultz,
1956) примерно те
же периоды, как и наблюдались у ребенка.
Половая зрелость наступает у самок шимпанзе к 8—10 годам, у самцов
позже, к 12 годам, а у горилл даже к 14 годам. Менструальные
явления у шимпанзе
наступают примерно один раз в 30 суток или даже через несколько
больший срок — до 40 дней.
Еще более резким отличием по сравнению с человеком служит то,
что при этом у самок шимпанзе (и горилл) формируются крупные
отечные
набухания
кожи в
виде так называемых менструальных подушек.
Подобное явление наблюдается у некоторых низших обезьян, а именно
павианов, макаков и мангобеев (Нестурх, 1946). У орангутанов
и гиббонов набухания
не образуются, хотя у беременных самок орангутанов нечто подобное
наблюдается.
66
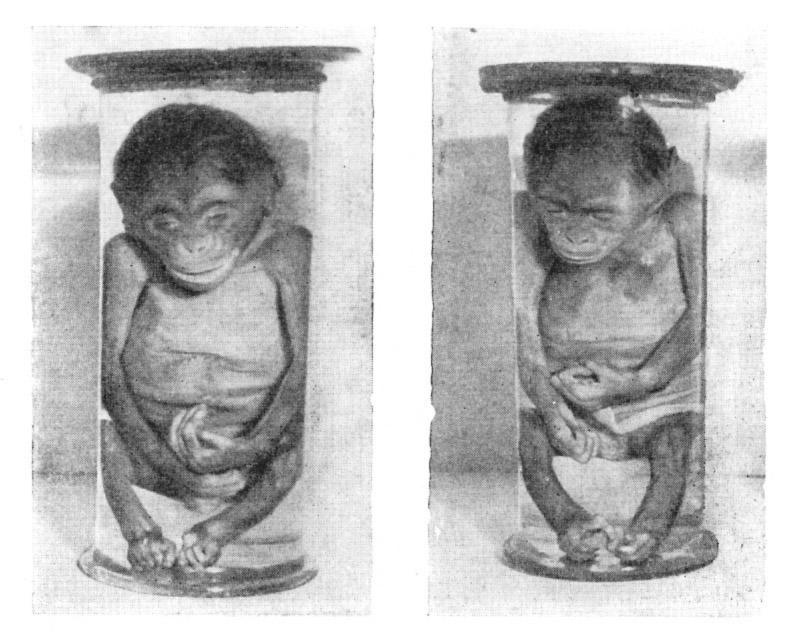
Рис. 17. Двойня шимпанзе: новорожденные самец (справа) и самка
(Московский зоопарк, 1939)
По препаратам экспозиции Института и Музея антропологии, Москва
Рис. 18. Новорожденный шимпанзе
По Р. и А. Йерксам, 1934
Рис. 19. Детеныш шимпанзе (слева) и гориллы
По Ф. Ч. Мерфилду,
1954
Рис. 20. Самка шимпанзе «Бубу» с детенышем «Джюбили»
14-ти дней и трех месяцев (Лондонский зоологический сад)
По С.
Цукерману, 1933

Какова длительность жизни антропоидов? Точные данные об их
долговечности в условиях неволи немногочисленны. Надо иметь
в виду, что человекообразные
обезьяны умирают преждевременно, главным образом от заболеваний
пищеварительного и дыхательного путей.
Об одной самке шимпанзе сообщали, что ей было около 60
лет (R. a. A. Jerkes, 1934). Достоверно, что один самец
шимпанзе,
привезенный
из
Африки, прожил
в неволе 39 лет и оставил большое потомство. Гориллы выживали
в неволе до 35 лет, а один орангутан прожил 31 год; такой
же срок
прожил один
гиббон,
о другом известно, что и спустя четверть века пребывания
в клетке, куда он попал уже взрослым, его мышцы были такие
же
крепкие,
зубы такие же
острые,
а характер столь же неукротимый, как и в первый день пребывания.
Павианы жили в неволе и свыше 50 лет. Даже лемуры довольно
долговечны: они
жили в неволе до 25 лет (Hill, 1953).
Отсюда мы можем сделать три заключения. Первый вывод: приматы
вообще долговечны по сравнению с большинством других
млекопитающих (кроме
слонов и китов).
Второй вывод: крупные антропоиды, по всей видимости,
живут на воле несколько десятков лет, а такие великаны среди
них, как
горилла
и орангутан, доживают,
может быть, и до возраста свыше 50—60 лет. Третий вывод:
современное долголетие человека в большой мере можно
объяснить довольно
обоснованным предположением
о значительной длительности жизни у его ближайших предков,
т. е. ископаемых крупных верхнетретичных антропоидов. Но, очевидно, долголетие
людей увеличилось вследствие
благоприятных влияний
общественных
условий и ныне долголетие людей заметно превосходит длительность
жизни антропоидов, доходя изредка даже до 100—150 лет
и выше.
Все приведенные нами факты не позволяют считать устаревшим
мнение Томаса Гексли о том, что человек находится в более
близком анатомо-физиологическом
родстве с человекообразными обезьянами, чем человекообразные
обезьяны с низшими, чисто четвероногими обезьянами.
Так как глубокое сходство между человеком и высшими обезьянами
обнаруживается также в системе органов размножения, то
уже довольно давно возникла
мысль о возможности успешного соединения половых клеток
высших приматов в искусственных
условиях.
Подобная идея высказана была семьдесят лет назад Гансом
Фриденталем на основании его опытов по сходству крови
человека и обезьян.
По отношению к другим млекопитающим
известны многие случаи межвидовой и более отдаленной
гибридизации. Известны довольно многочисленные случаи скрещивания приматов
в неволе среди полуобезьян,
среди обезьян Нового Света, а также обезьян Старого Света,
причем получались не только межвидовые, но изредка и
межродовые
гибриды
(Gray, 1954).
Замечательный случай получения межродового гибрида наблюдался
в начале 1949 г. на Сухум-
69
ской медико-биологической станции: самка павиана гамадрила
принесла детеныша женского пола Приму от самца макака,
бывшего в свою
очередь гибридом
от скрещивания свинохвостого макака лапундера с макаком
резусом. Прима наглядно совмещает
в себе признаки отца и матери. Вскоре от тех же родителей
появился еще один детеныш, гибрид женского пола Маркиза.
В результате межвидовой и более отдаленной гибридизации
среди обезьян на этой станции (ныне Институт экспериментальной
патологии
и терапии)
получаются
помесные обезьяны. Здесь изучаются не только их морфология,
физиология и патология, но и наследственные особенности.
Теоретически возможно предполагать, что гибридизация
между антропоидами и гоминидами могла бы быть осуществлена,
например, с самкой шимпанзе (Rohleder, 1918; R. and D.
Morris, 1966).

Рис. 21. Детеныш шимпанзе «Джорджи» (зоологический сад в г. Сан-Диего,
США)
По Б. Дж. Бенчли, 1944
|
Однако такого рода опыты сопряжены, конечно, с большими
затруднениями. Антропология и без этого располагает,
как мы уже отчасти видели,
очень большим количеством
данных, подтверждающих учение Дарвина о естественном
происхождении человека из мира животных. К ним присоединяются
многочисленные
находки костных
остатков человекообразных и низших обезьян третичного
и четвертичного периодов.
Ископаемые антропоиды
Полагают, что обезьяны Старого Света возникли из местных
полуобезьян в первую половину третичного периода, который
принято делить
на пять эпох. Из них
три более древние, а именно палеоцен, эоцен и олигоцен,
длились соответственно 10, 20 и 15 млн. лет (числа приблизительные)
и все вместе составляют
так называемый палеоген, а две более поздние эпохи —
миоцен и плиоцен, длившиеся
примерно 14 и 10 млн. лет,— образуют неоген. (Bomer,
1968).
Всего, таким образом, третичный период имел продолжительность
около 70 млн. лет. За этот огромный промежуток времени
произошло развитие
обезьян
из полуобезьян
и их дальнейшая эволюция. В ходе эволюции приматов образовались
многие древесные, а позже и наземные виды человекообразных
обезьян, среди
которых и развились
ближайт предки человека.
Об эволюции обезьян приходится судить в первую очередь
по их
70
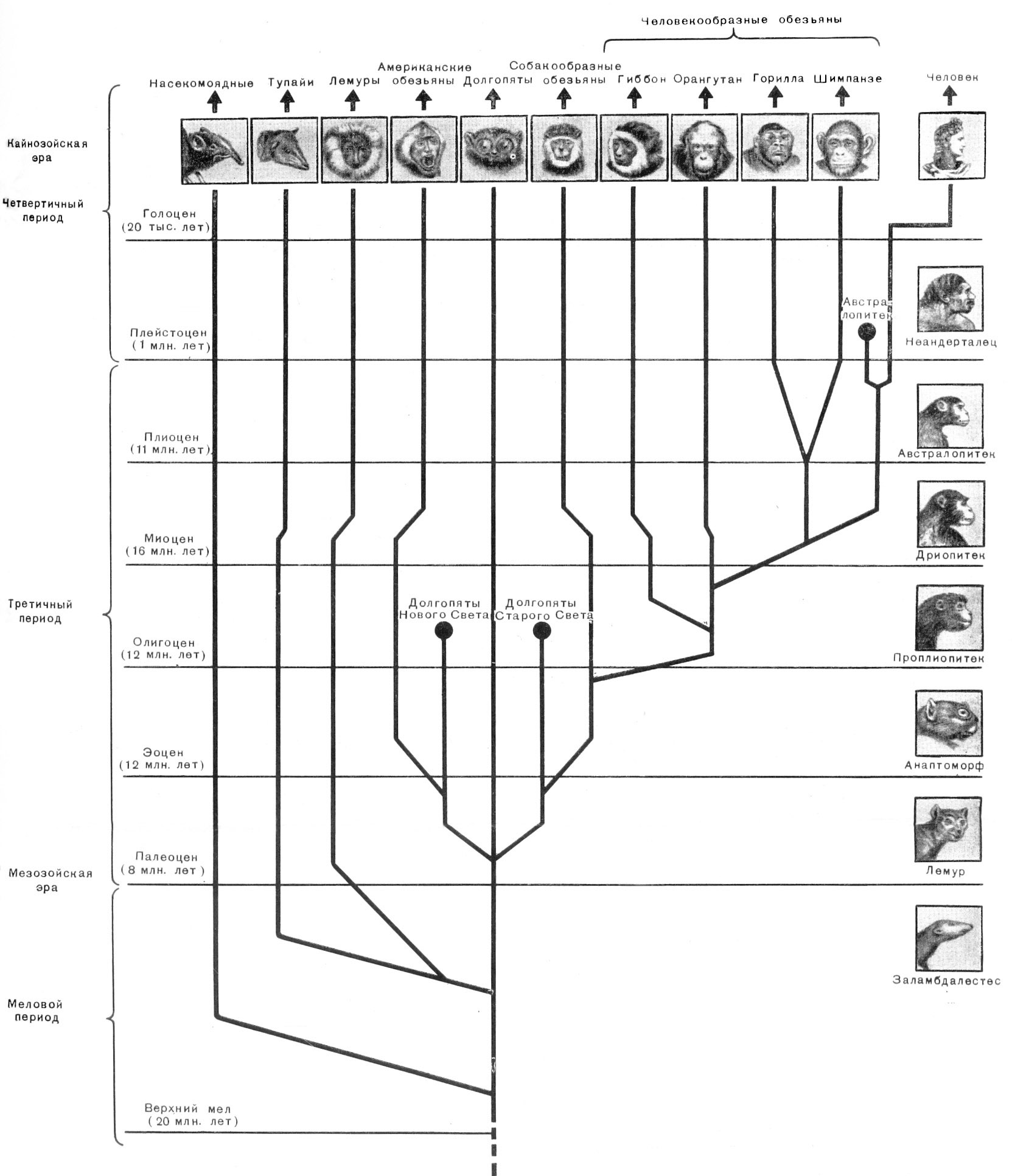
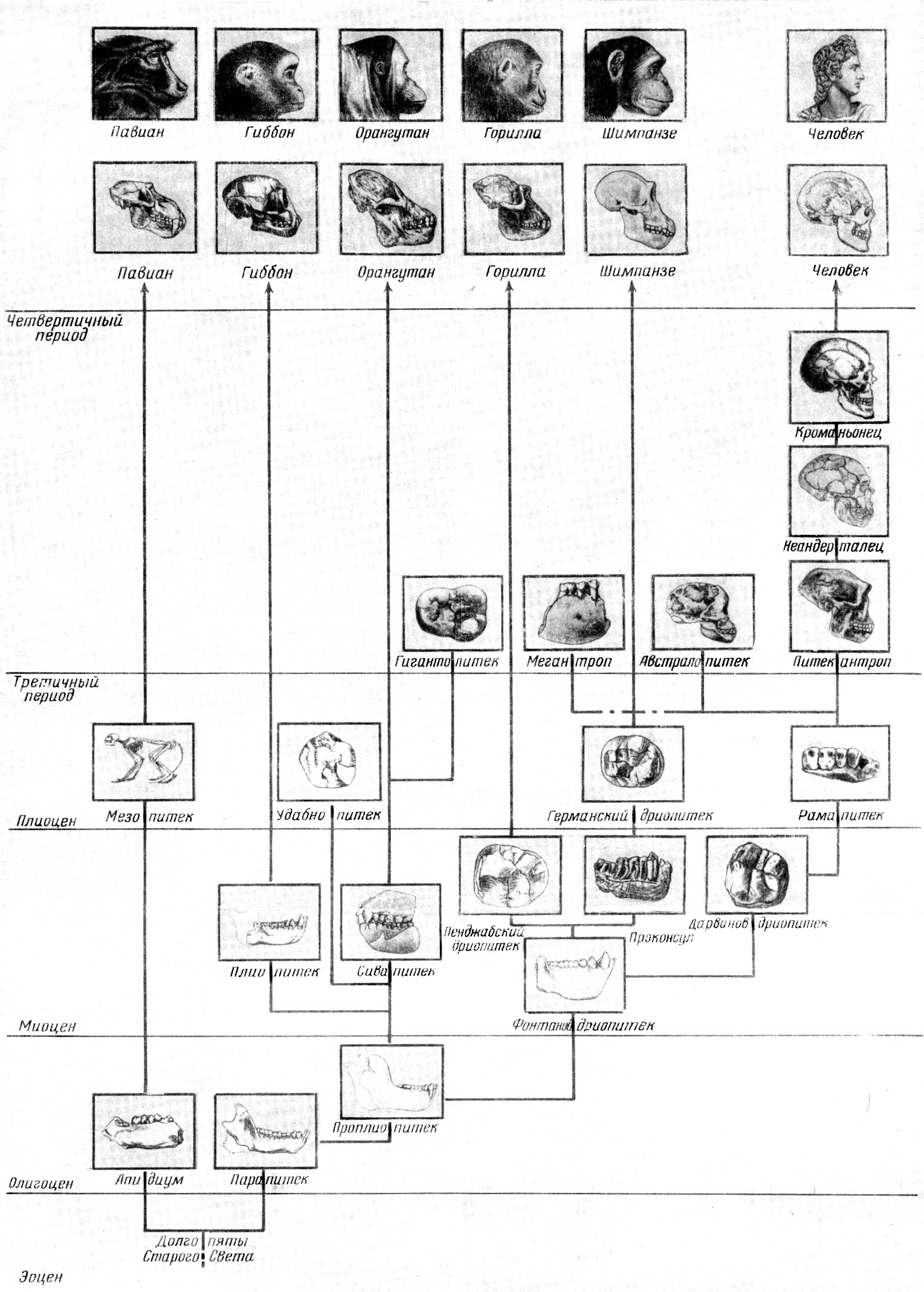
Рис. 22. Схема эволюции человека и приматов
По М. Ф.
Нестурху и С. А. Сидорову, 1954
Рис.23. Схема эволюции человека и обезъян
По М. Ф. Нестурху и С. А. Сидорову, 1954
(Те же рисунки, но выполненные на отдельном листе несколько большего размера)
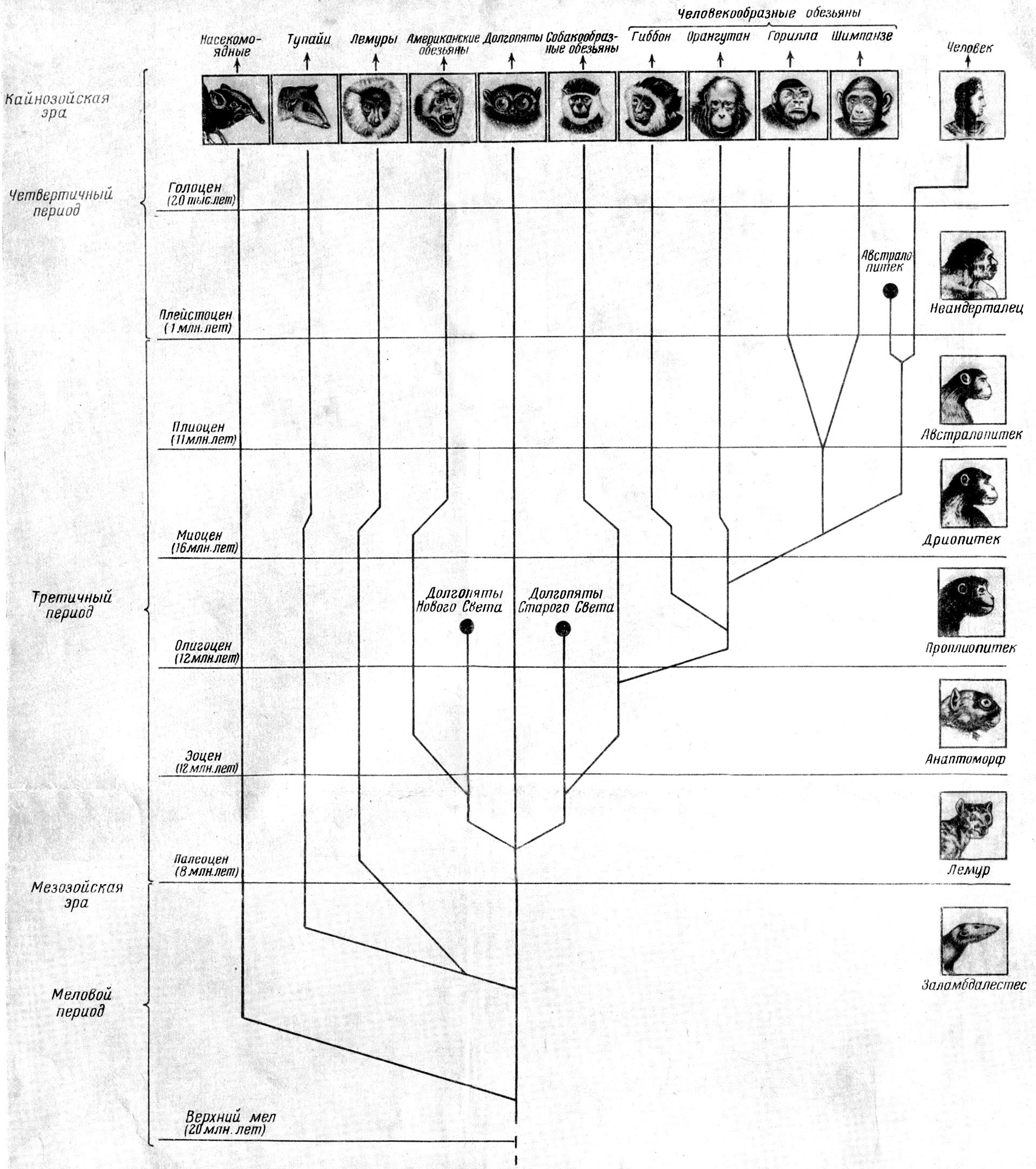
Рис. 22. Схема эволюции человека и приматов.
По М. Ф.
Нестурху и С. А. Сидорову, 1954.
Рис. 23. Схема эволюции человека и обезьян.
По М. Ф.
Нестурху и С. А. Сидорову, 1954.
костным остаткам, которые, к сожалению, представлены
до сих пор почти исключительно челюстями и
зубами; реже попадаются черепа и отдельные кости,
а тем более
скелеты. Все-таки
ископаемого материала
по обезьянам
теперь достаточно
для того, чтобы составить хотя бы общее понятие о путях
развития высших обезьян в третичном, а затем и четвертичном
периодах
(рис. 22—23).
Здесь большое
значение имеют форма, строение и размеры коренных
зубов.
Но как различить коренные зубы человекообразной обезьяны
и низшей? Для этого нужно внимательно рассмотреть их
жевательную поверхность,
так как
имеющиеся
бугорки на ней у разных групп обезьян неодинаково связаны
между
собой.
Так, у низших обезьян Старого Света, например, на нижних
коренных зубах четыре главных бугорка соединены попарно
(впереди — протоконид
с метаконидом,
а
позади — гипоконид с энтоко-нидом) высокими гребнями
эмали, идущими в поперечном направлении. Этого нет у
высших обезьян,
у которых
рисунок здесь другой,
а именно: высокие поперечные перегородки отсутствуют
и можно подметить лишь один, косо идущий невысокий валик.
По характеру расположения, или, как говорят, по узору
бугорков, человек явно относится к типу антропоидов.
Однако этот
узор у человека имеет
и свои заметные
особенности, развившиеся в ходе его эволюции после
ответвления от общего ствола верхнетретичных крупных человекообразных
обезьян. Именно на
нижних коренных
зубах современного человека основные четыре бугорка почти
равны друг другу по площади и поэтому ложбинки между
ними, или борозды,
образуют
более или
менее явно выраженный крестообразный узор, иначе называемый
«плюс-узором».
Между тем иногда вместо такого рисунка можно видеть на
некоторых современных человеческих коренных зубах другой
рисунок, обусловленный
тем, что
первый внутренний бугорок (метаконид) занимает большую
площадь, чем второй.
Вследствие такого его разрастания бороздка между упомянутым
бугорком и вторым внутренним
(энтоконидом) проходит дальше к заднему концу зуба, чем
бороздка между обоими главными наружными бугорками (про-токонидом
и
гипоконидом).
Подобный узор свойствен современным и ископаемым антропоидам
и сразу «выдает» обезьяний подслой в строении человеческих
зубов. А так как
подобное отношение
бугорков отмечено было у дриопитеков, составляющих
важное звено в человеческой родословной, то его назвали «узором
дриопитека».
Будучи обнаружен на
зубах человека, узор дриопитека является одним из самых
убедительных доказательств
происхождения человека от обезьян. В новейшее время
К. Коренгоф (Korenhof, 1960), применив оригинальный метод
изучения внутреннего
рельефа верхних
моляров, сумел обнаружить новые следы родства человека
с высшими обезьянами.
Зубная система человека и всех обезьян Старого Света
состоит из 32 постоянных зубов. В каждой
71
половинке челюсти сидят 2 резца, 1 клык, 2 малых коренных
зуба (иначе их еще называют предко-ренными, премолярами)
и 3 больших
коренных зуба,
или
моляра. Зубы у человека сидят тесно, без характерных
для обезьян промежутков между верхними клыками и резцами,
а
также между
нижними клыками и премолярами.
Отсутствие таких промежутков, или диастем, у человека
объясняется, в частности, тем что клыки у него невелики
(они не выдаются
из ряда зубов).
Но корни у
клыков длинные, очевидно, потому, что у наших предков
и коронка клыка была гораздо крупнее, вследствие чего
выдавалась
из зубного
ряда, как
у современных
антропоидов.
Последние третьи коренные зубы человека нередко сильно
задерживаются в ячейках и прорезываются поздно: их
называют «зубами мудрости».
Они имеют менее оформленный
характер, чем первый и второй моляры, и носят в значительной
мере черты рудиментарности. У некоторых людей один-два
зуба мудрости
могут совсем
не появляться из десен,
а в более редких случаях остаются в ячейках и все четыре
последних моляра.
Доказательства происхождения человека от обезьяны можно
найти и при изучении молочных зубов, которых у человека,
как и у
всех обезьян
Старого
Света,
лишь двадцать: в каждой половине челюстей у него прорезывается
два резца, один
клык и два коренных зуба. Очень интересно отметить
два обстоятельства: на молочных коренных зубах у каждого
ребенка видны мелкие бороздки
между бугорками, весьма сходные с теми, какие свойственны
шимпанзе, а на первых нижних коренных зубах можно видеть особый
бугорок (параконид),
располагающийся впереди всех остальных и представляющий
собой
наследие
от очень древних
предков
человека, именно от раннетретичных полуобезьян. Одной
из наиболее примитивных форм ископаемых антропоидов
является маленькая
обезьянка парапитек.
От нее известна только нижняя челюсть с зубами, найденная
Шлоссером в 1911
г. в
нижнеолигоценовых слоях близ г. Файюм (Египет), вместе
с челюстью несколько более крупной: человекообразной
обезьяны
проплиопитека
(рис. 24—25).
Если парапитек был величиной с кошку, то проплиопитека
по размерам тела можно
сравнить с небольшой собакой.
Судя по форме и другим особенностям челюсти и зубов,
парапитек был примитивнее, а проплиопитек более развит.
Однако узор
расположения бугорков на нижних
молярах у них такой же, как и у более поздних антропоидов
вплоть до
современных, т. е. с узором дриопитека.
Сравнительно большая примитивность парапитека следует,
в частности, из того, что клык его не выдается из зубного
ряда, так: что
сам Шлоссер за клык по
ошибке принял соседний резец, выступающий своей коронкой
выше соседних зубов. Шлоссер и посчитал зубную формулу
так: 1.1.3.3.
Он сближал
в этом
отношении
парапитека с одним из низших приматов, а именно с долгопятом,,
с которым у парапитека есть черта сходства в виде значительного
72
угла (33°) между половинками челюсти. Однако позднейшие
исследования челюсти парапитека, осуществленные крупным
сравнительным анатомом
и антропологом Густавом Швальбе, показали, что зубная
формула этой файюмской
обезьянки
такая
же, как у всех обезьян Старого Света: 2.1.2.3. К числу
примитивных форм высших приматов следует прибавить
недавно открытого альмогавера
из эоценовых
слоев
Испании, южный склон Пиренеев. Фрагмент нижней челюсти
с зубами изучили и описали Крусафонт и Вильяльта (1954).
Раньше был
открыт еще амфипитек:
фрагмент
нижней челюсти найден в эоценовых слоях в 60 км от
г. Могаунга (Бирма) в 1937 г. Эта форма тоже важна
для понимания
генезиса
обезьян1.
Проплиопитек с его заметно выдающимся клыком представляет
следующую ступень развития ископаемых антропоидов.
Он был несколько сходен
с гиббоном,
но уступал ему по размерам тела. Некоторые считают,
что проплиопитек обозначает
гиббоноидную стадию в эволюции человекообразных
обезьян и человека. Другие специалисты достаточно
единодушно признают проплиопитека
в качестве отдаленного общего
предка современных антропоидов и человека. Из
группы, представленной парапитеком и проплиопитеком, возникли
разные линии развития
ископаемых
антропоидов.
На линии, ведущей от проплиопитека к современным
гиббонам, находится плиопитек, челюсть которого
впервые была
найдена более 130 лет
назад (1837 г.) во Франции.
Это была вообще самая ранняя находка ископаемого
антропоида, и честь его открытия принадлежит
французскому палеонтологу
Эдуарду Ларте. Как
и на челюстях
других антропоидов, подбородочный отдел у плиопитека
отступает далеко назад. Клыки выдаются еще сильнее,
чем у проплиопитека.
В общем челюсть
обладает
столь заметным сходством с челюстями гиббонов,
что, несомненно, плиопитек, известный по челюстям
и зубам
из Франции,
Германии, Швейцарии и Монголии,
составляет одно из существенных звеньев родословной
этих малых антропоидов.
Проплиопитеки и плиопитеки населяли тропические
леса, покры-
1 В районе Файюма в 1963
г. продолжались палеонтологические раскопки, в процессе которых были сделаны
новые находки ископаемых человекообразных обезьян олигоценовой эпохи. Упомянем
здесь о наиболее важных, описанных Э. Саймонсом (Simons, 1965). В слоях кантраниевой
формации открыто 11 нижних челюстей и свыше 30 зубов примитивных узконосых обезьян.
К кругу крупных человекообразных Саймоне относит египтопитека (Aegyptopithecus
zeuxi), судя по фрагменту левой ветви нижней челюсти с премоляром и двумя молярами.
К более мелким формам тот же автор причисляет эолопитика (Aeolopithecus chirobates)
как гиббоноида: от него обнаружены правая и левые ветви челюстей с зубной формулой
2.1.2.3.
В последнее время среди палеонтологов появилась тенденция к более раннему отведению
родословных человека и антропоидов. Человеческую линию начинают вести чуть не
от проплиопитека через миоценового рамапитека, а крупных человекообразных — от
египтопитека через разные формы проконсулов, которых сближают с дриопитеками.
Для человеческой родословной все более выявляется значение находки рамапитека
из миоценовой эпохи — века расцвета человекообразных (Рейнолдс), так что более
вероятно все-таки, что разделение ветвей к человеку и к крупным антропоидам произошло
где-то около середины миоцена.
73
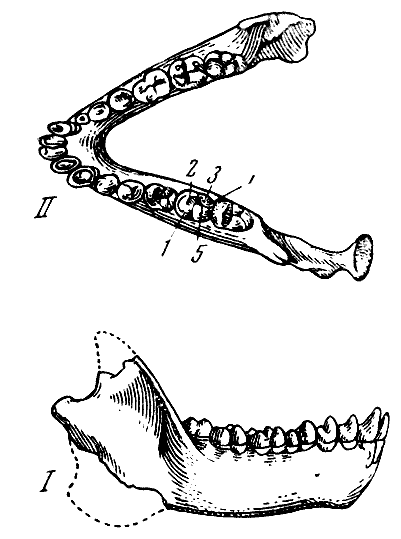
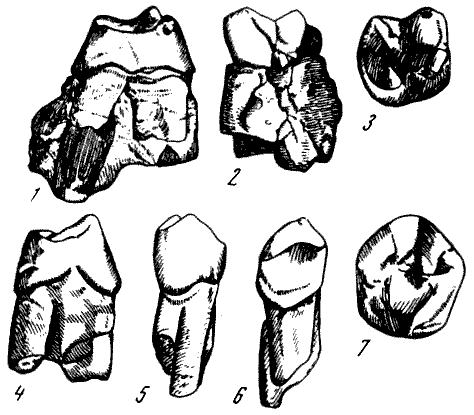
Рис. 24. Нижняя челюсть парапитека фраасова (Parapithecus fraasi
Schlosser) из нижнеолигоценовых слоев Файюма, Египет
I и II — вид справа
и сверху; 1 — протоконид; 2 — метаконид; 3 — энтоконид; 4 —
гипоконулид; 5 — гипоконид. По О. Шлоcсеру из О. Абеля 1931, 5/4
нат. вел. (ширина нижнего изображения 44 мм)
|
Рис. 25. Нижняя челюсть
проплиолитека геккелевского (Propliopithecus haeckeli Schlosser)
из нижнеолигоценовых
слоев Файюма, Египет
Вид справа,
частично реконструирована. Вверху зубы от клыка до последнего
моляра, вид сверху
По О. Шлоссеру из О. Абеля, 1931, 1/1, и зубы 4/3 нат. вел. (ширина нижнего изображения
58 мм)
|
вавшие в те времена (несколько десятков миллионов лет назад) огромные области
материков Европы, Азии и Африки. Изобилие пищи и сравнительно
небольшое количество хищников на деревьях благоприятствовали биологическому
процветанию высших
и низших обезьян (в то же время прогрессивное развитие
испытывали и обезьяны Южной Америки).
Можно предполагать, что прогресс высших обезьян выразился в следующем:
1) в освоении новых способов передвижения, или типов
локомоции, например путем
перелезания, перебрасывания или перелетания с помощью
рук по ветвям (брахиация) и путем хождения на двух конечностях
по более толстым
ветвям (круриация);
2) в развитии тонкости органов зрения и осязания
(преимущественно на ладонной поверхности рук); 3) в усиленном размножении;
4)
в развитии явлений стадности;
5) в интенсивном расселении по обширным областям
материков; 6)
в возникновении новых видов; 7) в появлении и усовершенствовании
новых черт приспособляемости
ко все изменяющимся условиям природы. У антропоидов,
вероятно, прогресс выразился, кроме того: 1) в усилении изменчивости под
влиянием новых условий среды;
2) в укрупнении размеров тела;
3) в развитии головного мозга;
4) в развитии высшей нервной деятельности, большей смышлености,
более сложного и разнообразного поведения; 5) в развитии
более сложной формы
материнского
инстинкта; 6) в появлении (у крупных антропоидов)
инстинкта строительства гнезд.
74
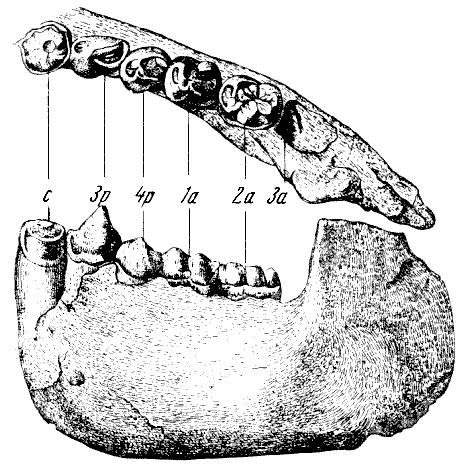
Рис. 26. Нижняя челюсть дриопитека фонтанова (Dryopithecus fontani
Lartet)
l — вид слева; 2 — вид сверху. По О. Абелю, 4931, 7/8 нат. вел. (ширина изображения
56 мм)
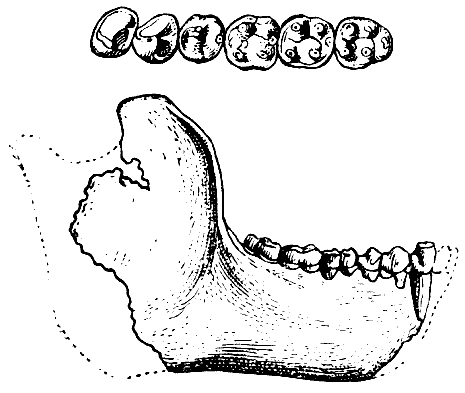
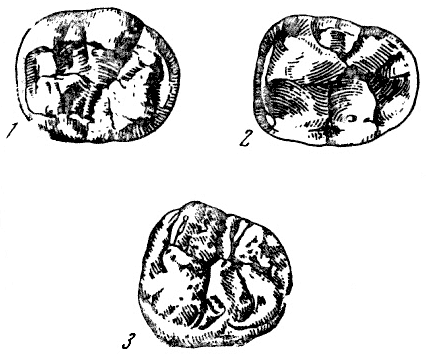
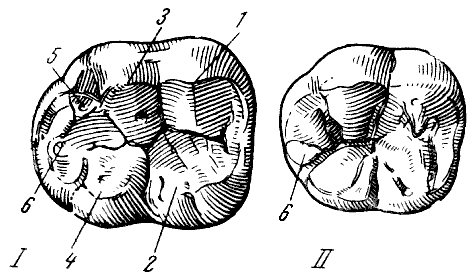
Рис. 27. Жевательная поверхность нижних моляров
I — с «узором дриопитека» на нижнем третьем большом коренном зубе чинджиева
дриопитека (Dryopithecus chinjiensis Pilgrim); II — с «плюс-узором» на соответствующем
зубе современного человека; 1 — протоконид; 2 — метаконид; 3 — гипоконид; 4 — энтоконид; 5 — гипоконулид (=мезоконид); 6 — шестой бугорок. По В. Грегори
из О. Абеля, 1931, 5/2 нат. вел. (ширина изображений 29 и 25 мм)
|
Об увеличении размеров тела в ходе эволюции человекообразных
обезьян свидетельствуют костные остатки антропоидов,
найденные в миоценовых
слоях древностью от 27
до 12 млн. лет. Здесь в первую очередь надо назвать
дриопитеков, известных по челюстям, зубам и фрагменту
плеча.
Впервые нижняя челюсть дриопитека (рис. 26) была
найдена в 1856 г. во Франции и описана Эдуардом Ларте.
Она
значительно крупнее,
массивнее
и выше, чем
у плиопитека; клыки представляют собой грозное оружие,
как
и у современных антропоидов; на жевательной поверхности
нижних коренных
зубов находится
тот самый узор, который сохранился до сих пор у потомков
дриопитеков, т. е. в
первую очередь у современных антропоидов (рис. 27).
Мы упоминаем об этом «узоре» потому, что он иногда
встречается не только на зубах ископаемых людей,
но и у современного
человека.
Повторим еще раз, что для человека в результате особого
пути эволюции, вследствие укорочения зубного ряда
и самих моляров,
стал характерным
другой рисунок
расположения бугорков на нижних молярах в виде «плюс-узора»:
все четыре главных бугорка занимают примерно равные
участки, будучи
крест-накрест
разделены
друг от друга поперечной и продольной бороздками.
В то же время у антропоидов передний внутренний бугорок,
располагающийся
с язычной стороны (метаконид),
крупнее соседнего заднего бугорка (энтоконида), в
связи
с чем
разделяющая их поперечная бороздка не приходится
точно напротив той, которая
разграничи-
75
вает два главных наружных бугорка (передний — протоконид
и задний — гипоконид), находящихся со щечной стороны.
Подобные довольно
сложные
детали (Korenhof,
1960) оказываются чрезвычайно важными для понимания
эволюции приматов и развития самого высшего из них
— человека1.
С какими же из современных антропоидов связан человек
наиболее тесными родственными узами по своему происхождению?
Идя
по способу исключения,
мы можем с уверенностью
сказать, что это не были гиббоны. Тем более это не
были орангутаны, которые отличаются многими чертами
чрезвычайной
специализации.
Оба эти антропоида
крайне приспособлены к жизни на деревьях, у них чрезвычайного
развития достиг особый тип передвижения по деревьям
с помощью одних рук
(быстрая либо медленная
брахиация).
Гиббоны — быстрые брахиаторы: они могут часами резвиться
меж ветвей, перелетая мгновенно с одной на другую,
и являют зрелище
необычайной
ловкости, соединенной
с удивительным изяществом движений.
Что же касается орангутана, то вследствие тяжелого
веса тела и хрупкости длинных костей конечностей
этот антропоид
обладает свойствами медленного брахиатора, осторожно пробующего
ветку, на которую он хочет перебраться.. Орангутаны
не перепрыгивают,
а перелезают с
ветки на ветку.
Медлительный орангутан с выделяющей его среди антропоидов
— рыжей
лохматой шерстью, щечными наростами, грудинной
железой неизвестного назначения,
отсутствием ногтя на рудиментарном большом пальце
стопы и другими чертами специализации
представляет собой совершенно уклоняющуюся форму:
этот антропоид прошел отдельный путь развития,
ушедший в сторону от путей
развития гиббонов,
шимпанзе и
горилл и еще гораздо более отдалившийся от линии
формирования
человека.
Коренные зубы орангутана заметно отличаются от моляров
гориллы или шимпанзе тем, что бугорки у них очень
низкие, есть несколько
добавочных
маленьких
бугорков, а бороздки, или морщинки, покрывающие
жевательную поверхность этих зубов, очень мелкие. У шимпанзе
бороздок много и они средней
глубины; у гориллы
их мало и они очень глубокие. Не удивительно,
что приматологи сразу различают зубы современных крупных
антропоидов
и еще легче опознают
зубы гиббонов.
Можно распознать и различить между собой и зубы
ископаемых антропоидов, что представляет, однако, довольно трудную
задачу, так как у
них тоже велика
индивидуальная изменчивость, а родов и видов гораздо
больше, чем у ныне живущих высших обезьян.
Судя по степени сходства в строении зубов и челюстей,
предковыми формами для орангутанов
1 Для
понимания процесса эволюции зубной системы человека важно учитывать процессы
редукции ее разных отделов в ходе антропогенеза (Зубов, 1968). У ближайших
предков человека, т. е. у прегоминид, ясно обозначалась редукция передного
отдела, в особенности клыка, исчезали и диастемы вверху и внизу, зубы становились
в более плотный ряд. А у гоминид изменения касаются уменьшения премоляров и
моляров, с переходом ключевого значения к первым молярам и с редукцией третьих
моляров.
76
могут служить следующие два антропоида: палеосимия
и сивапитек. Верхний правый третий коренной зуб
палеосимии был найден
в 1915 г. в богатом
скоплении костей и зубов ископаемых обезьян в
миоценовых слоях Сиваликских холмов в
Индостане, на склоне Гималайского хребта. Поверхность
зуба покрыта характерными мелкими морщинками,
почему эту обезьяну
и назвали
морщинистозубой палеосимией.
Начиная с 1879 г. в тех же слоях обнаружены также
челюсти разнообразных сивапитеков. Но лишь сравнительно
недавно
Мило Хелльману
(1938) сделал реконструкцию
зубных дуг сивапитека. На основании того, что
зубы ископаемых орангутанов неоднократно находили на
материке Азии —
в Южном Китае, где они
жили еще в четвертичном периоде, можно предполагать,
что орангутаны родом
из юго-восточной
Азии.
По сравнению с гиббонами и орангутанами гораздо
большую генеалогическую близость к человеку обнаруживают
шимпанзе и гориллы, развившиеся
из группы дриопитеков
середины третичного периода. Из нее же возникли
и
более поздние предки человека.
Без сомнения, дриопитеки были анцестральной группой
для современных крупных антропоидов, пишут Дж.
Харрисон, Дж. Уайнер (1968),
допуская наличие
ясных указаний на дифференциацию ее в направлениях
к
понгидам и к гоминидам.
Однако надо признаться, до сих пор еще, к сожалению,
невозможно судить точно о строении тела дриопитеков.
От них, правда,
известно свыше десятка
фрагментов
нижних челюстей вместе с несколькими десятками
нижних зубов, но (ерхние зубы известны лишь в
единичных экземплярах. Кроме
того,
верхнечелюстных
костей дриопитеков еще не найдено, как и черепов,
а из остального скелета известна
только плечевая кость, да и то в виде фрагмента.
Все-таки ученым удалось выяснить как по узору
дриопитека, так и по другим признакам, что германский
дриопитек
(Германия) мог
быть предком
для
шимпанзе, фонтанов дриопитек (Франция) или пенджабский
(Индия) — для гориллы, дарвиновский
же дриопитек (Австрия) относится к предкам человека
(рис. 28). Но, оказывается, есть еще и другие
ископаемые антропоиды,
более
или менее
подходящие для
того, чтобы их считать предками шимпанзе, гориллы
и человека.
Так, около 1933 г., в восточной Африке был найден
обломок верхней челюсти с клыком и всеми коренными
зубами от
антропоида, жившего
в начале миоцена,
около 25 млн лет назад. Обнаружен был и фрагмент
нижней челюсти. По кличке одного шимпанзе «Консул»
эту обезьяну
назвали проконсулом.
Новейшие находки костных остатков проконсула на
острове Русинга, оз. Виктория, в том числе черепа,
показывают
промежуточное
положение этой
обезьяны между
примитивными мартышкообразными и более поздними
человекообразными. На черепе отмечаются свойственные
исходной для низших
и высших узконосых обезьян, или
«протокатарриновой», группе признаки: например,
симфиз нижней челюсти короткий,
77
Рис. 28. Нижние моляры дриопитеков
1 — германского
(Dryopithecus germanicus Abel) из нижнеплиоценовых слоев Бонерца
в Трехтельфингене, Швабия, левый
второй моляр; 2 — пенджабского (Dr. punjabicus Pilgrim) из
миоценовых слоев Сиваликских холмов в Симле на южном клоне Гималаев,
правый
второй моляр; 3 — дарвиновского (Dr. darwini Abel) из среднемиоценовых
виндободиевых
слоев из Нейдорфа у Марха в Венском бассейне, левый последний
моляр. По О. Абелю, 1931, 2/1, 3/2 и 3/2 нат. вел. (ширина
изображений 21, 21 и 20 мм)
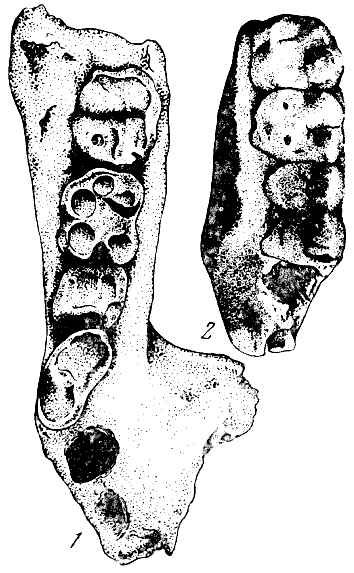
Рис. 29. Фрагменты челюсти рамапитека короткомордого
(Ramaplthecus brevirostris Lewis)
1 — правая половина нижней челюсти; 2 —
правая половина верхней челюсти. По Г. Льюису, 1934. Нат.
вел. (высота изображения 69 мм)
|
нижние резцы малы и стоят вертикально, отдел
межчелюстных костей узок, носовое отверстие имеет
грушевидную
форму, надглазничный
валик отсутствует;
на слепке
мозговой полости видно, что центральная (роландова)
борозда сдвинута вперед; уровень, церебрализации
невысок, т.
е. размеры головного
мозга сравнительно
невелики; задний внутренний бугорок верхних моляров
происходит не из ободка эмали, а из протоконуса.
В качестве предков человека некоторые указывают
на рамапитека. Два фрагмента верхней и нижней
челюсти от двух разных
особей рамапитеков (рис. 29)
были извлечены в Сиваликских холмах в 1934 и
1935 гг. из слоев нижнеплиоценового
возраста. Важно отметить, что в верхней челюсти
между
ячейкой клыка и вторым
резцом (I2) отсутствует диастема для вхождения
нижнего клыка при смыкании челюстей, а это свидетельствует
скорее в пользу
включения
рамапитека
в человеческую родословную (Simons, 1968), чем
против
его признания предком человека.
В жизни ископаемых антропоидов и множества других
животных в миоценовую и плиоценовую эпохи произошли
очень большие
изменения вследствие
великих процессов,
приведших к сильному преобразованию материков.
В течение этих миллионов лет в Старом Свете воздвигались
колоссальные
горные
цепи; климат
во многих местах
становился заметно суше, континентальнее; глухие
дебри сплошных тропических лесов, джунглей начали
просветляться,
редеть, а
затем
и исчезать.
78
Как многие другие тропические животные, обезьяны,
привыкшие к жизни в лесу, на деревьях, и не успевшие
приспособиться
к новым
условиям
на вновь
возникавших
огромных открытых пространствах, в большинстве
вымерли, часть отступила на юг и только сравнительно
немногие,
вроде павианов
или человеческих
предков
— антропоидов, перешли на исключительно
наземный образ жизни. При этом павианы укрупнились,
стали
жить большими
стадами и
приобрели на
пути
особой бестиализации очень сильные клыки и резкую
злобность. Двуногие же антропоиды
были менее устойчивы и более медленны при локомоции
в условиях открытой местности, лишены густого
волосяного покрова,
жили небольшими группами,
слабо размножались
и поздно созревали. Если даже они, будучи довольно
беспомощны,
и обращались к помощи природных предметов вроде
камней для добывания пищи или защиты
от врагов, то все же в конце концов все вымерли
(пав жертвой прежде всего кошачьих
хищников), кроме более умного и гибко приспособленного
ближайшего предка людей (Нестурх, 1964).
В Старом Свете остались, по сравнению с Южной
Америкой, не столь крупные массивы лесных тропических
зарослей,
уцелевшие со времени
геологического
переворота и последовавшего сильного преобразования
поверхности материков Афревразии. Поэтому низшие
и высшие узконосые
обезьяны
в большинстве
стали нередкими посетителями земли в поисках
добавочной пищи, чего совершенно нельзя
сказать о капуцинах,
игрунках и прочих обезьянах, оставшихся чисто
древесными животными в колоссальном лесном массиве,
обнимающем
бассейн реки Амазонки
и соседние области, и в
лесах центральной Америки.
О громадных переменах в жизни обезьян того времени
наглядно свидетельствует открытие остатков, ископаемого
антропоида
в юго-восточной Грузии,
сделанное советскими учеными. В Кахетии, в местности
Удабно, близ монастыря Гареджи,
в 1939 г. производились, палеонтологические раскопки,
организованные Бакинским университетом и Государственным
музеем Грузии.
В конце раскопок участница
эспедиции Е. Г. Габашвили нашла очень близко
от поверхности земли два зуба ископаемого антропоида.
Хотя они лежали
неглубоко, но
геологическая
древность
слоя оказалась значительной: конец миоцена или
начало плиоцена. Следовательно, обезьяны жили
на Кавказе
около 12 млн. лет назад.
Е. Г. Габашвили вместе с другим участником экспедиции,
палеонтологом Н. О. Бурчак-Абрамовичем, тщательно
изучили и описали эту редкую
находку (Бурчак-Абрамович,
Габашвили, 1945). Научное значение этой находки
очень велико, так как до 1939 г. остатков ископаемых
антропоидов
на территории
Советского
Союза обнаружено
не было. Только очень ограниченное количество
зубов и черепов низших обезьян было найдено в
первую
половину XX в. в Бессарабии
и ближайших
районах юго-западной
области Европейской части Советского Союза.
79
Рис. 30. Правые верхние зубы удабнопитека гареджинского (Udabnopithecus
garedziensis Burchiak-Abranovich et Gabashvili)
1—3 — первый моляр и 4—7 —второй премоляр; 1, 4 —
оральная сторона; 2, 5 — латеральная; 6 —
медиальная; 3, 7 —
жевательная поверхность. По Н. О. Бурчаку-Абрамовичу и
Е. Г. Габашвили, 1945, 7/6 (1—6) и 7/4 (7) нат. вел. (общая ширина рисунка 58 мм)
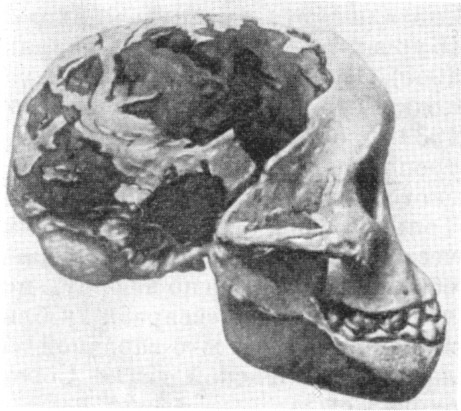
Рис. 81. Череп австралопитека африканского
(Australopithecus africanus Dart). Вид справа.
Нижний край нижней челюсти дополнил
В. Абель
По О. Абелю, 3/5 нат. вел. (ширина изображения 60 мм)
|
В Грузии были найдены верхний второй премоляр
и соседний первый моляр (рис. 30). В момент открытия
они были
соединены костной
тканью, следовательно,
принадлежат одной особи. Судя по тому, что на
моляре
передний внутренний бугорок (протоконус) соединен
с задним наружным
(метаконусом) с помощью гребешка, это несомненно
зуб антропоида, не отличающегося
очень крупными
размерами тела, так как размеры зуба невелики,
как у шимпанзе. Премоляр замечателен,
в частности, присутствием трех корней: у обезьян
эта особенность встречается редко, так как обычно
у них
на верхних премолярах
два
корня развиваются
не часто; на нижних премолярах наличие двух корней
составляет исключение.
Найденную ископаемую человекообразную обезьяну
авторы находки назвали гареджинским удабнопитеком.
Этот
вид вымер в те
времена, когда
вздымался Кавказский хребет.
До того в тропических лесах на территории Кавказа,
возможно, жили и другие высшие и низшие обезьяны.
Костные остатки,
черепа и зубы
ископаемых
обезьян
и других приматов, наверное, сохранились в третичных
напластованиях разных областей территории Советского
Союза.
Пока еще трудно определить истинное положение
удабнопитека среди ископаемых антропоидов и его
отношение к
современным (Гремяцкий,
1957), а также
к человеческой родословной. В этом смысле вопрос
представляется несколько менее трудным
по отношению к серии находок южноафрикан-
80
ских антропоидов из верхнеплиоценовых и нижнеплейстоценовых
слоев. Речь идет об австралопитеках и близких
к ним формах, обнаруживающих
ближайшее
морфологическое
сходство с шимпанзе, гориллой и человеком. Многие
авторы высказываются в пользу более тесных филогенетических
отношений между австралопитеками
и человеком.
Первая находка южноафриканских антропоидов была
сделана в 1924 г. в восточной части пустыни Калахари,
в Бечуаналенде.
Здесь,
близ железнодорожной
станции
Таунг, к северу от г. Кимберлея, в известняковых
разработках были обнаружены два черепа взрослых
павианов и один
череп детеныша человекообразной
обезьяны. Первоначально предполагали,
что геологическая древность находок не очень
велика, что они относятся к первой половине четвертичного
периода, возможно,
даже ближе к
его середине, с связи с чем хронологическая давность
их
исчисляется в 500—800
тыс.
лет.
Позже некоторые ученые считали эту находку более
древней, относя ее, например, к концу или даже
середине плиоцена.
Найденный неполный череп южноафриканского антропоида
принадлежал молодой особи в возрасте около пяти
лет (рис. 31—34). Об
этом свидетельствует полный набор молочных зубов,
позади которых
видны уже прорезавшиеся
первые постоянные
большие коренные зубы. На черепе хорошо сохранились
лицевой отдел и
лобная кость. Части черепа состав-
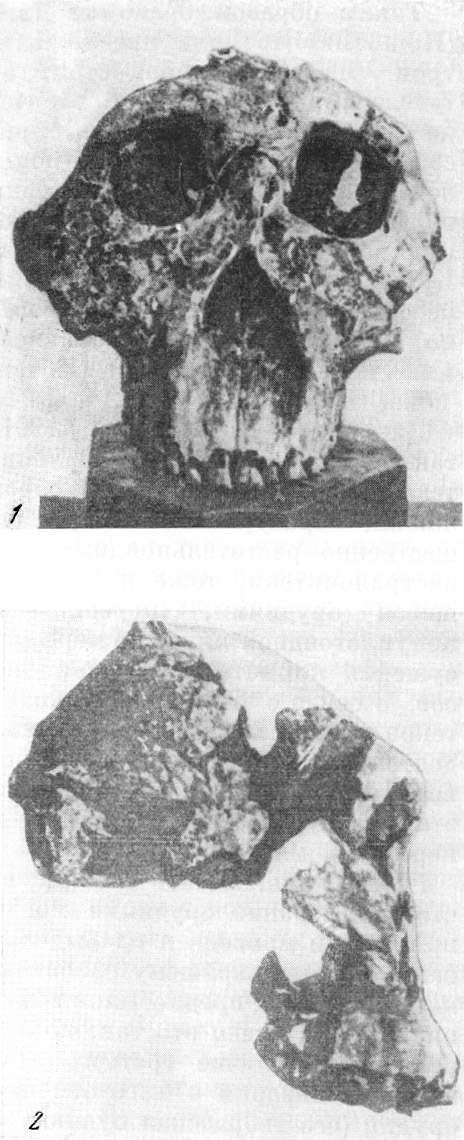
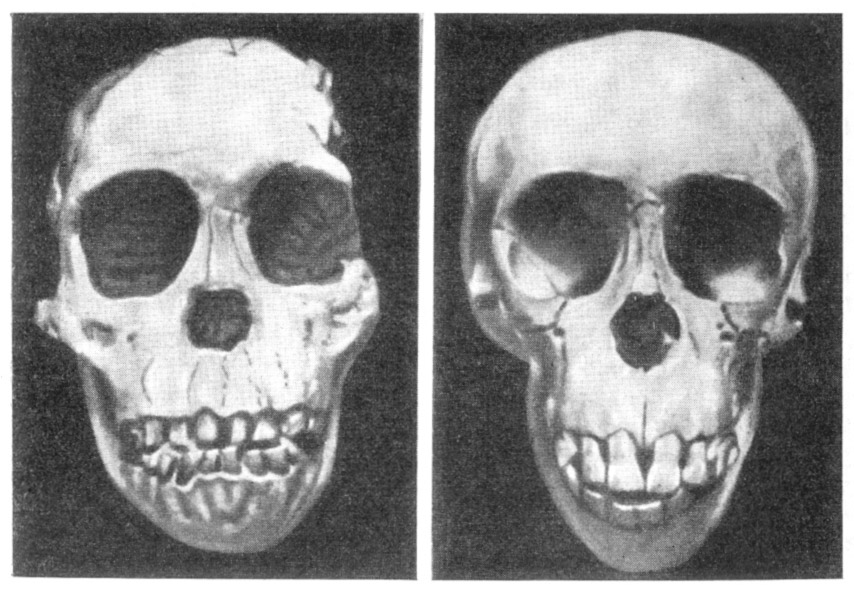
Рис. 32. Черепа африканского австралопитека
(по гипсовому слепку) и шимпанзе обыкновенного
(справа),
оба одного
возраста
По О. Абелю, 1931, 1/2 нат. вел. (ширина изображения 106 мм)
81
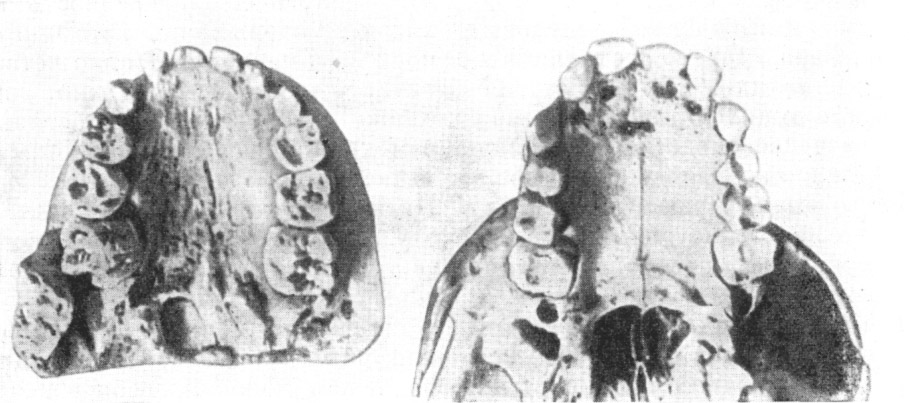
Рис. 33. Верхний зубной ряд и нёбо австралопитека
африканского (слева) и шимпанзе обыкновенного,
оба одного возраста
По О. Абелю, 1931, 3/4 нат. вел. (ширина изображения 115 мм)
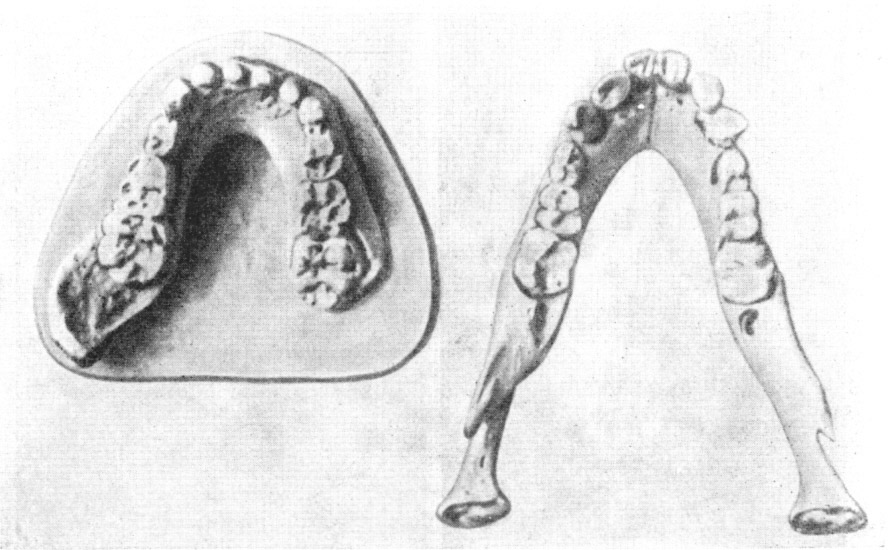
Рис. 34. Нижние челюсти африканского австралопитека
(слева) и шимпанзе обыкновенного,
оба одного возраста
По О. Абелю,
1931,
5/7 нат. вел. (ширина изображения 112 мм)
ляют неразрывное целое с минеральной массой,
заполняющей полость черепной коробки.
Череп был доставлен южноафриканскому биологу
Раймонду Дарту. Он изучил череп и опубликовал
его краткое
описание, в котором
предложил
назвать
найденную
обезьяну африканским австралопитеком (т. е.
южной обезьяной).
Находка «таунгской обезьяны» возбудила массу
споров. Одни ученые, например Отенио Абель,
приписывали череп детенышу
ископаемой
гориллы. Другие,
как Ганс Вейнерт, видели в нем гораздо больше
сходства
с черепом шимпанзе и свое
мнение основывали, в частности, на вогнутости
профиля лицевого отдела, а также на форме носовых
косточек
и глазниц.
Третья группа ученых, к числу которых относятся
Дарт, а также Вильям Грегори и Мило Хелльман,
полагала,
что у
австралопитека
больше
сходства с дриопитеком
и человеком. Расположение бугорков на нижних
молярах является не очень сильно измененным
узором зубов
дриопитека.
Надглазничный валик на черепе развит слабо,
клыки почти не выдаются из зубного ряда, лицо
в целом,
по мнению
Грегори, поразительно
предчеловеческое.
Четвертые, как Вольфганг Абель, обращали внимание
на черты специализации, уводящие австралопитека
в сторону
от человеческой
родословной.
Так, первые постоянные моляры австралопитека,
в отличие от человеческих, шире в задней
своей половине.
Перейдем к вопросу о вместимости мозговой коробки
описанного Дартом австралопитека. В 1937
г. советский антрополог В. М. Шапкин, применив предложенный
им точный метод,
получил цифру
420
см3, которая
недалека от определенной В. Абелем: 390 см3.
Раймонд Дарт определил вместимость мозговой
коробки в 520 см3, но эта цифра,
несомненно, преувеличена. Принимая во внимание
молодой
возраст найденного
экземпляра, можно было
предполагать, что вместимость
мозговой коробки взрослых особей австралопитеков
равна 500— 600 см3.
Представления о типе австралопитека заметно
обогатились, когда летом 1936 г. в Трансваале
был обнаружен
череп ископаемого антропоида. Он
был найден
в пещере у с. Штеркфонтейн, близ г. Крюгерсдорпа,
в 58 км к
юго-западу от г. Претории. Принадлежит этот
череп взрослой особи и очень
похож на череп
шимпанзе, но зубы сходны с человеческими.
Череп имеет удлиненную форму: длина мозговой
коробки
равна 145
мм, ширина 96 мм,
следовательно, черепной
указатель
низкий. Он составляет 96 X 100 : 145 = 66,2
(ультрадолихокрания).
Южноафриканский палеонтолог Роберт Брум, примерно
сорок лет работавший в Южной Африке в качестве
специалиста по млекопитающим
и их эволюции,
изучил
череп штеркфонтейнской ископаемой обезьяны
и отнес ее к роду австралопитеков, к виду
трансваальского австралопитека.
Однако
изучение позднее
найденного там же (в Штеркфонтейне) нижнего
последнего
моляра, оказавшегося весьма
крупным и сходным с человеческим, заставило
Брума ус-
83
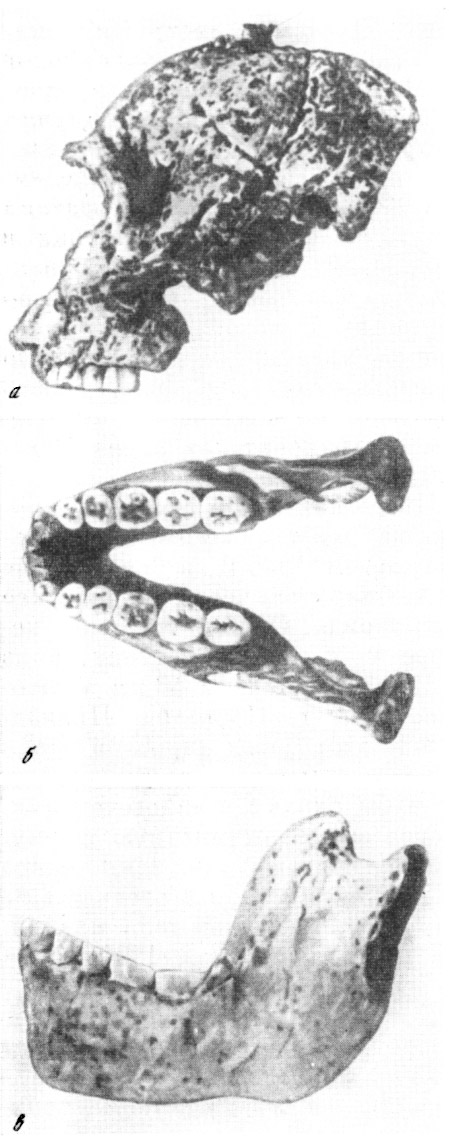
Рис. 35. Череп парантропа (а) крупнозубого (Paranthropus crassidens
Broom), находка 1938 г., и нижняя челюсть, находка 1948 г., вид слева
(б) и сверху (в)
По Р. Бруму из Г. Улльриха, 1953, 2/5 нат. вел. (ширина изображения 56
мм)
|
тановить новый род — плезиантропов, т. е.
обезьян, более близких к человеку. Поэтому
штеркфонтейнский
антропоид
получил и новое
видовое
название
— трансваальского плезиантропа.
Глубоко заинтересовавшись находками африканских
ископаемых антропоидов и проблемой антропогенеза,
Брум приложил
много энергии на дальнейшие
поиски
их остатков. С 1936 по 1947 г. было обнаружено
свыше 10 неполных черепов и 150 изолированных
зубов, а также некоторые кости скелета плезиантропов.
В
1938 г.
Бруму удалось найти замечательный
череп ископаемого антропоида
(рис, 35). История этого открытия такова.
Один школьник из с. Кромдраай добыл череп
обезьяны
из породы на
склоне холма
близ своей деревни и,
разбив на
куски, взял несколько вывалившихся зубов
для игры. О найденных зубах случайно узнал
Брум,
который поспешил
на место находки
и с помощью
школьника, отдавшего
ему зубы обезьяны, нашел куски черепа. Геологическая
древность находки падает, по-видимому, на
середину четвертичного
периода.
Сложив части черепа, Брум был поражен чертами
его сходства с человеческим, как, например,
в форме
височной кости,
в строении области слухового
прохода, в расположении затылочного отверстия
ближе к середине основания черепа,
чем у современных антропоидов. Зубная дуга
широка, клык невелик, зубы заметно
похожи на человеческие.
В итоге исследования Брум назвал кромдраайского
антропоида парантропом, т. е. обезьяной,
сто-
84
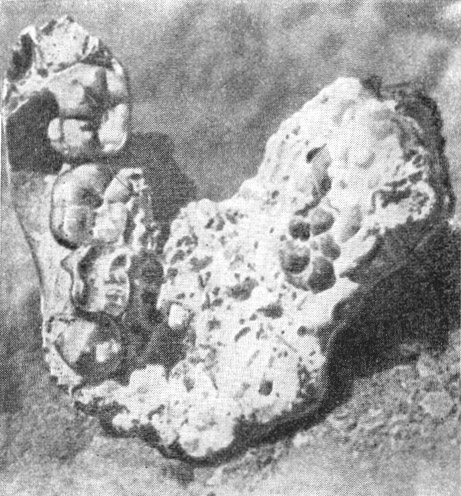
Рис. 36. Нижняя челюсть австралопитека прометея (Australopithecus prometheus
Bart) в процессе расчистки брекчии
По Р. А. Дарту, 1948, 5/8 нат. вел. (ширина изображения 59 мм)
|
ящей рядом с человеком. В 1939 г. были найдены
также некоторые кости скелета парантропа,
который обнаружил
сильное сходство
с плезиантропом.
Обе обезьяны
обладают большой близостью к австралопитеку.
В 1948—1950 гг. Брумом были сделаны новые
находки южно-африканских антропоидов — парантропа
крупнозубого
и австралопитека
прометея (рис. 36). Отсюда
можно сделать вывод, что Африка должна быть
очень богата остатками других, до сих
пор еще не открытых обезьян (Якимов, 1950,
1951; Нестурх, 1937, 1938), тем более что
в 1947 г.
английский ученый
Л. Лики нашел,
как мы уже
упоминали, череп африканского проконсула
(обладающего чертами сходства с шимпанзе)
в
области Кавирондо (Якимов, 1964, 1965).
На основании приведенных фактов можно считать
очень вероятным, что в первой половине четвертичного
периода и ранее,
в верхнем отделе третичного
периода,
в Африке уже сформировались несколько разных
видов
крупных высокоразвитых человекообразных обезьян
(Зубов, 1964).
Объем их мозговой коробки
равен 500 — 600 см3 и даже несколько больше
(при весе 40—50 кг), а челюсти
и
зубы, обладая
типично антропоидными чертами, в то же время
обнаруживают значительную близость и к человеческим
зубам. Австралопитеков
многие рассматривают
как «моделей»
предков человека.
Геологическая древность части подобных австралопитековых
уходит в нижний плейстоцен, который ныне
хронологически датируется
глубиной до 2 млн.
лет, вмещая виллафранкские слои (Иванова,
1965).
Некоторые из ископаемых африканских антропоидов
передвигались на двух ногах, о чем свидетельствуют
форма и строение
различных найденных
костей,
например
от таза австралопитека прометея (1948) или
плезиантропа (1947). Возможно, что они также
применяли находимые
в природе палки
и камни в качестве
орудий. Обитавшие в довольно сухих, степных
или же полупустынных областях (рис. 37),
австралопитеки употребляли также и животную
пищу. Они занимались охотой на зайцев, павианов.
Южноафриканский ученый Р. Дарт приписывает
ископаемым антропоидам, вроде австралопитеков,
умение пользоваться
огнем и речью.
Но фактов в пользу
такого
85

Рис. 37. Австралопитек. Реконструкция В. Е. Ле Грос Кларка, 1952
По Г. Улльриху, 1953, 2/5 нат. вел. (ширина изображения 59 мм)
|
предположения не имеется (Коenigswald, 1959).
Попытки представлять антропоидов Южной Африки
в виде настоящих
гоминид лишены оснований.
Недостаточны
также и доказательства, что эти обезьяны
были предками всего человечества или какой-нибудь
его части. То же относится и к найденному
в
Италии ореопитеку, остатки которого были
обнаружены в Тоскане у горы Бамболи.
Известны его зубы,
челюсти, фрагменты
костей предплечья, найденные в слоях среднемиоценового
и раннеплиоценового возраста. Судя по костным
остаткам, бамболиев ореопитек значительно
приближается к антропоидам (Hurzeler, 1954).
В 1958 г. в Тоскане близ селения
Бакчинелло в слоях лигнита, датируемых верхним
миоценом, на глубине около 200 м был
обнаружен почти целый
скелет ореопитека.
Это
безусловно одно
из крупнейших открытий в области палеонтологии
человека.
Скорее ореопитека надо трактовать как «неудачные
попытки» природы: эти обезьяны вымерли. Человеку
дала начало,
вероятно, одна
из южноазиатских форм антропоидов,
развивавшихся из раннеплиоценовых человекообразных
обезьян типа рамапитека и, вероятно, сходных
с австралопитековыми.
Большой интерес, конечно, представляют открытия
1959, 1960 и позднейших годов в Олдовайском
ущелье, Танзания,
сделанные
Луисом
Лики (Louis
Leakey) и его
женой Мэри: это были костные остатки человекообразных
обезьян — зинджантропа (рис. 38) и презинджантропа
(Реглетов, 1962,
1964, 1966). По радиокарбоновому
методу их древность была исчислена примерно
в 1 млн. 750 тыс. лет.
Первоначально Лики приписал череп зинджантропа
(Zinjanthropus) с его хорошо выраженными
сагиттальным и затылочным валиками предку
человека, но позже сам отказался от такого
мнения (Нестурх,
Пожарицкая, 1965):
сходство здесь больше
с парантропом, чем с австралопитеком.
Более близкой к человеку, по-видимому, оказалась
находка презинджантропа, сделанная Лики:
судя по скелету левой
стопы взрослого с довольно
выраженным продольным сводом, это существо
обладало двуногой походкой; а судя
по теменным костям молодой особи
86
Рис. 38. Череп зинджантропа бойсовского (Zinjanthropus boisei)
По Л. Лики, 1959. 1 — вид спереди, 2 — справа, ок. 1/4 нат. вел. (ширина
изображений 59 мм)
|
объем полости мозговой коробки составлял
бы свыше 650 см3. Поэтому
презинджантропа назвали
«человек
умелый»
— Homo
habilis (Leakey,
Tobias, Napier, 1964).
Ему же были приписаны находившиеся поблизости
несколько небольших камней со следами
огранения (Якимов, 1965), которые, однако,
могли получиться и случайно при попытках
убить какое-нибудь
мелкое животное
на твердой
почве.
Последние годы ознаменовались новыми находками
ископаемых антропоидов. Например, К. Арамбур
и И. Коппенс (Arambourg,
Coppens) найденную
в долине Омо, западная
Эфиопия, нижнюю челюсть приписали форме,
более примитивной, чем австралопитеки, и
назвали
ее «Паравстралопитек эфиопский» (Paraustralopithecus
aethiopicus).
Исследователи считают этого антропоида из
нижнего виллафранка более примитивным, чем
австралопитеки,
которые, правда,
встречаются и
в нижнеплейстоценовых слоях.
Плейстоцен углубился по международному соглашению
геологов за счет прибавления к нему виллафранкской
эпохи верхнего
плиоцена и составляет
примерно 2
млн. лет. Количество находок австралопитеков
увеличивается (в
Гаруси и Пелинджи
на озере Нейтрон в Танзании; у озера Чад;
в Канапой, Кения и других местах). Очень
удачна
богатая
находка остатков
от двенадцати
особей
австралопитеков,
сделанная Ч. Брэйном (1968 г.) в брекчиях
Сварткранса из старых раскопок 1930—1935
гг.; в том числе
оказалось возможным
получить
полный слепок
эндокрана одного из них.
87
Рис. 39. Скелет левой
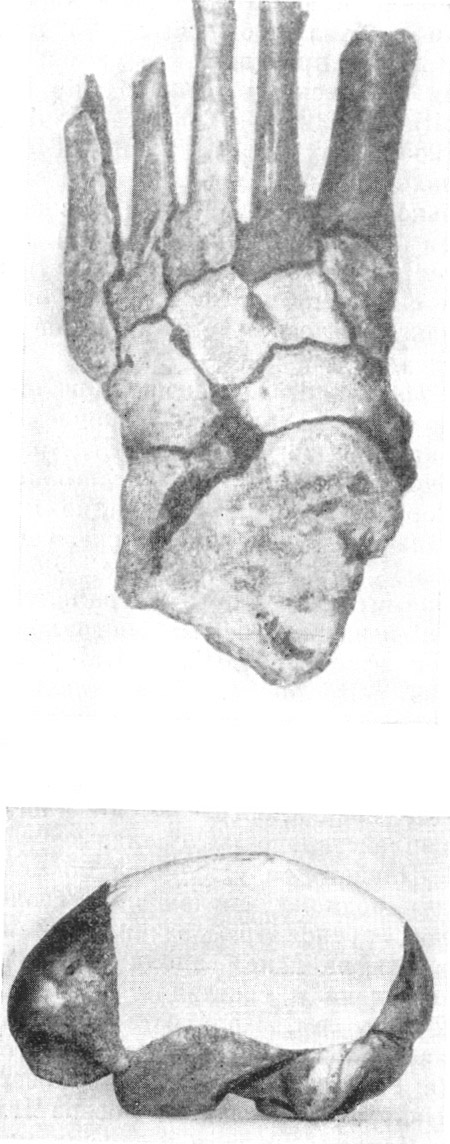
стопы Гомо габилис (по Э. Влчеку, 1965) и реконструкция его мозга,
вид слева
(по В. И. Кочетковой, 1969)
|
Таким образом, Гомо габилис (Homo habilis),
или презинджантроп (рис. 39), теперь стоит
не так
изолированно, как
это многим казалось раньше,
и можно
присоединиться к тем палеоантропологам, которые
считают его за один из географических вариантов
популяций
вида австралопитека.
Кроме того,
его
головной мозг был
не так велик, не 680 см3, а 657,
по мнению самого Ф. Тобайяса, или даже меньше
— 560
(Кочет-кова, 1969).
Дж. Робинсон (Robinson, 1961) так рисует
радиацию австралопитековых. Ведя двуногий
образ жизни,
парантропы были преимущественно
растительноядными, а австралопитеки, тоже
пользовавшиеся орудиями, перешли к полуплотоядной
пище по мере иссушения климата и поредения
лесов. В связи с этим у австралопитеков прогрессировала
орудийная
деятельность
и повышался
уровень интеллекта.
Значит,
первый этап — двуногость, а второй — переход
к
мясной пище.
Естественно, пишет Робинсон, что пользование
орудиями могло привести и привело к их выделыванию
и к дальнейшему
развитию потенциальных
предпосылок гоминизации.
В общем это так, но качественное отличие
третьего этапа гоминизации — изготовление
орудий (его
творческая сущность)
осталось для
Робинсона неподчеркнутым. Что
же касается парантропов, то они испытали
биологический регресс и вымерли.
Любопытны соображения Робинсона относительно
родословной гоминид, которую он рисует как
самостоятельную с большой геологической
древности. По
его мне-
88
нию, австралопитеки ведут свой род независимо
от раннемиоценовых понгид вроде проконсулов,
а может
быть, даже, учитывая
пример амфипитека, от
родословной линии, независимой со стадии
полуобезьян и медленно развивавшейся на большей
части своей истории.
Подобная идея о древности отделения человеческой
ветви не раз появлялась в истории науки.
Например, известный
австрийский
палеонтолог Отенио
Абель считал парапитека исходным представителем
человеческой ветви развития
с начала олигоцена. Чарлз Дарвин (1953, стр.
265) писал: «Мы далеки от того, чтобы
знать, как давно человек впервые отделился
от ствола узконосых; но это могло иметь место
в
такую отдаленную
эпоху, как
эоценовый период,
потому
что высшие
обезьяны были уже отделены от низших столь
рано, как в верхний миоценовый период, о
чем говорит
существование дриопитека».
Однако современная
палеонтология высших обезьян считает, что
отделение предчеловеческой ветви произошло,
скорее всего, в миоцене, а древнейшие люди
появились в
течение нижнего плейстоцена
(см. также: Бунак, 1966).
В течение третичного и в начале четвертичного
периодов, по теории В. П. Якимова об адаптивной
радиации
человекообразных высших
обезьян (1964),
одни из них
пошли по линии укрупнения размеров тела;
между тем у других в связи с развитием орудийной
деятельности и сложности
поведения
обозначился
более
прогрессивный
путь, на который и вступили австралопитеки
и предшественники древнейших гоминид (Урысон,
1969).
К числу форм, родственных австралопитекам,
относится еще одна находка черепа, но в центральной
части
Африки. Это
так называемый
чадантроп
(Tchadanthropus), открытый французским палеонтологом
Ивом Коппенсом (Coppens, 1965) в начале
1961 г. Речь идет о фрагменте черепа с лобной,
орбитальной, скуловой и верхнечелюстной
частями; лоб покатый, с сагиттальным утолщением;
надглазничный валик хорошо выраженный; скуловые
кости массивные;
глазницы крупные. Коппенс
склонен ставить
чадантропа ближе к питекантропам, но советский
антрополог М. И. Урысон (1966) на основании
своего анализа черепа
причисляет
его
к кругу прогрессивных
австралопитековых
начала плейстоцена.
Африканские находки человекообразных подверг
тщательному пересмотру В. Ле Гро Кларк (Le
Gros Clark, 1967).
Он считает, что плезиантроп,
зинджантроп,
презинджантроп и телантроп относятся к одному
роду австралопитеков подсемейства австралопитековых
семейства гоминид, другими
словами, что все это наиболее
примитивные гоминиды, но не относящиеся к
более высокоразвитым людям, образующим
род гомо. В роде австралопитеков Ле Гро Кларк
выделяет
лишь два вида — африканского и массивного.
По его мнению, стопа их
вряд
ли была хватательной,
хотя они
еще не очень хорошо передвигались на двух
ногах благодаря недостаточно развитому тазу.
Но в
кисти первый палец
был хорошо развит и
возможно, что австралопитеки
89
вo время охоты на животных применяли оружие
из кости, рога или зуба, поскольку природными
орудиями
своего
тела они
не располагали.
У австралопитеков
была
стадная организация и некоторый уровень первоначального
общения, звуковой коммуникации, в связи с
их довольно развитым интеллектом.
В новейшее время многие исследователи относят
к семейству гоминид (Hominidae) не только
людей собственно
начиная
с питекантропов,
но также австралопитеков
и близких к ним ископаемых человекообразных
крупных обезьян. Между тем современные и
ископаемые крупные
антропоиды
обычно относились
к семейству
понгид (Pongidae).
Теперь же существует тенденция объединять
оба эти семейства в надсемейство гоминоидов
(Hominoidea),
или человекоподобных
высших
приматов.
И нам представляется, что было бы правильнее
австралопитеков
и близкие к
ним формы помещать в семейство
понгид в качестве подсемейства австралопитецин
(Australopithecinae), или австралопитековых
(см. также: Зубов, 1964). Передвижение
на двух ногах и
манипулирование предметами из числа плейстоценовых
австралопитековых понгид перешло в искусственное
изготовление орудий лишь
у предкового вида для человека,
для гоминид.
Цепь находок древних человекообразных обезьян
продолжается и в Передней Азии. Так, в Израиле,
близ холма Убейдия
в Иорданской долине, в 1959
г. были открыты
два фрагмента массивной лобной кости неизвестного
крупного гоминоида.
Израильский
археолог М. Стекелис рассматривает найденные
тут же расколотые гальки и другие камни со
сколами как его
орудия, но,
скорее, это природные
осколки. Древность
крупного антропоида из Убейдии — нижнечетвертичная
эпоха. Другая, более крупная, можно сказать,
гигантская,
обезьяна
стала известной
по нижней
челюсти, обнаруженной
в 1955 г. близ г. Анкары, при раскопках на
горе Синап. Она отличалась некоторыми чертами,
сближавшими
ее
с древнейшими людьми, в частности
зачаточным выступом
на передней челюсти. Эта находка свидетельствует
о том, что
число крупных антропоидов в Азии было, вероятно,
не меньшим, чем в
Африке. Геологический
возраст анкаропитека — верхний миоцен.
Находки представителей авст-ралопитековой
группы южноафриканских антропоидов (рис.
40) заставили
многих ученых снова
задуматься над вопросом о географическом
ареале обитания предкового вида для человека,
о прародине человечества. Дарт провозгласил
южную Африку колыбелью
человечества, к мнению
Дарта примкнули
Брум, а также Артур Кизс.
Мысль об Африке как о вероятной родине человечества
не нова. Еще в 1871 г. Чарлз Дарвин указывал
на африканский материк
как на возможное
место
возникновения
первых людей из обезьян. Он ссылался, в частности,
на
то немаловажное обстоятельство, что горилла
и шимпанзе живут
именно здесь,
а они являются
ближайшими родственниками
человека. Известно, что живущие в пределах
достаточно обширной
90
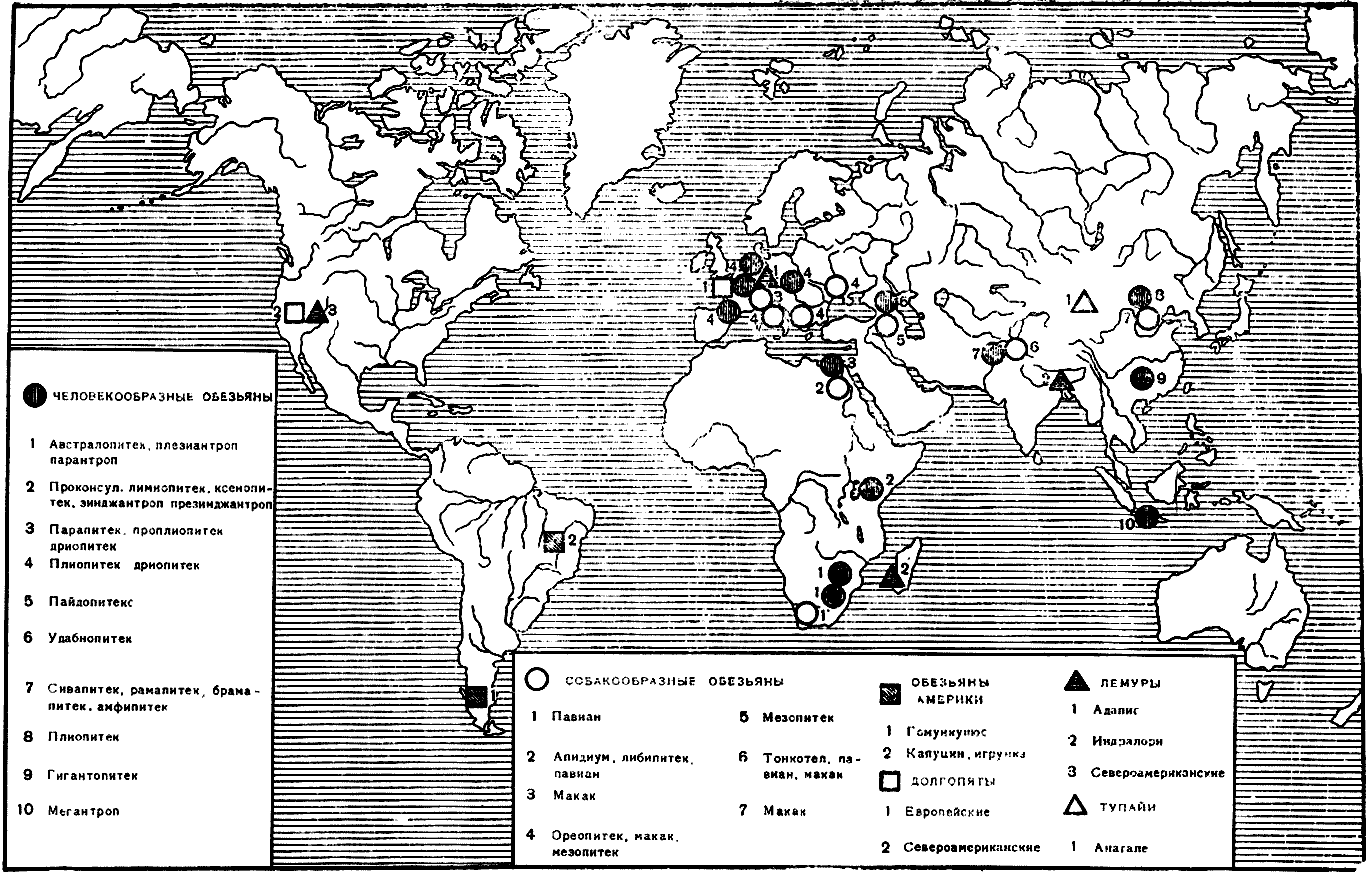
Рис. 40. Карта находок ископаемых приматов
области современные млекопитающие состоят
большей частью в отношениях филогенетического
родства
с вымершими формами той
же территории,
писал Дарвин. Напомним, однако,
что на северо-востоке Африки, в Египте,
были обнаружены остатки таких общих предков современных
антропоидов,
как парапитек
и проплиопитек. Поэтому более
вероятным являлось бы включение в прародину
человечества скорее северо-восточной
Африки, чем южной.
Совокупность других данных позволяет искать
прародину человечества не в Африке, а
в южной половине
Азии. С 1879 г. здесь,
на севере Индостана, в Сиваликских
холмах, найдено много остатков антропоидов
и низших узконосых обезьян; в Монголии
в 1924 был
найден
зуб плиопитека;
удабнопитек связывает
южноазиатских
ископаемых антропоидов с европейскими
плиопитеками и дриопитеками.
Очень длинная и довольно широкая зона распространения
многих уже известных в науке видов ископаемых
антропоидов тянулась
в миоцене
и плиоцене
от европейских берегов Атлантического
океана на восток-юго-восток вплоть до Малайского
архипелага (с островом Явой в его центре)1.
В пределах
обозначенной зоны наиболее вероятной
прародиной человечества
разные
ученые считают
ту или
иную значительную
область
центральной или южной Азии, к
которой, очень возможно, примыкала также
и часть территории в юго-восточной Азии.
Здесь
можно
упомянуть мнение
русского ученого
П. П. Сушкина
(1928), а также гипотезу Осборна. Может
быть, область прародины человечества
захватывала и часть северо-восточной
Африки (Нестурх, 1964).
В пользу предположения об азиатской прародине
человека свидетельствует также нахождение
остатков древнейших
людей в юго-восточной
Азии. Южноафриканское происхождение питекантропа
и синантропа представляется
нам недостаточно
обоснованным:
трудно и даже невозможно допустить мысль
о передвижении первых людей в течение короткого
срока на расстояние
почти 15 000
км из южной Африки
вдоль сильно
изрезанных берегов Индийского океана и
по местам
суши до самой Явы или сквозь южную Азию
на восток до места
обитания
синантропов.
Приводились еще гипотезы о происхождении
человека на севере Азии, где в третичном
периоде росли
тропические леса. По
гипотезе Л.
Вильзера
(Wilser, 1905), предки
человека под влиянием наступивших холодов
продвигались по материку Азии все более
и более на юг, где
в одной из областей и произошло
преобразование обезьяны
в древнейшего человека. Однако полное отсутствие
доказательств
лишает подобную гипотезу всякой убедительности.
Маловероятно также искать прародину человека
в Европе, представляющей собой по существу
полуостров материка
Афревразии. Совершенно
невероятно ис-
1 В недавнее время идею южноазиатской прародины
групп высших приматов и человека обосновал
многими геологическими
и
палеоприматологическими фактами Ю.
Г. Решетов (1966).
92
кать прародину всего человечества,
как это делал Отто Шётензак, в Австралии
с
ее архаическим
миром
сумчатых
млекопитающих,
или, как Флорентино
Амегино, в Америке, куда человек
проник впервые из Азии через Берингов перешеек
25—30 тыс. лет назад.
Совершенно очевидно, что ход эволюции
обезьян был тесно связан с огромными
тектоническими
процессами преобразования
поверхности
Земли в миоцене
и плиоцене. Тогда формировались
мощные горные складки,
климат изменялся с тропического
влажного на подтропический, умеренный,
а кое-где и на холодный, притом
во множестве областей
на более сухой,
с сильным
поредением и даже исчезновением
лесных массивов. Часть фауны должна
была погибнуть, другая приспособиться
на месте,
третья мигрировать.
Яркие следы
подобных миграций верхнетретичных
неогеновых млекопитающих можно
усмотреть в миграции многих азиатских животных,
например ископаемых лошадей, слонов,
страусов, обезьян, в Европу и Африку.
По богатым
залежам
костных остатков
этих животных
в местности Пикерми
(Греция), послужившей промежуточным
пунктом на пути их переселения,
палеонтологи называют
совокупность мигрировавших азиатских
животных того времени фауной Пикерми.
В современной
фауне
Африки
существует
значительная примесь
из потомков
пикермийской фауны1.
Одни обезьяны, как пентеликов мезопитек,
мигрировали дальше, другие, как
гареджинский удабнопитек,
вымерли по пути,
третьи преобразовались
на месте
в плейстоценовые формы азиатских
обезьян, дав начало их современным
формам, а
также человеку
Эти обезьяны
в большинстве
были вынуждены
перейти на
смешанный древесно-наземный образ
жизни, некоторые же на чисто наземный,
как
многие павианы. Предки человека
должны были приспособиться
к жизни на земле, но в отличие
от павианов перешли к передвижению
на двух
ногах. Условия
и возможности
возникновения человека кроются
прежде всего в особенностях строения и экологии
верхнетретичных
ископаемых
антропоидов (Гремяцкий,
1961, 1962).
Человек,
будучи узконосым обезьяноподобным
приматом, не
произошел из американских обезьян.
Последние во многом представляют
собой
противоположность
обезьянам Афревразии. Так, например,
положение вниз головой для очень
многих из них наиболее высокоразвитых
цепкохвостых
форм является довольно обычным,
нормальным.
Ожидать у них развития двуногости
одновременно с редукцией наружного
хвоста, предполагать
возможность
1 Сторонники африканской
прародины человека привлекают
различные факты
в пользу
своей гипотезы. В
новейшее время Т. Кук
(Cooke, 1968), рассматривая ископаемую
фауну млекопитающих, пишет, что
предки предполагаемых
иммигрантов из
Азии, в том числе приматов,
обнаружены
в более
ранних напластованиях континента
Африки и что сам человек, вероятно,
может
представлять собою
один
из существенных
результатов эволюции
местной фауны
высших приматов. Но тот же Кук
признает, что проблема эволюции фаун млекопитающих
на южных
континентах
все еще представляется
очень сложной.
93
особо высокого развития у них
большого пальца кисти и коры
головного мозга,
ожидать появления
совокупности
необходимых
предпосылок
к очеловечению обезьяны
безусловно не приходится.
Игрунки в этом случае тоже вовсе не
подходят. Гипотезы
о происхождении
человека
из американских
обезьян
несостоятельны.
Достаточно привести хотя
бы несколько черт их резкого
анатомического
несходства с
обезьянами Старого
Света и тем самым с человеком.
Так, у американских обезьян
вместо двух малых коренных
зубов имеется три; наружный слуховой проход
не длинный,
а короткий; горловые
мешки развиваются
не из
морганьевых
желудочков.
Еще менее допустима мысль
о непосредственном возникновении
человека от еще
более низкоорганизованных
приматов,
т. е. от долгопятов
или же лемуров.
Однако
в научной литературе встречаются
и подобные гипотезы, резко
противоречащие дарвинизму.
Борьбе с антидарвинистическими
гипотезами антропогенеза
посвящена следующая глава.
Оглавление
| Предисловие |
5
|
Часть I. Дарвиновская и другие гипотезы антропогенеза
|
7
|
| Глава первая |
Дарвин о происхождении человека |
8
|
| |
Идея антропогенеза до Дарвина
|
8
|
| |
Дарвин об эволюции животного мира
|
13
|
| |
Человеческая родословная по Дарвину
|
17
|
| |
Очерк развития знаний о приматах
|
29
|
| |
Развитие приматологии в СССР |
40
|
| Глава вторая |
Человекообразные обезьяны и их происхождение |
56
|
| |
Современные антропоиды
|
56
|
| |
Ископаемые антропоиды |
70
|
| Глава третья |
Позднейшие гипотезы происхождения человека
и их критика
|
95
|
| |
Религиозные трактовки антропогенеза
|
95
|
| |
Тарзиальная гипотеза
|
99
|
| |
Симиальные гипотезы
|
104
|
| |
Гипотеза антропогенеза Осборна
|
111
|
| |
Гипотеза антропогенеза Вейденрейха
|
119
|
| |
Некоторые факторы гоминизации и вымирания ископаемых антропоидов плиоцена
и плейстоцена |
124
|
| Часть II.Особенности строения человеческого тела
и возникновение древнейших людей |
133
|
| Глава первая |
Человек как примат |
134
|
| |
Черты приспособленности человеческого тела к прямохождению
|
134
|
| |
Характерные отличительные особенности человеческого тела, не стоящие
в непосредственной связи с прямохождением
|
140
|
| |
Специальные черты сходства между человеком и антропоидами
|
145
|
| |
Рудименты и атавизмы у человека |
148
|
| Глава вторая |
Роль труда и прямохождения в антропогенезе
|
154
|
| |
Роль труда
|
154
|
| |
Способы передвижения у человекообразных обезьян
|
157
|
| |
Вес тела в центр тяжести у человека и обезьян
|
165
|
| |
Нижние конечности
|
168
|
| |
Костный таз, позвоночник и грудная клетка
|
173
|
| |
Верхние конечности
|
180
|
| |
Пропорции тела и асимметрии
|
185
|
| |
Череп |
189
|
| Глава третья |
Головной мозг и высшая нервная деятельность
человека и обезьян
|
199
|
| |
Головной мозг и анализаторы человека и обезьян
|
199
|
| |
Развитие периферических частей анализаторов
|
210
|
| |
Высшая нервная деятельность обезьян
|
226
|
| |
Вторая сигнальная система — характерное отличие мышления человека |
246
|
| Глава четвертая |
Стадность у обезьян и зачаточные формы труда
|
249
|
| |
Стадность у обезьян
|
249
|
| |
Зачаточные формы труда
|
263
|
| |
Антропогенез и его факторы |
272
|
Часть III. Формирование человека по данным палеоантропологии
|
233
|
| Глава первая |
Первая ступень: древнейшие люди (питекантропы)
|
284
|
| |
Питекантроп острова Явы
|
284
|
| |
Синантроп
|
292
|
| |
Гейдельбергский человек |
305
|
| Глава вторая |
Вторая ступень: древние люди (палеоантропы)
|
309
|
| |
Ледниковая эпоха
|
309
|
| |
Неандертальцы и их физический тип
|
314
|
| |
Находки неандертальцев в СССР
|
322
|
| |
Палестинские неандертальцы
|
330
|
| |
Образ жизни древних людей
|
339
|
| |
Развитие мозга у ископаемых людей |
343
|
| Глава третья |
Современные люди (неоантропы)
|
356
|
| |
Человек верхнего палеолита
|
356
|
| |
Ошибочные гипотезы происхождения современного человека и их критика
|
370
|
| |
Человеческие расы
|
380
|
| |
Факторы формирования рас
|
387
|
| |
Наука против расизма |
394
|
| Summary |
|
403
|
| Приложение 1. |
Предложения по биологическим аспектам
расовой проблемы
|
406
|
| Приложение 2. |
Декларация о расе и расовых предрассудках
|
410
|
| Литература |
|
414
|