Глава третья
Позднейшие гипотезы происхождения человека
и их критика
Религиозные трактовки антропогенеза
Из предыдущего следует, что учение Дарвина об антропогенезе в той его части,
которая трактует о человеческой родословной и о ближайших предках людей на
Земле, вполне подтверждается современной биологической наукой, раскрывающей
все глубже картину эволюции животного мира и факторов развития организмов.
Дарвинизм нанес сокрушительный удар по религии, которая приводит к телеологическим
ложным представлениям об извечной предустановленной гармонии в природе либо
о развитии в живой и неживой природе «по плану божественного творения». До
сих пор в умах очень многих людей господствует идея о том, что изменения,
происходящие в мире, не являются связанными между собой.
Метафизически мыслящие люди считают, что мир остается неизменным и управляется
создавшим его божественным существом. Идеалистическое метафизическое мировоззрение,
лежащее в основе религии, составляет полную
противоположность диалектичен кому материализму как подлинно научному мировоззрению.
Религиозное миропонимание не позволяет понять появление новых качеств из
прежних, оно не допускает перехода количества в качество. Поэтому метафизики
отвергают происхождение человека от обезьяны.
Для очень многих западных антропологов проблема антропогенеза остается не
связанной с ведущей ролью труда. Именно этот фактор стоит на первом плане
в марксистско-ленинском объяснении качественно особого процесса формирования
человека.
Капиталистический строй мешает многим буржуазным ученым правильно понимать
процессы развития в природе и обществе, как совершающиеся диалектически;
он нередко диктует им такие выводы из фактов, которые находятся в противоречии
с последними. Своеобразным представителем таких ученых является крупный английский
биолог-эволюционист Альфред Уоллес (1823—1913).
Он не допускает, что тело человека развилось из тела предка,
95
похожего на человекообразную обезьяну, и чтоб крупный головной мозг человека
смог развиться из мозга обезьяны на основе таких факторов эволюции, как
изменчивость и естественный отбор.
Особенно не согласен Уоллес с возможностью развития человеческой психики
на основе высшей нервной деятельности животного. «Так, — говорит Уоллес,
— у нас есть умственные и нравственные «способности, которые не могли развиться
этим путем, но должны были иметь другое происхождение; и для этого происхождения
мы можем найти достаточную причину только в невидимом духовном мире» (1918,
стр. 729— 730). В связи с этим мнением Уоллеса напомним, что Линней, признавая
значительное сходство человека с обезьянами, провозглашал его душу частицей
божественного духа.
Уоллес выступает как яркий представитель метафизического мировоззрения. Он
лишь высказывает откровенно то, о чем обычно умалчивали многие другие эволюционисты
его времени. Пытаясь возможно теснее увязать эволюционизм с религией, он
писал, что в развитии человечества высший разум руководил для более благой
цеди законом естественного отбора, т. е., очевидно, для того, чтобы человек,
как наиболее высоко развитое животное, мог думать о боге!
Другие ученые в противовес дарвинизму неправильно считали, что строение обезьяны
должно быть выведено из строения человека как прообраза всех животных. Так,
по словам Иоганна Ранке (Ranke, 1897), наивысшая форма образования черепа,
именно человеческая, есть общий исходный пункт для развития черепа по всему
ряду млекопитающих.
Подобный же взгляд был развит позднее И. Кольманом (Kollmann, 1938), по мнению
которого общий предок людей и человекообразных обезьян обладал более высоким
черепом с округленной формой в отличие от низких черепов современных антропоидов.
Кольман основывался на том факте, что новорожденная обезьяна имеет по форме
черепа большее сходство со взрослым человеком, чем со взрослой обезьяной.
Однако мнение Кольмана и Ранке несостоятельно, так как в утробном развитии
череп более округлый и высокий не только у обезьян, но и у других млекопитающих,
и заключение, что млекопитающие произошли от людей, явно не соответствует
фактам палеонтологии и других биологических наук.
Некоторые авторы XX в., например Е. Даке (Dacque, 1935) и О. Клейншмидт (Kleinschmidt,
1931), пытаются под флагом науки воскресить идеи Кольмана. Даке пытался согласовать
научные данные о происхождении человека с библейским мифом о творении первых
людей.
Что же касается Клейншмидта, то он ошибочно считает всех ископаемых людей
и австралопитека за расы современного человека; кроме того, он отводит линии
других антропоидов далеко от родословной человека, объединяя человекообразных
обезьян в отдельный круг форм, чуждых человеку.
Известный антиэволюционисг М. Вестенгёфер (Westenhofer, 1935) старается
доказать, что че-
96
ловек произошел непосредственно от низших млекопитающих, минуя стадию обезьяны.
В подтверждение своей гипотезы он пытается опереться на факт сложного
строения человеческой почки, состоящей примерно из десятка долек или почечных
пирамид.
В этом действительно состоит одно из заметных отличий в строении тела
человека от обезьян и других приматов.
Однако что касается человекообразных обезьян, то у разных экземпляров
шимпанзе насчитывается до семи долек, у гориллы одна, у гиббонов до четырех,
у орангутана
четыре пирамиды слиты вместе и открываются общим сосочком в почечную
лоханку. У мартышек, макаков, павианов и прочих собакообразных обезьян
Старого Света
имеется лишь по одной дольке. У обезьян Нового Света тоже одна долька,
но у некоторых она имеет сложное строение, срастаясь из 4—6 пирамид.
В противовес гипотезе Вестенгёфера следует указать на факт большой изменчивости
числа долек у человека (от трех до двадцати). Почка явно подразделена
еще у человеческого плода. Сложность ее состава из пирамид отмечается
и у обезьян.
Хотя подразделенность почки у человека и исключительно сильная, но можно
показать способ ее развития из типа почки антропоидов.
Вестенгёфер, будучи последователем немецкого патолога Рудольфа Вирхова,
выступавшего против учения Дарвина и отвергавшего взгляд на питекантропа
как на древнейшего
человека, является ярым антидарвинистом и в своей концепции антропогенеза.
Стоя на позиции непознаваемости сущности этого процесса, Вестенгёфер
предпочитает научному знанию «поэтически интуитивный дух».
Механическое «соединение» науки и религии, эволюции и чуда в объяснении
антропогенеза свидетельствует о бессилии попыток реакционных биологов
примирить естествознание
с религией, что, как известно, делалось неоднократно раньше и делается
теперь.
Так, реакционный ученый Франц Кох (Koch, 1929) говорит следующее: «В
случае дарвинизма речь идет о чисто естественнонаучной проблеме, именно
о проблеме
внешнего мира, которая не имеет ничего общего с проблемами нашего внутреннего
мира, т. е. с вопросами религии или философскими убеждениями. Лишь невежество
или нетерпимость могут безоговорочно ставить дарвинизм в тесную связь
с материализмом или атеизмом».
Впервые полностью опубликованная С. Л. Соболем автобиография Ч. Дарвина
(1957), в которой великий ученый ясно выразил свои материалистические
и атеистические
взгляды, лишний раз показывает, что «невежество и нетерпимость» Коху
надо было искать не в лагере материализма, а у самого себя и у своих
единомышленников.
В таком же духе высказываются Г. Вейнерт (1935) и В. Бавинк (Bavink,
1933). По их мнению, вопрос о происхождении человека якобы не имеет никакого
отношения
к морали и религии.
Но все это только бесплодные усилия примирить два непримиримых миропонимания:
идеалистическое и материалистическое. Нау-
97
ка и религия несовместимы. В основе религии лежит вера в бога — творца
мира. Основу науки составляет метод познания природы и общества, метод исследования
закономерностей их развития.
Буржуазия культивирует религию. Реакционные буржуазные ученые, выполняя заказ
господствующих классов, оберегают вместе с церковниками религию от ударов,
наносимых ей наукой. Ленин неоднократно указывал, что угнетающие классы нуждаются
для охраны своего господства в социальной функции религии.
Проблема происхождения человека вовсе не есть какой-нибудь отвлеченный научный
вопрос, возбуждающий лишь кабинетные, теоретические споры. Вокруг него, наоборот,
разыгрываются схватки, являющиеся отражением классовой борьбы. Достаточно
напомнить об «обезьяньем процессе» в Америке.
В 1925 г. в г. Дайтоне, штат Теннесси, судили молодого учителя Джона Скопса
— приверженца эволюции — за то, что он, несмотря на запрещение, рассказывал
в школе ученикам о теории Дарвина и его гипотезе о происхождении человека
от обезьяны (Полетика, 1926).
Известно, что в разных штатах США существует на основе местного законодательства
запрещение преподавания эволюционного учения. В штате Теннесси 23 марта 1925
г. губернатором был утвержден следующий закон: «Да будет установлено генеральным
собранием штата, что будет незаконно для каждого учителя в каждом колледже,
в каждой нормальной или другой общественной школе штата, содержащейся целиком
или
частично на общественные школьные фонды штата, обучать всякой теории, которая
отрицает рассказ о божественном творении человека, как учит библия, и обучать
вместо этого, что человек произошел от животных низшего порядка». Позже закон
подвергался очередному обсуждению в законодательном собрании штата в конце
1934 г. и по решению большинства членов собрания его действие было вновь
продлено.
Суд над Скопсом происходил при громадном стечении народа, так как кроме местных
жителей присутствовало очень много приезжих. В качестве главного обвинителя
выступал кандидат в президенты бывший министр Уильям Дженнингс Брайан: в
руках он держал библию как «фундамент жизни».
Однако библия вовсе не помогла Брайану: пытаясь отвечать на ядовитые вопросы
защитников Скопса, например по поводу кита, якобы проглотившего пророка Иону,
насчет Навина, якобы остановившего Солнце, Брайан публично обнаружил свое
полное невежество в естествознании и истории, а заодно и незнание самой библии.
По выражению крупнейшего прогрессивного английского писателя Бернарда Шоу,
континент Америки в результате «обезьяньего процесса» оказался посмешищем
в глазах всего цивилизованного мира. Но, несмотря ни на что, классовый буржуазный
суд вынес обвинительный приговор и присудил Скопса к штрафу в сто долларов.
Религия продолжает быть верной пособницей капитализма. Ка-
98
питалистические государства всемерно поддерживают религию и утверждают
метафизическое мировоззрение в целях обеспечения своего господства. Миру
строящегося социализма
и коммунизма принадлежит диалектико-материалистическое мировоззрение, которое
является непримиримым противником религии.
Диалектический и исторический материализм служит могучим орудием в глубоком
преобразовании человеческого общества: при построении социализма и далее
коммунизма коренным образом меняется социально-экономическая основа, глубоко
изменяется мировоззрение широчайших масс трудящихся, освобождающихся от пут
религии и тем самым от старого метафизического, идеалистического мировоззрения.
Распространение правильного понимания явлений природы и общества, в частности
процесса антропогенеза, ускоряет смену мировоззрения.
Тарзиальная гипотеза
Представления Дарвина и Энгельса о развитии человека из ископаемой человекообразной
обезьяны различные биологи пытаются заменить идеями о его происхождении от
других приматов. Одним из таких ученых является английский биолог и анатом
Фредерик Вуд Джонс, создавший подробно разработанную гипотезу о происхождении
человека непосредственно от древнетретичного долгопята.
Единственными представителями современных долгопятов (рис. 41) являются маленькие
зверьки,
живущие в глухих тропических лесах на островах Индо-Малайской области и
составляющие характерный элемент местной фауны. По внешнему виду долгопяты
напоминают тушканчиков. У них огромные глаза, типичные для ночного животного,
очень длинные задние и очень короткие передние конечности, а также длинный
хвост, который служит им не только рулем и противовесом при ловких прыжках
с ветки на ветку, но и опорой при сидении. Передвигается долгопят в более
или менее вертикальном положении с помощью задних конечностей. Пяточная и
ладьевидная кости стопы у него очень длинные, откуда и название долгопят,
или тарзий.
Название тарзиальной гипотезы происходит от той же особенности. По Буду Джонсу,
долгопят сходен с человеком по многим чертам строения сильнее, чем последний
с антропоидами. Вуд Джонс утверждает, что вертикальное положение тела долгопята
при передвижении является исходным пунктом развития прямохождения у человеческих
предков.
Единственное зерно истины здесь заключается только в том, что долгопяты по
сравнению с лемурами несколько больше похожи на обезьян, и многие ученые
полагают, что именно группы ископаемых долгопятов Старого и Нового Света
дали начало соответствующим группам обезьян. Укажем, в частности, на такую
черту, как наличие стенки между глазной и височной ямами, в то время как
у лемуров в этом месте находится сквозное отверстие, окаймленное костным
кольцом (Hill, 1955).
99

Рис. 41. Долгопят привидение (Tarsius
spectrum Storr)
По Э. Г. Буленжеру, 1936
|
Но по некоторым признакам долгопяты примитивнее лемуров и сближаются с насекомоядными
млекопитающими, например в зубной системе (наличие переднего бугорка на нижних
молярах, т. е. параконида, отсутствующего у всех прочих приматов уже с ранних
времен третичного периода) и в кишечнике (очень простая форма толстой кишки).
Таким образом, вопрос о месте долгопятов в филогении приматов оказывается
достаточно сложным.
В настоящее время имеется каких-нибудь два-три вида долгопятов. Они представляют
собой специализированных потомков группы, некогда, в первые эпохи третичного
периода, бывшей весьма многочисленной и населявшей обширные области в Европе,
Азии и Северной Америке. Многие долгопяты,
жившие в самом начале третичного периода — в эпоху палеоцена, были по строению
тела более близки к лемурам, с которыми имели общих предков в виде верхнемеловых
насекомоядных млекопитающих. Некоторые примитивные долгопяты эпохи эоцена
дали начало обезьянам как в Новом, так и в Старом Свете (Гремяцкий, 1955).
В группе же обезьян Старого Света в последнюю эпоху третичного периода развились
ближайшие предки людей в виде наземных двуногих высокоразвитых человекообразных
обезьян. Но из этого, конечно, нельзя делать того заключения, которое стремится
доказать Вуд Джонс, что человек произошел непосредственно от древнетретичного
долгопята.
Вуд Джонс выступил с гипотезой о происхождении человека от долгопятов еще
в 1916 г. Однако эта гипотеза не встретила поддержки среди ученых. В 1929
г. вышла в свет вторая монография Вуда Джонса, в которой он собрал воедино
весь арсенал своих доказательств происхождения человека не от обезьяны, а
от долгопята. Вуд Джонс в этой монографии подробно разбирает ряд особенностей
строения долгопята, сопоставляя его с человеком и остальными приматами. В
частности, он старается доказать, что все черты сходства между человеком
и антропоидами являются лишь результатом параллельного развития, а вовсе
не выражением кров-
100
ного родства, как это считает большинство антропологов и других ученых.
Он считает, что человекообразные обезьяны ближе к низшим обезьянам, чем к
человеку.
В качестве важного аргумента Вуд Джонс приводит тот факт, что пропорции тела
у человека (короткие руки и длинные ноги) отличны от характерных для всех
человекообразных обезьян (длинные руки и короткие ноги), между тем у долгопята
пропорции тела якобы сильнее напоминают человеческие. Вуд Джонс присоединяет
также некоторые особенности волосяного покрова (в частности, характер расположения
волосяных токов на теле), укороченность лицевого отдела черепа, отсутствие
косточек в половом члене и клиторе, особенности формы и строения наружных
половых органов у самки и другие черты видимого сходства человека с долгопятом.
По мнению Вуда Джонса, у человека есть черты весьма примитивного характера,
которые указывают на очень древние стадии эволюции млекопитающих животных,
послуживших предками человеку: таковы особенности в строении ключицы и некоторых
мышц, которые встречаются кроме человека только у яйцекладущих млекопитающих,
например у утконоса; по строению кисти человек будто бы напоминает таких
древнейших животных, как амфибии каменноугольного периода палеозойской эры.
Гипотеза Вуда Джонса встретила резкую критику отечественных и зарубежных
авторов. Одни из приведенных Вудом Джонсом данных не соответствуют истинному
положению вещей, другие допускают иное объяснение. Так, например, Вуд Джонс
особенно подчеркивает пункт, касающийся формы корней у человеческих премоляров.
Известно, что у человека нижние предкоренные зубы имеют обычно по одному
корню вместо двух, как это типично и для всех узконосых обезьян. При этом
корни премоляров у последних располагаются вдоль зубного ряда. Когда же у
человека их два, то один корень располагается на щечной стороне, а другой
на язычной.
Верхние предкоренные зубы у узконосых обезьян нормально имеют три корня,
а у человека их только один-два, или корень подразделен надвое не полностью,
причем и здесь корни у человека стоят в щелочно-язычном положении, на что
особое внимание обращает Вуд Джонс. Он допускает, что расположение корней
верхних премоляров у человека еще можно было бы вывести из зубной системы
крупных человекообразных обезьян, ибо при наличии трех корней два стоят в
щечно-язычном положении. Но нижние предкоренные зубы человека остаются без
удовлетворительного объяснения, так как, по утверждению Вуда Джонса, у узконосых
обезьян они никогда не имеют двух корней, расположенных в щечно-язычном направлении,
а равно и трех корней, что, однако, иногда наблюдается у нижних премоляров
человека.
В этом случае Вуд Джонс ошибается, так как подобное расположение корней было
найдено и у некоторых высших узконосых обезьян.
101
В научной литературе описаны нижние премоляры с тремя корнями у шимпанзе.
Шарль Беннежан (1936) приводит описание и фотографию нижней челюсти шимпанзе
с таким премоляром.
Франц Вейденрейх (1937), исследовавший строение премоляров и их корней у
китайских древнейших людей, или синантропов, решительно утверждает, что они
представляют собой развитие типа предкоренных зубов ископаемых человекообразных
обезьян. Грегори (1934) объясняет изменение в положении корней премоляров
у человека процессом укорочения челюстей в ходе антропогенеза.
Изучение зубов на черепах людей и антропоидов в Институте и Музее антропологии
(Москва), произведенное нами, приводит, однако, к возможности еще одного
объяснения, которое заключается в том, что из трех корней премоляров нижней
челюсти в зубной системе предков человека передний не сохранился, а остались
два, стоявших в поперечном направлении. Такое предположение тоже свидетельствует
против гипотезы Вуда Джонса.
Существует еще очень много фактов, которые противоречат тарзиальной гипотезе.
Так, вопреки утверждению Вуда Джонса кровь долгопята вовсе не родственна
человеческой. Однако между кровью человека, шимпанзе и других антропоидов
отмечена очень большая близость, иногда почти тождество. Фредерик Тильней
(Tilney, 1928) сообщает о том, что якобы по реакции преципитации долгопят
ближе к человеку, орангутану и гиббону, чем к макаку и американским обезьянам.
Но это
указание явно ошибочно: по данным Ле Грос Кларка (Clark, 1934), опыты преципитации
с кровью долгопята и лори дали отрицательный результат при испытании на родство
с человеческой кровью: не получилось ни осадка, ни помутнения, ни кольца.
Вуд Джонс приводит следующие черты сходства между низшими узконосыми и человекообразными
обезьянами, отличающие их от человека;
1) носовые косточки рано срастаются между собой;
2) лобная и височная кости сближены, особенно у гориллы и шимпанзе1;
3) верхние предкоренные зубы имеют три корня;
4) существуют заметные половые различия в величине клыков;
5) имеется длинный наружный костный слуховой проход (только у орангутана
он короткий);
6) от дуги аорты у всех этих обезьян отходят лишь два арте-
1 Когда имеется соединение лобной и височной костей на определенном
протяжении с помощью шва, то говорят о лобно-височном птерионе. Для лемуров
и долгопятов,
американских обезьян, гиббона, орангутана и человека характерен другой птерионе
преимущественным контактом крыла основной кости с теменной костью: алисфеноидно-париетальный,
или крыловидно-теменной, птерион в виде лежачей буквы «Н» (эн), а не стоячей,
как при лобно-височном способе соединения костей в этой области (у многих
американских обезьян здесь принимает участие еще скуловая кость рядом с крылом
и получается скулово-алисфеноидно-теменная форма птериона). Некоторые авторы
ставят характер области птериона и вариации его формы в связь с развитием
у приматов височной доли как характерного для них участка больших полушарий
мозга. Во всяком случае форма птериона характеризует ту или иную группу приматов
в помощь систематикам.
102
риальных ствола (у гориллы и шимпанзе встречается форма, свойственная человеку);
7) имеется поверхностная плечевая артерия;
8) в задней конечности есть крупная сафеновая артерия;
9) развиты гортанные воздушные мешки (у большинства гиббонов они не развиты);
10) в составе почки у большинства есть лишь одна пирамида;
11) в теменно-затылочной части коры головного мозга сильно развита обезьянья
борозда.
К этим признакам можно было бы еще присоединить то, что, например, шимпанзе
и орангутан, по Чарлзу Зоннтагу (Sonntag, 1923), имеют гораздо менее развитую
систему лимфатических желез, чем человек: у шимпанзе 21 группа, у орангутана
20, а у человека 48. Отметим еще одно различие в кровеносной системе. У человека
подключичная артерия при вхождении в верхнюю конечность приобретает название
плечевой и идет в составе плеча глубже его самого крупного нерва, а именно,
срединного, или медианного. В отличие от человека у низших и человекообразных
обезьян плечевая артерия лежит поверхностно по отношению к нерву. Этот факт
объясняется тем, что в зародышевом состоянии закладываются два сосуда плеча:
один, идущий более поверхностно, и другой, лежащий глубже от срединного(медианного)
нерва. По мере дальнейшего развития у человека атрофируется поверхностная
ветвь, а у человекообразных обезьян — глубинная.
Приведенные выше черты сходства между человекообразными и мартышкообразными
обезьянами
сильно уступают, однако, тем общим чертам строения, которые были установлены
еще Томасом Гексли для человека я человекообразных обезьян и получили свое
краткое выражение в его известном выводе об их большем сходстве. По Гексли,
человекообразные обезьяны более сходны с человеком, чем с низшими обезьянами.
Он пишет: «Итак, какой бы ряд органов ни взялись мы изучать, сравнение
уклонений, коим они подвергаются у различных родов обезьян, приводит нас
постоянно к
одному и тому же заключению, а именно, что анатомические различия, отделяющие
человека от гориллы и шимпанзе, не так еще велики, как те, которые отделяют
гориллу от низших обезьян» (1864, стр. 117).
Низшие узконосые обезьяны являются своеобразно специализированными животными:
они обладают седалищными мозолями и защечными мешками; имеющиеся на коренных
зубах четыре бугорка характерно соединены с помощью поперечных гребней.
Мартышкообразным свойственно также более примитивное строение головного мозга,
кровеносные
сосуды которого устроены по иному типу, чем у человека и антропоидов. Именно
при так называемом обезьяньем типе имеется лишь одна передняя мозговая
артерия, напоминающая по своему внешнему виду основную артерию. Между тем
у человека
и антропоидов, как и у большинства других млекопитающих, имеются две передние
мозговые артерии, связанные между собой передней соединительной артерией
(Гиндце, Федотова, 1932, стр. 107—112). Но так как предки у
103
низших и высших обезьян были общие, то этим объясняются черты более близкого
сходства между ними. Многочисленные же черты сходства человекоподобных
обезьян с человеком служат одним из лучших доказательств происхождения
человека от
одного из ископаемых антропоидов.
Все указанные факты подтверждают симиальную гипотезу Дарвина и противоречат
тарзиальной гипотезе Вуда Джонса. Укороченность мордочки долгопята в значительной
мере объясняется очень сильным развитием органа зрения и редукцией органа
обоняния. Пропорции конечностей долгопята и человека не более, а даже менее
сходны между собой, чем у человека и лемура или обезьяны. Наличие у долгопята
очень длинного хвоста с разными специальными функциями, вплоть до отталкивания,
свидетельствует тоже не в пользу тарзиальной гипотезы. Будучи величиной
лишь с крысу, долгопят по размерам тела далеко уступает человекообразным
обезьянам.
Среди же ископаемых долгопятов ученым неизвестны ни средние, ни крупные,
ни тем более гигантские формы.
Против гипотезы Вуда Джонса свидетельствует также то, что долгопят ни в
коей мере не является общественным животным. Долгопяты встречаются только
в одиночку
или парами, но никогда не живут стадами: последнее характерно для многих
лемуров и для большинства обезьян. Этот факт весьма важен, так как человек
в первую очередь представляет собой общественное существо, производящее
орудия труда и употребляющее их.
Существенные возражения, которые вызывает тарзиальная гипотеза антропогенеза
Вуда Джонса, и ее явное несоответствие многочисленным фактам сравнительной
анатомии, приматологии, палеонтологии и антропологии приводят к выводу
о полной несостоятельности этой гипотезы (см.: Рогинский, 1948).
Симиальные гипотезы
Разные авторы (Карл Фогт, Флорентино Амегино, Джузеппе Серджи) пытались
доказать, что люди произошли от американских обезьян. Рассмотрим гипотезу
южноамериканского
ученого Амегино. Ему удалось найти остатки черепов и костей от скелетов
ископаемых приматов и других млекопитающих в древних (миоценовых) слоях
Южной Америки.
На основании изучения костных остатков Амегино построил новую родословную
человека, в которой поставил в число предков человека некоторых из обнаруженных
им ископаемых животных.
Однако критическое изучение черепов и костей из коллекции Амегино другими
учеными, в особенности Bluntschli (1913), привело совсем к иным результатам.
Некоторые остатки, отнесенные Амегино к приматам, оказались принадлежащими
другим млекопитающим, а представители действительно ископаемых приматов
вроде патагонского гомункулюса не отличались существенно от некоторых
современных американских обезьян. Гипотеза Амегино потерпела полное поражение,
хотя
в
других областях южноамерикан-
104
ской палеонтологии труды Амегино оказались весьма ценными.
К подобным «открытиям» некоторых американских ученых относится и находка
гесперопитека. Единственный зуб гесперопитека был найден Гарольдом Куком
в 1922 г. в нижнеплиоценовых слоях Вайоминга (Северная Америка). Находка
произвела сенсацию в ученом мире вследствие того, что такие крупные американские
палеонтологи, как Генри Осборн, Уильям Грегори и Мило Хелльман, определили
зуб как принадлежащий ископаемой человекообразной обезьяне, впервые открытой
на материке Америки. В печати появились далеко идущие догадки о родственных
связях и миграциях гесперопитеков. Такова, например, статья известного
английского биолога Дж. Эллиота Смита в одном из английских журналов, на
обложке которого
были изображены самец и самка гесперопитека, а в тексте гесперопитек был
назван «примитивным членом семейства людей».
Тщательное дополнительное исследование зуба (Gregory, 1927) и новые находки
показали, что была открыта не обезьяна, а одна из ископаемых форм североамериканских
свиней пекари из рода простенопс.
Оригинально использовал находку гесперопитека реакционный немецкий ученый
Франц Кох (Koch, 1929). В своей монографии он поместил любопытную родословную
человека, в которой гесперопитек изображен в качестве предка современного
человека. А так как в монографии Кох отвел центральное место североевропейской,
или нордической, расе как
высшему типу человечества, то получилось, что в предках арийской расы оказалась
ископаемая североамериканская свинья.
Несомненно требуется сугубо осторожное отношение к очень многим гипотезам
и теориям антропогенеза, в таком изобилии предлагаемых реакционными буржуазными
учеными. В самом деле, возможность находки зуба человекообразной обезьяны
в Северной Америке маловероятна сама по себе, потому что таких обезьян
там, очевидно, никогда и не было. Кроме того, маммалиологам известно, что
коренные
зубы обезьян и свиней обнаруживают подчас значительное сходство в форме
и строении1.
Итак, американские обезьяны не являются предками человека. Полагают, что
коренные обитатели Америки попали туда сравнительно недавно, около 25 тыс.
лет назад. Их предки, вероятно, перебрались из Азии через перешеек, на
месте которого впоследствии, после таяния ледников, образовался Берингов
пролив.
Индейцы представляют собой древнюю ветвь монголоидной расы, первоначально
развившейся в Азии. Европейцы проникли в Америку гораздо позже индейцев.
Они прочно обосновались там лишь после 1492 г., т. е. после открытия этого
материка Христофором Колумбом.
1Известный австрийский палеонтолог Отенио Абель (Abel, 1931),
анализируя особенности зубной системы приматов, даже относит человеческие
зубы к суидному
(свиному)
типу, характеризующемуся наличием отдельных конусовидных округлых бугорков
на жевательной поверхности коренных зубов, иначе бунодонтностью.
105
Некоторые реакционные буржуазные ученые, очевидно, стремились создать видимость
доказательств в пользу происхождения индейцев от местного антропоида типа
гесперопитека. Особенно выгодно это было бы приверженцам учения о полигенизме,
т. е. о множественности предков человечества. Полигенисты считают, что
различные человеческие расы произошли от разных видов обезьян. Так, по
представлениям
антропологов Теодора Арльдта и Джузеппе Серджи, современное человечество
составляет не один вид, а несколько видов или даже родов. Например, Серджи
(Sergi, 1911) принимает главные человеческие расы за роды, включающие в
свой состав одиннадцать видов и свыше сорока более мелких систематических
групп.
Однако уже давно было известно, что человеческие расы при всех своих различиях
близко родственны: это следует уже из того, что они достаточно сходны
между собой. Европеоиды смешиваются с неграми, монголами, готтентотами, японцы
с малайцами, полинезийцами, негры с монголами. Оказывается, что потомство
от подобных браков вполне нормальное, здоровое и обладает в свою очередь
способностью к дальнейшему размножению. Подобные факты составляют одно
из важнейших доказательств неправоты полигенистов. Человеческие расы
настолько
близки между собой по анатомическим и физиологическим особенностям, что
до
сих пор не найдено существенных различий между ними, например в строении
головного мозга, в свойствах крови, функциональной деятельности желез
внутренней секреции. Отсюда вытекает правота дарвиновского моногенизма, по
которому человечество произошло от одного вида человекообразных обезьян,
а не от нескольких.
Полигенисты подбирают всякие научные и ненаучные материалы для построения
своих расовых теорий. Их гипотезы предназначены для оправдания всевозможного
угнетения и эксплуатации трудящихся масс своих и колониальных стран.
Весьма ярким представителем расистов и полигенистов можно считать реакционного
немецкого антрополога Германа Клаача (Klaatsch, 1922), по мнению которого,
«современная наука не может поддерживать то чрезмерное человеколюбие,
которое
хочет видеть наших братьев и сестер во всех низших человеческих расах».
Своеобразная позиция отражается в трудах французского реакционного антрополога
Жоржа Монтандона (Montandon, 1928), который развил в применении к человеку
гипотезу итальянского биолога Даниэля Розы (1918) о происхождении и
развитии жизни на Земле. Роза считает, что живые существа зародились сразу
на всей
поверхности земного шара. Это были бесчисленные крошечные одноклеточные
организмы самой примитивной организации, но относящиеся все к одному
и тому же виду.
Современные крупные и мелкие подразделения животного мира имеют самостоятельное
происхождение. Мало того, Роза предполагает, что появившиеся в разных
местах Земли первые живые существа могли путем самостоятельного развития
дать
начало
106
очень сходным организмам на различных континентах.
До какой крайности доходит Роза в своих построениях, видно из того, что,
по его мнению, даже два человека одной и той же деревни могут не иметь
ничего общего не только со времени возникновения человека, но и с самого
начального
этапа развития жизни на Земле.
Монтандон, применяя гипотезу Розы к человеку, объявляет, что люди возникли
на материках независимо друг от друга и, следовательно, не связаны между
собой родственными узами. Моптандон пытался даже доказать, что и в Америке
человек мог произойти от американской человекообразной обезьяны.
В поисках подтверждений своей гипотезы Монтандон натолкнулся на сведения
об одном любопытном случае, происшедшем в 1920 г. с геологом Фрэнсисом
де-Луа в Южной Америке. Путешествуя в обширном тропическом лесу на границе
государств
Венесуэлы и Эквадора, де-Луа вместе со своими спутниками якобы подвергся
нападению двух крупных обезьян.
По описанию де-Луа, это произошло на берегу реки. Послышался сильный
шум, и путешественники выскочили из лагеря. Две довольно большие обезьяны,
испуская
крики и ломая по пути ветки, бежали к лагерю. Подпустив обезьян на близкое
расстояние, путешественники выстрелами убили одну из них, оказавшуюся
самкой (рис. 42). Другая обезьяна была ранена, но убежала, скрывшись
в лесной
чаще (де-Луа полагает, что то был самец). Убитая обезьяна была посажена
на ящик
и сфотографирована. Де-Луа утверждает, что обезьяна была без хвоста,
но на фотографии не видно, действительно ли это так. Кроме того, по фотографии
трудно проверить
его заявление, что рост самки был около 160 см. Череп обезьяны де-Луа
взял с собой, равно как и нижнюю челюсть и шкуру. К сожалению, в трудных
условиях
очень длительной экспедиции были утеряны и шкура, и нижняя челюсть (насчитывающая
якобы 16 зубов и хранившаяся у начальника экспедиции де-Луа). Череп попал
к повару экспедиции, который стал держать в нем соль: от сырости и под
воздействием соли череп распался на части и был выброшен. Таким образом,
единственным
документом является фотография обезьяны и данное де-Луа описание.
Монтандон извлек рассказ де-Луа из забвения и начал оперировать им как
доказательством правоты гипотезы ологенизма, трактующей о самостоятельном
происхождении человека
в Америке. Он доказывал, что убитая самка представляет собой настоящую
американскую человекообразную обезьяну: крупные размеры тела, отсутствие
хвоста, человекообразность
лица — все для Монтандона служило положительной аргументацией.
После сообщения, сделанного Монтандоном в Парижской Академии наук, в
ряде журналов появились статьи об амерантропоиде. Напечатана была и статья
де-Луа,
в которой автор в свою очередь поддерживал гипотезу ологенизма. Но появился
и ряд критических заметок. В результате аме-
107

Рис. 42. Амерантропоид луасов
(Amerantropoides loysi. Montandon)
По Ф. де-Луа, 1931
|
рантропоид оказался не чем иным, как экземпляром крупной паукообразной
обезьяны коаты. может быть даже представителем вида золотолобых коат.
У убитой самки
на каждой руке было лишь по четыре пальца, а большой палец был атрофирован,
как и у коаты. Лицо сходно с лицом коаты. Носовая перегородка широкая,
как и у большинства американских обезьян. Туловище и конечности достаточно
тонкие,
гранильные. Хвост, надо полагать, у убитого экземпляра длинный и цепкий,
но, по-видимому, был спрятан за ящиком. По де-Луа и Монтандону, рост
сидя около 70 см, что не очень превосходит рост сидя (до 65 см) у современных
коат.
Вся совокупность данных приводит к заключению, что гипотеза ологенизма
и в данном случае потерпела поражение. Другие факты, приводимые Монтандоном
в защиту ологенизма, могут быть
объяснены иначе (Гремяцкий, 1933, стр. 64—82; 1934, стр. 55—67).
В основе гипотезы ологенеза Розы лежит механистическое предположение
о самопроизвольном; развитии видов лишь с помощью дихотомии, т. е. путем
подразделения на две
ветви: раннюю и позднюю. Из них ранняя, по утверждению Розы, обычно дальше
уже не делится и в конечном счете вымирает; запоздалая же имеет шансы
на
дальнейшее развитие опять-таки путем дихотомии и прогрессирует, эволюционирует.
Дихотомия, однако, представляет собой совершенно произвольное допущение:
один вид может дать начало не только двум, но и большему числу подвидов
или разновидностей. Из восьми главных современных рас, насчитываемых
Монтандоном, европейская, монгольская и негрская, по его мнению, оказываются
запоздалыми,
т. е. способными к дальнейшей эволюции и усовершенствованию; все же остальные
расы якобы являются ранними, т. е. обреченными на вымирание. Для антропологов
гипотеза ологенизма Монтандона в корне неприемлема.
Следующая группа гипотез антропогенеза трактует о происхождении человека
из группы обезьян Старого Света. Это так называемые симиальные гипотезы
в отличие от тарзиальной. Почти никто из биологов не производит человека
из
группы низших обезьян с их четвероногим хождением, более или менее длинным
хвостом, их защечными мешками и седалищными мозолями, хотя недавно в
предки человеку начали ставить, проконсула.
108
Обычно авторы симиальных гипотез анализируют вопрос о том, какой из
современных антропоидов обладает наиболее существенными чертами родства
с человеком.
В зависимости от такого подхода есть гиббоноидная, орангоидная, гориллоидная
и шимпанзоидная гипотезы антропогенеза или их комбинации.
Приверженцы гиббоноидной гипотезы пытаются основываться, например, на
том, что проплиопитек, обладавший, по-видимому, гиббоноидными особенностями,
дал начало плиопитекам, а через них и гиббонам, но проплиопитек был
общим
предком и для прочих антропоидов, а также для человека (Gregory, 1933).
Идею о гиббоноидных филогенетических связях человека высказывали палеонтолог
Гюи Пильгрим и антрополог Ганс Верт.
Однако двуногая походка у человека развилась под влиянием иных причин,
чем у гиббона. Последний с его чрезвычайно длинными руками перешел к
прямохождению в связи не с наземным, а с древесным образом жизни, с
привычкой брахиации,
или руколетания, при котором туловище животного принимает вертикальное
положение.
Человек не прошел через стадию сверхспециализации, как гиббон, иначе
это в более сильной степени отразилось бы на пропорциях конечностей человека
во время его утробного развития и в дальнейшие периоды жизни.
Адепты гиббоноидной гипотезы (к числу которых относились, в частности,
некоторые представители школы анатомов университета имени Джонса Гопкинса
в
США) указывают на то, что гиббоны сильнее, чем другие обезьяны, напоминают
человека по некоторым особенностям, каковы: расположение внутренних органов;
форма сжатой в спинно-брюшном направлении грудной клетки с ее очень
широкой и короткой грудинной костью; сильное развитие большого пальца
стопы и второго
пальца кисти; слабость наружного рельефа мозгового отдела черепа; форма
коренных зубов; строение нижней челюсти, которая у сростнопалых гиббонов,
или сиамангов,
имеет сравнительно хорошо выраженную подбородочную часть с зачаточным
подбородочным выступом.
Но такие особенности гиббонов, как малые размеры тела, отсутствие лобных
пазух, седалищные мозоли, более примитивный головной мозг, свободная
центральная косточка в запястье, не позволяют, по мнению Грегори и других
авторов,
принимать гипотезу о непосредственном происхождении человека из группы
гиббонов. Своеобразную
вариацию гиббоноидной гипотезы развивает Шульц (Schultz, 1936), который
приходит к выводу, что гиббоны и человек во многих существенных особенностях
удержали
более тесную близость между собой, чем та, которая в отношении тех же
самых черт существует между человеком и крупными человекообразными обезьянами.
Предки гоминид и гиббонов ответвились, по мнению Шульца, от общего ствола
человекообразных высших приматов даже раньше, чем предки орангутана отошли
от ствола крупных антропоидов (одновременно с гориллой и шимпанзе либо
не-
109
много раньше). Более близкие черты сходства и родства человека с крупными
современными и ископаемыми антропоидами не позволяют нам принять гиббоноидной
гипотезы Шульца.
Некоторые ученые выдвигали гипотезу о происхождении людей от орангутанов.
По Герману Клаачу (Klaatsch, 1936), ориньякский человек ведет начало
от орангутана, а неандерталец сродни горилле. Ганс Фриденталь (Friedenthal,
1926) указывал на относительно более высокий череп орангутана как на
один
из признаков «сходства» этой обезьяны с представителями монголоидной
расы.
Однако длиннорукий орангутан должен быть отведен из числа ближайших
сородичей человека из-за множества особых черт специализации. Таковы,
например:
щечные наросты у самцов; редукция большого пальца стопы, выражающаяся,
в частности,
в отсутствии на нем ногтя; оголенные места с грубой кожей в области
седалищных мозолей; неподразделенные на доли легкие; сильная изборожденность
эмали
почти на всех зубах; отсутствие язычка на нёбной занавеске; вогнутый,
«бульдогообразный» профиль лица, или так называемый «симогнатизм»,
в меньшей степени характерный
и для шимпанзе в отличие от гориллы с его ровным профилем лица и лицевого
отдела черепа; волнистый контур расположения пигмента на поперечном
разрезе рыже-коричневых волос орангутана («гофрированный» край пигментного
слоя);
громадные гортанные мешки; грудинная железа, встречающаяся у плодов
и молодых особей, гораздо чаще у самцов.
Что касается гориллы, то некоторые авторы, как, например Артур Кизс,
склоняются к признанию этого огромного антропоида за ближайшего сородича
человека.
Головной мозг гориллы крупнее и более сложно устроен, чем у шимпанзе
или орангутана.
Некоторые черты сходства в мужских органах размножения у гориллы и
человека по сравнению с шимпанзе поразительны; мужские половые клетки
у гориллы
чрезвычайно похожи на человеческие. Строение черепа самки гориллы,
по Вейденрейху (Weidenreich,
1943), наиболее полно отвечает представлению о форме черепа ближайшего
предка питекантропов.
Против гориллоидной гипотезы антропогенеза могут быть, однако, представлены
и серьезные возражения, так как череп самца гориллы, снабженный мощными
гребнями, наоборот, уводит этого гигантского антропоида и его ближайших
предков в сторону
от человеческой родословной. Кровь гориллы в реакции преципитации показывает
меньший осадок, чем кровь шимпанзе или даже гиббона, хотя и гораздо
более крупный, чем у орангутана, который в данном отношении стоит
ближе к
низшим узконосым обезьянам типа павианов.
Наиболее распространенной является шимпанзоидная симиальная гипотеза
антропогенеза. Такие знатоки, как Густав Швальбе, Вильям Грегори, Ганс
Вейнерт, наделяют
дриопитека и более поздних верхнетретичных предков человека вплоть
до австралопитека чертами, сближающими их с шимпанзе. Черты поразительного
сходства между
шимпанзе и чело-
110
веком подмечены в строении больших полушарий мозга. В общем облике
шимпанзе проглядывают черты гораздо меньшей специализации. У шимпанзе
нет столь
сильных половых различий между самцами и самками, как у горилл
или же у орангутанов.
Грегори (Gregory, 1933) даже называет шимпанзе «живым ископаемым».
Если говорить о ближайших предках человека, то совершенно очевидно,
что в результате накопления анатомо-физиологических данных по
антропоидам и человеку
тезис Дарвина о ближайшем родстве человека, в частности, с африканскими
антропоидами остается незыблемым. Но попытки поколебать такой вывод
продолжаются
и ныне.
Многие буржуазные ученые стремятся вообще устранить не только антропоидов,
но и прочих приматов из родословной человека, изобразить предка
человека не в виде обитателя деревьев, а в форме маленького двуногого
наземного
человечка эоантропа, который будто бы непосредственно возник из
примитивных низших
млекопитающих в самой глубине третичного периода. Марселен Буль,
не отрицающий родства человека и обезьяны, рисует этого неизвестного
науке,
загадочного
«предка» человека, как маленького представителя семейства людей,
обладающих более или менее вертикальным положением тела, очень
большой черепной
коробкой и соответственно очень крупным мозгом по сравнению с размерами
тела.
Буль не признает дриопитека за предка человека. Воззрение о
существовании третичного человека, близкое к гипотезе Буля об эоантропе,
высказывает также Осборн.
Гипотеза антропогенеза Осборна
Крупнейший американский палеонтолог Генри Осборн (1857—1935) является
ярким представителем тех реакционных буржуазных ученых, которые
стремятся примирить
науку с религией, провозглашая, что творение органического
мира осуществляется путем целенаправленной эволюции. Метафизическая
формула «творение
путем эволюции» особенно в ходу у некоторых американских биологов.
Осборн полагает, что третичные предки человека в виде эоантропов,
или «людей зари», обладали почти всеми качествами, свойственными
современному человеку.
По Осборну, предки людей не были связаны с предками человекообразных
обезьян,
т. е. человек произошел не из круга обезьян; люди развились
первоначально в Центральной Азии и их предки вели наземный
образ жизни. Осборн
допускает существование людей не только в плиоцене, но и в
более ранние эпохи
третичного периода.
Построения Осборна носят чисто гипотетический характер, они
не основаны на фактах. Его идеи вытекают лишь из общих ложных
посылок
созданной
им же гипотезы
аристогенеза. Осборн постулирует, что эволюция совершается
не столько в процессе естественного отбора или под влиянием
внешней
среды,
сколько благодаря
111
заложенной в организмах чудесной способности к самопроизвольному прогрессивному
развитию. Гипотезе Осборна противоречит все, включая данные, доказывающие
родство человека с обезьянами, и находки ископаемых обезьян и людей.
Равно несостоятельна и его попытка опереться на «третичные эолиты», т.
е. камни,
имеющие вид примитивнейших орудий, как на доказательство существования
третичных людей.
Взгляды Осборна на проблему происхождения человека претерпели значительные
изменения на протяжении его долголетней деятельности в области палеонтологии,
общей биологии и антропологии. По собственному признанию Осборна, он
до 1923 г. вполне разделял мысль о происхождении человечества от
одного вида ископаемых
человекообразных обезьян согласно учению Дарвина об эволюции. Однако
у него как у палеонтолога были и некоторые сомнения в правильности
обезьяньей гипотезы
антропогенеза. Они заключались в следующих трех главных пунктах: 1)
пропорции конечностей у ископаемых людей являются не антропоидными,
а чисто человеческими,
так как нижние конечности у питекантропа и неандертальца длинные, а
верхние конечности у последнего короткие; 2) строение кисти у неандертальцев
вполне
человеческое, так как первый палец развит не слабее, чем у современных
людей, а прочие пальцы не менее гибкие; 3) некоторые кремневые орудия,
например
из Ред-Крега (юго-восточный берег Англии), насчитывают 1200000 лет
древности.
Отсюда Осборн приходит на путь признания очень глубокой древности человека,
который мыслится им как существо с выпрямленным положением тела и со
способностями к труду и искусству, т. е. уже в значительно развитом
состоянии. Путешествие
с Чарлзом Уильямом Эндрюсом в пустыню Гоби явилось толчком к созданию
Осборном гипотезы об извечности человеческого типа и о прародине человечества
в Монголии
и Тибете. Во время экспедиции в Монголию с ее ландшафтами горного плато
и обилием диковинных вымерших гигантских пресмыкающихся Осборн представил
себе
именно здесь колыбель человечества, которая рисовалась ему как «область
с извилистыми потоками и редкими лесами вперемежку с равнинами, покрытыми
лугами»
(1929). Именно тут, утверждал Осборн, могли быстро развиться четвероногие
и двуногие типы млекопитающих, в том числе предки так называемых «обезьянолюдей».
Они могли здесь легче наблюдать окрестности, скрываться вовремя от
врагов, употреблять имеющиеся под рукой камни в качестве орудия. За
несколько лет
до Осборна сходная идея о прародине человечества в виде полупустынного
плато-плоскогорья была высказана американским палеонтологом Жозефом
Баррелем, но Осборн утверждает,
что он пришел к своей гипотезе самостоятельно. Осборн первоначально
опубликовал ее в печати в октябре 1923 г. В последующие годы он неоднократно
возвращается
к отдельным сторонам проблемы антропогенеза. Наконец, в 1929 г. Осборн
развивает
112
свою гипотезу в широко обоснованную «теорию полупустынного плато»,
выступая на съезде в честь 200-летия Американского философского
общества.
Трактуя о прародине человечества и о предполагаемых экологических
условиях жизни предков человека, Осборн полемизирует с утверждением
Дарвина
о том, что это была страна с теплым или жарким климатом, покрытая
лесами. В противовес
мнению Дарвина Осборн считает, что человек, уже давно приспособленный
к прямохождению, не мог развиться в обстановке тропического леса,
где не нашел бы и материала
для изготовления каменных орудий. В доказательство правоты своего
взгляда Осборн приводит соображение о том, что якобы «интеллектуально-прогрессивные
и самостоятельно-приспособительные, самоадаптивные типы человечества»
смогли формироваться на ровных плато, как в Монголии и Тибете»
(см.: Сушкин, 1928).
Изобретательность в способах добывания средств существования у
третичного человека должна была, по Осборну, вызываться тем, что
растительной
пищи тут было немного, а дичи достаточное количество. Тяжесть и
обостренность борьбы
за выживание в таких природных условиях, говорит Осборн, должны
были энергично содействовать употреблению деревянного и каменного
оружия
в целях охоты.
Наконец, суровость климата на горном плато заставила уже древнейших
гоминид перейти к использованию огня для согревания тела и изготовления
пищи. Охота
на животных не
могла не содействовать развитию у предков человека силы и проворства
ног, органов зрения и легких. Так рисует Осборн в своих представлениях
экологию
и образ жизни представителей «зари человечества».
Посмотрим теперь, каковы, по Осборну, физический тип, строение
и черты поведения третичных людей, живших раньше питекантропов.
Он
считает
питекантропов зашедшими
в тупик развития, реликтом от наидревнейшего человечества, остатком
группы, вытесненной более прогрессивными формами гоминид из южной
части материка
Азии в условия островной жизни Океании. По мнению Осборна, для
«людей зари», эоантропов, были уже издавна характерны следующие
черты:
1) прогрессивное развитие сознания, быстрое развитие лобного отдела
мозга;
2) двуногое хождение с развитием стопы и ее большого пальца, приспособленных
к хождению и беганию;
3) укорочение рук и удлинение ног;
4) развитие большого пальца кисти, приспособленной к выделке орудий;
5) умение выделывать орудия;
6) применение и намеренное изготовление орудий из разных сортов
дерева, кости и камня;
7) способность к планированию действий и изобретениям, направляемая
лобными долями мозга;
8) использование рук и орудий при нападении, защите и других проявлениях
жизнедеятельности;
9) передвижение на двух ногах при ходьбе, беге, странствованиях
и убегании от врагов.
113
10) лазанье по деревьям на манер медведя (обхватывается главный
ствол при помощи рук и ног).
Для предков современных обезьян, по Осборну, характерны следующие
отличительные признаки:
1) остановка в развитии рассудочных способностей и головного
мозга;
2) жизнь преимущественно на деревьях, вызывающая крайнюю приспособленность
обезьян к лазанью; привычка захватывать ветви всеми четырьмя
конечностями;
3) четвероногий тип передвижения по земле;
4) удлинение передних и укорочение задних конечностей;
5) редукция большого пальца кисти и отсутствие у него способности
оказывать помощь другим пальцам при выделывании орудий, а также
развитие приспособленности
к хватанию у большого пальца стопы;
6) способность кисти к обхватыванию ветвей;
7) приспособление всех четырех конечностей к искусству лазанья
по деревьям;
8) ограниченная способность к планированию действий, которая
выражается в постройке весьма примитивных гнезд на деревьях;
9) применение рук главным образом для лазанья по деревьям,
а затем уже для добывания пищи и схватывания врага;
10) употребление ног для лазанья по деревьям и для обхватывания
ветвей;
11) спасение от врагов путем убегания по ветвям деревьев;
12) лазанье по деревьям всегда поверх ветвей, а не с помощью
обхватывания толстых ветвей (или
ствола дерева), как это делает человек.
Таким образом, Осборн проводит самое резкое различие между
людьми и человекообразными обезьянами — не только современными,
но и
древними, третичными. Тем самым
он пытается подорвать дарвиновский тезис о происхождении человека
из
группы обезьян Старого Света. Антидарвинистическая гипотеза
антропогенеза Осборна,
видного американского палеонтолога, основывается на той общебиологической
теории аристогенеза, которую он развивал и защищал на протяжении
полувека своей научной деятельности.
Совершенно законно поставить вопрос, в чем именно заключается
связь гипотезы антропогенеза Осборна с его теорией аристогенеза.
Связь
между ними органическая
и неразрывная: первая представляет собой часть второй. Законом
аристогенеза, или ректиградации, Осборн постулирует постепенное,
на протяжении
долгих веков, появление у организмов новых черт адаптивного
характера, предшествующих
в
зародышевой плазме и на начальных стадиях развития независимых
от естественного отбора.
В монументальной монографии Осборна (Osborn, 1936) о хоботных
млекопитающих, опубликованной уже после его смерти, мы читаем
следующие строки:
«Из длительного изучения автором титанотериев и хоботных на
протяжении тридцати пяти лет
следуют не только принципы, управляющие классическими модифицирующими
способами эволюции и известные Ламарку и Дарвину (изменчивость,
развитие, вымирание),
но также вновь открытые и
114
до сих пор непризнанные принципы и способы возникновения новых
признаков путем аристогенеза, или творческой биомеханической
ректиградации» (т. I, стр. XIV). Принцип ректиградации
и положен Осборном в основу
объяснения эволюционного
процесса, который, по его мнению, имеет целенаправленный
«творческий» характер.
Следующий пример дает нам представление о явно идеалистическом
характере воззрений Осборна на эволюцию животного мира,
воскрешающих идеи
о психологическом творчестве животных. Изучив серию черепов
детенышей и взрослых особей эоценовых
парнокопытных — титанотериев, Осборн нашел, что у них
«рога возникают определенным, предуготованным образом». Произведя
исследование
серии голов и черепов современных
быков, Осборн установил, что задолго до появления костных
элементов на месте рога возникает округлое утолщение кожи
с усиленным
разрастанием волосяного
покрова и накоплением кератина. В связи с этим Осборн задает
вопрос, что же сперва возникает: психическое побуждение
к использованию рога, либо эпидермальная
кератиновая защита костного центра рога, либо сам рог?
Поразителен ответ Осборна: «Кажется, что психическая тенденция должна
предшествовать эпидермальной и что последняя предваряет
костное разрастание».
Таким образом получается, что в основе эволюционного взгляда
Осборна лежит неприкрытая автогенетическая идея. Он является
ортогенетиком,
поскольку признает в организмах скрытую, предопределенную
тенденцию к развитию в том или ином направлении. Самопроизвольные
изменения
зародышевой
плазмы,
имеющие
с момента возникновения адаптивный характер по отношению
к будущим изменениям среды,— это, по Осборну, ортогенетические
«биопризнаки»,
которые развиваются
самостоятельно, целесообразно и последовательно. Не довольствуясь
внесением в понятие ректиградации элемента «творчества»,
автор
теории аристогенеза
вкладывает сюда еще и элемент «улучшения»: он дает ректиградациям
новое название «аристогенов», обозначающее и лучшую наследственность
и одновременно
лучшую приспособленность организмов для предстоящей им
дальнейшей жизни. Следовательно, эволюционные воззрения
Осборна носят
явно выраженный телеологический характер: в мире организмов
идет творческая
эволюция,
направленная к определенной
цели.
Опираясь на свою полувековую научную и научно-пропагандистскую
деятельность, Осборн высоко ценил свои научно-философские
взгляды. В упомянутой
монографии о хоботных он пишет: «Благодаря ясному разделению
между изменениями
в пропорциях (аллойометрия) и возникновением новых биопризнаков
(аристогенезис), а также
благодаря недавним открытиям у хоботных многочисленных
линий предков и потомков — сорока одной или больше, изучение
этой
группы влечет
за собой полный переворот
в нашей философии биологии, в наших взглядах на природу
и
115
причины эволюции» (т. I, стр. XV). В других местах монографии
Осборн считает возможным безоговорочно прилагать к
человеку те же биологические
закономерности
или факторы, как и к хоботным, без всякого учета могущественного
влияния социальных факторов на формирование человечества,
представляющее качественно
особый процесс по сравнению с эволюцией всего остального
мира живых существ на Земле.
Подгоняя свою гипотезу происхождения человека под общебиологическую
теорию аристогенеза, Осборн не только сделал методологические
и фактические ошибки,
но и пошел значительно дальше, чем этого требовала
даже его теория аристогенеза. Основной постулат Осборна
гласит,
что
предковая
форма человека должна была
обладать на все 100% особенностями и потенциальными
возможностями развития, которые наблюдаются у потомков.
Но здесь перед
нами лишь иная формулировка
старого взгляда преформистов, утверждавших возможность
закладки будущих поколений в предыдущих: тезис Осборна
является чисто
метафизическим построением. Для
чего понадобилось Осборну воскрешать преформизм? Очевидно,
Осборну было важно как можно дальше развести друг от
друга линии эволюции
человека и человекообразных
обезьян, уменьшить явные противоречия между научной
концепцией антропогенеза
и религиозным мифом о чудесном творении человека в
готовом виде «по образу и подобию» бога-творца согласно
библии.
Не будучи в состоянии опровергнуть множество фактов,
свидетельствующих о кровном родстве и теснейшей анатомо-физиологической,
а также психологической близости между человеком и
обезьянами, Осборн
предпочитает держаться
в рамках
палеонтологии, где он чувствует себя авторитетом. На
основе
своих теоретических взглядов он, как уже сказано выше,
представляет
себе третичного предка
человека в виде эоантропа — небольшого двуногого наземного
человекообразного
примата
с проточеловеческими пропорциями конечностей, крупным
мозгом и сравнительно развитым интеллектом. Эоантроп
не испытал
древесной фазы эволюции,
разве лишь в форме кратковременного пребывания на деревьях
в начале
третичного
периода. Осборн даже близок к тому, чтобы сблизить
эоантропа с формой, близкой к какому-нибудь наземному
долгопяту
из эоцена, лишь бы отдалить
человека
от антропоидов. Он отводит эоантропа от общего ствола
приматов еще в олигоцене и не связывает его с антропоидами.
Сходство
же человека
с последними Осборн
объясняет параллелизмом и аналогией в адаптации, якобы
имевшими место в процессе развития человека, всех антропоидов
и некоторых
низших
обезьян.
В аристогенетической эоантропоидной гипотезе нельзя
не усмотреть связи с тарзиальной гипотезой. Вуд Джонс
тоже
объясняет черты
сходства между
человеком
и антропоидами гомеоморфией, т. е. «уподоблением формы»,
вроде того, что наблюдается между обезьянами Старого
и Нового Све-
116
та, которые обладают многими чертами сходства, хотя,
по-видимому, развивались независимо друг от друга
на отдельных материках
в течение десятков
миллионов лет. Однако внешнее сходство, о котором
говорит Вуд Джонс, носит несравненно
более отдаленный характер аналогии, в то время
как сходство между человеком и антропоидами не только
относится к
категории гомологии,
но и объяснимо
лишь филогенетическим родством. Многочисленные
палеонтологические и палеоантропологические факты доказывают правильность
дарвиновской гипотезы антропогенеза.
Мнение Дарвина о дриопитеке как об общем миоценовом
предке гориллы, шимпанзе и человека выдержало проверку
фактами
почти на протяжении
столетия. Более
того, открытие рамапитека и австралопитекоидных
африканских человекообразных обезьян пролило свет
и на вопрос о
морфологических особенностях
плиоценовых антропоидов, близких к человеческой
родословной. Открытие зубов ископаемого
антропоида удабнопитека в Грузии (1939), сделанное
советскими исследователями Н. О. Бурчак-Абрамовичем
и Е. Г. Габашвили,
подтверждает широкое
географическое распространение и большое разнообразие
формы и строения близких к человеку
высших обезьян. Наконец, новейшие находки костных
остатков гигантских ископаемых плиоценовых и плейстоценовых
антропоидов, например
гигантопитека в Южном
Китае (1935) и мегантропа на острове Ява (1943),
свидетельствуют
о том, что антропоиды, близкие к предкам человека,
были распространены и на юго-востоке Азии.
Таким образом, нет сомнения, что в неогене было
много форм антропоидов, из которых довольно многие
были
близки к позднее
развившимся
первым людям —
питекантропам. Идея о возникновении древнейших
представителей человеческого рода из круга именно
подобных форм
получает ныне достаточно крепкую
основу. Отсюда, конечно, еще не следует, что можно
признать нашими предками гигантопитека
или мегантропа, как это предлагает Франц Вейденрейх
(Weidenreich, 1945) в своей гигантоидной гипотезе
антропогенеза. Он,
как и Осборн, стоит
на позициях
ортогенеза в эволюции приматов и человека в противовес
материалистическому учению дарвинизма.
Рассматривая ископаемых антропоидов и гоминид,
Осборн стремится отвести из человеческой родословной
древнейших
людей и
неандертальцев: он
ведет линию
от эоантропов прямо к типу современного человека,
древность которого резко преувеличивает1. Основа
теории Осборна
в виде костных
остатков даусоновского
эоантропа в Англии, найденных с 1908 по 1915 г.,
ныне лишена всякой научной достоверности
1 Изложение и критика гипотезы
Осборна основаны на работах автора (М. Ф. Нестурх). Гипотеза
антропогенеза
Осборна
и ее критика.—
Успехи современной
биологии, 1940. т. XIII, вып. 2, стр. 347—353;
Ортогенетическая
гипотеза Осборна о происхождении человека и ее
критика.— Естествознание в
школе, 1948,
№ 3, стр. 3—7. См. также некролог, посвященный
Г. Ф. Осборну и развенчивающий его реакционные
идеи
в науке
о человеке.—
Антропол. журнал, 1936,
стр. 371—375.
117
вследствие новейших сведений, по которым эти
находки были частично подброшены: так,
нижняя челюсть оказалась
принадлежащей
современному
шимпанзе и была
искусственно подкрашена. Таким образом,
находка эоантропа, бывшая для советских антропологов
всегда очень сомнительной,
является
фальшивкой (Рогинский, 1951,
стр. 153—204; Гремяцкий, 1954, стр. 154—157).
Осборн строит свои предположения об извечном
мастерстве предков человека на утверждениях
о существовании
третичных эолитов.
Но упоминаемые
им эолиты из Ред-Крега вовсе не являются
столь древними, как он полагает.
Кроме того,
очень трудно бывает иногда отличить искусственно
изготовленное каменное орудие от камня,
образовавшегося путем обивки
и ударов о другие камни
где-нибудь
в каменных осыпях или в бурных водоворотах.
Однако весьма возможно, что некоторые эолиты
четвертичного
периода
обязаны своей формой
первым людям, которые наряду
с искусственными орудиями, вероятно, еще
долгое время пользовались и необработанными
камнями,
выбирая из
них наиболее подходящие
и лишь слегка их подправляя.
Нельзя отвергать все четвертичные эолиты
только на том основании, что камни, похожие
на них
по форме,
известны
также и из миоценовой
эпохи,
так как возможно,
что первые искусственные орудия вообще едва
ли были отличимы от природных камней.
Орудия китайского древнейшего человека, синантропа,
свидетельствуют о том, что им должны
были предшествовать еще более простые орудия,
среди которых могли быть и эолиты.
Но целью Осборна было с помощью легенды об
«извечности» древнего человека-мастера так
или иначе примирить
науку с религией
именно в проблеме антропогенеза.
Не удивительно, что подобные «научные» построения
встречают сочувственное отношение среди духовенства.
Можно было бы удивляться тому, что такой
крупный ученый в области геологии, палеонтологии
и
общей биологии,
как Генри
Осборн,
широко известный
также своей книгой о среде, жизни и искусстве
человека древнего каменного века
(Осборн, 1924), был всю жизнь и верующим.
Но подобное противоречие при господстве в
США
религиозного духа фундаментализма достаточно
объяснимо.
Для богословов самая главная задача — не
допускать признания того, что человек развился
естественным
путем из своих
предшественников — обезьян, так или
иначе отгородить человека от всего остального
животного мира, даже
от таких форм, которые, собственно говоря,
по телесному строению уже являются людьми.
Все это для того, чтобы удержать догмат
о бессмертии души, которую бог якобы вдохнул
в первого человека
при чуде творения.
Заодно
богословы поддерживают
антинаучные тенденции, чтобы доказать,
что тип современного человека гораздо древнее,
чем даже
питекантроп, и
что, следовательно, люди не произошли от
обезьян. А раз так, то облегчается возможность
объявить тип современного
человека чрезвычайно
118
древним и появившимся независимо от животного
мира, т. е. путем чуда. Но все подобные
попытки с каждым
новым
открытием делаются
все более
несостоятельными.
Стараясь придать известную наукоподобность
своим писаниям, богословы всячески
используют работы
реакционных буржуазных
биологов и
других ученых. В качестве
доводов против представления об эволюции
как процессе, совершающемся исключительно
но законам
природы,
богословы используют всякие
гипотезы идеалистически настроенных
буржуазных ученых, развивающих идею
о том, что эволюция есть лишь развертывание
божественного
плана творения.
Особенно удобны для богословов гипотезы,
по которым эволюция основывается на
вечном перекомбинировании
одних и тех
же неизменных наследственных
частиц, генов. В качестве яркого представителя
такого направления в буржуазной билогии
можно назвать Яна Лотси (Lotsy, 1927),
антиэволюционное учение об организмах
как комбинациях генов
он
распространяет и на
человека. Механистичность
концепции Лотси, родственной аристогенезу
Осборна с его мозаикой генов, обнаруживается
из следующего вывода: «Важно то, что
даже если
виды постоянны, то
эволюция, по меньшей мере, оказывается
вполне мыслимой. И моя гипотеза имеет
полную аналогию в неживой природе:
гены соответствуют
элементам, а хромосомы — постоянным
химическим соединениям». Гипотеза антропогенеза
Лотси превращающая человека в скопление
неизменных генов и приводящая к идеалистическим взглядам на человеческие
расы, оказывается
научно несостоятельной, противоречит
дарвиновской концепции антропогенеза,
приводит по существу
к идее неизменности
человеческого типа.
Гипотеза антропогенеза
Вейденрейха
Видный немецкий антрополог Франц Вейденрейх
на протяжении долгих лет развивал
автогенетическую идею эволюции в применении
к животному миру
и в особенности
к человеку. Обратимся к разбору
гипотезы антропогенеза Вейденрейха. По его
словам, ход человеческой
эволюции, насколько он обнаруживается
в зубной системе,
является типичным примером ортогенетического
развития, влиявшего на всякую отдельную
систему органов тела
и на все строение
тела человека по пути преобразования
в одном и том же направлении.
В согласии со своей идеалистической
концепцией эволюции Вейденрейх
утверждает, что все,
происходящее с зубной
системой, есть
лишь следствие общего изменения
тела и не может быть результатом
какого-нибудь прямого или случайного
влияния на зубы,
как, например, со
стороны перемены
способа питания
или же со стороны
отбора неопределенных случайных
вариаций. Но в свете дарвинизма
лишается силы
не только идеалистическая гипотеза
эволюции
Вейденрейха, но
и его ортогенетический взгляд на
ход
119
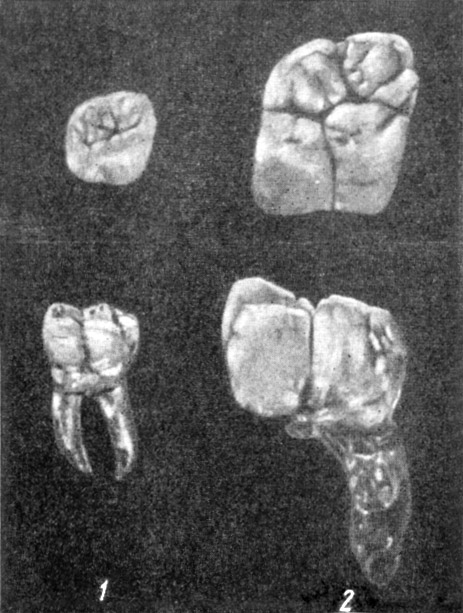
Рис. 43. Верхний и нижний моляры человека (1) и гигантопитека
(2) блэковского (Gigantopithecus blaeki Koenigswald), вид сверху
и со щечной стороны (по Г. Г. Р. Кенигсвальду, 1947, 6/5 нат. вел.).
По Э. Влчеку, 1965 (ширина изображения 59 мм)
|
эволюции физического типа гоминид
и их предков.
Вейденрейх пробует доказать,
что изменения телесной организации
живых существ
носят самопроизвольный характер
в силу внутренней тенденции
развития. Подобное
реакционное воззрение есть
не что
иное, как возвращение
к учению о жизненной силе или
попытка протянуть руку религии,
ничего
общего не
имеющая с материалистическим
учением дарвинизма. Мы уже
видели, что
перемена способа питания
и внешние факторы оказали реальное
влияние на эволюцию черепа
и зубов приматов.
Предложенная в 1945 г. Францем
Вейденрейхом гигантоидная гипотеза
происхождения
человека является, казалось
бы, наиболее солидно
аргументированной из идеалистических
гипотез антропогенеза. Ее фактическую
основу составляют
находки остатков
ископаемых огромных антропоидов
в юго-восточной Азии. Вейденрейх
принимает
их за гоминид, древнейших представителей
человечества. Основная идея
Вейденрейха заключается в том,
что
гигантизм есть характерная
особенность
ближайших предков
человека.
На первое место Вейденрейх
ставит гигантопитека, который
назван
блэковским в честь английского
анатома Дэвидсона
Блэ-ка, известного
по исследованиям
синантропов. Новый род ископаемых
антропоидов был установлен
в 1935
г. Г. Г. Р. Кенигсвальдом
по одному
сильно стертому
нижнему правому
последнему
коренному зубу (рис. 43). Интересна
история происхождения этого
моляра. Кенигсвальд
не нашел его в земле,
а приобрел в одной
из
китайских
аптек в г. Гонконге,
в которых он купил около 1500
зубов ископаемых орангутанов.
Судя по
зубам и костям других
ископаемых животных,
вместе с которыми
зубы поступили
в продажу
для изготовления лекарств либо
в качестве амулетов, зубы орангутанов
происходят
из пещер, находящихся
на территории
Юннаня и Гуанси
в южном Китае, и имеют
сравнительно молодой, плейстоценовый
геологический возраст. Среди
купленных зубов орангутанов
Кенигсвальду позже
удалось найти
еще два больших
коренных зуба гигантопитека
(последний нижний
120
левый и первый верхний правый),
а затем и еще один. Известно
уже около
тысячи
зубов
гигантопитеков
(Дун Ти-чен, 1963).
Длина коронки нижних коренных
зубов гигантопитека составляет
22 и 22,3
мм, у гориллы —
18—19,1 мм, у современного
человека — 10,7
мм,
т. е.
почти в
два раза меньше. Объем же коронки
у гигантопитека в среднем равен
4420 мм3, у гориллы
— 2356, т. е. почти
в два раза
меньше. А
так как у человека
объем
коронки третьего нижнего моляра
составляет всего лишь 723 мм3,
то такой же зуб
гигантопитека оказывается
по объему
в шесть
раз крупнее
человеческого.
Перейдем теперь к другим особенностям
формы и строения зубов гигантопитека
и укажем на
те,
которые, по
нашему мнению,
резко противоречат утверждению
Вейденрейха о человеческой
природе южноазиатских древних
великанов
обезьяньего мира. Что
касается пропорций
коронки, то оказывается,
что задний отдел нижнего
моляра, талонид, или пятка,
по ширине уступает переднему
отделу,
т. е.
тригониду, имевшему
у предков обезьян
треугольную форму
расположения бугорков: параконида
(исчезнувшего у большей части
приматов), протоконида и метаконида.
В таком
важном пункте гигантопитек
явно близок
к антропоидам,
а не к гоминидам, у которых
талонид, как правило, шире
тригонида. О том же свидетельствуют
и другие признаки,
например наличие
добавочных
бугорков на
жевательной поверхности
нижних моляров. Тут особо отметим
присутствие так
называемого шестого бугорка,
располагающегося на задне-внутреннем
крае между энтоконидом и гипоконулидом
и столь
характерного для дриопитеков,
с
которыми гигантопитека
роднит также
характер расположения главных
бугорков. Наконец, третьи нижние
моляры
гигантопитека
не обнаруживают
никаких
признаков редукции,
которая
как раз характерна для
последних моляров, или зубов
мудрости, у гоминид. Кенигсвальд
считает
возможным сближать
гигантопитека
даже с сивапитеками,
т. е. с группой,
более близкой
к предкам орангутанов, и такое
мнение может быть ближе к истине,
чем гипотеза
Вейденрейха
(Дун-Ти-чен,
1963).
Во всяком случае тезис Вейденрейха,
что гигантопитек — не антропоид,
а типичный
гоминид,. должен
быть совершенно отвергнут
как научно
несостоятельный.
Гигантопитек — не гоминид и
не предок человека. Это одна
из
специализированных форм антропоидов.
Огромный
череп
гигантопитека, вероятно,
был крупнее, чем у современного
человека.
Судя по найденным трем нижним
челюстям
с полным набором зубов (Пэй
Вэнь-чжун, 1957),
череп гигантопитека обладал
грубым наружным костным рельефом1.
Ог-
1 До сих пор гигантопитеки
были известны только из Китая.
Но
весною 1968 г.
американский палеонтолог
Э.
Саймонс
и индийский С. Чопра
(Simons, Chopra)
нашли в Сиваликских холмах,
к северу от Симлы, почти полную
нижнюю
челюсть гигантопитека в слоях
среднеплиоценовой древности.
Находка сделана недалеко
от области, где были обнаружены
челюсти рамапитека, и дриопитека,
живших
значительно ранее, 10—12
млн. лет
назад (фауна Нагри),
а индийский гигантопитек относится
к фауне Док-Патан, характерной
для эпохи
5—
10 млн. лет назад. Как и китайские
гигантопитеки, индийский
вряд ли достигал величины горного
гориллы, по мнению
Саймонса (Simons,
1968).
Рис. 44. Фрагмент нижней
челюсти мегантропа древнеяванского
(Meganthropus palaeojavanicus
Koenigswald) и
(на заднем плане) реконструкция
части его челюсти по
Ф. Взйденрейху (слева — челюсть человека,
справа
— челюсть гориллы)
По Г. Г. Р. Кенигсвальду,
1947, 2/7 нат. вел. (ширина изображения 117 мм)
ромная физическая сила
гигантопитеков создавала
им превосходство
в межвидовой борьбе:
обращаться к орудиям,
как оружию,
у них не было нужды.
Они охотились на животных и
уносили части
их
трупов в пещеры,
где обитали. Однако
нет
никаких данных предполагать,
что они могли
бы вступить на путь
очеловечения. Вторым
аргументом в руках
Вейденрейха является находка «фрагмента
нижней челюсти
древнеяванского мегантропа
(рис. 44), которую
один из сборщиков
Кенигсвальда
обнаружил
в 1941
г. во впадине
сбросового
происхождения в Сангиране
на острове Ява. От
челюсти сохранился
средне-передний
участок правой
половины ее тела.
По размерам фрагмента
и зубов можно судить
о том,
что ее
обладатель несомненно
был существом с очень
крупными
размерами
тела: высота
на уровне подбородочного
отверстия 48 мм,
а у гориллы
39—49
мм. Сохранились первый
коренной зуб, два
предкоренных, ячейка
клыка.
На жевательной поверхности
большого коренного
зуба меган-
122
тропа Вейденрейх констатирует
типичный узор
дриопитека; сюда присоединяется
шестой бугорок,
усугубляющий
впечатление об
обезьяньей природе
мегантропа. Жевательная
поверхность моляра
покрыта морщинками,
как и у антропоидов.
Судя по малой
глубине ячейки
клыка, его коронка
была невелика.
В согласии с таким
предположением
находится форма премоляров, напоминающих
человеческие.
А отсюда
можно сделать
вывод, что
и верхние клыки
были невелики. Один клык
был найден.
Исходя из высоты
челюсти на уровне
подбородочного
отверстия,
можно
заключить, что
по размерам тела
мегантроп уступал
только гигантопитеку
или горилле,
сближаясь с орангутаном.
Из группы гоминид
мегантропа
выводят
особенности
не только
моляров, но и
самой челюсти.
Например,
на язычной стороне
тела челюсти
в области сращения
располагается
загадочная впадина
двубрюшной мышцы
глубиной 15 мм.
Вейденрейх считает
впадину аномалией.
Тут же недалеко
располагается
указываемая Вейденрейхом
ямка
двубрюшной мышцы,
резко
отличающаяся
своей
треугольной формой
от удлиненной
плоской области
той же
ямки у человека.
Нам представляется,
что
впадина и ямка
двубрюшной мышцы
мегантропа
— это углубления
естественного
происхождения,
возникшие, возможно,
в связи с
особой специализацией
мышц и костных
участков в данной
области нижней
челюсти. Во
всяком случае
подобные морфологические
уклонения уводят
мегантропа
в сторону
от человеческой
родословной.
Далеко от человека
отстоит мегантроп
также по особенностям
контура
вертикального
разреза, проведенного
спереди
назад в срединной
(медианной) плоскости
реконструированной
челюсти. Судя
по
контуру,
мегантроп должен
быть помещен
между дриопитеком
и парантропом.
Наконец,
обратим
внимание на положение
подбородочного
отверстия у мегантропа.
Известно, что
у обезьян оно
лежит
низко,
а у людей
высоко.
Подбородочное
отверстие располагается
примерно
на уровне границы
между премолярами
и молярами, на
высоте
середины
горизонтальной
ветви тела
челюсти. У мегантропа
оно лежит на
середине высоты
тела челюсти,
т.
е. как у представителей
гоминид.
Однако нижнечелюстной
канал,
как
это видно на
поперечном разрезе
челюсти
мегантропа, идет
низко,
скорее, как у
антропоидов,
а не так
высоко,
как
у гоминид.
Таким образом,
и здесь мегантроп
проявляет
себя
больше как антропоид.
Придавая большое
значение указанным
питекоидным
особенностям
по сравнению
с гоминоид-ными,
которые у мегантропа,
по-видимому,
тоже
обнаруживаются,
хотя и в слабой
степени, мы считаем,
что мегантроп
скорее
антропоид,
чем гоминид.
Благодаря же
обладанию уникальными
чертами специализации
и крупными
размерами тела
мегантроп не
мог быть предком
человека: он должен быть
исключен из человеческой
родословной
(см.,
однако:
Гремяцкий,
1966).
Гигантизм в отряде
приматов представляет
собой исключение,
а не правило:
в Индии жил ги-
123
гантский дриопитек,
на юго-востоке
Азии обитали
гигантопитек
и мегантроп,
в восточной
Африке
— большой
проконсул, на острове
Мадагаскар жили
колоссальные
лемуры —
мегаладапис и
другие, но
и только. Гораздо
больше
известно
ископаемых
антропоидов с прегоминоидными
особенностями,
и невозможно
согласиться
с Вейденрейхом,
что будто
бы нашими
предками
были великаны
вроде каких-нибудь
мифических
гигантов
Адама и Евы. Гипотеза
Вейденрейха
не
может ответить
на такой
вопрос: как согласовать
с автогенетической
концепцией
развития
животного
мира предполагаемый
ею поворот
процесса эволюции вспять
с измельчанием
гигантских
антропоидов
и их превращением
в людей,
сильно уступавших
по размерам тела
своим
предкам?
В своей научно несостоятельной
гигантоидной
гипотезе
антропогенеза Вейденрейх,
если можно так выразиться,
извратил
свою
же собственную
ортогенетическую
концепцию.
Идея
Вейденрейха
несостоятельна
еще и потому,
что геологическая
древность
мегантропа
и гигантопитека
не
так велика,
чтобы ставить
их в предки
гоминидам (Нестурх,
1954,
стр. 29-46).
Некоторые
факторы гоминизации
и вымирания
ископаемых
антропоидов
плиоцена
и плейстоцена1
Всех крупных
человекообразных
обезьян
миоцена
и плиоцена,
включая
предков
гоминид,
следует
включать в семейство
понгид
(Pongidae).
Австралопитеки
и близкие
к ним формы
тоже относятся
к понгидам.
Распространять
на них
термин
гоминиды
(Hominidae)
неправильно,
так как
люди, в
собственном
смысле
этого слова,
качественно
отличны
даже
от наиболее
им близкородственных
животных.
Общее филогенетическое
родство
между гоминидами
и понгидами
заставляет
биологов
искать
специфические
черты сходства
с первыми
в структуре
черепа,
зубов,
посткраниального
скелета
у
разных
неогеновых,
в частности
у плиоценовых,
крупных
антропоидов.
Многие
антропологи
и палеонтологи
применяют
термин
«гоминизация»
к процессу
накопления
подобных
признаков
у
некоторых
крупных
антропоидов
верхней
половины
третичного
периода,
хотя
скорее
такое его
понимание
было
бы более
подходящим
к превращению
обезьяны
в человека
в
ходе качественно
особого
процесса
антропогенеза
от архантропов
до неоантропов.
Гоминизация,
если ее
применять
к
предчеловеческому
этапу
антропогенеза,
происходила
прежде
всего в
ходе эволюции
довольно
многих
видов ископаемых
ортоградных
антропоидов
плиоцена
и плейстоцена,
очутившихся
вместо
привычного
леса в
условиях
более или
менее
открытой
равнинной
или скалистой
местности.
Однако
некоторые
гоминоидные
особенности
проявлялись
и у других
антропоидов
независимо
от характера
их
образа
жизни.
Здесь
примером
может служить
нижнеплиоценовый
рамапитек
коротко-
1 Нестурх, 1964.
124
мордый
(Ramapithecus
brevirostris)
с его
параболоидной зубной
дугой.
С другой
стороны,
сходством
с
гоминидами
по
широкой форме
таза обладает
ореопитек
бамболиев
(Oreopithecus
bambolii),
который,
однако,
вряд
ли
вел наземный
образ
жизни.
Для большинства
случаев
расширение
костного
таза
и формирование
других
черт
адаптации
к жизни
на
открытой местности
в связи
с
развитием
ортоградного
типа
локомоции
свидетельствует
о том,
что
двуногое хождение
послужило
первоначальным
основным
фактором
прогрессивного
развития
на
первых ступенях
гоминизации
у некоторых
верхнемиоценовых
и
нижнеплиоценовых
антропоидов
(Chiarelli,
1968).
Если
под
гоминизацией
—
а это
будет
шире
и
правильнее — подразумевать
приближающие
к
типу
гоминид
анатомо-физиологические
адаптационные
преобразования
организма,
включая
сюда
коррелятивно
связанные
изменения
в
области высших
нервных
функций
и
в коре
больших
полушарий
предков
человека,
то
выявляется
большая
сложность
и
вариабильность этого
важного
процесса.
Таков
был
ход
эволюции
довольно
многих
высших
приматов
на
протяжении
последних
геологических
эпох
вплоть
до
начала
четвертичного
периода.
Здесь
нужно
подчеркнуть
материалистическую
сущность
всех
факторов
гоминизации.
Характер
и
результаты
взаимоотношений
организма
человеческих
предков
на
уровне
двуногих
антропоидов
с
окружающей
природной
средой
и
в его
итоговых
преобразованиях
на
путях
онто-
и
филогении противоречат
телефинализму
и
другим идеалистическим
концепциям
эволюции
животного
мира
и
процесса антропогенеза.
В неогеновые
эпохи
семейство
крупных
антропоидов
испытывало
интенсивное
развитие.
Адаптация
к
жизни в условиях
леса
и
открытой местности
вела
к
дальнейшей
дифференциации
понгид,
число
их
видов
и
родов
было
велико
(Гремяцкий,
1961).
Тогда
намечались
три
главных
пути
овладения
понгидами
средой:
по
линиям
образа
жизни
на
деревьях
—
дриопитеки
и
проконсулы,
древесно-наземного
—
ореопитеки
наземного
—
предки
австралопитеков
и
гоминид.
Если
считать,
что
первичным
для
предков
людей
был
древесный
образ
жизни
либо
что
он,
как
и наземный,
возник
из
смешанного
типа
древесно-наземного
существования,
какое
приписывают
африканскому
проконсулу
(Proconsul
africanus),
то
так
или
иначе
следы
прежней
приспособленности
тела
к лазанью
по
деревьям
с обхватыванием
ветвей
конечностями
в достаточной
мере
проявляются
в
строении
таких
наземных
приматов,
как
австралопитеки
и гоминиды
(Campbell,
1967).
Можно
полагать,
что
ортоградный
тип
локомоции
мог
возникнуть
также
у
тех крупных
древесных
антропоидов,
которые
еще
не
испытали
особой
специализации
по
линии
брахиирования
благодаря
переходу
к
семиортоградной или
ортоградной
круриации
по
нижним
ветвям
деревьев,
где
они
находили
себе
убежище
от
хищных
зверей.
Тип
локомоции
на
двух
ногах
появился
во
всех
трех
названных
125
выше
группах крупных
антропоидов, вероятно,
довольно независимо
от пропорций
передних и задних
конечностей, о чем
можно судить
по современным
крупным антропоидам
и по
примеру малых,
т. е.
гиббонов, или
гилобатид (Walker,
1968).
Однако
при передвижении
по земле
на двух
конечностях гиббоны
осуществляют тип
локомоции, несколько
отличный от
того, который,
надо полагать,
был свойствен
наземным ископаемым
антропоидам с более
благоприятным интермембральным
индексом
и более каудально
располагавшимся центром
тяжести, а также
носил в потенции
более прогрессивный
характер.
В
чем именно
лежала основная
причина, каков
был главный
фактор возникновения
ортоградности в
разных —
первоначально, возможно,
еще и семиортоградных
— видах крупных
наземных антропоидов?
Надо полагать,
что основным
фактором здесь
служила их
борьба против
новых врагов
в виде
многих хищных
зверей и птиц.
При
более высоком
положении головы
бинокулярное и цветное
зрение способствовало
большей безопасности
и сохранности стад
выпрямлявшихся наземных
антропоидов, а в
случае нападения—
инстинктивному употреблению
камней, палок,
костей, раковин
для активной
защиты с помощью
опосредованных оборонительных
и наступательных
действий.
К
развитию прямохождения
у ископаемых наземных
антропоидов послужили
и другие факторы,
возможно, в
связи с питанием,
когда, например
в условиях травянистых
равнин с рощами,
перелесками и кустарниками,
обезьяны могли
бы срывать
добавочные ягоды,
побеги и плоды,
стоя на
двух ногах,
если судить
по аналогии
с горными гориллами.
Вспомним здесь
о знаменитой африканской
антилопе генеруке
(Litocrahius walleri),
у которой в связи
с подобным способом
добывания пищи
в позвоночнике появился
поясничный изгиб.
Надо
также учитывать,
что самки
обезьян, беременные
или с малыми
детенышами, вероятно,
должны были
нередко поддерживать
равновесие на
двух ногах,
скорее, в
несколько более
выпрямленном положении
тела.
Трудно
допустить, чтобы
все виды
подобных ископаемых
антропоидов с еще
слабо развитым
прямохождением обратились
к постоянному, а не
только к спорадическому
использованию природных
предметов в качестве
орудий защиты
или нападения.
Важнее было
для некоторых
видов семиортоградных
или ортоградных
высших обезьян
овладение способами
применения камней
и других
подходящих предметов
к ежедневному и даже
целодневному систематическому
добыванию
всевозможной растительной
и животной пищи.
Частичную
аналогию подобной
беспрерывной деятельности
можно найти
у павианов, причем
их ногти
приобрели когтевидную
форму в связи
с обитанием
в скалистой местности
и частым
переворачиванием камней
для поимки
прячущихся там
червей и жуков.
Правда, имея
мощные клыки
и обладая большой
силой, павианы
камнями животных
не убивают.
126
Но
яванские макаки-крабоеды
ежедневно применяют
камешки для
разбивания панцирей
ракообразных и моллюсков,
хотя мозг
этих мелких
и слабых обезьян
далеко уступает
по своим
размерам мозгу
павианов и
тем более
антропоидов.
В
сфере постоянного
удовлетворения насущнейшей
жизненной потребности
— голода — следует
искать тот
узловой пункт,
который привел
к сочетанию двуногой
походки и употребления
предметов окружающей
природы как
орудий. А это
влекло накопление
в организме
разнообразных черт
в виде своеобразных
анатомо-физиологических
предпосылок
очеловечения
обезьяны в
ходе антропогенеза.
Действительно,
в то время
как у одних
видов прямохождение
требовало общего
повышения уровня
высшей нервной
деятельности в
связи с обостренной
борьбой за
выживание, но
без обязательного
применения опосредованных
действий, у других
видов примешивался
новый фактор
в виде
систематического употребления
орудийных
предметов, что,
безусловно, должно
было поставить
этих двуногих
высших обезьян
«на лестнице
эволюции» выше
первых.
Предтрудовая
орудийная деятельность
и оказалась
тем важнейшим
стимулом, который
повысил жизненную
устойчивость ею
овладевших видов
и придал новую
биологическую силу
плохо защищенным
от природы
обезьянам —
без мощных
острых клыков,
когтей и прочего
специального природного
вооружения.
Там,
где камней
кругом было
много, обезьяны
могли их
и не таскать
с собой в руках
или под
мышками, как
это делали
дикие шимпанзе
В. Кёлера.
В других местностях
обезьяны должны
были носить
камни с собой
либо чаще
обращаться к палкам.
А всякие дубинки
и сучья,
вероятно, использовались
нашими предками
в достаточной мере,
чтобы как-то
оправдывать догадки
ученых о «деревянном
веке древнейшей
культуры».
Освоение
предтрудовой орудийной
деятельности означает
появление новых
и новых условных
рефлексов, усложнение
высшей нервной
деятельности, разрастание
структур головного
мозга и обогащение
нервных связей,
особенно между
глазами и руками,
совместно прилаживавшимися
к новой интенсивной
трудовой деятельности.
Таким
образом, нервно-психический
фактор начинает
у предков человека
постепенно выходить
на первый
план и интенсивно
влиять через
анатомо-физиологические
механизмы
на
формирование многих
черт гоминизации,
не связанных
непосредственно с прямохождением,
хотя
в известной степени
этот важный
фактор способствовал
также и
дальнейшему прогрессивному
развитию двуногой
локомоции.
Как
же понимать
движущие причины
гоминизации, лежащие
в окружающей природе?
Сюда относятся
прежде всего
недостаток пищевых
ресурсов в условиях
открытой, нередко
скалистой или
пустынно-степной местности,
а с другой
стороны, избыток
хищных животных.
Оба эти
обстоятельства в первую
очередь и
вынудили неогеновых
наземных антропоидов
127
обратиться
к прямохождению, а
многих
из них
— также к пользованию
камнями и другими
предметами как
орудиями, т.
е. к орудийной
деятельности.
Орудийная
деятельность неогеновых
антропоидов при
добывании пищи,
защите и нападении
может быть
в широком смысле
слова обозначена
как предтрудовая,
хотя такой
термин в узком
смысле следовало
бы относить
только к предкам
гоминид.
Понятие
об орудийной
деятельности можно,
однако, применять
:и к другим
обезьянам, использующим
предметы природы
при добывании
пищи. В этом
отношении очень
интересны новейшие
длительные наблюдения
над шимпанзе
в восточной Африке,
которые проделала
Джейн Ловик-Гудолл
(Jane Lawick-Goodall,
1967). Она
не раз
видела и
фотографировала
диких
шимпанзе, когда
они добывали
термитов на
тростинку или
стебель растения.
Вырвав из
земли подходящий
стебелек, обезьяна
обрывала лишние,
мешающие листочки
и, если
нужно, укорачивала
его; после
этого шла
к термитному гнезду,
всовывала стебелек
в его
входное отверстие
и начинала
скрести около
него пальцем.
Встревоженные термиты
вылезали на
стебелек и для
шимпанзе оставалось
лишь провести
им вдоль
приоткрытого рта,
чтобы слизнуть
и съесть эту
живую пищу.
Примеры
применения предметов
в качестве орудий
известны также
среди птиц.
Так, на
Галапагосских островах
вьюрки, не
будучи в состоянии
пробить отверстие
в каком-нибудь дереве
своими клювами,
отламывают очень
твердые
шипы и,
продолбив с
их помощью
кору, добывают
себе жуков
и их личинок
(Eibe Eibesfeldt,
1963).
Новая
для обезьян
и малоблагоприятная
для
них среда
открытой местности
создавала для
этих медленно
размножающихся
приматов
совершенно реальную
угрозу быстрого
или затяжного
вымирания, так
как они
попали, несомненно,
в экологические
условия
не столько
нормы, сколько
близкие к пессимуму.
Следовательно,
для наземных
крупных антропоидов
началась длительная
и довольно безнадежная
битва за
выживание в неблагоприятных
условиях среди
открытой местности,
шла нелегкая
адаптация к
ним. Эти
довольно смышленые
животные даже
обратились к
активной
орудийной деятельности
в заметных масштабах,
если говорить
о количестве
овладевших
ею видов
наземных антропоидов.
Такая
форма приспособления
и в подобных
размерах составила
качественно особое
событие во
всей длиннейшей
истории животного
мира.
Какие
минусы общебиологического
характера были
у неогеновых
наземных
антропоидов?
Сюда
кроме черт
природной беззащитности,
а также медленности
бега на
двух ногах
надо отнести:
слабые темпы
естественного
прироста
вследствие длительной
беременности,
вероятно
со сроками
от 7 до
10 лунных
месяцев; появление
на свет,
как правило,
лишь одного
детеныша, как
и при
жизни на
деревьях; долгую
лактацию на
протяжении 1—2
лет; продолжительное
младенчество
до
полугода;
128
многолетнее
формирование
организма
с наступлением полового
созревания к
8—12
годам и позже
как у самцов
современных горных
горилл (Merfield,
1954).
Хотя
стада наземных
антропоидов
в
условиях открытого
ландшафта и могли
бы несколько
укрупниться,
общая
численность
особей,
как и у
других тропических
или подтропических
видов этих
приматов, вероятно,
оставалась
примерно
той же,
т. е.
не очень
значительной.
Но
и при
жизни в несколько
более крупных
группах (семейного
или стадного
типа) антропоиды,
наверное, представляли
собой добычу,
довольно подходящую
по размерам
тела. И овладение
их мясом,
жиром и внутренностями
вряд ли
составляло
для
такого умного,
сильного и ловкого
зверя, как
леопард, особенно
трудную задачу.
В
условиях
леса
обезьяны
гораздо
легче могут
спастись
от
хищника,
который
там довольствуется
одной жертвой
вместо
нескольких
на открытой
местности.
Ведущие
наземный
образ
жизни горные
гориллы
подвергаются
неустанным
преследованиям
со стороны
леопардов,
которые
иногда
утаскивают
их детенышей
и сокращают
численность
этих гигантов.
Надо
полагать,
что
при всей
генерализованности
и пластичности
организма
наземных антропоидов
неогена, особенно
же в плейстоценовую
эпоху, их
прогрессивно
повышавшийся
уровень приспособленности
к экологическим
и иным
условиям внешней
среды продолжал
колебаться
между
нормой и
пессимумом,
с
уклоном к
последнему.
Этим
и объясняется
заметное
поредение
родов
наземных высших
обезьян с
вымиранием
довольно многих
видов к концу
третичного
периода.
Правда, погибли
и разные антропоиды,
обитавшие
в лесах,
но все
же от
них осталось
поныне пять
родов, а
наземные в
результате
вымерли все
к середине
плейстоцена
и позже.
Следовательно,
открытая
местность
оказалась
для привычных
лесных обитателей
— обезьян
— труднопреодолимой
новой
средой. Не
надо забывать,
что ни
одна низшая
или тем
более высшая
обезьяна не
перешла к рытью
нор или
других подземных
убежищ либо
к жизни
в пещерах,
как
это сделали,
возможно,
лишь
один-два вида
австралопитеков.
И
даже орудийная
деятельность,
которая
смогла
приобрести
более систематический
характер,
вероятно,
только
у
нескольких
видов
наземных
двуногих
антропоидов,
не
оказалась
панацеей
от совокупности
тех зол,
какие таит
в себе
для
обезьян
наземная
среда,
кишащая
хищниками,
всякими
ядовитыми
животными
и множеством
внешних
и внутренних
опасных паразитов.
Изощрение
нервно-психических
особенностей
с наибольшей
силой проявилось
среди позднеплиоценовых
предков группы
австралопитековых,
некогда
широко распространенной
на материке
Африки примерно
к югу от
30° северной
широты и почти
до 30°
южной, а
в Азии — до
0°. Именно
среди позднеплиоценовых
австралопитеков
должна
была интенсивнее
развиваться
орудийная
деятельность.
Максимальной
высоты и сложности
предтрудовая
деятельность
достигла у
предкового
вида гоми-
129
нид,
который
далеко
превосходил
все
прочие смышленостью
и приспособляемостью,
прогрессивным
развитием
головного
мозга
и ловкостью
рук,
а также более
крупными
размерами
и весом
тела. Именно
этот вид
сумел перейти
рубиконы мозга,
мышления
и
труда,
оказавшиеся
недоступными
для других
австралопитеков,
которые
представляли
собой
неудавшиеся
попытки
животного
мира
вступить на
гораздо
более
трудный путь
истинной
гоминизации,
или подлинного
очеловечения,
обезьяны
под решающим
влиянием трудовой
деятельности.
Австралопитеки,
пользовавшиеся
природными
орудиями,
продолжали
развиваться
в плейстоценовую
эпоху,
но, очевидно,
мало
прогрессировали
в своей
предтрудовой
деятельности.
Она
стала терять
свою
спасительную
роль при
ухудшении
климатических
условий
и
появлении
новых
хищников.
Не
выдержали
они
борьбы
и
против
резко
прогрессировавшего
вооруженного
конкурента,
каким
оказался
древнейший
человек,
а затем
и древний.
Формировавшихся
гоминид,
которые
изготовляли
эффективные
искусственные
орудия
и
постоянно
охотились
на
животных
из-за
мяса,
жира
и
костного
мозга,
можно
было
бы
трактовать
как
новые
и
необычайно
хищные
существа,
принесшие
опаснейшую
угрозу
существованию
многих
видов
животных,
в
том
числе
и
обезьян.
Можно
вполне
допустить,
что
некоторые
виды
австралопитеков,
включая
парантропов
и
мегантропов
с
более
крупными
размерами
тела,
после
долгого
и
тягостного
существования
на
протяжении
сотен
тысяч
лет
виллафранкской
и
дальнейших
плейстоценовых
эпох
все-таки
все
были
истреблены
не
только
кошачьими
и
собачьими
хищниками,
но
и
гоминидами.
Одним
из
факторов
вымирания
верхнеплейстоценовых
крупных
наземных
двуногих
антропоидов
была,
таким
образом,
охотничья
деятельность
первобытных
человеческих
стад.
Вероятно,
древним
гоминидам
можно
приписать
и исчезновение
таких
крупных
и сильных
древних
азиатских
антропоидов,
как,
например,
гигантопитеков
и орангутанов
на
территории
южного
Китая,
а также
сравнительно
более
мелких
орангутанов
и достаточно
крупных
мегантропов
на
острове
Ява.
Конечно,
проблема
факторов
вымирания
и
гоминизации
крупных
антропоидов
плиоцена
и плейстоцена
гораздо
сложнее,
чем
это
нами
обрисовано.
Кроме
того,
наш
анализ
коснулся
преимущественно
наземных
форм
высших
обезьян.
Но
и здесь
остается
очень
много
неясностей.
Поэтому
проблема
факторов
эволюции
верхнетретичных
антропоидов
будет
еще
долго
занимать
напряженное
внимание
приматологов,
антропологов
и других
биологов.
Лишь
накопление
гораздо
большего
количества
палеонтологической
документации,
прежде
всего
в виде
костных
остатков
этих
интереснейших
наивысших
приматов,
может
привести
к более
твердым
выводам
о путях
и тупиках
эволюции
антропоидов.
В
конце
разбора
некоторых
антидарвинистических
гипотез
происхождения
человека
можно
вы-
130
сказать
предположение
о
том,
что
верхнетретичных
антропоидов
следует
для
удобства
изучения
разделить
на
две
группы.
В
одну
группу
должны
войти
предшественники
гоминид
и морфологически
близкие
к ним
формы
человекообразных
обезьян:
это
была
бы
группа
«прегоминоидных
антропоидов».
К
ним
следует
причислить,
например,
дарвиновского
дриопитека,
ореопитека,
рамапитека,
австралопитека,
парантропа,
плезиантропа.
Другие
антропоиды
миоцена
и
плиоцена
отходят
тогда
в группу
«пресимиоидных
антропоидов».
К
ним
относятся
также
предки
современных
гиббона,
орангутана,
шимпанзе
и
гориллы.
Родов
и видов
верхнетретичных
антропоидов
было
гораздо
больше,
чем
современных,
и они
были
расселены
гораздо
шире.
Подобная
рабочая
гипотеза
позволяет сосредоточить
особое
внимание
на
прегоминоидных
антропоидах
и
может
помочь
точнее
определить
ближайшего
предка
человека.
Вероятнее
всего,
таким
предком
был
родственный
австралопитекам
верхнеплиоценовый
антропоид
южной
Азии,
где
и
надо
искать
его
костные
остатки.
Там
скорее
всего
была
прародина
человечества.
В
некоторой
степени
в
пользу
такого
мнения
свидетельствуют
находки
остатков
ископаемых
антропоидов
в
Индостане,
Китае,
Индокитае
и
на
острове
Ява.
Из
южной
Азии
древнейшие
люди
стали
расселяться
в
окружающие
области:
перед
ними
открылись
новые
возможности
освоения
иных
ареалов,
благодаря
сплоченности
первобытного
стада
людей,
коллективной
трудовой
деятельности,
орудиям.
Антидарвинистические
гипотезы
антропогенеза терпели
и
терпят
поражение
в
свете
антропологических
и
приматологических,
а
также
общебиологических
данных.
Дарвиновское
учение
об
антропогенезе
уже
давно
легло
в
основу
разработки
проблемы
происхождения
человека.
Им
воспользовался
Энгельс
при
создании
фундамента
новой,
марксистской
теории
антропогенеза,
основывающейся
на
методах
диалектического
и
исторического
материализма.
В
следующей
части
книги
делается
попытка
ближе
осветить
некоторые
стороны
того
переломного
периода
в
истории
развития
животного
мира,
когда
возникал
человек.
Но
уже
здесь
представляется
целесообразным
высказать
несколько
общих
соображений
по
вопросу
о
разделении
человеческой
родословной
и
всего
процесса
антропогенеза
на
две
неравные
части.
Первая,
необычайно
длительная,
от
первичных
микроскопических
маленьких
живых
существ
и
до
ближайших
предшественников
питекантропов,
тянулась
чуть
ли
не
два
миллиарда
лет.
Весь
ход
развития
предков
человека зависел
от
беспрерывно
менявшихся
реакций
целостного
организма
того
или
иного
предка
на
изменение
внешних
условий
среды,
под
влиянием
одних
только
природных,
в
том
числе
биологических
факторов,
начиная
с
естественного
отбора.
В
известном,
самом
широком,
даже
философском
смысле,
в
ходе
развития
животного
мира,
вернее,
той
линии,
которая
в
конце
131
концов
привела
к
питекантропам,
шли
беспрерывные
количественные
накопления
новых
качеств.
Имела
место
все
время
такая
эволюционная
изменчивость,
которая
в
предковом
виде
для
гоминид
неизбежно
создала
еще
одно
новое
качество,
дотоле
и
потом
более
не
появлявшееся
в
других
подразделениях
класса
млекопитающих.
Это
совершенно
особое
качество
характеризует
новое
существо
—
наидревнейшего
человека.
Благодаря
труду
в
смысле
регулярного
изготовления
орудий
и
их
использования
в
практике
жизни
совместно
с
другими
сочленами
первобытнейшего
стада
возникли
и
качественно
новые,
социальные
закономерности,
новые
факторы
формирования
человеческого
тела
и
человека
в
целом.
Оглавление
| Предисловие |
5
|
Часть I. Дарвиновская и другие гипотезы антропогенеза
|
7
|
| Глава первая |
Дарвин о происхождении человека |
8
|
| |
Идея антропогенеза до Дарвина
|
8
|
| |
Дарвин об эволюции животного мира
|
13
|
| |
Человеческая родословная по Дарвину
|
17
|
| |
Очерк развития знаний о приматах
|
29
|
| |
Развитие приматологии в СССР |
40
|
| Глава вторая |
Человекообразные обезьяны и их происхождение |
56
|
| |
Современные антропоиды
|
56
|
| |
Ископаемые антропоиды |
70
|
| Глава третья |
Позднейшие гипотезы происхождения человека
и их критика
|
95
|
| |
Религиозные трактовки антропогенеза
|
95
|
| |
Тарзиальная гипотеза
|
99
|
| |
Симиальные гипотезы
|
104
|
| |
Гипотеза антропогенеза Осборна
|
111
|
| |
Гипотеза антропогенеза Вейденрейха
|
119
|
| |
Некоторые факторы гоминизации и вымирания ископаемых антропоидов плиоцена
и плейстоцена |
124
|
| Часть II.Особенности строения человеческого тела
и возникновение древнейших людей |
133
|
| Глава первая |
Человек как примат |
134
|
| |
Черты приспособленности человеческого тела к прямохождению
|
134
|
| |
Характерные отличительные особенности человеческого тела, не стоящие
в непосредственной связи с прямохождением
|
140
|
| |
Специальные черты сходства между человеком и антропоидами
|
145
|
| |
Рудименты и атавизмы у человека |
148
|
| Глава вторая |
Роль труда и прямохождения в антропогенезе
|
154
|
| |
Роль труда
|
154
|
| |
Способы передвижения у человекообразных обезьян
|
157
|
| |
Вес тела в центр тяжести у человека и обезьян
|
165
|
| |
Нижние конечности
|
168
|
| |
Костный таз, позвоночник и грудная клетка
|
173
|
| |
Верхние конечности
|
180
|
| |
Пропорции тела и асимметрии
|
185
|
| |
Череп |
189
|
| Глава третья |
Головной мозг и высшая нервная деятельность
человека и обезьян
|
199
|
| |
Головной мозг и анализаторы человека и обезьян
|
199
|
| |
Развитие периферических частей анализаторов
|
210
|
| |
Высшая нервная деятельность обезьян
|
226
|
| |
Вторая сигнальная система — характерное отличие мышления человека |
246
|
| Глава четвертая |
Стадность у обезьян и зачаточные формы труда
|
249
|
| |
Стадность у обезьян
|
249
|
| |
Зачаточные формы труда
|
263
|
| |
Антропогенез и его факторы |
272
|
Часть III. Формирование человека по данным палеоантропологии
|
233
|
| Глава первая |
Первая ступень: древнейшие люди (питекантропы)
|
284
|
| |
Питекантроп острова Явы
|
284
|
| |
Синантроп
|
292
|
| |
Гейдельбергский человек |
305
|
| Глава вторая |
Вторая ступень: древние люди (палеоантропы)
|
309
|
| |
Ледниковая эпоха
|
309
|
| |
Неандертальцы и их физический тип
|
314
|
| |
Находки неандертальцев в СССР
|
322
|
| |
Палестинские неандертальцы
|
330
|
| |
Образ жизни древних людей
|
339
|
| |
Развитие мозга у ископаемых людей |
343
|
| Глава третья |
Современные люди (неоантропы)
|
356
|
| |
Человек верхнего палеолита
|
356
|
| |
Ошибочные гипотезы происхождения современного человека и их критика
|
370
|
| |
Человеческие расы
|
380
|
| |
Факторы формирования рас
|
387
|
| |
Наука против расизма |
394
|
| Summary |
|
403
|
| Приложение 1. |
Предложения по биологическим аспектам
расовой проблемы
|
406
|
| Приложение 2. |
Декларация о расе и расовых предрассудках
|
410
|
| Литература |
|
414
|