| Главная | Библиотека | Форум | Гостевая книга |
|
Глава
9
Судьба
анализаторов
В связи с глубоким погружением китообразных сразу же возникает вопрос, как
они ориентируются и ловят добычу в малопрозрачной водной толще, а иногда
и в царстве вечной тьмы, ночью и днем, при далеких миграциях, как определяют
путь и точно выходят к цели, отдаленной на тысячи километров? Каковы механизмы
этой изумительной подводной ориентации и навигации?
Каждому ясно, что в этом им помогает высокоразвитая нервная система. Деятельность
мозга невозможна без непрерывного притока информации из окружающей внешней
и внутренней среды. Этот приток поступает в организм через органы чувств
(анализаторы), которые справедливо называют «окнами в мир». Без органов чувств,
доставляющих информацию о сезонных и суточных изменениях окружающей обстановки
и о многих других внешних раздражителях, не было бы у китообразных ни великолепной
ориентировки в пространстве, ни правильных миграций, ни случаев возвращения
в одни и те же бухты после грандиозных странствий.
В новой среде понадобилось коренное переустройство всех анализаторов, полученных
в наследство от наземных предков. Одни анализаторы приобретали первостепенное
значение, другие — второстепенное, третьи — исчезали.
Редукции у китообразных подверглось обоняние — ощущение запахов посредством
органа, расположенного в носу. Сама по себе жизнь в воде еще не ведет к исчезновению
органа обоняния. Вспомним хотя бы акул, которые идут на запах крови с далеких
расстояний, и у них сильно развиты обонятельные доли переднего мозга. У рыб
орган обоняния все время развивался и приспосабливался к улавливанию запахов
в воде, а у наземных предков китообразных запахи проникали через ноздри вместе
с вдыхаемым атмосферным воздухом. С переходом в воду потомки этих предков
утратили обоняние, так как молекулы пахучих веществ могли проникать в носовой
канал лишь в момент очень короткого вдоха, после длительной дыхательной паузы,
проводимой под водой с закрытыми ноздрями. Воздух над океаном был чистым,
и запахи в нем потеряли всякое значение для живущих в воде китов. Поэтому
обонятельные луковицы мозга, обонятельные нервы и обонятельный эпителий в
решетчатораковинном отделе черепа либо полностью исчезли (зубатые киты),
либо сохранились лишь в зачаточном виде, притом лучше у зародышей (усатые
киты). Растворенные же в воде химические вещества, в том числе и запахи пищи,
кала и мочи сородичей, стали раздражителями другого органа чувств — вкуса.
Вкусу китов считали слабо развитым или даже исчезнувшим на основании тонкого
вкусового нерва и находок в их желудках несъедобных предметов (булыжники,
галька, куски дерева и многое другое). Но камни, по-видимому, заглатываются
для перетирания пищи и потому не могут свидетельствовать о слабом развитии
вкуса. Другие инородные предметы заглатываются случайно при процеживании
цедильным аппаратом всего, что попадает в сферу фильтрации, когда невозможно
отделять съедобное от несъедобного. Ученые, например Э. И. Черный и А. В.
Яблоков, полагают, что китообразные способны тонко различать разную соленость
воды и обнаруживать своих сородичей по их «химическим следам» — по моче и
фекалиям. Опресненные устья рек посещают многие морские свиньи, белухи, дельфины-соталии,
орцеллы, малые полосатики, а серые киты находят полупресные лагуны, чтобы
очищать кожу от вшей, погибающих от опресненной воды.
Далеко не все китообразные могут долго пребывать, в пресной воде. Некоторые
океанские дельфины тяжело переносят недостаточную соленость воды: они заболевают
прыщами, рожистыми воспалениями, грибковыми поражениями кожи и погибают,
как это было с косаткой Моби-Долл. Калифорнийские зоологи Сибеналер, Мельба
и Давид Колдуэллы описали, как в Мексиканском заливе афалины по нескольку
недель остаются в опресненных заливчиках, кормясь сельдевыми рыбами, которые
здесь мечут икру. В это время на коже дельфинов выступает сыпь. Соленость
Черного моря вдвое меньше, чем океана. Не этим ли объясняется отсутствие
здесь китов и некоторых видов дельфинов, встречающихся в Средиземном море?
Лишь три вида дельфинов смогли приспособиться к жизни в Черном море, обособившись
в самостоятельные подвиды: дельфин-белобочка, афалина и морская свинья. Первый
заселил зону открытого моря, вторая и третья — прибрежную зону, причем морская
свинья заняла также мелководное и наиболее опресненное Азовское море (11‰).
Черноморские дельфины, приспособившись к условиям пониженной солености (18‰),
вероятно, не выходят через проливы и не общаются со средиземноморскими популяциями,
а средиземноморские китообразные, привыкшие к океанской солености (35‰),
не заходят в Черное море.
Физиолог США Джон Лилли описал случай, как одна афалина, находясь в бассейне
его лаборатории на Вирджинских островах, пришла в сильнейшее возбуждение,
когда вблизи проплывала стайка сородичей. Ученый полагает, что дельфин узнал
о близости стайки по «вкусу» (химическому составу) той воды, которая по шлюзу
вливалась из моря в бассейн. Вкусовые ощущения, по-видимому, воспринимаются
валиковидными сосочками, расположенными по краю языка. Вообще орган вкуса
у китообразных изучен еще очень слабо. За последние годы в этой области сделаны
интересные находки.
Советский морфолог А. В. Яблоков в 1956 году нашел у корня языка белухи,
а потом и у других видов зубатых китов несколько ямок, покрытых призматическим
эпителием. Этот исследователь связал функцию ямок с хеморецепцией, специализированным
восприятием вкусовых ощущений (лишь недавно в этих ямках морфолог Л. И. Суховская
у дельфина-белобочки обнаружила и вкусовые луковицы). Московские морфологи
В. Е. Соколов и О. В. Волкова также считают, что ямки на языке дельфинов
несут сенсорно-вкусовую функцию, а обильные здесь слизистые железы выносят
из организма китообразных соли, попадающие в их пищеварительный тракт вместе
с морской водой. Сотрудник Московского университета В. Б. Кузнецов экспериментально
показал разную реакцию на разные вещества в растворе, которым промывали ротовую
полость подопытных дельфинов-белобочек и морских свиней. Ответственными за
эту хеморецепцию считают упомянутые выше ямки на языке дельфинов.
Осязание китообразных, как показывают наблюдения над живыми особями, развито
достаточно хорошо. Кожа играет важнейшую роль в восприятии раздражений, поступающих
из внешнего мира. Она не ороговевает сверху (роговой слой быстро слущивается),
и это способствует сохранению высокой кожной чувствительности. Кожные рецепторы
(тактильные, температурные, болевые) сигнализируют животному о прикосновениях
к телу, о температуре среды, о болевых ощущениях, о меняющемся давлении воды.
Водная среда с ее сравнительно стабильными окружающими условиями как будто
мало способствует развитию высокой кожной чувствительности китообразных.
Однако данные показывают другое. Киевская группа исследователей (проф. Г.
Б. Агарков с сотрудниками) нашла в коже дельфинов (особенно на голове) богатые
пучки нервных волокон с их окончаниями, сложные клубочковые рецепторы и инкапсулированные
чувствительные аппараты типа колб Краузе и фатер-пачиниевых телец. Чувствительные
окончания этих нервных структур осуществляют, по мнению ученых, трофо-, баро-
и механорецепцию и служат рецепторами-датчиками, которые выполняют прямую
и обратную связь с мозгом. Возможно, что очень высокие сосочки дермы, помимо
выполнения своей демпферной функции, предотвращающей развитие вихревых потоков,
играют также роль тактильных органов.
Редукция волосяного покрова, потовых и сальных желез не притупила у китообразных
кожной чувствительности. Смена среды, ощущаемая при их выныривании, служит
им сигналом для открывания дыхала и дыхательного акта (поэтому к лобному
выступу дельфинов, выставляющемуся из воды первым, подходит толстый чувствующий
нерв). В ответ на легкое прикосновение к коже дельфины открывают и закрывают
глаза. Хорошо иннервированная кожа китообразных способна управлять пограничным
слоем воды с помощью тонких двигательных механизмов, тесно связанных с тактильной
чувствительностью наружного покрова. Двигательные реакции кожи способствуют
сохранению обтекаемых потоков вокруг тела быстро передвигающихся дельфинов.
Высокая тактильная чувствительность их кожи смогла развиться при жизни в
условиях относительной невесомости погруженного в воду животного. Присутствие
на морде усатых китов десятков волосков, луковицы которых богато оплетены
нервными окончаниями, показывает высокую кожную чувствительность в этой части
тела. Такие волоски — своеобразные вибриссы: при прикосновении к ним они
действуют, видимо, как рычаги, увеличивающие силу внешнего раздражения на
нервные окончания. Усатые киты, наталкиваясь в воде на крошечных рачков,
легко определяют густоту скопления корма при любом освещении и часто кормятся
ночью.
Зубатые киты, питающиеся одиночной и сравнительно крупной добычей, в осязательных
волосках не нуждаются и утрачивают их еще до или вскоре после рождения. На
месте выпадения волосков остаются ямки специфического строения. Иннервация
их оказалась настолько богатой, что английские ученые Э. Палмер и Г. Уэдделл
рассматривают эти ямки как спидометры — чувствительные приборы дельфинов
для измерения их скорости движения и восприятия низкочастотных колебаний.
Лишь речные дельфины с недостаточно развитым зрением, живущие в мутной воде
и роющиеся в илистом дне, сохраняют волоски на клюве в течение всей жизни.
Китообразным во время миграций и при нырянии приходится испытывать то тепло,
то холод, и можно полагать, что они тонко ощущают температуру среды, о чем
косвенно свидетельствуют их превосходная регуляция тепла и тот факт, что
многие киты избегают тропических вод.
Кожу китообразных называют волшебной, так как с кожным раздражением связано
не только их осязание, но и исключительная быстроходность, теплорегуляция,
проявление инстинкта сохранения вида, поведение во время сна и преследования
судов, поощрения во время дрессировки и многое другое.
Очень мало известно о проприоцептивной чувствительности китообразных (раздражение
нервных окончаний при сокращении, растягивании скелетных мышц). Однако то
совершенство, с которым они двигаются, позволяет думать, что у них хорошо
развит проприоцептивный аппарат.
Зрительный анализатор — важнейший среди анализаторов у дневных наземных млекопитающих.
С его помощью человек, например, получает 80 — 85% информации из внешней
среды. Зрением воспринимается окружающий мир — величина, форма, цвет, яркость,
освещение и отдаленность предметов, от которых отраженный свет поступает
в глаз. Море, как малопрозрачная среда, не благоприятствует сильному развитию
зрения водных млекопитающих. Свет здесь быстро поглощается и уже на глубине
500 м практически наступает полная темнота.
Глаз крупных китов весит около 1 кг, а у мелких дельфинов достигает величины
глаза собаки. Глазное яблоко, спереди уплощенное, напоминает форму несколько
сдавленного шара. Щель глаза закрывается веками, лишенными ресничек. Слезные
железы отсутствуют, а поверхность глаза смазывается прозрачным и густым белковослизистым
выделением гардеровых желез. Оно предохраняет глаз от вредного действия морской
воды и не мешает прохождению лучей. Белковая оболочка (склера) — толстая
и плотная. Родятся китообразные с открытыми глазами. В воде они видят лишь
на коротком расстоянии. Раньше думали, что на воздухе они близоруки, так
как их хрусталик шаровидный, а ресничные мышцы исчезли. Однако у них оказалось
довольно острое зрение и хорошо развитый зрительный нерв. В 1 мм2 сетчатки
обнаружено у морской свиньи 200 000, у белухи — 150000, у бутылконоса — 143000
и у финвала — 62000 палочек. Колбочки в сетчатке китообразных либо отсутствуют,
либо крайне малочисленны. По этой причине считают, что китообразные не различают
цветов.
Благодаря особой отражательной оболочке, содержащей большое количество кристалликов
гуанина, глаза дельфинов могут отражать свет подобно зеркалу. Поэтому ночью
глаза у дельфинов светятся, как у кошек.
Предполагают, что аккомодация глаза китообразных достигается за счет «игры»
радужины, которая придает зрачку форму то узкой щели (при ярком свете — в
воздухе), то круглого отверстия (при тусклом освещении — в воде). В первом
случае свет, пропускаемый через узкую, но высокую щель зрачка, проходит словно
через двояковогнутые очки, так как кривизна и толщина роговицы бывает наименьшей
в центре и наибольшей у краев. Во втором случае свет, пропускаемый через
круглое отверстие радужины, проходит через центральную (тонкую) часть роговицы
и минует ее толстую часть. Кроме того, роговица способна усиливать свою кривизну
под действием глазных мышц. Все это обеспечивает разную преломляющую способность
глаза, и китообразные могут хорошо видеть как в воде, так и на воздухе. Косатки,
кашалоты, полосатики и серые киты осматриваются, поднимая голову над водой;
дельфины в неволе следят за бросаемой рыбой и кидаются туда, куда она должна
упасть, а иногда схватывают ее даже в воздухе. Они точно хватают рыбу из
рук дрессировщика с высоты 5 м. Дельфины свободно поворачивают глаз и мигают,
если перед ними мелькает какой-либо мелкий предмет. Зрение китообразных преимущественно
монокулярное и в меньшей мере стереоскопическое. Поскольку глаза их расположены
по бокам вытянутой головы, далеко от переднего кончика морды, кашалоты, многие
киты и часть дельфинов не могут видеть предметов впереди своего рыла. Лишь
у дельфинов с маленькой мордой поле зрения левого глаза может частично перекрываться
полем зрения правого глаза, и в этом случае их зрение будет частично стереоскопическим.
Многие дельфины и даже косатки при разглядывании предметов в бассейне ложатся
на бок и исследуют их при помощи то одного, то другого глаза.
Капитан-китобой начала XIX века Вильям Скорсби описал, как гренландские киты,
проходя под килем его корабля, поворачивались на бок, чтобы рассмотреть,
что лежит над ними.
Ученые США Уинтроп Келлог и Чарлз Раис в небольшом бассейне во Флориде испытывали
на восьмилетней ручной афалине Пэдди ее способность к распознаванию различных
фигур с помощью зрения. Фигуры, выставляемые на мишени, состояли из светящихся
кружков, треугольников, квадратиков, крестиков, ромбиков и т. д. Когда дельфин
обучился распознавать эти фигуры, ученые комбинировали их в пары и показывали
ему под водой и на воздухе. В опытах под водой из 25 пар фигур-раздражителей
дельфин успешно различал 21 пару (84%). Результат был отрицательный, если
фигуры помещались на воздухе, а рассматривались из-под воды. Распознаванию
мешала рябь на поверхности воды, искажавшая изображение фигур при разглядывании
их из-под воды. Острота зрения у Пэдди оказалась наилучшей на расстоянии
1,8 м от мишени.
Насколько велика роль зрения у дельфинов при лове добычи и распознавании
препятствий?
Биологи француз Р. Бюснель, датчанин А. Дзидзик и норвежец С. Андерсон провели
серию опытов над морской свиньей в бассейне, имевшем диаметр 54 м. В бассейн
погружали вертикально в шахматном порядке 30 сетей из проволоки толщиной
от 0,2 до 4 мм. Сюда выпускали морскую свинью и за 20 минут свободного плавания
подсчитывали, сколько раз она натыкалась на сети и сколько раз их обходила.
В одной серии опытов она плавала с открытыми глазами, а в другой — с глазами,
закрытыми резиновыми колпачками. Подсчет показал: чем тоньше была сеть, тем
больше было сталкиваний. Сети из проволоки диаметром 0,2 мм «зрячая» морская
свинья избегала в 77% случаев, а «ослепленная» — в 46%. Сети из нейлоновой
лески толщиной 1,5 мм «зрячее» животное избегало в 95%, а «ослепленное» —
в 72%. Ясно, что зрение помогает избегать препятствий «зрячим» особям. Но
с помощью чего делают то же самое «ослепленные» дельфины?
Хорошо известно, что некоторые и зубатые и усатые киты кормятся по ночам,
когда зрение не помогает. У речного дельфина, живущего в мутной реке Ганге,
зрение редуцировалось из-за потери хрусталика. Сильно ослаблено оно и у китайского
речного дельфина-липотес. Во всех этих случаях китообразные при лове добычи
обходятся без помощи глаз.
Какой же орган в таком случае заменяет им зрение? Натуралисты уже давно заметили,
что если среда не позволяет млекопитающим использовать зрение, оно компенсируется
другим органом чувства. Таковым у китообразных оказался слух. Мир дельфинов
и китов — это мир звуков. Именно слух оказался в их жизни самым важным анализатором,
исключительное совершенство которого выдвинуло китообразных по уровню развития
на первое место среди животных Мирового океана.
Море оказалось исключительно благоприятной средой для развития тонкого слуха
у водных млекопитающих: ведь звуки в воде распространяются почти в 5 раз
быстрее, чем в воздухе, и на гораздо большее расстояние. Однако одного чуткого
слуха оказалось недостаточно, и у зубатых китообразных развился очень совершенный
гидролокатор, позволяющий ориентироваться в окружающей среде при помощи звуковых
лучей. Для исследования водного пространства звуковыми лучами китообразные
вначале издают звуки, а потом улавливают эхо посланных звуковых волн от окрестных
предметов. По такому эху китообразные отлично ориентируются в воде независимо
от освещенности и глубины и получают весьма полную информацию об окружающем
— о пище, сородичах, близости берега и о многом другом. Ориентация по отраженным
звукам и называется эхолокацией. Эхолокация — одно из средств обнаружения
препятствия и определения его положения в пространстве с помощью посланного
и отраженного звукового сигнала. Этим средством, как теперь установлено,
владеют многие млекопитающие, а также птицы, например, стрижи-саланганы,-
кроншнепы, живущие в пещерах американские козодои. Эхолокацией пользуются
некоторые ластоногие (тюлени, например), но лучше всего она развита у рукокрылых
(летучие мыши) и зубатых китов.
Рис. 35.
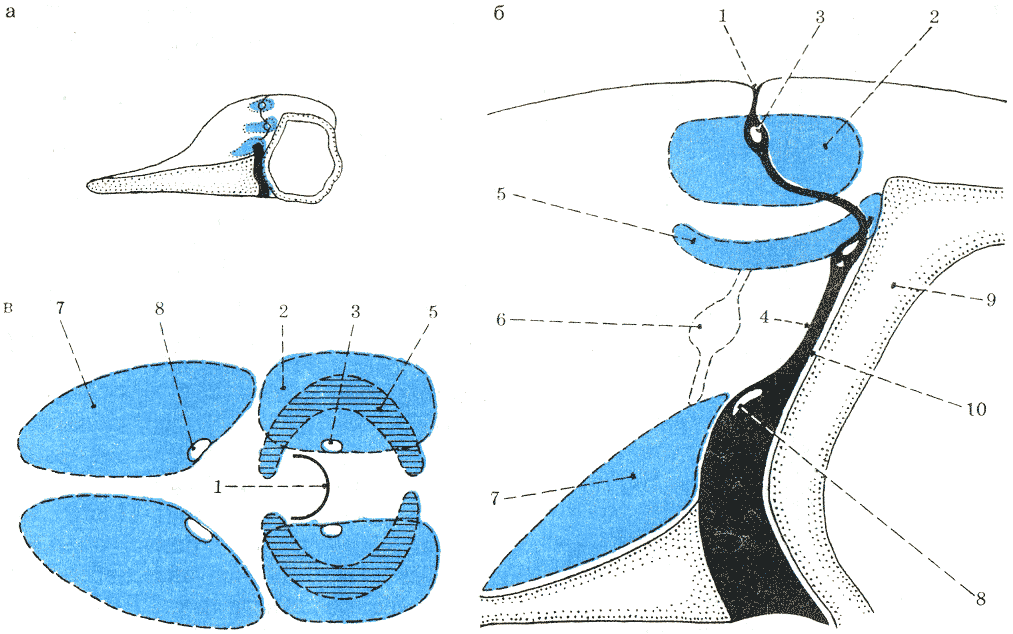
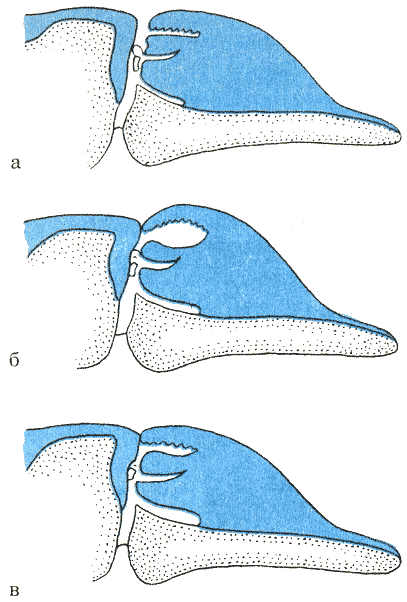
Схема расположения воздушных мешков у обыкновенного дельфина: а, б — продольный разрез головы; в — проекция сверху: 1 — щель дыхала; 2 — верхний мешок; 3 — его отверстие в носовой канал; 4 — мускульная пробка, запирающая носовой канал; 5 — трубчатый мешок; б — соединительный проход с расширением; 7 — нижний (надмежчелюстной) мешок; 8 — его отверстие в носовой канал; 9 — кости черепа; 10 — носовой канал. По Е. В. Романенко, А. Г. Томилину и Б. А. Артеменко.
В гидролокаторе китообразных имеются как бы два тесно связанных устройства: передающий механизм и приемный механизм. Передающее устройство включает три пары воздушных мешков, тесно связанных с носовым каналом, гортань, лобно-носовую подушку или «жировую линзу» и рефлектор, образованный вогнутой передней поверхностью черепа. Приемное устройство, к которому возвращаются отраженные звуки, включает органы слуха, нижнюю челюсть и, возможно, механорецепторы головы.
Рис. 36.
Схема действия воздушных мешков при эхолокации: а — перед началом щелканий; б — в момент щелканий; в — к концу щелканий. Рис. К. Норриса. |
Передающий механизм эхолокационного аппарата очень сложен.
Главную роль в нем играют воздушные мешки, связанные с носовым каналом и
расположенные
в мягких тканях головы над костными ноздрями. У разных видов дельфинов воздушные
мешки устроены по-разному. У большинства видов их три пары (рис. 35). Первая
верхняя пара — вестибулярные мешки — открывается в носовой канал на 1 см ниже
наружной щели дыхала. К ним прикрепляется несколько слоев радиально расположенных
мышц, способных при сокращении изгонять воздух из этих резервуаров.
Вторая пара — трубчатые мешки — залегает ниже первой пары в виде двух подкововидных
трубок вокруг ноздри. Они открываются с задне-боковой стороны носового канала,
ниже основной мускульной пробки. По-видимому, воздух, перекрытый пробкой,
может циркулировать в них без выхода наружу. Третья пара — нижние мешки —
залегает на межчелюстных костях и открывается как раз впереди наружного края
костных ноздрей, у самого основания мягкой части носового канала. Эта пара
связана со второй соединительными проходами. Все мешки работают с помощью
сложной системы носо-челюстных мышц. Выяснение роли каждого мешка в механизме
звукообразования — сложная задача, требующая специальных экспериментов и
особой методики.
Верхние мешки при издавании звуков могут играть роль вместилищ, из которых
воздух под давлением мышц в одних случаях вырывается наружу через узкую щель
едва приоткрывающегося клапана дыхала, а в других — поступает в нижележащие
мешки.
В первом случае в воде видно, как из дыхала вылетает струйка мелких воздушных
пузырьков, и края сомкнутой щели дыхала слабо шевелятся. Когда стягивают
кошельковый невод с окруженными дельфинами, вода словно вскипает от таких
пузырьков.
Во втором случае пузырьки в воде не видны, так как воздух перегоняется в
более глубокие мешки (во вторую и третью пары), что регулируется мускулатурой
дыхала, мускульной пробкой и ее выступом в виде губы. Этот выступ В. Эванс
и Д. Прескотт назвали «дребезжащим органом», так как он, открывая и закрывая
вход в трубчатые мешки, обусловливает щелканья и взрывы щелканий. По гипотезе
Кеннета Норриса, мешки работают так (рис. 36). В момент дыхательного акта
мускульная пробка носового канала втягивается, носовой проход и дыхало открываются,
а верхние мешки спадают. В конце дыхательного акта дыхало плотно закрывается,
верхние мешки наполняются воздухом, и носовой канал блокируется мускульной
пробкой так, что выше ее оказываются верхние и средние, а ниже ее — нижние
мешки. Ряд щелканий производится, как только воздух из верхних мешков наполняет
средние и отсюда начинает поступать через соединительные проходы в нижние
мешки. К концу ряда щелканий запас в верхних мешках истощается, и мускульная
пробка готова открыться, чтобы позволить вновь заполнить эти мешки для следующей
серии щелканий. Дыхательный акт при этом не обязателен, так как верхние мешки
могут перезаполняться воздухом из легких под давлением диафрагмы и мышц грудной
клетки.
Очевидно, функции мешков в звукообразовании, дифференцированы. Д. Лилли и
Э. Миллер в 1962 году наблюдали, как одна афалина щелкала левыми воздушными
мешками, а свистела правыми, но могла делать и то и другое одновременно.
Левая и правая части звукосигнального аппарата могут действовать независимо:
правая — издавать свист в воздухе или под водой, а левая — щелкать.
Механизм звучания ученые связывают не только с воздухоносными мешками, но
и с пневматическими синусами слуховой области: произведенный щелчок возбуждает
в системе воздушных синусов и мешков резонирующие колебания высоких частот.
А так как мешки под влиянием сокращения мышц дыхала способны изменять размер
и форму, то частоты свиста и щелканий меняются (частотные модуляции). Это
создает многообразие звуков.
Многое в механизме подачи звука еще не ясно, в том числе и вопрос, как дельфины
издают звуки при открытом дыхале. Чтобы показать участие воздушных мешков
в генерации сигнала, американские ученые Эванс и Прескотт пропускали воздух
под давлением через носовой канал отрезанной головы дельфина. Полученные
таким образом звуки напоминали звуки живых дельфинов. Ученые предположили,
что эхолокационные щелканья производят «дребезжащий орган» и трубчатые воздушные
мешки, в которых возможен непрерывный поток воздуха.
Может быть, звук возникает при вибрации тонкой перегородки между двумя ноздрями.
Поскольку некоторые сигналы дельфины издают с открытым дыхалом, не исключено,
что звуки генерируют также мелкие складки слизистой гортани. Норвежец О.
Юнсгор, а потом и американец В. Эванс получили визги, такие, как у дельфинов,
пропуская воздух через туго натянутый верхний конец отрезанной гортани дельфина.
Возможно, в генерации звуков участвуют и голосовые связки, которые обнаружила
недавно в гортани одной афалины анатом М. С. Грачева, если только ее находка
не относится к случаю атавизма.
Познакомимся с приемным механизмом гидролокатора китообразных.
Их отличный слух не снижается ни отсутствием наружного уха, ни суженным,
заросшим слуховым проходом. Дельфины воспринимают не только звуки, но и инфра-
и ультразвуки, лежащие далеко за пределами человеческого слуха. Они прекрасно
разбираются в многоголосом хаосе моря и очень точно определяют, с какого
направления поступает звук. В воде это сделать не может ни одно наземное
млекопитающее. Вибрация черепа под воздействием звуковых волн мешала бы такому
определению, если бы правое и левое ухо китообразных не были изолированы
от костей черепа.
Среднее и внутреннее ухо помещаются в двух очень плотных костных образованиях:
первое в барабанной, а второе — в околоушной кости (рис. 37). Обе кости жестко
связаны и подвешены на короткой сухожильной связке одна над другой в обширной
барабанной полости черепа и со всех сторон окружены воздушными камерами,
заполненными пеной из жировой эмульсии. Эта пена, содержащая миллионы воздушных
пузырьков, поглощает все звуковые колебания, которые проводят череп, мускулатура
и сало. Остается единственный путь для звука попасть во внутреннее ухо —
через наружный слуховой проход и цепь косточек среднего уха. Но в 1964 году
американский ученый К. Норрис обнаружил и другой путь передачи звука — через
нижнюю челюсть. Значение этого волновода экспериментально подтвердили в 1968
году электрофизиологи США и Японии Т. Буллок, А. Гринелл, Е. Икезоно и другие.
Нижняя челюсть своим задним концом вплотную подходит к ушной области дельфинов
и в задней части имеет очень тонкие наружные костные стенки. Этот участок
Норрис и рассматривает как «акустическое окно» для прохождения звука. При
этом ход акустических лучей во время эхолокации таков (рис. 38): локационный
пучок, произведенный воздушными мешками, а затем отраженный от костной стенки
черепа и преломленный жировой подушкой (акустической линзой), встречает на
своем пути, например, рыбу. Отраженные от рыбы акустические лучи возвращаются
в ухо дельфина: сначала через кожу к самой тонкой части нижней челюсти —
костной мембране (0,3 мм толщиной), отсюда к внутричелюстному жировому телу
и, наконец, в ухо. При всем том имеет значение угол, под которым отраженные
звуковые волны падают на нижнюю челюсть и в зависимости от этого проводятся
с различным эффектом. Коэффициент звуковой передачи будет высоким, если угол
падения звуковой волны к нижней челюсти будет не менее 30°, точнее — от 30
до 90°. В связи с этим обстоятельством и рассматривается привычка дельфина
покачивать («сканнировать») головой при приближении его к добыче.
Сильное развитие нервно-рецепторного аппарата в нижней челюсти дельфинов
может служить дополнительным аргументом к концепции К. Норриса о поступлении
акустической информации через нижнюю челюсть. Дельфины, слуховые отверстия
которых закрывались присосками из латекса, продолжали спокойно пользоваться
гидролокацией. В опытах американских и японских исследователей восприятие
звуков через нижнюю челюсть оказалось в 6 раз выше, чем через слуховой проход.
Вот почему теперь думают, что наружный конец слухового прохода не играет
большой роли в проведении звуковых волн.
Рис. 37.
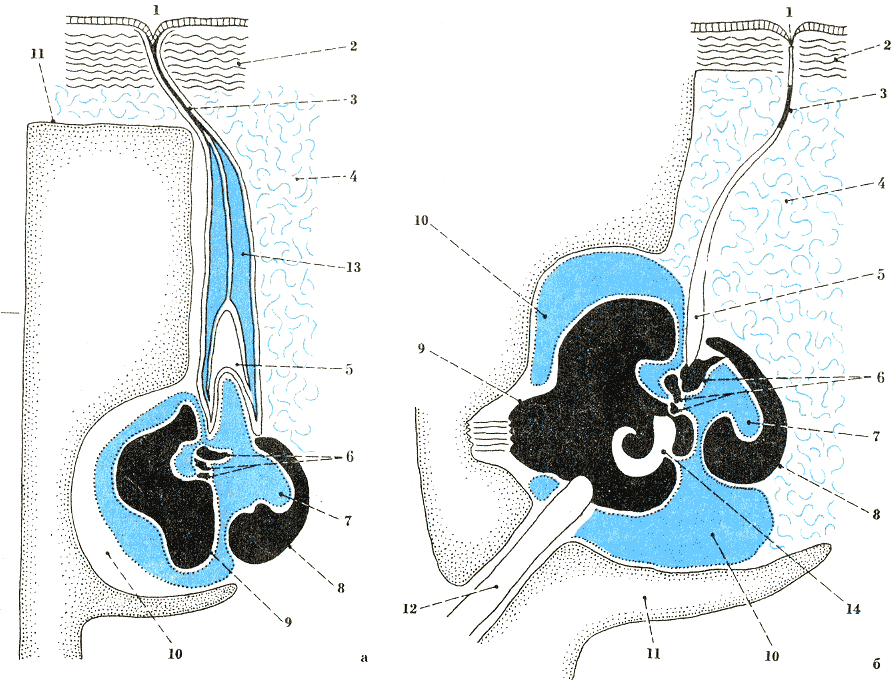
Ухо китообразных: а — усатых китов; б — зубатых китов; 1 — наружное ушное отверстие; 2 — слой кожного сала; 3 — слуховой проход и его зарастающая часть; 4 — соединительная ткань; 5 — барабанная перепонка; 6 — ушные косточки (молоточек, наковальня, стремечко); 7 — барабанная полость; 8 — барабанная кость (булла); 9 — каменистая кость; 10 — пространство, заполненное пеной; 11 — кости черепа; 12 — слуховой нерв; 13 — ушная пробка; 14 — улитка. По Рейсенбаху де Хаан и Пурвесу.
Ушные проходы дельфинов — тонкие, нитевидные, чуть изогнутые каналы — открываются
на обеих сторонах головы позади и чуть ниже уровня глаза. На середине пути
канал зарастает, но вновь появляется, входя в полость среднего уха. Барабанная
перепонка напоминает сложный зонтик, вершиной упирающийся в укороченную рукоятку
молоточка. Молоточек тонким концом связан с барабанной костью, толстым —
с наковальней. Все три слуховые косточки (молоточек, наковальня и стремечко)
плотные, миниатюрные, специализированные для передачи звуков и ультразвуков.
Удельный вес их почти в 1,5 раза больше, чем у человека. Амплитуда движения
стремечка, примыкающего к овальному окошечку, увеличивается по сравнению
с амплитудой движения барабанной перепонки почти в 30 раз, но во столько
же раз теряет в силе. Это важное приспособление к передаче звука в воде,
где амплитуда той же самой звуковой волны по сравнению с воздушной средой
уменьшена почти в 60 раз, но во столько же раз усилено ее давление. Барабанная
кость (булла) — плотная и полая внутри камера, у усатых китов — яйцевидная,
а у дельфинов — полуцилиндрическая. С помощью мышц эта кость может совершать
небольшие перемещения относительно черепа, обеспечивая стереофонический или
объемный прием отраженных локационных щелчков.
Строение внутреннего уха сложное. Жестко подвешенная околоушная кость массивная
и плотная, с минимумом пустот. Улитка и сильно уменьшенный вестибулярный
аппарат занимают в этой кости очень скромное место. В улитке развиты вторичная
спиральная пластина и относительно маленький, жестко подвешенный кортиев
орган. Слуховой нерв достигает исключительно мощного развития. У афалины,
например, улитка почти такой же величины, как у человека, но слуховой нерв
много толще: он достигает в сечении 5 мм.
У китообразных, по мнению голландского ученого Рейсенбаха де Хаана, выполнены
все условия для развития оптимального подводного слуха и восприятия ультразвуковых
частот. Советский исследователь В. П. Зворыкин показал, что у дельфинов гораздо
сильнее, чем у человека, развиты подкорковые слуховые центры, что поставлено
в связь со способностью зубатых китов воспринимать ультразвуки. Височная
же доля коры больших полушарий лишь по глубине и количеству извилин превосходит
височную долю мозга человека, но по микроструктуре такого превосходства нет.
Видимо, справедливо мнение, что эхолокационная деятельность у дельфинов протекает
больше на уровне подкорки, чем на уровне слуховой коры мозга.
В океанариумах методом условных рефлексов установили частоты, на какие отвечают
дельфины. Диапазон воспринимаемых частот оказался от нескольких десятков
герц до 196 кгц. Это в 10 раз выше верхнего предела слышимости человеческого
уха. Американские биологи Барбара Лоренс и Вильям Шевилл обучили одну афалину
подплывать к кормовой площадке в ответ на звуковой сигнал, и каждый раз награждали
ее рыбой. Скормив ей 1200 рыб, они узнали, что афалина отвечает на звуки
от 150 гц до 153 кгц. Однако подопытное животное отвечало на 100% сигналов
только в пределе частот от 150 гц до 120 кгц: с повышением частоты процент
ответов резко снижался.
Наши гидроакустики В. П. Морозов, А. И. Акопиан, В. И. Бурдин и другие в
1971 году показали, что максимальная чувствительность слухового анализатора
дельфинов-афалин располагается в полосе 20 — 80 кгц.
Смещение чувствительности слуха в сторону высоких частот связано с тем, что
в спектре звуков, издаваемых дельфинами, имеются высокочастотные сигналы.
А для наземных млекопитающих известно: какую частоту они излучают, такую
и воспринимают.
Дельфины способны воспринимать также инфразвуки, например, «голос шторма»:
они обычно уходят из прибрежной зоны перед надвигающейся бурей, угрожающей
выбросить их на отмель. Очевидно, ими улавливаются шум волн, омывающих их
тело, всплески на поверхности моря, гул, издаваемый косяками рыб, ракообразными,
водными млекопитающими, а также отраженные волны разных частот (эхо).
Рис. 38.
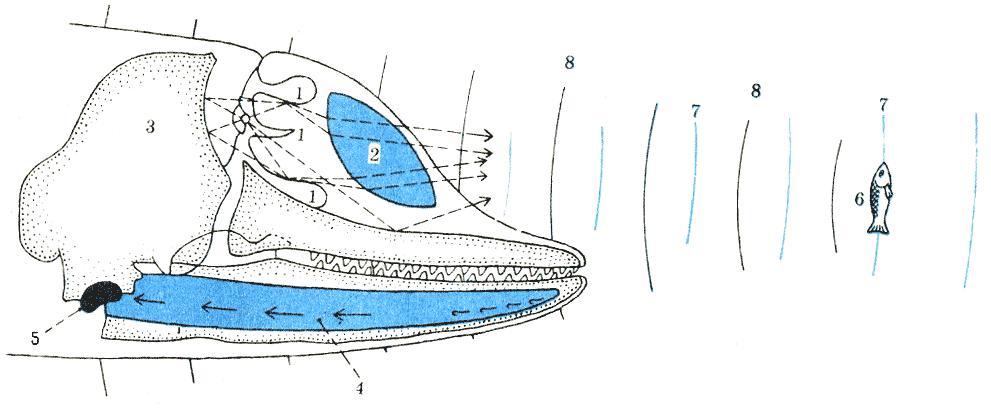
Схема эхолокации дельфина (ультразвуковой прожектор и акустическая линза в голове дельфина): 1 — воздушные мешки, излучающие звуки; 2 — жировая подушка; 3 — череп; 4 — нижняя челюсть, проводящая звуки в ухо; 5 — барабанная кость и внутреннее ухо; 6 — предмет лоцирования (рыба); 7 — излученные звуковые волны; 8 — отраженные звуковые волны. По Норрису, с изменениями.
Способность воспринимать звуки в очень широких пределах частот позволяет
китообразным получать исключительно широкую информацию из окружающей среды,
что вполне компенсирует недостаточное развитие их зрительного и вкусового
анализаторов.
В соответствии с высоким развитием слуха у китообразных сформировались превосходный
голосовой аппарат и весьма богатая звуковая сигнализация.
О том, что эти животные издают различные звуки, люди знали давно. Северный
дельфин — белуха — даже получил название «морской канарейки» за свою способность
хрюкать, стонать, свистеть, издавать звуки, напоминающие плач ребенка, женский
пронзительный крик, шум толпы, игру на флейте, щебетание птиц. Однако вокальные
способности китообразных не привлекали к себе внимания. Положение резко изменилось
в годы войны, после того, как звукометрические станции американского флота,
определявшие с помощью гидрофонов приближение вражеских подводных лодок,
поднимали напрасную тревогу из-за шума морских животных.
Точное определение источников звуков, в том числе и биогенных, приобрело
особое значение. Слух и голос китообразных стали предметом тщательных экспериментальных
исследований, и в этой области были сделаны важные открытия.
Каждый вид китообразных пользуется несколькими типами звуков. Для чего и
при каких обстоятельствах используют свой голос дельфины и киты? Каково при
этом их поведение и ответные реакции сородичей на эти сигналы?
Чтобы разобраться во множестве сигналов китообразных, необходимо классифицировать
издаваемые ими звуки в зависимости от физической характеристики и от биологического
значения сигналов для самих китообразных.
По физическим свойствам сигналы различают непрерывные и прерывные, состоящие
из серии импульсов или щелчков. Нейрофизиологи из США Джон Лилли и Элис Миллер
объединили все звучания дельфинов по физическим свойствам в три класса: I
— эхолокационные ультразвуковые щелканья (до 170 кгц); II — непрерывные звуки
— свисты частотой от 4 до 18 — 20 кгц и III — звуки сложного спектра — комплексные
волны высокой амплитуды, называемые по внешней аналогии с сигналами других
животных кряканьем, мяуканьем, лаем, воем, жужжаньем, ревом и т. д. Лилли
и Миллер открыли, что один и тот же дельфин способен издавать акустические
колебания двух или даже всех трех классов одновременно. Это значительно увеличивает
возможные комбинации сигналов и сильно усложняет их звучание.
По биологическому значению акустические сигналы зубатых китов можно условно
подразделить на две группы: одни издаются преимущественно для ориентации,
навигации, рекогносцировки, для разыскания пищи (эхолокационные сигналы I
класса), а другие — для связи или общения со своими сородичами (коммуникационные
сигналы II класса). Акустические же сигналы III класса, по-видимому, используются
и как коммуникационные и как эхолокационные. Рассмотрим вначале эхолокацию
как способ ориентации зубатых китов.

Глава 1. Нашествие сенсаций 8
Глава 2. «Помощники» и друзья человека в море 12
Глава 3. С суши в воду 18
Глава 4. Человек и дельфины 34
Глава 5. Поведение узников 46
Глава 6. «Тираны» китов — любимцы зрителей 62
Глава 7. Скороходы морей 70
Глава 8. Покорители морских глубин 84
Глава 9. Судьба анализаторов 94
Глава 10. «Акустические глаза» дельфинов 110
Глава 11. Поющие киты и «морские попугаи» 122
Глава 12. Обладают ли дельфины речью? 140
Глава 13. Мышление дельфинов конкретно, а действия
трафаретны 148
Глава 14. Рефлексы выныривания и инстинкт сохранения
вида 152
Глава 15. Сознательно ли дельфины спасают людей
160
Глава 16. «Самоубийства» среди китообразных или
жертвы инстинкта 164
Глава 17. Странные попутчики или бездумные лоцманы
172
Глава 18. Дельфины и наука 176
Глава 19. Сигналы дальнего действия и звуковые
маяки в океане 184
Глава 20. «Звезды моря» служат человеку 188
Глава 21. Будущее дельфинов 198
Систематический список видов отряда китообразных 202
Алфавитный список видов китообразных 205