По техническим причинам в этой и последующих главах при переводе текста книги в HTML форма написания зубной формулы млекопитающих была изменена по сравнению с оригиналом.
В. П.
ГЛАВА 10
НОТОУНГУЛЯТЫ
Древние греки и римляне были склонны к антропоморфизму и персонифицировали
многие явления природы. Южный ветер носил у греков имя Нотос, а римляне
называли его Нотус. В научной номенклатуре имена латинизируются даже в
тех случаях, когда они происходят из греческого или из варварских языков
(любой язык, кроме латинского и греческого, прежде считался варварским).
Это название южного ветра, звучавшее почти одинаково по-гречески и по-латыни,
в форме noto- часто использовали как приставку, означающую просто «южный».
Таким образом, название Notoungulata буквально значит «южные копытные [млекопитающие]».
Когда в 1903 г. Сантьяго Рот придумал это название, нотоунгуляты были известны
только из Южной Америки. Теперь, как уже говорилось в гл. 5, некоторые
из них найдены также в Северной Америке и Азии, но название Notoungulata
остается пригодным для группы, которая на протяжении всей своей известной
истории была преимущественно южноамериканской.
По довольно консервативной системе, которой мы здесь следуем, выделяют
всего 14 семейств нотоунгулят, в том числе сем. Arctostylopidae, не известное
из Южной Америки. В риочикии и касамайории, т. е. примерно в позднем палеоцене
и раннем эоцене, можно различить 8 семейств. В то время все они были еще
примитивными, и различия между ними были не очень велики. Еще одно семейство
(Notohippidae), эволюционно более продвинутое и сильнее отличавшееся от
других, впервые становится известным в мастэрсии (примерно средний эоцен).
В открытых до сих пор десеадских фаунах обнаружено 9 семейств нотоунгулят
— больше, чем в каком-либо ином веке наземных млекопитающих; из них 5 появляются
в десеадии впервые. К десеадию, т. е. примерно к раннему олигоцену, семейства
нотоунгулят стали более заметно отличаться друг от друга и явно заняли
разнообразные экологические ниши. В дальнейшем происходило дифференцированное
вымирание — своего рода прореживание. Из сантакрусия известно всего 5 семейств,
а от сантакрусия до часикия, т. е. для времени, примерно соответствующего
миоцену в целом, — только 6. Из плиоцена (от уайкерия до чападмалалия)
известно только 3 семейства; все они дожили до плейстоцена, но затем вымерли.
Древнейшие южноамериканские фауны, в которых присутствуют нотоунгуляты,
— риочикская, касамайорская и мастэрская, особенно интересны из-за происходивших
в них эволюционных процессов и событий. Здесь мы застаем данную группу
в разгаре того, что вполне можно назвать эволюционным взрывом.
Это проявляется в двух разных аспектах. Во-первых, нотоунгуляты, которые
в масштабах геологического времени, вероятно, еще недалеко ушли от своих
истоков, уже достигли значительного разнообразия. В 8 семействах нотоунгулят
из этих трех фаун по современным системам насчитывается более 35 родов
и около 70 видов. И это — с довольно ограниченной территории: к ним следует
добавить находки из других районов, полностью еще не описанные, а также
те, которые будут сделаны в будущем. Сборы из районов, которые в те времена,
как и сейчас, были окраинными частями Южной Америки, ясно показывают, что
нотоунгуляты, используя любую благоприятную возможность, бурно распространялись
по огромному континенту, на котором жило относительно мало других копытных,
а те, которые там были, как показано в предшествующей главе, постепенно
все больше специализировались, оставляя свободными многие экологические
ниши, пригодные для копытных.
Во-вторых, ранний расцвет данной группы отличался тем, что по крайней мере
некоторые из входивших в нее видов все еще оставались необычайно изменчивыми.
Это, вероятно, относится ко многим видам, но наиболее объективные данные
имеются по одному сравнительно примитивному виду с довольно устрашающим
научным названием Henricosbornia lophodonta Ameghino, 1901. Этот род долгое
время оставался настолько непонятным, что Генри Ферфилд Осборн (Henry Fairfield
Osborn), в честь которого он был назван, исключил его из своей авторитетной
классификации 1910 г. и из других работ. Однако во время моей первой экспедиции
в Южную Америку в 1930 — 1931 гг. моим спутникам и мне удалось собрать
значительно больше ста экземпляров одного этого вида. Хотя среди них не
было хорошо сохранившихся черепов или скелетов, зубы этого вида сейчас
изучены очень хорошо, что позволило твердо установить его широкую изменчивость.
Изучение наших сборов и материалов братьев Амегино показало, что Флорентино
Амегино, располагавший меньшим, чем мы, числом экземпляров данного вида,
разбил их на 16 видов, 8 родов, 4 семейства и 3 отряда. Мы приводим эти
данные не в порядке критики работы Амегино, а как яркое свидетельство того,
каким исключительным разнообразием отличаются зубы особей этого вида. Амегино
указывал, что все его различные «виды» этой группы произошли от кондиляртр,
что, очевидно, совершенно справедливо, если говорить о том единственном
виде, к которому их теперь относят. Однако он считал разные варианты этого
вида предками таких различных более поздних групп, как лемуры и носороги.
Зубы этого и некоторых других столь же древних видов в то время были подвержены
индивидуальной изменчивости, позднее некоторые вариации фиксировались и
становились характерными признаками различных родов и даже семейств (но
не отрядов).
В группе, находящейся в состоянии экспансии, еще не обосновавшейся окончательно
в какой-либо узкой экологической нише, вполне возможна такая изменчивость,
или, как можно было бы сказать, нечеткость генетической регуляции морфологических
признаков. Она обеспечивает лабильность популяции и ее способность к разнообразным
адаптациям и к освоению разнообразных образов жизни. Как только отклонившаяся
популяция становится более строго приспособленной к данному образу жизни
или нише, центростремительный (стабилизирующий) естественный отбор обычно
начинает препятствовать ее дальнейшему изменению, снижает скорость этого
процесса или даже останавливает его путем выбраковки более сильно отклоняющихся
генетических вариантов. В этих древних фаунах лишь немногие из нотоунгулят
достигли такой стадии. Как мы увидим дальше, в следующем веке, десеадии,
число их возросло.
Весьма характерное строение зубной системы нотоунгулят в его наиболее примитивной
форме можно наблюдать у таких древних родов, как Henricosbornia или Oldfieldthomasia;
морфология последнего изучена лучше, хотя изменчивость известна не так
хорошо, как у Henricosbornia. Первоначально нотоунгуляты имели
следующую зубную формулу, очень рано закрепившуюся у примитивных плацентарных
млекопитаюших:
3/3•1/1•4/4•3/3;
это означает, что у них было по три резца, одному клыку, четыре премоляра
и три моляра на каждой стороне верхней и нижней челюстей. У древнейших
форм все эти зубы в каждой челюсти образовывали непрерывный, постепенно
изменяющийся ряд: клык имел простое строение и весьма напоминал прилегающие
к нему резец и премоляр, а последний премоляр был очень похож на более
сложный первый моляр.
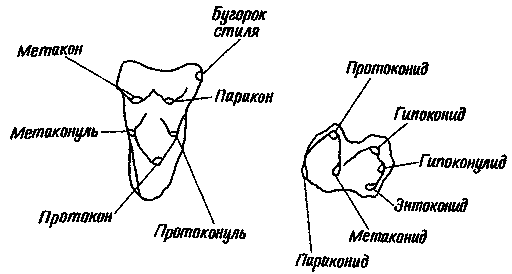
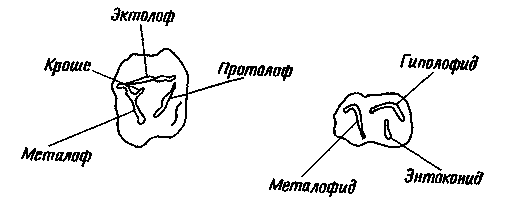
Моляры нотоунгулят по основным чертам строения отличаются от моляров всех
остальных млекопитающих. Они схематически изображены на рис. 19, В, основанном
главным образом на зубах Henricosbornia. На верхних молярах было по три
основных гребня, или лофа, показанных и обозначенных на рисунке. Имелся
еще один гребень меньшего размера, или шпора, отходящий от одного из основных
гребней и известный под названием кроше. Для строения коронки нижних моляров
наиболее характерно наличие двух гребней, называемых здесь лофидами. (В
системе номенклатуры зубных бугорков, разработанной Г. Ф. Осборном и ставшей
теперь, с соответствующими дополнениями, общепринятой, суффикс -ид используется
для большинства элементов нижних моляров.) Кроме того, один из бугорков
(энтоконид; см. рис. 19, А, В) превратился в короткий поперечный гребень,
отделенный от двух других; на этот признак мы уже указывали в предыдущей
главе как на одно из доказательств того, что моляры пиротерия не могли
произойти от моляров одного из нотоунгулят.
А
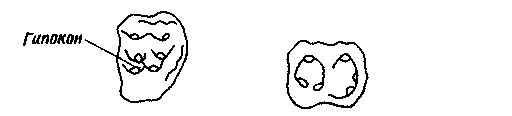
Б
В
Рис. 19. Правые верхние (слева) и нижние (справа) моляры трех групп млекопитающих.
A. Строение зубов, примитивное для плацентарных млекопитающих вообще. Это
не изображение зубов представителя какого-то конкретного рода, а обобщенная
схема структуры, с теми или иными вариациями имеющейся почти у всех меловых
плацентарных; на схеме указаны названия основных бугорков. Б. Зубы примитивного
дидолодонтида. Рисунок несколько стилизован, но напоминает, например,
зубы Ernestokokenia из риочикия и касамайория. Гипокон — новый бугорок;
его не было у большинства примитивных плацентарных, и он до сих пор отсутствует
у некоторых современных плацентарных. Остальные бугорки лишь слегка изменились
по сравнению с примитивным для плацентарных типом. B. Зуб примитивного
нотоунгулята. Рисунок сделан в основном по зубам Henricosbornia из риочикия
и касамайория. большая часть бугорков, имевшихся у примитивных плацентарных,
объединилась в гребни (лофы на верхних зубах и лофиды на нижних); появился
также новый гребень — кроше, отходящий от металофа.
Такие явно незначительные или скрытые от глаз непосвященных детали строения
зуба дают весьма существенную, хотя и не всегда достаточную, информацию
для определения и классификации многих млекопитающих, как ныне живущих,
так и вымерших. Основной примитивный нотоунгулятный тип строения зубов,
описанный в предыдущем абзаце и изображенный на рис. 19, наблюдается у
всех несомненных нотоунгулят, хотя, как мы отмечали в предыдущей главе,
время от времени к нотоунгулятам относят (правомерность этого весьма сомнительна)
некоторые группы с иным строением зубов. Эти черты строения моляров, характерные
для примитивных нотоунгулят, сохраняются у всех поздних и более специализированных
представителей этой группы, даже если в остальном их моляры сильно изменяются
и приобретают новые черты, а зубная система в целом становится более специализированной.
О своеобразии в строении ушной области у нотоунгулят уже упоминалось в
предыдущей главе. Из древнейших нотоунгулят оно наиболее подробно изучено
у касамайорского рода Oldfieldthomasia и проиллюстрировано на серии распилов.
Его наиболее замечательная особенность, присущая всем нотоунгулятам, —
наличие двух окруженных костями пузыревидных полостей, открывающихся в
среднее ухо, одна сверху, а другая снизу. Среди прочих специфических черт
строения отметим, что барабанная кость, помимо того, что она окружает нижнюю
(гипотимпанальную) полость, образует трубку, ведущую от среднего уха к
наружному (которое не окостеневает и поэтому в ископаемом состоянии неизвестно);
вдоль нижней, или вентральной, стороны этой трубки тянется гребень. На
нижней поверхности черепа, позади стенки нижнего пузыря (буллы, или гипотимпанальной
полости), имеется ямка, в которую входит отросток подъязычной кости, или
гиоида.
Подобное строение среднего уха встречается не только у нотоунгулят. Оно
независимо возникало у сумчатых по крайней мере трижды, в частности в Южной
Америке у вымерших форм Argyrolagus и Sparassocynus (см.
гл. 7). Также не менее трех раз оно возникало у грызунов. Однако происхождение
этой структуры
у всех названных групп и у нотоунгулят неидентично. Очевидно, что это пример
многократной конвергентной адаптации, но адаптации в чему? Эксперименты,
проведенные на некоторых грызунах, показывают, что подобное строение повышает
чувствительность уха к звукам определенных частот, возможно к голосам других
особей своего вида или же к звукам, издаваемым хищниками. Эксперименты
с моделями ушной области нотоунгулят, быть может, позволили бы выяснить
частоты, на которые были настроены их уши, но таких экспериментов пока
никто не ставил, да и едва ли можно узнать, какие звуки издавали друзья
или враги нотоунгулят.
Одно из древних и в общем примитивных семейств нотоунгулят, Notostylopidae,
все же стало умеренно специализированным в некоторых отношениях. Остатки
этого семейства редки в известных фаунах риочикия, обильны в касамайории,
снова редки в мастэрсии, а позже не встречаются. Типовой род Notostylops (рис. 20) обычен в касамайорских отложениях, которые братья Амегино называли
«нотостилопсовыми слоями» или «нотостилопием». Особенно обилен один вид,
Notostylops murinus. И снова последующие более обширные сборы, более точно
документированные, чем это было возможно во времена Карлоса Амегино, выявили
необычайную изменчивость в пределах этого единого вида. Снова, как и в
случае с Henricosbomia lophodonta, такая изменчивость ввела в заблуждение
Флорентино Амегино, который отнес разные экземпляры одного этого вида к
14 видам 6 родов, но на этот раз объединил их всех в одно семейство, Notostylopidae,
представляющее собой реально существующую группу, название которой таксономически
валидно.
Наиболее отчетливый признак специализации у этого семейства выражается
в том, что задние (третьи) резцы, клыки и передние (первые) премоляры всегда
мелкие и часто отсутствуют вовсе, так что между сохранившимися резцами
и сохранившимися премолярами образуется диастема. Зубная формула варьирует
от
предковой 3/3•1/1•4/4•3/3 до 2/2•0/0•3/3•3/3; даже в пределах
одного вида. Столь широкая внутривидовая изменчивость зубной формулы среди
млекопитающих
вообще
встречается крайне редко. У Notostylops один из верхних резцов (первый)
и один из нижних (второй) увеличены и при кусании смыкаются друг с другом.
Это придает животным некоторое сходство с грызунами, но большие резцы у
них кусающие, а не грызущие. Моляры незначительно изменены по сравнению
с самым примитивным нотоунгулятным типом. Наиболее выраженное их отличие
заключается в том, что кроше на верхних молярах, которое у Henricosborniidae
представляло собой всего лишь шпору, удлинилось, почти пересекая промежуток
между протолофом и металофом, а его гребень на ранних стадиях стирания
несет ряд крошечных бугорков.

Рис. 20. Череп и реконструкция головы Notostylops — раннеэоценового (касамайорского) нотоунгулята, имевшего примитивный общий облик, но несколько специализированную зубную систему.
По-видимому, Henricosborniidae, по крайней мере по строению зубной системы,
были наиболее примитивными из известных нотоунгулят, являя собой остатки
ствола, возможно, предкового для всех остальных нотоунгулят, сохранившиеся
примерно до касамайория (ранний эоцен). Notostylopidae представляет собой
умеренно специализированную ветвь, сохранившуюся только до следующего века,
мастэрсия. Эти два семейства не укладываются в рамки основных групп нотоунгулят,
существовавших в одно с ними время и сохранившихся в более поздние века,
а поэтому здесь они помещены в отдельную группу — подотряд Notioprogonia;
это название означает по-гречески «южный предок». (Известные нотоунгуляты
Северной Америки и Азии, Arctostylopidae, также относятся к этому подотряду.)
Остальные семейства, присутствующие в этих древних фаунах, можно распределить
по трем крупным группам; эти группы рассматриваются здесь как подотряды
отряда Notoungulata, и первоначально они были выделены на основании изучения
ископаемых форм из более поздних фаун. Подотряд Typotheria получил свое
название по одному из самых поздних представителей нотоунгулят, которого
долгое время называли Typotherium, т. е. «типовой зверь»; по-видимому,
это название не имело большого смысла. В качестве родового названия оно
оказалось невалидным — этот род следует называть Mesotherium («средний
зверь»), что, по-видимому, не более осмысленно, чем предыдущее название.
Но более поздних представителей этого подотряда почти всегда называли типотериями,
и название Typotheria для него вполне валидно. Этот подотряд уже на ранних
этапах своей истории дифференцировался на три семейства.
Подотряд Hegetotheria также назван по некоему роду Hegetotherium («главный
зверь» — еще одно ничего не говорящее название), а подотряд Toxodonta —
по роду Toxodon («дугозуб» — название, довольно удачно характеризующее
его верхние моляры). К подотряду Typotheria в трех древнейших фаунах относится
только одно семейство, тогда как в подотряде Toxodonta их различают уже
два, одно из которых появляется в летописи только в мастэрсии. Во всех
трех подотрядах есть и более прогрессивные семейства, известные из более
поздних фаун; по одному семейству из каждого подотряда дожило до плейстоцена:
Mesotheriidae из Typotheria, Hegetotheriidae из Hegetotheria и Toxodontidae
из Toxodonta. Хегетотерииды сохраняются только до самого раннего плейстоцена
(укия), но другие дожили почти до голоцена.
Представители подотрядов Typotheria и Hegetotheria имели размеры от мелких
до средних, подотряда Toxodonta — от средних по гигантских. У всех этих
трех подотрядов, как и у многих других, но безусловно не у всех групп млекопитающих
наблюдается увеличение средних размеров с течением времени. Во всех трех
подотрядах древнейшие представители — риочикские и касамайорские — имели
довольно примитивную зубную систему, обычно с полной зубной формулой: 3/3•1/1•4/4•3/3.
Все
зубы были низкокоронковыми (брахиодонтными), а моляры не так уж сильно
отличались от моляров примитивных Notioprogonia. Однако даже на этом эволюционном
уровне верхние моляры имели тенденцию к усложнению; оно проявлялось в дальнейшем
удлинении кроше и возникновении на основных гребнях, или лофах, добавочных
выступов. Как правило, это был выступ, называемый гребешком (crista), отходивший
от наружного гребня (эктолофа) внутрь и часто соединявшийся с кроше; таких
гребешков могло быть несколько. Иногда возникал еще один выступ, называемый
антекроше и отходящий назад от переднего гребня (протолофа). Антекроше
может достигать гребешка или кроше и соединяться либо с одним из них, либо
и с тем и с другим, но на том эволюционном уровне, о котором идет речь,
этого обычно не происходило.
Наиболее хорошо изученные ранние представители подотряда Typotheria — это
касамайорские роды Oldfieldthomasia, выделяемый в самостоятельное семейство,
и Notopithecus (рис. 21), входивший в сем. Interatheriidae, в основном
более позднее (олигоцен-миоценовое). Строение черепа и зубная система этих
двух родов хорошо известны и подробно описаны. Их зубы обладают общими
признаками, указанными в предыдущем абзаце. Черепа их были все еще довольно
примитивными, сохраняя почти полностью исходные нотоунгулятные черты, особенно
в строении ушной области.
В десеадии и в более поздние эпохи интератерии, родственные нотопитекусу
и, возможно, происходившие от него или от одной из близких к нему форм,
явно начинают специализироваться. Некоторые из них по строению черепа и
челюстей не отличаются заметным образом от своих более ранних родичей или
предков. У других, в особенности у самого Interatherium из сантакрусия,
череп и челюсти стали заметно короче, шире и выше. Еще более резко изменилась
зубная система: зубы стали высококоронковыми (гипсодонтными), а медиальная
(передняя) пара верхних резцов увеличилась в размере и затачивалась о две
пары нижних резцов, не особенно крупных. Как это часто бывает, все найденные
зубы оказались сильно стертыми. В таком состоянии жевательные поверхности
моляров и задних премоляров имеют довольно простой рисунок, а именно двулопастной,
что особенно четко видно на внутренней (лингвальной) стороне верхних и
наружной (буккальной) стороне нижних зубов. В результате у некоторых исследователей
создалось впечатление, что у этих типотериев нотоунгулятный тип строения
зубов, описанный выше, был утрачен и заменен более простым. На самом же
деле коронки этих зубов, когда они только что сформировались и еще не успели
износиться, сохраняли примитивный нотоунгулятный тип строения моляра, на
который просто наложились некоторые более поздние усложнения, но вскоре
первоначальный рисунок жевательной поверхности стирался.

Рис. 21. Череп и реконструкция головы раннеэоценового (касамайорского) примитивного интератерия Notopithecus.
Почти полные скелеты известны для двух сантакрусских типотериев — Protypotherium и Interatherium. Строение их конечностей весьма своеобразно. По крайней мере у некоторых ранних нотоунгулят, а возможно, и у всех конечность была пятипалой, т. е. имела примитивное для млекопитающих строение; третий, или средний, палец был немного длиннее остальных, что также обычно для примитивного строения лапы. Названные сантакрусские типотерии четырехпалые, первый палец у них утрачен. На передних конечностях первый палец отсутствует, пятый редуцирован как по величине, так и по функции, а второй — четвертый пальцы вполне функциональны; третий палец немного длиннее второго, а четвертый заметно короче остальных. На задних конечностях третий и четвертый пальцы одинаковой длины и ось конечности проходит между ними, т. е. нагрузка равномерно распределена между этими пальцами. Второй и пятый пальцы короче двух других, и длина их примерно одинакова у несколько более позднего интератерия из фриасия Колумбии эта странная специализация конечностей зашла еще дальше, у этого рода, Miocochilius, передняя конечность трехпалая, причем второй и третий пальцы (из первоначальных пяти) — функционирующие и равны по длине, а четвертый хотя и имеется, но гораздо короче и, вероятно, не несет нагрузки (рис. 22, Г). На задней конечности всего два пальца одинаковой длины — третий и четвертый из пяти, имевшихся у предковых форм. Таким образом, у этого животного весь вес (или большая его часть) падал на два пальца каждой конечности, причем на передней ноге ось конечности проходила между пальцами II и III (так принято указывать, о каком из первоначальных пяти пальцев, I — V, идет речь), а на задней ноге — между пальцами III и IV.
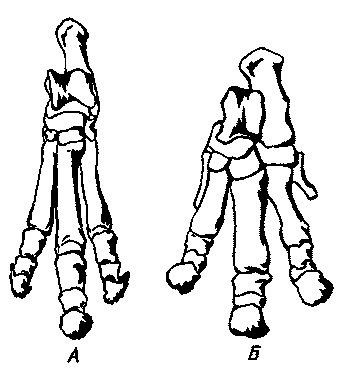 |
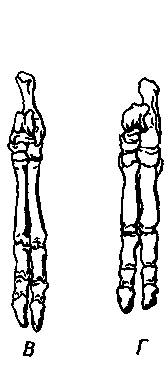 |
Рис. 22. Скелеты задних конечностей двух современных южноамериканских
копытных североамериканского происхождения и двух нотоунгулят южноамериканского
происхождения.
А. Современный тапир (Tapirus); функционируют три пальца (II, III
и IV), а ось конечности проходит по среднему пальцу (III). Б. Scarrittia — раннеолигоценовый
(десеадский) нотоунгулят из леонтиниид; функционировали те же пальцы, что
и у тапира, и ось конечности проходила там же. В. Современный пекари (Tayassu);
функционируют два пальца (III и IV), а ось конечности проходит между ними.
Г. Miocochilius — среднемиоценовый (фриасский) интератерий, функционировали
два пальца (III и IV), а ось конечности проходила между ними, как у пекари
(на передней конечности несли нагрузку пальцы, гомологичные функционирующим
пальцам передней конечности пекари).
Другие нотоунгуляты, у которых на задних конечностях было бы по два пальца
или ось конечности проходила бы между двумя пальцами, неизвестны. Остальные
группы нотоунгулят, конечности которых известны, могут иметь по пять, по
четыре или по три пальца, но во всех случаях эта линия проходит по среднему
(III) пальцу. За пределами островного материка Южной Америки большинство
копытных земного шара четко делятся на две обширные группы, определяемые
по положению оси конечности; У Artiodactyla, к которым относятся коровы,
овцы, свиньи и множество других животных, эта ось как на передней, так
и на задней конечности проходит между равными по величине пальцами III
и IV. У Perissodactyla, к которым относятся лошади, носороги и тапиры (рис.
22, А), эта ось совпадает с доминирующим или в конечном счете единственным
(у лошадей) средним пальцем (III). Столь странное разнообразие в пределах
одного южноамериканского отряда Notoungulata — замечательный пример гибкости
эволюционного процесса или, быть может, его способности использовать любые
благоприятные возможности.
В десеадии, т. е. примерно в раннем олигоцене, загадочным образом появляется
еще одна группа типотериев — подсемейство Trachytheriinae. Они явно родственны
интератериям, но более специализированы и их предок точно не известен.
У этих животных высококоронковость зубов дошла до предела: зубы продолжают
расти на протяжении всей жизни и никогда не образуют корней. Это явление
называется гипселодонтией и представляет собой конечную фазу развития в
сторону гипсодонтии (при гипсодонтии зубы имеют высокие коронки, но в конце
концов образуют корни, и выталкивание зубов из их ячеек прекращается).
Моляры у Trachytheriinae имели почти плоские наружные (буккальные) поверхности;
после того как вследствие изнашивания коронки рисунок жевательной поверхности
упрощался, становились видны три внутренние (лингвальные) лопасти неравной
величины. Эти животные — почти идеальный предковый тип для классических
типотериев (Mesotherium и его близких родичей, живших в позднем кайнозое),
и мы помещаем их здесь в сем. Mesotheriidae. Однако с ними связана еще
одна загадка: промежуточные формы, которые могли служить связующими звеньями
между трахитериями и настоящими мезотериями, не известны, а ведь их разделяет
интервал времени в несколько миллионов лет — от десеадия до постсантакрусия.
Возможно, что эти промежуточные формы жили к северу от Патагонии, в которой
главным образом изучалась эта часть ископаемой летописи.
Так или иначе, Mesotheriidae — классические типотерии — все-таки появляются
где-то в середине миоцена и сохраняются до плейстоцена, где в богатых ископаемыми
поверхностных слоях обширной аргентинской пампы часто встречаются остатки,
относящиеся к роду, который называли то Typotherium, то Mesotherium. Эти
животные были наиболее специализированными из всех типотериев, чего и следовало
ожидать для самого последнего из них, хотя это и противоречит концепции
«выживания менее специализированного». У него был широкий, низкий, массивный
череп, который сравнивали с черепом бульдога или, что более удачно, с черепом
гигантского бобра, поскольку это животное гораздо больше напоминало грызуна,
чем собаку. Ею зубная формула редуцировалась до 1/2•0/0•2/1•3/3.
Все
зубы были постоянно растущими (гипселодонтными). Крупные резцы были похожи
на резцы грызунов, а между ними и редуцированными премолярами и
в верхней, и в нижней челюстях была длинная диастема.
На протяжении многих лет типотериев и хегетотериев путали друг с другом
и обычно объединяли под первым названием, однако они были различимы уже
в риочикии. В некоторых отношениях, хотя и не во всех, эволюция хегетотериев
и типотериев действительно шла параллельно. Самые примитивные представители
хегетотериев, которые нам известны, — Archaeohyracidae — находились примерно
на том же эволюционном уровне, что и самые ранние типотерии, но, как мы
уже упоминали в гл. 6, у них быстрее возникли высококоронковые зубы.
Древнейшие хегетотерии известны только по нескольким зубам и неполным челюстям,
но начиная с десеадия (т. е. примерно с раннего олигоцена) и до раннего
плейстоцена они изучены довольно хорошо. Для длительного промежутка времени
приблизительно от раннего олигоцена до позднего плиоцена известны фактически
полные скелеты для одного десеадского рода (рис. 23), двух сантакрусских
и одного чападмалальского. (Были также созданы реконструкции некоторых
других форм по менее полным материалам.)

Рис. 23. Реконструкция раннеолигоценового (десеадского) хегетотерия Propachyrucos.
Так же как у типотериев, зубы хегетотериев стали постоянно растущими (гипселодонтными);
их число редуцировалось в этом семействе до 1/2•0/0•3/3•3/3, а оставшиеся
резцы увеличились в размере и стали напоминать резцы грызунов. Задние конечности
были значительно длиннее передних: в особенности длинной
была их стопа, опиравшаяся на землю всей своей поверхностью в состоянии
покоя и пружинившая при быстром движении. Ясно, что эти животные передвигались
прыжками и скачками и весьма походили на кроликов по образу жизни и по
внешнему виду с той разницей, что у некоторых из них были довольно длинные
хвосты. Интересно, что в то время, когда у них развился такой кроликоподобный
габитус, в Южной Америке кроликов не было, хотя, как мы увидим в следующей
главе, там обитали некоторые настоящие грызуны. (В систематическом отношении
кролики не относятся к грызунам, т. е. не являются представителями отряда
Rodentia, хотя грызть они, конечно, умеют; то же самое можно сказать и
о поздних типотериях и хегетотериях.) К тому времени, когда настоящие кролики
достигли Южной Америки, эти псевдокролики уже вымерли.
Третий подотряд нотоунгулят, Toxodonta, далек от внутреннего единства,
наблюдаемого у типотериев и хегетотериев. Здесь мы делим его на пять заметно
дивергировавших семейств. На протяжении большей части раннего кайнозоя
это была исключительно многочисленная группа, доминировавшая среди нотоунгулят,
но в конце концов в плейстоцене от их многообразия осталась лишь горстка
видов, которые впоследствии также все вымерли.

Рис. 24. Реконструкция раннеэоценового (касамайорского) изотемнида Thomashuxleya — примитивного представителя подотряда Toxodonta.
Все представители этого подотряда, известные из древнейших фаун, риочикской
и касамайорской, принадлежат к наиболее примитивному сем. Isotemnidae.
Единственная на сегодняшний день реконструкция целого скелета додесеадского
нотоунгулята, да и вообще столь древнего южноамериканского млекопитающего,
основана на экземплярах рода Thomashuxleya (рис. 24), собранных моей первой
экспедицией в Патагонию в 1930 — 1931 гг. Некоторые другие представители
этого семейства были столь же или даже более многочисленны и изучены довольно
хорошо, но не по целым скелетам. Все изотемниды были все еще весьма примитивны.
Фактически семейство трудно охарактеризовать по каким-либо признакам, кроме
примитивных, а именно таких признаков при построении системы желательно
по возможности избегать. Их зубы отличаются от зубов еще более примитивных
форм, например Henricosbornia, наличием на верхних молярах одного или нескольких
добавочных гребешков. Различие между ними состоит также в том, что у изотемнид
довольно характерные клыки, более похожие на настоящие клыки, но в этом
отношении данное семейство, весьма вероятно, еще примитивнее, чем даже
генрикосборнииды. Изотемниды были также крупнее других нотоунгулят, существовавших
в древних фаунах. Они отличаются от некоторых современных им и практически
от всех более поздних нотоунгулят именно своей большей примитивностью.
Это распространяется и на их конечности, у которых примерно такие же пропорции,
как у остальных примитивных копытных, и по пяти функционирующих пальцев
на каждой конечности, из которых средний (III) немного крупнее остальных.
Здесь, как и у некоторых других примитивных нотоунгулят, наблюдается значительная
внутривидовая изменчивость, и это нашло отражение в системе Амегино. Например,
то, что сейчас считают одним изменчивым видом Isotemnus primitivus (удачное
видовое название, выбранное Амегино), Амегино разбил на семь видов, относящихся
к пяти родам, хотя остатков этой формы в сборах было не особенно много;
однако всех их он считал представителями одного и того же семейства.
Другое семейство, Notohippidae, появляется в мастэрсии неожиданно; у него
нет твердо установленных предшественников, хотя оно, возможно, происходит
от довольно ранних изотемнид. Наиболее отчетливое его отличие от современных
ему изотемнид состоит в том, что хотя его моляры и имеют корни, их коронки
значительно выше. В последующие века коронки продолжали становиться все
выше, и у слабо изученного сантакрусского рода Notohippus — типового рода
данного семейства — были покрыты толстым слоем цемента. В этом отношении
его моляры приобрели сходство с зубами прогрессивных (травоядных) лошадей
(рис. 25), в том числе всех ныне живущих представителей сем. лошадиных.
Название, данное этому роду Амегино, означает по-гречески «южная лошадь».
Амегино после некоторых колебаний все же решил, что ранние нотохиппиды
были предками лошадей. На самом деле их сходство лишь отражает конвергентную
эволюцию, обусловленную адаптацией к питанию травами на пастбищах. Кроме
высоты коронок и чехла из цемента моляры имеют все диагностические признаки
нотоунгулят и ни одного признака лошадей; то же самое относится и к ушной
области.
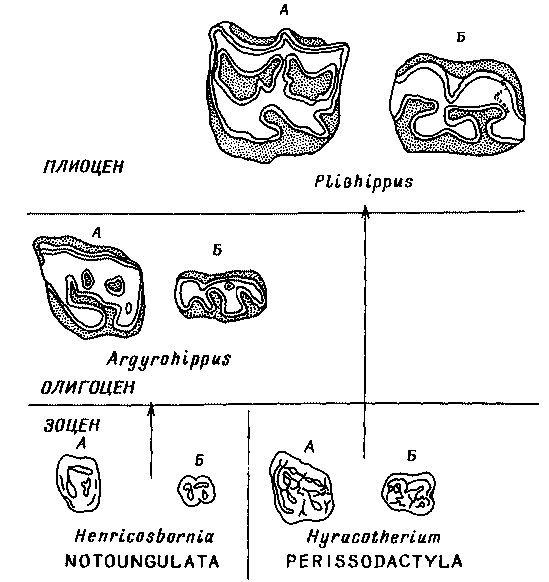
Рис. 25. Левые верхние (А) и правые нижние (В) моляры некоторых нотоунгулят,
местных копытных Южной Америки, и непарнокопытных, происходящих из северного
полушария.
Моляры Henricosbornia из нижнего эоцена (касамайория) имели примерно такое
же примитивное строение, какое было характерно для нотоунгулят в целом,
а моляры Hyracotherium из раннего эоцена Северной Америки и, Европы — примитивное
строение, характерное для непарнокопытных вообще и для сем. лошадиных (Equidae)
в частности. У Argyrohippus — нотохиппида из раннего олигоцена
(десеадия) — уже были развиты высококороиковые жевательные зубы, покрытые
слоем цемента
(на рисунке показан точками) и приспособленные для перетирания жестких
растений. У некоторых лошадей в процессе эволюции развиваются перетирающие
зубы, подобные изображенным здесь зубам Pliohippus из Северной Америки,
но лошади достигают этой стадии значительно позднее, чем южноамериканские
нотохиппиды, и несомненно приходят к этому совершенно независимо.
Сем. Leontiniidae, которое Флорентино Амегино трогательно назвал в честь
своей жены и помощницы, урожденной Леонтины Пуарье, — типично десеадская,
т. е. раннеолигоценовая, группа, хотя ее представители известны и из более
позднего колуапия. Леонтинииды — обычно самые заметные млекопитающие в
десеадии, хотя Амегино назвал эти слои по пиротерию. Pyrotherium приурочен
только к десеадским слоям, но его остатки нигде не встречаются в большом
количестве, а в некоторых богатых десеадских фаунах и вовсе отсутствуют.
У леонтиниид сохраняется полная примитивная зубная формула 3/3•1/1•4/4•3/3;
зубы, низкокоронковые и довольно простые по строению, имеют корни и образуют
непрерывный ряд с постепенным переходом от одного типа к другому. Клыки
маленькие, но первый верхний и третий нижний резцы увеличены в размере
и напоминают клыки или небольшие бивни. Голова тяжелая, тело высокое в
плечах. У Scarrittia, наиболее хорошо изученного представителя
этого семейства, передние конечности четырехпалые, палец I отсутствует,
палец V редуцирован
и не функционирует, пальцы II — IV функционирующие, а ось конечности проходит
через палец III. Задняя конечность пятипалая, но пальцы I и V редуцированы
и, очевидно, нефункционирующие; пальцы II — IV устроены примерно так же,
как на передней конечности (см. рис. 22, Б). Единственный хорошо известный
представитель сем. Homalodotheriidae — это род Homalodotherium (рис. 26),
известный также под неверным названием Homalodontotherium. Этот восьмисложный
термин выглядит монстром даже в палеонтологии, перегруженной массой многосложных
названий. Как то, так и другое название по-гречески означает «равнозубый
зверь» и отражает наличие у этого животного полного набора плотно примыкающих
друг к другу зубов, различные типы которых связаны постепенными переходами.
Строение зубов, несмотря на ряд мелких различий, в общем такое же, как
у леонтиниид; череп и скелет, за исключением конечностей, также весьма
сходны с таковыми у Scarrittia, хотя более мощные. Однако их конечности
различаются очень сильно. На передних конечностях по четыре крупных функционирующих
пальца, II— V, примерно одинаковой длины, причем каждый из них несет не
копыто, а большой коготь. Первый палец рудиментарен и совершенно не функционирует.
На задних конечностях также по четыре функционирующих пальца, II — V, причем
все они короче и массивнее соответствующих пальцев передней конечности;
особенно массивен короткий, похожий на обрубок пятый палец. Остается неясным,
несли ли они когти; если да, то эти когти были, вероятно, меньше, чем на
передних конечностях. Передние конечности были несомненно пальцеходящими,
т. е. животное при передвижении опиралось на основания пальцев; задняя
же конечность была стопоходящей, т. е. опиралась на землю всей стопой.
Вследствие такого различия в положении передних и задних конечностей плечевой
пояс был более приподнят над землей, чем тазовый.
Хомалодотерии появляются в палеонтологической летописи в то же самое время,
что и леонтинииды, но сохраняются дольше, вплоть до часикия. Homalodotherium известен из сантакрусия. Едва ли можно сомневаться в том, что эти два семейства
имели общее происхождение; вероятно, их предком был какой-то неизвестный
нам додесеадский леонтиниид, поскольку из этих двух семейств леонтинииды
явно более примитивны.
Для того чтобы представить себе, что животное, несомненно относящееся к
копытным (Ungulata), имело когти, нужно обладать изрядной долей фантазии;
тем не менее этот случай не единичен. Для периода времени, перекрывающегося
с временем существования в Южной Америке хомалодотериев, известна обширная
группа когтистых Ungulata, названных халикотериями и принадлежащих к тому
же отряду Perissodactyla, что и лошади. Халикотерии были распространены
по всем континентам, кроме Южной Америки, Австралии и Антарктиды. С ними
связан знаменитый эпизод в истории палеонтологии. Кювье, считающийся основателем
научной палеонтологии (хотя у него были менее известные предшественники),
был убежден в том, что между всеми частями данного организма имеется такая
тесная корреляция, что по одной части можно предсказать, какой должна быть
другая, в частности по одному зубу можно предсказать строение пальцев.
Этот вывод, который иногда называют законом Кювье, породил миф о том, что
палеонтолог может восстановить и восстанавливает облик целого животного
по одному зубу или кости (см. приложение). Открытие халикотериев заставило
отказаться от закона Кювье, во всяком случае палеонтологов; к сожалению,
многие непалеонтологи все еще продолжают в него верить. Согласно этому
закону, животное с халикотериевыми зубами должно было иметь копыта и никак
не могло иметь когтей. Вскоре было обнаружено, что у них действительно
были когти.

Рис. 26. Реконструкции раннемиоценового (сантакрусского) представителя рода Homalodotherium — странного «копытного» с когтями, относящегося к подотряду Toxodonta отряда Notoungulata.
Теперь уже не покажется удивительным, что Амегино считал примитивных хомалодотериев
(он называл их «хомалотериями») предками халикотериев. Последнее слово
в этом вопросе принадлежит У. Б. Скотту. Он писал, что изучение некоторых
халикотериев Северной Америки «убедительно показывает, что эти животные
— всего-навсего аберрантные непарнокопытные и не могут быть родственны
своим южноамериканским аналогам» (Scott, 1930, с. 331). Итак, снова адаптивная
конвергенция, но опять-таки адаптивная по отношению к чему? Поскольку животных,
подобных этим, в настоящее время вообще не существует, воображение порождало
всевозможные гипотезы в отношении как халикотериев, так и хомалодотериев.
Э. С. Риггс (Е. S. Riggs), собравший самые полные экземпляры Homalodotherium,
в 1937 г. с теми же основаниями, как и все прочие, рассуждал, что мощные,
снабженные когтями передние конечности этого животного «были хватательными,
способными в поисках пищи рыть землю или пригибать ветви деревьев для добывания
плодов или листьев».
Наконец, в нашем рассказе о нотоунгулятах мы подходим к величественному
семейству Toxodontidae. Первым известным ископаемым остатком этого семейства
и вообще первым остатком животных, которых много позднее стали называть
нотоунгулятами, был череп, купленный Дарвином 26 ноября 1833 г. за 18 пенсов
у фермера в Уругвае (носившем тогда название «Banda Oriental»). 19 апреля
1837 г., спустя несколько месяцев после возвращения Дарвина из путешествия
на «Бигле», Ричард Оуэн сделал на заседании Лондонского геологического
общества доклад об этом черепе. Он дал этому животному название Toxodon и, сравнив его с огромным множеством прочих млекопитающих, сделал вывод,
что это животное «служит доказательством наличия еще одной ступени в ряду
млекопитающих, ведущем от Rodentia через Pachydermata к Cetacea» (цит.
по Darvin, 1839, с. 180; см. литературу к гл. 2). Это замечание не следует
воспринимать как указание на эволюционную последовательность. В то время
Дарвин еще только становился эволюционистом и не высказывая своих взглядов
открыто; откровенным эволюционистом он стал лишь спустя примерно 20 лет,
а Оуэн не стал им никогда. Место в системе, отведенное Оуэном роду Toxodon,
кажется совершенно нелепым; однако нельзя забывать, что он исходил из существования
scala naturae, или «великой цепи бытия», лежащей в основе божественного
созидания, и что он помещал Toxodon на этой лестнице или цепи между Rodentia
и Pachydermata, к которым в те времена относили любых крупных четвероногих
копытных млекопитающих с толстой кожей. В сущности это было четкое указание
на адаптивные признаки рода, не имевшее, конечно, никакого отношения к
его филогенетическому положению.
Название Toxodon вполне уместно, поскольку оно означает «дугозуб», а верхние
моляры у этого рода имеют очень высокие коронки, сильно изогнутые в вертикальной
плоскости, что делает их похожими на дугу. Приведем еще один забавный исторический
штрих. Череп, который Дарвин приобрел за 18 пенсов (хотя эта сумма по тем
временам была значительной, но тем не менее сама сделка оказалась одной
из выгоднейших в истории), был без зубов. Как рассказывает Дарвин, дети
фермера «выбили зубы камнями, а затем использовали голову в качестве мишени».
Но в 180 милях от фермы Дарвин «нашел прекрасный зуб, точно соответствовавший
одной из ячеек в этом черепе», который и заставил Оуэна дать черепу название
Toxodon. Поместил же он их между Rodentia и Pachydermata, потому что две
пары верхних и три пары нижних резцов (верхние определялись по ячейкам
в типовом черепе) имели увеличенные размеры, обладали постоянным ростом
и отдаленно напоминали зубы грызунов, тогда как общие размеры («как у гиппопотама»,
отмечал Дарвин) и общий облик были, как тогда говорили, пахидермного типа.
Полная зубная формула Toxodon следующая: 2/3•0/1•4/3•3/3, все зубы постоянно
растущие, гипселодонтные. Как и в других подобных случаях, бугорки на коронках
вскоре снашиваются, а получающийся при этом рисунок
поперечного сечения моляров удивительно специфичен у разных родов и видов.
Нижние моляры не изогнуты в виде дуги, а в вертикальной плоскости почти
прямые. Рисунок жевательной поверхности после изнашивания может оказаться
довольно занятным, о чем свидетельствует название Gyrinodon, «головастикозуб»,
данное одному из менее обычных родов этого семейства. Череп у токсодонта
был тяжелый, с короткими носовыми костями, что заставляет предполагать
наличие у него подвижного мясистого носа; (рис. 27); скелет был также тяжелым,
мощным, приземистым, с небольшим горбом на уровне плеч; передние конечности
были короче задних. И передние, и задние конечности были трехпалыми (II
— IV), все пальцы были функционирующими, причем третий был самым длинным
и лежал по оси конечности.
Первые известные токсодонтиды происходят из десеадия, но, как и некоторые
другие группы, по-настоящему хорошо известны они только начиная с сантакрусия.
Из этих отложений описаны полные скелеты двух родов. Виды обоих родов были
значительно мельче, чем виды Toxodon, и имели примитивную зубную формулу:
3/3•1/1•4/4•3/3.
Все зубы имели очень высокие коронки, но в конце концов образовывали корни
и прекращали рост. Медиальная пара верхних резцов и самая задняя, третья,
пара нижних уже увеличились в размере и до некоторой степени напоминали
резцы грызунов. Моляры сохраняли лишь слегка измененный нотоунгулятный
тип строения на всех стадиях изнашивания, кроме самых поздних.
На более поздних этапах кайнозоя, от фриасия до монтеэрмосия, от этого
семейства отошла любопытная боковая ветвь, представители которой несли
на лбу один рог, подобный носорожьему. Сам этот рог, не имевший костного
стержня, не сохранился в ископаемом состоянии, но, как и у ныне живущих
носорогов, на его наличие ясно указывает шероховатый бугор на лежавших
под ним костях.
По крайней мере некоторые из более поздних токсодонтид, возможно, вели
частично земноводный образ жизни, и, по-видимому, претерпели некоторую
адаптивную конвергенцию в направлении как носорогов, так и бегемотов. С
некоторым удивлением приходится отметить, что Амегино не считал их предками
ни одной из этих групп, а выделил в самостоятельный отряд Toxodontia, не
давший никаких потомков; они и сейчас сохраняют самостоятельное положение
в системе в качестве подотряда Toxodonta.
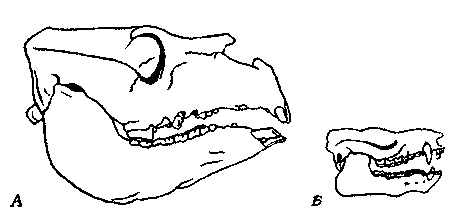
Рис. 27. Черепа одного из последних представителей и одного из древнейших
членов подотряда Toxodonta отряда Notoungulata.
А. Toxodon, поздний плейстоцен. Б. Thomashuxleya, ранний эоцен. Черепа
изображены в одном и том же масштабе. Различия в размерах и специализации
формы черепа и зубов очевидны.
ЛИТЕРАТУРА
Chaffee R. G. 1951. The Deseadan vertebrate fauna of the Scarritt pocket,
Patagonia. Bulletin of the American Museum of Natural History, 98,
503 — 562 (Содержит наиболее полное описание леонтиниида Scarrittia).
Lavocat R. 1958. Notoungules. In: J. Piveteau, ed., Traite de Paleontologie.
tome 6, 2, 60 — 121, Paris, Masson
Riggs E. S. 1937. Mounted skeleton of Homalodotherium. Geological Series
of Field Museum of Natural History, 6, 233 — 243.
Scott W. B. 1912. Toxodonta of the Santa Cruz beds. Entelonychia of the
Santa Cruz beds. Reports of the Princeton University Expeditions to Patagonia,
6, 111 — 300.
Scott W. B. 1930. A partial skeleton of Homalodontotherium. From the
Santa Cruz beds of Patagonia. Field Museum of Natural Nistory, Geology.
Memoirs
1, 7 — 33.
Sinclair W. J. 1909. Typotheria of the Santa Cruz beds. Reports of the
Princeton Expeditions to Patagonia, 6, 1 — 110.
Stirton R. A. 1953. A new genus of interatheres from the Miocene of Colombia
University of California Publications in Geological Sciences, 29, 265
— 348.
См. также ссылки, приведенные выше, в особенности на работы Паттерсона
и
Паскуаля (1972), Скотта (1937) и Симпсона (1948, 1967).
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ...............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
................................................ 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ .....................................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .............................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
...............................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ........................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252