ГЛАВА 7
СУМЧАТЫЕ
Австралия с давних пор известна как страна сумчатых, но и Южная Америка
тоже была страной сумчатых. Сумчатые стали особенно многочисленны и разнообразны
именно на этих двух континентах по одной и той же причине. В Австралии в
раннем кайнозое, по всей вероятности, не было плацентарных млекопитающих.
Сумчатые претерпели здесь адаптивную радиацию, позволившую им освоить многие
из тех экологических ниш, которые на большинстве других континентов были
заняты плацентарными. На значительно более поздних этапах кайнозоя в Австралию
проникли плацентарные грызуны сем. мышиных (Muridae). Возможно, что они сдерживали
эволюцию австралийских сумчатых в направлениях, конвергентных эволюции грызунов;
только вомбаты, в сущности мало похожие на мышей, приобрели адаптивное сходство
с некоторыми крупными грызунами, обитающими в других областях земного шара.
Единственное, кроме мышиных, местное наземное плацентарное Австралии — это
динго; но это почти наверное одичавшая форма полуодомашненных собак, завезенных
аборигенами. Конечно, нельзя считать простым совпадением, что после того,
как динго широко распространились на австралийском континенте, там вымер
так называемый тасманийский волк, или точнее тилацин — сумчатое, наиболее
сходное с собаками по внешнему виду и образу жизни. Несколько тилацинов уцелели
на острове Тасмания; возможно, что они живут там еще и до сих пор, хотя в
последнее время их никто не встречал.
В Южной Америке, как уже отмечалось, ранние сумчатые жили бок о бок с плацентарными,
но последние были представлены там лишь причудливыми ксенартрами и многочисленными
копытными. Особенно заметно отсутствие здесь плацентарных насекомоядных,
грызунов и хищников, которых в Северной Америке, Евразии и Африке было очень
много. Таким образом, и в Южной Америке сумчатые претерпели адаптивную радиацию,
что позволило им занять — в отношении образа жизни — те места, которые на
северных континентах и в Африке занимали плацентарные млекопитающие. Как
станет ясно из дальнейшего, плацентарные грызуны достигли Южной Америки и
притом довольно рано. Вероятно, именно по этой причине начинавшаяся эволюция
сумчатых, действительно похожих на грызунов, была ограничена.
По консервативной системе, используемой в этой книге (см. гл. 4), известные
южноамериканские сумчатые делятся на 8 семейств, условно объединяемых в 4
надсемейства. Одно семейство, Necrolestidae, не отнесено здесь ни к одному
из надсемейств. Единственный известный в нем род, Necrolestes, можно было
бы выделить в самостоятельное надсемейство, но в настоящее время это сомнительно.
Некоторые исследователи выделяют сверх этих восьми еще несколько семейств
южноамериканских сумчатых. Наше отношение к такому распределению и причины,
по которым оно здесь не было принято, излагаются в соответствующем месте
книги.
История сумчатых Южной Америки начинается, как и следовало ожидать, с сем.
Didelphidae — опоссумов в широком смысле слова. Это древнейшие сумчатые,
известные в Южной Америке и вообще на Земле; наиболее примитивные члены этого
семейства близки к предкам или даже являются предками всех остальных южноамериканских,
а, возможно, также и австралийских сумчатых. По своему обилию в Южной Америке
в наши дни дидельфиды не идут ни в какое сравнение с другими сумчатыми, из
которых там сохранилось еще только одно семейство, Caenolestidae, содержащее
по наиболее дробной классификации всего три рода. Ведущий исследователь дидельфид
Освальдо Рейг (Osvaldo Reig) различает в современной фауне 14 родов и 71
вид. Другим исследователям такая система кажется непомерно раздутой, но никто
не сомневается в том, что ныне живущие дидельфиды весьма многочисленны и
в высшей степени разнообразны. Они населяют огромную область с весьма разнообразными
условиями обитания: почти всю Южную Америку и значительную часть юга Северной
Америки. Виды и роды имеют ограниченные ареалы, к которым они специфически
приспособлены; их число в каждой отдельной местности, конечно, гораздо ниже,
чем приводимое для области в целом.
Примитивные дидельфиды были преимущественно всеядными, как и большинство
ныне живущих форм, которые могут питаться насекомыми, червями, улитками,
ракообразными, яйцами, амфибиями, рептилиями, птицами, млекопитающими, падалью,
а также плодами и другой сочной растительной пищей, причем столь интенсивно,
что в некоторых районах они даже наносят серьезный вред урожаям. Одни виды
потребляют больше животной, а другие больше растительной пищи; у некоторых
вымерших родов специализация достигала крайней степени. Из ныне живущих дидельфид
экологически наиболее своеобразен водяной опоссум, или плавун (Chironectes
minimus) — хищник, живущий в воде и питающийся в основном раками и рыбой.
Chironectes означает по-гречески «рукоплав», хотя животное использует при
плавании главным образом свои задние лапы, снабженные перепонками. Когда
этот вид впервые был обнаружен, его по ошибке приняли за выдру.
В гл. 6 уже говорилось о необычайном разнообразии дидельфид в риочикской
фауне Итабораи в Бразилии. Создается впечатление, будто эти животные, изолированные
на континенте, где не было им подобных, как бы пытались экспериментировать
с зачатками самых разнообразных экологических специализаций. Менее антропоморфное
объяснение состоит в том, что они подверглись дизруптивному естественному
отбору, действовавшему вначале на многочисленные мелкие разобщенные и периферические
популяции; а затем те популяции, которые преуспели, расселились так широко,
что многие из них стали занимать перекрывающиеся территории (по научной терминологии
— становились симпатрическими), как в Итабораи в риочикское время.
В касамайории кроме по крайней мере одного вполне обычного дидельфида существовал
еще один, более своеобразный, которого Флорентине Амегино назвал Caroloameghinia в честь своего брата — сборщика ископаемых. Основанием для этой дани одного
брата другому были две неполные нижние челюсти с несколькими зубами и одна
неполная верхняя челюсть, также с небольшим числом зубов. (Впоследствии я
восстановил справедливость, назвав другое ископаемое того же возраста из
Патагонии Florentinoameghinia.) Интерпретация, которую дал этой находке дон
Флорентине, была, как и обычно, и верной, и неверной: он правильно установил
происхождение рода от Didelphidae, но совершенно неверно истолковал его сходство
с плацентарными копытными, которых он считал потомками Caroloameghinia. По
таким признакам, как отсутствие режущих гребней на зубах и несколько большее
развитие основных бугорков примитивных сумчатых, это животное действительно
сходно с наиболее примитивными копытными, какими-то кондиляртрами, но это
лишь адаптивное и конвергентное сходство. Caroloameghinia был, конечно, сумчатым,
пусть менее плотоядным, чем большинство дидельфид, но настолько близкородственным
последним, что теперь его с общего согласия выделяют не в особое семейство,
как это делал Амегино и другие долгое время после него, а лишь в подсемейство
Caroloameghiniinae сем. Didelphidae.
Следующая фауна, из которой хорошо известны дидельфиды, — сантакрусская (ранний
миоцен), хотя они встречаются и в промежуточных фаунах. Довольно обычен в
сантакрусии один из дидельфид, которого Амегино назвал Microbiotherium. Это
название означает «коротко живущее млекопитающее». Нет данных, позволяющих
считать, что продолжительность жизни у этого животного была короче, чем у
других млекопитающих таких же размеров, но нет и правила, которое бы требовало,
чтобы научные названия животных имели определенный смысл; достаточно, чтобы
названия отличались друг от друга и чтобы было ясно, к чему именно они относятся.
Амегино понял, что Microbiotherium родствен дидельфидам, но выделил его в
самостоятельное семейство. Наиболее явное его отличие состоит в том, что
маленькие внешние (щечные) бугорки верхних моляров развиты у него слабее,
чем у дидельфид. В 1906 г. Синклер (Sinclair) — первый исследователь, занявшийся
этим родом после Амегино, — решил, что он относится к сем. Didelphidae; так
же думали и большинство более поздних исследователей, сохраняя в то же время
подсемейство Microbiotheriinae. Это подсемейство долгое время считали вымершим,
но еще в 1955 г. Рейг указал, что ныне живущий род Dromiciops очень сходен
с Microbiotherium и, возможно, даже идентичен ему. С тех пор он рассматривает
Microbiotheriidae как семейство, отличное от Didelphidae и представленное
в наши дни родом Dromiciops = Microbiotherium (?), a возможно, и некоторыми
другими родами.
Вопрос этот может показаться несущественным, чем-то вроде бури в таксономическом
стакане воды. В какой-то мере так оно и есть, но тем не менее этот вопрос
имеет отношение к некоторым более общим проблемам. Не так уж удивительно,
что род млекопитающих, впервые описанный и названный по ископаемым остаткам
и считавшийся вымершим, вероятно, 10 тыс., а может, и миллион лет назад,
на самом деле существует и поныне. Знаменитый среди зоологов пример — один
из родов мелких австралийских сумчатых, который полагали вымершим по крайней
мере 10 тыс. лет назад. Роберт Брум (Robert Broom) назвал его Burramys на
основании ископаемых костей, обнаруженных им в пещере в 1894 г. В 1966 г.
один из этих зверьков был найден в хижине для лыжников в так называемых (явное
преувеличение) «австралийских Альпах» и, как пишут, оказался настроенным
весьма дружелюбно. Dromiciops был открыт и получил название в 1894г., спустя
всего семь лет после Microbiotherium, поэтому сделанное в 1955 г. замечание,
что Dromiciops — это возможно доживший до наших дней микробиотерий, не столь
сенсационно, как история буррамиса. Однако в одном отношении наш случай,
возможно, еще более примечателен: если признать, что род Dromiciops идентичен
роду Microbiotherium, то, следовательно, данный род как таковой существовал
на протяжении примерно 20 млн. лет. Это может быть мировой рекорд для рода
млекопитающих.
Наш пример затрагивает еще одну проблему, имеющую более широкое значение.
Дело в том, что некоторые специалисты относят род Microbiotherium (вместе
с Dromiciops или без него) к подсемейству Didelphinae, другие (таких, вероятно,
в настоящее время большинство) выделяют его в отдельное подсемейство Microbiotheriinae
сем. Didelphidae, а третьи — в самостоятельное ceм. Microbiotheriidae. Когда
какая-либо группа организмов получает таксономическое название и становится
тем, что на научном языке называют таксоном, то суждение о том, является
ли эта группа родом, подсемейством, семейством или таксоном какого-нибудь
другого ранга, нельзя рассматривать как «верное» или «неверное». Хотя для
определения ранга таксонов предложены довольно конкретные критерии, ни один
из них нельзя использовать как незыблемое правило. Обычно на решение этого
вопроса влияют установившаяся практика или традиции, но в каждом отдельном
случае оно зависит в общем от личного мнения, вкуса и суждения автора о том,
насколько это удобно, гармонично и ясно.
Что касается данного примера, то я временно поместил Microbiotherium в подсемейство
Didelphinae. Это мое суждение основывается на приводимом Рейгом перечне и
анализе многих признаков современных и ископаемых опоссумов и их родичей,
хотя сам Рейг выделяет данную группу в отдельное семейство. Ни у Microbiotherium,
ни у Dromiciops нет ни одного известного нам признака, который не встречался
бы также у родов, относимых самим Рейгом к подсемейству Didelphinae. Но у
этих двух родов имеются сочетания признаков, не идентичные сочетаниям, встречающимся
у других родов Didelphidae. Именно так определяются роды и родовой ранг:
для каждого рода характерно свое особое сочетание признаков, которые встречаются
независимо в разных сочетаниях то там, то тут в пределах семейства. По моему
мнению в данном случае ранг подсемейства или более высокий не оправдан: такие
соображения приложимы и к другим случаям установления ранга в систематике.
Один исследователь подобных проблем однажды цинично заметил, что некоторые
систематики, по-видимому, думают, что если вы можете отличить двух животных
друг от друга, то значит, они относятся к разным родам, а если нет — то к
разным видам.
Помимо продолжения линии Didelphinae, приведшей к их современным процветающим
популяциям, в позднем кайнозое известна другая группа родичей опоссумов,
которая бесспорно заслуживает выделения по крайней мере в особое подсемейство,
хотя не удивительно, что иногда эту группу выделяют в самостоятельное семейство.
Здесь она рассматривается как подсемейство дидельфид, названное Sparassocyninae
по его единственному в настоящее время четко очерченному роду Sparassocynus.
Представители этого подсемейства встречаются в интервале времени от уайкерия
до укия, или, другими словами, приблизительно от раннего плиоцена до раннего
плейстоцена, но лучше всего оно известно по великолепно сохранившимся черепам
и нижним челюстям чападмалальского возраста, найденным в обрывах на берегу
моря около города Мар-дель-Плата в Аргентине. Коренные зубы у его представителей
были более «хищническими», чем у других дидельфид, что выразилось в развитии
мощных режущих гребней и редукции более обычной для дидельфид комбинации
гребней с дробящими и перетирающими участками. Еще более своеобразна ушная
область: среднее ухо разделено на две большие полости — нижнюю и верхнюю
(нижнебарабанную и верхнебарабанную), окруженные костными стенками. Эти животные
были мелкими, в высшей степени хищными плотоядными и занимали экологическую
нишу, свободную в то время от других южноамериканских млекопитающих.
Другую группу плотоядных млекопитающих, существовавших в Южной Америке начиная
по крайней мере с риочикия и вымерших примерно в конце плиоцена, относят
к сем. Borhyaenidae (рис. 7). Боргиениды, очевидно, произошли от ранней ветви
дидельфид, которая перешла к менее всеядному и более хищному типу питания;
предполагается, что они по большей части охотились на исключительно многочисленных
местных копытных. Их типичные представители относились к догоняющим хищникам,
но в процессе дивергенции среди них возникло много форм, различающихся по
величине, весу и морфологическим признакам. Амегино сгруппировал боргиенид
в семь разных семейств, но теперь все согласны с тем, что их следует объединить
в одно семейство. Однако они и в самом деле достигли такого разнообразия,
что последний исследователь боргиенид, Л. Г. Маршалл (L. G. Marshall), разделяет
их на четыре подсемейства, которые хорошо различимы и по которым таксоны
распределены обоснованным образом.
Члены подсемейства Hathlyacyninae сохраняли размеры от мелких до средних
и, очевидно, были менее плотоядными и хищными, чем большинство их более крупных
родичей. Маршалл полагает, что некоторые из них вели полудревесный образ
жизни. Все Proborhyaeninae и Borhyaeninae были крупными наземными хищниками,
отличаясь друг от друга по некоторым анатомическим признакам. Prothylacininae
также были крупными животными, одни, очевидно, полностью плотоядными, другие,
вероятно, почти всеядными; возможно, что это был вторичный переход. ко всеядности
у форм, происходивших от плотоядных предков, подобно тому, как это (в еще
большей степени) произошло с медведями, если говорить о плацентарных хищниках.

Рис. 7. Реконструкция Prothylacinus — раннемиоценового (сантакрусского) сумчатого хищника из сем. Borhyaenidae.
По общему облику и несомненно по образу жизни многих видов эти животные
несколько походили на собак. В меньшей степени они обладали внешним сходством
с кошками, однако, по всей вероятности, они несли при этом некоторые из экологических
функций, выполнявшихся на других континентах кошками. Эта группа стала классическим
примером эволюционной конвергенции, классическим еще и потому, что, несмотря
на поразительное адаптивное сходство, ни один квалифицированный морфолог
не смог бы принять какого-либо из этих сумчатых хишников за плацентарного
хищника. Амегино признал их своеобразие и поместил их в отдельный отряд (назвав
его Sparassodonta); он полагал, что этот отряд происходит от примитивных
опоссумов, что по современным представлениям верно, но опять-таки ошибся
в отношении их потомков: он считал их предками плацентарных хищников.
Сходство боргиенид с тасманийским волком, или тилацином, Австралийской области,
больше, чем с любым из плацентарных хищников. Это сходство столь велико,
что даже хороший морфолог мог бы принять его за указание на особо тесное
родство между ними, более тесное, чем могла бы обусловить принадлежность
всех этих животных к очень обширной группе Marsupialia. Синклер, который
был способным морфологом, отнес всех животных, включенных по нашей системе
в сем. Borhyaenidae, к австралийскому сем. Thylacinidae. В течение ряда лет
это принималось за доказательство особой и тесной связи между Южной Америкой
и Австралией. Несмотря на некоторые разногласия и остатки сомнений на этот
счет, представляется в высшей степени вероятным, что боргиениды и тилациниды
развивались независимо, каждые на своем континенте. То обстоятельство, что
сходство между ними больше, чем во многих других несомненных примерах конвергенции,
объясняется происхождением и той, и другой группы от примитивных дидельфидных
или дидельфидоподобных предков. Таким образом, здесь налицо не только конвергенция,
но и элемент параллельной эволюции.
Южноамериканские сумчатые сем. Thylacosmilidae дают еще один классический,
очевидный и даже еще более поразительный пример конвергенции. У Thylacosmilus (рис. 8) из уайкерия северной Аргентины были большие саблевидные клыки; по
всему облику черепа, челюстей и зубов он поразительно напоминает знаменитых
плацентарных «саблезубых тигров», которые были кошками в широком смысле,
но не были собственно тиграми. Допустимо предположить, что Thylacosmilus развился от какой-то ранней ветви Borhyaenidae, но появляется он в палеонтологической
летописи внезапно, и его предки пока не известны или еще не распознаны.
Второе ныне живущее семейство южноамериканских сумчатых теперь почти всегда
называют Caenolestidae. Относительно номенклатуры этого семейства и его подсемейств
существуют некоторые сомнения, но в данном контексте это не имеет значения,
и мы будем придерживаться общепринятых названий. Первая заметка о представителе
этого семейства была опубликована в Англии в 1860 г., позднее с некоторыми
интервалами было поймано еще несколько экземпляров. Их считали большой редкостью,
и едва ли кто-нибудь видел их в природе, кроме неграмотных туземцев, пока
в конце 1960-х годов Дж. А. Кирш (J. A. W. Kirsch), исследователь австралийских
и американских сумчатых, не провел более тщательные поиски и не обнаружил,
что некоторые из этих животных довольно обычны в отдаленных главным образом
горных районах — местах, где они водятся. Сейчас принято объединять ныне
живущих представителей этого семейства в три рода, в одном из которых пять
видов, а в двух других — по одному. Они не имеют настоящих народных названий.
По-английски их обычно называют rat opossums («крысы-опоссумы»), но это выдуманное
книжное имя, и к тому же бессмысленное, поскольку они и не крысы, и не опоссумы
и не обладают близким сходством ни с теми, ни с другими. По-испански их иногда
называют ratones runchos, т. е. «мыши-опоссумы», в равной степени нелепое
название. Runcho — название опоссума на колумбийском диалекте, которое не
понимают в других испаноязычных странах.
Моляры этих животных отличаются от примитивного для сумчатых типа более четырехугольной
формой, но самая примечательная черта зубной системы — наличие двух очень
крупных нижних резцов, направленных почти прямо вперед. Сходные увеличенные
и направленные вперед нижние резцы имеют многие австралийские сумчатые. По
этому признаку большую группу австралийских форм, в том числе кенгуру и родственных
им животных, часто называют Diprotodonta — «имеющие два первых (или передних)
зуба».
А |
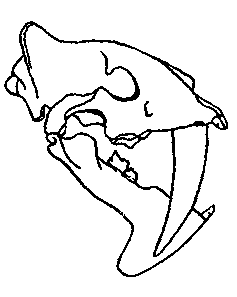 Б |
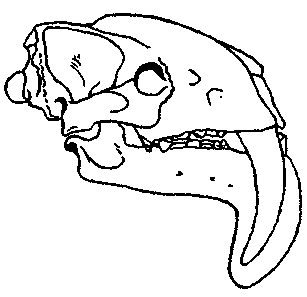
Рис. 8. A. Thylacosmilus — раннеплиоценовый (уайкерский) сумчатый саблезуб (сем. Thylacosmilidae). Б. Smilodon — плейстоценовый плацентарный саблезуб (сем. Felidae, подсемейство Machairodontinae).
На протяжении ряда лет исследователи этих животных полагали, что ценолестиды
и австралийские дипротодонты находятся в особом близком родстве; это была
еще одна предполагаемая связь между двумя континентами. Теперь, однако, все
согласны с тем, что это еще один пример эволюционной конвергенции, такой
же, как и между Borhyaenidae и Thylacinidae, т. е. конвергенции между двумя
группами, которые обе относятся к сумчатым, но имеют разное происхождение.
По образу жизни и питанию ценолестиды обладают еще большим конвергентным
сходством с плацентарными насекомоядными. Подобно последним, ценолестиды
питаются не только насекомыми, но и червями, и другими беспозвоночными, а
возможно, и мелкими позвоночными, если их удается добыть. Кирш обнаружил,
что эти зверьки с удовольствием едят мясо, а когда им приносят новорожденных
грызунов, убивают и поедают их. Он предполагает, что адаптивный отбор в сторону
увеличения нижних резцов был связан не только и не столько с захватыванием
пищи, сколько с закалыванием жертвы. Еще в 1897 г. Амегино правильно указал,
что ныне живущие ценолестиды явно родственны некоторым ископаемым формам,
которых он делил на несколько семейств под другими названиями и помещал в
отдельный отряд Paucituberculata; это название используют также некоторые
современные авторы, делящие сумчатых на несколько отрядов. Синклер относил
всех их к сем. Caenolestidae, разделяя его на три подсемейства. Ныне живущее
подсемейство, Caenolestinae, существовавшее, вероятно, на протяжении всего
кайнозоя, имеет наименее специализированную зубную систему. Теперь эти животные
обитают в районах Южной Америки с относительно прохладным климатом, довольно
высоко в Андах и на острове Чилоэ; возможно, что своим выживанием они обязаны
этой устойчивости к холодному климату. Однако в раннем кайнозое представители
того же подсемейства жили, очевидно, в теплом, даже тропическом климате.
Члены подсемейства Palaeothentinae, известные от десеадия до сантакрусия,
имели увеличенный передний нижний моляр с низким, но хорошо развитым режущим
гребнем. В третьем подсемействе, Abderitinae, передняя часть этого моляра
превратилась в высокое зазубренное лезвие. Сем. ценолестид дает интересный
пример вымирания ветвей, в некоторых отношениях отчетливо более продвинутых
или специализированных, при выживании наименее специализированных представителей
данного семейства. Пример этот не единственный, и некоторые эволюционисты
даже говорили о законе выживания менее специализированных форм, но этот «закон»
имеет много исключений.
В сем. Polydolopidae также имеются два увеличенных и направленных вперед
нижних резца, а у наиболее примитивных форм таких зубов несколько. Полидолопиды
также имеют зуб с сильно развитым режущим лезвием, что прежде рассматривали
как специализацию первого моляра, зашедшую дальше, чем у Abderitinae. Однако
один из самых ранних и примитивных представителей семейства — Epidolops из
риочикия Итабораи — явно свидетельствует в пользу того, что у ценолестид
и полидолопид режущими стали не одни и те же, т. е. не гомологичные, зубы.
Некоторые исследователи впали в крайность, высказав предположение, что эти
два семейства возникли независимо от разных дидельфид. Однако в других отношениях
Epidolops вполне сходен с ценолестидами, причем в большей степени, чем более
поздние, касамайорские, полидолопиды. Это означает, что эти два семейства
действительно имели общего предка, происходящего от дидельфид, и что их объединение
в одно надсемейство филогенетически вполне оправдано. Можно даже допустить,
что режущие зубы у обоих семейств все же гомологичны, хотя это довольно смелое
предположение. В любом случае из этих двух семейств полидолопиды явно более
специализированы, и они вымерли значительно раньше. Полидолопиды довольно
обычно в касамайории, редки в мастэрсии и десеадии, а позже не известны.
Большинство разнообразных полидолопид в адаптивном отношении были вполне
сходны с ценолестидами из подсемейства Abderitinae. Интересно, что эти две
потенциально конкурентные группы в десеадии встречаются совместно, но полидолопиды
после этого времени не известны. Возможно, что они сосуществовали и в додесеадские
времена, но, вероятно, в разных районах; этим, возможно, объясняется также
и то, что нам пока не известны додесеадские абдеритины. Обе эти группы в
какой-то степени сходны с некоторыми из самых мелких австралийских кенгуру,
например с беттонгиями. Последние относятся к дипро-тодонтам, и у них в нижней
челюсти также имеется один зазубренный лезвиевидный зуб. В целом семейство
кенгуру — это в основном растительноядные животные, но есть сообщения, что
беттонгии частично плотоядны и с жадностью поедают мясо, по крайней мере
в неволе. Нет никакого способа выяснить, имели ли вкус к мясу и эти вымершие
южноамериканские сумчатые, несколько похожие на кенгуру. Едва ли они могли
конкурировать с более крупными боргиенидами, но, возможно, конкурировали
с всеядными и частично плотоядными опоссумами, хотя в конечном счете безуспешно.
Следующее семейство сумчатых, которое нам предстоит рассмотреть, — одно из
наиболее странных, даже для этих фаун, в которых сегодня нам почти все кажется
странным. Кроме того, в настоящее время это одно из наиболее редких семейств
млекопитающих в коллекциях, известное только по двум экземплярам. Семейство
носит название Groeberiidae, а оба известных экземпляра относятся к роду
Groeberia, названному Брайеном Паттерсоном в честь Пабло Гробера (Pablo Groeber)
— аргентинца немецкого происхождения, посвятившего большую часть своей жизни
изучению геологии Аргентины, хотя он и не имел прямого отношения к открытию
ископаемых, названных его именем. Оба, экземпляра происходят из одной и той
же геологической формации — слоев дивисадеро-ларго, выходящих в предместье
города Мендоса в Аргентине, но неизвестно, имеют ли они совершенно одинаковый
геологический возраст; разница в размерах взрослых экземпляров столь велика,
что их рассматривают как два разных вида. Оба экземпляра, несмотря на плохую
сохранность, имеют вполне определимые признаки, радикально отличающие их
от любых других известных сумчатых и в сущности вообще от всех известных
млекопитающих.
В нижней челюсти у них имеется одна пара (по одному зубу на каждой стороне),
а в верхней — две пары увеличенных, изогнутых, постоянно растущих резцов,
покрытых эмалью на передней (губной) поверхности. Это бесспорно мощные грызущие
зубы, функционально сходные с резцами грызунов, однако ни одного грызуна
с подобным набором грызущих зубов не существует. Позади этих резцов имеется
диастема, а затем короткий ряд коренных зубов с низкими коронками и замкнутыми
корнями, с мелкими бугорками, расположенными иначе, чем у примитивных сумчатых.
Лицевая часть черепа этих животных была очень короткой и высокой.
Эти сильно специализированные животные появляются в палеонтологической летописи
без всяких предшественников, после чего мгновенно (в геологическом смысле)
исчезают навсегда. Возможная гипотеза, а вернее более или менее обоснованная
догадка заключается в том, что их эволюция протекала в течение некоторого
времени либо в палеонтологически не изученном районе, либо при каких-то особых
обстоятельствах. Некоторое правдоподобие этой догадке придает тот факт, что
вся фауна, в которой они были обнаружены, обладает рядом своеобразных особенностей,
непохожа ни на какие другие и, очевидно, представляет собой некое особое
экологическое сообщество. По возрасту она относится приблизительно к десеадию,
возможно к его началу, или, вероятно, даже несколько предшествует ему. Это
было, очевидно, как раз перед тем, как в Южную Америку проникли плацентарные
грызуны, а возможно, сразу после этого, что порождает еще одну или даже две
догадки. Возникновение этих сумчатых грызунов, вероятно, стало возможным
благодаря тому, что до этого не существовало плацентарных грызунов, которые
могли бы с ними конкурировать, а их вымирание могло быть обусловлено конкуренцией
с более эффективными плацентарными грызунами после появления последних.
До тех пор пока Groeberia была представлена только одним плохо сохранившимся
обломком нижней челюсти, предполагали, что этот род происходит от ранних
ценолестоидов. Но после дальнейшего изучения и дополнительной информации,
полученной по лучше сохранившемуся второму экземпляру, это предположение
стало казаться невероятным. И ценолестиды, и полидолопиды очень сильно отличались
от Groeberia почти по всем своим признакам, не носившим характера примитивных;
их эволюция шла не в том направлении, которое могло бы привести к Groeberia,
а в противоположную сторону. Более вероятно, что гроберииды представляли
собой ветвь, которая рано дивергировала от древнего ствола сумчатых, а затем
специализировалась, но окончательно решить этот вопрос удастся только если
будут обнаружены более древние гроберииды или прегроберииды.
Сем. Argyrolagidae, известное гораздо лучше, чем Groeberiidae, представляется
столь же странным, хотя и совсем в другом плане. Первые экземпляры аргиролагид
были найдены братьями Амегино в Монте-Эрмосо, но это были столь разобщенные
и загадочные фрагменты, что способность Флорентино устанавливать родственные
связи ископаемых форм с их предками (но не с потомками) в данном случае не
сработала. Он сильно ошибся и в отношении предков, и в отношении потомков.
Сейчас об этом легко говорить, но это не означает, что мы умнее братьев Амегино.
Дело просто в том, что мы располагаем великолепной серией ископаемых остатков,
которое не были известны братьям Амегино; они найдены главным образом в обнажениях
на берегу моря вблизи Мар-дель-Плата, столь изобилующих ископаемыми остатками,
и среди них есть даже значительная часть одного скелета аргиролагида в сочленении.
Судя по скелету, эти животные передвигались прыжками на двух больших мощных
двупалых задних конечностях, а их длинный тяжелый хвост играл роль балансира.
Для тех, кто привык иметь дело с черепами различных млекопитающих, череп
аргиролагида представляется совершенно диковинным; его огромные глазницы
сдвинуты так далеко назад, что позади глаз, где у других млекопитающих, в
том числе и у человека, находится часть челюстных мышц, для этих мышц практически
нет места. Странный облик черепа усугубляется тем, что нос продолжается костной
трубкой, выступающей вперед значительно дальше ротового отверстия. Своеобразны
и зубы аргиролагид: у них в каждой челюсти по две пары крупных резцов, причем
в верхней челюсти медиальные резцы несколько мельче и загнуты вниз и назад,
а в нижней челюсти они явно крупнее и загнуты вперед и вверх. Позади резцов
имеется диастема, а дальше как в верхней, так и в нижней челюсти — ряд из
пяти столбчатых зубов, сидящих вплотную друг к другу. Очертания поперечного
сечения зубов несколько варьируют у разных известных видов и родов. Все зубы
постоянно растут по мере стирания, никогда не образуя замкнутых корней.
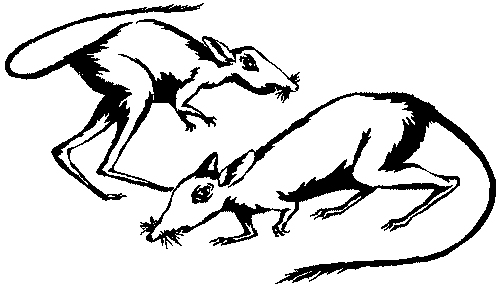
Рис. 9. Реконструкция Argyrolagus — сумчатого из сем. Argyrolagidae, жившего в конце плиоцена — начале плейстоцена и передвигавшегося на задних лапах прыжками.
В Аргентине аргиролагиды известны от уайкерия до укия, т. е. приблизительно
от раннего плиоцена до раннего плейстоцена.
Единственный экземпляр, явно относящийся к тому же семейству, был найден
также в Боливии, в слоях монтеэрмосского или чападмалальского возраста. Аргиролагиды
имеют отчетливо грызуний облик (рис. 9), причем в тех же самых фаунах обнаружено
много плацентарных грызунов. Ни один из них, однако, не имеет большого сходства
с аргиролагидами, которые, очевидно, адаптировались к своеобразной и новой
для Южной Америки экологической нише. Хотя аргиролагид легко распознать по
морфологическим признакам и их принадлежность к сумчатым не вызывает сомнений,
они обладают замечательным конвергентным сходством с некоторыми плацентарными
грызунами других континентов: передвижение на двух задних лапах, длинными
прыжками (сальтаторная или рикошетная система локомоции), часто в сочетании
с другими особыми признаками, например сильно развитыми слуховыми барабанами
в среднем ухе (они имеются и у аргиролагид). Такая система передвижения возникала
неоднократно и независимо в разных группах. В частности, она характерна для
«кенгуровых крыс и мышей», которые не кенгуру, не крысы и не мыши, а грызуны
сем. Heteromyidae*, живущие в Северной Америке, и для Dipodidae — тушканчиков
Африки и Азии. Интересно, что в Австралии такой локомоторной системой обладает
не сумчатое, а местный представитель мышиных (Muridae), относящийся к роду
Notomys. Все это — плацентарные грызуны, но они происходят от разных предков
и относятся к нескольким разным семействам. Большинство их особенно хорошо
чувствует себя в степях и более или менее аридных районах; аргиролагиды тоже,
вероятно, обитали в степи или пампе. В Южной Америке все еще имеются обширные
районы, причем именно в тех областях, где найдены ископаемые остатки аргиролагид,
которые, казалось бы, предоставляют превосходные условия для их существования.
Любопытно, что на этом континенте аргиролагиды вымерли, тогда как их близкие
аналоги из грызунов выжили на других крупных массивах суши. Теперь в Южной
Америке этот адаптивный тип отсутствует. Там живут несколько гетеромиид,
но это — поздние пришельцы, ограниченные самыми северными районами континента;
вероятно, они никогда не соприкасались с аргиролагидами и не замещали их.
| * В русской литературе их принято называть мешотчатыми прыгунами. — Прим. ред. |
Подобно Groeberia и некоторым другим, аргиролагиды появляются в ископаемой
летописи опять-таки неожиданно, не имея известных нам предшественников, и
после геологически короткого промежутка времени исчезают без видимой причины.
Последнее семейство сумчатых, которое будет рассмотрено в рамках принятой
здесь системы этого отряда, — снова высокоспециализированная, чрезвычайно
своеобразная группа; она появляется в палеонтологической летописи внезапно,
без предшественников, и спустя некоторое время, по геологическим масштабам
исключительно короткое, загадочно исчезает. Это сем. Necrolestidae, известное
по единственному роду и виду, ограниченному сантакрусием, т. е. примерно
ранним миоценом. Первые экземпляры были найдены Карлосом Амегино и описаны
Флорентино в 1891 г. под названием Necrolestes patagonensis. Амегино
относил этот вид к насекомоядным, но полагал, что он произошел от предков,
которых
мы называем сумчатыми (сам он их так не называл), а затем дал начало всем
остальным Insectivora. Хэтчер нашел несколько черепов, челюстей и других
частей скелета этих мелких животных (длина их черепа достигала всего 3,5
см), а в 1905 г. Скотт опубликовал их описание. Он согласился с ошибочной
частью обычной для Амегино двоякой интерпретации, решив, что Necrolestes не
имел ничего общего с сумчатыми, а был плацентарным насекомоядным, родственным
златокротам Южной Африки, как когда-то заявил Амегино. Скотт согласился с
Амегино также в том, что это указывает на непосредственную связь между Африкой
и Южной Америкой. Хотя высказывались и другие предположения, авторитет Скотта
оказал должное влияние. Никто не предпринял дальнейшего изучения самих ископаемых
остатков до 1958 г., когда Паттерсон провел дополнительную препаровку принстонских
экземпляров и изучил их заново, используя инструменты, которыми Скотт, работавший
более полувека назад, не располагал. Он представил доказательства того, что
Necrolestes был сумчатым, и с этим согласились очень многие. В таком положении
этот вопрос и остается, во всяком случае в настоящее время. Паттерсон поместил
Necrolestidae в надсемейство Borhyaenoidea, но признал, что это весьма условно,
так как основано лишь на нескольких действительно специфичных (производных)
боргиенидных признаках, если таковые вообще имеются. Раньше подобное предположение
высказал эксцентричный датский гений Херлуф Винге (Herluf Winge), но это
был всего лишь домысел. Винге никогда не видел ни одного остатка этого животного,
а его так называемые доказательства были далеко не убедительными.
Как же выглядело это таинственное животное? У него был вздернутый нос, многочисленные
резцы (пять сверху и четыре снизу), острые клыки и моляры с треугольными
коронками. Кости передней конечности ясно указывают, что это было роющее
животное, которое вело настоящий подземный образ жизни и было очень похоже
на крота. Этот необычайно яркий пример эволюционной конвергенции в направлении
златокротов Южной Африки (Chrysocloridae) и так называемых сумчатых кротов
Австралии (Notoryctidae) снова ввел в заблуждение квалифицированных морфологов.
В последнем случае, так же как в примере с боргиенидами и тилацинидами, сходство
было особенно велико, поскольку эффект конвергенции усиливался параллельной
эволюцией от далекого общего предка, которого следует искать среди наиболее
примитивных сумчатых. Сходство между этими тремя независимо развивавшимися
группами кротовидных животных зашло настолько далеко, что привело даже к
странному вторичному развитию треугольных моляров.
ЛИТЕРАТУРА
Simpson G. G. 1971. The evolution of marsupials in South America. Anales
Acad.
Brasileira Ciencias, 43 (supplement), 103 — 118.
Stonehouse В., Gilmore
D. 1977. The biology of marsupials. Baltimore, London, Tokyo, University
Park
Press.
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
................................................ 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ .....................................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .................................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
...............................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ........................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252