ГЛАВА 8
КСЕНАРТРЫ — СТРАННО
СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
Название Xenarthra происходит от греческих слов, означающих «странные
сочленения». Такое название было придумано для этой группы млекопитающих,
поскольку в их позвоночнике между некоторыми позвонками имеются дополнительные
сочленения, которых нет у других млекопитающих: они сочленены странным
образом.
Среди современных животных броненосцы, муравьеды и ленивцы столь не похожи
друг на друга, их образ жизни и поведение настолько различны, что едва
ли кому-нибудь пришло бы в голову без дополнительного морфологического
изучения объединить их в одну группу. При изучении же морфологии оказывается,
что их объединяют не только странные сочленения, но и другие особенности,
которые несомненно указывают на общее происхождение, хотя и в далеком прошлом.
У всех этих животных резцов мало или нет совсем, а некоторые (муравьеды)
вообще лишены зубов. Но при этом у некоторых броненосцев зубов больше,
чем у их примитивных плацентарных предков. Только у самых ранних форм были
остатки эмали; у более поздних все зубы состояли из одного только дентина,
а у некоторых — из двух видов дентина различной твердости. Даже древнейшие
из известных ксенартр уже утратили бугорчатое строение моляров, примитивное
для всех плацентарных млекопитающих; в тех случаях, когда моляры сохраняются,
они превращаются в постоянно растущие столбчатые зубы. У всех ксенартр
мощные конечности с хорошо развитыми когтями, причем коготь на третьем
пальце передней конечности часто бывает очень большим. У ксенартр есть
еще одна своеобразная черта; в то время как у остальных плацентарных млекопитающих,
как правило, только одна из трех частей, образующих таз, а именно подвздошная
кость соединяется с позвоночником в области крестца, у ксенартр и второй
элемент таза — седалищная кость — также соединен с крестцом.
Сейчас никто серьезно не сомневается в том, что Xenarthra, несмотря на
их значительное разнообразие, действительно образуют естественную группу
и имеют единое происхождение. Вопрос о том, кто именно был их предком и
когда и где он существовал, связан с большими сомнениями и еще не разгаданными
загадками, как это было показано в гл. 5. Здесь мы частично обходим эти
проблемы, рассматривая Xenarthra как самостоятельную единицу, а не как
часть спорной и по-разному определяемой группы, называемой Edentata (неполнозубые).
| * Находки примитивного представителя ксенартр Ernanodon antelios в верхнем палеоцене Китая и примитивного муравьеда Eurotamandua joresi в среднем эоцене Западной Германии поставили под сомнение предположение о южноамериканском происхождении этого отряда млекопитающих. Интересно, что европейский муравьед найден в одном местонахождении (мессель) с европейским панголином (Storch G. 1978. Eomanis waldi, eiri Schuppentier aus dem Mittel-Eozander "GrubeMessel"bci Darmstadt (Mammalia, Hiohdota), Sencken-bergiana lethaea, 59, 4 — 6, 503 — 529; Storch G. 1981. Eurotamandus joresi, em Myrmecophagidae aus dem Eozan der "Grube Messel" bei Darmstadt (Mammalia, Xenarthra), Senckenbergiana lethaea, 61, 3 — 6, 247 — 289). — Прим. ред. |
Таким образом, Xenarthra — это отряд млекопитающих, в высшей степени характерный
для всех южноамериканских кайнозойских фаун и в течение длительного времени
распространенный только на этом континенте*. Лишь к концу кайнозоя они
расселяются из Южной Америки в Центральную; меньшее число форм проникает
далее в Северную Америку и на некоторые из островов Вест-Индии. Ксенартры
очень рано разделились на три разные ствола, которые можно возвести в ранг
подотрядов, если рассматривать Xenarthra как отряд. Каждый подотряд в процессе
собственной адаптивной радиации приобрел широкое разнообразие. Мы придерживаемся
здесь общепринятых названий этих подотрядов: Cingulata, Pilosa и Vermilingua.
Название Cingulata означает «подпоясанный» или «опоясанный» и связано с
тем, что у членов этой группы большая часть тела покрыта костным панцирем.
Pilosa по-латыни означает «волосатый»; это название было предложено для
разнообразных и многочисленных ленивцев в несколько негативном смысле,
просто потому что они не покрыты панцирем. На самом деле Vermilingua столь
же волосаты, как и Pilosa, а все Cingulata имеют и всегда имели волосы,
которые даже торчат у них из панциря. Vermilingua — название, составленное
зоологами из латинских корней, означающих «червь» и «язык», т. е. «червеязычный»,
что связано с необычайно длинным и клейким языком муравьедов. Все три подотряда
в большем или меньшем обилии сохранились до сих пор.
Большинство Cingulata
принадлежит к двум хорошо различимым семействам: броненосцев (Dasypodidae)
и глиптодонтов (Glyptodontidae). Как уже упоминалось,
броненосцы, насколько это сейчас известно, — первые ксенартры, появляющиеся
в ископаемой летописи начиная с риочикия. Возможно, тем не менее, что к
тому времени уже возникли глиптодонты, что уже существовали предковые формы
муравьедов и практически наверное — предко-вые формы ленивцев. Очевидно,
они просто были редки или отсутствовали в тех небольших областях и ограниченном
наборе биотопов, из которых на сегодняшний день имеются более или менее
полные коллекции ископаемых млекопитающих. Ясно, что из ныне живущих ксенартр
броненосцы обладают наибольшей приспособляемостью или, другими словами,
менее разборчивы в отношении местообитаний и условий жизни. В настоящее
время их ареал простирается дальше, чем у любых других ксенартр, — и к
югу, в Патагонию, и к северу, где он заходит в США. Несмотря на то что
их зубная система представляется нам несколько неполноценной, они едят
практически все, что попадается. Большинство броненосцев предпочитают муравьев,
но не отказываются и от других насекомых, яиц, падали; некоторые даже убивают
и едят небольших змей, птенцов и мелких грызунов. Такая смешанная пища
очень напоминает пищу некоторых опоссумов. Некоторые виды броненосцев стали
в большей степени плотоядными, а другие, хотя это наблюдается реже, в какой-то
мере растительноядными. Как известно, у броненосцев голова покрыта костным
щитом, тело — панцирем, который частью фиксирован, а частью состоит из
подвижно сочлененных колец, а хвост заключен в гибкую костную трубку. На
костных щитках, которые соединяются в панцирь наподобие черепицы различной
формы, лежат тонкие роговые чешуйки, покрывающие те же части, что и костный
панцирь, но образующие другой рисунок. Многие думают, что броненосцы в
случае опасности сворачиваются в шар, однако такое поведение наблюдается
только у одной из современных форм, но не у огромного большинства других
броненосцев, древних и современных. Броненосцы не лазают по деревьям, однако
в остальном встречаются почти повсеместно и являются исключительно активными
землероями. Броненосец успевает зарыться в землю прежде, чем его удается
схватить: крошечные плащеносные броненосцы северной Аргентины ведут почти
такой же подземный образ жизни, как кроты. Кроме того, в задней части панциря
у них есть пластина, которой они затыкают свои ходы.
В касамайорское время броненосцы были уже довольно разнообразны, но из
отложений этого возраста они известны, за единственным исключением, лишь
по отдельным щиткам и не могут быть хорошо охарактеризованы. Более удовлетворительная
информация начинается с сантакрусия. В типовой фауне сантакрусия Патагонии
имеются пять довольно хорошо изученных родов, столь различных, что их обычно
относят к трем разным подсемействам. Одно из этих подсемейств (Euphractinae)
сохранилось до наших дней; к нему относится обычный патагонский броненосец
Zaedyus, или «пичи». На языке арауканских индейцев «пичи» означает «маленький»,
и хотя пичи гораздо крупнее плащеносных броненосцев, размеры его меньше
средних для этого семейства. Довольно странно, что другая ветвь того же
семейства в позднем кайнозое (в уайкерии и монтеэрмосии, т. е. в плиоцене)
дала гигантских броненосцев с длиной черепа около 27 см.
Остатки другого подсемейства, с некоторым сомнением определяемые из десеадия,
безусловно присутствуют в колуапии. Выделение этого подсемейства основывается
главным образом на сантакрусском роде Stegotherium (рис. 10), у которого
была длинная беззубая морда и только до пяти крошечных рудиментарных штифтовидных
зубов на каждой стороне верхней и нижней челюстей. Он, очевидно, адаптировался
к питанию одними лишь муравьями и термитами. Поскольку в это время уже
существовали настоящие муравьеды, еще лучше приспособленные к жизни за
счет этого обильного источника пищи, не удивительно, что Stegotheriinae
не выдержали конкуренции и после сантакрусия уже не известны.
Самый своеобразный из сантакрусских броненосцев — это единственный известный
нам рогатый представитель ксенартр Peltephilus, у которого на носу было
два рога. Его своеобразие заключается также в том, что передние зубы, занимающие
положение медиальных резцов, вероятно, не гомологичны настоящим резцам.
Была высказана догадка, что члены этого подсемейства (Peltephilinae), которое
часто возводится в ранг семейства, были плотоядными или питались падалью.
Это подсемейство известно начиная с десеадия и вплоть до часикия (присутствие
в последнем веке несколько сомнительно), т. е. примерно с раннего олигоцена
до позднего миоцена.
Stegotherium и Peltephilus служат одним из многих примеров того, как отдельные
моменты истории южноамериканских млекопитающих проливают свет на гораздо
более общие принципы и процессы эволюции. Эти два рода довольно близко
родственны: их обычно относят к одному и тому же семейству и всегда — к
одному и тому же подотряду. Тем не менее между ними имеются коренные различия
в отношении разных адаптивных признаков, особенно связанных с защитой и
питанием (рис. 11). Их общий предок был почти наверное в основном всеядным,
а эти потомки специализировались к совершенно разным составным частям его
более разнообразной пищи. Вместе с тем Stegotherium и такой муравьед, как
ныне живущий Myrmecophaga, весьма сходны в отношении формы черепа и редукции
зубов, далеко зашедшей у Stegotherium и полной у Myrmecophaga; тем не менее
они принадлежат к разным подотрядам и далеко не столь близко родственны,
как Stegotherium и Peltephilus.

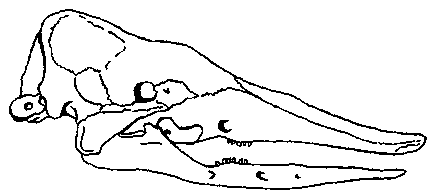
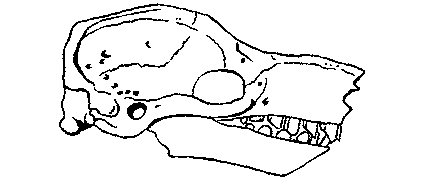
Рис. 10. Реконструкция Stegotherium — раннемиоценового (сантакрусского) броненосца, питавшегося муравьями.
А

Б
В
Рис. 11. Дивергенция двух раннемиоценовых броненосцев (А и В) по строению
черепа и зубной системы и конвергенция одного из них в направлении ныне
живущего муравьеда (Б).
А — Stegotherium — раннемиоценовый броненосец, питавшийся муравьями Б.
Myrmecophaga — ныне живущий муравьед. В. Peltephilus — раннемиоценовый
броненосец, по-видимому, хищный или питавшийся падалью. (Здесь не показаны
костные щитки, покрывавшие черепа броненосцев, на которых у Peltephilus были роговидные выросты.)
Здесь необходимо коснуться нескольких общих вопросов относительно моделей
эволюционного развития. Во-первых, сходные адаптивные. признаки могут возникать
совершенно независимо в разных эволюционных линиях. Пытаясь установить
степень генетического родства, не следует опираться на такие признаки и
остерегаться от включения их в набор признаков, используемых для оценки
сходства при определении положения в системе различных животных, т. е.
при процедуре, на проведении которой все же настаивают крайние сторонники
одной школы систематиков. Одновременно этот пример показывает, что у животных,
несомненно весьма близких генетически, могут развиваться резко различные
признаки и структуры, которые в свою очередь могут вводить в заблуждение,
если включать их в общий набор сходств и отличий, используемых для оценки
родства. Таким образом, внимательный исследователь эволюционной истории
сталкивается с ограничениями с двух разных, по существу противоположных,
сторон и должен их соответствующим образом учитывать.
Во-вторых, пример Stegotherium и Myrmecophaga до некоторой степени проясняет
вопрос о трудно выявляемом и во многих случаях неопределенном различии
между конвергенцией и параллелизмом. Эти роды родственны, оба относятся
к ксенартрам и несомненно имели общих предков. Все это — необходимые условия
параллелизма, когда он действительно имеет место. Но в данном случае общие
предки этих родов те же, что и у всех остальных Xenarthra, и среди других
потомков этих предков есть множество животных, отнюдь не обладавших теми
особыми признаками, которые имеются и у Stegotherium, и у Myrmecophaga.
Таким образом, вполне обоснованным представляется вывод о том, что их сходство
в той мере, в какой оно выражено, целиком обусловлено конвергенцией и вовсе
не зависит от их (довольно далеких) общих предков. Что же касается сумчатых
и, в частности, случая с боргиенидами и тилацинидами, которые гораздо больше
похожи друг на друга, чем стеготерии и муравьеды, то есть основания считать,
что их сходство до некоторой степени определяется общим происхождением,
т. е. имеет генетическую основу. Таким образом, наряду с конвергенцией
здесь, вероятно, имело также место явление параллелизма, а по мнению некоторых
исследователей, параллелизм был преобладающим или даже единственным фактором.
Следует также упомянуть о нескольких подсемействах, которые появляются
в известной мам палеонтологической летописи значительно позже. Dasypodiinae
известны только начиная с монтеэрмосия. Они представляют особый интерес
для североамериканцев, поскольку к ним отноосится девятипоясный броненосец
(Dasypus novemcinctus) — южноамериканский вид, расширивший свой ареал,
который захватывает теперь значительную часть США, где в настоящее время
это единственный местный броненосец. Родственный ему, то более крупный
вид — ныне вымерший Dasypus bellus — в течение плейстоцена был широко распространен
в районах, соответствующих нынешним южным штатам, вплоть до места, где
сейчас стоит город Сент-Луис; это самая северная находка броненосца в естественном
местообитании. Вымершие Pampatheriinae известны в Южной Америке с мастэрсия
(приблизительно средний эоцен) вплоть до плейстоцена. В плейстоцене они
достигали территории современных США и род Holmesina был распространен
по всему югу — от Техаса до Флориды. К нему относились некоторые из самых
крупных броненосцев, длина черепа которых достигала 30 см. Зубы пампатериев
разделены на три лопасти; в этом и в некоторых других отношениях они больше,
чем какие-либо другие броненосцы, напоминают глиптодонтов. Было высказано
предположение, что глиптодонты, возможно, возникли от ранних пампатериев
(самое позднее в палеоцене), но это чисто умозрительное заключение, и нам
оно представляется маловероятным.
Самые крупные из ныне живущих броненосцев относятся к роду Priodontes,
распространение которого ограничено тропической частью Южной Америки. Обычно
этот род выделяют в самостоятельную трибу или подсемейство, к которому
с известными сомнениями относят несколько средне-позднекайнозойских ископаемых
форм. Упоминавшиеся выше плащеносные броненосцы образуют подсемейство Chlamyphorinae,
неизвестное в ископаемом состоянии.
Несколько экземпляров из мастэрсия и десеадия и один — неизвестного возраста,
но лучшей сохранности, чем большинство других, — обычно относят к сем.
Palaeopeltidae. Членов этого гипотетического семейства относили то к броненосцам,
то к глиптодонтам, а то ни к тем, ни к другим. Они вызывают столько сомнений
и бесплодных дискуссий, что едва ли имеет смысл рассматривать их здесь
более подробно.
Среди ископаемых остатков млекопитающих, собранных в Аргентине Дарвином
в 1833 г., был фрагмент глиптодонта; в 1838 г. Ричард Оуэн (Richard Owen),
в ту пору друг Дарвина, но позднее его ожесточенный оппонент, ввел родовое
название Glyptodon, давшее имя всей группе животных, рассматриваемой теперь
как сем. Glyptodontidae. Это название составлено из древнегреческих корней,
означающих «резные зубы». Зубы действительно уникальные, и даже один отдельный
зуб можно безошибочно узнать. У всех глиптодонтов отсутствуют передние
зубы, резцы или клыки, но имеется ряд близко посаженных перетирающих зубов,
которых обычно восемь. Они столбчатые, растут на протяжении всей жизни
и состоят из трех лопастей каждый. У древнейших форм боковые стороны этих
лопастей округлые, но у относительно поздних они более угловатые. Зубы
лишены эмали, но у всех форм, кроме самых древних, на зубах развиты полоски
из более прочного дентина: продольная, проходящая посредине, а поперек
нее в каждой из трех лопастей — поперечная. При изнашивании зубов эти полоски
возвышаются над остальным дентином в виде небольших гребней. Зубы в самом
деле выглядят как резные.
Наиболее замечательная особенность глиптодонтов — не их зубы, но их броня;
это были самые тяжело бронированные млекопитающие, когда-либо жившие на
свете, поистине ходячие танки (рис. 12). Голова их была увенчана костным
шлемом, а тело сверху и с боков полностью покрыто костной мозаикой; в сантакрусское
время в ней еще были участки, обладавшие умеренной подвижностью, но позднее
она стала совершенно неподвижной. Хвост был целиком покрыт панцирем, состоявшим
у некоторых видов в основном из подвижно соединенных колец, что придавало
ему ограниченную гибкость, тогда как у других подвижными были только несколько
проксимальных или передних колец, а за ними следовал совершенно сплошной
конец, похожий на булаву, иногда, по-видимому, с роговыми шипами, хотя
последние и не сохранились. У всех глиптодонтов хвост явно служил грозным
оружием защиты. Казалось бы, благодаря такому оружию и броне они были надежно
защищены от нападения, и тем не менее по неизвестным причинам все они вымерли.
Несмотря на то что глиптодонты известны уже почти полтора столетия, что
они присутствуют и обычно обильны во всех южноамериканских фаунах начиная
е мастэрсия и почти до современности, что им посвящена поистине обширная
литература, как научная, так и популярная, их систематика не может нас
полностью удовлетворить. Амегино разбил их на четыре семейства, и хотя
сейчас их объединяют в одно-единственное сем. Glyptodontidae, те же самые
четыре группы нередко все еще рассматривают как подсемейства. В пределах
семейства разными авторами выделено более пятидесяти родов, хотя валидность
некоторых из них сомнительна. Кажется, что форма хвоста варьирует почти
бесконечно, но другие различия касаются лишь мелких деталей, а общая морфология
почти полностью стереотипна, по крайней мере у поздних форм. К уже упомянутым
признакам можно добавить одинаковый у всех высокий и короткий череп и нижнюю
челюсть; наличие на скуловой кости, под глазницей, направленного вниз шиповидного
отростка; сильные с мощными мышцами конечности, задние длиннее передних;
толстые, тупые, почти копытовидные когти. Все глиптодонты были травоядными
и, по-видимому, питались жесткими растениями, быстро стирающими зубы, такими,
как богатые кремнием травы пампы.
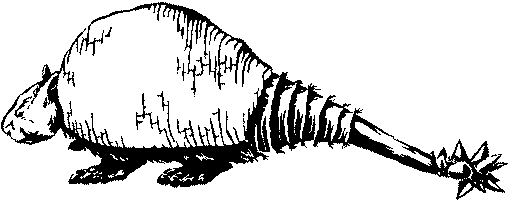
Рис. 12. Реконструкция Doedicurus — плейстоценового глиптодонта (по рисунку Хорсфолла для книги Скотта, с изменениями).
Как уже говорилось, первые глиптодонты известны из мастэрсия и десеадия,
и их зубы несколько отличаются от зубов более поздних глиптодонтов. В отложениях
этого возраста найдены лишь фрагменты панциря; самые ранние действительно
удовлетворительные экземпляры происходят опять-таки из сантакрусия. Сантакрусские
глиптодонты известны под общим названием Propalaeohoplophorinae. Это слово
из 22 букв и девяти слогов кажется почти комически длинным даже для тех,
кому по своей профессии приходится иметь дело с подобными названиями. Однако
возникло это название в результате неразумного, хотя, по-видимому, неизбежного
наращивания. Некто — им оказался Петер Вильгельм Лунд (Peter Wilhelm Lund)
— в 1838 г. первым назвал позднеплейстоценового глиптодонта Hoplophorus — «носитель панциря» — просто, благозвучно и вполне уместно. Некто другой
— не удивительно, что это был Амегино, — в 1887 г. назвал более древнего,
по-видимому предкового, глиптодонта из миоцена Palaeohoplophorus — «древний
носитель панциря». Кроме того, у Амегино в том же 1887 г. была еще более
древняя форма из сантакрусия, которую он считал предком Palaeohoplophorus;
поэтому он назвал ее Propalaeohoplophorus — «предшественник древнего носителя
панциря». Если же заменить окончание -us двусложным окончанием -inae, положенным
подсемейству, то и получится это чудовище — Propalaeohoplophorinae. В это
подсемейство, по-видимому, объединили животных просто потому, что они находились
примерно на одной и той же эволюционной стадии, а не по какому-либо другому
критерию. На этой стадии части покрывавшего тело панциря все еще обладали
некоторой подвижностью. Виды были также существенно мельче более поздних
глиптодонтов, но по общей морфологии они уже как бы представляли собой
миниатюрные облегченные модели, характерные для семейства в целом. Крупнейший
из плейстоценовых глиптодонтов достигал в длину около 3 2/3 м.
В плейстоцене глиптодонты тоже расселились в Северную Америку, проникнув
в районы, которые сейчас относятся к США, и стали довольно обычными в некоторых
частях нынешних южных штатов, от Аризоны до Флориды. Один род был близко
родствен или даже идентичен роду, известному из Урагвая, но три другие,
отличавшиеся более значительно, были, насколько это известно, распространены
только в области, относящейся сейчас к США и Мексике.
Все ксенартры — необычные и интересные животные, но ленивцы представляют
собой во многих отношениях самую необычную и интересную группу. Они прошли
в Южной Америке длительную историю и в конце концов расселились по всему
континенту — от северного конца до южного и от западного края по восточного.
Они были первыми ксенартрами, а фактически первыми из коренных южноамериканских
млекопитающих, которые достигли Северной Америки, где они также расплодились
и широко расселились. Они играют заметную роль в истории науки, а также
в современном изучении таких интересных вопросов, как время появления человека
в Западном полушарии или причины вымирания животных.
Гигантский ленивец был первым ископаемым млекопитающим, полный скелет которого
был собран и выставлен для обозрения. Этот скелет был найден около Лухана
в Аргентине, что само по себе примечательно, поскольку едва ли можно считать
простым совпадением, что мальчики Амегино, впоследствии прославившиеся
сборами и описаниями ископаемых млекопитающих, выросли в Лухане. Несомненно,
к этому занятию их притягивали местные находки, начавшиеся почти за сто
лет до их рождения. Тот скелет, о котором шла речь, был отослан в 1789
г. в Мадрид, где он находится и по сей день. Он был описан и изображен
в большом труде Жоржа Кювье, который был первой вехой в истории палеонтологии
позвоночных и назывался «Исследования ископаемых костей четвероногих, в
которых воссоздаются особенности нескольких видов животных, по-видимому,
уничтоженных происходившими на земном шаре революциями». Описание скелета
приводилось в главе, названной «О мегатерии, другом животном из семейства
ленивцев, но размером с носорога, почти полный ископаемый скелет которого
хранится в королевском кабинете естественной истории в Мадриде». Труд этот
вышел в свет в 1812 г.
В Северной Америке изучение ископаемых ленивцев начал Томас Джефферсон
(Thomas Jefferson), посвятив им доклад, прочитанный в Американском философском
обществе в 1797 г. и опубликованный в 1799 г. Среди имевшихся у него ископаемых
остатков был большой коготь, а точнее кость, которая находилась внутри
несохранившегося рогового когтя (концевая, или копытная, фаланга), и Джефферсон
назвал ее обладателя мегалоникс, или «большой коготь». Он полагал, что
эта кость принадлежит дикой кошке размером примерно втрое крупнее льва.
Джефферсон обладал блестящим умом и огромной эрудицией, но был слабым натуралистом
и еще более слабым морфологом. Одной из его странностей как натуралиста
была твердая вера в то, что господь бог не допустил бы, чтобы какое-нибудь
животное вымерло; поэтому «большой коготь» должен по-прежнему обитать где-нибудь
в диком состоянии. Друг и соратник Джефферсона, Каспар Уистар (Caspar Wistar),
несравненно более умудренный в подобных делах, опубликовал в 1799 г. заметку,
в которой показал, что Megalonyx (название, теперь уже закрепленное в номенклатуре)
был ленивцем. Оттуда Кювье и позаимствовал это название.
После такого раннего и славного начала и постоянно нараставшей интенсивности
последующих исследований хотелось бы сказать, что теперь мы располагаем
всем, что мы желали бы знать об истории ленивцев, но это, конечно, далеко
не так. Исследования продолжаются. В наших знаниях все еще остается немало
неясностей и пробелов. Удовлетворительной системы все еще нет. Обычно используемая
система, вынужденная довольствоваться имеющимися сведениями, делит вымерших
наземных ленивцев, некогда самой разнообразной и широко распространенной
группы из всех ксенартр, на три семейства. Их называют наземными ленивцами,
поскольку они вели, очевидно, наземный, а не древесный образ жизни, в отличие
от ленивцев, сохранившихся до наших дней и помещаемых (возможно, ошибочно,
как будет показано дальше) в одно, четвертое, семейство. Есть еще и пятое
семейство, плохо изученное и спорное, которое, как родственное ленивцам,
мы относим к Pilosa, но оно, вероятно, отличается от них столь сильно,
что, называя его представителей ленивцами, мы, возможно, лишь создаем путаницу.
Большая часть наших знаний об истории ленивцев касается трех семейств,
называемых Mylodontidae, Megalonychidae и Megatheriidae. Амегино отметил
находку двух экземпляров наземных ленивцев в касамайории. Позднее эти экземпляры
были затеряны, а о самой находке упоминалось в нескольких более поздних
обзорах. Но в 60-х годах образцы Амегино нашлись, и повторное изучение
показало, что они, по-видимому, вообще не относятся к ксенартрам. Возможно,
что из мастэрсия происходит таранная кость (астрагал) наземного ленивца,
найденная Амегино, но с равной вероятностью она может происходить из залегающих
выше по разрезу десеадских отложений. Из мастэрсия известен также коготь
'(когтевая фаланга), вероятно, но не наверняка принадлежащий наземному
ленивцу.
Первые известные наземные ленивцы, положение в системе и возраст которых
твердо установлены, происходят из десеадия. Найденные экземпляры неполны,
но они указывают на то, что основное разделение на Mylodontidae и Megalonychidae
к тому времени уже произошло. В сантакрусии присутствовали все три семейства
наземных ленивцев, но Megalonychidae гораздо более обычны в коллекциях
и сохранность их лучше. Тогда это были довольно мелкие и, возможно, полудревесные
животные, хотя разговорное название «наземные ленивцы» распространяется
и на них. Их более поздние потомки были крупнее, однако не были настоящими
гигантами. Их передние зубы, до некоторой степени похожие на клыки, не
происходили из клыков примитивных плацентарных млекопитающих. На это указывает
то обстоятельство, что при смыкании челюстей нижний зуб оказывался позади
верхнего, тогда как настоящие нижние клыки выступают вперед по сравнению
с верхними, что характерно и для правильного прикуса у человека. Немногочисленные
задние, или коренные, зубы мегалонихид были округлыми или овальными в поперечном
сечении. Сам Megalonyx известен главным образом из плейстоцена Северной
Америки; ареал этого животного на севере доходил до Аляски, но его остатки
найдены и в бассейне Амазонки. Вопреки убеждению Джефферсона он полностью
вымер примерно в конце плейстоцена, т.е. около 10 тыс. лет назад, как и
почти все наземные ленивцы. Другой мегалонихид, Nothrotherium, в плейстоцене
был обилен как в Северной, так и в Южной Америке. Именно мегалонихиды достигли
Северной Америки еще до возникновения Панамского моста суши и, таким образом,
стали первыми коренными южноамериканцами, которым это удалось сделать.
Мы знаем, когда именно, но, вероятно, примерно в это же время некоторые
мегалонихиды достигли и Больших Антильских островов, бывших и тогда островами.
Они попали туда путем дрейфа на плавнике и образовали там несколько видов
размерами от мелких до средних. Не исключена возможность, что эти виды
дожили до современности в геологическом смысле и были последними на нашей
планете наземными ленивцами, но все они вымерли до того, как на эти острова
пришли европейцы.
Уже в сантакрусии Megatheriidae были крупнее, чем Megalonychidae, а в плейстоцене
они стали настоящими гигантами, действительно достигая размеров носорога,
как писал Кювье, но других признаков сходства с носорогами у них не было
(рис. 13). Два самых крупных вида, обычных для плейстоцена обоих континентов,
теперь часто помещают в два рода — Megatherium («большой зверь») и Eremotherium («пустынный зверь», хотя он жил не только в пустынях и, вероятно, не был
для них типичен). Эти два вида столь похожи друг на друга, что, по-видимому,
могут служить иллюстрацией к приведенному выше суждению, что если двух
животных можно отличить друг от друга, то они принадлежат к разным родам.
Коренные зубы мегатериид были почти квадратными в поперечном сечении, и
у более поздних форм на них при стирании образовывалось по два поперечных
гребня. При таких крупных размерах эти животные несомненно не могли вести
даже полудревесный образ жизни. Возможно, что, имея очень крепкие, но относительно
короткие задние ноги, они иногда, а может быть и постоянно, ходили на двух
ногах.
Коренные зубы Mylodontidae, которых обычно бывает три или четыре, с очень
причудливым поперечным сечением, несколько напоминающим то, что дизайнеры
называют свободными формами, не похожи на зубы представителей других семейств.
Милодонтиды рано дифференцировались на две группы, иногда трактуемые как
подсемейства. Распространение одного из них для представителей которого
характерна относительно длинная и узкая морда, ограничивалось Южной Америкой;
представители другого, с короткой и широкой мордой, были многочисленны
в плейстоцене обеих Америк. Один из последних хорошо известен многим североамериканцам
благодаря находке огромного количества его остатков в Ранчо-ла-Бреа, в
Голливуде, Калифорния. Номенклатура этих животных оказалась довольно запутанной,
поскольку остается неясным, какому из двух родственных видов принадлежит
название Glossotherium, а какому — название Mylodon;
оба эти родовых названия были предложены в одно и то же время (в 1840 г.)
Ричардом Оуэном, частично
на основании экземпляров, собранных Дарвином. Вид из Ранчо-ла-Бреа долгое
время был известен как Mylodon; в последнее время его обычно называют Paramylodon,
но иногда его трактуют как вид или самое большее подрод рода Glossotherium.
Эта проблема не представляет интереса ни для кого, кроме специалистов,
у которых она навязла в зубах. Как бы их ни называли, все это — члены одной
группы близкородственных животных.

Рис. 13. Реконструкция Megatherium — самого крупного наземного ленивца. Это животное несомненно могло стоять, а возможно, и ходить как выпрямившись на задних лапах, так и на всех четырех конечностях. В последнем случае пальцы его передних лап загибались внутрь, так что лапы опирались на межфаланговые суставы.
Все наземные ленивцы были растительноядными животными; они питались листьями
и побегами. В плейстоцене многие из них на том, и на другом континенте
часто посещали пещеры; в некоторых они оставили мощные залежи экскрементов.
Последние позволили подробно изучить обычную пищу этих животных. Например,
экскременты Nothrotherium, жившего в районах, относящихся сейчас к юго-западу
США, содержат остатки таких пустынных кустарников, как древовидная юкка,
эфедра и креозотовый куст (Larrea, род, произрастающий также в районах
Южной Америки с сухим климатом).
Ныне живущие ленивцы относятся к двум родам: Bradypus с несколькими видами,
которые в просторечии называют аи, или трехпалыми ленивцами, и Choloepus с двумя видами, известными в просторечии как унау, или двупалые ленивцы
(имеется в виду число пальцев на передних конечностях; на задних у обоих
родов по три пальца). Все виды ведут строго древесный образ жизни и обычно
висят спиной вниз под ветвями, уцепившись за них огромными, сильно загнутыми
когтями. Если этих животных не беспокоят, они передвигаются очень медленно,
за что и получили название ленивцев, перенесенное затем на всех их вымерших
родичей, причем, по-видимому, незаслуженно, поскольку наземные ленивцы,
вероятно, были не столь уж ленивыми. На самом деле, защищаясь от нападения,
древесные ленивцы способны передвигаться очень быстро, а их беспощадные
когти придают им устрашающий вид. Несомненные предки древесных ленивцев
или весьма близкородственные им ископаемые формы не известны; древесные
животные, жившие в тропических лесах, вообще плохо представлены в палеонтологической
летописи. Два ныне живущих рода помещают в одно и то же сем. Bradypodidae;
но некоторые детали, понятные лишь специалистам, позволяют предположить,
что Choloepus, возможно, произошел от какого-то примитивного представителя
сем. Megalonychidae, a Bradypus — от одного из ранних мегатериид, близкого
к нотротериям. Если бы нашлись факты, подтверждающие такое предположение,
эти роды следовало бы поместить в разные семейства, однако, если они действительно
возникли от столь разных корней, то для возникновения существующего между
ними очень близкого сходства потребовалась бы очень сильная конвергенция.
Группа, называемая здесь сем. Entelopsidae, окутана туманом незнания и
разногласий. Описание ее основано главным образом на двух фрагментарных
нижних челюстях, происходящих, вероятно, из типового разреза сантакрусия.
По-видимому, они принадлежали животному, похожему на наземного ленивца,
но их своеобразие состоит в том, что у них имеется ряд плотно примыкающих
друг к другу передних (или медиальных) зубов, а задние (или дистальные)
зубы — двулопастные. До сих пор все более или менее понятно: это животное,
чрезвычайно плохо известное, вероятно, достаточно своеобразно и заслуживает
выделения в собственное семейство. Но в десеадии найдено несколько двулопастных
зубов, описанных под названием Orophodon, которые весьма схожи с задними
зубами Entelops — единственного рода, уверенно относимого к Entelopsidae.
Так почему же не поместить Orophodon в сем. Entelopsidae и перестать
ломать себе голову? А потому, что, по мнению двух авторитетов, лучше всех
знакомых
с этими ископаемыми остатками, Паскуаля и Роберта Хоффстеттера, Entelops и Orophodon принадлежат
не только к разным семействам, но и к разным надсемействам. Путаница, однако,
еще больше возрастает ввиду наличия некоторых (хотя совершенно
неубедительных) данных о том, что своеобразные костные щитки, описанные
как Palaeopeltis, возможно, принадлежат орофодону или какому-то
близкому к нему роду, известному по зубам. Это предположение, впервые сделанное
Турнуэром и Годри, в 1956 г. было поддержано Хоффстеттером, назвавшим орофодона
и близкие к нему формы панцирными наземными ленивцами (как он писал, «gravigrades
cuirasses»). Однако позже Хоффстеттер по существу отказался от этой точки
зрения и рассматривал Palaeopeltis как представителя ксенартр, положение
которого в системе остается неясным, а не как члена выделенного им сем.
Orophodontidae.
Эта путаная история, изложенная здесь в весьма сокращенном виде, служит
примером того, к чему может привести неполнота наших знаний, а в сущности
примером путаницы, возникающей в результате полного отсутствия знаний.
Она предостерегает также от рассмотрения принятой здесь системы, по которой
Palaeopeltidae отнесены к Cingulata, a Entelopsidae — к Pilosa, в качестве
окончательной или просто хорошо обоснованной. Эта система всего лишь следует
некоторым авторитетам, а именно Паттерсону и Паскуалю.
К третьему подотряду Vermilingua относится только одно известное семейство,
Myrmecophagidae, первоначально основанное на современном роде Myrmecophaga — гигантских, или трехпалых, муравьедов. Это название буквально означает
«пожиратель муравьев» и кажется вполне уместным, если не считать того,
что основной пищей этих животных служат термиты, которые, хотя их часто
и называют белыми муравьями, на самом деле вовсе не муравьи. Существуют
еще два современных муравьеда: четырехпалый муравьед Tamandua и карликовый,
или двупалый, муравьед Cyclopes. Гигантский муравьед ведет наземный образ
жизни; его украшенный пышными длинными волосами хвост не приспособлен к
цеплянию за ветви. Четырехпалый муравьед ведет частично, а карликовый —
почти полностью древесный образ жизни, и оба имеют хватательные хвосты.
Ископаемые данные по муравьедам крайне неполны. Эти животные известны начиная
с сантакрусия, но, должно быть, возникли гораздо раньше. Известные ископаемые
формы уже не имели зубов, были меньших размеров, чем Myrmecophaga, и морда
у них была короче, но все же удлиненная. Происхождение их от очень примитивного
броненосца, лишенного щитков или сбрасывающего щитки, допустимо, но недоказуемо.
Общее происхождение с ленивцами, независимое от броненосцев, едва ли допустимо.
ЛИТЕРАТУРА
Hoffstetter R. 1958. Xenarthra. In: Traite de paleontologie, ed. J. Piveteau,
tome 6, 2, 535 — 636, Paris, Masson. (Эта большая статья содержит обширную
библиографию.)
Patterson В. 1967. Xenarthra. In: The fossil record, ed. L. В. Н. Tarlo,
London, Geological Society of London, 771 — 772.
Scott W. B. 1903. Mammalia of the Santa Cruz beds, P. I., Edentata. Reports
of the Princeton University Expeditions to Patagonia; 5, 1 — 227.
См. также ссылку на работу Patterson, Pascual, 1972 в литературе к гл.
1.
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ...............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ
МЛЕКОПИТАЮЩИЕ ....................................... 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ .....................................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .................................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
...............................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ........................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252