Устрашающие термины «гистрикоморфный» и
«гистрикогнатный» имеют довольно простой смысл. Ими обозначают функциональные
и морфологические
признаки, интересные и важные в нескольких разных аспектах. Так,
например, один из вопросов, для решения которого имеют значение эти признаки,
относится к области географии: откуда происходит предок кавиоморф
— из Северной Америки или из Африки? Этого вопроса мы кратко
касались в предыдущей
главе, а здесь снова вернемся к нему в другом контексте.
Разберемся сначала в том, что такое гистрикоморфность. у всех
млекопитающих имеется мощная мышца, называемая жевательной (m.
masseter). Она
участвует в жевании, в кусании, а у грызунов еще и в грызении.
У примитивных
млекопитающих жевательная мышца прикрепляется на черепе к скуловой
кости (скуловой
дуге); такое прикрепление этой мышцы сохранилось и у человека
(жевательную мышцу
можно нащупать у себя, приложив пальцы к щеке и крепко сжав челюсти).
Подобное расположение жевательной мышцы характерно для большинства
примитивных грызунов,
особенно в эоцене. В настоящее время оно наблюдается лишь у одного
из современных грызунов, горного бобра* (родственного настоящему
бобру лишь
постольку,
поскольку и тот, и другой относятся к грызунам). У всех остальных
современных грызунов и у большинства постэоценовых ископаемых
форм строение жевательного
аппарата стало более специализированным благодаря смещению места
прикрепления этой мышцы вперед, ближе к передней части черепа.
Независимо от того,
каким способом достигалось такое смещение, его функциональное
значение связано
с передне-задним движением нижней челюсти при грызении и жевании.
Перестройка осуществлялась в разных группах грызунов путем совершенно
различных
морфологических изменений.
* Речь идет о североамериканском Aplodontia rufa. — Прим. ред. |
У кавиоморфных грызунов и некоторых других форм эта функциональная специализация
происходила по сравнению с большинством млекопитающих крайне своеобразным
путем. Жевательная мышца имеет сложное строение; ее более мощная часть
прикрепляется к боковой поверхности лицевой части черепа, впереди глазницы
и скуловой кости. Другая часть прикрепляется к задней части скуловой кости.
Часть жевательной мышцы, прикрепляющаяся к лицевому отделу черепа, направляется
не прямо к нижней челюсти, а проходит сначала через большое отверстие в
передней стенке глазницы, а уже потом спускается к нижней челюсти (рис.
32). У всех млекопитающих на этом месте имеется подглазничное отверстие,
но, за исключением кавиоморф и некоторых других грызунов, оно очень невелико.
Через него проходит ветвь лицевого нерва (также и у человека), но ни одна
мышца, кроме названных исключений, через него не проходит.
Эта особенность была еще в давние времена подмечена анатомами, в частности
у дикобразов Старого Света (африканских, средиземноморских и азиатских),
относящихся к роду Hystrix; соответственно такое расположение жевательной
мышцы получило название гистрикоморфного («такой формы, как у Hystrix»).
Систематики долгое время полагали, что эта необычная черта возникла в
процессе эволюции только один раз, а следовательно, все грызуны, которым
она свойственна,
одного происхождения и составляют естественную группу, названную Hystricomorpha.
Однако сейчас специалисты, изучающие грызунов, пришли к единодушному
мнению, что в нескольких группах, имеющих гистрикоморфное строение, этот
признак
возник независимо от предков рода Hystrix и близкородственных ему форм.
Таким образом, анатомически гистрикоморфные грызуны вовсе не обязательно
относятся к группе, называемой Hystricomorpha в систематике. Поэтому
разногласия относительно места происхождения Caviomorpha, упоминавшиеся
в предыдущей
главе, можно сформулировать здесь несколько иначе. Те, кто уверен, что
предок кавиоморф происходит из Северной Америки, рассматривают кавиоморф
как одну из групп грызунов, которые независимо приобрели гистрикоморфное
строение. Те же, кто уверен в африканском происхождении кавиоморф, полагают,
что они унаследовали гистрикоморфное строение от предковых Hystricomorpha
Старого Света.
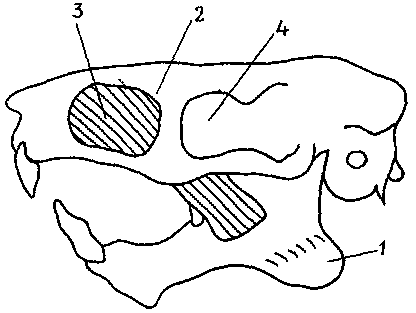
Рис. 32. Несколько обобщенная схема черепа, нижней челюсти
и одной из челюстных мышц кавиоморфного грызуна.
1 — угловой отросток нижней челюсти (он отогнут наружу, т. е. в сторону
читателя); 2 — край большого подглазничного отверстия; 3 — часть большой
жевательной мышцы у своего начала на боковой поверхности лицевой части
черепа; отсюда она проходит через подглазничное отверстие и прикрепляется
на боковой поверхности нижней челюсти (другие части большой жевательной
мышцы на схеме не показаны); 4 — глазница; подглазничное отверстие
открывается кзади в передней части глазницы.
Хотя я по возможности упростил здесь этот довольно сложный вопрос, мне
придется коснуться еще одного усложняющего момента, имеющего решающее значение.
Жевательная мышца обычно прикрепляется к наружной поверхности нижней челюсти,
в задней ее части. У некоторых видов, в том числе и у человека, место ее
прикрепления представляет собой просто довольно уплощенный участок челюсти,
без каких-либо особенностей. Однако у большинства млекопитающих, в том
числе и у грызунов, нижний край челюсти образует в этом месте своего рода
выступ, направленный кзади; в сравнительной анатомии этот выступ называют
угловым отростком. Обычно угловой отросток лежит в той же вертикальной
плоскости, что и вся челюсть и сидящие в ней зубы. Такое строение является
примитивным для плацентарных млекопитающих вообще и для грызунов в частности.
Однако у Hystrix и у ряда других грызунов угловой отросток сильно выступает
наружу (латерально), что создает функциональное отличие в работе жевательной
мышцы. Такое строение называют гистрикогнатным («с челюстью, как у Hystrix»).
Большинство гистрикогнатных грызунов одновременно и гистрикоморфны; это
относится и ко всем кавиоморфам, за одним возможным исключением. Как отмечалось
выше, Platypittamys, быть может, не был вполне гистрикоморфным. Это связано
с тем, что подглазничное отверстие у него было, по-видимому, относительно
маленькое и часть жевательной мышцы через него, возможно, не проходила.
Это мнение, однако, может оказаться ошибочным, поскольку все известные
экземпляры этого грызуна расплющены, так что нельзя с уверенностью утверждать,
что хотя бы маленькая часть жевательной мышцы не проходила через подглазничное
отверстие, как у всех остальных известных кавиоморф (А. Е. Вуд, относившийся
к названиям несколько игриво, дал ему имя Platypittamys, означающее нечто
вроде «грызун-блинчик», поскольку экземпляры эти были расплющены под давлением
перекрывших их пород; между прочим, они найдены в том же древнем кратере,
что и Scarrittia, упоминавшаяся в гл. 10.)
Сейчас некоторые исследователи, в особенности Лавока, выдающийся французский
специалист по ископаемым грызунам, придерживаются мнения, что, хотя гистрикоморфность
возникала несколько раз независимо, этого нельзя сказать о сочетании гистрикогнатности
с гистрикоморфностью и что те грызуны, которые обладают одновременно и
тем, и другим признаком, имели общее происхождение. Представители сохранившейся
до наших дней группы слепышовых (Spalacidae) гистрикогнатны, но не гистрикоморфны.
Лавока объясняет это тем, что слепышовые имели какого-то гистрикоморфного
предка (какого именно — неизвестно), а позднее утратили гистрикоморфность.
Разумеется, концепция единства групп, обладающих одновременно и гистрикоморфностью,
и гистрикогнатностью, стала одним из доводов в пользу африканского происхождения
Caviomorpha. Тем не менее, как указывает Вуд, также выдающийся авторитет
по ископаемым грызунам, и другие исследователи, существуют грызуны, у которых
два эти признака не встречаются вместе. Эти исследователи полагают, что
оба признака возникали в процессе эволюции независимо и неоднократно, а
поэтому нет причин, по которым сочетание этих признаков не могло бы возникнуть
независимо в соответствующих группах южноамериканских и африканских грызунов.
Более того, некоторые грызуны из эоцена Северной Америки вполне могли бы
дать начало кавиоморфам, которые в таком случае не происходили от африканских
грызунов, (возникших от сходных и родственных им более примитивных эоценовых
форм Старого Света), а развивались параллельно им.
В более широком аспекте даже детали, подобные только что затронутым, приобретают
общее значение, поскольку они служат примером вопросов методики и логики,
возникающих при решении проблем систематики, функциональной анатомии, эволюционного
происхождения, палеогеографии, современного распространения животных и
некоторых других проблем, которые мы здесь больше обсуждать не будем.
Все те шесть семейств Caviomorpha, которые появляются в летописи в десеадии
(ранний олигоцен), сохранились и в сантакрусии (ранний миоцен). Они стали,
более четко отличаться друг от друга, и их связи с более поздними и ныне
живущими семействами кавиоморф выступили яснее. Пять из шести десеадско-сантакрусских
семейств существуют и в настоящее время, а поэтому их разнообразные признаки
и экологические роли удобнее рассматривать на примере их многочисленных
и по большей части все еще процветающих современных представителей.
Одно семейство, а именно американские дикобразы (Erethizontidae) знакомо
всем жителям Северной Америки; его представители появляются здесь в плейстоцене
и все еще широко распространены на этом континенте. В Южной Америке сохранились
три рода, один из них — со многими видами, в большинстве своем не такими
колючими, как североамериканский дикобраз. Все дикобразы ведут, по крайней
мере частично, древесный образ жизни и питаются плодами, орехами, листьями
и корой.
Сохранившиеся до наших дней члены сем. Chinchillidae, распространение которого
по-прежнему ограничено Южной Америкой, — это не только шиншиллы, но также
и весьма похожие на них, хотя и несколько более крупные, вискаши. Все они
— наземные бегающие и прыгающие животные с довольно большими ушами, по
внешнему виду и поведению довольно сходные с длиннохвостыми кроликами.
Они становятся все более редкими, и после столь многих миллионов лет существования
это семейство, видимо, близко к вымиранию, по крайней мере в природе. Шиншилл
истребляют из-за их меха и по этой же причине выращивают в неволе. Вискаш
преследуют ради мяса; кроме того, едва они становятся мало-мальски многочисленными,
их начинают истреблять как вредителей.
Ныне живущие агути и пака составляют три рода в сходных и иногда объединяемых
сем. Dasyproctidae (рис. 33) и Cuniculidae; они широко распространены в
Южной Америке и в тропиках Северной Америки. Это довольно крупные животные
весом примерно до 10 кг. У них необычайно длинные задние ноги, и они очень
быстро бегают. По общему адаптивному типу они сходны с поздними хегетотериями
(Notoungulata); возможно, что это как-то повлияло на вымирание последних.
Мясо пака и агути обладает превосходным вкусом, поэтому на них интенсивно
охотятся.

Рис. 33. Реконструкция Neoreomys — раннемиоценового (сантакрусского) кавиоморфного грызуна (сем. Dasyproctidae).

Рис. 34. Реконструкция Platypittamys — раннеолигоценового (десеадского) кавиоморфного грызуна (сем. Octodontidae).
Сем. Octodontidae получило свое имя от ныне живущего рода Octodon («восьмизуб»),
названного так не потому, что у него восемь зубов, а потому, что поперечное
сечение его постоянно растущих моляров напоминает цифру 8. Самого примитивного
из известных кавиоморф Platypittamys (рис. 34) в настоящее время
относят к этому семейству, хотя его низкокоронковые зубы еще не были
похожи на
восьмерку. У рода Acaremys из сантакрусия моляры были высококоронковыми
и при стирании действительно напоминали цифру 8. Сейчас распространение
этого семейства ограничено югом Южной Америки. Оно содержит пять несколько
различающихся современных родов с многочисленными местными народными
названиями, малоизвестными за пределами Южной Америки. Виды самого рода Octodon называют
дегу. Животные, принадлежащие к этому и ко всем остальным ныне живущим
родам, за исключением одного, выглядят как крупные массивные крысы с
непропорционально большими головами. Все они искусные землерои, а многочисленные
виды рода
Ctenomys проводят под землей большую часть жизни. По внешнему
виду и поведению представители этого рода очень похожи на североамериканских
гоферов; они
примерно таких же размеров, но у них нет свойственных гоферам защечных
мешков. Народное название Ctenomys — туко-туко, по издаваемым ими звукам;
когда эти звуки раздаются из-под земли, прямо под ногами, они вызывают
жутковатое ощущение.
Наконец, среди всех этих семейств, существующих на протяжении уже почти
35 млн. лет, следует назвать сем. Echimyidae, отличающееся наибольшим разнообразием
и наивысшей численностью отдельных видов из всех ныне живущих кавиоморф.
Его виды широко распространены в Южной Америке и в тропической части Северной
Америки и имеют множество народных названий. Внешне все они более или менее
похожи на крыс; у многих, хотя и не у всех, в волосяном покрове есть короткие,
гибкие щетинки, за что они получили название колючих шиншилл. У некоторых
из них высота коронки моляров несколько увеличена, однако даже у современных
видов моляры сохраняют корни, тогда как у многих кавиоморф развились постоянно
растущие зубы. В какой-то момент своей сложной истории представители этого
семейства достигли островов Вест-Индии, дрейфуя на плавнике, и на островах
Пуэрто-Рико, Гаити и Куба эволюционировали в самостоятельные роды. Во всяком
случае, на Гаити они, очевидно, еще существовали, когда туда пришли европейцы,
но сейчас все островные роды вымерли.
Единственное семейство кавиоморфных грызунов, известное в Южной Америке
с десеадия, но не известное после фриасия, и, следовательно, номинально
вымершее (Eocardiidae), возможно, вымерло только как название. Два сохранившиеся
до наших дней семейства, впервые появляющиеся во фриасии, вероятно, были
потомками пока неизвестных или нераспознанных более ранних представителей
десеадско-фриасских эокардиид. Оба современных семейства, о которых идет
речь, хорошо известны — это Caviidae и Hydrochoeridae. Типичный для первого
семейства род — Cavia, или обычная морская свинка, по-английски названная
в свое время «гвинейской свинкой»; в этом семействе есть еще три рода,
называемых гвинейскими свинками или кавиями, хотя они имеют другие, местные,
названия. Есть старая шутка, что гвинейских свинок называют так именно
потому, что они не из Гвинеи и не свинки. Высказывались догадки, что их
назвали гвинейскими свинками, так как они похожи на детенышей настоящих
диких свиней, живущих в Гвинее, или потому, что их привозили в Англию на
кораблях, возвращавшихся из Южной Америки после путешествия в Гвинею за
рабами, или, наконец, просто потому, что Гвинеей называли любое далекое
и неизвестное место; ни одна из этих версий не кажется особенно убедительной.
Лучше называть этих животных кавиями, но происхождение этого названия также
неясно.
Каждый знает, что морские свинки — это унылые маленькие создания с относительно
большой головой и едва заметным хвостом. Прежде их широко использовали
в качестве лабораторных животных, так что «морскими свинками» стали называть
любое животное или человека, используемых в экспериментах, но сами морские
свинки теперь редко служат подопытными животными. Эти симпатичные домашние
зверьки относятся к числу тех немногих видов, которых впервые одомашнили
американские индейцы, употреблявшие их в пищу.
Совершенно отличается от морских свинок на первый взгляд другой представитель
сем. Caviidae — мара, или патагонский заяц, который хотя и патагонский,
но не заяц. Его называют также патагонской кавией, но он ни по внешнему
виду, ни поведением не напоминает кавий, т. е. морских свинок. Это крупный
зверек с более заостренным носом и более длинными ушами; конечности у него
также длиннее и передвигается он быстрее, при этом его прыжки действительно
больше похожи на заячьи. Его мясо, как и у морской свинки (и практически
у всех кавиоморф), имеет приятный вкус.
И у морской свинки, и у мара коренные зубы с очень высокой коронкой (гипселодонтные),
при стирании которой на жевательной поверхности выступает простой двух-
или трехлопастный рисунок.
Другое семейство, впервые появляющееся во фриасии и происходящее, возможно,
от более ранних эокардиид, — это Hydrochoeridae, к которому относится один
ныне живущий вид — водосвинка, или капибара, и его более многочисленные
вымершие родичи. Современный вид, ареал которого простирается от восточной
части Панамы в тропические области Южной Америки, — самый крупный из ныне
живущих грызунов; тело его достигает в длину примерно 1,3 м, в высоту —
примерно 50 см, а вес — до 50 кг. У водосвинки большая тяжелая голова с
тупым носом и маленькими ушами, короткий как бы обрубленный хвост; между
пальцами имеются неполные плавательные перепонки. Это полуводные животные,
всегда обитающие вблизи постоянных водоемов. Некоторые из их вымерших родичей
несомненно вели другой образ жизни. Например, у плиоценового Protohydrochoerus (рис. 35) конечности были длиннее, и он, по-видимому, был бегающим животным,
приспособленным к жизни в открытой пампе.

Рис. 35. Реконструкция Protohydrochoerus (он был, вероятно, бегающим) — кавиоморфного грызуна (сем. Hydrochoeridae).
Как и у морских свинок, зубы у водосвинок постоянно растущие (гипселодонтные).
Коренные зубы древнейших представителей Hydrochoeridae — один премоляр
и три моляра с каждой стороны — напоминали зубы некоторых Caviidae, но
у более поздних, более специализированных представителей семейства нижний
премоляр и в особенности последний верхний моляр становятся весьма усложненными.
Последний верхний моляр — очень крупный, с девятнадцатью поперечными эмалевыми
пластинками, которые разделены цементом и при стирании превращаются в перетирающие
гребни.
В эпоху Великого американского обмена, которую мы в дальнейшем рассмотрим
специально, водосвинки расселились далеко в глубь Северной Америки, и их
ископаемые остатки находят на юге США, особенно во Флориде, Техасе и Аризоне.
Некоторые ископаемые виды, как северо-, так и южноамериканские, были еще
крупнее, чем ныне живущий вид.
Но самым крупным из всех известных грызунов была, по всей вероятности,
не водосвинка, а представитель другого семейства, Dinomyidae, которое также
известно начиная с фриасия (миоцен) и содержит только один ныне живущий
вид под названием ложная пака, или пакарана (рис. 36). Она действительно
сильно напоминает паку, вплоть до рядов белых пятен по бокам туловища,
но еще больше она похожа на достигшую чудовищных размеров — почти с маленького
медведя — морскую свинку, хотя и не такую большую, как современная водосвинка.
Самым крупным из ее известных родичей и, возможно, самым большим грызуном,
когда-либо жившим на Земле, был позднеплейстоценовый вид из Аргентины,
названный Telicomys gigantissimus. Аргентинский палеонтолог Л. Краглевич
создал название Telicomys из греческих корней (смысл его можно более или
менее точно передать как «всем грызунам грызун», хотя сам автор об этом
и не писал). Это животное, видовое название которого означает «самый гигантский»,
было величиной почти с носорога. Его более мелкий, но все еще достаточно
крупный ныне живущий родич стал типом рода Dinomys («ужасный грызун»).
Те, кому знакомы эти животные, называют их «добродушными и миролюбивыми»,
поэтому их можно считать ужасными лишь в том смысле, что они ужасно большие
по сравнению с мышью (некоторые динозавры — «ужасные рептилии», — насколько
мы можем судить, также, вероятно, были добродушными).
Пакараны, а также настоящие паки, палеонтологическая история которых неизвестна,
произошли, вероятно, от того же предка, что и агути (сем. Dasyproctidae),
или же представляют собой одну из ветвей, рано отпочковавшихся от этого
семейства.

Рис. 36. Dinomys — единственный ныне живущий представитель сем. Dinomyidae.
Чтобы завершить этот обзор Caviomorpha, следует еще коснуться сем. Heptaxodontidae
и Capromyidae, о которых не упоминается ни в системе, приведенной в гл.
4, ни в первой части этой главы, поскольку, согласно имеющимся сведениям,
они никогда не встречались как таковые в Южной Америке. К сем. Heptaxodontidae
относятся шесть или семь родов с Ямайки, Гаити, Пуэрто-Рико, Сен-Мартена
и Ангилья (Антильские острова). Зубы и кости этих животных напоминают зубы
и кости диномиид; возможно, что сем. Heptaxodontidae — результат протекавшей
на этих разных островах эволюции группы, которая была занесена на них из-за
моря и первоначально, в Южной Америке, была диномиидной. Их часто относили
к Dinomyidae или путали с ними, но они, по-видимому, в самом деле представляют
собой группу, имевшую единое происхождение и претерпевшую ограниченную
радиацию на разных островах. Все эти животные сейчас вымерли, но их известные
остатки в геологическом смысле относятся к современной эпохе и их иногда
находят в том, что по-видимому, представляло собой кучи кухонных отбросов.
Некоторые виды наверняка еще существовали на этих островах, когда до них
добрались индейцы, а возможно, даже и тогда, когда там высадились европейцы.
Сем. Capromyidae (в его современном понимании) также ограничено в своем
распространении Антильскими островами; в него входят три вымерших рода
и четыре ныне живущих Современные роды живут на Кубе, Ямайке, Гаити и Багамских
островах; вымершие жили на Гаити, Пуэрто-Рико и Виргинских островах. Современные
формы известны под общим названием хутий; в этом индейском по происхождению
названии буква h произносится, поэтому по-испански его можно писать как
jutia. Хутии похожи на переросших крыс; несмотря на это, на них интенсивно
охотятся ради их мяса, так что шансов выжить на этих островах с их плотным
и голодным населением у них мало.
Хутии сходны и, по-видимому, до некоторой степени родственны нутрии, или
койпу, происходящей из Южной Америки и все еще живущей там; сейчас, однако,
в некоторых районах Северной Америки и Европы в природе живут нутрии, убежавшие
с пушных ферм и ставшие вредителями сельского хозяйства. В ископаемом состоянии
нутрии известны в Аргентине из позднего плиоцена (чападмалалия). Ныне живущие
нутрии ведут водный образ жизни и питаются главным образом водными растениями,
тогда как хутии — наземные и полудревесные животные. Между ними существует
и ряд морфологических различий; одно из самых интересных различий заключается
в том, что у самки нутрии шесть пар молочных желез, которые сильно смещены
вверх, располагаясь по бокам туловища, так что детеныши могут сосать мать,
когда она плывет. У хутий две пары желез, расположенных на обычном месте.
Хутий и нутрий часто относят к одному и тому же семейству, обычно к сем.
Capromyidae, а иногда рассматривают их как подсемейства сем. Echimyidae.
Представляется вероятным, что они возникли независимо друг от друга от
ранних эхимиид, а поэтому мы помещаем их здесь в разные семейства: нутрий
— в сем. Myocastoridae, ограниченное в своем распространении Южной Америкой,
а хутий — в сем. Capromyidae, ограниченное Антильскими островами.
Окидывая взглядом всю историю кавиоморф, мы видим, что в раннем олигоцене,
когда эти животные впервые появляются в палеонтологической летописи,
они уже находились на гораздо более продвинутой стадии эволюции, чем стадия
примитивных
плацентарных млекопитающих. Они уже приобрели не только исходную специализацию,
свойственную грызунам вообще, но и собственную дальнейшую специализацию,
кое в чем общую для них, для некоторых эоценовых грызунов Северной Америки
и для некоторых форм, известных в Евразии и Африке начиная с олигоцена.
В Южной Америке кавиоморфы также успели дифференцироваться на шесть адаптивных
групп, таксономически различаемых как семейства. Потомки пяти (а возможно,
и шести) групп все еще входят в состав современной фауны Южной Америки.
В ходе своей истории эти группы становились все более специализированными,
а также все более разнообразными, давая начало новым адаптивным группам
такого высокого систематического ранга, как семейство. Число семейств
возросло
от шести в начале их известной истории до девяти в раннем плиоцене, десяти
в позднем плиоцене и раннем плейстоцене и одиннадцати в голоцене. На
Антильских островах возникают еще два семейства; оба они существовали в
геологическую
эпоху, называемую современностью, или голоценом, хотя одно из них вымерло,
вероятно в XVI в. н. э.
Будучи в основном растительноядными (хотя некоторые из них разнообразят
свой рацион небольшим количеством животной пищи), кавиоморфные грызуны
на протяжении всей своей истории в Южной Америке входили в состав тех же
сообществ, что и растительноядные копытные, в частности до плейстоцена
— в одни сообщества с нотоунгулятами. Некоторые адаптивные эволюционные
тенденции проявлялись сходным образом в обеих группах, хотя и не у всех
их членов. В некоторых линиях кавиоморф и нотоунгулят отмечается тенденция
к увеличению общих размеров. В обеих группах жевательные зубы становились
все более высококоронковыми, а в конечном счете — непрерывно растущими
на протяжении всей жизни. В процессе этого изменения обе группы приобрели
моляры с перетирающими эмалевыми гребнями, хотя строение зубов у них резко
различалось. Кроме того, у обеих групп перетирающие зубы имели тенденцию
к развитию цемента, так что жевательная поверхность складывалась из трех
веществ различной твердости: самой твердой была эмаль, затем шел дентин,
а цемент был самым мягким.
Нотоунгуляты достигли расцвета в раннем олигоцене. Впоследствии они становятся
более специализированными, выживающие семейства дивергируют все сильнее,
но число их уменьшается. Из десяти олигоценовых семейств нотоунгулят в
миоцене остается только шесть, а в плиоцене и плейстоцене — всего три.
Сейчас все они вымерли. Угасание нотоунгулят сопровождается противоположной
тенденцией среди кавиоморфных грызунов. Последние достигли максимального
расцвета в плиоцене, когда в разных группах размеры особей варьировали
от самых мелких до гигантских и когда среди них были специализированные
группы, способные занять огромное число экологических ниш — от водоемов
до вершин деревьев, и имевшие самые разнообразные узкоспециализированные
пищевые потребности. Правда, некоторые из наиболее узкоспециализированных
плиоценовых кавиоморф вымерли, но все плиоценовые семейства сохранились,
причем некоторые из них представлены большим числом видов с высокой численностью.
Мы предполагаем, но предположение это обоснованно, что упадок нотоунгулят
и других древних южноамериканских копытных и расцвет древних местных грызунов
— явления взаимосвязанные и что грызуны все больше и больше приспосабливались
к некоторым экологическим нишам которые прежде занимали копытные.
ЛИТЕРАТУРА
Fields R. W. 1957. Hystricomorph rodents from the late Miocene of Colombia
South America. University of California Publications in Geological
Sciences 32, 273 — 404.
Scott W. B. 1905. Glires (of the Santa Cruz beds). Reports of the Princeton
University Expeditions to Patagonia, 5, 384 — 489.
Walker E. P., Warnick F., Hamlet S. E., Lange K. I., Davis M. A.,
Uible H. E. Wright P. F., 1975. Mammals of the world. 3d ed., rev. J. R.
Paradiso, Baltimore and London, Johns Hopkins University Press. (В
томе 2 на с. 1009
— 1067 описаны современные кавиоморфы, хотя и не под этим названием,
и приведены изображения представителей каждого рода.)
Wood A. E., Patterson В. 1959. The rodents of the Deseadan Oligocene
of Patagonia and the beginnings of South American rodent evolution.
Bulletin
of the Museum of Comparative Zoology, 120, 281 — 428.
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ...............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
................................................ 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ .....................................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .................................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
...............................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ...................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252