ГЛАВА 9
НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ
Нотоунгуляты столь многочисленны и по числу форм, и по числу особей и
столь заметно преобладают в большинстве ископаемых фаун Южной Америки,
что им следует посвятить отдельную главу. В данной же главе рассматриваются
еще более разнообразные, но значительно менее многочисленные копытные млекопитающие,
которые рано появляются в палеонтологической летописи Южной Америки и не
относятся к нотоунгулятам (или, во всяком случае, не относятся к ним наверное).
Здесь мы их делим на шесть отрядов, хотя, как будет показано дальше, относительно
ранга некоторых из них возникают сомнения. Эти отряды — Condylarthra, Litopterna,
Pyrotheria, Astrapotheria, Trigonostylopoidea и Xenungulata. За исключением
кондиляртр, которые, как указывалось в гл. 5, вероятно, возникли не в Южной
Америке, распространение этих номинальных отрядов было ограничено, насколько
известно, Южной Америкой; почти наверное там они и возникли.
В древнейших фаунах кондиляртры не отличаются обилием, но довольно разнообразны.
В риочикии встречаются три по нынешним представлениям валидных рода. Два
из них известны также из касамайория, где к ним прибавляется еще пять других
родов. Большинство их известно по отдельным зубам и нескольким неполным
челюстям, за одним приметным исключением: раздавленный, но почти полный
череп с нижней челюстью, найденный в риочикской фауне Итабораи. Все эти
ранние южноамериканские кондиляртры очень сходны с североамериканскими
палеоценовыми и раннеэоценовыми кондиляртрами, хотя и отличаются от них
на родовом уровне, и едва ли можно сомневаться в том, что эти животные
на обоих континентах имели общих предков. Большинство южноамериканских
родов особенно сходно с североамериканскими Phenacodontidae, и выделение
южноамериканцев в отдельное семейство, Didolodontidae, свидетельствует,
возможно, о переоценке степени различия между ними. Паула Коуто относил
наиболее изученный род Asmithwoodwardia к сем. Hyopsodontidae, в остальном
известному исключительно из Северной Америки и Евразии, явно подразумевая,
что в фаунистическом обмене между Южной Америкой и северными континентами
участвовали две разные ветви кондиляртр. Хотя зубы Asmithwoodwardia действительно
очень сходны с зубами североамериканского Hyopsodus, они весьма напоминают
также зубы другого южноамериканского рода, Ernestokokenia, который сам
Паула Коуто относит к сем. Didolodontidae. Существует обоснованная гипотеза,
что известные южноамериканские кондиляртры представляли собой первоначально
единую ветвь; в процессе дивергенции эта ветвь дала род, который в результате
параллельной эволюции стал походить на Hyopsodus. Такая точка зрения была
высказана Мальколмом Мак-Кенна (Malcolm McKenna) в 1956 г., а затем поддержана
Паттерсоном и Паскуалем. Справедливо также и то, что Hyopsodontidae и Phenacodontidae
не так уж сильно отличаются друг от друга, чтобы было явно необходимо считать
их разными семействами.
Зубная система всех дидолодонтид крайне примитивна для копытных. Моляры
с низкой коронкой, с более или менее коническими бугорками и со слаборазвитыми
гребнями (лофами и лофидами, по научной терминологии) или вовсе без них;
сохраняются все бугорки, примитивные для плацентарных млекопитающих в целом,
с добавлением лишь нескольких новых. И верхние, и нижние моляры стали почти
квадратными — обычная для копытных незначительная специализация, встречающаяся
также в некоторых других группах (см. рис. 19, Б).
После касамайория кондиляртры становятся редкими в аргентинских разрезах,
а после десеадия, т. е. приблизительно раннего олигоцена, не известны вовсе.
В Колумбии, однако, была сделана одна находка в значительно более поздних
отложениях фриасия, т. е. приблизительно среднего миоцена. Это пример дифференцировки
сильно разобщенных местных фаун внутри общей фауны данного континента.
Остатки Litopterna, известные из риочикия, касамайория и мастэрсия, представлены
в основном отдельными зубами, хотя по каждой из этих трех фаун имеется
несколько неполных челюстей с некоторым числом зубов. Вообще зубы литоптерн
на этих стадиях все еще напоминают зубы кондиляртр, и в этом отношении
ранняя эволюция литоптерн была гораздо более консервативной, чем в остальных
отрядах, не относящихся к кондиляртрам. У. Б. Скотт помещал Didolodontidae
в отряд Litopterna, а когда в 1914 г. Фредерик Лумис (Frederic Loomis)
опубликовал гипотетическую реконструкцию скелета десеадского литоптерна,
который на самом деле был смесью частей скелетов кондиляртр и литоптерн,
путаница еще больше возросла. Более поздние исследователи решили разделить
эти два отряда, признавая при этом, что один из них (Litopterna) произошел
от другого (Condylarthra). Несмотря на сходство моляров у кондиляртр и
литоптерн, уже в риочикии их обычно можно четко различить, а в отряде литоптерн
уже обособились два семейства: Proterotheriidae и Macraucheniidae, в пределах
которых в основном происходила его последующая эволюция. Наиболее отчетливое
отличие древнейших известных нам литоптерн от современных им кондиляртр
состояло в том, что у первых гребни на молярах становились все более выраженными
(зубы становились более лофиодонтными), причем на нижних молярах развивалось
по два полулунных гребня.
Так же как ряд других групп южноамериканских млекопитающих, литоптерны
по-настоящему хорошо известны лишь начиная с сантакрусской фауны. У нескольких
родов сантакрусских протеротериев наблюдаются два несколько различных адаптивных
типа. У обоих типов зубы имеют одинаково низкие коронки и сильно развитые
гребни, а нижние моляры — по два полулунных гребня. У родов Diadiaphorus и собственно Proterotherium, относящихся к первому адаптивному типу, конечности
трехпалые, средний палец довольно тонкий, но мощный, а боковые одинаковой
длины и такие тонкие и короткие, что вряд ли они были функциональными.
У рода Thoatherium (рис. 14) только один палец, а имевшиеся у предковых
форм боковые пальцы редуцированы и имеют вид лишь небольших шишечек.
Эти литоптерны по общему облику и адаптивным признакам были удивительно
похожи на лошадей (хотя и сильно отличались от них по многим морфологическим
признакам). И еще раз Амегино был введен в заблуждение этими конвергентными
признаками. Он полагал, что эти литоптерны особенно близки к лошадям, хотя
и не были их предками; как будет сказано ниже, предками лошадей он считал
одно из семейств нотоунгулят. Конечности трехпалых протеротериев внешне
и по своей механике сходны с конечностями некоторых вымерших трехпалых
лошадей. Зубы различаются более явно и все же функционально сходны с зубами
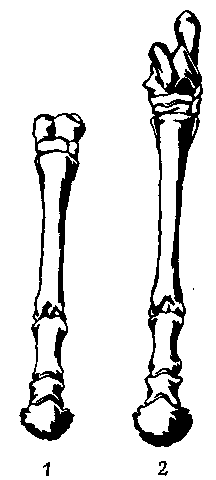
некоторых листоядных, но не травоядных трехпалых лошадей. У Thoatherium однопалость выражена еще сильнее, чем у однопалых лошадей, в том числе
и у всех ныне живущих представителей сем. Equidae. Остатки боковых пальцев
у него даже еще более рудиментарны, чем у Equus (рис. 15); вероятно, он
был самым совершенным однопалым животным, которое когда-либо жило на свете.
Примечательно, кроме того, что в этом семействе литоптерн стадия однопалости
была достигнута примерно в раннем миоцене, тогда как в семействе лошадиных
это произошло только в позднем плиоцене, т. е. спустя миллионы лет. Очевидное
и важное различие между ними состоит в том, что у Thoatherium зубы все
еше были относительно примитивного листоядного типа, тогда как все однопалые
лошади имели и имеют высокоспециализированную травоядную зубную систему.
Протеротерии сохранились до чападмалалия, т. е. почти до самого конца плиоцена,
но затем вымерли.
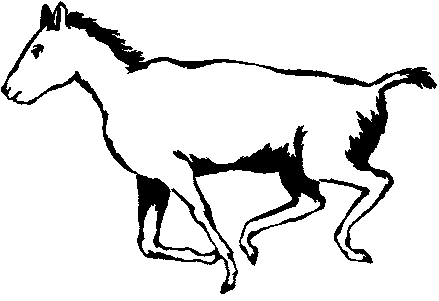
Рис. 14. Реконструкция раннемиоценового (сантакрусского) литоптерна Thoatherium, похожего на лошадь.
 |
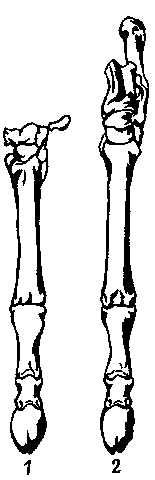 |
Рис. 15. Сравнение строения передней (1) и задней (2) конечностей современной лошади (Equus, слева) и раннемиоценового литоптерна (Thoatherium, справа), У которого однопалость достигла наивысшей степени по сравнению со всеми другими известными млекопитающими. Ьлизкое сходство этих двух ног обусловлено исключительно конвергенцией.
Второе семейство литоптерн, Macraucheniidae, представлено в сантакрусии
родом Theosodon. Представители этого рода были крупнее, чем протеротерии,
и имели трехпалые конечности, причем их боковые пальцы были более развиты
как по величине, так и в функциональном отношении, чем у трехпалых протеротериев.
Зубы сходны с зубами протеротериев, но имеют несколько более сложное строение.
Шея длинная, похожая на шею верблюдов, а носовая область черепа сдвинута
назад, указывая на то, что у этих животных имелся короткий хоботок, более
или менее сходный с хоботком тапиров. У последнего представителя этого
семейства, плейстоценовой Macrauchenia, шея была еще длиннее; судя по некоторым
данным, хобот у нее был еще более развит, хотя и не в такой степени, как
у слонов. Она была также гораздо крупнее, достигая размеров современного
верблюда. Macrauchenia означает по-гречески «длинная шея», но это слово
можно также перевести как «большая лама», поскольку Auchenia — название,
ранее присвоенное ламе (как сейчас выясняется, невалидное). У Дарвина,
собравшего остатки, на основании которых Оуэн описал Macrauchenia, первоначально
создалось впечатление, что это «лама, или гуанако величиной с верблюда».
Ламы и гуанако — мелкие безгорбые верблюды, и, если отбросить ее хобот,
Macrauchenia обладает полным конвергентным сходством с верблюдами.
Еще одно выделяемое в настоящее время семейство литоптерн носит название
Adianthidae. Это были мелкие изящные животные, существовавшие от десеадия
до сантакрусия; они плохо известны, за исключением одного почти полного
черепа, происходящего из слоев, вероятно, раннедесеадского возраста около
Мендосы в Аргентине.
Pyrotheria впервые стали известны по роду Pyrotherium (Ameghino, 1888)
и до сих пор лучше всего известны по этому роду. Он характерен для слоев
и фауны, которые братья Амегино назвали пиротериевыми, а мы сейчас, с небольшой
поправкой следуя Годри, называем десеадием (приблизительно ранний олигоцен).
Амегино полагал, что это животное относится к хоботным, т. е. родственно
мастодонтам и слонам, и рассматривал ранних представителей сем. Pyrotheriidae
как предков последних. Он считал, что пиротерии произошли от кондиляртр,
что, по-видимому, верно, хотя промежуточные формы между ними не известны;
однако предполагавшееся их общее происхождение с более поздними хоботными
основывалось опять-таки на конвергентном сходстве, причем в данном случае
сходство в действительности было невелико. Годри посвятил статью экземплярам
Pyrotherium, собранным для него Турнуэром, и был прав, отрицая их родство
с Proboscidea. Он писал: «В действительности Pyrotherium весьма отличается
от всех крупных животных, описанных до сих пор. Он не вписывается ни в
один известный отряд».
В 1911 — 1912 гг. Ф. Б. Лумис возглавил экспедицию в Патагонию, которую
финансировали выпускники колледжа Амхерст, окончившие его в 1896 г., в
рамках программы, посвященной их пятнадцатой встрече. Самой ценной находкой
этой экспедиции был череп Pyrotherium. Исследовавший его Лумис пришел к
выводу, что этот род и все семейство действительно относятся к хоботным,
как считал Амегино. Недавно Брайен Паттерсон допрепарировал экземпляр Лумиса
— единственный известный череп представителя этого семейства — и, изучая
его наново, обнаружил, что описание и рисунок Лумиса очень неточны, а выводы
совершенно неверны, В связи с заключением самого Паттерсона, основанным
на четких и несомненно тщательных наблюдениях, тем не менее возникает серьезное
затруднение, которое будет рассмотрено после обзора остальных известных
или вероятных представителей отряда Pyrotheria.
В касамайории обнаружен очень редкий и очень плохо известный род Carolozittelia,
который, вероятно, хотя и не абсолютно твердо, можно считать пиротерием.
Столь же фрагментарные, но несомненные остатки пиротериев были найдены
в мастэрсии Патагонии. Экземпляры собственно Pyrotherium происходят из
десеадия Аргентины и Боливии; никакие более поздние пиротерии не известны.
Имеется один экземпляр из Перу, отнесенный к раннему кайнозою (определить
его возраст более точно не удалось). Возможно, что, несмотря на значительное
отличие от Pyrotherium, он относится к сем. Pyrotheriidae, хотя утверждать
это с уверенностью нельзя.
Недавно Хоффстеттер (1970 г.) и Паттерсон (1977 г.) описали два похожих,
но различных рода: Colombitherium Hoffstetter из Колумбии и Proticia Patterson
из Венесуэлы. Эти роды родственны между собой, и Proticia, возможно является
предком Colombitherium. Они родственны также и Pyrotherium, но отличаются
от него так сильно, что выделены в отдельное семейство Colombitheriidae.
Colombitherium, вероятно, относится к эоцену; Proticia может быть также
эоценовой, а возможно, она древнее и относится к палеоцену. У пиротерия
верхние и нижние моляры снабжены двумя острыми поперечными гребнями. Верхние
премоляры и моляры коломбитерия — единственные известные его зубы — также
имеют по два поперечных гребня, но последние менее острые, с четко выраженными
бугорками на наружном и внутреннем концах, а характер их стирания заставляет
предполагать какой-то необычный механизм пережевывания пищи. На нижних
премолярах и первом моляре протиции (единственные найденные ее зубы) нет
вполне сформированных поперечных гребней, но имеются крупные, четко выраженные
тупые бугорки, расположенные таким образом, что из них могли бы развиться
(и, вполне вероятно, развились) такие гребни.
Теперь о проблеме, возникшей в связи с переизучением Паттерсоном черепа
Pyrotherium (рис.16). У Notoungulata имеются многочисленные признаки, обычные
для более примитивных членов этого отряда и видоизменяющиеся, но не исчезающие
полностью у более специализированных форм. Наиболее очевидный из этих признаков
— сложное строение ушной области, в своих деталях не встречающееся ни у
каких других животных, а также столь же поразительное и столь же неповторимое
и даже еще более бросающееся в глаза строение зубов, в частности моляров,
хотя и не только их. Паттерсон обнаружил, что строение ушной области у
Pyrotherium сходно с ее строением у нотоунгулят, а поэтому делает вывод,
что «по всей вероятности, Pyrotheria были нотоунгулятами». О зубах он пишет:
«Коренные зубы этих ранних пиротериев не дают никаких определенных указаний
на происхождение и родственные связи этой группы». Кое-какие указания на
происхождение группы они все-таки дают, не обязательно определенные, но
безусловно наводящие на размышления. Как уже отмечал Амегино, а вслед за
ним и Паттерсон, коренные зубы пиротерия вполне могли бы происходить от
кондиляртр или, точнее, от дидолодонтид. Это особенно ясно в случае Proticia,
имеющей самое примитивное среди пиротериев строение коренных зубов.
Паттерсон предполагает, что между коренными зубами пиротериев и примитивных
нотоунгулят существует, возможно, некоторое сходство, но мне это сходство
кажется обманчивым. В частности, Паттерсон пишет, что зубы примитивных
нотоунгулят имели тенденцию к образованию на задней части нижних моляров
поперечного гребня, включавшего в себя бугорки, именуемые в научной терминологии
энтоконидом и гипоконидом, и не включавшего бугорок, называемый гипоконулидом.
Если согласиться с этим, то можно было бы говорить о его сходстве с задним
гребнем нижних моляров пиротерия. Но поперечный гребень в задней части
моляров примитивных нотоунгулят (талонид) образуется не за счет энтоконида
и гипоконида, а за счет одного только энтоконида. Иногда он упирается в
основание гипоконида, но не захватывает этот бугорок. Наиболее примечательная
черта этой части нижних моляров нотоунгулят — длинный полулунный гребень,
охватывающий гипоконид и гипоконулид; представить же себе наличие у предков
пиротериев такой структуры почти невозможно. И в других отношениях о таком
происхождении зубной системы пиротериев, по-видимому, не может быть речи.

Рис. 16. Череп Pyrotherium (типовой род пиротериев) из раннего олигоцена,
с реставрацией недостающих частей, выполненной Паттерсоном, и реконструкция
его головы.
Хотя некоторые кости посткраниального скелета известны, для реконструкции
туловища их недостаточно.
Итак, к чему же мы в итоге приходим? К тому, что у пиротерия был, возможно,
предок, у которого ушная область была устроена, как у нотоунгулят, а зубы
— как у кондиляртр. Такой предок мог бы дать начало также и нотоунгулятам,
если бы в результате эволюции по совершенно иной линии у него возникли
зубы нотоунгулятного, а не пиротериевого типа. Как же мы должны называть
этого предка, если таковой существовал? Кондиляртром, поскольку его зубы
были кондиляртрового типа? Нотоунгулятом, поскольку по строению ушной области
он сходен с нотоунгулятами? Пиротерием, поскольку пиротерии стали самыми
крупными и самыми удивительными из его ранних потомков? Или же, раз выбор
столь широк, не воспользоваться ни одним из этих названий? Лично я считаю,
что не следует брать ни одно из этих названий, до тех пор пока не удастся
действительно найти более далеких предков пиротериев и нотоунгулят, чем
известные нам сейчас. Следует отметить также, что имеется еще одна возможность.
И пиротерии, и нотоунгуляты могли произойти от кондиляртр и в то время
еще не имели ни таких зубов, как у пиротериев или нотоунгулят, ни такого
строения ушной области, как у нотоунгулят. Затем пиротерии и нотоунгуляты
могли развиваться дивергентно в отношении зубов, но параллельно в отношении
строения ушной области. Пока же я предпочитаю условно оставить пиротериев
самостоятельным отрядом. Эта проблема, конечно, не относится к числу тех,
которые могут потрясти мир, и была изложена здесь с некоторыми подробностями
лишь в качестве примера того, с какими трудными и пока еще неразрешимыми
проблемами приходится иногда сталкиваться исследователям истории жизни.
В отношении отряда Astrapotheria подобных проблем не возникает. В строении
их коренных зубов имеются некоторые признаки сходства с нотоунгулятами,
но таких признаков немного, и они, по всей вероятности, конвергентны. В
строении ушной области или других структур никакого особого сходства с
нотоунгулятами нет, и в настоящее время практически все согласны с тем,
что эту группу следует считать отдельным отрядом. Отряд этот никогда не
отличался большим разнообразием, но он известен от касамайория до фриасия,
т. е. примерно от раннего эоцена до среднего миоцена, а ареал его простирался
от Южной Аргентины до северной Венесуэлы и Колумбии.
Древнейший астрапотерий — Scaglia из касамайория — известен лишь по единственному
экземпляру, представляющему собой часть черепа и верхнюю челюсть молодой
особи с молочными зубами. По имени астропотерия Astraponotus Амегино назвал
время, соответствующее нашему теперешнему мастэрсию, но данный род редко
встречается в сборах из отложений этого века и недостаточно хорошо известен.
Астрапотерии десеадия и колуапия изучены довольно хорошо, а из сантакрусия
известен почти полный скелет собственно Astrapotherium (рис. 17). Об этих
поздних представителях семейства Паттерсон и Паскуаль говорят, что «они
были двух размеров — крупного и очень крупного».
В черепе крупных олигоценовых и миоценовых астрапотериев над глазницами
выдавался вперед конусообразный лоб; очень короткие носовые кости позволяют
предполагать, что животное имело небольшой хобот; большие клыки в течение
всей жизни оставались острыми, затачиваясь о несколько более короткие нижние
клыки. Возможно, что нижние клыки Использовались для выкапывания корней.
Верхних резцов не было, но зато имелось шесть нижних резцов с изящными
двулопастными коронками. Коренные зубы (обычно пять верхних и четыре нижних)
имели довольно высокие коронки, но при этом несли простые гребни, характерные
для форм, питающихся листьями и побегами. Туловище было вытянутым, а ноги,
в особенности задние, были неожиданно слабыми для столь крупных зверей.
Лапы были относительно маленькими, причем задние — явно стопоходящего типа
т. е. при ходьбе животное опиралось на всю стопу. Скотт высказал предположение,
что явно неуклюжее, непропорциональное телосложение этих животных, возможно,
указывает на то, что они вели земноводный образ жизни. Астрапотерии обладали
некоторым конвергентным сходством с вымершей группой носорогов северного
полушария, называемых аминодонтами, которые также, возможно, были земноводными.
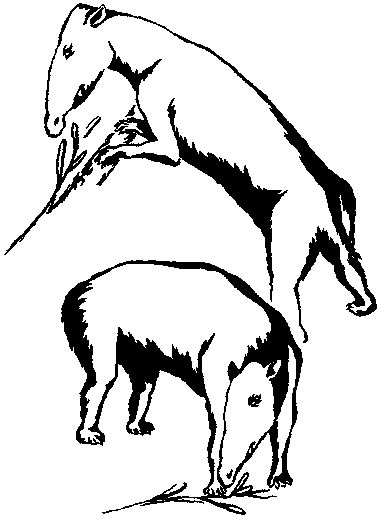
Рис. 17. Реконструкция типичного астрапотерия (Astrapotherium) из раннего миоцена (сантакрусия).
Остатки Trigonostylopoidea обильны в сборах из риочикия Бразилии и касамайория
Аргентины, но присутствие их в мастэрсии уже сомнительно, а позже они и
вовсе не известны. Долгое время их считали довольно близкими родичами астрапотериев,
но дополнительное изучение материала, в том числе ранее не известного хорошо
сохранившегося черепа (рис. 18), показало, что по многим морфологическим
признакам они совершенно не похожи на астрапотериев, в частности по строению
ушной области, которая у них совсем не такая, как у нотоунгулят. Сходство
с астрапотериями сводится к тому, что у них тоже были верхние и нижние
клыки, но небольшие и отличающиеся в деталях от клыков астрапотериев. Верхние
моляры с низкой коронкой, с гребнями, по очертаниям более или менее треугольные
(у риочикских форм в меньшей степени) и не похожие на характерные моляры
астрапотериев с четырьмя неравными сторонами. Почему эти животные процветали
именно в данном месте и именно в данное время и почему вскоре после этого
они вымерли, остается одной из многих загадок истории.
В заключение этой главы рассмотрим отряд Xenungulata, известный только
из риочикия. Его представители были самыми крупными и, как подразумевает
их название («странные копытные»), одними из самых необычных зверей своего
времени. Единственный известный род, Carodnia, был впервые обнаружен в
Патагонии, но представлен значительно полнее в бразильском риочикии, в
Итабораи. У него были долотообразные резцы, небольшие клыки и моляры (верхние
и нижние) с двумя острыми гребнями, поперечными, хотя и изогнутыми. Известна
его передняя лапа — тяжелая, напоминающая слоновью, с пятью пальцами, заканчивающимися
относительно маленькими, широкими и плоскими копытами.
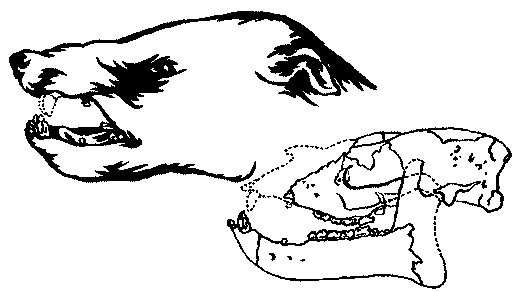
Рис. 18. Реставрированный череп и реконструкция головы типичного тригоностилопоида (Trigonostylops) из раннего эоцена (касамайория).
Паула Коуто, который описал экземпляры из Итабораи и на этом основании
выделил отряд Xenungulata и дал его характеристику, отметил некоторое сходство
этих животных с давно вымершими раннекайнозойскими копытными Северной Америки
и Азии — диноцератами (что означает «ужаснорогие»; в отличие от них ксенунгуляты
не имели рогов). Он думал, что это, возможно, указывает на филогенетическое
родство, но лишь через общего предка, относившегося к отряду Condylarthra.
В результате некоторые палеонтологи, сами не изучавшие названные группы,
например А. Ромер (A. Romer), сделали Xenungulata и Dinocerata подотрядами
одного и того же отряда, что почти неизбежно подразумевает северное происхождение
ксенунгулят, однако подобная точка зрения не получила общего признания.
Остается вероятным, что ксенунгуляты возникли в Южной Америке и что их
поистине весьма ограниченное сходство с диноцератами было обусловлено довольно
умеренной конвергенцией.
Паттерсон повторно изучил экземпляры Trigonostylopoidea и Xenungulata из
Итабораи. Недавно в частном письме он высказал мнение, что вопреки его
прежним взглядам обе эти группы копытных следует включать в отряд Astrapotheria.
Все, в том числе сам Паттерсон, согласны с тем, что группы, называемые
Trigonostylopoidea, Xenungulata и Astrapotheria, — валидные самостоятельные
группы (таксоны, на языке систематики), причем различия между ними столь
велики, что они заслуживают более высокого ранга, чем семейство. Это —
наиболее важный момент; менее важным представляется решение вопроса о том,
считать ли их тремя подотрядами отряда, который, возможно, следует называть
Astrapotheria, или тремя отдельными отрядами. До тех пор пока не опубликованы
доказательства того, что эти группы представляют собой эволюционные единицы,
связанные друг с другом теснее, чем просто общим происхождением от кондиляртр,
здесь временно сохраняется ставший привычным статус трех отрядов. Если
бы было показано, что они имели общего предка, принадлежащего им одним,
то возник бы вопрос, считать ли, что этот предок был астрапотерием, тригоностилопоидом,
ксенунгулятом или, что, по-видимому, более вероятно, ни тем, ни другим,
ни третьим.
В связи с этим укажем на один факт, ранее не публиковавшийся, а именно
что характерные кости стопы (таранная и пяточная), почти наверное принадлежащие
Trigonostylops, коренным образом отличаются от этих же костей астрапотериев.
У ксенунгулят эти кости не найдены.
ЛИТЕРАТУРА
Patterson В. 1977. A primitive pyrothere (Mammalia, Notoungulata) from
the early Tertiary of northwestern Venezuela. Fieldiana Geology, 33,
397 — 422. (Эта статья содержит также переописание черепа Pyrotherium
из Аргентины.)
Paula Couto С. de. 1952. Fossil mammals from the beginning of the Cenozoic
in Brazil: Condylarthra, Litopterna, Xenungulata and Astrapotheria. Bulletin
of the American Museum of Natural History, 99, 355 — 394.
Paula Couto C. de. 1963. Um Trigonostylopidae do Paleoceno do Brasil.
Anales Academia Brasileira de Ciencias, 35, 339 — 351.
Gaudry A. 1909. Fossiles de Patagonie: le Pyrotherium. Annales de Paleontologie,
Paris, 4, 28 p.
Scott W. B. 1910. Litopterna of the Santa Cruz beds. Reports of the Princeton
University Expeditions to Patagonia, 7, 1 — 156.
Scott W. B. 1937. The Astrapotheria. Proceedings of the American Philosophical
Society, 77, 309 — 393.
Simpson G. G. 1933. Structure and affinities of Trigonostylops. American
Museum Novitates, 608, 1 — 28.
См. также литературу к гл. 1 и ссылку на работу Simpson, 1948, 1967 в
конце гл. 6.
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..............................................................................................................................
5
Предисловие ................................................................................................................................................................
8
Глава 1. ПОЧЕМУ, ЧТО И КАК? .............................................................................................................................
11
Глава 2. ВЗГЛЯД В ПРОШЛОЕ: КАК ЗАКЛАДЫВАЛИСЬ
ОСНОВЫ ...............................................................16
Глава 3. ВРЕМЯ И ФАУНЫ .....................................................................................................................................
29
Глава 4. ДЕЙСТВУЮЩИЕ ЛИЦА ...........................................................................................................................
38
Глава 5. КОРНИ ........................................................................................................................................................
43
Глава 6. ПЕРВАЯ ФАЗА: ОДНИ СТАРОЖИЛЫ ...................................................................................................
58
Глава 7. СУМЧАТЫЕ ...............................................................................................................................................
67
Глава 8. КСЕНАРТРЫ — СТРАННО СОЧЛЕНЕННЫЕ МЛЕКОПИТАЮЩИЕ
................................................ 82
Глава 9. НЕКОТОРЫЕ СТАРОЖИЛЫ ИЗ КОПЫТНЫХ
..............................................................................
98
Глава 10. НОТОУНГУЛЯТЫ .................................................................................................................................
110
Глава 11. ВТОРАЯ ФАЗА: ПОЯВЛЯЮТСЯ ЧУЖЕЗЕМЦЫ
...............................................................................
131
Глава 12. ДРЕВНИЕ МЕСТНЫЕ ГРЫЗУНЫ ........................................................................................................
150
Глава 13. ПРИМАТЫ ..............................................................................................................................................
163
Глава 14. ТРЕТЬЯ ФАЗА: ВЕЛИКИЙ АМЕРИКАНСКИЙ
ОБМЕН ....................................................................
174
Глава 15. ВЕЛИКИЙ АМЕРИКАНСКИЙ ОБМЕН: ЧАСТНЫЕ
АСПЕКТЫ И АНАЛИЗ .................................. 187
Глава 16. НОВИЧКИ И СОВРЕМЕННАЯ ФАУНА ..............................................................................................
205
Глава 17. ИСТОРИЯ ЮЖНОАМЕРИКАНСКИХ МЛЕКОПИТАЮЩИХ
КАК ЭКСПЕРИМЕНТ И ДОКУМЕНТ ...................................................................................................................
228
Приложение. ЗАМЕТКА О ВОССТАНОВЛЕНИИ ОБЛИКА
ВЫМЕРШИХ ЖИВОТНЫХ .............................. 242
Предметно-именной указатель ................................................................................................................................
246
Указатель латинских названий ................................................................................................................................
252