
"Лошадиная фамилия"
Часть II
| Главная |
|
"Лошадиная фамилия" Часть II |
|||
6. Вымерший Hyracotherium (Eohippus) был почти идентичен по строению тела, ногам, пальцам и размерам ныне живущему даману, исключая череп и хвост, и является предком лошади не больше, чем предком носорога!
Мы не доказываем, что Hyracotherium - то же самое животное как Hyrax, но есть примечательные черты сходства.
Действительно ли Hyracotherium (Eohippus) был лошадью? Hyracotherium был обнаружен в Европе прежде, чем “Eohippus” был открыт в Северной Америке, и название рода Hyracotherium дано известным британским анатомом и палеонтологом, Ричардом Оуэном, который был также его первооткрывателем. Позже в Северной Америке были обнаружены другие экземпляры, и роду было дано название Eohippus. Впоследствии было выяснено, что североамериканские экземпляры принадлежали фактически к тому же самому роду, что Hyracotherium. Последнее название, таким образом, имеет приоритет, так что Eohippus - не обоснованное название для этих существ. Оно, однако, наиболее часто используется, несомненно потому, что название Eohippus означает “ранняя лошадь”, в то время как имя Hyracotherium было выбрано Оуэном из-за подобия этого существа животным рода Hyrax (даман). (The Origin Of Mammals, ICR Impact No. 87 Duane T. Gish, Ph.D, 1980)
Nilsson указал, что, в то время как Hyracotherium имеет мало черт сходства с лошадям, или вовсе никаких, он имеет явное морфологическое сходство и сходство в среде обитания с ныне живущими животными рода Hyrax. (Nilsson, H., Synthetische Artbildung, Verlag CWE Gleerup, Lund, Sweden, 1954. [See F.W. Cousins, Ref. 16, for a summary on the horse.]) Hyrax, подобно Hyracotherium, размером с кролика или с лисицу. Hyrax, также подобно Hyracotherium, имеет четыре пальца на передних ногах и три на задних. Щёчные зубы у этих двух существ имеют много сходства, и скорее похожи на таковые у носорогов, чем у лошадей. Среда обитания и образ жизни Hyrax также подобны тем, что предполагаются для Hyracotherium. Таким образом, Nilsson поддерживает мнение, что, хотя Hyracotherium никаким образом не напоминает современных лошадей, он был, очевидно, замечательно подобен современному Hyrax. Другие также сомневаются относительно того, был ли Hyracotherium связан с лошадью. Например, Kerkut утверждает, что “во-первых не ясно, что Hyracotherium был предковой лошадью. Таким образом утверждал Симпсон (1945), ‘Мэтью показал и упорно утверждал, что Hyracotherium (включая Eohippus) настолько примитивен, что это не более определенно лошадеобразное, нежели представитель тапиров, носорогов, и т.д., но было общепринято помещать его в основании группы лошадей.’” (Kerkut, G. A., Implications of Evolution, New York: Pergamon Press, 1960, p. 149) Другими словами, Hyracotherium не больше подобен лошади, чем тапиру или носорогу, и таким образом столь же оправданно он мог быть выбран как предок носорога или тапира. Тогда кажется, что объективность тех, кто был вовлечён в строительство филогенетического дерева лошади, была сомнительна с самого начала, и что “лошадь”, на которой целиком зиждется родословное дерево лошади, вообще не была лошадью. После публикации книги Керкута не было издано никакой достаточно полной работы о лошадях, которая существенно повлияла бы на его заключение, что “в определённой мере всё выглядит, как будто путь развития лошади мог быть даже столь же хаотическим, как предложенный Осборном (1937, 1943) для развития Proboscidea, где “почти ни в каком случае никакая известная форма не рассматривается как потомок любой другой известной формы; для каждой подчиненной группы принято предположение о происхождении, весьма изолированно и обычно совсем без известной промежуточной стадии, от гипотетических общих предков в раннем эоцене или позднем мелу (Romer 1949)”. (Kerkut, G. A., Implications of Evolution, New York: Pergamon Press, 1960, p. 149) Если действительно “эволюция лошади” столь же хаотична и фрагментарна, этот классический случай не имеет реальных достоинств для эволюции. Фактические свидетельства, с другой стороны, аккуратно соответствуют Модели Сотворения. (The Origin Of Mammals, ICR Impact No. 87 Duane T. Gish, Ph.D, 1980)
Собственно говоря, то, что гиракотерий (он же эогиппус) был,
возможно, достаточно близок к предкам носорогов и тапиров, как к предкам лошадей,
ещё не означает того, что он не мог быть предком лошади. Если взглянуть на эту
проблему с другого конца эволюционного древа, “снизу вверх”, то можно сказать,
что носороги и тапиры сохранили больше, нежели лошади, предковых черт, которыми
обладал гиракотерий. Вот если бы у гиракотерия нашли уникальные черты носорога
или тапира, которых заведомо не могло быть у лошади, тогда было бы совсем другое
дело – гиракотерия пришлось бы исключить из родословного древа лошадиных. Если
от гиракотерия (или родственных ему форм) произошли тапиры или носороги, то
это ещё не исключает возможности происхождения лошадей от представителя этого
же рода.
Само собой, гиракотерий был не полностью лошадью, сохраняя черты предковых видов.
Но с такой же долей уверенности мы можем утверждать, что это не был типичный
носорог или тапир. С такой же долей уверенности его можно исключить из родословных
носорогов и тапиров. Тем более, что современником гиракотерия был Hyrachius
– предок (или, по крайней мере, один из древнейших представителей) носорогов.
Что же касается сходства гиракотерия (эогиппуса) с даманом, то я боюсь, что
господа креационисты не столь единодушны в суждениях. Вот что пишет про это
животное книга “Жизнь - как она возникла? Путём эволюции или путём сотворения?”
(автор не указан), WATCH TOWER BIBLE AND TRACT SOCIETY OF NEW YORK, INC., страницы
66-67:
“Эогиппус, предположительно древнейшая лошадь, которая, по мнению экспертов, давно вымерла и известна нам только по ископаемым остаткам, может быть, жив и здоров и вообще является не лошадью, а даманом, пугливым животным размером с лисицу, обитающим в чащобах Африки”.
Оп-паньки! Прямо и открыто, чёрным по белому книга креационного
содержания (а что ещё можно ждать от иеговистов?) равняет гиракотерия (эогиппуса)
с даманом. Причём это сказал один из противников теории эволюции, Хитчинг (см.
анализ возражения № 11), которого не назвавшие себя авторы почему-то записали
в эволюционисты…
А как же обстоят дела на самом деле? Если сравнивать дамана и гиракотерия (эогиппуса),
можно найти ряд существенных черт различия.
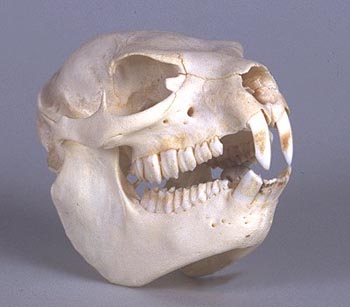
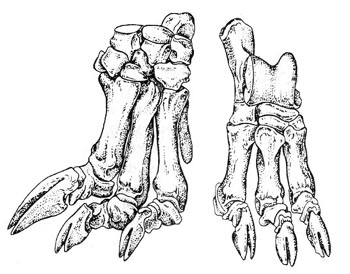
1. Даман является откровенно стопоходящим животным. У него на задних ногах есть одна замечательная особенность – стопа образует своего рода “присоску”, позволяющую лазить по скалам. На внутреннем пальце стопы растёт длинный искривлённый коготь. Гиракотерий, он же эогиппус – пальцеходящее животное с короткими копытообразными когтями.
2. У дамана практически нет хвоста, заметного снаружи. У примитивных лошадей хвост есть, причём явно заметный и довольно длинный.
3. Разница в зубной системе у современного дамана и эогиппуса весьма значительна. Зубная формула современных даманов 1/2-0/0-4/4-3/3. Позади резцов присутствует диастема – лишённый зубов промежуток. Резцы самозатачивающиеся и постоянно растущие, как у грызунов: это приспособление для питания грубой высокоабразивной пищей. Стоит сказать также, что некоторые палеонтологи отмечают, что рисунок бугров и гребней на жевательной поверхности зубов дамана напоминает таковой у непарнокопытных (это упоминает Р. Кэрролл, не уточняя, о каких непарнокопытных идёт речь). Справедливости ради отмечу, что утрата боковых резцов и клыков произошла лишь у поздних, плейстоценовых и современных видов (Р. Кэрролл). Но ведь с ними-то как раз идёт сравнение! У гиракотерия (эогиппуса) зубная формула такова: 3/3-1/1-4/4-3/3. Клыки, кстати, не утрачены и у современных лошадей. Зубы примитивнейших лошадей низкие и бугорчатые, приспособлены для питания мягкой растительной пищей. Что же до сходства зубов дамана и гиракотерия (эогиппуса) с зубами носорога – так, может быть, это носорог сохранил больше черт примитивных копытных, нежели лошади с их специализированными зубами?
 |
 |
 |
|
4. Лицевые кости дамана укорочены: глазницы в передней части черепа, и зрение бинокулярное. У гиракотерия (эогиппуса) глазницы – примерно на середине длины черепа, о чём говорят сами креационисты на этом же сайте в описаниях древних лошадиных. Глаза гиракотерия (эогиппуса) смотрят большей частью в стороны, и их поле зрения лишь слегка перекрывается.
5. Биохимические признаки объединяют даманов с хоботными – этот факт широко известен и освещён даже в популярной литературе. А непарнокопытные (среди которых числятся лошади) по биохимическим показателям стоят особняком от всех современных отрядов копытных. Даже парнокопытные биохимически (а по данным палеонтологии – и анатомически) ближе к китообразным, чем непарнокопытные к какому-либо иному отряду современных копытных.
6. Креационисты – большие любители считать рёбра и прочие кости (см. аргументы №№ 8 и 9). Так вот, у даманов позвоночник состоит из 20-23 грудных и 4-9 поясничных позвонков. У гиракотерия (эогиппуса), согласно материалам обсуждаемого сайта, 18 пар рёбер (и, соответственно, грудных позвонков). О количестве поясничных позвонков этого животного креационисты умалчивают. Возможно, информация о шести поясничных позвонках (см. аргумент № 9) относится как раз к этому зверю.
7. Экология дамана и гиракотерия (эогиппуса) сильно различается. Даманы – типичные обитатели открытых сухих мест (скалы, саванны и полупустыни). “Чащобами” в местах обитания даманов даже не пахнет (не исключено, конечно, что здесь просто неверно переведено слово "bush", обозначающее просто дикую местность). Даманы могут ловко лазить по деревьям и камням, пользуясь присосками на ступнях. Гиракотерий (эогиппус), судя по ископаемым находкам и особенностям зубов, жил в лесу и питался мягкой растительностью. Поскольку он был пальцеходящим, ждать от него способностей верхолаза бессмысленно: его ноги явно не хватательного типа, а цепляться копытообразными когтями за кору и камни нельзя.
Подробно о сравнении гиракотерия (эогиппуса) с даманом можно
прочитать в статье Майкла Хопкинса “Является ли “ранняя лошадь” даманом?”, которую
можно найти здесь:
http://www.talkorigins.org/faqs/horses/eohippus_hyrax.html
В этой статье приводится сравнение черепов и зубов данных животных, и перечисляются
сайты креационистов, которые “пудрят мозги” доверчивым посетителям заведомо
ложной информацией по этому вопросу. Среди них, кстати, указан сайт, с которого
были взяты обсуждаемые здесь пятнадцать тезисов креационистов. Также в данной
работе упоминаются другие “ляпы” креационистов.
Здесь же говорится о том, что даман был не единственным зверем, с которым сэр
Ричард Оуэн сравнил гиракотерия: в оригинальном описании упоминаются также заяц
и свинья (в оригинале сказано “hog”). Просто название ввело креационистов в
искушение чуть-чуть соврать. Даман ведь упоминается в Библии – Великом Первоисточнике
вдохновения.
О том, была ли родословная древних лошадиных кустом или деревом, и достойна
ли она быть примером эволюции, будет сказано дальше.
7. Собранные вместе ископаемые из многих местонахождений, как работа наугад, отягощённая предубеждением.
-“Свидетельства” добывались из разных геологически изолированных областей. Ряд начинается в Северной Америке, внезапно скачет в Европу, и затем назад в Америку. Wysong, R.L., The Creation-Evolution Controversy, Inquiry Press, Midland, Michigan, 1981, p. 455)
-Полный ряд окаменелостей в правильном эволюционном порядке не существует нигде в мире. (Kofahl, R.E., Handy Dandy Evolution Refuter, Beta Books, San Diego, California, 1997, p. 159)
-Ряд лошади был построен из окаменелостей, найденных в множестве различных частей мира, и нигде эта последовательность не оказывается в одном местоположении. Развитие лошади смонтировано путём расстановки различных окаменелостей, найденные в Индии, Южной Америке, Северной Америке и Европе, в ряд от самой маленькой до самой большой. (Современные лошади варьируют от 17 до 80 дюймов в высоту)
Вообще, если мы посмотрим (уже в какой раз!) на родословные
древа лошадей, любезно предоставленные нам креационистами, то заметим, что именно
североамериканские остатки древних лошадиных, и только они образуют непрерывный
ряд, и “скачков” основной эволюционной линии лошадей из Европы в Америку не
происходило! А вот из Америки в Европу лошади действительно расселялись неоднократно
и в разное время. Поэтому в Европе формы доисторических лошадей не образуют
непрерывной последовательности. Это отражено, например, в написанной в 1898
году книге Мельхиора Неймайра “История Земли”. Поэтому лошади из Европы, Южной
Америки и Индии стоят на боковых ветвях, вне “магистральной” линии, что привела
к формированию современной лошади. Конечно, они дополняют родословное древо
лошадей, и их нельзя сбрасывать со счетов, но они – лишь производные от основного
ствола.
Другое дело, что все эти находки не представляют собой некоего единого месторождения,
где прямо один на другом лежат скелеты – от примитивного до современного. Что
ж, такого местонахождения и вправду нет. Это связано с тем, что районы накопления
осадков, где происходило захоронение останков лошадей и других доисторических
животных, являются локальными, и существуют относительно недолго. Поэтому аргумент
креационистов в данном случае – “хочу то, чего нет”.
Два следующих аргумента были получены путём простых арифметических действий:
8. Изменения количества рёбер.
-Количество рёбер сначала уменьшается, затем внезапно увеличивается, и затем снова уменьшается. Hyracotherium имел 18 пар рёбер, Orohippus имел 15, Pliohippus имел 19, лошадь имеет 18 пар.
9. Изменение числа позвонков.
-Число поясничных позвонков также изменяется от шести до восьми и затем назад к шести.
“Кроме того, если лошадок расставить в воображаемой эволюционной последовательности, то окажется такая “эволюция” количества ребер: сначала 18, потом 15, потом 19, наконец снова 18. Подобные же вариации наблюдаются и в количестве позвонков”. (Православное Мiровоззрение и современное естествознание)
В русском языке есть выражение “пересчитать рёбра”. Означает
оно, правда, совсем не то, что хотят показать наши креационисты, но, если они
так желают, можно заняться и такой арифметикой.
Насколько существенно для родословного ряда лошадей количество рёбер? Не столь
уж и существенно. Дело в том, что количество костей в скелете даже у человека
(“венец творения”, “образ и подобие”, и прочая, и прочая…) может меняться, и
точно не определено – в разных источниках называют разные значения, которые
колеблются в некоторых пределах. А ведь человек – это один биологический вид
одного рода! Так, те же самые дополнительные рёбра у человека могут появляться
и исчезать. Не исключено, что у древних лошадей мог существовать некоторый полиморфизм
в числе рёбер, и в некоторых случаях крайний вариант мог закрепляться как норма.
Кстати, господа креационисты, скажите “спасибо!” мистеру Джонатану Сарфати за
такую информацию:
“Современные лошади могут иметь 17, 18 или 19 пар ребер”. (Джонатан Сарфати “Non-evolution of horse”)
Желающие лично убедиться в этом могут посетить соответствующую
страничку в Интернете:
http://www.answersingenesis.org/creation/v21/i
3/horse.asp
и собственными глазами увидеть эти строки в разделе “Lots of different horses”
его статьи. Думаю, все поняли, что этим признанием он сделал пересчёт лошадиных
рёбер бессмысленным занятием и свёл один из аргументов к нулю.
А что касается количества позвонков – здесь есть пример ещё более потрясающий.
Известно (и приводится как хрестоматийный пример), что у всех зверей – от кита
до жирафа – семь шейных позвонков. Но у трёхпалого ленивца, обитающего в наши
дни в Южной Америке, их количество колеблется (в пределах одного вида!) от шести
до девяти.
“На уровне мутаций трудно провести грань между небольшими скачками и постепенными (градационными) изменениями. Однако известны и случаи, когда относительно крупные перестройки происходят скачком. Такие сальтации встречаются у птиц при переносе сосудов с одного ствола на другой. У большинства птиц таким путем исчезает правая сонная артерия. Это скачок. Путем своеобразных сальтаций изменяется число позвонков, образуется новые анастомозы – перемычки между сосудами”. (Л. П. Татаринов “Суждение палеонтолога об эволюции”)
Итак, является ли в свете этих примеров пересчёт рёбер и позвонков
лошадей существенным возражением против эволюции? Думаю, нет. Ведь изменение
сегментации позвоночника, хоть и происходит на очень ранних стадиях развития
зародыша, является результатом немногих изменений в генотипе, которые вполне
могут происходить на видовом уровне.
Далее, ещё можно сказать, что учёные не всегда находят представителей именно
той самой “стволовой линии”. Ведь те названия, что пишутся в родословных линиях
разных животных – это родовые названия. А внутри каждого рода в пространстве
и времени сменяли друг друга разные виды животных. Так, если от рода А произошёл
род Б, то это ещё не означает, что все виды рода А превратились в виды рода
Б. Это сделал всего один какой-то вид, и это событие не мешало прочим видам
рода А развиваться своим путём: по определению, виды являются совокупностями
особей, достаточно (а в ряде случаев – полностью) изолированными от иных подобных
совокупностей. И вряд ли все популяции всех видов некоего рода живых существ
сохранятся в палеонтологической летописи. Часто просто нет уверенности, найден
ли представитель “стволовой линии”, или это несколько специализированная и уклонившаяся
форма. Вот, кстати, один из источников различий во взглядах авторов на лошадиное
родословное древо, о чём, разумеется, не могут умолчать креационисты:
10. Предложено множество различных родословных древ лошади.
-Нет никакого согласия по поводу родословной лошади. Вместо единственного ряда, который изображен в учебниках, было изобретено более 20 различных “древ лошади”, чтобы силой протянуть теорию через противоречащие свидетельства. Это доказывает, что родословные древа – догадки и спекуляция. (cf. Bowden, M., The Rise of the Evolution Fraud, Creation-Life Publishers, San Diego, California, 1982, p. 117)
Различие в родословных древах лошадей, представленных разными
авторами, может происходить от того, что авторы по-разному понимают механизмы
формирования каких-либо признаков, присутствующих у ископаемых видов. Ведь один
и тот же признак, имеющийся в наличии у двух родственных форм, может быть истолкован
двояко: как результат происхождения от общего предка, уже имевшего этот признак,
либо как результат независимого формирования под воздействием сходных условий
жизни. Во втором случае такое явление носит название параллелизма.
При восстановлении направлений эволюции любой группы живых существ важно как
раз различить, что есть итог параллелизма, а что – происхождения от общего предка.
“При них [параллелизмах в эволюции – В. П.], кроме зависимости от функции, отчетливо проявляется зависимость приобретаемого признака от внутренних особенностей организма. Самая частая форма параллелизма – многократное появление сходных особенностей строения в группах родственных организмов. Параллелизмы могут обнаруживаться на очень глубоких уровнях, - вплоть до уровня генотипа. Генетические параллелизмы, например, являются важной составляющей в знаменитой “Периодической таблице признаков” - гомологических рядах Николая Ивановича Вавилова”. (Л. П. Татаринов “Суждение палеонтолога об эволюции”)
Каждая новая находка, естественно, меняет представления исследователей о процессе эволюции, поэтому в разное время предлагались разные варианты родословных древ лошадей (или любых других животных), отражавшие взгляды, господствовавшие в то время, при том уровне знаний. Соответственно, по мере пополнения нашего знания об эволюции изучаемой группы организмов меняется облик её родословного древа – от приблизительного и умозрительного до всё более и более точного и близкого к истине.
“Как сказал биолог Хериберт-Нилссон, “родословное дерево лошади красиво и непрерывно только в учебниках”, и известный палеонтолог Найлз Элдредж назвал учебник “плачевным” и “классическим случаем палеонтологического музееведения”. Как показано в подробной диссертации Уолтера Барнхарта, “ряд лошади” – это интерпретация данных. Он информирует о том, как различные картины эволюции лошади были нарисованы различными эволюционистами на основе одних и тех же данных, как “эволюционировала” сама концепция эволюции”. (Джонатан Сарфати “Non-evolution of horse”)
Собственно говоря, эти слова креационистов также можно считать
не фактом, а просто одной из множества возможных интерпретаций, основанных на
этих же самых находках. Единственное, чем креационисты могут подтвердить свои
слова – это текст Библии. И в свете их собственных слов я могу спросить: а правильно
ли креационисты интерпретируют Библию? Может быть, перевод был сделан не совсем
точно, и на деле всё было совсем не так, как предполагают сами креационисты?
Иными словами, “от дурака слышу!”
Конечно, любой факт, полученный в настоящее время, можно счесть интерпретацией.
Мы не ощущаем радиоактивность, не видим инфракрасных и ультрафиолетовых лучей,
не слышим инфразвуков и не чувствуем магнитных полей. Но это не мешает нам изучать
их и делать выводы, основываясь на субъективно интерпретируемых показаниях приборов.
Точно так же и окаменелости. Мы не можем непосредственно наблюдать процесс эволюции,
и останки лошадей, равно как и множества других видов живых существ, лишь приоткрывают
завесу над этим процессом. Это как портреты одного и того же человека в разном
возрасте – в детстве, юношестве, зрелости, старости. Глядя на эти портреты,
и не зная человека лично, мы не утверждаем, однако же, что в промежутках между
временами, когда были сделаны портреты, этот человек не существовал.
Да, вывод об эволюции есть интерпретация результатов множества наблюдений, находок
и экспериментов. Но примечателен тот факт, что эти результаты непротиворечивы
или могут быть объяснены с позиций теории эволюции. В то же время подход креационистов
отличается большим количеством “натяжек” и недоговорок, в чём, думаю, читатели
уже убедились. Также сами креационисты противоречат друг другу, даже толкуя
(интерпретируя!) слова Библии – “единственно верного” первоисточника.
“Единственной цепочкой, которой могут похвастаться эволюционисты, является так называемая лошадиная серия. В учебниках приводится последовательность из четырех картинок древних лошадок, различающихся ростом и формой копыт (…). Но следует помнить, что такая схема выстроена довольно условно и искусственно. На самом деле в мире найдено не 4, а до 300 видов различных лошадей, причем в пределах одной территории никак не удается найти последовательности эволюционных изменений: внизу мелкие лошадки, далее все крупнее и крупнее вплоть до современных. Напротив, останки многих разновидностей встречаются в одних и тех же захоронениях. Как одни из них могли быть предками других?” (Православное Мiровоззрение и современное естествознание)
Боюсь, автор этих слов посчитал, что все вымершие виды лошадей
должны каким-то образом принять непосредственное участие в эволюции современной
лошади. В противном случае, почему он поминает общее число их видов как возражение?
Да, в родословной современной лошади приводится совсем небольшая цепочка видов,
гораздо меньшая, чем общее число найденных форм. Но сами же креационисты говорят,
что родословное древо лошадей не было столь прямолинейным! И в учебниках по
теории эволюции кроме упомянутой цепочки лошадей приводится рисунок весьма ветвистого
родословного древа. Разница в фактическом многообразии доисторических лошадей
и длине родословной цепочки, ведущей к современным лошадям, происходит от того,
что далеко не все древние лошади были предками современных!
Об эволюции семейства лошадиных хорошо сказал отечественный палеонтолог Л. П.
Татаринов:
“В примере с лошадиными ортоселекция на протяжении десятков миллионов лет действовала в сторону их приспособления к жизни в степных ландшафтах (они развивались в то же геологическое время) – к быстрому бегу и травоядности.
То, что в этом случае мы имеем дело с направленным отбором, а не направленной изменчивостью, подтверждают факты постоянного ответвления от магистрального ствола развития лошадиных разнообразных боковых ветвей. Вымирание – отпадение этих боковых линий – придало картине эволюции лошадиных направленный характер”. (Л. П. Татаринов “Суждение палеонтолога об эволюции”)
Таким образом, прямота эволюционной цепочки лошадей – лишь
иллюзия. Если бы хотя бы гиппарионы продержались чуть-чуть дольше (последние
африканские виды вымерли в среднем плейстоцене), ни о какой прямоте и речи не
было бы.
Но мало того, что лошади последовательно появлялись – они ещё последовательно
вымирали:
“Вопрос для креационистов: креационисты, желающие отрицать свидетельства эволюции лошади, должны взвешенно обдумать вот что: как еще Вы можете объяснять последовательность окаменелостей лошади? Даже если креационисты настаивают на игнорировании переходных окаменелостей (многие из которых были найдены), с другой стороны, как может быть объяснена безошибочная последовательность этих окаменелостей? Неужели Бог сотворил Hyracotherium, затем уничтожил Hyracotherium и сотворил некоторые промежуточные звенья Hyracotherium – Orohippus, затем уничтожил промежуточные звенья и сотворил Orohippus, затем уничтожил Orohippus и сотворил Epihippus, затем позволял эпигиппусу “микроэволюционировать” в Duchesnehippus, затем уничтожил Duchesnehippus и сотворил Mesohippus, затем сотворил некоторые промежуточные звенья Mesohippus – Miohippus, затем сотворил Miohippus, затем уничтожил Mesohippus, и т. д. … каждую разновидность, случайно подобную разновидности, которая появилась как раз перед ней, и появилась сразу после?” (Кэтлин Хант “Эволюция лошади” )
Превосходную статью Кэтлин Хант “Эволюция лошади” (Kathleen
Hunt “Horse evolution”), из которой взяты эти строки, можно найти здесь:
http://www.talkorigins.org/faqs/horses/horse_evol.html
Статья на английском языке отличается полнотой, имеются характеристики основных
родов вымерших лошадей, этапов эволюции в семействе лошадиных, а также ответы
на некоторые вопросы креационистов.
Впрочем, на вопрос Кэйтлин Хант креационисты ответили. В статье некоего Professor
Enigma (они что, фильмов про Бэтмена насмотрелись? Или просто стыдно за враньё?),
которая называется “More Fallacies of Evolution ~ More Reasons That Evolution
Didn't Occur”, и которую можно найти здесь: http://www.reachingforchrist.org/apologetics/fallacies2.php
, говорится следующее:
“What sequence of horse fossils? They are NOT found in order on the same continent. (R.E. Kofahl, Handy Dandy Evolution Refuter, p.65) Moreover, the similarities of all of these horses could be easily explained as variation within the same "biblical kind," or "microevolution," not "macroevolution." A horse is still a horse. Assuming evolution occurred, which this author has gratuitously done, is not the only way to explain it. There are further problems with the horse series”.
(“Какая последовательность окаменелостей лошади? Они НЕ найдены в порядке на одном и том же континенте. (R.E. Kofahl, Handy Dandy Evolution Refuter, p.65) Кроме того, черты сходства всех этих лошадей можно было бы легко объяснять как вариации в пределах одного и того же “ библейского вида” или “микроэволюции”, но не “макроэволюции”. Лошадь - все еще лошадь. Предположение, что произошла эволюция, который этот автор безосновательно сделал, - не единственный способ объяснить это”.)
Дальше “Профессор Загадка” набросился на явление, которое
в этом списке нелепостей значится под номером
4, а по данному вопросу больше ничего не сказал.
И слово “лошадь” в данном случае извольте понимать как “представитель семейства
лошадиных”, то есть, “не-носорог” или “не-тапир”. Действительно, если исключить
рано отделившееся от лошадей семейство палеотериев, то окажется, что эволюция
(или, если угодно, изменение внутри “библейского вида” (в Синодальном переводе
Библии это понятие называется “род”, у моррисиан – “барамин”)) происходила внутри
этого семейства. Но если понимать слово “лошадь” в узком бытовом смысле, только
как Equus caballus и близкие к ней дикие виды (тарпан и лошадь Пржевальского),
то окажется, что осёл, кулан, онагр, зебра и квагга окажутся уже вне этого “рода”.
Тот же осёл, хоть чисто биологически является лошадью (представителем род Equus),
действительно вовсе не является арабским скакуном.
Так вот, к примеру, гиппарион (представитель рода Hipparion) – это лошадь или
не-лошадь? Входит он в “род” лошадей или не входит? Если вы ответили на этот
вопрос адекватно, задайте себе другой вопрос: гиракотерий (представитель рода
Hyracotherium (опять специально подчёркиваю, что не Equus)) – это лошадь, или
как? Думаю, “Профессор Загадка” не зря предпочёл не обсуждать тему далее.
“Были ли лошади изначально разными видами в пределах одного семейства или относились к одному роду, - в любом случае лишнее ребро или лишнее копытце - это не замена чешуи на шерсть или перья. Общее правило, установленное всеми палеонтологами, таково: чем выше таксономическое деление от семейств и отрядов к классам и типам, тем меньше вероятность встретить какую-либо форму, которую с какой угодно натяжкой еще можно толковать как переходную. Между классами еще встречаются мозаичные формы, между типами их уже нет, тем более форм переходных.” (Православное Мiровоззрение и современное естествознание)
Креационисты очень любят проводить черту между микро- и макроэволюцией. Но в этом как раз состоит их большая ошибка, поскольку радикальных различий между механизмами микро- и макроэволюции просто нет! Примеры, описанные исследователями, подтверждают это:
“Уилсон (Wilson, 1971) описал прогрессивную эволюцию примитивных селенодонтных парнокопытных, у которых популяции одного раннего вида дали представителей разных родов, а затем и семейств”. (Р. Кэрролл “Палеонтология и эволюция позвоночных” М., Мир, т. 3, стр. 151)
“... радиация перспективных отрядов копытных, например парнокопытных и непарнокопытных Северной Америки и нотоунгулат Южной Америки протекала в общих чертах по одной и той же схеме (...). Признаки, ранее сохранявшие внутривидовую изменчивость, стабилизировались и усиливались, признаваясь в итоге отличительными чертами семейств и отрядов”. (Р. Кэрролл “Палеонтология и эволюция позвоночных” М., Мир, т. 3, стр. 151)
В данном случае явления микроэволюционного плана переросли в макроэволюционные из-за успеха отдельных видов (и даже не их самих, а их потомков) на арене жизни. Кажущаяся пропасть между представителями сколь угодно высоких таксономических единиц – следствие их независимого развития в течение длительного времени. Если посмотреть на “исходный материал” в этих группах, то окажется, что макроэволюционные преобразования сводятся к микроэволюционным.
11. Equus и Hyracotherium сосуществовали.
Две современных разновидности лошади, equus nevadenis и equus occidentalis, найденные в той же самой скальной формации в штате Небраска США, что и Eohippus, доказывают,, что обе они жили в одно и то же время. (cf. Wysong, R.L., The Creation-Evolution Controversy, Inquiry Press, Midland, Michigan, 1981, p. 455, см. также Hitching)
Pettingrew, палеонтолог, говорит, что современная лошадь предшествовала Hyracotherium 70 миллионов лет. Согласно ему, лошадь жила в мезозойскую эру, на 120 миллионов лет раньше, чем Hyracotherium, который появился в эоценовую эпоху, 50 миллионов лет назад.
Находки лошадей современного рода в мезозойских и раннекайнозойских
отложениях можно истолковать как результат неправильного определения возраста
пород, либо как заведомый подлог. Конечно, это вполне согласуется с креационной
теорией, утверждающей, что все животные появились на Земле как результат единого
акта творения, и были современниками друг друга.
Подобные находки известны науке. Как пример можно привести знаменитый череп
современного человека из Калавераса, “найденный” в отложениях возрастом 25 миллионов
лет. Креационисты носятся с этим черепом, словно язычники с идолом, не зная
(или не желая знать, или сознательно умалчивая) обстоятельства появления этой
находки. Так, калаверасский череп был найден на дне карьера во время взрывных
работ. Его возраст был определён как раз тем способом, который креационисты
любят приписывать палеонтологам: по возрасту окружающих слоёв. И, когда череп
из старой индейской могилы, разрушенной взрывом, съехал вниз (в более старые
породы), его ошибочно посчитали происходящим из этих пород. Так новое окружение
черепа изрядно “состарило” его.
Указания господина Петтингрю на находки лошадей в отложениях мезозойской эры
больше смахивают на нездоровую сенсацию типа открытия на Тибете пещеры с сушёными,
но живыми лемурийцами, атлантами и пришельцами с Тау Кита или Большой Медведицы.
Если животные современной эпохи сосуществовали с видами эпохи мезозоя (что должно,
по идее, являться доказательством единовременного сотворения жизни), то в отложениях
мезозоя должно быть найдено гигантское количество ископаемых остатков животных
эпохи кайнозоя или вообще видов, являющихся нашими современниками. И они должны
быть скорее устойчиво повторяющимся правилом, нежели единичным исключением.
Пока же остатки современных лошадей, найденные в мезозое, теряются среди явно
фальсифицированных или неверно определённых образцов и выглядят нонсенсом с
точки зрения науки.
“Следствие по делу” двух странных лошадей из Америки провёл Jon Barber в статье
“Did Hyracotherium and Equus live at the Same Time?”
Эту подробную работу на английском языке можно найти здесь: http://www.talkorigins.org/faqs/horses/eohippus_equus.html
Приведу кратко те аргументы, что изложены в статье:
В известной и активно цитируемой работе креациониста Хитчинга “Шея жирафа” (“The Neck of the Giraffe”) это утверждение впервые появилось в 1982 году. Источник не был указан.
Более раннее упоминание об этом явлении было приведено в работе Wysong “The Creation-Evolution Controversy” в 1976 году (Хитчинг цитирует эту работу в “Шее жирафа”), и сказано в ней следующее:
“Two modern type horses, Equus nevadensis and Equus occidentalis, have been found in the same geological strata as Eohippus. Thus we have modern day type horses grazing side by side with their precursors”.
(Две лошади современного типа, Equus nevadensis и Equus occidentalis, были найдены в том же геологическом слое, что Eohippus. Таким образом, мы видим лошадей современного типа, пасущихся бок о бок с их предком.)
Но это – не оригинальная идея (точка зрения? глупость? – В.
П.) Wysong. Она заимствована из пестрящей ошибками и нелепостями книги Риммера
“The Theory of Evolution and the Facts of Science”, впервые изданной в 1935
году (в 1966-м вышло 14-е издание, которое и отыскал Барбер). В этой книге Риммер
упоминает, что лошадь ввезена в Америку европейцами, и критикует палеонтологов
за то, что они используют находки в Америке, чтобы продемонстрировать происхождение
европейского животного. Также он критикует палеонтологов за попытку “заткнуть”
разрыв в американской палеонтологической летописи с помощью европейского вида
(гиракотерий, напомню, описан по европейским образцам, а “эогиппусом” чаще называют
американских животных – В. П.). Ошибка Риммера состоит в том, что он пытается
представить, что все виды, из которых якобы “сконструирована” родословная лошади,
жили одновременно в наши дни.
В книге Риммера есть и такие строки:
“We feel
that the case against the horse demonstration would not be complete
without a mention of the
paleontological fact, that all the evolutionary
writers and text books seem so eager to suppress, and that is that there
are true fossil horses known to science today! Do we ever hear about
them? Indeed, we do not, and for the simple reason that they spoil
the "demonstration." How
can you show the evolution of a four-toed, rodent-like animal, the size
of a cat, into the horse, that weighs a ton, if there was a true horse
eating grass side by side with the Eohippus that was just starting in
to evolve into a horse thirty million years later? That simply can't
be done:
so they just suppress any mention of the true horse of fossil ages in
North America”. |
“Мы чувствуем, что дело против лошадиных доказательств не
будет полным без упоминания о палеонтологическом факте, что все эволюционистские
авторы и учебники, кажется, жаждут умолчать правду, то есть, есть истинные
ископаемые остатки лошадей, известные науке сегодня! Мы когда-либо слышали
о них? Несомненно, нет, и по той простой причине, что они портят “доказательства”.
Как вы можете показывать эволюцию четырёхпалого, подобного грызуну животного
размером с кота, в лошадь, которая весит тонну, если истинная лошадь ела
траву бок о бок с Eohippus, который только начал развиваться в лошадь тридцатью
миллионами лет позже? Это просто не может быть сделано: поэтому они только
замалчивают любое упоминание о настоящей лошади доисторических времён в
Северной Америке”. |
“There are at least two of them, the Equus nevadensis, and the Equus
occidentalis. Did the reader ever hear of them? Not if his reading has
been confined to evolutionary authors. We desire to stress the Equus occidentalis
especially, as we are personally familiar with that variety. This horse
(and it was a true horse) roamed the western slope of what is now known
as the United States, especially the Pacific Southwest. It was the contemporary
of the elephant, the camel and the so-called Saber-tooth Tiger, with all
of whose bones the remains of this fossil horse are found in profusion.
Long before man appeared on this continent the great creatures that were
the companions of the horse disappeared, and the horse likewise vanished
with them. But today in profusion we are recovering his fossil remains,
and his bones rise up to confront the dogma of science whose basis is prejudice,
and to refute the supposed demonstration of his evolution from a creature
with whom he was on grazing terms! It is apparent to the most unlearned
that the case collapses: If the creature that evolved out of a tiny ancestor
millions of years after that ancestor died out, really lived with that
ancestor side by side, the supposed demonstration becomes a joke”. [emphasis
in original] |
“Среди них есть по крайней
мере две, Equus nevadensis, и Equus occidentalis. Слышал ли читатель о
них вообще? Нет, если круг его чтения был ограничен только эволюционистскими
авторами. Мы хотим особо отметить Equus occidentalis, поскольку мы лично
знакомы с этой разновидностью. Эта лошадь (и это была настоящая лошадь)
бродила по западным склонам того, что теперь известно как Соединённые
Штаты, в частности, на Тихоокеанском Юго-западе. Она была современником
слона, верблюда и так называемого саблезубого тигра, вместе с костями
которых останки этой лошади найдены в изобилии. Намного раньше, чем человек
появился на этом континенте, огромные существа, которые были соседями
лошади, исчезли, и лошадь исчезла вместе с ними. Но сегодня мы в изобилии
открываем её окаменелые остатки, и её кости заставляют противостоять догме
науки, чьё основание – предубеждение, и опровергать предполагаемую демонстрацию
её эволюции от существа, с которым она паслась одновременно! Даже самым
неискушённым очевидно, что дело разваливается: если существо, которое
развилось из крошечного предка спустя миллионы лет после того, как этот
предок вымер, действительно жило с тем предком бок о бок, предполагаемая
демонстрация становится шуткой”. [соответствует оригиналу] |
Очевидно, идея Риммера развилась из предположения, что ископаемые остатки принадлежат
животным, которые все обитали в одно время.
В единственном источнике, который упоминает Риммер, "Guide Book Leaflet
No. 36, June, 1927", опубликованный Американским музеем естественной истории,
упомянутые виды лошадей не значатся.
Местонахождение, из которого были описаны эти образцы, также не уточняется.
В работе МакФаддена от 1992 года вид Equus nevadensis не упоминается, а про
Equus occidentalis сказано, что это имя заменено на Equus laurentius (со ссылкой
на первоисточник).
В иных источниках сказано, что вид Equus nevadensis был описан по пяти зубам
и части плюсны (господа креационисты, в этом месте пошутите, пожалуйста, над
собой!). К ним добавились другие фрагменты, часть из которых, возможно, была
ранее описана как Equus occidentalis, а также фрагменты, для которых не были
указаны место находки и породы, в которых они были обнаружены. В более поздних
публикациях было рекомендовано считать название Equus nevadensis невалидным.
Риммер был несведущ в геологии, и его предположение о том, что лошади современного
типа и гиракотерий должны быть найдены в отложениях одного возраста, является
ошибочным. Тем не менее, оно было взято креационистами на вооружение, поскольку
соответствовало их теории.
Такова, вкратце, суть статьи Jon Barber. Как говорится, вопросы есть?
12. Современные лошади составляют большое разнообразие по размерам.
Правило Копа выстраивает лошадей в ряд от меньшей до большей. Все же аргентинская лошадь породы фалабелла полностью вырастает до 43 сантиметров (17 дюймов) в высоту. Это такой же размер, как у Hyracotherium. Размер лошадей значительно изменяется: от крошечной фалабеллы до массивного клейдесдаля [шотландская порода лошадей-тяжеловозов – В. П.]. Обе они лошади. Это доказывает, что выстраивать предков лошади согласно предположению об увеличивающемся размере неправильно.
Упоминавшаяся выше книга иеговистов “Жизнь – как она возникла…” тоже затрагивает этот вопрос (стр. 67):
“... не удивительно, что ископаемые остатки лошадей отличаются по размеру и форме. Даже сегодня лошади варьируют от карликовых пони до рослых рабочих лошадей. Все они - разновидности внутри рода лошадей”.
Большое спасибо господам креационистам за то, что они слегка
придушили собственную же аргументацию. Действительно, в отличие от фильма про
Годзиллу, размер для лошадей не имеет такого особого значения, как ему приписывают
те же самые, кстати, креационисты. В книгах эволюционистского плана увеличение
размеров внутри семейства также отмечается, но этот признак ставится далеко
не на первое место.
Среди самых древних животных надсемейства Equoidea были представители бесследно
вымершей ветви палеотериев (Palaeotherium, семейство Palaeotheriidae, близкое
к лошадиным) - животные, массивным телосложением и размерами напоминавшие тапира
или небольшого носорога. Они обитали в лесах древней Европы наряду с эогиппусом
и были весьма похожи на него по строению (с поправкой на массивное сложение).
А в миоцене наряду с довольно крупными Merychippus и Anchitherium обитал маленький
Archaeohippus. От Merychippus также эволюционировали более мелкие Protohippus
и Calippus. Поэтому разговоры креационистов о том, что эволюция лошади в представлении
эволюционистов сводится лишь к увеличению размеров потомков по сравнению с предками,
по меньшей мере безграмотны.
Правило, выведенное эмпирически палеонтологом Эдвардом Дринкером Копом, гласит,
что в процессе эволюции представители развивающейся линии живых организмов увеличиваются
в размерах, и затем вымирают. Это видно во многих случаях, причём не только
среди позвоночных, но и среди самых разных беспозвоночных. Но всегда ли лошади
следовали этому правилу?
“В эволюции Equidae не было таких направлений, которые продолжались бы на всём протяжении истории какой-либо филетической линии или имели бы место во всех линиях одновременно. Увеличение размеров тела было подвержено реверсиям, происходившим в некоторых линиях в разные периоды”. (В. Грант “Эволюция организмов”, М., Мир, 1980, стр. 264)
“Тенденция к увеличению размеров никогда не была всеобщей. В некоторых филетических линиях наблюдается такое увеличение, тогда как в других родственных им линиях на протяжении того же периода отмечается, напротив, уменьшение размеров. Известны и такие случаи, когда у одной и той же филетической линии в один период времени эволюция была направлена в сторону увеличения размеров, а в другой – в сторону их уменьшения. Оба эти отклонения от правила Копа обнаружены в семействе лошадей, а также у слонов, оленей и в других группах”. (В. Грант “Эволюция организмов”, М., Мир, 1980, стр. 262)
Итак, эволюционисты прямым текстом признают,
что размер для эволюции лошадей – не главное, и что на протяжении своей истории
лошади могли
даже уменьшаться
в росте. Это, вроде бы, было одним из возражений креационистов? Или
в свете цитат из Гранта его уже можно не считать возражением?
Самое главное в эволюции лошадиных - это изменение числа пальцев
на ноге (с 4 до 1), изменение формы зубов - от бугорчатых до складчатых,
изменение
мозга
– от маленького до относительно (не только абсолютно!) более крупного
и сложного. Размеры тела, конечно, также увеличивались, причём это
шло параллельно
приспособлению
к обитанию в открытой местности. Разные рода древних лошадей независимо
друг от друга переходили к жизни в открытой местности, где независимо
друг от
друга развивали сходные признаки (в данном случае это явление называется
“параллелизм”,
поскольку, в отличие от конвергенции, происходит в близкородственных
группах). Это, кстати, и затрудняет установление родства между разными
родами лошадиных,
поскольку часто бывает неясно, является ли какой-то признак результатом
родства или параллелизма в развитии. Поэтому правильнее проводить
сравнение
не по
одному произвольно выбранному признаку (например, по размеру тела,
как это делают
креационисты), а по целому комплексу признаков.
В эволюционном ряду лошадиных менялись пропорции черепа: увеличивалась
лицевая часть, становились прочнее челюсти, расширялся объём черепной
коробки. Разительно
менялись зубы. У древнейших родов семейства лошадиных зубы были мелкие
и слабые, с небольшой складчатостью. А у более поздних лошадей увеличивается
размер зубов
(не только просто линейный, но и по отношению к размеру черепа),
усиливается их складчатость, развивается слой твёрдого цемента, расширяются
жевательные
поверхности зубов. Меняются и корни зубов - у современных лошадей
зубы растут
всю жизнь, одновременно стираясь на вершинах. Эти признаки не менялись
все сразу - в разных родах и в разные геологические эпохи шло изменение
зубов
в разных направлениях. Изменения зависели и от среды обитания - у
лесных лошадей,
обитавших одновременно со степными, зубы оставались бугорчатыми и
не имели цемента.
Все изменения у представителей древних лошадиных связаны с приспособлением
к обитанию на открытой местности. Это стало выгодно для выживания
в связи с исчезновением лесов в раннем кайнозое. А некоторые изменения
в механике
ног
и строении зубов явились прямым следствием увеличения размеров тела животных.
13. Изменение количества зубов.
-Тот же самый беспорядок виден в числе зубов воображаемых предков лошади и числе зубов лошади наших дней.
-Единственные типы зубов, известные для лошадей, или листоядного, или травоядного типа. Никаких других типов зубов не было обнаружено. Так, даже переходные типы зубов не существуют. (Moore, John, N., and Harold S. Slusher, Eds., Biology: A Search for Order in Complexity, Zondervan Publishing House, Grand Rapids, Michigan, 1970, p. 548)
Что ж, дарёному коню, как известно, в зубы не смотрят. Но
в данном случае это нужно сделать.
Я уже говорил выше по поводу арифметики на службе креационизма, но в данном
случае хочу рассмотреть именно этот аргумент особо.
Особенности строения самих зубов, а также зубная формула являются в достаточной
степени значимыми чертами, характеризующими какую-либо группу млекопитающих.
Но из этих двух характеристик строение зубов всё же важнее, нежели их количество.
Если мы припомним горячо любимых Дуэйном Гишем южноамериканских копытных, то
можно привести один чудесный пример, описанный в книге Р. Кэрролла “Палеонтология
и эволюция позвоночных” (глава “Южноамериканские копытные”):
“Вид Henricosbornia lophodonta известен по зубным системам сотен экземпляров. Статистически обработав данные по количественным и качественным признакам этих зубов, Симпсон (Simpson, 1948) показал, что все они принадлежат единой биологической группе с непрерывной изменчивостью. Амегино (Ameghino, 1906), исходя из топологической концепции, выделял по этому материалу 16 видов из восьми родов, четырёх семейств и трёх отрядов”.
“Как и Henricosbornia, род Notostylops отличается широчайшей изменчивостью зубной формулы (Simpson, 1980), которая в рамках одного вида может колебаться от 3/3-1/1-4/4-3/3 до 2/2-0/0-/3/3-3/3”.
В чём состоит главная ошибка креационистов? В том, что животное
– это целостный организм, а они предпочитают рассматривать его в расчленённом
виде. Отдельно зубы, отдельно позвонки, отдельно рёбра не живут. Живёт организм
в целом. И определение вида точнее всего можно сделать, располагая полным набором
признаков, характеризующих его. Спору нет, зубная система животного очень важна
для определения видовой принадлежности животного (в частности, млекопитающего).
Но дело здесь не ограничивается простой арифметикой. Зоолог тщательно обмеряет
сами зубы, обращая внимание на их форму, складчатость, расположение бугров и
множество других признаков. Можно даже с большой долей уверенности установить
видовую принадлежность отдельного зуба или обломка челюсти. Готов побиться об
заклад, автор “зубного” тезиса не смог бы отличить зуб летучей мыши от зуба
землеройки (даже современных видов!), и, возможно, даже зуб быка от зуба носорога.
Если бы он мог это сделать, то такой “убийственный тезис” просто не появился
бы в этом списке.
Изменение лошадиных зубов – очень интересная тема. По мнению эволюционистов,
по этому признаку можно прямо судить об изменении рациона, и косвенно – об изменении
образа жизни. Впрочем, у креационистов на этот счёт своё мнение:
“Конечно, форма зуба может широко варьировать в пределах вида, означая этим, что неблагоразумно предполагать, что различные окаменелости зубов показывают эволюцию. Также неблагоразумно мыслить догматически о рационе, основываясь на форме зуба. Мы показывали это на примере с летучими мышами [эту ссылку можно найти здесь: http://www.answersingenesis.org/creation/v21/i3/horse.asp только пользы от этого никакой: там ничего не говорится про зубы летучих мышей – В. П.], и недавние свидетельства перевернули предыдущую мысль о рационе древних лошадей, основанную на форме зубов. Эволюционист-палеонтолог Брюс МакФадден анализировал зубы от шести “видов” лошади (более вероятно, разновидностей в пределах вида), “датированных” временем пять миллионов лет назад.
Предыдущие эволюционные теории утверждали бы, что они, должно быть, питались травой, потому что они все имели высоко-коронковые зубы. Но количество устойчивых изотопов углерода 12C и 13C, находившихся в зубах, указали, что лошади питались листьями, а не травой”. (Джонатан Сарфати “Non-evolution of horse”)
Креационисты хотят видеть в качестве “переходной формы” некий орган, совмещающий в определённой пропорции черты исходного и конечного состояний. Вот, например, что пишет сам Джонатан Сарфати в статье “Пятнадцать способов опровергнуть материалистский вздор: подробный ответ журналу “Scientific American””:
“Креационисты же всего лишь хотят, чтобы им показали непрерывный ряд существ с последовательно изменяющимися признаками: например, 100% ноги – 0% крыла; 90% ноги – 10% крыла… 50% ноги – 50% крыла… 10% ноги – 90% крыла; 0% ноги – 100% крыла”.
А если эволюция в принципе шла не
так, как ХОТЯТ УВИДЕТЬ креационисты? Корректно ли считать такое несоответствие
опровержением теории эволюции и доказательством правоты сторонников теории креационизма?
Эволюция зубов от Hyracotherium к Equus проходила не путём “разбавления” примитивных
признаков прогрессивными, как представлял бы, например, тот же Дж. Сарфати.
Верне Грант в книге “Эволюция организмов” приводит 6 различий в зубной системе
между гиракотерием и современной лошадью, и описывает характер эволюции зубной
системы лошадей:
“Зубы Equus обладают следующими прогрессивными признаками: 1) дифференцировка режущих передних зубов для откусывания и коренных зубов для пережёвывания; 2) большие размеры коренных зубов; 3) высокая коронка у коренных зубов; 4) увеличение числа функциональных (жевательных) коренных зубов – по шести на каждой стороне каждой челюсти; 5) сильное развитие на жевательной поверхности складок (лофодонтность), состоящих из эмали; 6) развитие между складками твёрдого цемента.
У Hyracotherium этим признакам соответствуют следующие примитивные особенности зубов: 1) более слабая дифференцировка передних и боковых зубов; 2) меньшие размеры зубов; 3) низкая коронка у коренных зубов; 4) отсутствие жевательных зубов (на каждой стороне каждой челюсти имелось по три коренных зуба, но это не были настоящие жевательные зубы); 5) складки эмали на жевательных поверхностях более простые и более низкие; 6) отсутствие цемента”. (В. Грант “Эволюция организмов”, М., Мир, 1980, стр. 255)
Но в эволюции лошадей от гиракотерия к современной лошади отсутствовали формы, у которых были зубы, демонстрирующие одновременно “усреднённое” состояние по всем этим признакам. Грант описывает эволюцию зубов лошадей таким образом:
“Эти изменения разных признаков зубов происходили не одновременно, а в разное время. Моляризация (направление 4) имела место главным образом в эоцене и олигоцене, а затем она более или менее прекратилась. Увеличение высоты коронки (направление 3) произошло в основном в миоцене, причём оно коснулось только таких вновь возникших травоядных групп, как Merychippus, тогда как у миоценовых лошадей, питавшихся листвой деревьев, коронки зубов оставались низкими”. (В. Грант “Эволюция организмов”, М., Мир, 1980, стр. 255)
Также Верне Грант описывает постепенное образование складок на зубах в линии от Mesohippus до Equus, и появление зубного цемента в ряду Parahippus – Merychippus – Nannippus, подробно изученное в работах Simpson:
“Известно несколько интересных примеров, когда тот или иной признак зубов появлялся в какой-либо древней полиморфной ископаемой популяции в качестве одного из вариантов, а затем закреплялся в качестве постоянного признака в более поздней популяции.
Один такой случай относится к определённой складке (известной под названием “кроше”) на верхних коренных зубах в линии Mesohippus — Equus. У Mesohippus эта складка обычно отсутствовала. Но в некоторых популяциях Mesohippus было по нескольку мутантных особей с небольшими кроше. У некоторых видов-дериватов Miohippus из олигоцена очень маленькое кроше представляло собой относительно постоянный признак; у других, более поздних видов-дериватов Parahippus из раннего миоцена небольшое кроше было постоянным признаком. Позднее в линии, ведущей к Equus, кроше достигло значительных размеров (Simpson, 1944; 1953), Зубной цемент впервые появляется аналогичным образом в линии Parahippus — Merychippus. У Parahippus цемент обычно отсутствует, но наблюдается в некоторых его популяциях в качестве полиморфного варианта, а позднее закрепляется в мономорфном состоянии у Merychippus”. (В. Грант “Эволюция организмов”, М., Мир, 1980, стр. 258 - 259)
В примере видно, как этот признак, представлявший собой случайное
отклонение в ранних популяциях, становится более заметным
и типичным в поздних
популяциях. Этот процесс происходил, судя по времени появления
указанных лошадиных,
в миоцене, параллельно увеличению высоты коронки зубов.
Поэтому утверждать, что у лошадиных не было переходных
типов зубов – неправильно. Эти переходы были, они доказаны
для
большого количества
ископаемого материала.
Другое дело – они могут не совпадать с представлениями
креационистов. Но это уже, извините, “дело десятое”.
В свете изложенных Грантом результатов исследований эволюции
лошадей аргументы Дж. Сарфати кажутся явно неуместными
– на эти вопросы
давно уже дан ответ.
Как видно из приведённого отрывка работы Гранта, высокая
коронка зубов появилась после того, как зубы приобрели
характерную форму (моляризовались),
но перед
образованием складок и параллельно появлению зубного цемента,
которые являются приспособлением для перетирания жёсткой
травы.
То есть,
форма зубов древних
лошадей совершенно не противоречит приведённым результатам
исследований МакФаддена, зато ясно показывает, что наш
“доброжелатель” Джонатан
Сарфати явно что-то
не договаривает, либо не столь сведущ в науке, как хочет показать.
14. Moropus, не включённый в родословную лошадиных.
“Moropus”, который жил в эпоху миоцена, не включен в ряд окаменелостей, хотя во многом напоминает лошадь, только потому, что это не служит цели эволюционистов. Как сказано в Энциклопедии Доисторических Животных, Moropus двухметровой высоты больше размером, чем Meryhippus того же возраста, и современные лошади.
В таком случае я мог бы присоединиться к креационистам, гневно
обличающим “нехороших и лживых” эволюционистов, и добавить от себя, что приведённый
пример с Moropus может быть ещё не самым вопиющим примером “эволюционистской
лжи”, основанной на умалчивании находок. Дело в том, что в гораздо более ранних
олигоценовых отложениях найден ещё один зверь – огромный Indricotherium, телосложением
также очень напоминавший лошадь, особенно если смотреть спьяну и издали. И мало
того, по размеру он гораздо крупнее, нежели упомянутый Moropus, живший после
него. И это обстоятельство, по идее, должно ещё нагляднее показывать фальшь
и лживость “нехороших эволюционистов”.
“Должно”, но не показывает. И не будет показывать. Дело в том, что упомянутый
индрикотерий не относится к семейству лошадиных: это носорог. А при чём тут
странный вымерший зверь моропус? В том-то и дело, что тоже абсолютно не при
чём: он принадлежит к совершенно другому семейству – халикотериев (Chalicotheriidae).
А это семейство принадлежит к подотряду Ancylopoda в отряде непарнокопытных.
Лошади же принадлежат, как я уже упоминал выше, к подотряду Hippomorpha. Халикотерии
мало похожи на лошадей, и вообще на копытных: телосложением они напоминают помесь
гориллы, медведя и ископаемого ленивца мегатерия, а на их лапах вместо копыт
росли длинные когти, иногда даже втяжные, как у кошек. Носороги, к слову, вместе
с тапирами являются представителями другого подотряда – Ceratomorpha, и являются
столь же отдалёнными родичами лошадей, как и халикотерии. Любой халикотерий,
включая моропуса, является гораздо более далёким родственником любой из лошадей,
нежели любые два рода лошадей внутри семейства лошадиных. Знакомый приём введения
“третьего лишнего”… Помнится, Дуэйн Гиш уже попробовал это сделать (см.
пример 1), только это вышло очень “криво”.
 |
 |
|
|

На картине художника Michael Rothman изображены
североамериканские
звери эпохи миоцена: свиньи, верблюды, лошади и моропус,
поднявшийся на задние лапы и объедающий листья.
Странный, однако же, приём опровержения применяют креационисты. Одно из двух: либо это личное невежество автора данного “аргумента”, либо злонамеренная ложь.
15. Hyracotherium часто находили на поверхности.
Ископаемые остатки лошадей не найдены одна под другой в породах. И напротив, кости Hyracotherium (Eohippus) часто бывали найдены в поверхности, и единственная причина для датировки этих слоёв эоценом – то, что окаменелости Hyracotherium были найдены в них!
А не приходит ли в голову уважаемым (и не очень уважаемым) креационистам, что простая глубина залегания слоёв в земной коре не есть точный и абсолютный критерий для определения их возраста? Если кости гиракотерия (или, если угодно, эогиппуса) найдены на поверхности земли, то это ещё не означает, что данный вид жил в позднем плейстоцене или голоцене. Просто из курса природоведения средней школы нам должно быть известно такое явление, как выветривание. Слои осадочных пород, лежащие поверх эоценовых (в эту эпоху жил гиракотерий) могут быть просто снесены, смыты, уничтожены силами природы, обнажив слой эоценовых отложений. И кроме того, какая-то область, где на поверхности земли лежат кости гиракотерия, не всегда была районом осадконакопления. Поэтому на ней могло просто не накопиться отложений последующих эпох. Так что ни колдовства, ни феноменальной продолжительности существования рода гиракотерий (сиречь эогиппус) здесь не наблюдается.
“Окаменелости не несут знаков, говорящих о том, каков их возраст. Их возраст, как правило, назначается им в зависимости от их относительной глубины захоронения. Тем, что лежат в самых глубоких слоях пород, назначили самый большой возраст. Основываясь на библейской позиции, мы должны ожидать, что многие, но не все окаменелости, должны быть захоронены в течение Всемирного Потопа, так что самые старые были бы в действительности только приблизительно 4500-летнего возраста. Окаменелости выше, возможно, могли быть захороненными местными катастрофами после Всемирного Потопа.
Вероятно, что многие из окаменелостей лошадей были послепотопными. Однако, даже если мы допустим эволюционную/длинную датировку возраста, они не показывают ясной прогрессии, представленной учебниками”. (Джонатан Сарфати “Non-evolution of horse”)
Уповать на Всемирный Потоп, как это делает Дж. Сарфати, думаю,
не следует. Просто нужно ещё доказать, что этот Потоп был. Будь он реальностью,
сама планета и расселение по ней живых существ выглядели бы совсем иначе, нежели
сейчас. Но это уже тема для совершенно отдельного разговора.
Но боюсь, что Дж. Сарфати несколько путает разные способы определения возраста
пород. Так, не всегда самые древние породы лежат ниже всех: иногда в результате
сдвигов земной коры один слой пород наползает на другой, и молодые породы оказываются
погребёнными под слоем более древних.
Что касается относительного возраста пород (то есть, принадлежности отложений
к какому-либо геологическому периоду), то его определяют по характерным для
него ископаемым – так называемым “руководящим ископаемым”. Такой способ точен,
но не даёт значений абсолютного возраста пород. А определение возраста пород
по радиоактивным изотопам даёт искомые цифры. Этот способ, вопреки утверждению
креационистов, не требует знания о первоначальном количестве определяемого изотопа
в породе: выбирается метод определения тех изотопов, которые полностью отсутствовали
в породе на момент её образования. Именно поэтому вулканические породы, побывавшие
в расплавленном состоянии, служат буквально “секундной стрелкой” для определения
абсолютного возраста отложений.
Если также учесть, что срубленное в начале ХХ века дерево сосны остистой достигало
возраста 5 тысяч лет, а некоторые колонии лишайников в Арктике вообще были современниками
мамонтов (их возраст оценивается до 12 тысяч лет), то библейская хронология,
поддерживаемая Дж. Сарфати, вообще не выдерживает критики.
16. Будет ли 16-й аргумент?
Пожалуй, вот и всё. Все 15 аргументов креационистов разобраны
на составные элементы, и их суть теперь ясно видна.
Одни из них оказались следствием невежества и незнания элементарных
вещей, которые должно знать человеку, вздумавшему опровергнуть теорию.
Другие оказались результатом того, что их авторы были явно не в курсе
достижений палеонтологии за последние полвека.
Самое обидное – это то, что третья группа аргументов является результатом
целенаправленной и злонамеренной лжи. И опровергнуть их оказалось легче
всего, хотя они казались столь же прочными, как все остальные.
В заключение я хотел бы ещё немного сказать о… лошадях.
Правда, не о настоящих, а о литературных, из “Путешествий
Гулливера”
Джонатана Свифта.
Думаю, некоторые
их жизненные принципы полезно знать креационистам:
“Я помню, что когда в беседах с хозяином о людях, их нравах и обычаях мне случалось упоминать о лжи и обмане, то он, несмотря на весь свой ум, лишь с большим трудом понимал, что я хочу сказать. Он рассуждал так: способность речи дана нам для того, чтобы понимать друг друга и сообщать друг другу полезные сведения о различных предметах. Поэтому если кто-нибудь станет утверждать то, чего нет, то назначение нашей речи совершенно извращается. Тот, к кому обращена речь, перестаёт понимать своего собеседника. Он не только не приобретает никаких новых сведений, но оказывается в гораздо худшем положении, ибо его стараются убедить, что белое – черно, а длинное – коротко. Этим и ограничивались все его понятия относительно способности лгать, которая пользуется таким распространением среди людей”.
“… долгие споры, ожесточённые пререкания, упорное отстаивание ложных или сомнительных мнений – пороки, неизвестные гуигнгнмам”.
В свете этих слов Свифта лошади (пусть даже вымышленные
гуигнгнмы) кажутся гораздо умнее иных людей. Поэтому
я считаю спор с
креационистами о “Лошадиной
фамилии” законченным.
Впрочем, автор этих строк ещё не сказал твёрдое “баста!”.
Если у прочитавших эту работу есть иные возражения
против эволюции
лошадей, пишите и заходите
на сайт:
sivatherium@yandex.ru – почта
sivatherium.narod.ru – сайт.
Спасибо за то, что дочитали до конца.
Волков Павел Иванович.
Владимир, 2004.