
"Лошадиная фамилия"
Часть I
| Главная |
|
"Лошадиная фамилия" Часть I |
|||
Поручик Ржевский приходит к врачу: Анекдот |
Если построить рейтинг современных и ископаемых животных, которых креационисты
особенно часто упоминают в своих произведениях, то за почётные первое, второе
и третье места по результатам подсчёта будут драться дарвиновы вьюрки, австралопитеки
и лошади, хотя мало кто из них сравнится по популярности с археоптериксом.
И это при том, что явления эволюции, в общем-то, хорошо прослеживаются на иных,
«неклассических» примерах: их проявление можно увидеть как в царстве рыб, так
и среди, например, головоногих моллюсков и насекомых.
Трудно сказать, почему у сторонников теории божественного творения особой любовью
пользуется именно лошадь со всей своей ископаемой роднёй. В любой их книге
можно найти рассказ о древних лошадях, снабжённый безграмотными комментариями
«юных натуралистов». Конечно, ряд ископаемых форм древних лошадиных часто упоминается
в работах по теории эволюции как наглядный пример эволюционных изменений. Причём
изменения эти описаны настолько хорошо и подробно, что просто уму непостижимо,
как креационисты умудряются писать про древних лошадей такие глупости, которые
иногда можно прочесть в их книгах. Судя по всему, их знакомство с сутью вопроса
было крайне поверхностным (как, впрочем, и везде). Ведь на многие, «убойные»
с их точки зрения, аргументы ответ легко находится в самих первоисточниках,
на которые креационисты порой ссылаются.
Редкостное однообразие аргументации заставляет меня думать, что на рассмотрение
и критику родословных древ иных животных (например, носорогов, слонов или китов)
у сторонников теории творения просто фантазии не хватает, и они просто списывают
«гневные отповеди нехорошим эволюционистам» друг у друга. В любом случае родословная
лошади встречается в книгах креационистов гораздо чаще прочих.
Пожалуй, наиболее полно и в наиболее чистом виде все аргументы креационистов,
старающихся опровергнуть данные об эволюции лошади, представлены в работе Biology
Textbook Fraud. The Horse Series. “The Second Piltdown man of Paleontology”
(«Мошенничество в учебнике биологии. Ряд лошадей. «Второй Пилтдаунский человек»
палеонтологии»). Если кто-то захочет познакомиться с текстом этой работы на
английском языке – милости просим вот сюда: http://www.bible.ca/tracks/textbook-fraud-dawn-horse-eohippus.htm Если
же по указанному адресу вы ничего не найдёте (например, если сайт просто ликвидируют
за ненадобностью) – просто почитайте то, что написано здесь дальше.
Думаю, после этого многим не захочется искать оригинальную работу.
Я решил опираться в своих выкладках именно на эту работу, поскольку, как я
уже сказал, это самый полный из всех известных мне первоисточников, где изложена
креационная точка зрения по данному вопросу. Время от времени я буду вставлять
в текст цитаты из других креационных работ, в чём-то перекликающихся с этим
материалом.
Итак, авторы, скромно решившие себя не называть, наскребли целых пятнадцать
доказательств того, что эволюционный ряд лошадей, представленный в учебниках
и книгах по теории эволюции, является ложным. Я постарался сделать перевод
этих тезисов, поскольку не все читатели владеют английским языком. После этого
я заглянул в первоисточники, которыми побрезговали креационисты, и нашёл в
них массу интересных вещей, которые им потребно знать по роду деятельности.
Итак, что же это за аргументы? Это, в принципе, «контрольный срез» мнений креационистов
относительно эволюции лошадей и её значения в теории эволюции вообще. На сайте
не приводится авторство того или иного тезиса (а жаль, героев надо знать в
лицо!), и все эти утверждения можно обнаружить в разном количестве в работах
иностранных и отечественных сторонников теории творения. Начать разбирать эти
аргументы лучше всего, разумеется, с самого начала.
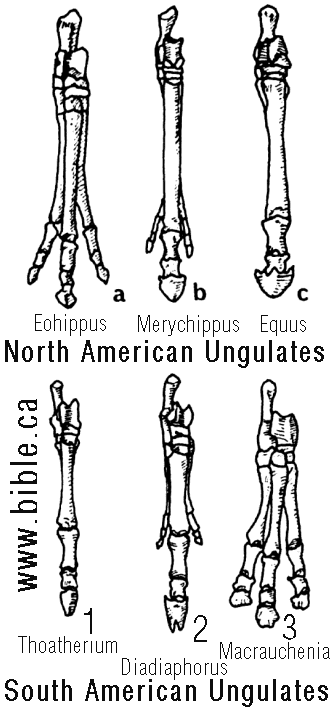
1. От трёх пальцев ноги к одному, или от одного к трём?
 |
|
Это - прямая противоположность предполагаемой последовательности событий, которые произошли с североамериканскими лошадями. Я не знаю никакого эволюциониста, кто предлагает такую эволюционную последовательность событий, но почему бы и нет? Возможно, потому что последовательность от трёхпалых до однопалых для североамериканских лошадей стала настолько популярной в эволюционных кругах, что никто не смеет предлагать обратный переход. Конечно, в Южной Америке реальных доказательств наличия переходных форм не больше чем в Северной Америке”. (The Origin Of Mammals, ICR Impact No. 87 Duane T. Gish, Ph.D, 1980; Cf. Romer, A. S., Vertebrate Paleontology, 3rd Ed., Chicago: Univ. of Chicago Press, 1966, p. 260-261)
В своей книге “Учёные-креационисты отвечают своим критикам” (текст этого произведения нетрудно найти в Интернете на “Сайте апологии христианства”) Дуэйн Гиш также обращается к этому же доказательству:
“Он [Футуяма - В. П.] заявляет, что в Южной Америке "псевдолошади", Diadiaphorus и Thoatherium, подвергаются параллельным эволюционным изменениям”; ... Все это целиком и полностью — ошибка, даже если принять теорию эволюции. Я описал это в разделе о лошадях в книге “Эволюция: вызов, бросаемый окаменелостями”, останки Diadiaphorus и Thoatherium одновременно появляются в миоценских скалах, что отнюдь не свидетельствует о переходе от тройного копыта к нераздвоенному. У Diadiaphorus боковые “пальцы” уменьшены, а третья разновидность, Macrauchenia, обладает полноценным “трехчастным” копытом. Macrauchenia, однако, была последней формой в соответствии со шкалой времени, которую эволюционисты применяют к плиоценовым скалам. Плиоцен — предполагаемая геологическая эпоха, следующая за миоценом. Фактически, считается, что Thoatherium с нераздвоенными копытами вымер до того, как “трехпалый” Macrauchenia появился на свет. Другими словами, последовательность развития в Южной Америке идет в ином направлении по сравнению с Северной Америкой: от лошади с нераздвоенным копытом — к трехпалой, а в Северной Америке, считается, было наоборот. ... эволюционисту было бы трудно объяснить, почему в Северной Америке эволюция превратила лошадь с тройными копытами в лошадь с нераздвоенными копытами, а в Южной Америке произошел обратный процесс”.
Готов поклясться
филейными частями самого дьявола, девяносто девять человек из ста поверят в эти
слова или просто не поймут, где здесь “собака зарыта”.
Всё правильно, именно на это и рассчитан ход Дуэйна Гиша. Но я попробую изменить
соотношение сил в обратную сторону, и даже довести его до полной победы над
теорией Гиша.
Что-то кажется мне, что г-н Гиш, защищая креационизм (и в первую очередь –
себя), совсем забыл о соблюдении точности в аргументации. Если обратиться к
такому авторитетному источнику, как “Палеонтология и эволюция позвоночных”
Р. Кэрролла, можно обнаружить удивительные вещи, которые полезно было бы знать
Дуэйну Гишу, принимающему в отношении эволюционистов характерную для креационистов
“позу скунса” со всеми основополагающими чертами сходства.
Главные “действующие лица” (точнее было бы сказать – “морды”) в этом примере
– вовсе не лошади (представители отряда непарнокопытных Perissodactyla), а
представители ныне вымершего отряда млекопитающих – литоптерн (Litopterna).
Вся история этого отряда проходила только на территории Южной Америки, которая
на протяжении всего кайнозоя была островным континентом. На территории этого
материка появились местные формы копытных (несколько отрядов!), внешне подобные
хорошо известным зверям Старого Света. Так, пиротерий был аналогом слона, токсодон
– бегемота, а быстроногие диадиафорус и тоатерий внешне были поразительно похожи
на лошадей. Но постепенно они вымирали: когда возник Панамский перешеек, североамериканские
формы быстро вытеснили южан, и их расселение привело к вымиранию местной фауны.
 |
 |
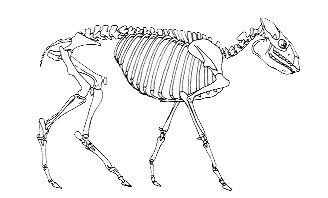
Скелет Diadiaphorus из книги Р. Кэрролла "Палеонтология и эволюция позвоночных". |
Реконструкция Toatherium из книги "Энциклопедический словарь юного биолога" |
В книге Кэрролла приведён подробный список известных к моменту выхода в свет данного издания книги родов позвоночных животных с указанием времени их жизни. Так, судя по этим данным, Diadiaphorus с не исчезнувшими до конца боковыми пальцами действительно появляется в палеонтологической летописи одновременно с однопалым Toatherium в нижнем миоцене, и даже переживает его, дожив до нижнего плиоцена, но это совсем не означает, что он был предком последнего. Они могли просто представлять потомков одного из более ранних родов с трёхпалой ногой (кто сможет поручиться за то, что все ископаемые виды уже найдены?). При этом редукция пальцев у Toatherium просто прошла быстрее, чем у Diadiaphorus в связи с различиями в среде их обитания. Весьма странными вещами выглядят применённое Д. Гишем для “доказательства” своей правоты введение в этот ряд “третьего лишнего”: Macrauchenia, животного ещё одного рода южноамериканских копытных Litopterna, и построение своей аргументации именно на этом факте. Дело в том, что Macrauchenia не является представителем семейства Proterotheriidae, к которому относятся оба упомянутых рода “ложных лошадей”. Она - представитель особого семейства Macraucheniidae, которое появилось в палеонтологической летописи задолго до того, как жил любой из упомянутых родов “ложных лошадей”. Семейство Proterotheriidae столь же древнее. Иными словами, непосредственные предки макраухении не имели ничего общего с “ложными лошадями” задолго до того, как те появились.

А вот "родственник" предыдущих видов
- макраухения.
Такой она предстала в сериале ВВС "Прогулки с чудовищами".
Ни Diadiaphorus, ни Toatherium не является предком Macrauchenia - они связаны в лучшем случае крайне отдалённым родством. Если говорить проще, Гиш пытается убедить нас, что, допустим, осёл – не из семейства лошадиных, ссылаясь на то, что у носорога есть рог. Поэтому убедительная просьба к нему - не бросаться умными словами столь опрометчиво.
2. Отсутствующий рудиментарный четвёртый палец ноги Hyracotherium.
-“Как Вы можете видеть от этой фотографии передней ноги Hyracotherium http://www.bible.ca/tracks/textbook-fraud-dawn-horse-eohippus.htm, он имел четыре пальца на ноге. Эволюционисты поднимают большой шум, указывая на рудиментарные остатки двух внешних пальцев ноги у современных лошадей http://www.bible.ca/tracks/textbook-fraud-dawn-horse-eohippus.htm ЗАДНЕЙ НОГИ. Все же, ни один из “эволюционных предков” Hyracotherium не имеет даже минимального “рудимента” на ПЕРЕДНЕЙ НОГЕ. Это абсолютно катастрофично для эволюционистов, пока они не призывают свою волшебную палочку прерывистого равновесия, чтобы уладить беспорядок.
-Итак, четвертый палец передней ноги Hyracotherium исчезает на 100 % у первого же трёхпалого “предка” (Mesohippus), но два наружных пальца ноги на задней ноге Hyracotherium, как предполагается, присутствуют как рудименты у “современной лошади”.
Господа креационисты! В свете этого утверждения хочу вас спросить:
а вы уверены, что при жизни гиракотерий (он же эогиппус) ДЕЙСТВИТЕЛЬНО не имел
даже следов рудимента этого пальца? Неужели вы имели редчайшую удачу анатомировать
свежий натуральный экземпляр этого зверя, добытого на охоте, чтобы столь безапелляционно
утверждать это? Если нет – можете ли вы быть уверенными в том, что на ископаемых
образцах сохранились все кости до единой? Рудимент мог быть представлен очень
тонкой косточкой, которая разрушается при фоссилизации, либо вообще хрящом,
который не сохраняется в ископаемом виде.
Однако допустим, что рудимент пятого пальца у гиракотерия и четвёртого пальца
у мезогиппуса действительно полностью исчезли. Это вполне возможно – у современных
китов, например, нет ушных раковин и задних ног (кроме тазовой кости). А что
удивительного в том, что у современных лошадей сохранились рудименты боковых
пальцев? Сами же креационисты очень любят приводить пример функциональности
рудиментов: аппендикса человека, тазовых костей китов, эпифиза наземных четвероногих.
Если рудиментарный орган имеет некоторое функциональное значение, он не исчезает
полностью, хотя сильно уменьшается и упрощается, и его функции меняются по сравнению
с исходным состоянием. Прежде, чем удивляться наличию у современной лошади рудиментов
боковых пальцев ноги, требуется сущий пустяк: доказать, что они НЕ НЕСУТ НИКАКОЙ
ФУНКЦИИ. Если это будет блестяще подтверждено, допустим, исследованиями Института
Муди (верите, нет, но такой институт существует! Не у нас, но где-то “за кордоном”),
то тогда можно будет смело удивляться наличию таких структур. Если же эти рудименты
служат местом прикрепления сухожилий или мелких мышц – тогда просьба забыть
о своих возражениях и принять всё, как есть. Впрочем, сами креационисты снова
и снова подбрасывают факты, разрушающие их собственные надежды. Так, в этой
же самой работе, призванной опровергать эволюционизм и подтверждать правильность
креационных взглядов, есть дивные строки:
“Эти структуры - не рудиментарные, а исполняющие важную задачу: они помогают лошади бежать в равновесии. Эти дополнительные боковые элементы не только усиливают ногу, но и помогают уравновесить её. Считайте их слоистым материалом, который усиливает ногу таким же образом, как слои фанеры делают её более прочной, чем цельная древесина. Эти три секции соединены вместе таким образом, чтобы сопротивляться перелому и увеличению сил кручения в ноге лошади. Без этого лошадь ломала бы свои ноги гораздо чаще”.
И после этого господа креационисты хотят, чтобы их теории
долго жили? Большое спасибо авторам этой интереснейшей странички за то, что
они сами “выплеснули ребёнка из купели”.
Что же касается перехода от гиракотерия с четырёхпалой передней ногой к мезогиппусу
с трёхпалыми ногами, то я должен опять задать своим оппонентам вопрос: господа
хорошие, а сами-то вы часто смотрите в те схемы, которые приводите в качестве
“неопровержимых аргументов”? Дело в том, что переход от гиракотерия к мезогиппусу
– это несколько упрощённый подход, принятый в школьных и институтских учебниках.
На схеме же (в том числе на той, что приводится на этом сайте “коневодов-любителей”)
ясно видно, что ПРЯМОГО перехода между гиракотерием и мезогиппусом просто не
существует! Этот “досадный” пробел заполняют последовательно два рода примитивных
лошадиных – Orohippus и Epihippus.
Между прочим, ниже по тексту на указанном сайте в очередной раз обнаруживаются
удивительные вещи! Например, господа креационисты любезно приводят характеристики
доисторических родов лошадей, чтобы читателю можно было представить себе героев
этого повествования. Вот здесь-то и начинаются “чудеса в решете”. Так, в характеристике
мезогиппуса чёрным по белому написано:
“Немного крупнее, чем Hyracotherium, 24 дюйма в плечах.
Mesohippus был пальцеходящим [в оригинале – “pad-footed”, то есть “опирающийся на подушечки пальцев” - В. П.] с тремя пальцами на всех четырех ногах, с крошечным рудиментарным 4-ым пальцем передней ноги”.
Волшебным способом в родословной лошади также появляются “потерянные”
орогиппус и эпигиппус. Это лишний раз говорит, что “из песни слов не выкинешь”.
Кстати, о ногах и пальцах лошадей… Креационисты утверждают, что переходных форм
между разновидностями лошадей, имеющими разное строение ног, не существует.
Вот информация с сайта www.answersingenesis.org – отрывок из статьи Джонатана
Сарфати “Non-evolution of horse” – весьма оригинальная попытка объяснения “кажущегося”
эволюционного механизма на примере преобразования ног лошадей.
“Важная часть библейской модели творения – тот факт, что различные виды существ были созданы с большим количеством генетической информации. Естественный отбор может сортировать эту существовавшую ранее генетическую информацию путём отсеивания существ, не соответствующих определённой окружающей среде. Таким образом много различных разновидностей может появиться в различных типах окружающей среды. Обратите внимание, что этот процесс отсеивания включает потерю информации, в чём не соответствует эволюции от частиц к людям, которое требует, чтобы лишённые разума процессы добавили новую информацию.
Также, многое из этой (сотворённой) генетической информации, возможно, было латентной (скрытой, то есть особенности, закодированные так, чтобы не быть выраженными в потомстве) в исходно сотворённых видах. Они также имели другие управляющие или регулирующие гены, которые “включают” или “выключают” другие гены. То есть, от них зависит, будет или не будет декодирована информация в гене, а признак, таким образом, будет выражен у животного. Это допустило бы очень быстрые и “скачкообразные” изменения, которые всё равно являются изменениями, включающими уже сотворённую информацию, но не генерацией новой информации.
Если применить эти принципы к лошади, то кодирование генетической информации для дополнительных пальцев ноги присутствует, но “выключено” у наиболее современных лошадей. Иногда в наши дни рождается лошадь, у которой гены “включены”, и, конечно, многие ископаемые лошади также имели “включённые” гены. Это объяснило бы, почему нет никаких переходных форм, показывающих постепенно уменьшающийся размер пальца ноги”.
Сарфати не одинок в своей теории. Её активно поддерживают и отечественные креационисты, не уточняя, однако, механизмов этого процесса.
“С другой стороны, в наше время иногда рождаются лошади и мулы с лишними пальцами, оканчивающимися карликовыми копытцами, совсем как у “первобытных” лошадей. Поэтому в существовании трех- и четырехпалых лошадей в прошлом нет ничего сверхъестественного. Просто первоначально сотворенный род лошадей имел достаточный запас наследственной изменчивости и первоначально это проявилось во множестве разновидностей, но впоследствии, видимо, часть этого разнообразия была безвозвратно утрачена.” (Православное Мiровоззрение и современное естествознание)
Очень соблазнительно было бы объяснить появление и исчезновение
признаков такими простыми вещами: “включил ген” - есть признак, “выключил” –
нет признака. Но такая теория имеет слабое место – как объяснить постепенное
изменение признака? Ведь в теории “включенных генов” есть только два состояния
гена – “есть” и “нет”. Но у ископаемых лошадей мы видим значительно большее
количество пальцев на ноге, чем проявляется при явлении атавизма, и форма этих
пальцев сильно варьирует. Например, самые ранние лошади типа гиракотерия (эогиппуса)
имеют четырёхпалую переднюю ногу, и опираются на подушечки пальцев, а не на
ногти (когти, копыта). В английских текстах про таких лошадей читаем: “pad-footed”,
то есть “опирающийся на подушечки пальцев”, где “pad” – подушечка (как, например,
на ноге кошки или собаки). Более поздние лошади также опираются на мягкие ткани
ноги, но у них уже три пальца. Далее в палеонтологической летописи появляются
трёхпалые лошади, опирающиеся уже на роговое копыто, а не на мягкие подушечки
пальцев. Далее появляются лошади, у которых боковые пальцы уже не постоянно
касаются земли, а за ними лошади с редуцированными, но заметными боковыми пальцами,
не касающимися земли. И в конце концов появляются полностью однопалые лошади.
В свете этих данных хочу спросить: сколько генов-“выключателей” было у гена,
отвечающего за форму ноги и количество пальцев?
Как пели в песенке герои одного из мультфильмов: “Ах, если бы, ах, если бы,
не жизнь была бы – песня бы!”. А как бы шла эволюция, если бы животные были
именно такими – с избыточным набором мудро и своевременно “включающихся” генов?
Тогда бы, как только появлялось подходящее место обитания, его сразу занимал
бы разумно подходящий для этого вид. И тогда небольшие свинообразные существа
не позволили бы неуклюжим болотным носорогам хилотериям валяться в грязи, а
сразу “включили” бы нужные гены и превратились в бегемотов. А крошечный Diacodexis
без лишних церемоний превратился бы за пару-тройку поколений в антилопу (чтобы
не было бегающих тапиров Lophialetes – экое баловство!) или жирафа (чтобы с
ним не тягался понапрасну носорог Indricotherium, который всё равно потом вымер),
и тогда жизнь приобрела бы современные черты задолго до нашего времени. Не было
бы на родословных древах длинных стволов, заканчивающихся вымиранием, не было
бы путей “проб и ошибок”. Но переходные формы всё равно оставались бы – только
здесь признаки появлялись бы один за другим резко и в полностью выраженном виде.
Разница была бы в выраженном “наборе” этих готовых признаков, примерно вот так:
абвг > Абвг > АБвг > АБВг > АБВГ. И последовательность таких форм
всё равно была бы найдена.
Креационисты любят утверждать, что теория эволюции недоказуема, и основана на
предположении. Но сами не стесняются выдвигать голословные и беспочвенные теории.
Как можно охарактеризовать теорию о сотворённых животных, имеющих, якобы, избыточный
набор генов? Точно такими же словами – как абсолютно недоказуемое предположение.
Это лишь теоретическая выкладка, не подкреплённая никакими материальными свидетельствами.
Да, в организмах живых существ есть огромное количество т. н. “молчащей” ДНК.
Но кодируют ли её последовательности какие-либо признаки, характерные для вымерших
предков или родственников некоего исследуемого вида? Доказательства – где они?
Кроме того, если предположить, что гены в процессе развития некоей группы живых
существ только утрачиваются, то выходит, что при циклическом изменении климата
(например, от сухого к влажному и опять к сухому) в глобальном масштабе эволюция
просто закончится на “втором витке”. Просто все носители нужных для выживания
генов вымрут при изменении климата в другую сторону, а новая генетическая информация,
нужная при обратном изменении климата, просто не появится.
Ещё одна проблема: делает ли эволюция скачки?
“Одним из хорошо исследованных примеров, на которые опирается концепция прерывистого равновесия, является эволюция озерных моллюсков Турканы, области на севере Кении. Их развитие в течение почти пяти миллионов лет было изучено на массовом материале. Оказалось, что новые виды или появляются внезапно, или преобразование видов происходит за пять – пятьдесят тысяч лет. Длительность стабильного стазиса, во время которого не происходит крупных изменений, составляет не меньше миллионов лет.
Но. Во-первых, практически во всех случаях новые виды моллюсков возникали после перерывов в накоплении озерных осадков, вслед за прослоями вулканических туфов. Значит, эволюционные события в перерывах постепенно развития моллюсков не документированы, а происходили за пределами исследованной территории. То есть речь не об отсутствии эволюции, а об отсутствии фактов о ней. [выделено мной – В. П.] Во-вторых, в некоторых случаях старые и новые виды моллюсков соединены рядом переходных форм (видообразование шло плавно).
Все это говорит о неполноте документации хода эволюции на уровне видов даже в классическом случае с моллюсками Турканы. И в других примерах “прерывистой эволюции” перерывы чаще всего отмечаются, когда есть основания говорить о перерывах времени наблюдения за последовательностью сменяющих друг друга поколений”. (Л. П. Татаринов “Суждение палеонтолога об эволюции”)
Действительно, эволюция лошади, как и эволюция приведённых в примере моллюсков, известна заведомо не полностью: условия далеко не всегда благоприятствуют захоронению, и даже напротив, полное уничтожение остатков мёртвого животного или растения является нормой. Да, в эволюции лошади есть перерывы. Другое дело – как их истолковывают сторонники разных научных теорий.
3. Неизвестный предок Hyracotherium.
-“Джордж Гейлорд Симпсон, после заявления, что нигде в мире нет никаких следов окаменелости, которая закрыла бы значительный промежуток между Hyracotherium (“Eohippus”), который, как полагают эволюционисты, был первой лошадью, и его предполагаемым предковым отрядом Condylarthra, продолжает говорить: “Это верно для всех тридцати двух отрядов млекопитающих … Самые ранние и наиболее примитивные известные члены каждого отряда уже имеют основные черты отряда, и ни в каком случае не известна более-менее непрерывная последовательность от одного отряда до другого. В большинстве случаев разрыв настолько резкий, и промежуток настолько большой, что происхождение отряда является умозрительным и весьма спорным”. (Simpson, G. G., Tempo and Mode in Evolution, New York: Columbia University Press, 1944, p. 105)
-Нигде, например, нет промежуточных форм, документирующих переход от не-лошадиного предка (возможно, кондилартра) с пятью пальцами на каждой ноге, к Hyracotherium с четырьмя пальцами на передней ноге и тремя на задней. Также нет переходной формы между четырёхпалым Hyracotherium и трёхпалым Miohippus, или между последним, имеющим зубы для объедания листвы, и трёхпалым Merychippus, имеющим высоко-корончатые зубы для ощипывания травы. Наконец, однопалые травоядные, типа Equus, появляются внезапно, без промежуточных звеньев, показывающих постепенное развитие от трёхпалых травоядных. (The Origin Of Mammals, ICR Impact No. 87 Duane T. Gish, Ph.D, 1980)
-Самый ранний член эволюционного ряда лошадей, Hyracotherium (Eohippus), не имеет никакой связи, ни в какой форме, с его предполагаемыми предками, кондилартрами. (Kofahl, R.E., Handy Dandy Evolution Refuter, Beta Books, San Diego, California, 1997, p. 159)
-“Нигде не могут быть найдены переходные промежуточные формы, показывающие переход от пятипалого предка к Hyracotherium (Eohippus), который имеет три пальца на своей задней ноге и 4 пальца на своей передней ноге. В силу этого обстоятельства единственный выход состоит в том, чтобы считать выступы на ногах Eohippus как атрофированный палец и предполагать, что это идёт от пятипалого предка”. (Dr. Ali Demirsoy)”.
Сразу же хочется ответить на первый аргумент креационистов.
Господа хорошие, вы когда-нибудь новости читаете? По-моему, вы “кормите” доверчивую
паству явно “несвежими” фактами. Дело в том, что цитируемый вами труд Джорджа
Гейлорда Симпсона датирован 1944 годом (ровнёхонько 60 лет назад)! Критикуемая
вами информация в то время принадлежала к числу субъективных и умозрительных
выводов, основанных на неполном знании ископаемых животных. Я не хочу сказать
этим, что Дж. Г. Симпсон был неучем (про Д. Гиша я готов сказать это хоть Богу,
хоть дьяволу). Просто в то время уровень изученности ископаемых животных был
гораздо ниже, чем сейчас, и видов было известно гораздо меньше.
Обратите внимание, Симпсон сказал “ИЗВЕСТНЫЕ члены каждого отряда”, внеся этим
долю субъективности в свой вывод. С того времени многое изменилось – большое
количество новых находок заставило пересмотреть некоторые теории, а что-то вообще
стало историей и данью прошлому.
Родство кондилартров и копытных признано безоговорочно. Мало того, палеонтологическая
летопись демонстрирует плавный переход от кондилартров к первым представителям
современных отрядов копытных:
“Относительно полно документирована крупномасштабная радиация копытных в самом конце мела и начале палеоцена. Тогда из единственного семейства древних кондилартр всего за 2-3 млн. лет дифференцировалось 16 самостоятельных отрядов (Van Valen, 1978). …
… Наиболее существенно здесь то, что многие виды и роды на следующих этапах радиации признавались представителями разных семейств и отрядов. На уровне особей эволюция ранних кондилартр выглядит не особенно отличающейся от типичного микроэволюционного процесса. Разница заключается не в самих видах, а в возможностях, реализованных их потомками” (Р. Кэрролл “Палеонтология и эволюция позвоночных” М., Мир, т. 3, стр. 151)
Прямое родство гиракотерия (эогиппуса) и фенакодуса признано далеко не всеми эволюционистами. Раньше, возможно, предположение о таком сходстве было выдвинуто на основе недостаточно изученных останков фенакодуса. В настоящее время фенакодус признаётся, можно сказать, “братом” эогиппуса – найден другой вид первичных копытных, Tetraclaenodon, который демонстрирует более примитивное строение конечностей, чем оба сравниваемых животных. Задняя нога фенакодуса имеет черты специализации, которые исключают отношение “предок – потомок” между ним и эогиппусом.
“Phenacodus из позднего палеоцена – раннего эоцена считался типичным кондилартром … Среднепалеоценовый род Tetraclaenodon известен менее полно, но занимает более близкое положение к предкам непарнокопытных. Среди всех фенакодонтов, которые ввиду отсутствия сильной специализации ещё могли дать начало непарнокопытным, он наиболее продвинут, но всё же гораздо примитивнее, чем древнейшие формы последних.
Среди примитивных непарнокопытных лучше других изучен Hyracotherium, обычный в нижнем эоцене. Радински (Radinsky, 1966) описал ряд изменений, произошедших при переходе к этому роду от тетракленодона; важнейшие из них затрагивают зубной и локомоторный аппараты”. (Р. Кэрролл “Палеонтология и эволюция позвоночных” М., Мир, т. 3, стр. 97).
Аргумент креационистов об отсутствии переходной формы нельзя
считать абсолютно исключающим процесс эволюции. Дело в том, что его можно понимать
двояко. Креационисты понимают это как то, что таких живых существ ВООБЩЕ НЕ
БЫЛО. Эволюционисты понимают так, что эти живые существа просто НЕ НАЙДЕНЫ.
А это, как говорят в Одессе, “две большие разницы”. Чтобы признать правоту креационистов,
нужно, по меньшей мере, доказать, что все виды ископаемых животных уже найдены.
Но разрушить это доказательство может лишь одна удачная экспедиция. Поэтому
я не рекомендовал бы доктору Али Демирсою особенно сильно настаивать на своей
точке зрения – любая новая находка выставит его на посмешище.
Понимание эволюционистами факта отсутствия искомой переходной формы основано
на том, что не все живые организмы могут быть представлены в палеонтологической
летописи. Эволюция ранних лошадей, в частности, происходила в условиях, не благоприятных
для захоронения: в лесах. Обилие почвенных кислот, многочисленные животные,
питающиеся падалью и невозможность быстрой консервации останков в бескислородной
среде (например, в болоте или озёрном иле) – вот факторы, губительные для фоссилизации
и сохранения образцов флоры и фауны.
Родство гиракотерия (эогиппуса) с лошадями и его непосредственная принадлежность
к семейству лошадиных прямо доказаны по особенностям его анатомии:
“МакФадден (MacFadden, 1976) показал, что слияние овального и среднего рваного отверстий, как и перемещение отверстия зрительного нерва к группе постеровентральных отверстий (или слияние с ними) представляют собой производные признаки, указывающие на принадлежность гиракотерия к той же монофилетической группе, что и лошадиные (Equidae)”. (Р. Кэрролл “Палеонтология и эволюция позвоночных”, М. Мир, 1993, том 3, стр. 99)
В свете этих слов, может быть, не стоило креационисту Kofahl,
R. E. называть свою книгу “Handy Dandy Evolution Refuter” (“Удобный и первоклассный
опровергатель эволюции”)?
Что же касается отсутствия переходных форм между разными видами лошадей, о которых
говорит Дуэйн Гиш, то ему лично я могу порекомендовать рассматривать и критиковать
эволюцию лошади на том уровне, что изложен в научных трудах, а не в школьном
учебнике (где материал заведомо упрощён из дидактических соображений). Также
можно посоветовать ему обратиться к изучению родословных древ лошадей, которые
критикуются в трудах самих же креационистов. Он с удивлением (или с удовлетворением?)
отметит, что “пропасти” между гиракотерием и миогиппусом, миогиппусом и мерикгиппусом,
а также между трёхпалыми и однопалыми лошадями оказываются заполненными разными
родами древних лошадей. Так что аргумент Гиша оказался беспочвенным.
4. Трёхпалый Neohipparion жил вместе с однопалым Pliohippus.
Стоит также отметить, что в формации Rattlesnake, John Day Country в северо-восточном Орегоне, трёхпалая лошадь Neohipparion найдена вместе с однопалой лошадью, Pliohippus. (cf. Nevins, S., Creation Research Society Quarterly, Vol. 10, 1974, p. 196) Никаких переходных форм между этими двумя видами не найдено. В других случаях “примитивные” виды рода, вроде таковых у Merychippus, найдены в геологических формациях, гораздо более молодых чем те, что содержат “продвинутые” разновидности. (cf. Gregory, J. T., University of California Publications in Geological Sciences, Vol. 26, 1942, p. 428) (The Origin Of Mammals, ICR Impact No. 87 Duane T. Gish, Ph.D, 1980)
Простите, уважаемые господа креационисты, но ведь вы сами
говорите, что родословное древо лошадей представляет собой не одну прямую линию,
а целый “куст” из нескольких эволюционных линий! Поэтому не представляется чем-то
удивительным, что в отложениях одной и той же эпохи найдены представители нескольких
родов лошадей.
В настоящее время на Земле обитает, например, много видов антилоп, среди которых
есть рогатые, есть те, у которых рога есть лишь у самцов, и есть вообще безрогие.
Есть антилопы с рогами длиной до метра, есть виды с витыми штопорообразными
рогами, и есть такие, у которых рога едва высовываются из шерсти на голове.
А есть один вид антилоп с четырьмя небольшими рогами на голове. И такое многообразие
антилоп, обитающих порой на одной территории и пасущихся бок о бок, ни у кого
не вызывает удивления. Почему же наличие в природе прошлой геологической эпохи
нескольких видов лошадей должно вызывать такие странные измышления? Например,
в конце миоцена на Земле жило, судя по ископаемым находкам, более 10 родов самых
разных лошадей.
Если рассмотреть именно приводимый креационистами пример, заодно взглянув на
любезно приводимые теми же самыми креационистами варианты родословного древа,
то выяснится одна почти удивительная вещь: все представители гиппарионов (как
собственно гиппарион, так и многочисленные производные роды, в том числе упомянутый
Neohipparion) не относятся к прямым предкам современных лошадей! Это представители
одной из крупных боковых ветвей родословного древа лошадей, на наличие которых
креационисты неустанно указывают эволюционистам. Все гиппарионы в лучшем случае
паслись рядом с прямыми предками лошадей, но не были этими самыми предками.
Neohipparion и Pliohippus – не родственные виды, а представители двух ветвей
эволюционного древа лошадиных; таким образом, они заведомо не связаны отношениями
преемственности. Переходные формы между этими представителями разных ветвей
бесполезно искать в отложениях одного возраста с рассматриваемыми видами: они
просто не существуют в этих отложениях. Родство между указанными видами следует
искать скорее во времени: это будет общий предок, живший заведомо раньше двух
указанных разновидностей лошадей.
Это можно объяснить, что называется, “на пальцах”: обычно никто, пребывая в
здравом уме, не пытается искать общих родителей у двоюродных или троюродных
родственников. Кстати, наличие у некоего человека двоюродного брата или сестры
вовсе не является условием, исключающим возможность существования на земле этого
субъекта. Так почему же трёхпалая и однопалая лошадь, не являясь близкими родственниками,
не могут существовать одновременно, и даже бок о бок?
Этот пример иллюстрирует излюбленный приём убеждения креационистов: ткнуть пальцем
в две произвольно выбранные формы животных, и съязвить по поводу отсутствия
между ними переходной формы.
Кроме того, у креационистов, чувствуется, вызывает удивление, почему одновременно
на Земле живут как “примитивный”, так и “прогрессивный” виды животных. В таком
случае пусть креационисты удивляются ещё больше: на Земле прямо сейчас, когда
они читают эти строки, одновременно живут как прогрессивные (правильнее сказать
– “специализированные”) кенгуру и вомбаты, так и сравнительно примитивные опоссумы
и сумчатые мыши. Вообще, кто сказал, что “примитивные” виды обязательно должны
вымирать? И ещё, что есть “примитивный”? Понятие “примитивный” ещё не означает
“плохо приспособленный” - оно означает “сохранивший больше черт сходства с предками
по сравнению с иными родственными формами”. Но если “примитивный” вид хорошо
приспособлен к своей среде обитания и находится в ней вне конкуренции, он не
вымрет, пока не появится другой, приспособленный ещё лучше. Поэтому наличие
“примитивных” форм в более молодых отложениях, чем ранее найденные, и сосуществование
их с “прогрессивными” сородичами не может служить опровержением эволюции.
5. Прерывистое, не постепенное “изменение”.
Хотя эволюционисты создают иллюзию медленного, постепенного, хорошо документированного изменения от Hyracotherium до Equus, фактически никаких эволюционных промежуточных звеньев не существует. Каждое из животных внезапно появляется в летописи окаменелостей (прерывистой), без физических признаков переходной разновидности. (Bowden, M., The Rise of the Evolution Fraud, Creation-Life Publishers, San Diego, California, 1982, p. 117)
Это утверждение креационистов базируется на имеющем место
быть факте заведомой неполноты палеонтологической летописи. Вид появляется в
ней лишь тогда, когда его численность становится достаточно высокой, чтобы сделать
вероятным даже такое событие, как успешное захоронение останков его представителей.
А это – весьма случайный процесс. Поэтому палеонтологам известны остатки в лучшем
случае самых массовых, “фоновых” видов.
Изменения при превращении одного вида в другой могут происходить в одной отдельно
взятой популяции, тогда как другие популяции могут стабильно сохранять свои
признаки. А изменившаяся популяция может какое-то время просто не встречаться
в палеонтологической летописи из-за того, что её численность слишком мала. Но
в это время в ней происходят разного рода изменения – идёт освоение новой экологической
ниши. Если эти изменения не привели к успеху – популяция просто вымрет, не оставив
следа среди окаменелостей. Если же она добивается успеха, её численность растёт,
и останки её представителей получают шанс на захоронение.
Таким образом, мы получаем следующее: вид появляется в палеонтологической летописи
значительно позже времени своего фактического появления в экосистеме.
Исчезновение вида из отложений тоже не совпадает с временем фактического вымирания.
Было общепринято, например, что все динозавры вымерли в конце мелового периода.
Но сейчас всё же есть единичные находки этих рептилий из отложений раннего палеоцена.
А кистепёрая рыба латимерия (Latimeria) до сих пор живёт на Земле, и мало того,
даже второй вид этого рода обнаружен в Индонезии. И это на фоне досадного пробела
в палеонтологической летописи длиной более 65 миллионов лет – ископаемые кистепёрые
рыбы моложе конца мелового периода неизвестны.
Впрочем, лошадям в этом плане всё же повезло. Твёрдые лошадиные зубы прекрасно
сохраняются в кайнозойских отложениях, демонстрируя широкий спектр изменчивости
как между видами внутри рода, так и от рода к роду. Например, в линии, ведущей
от Mesohippus к Equus, чётко прослеживаются переходы в строении зубов (подробнее
см. в комментарии к возражению № 13)
Впрочем, креационисты далеко не согласны с тем, что это наглядный пример… Их
аргументация в данном случае весьма оригинальна, и состоит в том, чтобы объявить
эти формы лошадей родственными, либо вообще одним видом (очевидно, “барамином”,
“родом” в широкой библейской трактовке). Такая участь уготована лошадям из перечисленных
родов, о чём сказано в характеристиках этих животных на том же сайте, где и
приведены упомянутые пятнадцать тезисов:
“Parahippus, Miohippus and Mesohippus are really the same animal. They are classified differently because different skeletons of the same animal can exhibit minute differences. They may be two closely related cross-fertile species like Mule deer and White Tail deer for example. The differences are also easily accounted for by if adult and juvenile specimens had been found. There is a huge amount of speculation among evolutionists and this is another example!
Teeth and leg differences between these three animals (Parahippus, Miohippus and Mesohippus) are no different that we find within men or dogs given "race and age" differences”.(Parahippus, Miohippus и Mesohippus являются в действительности одним и тем же животным. Они классифицированы по-разному, поскольку различные скелеты одного и того же вида животных могут показывать несущественные различия. Они могут быть двумя близкородственными, плодовитыми при скрещивании видами, подобно чернохвостому и белохвостому оленям, например. Различия легко объясняются тем, что могут быть найдены взрослые и молодые экземпляры. Из этого происходит множество спекуляций среди эволюционистов, и вот ещё один пример!
Различия зубов и ног между этими тремя животными (Parahippus, Miohippus и Mesohippus) не отличаются от тех, что мы находим у людей и собак как “расовые и возрастные” отличия).
Обвиняя эволюционистов в умозрительных выводах, креационисты
сами не стесняются делать ровно такие же, и даже более беспочвенные. Никто из
ныне живущих людей (и даже вообще никто из людей) не видел спаривания между
лошадями этих родов, поэтому вывод о возможности гибридизации между представителями
разных родов лошадей можно считать чистейшей фантазией. Столь же необоснован
вывод о плодовитости гибридов между этими видами. В природе такое происходит
далеко не всегда, даже если виды, образующие гибрид, принадлежат к одному роду
и живут рядом.
Если предположить, что один из названных родов лошадей представляет собой гибрид,
то стоит на секунду задуматься: какова вероятность образования такого гибрида?
Если такой гибрид появился в природе, следовательно, оба родительских вида обитали
в одной и той же области хотя бы на части ареала. Каждый вид – это совокупность
популяций, сравнительно изолированная от других ей подобных. Два близких вида,
живущих совместно, обычно вырабатывают механизмы, препятствующие гибридизации
и “растворению” друг в друге: разное поведение, разные сроки размножения, либо
бесплодие гибридов на генетическом уровне. Современные виды семейства лошадиных
пошли как раз по последнему пути.
Что мы наблюдаем на уровне ископаемого материала?
Во-первых, сами по себе факты находки тех существ, которых креационисты считают “гибридами”. В природе гибриды – единичное явление. Шансов на то, что сохранится именно этот единственный гибрид, практически нет. А ведь палеонтологи находят множество костей животных разных возрастов. Это говорит о том, что появление особей такого строения носило массовый характер. Неужели в прошлом существовало по крайней мере несколько популяций лошадей-“извращенцев”, отличавшихся противоестественной склонностью к спариванию с особям других видов?
Во-вторых, известно большое количество популяций этих “сомнительных” родов, в которых наблюдается спектр изменчивости, в том числе во времени. Эти признаки обработаны статистически и показывают единство популяции, а не случайный подбор отдельных особей (подробнее см. в книге В. Гранта “Эволюция организмов”, гл. 26). Это говорит о том, что палеонтологи имеют дело не с гибридами, а с непрерывно меняющимся и широко распространённым родом, внутри которого происходят изменения, подобные тем, что наблюдаются у современных видов животных и растений.
В-третьих, внутри “сомнительного” рода выделяется
ряд чётко очерченных видов. Гибрид же во многих случаях просто не сможет дать
второго поколения. Возможность видообразования при таком раскладе не стоит даже
обсуждать.
Может быть, мы имеем дело с животными разного возраста? Определить относительный
возраст животного по состоянию головок суставов и степени стёртости зубов можно,
причём очень легко. Так что палеонтолог легко отличит старую лошадь мелкого
вида от молодой лошади крупного вида.
“Pliohippus, Dinohippus and Equus are all cross-fertile horses. They are one toed with hoofs. Dinohippus and Pliohippus are merely an extinct variety of true horse.
Pliohippus is classified distinct from Equus because of very its curved teeth whereas Equus's teeth are straight”.
(Pliohippus, Dinohippus и Equus – плодовитые при скрещивании лошади. Они однопалые с копытами. Dinohippus и Pliohippus – просто вымершие разновидности настоящих лошадей.
Pliohippus классифицирован как отличающийся от Equus из-за своих очень изогнутых зубов, тогда как зубы Equus прямые).“Dinohippus is classified distinct from Equus because of teeth curved midway between Equus and Pliohippus. (Equus had straight and Pliohippus has very curved teeth)”.
(Dinohippus классифицирован как отличающийся от Equus из-за зубов, изогнутых в промежуточном положении между Equus и Pliohippus. (Equus имеет прямые, а Pliohippus – очень изогнутые зубы).“Modern horses are merely those horses that are not extinct. These include onagers, Quagga, kiang and various Zebra, asses & donkeys. All horses are cross-fertile”.
(Современные лошади – это просто те лошади, которые не вымерли. Они включают онагров, кваггу, кианга и различных зебр и ослов. Все лошади плодовиты при скрещивании).
Как говорила Алиса в Стране Чудес, “всё любопытственнее и любопытственнее…”.
Снова и снова повторяются одни и те же тезисы. У меня есть рекомендация к господам
креационистам, и даже не одна, а две сразу.
Во-первых, поделитесь, пожалуйста, со всем учёным миром методикой реконструкции
поведения давно вымерших животных. Если методика верна, её первооткрыватель
станет очень известной “шишкой” среди учёных.
Во-вторых, господа, обратите внимание на современных лошадей и на их гибриды.
Дело в том, что даже школьники прекрасно знают, что мул и лошак (продукты скрещивания
ослов и лошадей в разных вариантах) бесплодны! Также бесплоден зеброид – гибрид
зебры с лошадью или ослом.
Поистине странно, что взрослые люди, именующие себя учёными, не знают вещей,
о которых написал ещё старик Альфред Брем более ста лет назад, не говоря уже
о том, что о бесплодии мулов было известно едва ли не с античных времён.
А теперь о вещах, о которых Брем, может быть, не знал, а античные скотоводы
даже не задумывались.
Сводя три рода ископаемых лошадей в один, креационисты, я считаю, пытаются применить
в жизни любимый принцип Иосифа Виссарионовича: “нет человека – нет проблемы”.
Только вместо человека здесь лошадка, причём уже давно неживая.
Креационисты пытаются доказать, что несколько родов ископаемых лошадей выделено
сторонниками теории эволюции для того, чтобы показать наличие “переходной формы”.
Правомерно или неправомерно такое выделение, соответствует или не соответствует
родовому уровню совокупность признаков, на основании которых выделены данные
роды лошадиных – это уже дело десятое. Самое главное – после таких переименований
изменится, разве что, надпись на табличке в музее. Анатомические признаки останутся
неизменными.
Креационисты также сетуют на то, что от лошадей рода Dinohippus (из которых
выводится современный род Equus) найдены, мол-де, лишь фрагментарные останки:
“A small part of the skull is all that this "separate species" is based upon!”
(Маленькая часть черепа – это всё, на чём базируется этот “отдельный вид”!)
У Кэтлин Хант про этих лошадей сказано следующее:
“Dinohippus (поздний миоцен, 12 млн. лет назад) - однопалый питающийся травой, с пружинистыми ногами. Очень “конские” ноги, зубы и череп, с более прямыми зубами и меньшими ямками. Сначала был D. spectans, сопровождаемый D. interpolatus и D. leidyanus. Более поздней разновидностью был D. mexicanus, с даже более прямыми зубами и даже меньшими ямками”.
Упоминаются четыре вида этого рода… Может ли им всем одновременно
принадлежать один обломок черепа?
Что изменится, если диногиппус или плиогиппус станут называться Equus? Просто
тогда они станут считаться примитивными представителями этого рода. Возможно,
тогда переход от одного рода к другому не будет числиться среди явлений макроэволюции
(чего, очевидно, добиваются креационисты), но переход между зубами разной формы
всё равно будет прослеживаться на ископаемом материале. Если учесть, что принципиальной
разницы между механизмами макро- и микроэволюции нет, то креационисты не смогут
ничего доказать этим переименованием.
Переименование плиогиппуса в Equus не означает, что в миоцене по Земле начали
бегать лошади современного вида. У этих прото-лошадей в любом случае останутся
зубы того типа, что характерен для “формы плиогиппуса”. И в любом случае будет
ясно заметен переход от примитивных изогнутых зубов “формы плиогиппуса” к прямым
зубам современных лошадей через стадию слабо изогнутых зубов “формы диногиппуса”.
Так что факты в очередной раз оказываются упрямой вещью, и сменой вывесок их
не отменишь.
А есть ли ещё какие-то переходы между видами лошадиных? Несомненно, есть. Об
этом свидетельствуют учёные, много лет исследовавшие эволюцию лошадей. Вот данные,
которые приводит Кэтлин Хант (Kathleen Hunt) в работе “Transitional Vertebrate
Fossils FAQ” (“Часто задаваемые вопросы об ископаемых переходных формах позвоночных”).
Эту статью можно найти здесь:
http://www.talkorigins.org/faqs/faq-transitional
В статье приводится множество фактов наличия переходных форм в основных группах
позвоночных, в том числе таких, о которых любят порассуждать креационисты. Также
указаны имеющие место в настоящее время пробелы, не заполненные палеонтологическими
находками. О лошадях она пишет следующее:
“Переходы от вида к виду.
Лошади: Gingerich (1980) документировал видообразование от Hyracotherium grangeri к H. aemulor. Prothero и Schoch (1989) упоминают некоторые промежуточные окаменелости, которые связываются позднего Orohippus с Mesohippus celer. MacFadden (1985) документировал многочисленные плавные переходы среди трехпалых лошадей, особенно среди Merychippus и различных гиппарионов. Hulbert (в Prothero и Schoch, 1989) показал, что Dinohippus плавно переходит в Equus в последовательных плиоценовых слоях отложений. Simpson (1961) описывает постепенную потерю боковых пальцев у Pliohippus через 3 последовательных слоя отложений раннего плиоцена”.
В этой же работе приводятся переходы древних лошадиных от рода к роду, причём рассматриваются они на ВИДОВОМ уровне:
“Loxolophus (ранний палеоцен) – Примитивный кондилартр с довольно низкокоронковыми зубами, возможно, предковый для фенакодонтидных кондилартров.
Tetraclaenodon (средний палеоцен) – Более продвинутый палеоценовый кондилартр из семейства фенакодонтид, и почти наверняка предковый для всех непарнокопытных (другого отряда). Длинные, но неспециализированные конечности; 5 пальцев на каждой ноге (№1 и №5 мельче). Более эффективно устроенное запястье.
ПРОБЕЛ: Почти неизвестно ископаемых остатков непарнокопытных из позднего палеоцена. Это в действительности небольшой пробел; он заметен лишь потому, что ископаемая летопись непарнокопытных в остальном очень полная. Последние открытия ясно показали, что первые непарнокопытные появились в Азии (плохо изученный континент), и есть надежда, что ожидающиеся новые раскопки в Азии заполнят этот маленький, но досадный пробел. Первый ключ к этому уже появился:
Radinskya yupingae (поздний палеоцен, Китай) – Недавно открытый кондилартр, подобный непарнокопытным. (McKenna et al., в Prothero & Schoch, 1989.)
Hyracotherium (ранний эоцен, около 55 млн. лет назад; первоначально “Eohippus”) – Знаменитая “лошадь рассвета”, мелкое, собакообразное непарнокопытное с выгнутой спиной, короткой шеей, зубами всеядного зверя и короткой мордой. 4 пальца на передних, и 3 на задних ногах. По сравнению с Tetraclaenodon имел более длинные пальцы, взаимосвязанные кости лодыжки и несколько отличающиеся бугры на зубах. Возможно, эволюционировал от Tetra. примерно за 4 – 5 млн. лет, возможно, через азиатский вид вроде Radinskya. Заметим, что Hyrac. отличается от других ранних непарнокопытных (таких, как предки тапиров/носорогов) только маленькими изменениями зубов и размером тела.
Hyracotherium vassacciense (ранний эоцен) – Обычный вид, который, возможно, был родоначальником лошадиных.
Orohippus (средний эоцен, ~50 млн. лет назад) -- Мелкий, 4/3-палый, развивающий характерные для листоядного гребни на зубах.
Epihippus (поздний эоцен, ~45 млн. лет назад) -- Маленький, 4/3-палый, хорошо выраженные гребни на зубах, листоядный.
Epihippus (Duchesnehippus) – Более поздний подрод с Mesohippus-подобными зубами.
Mesohippus celer (самый поздний эоцен, 40 млн. лет назад) – Трёхпалый на всех ногах, листоядный, немного более крупный.
Mesohippus westoni (ранний олигоцен) – Более поздний, более продвинутый вид.
Miohippus assiniboiensis (средний олигоцен) – Этот вид откололся от ранних Mesohippus через кладогенетическую эволюцию, после которой Miohippus и Mesohippus сосуществовали следующие 4 млн. лет. Заметно крупнее, более длинный череп, лицевая ямка глубже и более выражена, слабо отличающийся сустав лодыжки, изменчивый наружный гребень на верхних коренных зубах. В раннем миоцене (24 млн. лет назад) Miohippus начал активное видообразование. Злаки только начали эволюционировать, и зубы начали меняться соответствующим образом. Ноги и т. д. начали меняться для быстрого бега.
Kalobatippus (поздний олигоцен) – Трёхпалый листоядный с ногами промежуточного типа между Mio. и Para.
Parahippus (ранний миоцен, 23 млн. лет назад) – Трёхпалый листоядный/травоядный, развивший “пружинистую ногу”. Permanent establishment наружного гребня, который был так многообразен у Miohippus. Более сильные гребни на зубах и более высокие коронки зубов.
'Parahippus' leonensis (средний миоцен, ~20 млн. лет назад) – Трёхпалый листоядный/травоядный с уклоном в травоядность. Развивается пружинистая нога и высококоронковые зубы.
'Merychippus' gunteri (средний миоцен, ~18 млн. лет назад) – Трёхпалое травоядное, полностью “пружинисто-ногое” с высококоронковыми зубами.
Merychippus primus (средний миоцен, ~17 млн. лет назад) – Слегка более продвинут.
Merychippus spp. Из среднего-позднего миоцена (16-15 млн. лет назад) – Трёхпалые травоядные, “пружинисто-ногие”, размером с маленького пони. Разделились по всем доступным нишам травоядных, дав начало, по меньшей мере, 19 успешным видам трёхпалых травоядных. Развились копыта конского типа, кости ног сросшиеся. Полностью высоко-коронковые зубы с толстым слоем цемента и такими же гребнями, как у Parahippus. Линия, которая впоследствии привела к Equus, развилась в таком порядке: M. primus, M. sejunctus, M. isonesus (эти два последних ещё имели смесь примитивных, гиппарионовых и лошадиных черт), M. intermontanus, M. stylodontus, M. carrizoensis. Эти два последних выглядели в значительной степени по-лошадиному, с довольно маленькими боковыми пальцами, и дали начало серии более крупных трёхпалых и однопалых лошадей, известных как “настоящие лошадиные”. Всё ясно, верно?
МАЛЕНЬКИЙ ПРОБЕЛ: неизвестно, какой вид Merychippus (stylodontus? carrizoensis?) дал начало первому виду Dinohippus (Evander, in Prothero & S 1988).
Dinohippus (поздний миоцен, 12 млн. лет назад) – Однопалый травоядный вид с пружинящими ногами. Очень “лошадиные” ноги, зубы и череп, с более прямыми зубами и меньшими ямками. Первым был D. spectans, сменённый D. interpolatus and D. leidyanus. Более поздним видом был D. mexicanus, с ещё более прямыми зубами и ещё меньшими ямками.
Equus (Plesippus), также называемый группой “E. Simplicidens” (плиоцен, ~4 млн. лет назад) – Три близкородственных вида однопалых, пружинисто-ногих травоядных с высокими зубными коронками. Ямок нет, очень прямые зубы. Размером с пони, полностью “лошадеподобное” тело – жёсткий позвоночник, длинная шея, длинные ноги, сросшиеся и не поворачивающиеся в стороны кости ног, длинный нос, подвижная морда, высокие челюсти. Мозг немного крупнее, чем у раннего Dinohippus. Ещё имели некоторые примитивные особенности, такие, как простые зубы и небольшие лицевые ямки, которые утратили поздние виды Equus. Эти виды “простых лошадей” быстро разделились на по меньшей мере 12 новых видов 4-х различных групп. В течение первых крупных оледенений позднего плиоцена (2.6 млн. лет назад), некоторые виды Equus проникли в Старый Свет. Везде в мире Equus заняли экологическую нишу “крупного равнинного бегуна, питающегося жёсткой травой”.
Equus (Hippotigris) (плейстоцен) – Подрод современных однопалых, питающихся травой зебр с пружинящими ногами.
Equus (Equus) (плейстоцен) – Подрод современных однопалых травоядных лошадей и ослов с пружинистыми ногами. [примечание: очень редко лошадь рождается с маленькими боковыми пальцами, показывая, что некоторые лошади сохранили гены боковых пальцев.]
Сравните Equus с Hyracotherium, и посмотрите, как много всего изменилось. Если вы считаете животных разделёнными на “роды”, думаете, Equus и Hyracotherium могут быть объединены в один “род”?”
Как видим, в эволюции лошади прослеживается судьба отдельных
видов внутри рода, и имеются ясно выраженные эволюционные связи.
Относительно однопалых лошадей можно добавить, что переход от трёх пальцев ноги
к одному развился параллельно в нескольких линиях. “Стволовая” линия от Merychippus
вела через род Dinohippus к современным лошадям Equus. В то
же время от Merychippus отделилась ещё одна, боковая ветвь лошадиных
– род трёхпалых лошадей Pliohippus, а от него произошёл однопалый Astrohippus.
Продолжение следует