Глава 10
ПАЛЕОФИТОГЕОГРАФИЯ ПЕРМСКОГО ПЕРИОДА
С.В. Наугольных
Введение
В современной фитогеографии используются два основных подхода
к ординации растительного покрова: флористический и физиономический. Первый
основан на
сравнительном
анализе конкретных флор, выявлении сходства и различий при сравнении таксономических
списков. Второй подход можно также назвать экологическим или ландшафтным, поскольку
он основан на анализе экологических типов растительности, изучении форм роста
и жизненных форм, а также сукцессионной динамики.
Палеофитогеографическое районирование пермской Земли на основе использования
флористического подхода проводилось разными палеоботаниками. Наиболее полно
полученные результаты изложены в работе С.В. Мейена [1987]. Мейен выделяет
несколько флор,
распределенных по палеофлористическим провинциям и областям и охарактеризованных
определенными наборами наиболее типичных таксонов высших растений, преимущественного
родового ранга. Ниже дана краткая характеристика этих флор и палеофлористических
провинций по Мейену [1987].
Мейен выделяет для пермского периода три палеофлористические области: Ангарскую
(северо-восток Евразии), Катазиатскую (Китай, Корея, Япония, Вьетнам, Лаос,
Таиланд, Индонезия, Малайзия) и Еврамерийскую (Западная и Центральная Европа,
восточная
часть Северной Америки, северная часть Африки), а также обособленное от них
Гондванское царство (Южная Америка, почти вся Африка, Индия, Австралия, Антарктида)
с высокой
степенью эндемизма таксонов, характерных исключительно для материков Гондваны.
Еврамерийская и Катазиатская флоры были приурочены к регионам с тропическим
и субтропическим климатом (экваториальный пояс). Ангарская флора рассматривается
как внетропическая бореальная, гондванская - как внетропическая нотальная.
Из таксонов,
наиболее характерных для раннепермских еврамерийских флор, Мейен указывает
Lebachia (Walchia), Ernestiodendron, Callipteris, для цехштейновой
флоры Западной Европы
- Ullmannia, Pseudovoltzia, Quadrocladus, Pseudoctenis, Sphenobaiera,
Taeniopteris, Lepidopteris, Neocalamites, Paracalamites (таксоны перечислены
по уменьшению степени присутствия в местонахождениях). Как своеобразная флора,
несколько
сходная с европейской нижнепермской флорой, Мейеном рассматривается североамериканская
флора формации Данкард. Для Катазиатской флоры характерны Lobatannularia,
Tingia,
Plagiozamites, Emplectopteris, Cathaysiopteris, для Ангарской - Rufloria,
Crassinervia, Lepeophyllum, Nephropsis, Cladostrobus, Kuznetskia, Tungussocarpus,
Sylvella, для Гондванской - Gangamopteris, Glossopteris. Флоры,
имеющие смешанный еврамерийско-ангарский состав, Мейен относил к Субангарскому
экотонному поясу, в юго-восточной части
которого присутствовали и катазиатские элементы.
Взгляды Мейена на палеофитогеографическое районирование перми позднее были
развиты его последователями [Есаулова, 1986; Дуранте, 1995].
Второй подход - физиономический - к изучению палеофитогеографии пермского периода
начал применяться только в последние годы [Ziegler, 1990; Wnuk, 1996; Красилов,
Наугольных, 2001]. Прежде чем перейти к рассмотрению картируемых единиц (биомов)
при физиономической интерпретации палеофитогеографии перми, необходимо сделать
несколько общих замечаний.
Фитогеография - наука во многом опирающаяся на статистику. Чем больше местонахождений
растительных остатков (= единичных или элементарных проб растительности) вовлечено
в анализ, тем более детально можно восстановить палеофитогеографические границы
и статус выделяемых единиц - провинций, областей или царств, биомов, и тем
корректнее в итоге будут общие выводы.
Однако нельзя сказать, что данные по флорам пермского периода очень благоприятны
для палеофитогеографического анализа. Местонахождения пермских флор распределены
по поверхности Земного шара очень неравномерно, что, скорее всего, было обусловлено
как физико-географическими, так и климатическими особенностями перми.
Так, например, основное количество местонахождений растительных остатков ассельского
и сакмарского возраста приурочено к районам Земного шара, относившихся к пермским
приэкваториальным зонам или значительно удаленным от этих зон регионам Гондваны
и Ангариды. Непосредственно севернее и южнее экваториального пояса и приэкваториальных
районов располагаются пустые пространства, в которых растительные остатки не
обнаружены. Флоры намечающихся областей, кратко рассмотренных в начале главы,
существенно отличаются друг от друга по общему таксономическому составу, но
где именно проходили границы между областями сказать очень трудно. В этой ситуации
особенно важным становится поиск и изучение немногочисленных флор по периферии
экваториального пояса. Значение таких периферийных флор для решения палеофитогеографических
проблем становится ключевым. Для сакмарского времени это, например, флора Нигера
переходного гондвано-еврамерийского типа или нижнекосьинская флора, обнаруженная
в сакмарских отложениях разреза по р. Кожим (Печорское Приуралье), ангарского
типа с еврамерийскими элементами.
В качестве второго примера можно рассмотреть имеющиеся данные для палеофитогеографического
районирования растительного покрова в казанском веке. В еврамерийской приэкваториальной
зоне местонахождений растительных остатков этого возраста почти нет. Это, видимо,
было связано с нараставшей во второй половине пермского периода аридизацией.
В приэкваториальной зоне располагался обширный аридный пояс. Вместе с тем появилось
огромное количество местонахождений растительных остатков на Русской платформе
и Приуралье. Снова возникает проблема репрезентативности: поскольку у нас нет
почти никаких сведений о флорах этого времени в приэкваториальной зоне Пангеи,
корректно нанести фитогеографические границы в этом случае нельзя. Основное
внимание при определении расположения границ должно быть сосредоточено на единичных
периферийных
флорах, таких, например, как флора Унайзах (Unayzah, поздний мургаб) [Broutin
et al., 1995], содержащая катазиатские
и еврамерийские элементы и имеющей, таким образом, смешанный характер.
Таким образом, при фитогеографическом анализе особенно большое значение приобретают
флоры, располагающиеся на периферии намечающихся областей или провинций и имеющих
переходный или смешанный таксономический состав. При составлении карт, предлагаемых
в настоящей работе, были использованы не столько данные о флорах как таковых
и их таксономическом составе, как это принято в традиционной палеофитогеографии,
сколько данные по типам растительности, биомам, с учетом сведений о формах
роста и экологической ординации высших растений. Представители наиболее типичных
высших
растений пермского периода, могущих служить палеофитогеографическими маркерами,
показаны на фототаблицах I-XVI (см. вклейку IV).
Использование физиономических критериев при палеофитогеографическом районировании
было предложено В.А. Красиловым [1979; 1985]. Биом как единица палеофитогеографического
районирования впервые был использован A.M. Зиглером [Ziegler, 1990] и позднее
был применен и в других работах по фитогеографии палеозоя [Wnuk, 1996; Красилов,
Наугольных, 2001; Naugolnykh, 2000].
10.1. Смешанные флоры пермского периода
Смешанная флора (mixed flora) - флора, включающая в свой
таксономический состав элементы двух или более фитогеографических провинций.
Анализ смешанных флор
представляется целесообразным начать с западной части Евразии.
Самая западная и, вместе с тем, одна из самых хорошо изученных смешанных флор
- раннепермская (предположительно, артинская или кунгурская) флора Иберийского
полуострова (Испания), известная из нескольких местонахождений. Ее также называют
флорой Гвадалканала. Эта флора известна благодаря циклу публикаций проф. Ж.
Брутэна, который в 1986 г. опубликовал монографию, включающую описания как
макроостатков
растений, так и миоспор из Гвадалканала [Broutin, 1986]. Выводы, к которым
пришел Брутэн, заключаются в следующем: 1) на общем фоне широкого развития
верхнекаменноугольных
(стефанских) и нижнепермских элементов во флоре Гвадалканала полностью отсутствуют
представители родов Alethopteris и Odontopteris (особенно
важно отсутствие вида О. minor, очень типичного для стефанских отложений
Европы; 2) отмечаются исключительно редкие находки реликтовых каменноугольных
птеридоспермов, относящихся к родам Callipteridium, Neuropteris и Linopteris;
3) во флоре присутствует
большое количество вальхиевых хвойных Walchia (= Lebachia), Ernestiodendron
и Feysia,
а также ксерофитных птеридоспермов - каллиптерид; 4) значительное количество
гинкгофитов Ginkgoites sp., Ginkgophytopsis aff. kidstonii (Seward)
Hoeg, Ginkgophyllum
boureaui Broutin, Psygmophyllum spp., Pelourdea sp., Rhipidopsis
baetica Broutin
(= Baiera baetica Broutin), Rhipidopsis spp.;
5) присутствие катазиатских элементов Protoblechnum wongii, Psygmophyllum
multipartitum, Fascipteris cf. robusta, Lobatannularia spp.;
6) присутствие ангарских элементов Phynadaeopteris anthriscifolia (Goepp.)
Radcz., Koretrophyllites crassinervis Broutin (эндемичный гвадалканалский
вид ангарского
рода), Annulina sp., Entsovia sp., Rhipidopsis cf.
ginkgoides Schmalh. Брутэн также отметил сходную гетерогенность споро-пыльцевых
комплексов из тех же отложений:
отсутствие или исключительную редкость спор Punctatisporites, Lycospora и
Thymnospora, характерных для верхнекаменноугольных отложений Европы;
присутствие многочисленной
пыльцы, типичной для европейского ротлигенда: Potonieisporites, Protohaploxypinus,
Gardenasporites: присутствие экзотических ангарских, гондванских и катазиатских
элементов.
В 1997 г. автор во время поездки во Францию, благодаря любезности проф. Ж.
Брутэна, имел возможность ознакомиться в лаборатории палеоботаники и палинологии
Университета
Пьера и Марии Кюри, Париж VI (Laboratoire de Paleobotanique et Palynologique,
Universite Pierre et Marie Curie, Paris VI) с прекрасной коллекцией растительных
остатков из Гвадалканала, в том числе и с типовыми материалами. Некоторые из
авторских наблюдений резюмируются ниже:
1. Высока вероятность принадлежности всех гинкгоподобных листьев из флоры Гвадалканала
одному естественному (= ботаническому) виду; старшим из предполагаемых синонимов
является Baiera baetica Broutin 1974, но относить этот вид, скорее всего, следует
к роду Ginkgophyllum Saporta. В основание листа всех гвадалканалских гинкгофитов
входит несколько (обычно 5-6) жилок, что несвойственно настоящим гинкговым,
всегда имеющим две базальные жилки.
2. Листья, сходные с Entsovia, имеют попарно сближенные дорзальные желобки
в основании, но в верхней части листа они расходятся и становятся сходными
с желобками
Rufloria.
3. Экземпляр папоротника, определенный как Prynadaeopteris anthriscifolia,
скорее всего к этому виду не относится. Он обнаруживает существенно большее
сходство
со стерильными перьями некоторых видов Oligocarpia.
Помимо гвадалканалской флоры на Иберийском полуострове известно еще одно очень
необычное местонахождение растительных остатков - Пена Сагра (Pena Sagra),
располагающееся в северо-восточной Испании, в Кантабрийских горах. В этом местонахождении
найдены
вайи Supaia, практически неотличимые от типичных северо-американских
представителей этого рода [Gand et al, 1997]. Возраст местонахождения определяется
как артинско-кунгурский.
Следующей смешанной флорой, располагающейся к юго-востоку от Гвадалканала является
раннепермская флора Центрального Марокко [El Vartiti et al., 1986; Aassoumi,
1994]. В этой флоре наряду с богатым комплексом типично еврамерийских растений
присутствуют катазиатские, ангарские и гондванские элементы. Еврамерийские
таксоны: Annularia spp., Asterophyllites cf. dumasii Zeller, Calamites cf.
cistii Brongn.,
C. gigas Brongn., C. suckowii Brongn., Pecopteris cf. aspidioides Sternberg,
P. cf. densifolia Goepp., Sphenopteris germanica Weiss, Callipteridium
marginatum Aassoumi, Neuropteris sp., Odontopteris gimmii Remy, Taeniopteris
abnormis Gutbier, Taeniopteris spp., Peltaspermum sp., Lodevia
nicklesii (Zeiller) Haubold
et Kerp,
Rhachiphyllum schenkii (Heyer) Haubold et Kerp, Rhachiphyllum sp., Dichophyllum cf. flabellifera Haubold
et Kerp, Autunia (al. Callipteris) conferta (Stemb.)
Kerp, Ginkgophyllum spp., Sphenobaiera digitata Florin, Sphenobaiera sp., Cordaites cf. angulosostriatus Grand'Eury, C. cf. principalis Germar, Cordaites sp.,
Poacordaites sp., Walchia piniformis Sternb., Ernestiodendron
filiciforme (Sternb.) Florin,
Otovicia (al. Walchia) hypnoides (Brongn.) Kerp et al., Culmitzscia
parvifolia (Florin) Clement-Westerhof, C. speciosa (Florin) Clement-Westerhof, C.
laxifolia (Florin) Clement-Westerhof, Feysia minutifolia Broutin
et Kerp, Cordaicarpus sp., Gomphostrobus
bifidus (Gein.) Zeiller et Potonie, Dicranophyllum spp.
Катазиатские таксоны: Annularia cf. hunanensis Gu et Zhi, Pecopteris cf. chihliensis Stockmans
et Mathieu, P. cf. lativenosa Halle, Sphenopteris cf. pseudogermanica Halle,
Protoblechnum cf. wongii Halle. Гондванские таксоны: Glossopteris
communis Feistmantel, Gangamopteris sp. Ангарские таксоны: Mostotchkia sp.
Сходство флоры Центрального Марокко и Гвадалканала очевидно. Разница заключается
лишь в большем количестве ангарских элементов во флоре Гвадалканала и в присутствии
листьев Glossopteris communis во флоре Центрального Марокко. Вместе с тем марокканская
флора имеет несколько
более
древний - ассельский или сакмарский возраст.
Автор имел возможность ознакомиться с типовой коллекцией растительных остатков
из Марокко в лаборатории палеоботаники и палинологии Университета Париж VI.
В коллекции, помимо перечисленных выше таксонов, присутствуют древесины с септированной
сердцевиной, ассоциирующие с вальхиевыми Feysia и другими кониферофитами
типа
Dicranophyllum-Mostochkia. Кроме этого, в той же коллекции есть фрагменты
крупных листьев, сходных с Psygmophyllum expansum, a также дланевидно
рассеченные листья типа Mauerites confertus Zalessky и спорофиллы
лепидофитов, сходные с Sadovnikovia belemnoides Naug.
В Марокко известна еще одна нижнепермская флора из района Демнат (Demnate,
Высокий Атлас). Эта флора типично еврамерийская и содержит в своем составе
виды, характерные
для позднекаменноугольных (стефанских) флор Европы, представленные в основном
гидрофильными элементами, а также ксерофитные виды, типичные для европейского
отэна [Doubinger, Roy-Dias, 1986]. Общий состав флоры: Pecopteris densifolia (Goep.)
Zeiller, P.
affinis Brongn., P. polypodioides (Sternb.) Doubinger,
Asterophyllites longifoilus (Sternb.) Brongn., Catamites cistii Brongn., Annularia
stellata (Schloth.)
Brongn., Sphenophyllum cf. oblongifoium Germar et Kaulfus, Odontopteris
dufresnoyi (Brongn.) Zeiller, O. cf. brardii Brongn., O.
subcrenulata (Rost), Aulacopteris sp. (побег одонтоптероидного
птеридосперма), Dichophyllum flabelliferum (Weiss) Haubold et Kerp, Autunia
conferta (Sternb.) Kerp. Walchia piniformis Sternb.,
Culmitzschia (Lebachia) speciosa (Florin) Clement-Westerhof, C.
laxifolia (Florin)
Clement-Westerhof, Otovicia hypnoides (Brongn.) Kerp et al., Walchia (?
Ernestiodendron) cf. germanica Florin, Walchianthus sp., Samaropsis sp., Dicranophyllum cf.
gallicum Grand'Eury [Doubinger, Roy-Dias, 1986] (список составлен с учетом
изменившейся
номенклатуры названий растений).
Пермская смешанная флора Нигера, известная из местонахождения Арлит [Arlit;
Rouvre, 1988], содержит следующие виды: еврамерийские Autunia (al.
Callipteris)
conferta,
Walchia piniformis, кордаиты еврамерийского облика Pachycordaites
lingulatus Grand'Eury, Dorycordaites sp., Artisia sp.,
семена Nucellangium sp., Cornucarpus acutus Lindley et Hutton, Samarospermum sp.,
Mucrospermum ovatum Rouvre, Phyllocarpus bifidus Rouvre, Pedicellocarpus
elongatus Rouvre, гондванские (?) Samaropsis surange Lele, S.
emarginatus Surange et
Lele, Cordaicarpus cf.
ovatus Lele, C. furcatus Surange et Lele. Таксономический список содержит как
типично еврамерийские, так и предположительно гондванские формы.
Самое известное из южно-европейских местонахождений смешанных флор - Хазро
в Турции, где присутствуют еврамерийские, гондванские и катазиатские элементы
[Wagner,
1962; Archangelsky, Wagner, 1983]. Флора Хазро включает в свой состав катазиатские
виды Bicoemplectopteris hallei Asama, Fascipteris hallei (Kawasaki)
Gu et Zhi,
Pseudomariopteris hallei (Stockmans et Mathieu) Wagner, Cladophlebis
tenuicostata (Halle) Archangelsky et Wagner, Pecopteris (Asterotheca?) calcarata Gu
et Zhi, Dizeugotheca sp., Sphenophyllum cf. koboense Kobatake, Lobatannularia
heianensis (Kodara) Kawasaki. Как еврамерийско-катазиатские элементы
могут рассматриваться пекоптериды Pecopteris nitida Wagner и P.
pirae Wagner.
Во флоре присутствуют
гондванские элементы: Glossopteris anatolica Archangelsky et Wagner; до работы
[Archangelsky, Wagner, 1983] этот вид был определен как Glossopteris cf. stricta Bunbury, a также простоперистые вайи Botrychiopsis sp. Листья G.
anatolica довольно обычны во флоре Хазро, встречаются в угленосных фациях, но никогда
не доминируют.
Возраст флоры Хазро по фораминиферм [Fontaine et al., 1980] определяется как
джульфинский, т.е. татарский по общей стратиграфической шкале.
Несколько исключительно интересных смешанных флор были открыты в последние
десятилетия в Малой Азии, в центральной части Аравийского полуострова и на
его восточном
побережье. Смешанная флора с доминирующими еврамерийскими, а также включающая
в свой состав катазиатские и гондванские элементы, была описана из центральной
части Саудовской Аравии (местонахождение Унайзах). Возраст местонахождения
первоначально определялся как раннепермский [Еl-Khayal et al., 1980], но позднее
был уточнен
и повышен до позднепермского. Флора содержит следующие таксоны: Annularia
mucronata Shenk, Lobatannularia cf. heianensis Kodaira, Pecopteris
phegopteroides Feistmantel,
P. cf. wongii Halle, Gemellitheca saudica Wagner,
Hill, El-Khayal, Qasimia
schyfsmae (Lemoigne) Hill, Wagner et El-Khayal, Cladophlebis aff. roylei Arber,
Fascipteris
hallei (Kawasaki) Gu et Zhi, Taeniopteris sp., Marattiopsis sp.
(=? Qasimia
schyfsmae), Gangamopteris sp. (это растение было ошибочно определено
И. Лемуанем как
Zamiopteris (?) sp.), Dadoxylon spp., Cordaites sp. [Lemoigne, 1981; El-Khayal,
Wagner, 1983; Hill et al., 1985; Wagner et al., 1985].
Одним из самых ярких событий последнего десятилетия было открытие смешанной
флоры Хариф (Ghariff) в Омане, на Аравийском полуострове [Broutin et al., 1995].
Возраст
флоры по подстилающим и перекрывающим слоям с морской фауной определяют как
кубергандинский (кунгуро-уфимский?). Из слоев с макромерными растительными
остатками были выделены
споры и пыльца, включающие в свой состав споры Laevigatosporites vulgaris,
Leiotriletes spp., Retusotriletes cf. simplex, Calamospora sp., Verrucosisporites sp.,
Cyclogranisporites arenosus, Polypodiisporites cf. mutabilis, Cristatisporites sp., Brazilea sp.,
одномешковую пыльцу Florinites pellucidus, F. eremus, Potonieisporites
novicus, Playfordiapora crenulata, Scheuringipollenites ovatus, неребристую
(небороздчатую,
лишенную тэний) двумешковую пыльцу Vitreisporites pallidus, Vesicaspora
schemeli, Falcisporites sp., Gardenasporites sp., Cedripotes
prisons, Platysaccus sp.,
ребристую двумешковую пыльцу Photohaploxypinus amplus, P. dvinensis, P.
hartii, P. limpidus, P. microcorpus, P. pennulatus, P. perfectus, Lunatisporites
cf.
pellucidus, Lueckisporites singhii, L. virkkiae, Corisaccites alutas, Striatoabieites
multistriatus, S. richteri, Striatopodocarpites cancellatus, S. fusus, Gutullapollenites
neohannonicus, Weylandites lucifer. В палиноспектре доминирует пыльца
Protohaploxipinus, сходная с Р. microcorpus - видом, характерным для перми
Гондваны и отмечаемом
в разрезе Соляного Кряжа [Salt Range, Пакистан].
Среди макроостатков растений во флоре Хариф присутствуют гондванские элементы
Glossopteris cf. occidentalis White, Glossopteris spp.,
женские фруктификации типа Arberia, еврамерийские хвойные Otovicia (al. Walchia) hypnoides (Brongn.)
Kerp et al., каламостахиевые Galamites gigas Brongn., гинкгофиты Baieroxylon
implexum (Zimmermann) Greguss, a также катазиатские цикадофиты Tingia aff.
hamaguchii.
В Средней Азии существует одно очень интересное местонахождение растительных
остатков предположительно кунгурского возраста - Сары-Тайпан. Таксономический
состав флоры: Vojnovskya stankevichii Sixtel, Nephropsis (Sulcinephropsis) spp., Lepeophyllum sp., Crassinervia spp., Rufloria (Alatoruflorid)
cf. derzavini (Neub.)
S. Meyen, Cordaites sp., Phyllotheca sp., Walchia sp.,
Samaropsis karamasarensis Sixtel (= S. rectialata Neub.).
Кроме перечисленных выше таксонов, но значительно реже встречаются фрагменты
древесин с септированной сердцевиной, семена Samaropsis (?) complanata Naug.,
Demetrocarpus sp. [Сикстель и др., 1975; Наугольных,
2001а],
а также определения автора по коллекциям Н.Н. Каландадзе, хранящимся в Геологическом
музее им. Вернадского и ГИН РАН, г. Москва.
Интерес, который вызывает к себе эта флора, в первую очередь определяется тем,
что она довольно удалена от региона с развитием типично ангарской растительности
(Сибирь), но, вместе с тем, имеет отличимо ангарский облик. Это может указывать
на существенно более сложный характер границы между фитохориями в этом регионе,
чем это было принято считать ранее [Меуеп, 1987; Мейен, 1987; Дуранте, 1995].
Типичная смешанная флора с доминированием катазиатских элементов характеризует
местонахождение Га'ара в западном Иране [Ga'ara; Ctyroky, 1973]. Возраст местонахождения
определяется как позднепермский. В состав флоры входят Lobatannularia heianensis (Kodaira)
Kawasaki, Plagiozamites
oblongifolius Halle, Pecopteris sp., Taeniopteris sp., Protoblechnum sp.
В песчаных прослоях той же формации встречены декортицированные стволы еврамерийских
лепидофитов, определенные как Sigillaria sp. (предположительно,
Subsigillaria, форма сохранности Syringiodendron). Тафономическая
избирательность, с которой распределены в разрезе катазиатские и еврамерийские
элементы, указывает
на то, что первоначально они входили в разные растительные ассоциации - околоводную
с лепидофитами и характерную для более возвышенных частей побережья ассоциацию
с катазиатскими элементами. Из располагающихся в том же регионе местонахождений
Афганистана С.В. Мейен определял еврамерийские (Pecopteris еврамерийского облика),
катазиатские (Lobatannularia) и ангарские (Phylladoderma) элементы [Мейен,
1981; Меуеп, 2002]. Смешение еврамерийско-гондванских элементов с примесью
ангарских
элементов отмечается в верхнепермских споро-пыльцевых комплексах Соляного Кряжа
в Пакистане.
Совершенно неожиданным и даже сенсационным является обнаружение смешанных раннепермских
гондвано-катазиатских флор на Тибете и в Кашмире [Li et al., 1991 и др.]. Долгое
время считалось общепризнанным, что Индия и остальная часть блоков, входящих
в настоящее время в Евразийский континент, в позднем палеозое были разделены,
что и обуславливало столь большой контраст между пермскими флорами Индии и
Китая.
Однако открытие смешанных катазиатско-гондванских флор в Кашмире и на Тибете
указывает на то, что уже в раннепермскую эпоху существовали широкие возможности
для миграционных обменов между индийской (гондван-ской) флорой и флорами Катазии.
Список таксонов, установленных по макроостаткам растений включает следующие
катазиатские элементы: Sphenophyllum thonii var. minor, S.
thonii var. archangelskyi,
S. thonii var. waltoni, Rajahia mamalensis, Pecopteris mamalensis,
Lobatannularia ensifolia, L. sinensis var. curvifolia, L lingulata,
а также гондванские элементы: Trizygia speciosa, Gangamopteris kashmirensis,
Glossopteris communis, G. cordatifolia,
G. dingriensis, G. intermittens, G. longicaulis, G. hishatbaghensis, G. taeniopteraides,
Ginkgophyllum (al. Psygmophyllum sensu Arber) haydenii, G.
sahnii [Singh
et al., 1982; Pant et al., 1984; Li et al., 1991; Chandra, Keqin, 1997].
В Таиланде помимо типично катазиатских флористических комплексов с гигантоптеридами
[Asama et al., 1968] известна флора Phetchabun, включающая в свой состав как
катазиатские, так и гондванские элементы. Общий список таксонов флоры Фетчабун,
имеющей возраст от кунгурского до казанского: Bowmanites sp., Sphenophyllum
trapaefolium Stockmans et Mathieu, S. phetchabunensis Asama, Alethopteris
thailandica Kon'no,
Glossopteris cf. angustifolia Brongn., Palaeovittaria
parvifolia Kon'no, Taeniopteris
hallei Kawasaki, T. cf. serrulata Kawasaki, T. thailandica Asama,
T. konnoi Asama, T. nystroemii Halle, T. hallei Kawasaki, T.
iwaii Asama, Taeniopteris sp., Cordaites principalis (Germar)
Gein., Poacordaites phetchabunensis Kon'no, P. linearis Grand'Eury, Pecopteris sp., Protoblechnum
wongii Halle, Psygmophyllum
komalarjunii Asama, Psygmophyllum (?) sp., Samaropsis sp. [Bunopas, 1981].
Одна из наиболее интересных и хорошо документированных смешанных флор описана
из западной части Новой Гвинеи [Irian Jaya; Rigby, 1996]. Флора включает в
свой состав как гондванские, так и катазиатские элементы, причем остатки индекс-таксонов
Glossopteris и Gigantonoclea иногда даже соседствуют на одних и тех же образцах.
Общий список флоры: Trizygia speciosa, Pecopteris spp., Ptychocarpus sp., Cladophlebis sp., Fascipteris
aidunae Rigby, Glossopteris irianii Rigby, Glossopteris sp.,
Vertebraria indica, Gigantonoclea irianii Rigby, Koraua
hartonoi Rigby.
Помимо перечисленных выше смешанных флор, следует назвать еще две экзотические
флоры, включающие в свой состав нетипичные элементы. Первая из них - флора
Мидткап [Midtkap]; [Wagner et al., 1982; 1999; 2002] происходит из верхнепермских
отложений северо-восточной Гренландии и содержит ангарский флористический комплекс.
Первоначально опубликованный таксономический список включал следующие таксоны:
Rhipidopsis spp., Pecopteris (? Asterotheca) helenaeana Zalessky,
Prynadaeopteris venusta Radczenko, Sphenophyllum cf. biarmicum Zalessky (скорее
всего, это
определение было сделано ошибочно, по небольшому фрагментарному экземпляру
Rhipidopsis),
Cordaites cf. sylovaensis (Neuburg) S. Meyen, Bardella sp. [Wagner et al.,
1982]. Однако после дополнительных сборов и ревизии старых определений, этот
список
был исправлен и значительно дополнен [Wagner et al., 1989; 2002]. В этих сообщениях
был предложен новый список таксонов флоры Мидткап: Viatscheslaviophyllum sp.,
Signacularia (?) sp., Prynadaeopteris anthriscifolia (Goepp.) Radczenko, Pecopteris sp. nov., Rhipidopsis
panii Chow, R. ginkgoides Schmalh., Pursongia sp., Cordaites
candalepensis (Zalessky) Radchenko, Lepeophyllum adzvense Neuburg, Sphenophyllum cf. verticillatum (Schlotheim). Кроме этого, в том же местонахождении встречены
крупные фрагменты окаменелой древесины.
Вторая флора с ангарскими элементами была кратко охарактеризована на материале
из конгломератов Mount Dall (Аляска). Флористический комплекс содержит Pecopteris
arborescens Schloth., P. hemitelioides Brongn., P. unita Brongn., циклоптериоидные
перышки (? Cyclopteris, Angaropteridium, Cardioneura spp.), Zamiopteris sp.
[Mamay, Reed, 1986]. К сожалению, сведения об этой флоре, возможно, сочетающей
еврамерийские
и ангарские элементы, пока еще очень отрывочны.
Весьма своеобразные смешанные флоры, содержащие в своем составе ангарские (Rufloria,
Zamiopteris, Comia, Psygmophyllum, Gaussia, Nephropsis), катазиатские
(листья членистостебельных типа Lobatannularia, а также Taeniopteris, близкие
катазиатским
видам рода), гондванские (Glossopteris, Gangamopteris) элементы, известны из
верхов нижней и верхней перми Дальнего Востока [Зимина, 1977].
После обзора основных смешанных пермских флор необходимо подробнее остановиться
на приуральских флорах нижней перми и, в частности, рассмотреть кунгурскую
флору Среднего и Южного Приуралья. В состав этой флоры входят компоненты, являющиеся,
с одной стороны, мигрантами из экваториального пояса (каламостахиевые Calamites
gigas Brongn., Annularia spp., Calamostachys dumasii (Zeil.)
Jongm. (? = Sachyogyrus
multifarius Zal.),
бовманитовые Sphenophyllum aff. thonii Mahr, Bowmanites
biarmensis Naug., близкий
еврамерийским представителям рода, мараттиевые папоротники Corsinopteris (= "Orthotheca"),
Ptychocarpus, Discopteris, редкие невроптериды Neuropteris sp.
и тригонокарповые Demetrocarpus
limbatus Naug.). Очень близки еврамерийским
таксонам кунгурские
вальхиевые хвойные (Kungurodendron sharovii S. Meyen (= Walchia
appressa s.
Zal.), Walchia bardaeana Zal.) и пельтаспермовые Peltaspermum
retensorium (Zal.) Naug. et Kerp, Rhachiphyllum spp. С другой
стороны, во флоре присутствуют автохтонно
развивавшиеся в Приуралье группы высших растений, некоторые из которых мигрировали
позднее в центральные районы Ангариды, как, например, псигмофиллоиды: Psygmophyllum из
группы видов, близких P.
expansum (Brongn.) Schimper, а другие оставались
на всем протяжении своего существования эндемичными для западной окраины Субангарской
области [Наугольных, 1998а, 19986]. Наиболее типичные эндемики кунгурской
флоры: Phyllotheca campanularis Zal. emend. Naug., Phyllotheca
stenophylloides Zal.,
Corynophyllites setiformis Zal., Equisetina magnivaginata Zal., Biarmopteris
pulchra Zal., Alternopsis stricra Naug., Bardia mauerii Zal., Kerpia
macroloba Naug., Bardospermum rigidum S. Meyen, Craspedosperma
bardaeanum Zal. и некоторые
другие. Кроме этого, в составе кунгурской флоры имеется ряд таксонов, мигрировавших
в Приуралье из Ангариды ("ангарские кордаиты" Rufloriapapillosa Gluch.,
R. laevis Glush., R. meyenii Gluch., R. unica Gluch., R.
olenekensis Gluch.,
R. recta (Neub.) S. Meyen, R. derzavinii (Neub.) Gluch., R.
lanata Gluch.,
войновскиевые) и космополитные таксоны, в основном представленные формальными
родами (Sphenopteris,
Pecopteris и др.).
Легко заметить, что многие крупные таксоны растений, представленные в кунгурской
флоре Приуралья, группируются в общности, объединяемые одним и тем же фитогеографическим
статусом. Так, например, все известные к настоящему времени представители мараттиевых
папоротников из кунгурского яруса Приуралья принадлежат родам, характерным
для экваториальных флор Еврамерийской фитохории. Войновскиевые представлены
почти
исключительно ангарскими родами и даже видами. Кунгурские членистостебельные
включают в свой состав эндемичные, ангарские и еврамерийские роды; при более
детальном рассмотрении выясняется, что каламостахиевые из кунгурского яруса
Приуралья (Calamites gigas Brongn., Annularia spp.) - мигранты
из экваториальных флор,
а
черновиевые (Equisetinostachys, Paracalamitina, Annulina и некоторые
приуральские представители родов Paracalamites и Phyllotheca) являются типично
ангарскими
таксонами.
Необходимо подчеркнуть, что перечисленные группы обладали существенно разными
экологическими амплитудами. Черновиевые явно были гигро- и гидрофитами, войновскиевые
и хвойные произрастали преимущественно на возвышенных участках побережья и
т.д. Очевидно, они принадлежали различным исходным растительным ассоциациям
и только
в виде "танатоценотической" смеси оказались погребенными в одних
и тех же местонахождениях. Относительное обилие этих групп, обладавших хорошо
обособленными
таксоценами (черновиевые, возможно, мараттиевые, хвойные семейства Walchiaceae),
меняется в довольно широких пределах от местонахождения к местонахождению,
что может указывать на различную удаленность исходных ассоциаций от формировавшихся
танатоценозов.
10.2. Механизм миграционных обменов и возникновения
феномена смешанных флор
Очевидно, что при образовании смешанных флор миграционные
процессы и встройка чуждых элементов-мигрантов в новые для них растительные
сообщества была облегчена
тем, что растения-мигранты либо занимали еще свободные ниши, либо реализовывали
экологические лицензии за счет физиологических преимуществ. Это очень важно
для общего объяснения феномена смешанных флор, поскольку трудно допустить простое
механистическое смешение гетерогенных компонентов в одних и тех же растительных
ассоциациях. Само проникновение, катистемия новых таксонов в неаутентичную
им
растительность указывает, с одной стороны, на экологическую природу мигрантов
(они были ценофобами или ценотолерантами) и, с другой стороны, на степень
организации сообществ, в которые происходило внедрение (они были далеки от
климаксного состояния).
10.3. Континентальные биомы
пермского периода
На палеобиогеографических картах, (рис. 69-72) автором показано
расположение основных континентальных биомов пермского периода. При характеристике
биомов
указывались основные (доминирующие) группы
растений, предположительно, создававших наибольшее проективное покрытие. Там,
где это представлялось наиболее хорошо обоснованным, приведены данные о формах
роста этих растений, их жизненных формах (по Раункиеру) и экологических предпочтениях.
Распределение биомов по континентам было обусловлено, прежде всего, климатическими
условиями произрастания высших растений.
Картируемые биомы:
1. Обедненная растительность тундрового
или лесотундрового типа; доминирующие группы растений: в Северном полушарии
- войновскиевые ("ангарские
кордаиты";
Vojnovskyales в субплакорных и плакорных ассоциациях), членистостебельные
семейства Tchernoviaceae (в околоводных растительных ассоциациях), листостебельные
мхи;
в Южном полушарии - представители порядка Dictyopteridales (sensu McLoughlin,
1992), "глоссоптериды"). Предельные формы роста - кустарники. Жизненные
формы - хамефиты и гемикриптофиты.
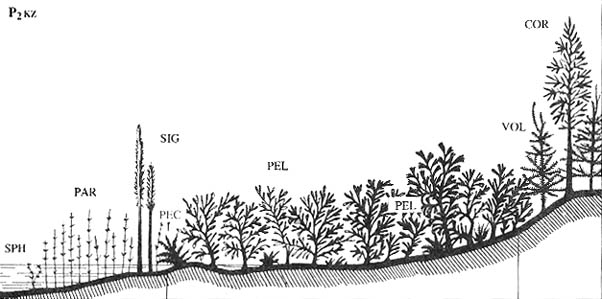
2. Типичная ангарская листопадная растительность (пермский аналог современных
бореальных лесов, "кордаитовая тайга"); доминирующая группа растений
- войновскиевые ("ангарские кордаиты"; Vojnovskyales) и членистостебельные
семейства Tchernoviaceae. Предельные формы роста - деревья. Основные жизненные
формы - фанерофиты. Растительность этого типа была приурочена к северному влажноумеренному
климатическому поясу [Наугольных, 1998а; Красилов, Наугольных, 2001].
3. Тропическая и приэкваториальная засушливые зоны, растительность средиземноморского
типа; доминирующие группы растений - пельтаспермовые птеридоспермы (Peltaspermales)
и хвойные семейства Walchiaceae (в раннепермской растительности) и семейства
Voltziaceae (в позднепермской растительности). Предельные формы роста - невысокие
деревья (хвойные) и кустарники (пельтаспермовые). Жизненные формы - хамефиты,
гемикриптофиты, фанерофиты.
4. Экваториальная и тропическая вечнозеленая растительность, сходная с современными
дождевыми лесами; доминирующие группы растений: реликтовые лепидодендровые
(лепидокарповые) Lehidodendron oculis-felis), птеридоспермы порядков
Trigonocarpales и Gigantonomiales
("гигантоптериды"), причем некоторые из последних по формам роста
были лианоподобными. О влажном и жарком климате этого биома говорит широкое
распространение
угленосных отложений в нижнепермских отложениях Катазии, где пермская растительность
с чертами экваториальных и тропических дождевых лесов была развита особенно
широко. Предельные формы роста - деревья. Доминирующие жизненные
формы - фанерофиты.
5. Ксерофитная растительность бессточных котловин; доминирующая группа растений
- пельтаспермовые птеридоспермы (Peltaspermales). Этот биом характерен для
внутренних районов приэкваториальной части Пангеи. Растительность этого биома
была образована
птеридоспермами родов Supaia, Zeilleropteris, Delnortea и близкими
формами [Mammay et al., 1988], а также, в меньшей степени, хвойными семейства
Walchiaceae
и сфенофиллами
Sphenophyllum gilmorei White. Остатки сфенофиллов, явно предпочитавших немногочисленные
в этом регионе экотопы с повышенной влажностью, встречаются в ин-ситном положении
в красноцветных отложениях русел временных водотоков, промоин и каналов прорыва,
затоплявшихся в короткие периоды сезонных дождей.
6. Нотальная листопадная растительность южного умеренного пояса; доминирующая
группа растений: представители порядка Dictyopteridales ("глоссоптериды").
Предельные формы роста - деревья (в северной части), кустарники (в южной части).
Жизненные формы - хамефиты, гемикриптофиты, фанерофиты. Растительность этого
типа была приурочена к южному влажно-умеренному климатическому поясу.
На картах также показаны зоны смешения растительности средиземноморского типа
(еврамерийский тип флор) и растительности южного умеренного или холодно-умеренного
биома (гондванский тип флор), а также зоны углеобразования.
10.4. Катены
Состояние палеофиоценологических исследований палеозойской
растительности, т.е. исследований, в фокусе которых находится не таксономия
и систематика ископаемых
растений, а растительные сообщества геологического прошлого, трудно признать
удовлетворительным. Сложилось две основных тенденции, одна из которых заключается
в грубом калькировании геоботанических методик и использовании их для классификации
ориктоценозов, типов местонахождений, а отнюдь не растительных сообществ.
Вторая
тенденция заключается в реконструкции палеофитоценозов путем типологических
экстраполяции с современной растительностью, которые далеко не всегда правомерны.
Очень большие трудности существуют в определении и корректном использовании
понятий, которые во многих случаях толкуются произвольно. Поэтому следует пояснить
основные
термины, используемые ниже.
Центральным и наиболее важным понятием при реконструировании древних растительных
сообществ является понятие катены [Красилов, 1972]. Катена - последовательность
растительных группировок (ассоциаций), сменяющих друг друга в зависимости от
удаления от бассейна или водотока по мере изменения градиента высотности. В
современной геоботанической литературе катены также называются сменно-доминантными
комплексами.
Ассоциация - звено катены, растительное сообщество или относительно гомогенная
пространственная отдельность растительности низшего ранга.
Далее следует подробнее остановиться на методах восстановления катениальных
последовательностей. Их можно разделить на две основные группы: визуальные и аналитические. Визуальные
методы применимы в исключительных случаях: когда мы имеем ин-ситные или гипавтохтонные
местонахождения растительных остатков и можем непосредственно в разрезе наблюдать
изменение характера флористических комплексов или типов палеопочв по латерали.
Из аналитических методов наиболее важными представляются следующие: 1) определение
соотношения и характера сохранности остатков растений различной таксономической
принадлежности; 2) распределение остатков растений по фациям, а также использование
литологических признаков-маркеров обстановок осадконакопления. В качестве примера
можно сказать, что укорененные, ин-ситные остатки членистостебельных в нижнепермских
отложениях Приуралья приурочены, как правило, к песчаникам с элементами косой
слоистости, что указывает на произрастание этих растений в мелководной прибрежной
части водоема, в зоне миграции знаков ряби [Осадочные породы..., 1990]. Остатки
вячеславий в уфимских отложениях Приуралья встречаются в различных фациях
- от прибрежно-морских до континентальных, что говорит в пользу эврибионтности
этого растения; 3) важным элементом аналитических методов реконструкции катениальных
последовательностей является анализ морфологических и анатомических особенностей
растений. Существуют экологически важные признаки-маркеры, которые действуют
практически безотказно. Так, например, наличие воздушных корней-пневматофоров
указывает на то,
что растение произрастало в гиперувлажненной среде. Развитие шипов, редукция
листьев и преобразование ветвей в мясистые листоподобные органы, приспособленные
для запасания влаги, говорит об аридных условиях, о существовании растения
в условиях стресса при недостатке влаги. Толстые кутикулы, многочисленные папиллы
и трихомы, глубоко погруженные устьица могут рассматриваться как ксероморфные
признаки. Однако необходимо подчеркнуть, что все перечисленные аналитические
методы должны рассматриваться в комплексе, поскольку в некоторых случаях один
и тот же признак может вызываться разными причинами. Ксероморфные признаки,
например,
могут присутствовать у растений, испытывающих физиологическую сухость, которая
не обязательно прямо коррелируется с атмосферными условиями [Busche et al.,
1978; Кегр, 1990].
Ниже рассматривается характер катениальных последовательностей, реконструированных
на основе использования разных методик для разных интервалов пермского периода.
К сожалению, данные о катениальных цепях ограничены пока только северным полушарием,
и не все катены восстановлены с одинаковой степенью детальности. Поэтому автору
пришлось выбрать только те из реконструкций катен, которые наиболее достоверны
и непротиворечивы. Для полноты обзора кратко рассмотрены позднекаменноугольные
и раннетриасовые катены.
Начать следует с позднекаменноугольно-раннепермской катены. К настоящему времени
опубликованы данные только для экваториального пояса и примыкающим к нему районам,
охватывающим значительную часть Северной Америки, Европу, Донбасс и, возможно,
Южное Приуралье.
Изучению и описанию катен позднего карбона и ранней перми этого региона было
посвящено несколько работ, из которых можно выделить две [Cridland, Morris,
1963; Щеголев, 1991], наиболее адекватно отражающие достигнутые результаты.
В реконструкциях катен, которые предлагаются этими авторами, много общего,
несмотря на то, что они были построены на разном исходном материале и с использованием
различных методик.
Самая нижняя часть катены, представляющая собой околоводное сообщество, состоит
из гидро- и гигрофитов (поименно-болотная растительная ассоциация, по А.К.
Щеголеву): членистостебельных - преимущественно каламитов нескольких видов.
Непосредственно
в прибрежной части водоемов произрастали
клинолистники - сфенофиллы. Другими важными элементами этой растительной ассоциации
являлись лепидофиты, в позднем карбоне и ранней перми представленные, в основном,
видом Subsigillaria brardii (Brongn.) Weiss. Кустарниковый синузий составляли
гигрофильные папоротники.
Второе звено катены было представлено ассоциацией кордаитов и мезофильных птеридоспермов,
преимущественно, тригонокарповых (Neuropteris spp., Odontopteris
subcrenulata (Rost) Zeill.). Эта ассоциация существовала в условиях умеренно увлажненных
или полусухих склонов.
Третье звено катены состояло из хвойных семейства Walchiaceae: Walchia
piniformis Sloth., Ermstiodendron filiciforme (Sloth.), Otovicia
hypnoides (Brongn.)
Kerp et al. и близких форм. Кроме хвойных, в этой же ассоциации присутствовали
и ксерофильные
птеридоспермы, в основном, пельтаспермовые - каллиптериды Autunia conferta (Sternb.) Kerp, Rhachiphyllum,
Dichophyllym и другие близкие растения.
Такой тип организации катены был характерен для большей части экваториального
пояса в течение позднего карбона и первой половины ранней перми, т.е. ассельского
и сакмарского веков. Судя по пока еще немногочисленным данным о гжельских,
ассельских и сакмарских флорах Южного Урала (разрезы Айдаралаш и Кондуровка),
которые включают
в свой состав побеги каламостахиевых, семена тригонокарповых птеридоспермов
и вальхиевые хвойные еврамерийского облика [Владимирович, 1986; Naugolnykh,
1999;
2000], очень вероятно, что подобные катениальные последовательности были характерны
и для самой ранней перми Южного Приуралья. Ангарская растительность ранней
перми, скорее всего, была организована иначе, но никаких данных о строении
катен раннепермских
фитоценозов Ангариды мы пока не имеем.
Конечно, следует признать, что растительные сообщества в геологическом прошлом,
так же, как и сейчас, были упорядочены нелинейно. На схеме, предложенной Г.
Керпом [Kerp, 1996] для нижней перми земли Саар-Пфальц (Германия), хорошо видно,
что
в организации растительности присутствует элемент мозаичности, определявшийся
характером рельефа и, как следствие, типом субстрата и характером его увлажненности.
На относительно крутом склоне развивается единственное катениальное звено с
доминирующими хвойными; на умеренно крутом слабо увлажненном склоне - две ассоциации,
относительно
возвышенных местообитаний с доминирующими хвойными и ассоциация мезофильных
птеридоспермов с многочисленными Odontopteris lingulata. В условиях наличия
полувлажной или обводненной пресными водами поймы добавляется еще одно
катениальное
звено - с доминирующими субсигилляриями.
К сожалению, данные о более молодых пермских флорах Европы и Америки, т.е.
экваториального пояса и прилегающих регионов, пока еще очень ограничены, поэтому
как были организованы
постсакмарские катены в Еврамерике остается неизвестным. Вместе с тем, именно
с этого интервала, т.е. начиная с артинского яруса, богатые флористические
комплексы начинают встречаться в Приуралье, в Субангарском экотонном поясе.
Флоры эти сохраняют в своем составе многочисленные еврамерийские элементы,
однако вместе с тем, содержат много эндемиков и растений, характерных для позднего
палеозоя
Ангариды.
Автор уже обращался к вопросу реконструкции катены артинского века в Среднем
Приуралье [Наугольных, 1997] по материалам из местонахождения Шайдуры, бассейн
р. Сылва. Это один из тех редких случаев, когда единственное местонахождение
дает возможность высказать предположение о характере катениальной цепочки.
Значительное отличие таксономического состава растений, обнаруженных в нетипичной
фации -
глубоководных аргиллитах с аммоноидеями, встречающимися в виде прослоев среди
преимущественно прибрежноводных или флишоидных отложений с преобладающими в
разрезе песчаниками, позволило предположить, что растительная ассоциация с
многочисленными
хвойными Coenoxylon, Tylodendron, Walchia, соответствующая шайдурскому
ориктоценозу, произрастала на большем удалении от бассейна, в более высоких,
предгорных районах
Палеоурала (рис. 73). Остатки растений этой ассоциации попадали в глубоководные
осадки в шлейфах пролювиальных выносов и мутьевых потоках. Нижнее околоводное
звено артинской катены состояло из членистостебельных в основном представленных
видом Paracalamites decoratus (Eichw.) Zal.
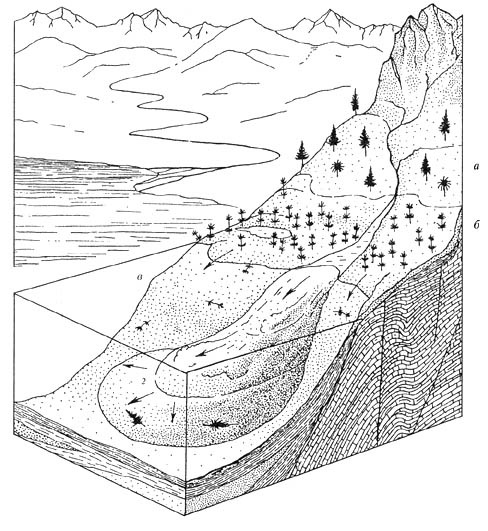
Рис. 73. Артинский век, катена Среднего Приуралья
а - ксерофильная ассоциация с доминированием хвойных семейства Walchiaceae; б
— гигрофильная ассоциация с доминированием членистостебельных Paracalamites;
в — зона формирования тафоценозов типичного верхнеартинского флористического
комплекса; г - зона формирования тафоценозов атипичного верхнеартинского
флористического комплекса
|
Далее перейдем к катениальной последовательности растительности
кунгурского века в Приуралье. Кунгурская катена была реконструирована с помощью
аналитических
методов. Во-первых, было выявлено изменение характера доминирующих элементов
от местонахождения к местонахождению; во-вторых, использовались признаки-маркеры,
указывающие на экологические предпочтения охарактеризованных ими растений;
в
третьих использовалась типологическая экстраполяция
данных по нижнепермским катенам Еврамерики, поскольку между растительностью
европейского ротлигенда и Приуральского кунгура наблюдается существенное сходство
и преемственность.
Нижнее, первое звено катены было представлено ассоциацией гидро- и гигрофитов,
образующих околоводную растительность (рис. 74). Гидрофитами, судя по характеру
сохранности, были клинолистники Sphenophyllum biarmicum Zal., (рис.
75, в). Этот вид был гетерофильным, что весьма характерно для растений, имеющих
полупогруженные
в воду побеги. В околоводной ассоциации произрастали черновиевые, представленные,
преимущественно видами Phyllotheca campanularis Zal. emend. Naug., P.
stenophylloides Zal. Гигрофиты были представлены паракаламитами и паракаламитинами
(Paracalamites frigidus Neub., P. striatus (Schmalh.) Zal., P.
kutorgae (Gein.)
Zal., Paracalamitina ignatievii Naug., Paracalamitina
sp.).
В этой же ассоциации в качестве травянистого синузия присутствовали гидрофильные
папоротники. Некоторые из этих папоротников обладали гидатодами - специальными
комплексами клеток в листьях, обеспечивающих выделение из растения капельно-жидкой
воды и солей. Гидатоды характерны для растений, произрастающих в увлажненных
условиях.
Следующее звено катены соответствует мезофильной ассоциации, образованной в
основном пельтаспермовыми птеридоспермами (каллиптеридами и псигмофиллоидами)
и ма-раттиевыми
папоротниками. Многие из этих
растений имели сильно дифференцированные эпидермы верхней и нижней сторон листа.
Верхняя эпидерма с мощными клеточными стенками несла толстую кутикулу и была
почти лишена устьиц; нижняя эпидерма была менее развитой с очень тонкой кутикулой.
Многочисленные устьица на нижней стороне листа были снабжены папиллами для
защиты от излишнего испарения влаги. Характер облиствения птеридоспермов этого
катениального
звена сильно менялся от сезона: в периоды кратковременного увлажнения эти растения
образовывали вайи с хорошо развитыми и обособленными крупными перышками (каллиптериды,
рис. 76) или большие глубоко рассеченные листья с широкими лопастями (псигмофиллоиды).
В более сухих условиях для уменьшения транспирации листовая пластинка уменьшалась
в размерах [Наугольных, 1999].
Третье звено катены было образовано ассоциацией с доминирующими вальхиевыми
хвойными с игловидными листьями, мощной кутинизацией устьичных аппаратов и
многочисленными
трихомными образованиями. Эти растения были хорошо адаптированы к аридным или
даже гипераридным условиям. Кроме хвойных, в этой же ассоциации произрастали "ангарские
кордаиты" (рис. 77; о таксономическом статусе группы
см. [Krassilov, Burago, 1981; Красилов, 1989; Наугольных, 2001 а]. Они были
представлены в основном родом Rufloria (этим же растениям принадлежали катафиллы
Lepeophyllum и Crassinervia, брактеи Nephropsis (Sulcinephropsis) и женские генеративные
органы Gaussia (= Paravojnovskya), Pholidophyllum (Scirostrobus) и Vojnovskya.
Наличие
у растений этой группы катафиллов, или почечных чешуи, защищавших молодые побеги
от перегрева и некомпенсируемых потерь влаги в аридных условиях, а также дорзальных
желобков, в которых располагались устьица, защищенные от высыхания хорошо развитыми
трихомами и папиллами, указывают на то, что по меньшей мере некоторые "ангарские
кордаиты" произрастали в условиях водного стресса и повышенной сухости.
Наличие в основании листьев кунгурских Rufloria характерных рубцов, образованных
отделительным слоем, указывает на то, что материнские растения были листопадными.
Опадение листвы, скорее всего, происходило в засушливые сезоны.
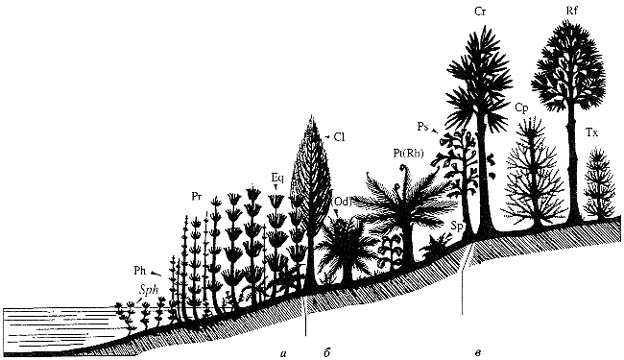
Рис. 74. Кунгурский век, катена Среднего и Южного Приуралья
Ассоциации: а - гидро- и гигрофильная, б - мезофильная, в - ксерофильная.
Растения: Sph - Sphenophyllum biarmicum Zal., Ph - Phyllotheca
campanularis Zal. emend. Naug., Pr - Paracalamitina ignatievii Naug., Paracalamites spp.,
Eq - Equisetina magnivaginata Zal., Cl - Calamites gigas Brongn.,
Pt (Od) - Pt(Rh) - пельтаспермовые птеридоспермы (каллиптериды) нескольких
типов, Ps - Psygmophyllum expansum (Brongn.) Schimp., Sp - Sphenopteris spp.,
Cr - Cordaites singularis (Neub.) S. Meyen, C. ex gr. lineatus (Neub.)
S. Meyen,
Cp - Cyparissidium (al. Walchia) appressum (Zal.) S.
Meyen, Rf - Rufloria spp., Tx - Taxodiella (al. Walchia) bardaeana (zal.) S. Meyen
|
|
 |
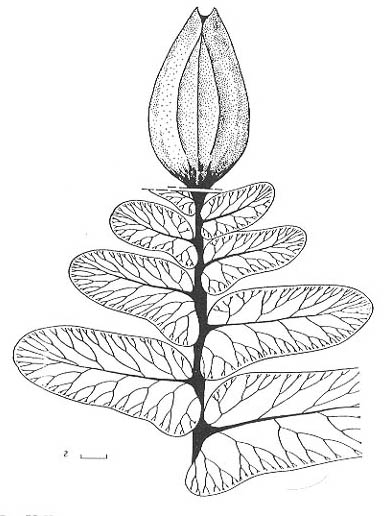 |
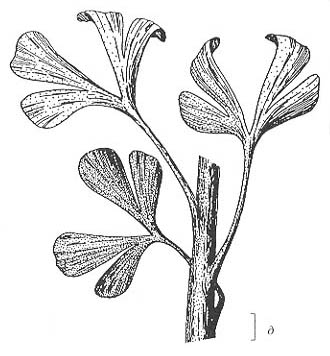
Рис.
75. Некоторые из наиболее типичных растений кунгурского (в-д) и уфимского-казанского
(а, б) веков, Приуралье
а, б — членистостебельное Paracalamitina striata (Schmalhausen) Zalessky
emend. Naug. (Naugolnykh, 2002), a - терминальная фертильная зона, б - форма
кроны; в - бовманитовое Sphenophyllum biarmicum Zalessky emend. Naug.;
г - тригонокарповый птеридосперм Demetrocarpus limbatus Naugolnykh;
д - гинкгофит Kerpia macroloba Naugolnykh
Длина масштабной линейки - 2 мм (а, г); 1 см (в, д); 5 см (б) |
 |
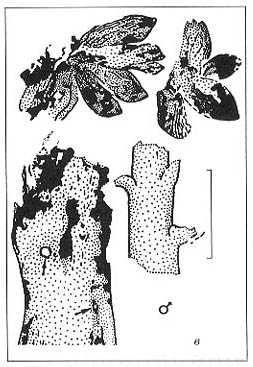 |
|
 |
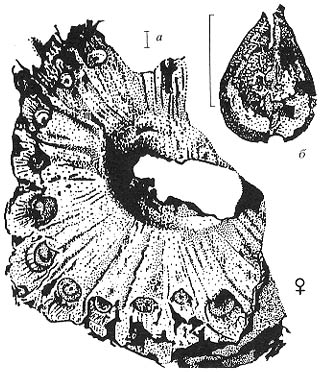
Рис. 76. Некоторые из наиболее типичных растений
кунгурского века, Приуралье; различные органы, возможно, принадлежавшие
одному растению,
описанному как Peltaspermum retensorium (Zalessky) Naugolnykh et Kerp (Naugolnykh, Kerp,
1996)
a - семеносный диск Peltaspermum sp.; б - семезачаток Cordaicarpus
sp.; в - микроспороклады
(андрофоры), состоявшие из розетковидных синангиев и несущих их побегов, Permotheca
disparts (Zalessky) Naugolnykh (Наугольных, 19986; см., также: Krassilov
et al., 1999); д - фрагмент вайи Rhachiphyllum (al. Callipteris) retensorium(Zalessky)
Naugolnykh в ассоциативной связи с семезачатком Cordaicarpus sp. (этот
же семезачаток показан на рис. (г) с большим увеличением; е - Rhachiphyllum (al.
Callipteris) retensorium (Zalessky) Naugolnykh, общий облик ювенильной вайи с
хорошо развитым окрылением рахиса и намечающимся перевершиниванием апекса.
Длина масштабной линейки - 1 мм (а); 2,5 мм (г); 5 мм (в); 1 см (б, д, е) |
|
Рис. 77. Вегетативный побег (д),
генеративные органы (в, г) и семена (а, б) "ангарских кордаитов" (порядок
Vojnovskyales)
а, б - Samaropsis danilovii Suchov; в - Vojnovskya sp.; г - V.
paradoxa Neub. [г - по: Мейен, 1982]; д - побег войновскиевого Cordaites
lancifolius Schmalhausen
[no: Schmalhausen, 1887, Taf. VII, 1; p. Сылва, пос. Суксун]. Экземпляры: а -
3737/274; б - 3737/201; в - 3773(11)/349(93); г - 3039/194. Местонахождения:
Чекарда-1, ел. 10 (а, б); Юлаево (в); Халмерьюсское угольное
месторождение, шахта 3 (г). Кунгурский ярус, верхнекунгурский подъярус, иренский
горизонт, кошелевская свита (а - в, д); уфимский ярус, нижнеуфимский подъярус,
Соликамский горизонт, интинская свита (е).
Длина масштабной линейки - 2 мм |
 |
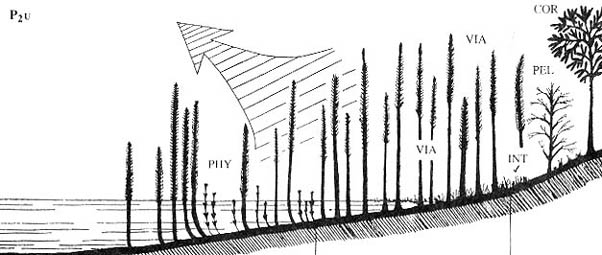 |
Рис. 78. Уфимский и казанский века, катены Русской платформы и Приуралья
Растения: Phy - Phyllotheca sp., Via - Viatcheslavia vorcutensis Zal.,
Int - Intia variabilis Neub., Pel - пельтаспермовые птеридоспермы,
порядок Peltaspermales,
семейство Cardiolepidaceae, Cor - Cordaites sp., Rufloria sp.,
Sph - Sphenophyllum stouckenbergii Schmalh., Par - Paracalamites
sp., Paracalamitina striata (Schmalh.) Zal., Sig - Signacularia
noinskii Zal., Pec
- Pecopteris spp.; пельтаспермовые птеридоспермы, семейство Cardiolepidaceae
(Phylladoderma) и Peltaspermaceae (Pursongia), Vol - хвойные семейства Voltziaceae
(Pseudovoltzia) |
Следующая катениальная последовательность - катена первой
половины уфимского века (рис. 78, нижняя часть), а именно - соликамского времени,
коренным образом
отличается
от катен, рассмотренных выше. Во-первых, таксономическое разнообразие
соликамской растительности неизмеримо беднее, чем предшествующей ей кунгурской.
Это всего три-четыре вида, из которых один принадлежит листостебельным мхам
- Intia variabilis Neub. Доминируют лепидофиты Viatcheslavia vorcutensis Zal.
Остатки
этого плауновидного растения встречаются в известняках прибрежно-морского генезиса
(местонахождения на реках Колве и Вишере), плитчатых мергелях со значительной
примесью глинистой фракции, образовавшихся, вероятно, в условиях мелководной
спокойной лагуны (местонахождения Полазна, Хохловка, Чумкасский карьер), а
также в неяснослоистых песчаниках, иногда обладающих
косой слоистостью (местонахождения Алебастрово и Ляды). Такой очень широкий
спектр
фаций от прибрежно-морских до аллювиальных указывает на то, что V. vorcutensis
произрастала на различном удалении от бассейна и занимала несколько биотопов.
Можно сказать, что раннеуфимская катена редуцировалась до одного звена, представленного
ассоциацией V. vorcutensis-Intia variabilis. Смена исключительно богатой
кунгурской растительности на однообразную растительность с доминирующей V.
vorcutensis очевидно маркирует региональный экологический кризис [Наугольных,
1992; 2001г.; 2001д], вызванный климатической флуктуацией (аридизацией).
В Приуралье на одном стратиграфическом уровне с остатками вячеславий часто
встречаются слепки кристаллов гипса и глиптоморфозы из каменной соли, что
указывает на аридный
климат с редкими относительно влажными сезонами. С последними, скорее всего,
были связаны репродуктивные циклы V. vorcutensis, на побегах которой
часто видны зоны со сближенными "листовыми подушками", которые на
самом деле являются следами формирования фертильных зон. Во время образования
органов
размножения
высшие растения почти перестают расти: все силы растения уходят на генеративные
процессы. Видимо с этим связано уменьшение расстояния между соседними основаниями
спорофиллов. То, что это именно рубцы спорофиллов косвенно подтверждается их
размером: они в два-три раза больше нормальных
листовых подушек. Основания спорофиллов V. vorcutensis также превышают по размерам
основания филлоидов в два-три раза (см. [Наугольных, 19986], рис. 9, C-G; Наугольных,
2001д]).
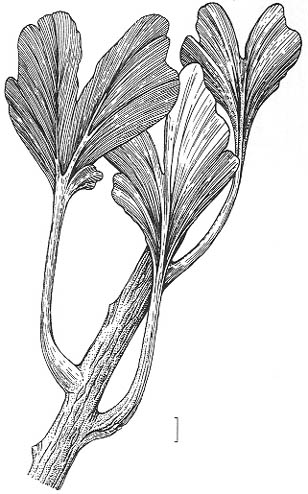
Рис. 79. Гинкгофит Kerpia belebeica Naugolnykh, казанский век, Приуралье
Длина масштабной линейки - 1 см |
Переходя к катениальным цепочкам второй половины уфимского и казанского веков,
необходимо отметить, что представления о их строении сформировались в основном
благодаря работам В.ГГ. Владимирович [1984] и Н.К. Есауловой [1986], придававших
большое значение в своих исследованиях тафономическим наблюдениям.
Как и в кунгурском ярусе, в казанских отложениях Приуралья и Русской платформы
встречаются ориктоценозы нескольких типов: с доминирующими клинолистниками
(соответствующим водным сообществам), с доминирующими паракаламитами и филлотеками
(полуводная
гигрофильная растительность побережий), с многочисленными пельтаспермовыми
(мезофильное сообщество). Кроме этих типов ориктоценозов, известны местонахождения
с большим
количеством хвойных, например Китяк с Pseudovoltzia и Слуда с остатками Quadrocladus [Meyen, 1997].
Казанская катена в целом напоминала кунгурскую (рис. 78, верхняя часть). Таким
образом, к этому времени уже восстановилась первоначальная структура растительности,
нарушенная во время раннеуфимского кризиса.
Нижнее звено казанской катены состояло из сообщества гидро- и гигрофитов, представленных
клинолистниками Sphenophyllum stouckenbergii Schmalh., паракаламитами
и аракаламитинами (о статусе рода Paracalamitina Zalessky см. [Naugolnykh,
2002]) и филлотеками.
Вероятно, к этой же растительной ассоциации тяготели Signacularia noinskii Zal.
- лепидофиты, являвшиеся прямыми потомками уфимских вячеславий. Одним из отличий
Signacularia от Viatcheslavia являются более широкие филлоиды,
характерные для первого рода, что может указывать на более влажный климат,
существовавший
в казанском
веке.
Второе звено катены соответствовало ассоциации мезофильных птеридоспермов:
кардиолепидиевых Phylladoderma meridionalis S. Meyen, P. septata Bogov, P.
volgensis Esaul. и
близких видов, а также каллиптерид Rhachiphyllum wangenheimii (Fischer)
Naug.,
Compsopteris salicifolius (Fischer) Naug., C. olgae Naug.
[Наугольных, 1999; 20016] и гинкгофитов (рис. 79). В этой ассоциации произрастали
мезофильные
папоротники Acitheca gigantea Esaul., Todites gracilis Esaul., Oligocarpia
camiensis Esaul.
[Есаулова, 1996].
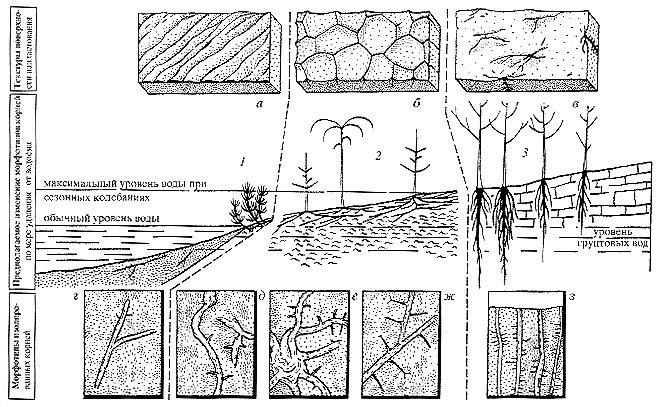 Рис. 80. Татарский век, катена Русской платформы и Южного Приуралья;
растительные ассоциации Рис. 80. Татарский век, катена Русской платформы и Южного Приуралья;
растительные ассоциации
1 - гидрофильная, 2 - гигрофильная и эфемероидов, 3 - ксерофильная ассоциация
фреатофитов. Текстуры поверхностей напластования: а - со следами течения и волноприбойными
знаками, б - с трещинами усыхания, в - поверхности карбонатных панцирей с проксимальными
частями корневых систем; г - з - различные морфотипы корневых систем и их распределение
по фациям
|
Третье звено катены было образовано ассоциацией ксерофильных
растений, состоящей из хвойных семейства Voltziaceae потомков раннепермских
вальхиевых (Pseudovoltzia
cornuta S. Meyen). Вероятно, в этой же ассоциации произрастали казанские
войновскиевые
Cordaites candalepensis и близкие формы.
Судя по общему сходству таксономического состава [Gomankov, 1995], раннетатарская
катена была организована сходным образом.
Реконструкция позднетатарской катены, первый вариант которой уже был опубликован
ранее [Арефьев, Наугольных, 1998; Наугольных, 2001 в], была получена при комплексном
использовании визуальных и аналитических методов. Очень важным и даже определяющим
было обнаружение остатков корневых систем высших растений in situ.
To, что изученные структуры действительно являются корнями высших растений,
доказывается
их анатомическим
строением. После тщательного анализа фактического материала выяснилось, что
остатки корней, принадлежащим различным морфотипам, закономерно распределены
по вмещающим
их фациям (рис. 80). Для отложений мелководий и прибрежных низменностей с чередовавшимися
субаквальными-субаэральными условиями были характерны горизонтально ориентированные
сложноветвящиеся тонкоосные корни;
для горизонтов каличей - карбонатных панцирей, образовавшихся в аридной обстановке
в субаэральных условиях, характерны хорошо развитые вертикальные, глубоко проникающие
в грунт корни типа R. sukhonensis Aref. et Naug. Эти две области можно рассматривать
как звенья единой катены, первое из которых соответствует ассоциации эфемероидов,
растений с сезонной вегетацией, экологическая стратегия которых сводилась к
развитию густой приповерхностной системы корней, наиболее эффективно поглощающей
влагу
в периоды сезонных увлажнений. Второе звено катены состояло из ассоциации фреатофитов,
экологическая стратегия которых была направлена на питание грунтовыми водами.

Рис. 81. Вегетативный побег пельтаспермового птеридосперма
рода Peltaspermopsis Gomankov emend. Poort et Kerp с листьями типа
Pursongia Zalessky, татарский
век, Приуралье и Русская платформа. Длина масштабной линейки - 10 см |
Ассоциация эфемероидов скорее всего была населена птеридофитами Dvinopteridium
edemskii Zal. и хвощами Paracalamites, Paracalamitina, Phyllotheca,
? Neocalamites. Возможно, этой же ассоциации принадлежали поздне-татарские
пельтаспермовые Peltaspermopsis с листьями Pursongia-Tatarina (рис.
81).
Ассоциация фреатофитов, по всей видимости, состояла из хвойных Geinitzia.
Эта модель катениальной последовательности для позднетатарской растительности
была предложена на материале по палеопочвам из бассейна рек Сухоны и Малой
Северной Двины [Арефьев, Наугольных, 1998]. Однако дальнейшие исследования
показали, что
сходные закономерности в распределении па-леопочв и остатков корневых систем
наблюдаются и в других регионах Русской платформы и Приуралья: в Поволжье и
Прикамье (разрезы Шереметьевка и Монастырский овраг), Оренбуржья (разрезы Кошка
и Вязовка),
в бассейне р. Вятка (разрез Котельнич) [Наугольных, 2001в] и Ветлуги. Это указывает
на то, что общая ландшафтная ситуация в позднетатарское время была довольно
однообразной по всей Русской платформе.
На разрезе у г. Котельнич следует остановиться подробнее. Котельнический разрез
является довольно протяженным и, несмотря на кажущуюся литологическую однородность,
представлен различными фациями. Основную массу пород составляют плотные красные
или малиновые глины, иногда переходящие в более плотные аргиллиты. В нижней
части разреза присутствуют светлоокрашенные прослои. В средней и верхней частях
разреза
встречаются песчаные линзы руслового и эолового генезиса (рис. 82, 83).
В светлых прослоях в нижней части котельнического разреза встречаются вертикально
ориентированные субцилиндрические конкреции. Эти конкреции уходят вглубь вмещающего
их глинистого матрикса, постепенно уменьшаясь в диаметре и иногда давая боковые
ответвления. После того, как несколько конкреций были разбиты, выяснилось,
что внутри они полые. Полость обычно заполнена красноватым глинистым материалом,
отличающимся по цвету от вмещающего конкреции матрикса (рис. 84).
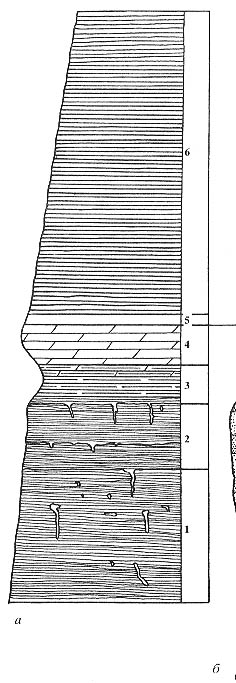 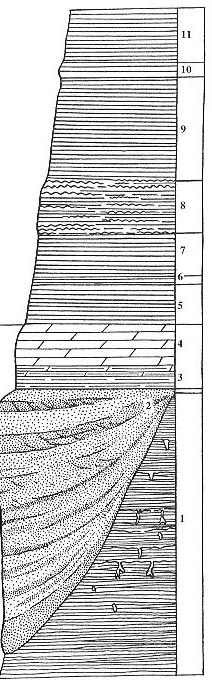
Рис. 82. Разрез Котельнич, стратиграфические колонки
а - правый берег р. Вятки, обнажение "Под кладбищем", первый овраг
с ручьем в 100 м ниже по течению от кладбища, г. Котельнич (в 150 м ниже по
течению от этой части разреза найдены прослои с хорошо сохранившимися Radicites
aff. sukhonensis Aref. et Naug.; 6 - правый берег р. Вятки, обнажение "Элеватор",
первая песчаная линза ниже г. Котельнич.
Длина масштабной линейки -1м |
Очень сходная сохранность наблюдается у остатков корневых систем Radicites
sukhonensis из бассейна реки Сухона. Природу R. sukhonensis можно
считать доказанной [Арефьев,
Наугольных, 1998]. Очень вероятно, что вертикальные трубчатые структуры в нижней
части Котельнического разреза также являются остатками корней, облеченных в
карбонатную рубашку (зачаточный калич), а сероцветные прослои в красных глинах
и аргиллитах,
или, по меньшей мере, некоторые из них - уровни с палеопочвами.
Собственно вмещающие отложения можно рассматривать как прибрежные и мелководные
отложения, накапливавшиеся в условиях периодического осушения. К этим слоям
приурочены остатки парейазавров Deltavjatia vjatkensis (Hartmann-Weinberg)
и других рептилий
из Котельнича, многие из которых обладали адаптациями к наземному образу жизни.
Парейазавры, по всей видимости, паслись в зарослях растений, образовывавших
вертикально ориентированные корни Radicites aff. sukhonensis, листьями
и побегами которых
они могли питаться. В следующую фазу образования местонахождения береговая
линия значительно придвинулась к области формировавшегося тафоценоза. Растения
с мощными
вертикальными корнями исчезли. Вместо них развилась прибрежная растительность,
соответствующая первому звену катены - ассоциации гигрофитов и эфемероидов.
Эта ассоциация была населена споровыми растениями и пельтаспермовыми птеридоспермами.
Многочисленные членистостебельные Phyllotheca, Paracalamites и Paracalamitina,
а также листья Pursongia широко встречаются в присутствующих на этом уровне
линзах серых аргиллитов. Песчаные линзы с элементами косой слоистости и изредка
встречающимися костями
земноводных образовались очевидно уже в условиях относительно постоянного водотока
(рис. 85).
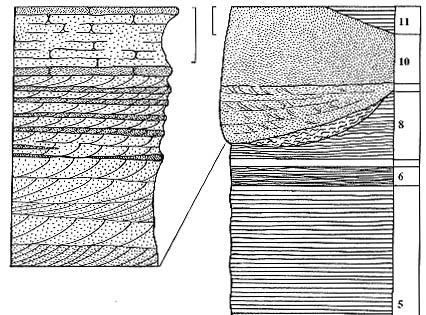
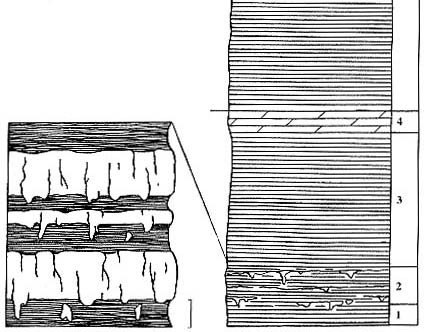
Рис. 83. Разрез Котельнич, стратиграфические колонки
Правый берег р. Вятки, обнажение "Шестаки", вторая песчаная линза
ниже
по течению от г. Котельнич, в 1 км выше по течению от ЛЭП.
Длина масштабной линейки -1м (правая колонка), 10 см (левые колонки) |
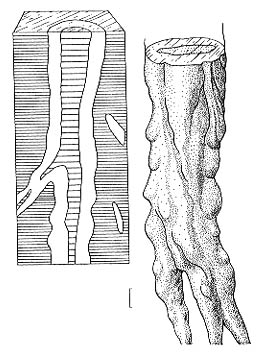 Рис. 84. Radicites aff. sukhonensis Aref. et Naug., остатки
корней. Местонахождение Котельнич, татарский ярус Длина масштабной линейки
- 1 см Рис. 84. Radicites aff. sukhonensis Aref. et Naug., остатки
корней. Местонахождение Котельнич, татарский ярус Длина масштабной линейки
- 1 см
|
О более молодых, чем верхнетатарские, пермских отложениях,
встречающихся в пределах Русской платформы, существуют только предварительные
данные. В местонахождении
Недуброво, располагающемся в Вологодской области, недавно был обнаружен макрофлористический
комплекс, более молодой,
чем все известные к настоящему времени местонахождения из верхнетатарского
подъяруса. Комплекс включает в свой состав листья пельтаспермовых Tatarina
lobata S. Meyen
(изображенные фрагменты также обладают большим сходством как на макро-, так
и
микроуровне с перьями последнего порядка Lepidopteris martinsii (Germar
Townrow), типичного птеридосперма европейского цехштейна), семеносные диски
Peltaspermum,
кардиолепидиевое Phylladoderma (Aequistomia) annulata S.
Meyen, хвойные Ullmannia
bronnii Goepp. и Quadrocladus solmsii (Gothan et
Nagalhard)
Schweitzer [Krassilov
et al., 1999; Lozovsky et al., 2001]. Как видно из этого списка, указанная
флора обладает значительным сходством с флорами европейского цехштейна [Weigelt,
1928;
1930; Gothan, 1907; Gothan, Nagalhard, 1922; Daber, 1960; Stoneley, 1958;
Schweitzer, 1960; 1963; 1968; Haubold, Schaumberg, 1985]. Точное строение недубровской
катены пока неизвестно. Можно предположить, что оно было близко катениальной
последовательности позднетатарского времени.
В нижнем триасе на Русской платформе, там, где обнаружены растительные остатки,
а именно в вохминском горизонте (индский ярус; еще не описанные спорофиллы
лепидофитов) и рыбинском горизонте (нижнеоленекский подъярус), флористические
комплексы очень
бедны в таксономическом отношении и содержат, как правило, единственный
вид - Pleuromeia (al. Lycomeia) rossica Neub. [Нейбург,
1960; Добрускина, 1980; 1982]. Безусловный доминант раннетриасовой растительности
Русской платформы
- лепидофит P. rossica - изучен довольно хорошо и к его морфологической
характеристике можно добавить лишь несколько частных замечаний. Однако тафономические
особенности
нахождения остатков P. rossica оставались почти неизвестными.
Автор предпринял специальные полевые работы на некоторых классических местонахождениях
P. rossica в бассейне р. Волги (разрезы в черте г. Рыбинска и в 10 км ниже
по течению на правом берегу р. Волги у сел Паршино и Тихвинское, рис. 86).
Во время
полевых наблюдений удалось установить следующие факты.
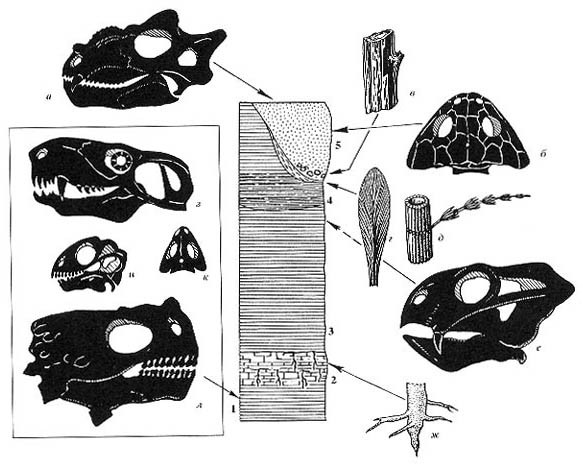 Рис.
85. Обобщенный разрез у г. Котельнич; распределение растительных остатков
и тетрапод Рис.
85. Обобщенный разрез у г. Котельнич; распределение растительных остатков
и тетрапод
а - парейазавр Deltavjatia vjatkensis (Hartmann-Weinberg), б - дромазавр
(аномодонт)
Suminia getmanovi Ivachnenko, в - тероцефал Karenites ornamentatus Tatarinov,
г - корни Radicites
aff. sukhonensis Aref. et Naug., д - дицинодонт, e - пельтаспермовый птеридосперм
Pursongia, ж — стегоцефал (Dvinosaurus)
(текст подписи полностью соответствует оригиналу в книге - прим. В. П.) |
 |
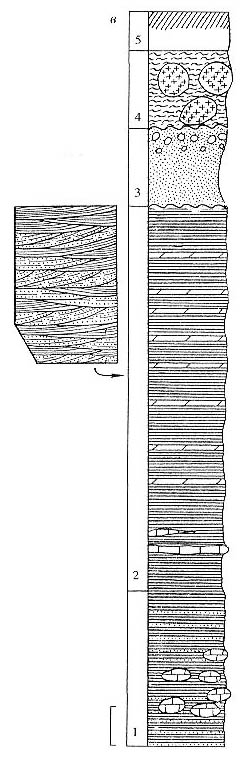 |
Рис. 86. Нижнетриасовые отложения, разрезы Паршино и Рыбинск
(р. Коровка и р. Черемуха), совместная колонка; морфология Pleuromeia (al.
Lycomeia) rossica Neub:
a - общий облик (форма роста) Р. rossica; внизу - характер строения
нижней части
побега с ризофором и корнями in situ; 6 — ризофор P. rossica с
сохранившимися корнями, указывающими на то, что растение было захоронено на месте
произрастания; в - генерализованная колонка отложений ветлужской серии у г. Рыбинск,
слева показан характер переслаивания песчано-глинистых пород ветлужской серии.
Длина масштабной линейки - 1 см (а, б), 10 м (в) |
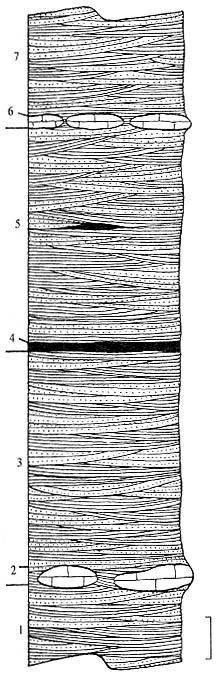 |
 |
 |
 |
Рис. 87. Разрез Паршино, детальная прорисовка части обнажения; справа
дана схема
образования гидроморфных почв с Pleuromeia rossica in situ
1 - моновидовые заросли Р. rossica; 2 - погребение растительных остатков
на месте
произрастания исходного фитоценоза, 3 - формирование тафоценоза.
Длина масштабной линейки - 10 см (для колонки), схема справа - вне масштаба |
В пределах толщи сероцветных, иногда слегка зеленоватых или
голубоватых мергелей и песчаников, располагающихся между двумя горизонтами
уплощенных карбонатных
конкреций, отмечаются несколько уровней с особенно высоким обилием остатков
P. rossica (рис. 87). Отдельные фрагменты побегов и спорофиллы рассеяны
по всей
толще в пределах нескольких метров мощности, но целые побеги с основаниями
и ризофорами встречены лишь на двух уровнях. Ризофоры нередко несут адвентивные
корешки и располагаются in situ, что указывает на автохтонность их
остатков. Судя по характеру косой слоистости, образованной при миграции знаков
ряби в
условиях
наклонной поверхности пляжа или
отмели, осадконакопление происходило в очень мелководных условиях. Очевидно
непосредственно в этой обстановке и произрастали плевромейи. Местонахождения
плевромей в ветлужском
надгоризонте распространены очень широко - от Углича до Костромы [Добрускина,
1982] и везде они носят характер монодоминантных ориктоценозов с P. rossica.
Все эти наблюдения косвенным образом указывают на ситуацию, очень сходную с
тем, что произошло в начале уфимского века: катениальная последовательность
редуцировалась
до одного звена, причем это единственное звено было представлено растительной
ассоциацией с исключительно бедным таксономическим составом. В завершение следует
сказать, что как по систематическому положению (представители порядка Isoetales),
так и по формам роста (небольшие неветвящиеся растения с терминальными стробилами
или фертильными зонами и игловидными филлоидами) раннеуфимские вячеславии
и раннетриасовые плевромейи очень близки. Также следует добавить, что монодоминантные
комплексы с Pleuromeia были распространены в нижнем триасе чрезвычайно широко:
от Европы [Fuchs et al., 1991] до Китая [Wang, 1993 и др.] и даже Австралии
[Retallack,
1980], причем во многих случаях их появление связывают с возникновением кризисных
ситуаций [Wang, 1989; 1992; 1993; 1996]. Совершенно очевидным является то,
что столь однообразный характер раннетриасовых флор указывает на экосистемный
кризис,
имевший место в наземных экосистемах в начале триаса. Это кризисное событие,
по всей видимости, было обусловлено пиком аридизации.
Таким образом, при анализе известных катениальных последовательностей на интервале
от начала пермского периода до начала триаса выявляются два события, которые
маркируются редукцией катениальных цепей и образованием своеобразных монодоминантных
сообществ лепидофитов [Наугольных, 2001 г]. Наличие этих эпизодов видимо было
связано с экосистемными кризисами (в раннеуфимское время - региональным, в
раннетриасовую эпоху - глобальным) в истории растительности и наземных биот
в целом. Кризисы
эти, по всей видимости, были связаны с аридизацией климата.
Литература
Арефьев М.П., Наугольных С.В. Изолированные корни из татарского
яруса бассейна рек Сухоны и Малой Двины // Палеонтол. журн. 1998. № 1. С. 86-99.
Владимирович В.П. Типовая казанская флора Прикамья. М., 1984. 92 с. Деп.
в ВИНИТИ. 12.06.84, № 4571-84.
Владимирович В.П. Высшие растения // Атлас характерных комплексов пермской
фауны и флоры Урала и Русской платформы. Л.: Недра, 1986. С. 32-38. (Тр.
ВСЕ-ГЕИ. Н.С.;
Т. 331).
Добрускина И.А. Стратиграфическое положение флороносных толщ триаса Евразии.
М.: Наука, 1980. 163 с. (Тр. Геол. ин-та РАН; Вып. 346).
Добрускина И.А. Триасовые флоры Евразии. М.: Наука, 1982. 182 с. (Тр. Геол.
ин-та РАН; Вып. 365).
Дуранте М.В. Реконструкция климатических изменений в позднем палеозое Ангариды
(на основе фитогеографических данных) // Стратиграфия. Геол. корреляция.
1995. Т. 3, № 2. С. 25-37.
Есаулова Н.К. Флора казанского яруса Прикамья. Казань: Изд-во Казан, ун-та.
1986. 176 с.
Есаулова Н.К. Новые виды папоротников из казанских отложений Прикамья //
Палеонтол. журн. 1996. № 4. С. 121-126.
Зимина В.Г. Флора ранней и начала поздней перми Приморья. М.: Наука, 1977.
127 с.
Игнатьев И.А., Наугольных С.В. Раннеказанская флора реки Сояны
и ее положение в ряду одновозрастных флор и растительных зон Ангариды //
Стратиграфия.
Геол. корреляция. 2001. Т. 9, № 3. С. 58-75.
Красилов В.А. Палеоэкология наземных растений: (Основные принципы и методы).
Владивосток: ДВНЦ АН СССР, 1972. 212 с.
Красилов В.А. Меловая флора Сахалина. М.: Наука, 1979. 182 с.
Красилов В.А. Меловой период: Эволюция земной коры и биосферы. М.: Наука,
1985. 240 с.
Красилов В.А. Происхождение и ранняя эволюция цветковых растений: М.: Наука,
1989. 264 с.
Красилов В.А., Наугольных С.В. Пермские биомы, экотоны и климатические
зоны // Экосистемные в истории Земли. М.: Палеонтол. ин-т РАН, 2001. Вып.
4. С.
53-63.
Мейен С.В. Следы трав индейских. М.: Мысль, 1981. 159с.
Мейен С.В. Фруктификации верхнепалеозойских кордаитантовых Ангариды //
Палеонтол. журн. 1982. № 2. С. 109-120.
Мейен С.В. Основы палеоботаники. М.: Недра, 1987. 404 с.
Наугольных С.В. Экологическая катастрофа в пермском периоде? // Природа.
1992. № 4. С. 64—66.
Наугольных С.В. Остатки растений анатомической сохранности из
урминской свиты (артинский ярус) Среднего Урала // Чтения памяти Сергея
Викторовича
Мейена:
Сб. тез. Геол. ин-т РАН, 1997. С. 14-17.
Наугольных С.В. Субангарская растительность ранней перми: Палеофитоценозы
и климат // Верхнепермские стратотипы Поволжья. Казань: Мастер Лайн, 1998а.
С.
127-128.
Наугольных С.В. Флора кунгурского яруса Среднего Приуралья. М.: Геос, 19986.
201 с. (Тр. Геол. ин-та РАН; Вып. 509).
Наугольных С.В. Новый вид рода Compsopteris Zalessky из верхней перми Пермского
Прикамья // Палеонтол. журн. 1999. № 6. С. 80-91.
Наугольных С.В. Морфология и систематическое положение представителей порядка
Vojnovskyales // Там же. 2001а. № 5. С. 95-105.
Наугольных С.В. Растительные остатки пермского возраста из коллекции Ф.Ф.
Вангенгейма фон Квалена в Геологическом музее им. В.И. Вернадского // VM-
Novitates. Новости из Геол. музея им. В.И. Вернадского. 20016. № 6. С.
1-32.
Наугольных С.В. Тайна Котельнича разгадана // Природа. 2001в. № 7. С. 20-27.
Наугольных С.В. Экологические кризисы в истории Ангарской растительности
// Биоразнообразие в истории Земли: Тез. докл. XLVII сес. Палеонтол. о-ва.
СПб.:
Изд-во СПбГУ, 2001г.
С. 66-68.
Наугольных С.В. Viatcheslavia vorcutensis Zalessky (плауновидные): Морфология,
систематика, палеоэкология // Палеонтол. журн. 2001д. № 2. С. 97-102.
Нейбург М.Ф. Pleuromeia Corda из нижнетриасовых отложений Русской платформы.
М.: Изд-во АН СССР, 1960. С. 65-90. (Тр. Геол. ин-та АН СССР; Вып. 43).
Осадочные породы: Состав, текстуры, типы разрезов). Новосибирск: Наука,
1990. 269 с. (Тр. ин-та геологии и геофизики СО АН СССР; Вып. 761).
Сикстель Т.А., Савицкая Л.И., Искандарходжаев Т.А. Растения среднего
карбона, верхнего карбона и нижней перми Ферганы. Ташкент: Фан, 1975. С.
77-143.
Щеголев А.К. Плауновидные и клинолисты позднего карбона. Киев: Наук, думка,
1991.
128 с.
Aasoumi M.H. Les paleoflores du Permien du Maroc Central: These du Dr.
de 1'Univ. P., 1994. 227 p.
Archangelsky S., Wagner R.H. Glossopteris anatolica sp. nov. from uppermost
Permian strata in south-east Turkey // Bull. Brit. Mus. (Natur. Hist.).
Geol. 1983. Vol.
37, N 3. P. 81-91.
Asama K., Jwai J., Vecraburas M., Hongnusonthi A. Permian plants from Loei,
Thailand // Geology and paleontology of Southeastern. Asia. Tokyo, 1968.
Vol. 4. P. 82-99.
Broutin J. Etude paleobotanique et palynologique du passage Carbonifere-Permien
dans le Sud-Oest de la Peninsula Iberique. P.: Ed. CNRS, 1986. 165 p.
Broutin J., Roger J., Platel J.P. et al. The Permian Pangea: Phytogeographic
implications of new paleontological discoveries in Oman (Arabian Peninsula)
// C.r. Acad. sci. Ser. Ha. 1995. Vol. 321. P. 1069-1086.
Bunopas S. Paleogeographic history of Western Thailand and adiacent parts
of South-east Asia: A plate tectonics interpretation // Geol. Surv. Pap.
Thailand.
1981. N 5: (Special issue). P. 192-197.
Busche R., Haas H., Remy R. Moglichkeiten und Grenzen der deutung Okologie
und Klima mit Hilfe disperser Kutikulen aus dem Autun des Nahe-Raumes (Pfalz)
//
Argumenta Palaeobot. 1978. Vol. 5. P. 149-160.
Chandra S., Keqin S. Evolution and comparison of the Gondwana flora and
the Cathaysia flora // Palaeobotanist. 1997. Vol. 46, N 3. P. 35-46.
Cridland A.A., Morris J.E. Taeniopteris, Walchia and Dichophyllum in
the Pennsylvanian System of Kansas // Univ. Kans. Sci. Bull. 1963. Vol.
44,
N 4. P. 71-85.
Ctyroky P. Permian flora from the Ga'ara region (Western Iraq) // Neues.
J. Geol. Palaontol. Monatsh. 1973. H. 1. S. 383-388.
Daber R. Palaeobotanische Bemerkungen zur Kupferschieferpalaeogeographik
// Geologie. 1960. Bd. 9. S. 930-934.
Doubinger J., Roy-Dias C. La paleoflore Autuno-Stephanienne de la region
de Demnate (est de Marrakech-Maroc) // Geobios. 1986. N 19, fasc. 6. P.
549-571.
El-Khayal A.A., Chaloner W.G., Hill C.R. Palaeozoic plants from Saudi Arabia
// Nature. 1980. Vol. 285, N 1. P. 33-34.
El-Khayal A.A., Wagner R.H. Upper Permian stratigraphy and megafloras
of Soudi Arabia: Palaeogeographic and climatic implications // C.r. X Congr.
Carbonifere.
Madrid, 1983. Vol. 3. P. 17-26.
El Wartiti M., Broutin J., Freytet P. Premiere decouvertes
paleontologiques dans les series rouges carbonatees permiennes du bassin
de Tiddas (Maroc Central) // C.r. Acad. sci. Ser. II. 1986. Vol. 303, N
3. P.
263-268.
Fontaine J.-M., Bruton C.H.C., Lys M. et al. Donnees nouvelles
sur la stratigraphie des formations paleozoiques de la plateforme Arabe
dans la region d'Hazro
(Turquie) // C.r. sci. D. 1980. Vol. 291. P. 917-920.
Fuchs G., Grauvogel-Stamm L., Mader D. Une remarquable flore a
Pleuromeia et Anomopteris in situ du Buntsandstein Moyen
(trias inferieur) de l'Eifel
(R.F.
Allemagne): Morphologic, paleoecologie et paleogeographie // Palaeontographica.
B. 1991. Bd. 222, Lfg. 4/6. S. 89-120.
Gand G., Kerp H., Passons C., Martinez-Garcia E. Palaeoenvironmental and
stratigraphie aspects of animal traces and plant remains in Spanish Permian
red beds. (Pena
Sagra, Cantabrian Mountains, Spain) // Geobios. 1997. Vol. 30, N 2. P.
295-318.
Gomankov A.V. Kitchkas flora from the Lower Tatarian of the Southern Urals
// Paleontol. J. (Moscow). 1995. Vol. 29, N 2A. P. 81-104.
Gothan W. Callipteris martinstii II Abbild. und Beschr. foss. Pflanzen,
1907. Lg. 5, N 96. S. 4.
Gothan W., Nagalhard K. Kupferschieferpflanzen aus dem niederrheinischen
Zechstein // Jb. preuss. geol. Landesanst. 1922. Bd. 42. S. 440-460.
Haubold H., Schaumberg G. Die Fossilien des Kupferschiefers. Wittenberg;
Lutherstadt: Ziemsen, 1985. 223 S.
Hill C.R., Wagner R.H., El-Khayal A.A. Qasimia gen. nov., an early Maram'a-like
fern from the Permian of Saudi Arabia // Ser, geol. 1985. Vol. 79. P. 1-50.
Kerp H. The study of fossil gymnosperms by means of cuticular analysis
// Palaios. 1990. Vol. 5. P. 548-569.
Kerp H. Der Wandel der Walder im Laufe des Erdaltertums // Natur
und Museum. 1996. Bd. 126, H. 12. S. 421-430.
Krassilov V.A., Afonin S.A., Lozovsky V.R. Floristic evidence of transitional
Permian-Triassic deposits of the Volga-Dvina region // Permophiles. 1999.
N 34. P. 12-14.
Krassilov V.A., Afonin S.A.,Naugolnykh S.V. Permotheca with in
situ pollen
grains from de Lower Permian of the Urals // Palaeobotanist. 1999. Vol.
48. P. 19-25.
Krassilov V.A., Burago V.I. New interpretation of Gaussia (Vojnovskyales)
// Rev. Palaeobot. and Palynol. 1981. Vol. 32. P. 227-237.
Lemoigne Y. Flore mixte au Permien superieur en Arable Saoudite // Geobois.
1981. N 14, fasc. 5. P. 611-635.
Li Xingxue, Rigby J., Wu Yiming. A biostratigraphical comparison between
the lower Gondwana floras of Southern Xizang (Tibet) and Kashmir // Gondwana
Seven
proceedings / Ed. by H. Ulbrich and A.C. Rocha Campos. Sao Paulo, 1991.
P. 533-538.
Lozovsky V.R., Krassilov V.A., Afonin S.A. et al. Transitional Permian-Triassic
deposits in European Russia, and non-marine correlations // Naura Bresciana.
Ann. Mus. Civ. Sci. Natur. Brescia. Monogr. 2001. N 25. P. 301-310.
Mamay S.H., Miller D.H., Rohz D.H., Stein W.E. Foliar morphology and anatomy
of the Gigantopterid plant Delnortea abbottiae from the Lower Permian of
West Texas // Amer. Journ of Botany. 1988. Vol. 75. P. 1409-1433.
Mamay S.H., Reed B.L. Permian plant megafossils from the conglomerate of
Mount Dall central Alaska range // US Geol. Surv. Circ. 1984. Vol. 868.
P. 98-102.
Meyen S.V. Fundamentals of palaeobotany. L.; N.Y.: Chapman and Hall, 1987.
432 p.
Meyen S.V. Permian conifers of Western Angaraland // Rev. Palaeobot. and
Palynol. 1997. Vol. 96. P. 351-447.
Meyen S.V. On the Subangara palaeofloristic area of the Permian // Сборник
памяти В.А. Вахрамеева. М.: Геос, 2002. С. 232-246.
Naugolnykh S.V. Paleobotany of the Upper Carboniferous / Lower Permian
of the Southern Urals. I. Seeds and enigmatics //Permophiles. 1999. Iss.
33.
P. 27-31.
Naugolnykh S.V. Paleobotany of the Upper Carboniferous / Lower Permian
of the Southern Urals. 2. Roots and woods // Ibid. 2000a. Iss. 36. P. 24-27.
Naugolnykh S.V. Mixed Permian floras of Eurasia // Paleontol. J. (Moscow),
2000b. Vol. 34, suppl. 1. P. 99-105.
Naugolnykh S.V. Paracalamitina striata - a newly reconstructed equisetophyte
from the Permian of Angaraland // J. Paleontol. 2002. Vol. 76, N 21. P.
377-385.
Nougolnykh S.V., Kerp H. Aspects of Permian palaeobotany and palynology.
XV. On the oldest known peltasperms with radially symmetrical o.vuliferous
discs
from the Kungurian (uppermost Lower Permian) of the Fore-Urals (Russia)
// Rev. Palaeobot. and Palynol. 1996. Vol. 91. P. 35-62.
Pant D.D., Nautiyal D.D., Srivastava P.C. The occurence of Cathaysian elements
in Glossopteris flora of Kashmir // Phyta. 1984. N4/5. P. 47-52.
Rigby J.F. The significance of a Permian flora from Irian Jaya (West New
Guinea) containing elements related to coeval floras of Gondwanaland and
Cathaysialand
// Palaeobotanist. 1996. Vol. 45. P. 295-302.
Rouvre L. Sur 1'age des flores paleozoiques nouvellement decouvertes sur
la bordure ouest de Г Air (Niger) // C.r. Acad. sci. Ser. II. 1988. Vol.
306.
P. 1057-1061.
Schmalhausen J. Die Pflanzenreste der artinskien und permischen
Ablagerungen im Osten des Europaischen Russlands // Mem. Comite geol. 1887.
Vol. 2,
N 4. P. 1-42.
Schweitzer H.-J. Makroflora des niederrheinischen Zechsteins // Senckenberg.
lethaea. 1960. Bd. 41. S. 37-57.
Schweitzer H.-J. Der weibliche Zapfen von Pseudovoltzia liebeana und seine
Bedeutung fur die Phylogenie der Koniferen//Palaeontographica. 1963. Bd.
113. S. 1-29.
Schweitzer H.-J. Flora des Oberen Perms in Mitteleuropa//Naturwiss. Rudsch.
1968. Bd. 21. S. 93-102.
Singh G., Maithy P.K., Base M.N. Upper Palaeozoic flora of Kashmir // Palaeobotanist.
1982. Vol. 30, N 2. P. 185-232.
Stoneley H. The Upper Permian flora of England // Bull. Brit. Mus. (Natur.
Hist.) Geol. 1958. Vol. 3. P. 293-337.
Wagner R.H. On a mixed Cathaysia and Gondwana flora from SE. Anatolia (Turkey)
// C.r. IV Congr. avanc. et etud. strat. et geol. Carbonifere, Heerlen.
Maastricht, 1983. T. 3. P. 745-752.
Wagner R.H., Hill C.R., El-KhayalA/A. Gemellitheca gen nov., a
fertile pecopterid fern from the upper Permian of the Middle East // Scr.
geol. 1985. Vol.
79. P. 51-74.
Wagner R.H., Lausberg S., Naugolnykh S.V. The Permian Angara flora from
North Greenland: A progress report // XIV Intern, congr. on the Carboniferous
and
Permian, 1999: Abstracts. Calgary. 1999. P. 150.
Wagner R.H., Lausberg S., Naugolnykh S. The Permian Angara flora from North
Greenland: A progress report // Carboniferous of the World / Ed. L.V. Hills
et al. Calgary,
2002. P. 382-392. (Canad. Soc. of Petrol. Geol. Mem.; N 19).
Wagner R.H., Soper N.J., Higgins A.K. A Late Permian flora of
Pechora affinity in North Greenland // Groenlands geol. unders. 1982.
Vol. 108. P. 5-13.
Wang Zi-qiang. Gigantic palaeobotanical events in North China // Acta Palaeontol.
Sinica. 1989. Vol. 28, N 3. p. 314-343.
Wang Zi-qiang. Successional trends of plant community during the Paleo-Mesophytic
transition in North China // Chinese Sci. Bull. 1982. Vol. 37, N 13. P.
1098-1103.
Wang Zi-qiang. Evolutionary ecosystem of Permian-Triassic redbeds in North
China: A historical recond of global desertification // The nonmarine Triassic.
New
Mexico. 1993. P. 471-476. (Mus. of Natur. History and Sci. Bull.; N 3).
Wang Zi-qiang. Recovery of vegetation from the terminal Permian mass extinction
in North China // Rev. Palaeobot. and Palynol. 1996. Vol. 91. P. 121-142.
Weigelt J. Die Pflanzenwelt des mitteldeutschen Kupferschiefers und ihre
Einschaltung ins Sediment // Fortschr. geol. Palaontol. 1928. Bd. 6, N
19. S. 395-592.
Weigelt J. Neue Pflanzenfunde aus dem Mansfelder Kupferschiefer//Leopoldina.
1930. Bd. 6. S. 643-668.
Wnuk C. The development of floristic provinciality during the Middle and
Late Paleozoic // Rev. Palaeobot. and Palynol. 1996. Vol. 0. P. 5-40.
Ziegler A.M. Phytogeographic patterns and continental configurations
during the Permian Period // Palaeozoic palaeo-geography and biogeography:
Boulder
(Colo.),
1990. P. 363-379. (Geol. Soc. Mem.; N 12).
ОГЛАВЛЕНИЕ
Введение (Н.М. Чумаков).......................................................................................................................................
5
Часть I
ЭПОХА ПЕРЕХОДА ОТ ТЁПЛОЙ К ХОЛОДНОЙ БИОСФЕРЕ: КЛИМАТ ПАЛЕОГЕНА
Глава 1. Климат Земного шара в палеоцене
и эоцене по данным палеоботаники (М.Л. Ахметьев)............. 10
Часть II
КЛИМАТ И ПЕРЕСТРОЙКИ В ТЁПЛОЙ БИОСФЕРЕ (МЕЛ, ЮРА)
Глава 2. Общий обзор позднемезозойского
климата и событий (Н.М. Чумаков)............................................
44
Глава 3. Палеогеографические
перестройки и седиментация мелового периода
(М.А. Жарков, И.О. Мурдмаа, Н.И. Филатова)....................................................................................................
52
Глава 4. Количественные палеоботанические
данные о позднемеловом климате
Евразии и Аляски (А.Б. Герман)............................................................................................................................
88
Глава 5. Климатическая зональность
и климат мелового периода (Н.М. Чумаков).........................................
105
Глава 6. Глобальная экспансия
планктонных фораминифер: триас, юра, мел
(К.И. Кузнецова, О.А. Корчагин)..........................................................................................................................
124
Глава 7. Динамика и возможные
причины климатических изменений в позднем мезозое
(Н.М. Чумаков)........................................................................................................................................................
149
Часть III
ЭПОХА ПЕРЕХОДА ОТ ХОЛОДНОЙ К ТЁПЛОЙ БИОСФЕРЕ (ПЕРМЬ И РАННИЙ ТРИАС)
Глава 8. Палеогеографические перестройки
и обстановки седиментации в перми и раннем
триасе (М.Л. Жарков)..............................................................................................................................................
158
Глава 9. Палеобиогеография пермских
фузулинид (Э.Я. Левен)........................................................................
181
Глава 10. Палеофитогеография
пермского периода (С.В. Наугольных).............................................................
194
Глава 11. Палеопочвы перми и
раннего триаса (С.В. Наугольных)...................................................................
221
Глава 12. Климат и климатическая
зональность перми и раннего триаса (Н.М. Чумаков).............................
230
Часть IV
ГЛАВНЫЕ КЛИМАТИЧЕСКИЕ И БИОСФЕРНЫЕ СОБЫТИЯ ПОЗДНЕГО ДОКЕМБРИЯ
Глава 13. Ледниковый и безледниковый
климат в докембрии (Н.М. Чумаков)...............................................
259
Глава 14. Проблема климатической
зональности в позднем докембрии. Климат и биосферные
события (Н.М. Чумаков, В.Н. Сергеев).................................................................................................................
271
Заключение (Н.М.
Чумаков)................................................................................................................................
290
Conclusion...............................................................................................................................................................
296