Глава 5
КЛИМАТИЧЕСКАЯ ЗОНАЛЬНОСТЬ
И КЛИМАТ МЕЛОВОГО ПЕРИОДА
Н.М. Чумаков
Рассмотренные в гл. 3 и
опубликованные ранее карты обстановок седиментации [Жарков и др., 1995,1998
а, б], а также
приложенные ниже палеобиогеографические схемы
для ряда климатически наиболее показательных организмов по векам мелового
периода (рис. 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, см.
вклейку II)
вместе с обобщением
качественных и количественных оценок климатических параметров, полученных
палеоботаническими, изотопными, геохимическими и литологическими методами,
позволили составить мелкомасштабные
глобальные схемы климатической зональности для 12 веков мелового периода
(рис. 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, см.
вклейку II).
Первые варианты
таких реконструкций для аптского, альбского, сеноманского и туронского
веков среднего мела были опубликованы ранее в черно-белом варианте [Чумаков
и др.,
1995]. Более подробно методика составления палеоклиматических схем представлена
в указанных публикациях и кратко во Введении к данной книге.
5.1. Климатические пояса
Из-за значительной продолжительности
большинства веков мелового периода на предлагаемых схемах отражена усредненная
картина климатической зональности,
поскольку внутривековые смещения границ климатических поясов в большинстве
случаев не могут быть уловлены и показаны. Такие перемещения проявляются
в виде зон, в которых встречаются разнородные индикаторы. Они протягиваются
вдоль границ выделенных климатических поясов и представляют что-то вроде "климатических
экотонов". Надо признать, что проведение границ климатических поясов
при этом производится в определенной степени субъективно, по самым ярким
климатическим индикаторам, таким как эвапориты, латериты, каменные угли,
ледовые отложения. Поэтому на картах показано максимальное в пределах века
распространение аридных, экваториальных и холодных поясов. Значительно точнее
внутривековые смещения границ поясов устанавливаются при сопоставлении разрезов
соответствующих ярусов, расположенных по обе стороны от границы. Однако такие
детальные исследования по своим масштабам выходят за рамки данной работы.
Представляемая серия палеоклиматических схем позволяет, со сделанными оговорками,
последовательно реконструировать меловую климатическую зональность от века
к веку и наметить ее общую эволюцию. Ниже дается характеристика климатических
поясов, начиная с высокоширотных, которые отличались в мелу особым своеобразием.
Высокоширотные умеренные климатические пояса. Как известно, полярные широты
являются областями наиболее чувствительными к климатическим изменениям, поэтому
они ярче других поясов отражают состояние климатической системы Земли и представляют
особый интерес для понимания специфики и динамики меловых климатов. По этой
причине в данной работе им уделено наибольшее внимание.
В интервале палеоширот 60°-85° меловые отложения охарактеризованы стратиграфически
и палеоклиматически в нескольких регионах: на севере Северной Америки и Азии,
в Южной Австралии, в Новой Зеландии, частично на Антарктическом п-ове. Судя
по многим признакам в течение мелового периода в этих регионах располагались
пояса с умеренным гумидным климатом. Как уже отмечалось в гл.
2, собственно
ледниковые отложения и следы многолетней мерзлоты здесь не обнаружены, а спорадическое
появление ледово-морских отложений (готерив-баррем Аляски) [Detterman et
al., 1975]; валанжин-готтерив и апт-альб Шпицбергена [Pickton, 1981]; валанжин-ранний
альб Австралии [Frakes et al., 1995] свидетельствует не об оледенениях, а лишь
о кратковременных похолоданиях, которые иногда и местами приводили к
сезонным замерзаниям морей. О кратковременных похолоданиях говорит также эпизодическое
образование глендонитов [Каплан, 1979].
Такой вывод в общем не противоречит мнению биостратиграфов о том, что моря
северного высокоширотного пояса (относимые обычно к Арктической биогеографической
провинции) были населены организмами, характерными для умеренно прохладных
вод. Впрочем, на этом общем фоне здесь временами происходили, по-видимому,
заметные похолодания и потепления, о чем говорят существенные изменения разнообразия
биоты (например, в неокоме) [Zakharov, 1992, Захаров и др., 2002].
Состав и облик наземной растительности и фауны подтверждают преобладание в
высоких и части средних широт Северного полушария пояса умеренного и гумидного
климата на протяжении всего мелового периода. Территориально этот пояс почти
совпадает с Сибирско-Канадской палеофлористической областью, выделенной В.А.
Вахрамеевым [1988]. В неокоме были распространены густые леса, состоявшие из
листопадных гинковых, чекановских и других голосеменных с папоротниковым и
хвощевым подлеском. На прибрежных равнинах большое распространение имели папоротниковые
марши [Красилов, 1985]. Последние, а также долины рек, открытые и полуоткрытые,
заболоченные пространства внутри континента, были не только областями интенсивного
углеобразования, но, судя по многочисленным палеонтологическим находкам, местами
обитания динозавров и насекомых (рис. 21, 23, 25, 27, см. вклейку
II).
В неокоме южная граница пояса располагалась в средних широтах и, протягивалась
с запада на восток, постепенно опускаясь от 50°-50° с.ш. в Северной Америке
до 40° с.ш. в Восточной Сибири. Еще восточнее, с приближением к Тихому океану,
она снова несколько поднималась. Суммарная ширина пояса почти достигала 45°
и не удивительно, что климат в его пределах не был однородным. Как уже отмечалось,
зимой в северной части пояса моря временами и местами замерзали, а на юге,
в Западной и Центральной Сибири, отлагались богатые органикой глины и иногда
образовывались каолинитовые (рис. 21-25, см. вклейку
II) и бокситовые коры
выветривания [Цеховский и др., 1995; Жарков и др., 1998 а, б].
Во второй половине раннего и самом начале позднего мела южная граница пояса
повсеместно отступила к 60° с.ш. и приобрела субширотное простирание. Это указывает
на общее потепление, особенно в пределах Восточной Сибири (рис. 29, 31, 33,
см. вклейку II). В целом с апта до начала турона в высоких северных широтах
преобладал весьма теплый климат, хотя на общем фоне этого потепления могли
происходить эпизодические кратковременные колебания климата.
В конце апта - начале альба появились первые покрытосеменные растения. С конца
альба в возрастающем количестве стали распространяться смешанные хвойно-широколиственные
леса, с листопадными покрытосеменными древесного и кустарникового типа. Среди
покрытосеменных были широко распространены формы с платановидными листьями
[Spicer, Corfield, 1992; Spicer et al, 1993; Герман, 1993; 1994], а среди хвойных
встречались многочисленные сосновые и секвой [Красилов, 1985; Вахрамеев, 1988].
Быстрой экспансии покрытосеменных в конце альбского - начале сеноманского века
очевидно способствовало кратковременное похолодание в конце альба ("амкинское"),
освободившее экологические ниши для новоселов [Лебедев, 1993]. Это похолодание
имело, возможно, глобальный характер, что выразилось в более широком распространении
сезонных ледовых отложений и расширении в южном Заполярье умеренного пояса
(рис. 31 и 32). Возможно также, что с ним были связаны сравнительно низкие
позднеальбские температуры, полученные методом CLAMP в Казахстане (гл.
4).
Ранее высказывались предположения, что во внутриконтинентальных полярных районах
Северо-Восточной Азии (Вилюйская впадина), где в составе растительности встречались
жестко- и мелколистные покрытосеменные и хвойные, климат был несколько иным,
более континентальным и сухим, с холодной зимой [Буданцев, 1979; Вахрамеев,
1988]. Однако широкое развитие здесь каолинитовых песков и присутствие остатков
теплолюбивых насекомых не согласуются с этими предположениями. Данные, полученные
методом CLAMP (см. гл. 4), также указали на сравнительно теплые зимы и достаточное
количество осадков в этих районах. В целом, с апта до начала турона в высоких
широтах преобладал весьма теплый климат, о чем свидетельствует повышение температуры
глубинных вод океана [Barrera, Savin, 1999]. Эти воды, как известно, формируются
в приполярных широтах Земли.
С туронского века северный высокоширотный умеренный пояс снова слегка расширился
(рис. 35, 37, 39, 41, 43). Это, по-видимому, было связано с начавшимся медленным
общим глобальным похолоданием, которое по разным признакам фиксируется во всех
широтах, и в том числе по устойчивому становлению, начиная с коньякского века
умеренного климата в южной Австралии. Умеренный характер растительности в пределах
рассматриваемых поясов, однако, сохраняется. Во второй половине позднемеловой
эпохи появляются кипарисовые, а роль папоротников постепенно сокращается [Вахрамеев,
1998], хотя в приморских районах с раннего мела сохраняются реликтовые элементы
гинкговых, чекановских, цикадовых и других групп. После некоторого коньякско-сантонского
расширения северный умеренный пояс в кампане и начале Маастрихта вновь слегка
отступил к северу.
Начиная с берриаса, на протяжении всего мелового периода в пределах рассматриваемого
пояса, от его южных границ вплоть до самых северных участков суши (палеошироты
80°-85°) шло интенсивное угленакопление. Это свидетельствует об очень большой
продуктивности растительного покрова. Резкий переход от полярного дня к холодной
полярной ночи, очевидно, тоже благоприятствовал угленакоплению, так как снижал
процессы биологической и химической деструкции отмершей растительной массы
и способствовал ее сохранности и дальнейшему захоронению. Можно, кроме того,
предполагать, что растительные сообщества высоких широт, не отличавшиеся высоким
видовым разнообразием, обладали относительно простой структурой и, следовательно,
характеризовались более высоким производством мортмассы [Krasilov, 1992]. В
результате совместного действия этих факторов в течение мелового периода сформировался
уникальный по своему положению, протяженности и запасам полярный угленосный
пояс.
Неизбежное в заполярных широтах чередование длительных полярных ночей и дней
должно было приводить к резкой сезонности климата, температурной и световой.
Признаки сезонности мелового высокоширотного климата хорошо устанавливаются
по палеоботаническим данным: по листопадности растений и структуре древесины.
Редко встречающиеся ложные кольца прироста в ископаемых стволах деревьев, большая
относительная ширина и крупные поперечные сечения трахеид у колец ранней (весенне-раннелетней)
древесины из альб-сеномана Северной Аляски указывают на достаточное и равномерное
увлажнение лесов в течение вегетационного периода [Spicer et al., 1992]. Это
подтверждается данными метода CLAMP (см. гл. 4).
В то же время тонкие линзовидные прослои ископаемого древесного угля (фюзинита),
отмеченные в некоторых каменных
углях, свидетельствуют о довольно частых, но, по-видимому, небольших по масштабам
лесных пожарах [Spicer et al., 1993]. Таким образом, длинный
(три-четыре месяца) полярный день, достаточно влажный и теплый, с большой долей
рассеянного света был, несомненно, благоприятным для произрастания растений
[Герман, 1994] и жизни животных.
Находки остатков ряда теплолюбивых и сравнительно теплолюбивых меловых животных
подтверждают выводы, сделанные на основании геологических и палеоботанических
данных о сравнительно теплом климате в пределах северного высокоширотного умеренного
пояса. Насекомые умеренно-теплолюбивого облика на севере Азии в апте, сеномане
и сантоне встречались до 75° с. палеошироты (рис. 29, 33 и 39). В неокоме остатки
динозавров известны на севере Азии (палеошироты 50°-55° [Несов, 1997]) и на
Аляске (палеошироты 60°-70°). В альбских и особенно в маастрихтских отложениях
остатки динозавров встречаются еще севернее: на северо-востоке Азии на палеоширотах
70°-80° [Несов, 1997], а на северной Аляске на палеоширотах 70°-85° с.ш. [Browers
et al., 1987; Clemens, Nelms, 1993 и др.]. На севере Канадского Арктического
архипелага, на палеошироте 70°-75° с.ш., в туронско-коньякских отложениях недавно
обнаружены остатки крупных хампсозавров и черепах [Tarduno et al., 1998]. В
южной части рассматриваемого пояса, в Северной Америке (на палеошироте более
60°), в кампанских и маастрихтских отложениях найдены остатки крокодилов [Markwick,
1998]. Подробнее на климатическом и экологическом значении всех этих находок
мы остановимся дальше, но в целом они указывают, как минимум на умеренный и
в основном умеренно-теплый климат мелового Заполярья.
Литологические и палеонтологические данные о преобладании в пределах высоких
северных широт умеренного и временами даже умеренно теплого климата хорошо
согласуются с количественными палеотемпературными оценками, как палеоботаническими,
так и изотопными. Изотопно-кислородным методом по белемнитам для валанжина
на палеошироте около 60° были получены средние палеотемпературы приблизительно
+8 °С (от 5° до 10 °С) [Ditchfield, 1997]. Если даже исходить из предположения,
что белемнитовые ростры росли лишь летом, эти температуры заметно выше соответствующих
современных средних летних температур на той же широте. Значительно больше
палеотемпературных данных существуют для среднего и позднего мела северного
высокоширотного пояса. Они получены методом CLAMP для Северо-Восточной Азии
и Аляски [Спайсер, Герман, 1998]; см. также гл. 4. Данные CLAMP показывают,
что на северо-востоке Евразии среднегодовые температуры в позднем альбе - сеномане
на палеошироте 62°-76° составляли 8°-13 °С, а средняя температура самого холодного
месяца от -2° до +3° - +6 °С, что соответствует в настоящее время климату Франции,
Германии, Крыма и Северного Кавказа. На Аляске на палеошироте 73° средняя температура
самого холодного месяца достигала +8 °С. В туроне-коньяке на северо-востоке
Евразии в пределах палеоширот - 72°-82°, т.е. несколько севернее температуры
были немногим ниже: среднегодовые составляли 7-9 °С, а самого холодного месяца
от -4° до 0 °С. На севере Аляски на широте 78° они были даже выше, соответственно
+13 °С и +6 °С. Таким образом, количественные определения палеотемператур тоже
подтверждают, что климат в пределах северного высокоширотного пояса был в меловой
период умеренно теплым. Довольно близкие температурные параметры получаются,
если оценивать физиологические ограничения толерантности крокодилов, в кампане
и Маастрихте проникавших в Северной Америке в южную часть рассматриваемого
пояса (до палеошироты свыше 60°). Для них минимальная среднегодовая температура
определяется в 14 °С, а средняя температура самого холодного месяца не ниже
+5 °С [Markwick, 1998]. Упоминавшаяся уже находка в туронско-коньякских отложениях
остатков хампрозавров, близких родственников крокодилов, со сходным образом
жизни, на севере Полярного канадского архипелага, палеошироте 75° [Tarduno
et al., 1998] дает дополнительные подтверждения приведенным выше количественным
оценкам. Не исключено, впрочем, что максимальные из них отражают достаточно
кратковременные потепления.
В средних и высоких палеоширотах южного полушария таксономический состав раннемеловой
флоры был иным, чем в Арктике: преобладали подокарповые и араукариевые хвойные
(Австральная палеофитогеографическая область В.А. Вахрамеева, [1988]). В оценках
раннемелового климата этой области между исследователями существуют некоторые
расхождения. В.А. Вахрамеев целиком отнес Австральную область к субтропикам,
отмечая в частности находки окремнелых стволов древовидных папоротников в Австралии
и стволов с широкими кольцами роста в Антарктике. В.А. Красилов [1985] и Р.
Спайсер с соавт. [Spicer, Corfield, 1992 и др.] рассматривают, однако, раннемеловую
южную Австралию как район с умеренным или холодноумеренным климатом. В нижнем
мелу растительность Южной Австралии характеризовалась хвойными, по-видимому,
несомкнутыми лесами с подокарповыми и араукариевыми доминантами и болотами,
в которых шло интенсивное углеобразование. С этими ландшафтами была связана
разнообразная фауна позвоночных, включавшая динозавров, лепидозавров, черепах,
лабиринтодонтных амфибий и др. [Константин и др., 1999]. Среди растений встречались
листопадные, но, по-видимому, преобладали деревья, не опадавшие зимой [Falcon-Lang,
Cantrill, 2001], частично мелколистные с толстой кутикулой, что указывает на
сравнительно низкие зимние температуры и некоторую сухость [Spicer, Corfield,
1992]. Данные выводы согласуются с присутствием в нижнемеловых отложениях южной
Австралии сезонных ледовых отложений и глендонитов [Frakes et al., 1995]. Учитывая
высокоширотное положение этого района в раннем мелу, можно рассматривать его,
как северную часть южного высокоширотного умеренного пояса. Северная граница
пояса здесь намечается для апта по находкам умеренно-теплых насекомых, а для
апт-альба по сравнительно более частой встречаемости ледово-морских отложений
(см. рис. 29, 31). Более холодный климат для апт-альба южной Австралии предполагается
также на основании анализа морфологии криотурбаций и изотопно-кислородного
анализа карбонатных конкреций. По криотурбациям, встречающимся на самом юге
континента, среднегодовые температуры оцениваются от -2 °С до +3 °С [Константин
и др., 1999], а согласно изотопии сначала в - 2 °С +/- 5 °С [Gregory et al.,
1989], а позднее от -8° до +6 °С [Fergusson, Gregory, 1999]. Возможно, что
эти данные, также как спорадически встречающиеся сезонно-ледо-вые отложения,
указывают на заметные, но эпизодические похолодания случавшиеся в данном регионе,
поскольку общий характер растительности свидетельствует о более теплом климате.
Он позволяет предположить, что среднегодовые температуры альба составляли здесь
приблизительно +5° - +8 °С [Spicer et al., 1993]. Сходные цифры получены математическим
моделированием [Barron, Washington, 1982]: зимние температуры около -18 °С,
а летние +27 °С (отсюда среднегодовые температуры ориентировочно можно оценить
в +4°-5 °С).
Вслед за В.А. Красиловым [1985] раннемеловой климат Южной Австралии в целом
можно квалифицировать как умеренный. По-видимому, позднее во время сеноманского
потепления, умеренный пояс отступал и располагался южнее Австралии, а в туроне
вновь продвинулась к северу, в пределы южной
Австралии [Чумаков и др., 1995]. Более уверенно южная умеренная зона устанавливается
с коньякского века. А с сантоннского века южный умеренный пояс уверенно выделяется
В.А. Вахрамеевым по появлению нотофагусовой пыльцы.
Изучение колец роста фоссилизированных стволов деревьев из раннемеловых отложений
бассейна Ероманга в южной Австралии позволяет различить присутствие двух популяций
деревьев, росших в несколько различных ландшафтных и климатических условиях.
Одна популяция, по-видимому, обитала на возвышенностях в более холодном климате,
подобном тому, в котором растут современные леса умеренно-холодной зоны на
западных склонах южных Анд и севере Огненной Земли, а другая - на низменностях,
в более теплом климате, подобном тому, в которых растут современные леса умеренно-теплой
зоны на юге Новой Зеландии и в Тасмании [Frakes, Francis, 1990]. Присутствие
во вмещающих отложениях горизонтов со следами ледового разноса указывает на
то, что зимой южная часть бассейна Ероманга иногда замерзала.
Леса близкие к южноавстралийским росли и на Антарктическом п-ове. Детальное
изучение найденных в прижизненном положении остатков ископаемого позднеальбского
леса вблизи основания Антарктического полуострова показало, что лес был средней
густоты и на 90% состоял из круглогодично зеленых широколиственных хвойных
деревьев (подокарповых и араукариевых). Этот лес имел определенное сходство
с современными влажными умеренными лесами южной Новой Зеландии [Falcon-Lang
et al., 2001a]. Изучение анатомии ископаемой древесины подтверждает, что климат
северной части Антарктического полуострова был умеренно теплым [Thorn, 2001].
В Новой Зеландии в это время были распространены леса, состоявшие из листопадных
и хвойных вечнозеленых растений, которые, были южными аналогами меловых высокоширотных
приморских лесов Арктики [Kelly, Moncrieff, 1992; Spicer, Corfield, 1992; Falcon-Lang,
Cantrill, 2001].
В кампанских отложениях Антарктического п-ова и Новой Зеландии были найдены
остатки крупных динозавров и птерозавров, частично сходные с раннемеловыми
формами Австралии [Molnar, Wiffen, 1994]. Зимы здесь были очевидно менее холодными,
чем в южной Австралии, и, в отличие от последней, местами (Новая Зеландия)
в значительно большем количестве росли цикадофиты [Вахрамеев, 1988; Мейен,
1987]. Этот вывод хорошо согласуется с данными анализа листового края и
методом CLAMP, которые дали для Южного острова Новой Зеландии среднегодовые
температуры для сеномана 10°-16 °С [Kennedy, 1996], а для Маастрихта 12°-15
°С [Kennedy et al., 2002]. Структура ископаемой древесины с Антарктического
п-ова свидетельствует о том, что возможные нижний и верхний пределы среднегодовых
температур (МАТ) составляли для альба 13°-25 °С, для коньяка 15°-21 °С, для
сеномана 17°-23 °С, а для Маастрихта 7°-10 °С [Francis, Poole, 2002]. Палеоботанические
данные хорошо согласуются с изотопными. Для апт-альба и альб-сеномана по раковинам
моллюсков с о-ва Росса изотопным кислородным методом получены среднегодовые
палеотемпературы соответственно 10° и 12 °С, для коньяка-раннего сантона 18
°С, для позднего сантона-кампана 14 °С, а для маастрихта - от 9° до 15 °С
[Dichfield et al., 1994]. Эти цифры весьма близки к полученным Е. Кеннеди для
Новой Зеландии
и свидетельствуют о том, что в среднем и позднем мелу высокие южные широты,
подобно высоким северным широтам, имели, как минимум умеренно-теплый климат,
а временами и еще более теплый.
Естественно, что донные воды в высоких южных широтах Атлантического и Индийского
океанов имели более низкие температуры, чем поверхностные: для кампана 7°-10
°С, а для Маастрихта 5°-8 °С, с небольшим их повышением до 11 °С в самом конце
Маастрихта (по d18O бентосных фораминифер [Barrera, Savin, 1999]). Сходные
данные приводятся в других сводных работах, при этом отмечается сближение температур
поверхностных и глубинных вод [MacLeod et al., 2000], что указывает на отсутствие
в то время оледенений в высоких широтах и стока холодных ледниковых вод в океан.
Для сравнения напомним, что в настоящее время температура глубинных вод в высоких
южных широтах составляет от -0,5° до +1,5 °С [Степанов и др., 1984].
Способ существования теплолюбивой биоты в меловых заполярьях зимой представляет
определенную проблему. Важнейшим условием периода зимнего покоя растений, насекомых
и холоднокровных наземных позвоночных является резкое замедление темпа метаболизма.
При низких зимних температурах это происходит почти автоматически. Значительно
труднее этим организмам войти в состояние зимнего покоя при сравнительно небольших
отрицательных или положительных температурах, которые предполагаются для мелового
высокоширотного пояса. Растения могли при этом сбрасывать листву или побеги,
а при достаточно низких температурах получали возможность оставаться вечнозелеными,
приобретая мелко- и жестколистность. И в тех и в других условиях они могли
быть, кроме того, однолетниками или сохранять подземные органы с запасом питательных
веществ. Значительно более сложную проблему представляет экологическая и физиологическая
интерпретация условий обитания крупных растительноядных и хищных динозавров
в полярную ночь. Вопрос состоит в том, могли ли динозавры быть постоянными
обитателями заполярий, как думают некоторые исследователи [Несов, Головнева,
1990; Несов, 1992; Clemens, Nelms, 1993] или динозавры были кочевыми животными,
совершавшими дальние миграции [Ногпег, 1997 и др.]. В подтверждение "оседлости" динозавров
исследователи ссылаются на то, что в сеноне вместе с остатками взрослых особей
были найдены зубы детенышей динозавров, в том числе только вылупившихся [Clements,
Nelms, 1993]. Длительные миграции, по мнению этих авторов, не оставляли бы
времени для выведения и выращивания детенышей, были для последних опасны, а
кроме того, отрывали бы травоядных динозавров от привычных и безопасных мест
обитания и питания. Однако в случае "оседлости" требуется объяснить,
как динозавры могли жить в длительные (до трех-четырех месяцев) полярные ночи.
Эти ночи, как было показано выше, характеризовались низкими, близкими к отрицательным
и отрицательными температурами, а также отсутствием зеленой растительной пищи.
Очевидно, что эти гигантские (до 10 м длиной) [Brouwers et al., 1987] и к тому
же стадные животные не могли впадать в спячку, хотя бы из-за невозможности
найти убежища для зимовок. Одни сторонники "оседлости" динозавров
предполагают, что полярные зимы в меловой период были достаточно теплыми или
что динозавры могли использовать для зимовок и инкубации яиц места, обогреваемые
вулканическим теплом* [Несов, Головнева, 1990; Несов, 1992]. Другие полагают,
что заполярные динозавры были теплокровными животными [Clemens, Nelms, 1993;
Константин, 1999 и др.]. О последнем, по мнению У. Клеменса и Г. Нелмса, свидетельствует
полное отсутствие в местонахождениях позднемеловых позвоночных на Аляске остатков
амфибий и нединозавровых пресмыкающихся, составляющих в одновозрастных более
южных местонахождениях почти 50% комплекса позвоночных. Это обстоятельство
- меньшее разнообразие позвоночных в местонахождениях Северной Аляски и преобладание
динозавров и млекопитающих (50% и почти 30% родов соответственно от состава
комплекса) говорит, по мнению авторов, о достаточно холодном климате в высоких
широтах и о теплокровности динозавров. Представления о гомойотермности динозавров
не кажутся сейчас многим исследователям, в том числе некоторым сторонникам "оседлости" полярных
динозавров, достаточно основательными [Татаринов, 1987; Раутиан, 1990]. Кроме
того, обе гипотезы не объясняют, чем питались стада растительноядных гигантов
в течение длинной полярной ночи.
| * Это объяснение не приложимо к самым северным кампанским и маастрихтским
динозаврам северной Аляски. |
Представления о миграциях динозавров, как уже отмечалось, тоже встречают ряд
возражений. Однако, если представить, что стада растительноядных динозавров
вели кочевой или полукочевой образ жизни, то легко объяснить, как они переживали
полярные ночи. Динозавры могли, например, пастись и за период дневного бодрствования,
не спеша, по 15-20 км в день, т.е. по 1-1,3 км/час (средняя скорость их движения
определяется значительно выше - в несколько км/час [Paul, 1994]), продвигаться
вместе с весной в более высокие широты (а осенью - обратно) вдоль морских побережий,
приморских низменностей или речных долин, не отрываясь от необходимых им пищевых
угодий и убежищ. Такие перемещения на 15-20 °С (1500-2000 км) по меридиану
из умеренно-теплых в высокие умеренно-холодные широты, туда и обратно, занимали
бы около половины года, а 3-5 месяцев полярного лета оставались бы для выращивания
детенышей (рис. 45). Предположения о тахиметаболической эндотермии [Paul, 1994]
и яйцеживорождении у динозавров [Несов, 1997] делает гипотезу миграций еще
более вероятной. В конце лета динозавры таким же образом, не торопясь, уходили
бы от полярной ночи в средние теплые широты, полностью использовав преимущества
полярного дня, весьма продуктивного для растений и благоприятного для гнездования
и накапливания жировых запасов. Тогда бы стада динозавров посещали большинство
пастбищ один или два раза в году с большим перерывом и пастбища успевали бы
восстанавливать после их нашествия свою растительную массу. Прекрасными путями
для таких миграций в Северной Америке в меловой период могли быть берега меридионального
Западного Внутреннего морского пролива, соединявшего Арктический океан с Тетисом,
или глубоко вдававшиеся в сушу протяженные заливы, сменявшие по временам этот
пролив. На северо-востоке Азии такими путями могла быть система многочисленных
речных долин [Филатова, 1995]. В южной Австралии путями миграции могли служить
берега морского бассейна Эроманга. Нельзя исключать, что отсутствие сквозных
пресноводных путей миграции было, кроме более холодного зимнего климата, второй
возможной причиной отсутствия амфибий на севере Аляски. Приведенное предположение
о мигрирующих стадах динозавров в общем вписывается в пределы температур, устанавливаемых
разными методами, и согласуется с расположением реконструируемых климатических
поясов (рис. 45).
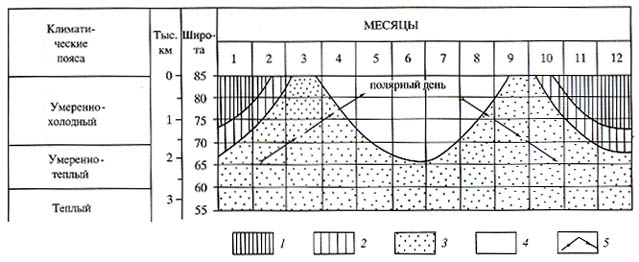
Рис. 45. Предполагаемый календарь сезонных миграций крупных динозавров
в высоких широтах
Условные обозначения: 1 — непрерывные полярные ночи; 2 — полярные сумерки; 3
— чередование дней и ночей; 4 — непрерывные полярные дни; 5 - миграции динозавров. |
Таким образом, сумма разных данных согласованно свидетельствует
о том, что в высоких широтах обоих полушарий на протяжении мела преобладал
климат, который
по температурам и влажности был несколько подобен современному умеренному.
По своему географическому положению этот климат можно назвать высокоширотным
умеренным, а соответствующие ему пояса - высокоширотными умеренными поясами.
Подобно современным умеренным поясам они характеризовались преимущественно
небольшими и средними положительными годовыми температурами (от нескольких
до 10°-14 °С), гумидностью, сезонностью и широким развитием лесов и болот.
Их спецификой были: расположение в значительно более высоких широтах и очень
большая световая и температурная сезонность, обусловленная чередованием непрерывных
заполярных дней и ночей. Спорадическое появление дропстоунов и глендонитов
указывает на то, что умеренно-теплый климат изредка сменялся умеренно-холодным.
Подобные климатические колебания были естественными при большой чувствительности
заполярий к небольшим климатическим колебаниям. Можно также полагать, что умеренно-теплый
климат преобладал во внешних приполярных зонах, а умеренно-холодный чаще проявлялся
в более высоких широтах, ближе к полюсам.
Среднеширотные теплые пояса. Климатические пояса, примыкавшие к описанным высокоширотным
поясам и охватывавшие главным образом средние широты, характеризовались в меловом
периоде весьма теплым и влажным, местами сезонно-влажным климатом, на что согласованно
указывают многочисленные литологические, палеонтологические и геохимические
индикаторы климата.
В Северном полушарии в этом поясе на суше были широко распространены сероцветные
терригенные угленосные отложения, бокситы, каолиновые коры выветривания и продукты
их переотложения (рис. с 21 по 43, нечетные). Их местонахождения преимущественно
тяготели к приморским и приокеаническим областям. Во внутриконтинентальных
районах формировались терригенные породы с почвенными комплексами, указывающими
на переменно-влажный климат [McCarthy et al., 1998], а местами (Южная Сибирь,
Центральная Азия) отлагались красноцветы, которые в отдельных небольших впадинах
содержали гипс (гл. 3). Это согласуется с мнением В.А. Вахрамеева об усилении
в неокоме в средних широтах Азии аридности в восточном направлении. В апте
и особенно в альбе увлажнение этих и многих других районов существенно увеличилось
[Вахрамеева, 1988].
В целом в рассматриваемом поясе произрастала богатая теплолюбивая флора, в
том числе беннеттитовые, цикадовые, с позднего альба - платанообразные, а с
турона - пальмы (Евро-Синийская флористическая область субтропической растительности
В.А. Вахрамеева [1988]). Преобладали вечнозеленые гумидные, семигумидные и,
по-видимому, сезонно влажные несомкнутые леса, главным образом хвойные с подчиненным
количеством древовидных папоротников и цикадофитов, обладавших маноксилическими
стволами, а также кустарники и возможно травы покрытосеменных, среди которых
доминировали растения с цельнокрайными листьями [Красилов, 1985; Wolfe, Upchurch,
1987; Spicer et al., 1993]. Очевидно значительную роль имели ландшафты сходные
с саванами. О сезонном увлажнении последних свидетельствует также характер
ископаемых почв [McCartty et al., 1998]. В центральной Европе, на богемском
микроконтиненте климат был теплый и в основном гумидный. На возвышенных участках
и прибрежных маршах преобладали хвойные, а в галерейных лесах - покрытосеменные
растения. И те и другие периодически выгорали во время короткого засушливого
сезона [Falcon-Lang et al., 200lb].
В пределах среднеширотного пояса обитали теплолюбивые насекомые, крокодилы
и многочисленные динозавры (рис. 21-24, см. вклейку II). Среднегодовые температуры
в южной части пояса (до 30°-45° с. палеошироты) по характеру растительности
оценивались в 20°С и более, а севернее - между 13° и 20 °С [Wolfe, Upchurch,
1987; Spicer, Corfield, 1992]. Заметно более низкие температуры для этих широт
получены методом CLMP (см. гл. 4). Для сеномана Центральной Европы - между
17° и 20 °С, Казахстана - около 15 °С, а позднего альба Казахстана - даже 10
°С. Последняя цифра вызывает определенные сомнения, если вспомнить, что для
позднего альба высоких широт Арктики (70°-68°) CLAMP дал среднегодовые температуры
от 8° до 13 °С. Возможно, что альбская тафрофлора Казахстана в основном отражает
высотную климатическую зональность или очень короткое позднеальбское похолодание.
Впрочем, не исключено, что тут проявилась ограниченность метода, которая была
недавно показана на примере современной флоры Южной Америки [Kowalski, 2002].
Исходя из широкого распространения бокситов (в Западно-Сибирском регионе вплоть
до 60° с. п.-ш., а в кампане и Маастрихте до 65°-70° с. п.-ш.) и актуалистической
интерпретации условий их образования, ряд исследователей склоняются к выводу
о том, что высокие среднегодовые температуры (не менее 20-22 °С) и значительная
влажность были
характерны для всего этого пояса, вплоть до его северных границ [Цеховский
и др., 1995]. Подтверждением этого мнения являются высокие среднегодовые температуры,
полученные методом CLAMP для коньяка северной Аляски (20 °С), турона Новосибирских
островов (19 °С, см. гл. 3) и находка теплолюбивых кампан-маастрихтских фораминифер
на междуречье рек Вилюй и Маркока [Черная и др., 2001]. В то же время нельзя
исключать, что все признаки очень теплого климата в высоких широтах отражают
лишь короткие эпизодические потепления, которые хорошо фиксируются по океаническим
осадкам.
В эпиконтинентальных морях пояса (Североморском, Центрально-Европейском, Московском,
Прикаспийском, Туранском, Западном Внутреннем бассейне Северной Америки) в
неокоме, аптском и альбском веках накапливались глауконитовые и фосфоритоносные
отложения, а с сеномана и особенно турона в европейских морях стали формироваться
мощные толщи писчего мела. Во внутреннем бассейне Северной Америки в это время
отлагались терригенные осадки и богатые органикой черные сланцы, а писчий мел
получил широкое распространение позднее, с коньякского века [Жарков и др.,
1995, 19986]. В Англии изотопно-кислородным методом по белемнитам температура
для среднего неокома оценивается в 12°-15,5 °С, за исключением раннего готерива,
для которого получена температура менее 9 °С [Price et al., 2000]. Температура
глубинных вод южной части северного среднеширотного пояса определена тем же
методом по донным фораминиферам для Маастрихта в 6-8 °С [Barrera, Savin, 1999].
Эпиконтинентальные моря этого пояса характеризовались в неокоме своеобразным
комплексом нано-планктона, который имел биполярное распространение и известен
как в северном, так и южном среднеширотном поясе [Mutterlose, Kessels, 2000].
Беспозвоночные, населявшие эти моря, принадлежали в основном к южным областям
бореальных палеобиогеографических царств [Hallam, 1973; Doyle, 1992] и частично
царству, переходному от бореального к тетическому [Koutsoukos, 1992]. На южной
окраине пояса, вдоль северной окраины центрального Тетиса были широко распространены
перикратонные карбонатные платформы (Иберийская, Армориканская, Карпато-Балканская,
Мизий-ская [Жарков и др., 1995; 1998 а, б], образованные гексакоралловыми банками
и биострома-ми, кораллово-рудистовыми постройками, водорослево-ламинитовыми
и строматолитовыми осадками, карбонатными илами с рудистами и фораминиферами,
органогенными песками [Masse et al., 1993; Philip et al., 1993]. Обитатели
этих морей преимущественно принадлежали уже к тетическому
(Hallam, 1973; Doyle, 1992] и переходному бореально-тетическому палеобиогеографическим
царствам [Koutsoukos, 1992]. Среди них заметное распространение имели герматипные
кораллы [Beauvais, 1992]. Все эти седиментологические и палеонтологические
данные указывают на равномерно теплый климат на территории всех эпиконтинентальных
морей, северных окраин Тетиса и прилегающих районов суши [Philip et al., 1993].
Здесь сказывалось, по-видимому, влияние теплых пассатных течений из Тихого
океана и Восточного Тетиса. Широтное расположение карбонатных платформ вблизи
30°-35° с.ш. хорошо иллюстрирует смещение на север теплых зон относительно
современной эпохи [Masse et al., 1993]. Ротационное вращение Земли в середине
мелового периода согласно расчетов Ллойда [Lloyd, 1982] превышало современное
на 3%, что могло привести к широкому развитию муссонно-го климата вдоль окраин
Тетиса и Центральной Атлантики и увеличению дождливых сезонов в среднеширотном
поясе [Lloyd, 1982; Parrish et al., 1982].
Для морей восточной окраины Азии кислородно-изотопные определения палеотемператур
по моллюскам показали, что в турон-кампанском интервале они колебались между
8° и 20 °С, хотя в основном были выше 15 °С и только в Маастрихте снизились
до 7°- 12 °С [Захаров и др., 2001]. На Хоккайдо (40° с.п.-ш.) по планктонным
и бентосным фораминиферам для кампанского века были получены еще более высокие
кислородно-изотопные температуры, соответственно около 26° и 18°С [Moriya et
al., 2003]. Для сравнения скажем, что современные субтропические моря имеют
среднегодовые температуры 15-20 °С, а моря южной части умеренного пояса 7-12
°С. Обращает на себя внимание, что в серии определений по моллюскам палеотемпературы,
полученные для туронского века в Корякин и на Хоккайдо, оказались близкими.
Авторы объясняют это существованием очень малого широтного температурного градиента.
Данное объяснение представляется маловероятным и не согласуется с уже упоминавшимися
более низкими палеотемпературными оценками, полученными методом CLIMAP для
северо-западной Камчатки (9 °С против 15-16 °С в Корякин).
По набору палеоботанических и литологических индикаторов рассмотренный пояс
часто именуют тропическим и субтропическим
[Ронов, Балуховский, 1981; Ясаманов, 1985], субтропическим [Вахрамеев, 1978;
1988], иногда "ослабленным тропическим", "почти тропическим" и "тропическим" [Синицын,
1980] или палеосубтропическим [Гольберт, 1987]. Эти названия не кажутся удачными,
поскольку прилагаются к поясу, весьма удаленному от тропиков и субтропиков.
В Северном полушарии данный пояс, то слегка расширяясь, то сильно сужаясь,
протягивался приблизительно между 60° и 30° с. палеошироты (в Южном полушарии
аналогичный пояс располагался между 40° и 65° ю. палеошироты). Поскольку в
течение геологической истории широтное положение климатических поясов сходных
по температурному режиму существенно менялось, неизбежно должно были изменяться
присущие им среднегодовые величины и амплитуды сезонных колебаний солнечной
радиации и температур, в большинстве случаев также величина и режим увлажнения,
направление преобладающих ветров, барические показатели. Все эти параметры
входят в число важнейших составляющих климата, поэтому нельзя называть палеоклиматические
пояса, исходя лишь из приблизительной аналогии средних температур, просто повторяют
названия современных климатических поясов*. Поэтому важно, чтобы названия климатических
поясов отражали реальное палеоширотное их положение. В этом случае станет понятным
не только место пояса на Земле, но и характер сезонности присущего ему климата.
С этой точки зрения рассмотренный сейчас меловой пояс и его аналог в южном
полушарии следует именовать "среднеширотными теплыми гумидными поясами" [Чумаков
и др., 1995]. Некоторые исследовали, исходя из широкого развития в этих поясах
латеритовых и каолиновых кор выветривания, предлагают именовать их "внетропическими
гумидными жаркими поясами" [Цеховский и др., 1995].
| * Или, что еще хуже, используются термины, производные
от современных названий климатических поясов, но значения которых в подавляющем
числе
случаев точно не определяются ("ослабленный тропический", "почти
тропический" или "палеосубтропический" климаты и др.). |
Ширина северного среднеширотного теплого пояса по простиранию
и в течение мелового периода изменялась. В неокоме он сужался с запада на восток
от 20° в Северной
Америке до 10° в Центральной Азии за счет расширения в этом направлении соседнего
аридного пояса. Временами среднеширотный теплый пояс почти прерывался в этом
районе (готерив, барем). В середине мелового периода пояс сильно расширился
в северном и южном направлении. Ширина его в это время была наибольшей (от
20° до 40°), хотя по-прежнему изменялась по простиранию.
В североамериканском и восточно-азиатском секторах были противоположные тенденции
изменения его ширины во времени. В американском секторе пояс от апта к турону
постепенно расширялся, а на востоке Азии за тот же период сузился, в результате
смещения к северу аридной зоны.
В Южном полушарии среднеширотный теплый гумидный пояс протягивался от юга Ю.
Америки и Африки до Австралии и характеризовался формированием на суше сероцветных
угленосных отложений и каолиновых кор выветривания (в пост-сеноманских отложениях
Австралии [Жарков и др., 1995, 1998 б]. В этом поясе произрастала теплолюбивая
флора, которую В.А. Вахрамеев [1988] отнес к австралийской провинции Австральной
(Нотальной) флористической области, и обитали довольно многочисленные динозавры
(рис. 21-44, см. вклейку II). По
характеру растительности и фауны, литологическим признакам и положению между
умеренным и аридным поясами большинство исследователей
называют этот пояс, как и аналогичный северный, "субтропическим" [Ронов,
Балуховский, 1981; Вахрамеев, 1985; 1988; Красилов, 1985; Ясаманов, 1985 и
др.]. Северная граница южного среднеширотного теплого пояса во времени несколько
смещалась (вследствие расширения или сужения смежной аридной зоны) и располагалась
на палеоширотах от 40° до 55° ю.ш. Положение же южной границы этого пояса в
неокоме не установлено, хотя отмечаемые в валанжине Южной Австралии дропстоуны
как будто бы достаточно определенно указывают на умеренно-холодный климат последней
[Fakes et al., 1995]. Положение этой границы в среднем мелу, как уже отмечалось
при описании высокоширотных поясов, вызывает некоторые разногласия. В.А. Вахрамеев
считал, что "субтропический" пояс в середине мелового периода распространялся
на всю Австралию и Южную Америку [1988]. Новые палеоботанические [Spicer, Corfield,
1992; Spicer et al., 1992, 1993], литологические [Frakes et al., 1995], палеотемпературные
[Ditchfield et al., 1994] и палеоэнтомологические (см. рис. 29) данные позволяют
присоединиться к мнению В.А. Красилова [1985] о том, что климат южной части
Австралии и, видимо, южного окончания Ю. Америки в меловом периоде в основном
был умеренным, а временами в Ю. Австралии даже умеренно-холодным [Fakes et
al., 1995; Spicer et al., 1993].
Это позволило наметить в данном регионе южную границу среднеширотного теплого
пояса по меньшей мере в апте и альбе (рис. 29-31) на палеоширотах 60-65 °С.
В сеномане и туроне южная граница теплого пояса, по-видимому, проходила южнее
Австралии, а в коньяк-ском веке возможно вновь продвинулась к северу и располагалась
на юге Австралии. С сантона южная граница южного теплого пояса достаточно уверенно
устанавливается по границе Австралийской и Антарктической провинций Нотальной
области В.А. Вахрамеева [1988].
Фауна морских беспозвоночных, населявших моря южного среднеширотного пояса,
имела свою специфику. Обычно эти регионы обособляются палеонтологами в качестве
Австрального (Нотального) палеобиогеографического царства и включаются, как
южная часть, в более широко понимаемое Южное умеренное царство [Hallam, 1973J.
Иногда эта фитохория рассматривается как самостоятельное царство, переходное
от Тетического к Австральному [Koutsoukos, 1992]. В пределах южного среднеширотного
пояса, к западу от Австралии, по пелагическим карбонатным осадкам и по их тонкозернистой
фракции изотопно-кислородным методом были определены средне- и позднемеловые
температуры поверхностных вод [Clarke, Jenkyns, 1999]. Температуры эти довольно
быстро повышались от 12 °С в позднем апте до 19 °С в среднем туроне, а затем
стали медленно снижаться до 12,5 °С в позднем компане. Температура глубинных
вод в средних южных широтах Атлантического океана, определенная тем же методом
по донным фораминиферам составляла 10-13 °С для кампана и 7-10 С для Маастрихта
[Barrera, Savin, 1999].
Обращает на себя внимание, что южный среднеширотный гумидный теплый пояс был
значительно уже, чем северный и был оттеснен аридными зонами в высокие средние
и частично в высокие широты. В Южном океане это было связано, видимо, с отклонением
к югу у берегов Африки южных субэкваториальных теплых пассатных течений, образовывавших
западную часть океанического круговорота. В конце раннего мела дополнительный
эффект мог давать сток теплых соленых вод из аптского Южно-Атлантического солеродного
бассейна в Южную Атлантику [Lloyd, 1982].
Аридные пояса (тропически-экваториальный пояс, северный и южный аридные пояса).
В неокоме и апте низкие и часть средних широт обоих полушарий Земли занимал
огромный аридный пояс, непрерывно простиравшийся приблизительно от 30°-40°
с. палеошироты до 45° ю. палеошироты (рис. 21-29). Аналога такого аридного
пояса, охватывающегй все экваториальные, тропические и значительную часть средних
широт обоих полушарий, на современной Земле нет и, соответственно,
нет соответствующего понятия и названия. Поэтому было предложено называть такие
пояса "тропическо-экваториальными аридными поясами" [Жарков и др.,
1995, Чумаков и др., 1995].
Этот пояс характеризовался широким развитием эвапоритов, которые в неокоме
отлагались по окраинам континентов, тяготея к северным и в меньшей степени
к южным тропическим широтам. Большие и устойчивые области аридного осадконакопления
в Юго-Восточной Азии и Южной Америке возникли в дождевой тени Катазиатского
и Тибетско-Индокитайских краевых вулканических поясов в первом случае и аналогичных
по природе позднемеловых Анд - во втором случае. Во внутриконтинентальных районах
пояса формировались терригенные красноцветы [Жарков и др., 1998а]. В апте эвапориты
и красноцветы стали интенсивно накапливаться в раскрывающемся рифте южной Атлантики
и в прилежащих частях Ю. Америки и Африки. Во внутриконтинентальных районах
Ю. Америки и Южной Африки на больших территориях продолжали формироваться красноцветные
[Жарков и др., 1998 б] и эоловые отложения [Scherer, 2000]. Об аридности пояса
свидетельствуют также приводимые ниже палеоботанические данные.
| * С позднего мела ее именуют иногда Пальмовой флористической
областью. |
Флористические ассоциации этого пояса, объединенные В.А. Вахрамеевым
[1988] в Экваториальную флористическую область*, довольно бедны и представлены
главным
образом палинологическими комплексами. Последние указывают на сухой и жаркий
климат (высокое содержание пыльцы Classopollis во многих регионах
- рис. 21-27). Пояс дождевых экваториальных лесов не был развит. По обе стороны
от экватора
в это время, а иногда даже в непосредственной близости от него отлагались эвапориты
(север Южной Америки, северо-восточная Африка, рис. 29). Несмотря на многие
признаки засушливости, в пределах этого пояса встречаются остатки динозавров,
изредка - теплолюбивых хвойных растений, стволы цикадофитов, единичные местонахождения
папоротников, насекомых, каолинитов. С апта в центральных частях Западной Гондваны
встречаются остатки крокодилов (рис. 29). Часть этих находок связана с внутриконтинентальными
областями суперконтинента. Поэтому следует полагать, что они не были совсем безжизненными
и, по-видимому, в них встречались оазисы. Наиболее значительные признаки увлажнения
тяготеют, однако, к прибрежным районам пояса. Так, на севере Аравийского п-ва
в барреме местами накапливались угли (рис. 27), а в апте - бокситы и железные
руды [Цеховский и др., 1995]. Эти местонахождения указывают на существование
области гумидного экваториального климата на подветренном восточном океаническом
побережье Западной Гондваны (рис. 29). По-видимому, это был зародыш будущего
экваториального пояса.
В морях и океанах тропическо-экваториального пояса большое распространение
имели отложения карбонатных платформ, барьерные рифы, черные сланцы (Северная
и Южная Атлантика [Жарков и др., 1998а, б]). На карбонатных платформах и рифах
Средиземноморского Тетиса в условиях островного климата сформировались многочисленные
месторождения карстовых бокситов [Цеховский и др., 1995]. В морях пояса обитала
богатая и теплолюбивая фауна. Для многих групп морских беспозвоночных (двустворок,
аммонитов, раннеаптских белемнитов, кораллов, фораминифер и др.) [Beauvais,
1992; Hallam, 1973; Doyle, 1992; Koutsoukos, 1992 и др.] тропически-экваториальный
пояс в значительной мере совпадает с Тетическим биогеографическим царством.
Начиная с альба, по мере зарождения и развития экваториального гумидного пояса
в Западной Гондване (см. следующий раздел), огромный
единый тропически-экваториальный аридный пояс распался на два тропических аридных
пояса - северный и южный (рис.
33 - 43). К первому относятся области эвапоритового осадконакопления на юге
Северной Америки, в Северной Африке и в Юго-Восточной Азии, а к южному - области
развития эвапоритов и красноцветов в Южной Америке, Центральной и Восточной
Африке и эпизодически в Индии. Следы альбского южного аридного пояса обнаружены
и на Тихоокеанской плите, располагавшейся тогда в низких южных широтах почти
в центре Тихого океана. Здесь на вулканическом основании развивались коралловые
атоллы с признаками периодически осолонявшихся лагун [Жарков и др., 1995].
Оба пояса хорошо трассируются по распространению споро-пыльцевых комплексов
с высоким (> 50%) содержанием пыльцы классополлис [Вахрамеев, 1988]. В пределах
континентов эти пояса располагались асимметрично по отношению к экватору.
Южный аридный пояс был на протяжении альб-коньякского интервала шире северного
и располагался дальше от экватора. Позднее они более или менее сравнялись по
ширине, но некоторое смещение южного аридного пояса к югу сохранилось. На юго-восточном
подветренном побережье Африки в Маастрихте в пределах южного аридного пояса
наряду с гипсоносными красноцветами местами формировались каолинитовые и латеритовые
коры выветривания. Это свидетельствует, очевидно, о периодическом формировании
здесь муссонного климата.
Экваториальный гумидный пояс. Как уже отмечалось в предыдущем
разделе статьи, в неокоме и апте экваториального гумидного пояса еще не существовало
и вблизи
экватора локальные признаки гумидного климата улавливаются только на северо-востоке
Аравии. В альбе признаки влажности в приэкваториальной зоне становятся более
многочисленными. На северо-западе Южной Америки известны альбские палинологические
спектры, содержащие лишь небольшое количество пыльцы Classopollis [Вахрамеев,
1988], а на северо-востоке Африки устанавливается зажатый между областями эвапоритовой
седиментации, еще довольно узкий, гумидный пояс, который прослеживается по
местонахождениям бокситов, каолинитовых пород и железных руд (рис. 31) [Цеховский
и др., 1995]. В сено-манский век экваториальный гумидный пояс расширяется и
трассируется по местонахождениям каменных углей, бокситов, каолинитовых пород
и железных руд от Аравии через Центральную и Западную Африку до северовосточных
районов Ю. Америки (рис. 13). Одновременно у западного побережья Экваториальной
Африки в палинологических спектрах отмечается значительное сокращение количества
пыльцы Classopollis - индикатора аридности. Это тоже указывает на
увлажнение климата [Вахрамеев, 1988]. По окраинам Западно-гондванских материков
и в Юго-Восточной
Азии в это время заметно сокращаются области эвапоритового осадконакопления,
хотя формирование красноцветов продолжалось в многочисленных континентальных
впадинах Африки, Южной Америки, Китая и Монголии [Жарков и др., 1998 б]. В
туроне экваториальный гумидный пояс прослеживается еще отчетливее, чем раньше,
по угленосным отложениям, бокситам, каолинитовым породам и железным рудам в
Северной и Западной Африке. Из спорово-пыльцевых комплексов Экваториальной
Африки в это время практически исчезает пыльца Classopollis. Именно в это время,
по мнению В.А. Вахрамеева [1988],
возник пояс влажных тропических лесов. Представляется, однако, что пояс мог
возникнуть еще в альбе-сеномане, в виде цепи крупных оазисов, когда на севере
центральной Африки широкое развитие получили латеритные и каолиновые коры
выветривания и началось угленакопление.
Становление экваториального гумидного климата в виде глобального широтного
пояса шло в Западной Гондване параллельно с раскрытием Южной Атлантики и прогрессивным
сокращением и раздроблением областей аридной седиментации. Представляется,
что процессы раскрытия Южной Атлантики и возникновения экваториального гумидного
климата были взаимосвязаны, поскольку в центре Западной Гондваны появлялся
новый огромный бассейн - источник влаги со своими при-океаническими зонами.
Одновременно поперечные размеры массивов суши в широтном направлении сократились
почти вдвое и стали проходимы для пассатных ветров, приносивших влагу. При
раскрытии Южно-Атлантического океана, по-видимому, гумидные приоке-анические
области образовались сначала на его берегах. Затем по мере расширения океана
они росли, сливались с возникшими ранее, внутриконтинентальными оазисами и
к сено-ману образовали более или менее единый пояс. В альбе-сантоне экваториальный
пояс располагался почти симметрично по отношению к экватору, а его ширина колебалась
от 10° до 20°. В кампанский век произошло заметное его смещение к югу и одновременное
в связи с общим потеплением расширение до 25°-30° (рис. 41). Раскрытие Южно-Атлантического
океана существенно изменило также циркуляцию и вертикальную структуру водной
толщи Атлантики и, как полагают некоторые, способствовало глобальному среднемеловому
потеплению на Земле [Poulsen et al., 2003].
Для неокома вообще и для неокома экваториальных широт, в частности, имеется
довольно мало кислородно-изотопных определений палеотемператур морских вод.
Значительно более многочисленны они для конца раннего и позднего мела. По сводным
данным в неокоме, апте и альбе приэкваториальные широты характеризовались температурами
от 25° до 30 °С, в раннем сеномане они резко снизились до 18-24 °С, а затем
снова возросли до 26 °С и более, а существенно стали снижаться только в маастрихте,
когда опустились до 17-19°С [Frakes, 1999]. Для позднего альба - начала сеномана
эти цифры подтверждаются новейшими детальными исследованиями в районе Саргассова
моря, которые показали, что температуры поверхностных вод колебались между
25° и 30 °С [Wilson, Norris, 2001]. Возможно, в позднем мелу они временами
достигали даже 28-32 °С [Pearson et al., 2001; Huber et al., 2002].
Эти определения хорошо согласуются между собой, но заметно расходятся с более
ранними данными, согласно которым повышение температур в средних и высоких
широтах сопровождалось их снижением в приэкваториальных широтах [Macleod et
al., 2000]. Считается, что такое занижение температур было связано с недостаточным
учетом диагенитических изменений в анализируемых материалах [Pearson et al.,
2001] и солевого состава морских вод [Huber et al., 2000]. Температура донных
вод в приэкваториальных широтах в конце мелового периода определяется для кампана
в 7°-11 °С и для маастрихта в 6°-9 °С [Banera, Savin, 1999].
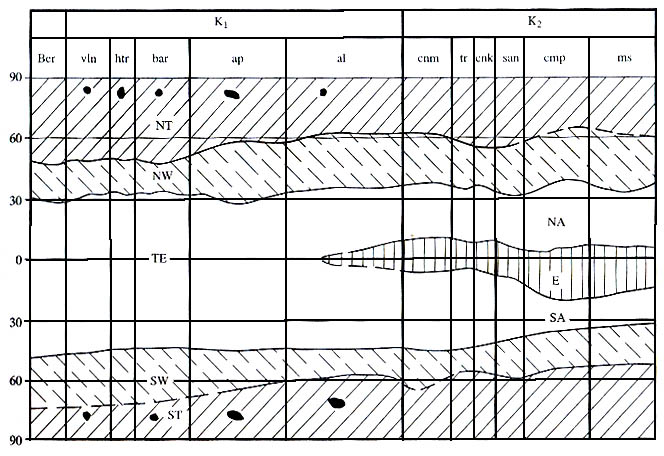
Рис. 46. Перестройки климатической зональности в меловой период
NT, ST - северный и южный высокоширотные умеренные пояса; черные включения -
следы сезонного ледового разноса обломочного материала; NW, SW - северный и
южный средне-
широтные теплые пояса; ТЕ - тропико-экваториальный аридный пояс; NA, SA - северный
и южный аридные пояса; Е - экваториальный пояс. |
Перестройки климатической зональности. Изменения в характере
и положении палеоклиматических поясов, вытекающие из сравнения рассмотренных
реконструкций, схематично суммированы
на рис. 46. Последний иллюстрирует, что в меловом периоде, длившемся около
80 млн лет, отчетливо различаются два временных интервала, почти равной
продолжительности, которые характеризовались разными типами климатической зональности.
В интервале, который продолжался от бе-риасского до начала альбского века,
отсутствовал экваториальный гумидный пояс. Поэтому северный и южный аридные
пояса слились в один огромный пояс. Средняя ширина его была около 80°-85°.
Этот тропическо-эквато-риальный пояс, охватывал все низкие и часть средних
широт. Океан Мезотетис, располагавшийся в низких широтах между двумя крупными
массивами суши (Евразией - на севере и Восточной Гондваной - на юге) создавал
благоприятные условия для возникновения мус-сонного климата. Однако проникновению
муссонов в краевые части южной и юго-восточной Евразии, по-видимому, препятствовали
горные цепи вулканических поясов, которые окаймляли активные окраины этого
континента. В их дождевой тени возникали многочисленные гипсоносные бассейны,
протягивавшиеся от Иберии до Китая. Аридность восточной Африки обуславливалась
существованием двух летних областей низкого давления над Восточной Евразией
и Восточной
Гондваной, которые отклоняли траектории тропических муссонов, соответственно
к северу и югу. Как уже отмечалось, в приэкваториальной зоне местное увлажнение
отмечается только на севере Аравийского блока. Сухость внутренних районов Африки
и Южной Америки объясняется их удаленностью от океанов.
С обеих сторон тропическо-экваториальный пояс окаймлялся теплыми среднеширотными
поясами переменной ширины (от 10° до 30°), которые в своих океанических секторах
и областях развития эпиконтинентальных внутренних морей имели отчетливо гумидный
климат. Он обуславливался тем, что в средних широтах перенос влаги и движение
циклонов происходили в восточных румбах. Кроме того, на их пути в северном
полушарии располагался ряд дополнительных источников влаги в виде эпиконтинентальных
морей, а в южном полушарии - также океанических бассейнов. Достаточное количество
тепла и влаги в среднеширотных поясах позволяло формироваться каолинитовым
и латеритовым корам выветривания, а в приокеанических секторах и приморских
областях благоприятствовало угленакоплению. В южные, удаленные от океанов
и морей внутриконтинентальные области Евразии, влаги с запада поступало меньше.
Муссоны проникали сюда в ослабленном виде, поэтому здесь климат был семигумидным
и характеризовался сезонным, хотя и достаточным увлажнением (см. гл.
4).
Высокие и частично средние широты в раннем мелу занимали высокоширотные умеренные
пояса. Особенно значительным по своим размерам был северный умеренный пояс.
Его ширина порой достигала 50°. Южный умеренный пояс в неокоме был значительно
более узким, чем северный, и выделялся в основном по присутствию следов сезонного
ледового разноса. Значительную часть высокоширотных умеренных поясов, особенно
в приокеанических секторах и приморских областях, занимали смешанные листопадные
широколиственно-хвойные леса, часто заболоченные, в которых шло интенсивное
углеобразование. Климат этих поясов был сезонным, в целом умеренный, с достаточно
мягкими зимами. Вблизи границ со среднеширотными поясами климат приближался,
по-видимому, к умеренно-теплому. Наоборот, в зонах, расположенных ближе к обоим
полюсам, моря, озера и реки в первую половину мелового периода иногда покрывались
льдом, что указывает на эпизодическое распространение умеренно-холодного климата.
В альбский век на Земле произошла перестройка климатической зональности. Во-первых,
зародился экваториальный гумидный пояс, который разделил тропическо-экваториальный
пояс на два аридных пояса, северный и южный. На протяжении позднего мела этот
пояс постепенно расширялся и несколько смещался к югу. Возникновение экваториального
гумидного пояса, как отмечалось [Жарков и др., 1995; Чумаков и др., 1995],
совпало с началом раскрытия южной части Атлантического океана. Дальнейшее его
расширение и смещение к югу шло параллельно с продолжавшимся расширением этой
части Атлантического океана. Окаймлявшие экваториальный
гумидный пояс северный и южный аридные пояса при этом сужались и дробились.
Особенно заметно сузился
южный аридный пояс. Среднеширотные теплые пояса во вторую половину мелового
периода немного расширились. Высокоширотные умеренные пояса также претерпели
изменения: северный заметно сузился и стал охватывать только высокие широты,
а южный, немного расширившись, также распространился на все высокие широты.
Примечательно, то с начала позднемеловой эпохи следы сезонного ледового разноса
в высокоширотных поясах перестали встречаться*. В общем, климатические пояса
после перестройки зональности в альбском веке стали располагаться более симметрично
по отношению к экватору. Суммарное увеличение площади теплых среднеширотных
поясов, небольшое сокращение площади высокоширотных поясов, а главное исчезновение
в них следов ледового разноса говорит о потеплении в середине мелового периода.
Почти одновременное появление и расширение экваториального гумидного пояса
и сужение аридных поясов указывают на то, что глобальный климат в эту эпоху
стал не только более теплым, но и более влажным.
* По устному сообщению А. Альберга, маленький валун был встречен в маастрихтских
глинистых породах в одном из обнажений на р. Гребенка (правый приток р.
Анадырь). А. Альберг склоняется к тому, что этот валун был принесен льдом.
Нам представляется, что ничтожная концентрация камней в этих отложениях
скорее свидетельствует в пользу их биогенного разноса. |
Разрешающая способность реконструкций климатической зональности определяется
продолжительностью тех стратиграфических подразделений, для которых они составляются.
В нашем случае это ярусы мелового периода, длительность которых колеблется
от 2 до 15 млн лет, составляя в среднем 6,75 млн [Harland et al., 1990]. Таким
образом, рассмотренная серия реконструкций отражает лишь медленные климатические
изменения, длительностью несколько млн лет и больше.
Более короткие климатические изменения на этих реконструкциях осредняются.
Поэтому при анализе реконструкций климатической зональности, составленных по
ярусам и эпохам, в том числе и для юрского периода [Rees etal., 2000], создается
впечатление о большой устойчивости климатической зональности. В действительности,
при детальном изучении и сопоставлении нескольких опорных разрезов выявляются
многочисленные более короткие климатические колебания, которые рассматриваются
нами в гл. 7.
5.2. Глобальный климат мелового периода
Совокупность приведенных выше палеоклиматических данных
свидетельствует о том, что на протяжении всего мелового периода глобальный
климат Земли был безледниковым,
а многолетняя гляциосфера отсутствовала. На это указывают три фундаментальных
факта.
Во-первых, существование поясов умеренного климата в обеих полярных областях
Земли. Это согласованно подтверждается литологическими, палеонтологическими
и геохимическими данными.
Во-вторых, отсутствие каких-либо материальных следов меловых ледниковых покровов,
многолетней мерзлоты или постоянных ледовых шапок. Следами последних не могут
считаться раннемеловые отложения с рассеянными камнями ("дропстоунами"),
которые единодушно квалифицируются как результат сезонного ледового или биогенного
разноса.
В-третьих, меловой океан был теплым. В нем отсутствовала психросфера - толща
холодных глубинных и придонных вод, формирующаяся в полярных областях и заполняющая
все океаны от дна до глубины 500-1000 м. Температура психросферы приблизительно
соответствует средней температуре воды в высоких широтах, поэтому температура
глубинных и придонных вод в современных океанах колеблется от -0,5° - +5 °С,
в меловых же океанах она составляет от 7°-11 °С до 20 °С, и лишь в маастрихте
снижалась до 5°-8 °С [Barrera, Savin, 1999; Huber et al., 2002 и др.].
Основные выводы, которые сделаны из рассмотренных выше данных о существовании
безледникового климата в мелу, характере климатической зональности, положении
климатических поясов и их климатических параметрах в целом хорошо согласуются
с последними результатами математического моделирования меловых климатов (см.
гл. 2).
Таким образом, не вызывает сомнений, что глобальный климат мелового периода
был безледниковым и следовательно меловая биосфера была теплой. Вопрос сейчас
состоит в том, в каких пределах климат мелового периода мог варьировать. Совершенно
очевидно, что он не был единообразным на протяжении всего мелового периода
и характеризовался трендом и, как будет показано в гл.
7, осцилляциям нескольких
рангов, однако при всех этих изменениях климатическая система Земли явно не
пересекала нижний порог, за которым начинали возникать ледники. Предположения
о возникновении ледниковых покровов в течение мелового периода и их осцилляциях
основаны на косвенных и достаточно спорных соображениях.
Имеющимся фактам более соответствует выделение на примере мела нескольких градаций
безледникового климата: "жаркого безледникового" (hot greenhouse), "теплого
безледникового" (warm greenhouse) и "прохладного безледникового" (cool
greenhouse), как это предлагает Б. Хубер с соавт. [Huber et al., 2002]. Основываясь
на изотопно-кислородных палеотемпературах среднебатиальных вод, они считают,
что с позднего альба до позднего сеномана климат был "теплым безледниковым" (усредненная
температура этих вод -16 °С), с позднего сеномана до раннего кампана климат
был "жарким безледниковым" (температура этих вод около 18°-19 °С),
а со среднего кампана до конца маастрихта - "прохладным безледниковым" (температура
этих вод 9°-11 °С). В общих чертах эти результаты согласуются со многими
приведенными выше палеоклиматическими данными, которые тоже свидетельствуют
о потеплении
в конце раннего мела, наиболее теплом климате в середине мелового периода и
похолоданиях в конце. В то же время отделить теплую градацию от жаркой другими
методами, кроме изотопных по глубоководным разрезам, весьма затруднительно.
Не слишком четко эти две градации различаются и по данным Б. Хубера с соавт.,
тем более что обе градации осложнены многочисленными осцилляциями, а изотопный
разрез состоит из ряда фрагментов. Необходимость выделять в безледниковом климате
градации назрела и очень интересное предложение Б. Хубера с соавт. является
хорошим доказательством тому. Но представляется, что практическая реализация
этого предложения дело будущего, а сейчас, видимо, реально выделять в глобальном
безледниковом климате две градации: теплого безледникового и прохладного
безледникового климатов. Это предложение ближе к тому, которое
сделал Л. Фрейкс с соавт. [Frakes et al., 1992], различая в мезозое теплые
интервалы (warm modes) и среднемезозойский прохладный интервал (cool mode).
В последнем, по представлениям Л. Фрейкса с соавт., в отличие от остальных
ледниковых "cool mode", тоже не было ледников, а только временами
в высоких широтах образовывались сезонные льды. Рассмотренная выше серия реконструкций
меловой климатической зональности позволяет произвести дальнейшее подразделение
выделенных градаций безледникового климата по степени развития гумидных и аридных
поясов. В пределах суши, занимаемая ими доля, в течение мелового периода довольно
сильно изменялась. В апте гумидные пояса занимали 55% суши, аридные 45%; в
Маастрихте - соответственно 74 и 25%. В первом случае их соотношение составляло
1,2, а во втором - почти 3. Если условно принять современное отношение площади
гумидных поясов к аридным (около двух) за границу между аридным и гумидным
глобальным климатами, то можно сказать, что в первом случае глобальный климат
был аридным прохладным безледниковым, а во втором - гумидным прохладным
безледниковым.
Начиная с альба, на Земле стала заметно возрастать роль поясов гумидного климата
и поэтому в середине мела глобальный климат стал гумидным теплым безледниковым.
5.3.
Климат и биотические события
Данные о биосферных событиях в меловой период были кратко
изложены в гл. 2. В этом разделе мы хотим отметить совпадение некоторых климатических
и биотических
событий между собой и высказать предположения о их возможной связи.
Прежде всего, обращает на себя внимание, что, если исключить кризис на границе
мела и палеогена, на протяжении весьма длительного временного интервала позднего
мезозоя (145 млн лет) и в том числе в меловом периоде (более 80 млн лет) в
морской биоте не было массовых вымираний. В меловом периоде произошло одно
вымирание второго порядка вблизи границы сеномана и турона. В то же время к
этому периоду приурочены сравнительно многочисленные слабые биотические события
третьего и четвертого порядка [Walliser, 1995]. Такое распределение контрастирует
с ледниковыми интервалами фанерозоя, к которым, преимущественно к их окончанию,
приурочены редкие, но крупные биотические события. Это наводит на мысль о
том, что в теплой биосфере климатические изменения имели меньшее влияние на
биоту, чем в холодной, и преобладали события, связанные с ее естественным эволюционным
развитием. Тем не менее, совпадение во времени ряда климатических и биотических
событий позволяет предполагать между ними существование причинно-следственных
связей.
Так, например, значительное сокращение разнообразия в раннем готериве Арктического
бассейна [Zakharov, 1992; Захаров и др., 2002] происходило в интервале широкого
сезонно-ледового разноса в этом регионе (рис. 45).
Сравнительно небольшое верхнеальбское амкинское похолодание совпадало с чрезвычайно
быстрой экспансией покрытосеменных. В этом случае климатические изменения,
по-видимому, послужили толчком (непосредственной причиной) для начала событий,
подготовленных эволюционными процессами. Предполагается, что похолодание освободило
экологические ниши для покрытосеменных [Лебедев, 1993].
Значительное снижение в конце сеномана - начале турона разнообразия морской
фауны, в том числе планктонных фораминифер (гл. 6, рис. 47, см. вклейку
III),
произошло на фоне быстрого потепления, а затем похолодания. По-видимому, это
событие второго порядка (Walliser, 1995] было обусловлено климатическими изменениями,
поскольку динамика этих событий была аналогичной.
Кризису на границе мела и палеогена предшествовало длительное, осложнявшееся
осцилляциями, неравномерное похолодание и сокращение таксономического разнообразия
ряда групп морской фауны в конце позднего мела [Алексеев, 1998]. Можно полагать,
что длительные климатические изменения в данном случае подготовили условия
для возникновения биотического кризиса, ослабив меловую биоту. Возможно, что
параллельно подготавливали кризис и эволюционные новации. Непосредственными
же причинами кризиса стали совсем другие события, в том числе и импактные.
Роль триггера в этом кризисе возможна была не столь большой, как представляется.
В открытых сложных системах, находящихся в квазиравновесном состоянии, как
биосфера, кризис может быть результатом сравнительно небольших, но длительно
накапливающихся изменений (бифуркаций).
В мезозое, в том числе в меловом периоде, происходили и бескризисные биотические
перестройки, связанные с постепенными климатическими изменениями. Например,
описанная в гл. 6 экспансия планктонных фораминифер,
была очевидно связана с мезозойским, потеплением и последующей меловой гумидизацией
климата на Земле. Эти климатические изменения обеспечили благоприятный тепловой
режим и повышенный континентальный сток питательных веществ в моря, которые
способствовали дивергенции и пространственной экспансии планктонных фораминифер.
Выводы
Реконструкции палеоклиматической зональности для веков мелового
периода дают представление о эволюции климата на протяжении 80 млн лет и позволяют
сделать
следующие выводы.
1. На протяжении мелового периода глобальный климат был безледниковым, что
подтверждается как характером климатической зональности, так и отсутствием
психросферы в океанах.
2. Высокие широты обоих полушарий Земли характеризовались в этот период гумидным
умеренным и частично умеренно-теплым климатом.
3. Средние широты обоих полушарий занимали гумидные теплые климатические пояса.
4. В раннемеловую эпоху до альбского века низкие широты обеих полушарий Земли
занимал единый аридный жаркий пояс.
5. В альбском веке в приэкваториальной зоне возник гумидный жаркий экваториальный
пояс, который постепенно расширяясь, прослеживается до конца мелового периода.
6. Начиная с альбского века площадь гумидных поясов на Земле расширялась.
Это в значительной степени было связано с раскрытием Южно-Атлантического и
Индийского океанов и с эпиконтинентальными трансгрессиями.
7. Безледниковый климат может быть сейчас подразделен на две градации: теплый
безледниковый климат и прохладный безледниковый климат. В каждой из этих градаций
могут различаться гумидная и аридная разновидности.
8. Безледниковый климат оказывал огромное воздействие на характер меловой биоты
и определял ее высокую продуктивность. Пояса наибольшей продуктивности располагались
в средних и высоких широтах.
9. За исключением массового биотического кризиса на границе мела и палеогена,
а также менее крупных событий в готериве и у границы сеноман-турона, в меловом
периоде отмечается целый ряд сравнительно небольших биотических событий, очевидно
эволюционной природы. Некоторые биотические события происходили одновременно
с климатическими событиями, что, возможно, указывает на их следственно-причинную
связь. Наряду
с этим на биоту оказывали воздействие медленные изменения климата.
Литература
Алексеев А.С. Массовые вымирания в фанерозое. М.: Изд-во
МГУ, 1998. 78 с.
Буданцев Л.Ю. Фитостратиграфические комплексы позднего мела Лено-Вилюйского
и Чульшо-Енисейского бассейна как основа для межрегионального сопоставления
континентальных отложений // Стратиграфия нижнемеловых отложений нефтегазоносных
областей СССР. Л.: Недра, 1979. С. 149-162.
Вахрамеев В.А. Климаты северного полушария в меловом периоде и данные палеоботаники
// Палеонтол. журн. 1978. № 2. С. 3-17.
Вахрамеев В.А. Фитогеография, палеоклиматы и положение материалов в мезозое
// Вести. АН СССР. 1985. № 8. С. 30-42.
Вахрамеев В А. Юрские и меловые флоры и климаты Земли. М.: Наука, 1988. 214
с.
Герман А.Б. Этапность и цикличность развития позднемеловой флоры Анадырско-Корякского
субрегиона // Стратиграфия. Геол. корреляция. 1993. Т. 1, № 1. С. 87-108.
Герман А.Б. Разнообразие меловых платанообразных Анадырско-Корякского
субрегиона в связи с изменением климата // Там же. 1994. Т. 2, № 4/5. С. 62-77.
Гольберт А.В. Основы региональной палеоклиматологии. М.: Недра, 1987. 222 с.
Жарков М.А., Мурдмаа И.О., Филатова Н.И. Палеогеография середины мелового периода
// Стратиграфия. Геол. корреляция. 1995. Т. 3, № 3. С. 15-41.
Жарков М.А., Мурдмаа И.О., Филатова Н.И. Палеогеография берриас-берремского
веков раннего мела // Там же. 1998а. № 1. С. 49-72.
Жарков М.А., Мурдмаа И.О., Филатова Н.И. Палеогеография коньяк-маастрихтского
времени позднего мела // Там же. 19986. № 3. С. 3-16.
Захаров Ю.Д., Игнатьев А.И., Борискина Н.Г. и др. Палеотемпературная кривая
для позднего мела Северо-Западной Пацифики (Хоккайдо, Сахалин, Восточная Корякия)
//Тихоокеан. геология. 2001. Т. 20, № 1. С. 15-24.
Захаров В.А., Шурыгин Б.Н., Курушин Н.И. и др. Мезозойский океан в Арктике:
Палеонтологические свидетельства // Геология и геофизика. 2002. Т. 43, № 2.
С. 155-181.
Каплан М.Е. Кальцитовые псевдоморфозы (псевдогейлюссит, яровит, тинолит,
глендонит, генойши, беломорские рогульки) в осадочных породах: Происхождение
псевдоморфоз
// Литология и полез, ископаемые. 1979. № 5. С. 125-142.
Константин Э., Чипсами А., Викерс-Рич П. и др. Перигляциальные обстановки и
полярные динозавры // Палеонтол. журн. 1999. № 2. С. 69-75.
Красилов В.А. Меловой период: Эволюция земной коры и биосферы. М.: Наука, 1985.
240 с.
Лебедев Е.Л. Амкинское похолодание и развитие флор на рубеже раннего
и позднего мела // Стратиграфия. Геол. корреляция. 1993. Т. 1, № 2. С. 78-84.
Мейен С.В. Основы палеоботаники, М.: Недра, 1987. 403 с.
Несов Л.А. Значение смен вертикальной циркуляции в океане для исторических
перемен в планетарном балансе тепла, региональных климатах и биосе // Вести.
СПб. ун-та. Сер. 7. 1992. Вып. 1. С. 3-10.
Несов Л.А. Неморские позвоночные мелового периода Северной Евразии. СПб.: Ин-т
земной коры СПб. ун-та, 1997.218с.
Несов Л.А., Головнева Л.Б. История развития флоры, фауны позвоночных и климата
в позднем сеноне на северо-востоке Корякского нагорья // Континентальный мел
СССР. Владивосток: ДВО АН СССР, 1990. С. 191-212.
Раутиан А.С. Эвристическая модель становления основ гомойотермии // X Всесоюз.
совещ. по эволюц. физиологии: Тез. докл. Л.: Наука, 1990. С. 31-32.
Ронов А.Б., Балуховский А.Н. Климатическая зональность материков и общая тенденция
изменения климата в позднем мезозое и кайнозое // Литология и полез, ископаемые.
1981. № 5. С. 118-136.
Синицын В.М. Введение в палеоклиматологию. Л.: Недра, 1980. 248 с.
Спайсер Р.Э., Герман А.Б. Меловой климат Азии и Аляски: Сравнение палеоботанических
свидетельств с компьютерной моделью // Палеонтол. журн. 1998. № 2. С. 3-18.
Степанов В.Н., Агапитов Е.Н., Гриценко A.M. Термогалинная стратификация вод
Мирового океана. М.: Наука, 1984. 140 с.
Татаринов Л.П. Очерки по теории эволюции. М.: Наука, 1987. 251 с.
Филатова Н.И. Эволюция меловых обстановок на Северо-Востоке Азиатского континента
// Стратиграфия. Геол. корреляция. 1995. Т. 3, № 3. С. 64-75.
Цеховский Ю.Г., Щипакина И.Н., Храмцов И.Н. Латеритный аллювий и продукты
его переотложения -важные индикаторы палеоклиматов апт-туронского времени //
Там же. 1995. Т. 3, № 3. С. 89-99.
Черная Т.А., Бугрова Э.М., Круговых В.В. Разнообразие ранее неизвестной
позднемеловой-палеогеновой биоты Восточной Сибири // Биоразнообразие в истории
Земли: Тез. докл. 67-й
сес. Палеонтол. о-ва. СПб.: ВСЕГЕИ, 2001. Р. 98-100.
Чумаков М.А., Жарков М.А., Герман А.Б. и др. Климатические пояса в середине
мелового периода // Стратиграфия. Геол. корреляция. 1995. Т. 3, № 3. С. 42-63.
Ясаманов Н.А. Древние климаты Земли. Л.: Гидро-метеоиздат, 1985. 295
с.
Barrera E., Savin S.M. Evoluton of the Campanian-Maastrichtian marine climates
and oceans // Geol. Soc. Amer. Spec. Pap. 1999. N 332. P. 245-272.
Barren E.J., Washington W.M. Cretaceous climat: Comparision of atmospheric
simulations with the geologic record // Palaeogeogr., Palaeoclimatol., Palaeoecol.
1982. Vol. 40. P. 103-133.
Beauvais L. Palaeobiogeography of the Early Cretaceous corals //Ibid. 1992.
Vol. 92, N 3/4. P. 233-247.
Brouwers E.M., Clemens W.A., Spicer R.A. et al. Dinosaurs on the North Slope,
Alaska: high latitude, Latest Cretaceous environments // Science. 1987. Vol.
237, N 4822. P. 1608-1610.
Clarke L.J., Jenkyns H.C. New oxygen isotope evidence for long-term Cretaceous
climatic change in the Southern Hemisphere // Geology. 1999. Vol. 27, N 8.
P. 699-702.
Clemens W.A., Nelm L.G. Paleoecological implications of Alaskan terrestrial
vertebrate fauna in Latest Cretaceous time at high paleolatitudes // Ibid.
1993. Vol. 5, N 6. P. 503-506.
Detterman RJL., Reiser H.N., Brosge W.P., Dutro J.T. Post-Carboniferous stretigraphy,
Northeastern Alaska // Geol. Surv. Professional paper 886. 1975. 46 p.
Ditchfield P.W. High northern palaeolatitude Jurassic-Cretaceous palaeotemperature
variation: New data from Kong Karls Land, Svalbard // Palaogeogr., Palaeoclimatol.,
Palaeoecol. 1997. Vol. 130. P. 163-175.
Ditchfield P.W., Marshall J.D., Pirre D. High latitude palaeotemperature variation:
New data from the Tithonian to Eocene of James Ross Island, Antarctica // Ibid.
1994. Vol. 107, N17. P. 79-101.
Doyle P. A review of the biogeography of Cretaceous belemnites //Ibid. 1992.
Vol. 92, N 3/4. P. 207-216.
Falcon-Lang H.J., Cantrill D.J. Leaf phenology of some Mid-Cretaceous
polar forests, Alexander Island, Antarctica // Geol. Mag. 2001. Vol. 138, N
1. P.
39-52.
Falcon-Lang H.J., Cantrill D.J., Nichols G.J. Biodiversity and terrestrial
ecology of a Mid-Cretaceous, high-latitude floodplain, Alexander Island, Antarctica
// J. Geol. Soc. London. 2001a. Vol. 158. P. 709-724.
Falcon-Lang H.J., Kvacek J., Ulicny D. Fire-prone plant communities and paleoclimate
of a Late Cretaceous fluvial to estuarine environment, Pecinov Guarry, Czech
Republic // Geol. Mag. 2001b. Vol. 138, N 5. P. 563-576.
Ferguson KM., Gregory R.T., Constantine A. Lower Cretaceous (Aptian-Albian)
secular changes in the oxygen and carbon isotope record from high paleolatitude
fluvial sediments, Southeast Australia: Comparisions to the marine record //
Geol. Soc. Amer. Spec. Pap. 1999. N 332. P. 59-72.
Frakes L.A. Estimating the global thermal state from Cretaceous sea
surface and continental temperature data//Ibid.
1999. N332. P. 49-56.
Frakes L.A., Alley N.F., Deynoux M. Early Cretaceous ice rafting and
climate zonation in Australia // Intern. Geol. Rev. 1995. Vol. 37. P. 567-583.
Frakes L.A., Francis I.E. Cretaceous palaeoclimates // Cretaceous resources,
events and rhythms / Ed. R.N. Ginsburg and B. Beaudoin. Dordrecht: Kluwer,
1990. P. 273-287.
Frakes L.A., Francis J.E., Syktus J.I. Climate modes of the Phanerozoic. Cambridge:
Cambridge Univ. press, 1992. 274 p.
Francis J.E., Poole I. Cretaceous and Tertiary climates of Antarctica; Evidence
from fossil wood // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2002. Vol. 182.
P. 47-64.
Gregory R.T., Douthitt C.B., Duddy I.R. et al. Oxygen isotopic composition
of carbonate concretions from the Lower Cretaceous of Victoria, Australia:
Implication for the evolution of meteoric waters on the Australian continent
in a paleopolar environments // Earth and Planet. Sci. Lett. 1989. Vol. 92.
P. 27-42.
Hallam A. (ed.). Atlas of palaeobiogeography. Amsterdam etc.: Elsevier, 1973.
672 p.
Hallam A. A review of Mesozoic climates // J. Geol. Soc. London. 1985. Vol.
142. P. 433-445.
Harland W.B., Armstrong R.L., Cox A.V. et al. A geologic time scale,
1989. Cambridge: Cambridge Univ. press, 1990. 263 p.
Horner J.R. Behavior // The encyclopedia of dinosaurs // Ed. P. Currie, V.
Padian. N.Y. etc.: Acad. press, 1997. P. 45-50.
Huber B.T., Macleod K.G., Wing S.L. (ed.). Warm climates in Earth history.
Cambridge: Cambridge Univ. press,
2000. 452 p.
Huber B.T., Norris R.D., MacLeod K.G. Deep-sea paleotemperature
record of extreme warmth during the Cretaceous // Geology. 2000. Vol. 30, N
2. P. 123-126.
Kelly S.RA., Moncrieff A.C.M. Marine molluscan constraints on the age of Cretaceous
fossil forests of Alexander Island, Antarctica // Geol. Mag. 1992. Vol. 129,
N 6. P. 771-778.
Kennedy E.M. A palaeoclimate perspective on two Cretaceous fossil
from New Zealand // Чтения памяти B.A. Вахрамеева. М.: Геос, 1996. С. 34-37.
Kennedy E.M., Spicer R.A., Rees P.M. Quantitative palaeoclimate estimates from
Late Cretaceous and Paleocene leaf floras in the northwest of the South Island,
New Zealand // Palaeogeogr., PalaeoclimatoL, Palaeoecol. 2002. Vol. 184, N3/4.
P. 321-345.
Koutsoukos E.A. Late Aptian to Maastrichtian foraminiferal biogeography and
palaeoceanography of the Sergipe Basin, Brazil // Ibid. 1992. Vol. 92, N 3/4.
P. 291-324.
Kowalski E.A. Mean annual temperature estimation based on leaf
morphology: Test from tropical South America // Ibid. 2002. Vol. 188. P. 141-165.
Krasilov V.A. Coal-bearing deposits of Soviet Far East // Controls on the distribution
and quality of Cretaceous coals / Ed. P.J. McCabe, J.T. Parrish. Boulder (Colo.),
1992. P. 263-267. (Geol. Soc. Amer. Spec. Pap.; N 267).
Larson R.L., Fisher A.G., Erba E., Premoli Silva I. (ed.). Apticore-Albicore:
A Workshop report on global events and rhythms of the Mid-Cretaceous, 4 - 9
October 1992. Perugia, 1993. 56 p.
Lloyd C.R. The Mid-Cretaceous Earth: Palaeogeography, ocean circulation
and temperature, atmospheric circulation // J. Geol. 1982. Vol. 90. P. 393-413.
Macleod K.G., Huber B.T., Ducharme M.L. Paleontological and geochemical constrains
on the deep ocean during the Cretaceous greenhouse interval // Warm climates
in Earth history / Ed. B.T. Huber et al. Cambridge: Cambridge Univ. press,
2000. P. 241-274.
Markwick P.J. Fossil crocodilians as indicators of Late Cretaceous
and Cenozoic climates: Implication for using pale-ontological data in reconstructing
palaeoclimate
// Palaeogeogr., PalaeoclimatoL, Palaeoecol. 1998. Vol. 137, N3/4. P. 205-271.
Masse J.P., Bellion Y., Benkhelil J. et al. Early Aptian (114 to 111 Ma) //
Atlas Tethys palaeoenvironmental maps / Ed. J. Dercourt et al. P.: Gauthier-Villars,
1993. P. 135-152.
McCarthy PJ., Martini IJ., Leckie D.A. Use of micromorphology
for palaeoenvironmental interpretation of complex alluvial palaeosols: An example
from the Mill Creek formation (Albian), southwestern Alberta. Canada // Palaeogeogr.,
PalaeoclimatoL, Palaeoecol. 1998. Vol. 143, N1/3. P. 87-110.
Molnar R.E., Wiffen J. A Late Cretaceous polar dinosaur fauna from New Zealand
// Cretaceous Res. 1994. Vol. 15, N 6. P. 689-706.
Mutterlose J., Kessels K. Early Cretaceous calcareous narmofossils from high
latitudes: Implications for palaeobio-geography and palaeoclimate // Palaeogeogr.,
PalaeoclimatoL, Palaeoecol. 2000. Vol. 160. P. 347-372.
Paul G.S. Physiology and migration of north slope dinosaurs // Proc.
Intern, conf. on Arctic margins, 1992. Anchorage: MMS, 1994. P. 405-418.
Parrish J.T., Ziegler A.M., Scotese C.R. Rainfall patterns and the distribution
of coals and evaporites in the Mesozoic and Cenozoic // Palaeogeogr., PalaeoclimatoL,
Palaeoecol. 1982. Vol. 40. P. 67-101.
Pearson P.N., Ditchfield P.W., Singdno J. et al. Warm
tropical sea surface temperatures in the Late Cretaceous and
Eocene epochs // Nature. 2001. Vol. 413, N 6855. P. 481-487.
Philip J., Babinot J.F., Tronchetti G. et al. Late
Cenomanian (94-92 Ma) // Atlas Tethys palaeoenvironmental
maps / Ed. I. Dercout et al. P.: Gauthier-Villars, 1993. P. 153-178.
Pickton C.A.G. Palaeogene and Cretaceous dropstones in
Spitsbergen // Earth's Pre-Pleistocene glacial record / Ed. M.J. Hambrey et al. Cambridge: Cambridge.
Univ. press, 1981. P. 567-569.
Price G.D., Ruffell A.H., Jones C.E. et al. Isotopic evidence for
temperature variation during the Early Cretaceous (Late Ryazanian-Mid-Hauterivian)
// J.
Geol. Soc. London.
2000. Vol. 157. P. 335-343.
Rees P.A., Ziegler A.M., Valdes P.J. Jurassic phytogeography and
climates: New data and model comparisions // Warm climates in Earth history
/ Ed. B.T.
Huber et al. Cambridge: Cambridge. Univ. press, 2000. P. 297-318.
Rich P.V., Rich Т.Н., Wagstaff B.E. Evidence for low temperatures
and biologic diversity in the Cretaceous high latitudes of Australia // Science.
1988. Vol.
242. P. 1403-1406.
Scherer C.M.S. Eolian dunes of the Botucatu Formation (Cretaceous) in southernmost
Brazil: Morphology and origin // Sediment. Geol. 2000. Vol. 137. P. 63-84.
Sellwood B.W., Valdes P.J., Price G.D. Geological evaluation of multiple
general circulation model stimulation of Late Jurassic palaeoclimate // Palaeogeogr.,
PalaeoclimatoL, Palaeoecol. 2000. Vol. 156, N 1/2. P. 147-160.
Spicer R.A., Corfield R.M. A review of terrestrial and marine climates in the
Cretaceous with implications for modelling the "Greenhouse Earth" //
Geol. Mag. 1992. Vol. 129, N 2. P. 169-180.
Spicer R.A., Parrish J.T., Grant P.R. Evolution of vegetation and coal-forming
environments in the Late Cretaceous of the North Slope of Alaska // Geol. Soc.
Amer. Spec. Pap. 1992. N 267. P. 177-192.
Spicer R.A., Rees P.McA., Chapman J.L. Cretaceous phytogeography and
climate signals // Philos. Trans. Roy. Soc. London. B. 1993. Vol. 341. P. 277-286.
Stevens G.R. Cretaceous belemnites // Atlas of palaeobiogeography
// Ed. A. Hallam. Amsterdam etc.: Elsevier, 1973. P. 385-401.
Tarduno J.A., Brinkman D.B., Renne P.R. et al. Evidence for extreme climatic
warmth from Late Cretaceous Arctic vertebrates // Science. 1998. Vol. 282,
N 18. P. 2241-2243.
Thorn V. Vegetation communities of a high palaeolatitude Middle Jurassic forest
in New Zealand // Palaeogeogr., PalaeoclimatoL, Palaeoecol. 2001. Vol. 168,
N 15. P. 273-289.
Walliser O.H. (ed.). Global events and event stratigraphy in Phanerozoic. В.:
Spinger, 1995. 335 p.
Wilson P.A., Norris R.D. Warm tropical ocean surface and global anoxia during
the Mid-Cretaceous period // Nature.
2001. Vol. 412, N 26. P. 425-428.
Wing S.L., Greenwood D.R. Fossils and fossil climate: The case for
equable continental interiors in the Eocene // Philos. Trans. Roy. Soc. London.
B.
1993. Vol. 341, N 1297. P. 243-252.
Wolfe J.F., Upchurch G.R., Jr. North American non-marine climates and vegetation
during the Late Cretaceous // Palaeogeogr., PalaeoclimatoL, Palaeoecol. 1987.
Vol. 61. P. 33-77.
Zakharov V.A. Climatic fluctuations and other events in the Mesozoic of Sibirian
Arctic // Proc. Intern, conf. on Arctic margins, 1992. Anchorage: MMS, 1994.
P. 23-28.
ОГЛАВЛЕНИЕ
Введение (Н.М. Чумаков).......................................................................................................................................
5
Часть I
ЭПОХА ПЕРЕХОДА ОТ ТЁПЛОЙ К ХОЛОДНОЙ БИОСФЕРЕ: КЛИМАТ ПАЛЕОГЕНА
Глава 1. Климат Земного шара в палеоцене
и эоцене по данным палеоботаники (М.Л. Ахметьев)............. 10
Часть II
КЛИМАТ И ПЕРЕСТРОЙКИ В ТЁПЛОЙ БИОСФЕРЕ (МЕЛ, ЮРА)
Глава 2. Общий обзор позднемезозойского
климата и событий (Н.М. Чумаков)............................................
44
Глава 3. Палеогеографические
перестройки и седиментация мелового периода
(М.А. Жарков, И.О. Мурдмаа, Н.И. Филатова)....................................................................................................
52
Глава 4. Количественные палеоботанические
данные о позднемеловом климате
Евразии и Аляски (А.Б. Герман)............................................................................................................................
88
Глава 5. Климатическая зональность
и климат мелового периода (Н.М. Чумаков).........................................
105
Глава 6. Глобальная экспансия
планктонных фораминифер: триас, юра, мел
(К.И. Кузнецова, О.А. Корчагин)..........................................................................................................................
124
Глава 7. Динамика и возможные
причины климатических изменений в позднем мезозое
(Н.М. Чумаков)........................................................................................................................................................
149
Часть III
ЭПОХА ПЕРЕХОДА ОТ ХОЛОДНОЙ К ТЁПЛОЙ БИОСФЕРЕ (ПЕРМЬ И РАННИЙ ТРИАС)
Глава 8. Палеогеографические перестройки
и обстановки седиментации в перми и раннем
триасе (М.Л. Жарков)..............................................................................................................................................
158
Глава 9. Палеобиогеография пермских
фузулинид (Э.Я. Левен)........................................................................
181
Глава 10. Палеофитогеография
пермского периода (С.В. Наугольных).............................................................
194
Глава 11. Палеопочвы перми и
раннего триаса (С.В. Наугольных)...................................................................
221
Глава 12. Климат и климатическая
зональность перми и раннего триаса (Н.М. Чумаков).............................
230
Часть IV
ГЛАВНЫЕ КЛИМАТИЧЕСКИЕ И БИОСФЕРНЫЕ СОБЫТИЯ ПОЗДНЕГО ДОКЕМБРИЯ
Глава 13. Ледниковый и безледниковый
климат в докембрии (Н.М. Чумаков)...............................................
259
Глава 14. Проблема климатической
зональности в позднем докембрии. Климат и биосферные
события (Н.М. Чумаков, В.Н. Сергеев).................................................................................................................
271
Заключение (Н.М.
Чумаков)................................................................................................................................
290
Conclusion...............................................................................................................................................................
296
