Глава 9
ПАЛЕОБИОГЕОГРАФИЯ ПЕРМСКИХ ФУЗУЛИНИД
Э. Л. Левей
Данные биогеографии не в меньшей степени, чем седиментационные обстановки,
описанные в предыдущей главе, свидетельствуют о характере климата прошлых
эпох, его поясной зональности и изменении во времени. Морские и особенно
наземные
организмы являются чуткими индикаторами среды обитания, и их распределение
по поверхности планеты определяется, в первую очередь, климатическими факторами.
В настоящей главе это будет продемонстрировано на примере биогеографического
анализа палеосообществ пермских фузулинид - широко распространенных и хорошо
изученных бентосных морских организмов, обитавших на небольших глубинах и
поэтому чутко реагировавших на температурные колебания.
Сейчас уже можно считать установленным, что современное распространение
разных типов палеозойских фаун и флор на земной поверхности не укладывается
в четкие
закономерности и ассиметрично по отношению к положению экватора и, соответственно,
климатических поясов. Картина становится более закономерной, если палеобиогеографические
реконструкции производить на палинспастической основе. В качестве таковой
с небольшими изменениями и дополнениями приняты карты, иллюстрирующие в предыдущей
главе палеоседиментологические обстановки.
Первые палеогеографические карты для различных эпох пермского периода, основанные
на распространении фораминифер, были составлены А.Д. Миклухо-Маклаем [1963].
Позже вопросы биогеографии фузулинид были затронуты в работах DJ. Gobett
[1967; 1973], Ch. A. Ross [1967a; 1982; 1990; 1995], Ch. A. Ross, J.R.P.
Ross [1983],
К. Ishii et al [1985], Т. Ozawa [1987], Э.Я. Левей [1994], Э.Я. Левей, С.Ф.
Щербович [1978], E.Ja. Leven [1994; 1997], F. Kobayashi [1997 a,b; 1999]
и др. При составлении предлагаемых в настоящем разделе монографии палеобиогеографических
карт использованы все содержащиеся в перечисленных публикациях данные; учтены
также
собственные материалы по Памиру, Афганистану, Ирану, Турции, Каракоруму,
Внутренней Монголии, Южному Китаю и Новой Зеландии. Карты составлены для
четырех временных
интервалов, отвечающих четырем эпохам, на которые, по нашему мнению [Leven,
2003], следует делить пермский период. Для них предлагается употреблять названия
уральская, дарвазская, янсинская и лопинская, в разное время использовавшиеся
для обозначения стратиграфических подразделений, примерно отвечающих по времени
предлагаемым эпохам (табл. 7).
Таблица 7
Стратиграфические шкалы пермской системы
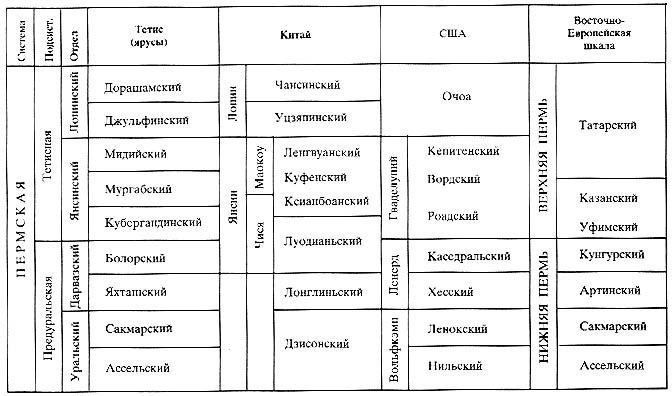
Достоверность палеореконструкций во многом зависит от точности возрастной
корреляции привлекаемых для этого материалов. Поскольку для расчленения перми
в разных
биогеографических областях используются разные стратиграфические шкалы, мы
сочли необходимым привести здесь схему корреляции главных из них (табл. 7).
Оговоримся, что схема отражает современное состояние пермской стратиграфии,
не во всем удовлетворительное. Поэтому в ряде случаев принимаемая нами корреляция
в той или иной степени условна. Это относится прежде всего к ярусам верхней
подсистемы пермской системы (подробнее см. [Левей, 2001]).
Фузулиниды были мелководными бентосными теплолюбивыми организмами, пределы
распространения которых на север и юг ограничивались 35°-40° северной и южной
широт. Биогеография фузулинид определялась положением их местообитания по
отношению к палеоэкватору; немаловажное значение имела степень изоляции бассейнов
и наличие
или отсутствие путей свободной миграции. В Перми обособились три главные
области их распространения, различающиеся характером фузулинидовых сообществ.
Наиболее
крупная из них охватывала территорию Палео-Тетиса и острова западной Палео-Пацифики.
Вторая область занимала западные побережья Северной и Южной Америки и часть
островов восточной Палео-Пацифики. Третья область
распространялась на Восточно-Европейский и Свердрупский бассейны. Последний
в начале перми имел связь с бассейном западного побережья Северной Америки.
Ч. Росс [Ross, 1995] рассматривал перечисленные области как провинции, присвоив
им названия Палео-Тетическая (paleo-Tethys), Мидконтинент-Андская (Midkontinent-Andean)
и Франклино-Уральская (Franklinian-Uralian), соответственно. В дальнейшем
мы будем пользоваться этими названиями, считая, однако, что обозначаемые
ими территории
следует понимать в более широком значении, чем провинция. Все три области
существовали в ранней Перми. В начале янсинской эпохи фузулиниды исчезли
во Франклино-Уральской
области, а в
начале лопинской - также и в Мидконтинент-Андской, сохранившись лишь в пределах
Тетиса. Ниже остановимся на биогеографии фузулинид в разные эпохи пермского
периода более подробно.
9.1. Уральская эпоха
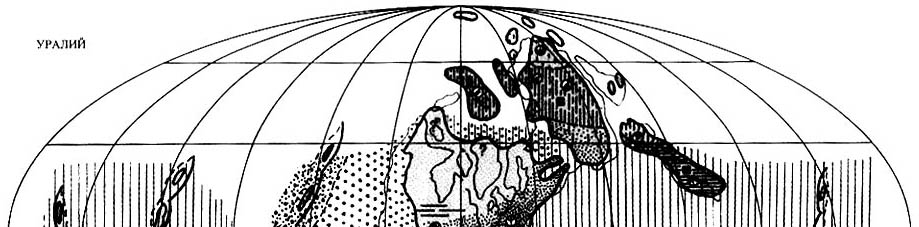  |
 |
Рис. 69. Палеобиогеография фузулинид и наземной флоры уральской эпохи
(ассельский и сакмарский века) (составили Э.Я. Левей и С.B. Наугольных) Условные обозначения к рис. 69-72. Биогеография фузулинид 1-8: 1 - Палео-Тетическая область; 2 - Южно-Тетическая провинция; 3 - Мидконтинент-Андская область; 4 - провинция Сономия; 5 - Франклино-Уральская область; 6 - Западно-Тетическая провинция; 7 - Восточно-Тетическая провинция; 8 - провинция Панталасса. Биогеография наземной флоры 9-15: 9 - обедненная растительность тундрового или лесотундрового типов; 10 - типично-ангарская листопадная растительность (пермский аналог современных бореальных лесов, "кордаитовая тайга"); 11 - тропическо-экваториальные засушливые зоны, растительность средиземноморского типа; 12 - экваториальная и тропическая вечнозеленая растительность, сходная с современными дождевыми лесами; 13 - ксерофильная растительность бессточных котловин; 14 - зоны смешения растительности средиземноморского типа (еврамерийский тип флор) и растительности южного умеренного и холодно-умеренного биома (гондванский тип флор); 15 - нотальная листопадная растительность с доминированием глоссоптерид. Жизненные формы и формы роста высших растений, типы ландшафтов 16-21: 16 - хамефиты и гемикриптофиты, кустарники, тундра; 17 - фанерофиты, древовидные листопадные формы Северного полушария (войновскиевые); 18 - фанерофиты; древовидные листопадные формы южного полушария ("глоссоптериды"); 19 - ксерофиты, представленные фанерофитами и хамефитами; 20 - аридные ландшафты, колонизированные пельтоспермовыми птеридоспермами и хвойными; приэкваториальные дождевые леса; 21 - хамефиты и гемикриптофиты; предполагаемая тундровая зона Нотальной области (Гондвана); 22 - зоны углеобразования. Элементы палеогеографии 23-26: 23 - границы моря и суши; 24 - границы континентов и микроконтинентов; 25 - границы палеогеографических провинций и растительных зон; 26 - островные дуги. |
Палео-Тетическая область распространялась в это время на
огромную территорию между Еврамерийским и Ангарским материками на севере
и Гондваной на юге (рис.
69). По современным представлениям большую часть этой территории занимал
океан Тетис с многочисленными островами и микроконтинентами, постепенно дрейфовавшими
в северном
направлении, вплоть до последующей коллизии с Ангаридой. По поводу их числа,
размеров и расположения пока нет единого мнения. На палеореконструкциях последних
лет, в том числе и на принятой за основу в настоящей монографии, акватория
Тетиса как бы "запирается" с востока микроконтинентами Северного
и Южного Китая и Индокитая, отделяющими ее от палео-Пацифики. Предполагается,
что в пределах последней также были разбросаны небольшие микроконтиненты
и вулканические острова, в прибрежных водах которых обитали фузулиниды тетического
типа. Часть этих микроконтинентов дрейфовала в западном, северо-западном
и
юго-западном направлениях, и их фрагменты сейчас можно наблюдать в аккреционных
комплексах Корякского нагорья, Приморья, Японии и Новой Зеландии. Другая
часть микроконтинентов перемещалась в восточном и северовосточном направлениях,
пока
не была при-членена к Северо-Американскому континенту [Ch. Ross, J. Ross,
1983; Ch. Ross, 1995; Ishii et al., 1985; Leven, Grant-Mackie, 1997; Kobayashi,
1999].
В уральскую эпоху фузулиниды были расселены почти на всей акватории Тетиса,
за исключением самых южных районов, где их существованию препятствовал суровый
климат. Особенно это относится к ассельскому времени, когда гондванское оледенение
достигало своего максимума и когда фузулиниды не заходили южнее 30° ю.ш.
Так, они полностью отсутствуют в районах, примыкавших к Австралии и Индии,
таких
как западный Таиланд и Бирма (Блок Сибумасу), Южный Тибет, Гималаи, Каракорум,
Южный Памир и Южный Афганистан. Обедненные фузулинидовые комплексы начинают
встречаться лишь в Центральном Иране [Baghbani, 1993; Leven, Taheri, 2003],
т.е. в районах, располагавшихся уже севернее тридцатой параллели. В связи
с сакмарским потеплением и таянием гондванских ледников происходила южная
экспансия
фузулинид, появление которых отмечено на всех перечисленных выше территориях,
за исключением Гималаев; не было их и в бассейнах, располагавшихся на Австралийских
шельфах.
Надо сказать, что в современных структурах перечисленные выше районы представляют
собой отдельные тектонические блоки, являющиеся лишь фрагментами существовавших
ранее тектонических структур. Поэтому истинные размеры располагавшихся на
этой территории морских бассейнов остаются неясными и вероятно они существенно
превышали
суммарную площадь тектонических блоков, в
пределах которых сейчас обнаруживаются морские осадки с фузулинидами. Скорее
всего, эти блоки являются частями северного края Гондваны и в начале перми
не были отделены от нее и друг от друга глубоководными бассейнами (бассейном)
океанического типа, как это показано на большинстве палинспастических палеогеографических
карт, в том числе и на опубликованных недавно картах Зиглера [Ziegler et
al., 1998]. К такому выводу приводит повсеместный мелководный характер пермских
отложений и отсутствие хорошо доказанных офиолитов этого возраста.
Почти вся рассматриваемая Палео-Тетическая область находилась в пределах
тропико-экваториального пояса. Соответственно, фузулинидовая биота, в целом,
отличалась здесь таксономическим
разнообразием, усиливавшимся многообразием условий обитания фузулинид, которые
не могли быть одинаковыми на столь огромной и геоморфологически неоднородной
территории. Среди фузулинид господствовал отряд Schwagerinida, представленный
четырьмя семействами - Triticitidae, Rogosofusulinidae, Schwagerinidae и
Pseudofu-sulinidae. Наиболее характерными были швагеринидеи, отличающиеся
вздутой субсферической
и сферической раковиной и свободным навиванием оборотов спирали (роды Sphaeroschwagerina,
Pseudoschwagerina, Paraschwagerina, Zellia, Robustoschwagerina и некоторые
другие). Обилие и разнообразие представителей этих родов может служить показателем
максимально благоприятных условий обитания. Не менее ярким свидетелем последних
являются также такие представители семейства Rugosofusulinidae как Rugosofusulina и,
в особенности, Dutkevitchia. Заметную роль в фузулинидовом сообществе
занимали род Quasifusulina семейства Fusulinidae и роды Pseudofusulina,
Monodiexodina и Eoparafusulina семейства Pseudofusulinidae.
Сейчас трудно говорить о каком-либо провинциализме в распространении фузулинид
в пределах Тетиса, главным образом, из-за неравномерной изученности их сообществ
в разных частях этой области. Наиболее разнообразно эти сообщества представлены
в разрезах Карнийских Альп (Kahler, 1985; Forke, 1995] Северного Афганистана
[Leven, 1971], Дарваза [Левей, Щербович, 1998], Ферганы [Бенш, 1962; 1972],
Синцзяня, Кунь-Луня, южной части Северного Китая [Чжан Линь-синь, 1963; Zhao
Zhixin et al., 1984; Lee, 1927 и др.]. Все эти районы располагались на севере
Тетиса. Не менее разнообразны ассельско-сакмарские фузулинидовые комплексы
во многих
разрезах Южного Китая, Восточного Индокитая и Японии. Что касается южных
пери-Гондванских районов Тетиса, то тут картина иная. Ассельские и сакмарские
фузулиниды известны
в разрезах Туниса, Тавра, Центрального Ирана и Эльбурса, но комплексы их
здесь, как уже говорилось, обеднены. Юго-восточнее, в полосе, захватывающей
Южный
Афганистан, Южный Памир, Каракорум, большую южную часть Тибета, Бирму и Западный
Таиланд, ассельские фузулиниды отсутствуют. Появляются они в сакмарских
отложениях и представлены почти исключительно эндемичными псевдофузулинами
(калакташский
комплекс); [Leven, 1993]. Встречаются монодиексодины и эопарафузулины, обитавшие
в условиях с повышенной гидродинамикой. Субсферические швагериниды, характерные
для более теплых тропических районов Тетиса, отмечены лишь на самом севере
обозначенной территории - на Центральном Памире [Leven, 1993]. Своеобразие
калакташского комплекса позволяет выделить район его распространения в отдельную
провинцию
- Южнотетическую.
Ассельско-сакмарские фузулиниды отмечены в некоторых террейнах аккреционных
комплексах Японии (террейн Акиоши) [Kobayashi, 1999], где их комплекс мало
отличается от такового Южного Китая, также характеризуясь разнообразием и
присутствием большинства родов, типичных для экваториально-тропического пояса
[Ozawa, Kobayashi,
1990].
Мидконтинент-Андская область охватывала западное побережье
Северной и Южной Америки и близлежащие острова (рис. 69). Хотя она, так же
как и Палео-Тетическая,
располагалась в пределах тропико-экваториального пояса, значительная степень
ее изоляции обусловила формирование здесь своеобразного фузулинидового сообщества,
существенно отличающегося от тетического. Его основу составляли разнообразные
псевдофузулины и ругозофузулины, в видовом отношении совершенно иные, чем
в Тетисе. Большая часть родов семейства Schwagerinidae с субсферической раковиной
отсутствует. Исключение составляют псевдошвагерины и парашвагерины, но, судя
по характеру ювенариумов, они имели автохтонное происхождение от туземных
видов
рода Triticites, очень характерного для нижней части разреза серии
Вольфкэмп. Потомками тритицитесов были также Leptotriticites и Dunbarinella -
роды, не встречающиеся за пределами рассматриваемой области. Кроме перечисленных,
существенно
подчиненное значение в сообществе имели такие роды, как Schubertella,
Oketaella, Waeringella, Pseudofusulinella и некоторые другие. Они известны
и в разрезах Тетиса, но, за исключением шубертелл, встречаются там крайне
редко.
Вероятно, к рассматриваемой области принадлежала и островная территория или
террейн Сономия (Sonomia), фрагменты которого разбросаны в пределах штатов
Калифорния, Орегон и Невада [Ch. Ross, J. Ross, 1983]. Наиболее полный комплекс
фузулинид из этого террейна описан Дж. Скиннером и Г. Уильди [Skinner, Wilde,
1965]. В целом, он близок к комплексу из соответствующих отложений Мидконтинента,
но отличается присутствием эндемичных родов Klamathina, Cuniculinella и Mccloudia,
а также родов Pseudofusulinella и Eoparafusulina, которые хотя и встречаются
в ассельско-сакмарских разрезах Мидконтинента, но для них не характерны.
Все это позволяет выделять террейн Сономия в самостоятельную провинцию в
пределах
Мидконтинент-Андской области.
Франклино-Уральская область охватывала Восточно-Европейский и Свердрупский
бассейны, и частично аляскинскую часть Северной Америки. На юго-востоке Восточно-Европейский
бассейн имел свободные связи с Тетисом. Последнее в значительной степени
определяло характер фузулинидового сообщества этого бассейна, которое в ассельском
веке
мало отличалось от тетического сообщества. По мере удаления от пролива, соединяющего
бассейн с Тетисом, характер сообщества несколько менялся, что выражалось
в уменьшении таксономического разнообразия и появлении родов, не характерных
для теплолюбивого тетического сообщества или вовсе в нем отсутствующих. К
ним,
прежде всего, следует отнести роды семейства Fusulinellidae, такие как Pseudofusulinella,
Kanmeraia, Waeringella, Eowaeringella и некоторые другие. Многие из них встречаются
в Мидконтинент-Андской области, что может служить свидетельством существования
связей между ней и рассматриваемой областью. Об этом же говорит и присутствие
сферошвагерин на северо-востоке Британской Колумбии (Канада) - районе, принадлежавшем
к континентальному шельфу Северной Америки [Ch. Ross, J. Ross, 1983]. Общим
является и присутствие эопарафузулин.
Р. Линь, У. Нассичук и Р. Торстейнссон [Lin et al., 1994[ сделали заслуживающее
внимание предложение - выделить территорию, охватывающую север Канады, Гренландию
и Шпицберген, где разнообразие ассельского фузулинидового сообщества минимально
и ограничивается четырьмя-пятью родами, в самостоятельную Арктическую провинцию.
По отношению к палеоширотам она занимает
то же положение, что и выделяемая в сакмарском веке Южнотетическая провинция,
также отличающаяся бедностью и однообразием фузулинидовых комплексов, характерным
элементом которых, как и в Арктической провинции, являются эопарафузулины.
Очевидно, подобное сходство не случайно и, по-видимому, обусловлено сходными
климатическими условиями существования ассельских фузулинид в Арктической
провинции и сакмарских - в Южнотетической. Напомним, что в ассельское время
фузулиниды в Южнотетической провинции еще отсутствовали. Отсюда вытекает,
что в самом начале перми южная и северная границы распространения фузулинид
были
неодинаковыми: первая находилась ближе к экватору, чем вторая. Такая асимметрия
была, вероятно, вызвана влиянием южного гондванского оледенения.
Начиная с сакмарского века, связь Восточноевропейского и Тетического бассейнов
была затруднена или вовсе прекратилась. Следствием явилось постепенно увеличивающееся
обособление фузулинидового сообщества рассматриваемой области от тетического.
В первую очередь, это нашло отражение в резком сокращении роли семейства
швагери-нидей. Лишь в первую половину сакмарского века здесь продолжали существовать
сферошвагерины
и парашвагерины, но и те вскоре исчезли. Такие характерные для Тетиса швагеринидеи,
как Zellia, Pseudoschwagerina, Robustoschwagerina, Acervoschwagerina в
сакмарских отложениях Франклино-Уральской области не отмечены. Доминирующее
значение
приобретают представители семейства псевдофузулинидей, представленные родом
Pseudofusulina и некоторыми близкими им родами, в видовом отношении,
как правило, отличающимися от тех же родов в Палео-Тетической области. Отмеченные
отличия проявляются
уже в разрезах Южного Урала, т.е. на юге Франклино-Уральской области. По
мере продвижения на север они усиливаются. Заметную роль начинают играть
эопарафузулины.
В Арктической провинции сакмарские фузулиниды пока не известны, что, возможно,
связано с плохой изученностью соответствующего интервала разреза.
Оценивая, в целом, особенности ассельско-сакмарского фузулинидового сообщества
Франклино-Уральской области, можно констатировать, что они определялись в
основном степенью открытости связей Восточноевропейского и Тетического бассейнов
и климатическим
фактором; определенную роль играла вероятная связь с Мидконтинент-Андской
областью.
9.2. Дарвазская эпоха
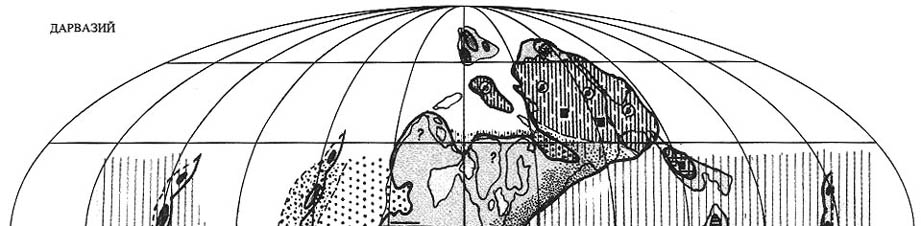 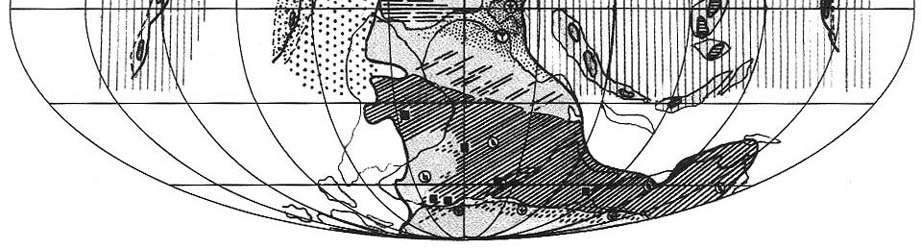 |
| Рис. 70. Палеобиогеография фузулинид и наземной флоры дарвазской эпохи
(яхташский и болорский века) Составили Э.Я. Левен и С.В. Наугольных. Условные обозначения см. на рис. 69 |
Палео-Тетическая область. Конец сакмарского века в рассматриваемой
области отмечен регрессией (рис. 70), пик которой приходился на начало яхташского
века [Левей, 1993; Leven, 1994]. С этим событием связана существенная перестройка
фузулинидовой биоты. Исчезли или резко сократились в численности такие характерные
для предыдущей эпохи роды отряда Schwagerinida, как Sphaeroschwagerina,
Pseudoschwagerina,
Paraschwagerina, Zellia, Rugosofusulina, Dutkevitchia и ряд других.
На смену им пришли Darvasites, Nagatoella, Chalaroschwagerina, Leeina,
Praeskinnerella,
первые Skinnerella, наряду с псевдофузулинами определявшие облик
дарвазского фузулинидового сообщества. Важным событием явилось появление
рода Pamirina,
а затем его прямого потомка - рода Misellina, являющегося первым представителем
высших фузулинид отряда Neoschwagerinida. Характерными для дарвазского сообщества
являются впервые появившиеся Mesochubertella и Toriyamaia, а также возникшие
раньше Biwaella. Широкая трансгрессия, сменившая регрессию во второй половине
яхташского века, способствовала быстрому освоению вновь появившимися таксонами
всего пространства Тетиса. Стирается своеобразие Южнотетической провинции,
поскольку многие из вновь появившихся родов проникли на относившиеся к ней
территории Южного Афганистана, Южного Памира, Каракорума, Южного Тибета.
По-видимому, этому способствовало и общее потепление климата, хотя последнее
не бесспорно,
поскольку смягчение климата на перечисленных территориях можно объяснить
перемещением соответствующих материковых блоков на север, в более низкие
широты.
Распространение вновь появившихся таксонов не ограничивалось только пределами
собственно Тетиса, а захватывало и острова Пацифики. Характерные для дарвазского
фузулинидового сообщества роды и виды отмечены в террейнах Японии, Корякского
Нагорья, запада Северной Америки (Калифорния) [Kobayasi, 1977a; Davydov et
al., 1996; Douglass, 1967].
Мидконгинент-Андская область продолжала оставаться изолированной
(рис. 70). Основу фузулинидового сообщества этой области по-прежнему составляли
псевдофузулины,
которые во вторую половину рассматриваемого отрезка времени начали сменяться
парафузулинами и скиннереллами, отличающимися от псевдофузулин появлением
куникул. До этого для фузулинидовых ассоциаций
были характерны также чаларошвагерины (особенно, в провинции Сономия), прескиннереллы,
а также редкие монодиексодины; продолжали существовать псевдофузулинеллы.
Все перечисленные роды, за исключением последнего, встречаются в дарвазских
фузулинидовых ассоциациях Палео-Тетической области, но, в целом, как и раньше,
фузулинидовое сообщество рассматриваемой области отличается своим таксономическим
однообразием. Вместе с тем обращает на себя внимание, по-видимому, одновременное
появление и там, и здесь морфологически близких таксонов. К ним можно отнести
обладающих свободной спиралью и характерной складчатостью септ чаларошвагерины,
а также парафузулины и скиннереллы у которых возникли куникулы. Казалось
бы, этот факт должен свидетельствовать о свободных связях между Палео-Тетическим
и Североамериканским бассейнами. Однако совершенно неясно, каким образом
они
могли осуществляться. Кроме того, сейчас уже можно считать несомненным нередко
наблюдаемое независимое возникновение сходных морфологических признаков у
различных, часто даже неродственных фузулинид. Они могут появляться в разных
филумах
и разное время. Например, появление куникул отмечено в отряде Fusulinida
(род Paraeofusulina) в московском веке, в отряде Schwagerinida (роды Parafusulina,
Darvasites, Chalaroschwagerina, Cuniculinella) в сакмарском и яхташском
веках
и в отряде Schubertellida (род Codonofusiella) в мидийском веке.
Чем ближе в родственном отношении находятся те или иные виды, тем ближе по
времени
происходит у них сходное морфологическое обновление. Это обстоятельство не
учитывалось
при разработке принимаемой сейчас систематики фузулинид, в основу которой
положены морфологические различия выделяемых таксонов. Поэтому многие из
этих таксонов,
в особенности роды, являются гетерогенными. К таким родам, несомненно, относятся
парафузулины и, вероятно, чаларошвагерины, которые в Мидконтинент-Андской
и Палео-Тетической областях могли возникнуть независимо, что подтверждается
присутствием и там и здесь переходных форм между этими родами и туземными
видами псевдофузулин. Изложенные факты крайне усложняют работу систематиков
и стратиграфов,
и их, конечно, надо иметь в виду и при биогеографических реконструкциях.
В свете рассматриваемых в настоящей статье проблем это означает, что американские
и тетические парафузулины (так же как и чаларошвагерины), строго говоря,
являются
разными, независимо возникшими родами, хотя и крайне близкими морфологически.
Поэтому нет необходимости искать пути проникновения американских парафузулин
в Тетис
или
наоборот.
Франклино-Уральская область. В дарвазское время Восточно-Европейский
и Палео-Тетический бассейны были полностью изолированы. Прекращение обмена
между их фузулинидовыми сообществами и начавшееся осолонение Восточно-Европейского
бассейна привели к разубоживанию в нем фузулинидо-вых ассоциаций, основной
составляющей которых стали псевдофузулины и появившиеся (независимо от североамериканских
и тетических) парафузулины. В видовом отношении они сильно отличались от
этих родов других областей. По-видимому, связи с Мидконтинент-Андской областью
также были прерваны, так как ничего не говорит об их существовании. Наиболее
западная точка, где были отмечены артинские фузулиниды восточноевропейского
типа, находится в штате Юкон Канады [Ross, 1967b]. В кунгурском веке все
фузулиниды
в рассматриваемой области вымерли и больше не появлялись.
9.3. Янсинская эпоха
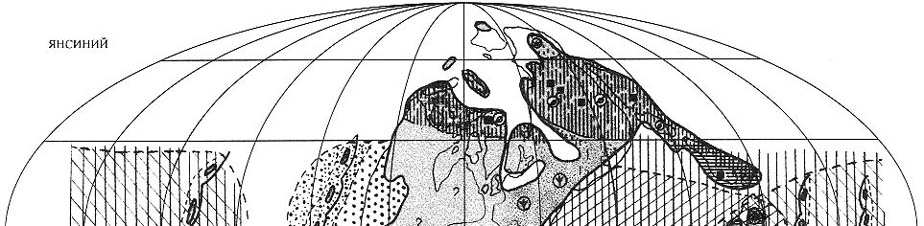 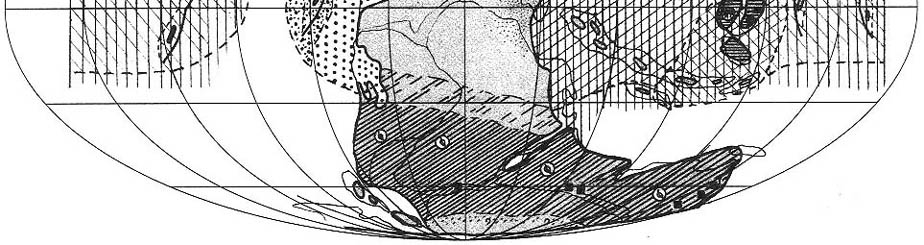 |
| Рис. 71. Палеобиогеография фузулинид и наземной флоры
янсинской эпохи (кубергандинский, мургабский и мидийский века) Составили Э.Я. Левен и С.В. Наугольных. Условные обозначения см. на рис. 69 |
Палео-Тетическая область. Начавшаяся в конце яхташского
- болорском веке трансгрессия, в кубергандинском веке, т.е. начале янсинской
эпохи охватила
все пространство
Тетиса (рис. 71). Это привело к повсеместному формированию карбонатных платформ
и существенному обновлению фузулинидовой биоты. Вымирают Toriyamaia,
Biwaella, Mesoschubertella, Quasifusulina, Darvasites, Chalaroschwagerina и
некоторые другие роды, определявшие облик дарвазского сообщества. На смену
им приходят
Yangchienia, Skinnerella, Chusenella, Eopolydiexodiana, а также
высшие фузулиниды отряда Neoschwagerinida, которые хотя и возникли в болорском
веке, но только
начиная с кубергандинского заняли главенствующую роль. Возросло общее разнообразие
фузулинидовых ассоциаций, максимум которого приходился на мидийский век.
Тогда же многие фузулиниды, особенно неошвагериниды, достигли наивысшей
специализации
[Левей, 2003].
В своем распространении, как и до этого, фузулиниды ограничивались 35-й-40-й
параллелями северной и южной широты. Правда, на современных картах местонахождения
типично янсинских комплексов фузулинид расположены в значительно более высоких
северных широтах - в Корякском нагорье и Британской Колумбии (Канада), а
также на
островах Новой Зеландии. Последние в перми также находились в высоких широтах,
но южных. Некоторые исследователи объясняют проникновение фузулинид в эти
широты теплыми течениями, что на наш взгляд маловероятно, поскольку в этом
случае
неизбежно происходило бы смешение тепло- и холодно-любивой фауны, что нигде
не наблюдается. Учитывая, что во всех случаях фузулиниды приурочены к тектоническим
блокам внутри сложных аккреционных комплексов, более вероятной, на наш взгляд,
является террейновая гипотеза происхождения этих блоков, допускающая первоначальное
положение террейнов в тропической области Палео-Пацифики [Ross Ch., Ross
J., 1983]. Это относится также к тектоническим блокам, составляющим Японские
острова [Kobayashi, 1997b], и, вероятно, к известным выходам перми в Приморье.
В распространении фузулинидовых ассоциаций в пределах рассматриваемой области
намечаются определенные закономерности, впервые отмеченные в работе [Ishii
et al., 1985]. Более подробно они затронуты Ф. Кобаяши [Kobayashi, 1997a],
который выделил для средней перми три провинции - Западно-Тетическую, Восточно-Тетическую
и провинцию Панталасса. С этими выводами, в целом, можно согласиться. Различия
между Западно-Тетической провинцией и провинцией Панталасса состоят в том,
что для первой из них характерно повсеместное распространение семейства Sumatrinidae
отряда Neoschwagerinida при отсутствии подсемейства Lepidolininae семейства
Neoschwagerinidae этого же отряда. Это подсемейство широко распространено
в пределах Панталассы, но суматриниды здесь не встречаются. Ареалы распространения
суматринидей и лепидолинин перекрываются на территории современного Китая
и Индокитая, и именно она рассматривается Кобаяши как самостоятельная Восточно-Тетическая
провинция. Внутри намеченных провинций вероятно существовала какая-то широтная
зональность, контуры которой пока весьма расплывчаты. Например, в отделившихся
от Гондваны блоках фузулинидовые ассоциации, в целом, менее разнообразны,
чем
севернее [Leven, 1997]. Интересно, что обратная картина наблюдается среди
мелких фораминифер, многие представители которых, такие как мидийские Shanita,
Rectostipulina,
Sphairionia и ряд других, обитали только на юге Тетиса. Наименьшим разнообразием
фузулинидовые ассоциации отличались в самых южных и северных точках их распространения.
В первом случае это Оман и Соляной Кряж, во втором - Внутренняя Монголия
Китая.
Мидконтинент-Андская область сохраняла свою обособленность (рис. 71). По-прежнему
абсолютно доминировало семейство Schwagerinidae, представленное, главным
образом, парафузулинами, скиннереллами и, в меньшей степени, псевдофузулинами.
Лишь
в конце рассматриваемого отрезка времени появились многоапертурные, не встречающиеся
за пределами области Polydiexodina, морфологически очень близкие к тетическим
Eopolydiexodina, которые ранее относились к полидиексодинам. В качестве самостоятельного
рода они стали рассматриваться лишь когда было установлено их существенно
более раннее появление и от иных предков. Здесь мы сталкиваемся еще с одним
ярким
примером независимого гомологичного развития фузулинид рассматриваемой области,
о котором говорилось ранее [Левен, Грабчак, 1986].
Многочисленные представители отряда неошвагеринид, наиболее характерного
для янсинского фузулинидового сообщества области Палео-Тетис, в Мидконтинент-Андской
области, практически, отсутствуют. Лишь в самом конце рассматриваемой эпохи
сюда проникает род Yabeina, представленный только одним примитивным и своеобразным
видом. Последовавшая затем регрессия привела к вымиранию всей фузулиновой
биоты,
и Мидконтинент-Андская область перестала существовать.
Франклино-Уральская область. Как уже говорилось, последние
фузулиниды исчезли здесь в кунгурском веке и вместе с ними перестала существовать
и сама область,
поскольку она изначально была обособлена на основании присущего ей характерного
фузулинидового сообщества. Очевидной причиной вымирания фузулинид было осолонение
и частичное осушение бассейна, что, в свою очередь, стало следствием потепления
и аридизации климата, а также регрессии. Однако, когда во время широкой казанской
трансгрессии здесь восстановился нормальный морской режим, фузулиниды сюда
не вернулись. Они могли проникнуть в бассейн лишь со стороны западно-американского
побережья, но обогнуть Аляскинский выступ, вдававшийся далеко на север в
область низких температур, оказались не в состоянии.
9.4. Лопинская эпоха
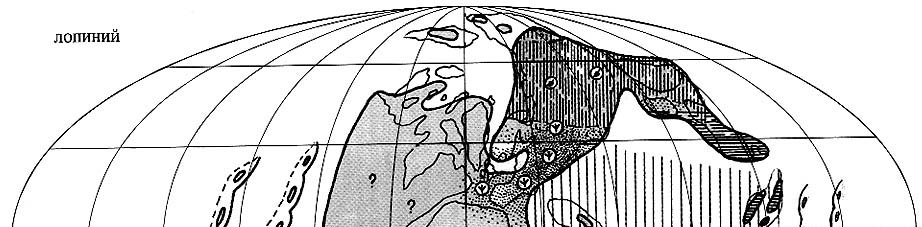 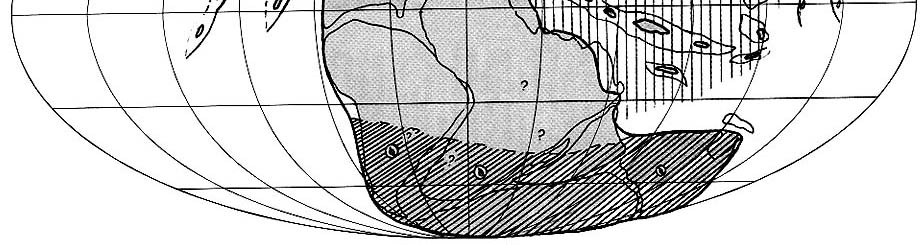 |
| Рис. 72. Палеобиогеография фузулинид и наземной флоры
лопинской эпохи (джульфинский и дорашамский века) Составили Э.Я. Левен и С.В. Наугольных. Условные обозначения см. на рис. 69 |
Палео-Тетическая область (рис. 72). На рубеже янсинской
и лопинской эпох произошел крупнейший кризис в развитии фузулинид, когда
вымерло 77% всех
родов и исчезли
до этого доминировавшие отряды неошвагеринид и швагеринид [Левен, Корчагин,
2001]. Причины
кризиса не ясны и, по-видимому, имели многофакторный характер. Определенную
роль, вероятно, сыграла послемидийская регрессия, которая однако проявилась
не везде. Примечательно, что во многих разрезах на смену мидийским мелководным
фациям приходят более глубоководные отложения (глины, мергели, микритовые
известняки) и, соответственно меняется характер всей биоты, существенным
элементом которой
становятся пелагические группы фауны, такие как конодонты и аммоноидеи.
В связи с послемидийским вымиранием разнообразие фузулинидового сообщества
существенно понизилось. Пережившие кризис отряды Schubertellida, Staffellida
и Ozawainellida представлены всего 10-15 родами, в числе которых наиболее
заметными были роды семейства Palaeofusulinidae, в особенности род Palaeofusulina.
Сравнительная
редкость находок фузулинид делает невозможным произвести какое-либо биогеографическое
районирование внутри рассматриваемой области. Ясно лишь, что в наибольшей
степени фузулиниды представлены в районах, располагавшихся вблизи экватора.
Фузулиниды полностью исчезли с лица Земли в самом конце перми. Этим для данной
группы фауны завершилось "великое пермское вымирание", первая фаза
которого приходилась на конец мидийского века.
Выводы
Представленная выше картина палеобиогеографии фузулинид для четырех временных отрезков имеет самый общий характер. Однако даже на таком уровне биогеографический анализ позволяет, во-первых, подтвердить в основных чертах справедливость палинспа-стических реконструкций для пермского периода, во-вторых, выявить, связанную с климатом, широтную зависимость таксономического разнообразия и численности фузулинидовых сообществ и, наконец, в-третьих, на примере Мидконтинент-Андской и Франклино-Уральской областей показать влияние на характер сообществ наличия или отсутствия свободных связей между палеобассейнами. Наблюдаемое в разных изолированных друг от друга бассейнах сходство отдельных элементов фузулинидовых сообществ позволяет ставить вопрос о широком проявлении среди фузулинид явлений параллельного гомеоморфного развития со всеми вытекающими отсюда таксономическими и стратиграфическими последствиями. Более детальному биогеографическому анализу фузулинид препятствует их неравномерная и, в общем, еще слабая изученность в пределах основной области их распространения - Палео-Тетической. Кроме того, подчас неразрешимые трудности создает сложное геологическое строение этой области, представляющее собой мозаику из тектонических блоков, испытавших с пермского времени значительные горизонтальные перемещения. При этом, как направленность этих перемещений, так и их масштаб, часто остаются неясными. Проблематично и существование самого обширного океана Тетис, поскольку, если в пределах области и имеются выходы пермских офиолитов, что пока точно не доказано, то в очень ограниченном количестве. В основном же пермские отложения имеют мелководный характер. Для решения всех этих проблем нужны широкомасштабные комплексные исследования, немаловажную роль в которых должен играть биогеографический анализ.
Литература
Бенш Ф.Р. Позднекаменноугольные и раннепермские фузулиниды Северной
Ферганы // Стратиграфия и палеонтология Узбекистана и сопредельных
районов. Ташкент:
Изд-во АН УзССР, 1962. С. 186-252.
Бенш Ф.Р. Стратиграфия и фузулиниды верхнего палеозоя Южной Ферганы.
Ташкент: ФАН, 1972. 140 с.
Левен Э.Я. Главные события пермской истории Тетиса и фузулиниды
// Стратиграфия. Геол. корреляция. 1993. Т. 1, № 1. С. 59-75.
Левен Э.Я. Трансгрессии и регрессии пермского моря Тетис
и их влияние на развитие фузулинидовой биоты // Экосистемные перестройки
и эволюция
биосферы.
М.: Недра,
1994. Вып. 1. С. 119-125.
Левен Э.Я. О возможностях использования глобальной ярусной шкалы
пермской системы в пределах Тети-са // Стратиграфия. Геол. корреляции.
2001.
Т. 9, № 2. С. 15-29.
Левен Э.Я. Динамика родового разнообразия и основные этапы развития
фузулинид // Стратиграфия. Геол. корреляция. 2003. Т. 11. № 3. С.
15-26.
Левен Э.Я., Грабчак О.Л. Об объеме и систематике подсемейства
полидиексодинин // Вопр. микропалеонтологии. 1986. Вып. 28. С. 24-28.
Левен Э.Я., Корчагин О.А. Пермо-триасовый биотический кризис и фораминиферы
// Стратиграфия. Геол. корреляция. 2001. Т. 9, № 4. С. 55-64.
Левен Э.Я., Щербович С.Ф. Фузулиниды и стратиграфия ассельского яруса
Дарваза. М.: Наука, 1978. 164 с.
Миклухо-Маклай А.Д. Верхний палеозой Средней Азии. Л.: Изд-во ЛГУ,
1963. 329 с.
Чжан Линь синь. Верхнекаменноугольные фузулиниды Кельпина и сопредельных
областей Синьцзяна // Acta Palaeontol. Sinica. 1963. Vol. 1. P. 219-227;
Vol. 2. P.
55-63.
Baghbani D. The Permian sequence in the Abadeh Region, Central Iran
// Occas. Publ. Earth Sci. Res. Inst. Univ. South Carolina. 1993.
N special
9B: Contributions
to Eurasian geology. P. 7-22.
Davydov V.I., Belasky P., Karavayeva N. Permian fusulinids from the
Koryak Terrane, Northeastern Russia, and their paleobiogeographic
affinity //
J. Foraminiferal Res. 1996. Vol. 26, N 3. P. 213-243.
Douglass R.C. Permian Tethyan fusulinids from California // Contributions
to paleontology. Wash. (D.C.), 1967. P. 1-13. (Geol. Surv. Prof.
Pap.; N 593-A).
Forke H. Biostratigraphie (Fusuliniden; Conodonten) und Mikrofazies
im Unterperm (Sakmar) der Karnischen Alpen (NaBfelgebiet, Osterreich)
//
Jb. Geol. Bundesanst.
1995. Bd. 138. S. 207-297.
Gobett D.J. Palaeozoogeography of the Verveekinidae (Permian foraminifera)
// Aspect of Tethyan biogeography. 1967. P. 77-91. (Syst. Assoc.
Publ.; N 7).
Gobett D.J. Permian fusulinacea // Atlas of palaeobiogeography
/ Ed. A. Hallam. Amsterdam etc.: Elsevier, 1973. P. 151-158.
Ishii K., Okimura Y., Ichikawa K. Notes on Tethys biogeography
with reference to Middle Permian fusulinaceans // The Tethys / Ed.
K.
Nakazawa, J.M.
Dickins. Tokyo: Tokai Univ. press, 1985. P. 139-155.
Kahler F. Obercarbon und Unterperm der Karnischen Alpen. Klagenfurt;
1985. 93 s. (Sonderheft der Carintia II; H. 42).
Kobayashi F. Middle Permian biogeography based on fusulinacean fauna
// Late Paleozoic foraminifera; their biostratigraphy, evolution
and paleoecology
and the Mid-Carbomiferous boundary / Ed. Ch.A. Ross et al. Wash.
(D.C.), 1997a. P. 73-76. (Cushman Found, for Foraminiferal Res.;
Special Publ.
36).
Kobayashi F. Middle Permian fusulinacean faunas and paleogeography
of exotic terranes in the Circum-Pacific // Ibid. 1997b. P. 77-80.
Kobayashu F. Tethyan uppermost Permian (Dzhulfian and Dorashamian)
foraminiferal faunas and their paleogeographic and tectonic implications
// Palaeogeogr.,
Palaeoclimatol., Palaeoecol. 1999. Vol. 150. P. 279-307.
Lee S.J. Fusulinidae of North China // Paleontol. Sinica.
Ser. B. 1927. Vol. 4, fasc. 1. P. 1-172.
Leven E.Ja. Les gisements Permiens et les Fusulinides de
['Afghanistan du Nord // Notes et memoires sur le Moyen-Orient. 1971.
Vol. 12.
P. 1-36.
Leven E. Ja. Early Permian fusulinids from the Central Pamir // Riv.
Ital. Paleontol. Stratigr. 1993. Vol. 99, N 2. P. 1-36.
Leven E.Ja. The Mid-Early Permian regression and transgression
of the Tethys // Pangea: Global environments and resources. Calgary,
1994.
P. 233-239.
(Canad. Soc. Petrol. Geol. Mem.; N 17).
Leven E.Ja.Permian stratigraphy and fusulinids of Afghanistan with
their paleogeographic and paleotectonic implications. Boulder (Colo.),
1997.
138 p. (Geol. Soc. Amer.
Spec. Pap.; N 316).
Leven E.Ja. The Permian stratigraphy and fusulinids of the Tethys
// Riv. Ital. Paleontol. Stratigr. 2003. Vol. 109, N 2. P. 267-280.
Leven E.Ja., Grant-Mackie J.A. Permian fusulinid foraminifera
from Wherowhero Point, Orua Bay, Northland, New Zealand // N.Z.J.
Geol.
and Geophys.
1997. Vol. 40. p. 473-486.
Leven E.Ja., Tacheri A. Carboniferous-Permian stratigraphy
and fusulinids of East Central Iran. 1. Gzhelian and / asselian deposits
of the
Ozbak Kuh Region
// Riv. Ital. Paleontol. Stratigr. 2003. Vol. 109. N 3. P. 267-280.
Lin R., Nassichuk W.W., Thorsteinsson R. The Lower Permian Fusulinacean
Sphaeroschwagerina in the Sverdrup Basin, Canadian Arctic Archipelago
// Pangea: Global environments
and resources. Calgary, 1994. P. 891-905. (Canad. Soc. Petrol. Geol.
Mem.; N 17).
Ozawa T. Permian fusulinacean biogeographic provinces in Asia and
their tectonic implications // Historical biogeography and plate
tectonic
evolution of Japan
and Eastern Asia / Ed. F. Taira, M.P. Tashiro. 1987. P. 97-110.
Ozawa Т., Kobayashi F. Carboniferous to Permian Akiyoshi Limestone
Group // Benthos'90: Guidebook for field trips: 4th Intern, symp.
on benthic
foraminifera. Sendai, 1990. P. 1-31.
Ross Ch.A. Development of fusulinid (Foraminiferida) faunas realms
// J. Paleontol. 1967a. Vol. 41, N 6. P. 1341-1354.
Ross Ch.A. Late Paleozoic Fusulinacea from Northern Yukon Territory
// Ibid. 1967b. Vol. 41, N 3. P. 709-725.
Ross Ch.A. Paleobiology of Fusulinaceans // Third North
Amer. Paleontol. Convention. Proc. 1982. Vol. 2. p. 441-445.
Ross Ch.A. Paleobiogeography of Fusulinacean foraminifera // Benthic
foraminifera: Benthos'90. Sendai: Tokai Univ. press, 1990. P. 23-31.
Ross Ch.A. Permian Fusulinaceans // The Permian of Northern Pangea.
В.; Heidelberg: Springer, 1995. Vol. 1: Paleogeography, paleoclimates,
stratigraphy.
P. 167-185.
Ross Ch.A., Ross J.R.P. Late Paleozoic accreted terranes
of Western North America // Pre-Jurassic rocks in Western North American
suspect
terranes
/ Ed. C.H.
Stevens. Los Angeles, 1983. P. 7-22.
Skinner J.W., Wilde G.L. Permian biostratigraphy and fusulinid faunas
of the Shasta Lake Area, Northern California // Univ. Kans. Paleontol.
Contrib.
1965.
Art. 6. P. 98.
Zhao Zhixin, Han Jianxiu, Wang Zegji. The Carboniferous strata and
its fauna from southwestern margin of Tarim Basin in Xinjiang. Beijin:
Geol.
Publ.
House, 1984. 187 p.
Ziegler A.M., Gibbs M.T., Hulver M.L. A mini-atlas of oceanic
water masses in the Permian period // The Permian system: Stratigraphy,
palaeogeography
and
resources / Ed. G.R. Shi et al. Melbourne, 1997. P. 323-343.
ОГЛАВЛЕНИЕ
Введение (Н.М. Чумаков).......................................................................................................................................
5
Часть I
ЭПОХА ПЕРЕХОДА ОТ ТЁПЛОЙ К ХОЛОДНОЙ БИОСФЕРЕ: КЛИМАТ ПАЛЕОГЕНА
Глава 1. Климат Земного шара в палеоцене
и эоцене по данным палеоботаники (М.Л. Ахметьев)............. 10
Часть II
КЛИМАТ И ПЕРЕСТРОЙКИ В ТЁПЛОЙ БИОСФЕРЕ (МЕЛ, ЮРА)
Глава 2. Общий обзор позднемезозойского
климата и событий (Н.М. Чумаков)............................................
44
Глава 3. Палеогеографические
перестройки и седиментация мелового периода
(М.А. Жарков, И.О. Мурдмаа, Н.И. Филатова)....................................................................................................
52
Глава 4. Количественные палеоботанические
данные о позднемеловом климате
Евразии и Аляски (А.Б. Герман)............................................................................................................................
88
Глава 5. Климатическая зональность
и климат мелового периода (Н.М. Чумаков).........................................
105
Глава 6. Глобальная экспансия
планктонных фораминифер: триас, юра, мел
(К.И. Кузнецова, О.А. Корчагин)..........................................................................................................................
124
Глава 7. Динамика и возможные
причины климатических изменений в позднем мезозое
(Н.М. Чумаков)........................................................................................................................................................
149
Часть III
ЭПОХА ПЕРЕХОДА ОТ ХОЛОДНОЙ К ТЁПЛОЙ БИОСФЕРЕ (ПЕРМЬ И РАННИЙ ТРИАС)
Глава 8. Палеогеографические перестройки
и обстановки седиментации в перми и раннем
триасе (М.Л. Жарков)..............................................................................................................................................
158
Глава 9. Палеобиогеография пермских
фузулинид (Э.Я. Левен)........................................................................
181
Глава 10. Палеофитогеография
пермского периода (С.В. Наугольных).............................................................
194
Глава 11. Палеопочвы перми и
раннего триаса (С.В. Наугольных)...................................................................
221
Глава 12. Климат и климатическая
зональность перми и раннего триаса (Н.М. Чумаков).............................
230
Часть IV
ГЛАВНЫЕ КЛИМАТИЧЕСКИЕ И БИОСФЕРНЫЕ СОБЫТИЯ ПОЗДНЕГО ДОКЕМБРИЯ
Глава 13. Ледниковый и безледниковый
климат в докембрии (Н.М. Чумаков)...............................................
259
Глава 14. Проблема климатической
зональности в позднем докембрии. Климат и биосферные
события (Н.М. Чумаков, В.Н. Сергеев).................................................................................................................
271
Заключение (Н.М.
Чумаков)................................................................................................................................
290
Conclusion...............................................................................................................................................................
296