| Главная | Библиотека | Форум | Гостевая книга |
105
Глава шестая
Характер роста и процессы
увеличения многообразия
Самая плодородная почва для распространения (эволюционного) синтеза, вероятно, лежит в развитии теории применительно к роли внутренних биотических факторов в установлении отличных друг от друга скоростей возникновения и вымирания видов, и в понимании того, как эти основополагающие макроэволюционные переменные складываются, давая в итоге собственно процесс увеличения многообразия. Дэвид Яблонски «Origination Patterns and Multilevel Processes in Macroevolution» |
Окаменелости обладают потенциальной возможностью давать прямое свидетельство
того, как изменения в стратегии роста могли оказывать воздействие на процессы
увеличения разнообразия с течением геологического времени. Новые стратегии
могли позволить некоторым видам воспользоваться новыми экологическими возможностями,
или же вносили свой вклад в их гибель. Характер эволюции групп организмов
в геологическом времени, обычно называемый динамикой клады, является предметом
интенсивных исследований в том сегменте сообщества палеонтологов, который
работает с беспозвоночными животными. Работа в этой области во многих случаях
обращается к обширным базам данных и математическим моделям, которые основаны
на нескольких переменных величинах.
Темпы изменения разнообразия, означающие, насколько быстро или медленно меняется
106
количество видов в группе с течением геологического времени, традиционно
связываются с внешними факторами окружающей среды. При объяснении эволюционного
события обычно идёт спор о двух альтернативных внешних факторах: Чёрная Королева
против Придворного Шута. В сценарии Чёрной Королевы главными направляющими
силами изменений являются биотические взаимодействия, а в сценарии Придворного
Шута самыми важными являются физические или абиотические возмущения, такие,
как изменения климата.1
«Внутренние» факторы, то есть те, которые связаны с физиологией и развитием
организма и являются результатом его эволюционной истории, также стали предметом
возрастающего интереса.
Исследования множества групп, которые жили в различные геологические эпохи,
показали, что противопоставление друг другу внешних и внутренних факторов
является ложным. Переменные компоненты истории жизни развиваются, находясь
в пределах, обусловленных внутренними особенностями включённых в этот процесс
организмов, которые изменяют окружающую среду, но также и подвергаются её
воздействию. Крупномасштабный пример такого рода – это появление фотосинтезирующих
бактерий, которые с течением геологического времени изменили атмосферу, высвобождая
кислород, что документируют минеральные отложения, возраст которых установлен.
Появление в атмосфере свободного кислорода также стало пусковым событием на
геологическом уровне, поскольку кислород вносит свой вклад в характер химического
выветривания горных пород. Некоторые из условий окружающей среды, с которыми
должны были сталкиваться на протяжении своей жизни ныне вымершие организмы,
крайне сильно отличались от тех, которые окружают нас сегодня. Например, содержание
кислорода в атмосфере в различные отдалённые периоды времени было намного
ниже, или же намного выше, чем сегодня.
Также стоит подумать над значением концепции «глубин времени» для понимания
эволюции индивидуального развития в крупных родословных ветвях. Как теоретические
ожидания, так и эмпирические данные показали, что для процессов роста биологического
многообразия простые корреляции являются исключением.
Предрасположенность к видообразованию или к устойчивости к вымиранию не может
107
сводиться случайным образом к единственной особенности; во всех случаях важно
рассматривать все переменные величины, относящиеся к истории жизни.
Некоторые различия в окружающей среде между нынешним миром и прошлыми эпохами
затрагивают переменные величины, которые постепенно изменялись по ходу геологического
времени. Другие же связаны с внезапными, катастрофическими событиями такого
масштаба, какой никогда не был зафиксирован на протяжении летописной истории
человечества. Некоторые из этих событий отмечены во времена, когда происходили
массовые вымирания, например, как вымирание динозавров. С ними связан исключительно
активный вулканизм. Последствия от удара астероида, который упал на полуострове
Юкатан, признаны многими. Возможно, столь же существенным было воздействие
событий, которые привели к появлению Деканских траппов на Индийском субконтиненте;
это одно из самых крупных проявлений вулканической активности на суше, когда
на континентальной коре образовались гигантские плато из лавы. Траппы плоскогорья
Декан представляют собой полмиллиона квадратных километров вулканических пород,
в десять раз больше, чем территория Швейцарии. Данный ландшафт состоит из
нескольких плоских областей, или траппов, каждая из которых была образована
поверхностью одного лавового потока и раскинулась на многие километры. Деканские
траппы являются последствиями вулканических явлений, масштаб которых не сравним
ни с чем из того, что известно нам из истории человечества. Даже извержение
Везувия и его хорошо известные последствия для Помпей бледнеют по сравнению
с ними.2
Радиометрическое датирование показало что извержения, которые привели к появлению
деканских траппов, охватывают отрезок времени примерно в 2 миллиона лет и
случились около 66 миллионов лет назад. Эти вулканические извержения не только
залили лавой обширные территории, но также выбросили в атмосферу большое количество
двуокиси серы и мелкой пыли. Климатические изменения, ставшие результатом
этих событий, оказали крупномасштабное воздействие на растительность и моря.
Морской планктон был, вероятно, в значительной степени уничтожен, что вызвало
целый каскад событий, распространявшихся по связям, характери-
108
зовавшим экосистемы того времени. Весьма вероятно, что хитросплетения экологических
взаимоотношений пришли в хаос. Во время ещё большего по масштабам среди известных
человеку эпизодов вымирания, на границе перми и триаса, вулканическая деятельность,
которая привела к образованию Сибирских траппов, продолжалась лишь около одного
миллиона лет, но охватывала гораздо большую площадь, чем та, которая связана
с Деканскими траппами. Образовавшиеся в результате этого вулканические породы
в настоящее время занимают территорию примерно в 2 миллиона квадратных километров,
меньше трети от оцениваемой первоначальной площади, и всё же почти равны по
площади территории Западной Европы. Изменения в химическом составе океана,
связанные с этой крупномасштабной вулканической деятельностью, нанесли огромный
удар по эволюции жизни в масштабах всего мира.
Крупномасштабные акты вымирания оказали воздействие на историю жизни на Земле,
и восстановление многообразия групп, которые не полностью вымерли, представляет
собой большой интерес и требует понимания особенностей окружающей среды и
видов организмов, характеризующих мир после вымирания.
Повышение кислотности океана
в прошлом и в наше время
После окончания пермского периода, когда произошло самое большое из когда-либо случавшихся событий массового вымирания, ранний триас характеризовался нестабильностью окружающей среды. Из работ геохимиков нам известно, что уровни кислотности океана и количества кислорода в атмосфере колебались. Изменчивые условия оказывали воздействие на метаболизм и в конечном счёте на характер роста у различных групп организмов, включая тех, кто строит скелет из карбоната кальция, например, рифообразующие кораллы (см. главу 10). Повышение кислотности океана – снижение значения pH – воздействовало на характер роста аммонитов и других моллюсков, кораллов
109
и большинства, если не всех остальных групп морских беспозвоночных. Отмеченная
вулканическая деятельность привела к достаточно резкому выбросу метана, а
также к увеличению концентрации двуокиси углерода не только в атмосфере, но
также и в океанах. Это снизило возможность выпадения в осадок морского карбоната
кальция и потенциальную возможность обызвествления у организмов, живущих на
всех уровнях толщи воды (бентос, планктон) – «кризис биокальцификации», который
исследовали мой коллега Хельмут Вайссерт из Швейцарской высшей технической
школы Цюриха и многие другие.
Исследование ископаемых морских беспозвоночных и истории их жизни зачастую
происходит параллельно с изучением изменений в океанах, что сводит воедино
геохимию и палеонтологию.
Точно так же, как в раннем триасе, в наше время происходит повышение уровня
кислотности морской воды по всему миру, поскольку океаны поглощают всё увеличивающееся
количество двуокиси углерода в атмосфере. Считается, что это закисление океана
является главной угрозой для морских экосистем в ближайшем будущем. Эксперименты
показали, что это изменение вредит способности к воспроизводству и росту по
крайней мере у некоторых видов веслоногих ракообразных, улиток, морских ежей
и офиур. Поскольку изменение происходит очень быстро, многие виды вряд ли
смогут выжить. Но не на все группы это воздействие оказывается одинаково.
Сэм Дюпонт и его коллеги из Швеции представили экспериментальные свидетельства
того, что повышение кислотности океанской воды оказывает положительное воздействие
на личинок морской звезды Crossaster: культивируемые при низких значениях
pH личинки растут быстрее, без видимого влияния на выживание или формирование
скелета. Это был первый отчёт о положительном влиянии повышения кислотности
океанской воды на темп роста у какого-либо беспозвоночного. Различающиеся
эффекты воздействия на историю жизни организмов должны отвечать за то, кто
выживет, а кто станет жертвой при наступлении события массового вымирания
и в ходе последующего восстановления биосферы. Как сказали Арно Брейяр и его
коллеги, «в выживании групп живых организмов играют свою роль и хорошие гены,
и хороший расклад дел».
110
Основные типы эволюции
личинок в океанах
Многие виды морских беспозвоночных, обитающие на морском дне, обладают свободноживущей
личиночной стадией, которая служит для расселения и очень сильно отличается
от взрослой особи морфологически и экологически. Большинство морских беспозвоночных
использует одну из двух различающихся между собой стратегий личиночного развития,
прежде чем осядет на дно и претерпит метаморфоз в ювенильную форму. При планктотрофии
личинки питаются в толще воды на протяжении недель или месяцев. При альтернативной
стратегии – не связанной с питанием лецитотрофии – личинки питаются желтком
яйца в течение гораздо более короткого времени, которое они тратят на жизнь
в толще воды. Как уже предсказало исследование распределения этих стратегий
по эволюционным древам ныне живущих видов, в ходе эволюции между ними должны
были происходить многократные смены, даже в пределах отдельно взятой группы.
Вопрос, который заставляет проводить множество исследований различных беспозвоночных,
состоит в том, чтобы понять, были ли эти изменения беспорядочно распределены,
или же концентрировались в определённые периоды времени. В целом принято считать,
что их движущими силами, вероятнее всего, были возникновение хищничества на
океанском дне, когда в процессе эволюции появились новые группы или новые
стратегии хищничества, или же изменения в окружающей среде. В дополнение к
академическому интересу к этому аспекту эволюции, целью данных исследований
также является изучение потоков питательных веществ и экологических взаимодействий
в океане, которое в конечном счёте оказывает влияние на рыболовство.
Работа с ископаемыми остатками организмов может дать прямую информацию о крупных
изменениях сообществ организмов в океанах прошлого, связанных с новыми стратегиями,
появляющимися в истории жизни организма. Хороший пример этого – возникновение
планктотрофии при переходе от кембрия к ордовику, примерно 490 миллионов лет
назад. Этот период времени очень важен, поскольку в это время зарегистрировано
резкое увеличение разнообразия жизни по всему миру и появление всех основных
групп ныне живущих организмов.
111
Прямое свидетельство эволюции стратегии личиночного развития получено для
ископаемых брюхоногих моллюсков. В древние времена, как и в наши дни, некоторые
виды имели прямое развитие и, как в случае лецитотрофных личинок, большее
количество желтка в их яйцах означало, что у них были более крупные оболочки.
Сами оболочки яиц редко сохраняются в ископаемом состоянии, потому что они
очень тонкие и мелкие (обычно менее 1 мм). Но их естественные слепки нередки.
По-немецки их называют Steinkerne (ядровые камни), и они во многих
случаях являются побочным продуктом работы палеонтологов, которые изучают
конодонтов, возможно, находящихся в самом основании родословного древа позвоночных,
растворяя породу химическими веществами и исследуя остаток. Поскольку размер
Steinkerne отражает количество желтка, мы можем с уверенностью делать
вывод о том, было ли животное планктотрофным. Александр Нутцель и его коллеги
измерили сотни этих Steinkerne из позднего кембрия и раннего ордовика
и напрямую зарегистрировали появление планктотрофных форм при переходе между
этими периодами приблизительно 490 миллионов лет назад.
Для раннего ордовика характерно многообразие бентосных форм, кормившихся взвесью
и прикреплявшихся к морскому дну. Некоторые исследователи предположили, что
эта особенность запустила эволюцию планктотрофии как стратегии избегания хищничества.
Очевидно, что этот вероятный сценарий подразумевает то, что снабжение питательными
веществами в мировом океане было достаточным для поддержания существования
плавающих личинок. Фактически, это стало результатом того эффекта, который
оказали на динамику океанских вод тектонические события в раннем ордовике.
Биологи, имеющие дело с океанической жизнью, начинают уделять пристальное
внимание работам палеонтологов в этой области, пробуя понять те условия, которые
оказывали влияние на организмы в прошлом, и тем самым предугадать, какое воздействие
могли бы оказать глобальные изменения окружающей среды на современные организмы.
Поток информации движется в обе стороны, и итогом этого становится тщательное
изучение истории жизни современных видов, которое
112
даст самое подробное понимание эволюции стратегий личиночного развития в летописи окаменелостей.
Климатические изменения и эволюция
индивидуального развития у млекопитающих
Исследования характера распределения биологического многообразия и изменений
окружающей среды в разрезе геологического времени показало, что в фаунах позвоночных
имели место эпизоды изменений, вызванные климатическими изменениями, но были
также и такие, при которых не зарегистрировано никаких следов изменений. В
ходе климатических изменений в неогене Северной Америки, которые привели к
преобразованию обширных областей в пустыни или в очень сухие местообитания,
возникло несколько новых групп млекопитающих. Была выдвинута гипотеза о том,
что это произошло путём схожих онтогенетических изменений в нескольких родословных
линиях одновременно. Передвижение скачками на задних конечностях возникло
в сухих местообитаниях по всему миру (напр. у кенгуру в Австралии). Этот вид
передвижения связан с различными пропорциями конечностей, которые появляются
в процессе эволюции путём возникновения различий в скорости или времени начала
роста у разных видов.3
Это означает, что климатические изменения в итоге могли оказать одновременное
воздействие на характер индивидуального развития в различных группах живых
организмов, которые приобретают в процессе эволюции схожие адаптации к новым
средам обитания.
Изменение климата может приводить к эволюционным изменениям размеров, которые
обычно связаны с изменениями в истории жизни. Виды лошадей, когда-то широко
распространённые и в изобилии представленные в североамериканских фаунах,
демонстрируют быстрое уменьшение в размерах перед своим исчезновением в плейстоцене
Аляски. Возможно, что естественный отбор оперирует переменными компонентами
истории жизни, ведущими к эволюционным изменениям размеров тела. Исследование
роста и продолжительности жизни, оставивших свой след в гистологии зубов,
может выявить такие изменения. С точки зрения морфологии, из-за аллометрических
отношений изменения в размерах во многих случаях означают изменения в
113
Рисунок 30. В процессе эволюции прыгающие млекопитающие независимо появились во многих пустынных областях по всему миру ближе к концу миоценовой эпохи. Пример такого рода – сумчатое млекопитающее Argyrolagus scagliai из Аргентины.4 Хвост, вероятно, был намного длиннее, и здесь изображены только сохранившиеся позвонки образца из Museo Municipal de Mar del Plata. У современных форм и, возможно, также у прыгающих аргиролагид большая разница в размерах конечностей развилась путём изменения индивидуальных скоростей дифференциации скелета. Из Simpson 1970, с изменениями. |
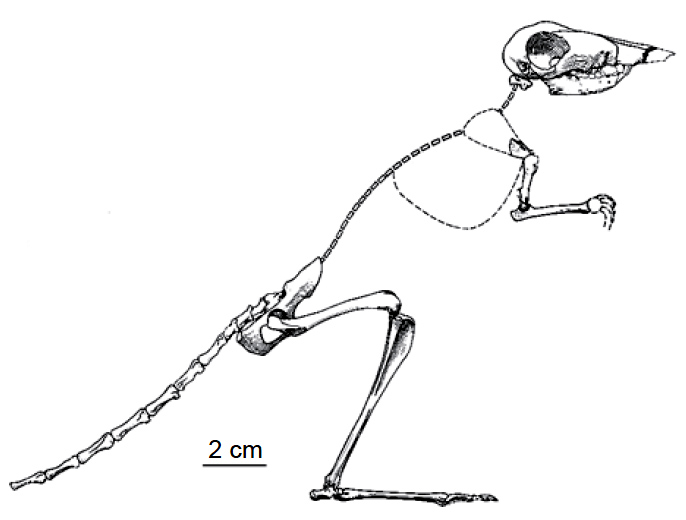
форме анатомических структур организмов. Многочисленные примеры этого процесса
возникли параллельно во многих группах животных в Африке около трёх миллионов
лет назад. Аналогичные явления, возможно, происходили в разное время на других
континентах.
Эволюция млекопитающих в Южной Америке – это благодатный объект для исследований
в области палеонтологии позвоночных, но лишь недавно знание хронологии и репрезентативность
выборок фауны стали достаточно хорошими, чтобы позволить нам уверенно исследовать
характер эволюции. Панчо Гоин, Фреди Карлини и их коллеги из Ла-Платы недавно
заметили, что значительные изменения в фауне млекопитающих совпадают с резким
снижением глобальных температур в самом конце
114
эоцена – раннем олигоцене. Они назвали это событие Bisagra Patagónica, или
«Патагонский шарнир», и оно было зарегистрировано в Южной Аргентине. Многие
группы сумчатых и родственников броненосцев вымерли в конце эоцена, и много
новых видов этих животных появилось в олигоцене.4
И что примечательно у раннеолигоценовых групп, так это их новая стратегия
в онтогенезе зубов – состояние, когда зубы постоянно растут, называемое гипсодонтией.
Эта особенность включает новую совокупность изменений в метаболизме и в истории
жизни, и, вероятно, возникла как ответ на износ зубов, вызванный главным образом
присутствием вулканического пепла в пище, как выяснил в ходе исследований
Рик Мэдден.
В противоположность вышеприведённым примерам, существенное снижение температуры,
которое произошло в Северной Америке в то же самое время, что и Bisagra Patagónica
в Аргентине (на границе эоцена и олигоцена, примерно 34 миллиона лет назад),
явно оказало лишь небольшое влияние на эволюцию млекопитающих. Опираясь на
превосходные коллекции многочисленных видов млекопитающих из отложений свиты
Уайт-Ривер с запада Соединёных Штатов, Дон Протеро и его коллеги показали,
что подавляющее большинство видов млекопитающих не изменилось морфологически,
несмотря на явно выраженные климатические изменения.
Изучение поверхности зубов показало, что должны были произойти изменения в
рационе, хотя форма зубов осталась той же самой. Возможно, млекопитающие Северной
Америки в том месте и к тому времени приобрели в процессе эволюции большую
фенотипическую пластичность, чем их современники в Южной Америке. С другой
стороны, хотя эоцен-олигоценовое изменение климата было глобальным, его последствия
могли отличаться на локальном уровне, при различной степени вулканической
активности на этих двух континентах, что в этом отношении является важным
фактором, который стоит учитывать.
Воздействие температуры на эволюцию индивидуального развития
115
Рисунок 31. Общая картина вымирания и роста биологического разнообразия в нескольких группах сумчатых млекопитающих во времена крупных климатических изменений, отмеченных для границы эоцена и олигоцена, которые Панчо Гоин и его коллеги назвали Bisagra Patagónica. По Goin, Abello и Chornogubsky, 2010, с изменениями. |
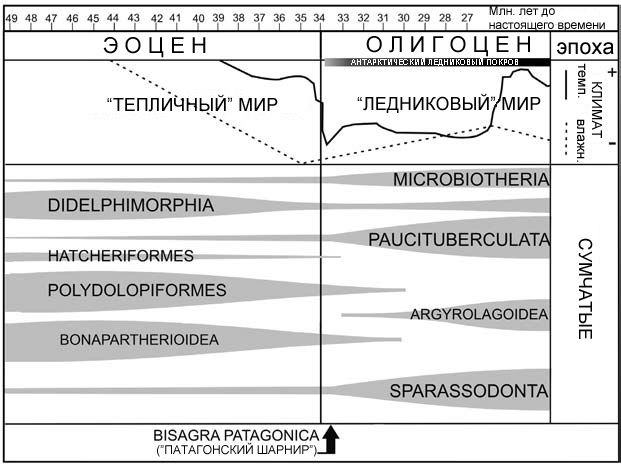
хорошо прослеживается во многих эмпирических исследованиях ныне живущих видов. Также существуют косвенные подсказки, которые предоставляет летопись окаменелостей. В эоцене, за миллионы лет до более холодного олигоцена, существовало множество разнообразных ископаемых фаун с такими же признаками, как у тропических фаун наших дней, даже в высоких широтах. В современной северо-восточной Колумбии примерно от 58 до 60 миллионов лет назад жила примечательная гигантская змея Titanoboa cerrejonensis, длина тела которой оценивается в 13 метров, а масса в 1140 килограммов. Поскольку температуры повышались и снижались с течением геологического времени, то же самое происходило с
116
верхним пределом размера холоднокровных существ, таких, как змеи, обмен веществ у которых находится под влиянием средней температуры окружающей среды. Джейсон Хэд с сотрудниками подсчитал, что для выживания змеи размером с Titanoboa требовалась бы круглогодично средняя температура от 30°C до 34°C. Для сравнения, среднегодовая температура сегодняшней Картахены на побережье Колумбии – около 28°C.
Юрские акулы
Способ воспроизводства и способность эволюционировать, адаптироваться в течение немногих поколений, могут изменяться на протяжении эволюционного времени как ответ на изменения окружающей среды абиотического и биотического характера совместно с внутренними изменениями. Пример этого явления показывает ранний этап увеличения видового разнообразия и адаптивной радиации современных акул и скатов в ранней юре, изученный Юргеном Криветом и его коллегами. После волны массового вымирания конца триаса начались оппортунистическое быстрое увеличение видового разнообразия и активная экологическая радиация у акул и родственных им форм, который были связаны с мелкими размерами тела, небольшой продолжительностью жизни и размножением путём откладывания яиц, и, вероятно, были вызваны этими же обстоятельствами. Эти особенности истории жизни позволили быстрее возникать экологическим новшествам в плане строения тела. Итоги исследования современных акул, скатов и химер показали, что у живородящих видов риск вымирания выше, чем у яйцекладущих, даже в случае пропорционально более высокого уровня выедания хищниками особей и яиц в случае яйцерождения. Так происходит из-за того, что эволюционная приспособленность живородящих видов оказывается очень низкой в случае гибели беременных самок, и это делает их более подверженными риску исчезновения. В ранней юре яйцерождение позволило акулам и их родственникам лучше приспособиться к изменению условий окружающей среды. Большая продолжительность жизни и живорождение у большинства ныне живущих видов акул делают их сильно уязвимыми в случае массового вымирания.
117
Выживание и рост многообразия
на раннем этапе эволюции млекопитающих
Очень немногие виды наших далёких зверообразных предков пережили пермо-триасовый
эпизод массового вымирания примерно 250 миллионов лет назад. Большинство родословных
линий прервалось, и многие из их представителей были плотоядными формами.
Выжила лишь малая часть этих ранних родственников млекопитающих, и среди них
был Lystrosaurus, наиболее многочисленный вид четвероногих позвоночных
во время раннетриасового периода восстановления фауны после массового вымирания.
Lystrosaurus принадлежит к числу дицинодонтов, получивших название
из-за своей специализированной зубной системы, включавшей лишь два клыка.
Эти травоядные были крупными, размером от крысы до быка, и разнообразными:
известно более семидесяти их родов. Lystrosaurus не только выжил,
но также широко распространился географически и был многочисленным в эпоху
раннего триаса.
На основании своих собственных палеонтологических исследований Дженнифер Бота-Бринк
установила, что у Lystrosaurus и других дицинодонтов была высокая
скорость роста по сравнению с другими современными им видами. Гистология более
чем сотни образцов костей дицинодонтов демонстрирует увеличенные каналы в
кортикальном слое, которые могли усиливать снабжение кислородом и питательными
веществами, способствуя более эффективной ассимиляции питательных веществ
и ускорению роста кости. Lystrosaurus и его ближайшие родственники
росли быстрее, чем его эволюционные «кузены». Исключительно высокая скорость
роста, вероятно, способствовала выживанию Lystrosaurus во время массового
вымирания в конце перми и его многочисленности в раннем триасе, в фазе восстановления
фауны после вымирания. Ускоренный рост дицинодонтов, возможно, позволял им
быстро достигать размеров тела, обеспечивающих безопасность от нападения хищников.
Кроме того, гистология костей предполагает быстрое достижение половой зрелости,
которое придало бы Lystrosaurus устойчивость к колебаниям условий
окружающей среды.
Это открытие, связанное с историей жизни, также соотносится с
118
обычностью ископаемых остатков этих животных, указывая на их многочисленность в отложениях, где отмечено наличие лишь немногих конкурирующих видов травоядных.
Острова как место экспериментов
в эволюции истории жизни
Острова – это любимый объект исследований для эволюционных биологов, поскольку
они стоят ближе всего к естественному эксперименту, с помощью которого проверяются
гипотезы. На ограниченном по площади месте с чётко очерченными границами значительно
легче документально зафиксировать ключевые физические и биологические переменные
величины и изучить, как они воздействуют на эволюционный процесс. Классические
исследования островных популяций – это исследования галапагосских земляных
вьюрков, ящериц рода Anolis с островов Карибского моря и плодовых
мушек с Гавайских островов. Озёра, такие, как Виктория в Африке, также являются
местами, где могут проводиться большие естественные эксперименты, поскольку
некоторые группы рыбы могут существовать только в своём озере.5
Острова также являются важным объектом исследований в области биологии развития
и изучения ископаемых форм. Многие из изучаемых организмов вымерли лишь недавно,
в течение последних сотен или тысяч, а не миллионов лет. В большинстве случаев
причиной их исчезновения было пагубное воздействие со стороны человека.
Существуют различные виды островов: старые и молодые, маленькие и большие,
океанические и континентальные. Континентальные острова – это такие, которые
в прошлом соединялись с соседними массивами суши. Они сами по себе находятся
на континентальном шельфе и могут соединяться, или же ранее соединялись с
материком посредством сухопутного перешейка. Бали, Куба и большинство островов
Средиземноморья являются континентальными. Океанический остров – это остров
в своём наиболее чистом виде. Он никогда не был связан с материком, и он образуется
посредством некоторых геологических процессов (чаще всего после извержения
вулкана), и поднимается со дна
119
океана. Галапагосские и Гавайские острова, Маврикий и Реюньон – это всё вулканические
острова. Что здесь касается моей темы, так это изоляция, которая имеет место
на островах, и вместе с ней специфические условия, которые привели к эволюции
различных видов истории жизни; многое из этой область может быть зафиксировано
в летописи окаменелостей. В отношении ископаемых млекопитающих нами изучены
одни из самых потрясающих или, по крайней мере, наилучшим образом известные
примеры эволюции на островах.6
В ходе одного из исследований подсчитано, что скорость морфологической эволюции
у млекопитающих, главным образом у грызунов, на островах выше, чем у материковых
популяций, вплоть до соотношения три к одному.7
Ускоренные изменения на островах предполагались на протяжении долгого времени,
и исследования их количественных характеристик включают, например, исследования
Адриана Листера, касающиеся популяции благородного оленя на острове Джерси.
Летопись окаменелостей показывает многочисленные резкие изменения размеров
в классических случаях слонов размером с пони, мышей размером с кролика, бегемотов
размером со свинью и оленей размером с собаку. После образования репродуктивной
изоляции в процессе эволюции возникают нетипичные размеры, тесно связанные
с изменениями в истории жизни.8
Из-за явления аллометрического роста итогом разницы в размерах являются различия
в формах организмов. Поэтому онтогенетический подход к изучению морфологической
эволюции островных млекопитающих является интуитивно наиболее верным.
Существует общая тенденция: мелкие млекопитающие эволюционируют в сторону
увеличения размера, а крупные виды – в сторону уменьшения размеров в течение
коротких отрезков геологического времени.9
Существует много примеров такого рода, вроде вымершего сицилийского слона
Elephas falconeri, у которого, по оценкам, рост взрослой особи был
менее 1 метра, а масса тела – 100 килограммов, лишь примерно 1 процент от
массы его материкового предка. Другие примеры родственных слонам животных
обнаружены на островах Ченнел, в двадцати милях от побережья Южной Калифорнии.
Там эволюционировал Mammuthus exilis, карликовый вид, произошедший
от Mammuthus columbi, одного из нескольких видов мамонтов, которые
бродили по Северной Америке на протяжении последнего миллиона лет.
120
Карликовые слоны были обычными обитателями Средиземноморья; они обнаружены
на Крите, Мальте, Кипре и Сицилии. Было высказано предположение, что черепа
этих животных, возможно, послужили первоисточником для греческих легенд о
циклопах: у слоновых черепов есть большое отверстие во лбу, с которым соединяется
хобот, и которое могло быть по ошибке принято за единственную огромную глазницу,
находящуюся в центре лба. В противоположность причудливой ассоциации носового
отверстия слона с единственным глазом мифического существа анатомически точный
подход делает альтернативный источник вдохновения, циклопическое состояние
уродливого младенца, менее вероятным. У мифического существа единственный
глаз расположен выше носа, а в случае уродства глаз развивается ниже его.10
Изучение млекопитающих с различных островов выявляет черты сходства у видов,
которые их населяют, и в адаптациях, которые они приобрели в процессе эволюции.
Многочисленные ископаемые остатки подтверждают существование в плейстоцене
вымерших слонов, оленей и бегемотов на многих островах Средиземноморья, а
также оленей и слонов на островах Японии и Юго-Восточной Азии. Таксономическое
сходство вероятнее всего объясняется способностями к плаванию в случае крупных
видов или способностью выживать на естественных плотах в случае мелких. Существуют
также экологические параллели в условиях, с которыми сталкиваются различные
млекопитающие в различных регионах. Каждый остров уникален, и в каждом случае
различные в силу естественных причин характер воздействия естественного отбора
и исторические обстоятельства играли свою роль в формировании фаунистических
и флористических сообществ. Однако, принципы и механизмы их воздействия, похоже,
универсальны.
Экологический контекст индивидуального развития подходит для понимания случаев
островных карликовости и гигантизма, явлений, в ходе возникновения которых
должны участвовать, переплетаться и взаимно влиять друг на друга биологические
переменные величины: размер, продолжительность жизни, возраст достижения половозрелости,
скорость роста, доступные ресурсы, конкуренция и потребности обмена веществ.
121
Рисунок 32. Череп карликового слона Elephas falconeri из позднего плейстоцена Сицилии. Изображение на основе образца, выставленного в Музее природы Зенкенберг, Франкфурт. Рисунок Клаудии Джоэл. |
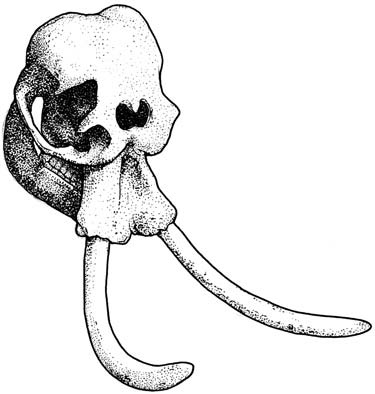
То, как организмы поддерживают процесс своего роста, зависит от темпов потребления
пищи и расхода энергии. Часть потреблённой пищи окисляется и используется
в энергетических превращениях, а остальное участвует в реакциях синтеза и
откладывается в виде биомассы. Были разработаны математические модели, основанные
на физиологических измерениях обмена веществ и процесса роста, и они помогли
установить, что у животных из различных групп работают одни и те же общие
принципы отношений.11
Это может объяснить, почему мы наблюдаем параллельное развитие морфологических
особенностей на различных островах.
Есть два пути, приводящих к быстрой эволюции изменений размера на островах.
Позднее наступление половозрелости может привести к появлению большего размера
тела. Такая картина развития связана с появлением немногочисленного потомства.
С другой стороны,
122
оптимизация метаболизма под влиянием иных условий с иным, более ограниченным
выбором ресурсов, может привести к появлению способности к воспроизводству
в раннем возрасте и к небольшому размеру.
Как ископаемые млекопитающие ведут себя в условиях иного, меньшего количества
ресурсов, с которым им пришлось столкнуться на островах? Вновь и вновь ископаемые
остатки предоставляют документальные свидетельства значительной эволюционной
пластичности в стратегиях, связанных с индивидуальным развитием и физиологией,
во многих группах. Примером этого является Myotragus, близкий родственник
овец и коз, который вымер около пяти тысяч лет назад. Предок Myotragus,
вероятно, добрался до Балеарских островов около шести миллионов лет назад,
в то время, когда Средиземноморье представляло собой группу солёных озёр.
Myotragus, вероятнее всего, эволюционировал в условиях, которые очень
сильно отличались от тех, которые существовали на материке: при небольшом
количестве ресурсов и при отсутствии хищников. Островной Myotragus
стал исключительно мелким животным: самые крупные взрослые экземпляры достигали
максимального роста от 45 до 50 сантиметров от земли до холки и, вероятно,
весили не более 50 – 70 килограммов.
Как они росли, какова была их энергетическая стратегия? Гистологическое исследование
длинных костей Myotragus показало наличие по всему кортикальному
слою ламеллярной кости – вида кости, обычного в прочих случаях только среди
современных рептилий вроде крокодилов, которые являются эктотермными животными.
Гистология костей показывает, что Myotragus рос медленно, не имел
единой скорости роста на протяжении жизни, и это было связано с существенной
задержкой достижения скелетной зрелости. У Myotragus также была гораздо
большая продолжительность жизни по сравнению с его эволюционными кузенами.
Не затронутый деятельностью человека, Myotragus вёл медленную и долгую
жизнь на острове в Средиземноморье.
В целом считалось, что явление педоморфизма (сохранение во взрослом состоянии
инфантильных или ювенильных признаков) способно описать многое в эволюции
островных млекопитающих. Но организм сам по себе мозаичен,
123
и в нём каждый орган или часть тела, даже будучи объединённым с остальными
частями, управляется различными правилами относительного роста. Так, хотя
островная форма может обладать общим сходством с ювенильной особью предкового
вида, оно может быть лишь поверхностным, а многие из особенностей строения
будут демонстрировать специальные адаптации к новой экологии и к новым метаболическим
потребностям. Пропорции тела у островных форм нельзя объяснить просто как
результат остановки или замедления развития, что придало им сходство с ювенильной
стадией предка.
Открытие нового вида гоминид из плейстоцена индонезийского острова Флорес,
знаменитого Homo floresiensis, даёт нам ясный пример необходимости
рассмотрения онтогенеза при исследовании ископаемых остатков и важности явления
«мозаичной» эволюции. H. floresiensis был лишь около одного метра
ростом и был полностью двуногим; для него был реконструирован мозг, похожий
на мозг шимпанзе, объёмом 417 кубических сантиметров для примерно 30-килограммового
тела. Такой размер мозга озадачивает, поскольку этот вид делал орудия труда
и жил совсем недавно, а его предок-гоминид обладал намного большим размером
мозга по сравнению с ним – возможно, это был H. erectus, объём мозга
у которого был примерно 900 кубических сантиметров. Вот, почему некоторые
учёные утверждали, что H. floresiensis был ничем иным, как популяцией
карликов с патологическим состоянием, которое включало микроцефалию.12
Но, если не брать во внимание эту опровергнутую альтернативу, то, рассматривая
аллометрические отношения размера мозга к размерам тела, можно увидеть, что
объём мозга H. floresiensis лежит вне рамок ожидаемого, вне зависимости
от того, какой именно вид гоминид из потенциальных кандидатов предполагался
в качестве его предка.
Примеры чрезвычайно сильного сокращения размера мозга, выпадающего за ожидаемые
значения, определяемые с позиций аллометрического подхода, известны для других
млекопитающих. Элеонор Уэстон и Адриан Листер показали это в количественном
отношении для ископаемых гиппопотамов с Мадагаскара. Мозговая ткань требует
очень больших затрат
124
в плане метаболизма, и потому была выдвинута гипотеза о том, что этот орган
уменьшится в размерах в эволюционных последовательностях, где было выгоднее
экономить энергию. Отсутствие простого онтогенетического масштабирования –
не редкость в случае быстрого эволюционного уменьшения размеров. Например,
Мейке Кёлер и Сальвадор Мойя-Сола сообщили о значительной редукции мозга и
органов чувств у Myotragus в ходе его географической изоляции на
Майорке. В другом примере Луиза Рот сообщила в 1980-х годах, что островные
карликовые виды ископаемых хоботных не просто мелкие с сохранившимися масштабными
соотношениями (педоморфными) в пропорциях их посткраниального скелета, но
также демонстрируют отклонения, которые, вероятнее всего, являются ответом
на локомоторные и метаболические ограничения и адаптации, что было также показано
для многих ископаемых видов с островов Средиземноморья.
Картина, наблюдаемая у гиппопотамов, Myotragus и, вероятно, у Homo
florensiensis, имеет свою параллель в виде процесса одомашнивания, при
котором происходит сокращение размеров мозга и редукция органов чувств, что
уже отметил Дарвин в своей вышедшей в 1868 году книге
об одомашнивании. Фактически, в одной заметке некоторые авторы ошибочно
заявили, что морфология Myotragus – это результат не адаптации к
новому островному местообитанию, а адаптаций, являющихся итогом одомашнивания.
Пример важной особенности в истории жизни, которую стараются отбирать разводчики
– это достижение половой зрелости при небольших размерах. Эта особенность
явно выглядит более отзывчивой на применение искусственного отбора, чем особенности
относительного роста или строения тела.13
К пониманию факторов, принимающих участие в процессе быстрой эволюции, которую
демонстрируют ископаемые остатки, найденные на островах, можно также прийти
при изучении воздействия содержания животных в неволе в зоопарках и в других
местах. Эта задача была бы аналогична той, которую поставил перед собой Дарвин,
когда изучал изменения под воздействием одомашнивания, желая получить понимание
процесса эволюции в целом.
Содержание в неволе оказывает значительное воздействие на основные аспекты
био-
125
логии животных, такие, как размеры тела и продолжительность жизни. Изменения, происходящие на протяжении времени жизни отдельной особи, конечно, не наследуются, но можно представить себе, как эти условия неволи, особый режим отбора, могут привести к эволюционным изменениям, если они сохраняются на протяжении ряда поколений. В условиях неволи четыре основополагающих аспекта жизни отличаются от естественных условий: доступность пищи, отсутствие хищников, ограниченное пространство для жизни и меньшее разнообразие или меньшее количество видов, сосуществующих в одном местообитании. Эти аспекты также отличают друг от друга островную и материковую популяции.
Содержание
Благодарности vii
Пролог xi
1. Окаменелости, онтогенез
и филогенез
1
2. Эво-дево, пластичность
и модули
34
3. Онтогенез позвоночных
в ископаемом состоянии
46
4. Кости и зубы под микроскопом
66
5. Пропорции, рост и таксономия
92
6. Характер
роста и процессы увеличения многообразия
105
7. Ископаемые остатки и генетика
развития
126
8. «Недостающие звенья» и
эволюция индивидуального развития
141
9. Развитие у млекопитающих
и у человека
158
10. О трилобитах, ракушках
и букашках
176
Эпилог: есть ли
мораль
в палеонтологии развития? 195
Примечания 197
Библиография 213