В кайнозое, по сравнению с предшествующими эрами
фанерозоя, климатические реконструкции по данным палеоботаники имеют более
высокую степень достоверности. Принадлежность
высших растений, особенно цветковых, к тем или иным рецентным семействам
даже для начала палеогена удается устанавливать с большой степенью надежности.
От
30 до 50% таксонов голосеменных и покрытосеменных раннего палеогена идентифицируются
с современными родами. Для олигоценовой эпохи эта цифра возрастает до 80-85%,
причем для многих вымерших видов можно доказать их секционную принадлежность
с выходом на близкородственные рецентные виды. Климатическая локализация
ареалов последних позволяет использовать актуалистический подход, оценивая,
хотя и
провизорно, экологические условия и климатические обстановки существования
растений в палеогене. Особенно благоприятными оказываются те случаи, когда
рецентные аналоги ископаемых форм имеет рифугиальную природу с ограниченными
ареалами распространения.
Наряду с такими важнейшими параметрами климата как среднегодовая температура
и влажность, анализ систематического состава ископаемых ориктоценозов растительных
мегафоссилий, а также палинологических комплексов и даже отдельных их спектров
позволяет оценивать сезонное распределение температуры и осадков и их возможные
экстремальные значения в самый холодный и самый
теплый месяцы года, а также связанные с климатом такие показатели как световой
режим, продолжительность вегетационного периода и др.
Очертив климатические параметры, которые являются определяющими для существования
растений, и могут быть восстановлены при анализе ориктоценозов, перечислим
кратко те морфолого-анатомические признаки самих растений, с помощью которых
последние могут выступать в качестве индикаторов климатических обстановок.
К таким признакам относятся прежде всего жизненные формы, а также преобладающие
размеры отдельных органов растений, морфология листовых пластинок и особенно
характер края листьев, типы строения древесины, корневой системы, кутикулы
и др. Используя ископаемые растения в качестве климатических индикаторов,
нужно иметь в виду, что их рецентные аналоги могут иметь и несколько иные
ограничения
по температурам, что связано с длительным периодом их адаптации к меняющимся
условиям окружающей среды.
Высшие растения как климатические индикаторы
Рассмотрим на
примере некоторых родов, характерных для палеогеновых флор, но присутствующих
и в современной флоре Земного шара, климатические параметры
их обитания, для того, чтобы использовать эти показатели при оценке климатических
обстановок в историческом интервале от 65 до 33 млн лет назад.
Большинство из широко представленных в палеогене таксодиевых ныне являются
реликтами и распространены в пределах небольших по площади рефугиумов. Климатическим
индикатором природной флоры как в геологическом прошлом, так и ныне, является
Sequoia. По данным Д. Мая [Mai, 1995], оптимальные среднегодовые температуры
обитания этого растения колеблются от 11° до 15 °С, наиболее холодного -
от 8° до 10 °С, а самого теплого - от 13° до 19,5 °С. Значительно большие
амплитуды
зимних и летних температур характерны для ареала другого представителя сем.
Taxodiaceae - рода Glyptostrobus - еще одного рецентного реликта, но обычного
растения палеогена Северного полушария. Среднегодовые температуры тех районов
Китая, где он ныне произрастает, +17,7 °С, среднезимние +6 °С, а наиболее
теплого месяца — +30 °С. Современное распространение Taxodium - болотного
кипариса
ныне не выходит за пределы среднегодовой изотермы +13,3 °С, а среднегодовые
температуры холодных месяцев колеблются от +4,9° до +9,9 °С, хотя в Европе
в интродуцированных посадках он переносит и более низкие зимние температуры
от -2,6° до +4,3 °С.
Принято считать, что наиболее надежными климатическими индикаторами среди
цветковых являются пальмы. Ареалы большинства видов их лежат в тропиках,
т.е. не выходят
за пределы среднегодовой изотермы +18 °С. Однако некоторые из пальм обитают
в условиях более низких температур (в Новой Зеландии +12 °С, в Китае и Японии
+14 °С, в Северной Америке +15 °С). Еще более впечатляет возможность пальм
переносить кратковременные экстремальные похолодания. Например, Trachycarpus может переживать понижения температур до -11 °С (в Китае до -14 °С), Sabal до -10,5 °С, a Chamaerops, Levistona, Phoenix до -9 °С.
Для большинства древесных цветковых ограничителями их распространения являются
среднезимние температуры. Северная граница обычных для палеогена Евразии
родов - Cinnamomum (коричный лавр) и Persea (авокадо) в современной флоре
лимитируется
среднегодовой изотермой +13 °С. Как и практически все лавровые, они не переносят
заморозков. Рецентные виды родов Mastixia и Symplocos, составляющих в палеогене
основу так называемых мастиксиевых флор, не пересекают январскую изотерму
+10 °С. Для такого весьма распространенного листопадного древесного растения
палеогена
Северного полушария как Liquidambar, достоверно устанавливаемого
по листьям, плодоношениям и
пыльце, северный предел распространения ограничен изотермой +11 °С. Существованию
многих листопадных мезофильных растений весьма способствуют высокие летние
температуры в пределах южных частей их ареалов на границе умеренно-теплой
и субтропической зон. Д. Май [Mai, 1995] для Северного полушария приводит
широкий
спектр таких растений (Liquidambar, Juglans, Pterocarya, Idesia, Platanus,
Aesculus, Syringa, Nyssa, Ailanthus, Koelreutheria, Carya, Catalpa, Celtis,
Phellodendron и др.).
Опыт интродукции в Европе выявил крайние температурные градации в распространении
многих субтропических растений. Среднегодовые температуры порядка +8 °С (при
экстремальных зимних до -15 °С) являются предельными для Castanopsis,
Meliosma, Ficus, Symplocos, Alangiun, Platycarya. Для другой группы растений, включающих
Cinnamomum, Engelhardia, Masixia, Koelreutheria, Tetrastigma, Zizyphus и
все пальмы, среднегодовые температуры зон произрастания колеблются в пределах
+15°-19°С,
при температурах самого холодного месяца порядка от +6° до +10 °С.
Как известно, современная тропическая зона ограничена изотермой самого холодного
месяца +18 °С. При этом большие трудности возникают при разграничении типичных
растений тропиков от субтропических, обитающих в горных районах тропических
областей. При оценке климатических условий палеогена эти трудности еще более
усугубляются, если интерпретировать ландшафтную позицию таких растений. Родовой
спектр их чрезвычайно широк, поэтому оперируя ископаемыми находками, обычно
приходится рассматривать роды того или иного семейства в их полном объеме
тропическими, не выходящими за пределы изотерм +19° ... +21 °С.
Перечисленные выше примеры в большей степени касались рецентных и более древних,
в том числе и палеогеновых растений Северного полушария. Подобный набор таксонов
на родовом уровне свойственен и ориктоценозам Южного полушария, где в качестве
растений-индикаторов климата обычно выступают некоторые секции рода Nothofagus (южных буков), причем мелколистные представители секций Fusca и Menziesii являются показателями влажного умеренного климата, сопровождающегося снежным
покровом,
a Brassi - субтропического климата. Не менее надежными индикаторами являются
пальмы, казуариновые, древовидные папоротники, саговники, ногоплодниковые,
миртовые, имбирные и др.
Хорошо известно, что в ископаемых ориктоценозах палеогена, особенно отражающих
рипарийные условия местообитания, обычно присутствуют остатки водных и околоводных
растений. Экологически они являются в большей степени консервативными во
времени. Прогревание поверхностных слоев воды в летние месяцы, необходимое
для обеспечения
их жизнедеятельности, должно быть для водных папоротников Salvinia и Azolla,
а также Trapa не ниже +16° ... +18 °С, примерно такой же для многих Nymphaceae,
а для Hydrocharis она выше +10 °С [Mai, 1995].
Кроме определенных температурных параметров, многие ископаемые растения,
как и родственные им рецентные, требуют для своего существования определенную
влагообеспеченность.
Упомянутые выше представители некоторых родов, в том числе и таксодиевых,
в их рефугиумах ныне обитают при следующих показателях среднегодовых осадков:
Glyptostrobus - 1400 мм/год, Тахоdium - 1000 мм/год, Sequoia - до 800 мм/год.
Близки по этим показателям некоторые индикаторные растения, имеющие в современной
флоре более обширные ареалы в тропиках и субтропиках - Cinnamomum -1300 мм/год,
Mastixia - 1000 мм/год. Многие растения, как известно, более успешно произрастают
при определенной сезонной ритмичности в поступлении влаги, предпочитая оптимальную
обеспеченность ею в летние или, наоборот, в зимние месяцы. Сезонность поступления
влаги достаточно надежно устанавливается для ископаемых форм по строению
вторичной
ксилемы и всей проводящей системы в целом, а также кутикуле. Многие растения,
ориктоценозы которых происходят из красноцветных и пестроцветных, включая
и соленосные толщи, требуют для своего жизнеобеспечения ограниченного количества
влаги. К ним относятся жестколистные наннофилы Palibinia из семиаридной зоны,
а также виды Nitraria, характерные для пустынных и полупустынных областей
палеогена
Евразии. В Южном полушарии индикаторами засушливости климата являются некоторые
протейные и миртовые (Dryandra, Eucalyptus). Показателями высокой или низкой
влагообеспеченности является и пыльца некоторых растений. Находки пыльцы
Classopollis, Ephedra или Chenopodiaceae обычно связывают с засушливостью
климата, хотя эти
растения могут выступать и как галофиты, являясь пионерными растениями при
расселении на засоленных субстратах прибрежной зоны отступающих морских бассейнов.
Пыльца Aquilapollenites, весьма обычная для маастрихтских и датских
угленосных отложений
Северного полушария, напротив, служит индикатором гумидного климата.
Листопадные древесные растения - обитатели высоких широт, особенно Северного
полушария, в условиях влажного климата палеогена характеризовались крупнолистностью,
чему способствовал летний световой режим, обеспечивающий активное функционирование
всех органов растения и в ночные часы.
При восстановлении климата палеогена большую роль играет общая экологическая
оценка флор этого возраста. Она базируется на сравнении составов ископаемых
ориктоценозов с составами современных растительных ассоциаций и группировок
родовой, а иногда даже и видовой состав которых оказываются близкими. Это,
в первую очередь, относится к восстановлению древних мангровых ассоциаций
тропиков, важнейшим компонентом которых была пальма Nypa, плоды
которой часто встречаются
в прибрежно-морских отложениях палеоцена и эоцена всей тропической зоны Земли.
Это же относится к прибрежной растительности теплоумеренной и субтропических
зон Северного полушария, одним из эдификаторов которой являлся вымерший в
палеогене род Macclintockia. Надежным показателем распространения
древней болотной растительности
влажных или сезонно-влажных субтропиков, включая южную окраину умеренной
зоны, являются совместные находки важнейших растений-углеобразователей: болотного
кипариса, ольхи и ниссы. Показательным примером возможности надежной реконструкции
климатической обстановки служат находки в пограничном интервале эоцена и
олигоцена
в вулканогенных разрезах горной гряды Дарры-Даг в окрестностях г. Джульфы
(республика Азербайджан). В совместном захоронении встречены остатки пальм
Trachycarpus и Sabal, папоротники Acrostychum и Blechnum,
листья Myricaceae,
Lauraceae, Fagaceae и Myrtaceae вместе с побегами и женскими
стробилами Taxodium и Pinus,
а также мелколистные Ulmaceae, при абсолютном доминировании папоротников
и веерной пальмы Sabal [Ахметьев, Запорожец, 1989; Ахметьев, 1993].
Рассмотренный выше состав совместно встреченных родов высших растений в значительной
степени
является тождественным тому, который обычен для современных болотных лесов
Флориды. Это позволяет заключить, что территория Южного Азербайджана в конце
эоцена входила в зону переменно-влажного субтропического климата, близкому
тропическому. Обращаясь к климату современной Флориды необходимо заметить,
что
к тропической зоне относится только южная оконечность полуострова, большая
же часть его из-за периодических заморозков в зимнее время не может быть
включена даже в субтропическую зону.
Прочие климатические индикаторы палеоцена и эоцена
Важными индикаторами климата палеоцена и эоцена нередко выступают
встречающиеся в одних и тех же ориктоценозах с растениями остатки наземных
и морских животных:
крокодилов, болотных черепах и других представителей герпетофауны, а также
насекомых - термитов, многих фитофагов, позвоночных, акул. Оптимальные средневековые
температуры обитания рецентных миссисипских и флоридских аллигаторов +20°
- +25 °С, китайских +16° - +17 °С, а китайских болотных черепах +20° - +22
°С,
при экстремально высоких температурах самого теплого месяца, необходимых
для их воспроизводства (до +35 °С). Соответственно, для приведенных выше
групп
животных пороговые значения зимних температур ограничивались +1° - +5 °С
для североамериканских и +3° - +7 °С для китайских аллигаторов. Распространение
китайских болотных черепах не выходит за пределы январской изотермы 0 °С.
Есть
все основания полагать, что и в палеогене климатические показатели условий
оптимального существования этих животных были близкими.
В данном разделе мы не рассматриваем многие морские организмы, в частности
микропланктон, пространственное распределение которого в открытых морских
и океанических бассейнах, тесно связано с климатической зональностью, а также
кораллы, моллюски, морские ежи и др., в расселении которых климатические
факторы
являются определяющими. Точно также мы оставляем без рассмотрения литологические,
эдафические и изотопные критерии климатических реконструкций. Однако все
они учитывались при подготовки климатических карт на отдельные интервалы
палеогеновой
истории.
Критические замечания по поводу использования актуалистических подходов при
интерпретации климата палеогена (аллохтонность ориктоценозов, использования
для сравнения рецентных реликтовых растений, обитающих ныне в рефугиумах
в экстремальных климатических условиях, недоучет конкуренции и т.п.) не исключает
всего сказанного выше как базы для реконструкций климатических условий палеогена.
К методике климатических реконструкций по палеоботаническим данным
В этом разделе мы ограничимся
лишь кратким рассмотрением различных методик климатических реконструкций по
палеоботаническим данным, так как их содержательная
часть подробно изложена в специальных публикациях авторов этих методик и
не нуждается в дублировании (см. например, [Wolfe, 1993; Mai, 1995; Wolfe,
Dilcher,
2001]). При построении палеоклиматических карт нами были использованы результаты
разных авторов по анализу конкретных ориктоценозов (или иными словами, локальных
флор (florule)), валидных по своей представительности для обработки.
Наиболее ранние заключения о палеоклиматах палеогена восходят к середине
XIX в. - времени расцвета классической палеоботаники, когда в результате
сравнения ископаемых таксонов с их рецентными аналогами делались выводы о климатических
обстановках прошлого. Эти подходы, далеко не безупречные, демонстрировались
О. Геером, К. Эттигсхаузеном, Г. де Сапорта и многими другими. Позже именно
они трансформировались в один из наиболее надежных методов палеоклиматических
реконструкций. Он основан на картографическом анализе ареалов рецентных таксонов,
близких ископаемым, и последующим построением климатодиаграмм по одной или
нескольким точкам максимального "сгущения" ареалов с подбором базовых
данных по ближайшим метеостанциям и их обработке. Этот метод нашел широкое
применение в палеофитогеографических и палеоклиматических реконструкциях
и в нашей стране был модифицирован В.П. Гричуком в применении к палинологическим
данным. В большей степени он использовался для интерпретации климата позднего
кайнозоя, так как ошибки в определении, по крайней мере родовой принадлежности
ископаемых таксонов, в этом случае минимальны [Ни, Chaney, 1940; Friedrich,
1966; Mai, 1995 и др.]. Однако следует отметить, что рассмотренный методический
подход может найти применение лишь для монографически изученных флор с установленным
положением в системе абсолютного большинства ископаемых растений, видовое
разнообразие
которых должно быть не менее нескольких десятков.
Другой метод, в основе которого используются морфологические параметры ископаемых
листьев, получил название CLAMP (Climate Leaf Analysis Multivariate Program).
Он начал разрабатываться с конца 1970-х гг. американским палеоботаником Дж.
Вулфом [Wolfe, 1993; 1994] и усовершенствованный в последние годы Р. Спайсером
в сотрудничестве с А.Б. Германом и другими специалистами (с использованием
статистического пакета "CANOCO") [Herman et al., 2002]. У истоков
этого метода стояло известное наблюдение ботаников начала прошлого века [Baily,
Sinnot, 1915] о доминировании в тропической и субтропической зонах листьев
с цельнокрайней листовой пластинкой и, наоборот, превалировании листьев с
зубчатым краем в умеренной зоне. Преимущество метода CLAMP заключается в
возможности
использования для климатических реконструкций ограниченного числа таксонов
цветковых растений (но не менее 25 - 30 видов в конкретных флорах), при этом
систематическое положение ископаемых растений может быть достоверно и не
установлено. Важна лишь возможность учета необходимых параметров листовых
пластинок и уверенность
в приуроченности растений к одному и тому же слою изучаемого разреза. Этот
метод в последнее десятилетие уже применялся для интерпретации климатов палеогена
Австралии [Green-wood et al., 1992 и др.], Аляски [Спайсер, Герман, 1998],
Канады и расположенной к югу от нее территории США [Wolfe, 1993, 1995; Wolfe,
Dilcher, 2001 и др.], Северо-Востока России, Западной и Центральной Европы,
юга Дальнего Востока России и Казахстана [Ахметьев и др., 2004]. Всего обработано
более сотни локальных флор в основном внетропических областей и опубликованные
данные учтены в настоящей работе.
К основным климатическим параметрам, которые удается получить с применением
метода CLAMP, относятся среднегодовая температура МАТ (С); средняя температура
самого теплого месяца - WMMT (С); средняя температура самого холодного месяца
- СММТ (С); среднегодовое количество осадков - MAP (mm); среднемесячное количество
осадков в вегетационный период - MMGSP (mm); общее количество осадков в вегетационный
период - MGSP (mm); количество осадков, приходящихся на три наиболее сухих
месяца - 3DRIM (mm); продолжительность вегетационного сезона в месяцах -
LGS (months). Нетрудно видеть, что соответствующие параметры стремятся выявить
и климатологи при оценке современного климата.
Важным звеном реконструкций является установление связей между климатическими
параметрами, полученными в результате обработки конкретных флор в сочетании
с анализом распределения в них растений - климатических индикаторов. Необходимо
также учитывать представительство во флорах тех или иных географических элементов,
установленных на основании изучения конкретных видов и их сравнения с рецентными
аналогами, распространенными ныне в различных регионах Земного шара. В итоге
возникает возможность очертить области распространения основных типов зональной
растительности и перейти к построению карт с выделением палеоклиматических
зон для определенных временных срезов. Эти карты в своей содержательной части
наиболее близки к современным климатическим картам Земного шара Б.П. Алисова
[Большая Советская..., 1973] с отражением климатических поясов, а также к
картам Л.С. Берга [1927], хотя первые построены на основании особенностей
общей циркуляции
современной атмосферы, а вторые - на основе смены ландшафтных зон.
Говоря о соотношении типов растительного покрова с климатом для палеогенового
периода, удается с достаточной достоверностью очертить области распространения
основных зональных типов растительности, особенно для средних и высоких широт
Северного полушария и некоторых регионов Южного (Австралия, Новая Зеландия,
отчасти Южная Америка) с восстановлением наиболее вероятных климатических
параметров существования этих типов. Зональные типы растительности в палеогене
были представлены:
а) тропическими и паратропическими дождевыми лесами; б) вечнозелеными лавролистными
широколиственными лесами;
в) вечнозелеными склерофильными лесами;
г) листопадными широколиственными и смешанными мезофильными лесами; д) хвойными
лесами. С каждым из перечисленных типов растительности хорошо корреспондируется
процентное содержание таксонов с цельнокрайней листовой пластинкой, которое
меняется от 60 до 75% для паратропических лесов, до 20-40% для смешанных
мезофильных листопадных.
Азональная растительность, которую чаще всего и отражает состав ориктоценозов,
образована сообществами водных растений, а также прибрежно-морских, рипарийных
и болотных лесов. Для палеогеновых флор зонального и азонального типов с
высоким процентом цельнокрайних листьев давно отмечено, что содержание листьев
лавровых
находится в противоположных соотношениях с содержанием листьев бобовых. Первые
характеризуют субтропический гумидный климат, вторые - семиаридный.
Климатические ограничения для зональной растительности палеогена
Евразии
[по: D. Mai, 1995] |
|||||||
| Тип растительности | Среднегодовые температуры, °С | Среднегодовые колебания, °С | Температура самого холодного месяца, °С |
Абсолютный минимум, °С | Среднегодовое количество осадков (мм/год) | Количество засушливых месяцев | Особые отметки |
| Паратропические дождевые леса |
25 - 20 |
< 18 |
+19 - +30 (+11) |
-1,8 |
До 1000 |
0 (-4) |
Осадков в самый засушливый месяц более 60 мм. Заморозки нерегулярны |
| Субтропические дождевые и лавровые леса | 20 - 15 |
< 24 |
+13 - +6 (+16) (+5) |
-10 |
До 1000 |
0 (-4) |
|
| Теплоумеренные вечнозеленые лавровые леса |
16,5 - 13 (12) |
< 27 |
+6 - +1 |
-15 |
1000 - 3000 (600) |
0 (-4) |
|
| Теплоумеренные дождевые леса | 13 - 9 (15,9) |
20 - 30 (19) |
Более -2 |
-25 |
1000 - 3000 |
0 (-3) |
Температура теплого месяца более 20 °С |
| Дубовые и каштановые смешанные леса |
13 - 10 (16) (9) |
17,5 - 34 |
+4 - +8 (+6) |
-30 |
> 500 - 2500 |
0 (-3) |
С хорошо выраженным непродолжительным холодным периодом |
| Буково-дубовые смешанные леса | 10 - 6 (13) (4,5) |
15 - 30 (13) |
+2,5 - +10 (+4) |
-40 |
> 500 - 2500 |
0 (-3) |
Температура самого теплого месяца более 15 °С; 120 - 130 дней более 10 °С |
Данные ботаников о климатических параметрах современных зональных типов растительности
для их использования при климатической интерпретации кайнозойских флор Западной
Евразии сведены в табл. 1 [Mai, 1995]. Эту таблицу следует лишь дополнить климатической
характеристикой тропических дождевых лесов, которых, по мнению Д. Мая, не было
в палеогене средних и высоких широт Западной Евразии. Эти леса в условиях влажного
климата произрастали при температурах +25° - +28 °С и их распространение ограничивалось
среднегодовой изотермой +20 °С (минимальные среднемесячные температуры таких
лесов по Кеппену +18 °С). Максимальные колебания годовых температур у экватора
до 5 °С, у тропиков до 13 °С, а дневные колебания температур были в пределах
3°-16 °С, никогда не опускаясь ниже 0 °С. Среднегодовое количество осадков
от 1000 мм/год и более, а минимальное количество осадков, приходящееся на самый
засушливый период 75 мм/год. Минимальная относительная влажность за полугодовой
период 65%.
Комментируя составленную таблицу, Д. Май отмечал, что полоса влажного тропического
климата в эоцене распространялась от Южной Европы, через Малую Азию и далее
на восток вдоль северного побережья древнего Тетиса. Значительно шире, чем
ныне, в палеогене были распространены лавровые леса в центральных и южных регионах
Западной Евразии. Что же касается жестколистных субтропических лесов (harwood),
то в палеогене Западной Евразии фиксируются лишь отдельные их элементы. Значительно
шире они были распространены в Северной Африке. Листопадные леса и соответствующая
им климатическая зона занимали значительную часть территории внетропической
Западной Евразии.
К сожалению, сведения о флорах Земного шара и общая климатическая картина палеогеновой
эпохи для различных регионов по
палеоботаническим данным может быть представлена с разной степенью достоверности.
Для Северной Америки, Западной, Центральной и отчасти Восточной Европы, Казахстана,
Дальнего Востока и Северо-Востока России, Японии, Восточного Китая, Австралии,
Новой Зеландии, Индии, крайнего юга Южной Америки и Антарктических островов,
Западной Приэкваториальной части Африканского побережья и Северной Африки построение
климатических карт возможно для эпох, а по некоторым регионам - даже для веков.
Для территорий, занятых древними щитами - Канадским, Бразильским, Африканским,
а также для севера Восточно-Европейской платформы и практически для всей Сибирской
платформы и Антарктического материала - области широкого пояса развития палеогеновых
морских отложений в пределах Тетиса - палеоботанической информации настолько
мало, что удается, в лучшем случае, дать климатическую характеристику только
для отдельных эпох. Несколько большую детализацию обеспечивают палинологические
данные, которые были также учтены при подготовке текста и карт.
По существу, все сходятся во мнении, что начало
датского века совпадает с глобальным похолоданием, первым наиболее заметным
в истории "теплой биосферы" мезозоя.
Однако о причинах этого похолодания мнения расходятся. Значительная часть
специалистов, в первую очередь американских, поддерживает гипотезу Альвариша,
связавшего похолодание и биотический кризис на рубеже мела и палеогена с
аномальным импактным событием (падением крупного метеорита) на юге Североамериканского
континента. Следствием данного события, по мнению сторонников этой гипотезы,
явилось быстрое массовое вымирание, зафиксированное во многих группах биоты,
особенно морского происхождения. Что же касается самого похолодания, то оно
связывается с непродолжительным по времени, но весьма заметным снижением
объема солнечной энергии, поступающей на поверхность планеты из-за пыли космического
и вулканического происхождения.
Более умеренные сторонники импактной гипотезы, не исключая влияния импактных
событий на кризис, отводят ему только триггерную роль, полагая, что падение
крупного метеорита могло явиться лишь резонатором, активизирующим спрединговые
зоны и вулканизм. Это, в свою очередь, могло повлечь за
собою и другие биосферные изменения, в том числе и биотические. При этом обычно
обращалось внимание на массовые изменения в составе бентосной биоты (фораминифер
и др.) в меридиональной спрединговой зоне Индийского океана на "подходе" к
Индостану. На самом же полуострове в это время формировалась трапповая серия.
Вместе с тем немало специалистов, которые считают, что даже если импактные
события и имели место в пограничном интервале мела и палеогена, они не сыграли
сколько-нибудь существенной роли ни в причинах биотического кризиса, ни в глобальном
похолодании. В качестве контраргументов импактной гипотезе выдвигается следующее.
1. Вымирание на рубеже мела и палеогена происходило не внезапно, а постепенно,
иногда скачкообразно. Причинами вымирания были как эволюционные процессы, так
и изменения условий внешней среды. Они связаны с глобальным похолоданием, вызванным
общепланетарной высокоамплитудной регрессией, а также активизацией вулканизма,
особенно в пределах Тихоокеанского кольца. Формирование вулканических цепей
по периферии Тихого океана было растянуто на несколько млн лет, захватывая
маастрихт и даний. Сокращение биоразнообразия континентальной биоты, в том
числе и элиминация динозавров, могли быть вызваны нарушением пищевых цепей
в конце Маастрихта, когда прибрежная растительность, формирующая основные пастбища
динозавров, оказалась уничтоженной. Что касается наземной флоры, то ее смена
началась сначала в высоких широтах (Корякско-Камчатский регион) и лишь затем
сместилась в средние широты (юг материковой части Дальнего Востока России,
Сахалин, Япония, Северо-Восточный Китай) [Головнева, 1994]. Судя по многочисленным
публикациям по межтрипповым флороносным слоям Индии, каких-либо внезапных изменений
в составе флоры на рубеже мела и палеогена в тропиках не происходило.
2. Смена биоты в пограничном интервале происходила весьма избирательно. Больше
пострадал морской бентос и планктон, особенно карбонатный (изменения в составе
органикостенного фитопланктона и диатомовых водорослей не носили критический
характер), причем в большинстве случаев эти изменения начались еще в Маастрихте.
В меньшей степени пострадали наземные моллюски, пресноводные рыбы и, как будет
показано, наземная флора.
3. Связь повышенных содержаний иридия только с импактными событиями не очевидна.
Во-первых, иридиевые "аномалии" явно тяготеют к регионам развития
вулканизма, а во-вторых, в ряде разрезов фиксируется несколько последовательных
иридиевых аномалий, что требует признания серии последовательных импактных
событий. Имеются примеры аномального содержания иридия (выше на два порядка)
в костях динозавров, т.е. заведомо в маастрихтских отложениях [Моисеенко и
др., 1997].
Проблема глобального похолодания на рубеже мела и палеогена требует дополнительных
исследований и возможно в недалеком будущем удастся добыть новые факты, раскрывающие
специфику природных явлений этого необычного в геологической истории рубежа.
Климатическая зональность в палеоценовую эпоху по сравнению с завершающим этапом
меловой истории отличалась большей широтной дифференциацией и контрастностью
среднегодовых и сезонных температур, особенно в средних и высоких широтах Земного
шара. Наиболее характерно это было для начала датского века в связи с глобальной
регрессией. На фоне сравнительно малоамплитудных колебаний климата Зеландия
и начала танета, во вторую половину танетского века произошло заметное потепление,
которое достигло оптимальных значений в конце танета и совпало с известным
углеродным изотопным эпизодом. Однако все более проявляющаяся тенденция изменения
положения границы палеоцена и эоцена в сторону ее омоложения в итоге может
привести к изменению датировки этого эпизода и в этом случае климатический
оптимум палеогена будет считаться раннеипрским. В настоящее время имеющиеся
данные позволяют в палеоцене выделить следующие климатические зоны.
Зона умеренного и умеренно-теплого влажного климата Северного полушария. К
началу датского века в высокоширотном секторе Северного полушария обособилась
крупнолистная мезофильная геофлора, территориально слабо дифференцированная,
южные границы которой определяют пределы распространения северной умеренно-теплой
зоны на планете. Большинство флор, характеризующих эту зону, приурочено к угленосным
бассейнам, в которых на протяжении всего палеогена, как в акваториях, так и
на суше, происходило терригенное осадконакопление с формированием угленосных
толщ в прибрежных зонах и внутренних бассейнах. Этот тип высокоширотной геофлоры
получил разное название. Пионеры ее изучения - Д. С. Гарднер [Gardner, 1879]
и А. Энглер [Engler, 1879] называли ее арктотретичной, не определяя четко возрастные
позиции. А.Н. Криштофович [1955] выделил ее как
гренландскую, по наиболее известной датской флоре Западной Гренландии, считая,
что время существования этой флоры не выходит за рамки палеоцена-начала эоцена.
Л. Ю. Буданцев [1983], много лет посвятивший изучению рассматриваемой флоры,
называл ее арктической, ограничивая ее распространение лишь частью пространства,
занятой Бореальной флористической областью и двумя провинциями последней: Берингийской
и Тулеанской, климатическая природа которых была единой.
Южная граница собственно умеренно-теплой климатической зоны в начале палеогена
проходила близ южной оконечности Гренландии, захватывая север Скандинавии.
Восточнее эта граница пересекала северную часть Западно-Сибирской плиты, опускаясь
к югу в пределах Сибирской платформы до широты слияния Лены и Алдана. К Тихоокеанскому
побережью она выходила в районе Охотска, а далее к востоку захватывала Корякию
и Камчатку (за исключением крайнего юга). На Американском континенте рассматриваемая
граница проходила через Южную Аляску и Алеутские острова, пересекала Северную
Канаду и, оставляя к северу Канадский Арктический Архипелаг, протягивалась
к Южной Гренландии.
В пределах умеренно-теплой зоны сосредоточена основная масса угольных бассейнов
палеоцена Северного полушария, в том числе Западно-Гренландский, Баренцбургский
(на Шпицбергене), а также впадины, выполненные угленосными отложениями в низовьях
Лены на севере Сибири: Кунгинская, Кенгдейская, Сого и др., угольные бассейны
Корякин и Камчатки (бухта Угольная, бассейн р. Напаны и др.). Не менее многочисленны
палеоценовые угольные бассейны на Аляске и на территории Приарктической Канады.
Климатические параметры многих флор умеренной зоны рассчитаны с помощью методов
CLAMP (табл. 2). В первую очередь, это относится к флорам Северо-Востока России
и Аляски, тогда как для многих флор они выявлены ареалогическим способом.
Таблица 2
Климатические параметры некоторых палеоценовых флор
умеренно-теплой зоны Северного полушария,
рассчитанные методом CLAMP [Golovneva, 2000] |
||||||
| Флора | Среднегодовая температура, °С | Средняя температура холодного месяца, °С | Средняя температура теплого месяца, °С | Среднегодовое количество осадков, мм | Среднемесячные осадки в вегетационный сезон, мм | Продолжительность вегетационного сезона в месяцах |
| Баренцбург | 12,6 |
6,5 |
19,8 |
1826 |
146 |
7,4 |
| Стурвольская | 9,5 |
1,5 |
18,4 |
1716 |
119 |
5,9 |
| Рарыткин | 11,5 |
4,6 |
19,4 |
1722 |
129 |
6,9 |
| Напанская | 11,8 |
5,8 |
19,1 |
2048 |
175 |
7,0 |
| Малл | 10,3 |
2,8 |
18,8 |
1739 |
126 |
6,3 |
| Ридинг | 12,3 |
5,0 |
20,1 |
1494 |
105 |
7,3 |
| Стандартное отклонение |
1,8 |
3,3 |
3,1 |
430 |
23 |
1,1 |
К наиболее хорошо изученным высокоширотным умеренным флорам
дания относятся флоры Агатдален и Атаникердлук п-ова Нугсуак, о-ва Диско Западной
Гренландии,
мыса Вашингтон Северной Гренландии, а также Баренцбургской угленосной толщи
Шпицбергена [Буданцев, 1983]. На Аляске флора этого возраста недавно была открыта
к северу от хребта Брукс [Herman, Spicer, Kvacek, 2002]. На Северо-Востоке
России наиболее богатой в систематическом отношении является флора рарыткинской
свиты низовьев Анадыря, а в низовьях Лены флора так называемого хараулахского
типа [Буданцев, 1983]. Главными компонентами этих флор являются хвощи, полиподиевые
папоротники (Osmunda, Onoclea), гинкго, древние сосны и разнообразные
таксодиевые и кипарисовые (Taxodium olrikii, Glyptostrobus nordenskioldii,
Metasequoia occidentalis (= M. disticha), Cupressinocladus).
В спектре цветковых доминирующее значение имели представители рода Trochodendroides,
а также семейств Hamamelidaceae, Platanaceae (Platanus, Beringiophyllum),
Betulaceae (Corylites), Fagaceae (Fagopsis), Tiliaephyllum, Viburnum, Nyssa (?)
и др. Сравнительно небольшое содержание среди листьев цветковых растений цельнокрайних
морфотипов (до 17-25%)
и крупных размеров листьев, характерных для высокоширотных мезофильных флор,
позволяют предположить следующие климатические параметры существования этих
флор: среднегодовая температура от 10°-12 °С до 14 °С, при средне-зимних до
-4° до -6 °С (-7,8 °С для флоры Агатдален), при кратковременных экстремальных
понижениях температуры до -15 °С и среднегодовом количестве осадков до 1400
мм.
В позднем палеоцене (в Зеландии и танете, особенно в позднем танете) арктическая
флора приобрела черты умеренно-теплой, переходной к субтропической. Область ее
распространения представляла собой, по существу, экотон, сформированный у южной
окраины Бореальной области. Весьма показательными в этом отношении являются флоры
Центральной Аляски (Чекалун) и о-ва Куприянова. Флора Чекалун конца танета наряду
с разнообразными папоротниками (Anemia, Dennstaedtia, Onoclea, Osmunda),
таксодиевыми, гинкговыми и листопадными цветковыми (Trochodendroides, Carya,
Pterocarya, Corylites),
обычными для флор умеренно-теплого влажного климата, содержала пальмы (Sabalites)
и некоторые другие растения (Macaranga, Melanolepis), более характерные для южной
субтропической зоны. Во флоре о-ва Куприянова, кроме пальм, были найдены и саговники
[Hollick, 1936; Wolfe, 1972]. Среднегодовые температуры, оптимальные для существования
этих флор, по оценке Дж. Волфа, находились в пределах 10°-12 °С, количество среднегодовых
осадков было не менее 2000 мм. Индекс цельнокрайности колеблется от 30 до 40%
[Wolfe, Dilcher, 2001].
Зона умеренно-теплого влажного климата протягивалась южнее высокоширотного сектора
распространения арктической геофлоры и в Евразии, занимая значительную территорию
Бореальной флористической области. Судя по среднегодовым и среднелетним параметрам
зоны ее климат был более близок умеренно-теплому, а по высоким среднезимним температурам
приближался к субтропическому. В Западной Евразии и Северной Америке южная окраинная
часть Бореальной области с позиций флористики представляла собой экотон со смешением
листопадных древесных растений с листопадными и вечнозелеными субтропического
и даже тропического родства. Этот неширокий экотон разделял зону умеренно-теплого
климата от так называемого "паратропического", влажного, с высокими
среднелетними и относительно высокими среднезимними температурами, но, в отличие
от тропического климата, не исключавшего
кратковременных заморозков. В Центральной и Восточной Азии, где южная граница
Бореальной флористической области заметно смещалась на юг, соответствующая ей
южная граница зоны умеренно-теплого климата также устремлялась в средние широты
и в палеоцене проходила через центральную часть Великобритании и северную часть
Европы. Далее она пересекала Средний Урал и заметно отклоняясь к югу (особенно
в дании) к южным границам Восточного Казахстана, где к этой климатической зоне
должны быть отнесены бассейн р. Иль и территория Призайсанья. Далее к востоку
рассматриваемая зона охватывала всю территорию Среднего и Нижнего Приамурья,
Северо-Восточный Китай до широты Шандунского п-ова и Северную Японию. На Американском
континенте в эту зону следует включить территорию Канады и отчасти пограничные
с ней районы США.
Типичными флорами Западной Евразии, характеризующими эту климатическую зону,
выступали флоры позднего танета и пограничного интервала палеоцена и эоцена Северной
Великобритании и Ирландии и прежде всего флора о-ва Малл и Фарерских о-вов [Boulter,
Kvachek, 1989]. На Урале роль южной пограничной выполняет флора р. Лозьва, описанная
А.Н. Криштофовичем [1928]. На Дальнем Востоке типичными флорами этого типа являются
цагаянские флоры Зейско-Буреинского осадочного бассейна [Ахметьев и др., 2002],
маломихайловская флора низовьев Амура, флора бошняковской свиты Сахалина и тахобинской
свиты Восточного Сихотэ-Алиня, геткилнинской свиты Западной Камчатки, а также
близкие им по типу флоры Зайсанской впадины (Кара-Бирюк, Тайжузген) и Монголии.
На Американском континенте к зоне умеренно-теплого климата принадлежат флоры
формации Эдмонтон Южной Канады и верхних слоев угленосной формации Форт Юнион,
распространенной на севере Мидконтинента.
Флора о-ва Малл и соседних участков Северной Ирландии и Шотландии из верхнетанетских
отложений отличается присутствием в ее составе разнообразных папоротников (Onoclea,
Dryopteris, Osmunda, Gleichenites, Dennstaedtia), Ginkgo, хвойных
(Amentotaxus,
Cephalotaxus, Cunninghamia, Cupressinocladus, Doliostrobus, Fokienia, Glyptostrobus,
Sequoia, Taxodium и др.) и широким спектром цветковых растений, в составе
которых
наряду с Hamamelidaceae, Platanaceae, Cocculus (= Trochodendroides)
и Corylites встречены более термофильные Ushia, Fagopsis, Debeya,
Musophyllum,
Macclintockia и др. Близкой по типу является и листопадная мезофильная флора
Лозьвы на Урале, имеющая, как выяснено в последние годы, зеландский возраст.
Датская цагаянская флора, являющаяся типовой для многих флор Дальнего Востока,
также мезофильная, листопадная, но в отличие от флоры о-ва Малл содержит меньше
термофильных форм при сохранении того же спектра хвойных и доминирующих цветковых,
к которым, кроме Platanaceae, Trochodendroides, добавляются Nyssa, Celtis,
Tiliaephyllumи Beringiophyllum. В некоторых датских дальневосточных флорах (бошняковской на
Сахалине, тахобинской в Приморье) [Красилов, 1976; Аблаев, 1974] еще встречается
как рифугиальный элемент Nilssonia, в датских флорах Японии - саговники и пальмы.
Для южной части распространения флор Бореальной области, входящих в зону умеренно-теплого
климата, характерны среднегодовые температуры до 14°-16 °С, относительно высокие
среднелетние (до 20°-22 °С) и среднезимние температуры (порядка -4°-6 °С для
датского века и выше на 2°-3 °С для флор позднего палеоцена, при высокой влагообеспеченности
(до 1500-2000 мм/год)). Характерно, что большинство местонахождений флор происходит
из угольных бассейнов.
Зона влажного субтропического и паратропического климата Северного полушария. Эта
климатическая зона занимает средние широты Северного полушария. В Европе она
протягивается от Южной Англии к югу и востоку, захватывая всю Западную Европу,
а в Восточной Европе прослеживается на Украину, Среднее и Нижнее Поволжье и Южный
Урал, в Центральной Азии она занимает Северный Казахстан и Северо-Западный Китай
(верховья Иртыша), где флоры носят смешанный характер, являясь переходными к
гумидным флорам тепло-умеренной зоны. Тип флоры, характеризующий зону паратропического
климата, был назван А.Н. Криштофовичем гелинденским, по типовому местонахождению
в Бельгии. В Западной Европе, кроме зеландской флоры Гелиндена, к этому типу
могут быть отнесены раннетанетская флора Мена и позднетанетская из травертинов
Сезанна во Франции, палеоценовые флоры (без уточнения возраста) Вейс-Эльстерского
грабена (Германия), а также флоры палеоцена Румынии и Сербии. В России наиболее
богатые местонахождения флор гелинденского типа известны из верхнесызранских,
нижнесаратовских и верхнесаратовских (= камышинских) отложений Поволжья в пределах
Ульяновско-Саратовского прогиба (Вольск, Никулино,
Ашут и др.) и на правобережье Волги у г. Камышина (конец зеландия-танет). На
Южной Урале и в Западном Казахстане имеются, по меньшей мере, еще два богатых
местонахождения флор этого типа (Романколь и Тыкбутак). Доминирующими компонентами
гелинденской и камышинской флор являются лавровые (Sassafras, Daphnogene,
Lauraceaephyllum,
Litsea, Ocotea), вечнозеленые буковые (Ushia, Dryophyllum), а также аралиевые,
пальмы (Trachycarpus, Nypa), мастиксиевые и другие растения тропического и субтропического
родства, при практически полном отсутствии хвойных. В камышинской флоре, а также
в одном из местонахождений Саратовской области, встречены плоды мангровой пальмы
Nyra. Климатические параметры описываемой зоны даны в табл. 1. Палеоценовые флоры
Центральной и Южной Европы более мелколистны, отражая, возможно, сезонность климата
[Ахметьев, 1993; Макулбеков, 1997].
В палеоцене от Каспийского и до Желтого моря через Среднюю Азию и районы Центрального
Китая прослеживалась полоса аридного климата. Ближе к Тихоокеанскому побережью
она делилась на две части. Северная, примыкающая к Шандунскому п-ову, по температурным
параметрам должна относиться к зоне умеренно-теплого климата, южная -к субтропическому.
Флористическим индикатором этой зоны является Palibinia, имеющая жесткие с толстой
кутикулой листья. Ее условно относят к семейству Anacardiceae. В субтропической
зоне Китая этому растению нередко сопутствуют узколистные лавровые и восковниковые.
Характерно, что местонахождения палибиний встречаются только в пестроцветных,
иногда в соленосных толщах. Слабую влагообеспеченность этой зоны подчеркивает
и ксероморфный облик этих растений.
В Северной Америке "паратропические", субтропические и близкие к тропическим
флорам получили распространение отчасти на территории Южной Канады, но, главным
образом, в западных и центральных районах США, а также в обрамлении морского
залива, отходящего к северу от современного Мексиканского и достигавшего района
слияния рек Миссисипи и Миссури. Климатические параметры, рассчитанные по большинству
из изученных флор методом CLAMP, свидетельствуют, что среднегодовые температуры
Зеландия Центрального Техаса (флоры серии Уилкокс) [Wolfe, Dilcher, 2001] около
59-60 млн лет назад были +19 °С, т.е. близкие современным для этого же района.
Однако климат был более мягким, с перепадом тем-
ператур в течение года не более 11 °С (сейчас около 20°С) и более влажным (более
2000 мм/год). Распределение осадков, как и ныне, было сезонным с соотношением
летних максимумов к зимним минимумам за три наиболее сухих и три наиболее "влажных" месяца
1200-1600 мм, т.е. климат имел все черты муссонного. В раннем танете (57-58 млн
лет назад) среднегодовые температуры возросли до +23° - +24 °С и климат стал
еще более мягким с перепадом годовых температур не более 9°С. Сезонность его
оказалась выраженной в еще большей степени. К середине танета среднегодовые температуры
еще более возросли, годовое количество осадков увеличилось не менее чем на 500
мм, но влагообеспеченность оставалась по-прежнему сезонной. В более низких широтах,
по крайней мере до 30° с.ш., климат носил признаки экваториального. Вдоль северной
окраины прибрежной равнины Мексиканского залива на территории штата Арканзаса
в это время происходило латеритообразование и формировались бокситы. Оптимальные
среднегодовые температуры, характеризующие флоры Беллингхэм Бэй и Слайд Мембер
на северо-западе США (штат Вашингтон) были около 15°-17 °С.
Значительно труднее интерпретировать климатическую обстановку в палеоцене внутренних
равнин США, которые в связи с начавшимися поднятиями Скалистых гор, после осушения
в дании Кэнненболского моря, оказались отделенными от притихоокеанских районов
США. Поэтому повсеместно фиксирующиеся в пределах внутренних равнинах более низкие
температуры могут скорее быть связаны с более высокими абсолютными отметками
возвышенных равнин. Среднеиюльские температуры по листовым флорам из формации
Поудер этого района по оценке Дж. Вулфа [Wolfe, Dilcher, 2001] методом CLAMP
определены в +31 °С, при очень низких перепадах среднегодовых температур (до
7°С).
Тропическая и экваториальная зоны. Районы распространения в палеоцене влажной
тропической флоры отчетливо выделяются в Центральной Америке [Graham, 1994],
а также в современной приэкваториальной зоне Южной Америки, где на восточном
побережье Бразилии обнаружены остатки мангровых (Nypa), а также Bombacaceae и
Psidium. В Африке влажные тропические леса, обрамляющие семиаридную
саванну, занимали территорию современной Сахары, протягивались в прибрежные части
Тетиса,
омывающего северное побережье Африки, а также были развиты в пределах современной
Нигерии,
Западного Конго и Либерии, а на востоке - от Большого Африканского Рога до Занзибара.
Если в составе палеоценовых флор Южной Америки доминировали неотропические элементы,
одновозрастная африканская флора была образована представителями Палеотрописа
и объединяла много тропических семейств Старого Света (Arecaceae, Ebenaceae,
Fabaceae, Lauraceae, Monimiaceae, Moraceae, Myrsinaceae, Rutaceae, Sterculiaceae,
Tenstroemiaceae и др.).
Палеоценовая флора Индии представлена исключительно таксонами тропических семейств.
В датских отложениях Ассама палинологами установлено присутствие представителей
семейств Nymphaceae, Meliaceae, Olacaceae, Rhizophoraceae, Myrtaceae и др. [Lakhampal,
1974]. Вдоль морских побережий, обрамляющих Декканское базальтовое плато, повсеместно
распространялись мангровы (Nypa, Rhizophora и Sonneratia). В палеоценовых отложениях
Индии обнаружено не менее двух десятков различных родов пальм. В Восточной Азии
северная граница влажной тропической зоны протягивалась к северу от побережья
Южного Китая, разделяя расположенные к югу бассейны угленакопления, от лежащих
к северу эвапоритовых бассейнов на территории провинций Гуандун и Гуаньси [Guo
Shuangxing, 1979].
Зона субтропического гумидного и семиаридного климата Южного полушария. В настоящее
время еще очень мало данных по флорам южной части Африканского континента, которые
могли бы служить основой для восстановления климатической обстановки палеоцена
этой территории. По данным М. Бэмфорд [Bamford, 2000], изучившей ископаемые древесины
Намибии, аридизация западного побережья Южной Африки стала проявляться с начала
палеогена, что связывается с существованием уже в это время холодного Бенгальского
течения.
В Южной Америке ряд палеоценовых флор (Фюнес, Сюр де Рио Дезеадо и др.) известны
из южных районов Аргентины и Чили. Ископаемые древесины из местонахождения Голф
Сан Джордж (46° ю.ш.) указывают на сходный характер этой палеоценовой флоры и
растительности с таковыми современной Южной Бразилии развитыми на 20° севернее
[Romero, 1986]. Эта флора носит аллохтонный характер и включает растения, обитавшие
в различных экологических условиях от мангровых болотных тропических лесов до
саванных редколесий. Здесь же были обнаружены и остатки крокодилов. Наряду с
неотропическими (южноамериканскими) элементами
флоры встречены некоторые австрало-азиатские - Dacricarpus, Dicranopteris,
Gunnera,
Anacolosa, Nypa. Флора лагуны дель Хунко Северо-Западной Патагонии насчитывает
несколько десятков видов, из которых около 70%, имеют цельнокрайнюю листовую
пластинку. Это свидетельствует о принадлежности ассоциации тропическому или субтропическому
дождевому лесу. Доминирующими компонентами флоры являются неотропические Annoba,
Cochlospermum, а также некоторые таксоны современного Голантарктического
царства (Lomatia, Peumus). Nothofagus - наиболее умеренный в климатическом отношении
голантарктический
элемент встречен не был [Romero, 1986].
Зона умеренного и умеренно-теплого влажного климата Южного полушария. Новая
Зеландия в палеоцене уже представляла собой массив суши, изолированный от Австралии
и
Антарктики, и находилась в пределах 50°-60° ю.ш. Ее географическое положение
не только в начале палеоцена, но и на протяжении всего палеогена обеспечивало
в ней наличие мягкого морского влажного климата. Раннедатское похолодание проявилось
лишь на юге острова [Kennedy, Raine, 2001]. Среднегодовые температуры Новой Зеландии
в конце Маастрихта, рассчитанные методом CLAMP, были в пределах 12°-15 °С, а
в дании снизились до 6°-11 °С. Одновременно произошло и значительное уменьшение
размеров листовых пластинок, вызванное неблагоприятными экологическими и климатическими
условиями, а возможно и сезонным распределением осадков, которых было не менее
1000 мм/год. В районе Оамару в дании происходило угленакопление. Из межугольных
глинистых пачек автором данного раздела во время рейса НИС "Витязь" в
Тихий океан в 1970-1971 гг. была собрана небольшая коллекция, основу которой
составляют остатки хвойных (Podocarpaceae, Araucariaceae). Климат палеоцена о-ва
Кинг Джордж и Антарктического п-ва [Torres, 1992] был переходным от умеренно-теплого
к субтропическому с большим количеством среднегодовых осадков (не менее 1600-1800
мм/год), относительно высокими среднезимними температурами (до +4° - +6 °С) при
непродолжительных заморозках. Наряду с древовидными папоротниками и протейными,
обычными компонентами развитых здесь флор являлись южные буки (Nothofagus),
ногоплодниковые и араукариевые. Палеоценовые флоры крайнего юга Американского
континента и Огненной
Земли, содержащие до 40-45% листьев с цельнокрайними пластинками, характеризуют
климатические условия как переходные от умеренно-теплых к субтропическим [Dusen,
1907; Romero, 1986].
Одной из характерных черт палеоценовой флоры Австралии, когда последняя еще входила
в состав суперконтинента, является сочетание растений, обитающих ныне в разных
климатических обстановках. Наряду с типичными раннепалеогеновыми компонентами
Нотальной области - Araucaria, Podocarpus, Dacrydium, Microcachris, Nothofagus,
Proteaceae, Myrtaceae и др., важную, возможно даже лесообразующую роль играли
саговники и древовидные папоротники. Поэтому даже в Южной Австралии флора палеоцена
близка к гумидной субтропической. Крупнолистность этой флоры, вероятно, связана
с особенностями светового режима, так как Австралия в то время находилась в пределах
50°-60° ю.ш. Климатические параметры этих флор были следующие: среднегодовые
температуры +15° - +20 °С, среднелетние до +25 °С, средне-зимние +6° — +8 °С,
при среднегодовых осадках свыше 1000 мм.
Климат высоких широт как Южного, так и Северного полушарий, характеризовался
чрезвычайно низким широтным температурным градиентом, небольшими амплитудами
сезонных, месячных и суточных температур [Greenwood et al., 2001].
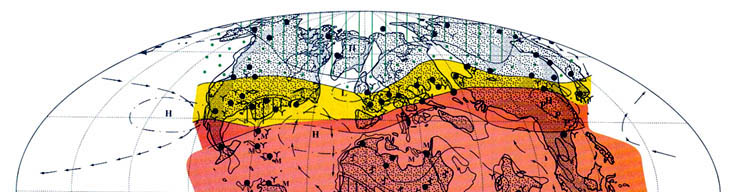
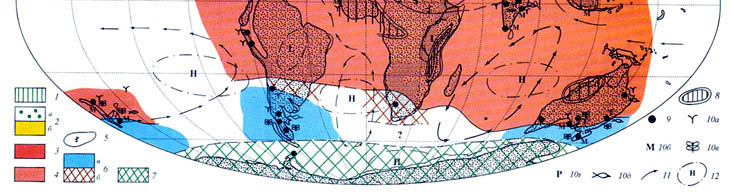
Эоцен был последней эпохой фанерозоя, когда на Земле еще продолжала сохраняться теплая биосфера. Глобальное проявление так называемого "углеродного эпизода" на рубеже палеоцена и эоцена, сопровождавшееся не только заметным снижением содержания изотопа углерода δ13C, но и существенным потеплением климата, которое оценивается в 3 °С, вызвало заметное расширение тропической и субтропической зон Земного шара с их экспансией в более высокие широты. Это в свою очередь сопровождалось сокращением умеренной и тепло-умеренной зон в приполярных областях и дальнейшим снижением широтного температурного градиента в системе "полюс-экватор". Одной из специфических особенностей эоценового (в особенности раннеэоценового) глобального климата, было широкое распространение так называемого "паратропического" влажного климата, иногда приобретавшего черты муссонного. Специфическими особенностями которого были низкая амплитуда сезонных и суточных климатических колебаний, относительно высокие среднегодовые температуры зимних месяцев при допустимых кратковременных слабых заморозках в приполярных регионах обоих полушарий. Основными биогенными показателями климатических обстановок в эоцене по-прежнему остаются такие растения-индикаторы климата как пальмы, имбирные, саговники, древовидные папоротники и обитатели мангровых, из представителей фауны индикаторами климатических условий являются ископаемые остатки крокодилов, колониальных кораллов, некоторых моллюсков (рис. 3, см. вклейку I).
Ранний эоцен
Зона умеренного и умеренно-теплого влажного климата
Северного полушария. В раннем эоцене по сравнению с палеоценовой эпохой эта зона значительно сократилась
по площади и охватывала главным образом приполярную область и только в Сибири
и Притихоокеанских регионах Азии и Северной Америки она опускалась к средним
широтам. Южная граница зоны проходила через Северную Атлантику, Печорский бассейн
и Северный Урал, а в Западной Сибири смещалась несколько к югу, проходя почти
параллельно современной береговой линии Северного Ледовитого океана южнее Полярного
круга. В Восточной Сибири эта граница отклонялась еще далее к югу и выходила
к Тихому океану южнее Магадана и Корякского нагорья, пересекая Камчатский перешеек.
На Американском континенте эта граница трассировалась через Южную Аляску. Во
внутренней части континента она отклонялась к югу почти до границы США и Канады,
выходя на побережье Атлантического океана между Гренландией и Ньюфаундлендом.
Наиболее представительные в систематическом отношении флоры этой зоны приурочены
к угленосным впадинам побережья Северного Ледовитого океана в низовьях Лены,
междуречье Яны и Индигирки, в Корякии и на Северо-Западной Камчатке. На Американском
континенте рассматриваемой зоне принадлежат флоры Аляски, расположенные севернее
хр. Брукса, а также флоры о-вов Акселя Хейберга и Элсмира в Северной Канаде,
которые находятся на 79° с.ш. Практически все эти флоры крупнолистны, что связано
с их высокоширотным положением и полярным световым режимом. Преимущественно положительные
среднезимние температуры и повышенная влажность воздуха объясняет совместное
нахождение в ориктоценозах многочисленных остатков хвощей, полиподиевых папоротников,
хвойных (Metasequoia, Тахоdium, Glyptostrobus, Pinus, Cupressaceae),
листопадных покрытосеменных, включая представителей сережкоцветных (ореховых,
березовых), платановых, гамамелиевых, ильмовых, листьев и фруктификаций Trochodendroides,
a также листьев Macclintockia (в прибрежных фациях), в сочетании с термофильными
вечнозелеными и листопадными (лавровыми, магнолиевыми, аралиевыми). Характерно,
что наряду со смешанной флорой в этой зоне известны находки остатков черепах
и крокодилов, а на севере Тихоокеанского сектора, в ряде местонахождений Аляски,
Камчатки и Анадырского района, кроме того остатки веерных пальм (Sabal) и даже
саговников [Ахметьев, 1993; Буданцев, 1983; Greenwood, Wing, 1995].
Переход от зоны умеренно-теплого климата к паратропическому в Западной Евразии
и Северной Америке, а также к субтропическому в Восточной Азии происходил по-разному.
В Евразии и Северной Америке эта смена осуществлялась в пределах не широкой экотонной
зоны, которая содержала смешанную флору. Напротив, в Восточной Азии, особенно
у Тихоокеанского побережья, подобные флоры с участием листопадных и вечнозеленых
элементов занимают гораздо более широкую экозону, охватывающую Приохотье, частично
Приамурье, Сахалин, Камчатку, Северо-Восточный Китай, север Корейского п-ова,
о. Хоккайдо. На формировании этой экозоны, несомненно, сказалось влияние существовавшего
на протяжении всего кайнозоя холодного течения, омывавшего с востока Тихоокеанское
побережье России и Кореи. Листопадные элементы в условиях расчлененного рельефа
океанической окраины проникали далеко на юг, а в зонах морских побережий вечнозеленые
растения, включая пальмы, - к северу. Находки последних отмечены к югу от устья
р. Анадырь, а также в двух пунктах у северо-западного побережья Камчатки. Остатки
Sabal и саговников обнаружены на о. Хоккайдо [Ахметьев, 1993].
Для характеристики описываемой зоны интересны климатические данные, рассчитанные
методом CLAMP для раннеэоценовых или ранне-среднеоэоценовых флор Канадского Арктического
архипелага [Greenwood, Wing, 1995]. Для флоры о-ва Акселя Хейберга (79°55' с.ш.),
предположительно произраставшей на высоте 200 м над уровнем моря, параметры рассчитаны
для 12 таксонов и представлены так: среднегодовая температура +9,3 °С, амплитуда
среднегодовых колебаний температур между самым холодным и самым теплым месяцами
13,6 °С, средняя температура самого холодного месяца - 0,8 °С. Близкие
значения получены по 15 таксонам и для флоры о-ва Элсмир. Предполагается, что
эта флора произрастала на высотах до 500 м над уровнем моря, при следующих климатических
показателях: среднегодовая температура +8,2 °С при амплитуде среднегодовых колебаний
температур 14 °С и средней температуре самого холодного месяца -2,0 °С. Существенно
иные климатические параметры получены на территории Южной Канады и приграничных
штатов США (45°-50° с.ш.) для раннеэоценовых и ранне-среднеэоценовых флор экотонной
зоны, разделявшей зональные типы лесной, преимущественно листопадной и "паратропической" растительности.
Флоры местонахождений Мак Оби, Бирс Поу, Кемельс Батт и др., рассчитанные по
весьма представительным выборкам (12-32 таксонов) характеризуются среднегодовыми
температурами 10°-11 °С. Близкие значения (9°-11 °С) получены ареалогическим
анализом. Колебания сезонных температур были от 9° до 13 °С, а температуры самого
холодного месяца варьировали от +0,6° до +3,5 °С. Характерно, что смешение листопадных
и вечнозеленых элементов в этих флорах в большей степени связано с абсолютными
отметками обитания растений, которые для большинства флор лежали в интервале
500-1000 м над уровнем моря. К описываемой группе флор необходимо отнести также
флоры Пьюджет и Репаблик раннего-среднего эоцена (50-48 млн лет) штата Вашингтон
[Wolfe et al., 1998]. Среднегодовые температуры для флоры Пьюджет оцениваются
+18° - +23 °С, а для расположенной на 150-180 км севернее флоры они были существенно
ниже - только +10 °С. Это, по-видимому, связано с различиями абсолютных отметок
обитания названных флор.
Все флоры умеренной высокоширотной зоны тяготели к влажным районам со среднегодовым
количеством осадков от 1000 до 2000, а местами до 4000 мм/год с их равномерным
выпадением. Лишь в экотонной зоне распределение осадков носило более дискретный
характер и здесь выделялись районы, в которых дожди выпадали в основном в летний
период. В этих случаях климат приобретал черты муссонного.
Зона влажного субтропического и паратропического климата Северного полушария. Эта
зона отчетливо очерчивается в Западной Евразии и Северной Америке, особенно в
пределах последней, где фиксируется большое количество местонахождений раннеэоценовой
теплолюбивой флоры. В Западной Евразии эта климатическая зона охватывала средние
широты Западной и Центральной Европы
(Лондонской и Гемпширский бассейны, территорию Франции, Германии, Польши) протягивалась
далее на восток через Украину и Западный Казахстан до бассейна Иртыш. К востоку
от оз. Зайсан флора этой зоны все более приобретает черты сезонно-влажной субтропической,
что нашло отражение в составе богатейшей райчихинской флоры Приамурья и некоторых
флор Северного Китая и Японии [Ахметьев, 1993; Mai, 1995; Guo Shuangxing, 1979].
На Американском континенте соответствующие флоры, определяемые Дж. Вулфом [Wolfe,
1995; Wolfe, Dilcher, 2001] как паратропические, известны из средних широт западной
части США. На юго-востоке страны флоры носят уже преимущественно тропический
характер, с практически повсеместно сезонным выпадением осадков, что придает
климату сходство с муссонным. На юге Европы рассматриваемая климатическая зона
постепенно сменялась тропической. Поэтому флора и растительность раннего эоцена
Южной Европы по климатическим параметрам (но не по составу!) мало отличались
от североафриканских, находившихся на противоположном побережье Тетиса. В Южном
Казахстане и Туркмении, судя по распространению нижнеоэоценовых соленосных и
пестроцветных глин, зона влажных субтропиков сменялась зоной сухих субтропиков
и граница между ними прослеживалась к востоку через центральные районы Китая
до Тихого океана.
Характерно, что большинство флор описываемой климатической зоны содержит остатки
мангровой пальмы Nypa, а также других перистых и веерных пальм, представителей
герпетофауны, особенно многочисленных на территории Запада США.
В Западной Европе флоры пограничных горизонтов палеоцена и нижнего эоцена, а
также базальных слоев нижнего эоцена известны из Лондонского, Парижского и Бельгийского
бассейнов, где они объединены Д. Маем [Mai, 1995] в комплексе Ридинг-Вервинс
по характерным местонахождениям Южной Англии и спарнакских слоев Парижского бассейна.
В составе флор известны представители многих тропических и субтропических семейств:
Anacardiaceae, Apocinaceae, Araliaceae, Ericaceae, Euphorbiaceae, Lauraceae,
Leguminosae, Mastixiaceae, Moraceae, Sterculiaceae, Symplocaceae, Zingiberaceae
и др. Вместе с тем присутствуют, хотя и в подчиненном количестве, остатки листопадных
платановых, троходендроидеса, абелии, бархатного дерева, а также сосны.
Еще более определенно "паратропический" тип флоры характерен для стратиграфически
более высокого комплекса середины ипра (зоны NP11 и NP 11/12 по наннопланктону).
Д. Маем [Mai, 1995] он назван комплексом Беллей-Шеппи по характерным флорам суассонских
и кюизских слоев Франции и Бельгии, а также подразделений "Д" и "Е" лондонских
глин у Кента. Паратропический характер флор этого комплекса подчеркивается присутствием
в ориктоценозах многих представителей мангровых (Nypa, Ceriops, Palaeobruguera (Rhizophoraceae)),
а также пальм, лавровых, диллениевых, флакортиевых и других представителей типичных
тропических семейств. Тем не менее, в орикоценозах по-прежнему
встречаются листопадные растения, что связано, по-видимому, с заметным расчленением
рельефа по периферии осадочных бассейнов. Для раннеэоценовых флор из лондонских
глин определены следующие климатические параметры: среднегодовые температуры
+16° - +19 °С, температуры самого теплого и самого холодного месяцев соответственно
+23 °С и 8° - +16 °С, при среднегодовом количестве осадков более 2000 мм [Van
Beuskom, 1971].
В Центральной Европе зона развития паратропического климата хорошо восстанавливается
по палинологическим данным. Восточнее эта зона прослежена через центральные районы
Украины, где А. Н. Криштофовичем [1928; 1955] и Ф. А. Станиславским [1956] описаны
находки из Каневских отложений мангровой пальмы Nypa и несколько видов Macclintockia.
Эта же климатическая зона, по данным Н.М. Макулбекова [1977] и М. А. Ахметьева
[1999], установлена в Примугоджарье (флоры Караколя и Сарколя), занимает значительные
части Западного Казахстана (Тыкбутак) и Северного Казахстана (Экибастуз), уходя
через Зайсанскую впадину (Кирши и Чакельмес) в Южную Монголию. В последнем регионе
состав растений характеризует более аридную обстановку, чем в прочих названных
регионах. Флоры всех перечисленных местонахождений, по сравнению с позднепалеоценовыми,
более мелколистны, имеют ксероморфный облик, что связано вероятно с более заметно
выраженной сезонностью климата. Это относится к листьям еще распространенной
в раннем эоцене Ushia, а также богато представленным в этих флорах представителям
Myricaceae и Ericaceae. В раннем эоцене в окрестностях оз. Зайсан появляются
мелколистные жестколистные ликвидамбры, а также такие характерные представители
субтропической и тропической флоры как Idesia, Ungnadia, Eugenia.
Продолжение зоны влажного субтропического и паратропического климата далее на
восток фиксируется находками райчихинской флоры восточнее г. Благовещенск, содержащей
характерные тропические папоротники (Lygodium, Anemia), имбирные, крушиновые,
анокардиевые, лавровые, магнолиевые и других представителей цветковых. На территории
Китая эта климатическая зона охватывает Южную Манчжурию с богатейшей флорой Фушуня,
в составе которой встречены пальмы (Sabal) и саговники (Cycas). По сравнению
с палеоценовой эпохой, зона влажных субтропиков существенно расширилась, оттеснив
к северу южную границу распространения смешанной флоры, содержащей наряду с вечнозелеными
разнообразные листопадные древесные растения [Guo, Shuangxing, 1979].
Описываемая климатическая зона отчетливо выражена на территории западных и внутренних
штатов США, примыкающих к границе с Канадой. С начала эоцена здесь существовали
крупные озерные системы, изолированные от Тихоокеанского побережья серией меридиональных
хребтов. Пространственно эти системы унаследовали ложе пролива, меридионально
пересекавшего Североамериканский континент до конца мелового периода [Wolfe,
Dilcher, 2001].
К наиболее северным раннеоэоценовым флорам района залива Якутат относятся флоры
Калтит и Куштака на южном побережье Аляски, а на юге Британской Колумбии флоры
формации Хэмтиндон [Wolfe, Dilcher, 2001]. Флора Калтит, ныне расположенная за
северным полярным кругом (70° с. ш.), по расчетам CLAMP, выполненным автором
метода Дж. Вулфом, рассматривается как близкая к паратропической. Она формировалась
при среднегодовых температурах +19,6 °С, средне-зимних +12,6 °С и среднелетних
+26,2 °С. Характерна весьма незначительная амплитуда сезонных колебаний - всего
13,6 °С [Wolfe, 1994]. В составе флоры Калтит и других флор залива Якутат был
установлен широкий спектр растений тропиков и субтропиков, включая и восточно-азиатского
родства. Родовой спектр этой флоры включает: Alangium, Barringtonia, Macaranga,
пять видов лавровых, пальмы, саговники, деревянистые лианы, мангровое Kandelia,
Meliosma, Parachorea, Platycarya, Tetracentron и др. Большинство растений
принадлежало вечнозеленым широколиственным цветковым с количеством цельнокрайних
форм до 70%.
Рецентные их аналоги близ морских побережий сосредоточены в тропиках, где температуры
никогда не снижаются до 0 °С. Высказывалось предположение
[McKaena, 1983], позже поддержанное Н. Фредериксеном [Frederiksen, 1988],
что
Якутатский блок представляет собой террейн, который в раннем палеогене занимал
значительно более южнее положение, чем ныне (возможно 48°-50° с.ш.). В противном
случае было бы трудно объяснить тот факт, что флора Терминал Док, расположенная
по широте на 10° южнее Якутатского блока характеризовалась среднегодовыми температурами
всего на 2°-3 °С выше соответствующих температур раннеэоценовых флор Юго-Восточной
Аляски.
Большое количество таксономически небогатых раннеэоценовых флор известно из южных
районов Британской Колумбии и пограничных с Канадой штатов США, а также южнее,
вплоть до 45° с.ш., главным образом, в пределах Скалистых гор. Обработанные методом
CLAMP, они относятся по составу к типичным флорам паратропического типа, но тем
не менее заметно отличаются по своим климатическим параметрам (особенно средне-зимними
температурами и амплитудами колебаний сезонных температур). Это прежде всего
связано с накоплением флороносных толщ в бассейнах, расположенных на разной высоте
над уровнем моря (от первых десятков метров до 1000 м и более) [Greenwood, Wing,
1995; Wolfe, Dilcher, 2001].
Зона сухих субтропиков Северного полушария. Отчетливые
признаки аридной зоны в Северном полушарии устанавливаются по распространению
эвапоритов в южных районах
Казахстана и Киргизии, Южной Монголии и восточнее в Центральных и Южных районах
Китая. Аридная обстановка этой зоны подчеркивается большим участием в палинокомплексах
пыльцы нитрарий, эфедры и маревых, а также присутствием эфедры и палибинии. Последняя
- характерное ксероморфное растение, условно сближаемого с семейством анакардиевых.
В Южной Гоби флора раннего эоцена представлена разнообразными мелколистными ильмовыми.
На территории Китая остатки палибинии известны из красноцветов и соленосных толщ,
распространенных к югу от Шандуньского п-ова, но особенно широко представленных
в южных провинциях Цзянси, Гуандун и др. Здесь имеются многочисленные впадины,
заполненные гипсоносными глинами, разнообразными по составу солями. В провинции
Хунань находится одно из крупнейших месторождений каменных солей этого возраста
- Хеньянское. На юге аридная зона быстро сменяется зоной влажного тропического
климата.
Зона влажного тропического и экваториального климата. Эта зона прослеживается
вдоль Южной Европы, находит непосредственное продолжение на Африканском континенте,
где она разделена семиаридной тропической зоной. По характерным флорам тропического
родства рассматриваемая зона устанавливается в Италии (флористический комплекс
Монтеболка) из пограничных слоев нижнего и среднего эоцена, а также в Сардинии,
на территории бывшей Югославии (флора Монте Промина) и Румынии. Кроме разнообразных
представителей мангровых и множества таксонов пальм, здесь присутствуют характерные
виды рода Ficus, а также разнообразные Lauraceae. Большинство таксонов являются
типичными представителями палеогеновой флоры Тетической области.
На Североамериканском континенте флоры этого типа весьма обстоятельно изучены
в бассейне р. Миссисипи в прибрежной зоне древнего Мексиканского залива, заходившего
в раннем эоцене вглубь континента вплоть до современных штатов Кентукки и Теннеси.
Интерпретируя палинологические и макрофлористические данные [Frederiksen, 1980;
Dilcher, 1973; Wolfe, Dilcher, 2001; Wolfe, 1985 и др.], большинство специалистов
считали, что климат северной оконечности древнего Мексиканского залива в эоцене
был сезонно сухим тропическим или гумидным субтропическим, близким муссонному.
Тропические леса, образованные анноновыми, лавровыми, луносемянниковыми и пальмами,
в раннем эоцене простирались до 50° с.ш. В начале раннего эоцена климат к югу
от 30° с.ш., вероятно напоминал экваториальный, что находит подтверждение в широком
распространении в прибрежной зоне Мексиканского залива мангровых растений, представителей
герпетофауны, а в морских карбонатных толщах - кораллов. Количество осадков здесь
возрастало до 5000 мм/год, но выпадение их оставалось сезонным. Вдоль побережья
древнего залива происходило формирование латеритных кор выветривания, а в Арканзасе
сформировались крупные месторождения бокситов. К концу раннего эоцена, по данным
Д. Дильчера [Wolfe, Dilcher, 2001], среднегодовые температуры в Центральном Техасе
оценивались в +22° - +23 °С. Заметно сократилось годовое количество осадков (до
1300 мм), но еще более отчетливо стала выражаться сезонность климата, так как
на три засушливых месяца приходилось всего 150 мм осадков.
На климат раннего эоцена Африки влияли два фактора: смещение континента к северу
относительно экватора на 5°-6° или примерно на 500-600 км, а также активно развивающаяся
морская трансгрессия, в результате которой море достигло центральных районов
современной Сахары, захватив широкой полосой значительную часть Марокко, Туниса,
Алжира, Ливии и Египта. Полоса тропических
дождевых лесов протягивалась узкой полосой к югу от морского побережья, что фиксируется
многочисленными находками мангровых растений, а также определенным по остаткам
древесин типичных представителей Палеотрописа, распространенных ныне только в
лесах такого типа (Pterocarpus, Entandra и др.). Вторая полоса тропических дождевых
лесов протягивалась в широтном направлении от Сенегала и Камеруна на западе до
верховьев р. Конго на восток. В береговой зоне морского бассейна, который заходил
от Атлантики вглубь континента и захватывал территорию Сенегала и все регионы,
примыкающие к Гвинейскому заливу, включая Нигерию и Камерун, повсеместно в это
время произрастали растения мангровых зарослей, включая пальму Nypa. Разделяющее
эти две полосы пространство, занимающее большую часть территории Северной Африки,
было занято саванным редколесьем, образованном бобовыми, эофорбиевыми, видами
рода Combretum и пр. Еще один пояс развития влажного тропического климата прослеживался
в раннем эоцене вдоль восточного побережья Африки от Большого Африканского Рога
до широты южной оконечности Мадагаскара. Западнее тропического пояса протягивался
семиаридный пояс с саванной растительностью, сливающийся на севере с широтной
полосой саванн, развитой к востоку от Сахары. В этом семиаридном поясе к югу
от экватора на протяжении всего эоцена происходило формирование мощного песчаного
комплекса, получившего на юге Африки название формации Калахари, а севернее в
бассейне Конго на территории Заира - песков Батеке [Maley, 1996].
Климат Индии в раннем эоцене, судя по доминированию представителей дождевых лесов,
был тропическим гумидным. Флора была исключительно богата, так как ныне известно
не менее 200 родов. Столь же богата флора верхних горизонтов трапповой серии,
также имевшая возможно раннеэоценовый возраст. Практически повсеместно древнее
морское побережье маркируется находками мангровых растений - Nypa, Rhizophora,
Sonneratia, Brownlovia. Исключительно разнообразными были пальмы, по остаткам
древесин которых установлено свыше двух десятков видов. Климат Ассама, Бирмы,
Таиланда, а также расположенных к востоку приэкваториальных регионов Индонезии
и Новой Гвинеи, судя по палинологическим данным, был также влажным
тропическим. Тропическая зона охватывала и южные районы Китая, причем местами,
как например, на Хайнани, происходило формирование угленосных толщ и в составе
флоры присутствовали разнообразные лавровые (Cinnamomum, Ocotea), а также пальмы
(Sabalites) и цитрусовые (Citrus niger) [Guo Shuangxing, 1979].
Несмотря на относительно высокоширотное положение Австралийского континента в
раннем эоцене (его южная оконечность находилось вблизи 60° ю.ш.), флористические
данные, полученные для юга материка, включая Тасманию, свидетельствуют о существовании
здесь в это время влажного субтропического или паратропического климата, иногда
близкого к типичному тропическому. Зимняя температура на приморских низменностях
Австралии всегда оставалась выше О °С, а распределение годовых осадков допускало
существование муссонного климата в более внутренних районах континента. Этот
вывод подтверждается находками в ряде местонахождений, в том числе и в Тасмании,
плодов мангровой пальмы Nypa, остатков пандановых, саговников, древовидных
папоротников
в сочетании с широким спектром южных хвойных (Libocedrus, Podocarpus, Dacrycarpus,
Acmopyle, а также зубов крокодилов [Pole, 1992]). Еще больше свидетельств
о былом широком распространении здесь мангровой растительности получено по данным
палинологии.
Большинство палинокомплексов, полученных из разрезов нижнего эоцена всех без
исключения окраинных бассейнов южного побережья континента и его восточного обрамления,
содержат пыльцу тех или иных мангровых растений в сочетании со спорами древовидных
папоротников (Cyathea, Dicksonia) [Truswell, 1990] и остатками кораллов.
Таким образом, температура поверхностных океанических вод, омывающих с юга и
востока
континент, была не ниже +20 °С. Ориктоценозы конкретных местонахождений на крайнем
юге Австралии (Брэнди Крик, Виктория) включают большое количество родов и видов,
принадлежащих тропическим семействам, особенно лавровым, кунониевым и элеокарповым
(Entandra, Cryptocarya, Beilschmeidia и др.) [Keefe, Greenwood, 2001].
Большинство рецентных видов перечисленных выше родов сосредоточено ныне во влажных
тропических
лесах Северо-Восточной Австралии. Предполагается, что основные климатические
параметры зоны обитания флоры Брэнди Крик были такими: среднегодовая температура
выше 15 °С, количество среднегодовых осадков выше 2000 мм, зимние температурах
выше 0 °С. Итоги последнего обобщения накопленных данных об эоценовых флорах
Австралии, развитых в окраинных частях осадочных бассейнов Джипсленд и Отуэй
[Greenwood et al., 2000;
Greenwood et al., 2001], несколько расширили представление о составе раннепалеогеновых
австралийских флор, выявив участие в них не только вечнозеленых тропических растений,
но также и разнообразных представителей южных хвойных - кипарисовых, ногоплодниковых,
араукариевых, включая недавно открытую в окрестностях Сиднея в современной флоре
воллемию. Кроме того, присутствие не только вечнозеленых, но и листопадных цветковых
(Nothofagus, Eucryphia и др.), позволило несколько по-иному интерпретировать
общую климатическую картину на юге континента в раннем эоцене и показать, что
паратропические дождевые леса были распространены только на приморских низменностях,
замещаясь в горах и на высоких равнинах лесами, которые современные ботаники
обычно рассматривают как горные тропические или субтропические. В этой связи
даны и уточнения климатических параметров Южной Австралии: среднегодовые температуры
были в пределах 16°-18 °С, среднезимние - выше 3 °С, а годовое количество осадков
превышало 2000 мм/год. Это не исключало наличия в зимнее время непродолжительных
снегопадов и даже кратковременного существования снежного покрова на высоких
плато и в горах. Важными показателями климата являлись сезонные колебания температур
(от 12° до 22 °С) [Greenwood, Wing, 1995]. Колебания в раннем эоцене по амплитуде
превосходили одновозрастные колебания на тех же широтах Северной Америки и Европы,
но значительно уступали амплитудам современной эпохи. В цитированной работе Д.
Гринвуда и С. Винга приведены климатические показатели для раннеэоценовой флоры
Дине западнее г. Мельбурна, произраставшей на предполагаемых абсолютных отметках
формирования около 500 м. Рассчитанные методом CLAMP по 18 таксонам эти показатели
таковы: среднегодовая температура 12,2 °С, амплитуда сезонных колебаний 13,3
°С и среднезимняя температура 4,6 °С.
Климат раннего эоцена Новой Зеландии, которая как и Австралия в палеогене находилась
в более высоких широтах по сравнению с ее нынешним положением, сохранялся неизменно
гумидным, влажным, в прибрежной зоне близким по основным показателям к тропическому,
что подтверждается находками остатков мангровых растений. Однако уже на высотах
200 м над уровнем моря среднегодовые
температуры снижались и скорее соответствовали характерным для южных влажных
субтропиков. Так, рассчитанная методом CLAMP по 13 таксонам из местонахождения
Livingstone на южном острове (44°51' ю.ш.) она составила +13 °С, при среднезимних
температурах до 13,6 °С и амплитуде сезонных колебаний 15,5 °С.
Информация о флорах раннего эоцена Южной Америки скудны, но тем не менее позволяют
воссоздать в общих чертах существовавшую здесь картину распределения основных
климатических поясов. Небогатые флороносные комплексы, представленные исключительно
неотропическими таксонами, описаны Э. Берри [Berry, 1927; 1929; 1937] из прибрежных
районов Западного Перу к югу от экватора (местонахождения Сюр де Талара и Белен),
а стратиграфическое положение флороносных слоев уточнено Р. Хоффстеттером [Hoffstetter,
1936a]. Присутствие в ориктоценозах плодов пальм, остатков сапиндовых, а также
Anacardium, Humuria, Cupanoides, Attalea olsoni и других растений тропического
родства позволило заключить, что климат раннего эоцена приокеанической равнины
севера Южной Америки в раннем эоцене был тропическим гумидным. Это заключение
находит подтверждение и в данных палинологии. Что же касается конкретных климатических
параметров (температуры и влажности) экваториальной зоны Южной Америки в эоцене
по изменению растительности в ответ на глобальное потепление на рубеже палеоцена
и эоцена и в раннем эоцене, то строгие данные на этот счет отсутствуют. Палинологические
данные, полученные по разрезам Колумбии, свидетельствуют лишь о заметном возрастании
биоразнообразия в растительном мире приэкваториальной зоны в раннем эоцене, достигшего
своего максимума на рубеже раннего и среднего эоцена. Если количество таксонов
в палиномацератах из верхнепалеоценовых толщ лежит в пределах 70-80, то в начале
раннего эоцена это количество возрастает до 100, а к его концу до 120, что несомненно
связано с благоприятной экологической и климатической обстановками во влажных
тропиках [Jaramillo, 2001].
Интересные находки ископаемых растений происходят из более южных районов континента,
расположенных в Чили на 37° ю.ш. (местонахождения Лота и Коронель). Изученные
отсюда более 100 видов покрытосеменных также оказались типичными представителями
неотропической флоры [Engelhardt, 1895; Berry, 1922]. По мнению Э. Берри, большинство
рецентных потомков этих раннеэоценовых
растений произрастает в тропиках Южной Америки к востоку от Анд, главным образом
в перуанской части Амазонского бассейна, распространяясь в Северную Боливию,
но не выходя южнее за пределы 10°-12° ю.ш. Это свидетельствует о существенном
расширении в раннем эоцене тропической зоны в Южной Америки. Э. Берри охарактеризовал
климат данной зоны как безморозный, очень теплый, с большим количеством солнечных
дней. Высокий процент видов цельнокрайних листьев (более 70%), в ориктоценозах
подтверждает точку зрения о распространении зоны раннеэоценового тропического
климата на юг по крайней мере до 35°-40° ю.ш.
Зона влажных и сухих субтропиков Южного полушария. Вся южная оконечность Африканского
континента в раннем эоцене входила в зону сухих субтропиков. Флора и растительность,
напоминающие современную капскую флору окрестностей Кейптауна, получили более
широкое распространение и, вероятно, достигали широты р. Оранжевая. Об этом свидетельствуют
находки ископаемых древесин на юге Намибии. В состав развитых здесь разреженных
лесов входили представители семейств Balanitaceae, Burseraceae, Euphorbiaceae,
Leguminosae, Myrtaceae [Bamford, 2000]. Облик растительности (бушленды) весьма
напоминали средиземноморские маквисы. Климат также напоминал средиземноморский,
характеризуясь жарким сухим летом и влажной зимой.
Южная граница тропической зоны в Южной Америке в раннем эоцене проходила около
40° ю.ш. В ориктоценозах двух богатейших раннеэоценовых флор Северо-Западной
Патагонии - Рио Пичелейфу и Рио Ченквинею (39° ю.ш.), отличающихся высоким биоразнообразием
(более 140 видов), наряду с неотропическими таксонами присутствуют и антарктические
элементы [в понимании Romero, 1986]. Это сближает их с позднепалеоценовой флорой
Лагуны дель Хунко этого же региона. Структурируя флору Рио Пичелейфу, Э. Берри
[1938] отмечал, что при высоком содержании таксонов с цельнокрайней пластинкой
(69%) в ее составе могут быть выделены тропические, субтропические, умеренные,
а также семиаридные элементы (Cupania, Schinopsis, Schinus) и растения,
обитающие ныне только в умеренных влажных лесах Южного Чили. Специфической чертой
этой
флоры является высокое содержание в ее составе лиан (14 видов). Характерно, что
во флоре Рио Ченквинею появляется нотофагус. Близкое сходство с флорой Лагуны
дель Хунко (42°05' ю.ш., 70° з.д.) позволяет распространять климатические
параметры этой флоры [Wilf, Johnson et al., 2001] и на две другие отмеченные
выше флоры. Они существовали при среднегодовых температурах до 16 °С и осадках
1000-2000 мм, характерных для влажных субтропиков. Учитывая небольшие амплитуды
колебаний сезонных и суточных температур, этот климат лучше называть паратропическим.
Высокая влагообеспеченность Северо-Западной Патагонии исключает существование
в раннем эоцене Андской горной цепи, так как в противном случае несомненно бы
сыграл эффект "дождевой тени". Влажный субтропический климат получил
распространение в расположенном южнее бассейне Голфо Сан Хорхе, где во флоре
Канадой Хондо [Romero, 1978] также отмечается смешение неотропических и антактрических
элементов.
Есть основание предполагать, что зона субтропического климата распространялась
в раннем эоцене до Антарктического п-ова, расположенного южнее Северо-Восточной
Патагонии на 2000 км. В нижнеэоценовой флоре о-ва Кинг Джорджа наблюдается смешение
антарктических (Laurelia, Nothofagus) и неотропических (Schinopsis,
Nectandra)
элементов, при этом листья семи из десяти присутствующих таксонов покрытосеменных
характеризуется цельнокрайней пластинкой [Orlando, 1963].
Таким образом, большая часть Южной Америки (к югу от 40° ю.ш.) в раннем эоцене
входила в зону тропического климата. Субтропический и паратропический климаты,
имевшие местами более отчетливые черты сезонного (муссонного), распространялись
в пределах остальной части южноамериканского континента вплоть до Антарктического
п-ова. Этот вывод основан не только на результатах из морфологического анализа
ископаемых листьев, но и на присутствии в тафоценозах остатков пальм, представителей
мангровых, остатков древовидных папоротников и крокодилов. На юге Австралийского
континента и в Новой Зеландии, субтропический климат, как уже отмечено выше,
был свойственен только высоким равнинам, плато и горным районам.
Зона умеренного и умеренно-теплого влажного климата Южного полушария. Для этой
зоны имеются крайне скудные палеоботанические данные, полученные по некоторым
местонахождениям Антарктического п-ова и примыкающих к нему островных архипелагов.
Здесь во флорах раннего эоцена при полном отсутствии неотропических компонентов
основное распространение получили Nothofagus sec. Fusca и Menziesii,
обитающие
ныне в горных лесах Южных Анд. Вероятно, к
этой зоне лесной растительности можно отнести и Антарктический континент, хотя
каких-либо палеоботанических данных по его раннему эоцену до сих пор не было
получено.
Заканчивая раздел, касающийся описания климата и климатической зональности в
раннем эоцене Земли, следует подчеркнуть основные его особенности. Во-первых,
происходило заметное расширение тропической и экваториальных зон, климат которых
был преимущественно влажным с большим количеством (до 2000 мм и более) годовых
осадков. Во-вторых, он характеризовался крайне низкой амплитудой сезонных температур
и незначительным широтным температурным градиентом, не повторявшимися в другие
эпохи кайнозоя. В зонах, примыкающих к Тихоокеанскому побережью, этот градиент
был 0,4 °С на 1° широты, а во внутренних частях континентов 0,8 °С, что в несколько
раз ниже современного температурного градиента как на окраинах, так и во внутренних
районах континентов. В-третьих, отсутствовали какие-либо признаки понижения температур
в тропической и экваториальной зонах, что нередко рассматривается как аргумент
в пользу вывода о равномерном распределении тепла на поверхности Земли в эоцене
по сравнению с современной эпохой, а также более благоприятных условий тепло-
и влагопереноса в раннеэоценовую эпоху из низких в высокие широты. В-четвертых,
формирование климата, близкого к тропическому и субтропическому муссонному в
Европе, Северной Америке, Австралии, Новой Зеландии и отчасти в Южной Америке.
Вместе с тем, можно определенно говорить о различиях в оценке климата раннего
эоцена, полученной с помощью компьютерных расчетов на аналоговых машинах и на
основании палеоботанических данных. Последними не подтверждается резкая континентализация
климата во внутренних районах материков и низкие зимние температуры. Среднегодовые
показатели климата для различных климатических зон Земли свидетельствуют, что
климат этой эпохи был в среднем теплее современного не менее чем на 6°-8 °С.
Средний эоцен
В среднем эоцене в основном продолжала сохраняться та же
климатическая зональность, что была прослежена на материалах палеоценовых и
раннеэоценовых флор. По сравнению
с раннеэоценовой эпохой более отчетливо выражена дифференциация климатических
зон, связанная с постепенным возрастанием температурного градиента в системе "полюс-экватор", произошло некоторое расширение аридных поясов обоих полушарий
и заметное сокращение широкой в раннем эоцене зоны влажного тропического и
экваториального климата.
Зона умеренного и умеренно-теплого влажного климата Северного полушария. Эта
зона, существовавшая с начала палеогена, занимала приполярную область Северного
полушария, хотя ее южная граница в среднем эоцене и сместилась несколько к
югу по сравнению с ее положением в ранним эоцене. Все основные местонахождения
среднеэоценовых
флор расположены в Восточной Азии и отчасти в Северной Америке. В первом из
этих регионов они известны в Муруктинской котловине Центральной Сибири, в Пенжинской
губе, хребте Рарыткин, на Западной Камчатке, Восточном Сихотэ-Алине, Южном
Приморье
и на Сахалине. Основными доминантами наиболее северных из этих флор являются
гинкго, сосновые (Pinus, Pseudolarix), таксодиевые (метасеквойя, глиптостробус,
тайвания), кипарисовые (туя), ореховые, березовые (ольха, палео-граб, береза),
крайгия, фагопсис, платаны, троходендроидесы, иногда сассафрас, а в южном направлении
постепенно происходит насыщение этих флор, особенно лютетских, субтропическими
растениями (в экотонной зоне). Климатические показатели наиболее северных флор:
среднегодовые температуры 10°-12 °С, среднезимние около или немного ниже 0
°С при среднегодовых осадках 1000-1500 мм. На юг, в направлении экотонной зоны
заметно
возрастало разнообразие термофильных растений семейств Anacardiaceae, Vitaceae,
Phamnaceae, Lauraceae, Fagaceae (цельнокрайние вечнозеленые и листопадные дубы,
каштаны), Myricaceae, Juglandaceae, Ericaceae и др. Расширяется спектр папоротников
Osmunda, Onoclea, Dennstaedtia и др. В горных районах Северо-Востока Азии и
Дальнего Востока (хребет Рарыткин, Сихотэ-Алинь) в составе среднеэоценовых
флор присутствуют
холоднолюбивые мелколистные ольхи, сосны, ели, лиственницы, рододендроны, мелколистные
брусничные, тополя. Для наиболее южных из флор этой группы (Восточный Сихотэ-Алинь)
методом CLAMP автором рассчитаны следующие климатические параметры при содержании
цельнокрайних форм в ориктоценозах около 35-45% среднегодовые температуры 12°-14
°С, среднелетние температуры 19°-21 °С, средне-зимние температуры +2 °С, при
среднегодовом количестве осадков 1200-1600 мм.
Зона влажного и переменно-влажного (сезонного) субтропического климата
Северного полушария. В среднем эоцене средние широты Западной Евразии в лютете входили
в зону влажного субтропического (паратропического) климата. В бартоне климат
этой же зоны существенно изменился и стал переменно-влажным (сезонным), местами
семиаридным с инверсией основных периодов выпадения осадков. Если в конце раннего
и в начале среднего эоцена климат средних широт обоих полушарий напоминал муссонный
с большим количеством осадков при относительно равномерном их распределении
в течение года или пиком в летние месяцы, то климат бартона, особенно в средних
широтах Евразии, отличался жарким засушливым летом и относительно прохладной
зимой, когда происходило основное выпадение осадков. Этот климат напоминал
современный
средиземноморский. Характерно, что признаки ксерофитизации в растительном мире,
особенно у южной границы описываемой климатической зоны, начали проявляться
в морфолого-анатомически особенностях строения листовых пластинок еще в лютете.
В бартоне эти особенности еще больше усилились. Переход от зоны переменно-влажного
климата к зоне семиаридного, особенно на востоке Евразии, заметить чрезвычайно
трудно. На юге Западной Европы, где зона сухих субтропиков в лютете отсутствовала,
влажная или переменно-влажная субтропическая зона сменялась влажной тропической,
которая охватывала и север Африканского континента. Благодаря систематическому
богатству как лютетских, так и бартонских флор и высокой точности определения
возраста локальных флор, климатическая картина среднего эоцена восстанавливается
с большой степенью достоверности и детальности. Флороносные слои залегают либо
среди морских осадков с остатками плантонных фораминифер, наннопланктона и
диноцист
(флоры Южной Англии, Венгрии, Украины, Северо-Восточного Казахстана), либо
содержат богатые комплексы различных групп наземной биоты и прежде всего млекопитающих
(флоры Германии, Монголии).
В лютетскую группу на западе и в центре Евразии входят флоры Сельси Южной Англии,
Месселя (ранний лютет) и Гейзельталя (поздний лютет) Германии, Монтеболка (пограничный
интервал ипра и лютета) Италии, Лабатлана (Венгрия), бучакской свиты Украины,
местонахождений Орь Западного Казахстана, Карасора Павлодарского Прииртышья
и Хайчин Улы Южной Гоби Монголии [Mai, 1995; Макулбеков, 1972; 1997]. На востоке
Евразии полоса мезофильных субтропических лютетских флор захватывает самый
юг
Приморья, Северо-Восточный Китай (флора верхних слоев Фушуня) и Центральную
часть Японии. Дальневосточные флоры, оставаясь мезофильными, являются субтропическими
лишь в полосе морского побережья. Во внутренних впадинах они насыщаются умеренными
компонентами гинкго-метасеквойя-троходендроидесово-платаново-ильмовой группы
таксонов.
Бартонские флоры лучше изучены в Восточной Европе и Казахстане (флоры Баки
Южного Урала, раннебартонские флоры Такырсора, Жамантуза, Кульболды и Селеты
Павлодарского
Прииртышья) [Макулбеков, 1972; 1997]. Отличительной особенностью их является
ксероморфность, мелколистность и жестколистность. Основными лесообразующими
компонентами являются лавровые, буковые и вересковые, из хвойных - долиостробус
и секвойя.
Переход от летневлажного субтропического климата лютета к зимневлажному, напоминающему
средиземноморский, в бартоне отразился и на составе спорово-пыльцевых комплексов
[Ахметьев и др., 2004]. Если основными компонентами палинокомплексов в лютете
средних широт были кастанопсисы и аралиевые с подчиненным количеством мелкой
пыльцы вечнозеленых дубов, то во второй половине бартона пыльца кастанопсиса
отходит на второй план, а к концу бартона почти исчезает, замещаясь пыльцой
дубов.
Субтропическо-паратропический характер лютетских флор подчеркивается присутствием
во всех комплексах прибрежной зоны остатков пальм и мангровых. Плоды пальмы
Nypa burtinii и часто сопутствующих ей папоротника Acrostichum известны из
Сассекса
и других местонахождений Южной Англии, а также Гейзельталя, Месселя и Экфельда
Германии, Лабатлана Венгрии. В английских местонахождениях они дополняются
находками других пальм (Chamaerops, Livistona, Phoenix, Sabal, Serenoa,
Trachycarpus).
Основными компонентами лютетских флор Западной Европы выступали Fagaceae (Dryophyllum,
Eotrigonobalanus, Pasania), Lauraceae (Cryptocarya, Daphnogene, Laurophyllum,
Lindera, Litsea, Ocotea), Moraceae (различные виды Ficus), Icacinaceae, разнообразные
травянистые однодольные и др. В Западной Европе мезофильные лютетские флоры
иногда
называют древнейшими "мастиксиевыми" [Mai, 1995]. Флора Лабатлана
и соседних с ним одновозрастных местонахождений Венгрии (Дорог, Татабанья,
Солемар),
расположенные несколько южнее немецких, содержит еще больше растений субтропическо-тропического
родства. Спектр
таких семейств и родов чрезвычайно широк (Myricaceae, Magnoliaceae, Hamamelidaceae,
Moraceae, Myrtaceae, Bambacaceae, Sterculiaceae, Myrsinaceae и др.).
Климатические параметры, рассчитанные для некоторых из наиболее представительных
в систематическом отношении лютетских флор таковы. Среднегодовые температуры
для Месселя выше +20 °С, Гейзельталя -+15°... +19 °С, Лабатлана - выше +22
°С, самого холодного месяца - для Месселя +10 °С, Гейзельталя +3°... +15 °С,
Лабатлана
+15 °С, количество среднегодовых осадков для Гейзельталя 1000-1600 мм, для
Лабатлана -1500-2000 мм.
В связи с существенным уточнением в последние годы возраста эоценовых отложений
Восточно-Европейской платформы многие флоры, которые прежде датировались поздним
эоценом, ныне относятся к среднему эоцену, как к лютету, так и к бартону. Прежде
всего это относится к флорам из "спондиловых" глин окрестностей г.
Киев, залегающих в нижней (карбонатной) части разреза киевской свиты. Лютетский
возраст имеют и бучакские пески, песчаники и угли Звенигородки и Волянщины, которые
и прежде рассматривались в объеме среднего эоцена [Станиславский, 1950]. Кроме
находок мангровых (нипа, акростихум), в их составе отмечены разнообразные восковницы,
широкий спектр лавровых, вечнозеленых буковых, а также фикусы, бобовые, сапотовые,
вересковые. Флора Волянщины существовала при среднегодовых температурах 18°-20
°С, а присутствие в ее составе жестколистных дубов и вересковых свидетельствует
об определенном сходстве среднеэоценовых лесов центральных районов Украины с
жестколистными лесами и маквисами современного Средиземноморья. Это позволяет
заключить, что перестройка климата в среднем эоцене Восточной Европы на среднеземноморский
начала происходить с конца лютета, а в бартоне лишь усилилась.
Лютетские и бартонские флоры Казахстана мало различались на западе и востоке
республики [Макулбеков, 1972; 1997]. В Примугоджарье и Актюбинском Предуралье
к лютету относятся флоры Шандинска и Ори. В их составе преобладают лавровые
Persea, Litsea, Laurus, буковые Castanopsis, вечнозеленые Quercus, реже - Sequoia,
Ericaceae,
Moraceae, Proteaceae. В богатейшей флоре Павлодарского Прииртышья - Карасорской
основной фон составляют крупнолистные виды Castanopsis, Quercus и Lithocarpus,
состав лавровых тот же, что и в западноказахстанских лютетских флорах - Persea,
Laurus, Cinammomum. Представители
других родов, в том числе и присутствующие в карасорской флоре единично, обитают
ныне, в основном, во влажных тропиках и субтропиках Старого Света (Ebenaceae,
Sapotaceae, Sterculiaceae, Cesalpiniaceae, Fabaceae, Proteaceae, Apocynaceae,
Musaceae, пальмы и др.). Н.М. Макулбеков [1972] полагает, что карасорская флора
обитала при среднегодовых температурах 15°-20 °С и среднегодовом количестве
осадков 800-1200 мм при их сезонном выпадении.
Бартонские (скорее раннебартонские) флоры Казахстана отличаются ксероморфностью.
К ним относится Баки Южного Приуралья и несколько флор Павлодарского Прииртышья
(такырсорская, жамантузская и др.). В растительном покрове Казахстана в это
время господствовали жестколистные леса, а также кустарниковые заросли, образованные
вересковыми, низкорослыми вечнозелеными дубами, миртовыми, протейными и апоциновыми.
B.C. Корнилова [1955] полагала, что флора Жамантуза произрастала при высоких
летних температурах (25°-30°С, возможно выше), среднезимних температурах не
ниже
+5°... +12 °С и среднегодовых осадках от 500 до 800 мм с их выпадением в зимний
период (рис. 2).
|
| Рис. 2. Кривые изменения климата в палеогене и неогене, построенные по палеоботаническим данным для Западного и Юго-Восточного Казахстана. |
Благодаря обнаружению в последнее время богатых комплексов
диноцист в морских отложениях верхнелюлинворской и нижнетавдинской подсвит
(соответственно поздний лютет-ранний бартон и поздний бартон), уточнен возраст
палеогеновых флор Павлодарского Прииртышья, расслаивающих флороносную континентальную
толщу эоцена. Это позволило скорректировать и особенности изменения климата
в средних широтах Западной Азии, учитывая однотипный состав флор и последовательность
их смен во всей полосе от Украины до Восточного Казахстана [Ахметьев и др.,
2004]. Смена доминирующих групп пыльцы буковых (Castanea castanaefolia и Castanopsis
pseudocingulum на Quercus gracilis и Q. graciliformis)
в палинокомплексах второй половины эоцена не является сугубо региональной,
а широко прослежена в разнофациальных
разрезах от Украины и Северного Кавказа до Сибирской платформы. При этом выделяется
три комплекса: 1) более древний с обилием пыльцы каштана и кастанопсиса и почти
не содержащий пыльцу дубов группы Quercus gracilis и Quercus graciliformis (вторая
половина лютета, включая пограничный интервал лютета и бартона); 2) средний
смешанный комплекс, содержащий пыльцу обеих групп, но с некоторым преобладанием
пыльцы дубов (бартон s.l.); 3) комплекс с доминированием последней группы при
редукции пыльцы кастанопсиса (терминальный бартон и приабон). Смена этих комплексов,
которая в зависимости от широты могла быть
асинхронной, отражала климатические перестройки эоцена. До появления в разрезах
пыльцы кастанопсиса и их остатков среди мегафоссилий (карасорский комплекс
Н.М. Макулбекова) в большинстве местонахождений флор палеоцена и раннего эоцена
растения отличались крупными листьями с характерными апикальными капельницами,
указывающими на влажный паратропический климат с относительно равномерным выпадением
осадков в течение года. Карасорская флора не является крупнолистной, а все
современные виды кастанопсиса, кроме одного калифорнийского, распространены
исключительно в пределах муссонной области Юго-Восточной Азии, т.е. в условиях
сезонного летневлажного климата. Рецентные потомки средне-позднеэоценовых ксерофитных
дубов, напротив, сосредоточены ныне в Среднеземноморье или Калифорнийско-Мексиканском
регионе с отчетливо выраженным зимневлажным климатом средиземноморского типа.
Это позволяет сделать вывод, что субтропический муссонный климат, распространенный
в средних широтах Западной Евразии в лютете и в начале бартона, уже в первой
половине бартона начал трансформироваться в сезонный климат средиземноморского
типа с жарким летом (отсюда ксероморфизм листьев местонахождений Баки, Жарлыкапа
и др.). Подобная трансформация климата в среднем и позднем эоцене характерна
не только для Западной Евразии, но и для средних широт Северной и Южной Америки
[Wolfe, Dilcher, 2001; Greenwood, Wing, 1995], а также Австралии и Китая [Greenwood,
1996; Guo Shuangxing, 1993]. В средних широтах Евразии климатическая перестройка
первой половины бартона совпала во времени с потерей связей Западно-Сибирского
моря с Арктическим бассейном к началу формирования тавдинской свиты.
Среднеэоценовые флоры Зайсанской впадины и Южной Гоби Монголии носили смешанный
характер с преобладанием вечнозеленых. Саргамысская флора Зайсана приурочена
к слоям, залегающим над пачками с обайлинским и саргамысским комплексами позвоночных,
формировавшимися, по данным Л.К. Габунии, в начале среднего эоцена. В составе
флоры присутствует целый ряд типичных субтропических растений (Hibiscus,
Ficus, Idesia, крупноплодный Ailanthus и др.), но она не несет
ясно выраженных ксероморфных признаков, хотя и не является крупнолистной
(рис. 2). В Южной Гоби в лютетской флоре Хайчин-Ула-2, в составе прибрежно-водной
группировки много однодольных, обычны обитатели рипарийных фаций - ксероморфные
тополь и ива. Им сопутствуют мелколистные восковница, ильм и ликвидамбр. Флора
существовала, по-видимому, в зоне перехода от сухих субтропиков к более влажной
теплоумеренной зоне.
В Западной Сибири зоны субтропического влажного и переменно-влажного климата,
судя по данным палинологии, распространялась на север по крайней мере до 50°-55
°С с.ш., так как в составе палинокомплексов верхнелюлинворской и нижнетавдинской
подсвит в лютете доминировала пыльца аралиевых и кастанопсиса, а в бартоне
- мелколистных дубов с подчиненным количеством кастанопсиса при крайне незначительном
содержании пыльцы хвойных. Судя по появлению на западе Сибирской платформы
в раннем и начале среднего эоцена бокситов, влажность климата возрастала в
пределах Западно-Сибирской плиты с запада на восток. Это было связано с направлением
течения в морском бассейне, занимавшим большую часть плиты. Вдоль западной
границы Сибирской платформы течение было ориентировано с юга на север и таким
образом переносило теплые воды Тетиса в более высокие широты.
К востоку от Байкала и на Дальнем Востоке сравнительно узкая зона влажного
и переменно-влажного субтропического климата к югу постепенно сменяла зону
переходного климата от умеренно-теплого к субтропическому (флористический экотон,
образованный лесной растительностью с участием вечнозеленых и листопадных элементов).
Зона субтропического климата в среднем эоцене распространялась от Зайсана до
крайнего юга Приморья, Корейского п-ова, Северной Японии и Северо-Восточного
Китая (до Шандунского п-ова на юге, где начиналась зона сухих субтропиков).
Индикаторами субтропического климата являются находки пальм Sabal (Хоккайдо,
Маньчжурия), а также представителей лавровых, магнолиевых, бобовых, гамамелиевых,
анакардиевых и др. семейств. Флоры бартона, особенно на юге Приморья, в целом
более мелколистны и узколистны (тополя, лавровые, буковые, мелколистные цельнокрайние
дубы, кастанопсисы и др.). В то же время почти всегда в захоронениях вечнозеленым
сопутствуют листопадные растения - дериваты раннепалеогеновых умеренно-теплолюбивых
флор: троходендроидесы, платановые, березовые, ореховые, восковниковые и др.
Смена гумидной субтропической флоры лютета на более засушливую в бартоне, отражающую
летневлажный климат муссонного типа, была вызвана закрытием сквозной связи
Арктического бассейна с Тетисом через Тургайский пролив и Западно-Сибирское
море, что привело к нарушению сложившейся ранее водной и атмосферной циркуляции.
Сохранившийся в это время в зоне Тургайского пролива обширный, далеко заходящий
к северу залив Тетиса, занятый теплыми тетическими водами, вызывал появление
своего рода "средиземноморского" эффекта, в результате которого климат
стал сезонным с инверсией основных периодов выпадения осадков. При этом общее
количество летних осадков заметно сократилось, а начиная с бартона пик осадков
стал приходиться на зимний сезон. В среднем эоцене, особенно в бартоне, как
в Евразии, так и, как увидим далее, в Северной Америке заметно возросла амплитуда
колебания сезонных температур, произошло снижение среднезимних температур,
началась пока еще слабо выраженная континентализация климата во внутренних
регионах материков.
В Северной Америке зона влажного и переменно-влажного субтропического климата
в среднем эоцене занимала все пространство средних широт, достигая наибольшей
ширины на западе континента. На севере она, как и в Евразии, постепенно сменялась
зоной умеренно-теплого климата, образуя подобие экотонного пояса со смешанным
типом флор. На западе Северной Америки известно несколько десятков флор, формировавшихся
в прибрежной зоне [Wolfe, Dilcher, 2001]. В Якутатском террейне к ним относится
флора хребта Шарлотты, на юге Британской Колумбии - флора Китсилано Бич, к
югу от Сиетла - Франклинийские флоры каньона Грин Ривер из разрезов серии Пюджет.
Наконец, на крайнем юго-западе США к ним относится флора Дель Map в районе
Сан Диего. Все перечисленные среднеэоценовые флоры, развитые от Аляски до Калифорнии,
также как и более древние палеоценовые и раннеэоценовые флоры этого континента
могли существовать только в условиях высокой влажности климата (от 2000 до
4000 мм среднегодовых осадков). Это относится даже к самой южной из флор -
Дель Map. Что касается среднегодовых температур, то этот параметр изменялся
в пределах от +9 °С для северных, до +24 °С - для южных районов, а амплитуда
сезонных колебаний изменялась от 8° до 19 °С. Среднезимние температуры районов
развития, перечисленных флор были выше 0 °С. С этим выводом согласуются находки
в отложениях прибрежной зоны палеоозера Грин Ривер остатков крокодилов и черепах,
саговников, имбирных и пальм. Зимние температуры здесь не могли спускаться
ниже +5 °С. Богатство систематического состава флор (от 25 до 50 и более видов),
ныне локализованных между 44° и 39° с. ш., позволило рассчитать для них температурные
параметры методом CLAMP. Среднегодовые температуры, среднезимние температуры
и амплитуда сезонных температур оказались соответственно равными для флоры
Кисинжер Лейке (43°48' с.ш.) - +15,7 °С, +6,7 °С, 17,5 °С, для флоры Аппер
Вилвуд (43°48' с.ш.) - +15,3 °С, +6,3 °С, 17,0 °С, для флоры Винд Ривер (43°42'
с.ш.) +16,5 °С, +8,5 °С, 17,1 °С, для флоры Грин Ривер (43° с.ш.) - +14,3 °С,
+6,1 °С, 18,7 °С, для флоры Чок Блаффс (39° 10' с.ш.) -+14,4 °С, +5,6 °С, 9,4
°С. Последняя, насчитывающая 67 видов цветковых, расположена в наиболее внутренней
части Североамериканского континента [Greenwood, Wing, 1995; Wolfe, Dilcher,
2001].
Зона сухих субтропиков Северного полушария. К середине среднего эоцена в Северном
полушарии произошло окончательное оформление аридной зоны, протягивавшейся
от Каспийского моря на западе до Желтого и Восточно-Китайского морей на востоке.
Северная граница этой зоны проходила через Северную Туркмению, Южный Казахстан,
Южную Монголию, а также Таримскую и Цайдамскую впадины внутренних районов Китая.
К морскому побережью эта зона выходила южнее Шандунского п-ова. В пределах
Казахстана она граничила на севере с зоной сезонно-влажных субтропиков, где
ископаемые флоры также несут ксероморфные признаки.
В Туркмении мелколистная ксерофитная флора изучена из трех местонахождений
в районе оз. Эр-Ойландуз, Западного (Акар-Чешме) и Юго-Восточного Бадхыза (г.
Монах). Повсеместно флороносные слои приурочены к морским отложениям туркестанского
региояруса (по О.С. Вялову) и их раннебартонский возраст устанавливался по
комплексам планктонных фораминифер и наннопланктона. В составе флоры известны
представители более 20 семейств, среди которых наиболее разнообразны папоротники,
однодольные, мелколистные лавровые, протейные, анакардиевые, вересковые, сапотовые,
эбеновые, диллениевые, сапиндовые. Доминирующими элементами флоры явились несколько
видов рода Palibinia и вид Rhus turcomanica. Оба таксона мелколистные, с толстой
кутинизированной тканью листа, принадлежали, по-видимому, вечнозеленым кустарниковым
ксерофитам. Н.Д. Василевская [1957], монографически изучившая флору, предполагает
следующие оптимальные условия ее существования: среднегодовая температура +15°-20
°С при сезонно выпадающих годовых осадках от 250 до 1000 мм. Восточное положение
зоны фиксируют многочисленные остатки палибиний из красноцветных, а иногда
и эвапоритовых отложений ряда впадин внутренних районов Китая. Особенно много
местонахождений в провинциях Хунань и Шаньси, где вместе с палибинией встречены
остатки восковниковых, лавровых, ильмовых с ксерофитными признаками. Что касается
климатических показателей внутренних районов Китая, входящих в аридную зону,
то они, вероятно, отличались от таких показателей западного продолжения зоны
лишь меньшим количеством осадков (200-500-600 мм) и их сезонным распределением.
Зона влажного экваториального и тропического климата (на Африканском
континенте, разделенная семиаридной тропической зоной). В среднем эоцене по сравнению с
ранним эоценом Африка сместилась на несколько сотен километров к северу, приблизившись
к ее современному положению относительно экватора. Береговая линия Тетиса на
севере этого континента также мигрировала к северу, в результате чего сократился
прибрежный пояс тропической растительности (гумидных тропиков). В то же время
аридный тропический пояс несколько расширился, распространившись частично на
территорию Нигера. Характерно, что на площадях харвудов Южной Европы (Сардиния,
Испания) во флорах встречаются отдельные выходцы из более южной саванны. Сама
же североафриканская саванна, простиравшаяся от Мавритании и Сенегала на западе
до районов Большого Африканского Рога на востоке, была ареалом развития Leguminosae,
Annonaceae, Combretaceae, Ebenaceae, Euphorbiaceae, Fabaceae, Sterculiaceae
и др. Бобовые вместе с лавровыми и пальмами являлись главными лесообразователями
более влажных тропических лесов, обрамляющих севернее саванные редколесья,
а непосредственно у морского побережья появлялись мангровы. Их остатки известны
из целого ряда местонахождений (Nipodites sickenbergeri из лютета Египта, Sonneratioxylon
aubrevillei и Rhizophora из верхнего лютета Ливии [Boureau et al., 1983 и др.].
Южная граница тропической саванны проходила от северной части Сенегала на восток,
охватывая бассейн верхнего течения р. Нигер, а к югу от Судана поворачивала
на юг, так как восточнее вдоль побережья Индийского океана вплоть до Занзибара
протягивалась полоса влажных тропических лесов. Вся приэкваториальная зона
Западной Африки, особенно примыкающая к акватории, вдававшейся в континент
в районе Гвинейского залива и Камеруна, была занята тропической гилеей и мангровой
растительностью, остатки которой фиксируются к западу от Сенегала на поднятии
о-вов Зеленого Мыса. Здесь Е.Д. Заклинской [Zaklinskaja, 1978] в керне скважины
глубоководного бурения была обнаружена пыльца ризофоры, соннератии, авиценнии
и пальмы нипа. Остатки плодов Nypa burtinii [Fritel, 1921] еще ранее найдены
в Сенегале. Распространение тропической растительности в пределах всей территории
Сенегала подтверждается находками ряда растений, характерных для вечнозеленых
и иногда полулистопадных тропических лесов [Koeniguer, 1989]. Кроме остатков
пальм, здесь встречались древесины сапиндовых, цезальпиниевых, а также форм,
близких к Irvingia и Placodiscus. Восточнее Сенегала полоса мангровых прослеживалась
до Нигерии и Камеруна, где вместе с остатками плодов и пыльцы нипы обнаружена
как пыльца растений тропической гилей (Bombacacidites и др.) и даже болотной
тропической растительности со схизейными и циатейными папоротниками, так и
пыльца растений саванного редколесья, расположенного севернее. О климате западного
побережья Африки (Ангола) и бассейна р. Конго сведений нет. Можно лишь отметить,
что к югу от бассейна р. Конго в направлении Намибии появляются пески формации
Калахари, накапливавшиеся на протяжении всего палеогена в условиях семиаридного
климата.
Климат Индии в среднем эоцене в целом оставался влажным тропическим и только
в районе Кача он был сезонно засушливым. Состав древесных тропической флоры
отличался чрезвычайной пестротой и, кроме представителей мангровых, включал
более десятка родов пальм, а также Musaceae, Sparganiaceae, Bombacaceae, Sterculiaceae,
Tiliaceae, Sapindaceae, Anacardiaceae, Leguminosae, Combretaceae, Myrtaceae,
Lecydiaceae, Lauraceae, Euphorbiaceae, Urticaceae и др. Calophyllum (Guttiferae)
- наиболее распространенное растение тропических побережий - известно из целого
ряда среднеэоценовых местонахождений Индии, а также на трех крупнейших островах
Индонезии - Ява, Суматра и Калимантане. Однако представители двух других родственных
родов пальм - Mesua и Garcinia встречены только в эоцене Индии.
Юго-Восточная Азия, лежащая в экваториальной зоне, судя по составу мегафоссилий
из Индонезии, изученных еще в конце XIX столетия К. Эттингсхаузеном, и по находкам
остатков Nypa и других пальм (Oncosperma, Calamus и Metroxylon),
также характеризовалась влажным тропическим климатом. Широкий спектр тропических
и субтропических таксонов
установлен для среднеэоценовых флор Малайзии [Bande, Prakash, 1986]. Здесь
отмечены Taxodium, Sparganium Nypa, Calamus, Calophyllum, Dipterocarpus,
Hopea, Durio, Cullenia, Pterospermum, Pentace, Jarandersonia, Abacolosa, Sapindus,
Gluta, Capanosperma, Dalbergia, Cassia, Terminalia, Ficus. В пределах
Японского архипелага граница тропической и влажной субтропической зон проходила
вероятно
между о-вами Хонсю и Кюсю, так как среднеэоценовая флора последнего представлена
исключительно вечнозелеными растениями.
В северную часть гумидной тропической зоны, как и в раннем эоцене, входили
все провинции крайнего юга Китая, включая о. Хайнань. Характерные флористические
комплексы с доминированием вечнозеленых растений (главным образом, лавровых)
были описаны из формации Синао провинции Гуаньси [Zjang, 1988]. Северная граница
тропической зоны китайскими палеоботаниками прослежена и западнее, вплоть до
окрестностей Лхассы, откуда были описаны остатки пальм (ливистона и сабаль),
а также других растений - показателей влажного тропического климата: Cyclosorus,
Magnolia, Annona, Litseaephyllum, Laurophtllum, Daphnogene, Ficus, Lithocarpus,
Melanirrhoea, Sapindus и др.
Климат южной части Австралии в среднем эоцене реконструируется по богатейшим
флорам, изученным в последнее десятилетие из периферических частей морских
бассейнов Отуэй, Джипсленд, Басе и Муррей, открывавшихся в океан. Большая часть
этих флор: Голден Гро, а также Мэслин Бэй и Морнингтон, расположенные во внутренних
частях континента (Пул Крик, Нелли Крик и Стюарт Крик) скорее лютетские, а
меньшая часть (Бартон Рэндж, Флиндерс, Трэрарден и одна из богатейших флор
континента - Энглезея - бартонские [Greenwood, 1987; 1992; 1996; Greenwood,
Wing, 1995; Greenwood, Conran, 2000; Greenwood et al., 2000; Christophel, Greenwood,
1987; Christophel et al., 1992]. Таксономический анализ этих среднеэоценовых
флор и их сравнение с составом основных эдификаторов современных тропических
дождевых лесов Северо-Восточной Австралии свидетельствуют об их широком распространении
в пределах упомянутых бассейнов и на периферии паратропических дождевых лесов,
приобретающих иногда характер муссонных.
Лишь во внутренних частях Австралии климат становился более отчетливо сезонным,
причем некоторые развитые здесь флоры имеют ксероморфный облик (Нелли Крик).
Количество таксонов с цельнокрайней простой листовой пластинкой в составе этих
флор колеблется от 60 до 80%. Листья обычно мезофильные, крупные, характерные
для дождевых лесов. Практически во всех местонахождениях (кроме Неррига) встречены
остатки пальм. Помимо Nypa, отмеченной во многих местонахождениях,
описаны представители других родов пальм, а также прочих однодольных тропического
родства:
Paracordyline, Piper и др. Обычны остатки растений мангровых, а также
представителей многих тропических родов и семейств древесных цветковых, растущих
вблизи морских
побережий, ареалы которых ныне практически не выходят за пределы тропической
зоны. Таковы Eleocarpaceae (Sloanea/Elaeocarpus), Sterculiaceae (Brachychiton),
Proteaceae (Banksiaephyllum), многие Lauraceae и др. Тропический климат дождевых
лесов подчеркивается и составом хвойных, особенно ногоплодниковых, ныне распространенных
в лесах Северной и Северо-Восточной Австралии. Для восстановления климата среднего
эоцена южных регионов Австралии Д. Гринвудом для некоторых местонахождений
были рассчитаны климатические параметры методом CLAMP (для лютета по флорам
Пул Крик, Нелли Крик, Голден Гров, Неррига, для бартона - Англезея). Количество
таксонов, использованных при подсчете флор прибрежной зоны континента колеблется
от 21 до 28, для более внутренних его частей 11-14. Для южной группы лютетских
флор (34°-35° ю.ш.) среднегодовые температуры оцениваются как +18°-+20 °С,
температуры наиболее холодного месяца +10°— +11 °С, амплитуда колебания сезонных
температур 16°-17 °С, количество среднегодовых осадков 1500-2000 мм. Для более
северных флор (29° ю.ш.) среднегодовые температуры несколько выше: +19°-+21
°С, также как и температуры самого холодного месяца (+12,4°— +14 °С), при среднегодовых
осадках 1100-1360 мм, с их сезонным распределением и сокращением в три наиболее
сухих месяца до 50 - 100 мм. Это придает климату муссонный характер, так как
основная масса осадков выпадала в летние месяцы. Возможно этим вызван ксероморфизм
листьев флор Нелли Крик и Пул Крик. Амплитуда колебания сезонных температур
у более северных флор также ниже, чем у более южных флор (12°-13 °С). Климатические
показатели бартона для южных районов Австралии по влагообеспеченности
и температуре по сравнению с лютетскими несколько изменились, хотя участие
в составе бартонской флоры мезофильных вечнозеленых тропических растений сохранилось.
Наиболее характерными элементами флоры Англезея в Виктории являются папоротник-лиана
Lygodium, Bowenia, ногоплодники тропического родства: Falcatifolium,
Dacrycarpus, Podocarpus platyphyllum, Prumnopitys lanceolata, Decussocarpus. Тропический
состав цветковых подчеркивается широким представительством семейств Loranthaceae,
Sapindaceae, Rutaceae, Lauraceae, присутствием Brachychiton, Sloanea, Gymnostoma,
Banksiaephyllum и пр. В составе бартонской флоры не менее трех родов пальм,
саговники и имбирные. Изменение климата сказалось на некотором понижении среднегодовых
температур (17,1 °С) и средне-зимних (8,2 °С) температур при одновременном
увеличении количества среднегодовых осадков (2220 мм) и ростом амплитуды распределения
их по сезонам (240 и 1400 мм). Это позволяет заключить, что во вторую половину
среднего эоцена полоса развития муссонного климата, по-видимому, сместилась
южнее и к северу от этой полосы климат стал переходным от гумидного тропического
к сезонному субтропическому. Вся северная часть Австралии в среднем эоцене
оставалась в тропической зоне.
На юго-востоке Северной Америки в начале среднего эоцена (флоры группы Клэйборн)
сохранялся морской залив, глубоко заходящий во внутренние районы континента
к северу от современного Мексиканского залива. Вся названная часть континента
по-прежнему лежала в пределах тропической зоны, однако сезонность климата здесь
стала проявляться в меньшей степени. Многие местонахождения среднеэоценовой
флоры в Техасе содержат остатки мангровых растений и пальм, включая Nypa из
формации Ларедо серии Клэйборн. Это позволяет заключить, что среднегодовые
температуры на территории этого штата в среднем эоцене были не ниже +19 °С,
а температура поверхностных морских вод +24 °С. К характерным представителям
тропического корня можно отнести встреченные во флорах этого возраста Annona,
Bombacaceae, Oreomunnea-Alfaroa, тропические рода семейств Eophorbiaceae и
Sapindaceae. При этом среднеэоценовые тропические флоры юго-востока Северной
Америки, в отличие от раннеэоценовых, все более сближались с неотропическими
флорами Центральной и Южной Америки, теряя, хотя и не полностью, связь с восточноазиатскими.
Центральная Америка, судя по материалам, полученным в Гватемале и Панаме, в
среднем эоцене также входила в зону влажного тропического климата [Graham,
1994]. Южнее в то время были развиты дождевые тропические леса, которые занимали
всю приэкваториальную часть Южной Америки и распространялись на юг до средних
широт. Однако, по мнению Э. Ромеро [Romero, 1986], южная граница тропической
зоны в среднем эоцене переместилась несколько к северу, по сравнению с ее положением
в раннем эоцене, когда тропики занимали практически всю территорию континента.
Среднеэоценовые флоры были открыты в первой половине XX в. в разных районах
Колумбии, Венесуэлы и Эквадора в осадках окраинных частей морских бассейнов,
омывавших Бразильский щит. Фруктификации покрытосеменных, принадлежащие Sapindoides и Celtis
bolivarensis, были описаны Э. Берри из департамента Боливар Колумбия
[Berry, 1924; 1929]. В Венесуэле на юго-восточном побережье Венесуэльского
залива из формации Эскуке был описан типичный представитель Неотрописа Entada
boweni [Berry, 1920]. Характерные растения прибрежных тропических дождевых
лесов были найдены в районе Анконы (п-ов Святой Елены, Эквадор) непосредственно
к северу от экватора. Находки происходят из среднеэоценовой толщи песчаников
Соккоро [Berry, 1932; Shepard, 1937; Hoffstetter, 1936b]. Э. Берри отсюда определены
Astrocarym shepparsii, Ventanea sheppardii, Palmacarpon bravoi, Annona
peruviana, Sapindoides peruvianus. Последние три таксона известны также из среднего эоцена
соседней Колумбии.
Э. Ромеро [Romero, 1986] отмечает, что в среднем эоцене неотропическая палеофлора
занимала все пространство континента по крайней мере до 20° ю.ш., замещаясь
южнее смешанной палеофлорой, состоящей из неотропических и антарктических элементов,
получившей распространение до 50° ю.ш., причем в северной части (до 30° ю.ш.)
в составе этой смешанной флоры Nothofagus отсутствовал. Можно заключить, что
граница между зонами тропиков и субтропиков в Южной Америке в среднем эоцене
проходила от широты г. Сант Яго на Тихоокеанском побережье через низовья Параны
до Атлантического океана на крайнем юге Бразилии.
Остается открытым вопрос о формировании в эоцене на Южноамериканском континенте
тропических саванн. Гонсалес-Гусман [Gonzalez-Guzman, 1967] считал, что уже
на рубеже раннего и среднего эоцена саванные группировки с участием Malvaceae
и Jussiae получили распространение на территории Колумбии в районе Тибу. Однако
более поздними исследованиями это не было подтверждено.
Зона сухих и влажных субтропиков Южного полушария. Зона гумидного и переменно
влажного субтропического климата в Южном полушарии на юге Австралийского континента
устанавливается лишь в конце среднего эоцена, когда в связи с похолоданием
произошло снижение среднегодовых и среднезимних температур на его южном побережье,
но особенно в Тасмании, где впервые в макроостатках был встречен Nothofagus.
В то же время в Новой Зеландии, все более удалявшейся от континентальных масс
бывшей Гондваны, флора стала все шире приобретать индивидуальные черты. По
палинологическим данным [Mildenholl, 1980], существенных различий в составе
флоры северного и южного острова не отмечается. Занимая позицию в средних широтах
(45°-55° ю.ш.) архипелаг входил в зону влажных субтропиков с очень низкой амплитудой
колебаний сезонных температур, относительно высокими среднезимними температурами
(выше +10 °С) и среднегодовыми температурами 15°-20 °С [Pole, 1992].
От Новой Зеландии пояс влажного субтропического климата через Южную Пацифику
трассировался в среднем эоцене к Южной Америке. Показателем субтропического
климата того времени в Патагонии является богатая флора Рио Турбио, изученная
Э. Берри [Berry, 1937], М. Хуникином [Hunicken, 1967], С.М. Архангельским и
Э. Ромеро [Archangelsky, Romero, 1974; Romero, 1986]. Флора насчитывала более
100 таксонов, которые образовывали смешанный комплекс с участием неотропических
и антарктических растений. Первые представлены Allophyllos, Annona, Bignonia,
Buettneria, Cupania, Nectandra, Phoebe, Styrax, Tetracera и др. Доля представителей
антарктической флоры более скромная. Кроме Nothofagus, этот тип флоры представляют
Embothriophyllum, Rhoohyllum и др. Содержание листьев с цельнокрайней пластинкой
достигает лишь 40%. По мнению Д. Дильчера [Dilcher, 1973], такое содержание
характерно для лесов гумидной теплоумеренной зоны, а по представлению Дж. Вулфа
[Wolfe, 1971] - скорее для субтропических дождевых лесов.
Зона умеренного и умеренно-теплого влажного климата Южного полушария. В среднем
эоцене эта зона ограничивалась регионами, лежащими в высоких широтах Южного
полушария, а именно Огненной Землей и Антарктидой с прилегающими островами,
откуда еще в начале прошедшего века были описаны
находки ископаемых растений (о-ва Сеймур и Кинг Джордж). Нельзя исключать возможность
вхождения в эту климатическую зону Тасмании (полностью или частично), так как
флора Сетхана Северной Тасмании содержит мелколистные нотофагусы, которые типичны
для умеренного и умеренно-теплого климата и заключает не более 35% листьев
с цельным краем.
Обнаруженные в среднем эоцене на о-ве Кинг Джордж остатки древесин принадлежат
семействам Araucariaceae, Podocarpaceae, Cupressaceae, Fagaceae (Nothofagus),
Monimiaceae, Cunoniaceae, Eucryphiaceae и Rubiaceae, а в сопутствующем палинокомплексе
присутствуют споры глейхениевых, полиподиевых и циатейных папоротников, ногоплодниковых,
а также протейных (Beaupreaidites), гуннеровых и буковых (Nothofagus)
[Torres, 1992]. Богатый комплекс растений неотропического и антарктического
родства
с участием ногоплодниковых, южных буков, протейных и лавровых (?) еще в начале
века описал П. Дузен [Dusen, 1907] с о-ва Сеймур. Большинство рецентных аналогов
этих ископаемых форм ныне обитает в прибрежных районах Южного Чили. Это позволяет
по аналогии дать такую оценку среднепозднеэонового климата района Антарктического
полуострова: среднегодовая температура +6 °С, среднелетние 8°-10 °С, при среднегодовом
количестве осадков более 500 мм/год.
В данной главе представлен реконструированный по палеоботаническим
данным глобальный климат Земли в раннем палеогене от 65 до 37-38 млн лет назад,
когда еще на
Земном шаре сохранялась "теплая" биосфера. Рассмотренное выше похолодание
на рубеже мела и палеоцена, хотя и носило глобальный характер, было связано
с общепланетарной регрессией, осушением морских рукавов, по которым осуществлялся
тепло- и влагоперенос из низких в высокие широты. Импактные события, если они
и имели место, вероятно лишь "спровоцировали" ряд событий исключительно "земного" происхождения,
нарушившие биоразнообразие на Земном шаре. Об этом свидетельствуют факты гетерохронного
вымирания различных групп организмов в пределах довольно длительного (не менее
3-5 млн лет) переходного интервала от мела к палеогену. Наземная флора высших
растений и образованный ими растительный покров - самая незащищенная часть
поверхностной биоты, в рамках этого интервала оказалась в
незначительной степени подвержена процессам элиминации. Более того, флоры позднего
маастрихта и раннего дания демонстрировали плавные широтные изменения таксономического
состава, фиксируя наличие на Земле определенных климатических поясов и континуальность
процессов климатических изменений. Зафиксированные аномальные содержания иридия
в костных остатках динозавров в среднемаастрихтских отложениях Приамурья [Моисеенко
и др., 1997], а также факты существования в некоторых разрезах нескольких слоев
с аномальным содержанием иридия в переходных слоях от мела к палеогену [Катастрофы
в истории Земли..., 1986] свидетельствуют, что эти аномалии имеют не космическую,
а скорее вулканическую природу.
Максимальное потепление на Земном шаре фиксируется в переходном интервале от
палеоцена к эоцену и охватывает значительный интервал раннего эоцена. Климат
Земли в это время был теплее современного на 6°-8 °С. Эффект потепления усиливался
и широкими возможностями осуществления тепло- и влагопереноса из низких в высокие
широты, благодаря наличию в раннем палеогене сквозных морских рукавов, связывающих
Тетис с Арктическим бассейном при отсутствии широтных орографических барьеров
в Евразии - горные цепи Альпид.
Одной из особенностей глобального климата в конце палеоцена было формирование
муссонной обстановки в средних широтах обоих полушарий с последующей трансформацией
муссонного климата в конце лютета в сезонный переменно-влажный, а в бартоне
местами и семиаридный. В Северном полушарии, где в раннем палеогене, как и
ныне, были сосредоточены континентальные массы, подобная трансформация климата
была связана с нарушением важнейших коммуникаций, связывающих низкие и средние
широты с высокими - замыканием в конце лютета или в начале бартона Западно-Сибирского
внутреннего эпиконтинентального моря, ранее осуществлявшего связь Арктического
бассейна с Тетисом. Поступление теплых вод Тетиса на пространства Западно-Сибирской
плиты с формированием в пределах эпиконтинентального бассейна их замкнутой
циркуляции привело к эффекту, очень напоминающему ситуацию в современном Средиземноморском
бассейне после потери его связей с Индийским океаном, но при сохранении их
с Атлантическим.
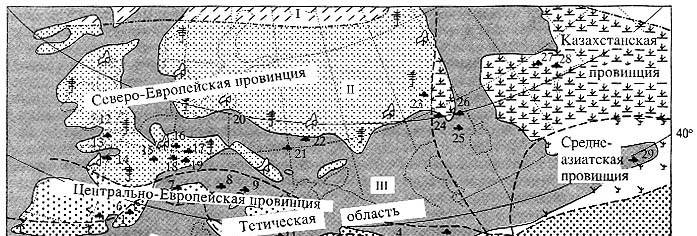 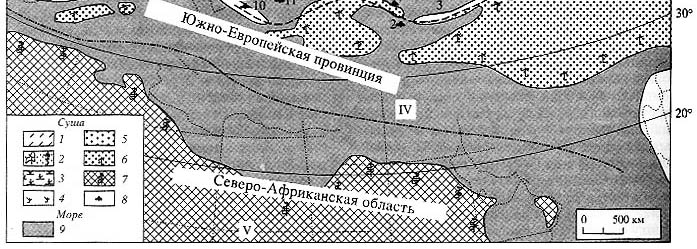 |
Рис. 4. Фитогеографическая и климатическая зональность Западной Евразии
в приабоне 1) Бореальная область; 2-6) Тетическая область; 2) вечнозеленые и летнезеленые хвойно-широколиственные, главным образом, дубово-лавровые леса; 5) дубово-лавровые жестколистные леса и маквис с анакардиевыми, вересковыми, бобовыми и восковниковыми, в прибрежной зоне — кустарниковые и кустарничковые ассоциации, образованные вересковыми, бобовыми и галофитами (маревые, эфедра и др.); 4) травянисто-кустарничковые формации, с участием галофитов; 5) жестколистные ксерофильные леса, образованные кипарисовыми, лавровыми буковыми и бобовыми; 6) вечнозеленые мезофильные субтропические и тропические леса с наземным покровом из папоротников, местами по морским побережьям мангровые ассоциации; 7) Северо-Африканская область: тропические леса, образованные лавровыми, бобовыми и буковыми, замещающиеся вглубь континента саванной; 8) главнейшие местонахождения позднеэоценовых флор: 1 — Селас, 2 — Дарры-Даг, 3 — Талыш, 4 — Ахалцихе, 5 — Эбро, 6 — Брив, 7 — Херинг, 8 — Брезно, 9 — Гирбоу, 10 — Овце Полье, 11 — Хвойна, 12 — Хэмпшир, 13 — о-в Уайт, 14 — Сарш, 15 — Мульхауз, 16 — Цейц, 17 — Мозель, 18 — Старо-Седло, 19 — Кучлин, 20 — Светлогорск, 21 — Кочетын, 22 — Тим, 23 — Орь, 24 — Баки, 25 — Шандинский грабен, 26 — Романова, 27 — Жамантуз, 28 — Такырсор, 29 — Гиссар. Климатические зоны: I — зона умеренно-теплого влажного климата; II — зона субтропического влажного климата; III — зона семиаридного субтропического климата; IV — зона влажного тропического климата; V — зона семиаридного тропического климата |
Детальное расссмотрение глобального климата и климатической
зональности позднего эоцена не входило в задачи автора данного раздела, так
как этим проблемам посвящено несколько специальных работ [Ахметьев, Запорожец,
1992; Геологические и биотические события..., 1996, 1998; Akhmetiev, 2000]
и разделы монографии, посвященной проблемам биогеографии и климата позднего
эоцена и олигоцена Северного Перитетиса [Popov et al., 2001, 2002] (рис. 4).
Поэтому ниже мы остановимся лишь на основных выводах, сделанных в перечисленных
публикациях, относительно глобального климата позднего эоцена. Прежде всего
надо подчеркнуть, что на протяжении этого периода постепенно возрастала континентализация
климата, что сопровождалось смещением климатических зон с севера на юг в Северном,
и с юга на север в Южном полушариях, а также возрастанием температурного градиента
в системе "полюс-экватор", существенным увеличением амплитуды сезонных
температурных колебаний на всех широтах и, главное, быстрым падением среднезимних
температур в средних и высоких широтах. Наиболее интенсивными планетарные изменения
климата происходили в конце позднего эоцена и на рубеже эоцена и олигоцена.
Это было связано с цепочкой последовательных событий, начавшихся с окончательной
изоляцией Антарктического континента от других континентов бывшей Гондваны
(образование пролива Дрейка между Антарктикой и Южной Америкой), одновременным
формированием циркумантарктического течения,
что привело к началу оледенения Антарктики, гляциоэвстатическому падению уровня
вод Мирового океана, общему нарушению ранее существовавшей системы океанической
циркуляции и глобальной регрессии. Эта регрессия сопровождалась уходом морских
вод из внутренних бассейнов Северного полушария – осушением Западно-Сибирского
моря и Тургайского пролива [Popov et al., 2001] равно как и глубокого морского
залива, протягивавшего от современного Мексиканского залива в направлении границы
Канады и США через внутренние районы Мидконтинента. Начавшиеся поднятия в широтной
Альпийской зоне Евразии, а особенно в пределах Китайской ветви этой зоны, привели
к некоторому нарушению меридионального атмосферного влагопереноса из низких
в высокие широты.
Сдвиг границ широтных климатических зон в сторону экватора на несколько сотен
км сопровождался в обоих полушариях экспансией умеренной флоры в более низкие
широты из высоких, что подтверждается многими десятками и первыми сотнями местонахождений
ископаемых растений на всех континентах. Одновременно происходило резкое усиление
континентализации климата с формированием снежного покрова в зимний период
не только в высоких, но отчасти и в средних (50°-60° с. и ю.ш.) широтах.
Таким образом, на рубеже эоцена и олигоцена по существу произошел достаточно
быстрый в геологическом масштабе времени (3 — 5 млн лет) переход от "теплой" (парниковой)
к "холодной" (криогенной) биосфере.
Работа выполнена при поддержке грантов РФФИ № 02-05-65170 и НШ-1615.2003.5.
Аблаев А.Г. Позднемеловая флора Восточного Сихотэ-Алиня и
ее значение для стратиграфии. Новосибирск: Наука, 1974. 180 с.
Ахметьев М.А. Фитостратиграфия континентальных отложений палеогена и
миоцена Внетропической Азии. М.: Наука, 1993. 142 с.
Ахметьев М.А. Казахстан и Центральная Азия (равнины и предгорья) // Палеоген
и изменение климата и ландшафтов за последние 65 млн лет. М.: Геос, 1999.
С. 168-178.
Ахметьев М.А., Запорожец Н.И. Новые находки пальм из олигоценовых отложений
Дарры-Дага (Нахичеванская АССР) // Бюл. МОИП. Отд. геол. 1989. Т. 64, вып.
6. С. 57-67.
Ахметьев М.А., Запорожец Н.И. Климаты олигоцена востока Крымско-Кавказской
области и Казахстана (по анализу макрофлоры, палинокомплексов и комплексов
фитопланктона) // Изв. АН СССР. Сер. геол. 1992. № 3. С. 5-23.
Ахметьев М.А., Запорожец Н.И., Макулбеков Н.М. Растительные мегафоссилии,
споры, пыльца и диноцисты эоцена из тавдинской свиты Павлодарского Прииртышья
// Стратиграфия.
Геол. корреляция. Т. 12. № 1.
Ахметьев М.А., Кезина Т.В., Кодрул Т.М., Манчестер С.Р. Стратиграфия и
флора пограничных слоев мела и палеогена юго-восточной части Зейско-Буреинского
осадочного бассейна // Сборник памяти члена-корреспондента АН СССР, профессора
Всеволода
Андреевича Вахрамеева: (К 90-летию со дня рождения). М.: ГЕОС, 2002. С.
275-315.
Берг Л.С. Основы климатологии. М.; Л.: Госиздат, 1927. 265 с.
Большая советская энциклопедия. 3-е изд. М.: Сов. энциклопедия, 1973. Т.
12. С. 305-308.
Буданцев Л.Ю. История арктической флоры эпохи раннего кайнофита. Л.: Наука,
1983. 156 с.
Василевская Н.Д. Эоценовая флора Бадхыза // Сборник памяти Африкана Николаевича
Криштофовича. М.; Л.: Изд-во АН СССР, 1957. С. 103-176.
Геологические и биотические события позднего эоцена – раннего олигоцена
на территории бывшего СССР. Ч. 1. Региональная геология верхнего эоцена
и нижнего
олигоцена / Отв. ред. В.А. Крашенинников, М.А. Ахметьев. М.: Геос, 1996.
314 с. (Тр. ГИН; Вып. 489).
Геологические и биотические события позднего эоцена-раннего олигоцена на
территории бывшего СССР. Ч. 2. Геологические и биотические события / Отв.
ред. В.А. Крашенинников,
М.А. Ахметьев. М.: Геос, 1998. 250 с. (Тр. ГИН; Вып. 507).
Головнева Л. Б. Маастрихтские и датские флоры Корякского нагорья. СПб.:
Наука, 1994. 148 с.
Катастрофы в истории Земли: Новый униформизм. М.: Наука, 1986. 420 с.
Корнилова B. C. О полтавской флоре Казахстана // Докл. АН СССР. 1955. Т.
104, № 1. С. 124-127.
Красилов В. А. Цагаянская флора Амурской области. М.: Наука, 1976. 92 с.
Криштофович А. Н. Гренландская третичная флора на Северном Урале и ботанико-географические
провинции третичного периода // Природа. 1928. № 5. С. 449-502.
Криштофович А. Н. Развитие ботанико-географических областей Северного полушария
с начала третичного периода // Вопросы геологии Азии. М.; Л.: Изд-во АН
СССР, 1955. Т. 2. С. 824-844.
Макулбеков Н. М. Эоценовая флора Северного Казахстана. Алма-Ата: Наука
КазССР, 1972. 177 с.
Макулбеков Н. М. Палеоценовые и эоценовые флоры Центральной Евразии: (Этапы
развития, корреляция морских и континентальных толщ): Автореф. дис. ...
д-ра геол.-минерал. наук. М., 1997. 67 с.
Моисеенко В. Г., Сорокин А. П., Болотский Ю. Л. Ископаемые рептилии Приамурья.
Хабаровск: АмурНЦ ДВО РАН, 1997. 53 с.
Спайсер Р. Э., Герман А. Б. Меловой климат Азии и Аляски: Сравнение палеоботанических
свидетельств с компьютерной моделью // Палеонтол. журн. 1998. № 2. С. 3-18.
Станиславский Ф. А. Палеогеновая флора с. Волянщины: Автореф. дис.... канд.
геол.-минерал. наук. Киев, 1950. 31 с.
Станиславский Ф. А. Об остатках макклинтокий и возрасте вмещающих их палеогеновых
отложений УССР // Ботан. журн. 1956. Т. 41, № 8. С. 1188-1193.
Akhmetiev M. A. The Late Eocene and Oligocene phytogeography of Western
Euroasia // Paleontol. J. 2000. Vol. 34, N2, suppl. 1. P. 106-115.
Archangelsky S.A., Romero E.J. Los registros mas antiguos de Nothofagus
(Fagaceae) de Patagonia (Argentina у Chile) // Bol. Soc. Bot. Mexico. 1974.
N 33. P.
13-30.
Baily I.W., Sinnot E.W. A botanical index of Cretaceous and Tertiary climates
// Science. 1915. Vol. 41. P. 831-834.
Bamford M. Tertiary angiosperm woods from the west coast of South Africa
// Abstr. VI Conf. of IOP. Quinhuangdao Hubei, 2000. P. 7-8.
Bande M.B., Prakash U. The Tertiary flora 6f Southeast Asia with remarks
of its palaeoenvironment and phytogeography of the Indo-Malayan Region
// Rev.
Palaeobot. and Palynol. 1986. Vol. 49. P. 203-233.
Berry E.W. Tertiary fossil plants from Venezuela // Proc. US Natur. Hist.
Mus. 1920. Vol. 59. P. 553-579.
Berry E.W. The flora of the Concepcion - Arauco coal measures of Chile
// John Hopkins Univ. Stud. Geol. 1922. Vol. 4. P. 73-132.
Berry E.W. The Middle and Upper Eocene floras of southeastern North America
// US Geol. Surv. Prof. Pap. 1924. Vol. 92. P. 1-206.
Berry E.W. Petrified fruits and seeds from Oligocene of Peru // Pan-Amer.
Geol. 1927. Vol. 47. P. 121-132.
Berry E.W. A revision of the flora of the Latath formation // US Geol.
Surv. Prof. Pap. 1929. Vol. 154-H. P. 225-264.
Berry E.W. A new palm from the Upper Eocene of Ecuador//J. Wash. Acad.
Sci. 1932. Vol. 22. P. 327-329.
Berry E.W. Eocene plants from Rio Turbio, in the territory of Santa Cruz,
Patagonia // John Hopkins Univ. Stud. Geol. 1937. Vol. 12. P. 91-98.
Berry E.W. Tertiary flora from the Rio Pichileufu, Argentina // Spec. Pap.
Geol. Soc. Amer. 1938. Vol. 12. P. 1-149.
Boulter M.C., Kvacek Z. The Palaeocene flora of the Isle of Mull. L.: Palaeontol.
Assoc., 1989. 149 p. (Spec. Pap. Palaeontol.; № 42).
Boureau E., Cheboldaeff-Salard M., Koeniguer J.-C. et al. Evolution des
flores et de la vegetation Tertiares Afrique, au nord de 1'Equateur //
Bothalia.
1983. Vol. 14, N 3/4. P. 355-367.
Christophel D.C., Greenwood D.R. A megafossil flora from the Eocene of
Golden Grove, South Australia // Trans. Roy. Soc. S. Austral. 1987. Vol.
111. P.
155-162.
Christophel D.C., Scriven L.J., Greenwood D.R. An Eocene megafossil flora
from Nelly Greek South Australia // Ibid. 1992. Vol. 116, N 2. P. 65-76.
Ditcher D.L. The Eocene floras of Southeastern North America // Vegetation
and vegetational history of Northern Latin America / Ed. A. Graham. Amsterdam:
Elsevier, 1973. P. 39-59.
Dusen P. Uber die tertiare Flora des Seymour Insel // Wiss. Ergebn. Nordsk
Sweden Sudpolar Exped., 1901-1903. 1907. N3. S. 1-21.
Engler A. Versuch einer Entwicklungsgeschichte der Pflanzenwelt seit der
Tertiarperiode Leipzig: Engelmann, 1879. 202 S.
Engelhardt H. Die Tertiarflora des Jesuitengrabens bei Kundratice in Nordbohmen
// Nova Acta Acad. Caes. Leop.-Carol. German. Nat. Cur. 1895. Bd. 48. S.
259-408.
Frakes A.L., Kemp E.M. Influense of continental position on Early
Tertiary climates // Nature. 1972. Vol. 240. P. 97-100.
Frederiksen N.O. Sporomorphs from the Jackson Groupe (Upper Eocene) and
adjacent strata of Mississippi and Western Alabama // US Geol. Surv. Prof.
Pap. 1980.
Vol. 1084. P. 1-75.
Frederiksen N.O. Sporomorphs biostratigraphy, floral changes and paleoclimatology
Eocene and Earliest Oligocene of the eastern Gulf Coast // Ibid. 1988.
Vol. 1448. P. 1-68.
Friedrich W.L. Zur Geologie von Brjanslaekur (Nordwest-Island) unter besonderer
Beriicksichtigung der fossilen Flora. Koln, 1966. 108 S. (Sonderveroff.
Geol. Inst. Univ.; Bd. 10).
Fritel P.H. Sur la decouverte au Senegal de deux fruits fossiles appartenant
aux genres Kigelia D.C. et Nipadites Bowerb//C.r. Acad. sci. 1921. Vol.
173. P. 245-246.
Gardner J.S. A monograph of the British Eocene flora. L., 1879. 86 p.
Golovneva L.B. Early Palaeogene floras of Spitsbergen and North Atlantic
floristic exchange // Acta Univ. carol. Geol. 2000. Vol. 44, N 1. P. 39-50.
Gonzalez-Guzman A.K. A palynological study on the Upper Los Cuervos and
Mirador Formation (Lower and Middle Eocene, Tibu Area, Colombia). Leiden:
Brill,
1967. 68 p.
Graham A. Neotropical Eocene coastal floras and 180/160 - extimated warmer
vs. Cooler Equatorial Water // Amer. J. Bot. 1994. Vol. 81, N 3. P. 301-306.
Greenwood D.R. Early Tertiary Podocarpaceae: megafos-sils from Anglesea
locality, Victoria // Austral. J. Bot. 1987. Vol. 35. P. 111-133.
Greenwood D.R. Taphonomic constraints on foliar physiognomic interpretations
of Late Gretaceous and Tertiary pale-oclimates // Rev. Palaeobot. and Palynol.
1992. Vol. 71. P. 142-194.
Greenwood D.R. Eocene monsoon forest in Central Australia? // Austral.
Syst. Bot. 1996. Vol. 9. P. 95-112.
Greenwood D.R., Collinson M.E. The origins and Palaeogene history of modern
plant communities // Palaeontol. Soc. Spec. Publ. 1992. N 6. P. 1-113.
Greenwood DR., Conrad J.G. The Australian Cretaceous and Tertiary Monocot
fossil record // Monocots: Systematics and evolution. Melbourne: CSIRO,
2000. P. 52-59.
Greenwood D.R., Vadal A.J., Douglas J.G. Victorian Paleogene and Neogene
macrofloras: A conspectus // Proc. Roy. Soc. Victoria. 2000. Vol. 112,
N 1. P. 65-92.
Greenwood D.R., Vadal A.J., Moss P., Keefe R. Early Paleocene
climates and vegetation in SE Australia // Climate and biota of the Early
Paleogene:
Intern, meet.
Wash. (D.C.): Smithsonian Inst. 2001. Abstr. Vol. P. 38.
Greenwood D.R., Wing S.L. Eocene continental climates and latitudian gradients
// Geology. 1995. Vol. 23. P. 1040-1048.
Guo Shuangxing. Late Cretaceous and Early Tertiary floras from the southern
Guangdong and Guangxi with their stratigraphical significance // Mesozoic
and Cenozoic red beds of South China. Beijing: Science press, 1979. P.
231-233.
Guo Shuangxing. The evolution of the Cenozoic tropical monsoon climate
and monsoon forests in Southwestern China // Evolving landscapes and evolving
biotas of East Asia since the Mid-Tertiary: Proc. 3th Conf. on the evolution
of the
East Asian environment / Ed. N.G. Jablonski. Hong Kong: Univ. of Hong Kong,
1993. P. 124-135.
Herman А.В., Spicer R.A., Kvacek J. Late Cretaceous climate of
Eurasia and Alaska: A quantitative palaeobotanical approach // Aspects
of Cretaceous
stratigraphy
and palaeobio-geography: Proc. 6th Intern. Cretaceous symp., Vienna, 2000.
Wien: Verl. Osterr. Acad. Wiss. 2002. Vol. 15. P. 93-108.
Hoffstetter R. Lexique stratigraphique international. P.: CNRS, 1936a.
Vol. 5: Amerique Latine, fasc. 5b: Peru. 16 p.
Hoffstetter R. Lexique stratigraphique international. P.: CNRS, 1936b.
Vol. 5: Amerique Latine, fasc. 5a: Ecuador. 16 p.
Hollick A. The Tertiary floras of Alaska // US Geol. Surv. Prof. Pap. 1936.
Vol. 182. P. 1-185.
Hu H. H., Chaney R.W. Miocene flora from Shantung Province, China//Palaeontol.
sinica. N.S. 1940. N 1. P. 1-82.
Hunicken M. Flora Terciaria de los Estratos de Rio Turbio, Santa Cruz //
Rev. Fac. Exact Fis. Nat. Univ. Cordoba. Secc. Natur. 1967. N 27. P. 139-227.
Jaramillo С. A. Response of tropical vegetation to the Early Paleocene
warming climate // Climate and biota of the Early Paleogene: Intern, meet.
Wash.
(D.C.): Smithsonian Inst., 2001. Abstr. vol. P. 48.
Keefe R.L., Greenwood D.R. Ecology of an Early Eocene rainforest at Brandy
Creek, Victorian High Plains, South-Eastern Australia // Ibid. 2001. Abstr.
vol. P. 50.
Kennedy E., Raine J.I. Terrestrial Paleocene climate and leaf flora of
New Zealand // Ibid. 2001. Abstr. vol. P. 51.
KoeniguerJ.-Cl. Les Paleosavanes du nord de 1'Afrique // Mem. Soc. Biogeor.
Ser. 3. 1989. N 111. P. 63-73.
Lakhampal R.N. Floristic evidence in the stratigraphical subdivision of
the Indian Tertiary // Aspects and appraisal of Indian palaeobotany. Lucknow,
1974. P. 516-525.
Mai D. Tertiare Vegetations - geschichte Europa. Jena: Fischer, 1995. 692
S.
Maley J. The African rain forest - main characteristics of changes in vegetation
and climate from the Upper Cretaceous to the Quaternary // Proc. Roy. Soc.
Edinburgh B. 1996. Vol. 104. P. 31-73.
McKaena M.C. Sweepstakes, filters, corridors Noah's Arks, and beached Viking
funeeral ships in palaeogeography // Implications of continental drift
to the Earth sciences / Ed. D.H. Tarling, S.K. Runcorn. L.; N.Y.: Acad.
press,
1983.
Vol. 1. P. 295-308.
Mildenholl D.C. New Zealand Late Cretaceous and Cenozoic plant biogeography:
A contribution // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1980. Vol.
31. P. 197-233.
Orlando H.A. La flora fossil en las inmediaciones de la Peninsula
Ardley, Isla 25 de Mayo, Islas Shetland del Sur. Buenos Aires, 1963. 17
p. (Inst.
Antarct.
Argent. Contrib.; N79).
Parrish J.T., Curtis R.L. Atmospheric circulation, uppwelling, and organic-rich
rocks in the Mesozoic and Cenozoic eres // Palaeogeogr., Palaeoclimatol.,
Palaeoecol. 1982. Vol.40. P. 31-66.
Pole M. Early Eocene coastal vegetation, Regatta Point, Tasmania // O.F.P.
Inform. 1992. N Special 16-B: IV Conf. Organis. Intern, de Palaeobot. P.
124.
Popov S.V., Akhmetiev M.A., Bugrova E.M. et al. Biogeography of
the Northern Peri-Tethys from the Late Eocene to the Early Miocene. 1.
Early Eocene
// Paleontol. J. 2001. Vol. 35, suppl. l.P. 1-68.
Popov S.V., Akhmetiev M.A., Bugrova E.M. et al. Biogeography of
the Northern Peri-Tethys from the Late Eocene to the Early Miocene. 2.
Early Oligocene
// Ibid. 2002. Vol. 36, suppl. 3. P. 185-259.
Romero E.J. Paleoecologia у paleofitogeografia de las tafofloras
del Cenofitico de Argentina у areas vecinas // Ameghiniana. 1978. Vol.
15. P. 209-227.
Romero E.J. Paleogene phytogeography and climatology of South
America // Ann. Missouri Bot. Gard. 1986. Vol. 73. P. 449-161.
Shepard G. The geology of South Western Ecuador, Muby. L., 1937. 121 p.
Torres T.G. Tertiary paleobotanical study in King George Island, Antarctiva
// O.F.P. Inform. 1992. N Special 16-B: IV Con. Organis. Intern, de Palaeobot.
P. 167.
Truswell E.M. Australian rainforest: The 100 million year record
// Australian tropical rainforest // Ed. L.J. Webb, J. Kikkawa. Melbourne:
CSIRO, 1993.
P. 7-22.
Van Beuskom C.F. Revision of Meliosma (Sabiaceae) section Lorenzanea excepted,
living and fossil, geography and phylogeny // Blumea. 1971. N 19. P. 355-529.
Wilf P., Johnson K., Cuneo R. et al. Fossil plants, climatic indications,
and early Eocene age of en exceptional continental deposits from Southern
South
America // Climate and biota of the Early Paleogene: Intern, mett. Wash.
(D.C.): Smithsonian Inst., 2001. Abstr. vol. P. 101.
Wolfe J.A. Tertiary climatic fluctuations and methods of analysis
of Tertiary floras // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1971.
Vol. 9. P. 27-57.
Wolfe J.A. An interpretation of Alaskian Tertiary floras // Floristics and
paleofloristics of Asia and Eastern North America // Ed. A. Graham. Amsterdam:
Elsevier, 1972.
P. 201-233.
Wolfe J.A. Distribution of major vegetational types during the Tertiary
// Geophys. Monogr. 1985. N 32. P. 357-375.
Wolfe J.A. A method of obtaining climatic parameters from leaf
assemblages // Bull. US. Geol. Surv. 1993. N 2040. P. 1-71.
Wolfe J.A. Tertiary climatic changes at middle latitudes of western North
America // Palaeogeogr., Palaeoclimatol., Palaeoecol. 1994. Vol. 108. P.
195-205.
Wolfe J.A. Paleoclimatic estimates from Tertiary leaf assemblages // Annu.
Rev. Earth and Planer. Sci. 1995. Vol. 23. P. 119-142.
Wolfe J., Dilcher D. Late Paleocene through Middle Eocene climate in Lowland
North America // Climate and biota of the Early Paleogene: Intern, meet.
Wash. (D.C.): Smithsonian Inst., 2001. Abstr. vol. P. 102-103.
Wolfe J.A., Forest C.E., Molnar P. Paleobotanical evidence of
Eocene and Oligocene paleoaltitudes in midlatitude Western North America
// Bull Geol.
Soc. Amer.
1998. Vol. 110, N5. P. 664-678.
Zaklinskaja E.D. Palynology of Paleogene clay from DSDP Site 368, Cabe
Verde rise // Init. Rep. DSDP. 1978. Vol. 41. P. 933-937.
Zjang Jihui. Discovery of old Tertiary flora from Pan Xian of Guizhou and
its significance // Pap. Stratigr. Palaeontol. Guizhou. 1983. N 1. P. 133-141.
In Chinese with English summary.
Введение (Н.М. Чумаков).......................................................................................................................................
5
Часть I
ЭПОХА ПЕРЕХОДА ОТ ТЁПЛОЙ К ХОЛОДНОЙ БИОСФЕРЕ: КЛИМАТ ПАЛЕОГЕНА
Глава 1. Климат Земного шара в палеоцене
и эоцене по данным палеоботаники (М.Л. Ахметьев)............. 10
Часть II
КЛИМАТ И ПЕРЕСТРОЙКИ В ТЁПЛОЙ БИОСФЕРЕ (МЕЛ, ЮРА)
Глава 2. Общий обзор позднемезозойского
климата и событий (Н.М. Чумаков)............................................
44
Глава 3. Палеогеографические
перестройки и седиментация мелового периода
(М.А. Жарков, И.О. Мурдмаа, Н.И. Филатова)....................................................................................................
52
Глава 4. Количественные палеоботанические
данные о позднемеловом климате
Евразии и Аляски (А.Б. Герман)............................................................................................................................
88
Глава 5. Климатическая зональность
и климат мелового периода (Н.М. Чумаков).........................................
105
Глава 6. Глобальная экспансия
планктонных фораминифер: триас, юра, мел
(К.И. Кузнецова, О.А. Корчагин)..........................................................................................................................
124
Глава 7. Динамика и возможные
причины климатических изменений в позднем мезозое
(Н.М. Чумаков)........................................................................................................................................................
149
Часть III
ЭПОХА ПЕРЕХОДА ОТ ХОЛОДНОЙ К ТЁПЛОЙ БИОСФЕРЕ (ПЕРМЬ И РАННИЙ ТРИАС)
Глава 8. Палеогеографические перестройки
и обстановки седиментации в перми и раннем
триасе (М.Л. Жарков)..............................................................................................................................................
158
Глава 9. Палеобиогеография пермских
фузулинид (Э.Я. Левен)........................................................................
181
Глава 10. Палеофитогеография
пермского периода (С.В. Наугольных).............................................................
194
Глава 11. Палеопочвы перми и
раннего триаса (С.В. Наугольных)...................................................................
221
Глава 12. Климат и климатическая
зональность перми и раннего триаса (Н.М. Чумаков).............................
230
Часть IV
ГЛАВНЫЕ КЛИМАТИЧЕСКИЕ И БИОСФЕРНЫЕ СОБЫТИЯ ПОЗДНЕГО ДОКЕМБРИЯ
Глава 13. Ледниковый и безледниковый
климат в докембрии (Н.М. Чумаков)...............................................
259
Глава 14. Проблема климатической
зональности в позднем докембрии. Климат и биосферные
события (Н.М. Чумаков, В.Н. Сергеев).................................................................................................................
271
Заключение (Н.М.
Чумаков)................................................................................................................................
290
Conclusion...............................................................................................................................................................
296