| Главная | Библиотека сайта | Форум | Гостевая книга |
Джордж Пойнар, мл. и Роберта Пойнар
Кто кусал динозавров?

Кто кусал
динозавров?
Насекомые, болезни и смерть в меловой период
ДЖОРДЖ ПОЙНАР, мл.
и РОБЕРТА ПОЙНАР
С ФОТОГРАФИЯМИ И РИСУНКАМИ
АВТОРОВ
Перевод Павла Волкова
ИЗДАТЕЛЬСТВО ПРИНСТОНСКОГО УНИВЕРСИТЕТА
ПРИНСТОН И ОКСФОРД
Copyright © 2008 by Princeton University Press
Опубликовано Издательством Принстонского Университета, 41 William Street, Princeton, New Jersey 08540
В Великобритании: Издательство Принстонского Университета, 3 Market Place, Woodstock, Oxfordshire OX20 1SYAll Rights Reserved
Библиографическая запись Библиотеки Конгресса США
Пойнар, Джордж О.
Кто кусал динозавров?: насекомые, болезнь и смерть в меловой период/
Джордж Пойнар, мл. и Роберта Пойнар; с фотографиями и рисунками авторов.
p. cm.Включает библиографические ссылки и предметный указатель.
ISBN-13: 978-0-691-12431-5 (щел. бумага)1. Палеоэкология-Кайнозой 2. Болезни динозавров. 3. Палеоэкология.
4. Насекомые, ископаемые. 5. Экология насекомых I. Пойнар, Роберта. II. Название.QE720. P65 2007
560_.45-dc22 2007061024Библиографическая запись Британской Библиотеки доступна
Эта книга была составлена в Палатино
Напечатана на бумаге без кислоты.
press.princeton.eduНапечатано в Соединённых Штатах Америки
10 9 8 7 6 5 4 3 2 1
Эта книга посвящается
вопрошающим умам
грядущих поколений.
Натуралистами открыты
У паразитов паразиты.
И произвел переполох
Тот факт, что блохи есть у блох,
И обнаружил микроскоп,
Что на клопе бывает клоп,
Питающийся паразитом,
На нем другой, ad infinitum...*
- Джонатан Свифт (1667-1745)
* Перевод С. Я. Маршака.
vii
| Предисловие | ix | |
| Благодарности | xi | |
| Введение | 1 | |
| 1. | Окаменелости: капсула времени | 7 |
| 2. | Меловой период: эпоха перемен | 17 |
| 3. | Травоядность | 37 |
| 4. | Динозавры, конкурирующие с насекомыми | 50 |
| 5. | Кто «изобрёл» цветковые растения – динозавры или насекомые? | 55 |
| 6. | Опыление | 57 |
| 7. | Инфекционное увядание и другие болезни меловых растений | 63 |
| 8. | Меловой период: время химер и других странностей | 72 |
| 9. | Санитарные службы мелового периода | 79 |
| 10. | Явление энтомофагии среди динозавров | 91 |
| 11. | Пожирающие динозавров | 102 |
| 12. | Мокрецы | 110 |
| 13. | Москиты | 116 |
| 14. | Комары | 122 |
| 15. | Мошки | 127 |
| 16. | Слепни и златоглазики | 131 |
| 17. | Блохи и вши | 135 |
viii
| 18. | 141 | |
| 19. | Паразитические черви | 147 |
| 20. | Открытие болезней мелового периода | 157 |
| 21. | Болезни и эволюция их возбудителей | 171 |
| 22. | Насекомые: абсолютные чемпионы по выживанию | 185 |
| 23. | Вымирание и мел-палеогеновая граница | 192 |
| ПРИЛОЖЕНИЕ A: Насекомые мелового периода | 203 | |
| ПРИЛОЖЕНИЕ B: Ключевые факторы, способствующие выживанию наземных животных | 219 | |
| ПРИЛОЖЕНИЕ C: Проблемы, возникающие при оценке летописи окаменелостей и вымирания | 221 | |
| Ссылки | 225 | |
| Предметный указатель | 253 |
ix
Сто миллионов лет назад Землёй правили динозавры... вот только
так ли это? В действительности же существовали миллионы крохотных животных,
которые не боялись этих могучих рептилий-великанов и не были обеспокоены их
ужасающим царствованием, потому что фактически выслеживали в качестве добычи
их самих.
Орды агрессивных кусачих насекомых нападали на величественного Tyrannosaurus
rex почти так же, как они досаждают людям в наши дни.
На протяжении мелового периода популяции насекомых, свободные от того воздействия
инсектицидов, какому они подвергаются сегодня, процветали и, несомненно, составляли
большую часть видового разнообразия и биомассы животных на земле. Наш мир, в
котором существует почти миллион описанных видов насекомых и, возможно, втрое
больше пока ещё неизвестных науке1 –
это, несомненно, мир насекомых. Просто представьте себе, как всё выглядело 100
миллионов лет назад, когда разнообразие насекомых было ещё больше, и подумайте,
что, возможно, всего лишь возможно, миром правили именно насекомые. И если вы
ещё не убедились в абсолютном превосходстве насекомых над динозаврами, просто
подумайте над таким фактом: насекомые существовали по всему миру до, во время
и после эпохи господства не-птичьих динозавров.
Нам хотелось бы взять вас в путешествие во времени, чтобы исследовать мир динозавров
и узнать, кто их кусал. Используя ископаемые остатки насекомых мелового периода,
мы будем наглядно представлять вероятные отношения, которые существовали между
насекомыми и динозаврами, и попытаемся предсказать, как они могли воздействовать
на популяции динозавров. Наши интерпретации повадок ископаемых насекомых будут
основываться на поведении и экологии их современных потомков2.
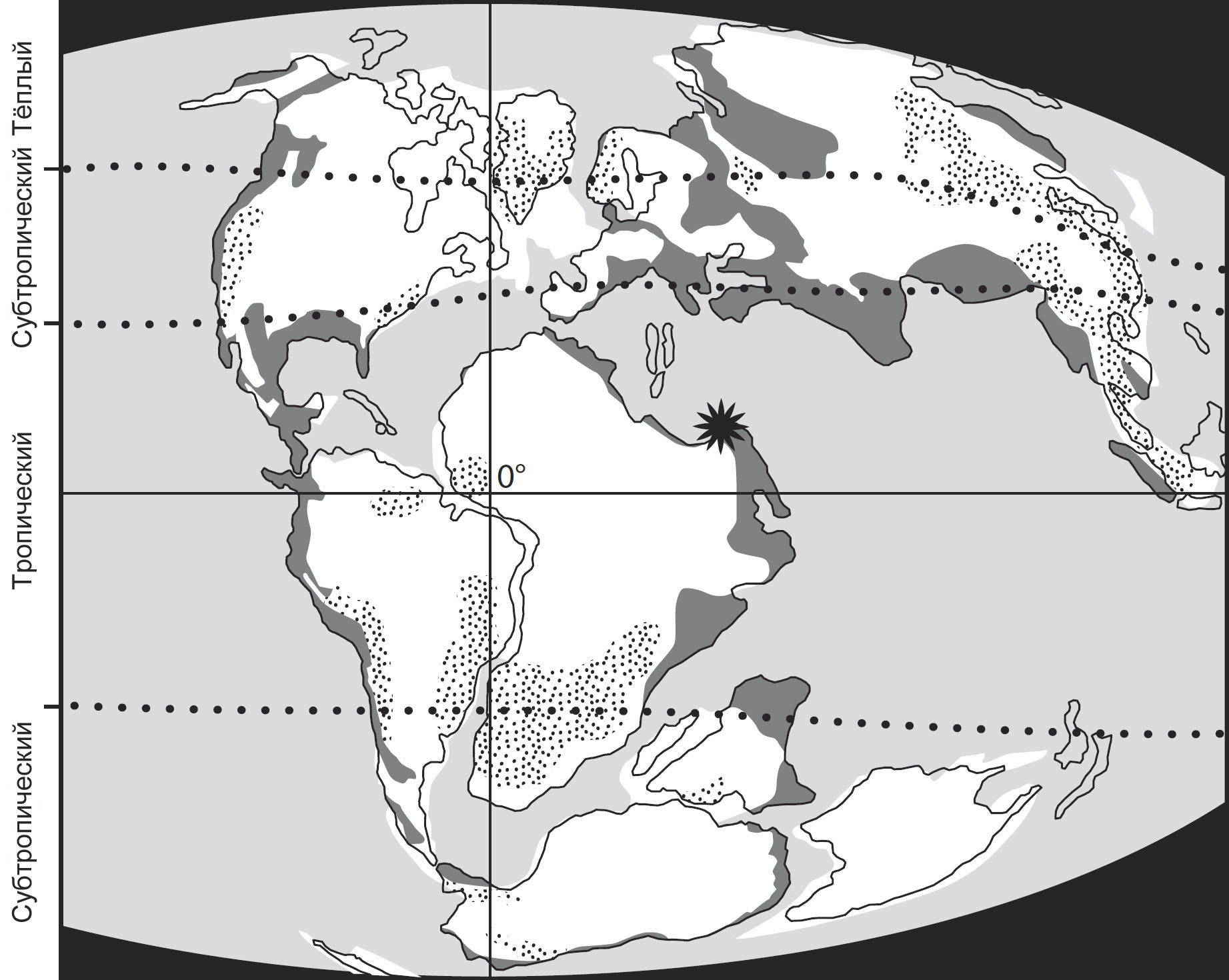
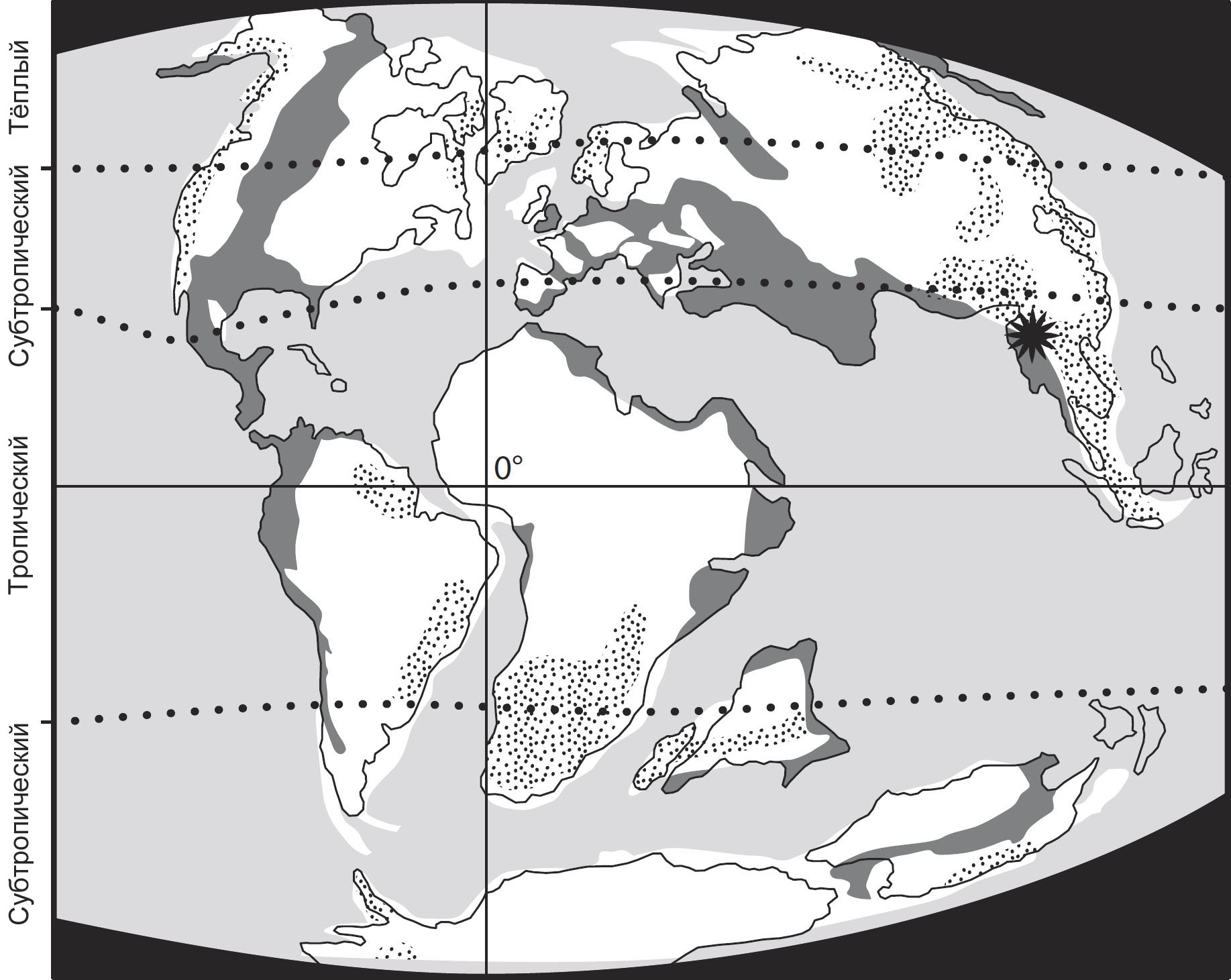
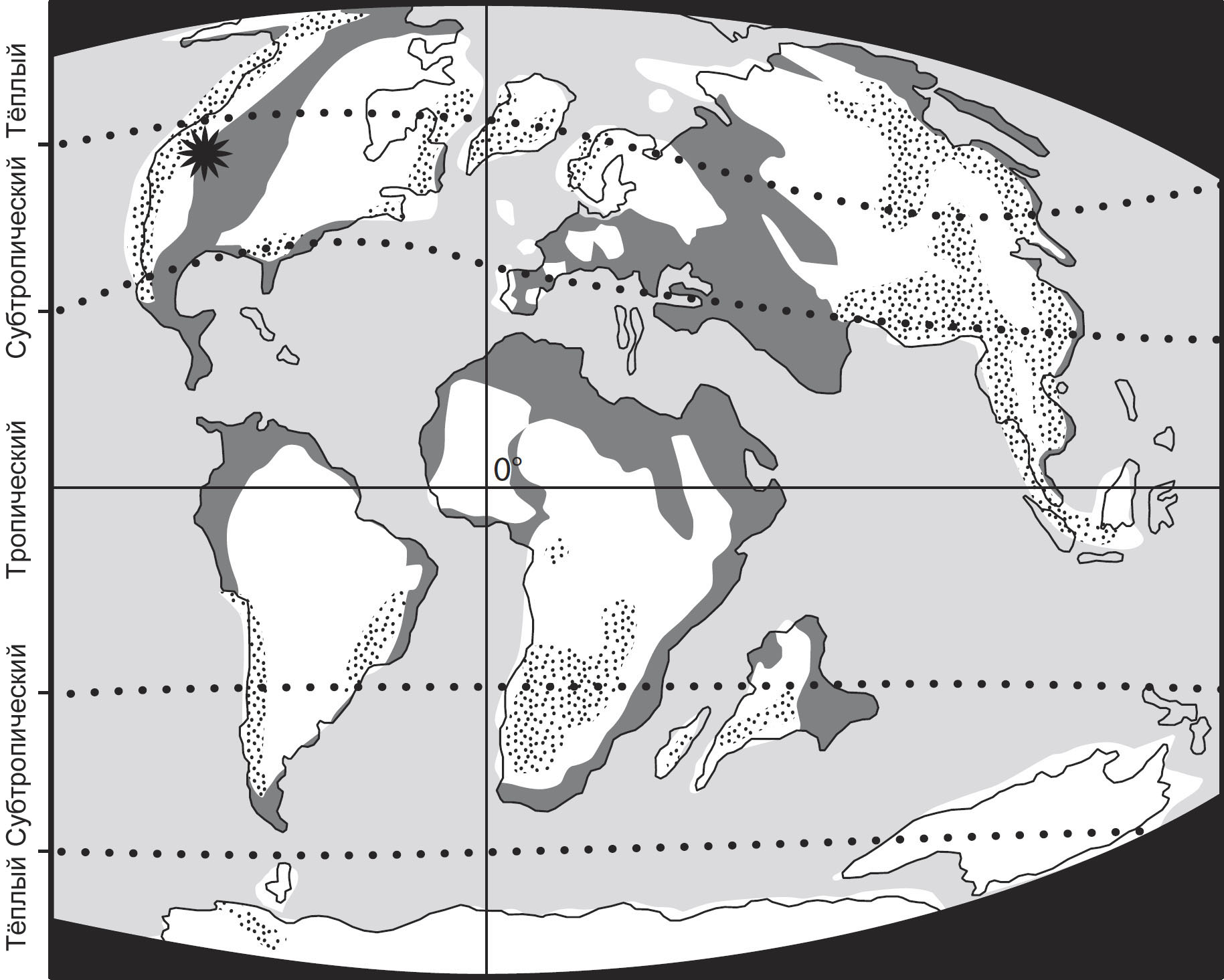
Ключевым моментом этого предприятия будут несколько важных месторождений янтаря,
которые позволяют нам взглянуть на насекомых, формировавших облик природной
среды на протяжении трёх важных периодов: раннемеловые ливанские отложения,
датируемые 130-135 миллионами лет назад (млн. л. н.), бирманские отложения середины
мелового периода, примерно 99-105 млн. л. н., и позднемеловые канадские отложения,
77-79 млн. л. н. Иные
x
мезозойские окаменелости также вносят свой вклад в наше представление
об этом исчезнувшем мире. И в конце мы рассмотрим гипотезу о том, что насекомые,
переносящие болезнетворных агентов, могли внести свой вклад в сокращение численности
популяций динозавров, уже находящихся под угрозой исчезновения, и привели, по
крайней мере, к локальному, и даже к глобальному их вымиранию.
Исследования прошлого часто полны противоречий во взглядах. Эта книга посвящена
не обсуждению нерешённых вопросов, принятию той или иной существующей точки
зрения, или выдвижению новой. Для нашего исследования роли насекомых в меловой
период не имеет значения, были ли динозавры холодно- или теплокровными, или
были на промежуточной стадии, или и теми, и другими. Неважно, были ли некоторые
динозавры предками птиц. И в действительности неважно, были ли некоторые из
окаменелостей неправильно идентифицированы, или оспариваются ли существующие
теории вымирания в конце мелового периода. Это история о том, как наглядно представить
себе древнюю сеть жизни. T. rex и зауроподы, выдернутые из прошлого
и размазанные по большому экрану, не были изолированными объектами. Они были
часть комплекса, который был неразрывно переплетён со всеми иными фаунами и
флорами в их среде обитания, а в действительности же – и со всей планетой. Чтобы
проиллюстрировать это, некоторые главы начинаются сценой, изображающей то, как
мы представляем себе жизнь в меловой период. Они напечатаны иным шрифтом.
Наша идея состоит в том, чтобы изобразить структуру этой сети из прошлого в
общих чертах. Тонкие объединяющие штрихи внесут уже другие люди; что-то из того,
о чём мы говорим, будет сочтено ошибочным и отвергнуто, а что-то окажется подходящим
и останется. В сущности, на холсте набросан всего лишь эскиз для грядущих поколений,
которые будут работать над ним, но маловероятно, что эта картина будет когда-либо
завершена. Сегодняшний мир напоминает непрерывно меняющийся, пульсирующий мегаорганизм,
сложность которого мы едва начали осознавать, хотя человек уже уничтожает его.
Экология мира мелового периода была бы, несомненно, ещё более сложной, чем у
современного мира, а флористическое и фаунистическое разнообразие – ещё большим.
Многие из тех растений и животных не дожили до конца мелового периода. И причина
их вымирания в итоге обнаружится в значительном нарушении глобальной экологии
– в разрыве в сети жизни.
xi
Мы хотели бы поблагодарить за разного рода помощь, оказанную в ходе подготовки этой работы, следующих людей: Джона Айтчисона, Норму Андерсон, Ника Арнольда, Денниса Брамана, Кентона Чамберса, Питера Крэнстона, Брайена Данфорта, Джима Дэвиса, Джейн Грей, Пенни Гуллан, Гранда Хуана, Р. Л. Джейкобсона, Александра Кирейчука, Боба Мейсона, Раифа Милки, Шерил О’Доннел, Теда Пайка, Барри Рота, Джона Рубена, Ричарда Шадзевского, Сэма Телфорда-младшего, Джека Ткача и Чарльза Веллмана. Особую благодарность выражаем Алексу Брауну за помощь в поиске образцов янтаря, Рону Бакли и Скотту Андерсону за предоставленные образцы из их коллекций бирманского янтаря, и Артуру Буко за плодотворное обсуждение. Все образцы, изображённые здесь, включены в коллекцию янтаря Пойнара, находящуюся в Орегонском Университете, за исключением следующих: образец с цветной вкладки* 6A из коллекции Скотта Андерсона; образцы с цветных вкладок 5E, 8A, 12D, 13C и 15A – из коллекции Рона Бакли; образцы с цветных вкладок 2C, 5A, 5B, 6B, 6D, 7C, 9B, 11D, 12B, 13A, 13B и 15B – из Королевского Тиррелловского палеонтологического музея, Альберта, Канада; и цветок на цветной вкладке 14A – из коллекции Гранда Хуана. Темы оформления проекта созданы Грегом Пойнаром.
* Здесь и далее по тексту: цветные вкладки отсутствуют по не зависящим от переводчика обстоятельствам. – прим. перев.
Пустая страница

Пустая страница
1
Оставшиеся в живых члены стада динозавров-орнитопод паслись
на краю хвойного леса мелового периода. Этот сухой сезон был особенно трудным
и долгим. Несколько особей время от времени поедали высохшие на солнце осоки
и хвощи, выросшие близ берега извилистой реки. Река пересохла, превратившись
в тонкий ручеёк, текущий среди сильно подмытых берегов. Болезнь сократила численность
этого стада, некогда насчитывающего сотни голов, до менее чем пятидесяти особей
– явно отсутствовали самые молодые и старые животные. Многие из величественных
животных выглядели апатичными и даже не обращали внимания на стаю хищных динозавров-теропод,
которые следовали за ними. Эти свирепые плотоядные, вооружённые острыми зубами
и серповидными когтями, были жирными и сытыми, потому что больные травоядные
становились лёгкой добычей.
Обычно эти растительноядные динозавры тратили около половины всего времени бодрствования
на пережёвывание нежной растительности, растущей вдоль потоков воды или на открытых
лугах. Однако, потеряв аппетит, заражённые животные не ели уже несколько дней
и с трудом ковыляли из-за истощения. Обычно они избегали прямых лучей полуденного
солнца, но сейчас многие из них неподвижно стояли на самой жаре. Они часто подходили
к краю воды, с трудом наклонялись и подолгу пили, явно забыв об опасности, которая
исходила от затаившихся там крокодилов. Постоянный понос обезвоживал их, и их
жажда была почти неутолимой. Краски окружающего пейзажа нарушались пятнами окрашенных
кровью испражнений, которые привлекали орды мух и жуков.
Один дрожащий орнитопод с сухой кожей, обтягивающей явственно выпирающие рёбра
и позвоночник, повалился набок, и его начало рвать струями слизи с кровавыми
пятнами, в которой извивались поблёскивающие круглые черви. Эта больная особь,
глаза которой сейчас превратились в узкие щёлочки, была слишком истощённой,
чтобы сгонять с себя голодные массы назойливых
2
насекомых, ползающих по тонкой чешуйчатой коже в поисках
мест, где им можно будет начать кормиться. Когда умирающее животное в конце
концов погибло, несколько членов стада подошли к нему и начали подталкивать
его, но ответа не было, и они отошли, уступая дорогу приближающимся тероподам.
Плотоядные начали терзать тушу, не понимая, что едят заражённое мясо и подвергаются
нападению тех же самых насекомых, которые до этого кормились на больных динозаврах.
Некоторые из них, однако, начинали показывать первые признаки заражения и отходили
в сторону от вакханалии кормёжки, чтобы прилечь и отдохнуть, откусив мясо всего
лишь по несколько раз. Пока другие особи пожирали остатки, на них высадился
целый десант различных клещей, не упустивших возможности сменить место жительства
с трупа на шкуру теропод.
Если бы можно было произвести вскрытие трупа этого орнитопода,
оно выявило бы множество паразитов и болезнетворных микроорганизмов, населяющих
ткани.
Некоторые из них, вроде амёбной дизентерии, малярии и круглых червей-аскарид,
вызывали бы появление язв кишечника, абсцессов печени и деформированных кровяных
клеток. Но действительная причина смерти нашего динозавра была бы отмечена как
лейшманиоз – болезнь, вызываемая простейшими. Точно так же, как и другие члены
стада, он был жертвой вновь появившегося болезнетворного организма, который
опустошал мир мелового периода. Примерно 100 миллионов лет назад некоторые из
этих микроорганизмов выработали новые отношения с кровососущими двукрылыми,
в ходе которых некогда безопасные симбионты мух превратились в возбудителей
смертельных болезней. Вступив в невиданный ранее союз, эти переносимые насекомыми
заразные болезни вкупе с уже давно сформировавшимися паразитами превратились
в нечто большее, чем могла выдержать иммунная система динозавров. Опустошительные
эпидемии начали нарушать равновесие между травоядными и плотоядными динозаврами,
существовавшее на протяжении тысячелетий. Снабжённые смертельным оружием, кровососущие
насекомые были верховными хищниками в пищевой цепочке и теперь могли влиять
на судьбу динозавров так же, как сегодня они придают облик нашему миру.
Даже когда оставшиеся члены этого стада пали жертвами болезни, насекомым нашлось
занятие: они обеспечивали распространение эпидемии. Кровососущие мухи, питаясь
кровью ослабленных особей, собирали пато-
3
генные микроорганизмы, чтобы ввести их другим жертвам. Благодаря
своей способности к полёту они могли разносить болезнь и заражать других восприимчивых
к ней динозавров в пределах своего ареала. Мухи, жуки и тараканы, посещающие
заражённые фекалии и трупы, собирали бактерий, простейших и нематод, становясь
затем их переносчиками и заражая других позвоночных. Динозавры, отобедав тараканами,
которые теперь носили яйца аскарид, в итоге зарабатывали поражение желудка.
Если же рассматривать события в большем масштабе, то, когда вспышка болезни
уничтожила травоядных динозавров, оказалось нарушено равновесие в их экосистеме.
Вначале плотоядные, возможно, могли быть в выигрыше, потому что мёртвые и умирающие
животные встречались в изобилии. Обратной стороной этого видимого изобилия было
то, что они также заболевали, как только истощались запасы их пищи. В последующие
месяцы, когда сообщество орнитопод распадётся полностью, им будет угрожать голод.
Сочетание голода и многочисленных заразных болезней ускорит их упадок. Растительность,
которой обычно питались орнитоподы, будет процветать, а вместе с ней и какие-то
травоядные, которые также использовали эти растения в пищу. Численность тех
специализированных видов, выживание которых зависело от орнитопод, снизится.
В конечном счёте те ниши, которые опустели после них, займёт кто-то другой,
и жизнь продолжится. Оправятся ли популяции в дальнейшем, или нет, зависит от
многих факторов, но в те времена, равно как и в наше время, переносимые насекомым
болезни могли поставить на грань вымирания любое животное.
Насекомые воздействуют на мир не только благодаря болезням, которые они передают,
но и неисчислимым множеством иных способов. Они могут быть маленькими существами,
но они – самая разнообразная группа живых организмов, и, вероятно, они были
самой существенной экологической силой на Земле с тех времён, как впервые возникли
примерно 350 миллионов лет назад. Насекомые составляют более 57% видового разнообразия
жизни на Земле и 76% всей животной жизни. В настоящее время существует более
990 000 известных видов, но ещё больше ожидает своего открытия. Сравнение их
количества с млекопитающими, которые составляют 0,35% от всех видов, лишь ещё
сильнее подчеркнёт их всеобъемлющую важность297.
Травоядные насекомые, которые включают 45% от общего числа их видов, составляют
примерно четверть от всех ныне живущих видов4.
Растительноядные формы потребляют значительно большее количество растительных
тканей, чем позвоночные,
4
в любом из исследованных биомов, за исключением травянистых
равнин. Указания со стороны летописи окаменелостей подтверждают, что так было
начиная с того времени, как сложились первые наземные экосистемы5.
И потому можно принять как факт то, что они были серьёзными конкурентами травоядных
динозавров на протяжении мелового периода.
Приблизительно треть всех живых организмов на Земле – это насекомые, которые
по своим пищевым предпочтениям являются хищниками и сапрофагами. И тот факт,
что плотоядные насекомые представляют примерно 20% от числа видов животных в
биоте, уже говорит нам о том, насколько они важны для сохранения контроля над
популяциями членистоногих. Хищные насекомые обычно питаются другими насекомыми,
но некоторые убивают более крупную добычу. Например, известно, что личинки слепней
убивают мелких лягушек, а крупные богомолы способны добывать мелких ящериц,
птиц и неосторожных мышей.
В число плотоядных насекомых также входят паразиты, существа, живущие внутри
или на поверхности тела своих жертв. Изрядное их число питается кровью животных.
В целом это такие существа, которые и очаровывают, и пугают людей. Они также
внушают нам страх, потому что один укус может потенциально привести к смерти.
Кусающие насекомые переносят вирусов, бактерии, простейших и нематод. Одно из
них, малярийный комар, убивает более миллиона людей ежегодно и является главной
причиной смерти детей моложе пяти лет во всём мире. Это означает, что каждый
год примерно один человек из пятидесяти шести умирает именно от этой переносимой
насекомым болезни. Именно поэтому мы неустанно хлопаем, давим, ставим сетки
и брызгаем аэрозолями, пытаясь избежать нападения этой заразы. Однако они всегда
умеют отыскивать нас, потому что у них были целые геологические эпохи для того,
чтобы приспособиться к питанию на всех наземных животных. И они наверняка мучили
динозавров так же нещадно, как и нас самих.
Обычно люди не слишком сильно задумываются о насекомых-сапрофагах – о тех неисчислимых
легионах, которые пожирают мёртвое и отмирающее органическое вещество. Но те
существа, которые поедают экскременты, падаль и детрит, являются важным и необходимым
компонентом нашего мира, составляя примерно 11% от всей биоты. Они – уборщики,
нацеленные на выполнение задачи по избавлению от побочных продуктов жизни, и
они перерабатывают отходы с удивительной эффективностью. Насекомые-сапрофаги
всегда были неотъемлемой частью любой экосистемы, и, несомненно, в меловой период
были так же важны, как и в наши дни.
5
Хотя насекомые питаются растениями и другими животными, они
и сами по себе являются важным источником пищи в пищевой цепочке. Мелкие и многочисленные,
они встречаются во всех наземных местообитаниях. Представляя собой удобно упакованный
белок и другие необходимые питательные вещества, которые уже готовы к употреблению
и сравнительно легко доступны, насекомые используются в пищу множеством живых
существ. В меловой период они могли быть важным компонентом рациона как для
молодых динозавров, так и для мелких животных, которыми питались динозавры.
Крохотные, но могущественные насекомые оказали огромное воздействие на всю экологию
Земли, несомненно определяя пути эволюции и вызывая вымирание наземных организмов.
Крупнейшие из наземных животных, динозавры, схлестнулись с ними в битве не на
жизнь, а на смерть ради выживания. Подробности этого состязания можно получить
из летописи окаменелостей. Ископаемые существа, интерпретируемые путём сравнения
с их современными аналогами2, рассказывают
нам о том, как насекомые могли оказывать воздействие на динозавров и на весь
мир мелового периода. Их сохранившиеся останки – это основной момент для путешествия,
которое уведёт нас в прошлое и откроет новые аспекты экологии и упадка динозавров.
Окаменелости будут теми ключами, которыми мы воспользуемся, чтобы отпереть дверь,
ведущую в тайны мелового периода.
6
Пустая страница
7
Вскоре после того, как хищники закончили пировать на трупе
больного орнитопода, упали первые капли дождя приближающейся бури. Многие из
животных прервали свою деятельность, приветствуя ветер, несущий прохладу, а
когда дождь полился в полную силу, некоторые из них бросились на поиски укрытий.
Кровососущие мухи, насыщавшиеся на телах ослабленных болезнью орнитопод, искали
защиты от надвигающейся бури и летали среди ветвей высоких деревьев араукарии
в ближайшем лесу. Несколько особей слегка задели поблёскивающие натёки смолы
и прилипли. Они отчаянно пробовали освободиться, но борьба лишь глубже затягивала
их в золотую глубину, и в конце концов ещё один поток смолы полностью покрыл
их и запечатал в янтарной могиле.
Тем временем в раскинувшейся неподалёку пойме небольшое стадо больных динозавров
продолжало вяло пастись, объедая сухие пучки растительности, покрывающие берега
внешне спокойной реки, и пребывая в блаженном неведении того, что выше по течению
вода, выпавшая в виде дождя во время бури, мчалась вниз по оврагам и ущельям
и быстро поднимала уровень реки, порождая потоп. Течение начало усиливаться,
и несколько больных орнитопод на маленьком островке посреди реки внезапно осознали,
что вода уже плещется вокруг их ног, а её уровень быстро повышается. В потоке,
который теперь стал очень бурным, добраться до берега не было возможности, и
из-за того, что они были ослаблены, течение снесло их и утопило. Их тела вынесло
ниже по течению к излучине реки, куда они приплыли, кружась в воде, и опустились
на дно. Листья, сорванные нисходящими потоками воздуха и подхваченные ветром,
попали в реку и утонули вокруг их тел. Ливень закончился так же быстро, как
и начался.
8
Илистая вода потекла медленнее, и слой осадка отложился поверх тел динозавров и листьев и веток, снесённых в мусор. Солнце показалось на небе и высушило капли дождя, а река вернулась в своё обычное русло. И вновь звуки насекомых наполнили воздух; жизнь продолжалась.
Эти события становятся известными через миллионы лет, когда
их жертвы покажутся нам в виде окаменелостей, застывших в горных породах и погребённых
в янтаре из леса вымерших араукарий. Именно по таким окаменелостям мы будем
пытаться распутать историю борьбы, ужаса и болезни мелового периода, где сплелись
насекомые, динозавры и растения, которыми они питались.
Многие люди спрашивали нас о том, почему нас так необычайно чаруют ископаемые
остатки организмов, особенно те, что встречаются в янтаре. Ответ на этот вопрос
сложен, но можно назвать несколько причин. Окаменелости интригуют, потому что
они помещают время существования человека в перспективу. Мало кто из нас хотя
бы иногда задумывался над первобытным прошлым, потому что мы проживаем свою
жизнь, привязанную к настоящему моменту, измеряя время часами и календарём.
Окаменелости напоминают нам о несущественности нашего собственного эфемерного
времени. Насколько важным является Homo sapiens, если наш вид украшает
собой эту планету лишь немногим более 200 000 лет – бесконечно малая вспышка
в хронологии жизни в целом? Существа, которых мы изучаем и о которых пишем,
жили в течение мелового периода – обширного отрезка времени, который начался
около 145,5 млн. лет назад и закончился около 65,5 млн. л. н.6
(рис. 1). Простое осознание продолжительности этого временного интервала освобождает
нас от довольно ограниченного мнения о нашей собственной важности.
Окаменелости – это редкость, которая дошла до нас вопреки непреодолимым трудностям,
и уже одно лишь это придаёт ей очарование. Получить возможность дотронуться
до кости огромного хищника, который жил и здравствовал 100 миллионов лет назад,
или заглянуть в кишечник насекомого, которое, возможно, кормилось на том самом
динозавре – это большая честь. Динозавры существовали на протяжении удивительных
165 миллионов лет. Но сколько их жило и умерло за это время – триллионы? Любое
число, о котором мы подумаем, будет всего лишь приблизительным.
Хотя обнаружены уже десятки тысяч фрагментов костей и зубов динозавров, составлено
довольно мало сочленённых
9
 |
|
| Рисунок 1. Геохронологическая шкала, показывающая меловой период, его подразделения и века, а также их примерный возраст в миллионах лет назад6. Пунктирные полосы показывают возрастные оценки для ливанских (L), бирманских (B) и канадских (C) отложений янтаря. |
скелетов, и просто коснуться одного из них – это ни с чем не
сравнимый дар. В 2000 году эксперт по динозаврам Дейл Расселл7
прокомментировал это так: «хотя палеонтологи собирают останки мезозойских животных
в течение более чем столетия, общее количество известных фрагментов скелетов
динозавров – всего лишь около 5 000». С того момента количество целых или частичных
скелетов, конечно же, возросло, но стоит ли удивляться тому, что за проданную
на аукционе «Сью», скелет T. rex, было выручено почти 8,4 млн. долларов – такова
окончательная сумма сделки8.
Хотя эти причины превращают работу с редкими окаменелостями в поистине замечательный
опыт, большая часть их обаяния заключена в
10
историях, которые они рассказывают о прошлом. Думая об ископаемых
насекомых, многие люди естественным образом представляют себе янтарь, хотя существует
также некоторое количество замечательных отпечатков на камнях. Для нас янтарь
– это коронная драгоценность среди всех окаменелостей из-за уникального, напоминающего
прижизненный, облика погребённых в нём живых существ и из-за его способности
захватывать чрезвычайно мелкие и хрупкие объекты вроде паутины и микробов (рис.
2). Янтарь также превращает насекомых в драгоценности потрясающей красоты. Увеличенные
микроскопом и озарённые сиянием янтаря, спины жуков с металлическим блеском,
украшенные бороздками и испещрённые ямками, превратились в скульптуры. Составленные
из бесчисленных фасеток глаза слепней с радужным блеском вечно таращатся в окружающую
смолу, и можно лишь представлять себе, какие сцены они видели миллионы лет назад.
Янтарь часто сохраняет последние муки жизни вроде самок насекомых, отчаянно
пытающихся дать жизнь будущим поколениям, откладывая яйца в своих янтарных гробах.
Но самые острые ощущения от работы с ископаемыми остатками в янтаре таит в себе
тот факт, что в каждом кусочке янтаря потенциально заключены тайны, ожидающие
своего открытия. Виктор Гюго писал: «Где кончается телескоп, там начинается
микроскоп. У кого из них поле зрения больше?»* Так что вам можно и не быть капитаном
звездолёта, чтобы «исследовать странные новые миры» или «пройти там, где ещё
не ступал ни один человек». Вам всего лишь нужно быть биологом с микроскопом
и набором образцов янтаря. И рано или поздно в одном из кусочков найдётся сюрприз.
Возможно, там обнаружится новый вид, род, или даже семейство организмов, ранее
неизвестных науке. Это может быть первое появление в летописи окаменелостей
животного или растения вроде самой древней пчелы или травы, или даже странное
химерическое существо, которое одновременно и заключает в себе тайну, и ставит
в тупик. Эти открытия – своего рода «джекпот» в науке, который ждёт палеонтолога,
словно выигрышный билет в лотерее, и они заставляют вспомнить одну из наших
любимых историй, в которой задействованы торговец янтарём и янтарный кулон,
который надела на вечер ничего не подозревающая женщина. Поскольку любой истинный
любитель янтаря в любое время желает заглянуть в любой кусочек янтаря, чтобы
увидеть, какие сокровища заключены внутри него, для него было вполне естественным
изучить кулон. То, что он увидел, было удивительно: две блохи. А разве мы не
сказали, что ископаемые блохи чрезвычайно редки, а здесь их было две в одном
куске янтаря? В итоге кулон оказался в музее! Это
* В. Гюго «Отверженные». – прим. перев.
11
|
12
просто говорит нам о том, что любая драгоценность из янтаря,
в том числе какая-нибудь бусинка в ожерелье, может содержать редкие научные
сокровища.
Можете ли вы поверить в то, что этот тёплый драгоценный камень органического
происхождения начал своё существование как смола, медленно сочащаяся из коры
древнего дерева миллионы лет назад? Янтарь – это сохранившаяся в ископаемом
состоянии смола, но процессы, которые превратили её в янтарь, весьма отличаются
от тех, которые сохранили насекомых, попавших внутрь неё. Поэтому янтарь – это
поистине универсальный склад окаменелостей, сохранивший для нас не только свидетельство
существования гигантских деревьев, из которых он появился, но также и существ
из той древней окружающей среды. После миллионов лет, проведённых в крепких
объятиях земли, или даже будучи погружённым в море, янтарь мелового периода
обнаружился в различных отложениях по всему миру. Похоже, весь янтарь того времени
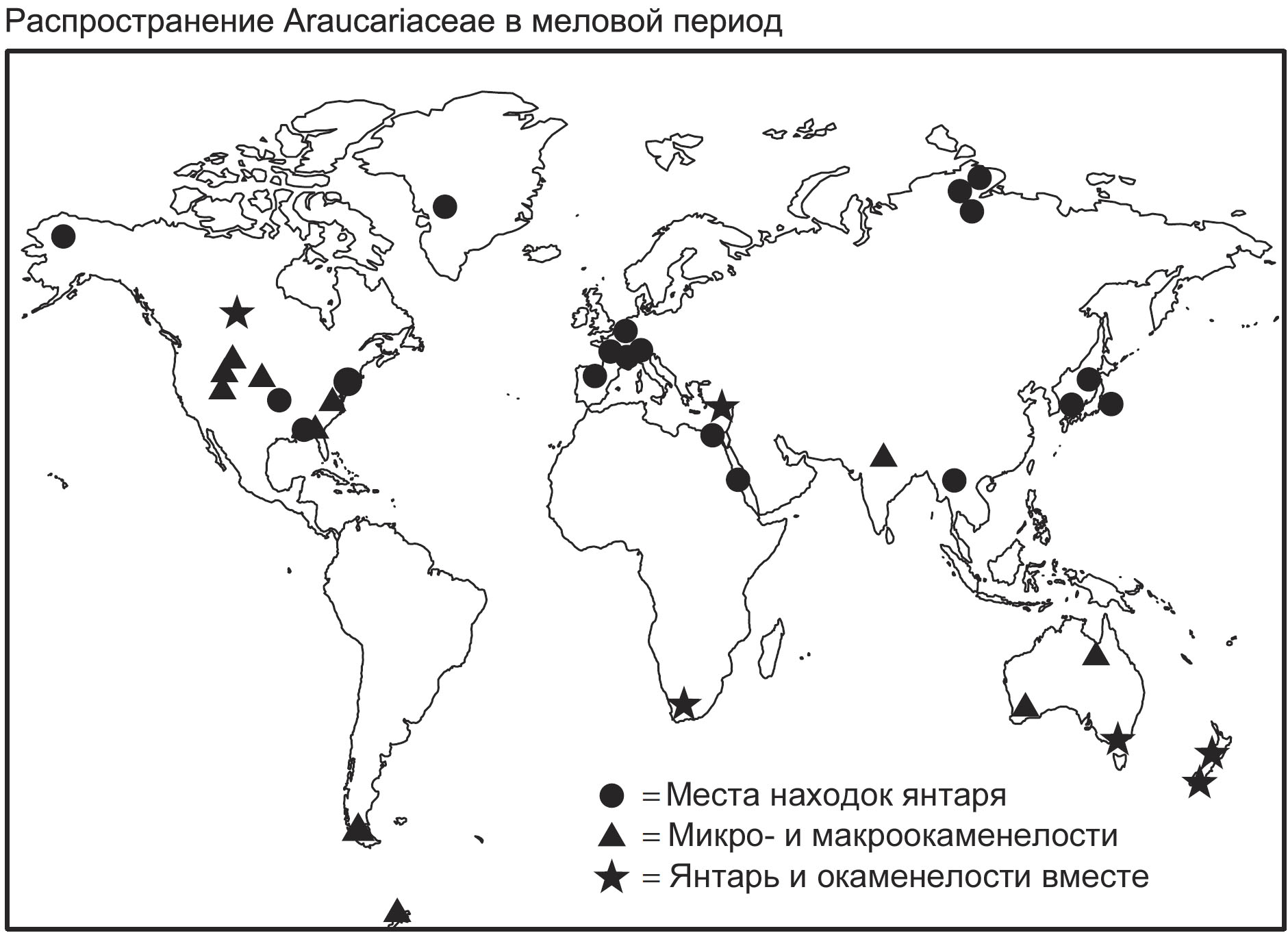
образовался благодаря древней группе хвойных деревьев – араукариевым (рис. 3).
В меловой период эти деревья были распространены всесветно, что видно по их
ископаемым остаткам (рис. 4), хотя сейчас они ограничены несколькими популяциями
в южном полушарии344.
В некоторых местах янтарь выкапывают из глубинных слоёв почвы, но в Южной Альберте,
Канада, меловой янтарь рассыпан по пустынной прерии, где ветер и дождь разрушали
почву и выпускали это сокровище из заточения в угле и сланце. Когда мы совершили
путешествие на те равнины в западной Канаде с Тэдом Пайком, в дальнейшем учившимся
в докторантуре в Университете Калгари, мы помогли собрать янтарные самородки,
рассеянные среди фрагментов покрытых лишайником костей динозавров. Размером
они были от слезинки до грецкого ореха, и многие из них были покрыты тёмной
окисленной корочкой.
Когда смотришь на них, лежащие рядом друг с другом, контраст между янтарём и
другими окаменелостями становится очевидным (рис. 5). В отличие от целых насекомых,
заключённых в янтаре, от динозавров нам остаются для истолкования лишь кости,
зубы, когти, немного яиц, копролиты и следы. Нам пришлось бы проехать лишь несколько
миль на север к Провинциальному Парку Дайносор и известному Тиррелловскому палеонтологическому
музею, чтобы увидеть одну из самых больших в мире коллекций ископаемых остатков
позвоночных мелового периода. Они образовались после того, как вода занесла
грязью или песком их останки, их пропитали растворённые в воде минеральные вещества,
13
|
14
|
и в конечном счёте они превратились в камень. Очень редко, когда осадочный
покров на динозавре был особенно тонкозернистыми, отпечатки демонстрируют
рельеф шкуры.
Ископаемые остатки растений привлекательны с несколько иной стороны, нежели
янтарь или кости. Если янтарь следует отполировать, чтобы он мог показать
скрытые сокровища, а остатки динозавров обычно нужно кропотливо выкапывать
и перевозить в лабораторию для исследования, то растения во всём их великолепии
можно лицезреть прямо на месте раскопок – хотя это тоже вовсе не лёгкая работа.
Мы тратили многие часы, взбираясь по горячей, скользкой поверхности утёсов,
покрытой беспорядочно разбросанными валунами. Пусть мы обливались потом и
должны были постоянно оставаться настороже из-за прячущихся гремучих змей,
но прекрасный образец породы мог сделать охоту стоящей затраченных усилий.
Поставленная на ребро и расколотая, плита песчаника идеально продемонстрирует
два отпечатка растения: один, показывающий верхнюю, а другой нижнюю поверхность
листьев. Иногда некоторые из этих спрессованных
15
|
16
окаменелостей могут сохранять участки зелёного пигмента или
даже свидетельства их повреждения насекомыми. Испытываешь восхитительное чувство,
освобождая листья из этой каменной могилы после того, как прошли миллионы лет.
Не только листья, но и окаменелая древесина, деревья, шишки и даже пыльца могут
рассказать о лесах прошлого. Окаменелый лес из формации Чинле в Аризоне включает
минерализованные пни араукарий и даже маленькие кусочки янтаря. Поездка туда
дала нам представление об огромных размерах смолоносных деревьев мезозоя. На
нас оказали громадное впечатление массивные брёвна длиной до 200 футов и диаметром
10 футов. Их состояние указывает на то, что они переносились на какое-то расстояние
по рекам перед тем, как в итоге оказались захороненными и окаменели. Эти брёвна
невероятно красивы: все их ткани были замещены разными разновидностями кварца,
в том числе аметистом и розовым кварцем. Доминирующим цветом, однако, являются
яшмовый и переходные оттенки от красных до жёлтых и коричневых. Хотя эти деревья
араукарии – впечатляющего вида макроокаменелости, микроокаменелости растений
вроде пыльцы и спор, пусть и не столь драматично выглядящие, зачастую могут
снабдить значительно большим количеством информации о типах растений, существовавших
в прошлом.
Это возвращает нас к одной из причин того, почему окаменелости настолько очаровывают
нас: каждая из них может рассказать историю. Исходя из их анатомии, мы можем
получить крупицы информации об образе жизни и средах обитания. На основе собраний
окаменелостей мы можем делать выводы о взаимодействии животных и растений, об
экологии и климате тех прошедших эпох, и если рассматривать их все вместе, о
том, на что была похожа жизнь миллионы лет назад, в течение конкретной эпохи
мелового периода. Вот то, что мы надеемся вам показать: картину жизни от 145,5
до 65,5 миллионов лет назад, когда насекомые и динозавры конкурировали за доступные
пищевые ресурсы, кормились друг другом и страдали от паразитов и вновь появившихся
в результате эволюции болезней. И в первую очередь нам следует объяснить, как
физические и биологические перемены на протяжении мелового периода влияли на
то, как насекомые кусали динозавров.
17
На протяжении мелового периода солнце восходило и заходило более 29 миллиардов раз. Каждые новые рассвет и сумерки видели рождение и смерть миллиардов живых организмов, и в течение каждого проходящего тысячелетии возникал или вымирал биологический вид. На протяжении этого периода истории планеты драматические физические и биологические изменения определяли пути эволюции насекомых, растений и динозавров. Различия между таксонами насекомых явно заметны на заключенных в янтаре ископаемых остатках, найденных в раннемеловых отложениях Ливана, отложениях середины мела в Бирме, и в позднемеловых отложениях Канады. Другие отложения, содержащие ископаемые остатки, говорят нам о том, что подобные же изменения также происходили у динозавров и растений.
Географические и климатические изменения
В течение этих 25-30 миллионов лет, отделяющих одно янтароносное
местонахождение от другого, континенты дрейфовали на сотни миль, формировались
горы, менялся уровень моря, извергались и гасли вулканы, смещались климатические
пояса, а землетрясения раздирали сушу. Все эти физические преобразования в облике
Земли были последствиями явления, известного как тектоника плит.
Геологи достигли больших успехов, расшифровывая тайны истории планеты путём
исследования тектоники плит. Они установили, что поверхность Земли сложена из
комплекса континентальных и океанических плит, которые постоянно расходятся
в стороны в некоторых областях, и сталкиваются вместе в других, и потому постоянно
скользят и ползут над, под, или навстречу друг другу. Даже сегодня Земля продолжает
изменяться, и в самых активных областях движения плит землетрясения и вулка-
18
19
|
Нам известно, что геологические изменения на протяжении мелового периода делали площадь поверхности суши всё меньше и меньше по мере того, как разделялись континенты и поднимался уровень моря. По оценке команды учёных, с начала и почти да самого конца мелового периода по всему миру имела место чистая потеря приблизительно 14% неморских областей, и потому 80 миллионов лет назад в мире было меньше всего поднимающейся над океаном суши по состоянию на последние 245 миллионов лет, и на удивительные 28% меньше, чем было 5 млн. лет назад.9
20
|
Климат – это ещё один физический фактор, который оказывал влияние на образ жизни представителей биоты прошлого. Изменения климата происходят по многим причинам, но размер и топография массивов суши наряду с характером ветров и океанскими течениями оказывают на градиент солнечной энергии, простирающийся от экватора до полюсов примерно такое же влияние, как озоновые слои и положение континентов. Поскольку возможно установить корреляцию большинства этих факторов с изменениями, вызванными тектоникой плит, из этого следует, что на протяжении мелового периода имели место некоторые колебания климата. В целом, однако, меловой период, как полагают, был значи-
21
|
тельно теплее, чем в наши дни, и маловероятно, что на протяжении большей части тех 80 миллионов лет имели место температуры ниже нуля. В арсенале различных дисциплин имеются методы получения информации о климатических условиях прошлого, вроде выявления распределения чувствительных к климатическим условиям минеральных отложений на поверхности земли или исследования распределения биот прошлого, показывающих специфические температурные предпочтения. Поскольку наши три местонахождения янтаря были обнаружены в угольных слоях, мы можем сделать вывод о том, что связанные с ними леса процветали в среде, условия в которой могли быть от сырых до влажных. Мы также знаем, что деревья араукарии,
22
Биотические изменения
Когда свет раннего утра начал разливаться по раннемеловым небесам Ливана,
ночные существа принялись искать укрытия.
Охотники и собиратели заползали под листья, забивались в трещины и пещерки,
поднимались на вершины деревьев или торопливо зарывались в землю. В лесу ненадолго
воцарился покой. Разгорающийся огонь восхода осветил почти чуждый растительный
мир, весьма сильно отличающийся от того, который существует в наши дни.
23
Архаичные растения, которые покрывали землю в те времена, представляли
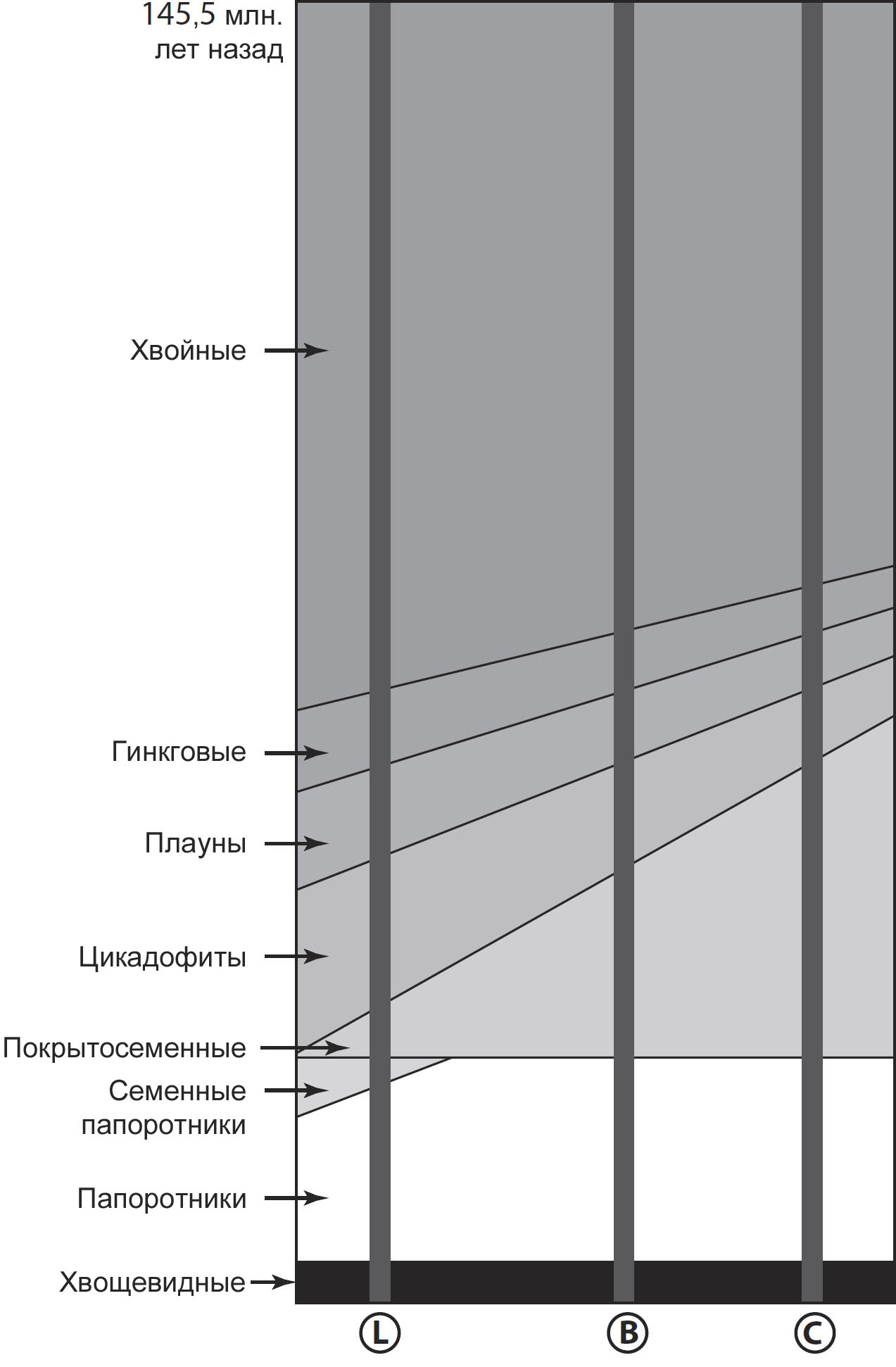
собой преимущественно голосеменные вроде хвойных, гинкговых и саговников (рис.
9). Важнейшим с нашей точки зрения хвойным был бы янтароносный представитель
араукариевых Agathis levantensis. Повсюду росли древние папоротники,
печёночники, плауны, хвощи и листостебельные мхи. Цветковые растения, покрытосеменные,
были редкими, крохотными и неприметными, и были представлены преимущественно
разновидностями с примитивными цветками первичного типа. Экзотичные семенные
папоротники сократились в численности до почти реликтовых элементов. Примитивные
кустарники вроде Caytonia с семенами, охваченными купулой, и родственные
цикадеоидеи с бочкообразными стволами, по которым были разбросаны репродуктивные
органы, создавали странную, таинственную атмосферу пейзажа.
Мир был незнакомым вечнозелёным местом, где по лесу плавали взвешенные в воздухе
и разносимые ветром пыльца и споры – залог появления следующих поколений. Ещё
не существовало никаких броских выставок ароматных цветков, которые украшали
бы открытые местности пёстрым цветочным ковром или испещряли бы полог леса блистательными
всплесками красок. Те немногие ископаемые остатки растений, которые были найдены
в янтарном местонахождении, конечно же, не представляют всего разнообразия флоры,
которое должно было существовать11-13.
Здесь, в Ливане, и по всему земному шару обитали последние выжившие представители
растительных сообществ, подобные которым к середине мелового периода исчезнут
с лица земли.
Состав флоры всей планеты уже стоял на пороге преобразований, потому что начали
процветать покрытосеменные растения, и начался стремительный рост их разнообразия,
конечной точкой которого станет их господство в растительном мире (рис. 9).
Уже к середине мела были налицо существенные различия. Очевидным изменением
на отрезке времени продолжительностью в 30 миллионов лет было появление в процессе
эволюции новых родов и видов растений. Возникающие местообитания изобиловали
хвойными более современного типа. В бирманских янтароносных араукариевых лесах
душистые кедры касались ветвями стройных сосен, высокие секвойи росли рядом
с изящными метасеквойями, а раскидистые кипарисы были соседями компактных можжевельников14-20.
В подлеске по-прежнему доминировали папоротники (рис. 10) и плауны.
Покрытосеменные растения, эти восходящие звёзды, теперь настойчиво конкурировали
за место повсюду, и их растущее
24
 |
|
| Рисунок 9. В течение 80 миллионов лет мелового периода состав мировой флоры значительно изменился. Самым важным событием была радиация покрытосеменных растений. Она сопровождалась снижением разнообразия других групп растений. Буквы L, B и C показывают примерное время, когда процветали янтароносные леса из ливанских (L), бирманских (B) и канадских (C) местонахождений, и иллюстрируют в самых общих чертах соотношение между типами растений, которые могли существовать в данных местностях. |
разнообразие явственно сопровождалась снижением разнообразия других растений18,348.
Из буйной зелени выглядывали редкие группы изящных бледных цветков, а пальмовые
орехи накапливались в подлеске вместе с шишками хвойных деревьев. Процветали,
по меньшей мере, два злака (рис. 11), родственных современному бамбуку17,
возможно, даже образуя постоянно разрастающиеся заросли, которые угрожали захватить
обширные пространства, поросшие папоротниками и хвощами. Рогатиковые грибы и
трутовики украшали собой гниющие стволы деревьев, а дождевики и негниючники
благоденствовали на лесной подстилке91,345
(рис. 12, цветные таблицы 12A, 12D).
К тому времени, когда стал образовываться канадский янтарь, леса, кустарниковые
заросли и луга больше напоминали те, что существуют в настоящее время. Наши
знания о растениях, произраставших в Альберте в течение
25
|
кампанского века позднего мела действительно весьма обширны благодаря совместным
усилиям многих палеонтологов, работавших десятилетиями21-27.
Теперь в лесах, вероятно, совместно произрастали подокарпусы и смолоносные
деревья каури (араукариевые). Количество покрытосеменных растений значительно
увеличилось, и в это время некоторые из растений, произрастающих в северных
широтах, почти наверняка были листопадными. Вопрос о размерах и габитусе тех
древних цветковых растений, и о том, какой вклад они могли вносить в экосистему,
открыт для обсуждения. Превратились ли некоторые из них к этому времени в
деревья, или они лишь покрывали землю или представляли собой низкорослые кустарники?
Как сказали Деннис Браман и Ева Коппельхас о позднемеловых ископаемых покрытосеменных
Канады, «если у них был древовидный габитус, можно было бы ожидать находок
крупных кусков фоссилизированной древесины. Однако в настоящее время древесина
покрытосеменных растений неизвестна»21.
Поэтому мы можем лишь строить предположения относительно того, какова была
их роль в формировании ярусов этих лесов.
Существует множество факторов, которые определяют, где будет расти растение,
в том числе температура, доступность воды из-за осадков и испарения, тип почвы,
дренирование, освещённость, географическая широта, ландшафт и высота над уровнем
моря. На любом из янтароносных участков эти параметры менялись со временем,
оказывая влияние на растения, которые могли процветать там непрерывно на протяжении
ряда тысячелетий. И параллельно тому, как постепенно изменялись распространение
и относительная важность растений различных типов на протяжении всего мелового
периода, когда одни семейства, роды и виды
26
|
|
| Рисунок 11. Колосок примитивного злака Programinis burmitis в бирманском янтаре17. |
27
|
28
29
|
30
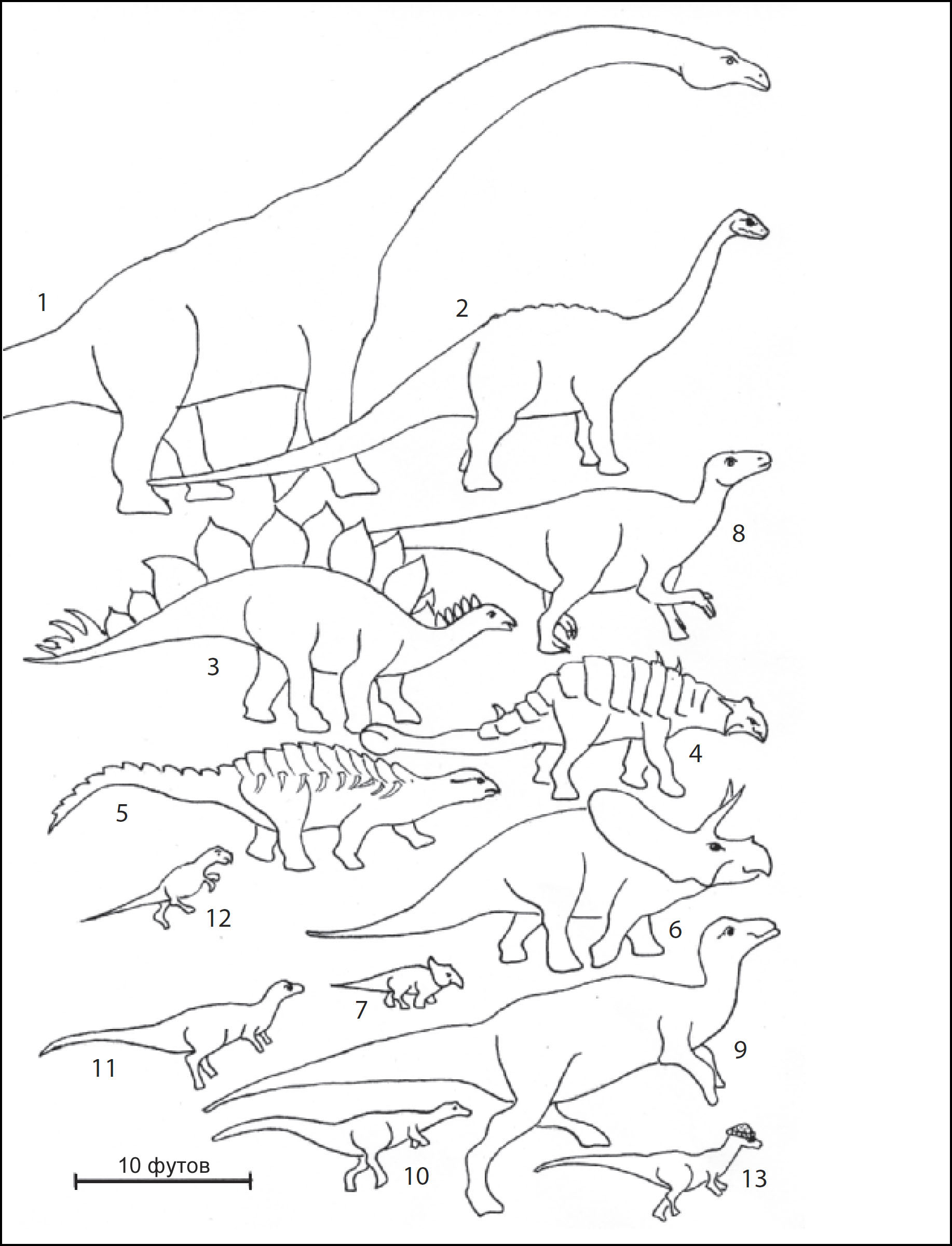
в Азии. Стада зауропод, крупнейших среди четвероногих травоядных,
с несоразмерно маленькими головами, венчающими длинные шеи, телами размером
с дом и ногами, подобными телефонным столбам, продолжали сотрясать землю неуклюжими
шагами на просторах Южной Америки. Вес этих титанов длиной от 80 до 100 футов
оценивается в 50 тонн. Однако в других уголках земного шара их можно было наблюдать
всё реже и реже. К позднему мелу в северном полушарии начинают впервые появляться
новые группы динозавров. И в это время мелкие пахицефалозавры, многие из которых
обладали ярко выраженной толстой крышей черепа, окаймлённой рядами костяных
бугров, и разнообразные цератопсы с рогами, костяными воротниками и непропорционально
крупными головами присоединились к стадам бронированных анкилозавров и двуногих/четвероногих
гадрозавров, которые могли достигать 40 футов в длину и весить более 4 тонн.
Как ни странно, но оказывается, что на южных континентах сохранились главным
образом раннемеловые типы динозавров.
По мере того, как травоядные из крупных и медлительных превращались в более
быстроногих существ меньшего размера, а некоторые вооружились защитными рогами,
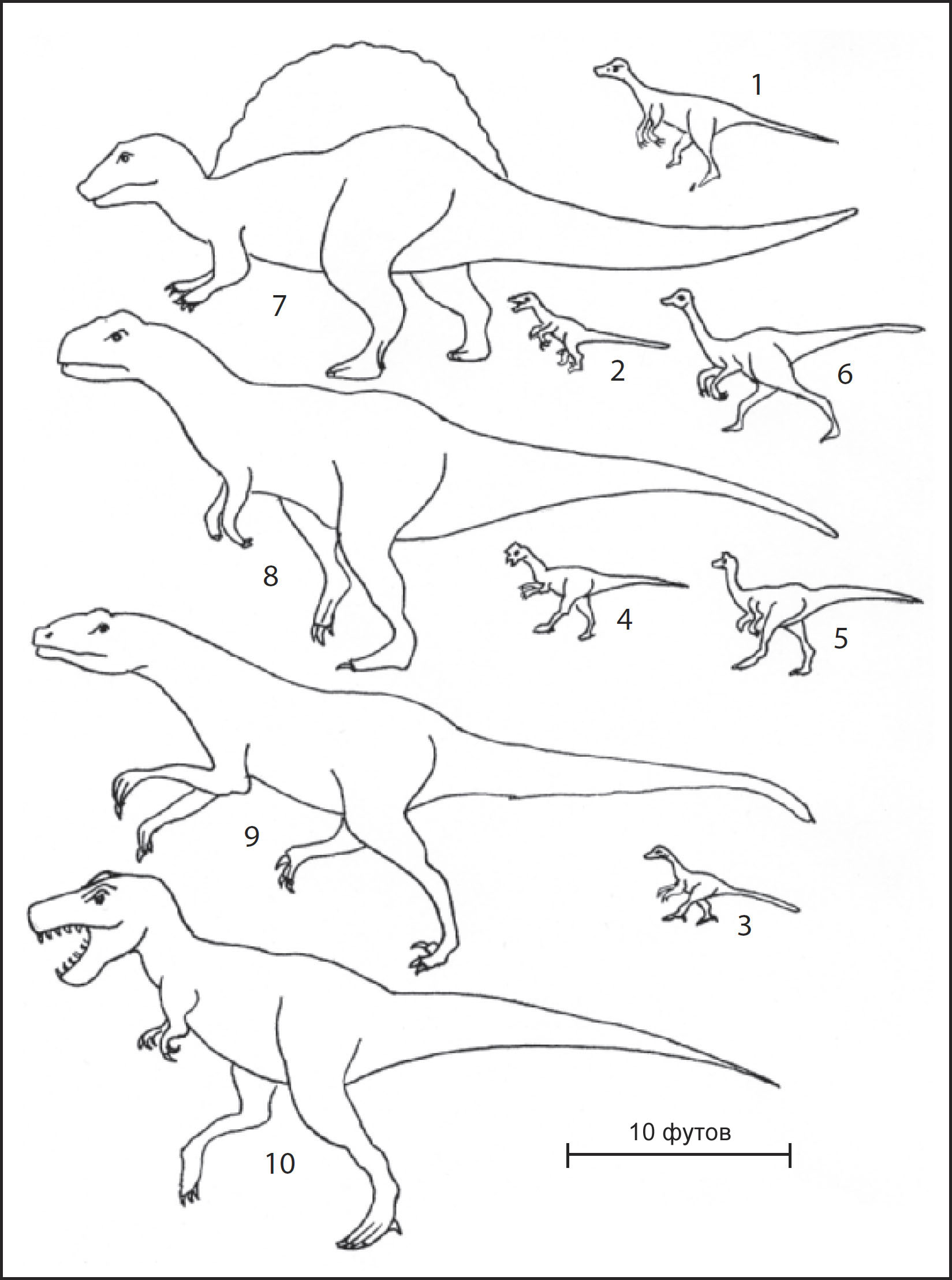
хищники, которые питались ими, тоже менялись (рис. 14). Первая половина мелового
периода была временем взрывного роста разнообразия теропод. Эти жестокие двуногие
машины, питающиеся мясом, становились всё крупнее, и, если было возможно, всё
более чудовищными. В северном полушарии появились дейнонихозавры, вооружённые
огромными серповидными когтями. Представители этой группы включали дромеозавров
и троодонов, которые обладали относительно крупными мозгом и глазами, и были
снабжены поднимающимися вторыми пальцами задних лап, заканчивающимися изогнутыми
когтями, которые могли выбрасываться вперёд, чтобы резать своих жертв. Теризинозавры,
также принадлежащие к этой группе, но явно ограниченные Азией*, не просто были
большими, но вдобавок каждая их передняя лапа заканчивалась тремя ужасающими,
напоминающими косу когтями.
К концу мелового периода Землёй правили самые крупные из всех наземных хищников
– тираннозавры. Любой ребёнок может с лёгкостью представить себе T. rex
с его характерными огромными челюстями, 10-дюймовыми изогнутыми клыками, головой
длиной 4 или 5 футов и телом длиной 45 футов. С их впечатляющим телосложением
несколько контрастировали необычно короткие передние лапы, заканчивающиеся крохотными
двупалыми кистями. Другими, менее известными, но столь же зловещими тероподами
были кархародон-
* Остатки теризинозаврид найдены и в Северной Америке, причём ещё до выхода в свет данной книги: в 2001 г. в Нью-Мексико был обнаружен Nothronychus. Но следует помнить, что, несмотря на родство с хищниками, теризинозавры были растительноядными животными. – прим. перев.
31
|
32
тозавры и гиганотозавры. Хотя их иногда считают падальщиками,
всем этим мегахищникам, вероятно, приходилось конкурировать за пищу с бродячими
стаями дромеозавров или троодонов.
Две другие появившиеся группы теропод, однако, могли кормиться в значительной
степени иной пищей. Орнитомимиды и овираптозавры отказались от рядов остроконечных
клыков в пользу торчащих вперёд беззубых клювов, которые, однако, сохранили
значительную режущую силу. Уникальная, с реликтовыми элементами, фауна динозавров
южного полушария включала абелизаврид, крупных рогатых теропод. Хотя чаще всего
в ископаемом состоянии сохраняются бесчисленные громко топающие стада тяжеловесных
травоядных и огромные хитрые хищники, также в изобилии встречались мелкие динозавры
размерами от курицы до большой собаки, и их важность не следует упускать из
виду.
Насекомые
В настоящее время существует примерно 762 семейства насекомых33;
если же вы умножите это число на четыре, то, вероятно, у вас будет представление
о разнообразии насекомых в меловой период, поскольку, даже если знать о том,
насколько мало существует меловых местонахождений ископаемых насекомых, для
этого периода известно уже примерно 490 семейств (приложение A).
Изменения в разнообразии насекомых можно продемонстрировать, сравнивая раннемеловые
ископаемые остатки с образцами из середины мела и позднего мелового периода.
Многие из раннемеловых насекомых сохранили архаичные черты, уникальные для каждого
вида физические признаки, от своих юрских предшественников, и это заметно на
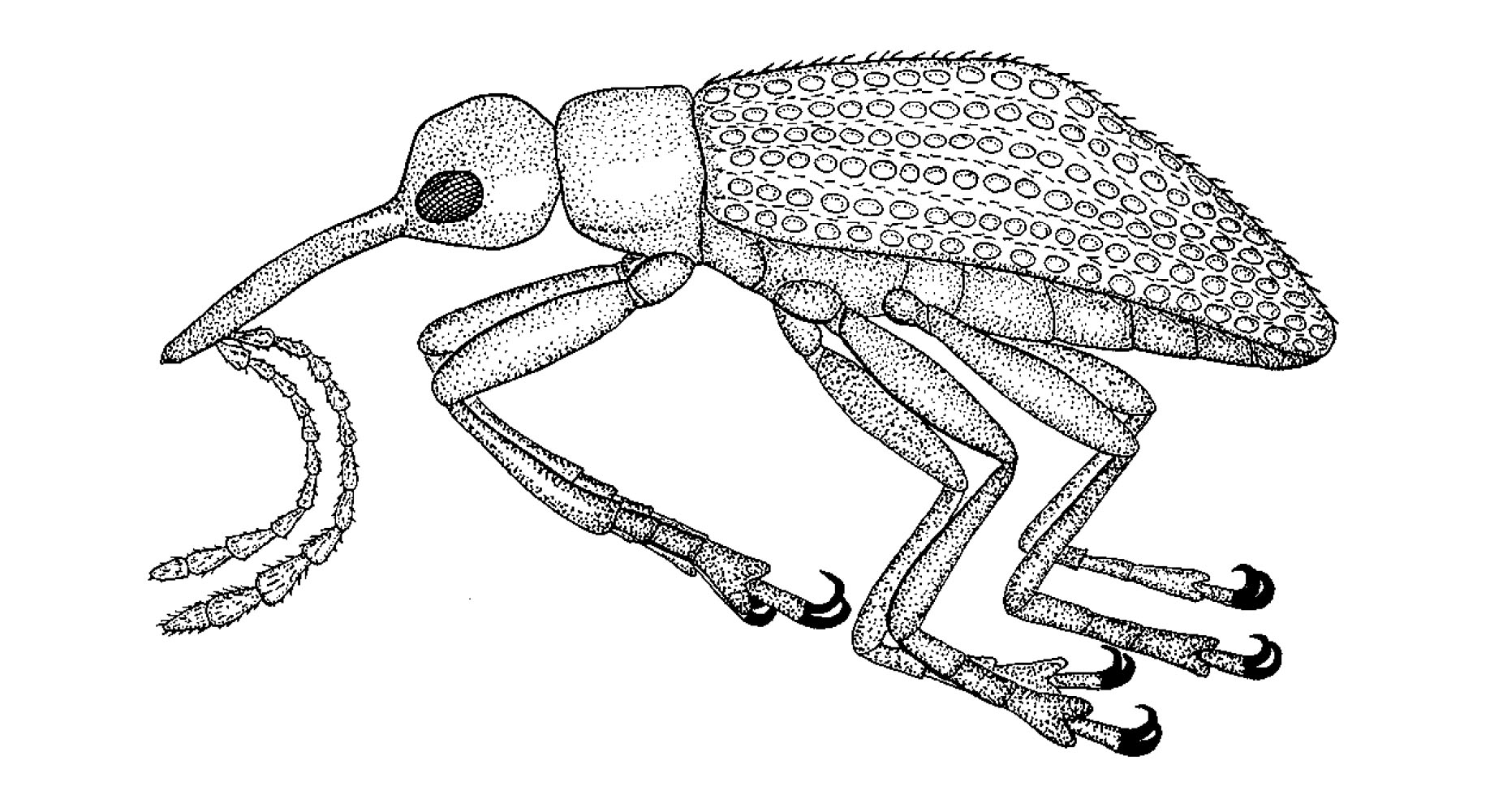
их представителях из ливанского янтаря. Они включают примитивных долгоносиков34,
которые развивались в шишках хвойных (рис. 15), щетинистых палочников, гигантских
кобылковидных насекомых и бурящих стебли пилильщиков – все они питались листьями
или пыльцой голосеменных35. Они делили
свой мир с шипастыми цикадами, кормящимися на гинкго, и с примитивными молями,
гусеницы которых поедали мох36. Мелкие
трипсы37 с ротовыми органами, видоизменёнными
для прокалывания и соскребания, поедали пыльцу и даже вызывали опадение листвы
с некоторых араукариевых, а тли с длинными хоботками38
высасывали соки хвойных, хвощей, папоротников и саговников.
Как раз в это время некоторые насекомые приобрели
33
|
вкус к крови позвоночных. В раннемеловую эпоху слепни, мошки,
комары35, мокрецы, москиты и ещё многие
другие менее известные роящиеся насекомые рыскали в поисках свежей крови любых
живых существ – от мелких лягушек до гигантских динозавров13.
Увеличение биологического разнообразия цветковых растений в середине мелового
периода обрекло на вымирание многих архаичных насекомых. Фактически же, в это
время имело место больше эпизодов исчезновения насекомых, чем даже на мел-палеогеновой
границе35. Середина мелового периода
была временем, когда древние, давно закрепившиеся на своих позициях насекомые
конкурировали с более современными за одни и те же местообитания. К некоторым
из примитивных типов принадлежали неприметные жучки, настолько мелкие, что голодные
ящерицы, обследующие кору, вероятно, не обратили бы на них внимания (рис. 16).
Один такой жук питался клещами и обладал дополнительным сегментом, который придавал
его передним ногам дополнительную степень гибкости, что помогало хватать добычу39.
Его соседом по местообитаниям был странный жук с кутикулярными образованиями
на голове и туловище, которые могли бы поспорить с воротниками, рогами и пластинами
какого-нибудь динозавра. У этого насекомого были не только щитовидные выросты,
защищающие выпуклые глаза, но ещё и похожий на плавник вырост на макушке головы,
плюс ряд складок, которые тянулись вдоль спины347.
Третий жук был настолько архаичным, что его пришлось описывать в составе нового
семейства40. Обладая редуцированными
ротовыми органами, это насекомое могло питаться лишь грибами, мхом или водорослями.
Другие жуки использовали для защиты химические вещества349.
34
Рисунок 16. Три потрясающих жука из бирманского янтаря. Справа вверху
представитель вымершего подсемейства похожих на муравьёв жуков-сцидменид
(Scydmaenidae)39. Уникальной
особенностью этого небольшого охотника на клещей является дополнительный
сегмент в его передних ногах. Вверху слева показан вымерший жук-сильванид
с выростами на голове и туловище, которыми он мог бы посоперничать со
многими динозаврами. |
35
Среди крупных насекомых, которые появились в процессе эволюции
за миллионы лет до этого времени, и чьё господство подходило к концу, были существа,
которые выглядели как гибрид между кузнечиком и сверчком (цветная вкладка 8A).
Эти элканиды мощного телосложения, с выпуклыми глазами и длинными усиками, вероятно,
питались голосеменными. Они непосредственно взаимодействовали с динозаврами,
став для них и источником пищи, и конкурентами за истощающиеся растительные
ресурсы. Однако их мелкие родственники, триперсты и прыгунчики, привлекали внимание
голодных гекконов и земноводных (цветная вкладка 7B).
Свидетельства существования более современных групп насекомых в бирманском янтарном
местонахождении середины мелового периода включают долгоносика с коленчатыми
усиками, похожего на тех, которые питаются цветковыми растениями в настоящее
время (цветная вкладка 5E)41. Повреждения,
сделанные насекомыми на цветках покрытосеменных растений из бирманского янтаря,
показывают, что растительноядные животные уже приспособились к рациону, состоящему
из цветковых растений18,19
(цветная вкладка 14A). Возможно, повреждения были нанесены мелкими узкотелыми
трипсами или личинками галлиц (цветные вкладки 3A, 3C). Наверняка существовала
целая орда насекомых, в том числе фонарницевые и тли, которая питалась примитивными
злаками, и они конкурировали с динозаврами за этот деликатес17,42.
Некоторые из этих тлей неплохо жили, обладая всего лишь двумя крыльями вместо
четырёх, имеющихся у тлей в настоящее время43.
И тогда эти сосущие насекомые могли переносить вирусов растений. Удивительной
находкой стала мелкая пчела, которая явно опыляла некоторые из покрытосеменных
растений в том местонахождении44 (цветная
вкладка 14B).
Наряду с типичными кровососами, в местонахождении бирманского янтаря также обнаружены
клещи45 (цветная вкладка 11E). Это
настоящий вызов для воображения – представить себе, какие ещё членистоногие
с колющим ротовым аппаратом кормились в том янтароносном лесу, насколько важную
роль играли они в передаче патогенных организмов, вызывающих болезни46,47
(цветная вкладка 10). Настоящей неожиданностью стали двукрылые скорпионницы
с клювами, внутри которых находились похожие на кинжалы мандибулы с зазубренными
краями (цветные вкладки 11A, 11B). Мы можем лишь гадать о том, сколько других
типов кусающихся членистоногих существовало в тех лесах мелового периода.
К позднему мелу ещё больше насекомых напоминало
36
живущих в наши дни, в том числе питающийся на пальмах жук с
канадского местонахождения янтаря26
(цветная вкладка 5A). Также должно было существовать значительное количество
новых растительноядных насекомых, родственных тем, которые в настоящее время
кормятся на клёнах и платанах, ольхе и вязе, или на лилиях и осоках – все эти
растения существовали в то время (цветная вкладка 12B). Многие листья, вероятно,
покрывали сладкие выделения и сброшенные шкурки тлей. Большое количество тлей
в канадском янтаре, составляющее примерно около 40% от всех ископаемых остатков
животных, указывает на то, что они были одной из доминирующих групп растительноядных
насекомых49 (цветная вкладка 2C). К
тому времени у некоторых самок выработалась способность отрождать копошащийся
молодняк прямо на растение и отказаться от стадии яйца – преимущество, которого
были лишены их предки. Поскольку ныне живущие тли переносят примерно 50% распространяемых
насекомыми вирусов растений, возможно, на них лежит ответственность за быструю
смену групп позднемеловых растений. Комары в канадском янтаре указывают на то,
что именно в данном конкретном месте эти кровопийцы были многочисленными – возможно,
они размножались в солёных болотах вблизи места образования янтаря191
(цветная вкладка 11D), тогда как ротовой аппарат мокрецов заставляет предполагать,
что они кормились на динозаврах51.
Различные группы насекомых появлялись и исчезали на протяжении всего мелового
периода, и многие из них были прямо или косвенно связаны с динозаврами. Некоторые
сосущие насекомые просто обеспечили себе источник пищи, а другие поедали растения
у динозавров или распространяли инфекционное увядание и другие болезни растений.
Падальщики утилизировали отбросы и трупы динозавров, а кровососущие виды, находящиеся
на вершине пищевой цепи, не только пировали на динозаврах, но ещё и вводили
в их тела возбудителей болезней. И то, о чём будет рассказано далее – это интригующая
история о борьбе между растениями, насекомыми, патогенными микроорганизмами
и динозаврами, которая происходила миллионы лет назад.
37
Солнце немилосердно жарило полог свежевымытого дождём леса,
заставляя пар подниматься над влажной растительностью, словно клубы дыма. Над
этой лиственной крышей возвышались кроны доминирующих деревьев – каури (из семейства
араукариевых). Будучи массивными деревьями с основанием ствола диаметром до
40 футов, они достигали высоты 120 или даже 200 футов. Их уплощённые эллиптические
листья трепетали в лучах солнца, демонстрируя заострённые кончики, выраженные
продольные жилки и повреждения, нанесённые насекомыми. Шишка каури, кишащая
маленькими гусеницами, оторвалась от своей ножки высоко в ветвях, вращаясь,
полетела вниз сквозь полог леса, сбивая листья и отскакивая от ствола, и закончила
свой путь в огромной груде опавшей листвы, скопившейся вокруг основания векового
дерева.
Шум насторожил группу молодых пахицефалозавров, которые искали корм в тускло
освещённых уголках подлеска. Один из них подошёл к тому месту, куда упала и
зарылась шишка, и сунул лапу в мусор. Он вытащил повреждённую шишку и получил
в награду нескольких сочных личинок, которых съел. Пахицефалозавры казались
карликами среди окружающих деревьев и пышной растительности. Изящные хвойные
деревья возвышались над подлеском, словно шпили собора, рисующиеся над городским
пейзажем, тогда как другие виды архаичных голосеменных образовывали густой полог.
Их узкие, остроконечные, а часто колючие листья весьма отличались от экзотичных,
покрытых рябью из жилок веерообразных листьев окружающих деревьев гинкго, которые
также тянулись вверх за своей долей света, попадающего в полог леса. Их густые
листья, из которых укусы насекомых вырезали странные скульптуры, трепетали на
ветру и создавали симфонию тонов зелёного цвета, тонко оттенённых случайными
вспышками красновато-коричневых, бурых и жёлтых красок.
38
Молодые двуногие динозавры выбрались из тускло освещённых
уголков мрачных галерей на залитые солнцем прогалины, созданные упавшими великанами,
и пошли вдоль берега ручья, прорезающего заросли нижнего яруса леса, состоящие
из вечнозелёных деревьев, перемежающихся с пушистыми древовидными папоротниками
и жёсткими пальмами. Они пировали, поедая папоротники, росшие повсюду в огромном
изобилии размеров и форм, с листьями, нежными, словно кружева, или жёсткими,
как щетина. Некоторые из них, вроде жёстких лазающих папоротников, украшающих
ветви могучих деревьев или росших на стволах, находились слишком высоко, чтобы
до них можно было дотянуться. Но более мелкие разновидности, густо растущие
вдоль речного берега или пышно разрастающиеся на лугах, были лакомством. Мужские
и женские растения саговников, напоминающие коренастые пальмы, соперничали за
место с крупными папоротниками, приземистыми хвойными и примитивными родственниками
кустовидных магнолий с мелкими мясистыми плодами, и повсюду можно было найти
и съесть лакомые кусочки. Любое пригодное для жизни место в подлеске было покрыто
обильной порослью папоротников, хвощей, мелких травянистых растений, а надеющиеся
на лучшее проростки обеспечивали жильём и пищей множество мелких позвоночных
и беспозвоночных. Влажные уголки кружевной тени, покрытые коврами, сотканными
из изящного папоротника-орляка, бледных, словно привидения, лишайников, и тёмных
бархатистых мхов, были домом для клещей, ногохвосток и пауков.
Лианы и эпифиты украшали все ярусы леса. Они успешно превратили полог леса в
висячие сады, где ползли по покрытым мхом ветвям и прорывались сквозь покров
из листвы. Ствол и ветви деревьев были драпированы прядями нежных листостебельных
мхов, полотнищами печёночных мхов и завесами тонколистных папоротников. Цепкие
лианы, тянущиеся вокруг стволов и членов и между ними, связывали воедино все
ярусы леса и служили чем-то вроде шоссе для тараканов и сверчков. Они переплетались
с массами спутанных корней, образуя в подлеске целую сеть, и извивались и закручивались
петлями, выкарабкиваясь из верхних ярусов полога леса. Эпифиты закреплялись
на каждом уровне, ложась бременем на ветви, давая густой обильный рост на любой
доступной поверхности, и обеспечивая укрытиями лягушек и птиц. Даже осоки росли
в развилках деревьев, а похожие на бамбук злаки с пушистыми колосками процветали
на открытых местах. На уровне земли мусор из опавших листьев, шишек, веток,
стволов, коры и примитивных плодов служил идеальным домом для сапрофитных грибов
разнообразнейшего
39
обличья, а также для многоножек, тысяченожек и скорпионов.
А глубоко в глинистой почве хвойные, образующие лес, раскидывали опорные корни,
которые сосуществовали с мицелием симбиотических грибов и снабжали пищей нематод,
жуков и равноногих ракообразных.
Эти зелёные джунгли изобиловали невидимой глазу жизнью. Пахицефалозавры были
не единственными животными, которые там кормились, но всего лишь одной из наиболее
заметных форм. Тысячи видов насекомых наряду со значительным контингентом прочих
членистоногих ползали по каждому дереву и кустарнику, жуя и прогрызая себе путь
по этому ломящемуся от изобилия столу. Жуки тайком точили ходы под корой или
поедали пыльцу. Гусеницы скрытно кормились на нижней стороне листьев, а фонарницевые
потихоньку сосали соки растений. Пели замаскировавшиеся сверчки, а голоса невидимых
стрекочущих кузнечиков наполняли воздух. Растительноядные насекомые существовали
практически на каждой поверхности растения на любом уровне по всему лесу. Листва
страдала от жуков-листоедов, скелетировавших листья, от минирующих личинок мелких
молей, от сверчков, выгрызавших в ней дыры, и от гусениц, обгрызавших листья.
Ящерицы, лягушки и мелкие млекопитающие лазили в поисках добычи по веткам и
через лианы, а другие теплокровные позвоночные в это же время обрывали с растений
питательные семена и съедобные листья.
В подлеске группа маленьких пахицефалозавров продолжала искать грибы, упавшие
плоды и раскрывшиеся шишки с семенами, чтобы набить себе брюхо. Один из них
раздробил гнилое бревно, и оказавшиеся внутри него жирные белые личинки жуков
были с жадностью проглочены этими оппортунистическими всеядными существами.
Внезапно насторожившись, они прервались и дружно взглянули вперёд, в ту сторону,
откуда вдруг донёсся треск. Словно безмолвные призраки, большое стадо живущих
в лесу цератопсов незаметно двигалось по подлеску, и теперь животные пользовались
своими массивными головами, чтобы валить молодые сочные деревья и лакомиться
их листвой. Потревоженный шумом, одинокий самец-анкилозавр, спокойно кормившийся
в зарослях хвоща, мощным рывком бросился прочь, яростно продираясь сквозь заросший
подлесок. Стая страусообразных орнитомимид, шагая по тропе, протоптанной ежедневным
движением множества крупных травоядных, вышла в подлесок и начала, пользуясь
своими клювами, обрывать
40
листву и собирать насекомых с окружающих кустов и низкорослых
травянистых растений. Их преследовали по пятам несколько молодых троодонов,
изгнанных из стаи новым вожаком. Они не ели почти неделю, а орнитомимиды выглядели
лёгкой добычей. Подкравшись поближе, более сильный и агрессивный троодон, движимый
голодом, прыгнул, пытаясь схватить ближайшую жертву. Оба динозавра подскочили
вверх, нанося друг другу удар задними лапами. Но орнитомимид был крупнее и опытнее,
чем нападающий, и его более длинные ноги с пальцами, вооружёнными когтями, оставили
кровоточащий шрам на грудной клетке неопытного хищника, заставляя его отступить
вместе со своими спутниками. Позже молодые тероподы согласятся на трапезу из
насекомых и нескольких кусков мяса змеи, на которую они случайно наткнутся.
Никто из травоядных не останется голодным, потому что этот пышный влажный тропический
лес в изобилии даёт растения для их пиршества.
Следы пищевого поведения насекомых были повсюду. Хвощи бледнели и съёживались
из-за личинок жуков, живущих в их стеблях. Гинкго лишались цветочков и листвы
из-за заражения личинками молей и пилильщиков. На саговниках и цикадофитах пищевое
поведение насекомых проявлялось в виде зияющих отверстий в их листьях, а их
стебли, покрытые бурыми пятнами, были полны жирных личинок молей и жуков. Вайи
чахлых древовидных папоротников отмерли, когда личинки долгоносиков скрытно
пробурили ходы сквозь их ткани. Мелкие папоротники потихоньку лишались зелени
из-за листогрызущих личинок пилильщиков и молей. Некоторые листья гигантских
деревьев каури свернулись и побледнели из-за многочисленных листовых мин, прогрызенных
крохотными личинками молей, а огромные отверстия в листьях свидетельствовали
о присутствии ощетинившихся пучками торчащих волосков гигантских долгоносиков,
осторожно ползавших по ветвям. Упавшие сучья демонстрировали сильно источенную
древесину, которая служила выводковыми камерами для короедов и долгоносиков.
Фактически, гибнущие деревья каури занимали целые области леса. Иглы деревьев
метасеквойи желтели из-за того, что на них кормилось множество щитовок и тлей,
а в это время крепко сложенные жуки-листоеды и длинноусые кузнечики питались
41
листвой. Участки игл на различных хвойных были покрыты
и заплетены паутинным покровом, созданным живущими в скоплениях личинками пилильщиков,
а деятельность прожорливых гусениц пилильщиков, палочников и кузнечиков частично
оголила крупные ветви.
Цветковые растения не избежали внимания со стороны насекомых. Отметины на краях
многих мелких цветков как на травах, так и на кустах, указывали, что грызущие
насекомые включили их в свой рацион. Листья на некоторых из них были «сшиты»
вместе нитями, которые выделяли шёлковые железы мелких гусениц, создававших
безопасное убежище, где они могли спокойно кормиться. Свернувшиеся листья пальм,
с которых соскребли внутренний эпидермис, и старые листья с огромными разрезами,
тянущимися от краёв до средней жилки, были свидетельством деятельности различных
стадий жизни зелёных гусениц с белыми полосками. По подлеску разбросаны поваленные
стволы пальм, ослабленных большими белыми личинками долгоносиков, прогрызающими
их ткани. В ближайшем пруду многие из листьев кувшинки были съедены популяциями
жуков-листоедов. Древний бамбук, ползущий вверх по стволам араукарий, страдал
от значительного повреждения листьев большими бурыми пластинчатоусыми жуками,
а его корни стали объектом нападения мелких гусениц. Скрытые от глаз орды насекомых
постепенно потребляли свою долю изобильной лесной флоры.
Зелёные растения – это важнейшие организмы в любой наземной
экосистеме, и так было с тех самых пор, когда они впервые возникли в процессе
эволюции. В плане биологического разнообразия они уступают лишь насекомым и
представляют примерно четверть от общего количества известных видов. Они улавливают
энергию Солнца и преобразуют её в растительные ткани, которые поддерживают существование
всех животных. Травоядные переносят эту энергию непосредственно в собственные
ткани, когда поедают растения, а плотоядные получают энергию опосредованно,
когда питаются травоядными. По этой причине растения находятся в самом основании
пищевой цепи.
Существует много факторов, которые ограничивают место жительства конкретного
вида насекомых или динозавров, но обеспечение пищей – самый важный из них. Количество
растений и их пищевые качества определяют, сколько
42
|
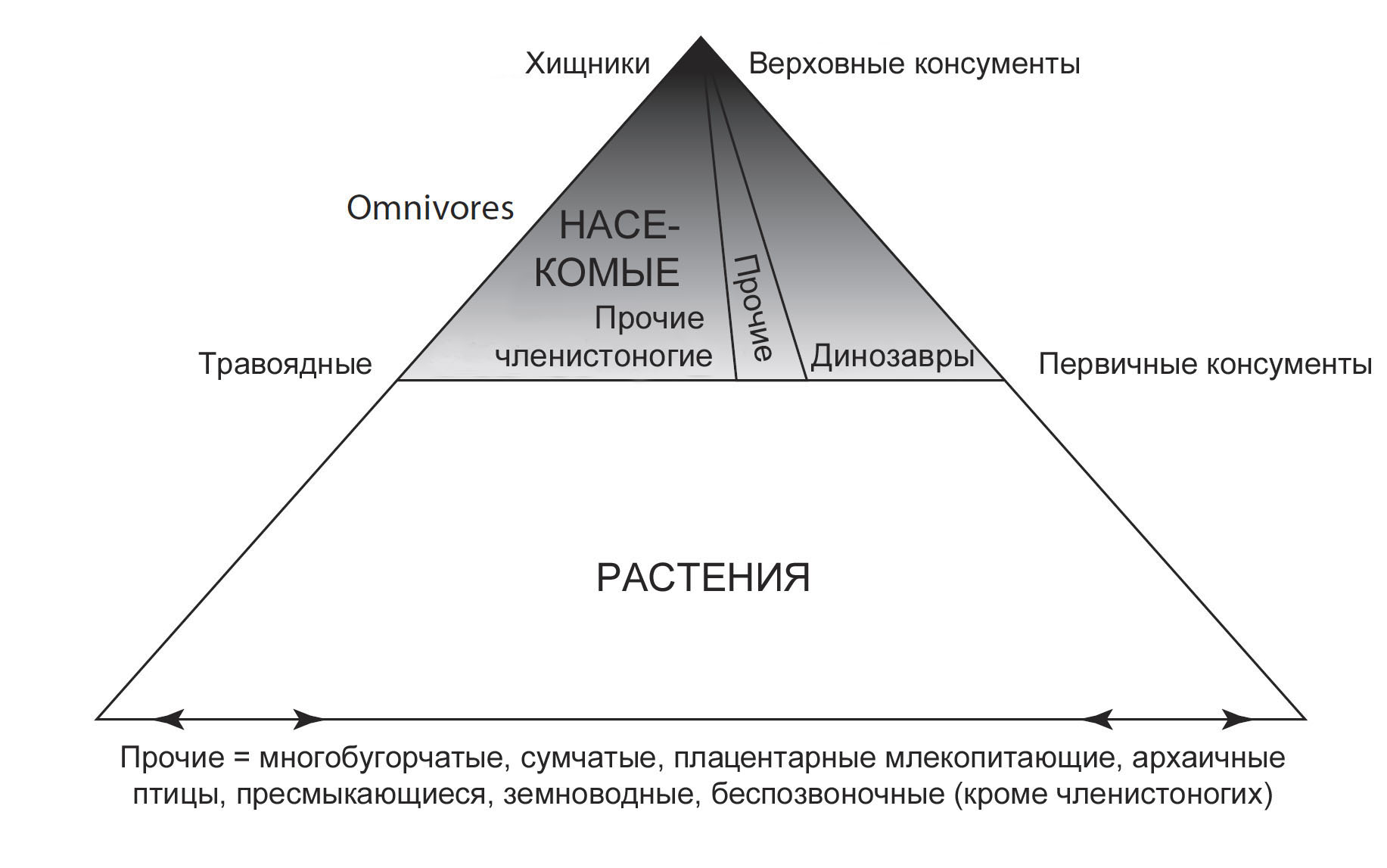
и какие типы травоядных, а в конечном счёте и плотоядных животных и где будут жить. Чтобы иллюстрировать этот принцип, биологи построили экологические пирамиды (рис. 17). Они существуют в нескольких вариантах: пирамида чисел (сколько организмов), пирамида биомассы (общий вес всех представленных живых организмов) и пирамида энергии (сколько энергии произведено, использовано и запасено в организмах). Пирамиды разделены на несколько уровней. Нижний и самый большой включает продуцентов (производителей) – растения. Последующие уровни заняты консументами (потребителями) – животными. Далее консументы подразделяются на типы или порядки. Первый тип, или первичные консументы – это травоядные (потребители растений), второй тип – плотоядные (потребители животных), которые питаются травоядными, и третий тип – плотоядные животные, которые питаются другими плотоядными, и так далее, пока не дойдёте до верховного хищника. Всеядные существа вписываются в вездесущую категорию, которая пронизывает все уровни консументов.
43
Многие люди не осознают того факта, что насекомые доминируют на уровне типичных
консументов любой экологической пирамиды, будь это пирамида чисел, биомассы
или энергии, и они, вероятно, занимают это положение с тех пор, как впервые
стали ходить по земле. Они составляют до 60% травоядных консументов в большинстве
экосистем. Помимо этого, насекомые и другие мелкие членистоногие потребляют
значительно большее количество растительных тканей, чем все позвоночные в любом
исследованном местообитании (кроме травянистых равнин), и это преобладание существовало
с самых ранних времён53,55.
Это означает, что остальные 40% из доли консументов в пирамиде 100 млн. лет
назад должны делить между собой динозавры и все другие меловые животные, такие,
как млекопитающие, рептилии, амфибии, птицы и беспозвоночные (рис. 17). Точное
соотношение долей, относящихся к каждой из этих групп – это вопрос, на который
невозможно дать ответ. Но в любом случае, какой бы большой ни была часть пирамиды,
которая принадлежала бы динозаврам, мы знаем, что она была бы значительно меньше,
чем часть, принадлежащая насекомым!
Экологическая пирамида должна рассматриваться как постоянно расширяющееся и
сужающееся построение. Её конфигурация меняется от одной экосистемы к другой.
Влажный тропический лес, обладающий величайшим видовым разнообразием на всей
Земле, представлял бы собой законченную пирамиду. В меловой период этот тип
местообитаний наверняка охватывал значительную часть земного шара. Половина
известных в нашем мире видов обитает во влажных тропических лесах, которые сейчас
охватывают около 6% поверхности суши66.
Количество видов насекомых оценивается в пределах от 5 до 10 миллионов. Поскольку
большая часть суши в меловой период представляла собой тропические местообитания,
из этого следует, что тогда имело место значительно большее разнообразие насекомых.
На противоположном конце спектра разнообразия находилась бы пирамида со скудной
растительностью вроде пустынной или тундровой. Фактически, между влажным тропическим
лесом и тундрой имеет место 75-кратное различие в биомассе растений, и 20-кратное
– между влажным тропическим лесом и болотом. К различным уровням биоты, от сообществ
до ниш, можно применить меньшие, специализированные пирамиды. В пределах каждый
экологической пирамиды существует один или больше ключевых видов, присутствие
которых является критически важным для её стабильности. Все другие виды в рамках
экосистемы обладают различным уровнем важности. Взаимосвязи могут
44
внезапно измениться, и пирамида может разрушиться в любое время.
Любое разрушение в дальнейшем сопровождается периодом восстановления, и пирамида
расширяется, пока вновь не достигнет равновесия между растениями и животными.
Но по итогам её восстановления популяции флоры и фауны могут оказаться вовсе
не теми же самыми. Уменьшение и расширение экологической пирамиды в обычных
условиях – циклический или сезонный процесс, но оно может стать катастрофическим
из-за пожара, засухи, наводнения, исчерпания ресурсов пришлыми видами и так
далее.
В границах каждой экосистемы существует множество разнообразных местообитаний
со специализированными нишами. В них животные конкурируют за пищу, пространство
и укрытия. В лесу эти местообитания распределены ярусами, выстроенными в вертикальном
направлении. Во влажном тропическом лесу имеются открытое воздушное пространство
наверху, а внутри леса пригодными для заселения местообитаниями являются ярус
эмергентов, полог леса, нижний ярус, ярус кустарников, ярус травянистых растений,
лесная подстилка и почва. Реконструируя течение жизни на древних ландшафтах,
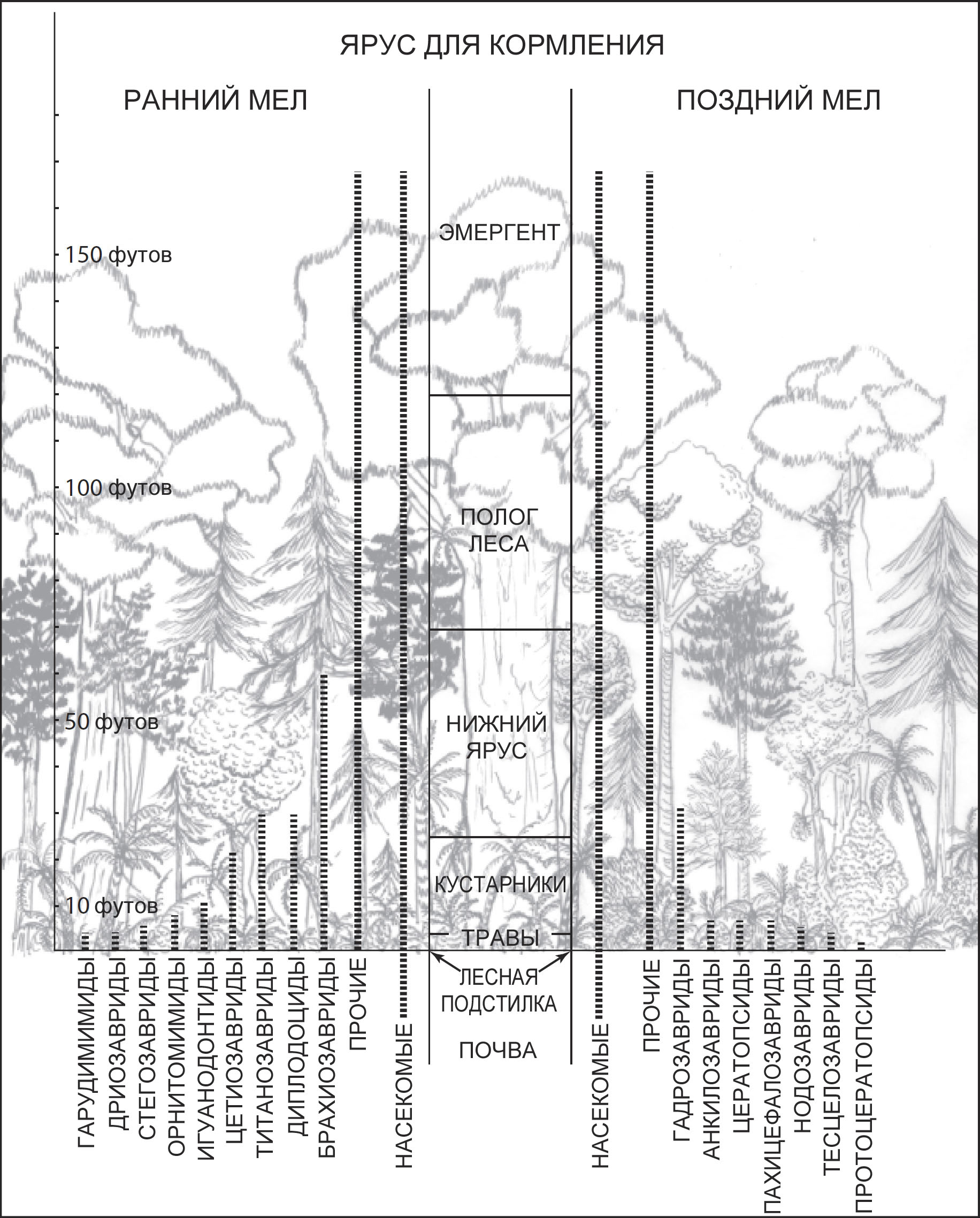
палеоэколог пробует соотнести фауну с этими ярусами (рис. 18). Воздух над лесом
мог быть домом для парящих птерозавров и архаичных птиц (перья опускались вниз
сквозь кроны деревьев, чтобы оказаться захороненными в янтаре во всех трёх янтарных
отложениях: цветные вкладки 15B, 15C). Птицы преследовали насекомых, роящихся
над кронами деревьев в жарком влажном воздухе, или садились на ветки, чтобы
схватить кормящихся там насекомых, пока растительноядные птицы питались семенами,
пыльцой и плодами. Мелкие птерозавры гонялись за стрекозами, а крупные, возможно,
пикировали вниз и срывали ничего не подозревающих млекопитающих с вершин деревьев.
На всех ярусах леса шныряли, оставляя после себя в смоле шерсть, мелкие млекопитающие
– многобугорчатые и сумчатые, которые перешли к лазающему образу жизни. В большинстве
своём они были растительноядными, всеядными или насекомоядными. Бок о бок с
ними, лазая по лианам или среди листвы, и даже скрываясь в эпифитах, жило несметное
множество других животных вроде древесных лягушек, ящериц, змей, гекконов, сотни
тысяч насекомых и бесчисленное множество иных мелких членистоногих (цветная
вкладка 15A). Каждый вид животных нашёл бы свою собственную, специфическую нишу
– физическое и биологическое пространство, а также тип окружающей обстановки,
удовлетворяющий видовые потребности. На уровне почвы, например,
45
|
46
могли бы жить насекомые, млекопитающие, и даже динозавры, приспособленные
к роющему образу жизни. Они выкапывали бы корни в листовой подстилке, рылись
в земле или копали норы. Клещи и тысяченожки, ногохвостки и сверчки, кольчатые
черви и нематоды, улитки и скорпионы, а также тараканы ползали или копошились
среди листьев, ели сами или становились чьей-то едой. Насекомые ползали и кормились
на каждом ярусе леса. Они пожирали листву, цветы, плоды, семена, древесину,
пыльцу, нектар, и даже смолы. Возможно, триста или более насекомых питались
на единственном виде деревьев, а многие из их, вероятно, включали в свой рацион
лишь три вида растений или меньше.
А чтобы представить себе динозавров в лесу, достаточно приложить лишь немного
воображения. Мы полагаем, что многие из динозавров, живших в лесах, были ограничены
в выборе ярусов, где могли кормиться, лишь самыми нижними из них. Похоже, никто
в настоящее время не предполагает, что нептичьи динозавры жили на деревьях или
хотя бы лазали по ним*. Размер мог ограничивать глубину проникновения крупнейших
видов динозавров в лес, и они, возможно, вынуждены были питаться по краям зарослей
или в более открытых редколесьях. Но наверняка по тёмным и густым влажным тропическим
лесам тихо бродили в поисках пищи мелкие и среднеразмерные двуногие и четвероногие
динозавры.
Животные, которые не летали и не лазали по деревьям, должны были питаться в
пределах вертикального диапазона, который диктовался их высотой, или искать
какой-то иной способ расширить этот диапазон. Предполагалось, что некоторые
четвероногие динозавры использовали для этого позу треножника и удерживали равновесие,
опираясь на свой хвост и задние ноги, чтобы принять позу для кормления, которая
увеличивала их высоту52. Вероятными
кандидатами на это считались зауроподы, уже снабжённые чрезвычайно длинными
шеями, и крепко сложенные стегозавры (рис. 19). Поэтому кормовые ярусы динозавров
обладали широким потенциалом (рис. 18); но нам никогда не будут известны их
истинные пределы. В наши дни многие наземные животные проводят значительную
часть времени, кормясь на уровне глаз или ниже, с некоторыми очень немногими
исключениями. В целом же мега-травоядные позднего мела кормились в более низком
ярусе, чем раннемеловые, по крайней мере, в северном полушарии. Раннемеловые
зауроподы могли конкурировать с насекомыми, успешно питающимися в нижнем ярусе
леса. Позднемеловые цера-
* В настоящее время динозавры семейства Scansoriopterygidae считаются древесными формами. На момент выхода книги в свет был описан лишь Scansoriopteryx, в настоящее время известны явно древесные Epidexipteryx и Yi.
47
|
48
|
49
топсы могли искать пищу лишь в пространстве от низкорослых
кустарников до уровня земли, а протоцератопсы сосуществовали с насекомыми на
уровне травяного покрова экосистемы. Но в араукариевых лесах всё пространство
над нижним ярусом доставалось насекомым, другим членистоногим и мелким позвоночным
(рис. 20).
Травоядные – это самые многочисленные и разнообразные животные в любой экосистеме.
Так как же конкурировали растительноядные насекомые и динозавры? Вероятно, почти
так же, как насекомые конкурируют с млекопитающими (в том числе и с нами самими).
Поскольку насекомые кормятся растениями на всех ярусах, мы предполагаем что,
чем бы ни питались динозавры, этим также питались и насекомые. Мы знаем, что
растительноядные насекомые оказывают большее воздействие как на рост, так и
на воспроизводство растений, чем млекопитающие53,
и что в экосистеме может существовать почти в десять раз больше беспозвоночных
потребителей растений, чем млекопитающих54.
Эти те же самые наблюдения, несомненно, применимы к древним экосистемам; различие
в воздействии на растения со стороны мега-травоядных по сравнению с насекомыми
заключается в большем размере динозавров. Тем не менее, следует помнить, что
в весовом отношении насекомые в большинстве экосистем превосходят динозавров.
Так, одно мега-травоядное, возможно, съело бы столько же, сколько 100 000 насекомых,
или даже больше, но общее воздействие многих миллионов насекомых было бы более
постепенным и распределённым по большей площади. Крупные травоядные со своим
более крупным размером тела склонны поедать более широкий ассортимент растений,
откусывать большие куски и передвигаться по большей области, в больших количествах
вытаптывая растения и оставляя навоз и мочу. Это за короткое время создаёт на
небольших участках ландшафта области с уничтоженной листвой и с нарушенной растительностью.
Насекомые, обладая небольшим размером, но невероятной численностью, могут съесть
столько же, или даже больше, но они обычно более равномерно распределены по
площади экосистемы. Благодаря своему небольшому размеру насекомые могут выборочно
питаться семенами и корнями, что оказывает совокупное воздействие на жизнеспособность
растений. Насекомые также переносят возбудителей заболеваний растений, которые
способны истребить целые популяции растений. Поэтому насекомые, пусть даже мелкие
и неприметные, вероятно, были серьёзными конкурентами динозавров, не только
поедая больше растений, но также оказывая воздействие на здоровье и распространение
растений в целом.
50
Исходя из поведения ныне живущих растительноядных насекомых,
мы можем делать выводы о том, как они могли конкурировать с динозаврами. Мы
знаем, что долгоносики-цветожилы (Nemonychidae), похожие на найденного в ливанском
янтаре, питаются пыльцой в мужских шишках деревьев каури, и предполагаем, что
в раннем мелу они обладали сходными особенностями поведения. И весьма вероятно,
что этот источник белка искали динозавры – так же, как птицы и ящерицы питаются
пыльцой в наше время56. Совпадение
интересов насекомых и динозавров в отношении шишек хвойных растений представляет
собой лишь одно из проявлений антагонизма, который имел место между этими группами.
Динозавры и насекомые конкурировали в нижнем ярусе леса и дальше вниз, и были
самой серьёзной угрозой для молодых деревьев и других новых растений. Насекомые
также питались в верхней части ярусов полога леса и эмергентов, где наносимые
ими повреждения влияли на здоровье и долговечность деревьев, а также на их способность
к воспроизводству.
Хотя известно мало видов насекомых, повреждающих хвощи, в меловой период на
разнообразном собрании этих грубых растениях наверняка пировала целая группа
видов этих существ. Некоторые насекомые, вроде живущих в стеблях жуков и питающихся
листьями гусениц, по-прежнему используют эти примитивные растения, и если у
их предков были сходные повадки, они могли бы досаждать травоядным динозаврам.
На протяжении всего мелового периода деревья гинкго, или «серебряный абрикос»,
были разнообразными и широко распространёнными, и периодическое появление их
пахучих плодов*, похожих на вишню, наверняка привлекало и насекомых, и динозавров.
Вероятно, протоцератопсы и пахицефалозавры никогда не упускали возможности съесть
эти сочные плоды, но они должны были конкурировать с целым рядом видов жуков
и мух.
* Гинкго – голосеменное растение, его «плоды» представляют собой семена с мясистой оболочкой. – прим. перев.
51
Динозавры, которые поедали листья, похожие на гвозди побеги и мужские цветы*
гинкго, должны были соперничать с гусеницами молей и пилильщиков, которые по
ночам пожирали маленькие цветки и листья, оставляя на следующее утро оголённые
ветви. Меловые тли, щитовки, фонарницевые, и ещё длинноклювые цикадоподобные
насекомые Palaeontinidae могли сосать соки из этих растений, заставляя листья
увядать или опадать раньше времени35.
Мелкие жуки, которые пировали на богатых пыльцой мужских цветках, могли уменьшать
репродуктивный потенциал этих деревьев.
Цикадофиты и саговники в меловой период были разнообразными группами, представители
которых варьировали по высоте от карликовых до колоссальных растений. Игуанодоны
и стегозавры, которые беспрепятственно кормились их перистыми листьями, вероятно,
тревожили пластинчатоусых жуков, пожирающих края листьев. Пыльца и семенные
шишки этих растений были бы лакомством, хотя тех видов, на которых растут орехи,
содержащие ядовитые химические вещества, они, возможно, избегали бы – но не
обязательно, поскольку павианы в Африке без проблем справляются с нежелательными
соединениями в питательных семенах саговников57.
Динозавры, несомненно, конкурировали с целой группой насекомых, питающихся мужскими
и женскими шишками, в том числе с личинками мелких долгоносиков, живущими в
семенах. Крахмалистые стволы саговников, бесспорно, давали пищу пахицефалозаврам
и гипсилофодонам, но они должны были соперничать с личинками жуков-усачей и
молей58. Другими насекомыми-конкурентами
на этих растениях, возможно, были сосущие сок тли или щитовки, которые оставляли
на гибнущих листьях слои липких беловатых отходов. И к середине мелового периода
муравьи уже начинали расселять этих потребителей растительного сока по всему
лесу.
Другой многочисленной группой растений, служивших пищей динозаврам, были папоротники.
Они были величиной от карликовых адиантумов и среднеразмерного орляка до высоких
древовидных и семенных папоротников. Фактически, некоторые из древовидных папоротников,
вероятно, росли такими плотными группами, что скрывали собой обедающих динозавров.
Стегозавры, вне всяких сомнений, были весьма охочими до нежных вай этих растений,
но, вероятно, конкурировали с личинками пилильщиков, гусеницами молей, долгоносиками
и личинками галлиц, живущими в камерах вдоль края листа. Возможно, тли, клопы-слепняки,
цикадки и фонарницы соперничали с динозаврами за
* Понятие «цветы» для голосеменного растения некорректно, но в оригинальном тексте употребляется слово “flowers” – прим. перев.
52
этот ресурс. Для динозавров наверняка были деликатесом их мягкие стебли и крахмалистые,
полные мякоти стволы, но и те, и другие места служат местами развития личинок
долгоносиков. Некоторые из этих примитивных долгоносиков по-прежнему существуют,
и один австралийский вид нанёс значительный ущерб древовидным папоротникам,
когда его случайно завезли на Гавайи59.
Многие из меловых папоротников, вероятно, обладали съедобными подземными побегами60,
который являлись деликатесом и для протоцератопсов, и для личинок жуков.
Динозавры, питающиеся хвойными деревьями, сталкивались с целой толпой насекомых-конкурентов.
Можно представить себе, как пахицефалозавры нетерпеливо собирались вокруг араукарий,
когда падали шишки. И мужские, и женские шишки очень питательны, обладают повышенным
содержанием углеводов, белков, жиров и микроэлементов. Здесь динозавры соперничали
с личинками долгоносиков, развивающимися в мужских шишках34,
и с личинками молей (Agathaphigidae) и жуков (Belidae), обитающими в женских
шишках61 (цветная вкладка 12C). В наши
дни гусеницы Agathiphaga могут уничтожить до 95 % семян каури в шишке182.
В них даже могут находиться орехотворки, пожирающие семена62
(цветная вкладка 5B). Диплодоциды и титанозавры, питающиеся листьями молодых
араукариевых, несомненно, делили свою пищу и с минирующими, и с листогрызущими
гусеницами, а также с трипсами, тлями и щитовками (цветные вкладки 3A, 3E, 3F).
Возможно, в это время уже могли существовать предки молей-пестрянок (Gracillariidae),
молодые особи которых минируют листья новозеландских деревьев каури63,
а также предки гигантских личинок долгоносиков, которые развивались в стволах
араукариевых64.
Бахромчатокрылые трипсы могли обитать на почках, цветках, листьях и стеблях
растений всех типов (цветная вкладка 3A). Эти мелкие насекомые обладают ротовым
аппаратом, приспособленным к сосанию, и демонстрируют два основных способа питания:
поверхностный, ограниченный главным образом эпидермальными клетками, и проникающий,
который достигает более глубоких тканей растения. Остающиеся после кормления
проколы, а также надрезы, сделанные для кладки яиц, приводят к механическим
повреждениям растительной ткани, и если насекомые многочисленны, может происходить
опадение листьев. Этот тип повреждений вызывают трипсы, которые нападают на
деревья каури в северо-восточной Австралии.
Другими важными вредителями, вызывающими опадение хвои в янтароносных лесах,
были пилильщики, многие из которых появляются массово. В наши дни широко распространённый
53
красноголовый сосновый пилильщик наносит весьма большой ущерб
соснам, лиственницам, кедрам и елям65.
Если подвергшиеся нападению деревья не погибли непосредственно от этого, повреждение
может тормозить рост и вызывать гибель ветвей. Другими пилильщиками, жившими
на хвойных деревьях, которые существовали в те времена, были листоядные настоящие
пилильщики (Tenthredinidae) и паутинные пилильщики, а также ксиелиды (Xyelidae),
которые размножались в шишках, почках и растущих побегах. Несомненно, эти насекомые
были серьёзными конкурентами динозавров, кормящихся на хвойных деревьях.
И хвойные, и покрытосеменные растения подвергались нападениям таких неспециализированных
едоков, как сверчки, кузнечики, элканиды (Elcanidae), эумастациды, короткоусые
прямокрылые, веты и хаглиды (Haglidae) (цветные вкладки 4A, 4B). По данным летописи
окаменелостей, в мезозое хаглиды были весьма разнообразны, и об их вероятном
поведении можно отчасти судить по тем видам, которые всё ещё обитают в Северной
Америке*. Те особи, которые питаются пыльцой хвойных – это поющие самцы, которые
предлагают за возможность спариться не только песню, но и мясистые складки своих
крыльев в качестве вкусного лакомства. Склонность к кормлению в группе у меловых
кузнечиковидных представителей семейств Locustopseidae и Elcanidae, вероятно,
уничтожила множество растений подобно тому, как орды саранчи начисто съедают
зерновые культуры в наши дни65. А короткоусые
прямокрылые, должно быть, уже тогда нападали на самые разнообразные растения
так же, как они поступают сегодня. Вызванное насекомыми опадение хвои, возможно,
уничтожало множество деревьев, поскольку они не могут отращивать новую хвою
и восстанавливаться, как это делают покрытосеменные66.
Молодые деревья метасеквойи наверняка были ещё одним видом растений, которое
предпочитали динозавры. Когда зауроподы пожирали нежные хвоинки, они наверняка
сталкивались с личинками жуков-листоедов, спрятавшимися между иглами, мучнистыми
червецами, скрывающимися внутри своих белых коконов, похожих на бумажные, и
с мелкими гусеницами, спрятавшимися в кончиках почек. А динозавры, которые выбирали
нижние ветви кипарисов, делили свой обед с мелкими гусеницами, повреждающими
почки, с личинками пилильщиков и щитовками, которых посещали муравьи. Сосущие
растительные соки мезозойские тли, которые являются родственниками современных
тлей семейства хермесов (Adelgidae), были вероятной причиной ослабления и гибели
множества хвойных деревьев в янтароносных лесах.
Когда тсуговый хермес был завезён в восточную часть Северной Америки, он распространился
на трети общей площади лесов, уничтожив на своём пути множество деревьев тсуги67.
Эти питающиеся на ксилеме сосущие насекомые вводят в ткани растения токсины,
заставляющие хвою желтеть и
* Haglidae считаются полностью вымершим семейством. Возможно, речь идёт о родственных семействах, иногда объединяемых с этим и включающих немногочисленные современные виды. – прим. перев.
54
опадать. Это делает растения более восприимчивыми к засухе
и нападению других насекомых и болезней растений. Благодаря сочетанию способности
распространяться при помощи ветра (они весьма лёгкие) и быстрого роста численности
популяций за счёт партеногенетических форм эти насекомые стали причиной массовой
гибели тсуги в Северной Америке. Массивы тсуги гибли примерно через четыре года
после первого нападения, а когда исчезали зрелые деревья, просветы в пологе
леса занимали более быстрорастущие покрытосеменные. Вполне разумно ожидать сходного
сценария в меловых янтароносных лесах.
Загадочный Brachyphyllum, вымершее мезозойское голосеменное растение,
был достаточно широко распространён в меловой период. Динозавры, поедающие листья
и опавшие мужские шишки растений, встретили бы при этом причудливых, стройных
кузнечикоподобных насекомых, а также необычных палочников и странного вида пилильщиков
– у них у всех в пищеварительном тракте открыли остатки листьев и пыльцы Brachyphyllum35.
В 1962 году один энтомолог предположил, что динозавров могли истребить гусеницы,
которые уничтожили их источники корма68.
Динозавры гибли от голода, когда их кормовые растения были съедены ордами личинок
молей. Хотя эта теория делает упор на конкуренции, которая возникала в ходе
потребления листвы насекомыми и травоядными динозаврами, невозможно представить
себе, что её результатом будет вымирание динозавров в том или ином виде.
Питались ли динозавры шляпочными или какими-то другими грибами? Искал ли кто-нибудь
из них трюфели? Сложно сказать, но, если существовали плодоядные динозавры,
у них наверняка был свой любимый вид пищи. Она включала разного рода шляпочные
грибы, трутовики, дождевики и трюфели (цветная вкладка 12A, рис. 12). Многочисленные
и самые разнообразные животные, от грызунов и свиней до оленей и банановых слизней,
и даже черепахи, ящерицы и птицы ищут грибы. Но независимо от того, какого рода
грибам отдаётся предпочтение, пластинчатым грибам, дождевикам, или рогатиковым,
к ним, вне всяких сомнений, прилагался широкий ассортимент насекомых-конкурентов,
включая разнообразных жуков и мух.
55
Хотя в начале мелового периода сформировалось относительно
немного покрытосеменных, к позднему мелу цветковые растения составляли, возможно,
половину всего многообразия растений. Благодаря удивительно высокой скорости
роста и сравнительно быстрому периоду воспроизводства эти растения были обречены
на успех. В их генетических программах была заложена способность к расселению
во множество разнообразных местообитаний, от топей и болот до трещин в камнях,
горных вершин и ветвей деревьев.
В своей книге о динозаврах52 Роберт
Бэккер выразил уверенность в том, что растительноядные динозавры могли «изобрести»
цветковые растения. Он пришёл к такому выводу, основываясь на том, что, в отличие
от позднеюрских листоядных животных, которые кормились листвой в пологе леса
и в лежащих ниже ярусах, меловые динозавры кормились главным образом на земле,
без разбора обстригая представителей флоры почти под корень. Покрытосеменные
растения, которые быстро растут и воспроизводятся, восстанавливаются после такого
объедания начисто быстрее по сравнению с голосеменными, получая, таким образом,
конкурентное преимущество, которое в итоге привело к их господству. Эту теорию,
однако, встретили критически69, и Пол
Барретт и Кэтрин Уиллис опровергли теорию Бэккера на том основании, что покрытосеменные
не были «ни достаточно обильными, ни достаточно широко распространёнными, чтобы
быть основным компонентом рациона динозавров на протяжении раннего мела». Кроме
того, вряд ли многие из меловых динозавров объедали растительность до самой
земли, и проростки как покрытосеменных, так и голосеменных выживали бы. Также
Бэккер сравнивал широко распространённую среди динозавров травоядность с таковой
у млекопитающих, пасущихся на травянистых
56
равнинах, но не существует никаких свидетельств в пользу того,
что такие местообитания возникали когда-либо в меловой период.
Раннемеловые растительноядные насекомые определённо играли важную роль в эволюции
покрытосеменных благодаря тому, что кормились на архаичных голосеменных растениях.
Нанесённые насекомыми повреждения встречаются во всех слоях отложений и обнаруживаются
от корней до кончиков листьев. Начисто объедая листву широко распространённых
саговников, хвойных и папоротников, пожирая заросли хвощей и ковры плаунов,
и избегая трогать покрытосеменные растения, они пропалывали ряды конкурентов,
и их избирательность в питании работало в пользу цветковых растений. Поэтому,
если деятельность травоядных в меловой период способствовала росту разнообразия
цветковых растений, то значительная часть заслуг в этом принадлежит насекомым.
Когда примерно в середине мела на сцене появились насекомые, отдающие предпочтение
покрытосеменным растениям в качестве источника пищи, они начали обгрызать листву
на ветках, бурить стебли и питаться цветами и плодами. Обитающие под землёй
существа вроде термитов, личинок жуков и мух, гусениц и тлей поедали корни и
подземные стебли65. Посредством избирательного
выедания определённых родов и видов покрытосеменных растений меловые насекомые
определяли то, какие родословные линии цветковых растений сохранятся до наших
дней.
Однако иной вид деятельности делает насекомых гораздо более важной по сравнению
с динозаврами движущей силой эволюции покрытосеменных, и он не имел ничего общего
с травоядностью, но всецело относился к воспроизводству растений. Насекомые
стали основными переносчиками генетического материала от одного цветка к другому,
заменяя случайный успех опыления ветром адресным переносом гамет.
57
В тёмном углублении у основания куста папоротника крохотная
пчёлка взвалила на себя трудное бремя материнства, начав постройку гнезда. Вначале
она очистила на земле небольшой участок, пользуясь своими зазубренными жвалами,
чтобы кусочек за кусочком убирать большие частицы почвы, а затем начала усердно
рыть землю передней и средней парами ног. Когда кучка выкопанной почвы стала
выше, занятое работой животное постепенно скрылось внутри тоннеля, пока, наконец,
присутствие перепончатокрылого стало возможно определить лишь по выбрасываемым
наружу частицам почвы. Работая день и ночь, крохотное насекомое построило камеру,
которая должна будет служить детской для её первого потомства. Она добавила
последние штрихи, уплотняя почву в камере кончиком притупленного брюшка.
Теперь, покончив с первой задачей, эта мать должна была приложить все усилия
для выполнения рискованной обязанности по заполнению камеры пыльцой, чтобы вывести
потомство. В тусклом свете наступающего утра одиночная пчела подготовилась к
полёту. Перед тем, как совершить первую вылазку из гнезда, она взлетела и сделала
три или четыре круга над этим местом, чтобы запомнить его расположение, а затем
стремительно полетела через лес. Пролетая извилистым маршрутом между огромными
деревьями араукарий, стволы которых поблёскивали, все в пятнах смолы, и облетая
свисающие лианы, крохотное насекомое заметило какие-то мелкие причудливые белые
цветки, торчащие среди пучков голубовато-зелёных листьев.
Множество разнообразных лазающих и вьющихся в воздухе насекомых уже было занято
кормлением на цветках. Жуки с металлическим блеском и похожими на расчёску усиками
пожирали пыльцу, длинноногие мухи в это время слизывали нектар, а крохотные
трипсы с поблёскивающими серебристыми крыльями соскабливали ткани с лепестков.
Избегая их соседства, маленькая пчела опустилась на группу недавно распустившихся
цветов и начала нагружать как можно больше пыльцы
58
на жёсткие волоски на своих задних ногах. Даже при полной
загрузке ей всё равно придётся сделать ещё пять или шесть вылетов, прежде чем
она наберёт достаточно пыльцы, чтобы выпестовать всего одну личинку до взрослого
состояния. Прибыв обратно в гнездо, самка быстро забралась в камеру для потомства
и начала соскабливать пыльцу. Улетая, она перекрыла вход в гнездо, частично
закрыв входное отверстие пробкой из грязи, чтобы помешать каким-нибудь вороватым
насекомым, которые могли искать даровую еду, а затем разложила поверх него немного
мусора в качестве камуфляжа. Затем она полетела обратно в лес, чтобы найти ещё
цветы.
На сей раз её путешествие привело её на поляну, где уже стоял шум от бурной
деятельности, но на сей раз не только насекомых. В подлеске паслись цератопсы,
поедая всё на своём пути, словно лесорубы, занятые прокладыванием просеки. Обрывая
без разбора папоротник, саговники, хвощи, низкорослые хвойные и кустарниковые
покрытосеменные растения, гигантские травоядные пожирали травы и кустарник,
которые отросли в этих местах после их прошлого визита.
Явно не имея здесь конкурентов, перепончатокрылое перелетало с цветка на цветок,
собирая пыльцу и останавливаясь время от времени, чтобы глотнуть нектара. Её
мохнатое тело было покрыто пыльцой и при каждом посещении очередного цветка
некоторые из этих зёрен оставались на женских частях цветка. Простой акт перекрёстного
оплодотворения гарантировал, что завяжутся семена и появится новое поколение.
Это был дар, полученный в обмен на пыльцу и нектар, которые были нужны пчеле
для продолжения рода.
Загрузив задние ноги до отказа, трудолюбивая пчела возвратилась к гнезду. Теперь,
когда был собран достаточный запас пищи для одного потомка, к пыльцевым зёрнам
было добавлено немного нектара, и смеси была придана форма небольшого шарика.
Этот источник корма был заботливо уложен так, чтобы не касаться сочащихся влагой
земляных стенок, поскольку поражение плесенью могло сделать его несъедобным
для молодняка. Удовлетворившись результатом, самка развернулась и отложила жемчужно-белое
яйцо на вершину массы пищи, а затем торопливо выбралась задом наперёд из камеры
и надёжно закрыла вход, утрамбовав поверх отверстия слой частиц почвы.
Самым существенным видом помощи насекомых в процессе закрепления и распространения цветковых растений была их деятельность, связанная с опылением – то, чего не могли сделать динозавры. Большинство
59
ранних растений были ветро- и водоопыляемыми, и ни динозавры,
ни насекомые не принимали активного участия в этом процессе. Тем не менее, это
не препятствовало некоторому числу насекомых, от жуков и трипсов до мух и ос,
питаться пыльцой саговников, цикадеоидей и хвойных. Эти древние ассоциации могли
быть сугубо односторонними, когда насекомые просто поедали пыльцу и улетали
прочь, но в то же самое время в некоторых из них пыльца могла случайно переноситься
с одного растения на другое. Мы знаем, что кузнечики и пилильщики поедали пыльцу
мезозойских голосеменных, потому что они сохранились с её зёрнами в своих кишках70.
Насекомые, которые обычно питались пыльцой некоторых растений, возможно, превратились
в главных, или даже единственных опылителей этих видов, когда ассоциации стали
более устойчивыми.
К середине мела многие из питающихся пыльцой насекомых добавили покрытосеменные
растения в свои списки кормовых растений. Хотя доля насекомоопыляемых цветковых
растений середины мела могла и не достигать тех же примерно 70%, как в наше
время, этот ресурс наверняка использовали в пищу многие жуки, мухи, осы и трипсы71.
Хотя многие из этих насекомых, посещающих цветки, были лишь случайными опылителями,
некоторые растения, несомненно, зависели от них. Вероятно, предки белид*, долготелов,
долгоносиков-молитин, а также жуков-ящериц (Languriidae), которые опыляют саговники
в настоящее время, уже в меловой период приняли на себя эту роль48,72.
Привлекая этих жуков, растение предлагало им защиту, пищу, место для спаривания
и для развития личинок. Долгоносики-белиды поедают богатые крахмалом структуры
(спорофиллы), которые поддерживают сами пыльцевые мешки, тогда как жуки-ящерицы
едят саму пыльцу. Таким образом, поделив между собой источники пищи, они могут
сосуществовать на одном и том же растении. Саговники других типов опыляются
мелкими долгоносиками подсемейства молитин, у которых взрослые особи кормятся
пыльцой, а личинки поедают крахмалистые стебли.
Другие голосеменные из янтароносных лесов, вероятно, частично опылялись долгоносиками:
цветожилами, белидами, каридами* и долготелами, которые в наши дни связаны с
араукариями, кипарисами и подокарпами. Считалось, что опылителями могли быть
даже жуки-листоеды, развивавшиеся стволах цикадофитов35.
Вполне возможно, что долгоносик-цветожил из ливанского янтаря мог опы-
* Автор упоминает белид, говоря о долгоносиках. Тем не менее, этих жуков часто считают самостоятельным семейством Belidae, родственным настоящим долгоносикам (Curculionidae). То же самое относится к жукам семейства карид (Caridae). – прим. перев.
60
лять смолоносное дерево, поскольку известно, что в настоящее
время они развиваются в мужских шишках араукарий и других хвойных34.
Существующие в наши дни ассоциации, отмеченные для долгоносиков и пальм, возможно,
начали своё существование в меловой период. Огромные количества африканских
долгоносиков из подсемейства Derelominae собираются для кормления, спаривания
и кладки яиц на мужских цветках масличной пальмы. Там они собирают, а затем
переносят на женские цветки липкие пыльцевые зёрна. Этот коэволюционный тип
поведения установился настолько прочно, что у клещей и нематод, кормящихся на
цветках пальмы, возникла зависимость от жуков, переносящих их с растения на
растение73. Если долгоносики внезапно
исчезнут, то же самое случится с клещами, нематодами, и, возможно, с пальмами.
По мере возрастания многообразия покрытосеменных та же тенденция наблюдалась
и у насекомых-опылителей. Ранние покрытосеменные конкурировали не только с голосеменными,
но также и друг с другом – за возможность выиграть внимание потенциальных опылителей.
В целях привлечения насекомых у них появлялась всё более вкусная награда, а
также подсказки для зрения и других чувств в виде привлекательных красок и ароматов.
И по мере того, как примитивные моли, мухи, осы и пчёлы становились всё более
успешными в своей роли, всё больший процент цветов оказывался оплодотворённым
и всё большее количество семян завязывалось и распространялось, гарантируя тем
самым, что при появлении новых доступных местообитаний быстрорастущие покрытосеменные
могли колонизировать их раньше, чем голосеменные.
Самые надёжные опылители – это те, кто использует данный ресурс, чтобы создать
запас белка при выкармливании потомства. Они постоянно посещают один за другим
множество цветков, оставляя пыльцу на рыльцах. Иметь курьера, разносящего пыльцу,
даже если частью её придётся пожертвовать в качестве пищи – это, конечно же,
более прогрессивный метод, чем рассеивать массы пыльцы и рассчитывать на то,
что ветер доставит её в нужное место.
Вероятно, первыми опылителями на постоянной основе были осы, которые заменили
для своего потомства добычу, состоящую из членистоногих, растительным белком.
Эти осы могли переносить пыльцу на различных частях своего тела – возможно,
на волосках на своей голове, поскольку именно так современная оса носит пыльцу
для своей молоди74. В действительности
метод переноски, возможно, зависел от размера и свойств пыльцы (клейкая или
сухая, слипающаяся в комки или сыпучая, и т. д.).
61
В какой-то момент в меловой период перепончатокрылые выработали
морфологические особенности, облегчавшие сбор пыльцы, наиболее существенной
из которых были разветвлённые волоски. Такие перистые волоски служили сетью
для пыльцы, позволяя удерживать её зёрна, и делали значительно проще её доставку
к местам жизни личинок. В наше время перистые волоски имеются у всех пчёл, и
переход от осы к пчеле, если судить по данному признаку, явно произошёл в раннем
мелу. На основании молекулярных исследований предполагается, что время появления
пчёл – около 125 миллионов лет назад75.
Эти ранние «протопчёлы», напоминающие экземпляр из бирманского янтаря44,
всё ещё сохраняли некоторые особенности ос (цветная вкладка 14B).
Меловые осы и примитивные пчёлы проводили значительную часть времени, посещая
и опыляя цветы, и растения отвечали на это, делая свои цветы более привлекательными.
Так появились броские и ярко окрашенные лепестки, ароматы, масла и нектарники;
последние располагались не только на самом лепестке или в его основании, но
и на цветоносах, стеблях и листьях. Такие особенности словно сигнализировали
и манили к себе, как ярмарочные зазывалы: «Иди сюда и загляни сначала ко мне».
В настоящее время пчёлы, которых существуют тысячи видов, являются важнейшими
в мире опылителями, и все они в своём выживании зависят от покрытосеменных растений.
Большинство пчёл – одиночные виды, каким был и меловой Melittosphex,
и они собирали пыльцу с конкретных растений или групп растений, или с растений
в определённых местообитаниях76,77.
Они зачастую занимают территорию для проживания внутри строго определённой экозоны
– такой, как торфяники, дюны, луга или пустоши. Выполнение всех задач зависит
от единственной самки, которая собирает пыльцу, уносит её в гнездо, сделанное
в земле или разрушающейся древесине, создаёт запас пыльцевых зёрен в камерах,
помещает в каждую камеру яйцо, а затем запечатывает камеры. Из каждого яйца
развивается личинка, пища для которой ограничена лишь тем, что запасено для
неё, и этого количества еды обычно хватает как раз чтобы достичь взрослого состояния.
Когда именно в прошлом возникли общественные пчёлы, сказать сложно, потому что,
хотя ранее сделанные сообщения о меловой общественной пчеле (Cretotrigona)
были признаны ошибочными78, примитивные
безжалые медоносные пчёлы могли возникнуть в процессе эволюции в позднем мелу.
Их хорошо развитые общества и потребность в нектаре и пыльце
62
могли сделать их успешными опылителями цветковых растений –
такими же, какими они являются в более тёплых областях мира в наши дни79.
Новые насекомые-опылители, появившиеся ближе к середине мелового периода, определённо
дали огромное преимущество покрытосеменным растениям. Это означало, что цветам
не нужно было производить пыльцу в огромных количествах, чтобы её разносили
потоки ветра. Малое количество пыльцы, доставленное надёжным насекомым-распространителем,
которое только что посетило цветок того же самого типа, было гораздо более успешной
системой, и Melittosphex – это доказательство того, что это случилось
около 100 миллионов лет назад44.
Только какое влияние могли бы оказать эти опылители на динозавров? Если ближе
к средине мела покрытосеменные растения были редкими и имели ограниченное распространение,
о чём уже было сказано69, то динозавры
могли кормиться главным образом голосеменными. Увеличивая разнообразие покрытосеменных
благодаря опылению, насекомые могли дать покрытосеменным растениям преимущество
в конкуренции с голосеменными, и динозавры могли пострадать от этого. Однако
в позднем мелу, когда по крайней мере некоторые динозавры питались покрытосеменными,
насекомые-опылители помогали им, став залогом ещё большего разнообразия и распространения
цветковых растений. Насекомые-опылители были своего рода краеугольным камнем:
они приносили пользу не только покрытосеменным растениям, но и всем растительноядным
насекомым и позвоночным, которые зависели от этих растений в плане питания,
а также хищникам и паразитам, которые в своём выживании всецело зависели от
травоядных.
63
Птерозавры, парящие над обширным араукариевым лесом, проносились
над полосой бронзового цвета, которая прорезала зелёный фон окружающего пейзажа.
Хвойные деревья, составляющие этот красновато-коричневый участок, стояли, лишённые
хвои, а их изящные ветви стали жёсткими и ломкими. По всей границе этого бросающегося
в глаза кладбища деревьев лес был заражён, и ему была уготована та же самая
судьба: листва деревьев уже окрашивалась в различные оттенки жёлтого и рыжего.
Маленькая самка жучка-короеда карабкалась по стволу пока ещё здорового дерева
каури в поисках места, где можно прогрызть кору и построить галереи для своего
потомства. Она проползала мимо жуков того же вида, которые с таким же рвением
были заняты поисками, но теперь задыхались в капельках смолы, излитой деревом
через пробуренные ими отверстия. Это был основной способ защиты дерева от таких
нападений. Но, как и многие другие, эта самка жука в итоге добилась успеха,
и каждое удачное завершение миссии отмечалось древесной пылью, разлетающейся
из маленького входного отверстия в коре.
Как только самка добралась до нежных растительных клеток под корой, она начала
вгрызаться в древесину, инстинктивно строя длинные, узкие тоннели, окаймлённые
пунктирной линией боковых галерей для яиц. Древесные опилки и буровая мука,
образовавшиеся в ходе строительства, сгребались назад в проходы или выбрасывались
наружу сквозь маленькие дырочки в коре. Ночью в воздухе стоял хруст, который
производили тысячи таких жуков, одновременно грызущих кору и древесину. После
окончания постройки главной галереи самка жука соорудила несколько боковых галерей
для яиц, в которые было отложено
64
несколько маленьких белых яиц. Пока она грызла дерево,
с её тела сваливались споры крохотных симбиотических грибков. За несколько дней
грибок вырастил гифы на поверхности всех туннелей и обеспечил молодняк питанием.
Пожирая пронизанную грибком древесину, безногие личинки быстро росли, а когда
их развитие почти завершилось, они построили под самой корой расширенные камеры
для окукливания. Жизнь в этих галереях, отрезанных от мира хищников снаружи,
оказалась безопасной и защищённой. Но другие жуки таких же форм и размеров,
как сами короеды, забрались в некоторые из тоннелей и съели много находящейся
там молоди на разных стадиях развития.
Те куколки, которые избежали нападения разбойников, превратились в мягких желтовато-коричневых
взрослых особей и ждали, пока затвердеет их экзоскелет, прежде чем прогрызть
выход из места своего проживания. Выбираясь наружу, они несли в щелях и бороздках
на своём теле споры симбиотических грибков, которые были так необходимы для
их развития. Грибковая инфекция, которая осталась после них в покинутых жилищах,
вызывала появление синей гнили древесины по мере того, как болезнь прогрессировала
и постепенно распространялась по всем тканям дерева, обрекая его на незавидную
судьбу. Листья поражённого дерева сначала желтели, а затем медленно увядали
и отмирали, пока не оставался лишь безжизненный ствол.
В другой части леса самка рогохвоста готовилась отложить яйца в ствол хвойного
дерева. Она использовала пилообразный яйцеклад, чтобы прорезать кору, а затем
отложила около дюжины кремово-белых яиц в глубокую щель, где они были бы защищены
от хищников. Во время кладки яиц были также высвобождены споры и фрагменты грибницы,
хранящиеся в мешочках в основании её брюшка. Выживание личинок рогохвостов зависело
от роста этого симбионта, который, по иронии судьбы, в итоге уничтожит дерево.
В течение следующих нескольких месяцев, пока последующие поколения рогохвостов
будут распространять болезнь, пожелтеют, побуреют и погибнут обширные массивы
хвойных.
Тем временем близ русел рек пальмы сбросили недозрелые плоды. Цветки почернели
и опали с цветоносов, а в это время их листья, начиная с самых нижних, один
за другим желтели, а затем бурели. Мелкие разноцветные цикадки, питаясь на больных
65
растениях, собирали в себе микроскопических возбудителей, которые были ответственны за возникновение этой вирусной болезни, и во время следующих кормлений невольно распространяли их ещё дальше и шире. Эти болезни растений были неотъемлемой частью жизненного цикла леса.
Меловой период был миром плесени, не слишком сильно отличаясь
от тропических областей нашего времени. Грибы паразитировали на других грибах,
а те, в свою очередь, были паразитами третьих345.
Пожив в тропиках, мы знаем, каково это – обнаружить массы длинных серых нитей,
высовывающиеся из обуви, оставленной на несколько дней в туалете, или когда
пятна распространяются по разным частям твоей кожи, когда споры забивают твою
дыхательную систему, а тонкие тяжи оставляют царапины на поверхности линз микроскопа.
Так вышло, что практически все микроскопы в нашей западноафриканской лаборатории
оказались бесполезными из-за вызванных грибками царапин на поверхностях линз,
которые делали изображение размытым. И особенно обидно то, что было невозможно
просто почистить линзы: их нужно было заново отшлифовать, прежде чем они вновь
станут рабочими – вовсе не выход в африканских полевых условиях!
Диапазон местообитаний, которые могут занимать грибы, поистине удивителен. Сделанное
наугад микроскопическое исследование листьев и стеблей растений, подобранных
в лесу мелового периода, выявило бы значительное многообразие грибов. Коричневого
цвета грибы, вызывающие пятнистость листьев, испещряли верхнюю сторону листьев
деревьев семейства араукариевых, а скопления спор ржавого цвета в это же время
окрашивали нижнюю. Веточки этих преобладающих в лесу деревьев были покрыты тяжами
белых грибков, а плесень иного рода въедалась в стебли, вызывая появление обширных
бесформенных язв, сквозь которые проникали древоразрушающие грибы других типов.
Даже молодые сеянцы страдали от грибов, вызывающих «чёрную ножку», которые разрушали
их корни ещё до того, как у них появлялся шанс достичь одного фута в высоту.
Если бы у нас была возможность совершить путешествие в одно и то же место в
меловом лесу до и после того, как грибковые болезни пройдут свой цикл развития,
мы сразу заметили бы отличия в облике растительного сообщества, проявившиеся
в результате гибели самых восприимчивых видов и взрыва численности более стойких
типов.
Насекомые, вне всяких сомнений, оказывали огромное воздействие на состав флоры,
66
перенося патогенные организмы, нападавшие на кормовые растения
динозавров. Суди по имеющимся у нас знаниям, грибные и вирусные заболевания,
переносимые насекомыми, были критически важны при выяснении того, какие растения
будут жить, а какие погибнут в мире мелового периода. Поскольку бактерии и простейшие
инфицируют лишь немногие растения, эти организмы не выработали таких обширных
патогенных ассоциаций с растениями, какие наблюдаются у грибов и вирусов.
Короеды – это мелкие тёмные насекомые размером с булавочную головку, но, будучи
вооружёнными своими симбиотическими грибами, они становятся предвестниками смерти
и разрушения. Эти насекомые прогрызают наружный слой коры вечнозелёных и листопадных
деревьев, чтобы строить выводковые галереи в заболони65,80.
К сожалению те же самые грибы, что кормят жуков, также убивают дерево-хозяина,
закупоривая его водопроводящие клетки81.
Несомненно, в меловой период араукариевые, подокарпы, кипарисы и Metasequoia
были столь же подвержены нападению жуков-короедов, как и в наши дни82,
и некоторые полагают, что араукариевые мелового периода были первыми деревьями,
на которые нападали жуки-короеды83.
Разрушительное воздействие грибных заболеваний, которые переносят эти жуки,
стало очевидным в течение прошлого столетия, когда в Северную Америку попали
два патогенных организма. Одним из них была печально известная голландская болезнь
вязов, которая появилась в 1930 году и за пятьдесят лет распространилась от
побережья до побережья. Успешное распространение смертельно опасного грибка
двумя видами жуков-короедов, по сути, уничтожило американский вяз в Северной
Америке повсеместно84.
Вторым примером был рак коры каштанов. Завезённая в Соединённые Штаты из Азии
около 1900 года, болезнь в течение пятидесяти лет уничтожила более 99% из нескольких
миллиардов деревьев американского каштана* в восточных лесах. Азиатские каштаны
были восприимчивы к инфекции, но не гибли от неё, тогда как американские каштаны,
никогда не сталкивавшиеся с болезнью, не имели никакой устойчивости к ней. Это
считается одним из самых разрушительных, если не самым разрушительным из известных
случаев заболевания растений. Бурящие древесину насекомые, в том числе жуки-усачи,
жуки-горбатки (шипоноски) (цветная вкладка 5D) и жуки-короеды способствовали
распространению этого болезнетворного организма85.
Когда именно жуки-короеды вступили в симбиотические отношения с грибами – это
интересный вопрос. Ископаемые жуки-короеды (Scolytidae) встречаются на протяжении
всего мела и, вероятно, зави-
* Также известен как каштан зубчатый (Castanea dentata). – прим. перев.
67
симость от грибов возникла на ранних этапах их существования.
Многие из ныне живущих короедов и плоскоходов (Platypodinae) переносят споры
и фрагменты грибницы этих грибов в специальных мешочках (микангиях) на разных
участках своего тела86.
Другой гибельный дуэт, который, вероятно, сложился в меловой период, составляют
рогохвост и его симбиотический гриб. Хотя большинство рогохвостов размножается
в мёртвой древесине, Sirex noctilio выбирает в качестве места размножения
живые сосны. В Европе, где это насекомое является местным видом, сосны приспособлены
и обычно не гибнут от заражения. Однако сосна лучистая из Калифорнии, которую
новозеландцы и австралийцы выращивают в промышленных масштабах ради древесины,
сильно страдает от него. Небольшие природные прибрежные популяции сосны лучистой
жили в благословенной изоляции от рогохвостов, но, когда они встретили этот
дуэт на австралийских плантациях, наступил хаос. В течение десяти лет погибло
4,8 млн. деревьев сосны лучистой, и, если бы не успешная программа биологического
контроля, рогохвост и грибок уничтожили бы сосну лучистую на этом континенте87.
Весьма вероятно, что рогохвосты мелового периода распространяли симбиотические
грибки для выкармливания своего потомства.
Жуки-усачи, разносящие сосновую стволовую нематоду и грибки, показывают, насколько
сложными и смертоносными могут быть союзы насекомого и возбудителя болезней
растений. Некоторые виды жуков-усачей, когда откладывают яйца заражают ствол
сосны и нематодой, и грибком. Молодь жука и нематода вместе поедают заражённую
грибком древесину, но нематоды быстро размножаются и закупоривают водопроводящую
систему деревьев. Заражённые деревья умирают в течение 30-40 дней после проявления
первых признаков заболевания. Эта болезнь дала очень серьёзную вспышку в Японии
в 1940-х годах, вызывая ежегодные потери примерно 400 000 кубометров древесины
в течение последующих 25 лет88. Местные
сосны Тунберга сильно восприимчивы к ней, тогда как сосны из Северной Америки,
где, возможно, возникла болезнь, обладают определённой устойчивостью к ней.
Жуки-усачи существовали в меловой период, а нематоды напоминают обитавшие в
древесине сосен виды, находимые в ливанском и бирманском янтаре.
Другой смертоносной группой паразитических грибов являются ржавчинные грибы,
которым служат переносчиками жуки и мухи. Пузырчатая ржавчина веймутовой сосны
является
68
одной из самых серьёзных болезней леса в Северной Америке,
и без контроля над возбудителем болезни было почти невозможно вырастить сосны89.
Пусть наши веймутовы сосны были спасены благодаря фунгицидам и отбору устойчивых
к болезни деревьев, но смогли бы химические вещества естественного происхождения
спасти хвойные деревья мелового периода от такого рода патогенных организмов?
Мы можем лишь догадываться о том, насколько серьёзную опасность представляли
новые разновидности ржавчинных грибов для архаичных голосеменных. Современным
ржавчинным грибам для завершения жизненного цикла требуются папоротники и ели,
и подобного рода ассоциации между папоротниками и хвойными, несомненно, существовали
в меловой период90.
Деревья каури подвержены поражению множеством грибов182,
и мы предполагаем, что грибы сходного типа заселяли меловые араукарии. Мы знаем
что на коре араукариевых росли грибы различных типов91,345
из бирманского янтаря (цветные вкладки 12A, 12D). Некоторые из этих видов, вроде
гриба спарассиса (Sparassis), вызывают инфекции корней и стволов у
современных хвойных92, тогда как другие
патогенны для покрытосеменных растений93.
Насекомые, обнаруженные в янтаре вместе с древним рогатиковым грибом, возможно,
распространяли споры так же, как в наши дни жуки переносят споры грибов внутри
своего кишечника94. Если рогатиковый
гриб из бирманского янтаря был патогенным, он наверняка оказывал воздействие,
как минимум, на одно из основных кормовых растений динозавров.
Комбинация из крайне восприимчивых к болезни деревьев, безотказно работающих
насекомых-переносчиков и вновь возникающих патогенов создаёт стимул для уничтожения
популяций растений, и, если грибы устанавливали новые симбиотические ассоциации
с меловыми насекомыми, то сценарии, подобные вышеописанным, возможно, были распространены
довольно широко.
Теперь обратим внимание на насекомых, переносивших вирусы растений и микоплазмы
(мелкие одноклеточные организмы) в меловой период. Хвоя метасеквойи пожелтела,
листья папоротника испещрил мозаичный узор из пятен, или на листьях араукарии
появились кольцевые пятна? Пейзаж усеян чахлыми саговниками или древовидные
папоротники демонстрируют съёжившиеся вайи? Какие-то примитивные покрытосеменные
растения внезапно пожелтели и завяли? Если это так, то причиной этого могли
быть вирусные инфекции89.
Тли, цикадки, фонарницевые, белокрылки, а фактически – все
69
сосущие растительный сок, и даже некоторые кровососущие насекомые,
жившие в янтароносных лесах, обладали потенциальными возможностями переноса
вирусов растений (цветные вкладки 2 и 3). Предполагают, что ассоциации между
вирусами, насекомыми и растениями возникли в ходе коэволюции примерно 200 миллионов
лет назад, задолго до формирования нашего самого раннего янтарного местонахождения
в Ливане95.
Меловые тли были весьма разнообразными, и, памятуя о том, что в наши дни они
переносят более трёхсот вирусов95,
мы предполагаем, что они передавали этих возбудителей болезней уже в те времена
(цветные вкладки 2C, 3E). Некоторые из самых ранних вирусов, переносимых тлями,
могли поражать папоротники96. Когда
инфицированные вирусом тли завершают акт кормления и извлекают свои длинные
и тонкие ротовые органы из тканей растения, они закрывают место кормления пробкой
из слюны, заражённой этими патогенами95.
Эти вирусы могут убить их растение-хозяина, и, хотя те из них, которые поражали
хвойные и другие голосеменные растения, были исследованы лишь вскользь, они,
вероятно, возникли в этих группах растений в меловой период. И поскольку переносчиками
этих инфекционных агентов стали тли, их перенос на покрытосеменные растения
был лишь вопросом времени. Пальмы – это группа ранних покрытосеменных растений,
которые, вероятно, поражались вирусами, возможно принадлежащими к той же самой
группе (потивирусы), которую тли переносят на африканскую масличную пальму97.
Если основываться на наших современных знаниях, весьма вероятно, что переносимые
насекомыми вирусы играли большую роль в формировании облика мира покрытосеменных
растений.
Цикадки и фонарницевые (цветные вкладки 2A, 2B, 3D, 3F) – это ещё две группы
насекомых из янтарей мелового возраста, которые известны как распространители
вирусов растений и микоплазм. Известно, что существует более 130 видов цикадок
из 8 подсемейств, которые в настоящее время передают этих возбудителей заболеваний,
размножающихся как в растениях, так и в насекомых95.
Многие из ныне существующих вирусов, переносимых фонарницами, поражают травы95,
поэтому некоторые из представителей этих насекомых из бирманского янтаря, возможно,
распространяли вирусы среди ранних представителей бамбуков, а также среди других
цветковых растений, произраставших в те времена17,18,348.
Мучнистые червецы, встречавшиеся на протяжении всего мелового периода98,99,
в настоящее время широко распространены в тропиках и субтропиках (цветная вкладка
3F). Эти мелкие сосущие насекомые могут убивать растения просто своими ядовитыми
выделениями, но многие из них переносят вирусы100.
Хотя некоторые предположили, что
70
мезозойские щитовки жили только на голосеменных98,
и потому могли вводить вирусы в саговники, кейтонии и хвойные, многие из них
наверняка начали питаться на покрытосеменных растениях, а также передавать этим
растениям вирусы. Состояние, известное как болезнь вздутых побегов какао, возникло,
когда дерево какао было завезено в Африку со своей родины – из Южной Америки.
Мучнистый червец перенёс вирус с туземных африканских растений на вновь завезённые
растения какао, и от этой новой болезни погибло или сильно пострадало множество
деревьев. Это ещё один пример того, как растение может быть уничтожено переносимым
насекомыми вирусом.
Белокрылки, представители семейства Aleyrodidae, распространены повсеместно
в тропиках и субтропиках и были хорошо представлены в янтароносных лесах. Они
довольно мелкие, достигают всего лишь примерно 3 мм в длину, и их можно узнать
по белому порошкообразному налёту на крыльях (цветная вкладка 3B). Неполовозрелые
особи, плоские существа овальной формы, обычно держатся на нижней стороне листьев,
где их длинные стилеты высасывают соки из растения-хозяина. Когда численность
популяций высока, не только их питание наносит ущерб листьям и тормозит рост,
но ещё и липкая падь, выделяемая на листья, закупоривает дыхательные поры растения
и служит питанием для сажистых грибков, которые мешают фотосинтезу. Белокрылки
переносят вирусы покрытосеменных и голосеменных растений101,
вызывая появление жёлтого мозаичного узора на поверхности листьев и жилок, а
также разного рода деформации листьев. Меловые белокрылки, вероятно, распространяли
вирусы, которые поражали папоротники, гинкго, хвойные и покрытосеменные растения.
Известно, что шесть вирусных заболеваний экономически важных тропических растений
переносятся насекомыми102. Эта болезнь
вздутых побегов какао, вирусная мозаика африканской кассавы, вирус полосатости
южноафриканской кукурузы, жёлтая крапчатость на кенийском и филиппинском рисе
и «верхушечный пучок» банана в Юго-Восточной Азии. Все они переносятся различными
цикадками, жуками-листоедами, белокрылками, мучнистыми червецами и тлями. Все
эти группы насекомых уже существовали в меловой период, и некоторые из них наверняка
распространяли вирусы как среди голосеменных, так и среди цветковых растений.
Интересно, что самые разрушительные болезни растений возникают, когда в установившихся
экосистемах появляются экзотические возбудители болезней, или когда заве-
71
зённые растения приобретают инфекции от заражённых эндемичных
видов. В обоих случаях ключевую роль в распространении болезней играют местные
насекомые. Вспышки болезней, вне всякий сомнений, происходили в янтароносном
лесу, когда сосущие насекомые только начинали образовывать ассоциации с вирусами
растений. Восприимчивость меловых растений к вирусным агентам могла повлиять
на состав растительного сообщества.
Итак, около 100 млн. лет назад переносимые насекомыми возбудители заболеваний
растений уже наверняка прореживали ряды некоторых архаичных голосеменных, от
которых динозавры зависели как от источника пищи, таким образом открывая новые
возможности для появления покрытосеменных. Лежит ли на переносимых животными
возбудителях болезней растений ответственность за исчезновение древних араукарий
из северного полушария? Большинство видов араукарий в наши дни произрастает
в нескольких реликтовых областях в Южной Америке, в северо-восточной Австралии,
на Северном острове Новой Зеландии, в Индонезии и на некоторых отдалённых островах
вроде Новой Каледонии и Фиджи. Избежали ли эти островные популяции болезней,
переносимых насекомыми, которые уничтожили это семейство деревьев в северном
полушарии? Новая Каледония – это изолированная, отдалённая территория, однако
она является домом для 5 видов Agathis и 13 видов Araucaria,
что составляет 45% от общемирового видового разнообразия этого семейства. Трагично
сознавать, что после выживания в течение целых геологических эпох в великолепной
изоляции эти новокаледонские популяции оказались в значительной степени уничтоженными
из-за разрушения среды обитания, и более 80% видов в настоящее время находится
под серьёзной угрозой исчезновения103.
Исчезновение к концу мелового периода целых групп голосеменных и покрытосеменных
растений могло принести серьёзные проблемы тем динозаврам, которые зависели
от них как от источника питания. Это лишь один из многих фундаментальных примеров
того, как мелкие и на первый взгляд незначительные насекомые оказывали опосредованное
воздействие на динозавров, перенося возбудителей заболеваний растений, которые
истребляли источники их пищи.
| Предисловие | ix | |
| Благодарности | xi | |
| Введение | 1 | |
| 1. | Окаменелости: капсула времени | 7 |
| 2. | Меловой период: эпоха перемен | 17 |
| 3. | Травоядность | 37 |
| 4. | Динозавры, конкурирующие с насекомыми | 50 |
| 5. | Кто «изобрёл» цветковые растения – динозавры или насекомые? | 55 |
| 6. | Опыление | 57 |
| 7. | Инфекционное увядание и другие болезни меловых растений | 63 |
| 8. | Меловой период: время химер и других странностей | 72 |
| 9. | Санитарные службы мелового периода | 79 |
| 10. | Явление энтомофагии среди динозавров | 91 |
| 11. | Пожирающие динозавров | 102 |
| 12. | Мокрецы | 110 |
| 13. | Москиты | 116 |
| 14. | Комары | 122 |
| 15. | Мошки | 127 |
| 16. | Слепни и златоглазики | 131 |
| 17. | Блохи и вши | 135 |
| 18. | 141 | |
| 19. | Паразитические черви | 147 |
| 20. | Открытие болезней мелового периода | 157 |
| 21. | Болезни и эволюция их возбудителей | 171 |
| 22. | Насекомые: абсолютные чемпионы по выживанию | 185 |
| 23. | Вымирание и мел-палеогеновая граница | 192 |
| ПРИЛОЖЕНИЕ A: Насекомые мелового периода | 203 | |
| ПРИЛОЖЕНИЕ B: Ключевые факторы, способствующие выживанию наземных животных | 219 | |
| ПРИЛОЖЕНИЕ C: Проблемы, возникающие при оценке летописи окаменелостей и вымирания | 221 | |
| Ссылки | 225 | |
| Предметный указатель | 253 |