«Зоология и ботаника остаются все еще собирающими факты науками, пока сюда не присоединяется палеонтология — Кювье,— а вскоре затем открытие клетки и развитие органической химии. Благодаря этому сделались возможными сравнительная морфология и сравнительная физиология, и с тех пор обе стали подлинными науками»*.
Ф. Энгельс
Глава XI
КАК ПРОИЗОШЛИ ОТДЕЛЬНЫЕ ГРУППЫ
РАСТЕНИЙ
1. ПРОИСХОЖДЕНИЕ БАКТЕРИЙ
| * Ф. Энгельс. Диалектика природы. М., Госполитизиат. 1952, стр. 146. |
Мы уже говорили, что среди бактерий можно указать такие, которые организованы
проще, чем какое бы то ни было другое живое существо. Они так мелки, что
проходят через тончайшие фильтры, невидимы или почти невидимы при самых
сильных увеличениях
и проявляют себя только способностью вызывать вокруг себя определенные
химические превращения и, кроме того, способностью размножаться с большой
быстротой.
Те из бактерий, которые при этом не нуждаются для своего питания ни в каких
других животных и растениях и существуют за счет химических реакций, вызываемых
ими в растворах газообразных и минеральных веществ, называются хемотрофными.
Они-то в настоящее время и вызывают в нашем уме представление о первичном
их зарождении на Земле, еще лишенной условий, необходимых для жизни обычных
растений
и каких бы то ни было животных. Последние безусловно нуждаются в наличии
вокруг них каких-то других организмов, вырабатывающих углеводы, жиры и
белки, составляющие
их пищу.
Палеонтология не противоречит нашим теоретическим представлениям в этом
вопросе. Один из наиболее древних остатков живых существ — это так называемые
железобактерии
железных руд в верхнеальгонских слоях Северной Америки. Железобактерии
и в настоящее время живут в ключах, вытекающих из земли и содержащих в
растворе
значительное количество солей закиси железа. Окисляя эти соли в соли окиси
и отлагая, как отработанный материал, вокруг себя гидрат окиси железа,
эти бактерии понемногу образуют значительные скопления железа и могут накопить
достаточное его количество для образования железных руд.
Альгонские слои относятся к более новым кристаллическим сланцам архейской
группы. В несколько более поздних отложениях таких сланцев во Франции (Бретань)
были
найдены скелеты корненожек, организмов, близких к амебам, в современных
своих представителях живущих в илах мореного дна. В более старых отложениях
не найдено
никаких следов жизни. Таким образом, указанные только что остатки железобактерий
древнее всех других остатков жизни. Между тем среди хемотрофных бактерий
они далеко не простейшие, и надо думать, что от появления на Земле первых
живых
существ и до времени отложения слоев, содержащих их остатки, прошел не
один миллион лет, ибо слои кристаллических архейских сланцев очень толсты.
Вопрос о происхождении бактерий осложняется тем, что среди них есть такие,
которые произошли, по всей вероятности, путем опрощения более сложных организмов.
Так, серные бактерии из рода Beggiatoa очень близки к окрашенным, имеющим
хлорофилл водорослям осцилляриям, а живущие в крови высших животных бактерии
спирохеты,
может быть, даже животного происхождения.
Дело в том, что образ жизни бактерий-паразитов способствует упрощению организации
благодаря своему однообразию и делает сходными организмы, различные по
своему происхождению.
Кроме железобактерий, в альгонских слоях Северной Америки были найдены
еще бактерии микрококки, жизнь которых связана с отложением известняков.
В более
поздних слоях бактерии встречаются все чаще, но здесь они уже чаще связаны
с процессами гниения растительных остатков, чем с минеральными процессами.
В слоях палеозойской группы были найдены серные бактерии, микрококки и
бациллы.
Когда развилась растительная и животная жизнь, то легкость питания органическими
веществами, понятно, способствовала развитию бактерий, питающихся за счет
животных и растений. Последними то времени появления должны быть те
бактерии, которые являются паразитами человека и погибают вне человеческого
организма.
Для более отчетливого представления о бактериях дадим группировку ныне
живущих бактерий в систему. Прежде всего они делятся на две основные группы:
настоящие,
или простые, бактерии и нитчатые бактерии.
I. Настоящие, или простые, бактерии (Eubacteria или Нарlobacteria). Одноклетные,
шарообразные или палочкообразные бактерии, выделяющие обильную слизь, благодаря
которой они объединяются легко в колонии в форме пленок, дисков, шаров,
цепочек или нитей. В молодом возрасте они по большей части имеют органы
движения, реснички
или жгутики. При неблагоприятных условиях образуют внутри клеток споры.
Клетки бесцветны, и если протоплазма их образует пигменты, то эти пигменты
отлагаются
вне клеток (пигментные бактерии, колонии которых имеют красную, фиолетовую,
желтую, синюю или зеленую окраску).
1. Семейство кокковых бактерий. Отличается шарообразными клетками. Сюда
принадлежат самые мелкие бактерии. Роды: стрептококк, микрококк, сарцина
и планококк.
2. Семейство бацилл. Цилиндрические или палочкообразные формы. Роды: бактериум,
бациллус, псеудомонас и астазия.
3. Семейство спирилловых. Одиночные более и менее изогнутые клетки, от
формы слегка изогнутой запятой до настоящей спирали. Роды: спириллум и
спирохета.
II. Трихо- или десмо-бактерии (Trichobacteria или Phycobacteria, Desmobacteria).
Нитчатые формы, причем отдельные клетки располагаются внутри плотного студенистого
футляра,
1. Семейство Rhodobacteriacea — пурпурные бактерии, протоплазма которых
развивает в себе пигмент розового, пурпурного или фиолетового цвета с помощью
пигмента
бактериопурпурина; кроме того, обычно в их протоплазме отлагаются еще зернышки
серы.
2. Семейство Beggiatoaceae, род беггиатоа или беджиатоа, клетки которого
соединены в плотные нити, изгибающиеся три движении, и содержат в своей
протоплазме обильные
отложения зернышек серы. Это так называемые серобактерии, разрушающие сероводород
и очищающие воду, испорченную процессами гниения.
3. Семейство Chlamydobactericeae — хламидобактерии. Клетки находятся в
плотных чехлах, образуя простые или вильчато-ветвящиеся нити. Роды: кладотрикс
и лептотрикс.
Кроме того, к бактериям относят еще семейства: 1) микобактерий, или актиномицетов,
представители которых обнаружены в корнях некоторых палеозойских растений;
2) миксобактерий, которые дают нечто вроде плодовых тел, и 3)
спиротриховых, куда относится характерная спиральная железобактерия галионелла.
Деление бактерий по физиологическим и биохимическим их особенностям на
хемотрофные, сапрофитные и паразитные было гораздо более определенным и
ясным, чем морфологическая
их система. Ясно, что хемотрофные бактерии, способные, хотя бы в некоторых
случаях, строить свою жизнь исключительно за счет неорганического мира,
первобытнее, чем сапрофиты и паразиты, зависящие часто от высокоорганизованных
животных
и растений. Морфологически примитивнейшими бактериями являются микрококки,
но среди них есть и хемотрофы, и сапрофиты, и паразиты. Физиологическое
деление не совпадает с морфологическим. Действительно простейшими являются,
конечно,
те бактерии, которые одновременно просты и физиологически и морфологически,
т. е. хемотрофные микрококки, например, микрококки, принимающие деятельное
участие в круговороте углекислого кальция.
Дальнейшее опрощение приводит нас к невидимым образованиям типа бактериофага
или «живого жидкого заразного начала» Бейеринка, а еще дальнейшее — к вопросу
о возникновении органических катализаторов-энзимов. Ферворн еще в 1903
г., выступая с мало обоснованной гипотезой биогена, т. е. единого энзиматичеекого
вещества, вызывающего все реакции жизни, должен был согласиться, что прижизненный
обмен веществ представляет собою цепь энзиматических реакций. Синтез энзимов
— едва ли не основная задача, которая стоит перед химией на пути к разрешению
загадки, чем отличается живой белок от запасного, протоплазма зародыша
от того
запаса протеиновых веществ, который ему обычно сопутствует. Бактерии с
их ничтожными морфологическими различиями нередко различаются только своими
энзимами, например,
бактерии, разжижающие и не разжижающие желатину, иа которой их культивируют.
Благодаря своей большой пластичности, своей способности изменять вырабатываемый
ими набор энзимов в соответствии с тем сырьем, которым они питаются, большинство
бактерий и сейчас обладает выдающейся возможностью приспособления к изменчивой
внешней среде. Они не только не вымирают, но наоборот, в течение всей истории
жизни на Земле овладевают все новыми и новыми источниками питания.
А. А. Рихтер, пользуясь методом Виноградского для прямого микроскопического
учета микробов в почве, получил следующий подсчет: «лесная почва заключает
в грамме около 3 млрд. индивидуумов, каштановая около 1 1/2 млрд. и песчаная
около 1 млрд.». При этом «главной и наиболее постоянной группой почвенных
микроорганизмов являются кокки; бациллы
преобладают в лесной почве, отходя на задний план в культурной черноземной
и песчаной почвах; азотобактеры богато представлены во всех почвах, но
особенно много их в почвах культурно-черноземных»*. А ведь азотобактер
— главный источник
вновь связываемого атмосферного азота. Если таковы цифры на 1 г почвы,
то сколько же получится особей на почву целого поля и сколько же их будет
на общую массу
всего поверхностного слоя почвы на всей Земле!
| * А. А. Рихтер. Записки Сарат. гос. унив., IV, 1925, вып. 1. |
В морях мы также встречаем массы бактерий как на дне мелководий, на остатках
отмерших животных и растений и минеральных средах, так и свободно плавающими
в воде. В морях Каспийском и Черном вся масса глубинных вод насыщена сероводородом,
который, несмотря на присущую газам способность к быстрой диффузии, не
проникает в поверхностные слои воды. Благодаря этому животная и растительная
жизнь этих
морей вся сосредоточена в верхних слоях воды до глубины 200 м. Объясняется
это тем, что на данной глубине держится масса серобактерий, окисляющих
серу сероводорода в серную кислоту. Сколько же особей содержит слой, густо
заселенный
серобактериями на протяжении всей глубоководной части обоиз морей!
Благоденствуя таким образом на всем протяжении геологической истории жизни
иа Земле, бактерии должны были от времени до времени отщеплять от себя
родоначальников высших по сравнению с ними типов.
Первым принципом, который мог, осуществляясь, превратить бактерию в высшую
форму, является принцип дифференциации. Выделение из протоплазмы морфологически
обособленного ядра, локализация пигментов, сначала в поверхностном слое
протоплазмы, затем в особых ее участках — пластидах, обособление целлюлозной
или хитиновой
оболочки, — таковы главные моменты превращения клетки бактерий в клетку
высшего типа. Ф. Энгельс рассматривает образование клеточного ядра как
явление поляризации
живого белка**.
| ** См. Ф. Энгельс. Диалектика природы. М., Госполитиздат, 1952, стр. 166. |
Принимая во внимание возможность такого процесса, лам легко установить
связи, соединяющие филогенетически древних бактерий с жгутиковыми организмами,
с циановыми
водорослями, с низшими грибами.
Большая часть бактерий осталась бактериями и продолжала приспособляться
к все изменявшимся условиям жизни, совершенствуя в своей способности использовать
внешнюю среду и защищаться от разрушающих факторов.
2. ПРОИСХОЖДЕНИЕ ВОДОРОСЛЕЙ
Так как простейшие водоросли отличаются от бактерий очень мало, то мы считаем,
что они тесно связаны с ними родством. Промежуточную группу образуют
жгутиковые организмы, стоящие на рубеже растительной и животной жизни.
Это одноклетные,
различно построенные мелкие обитатели вод, с различным типом питания.
Одни из них имеют внутреннюю полость (пищеварительную вакуолю) и ротовое
отверстие
и заглатывают пищу, например, бактерий; их можно считать прародителями
животных организмов. Такие жгутиковые не имеют окраски, не содержат
хлорофилла и не
могут существовать без готовой органической пищи. Другие жгутиковые
имеют хлорофилл, лишены постоянной пищеварительной полости (вакуоли) и питаются
как растения
за счет растворенной в воде углекислоты, солнечных лучей и пр. Это
и
есть родоначальники настоящих водорослей.
Палеонтология указывает остатки водорослей, начиная с кембрийских слоев.
Известны и кембрийские жгутиковые. Для каменноугольных отложений точно
установлен уже
целый ряд низкоорганизованных водорослей. Высокоорганизованные крупные
формы известны, как появившиеся впервые в силурийских слоях. Это были
так называемые
сифонники, или сифонные водоросли, часто имеющие сложный известковый
скелет. В девонских слоях изучены А. П. Карпинским окаменелые плодики
лучицевых
водорослей трохилисков, тогда как веточки лучиц из средних и верхних
девонских слоев описаны
английскими учеными Кидстоном и Лангом.
Предки крупных океанских бурых водорослей описаны из силурийских и
девонских слоев. Водоросли эти играют большую роль в экономике моря
и во многих
странах являются предметом промысла: частью идут в пищу, частью для
добычи из их
золы иода и калийных солей. В море они образуют большие заросли и привлекают
к себе
разнообразное животное население. Красные морские водоросли, или багрянки,
появились также в силурийское время, но полного развития достигли только
в мелу, как и предыдущая группа.
Водоросли сыграли и теперь играют большую роль в деле накопления органического
вещества и минеральных отложений. С одной стороны, они питают массы
животных организмов, с другой — способствуют образованию крупных пластов
нуллипоровых
известняков, причем остатки подобных водорослей — литотамниев — известны
с нижнего мела.
Современный нам мир водорослей представляет собою пеструю картину организмов
весьма различной древности. Безусловно, некоторые их типы должны были
существовать и
в докембрийское время, но только с силура начинают попадаться достоверные
их остатки. Стало быть, одни из современных нам водорослей возникли
в докембрийское архейское время, другие — в силурийское, третьи — в
каменноугольное.
И
позже
продолжали появляться новые и новые формы, пока в третичное время не
сложился окончательно современный нам мир водорослей, перегруппировавшийся,
однако,
еще раз по климатам в ледниковую эпоху.
Если расчленить водоросли на отдельные группы, то получится следующая
картина:
1. Циановые или синезеленые водоросли, иначе водоросли дробянки (Schizophyceae
или Cyanophyceae), по строению клетки и многим другим особенностям
ближайшие к бактериям. Живут в водоемах, богатых органическими веществами,
а также
в водоемах, богатых минеральными солями, в горячих ключах, в сырой
почве, на
затененных скалах и пр.; к ним принадлежат также и некоторые виды,
сверлящие известняки.
| * М. Нinner. Handbuch der Palaobotanik, 1. 1927. ** Gruss. Palaobiologica, 1928. |
В сводке Хирмера* приводятся циановые водоросли: из альгонских слоев
6, установленных условно; из кембрийских 2, из ордовицких 6, из силурийских
2, из девонских
2, из каменноугольных 5, из пермских 1, из триасовых 2, из юрских
4, из
меловых 1, из эоценовых 1 и из миоценовых 3, итого 35. Позднее Грюсс**
описал из девона
как ископаемую циановую водоросль, Nematorites oscillatoriiformis;
если бы он был прав, относя ее к циановым, то это была бы циановая
водоросль
высшего
типа, так как он говорит, что это нитчатка с клеточными ядрами в
клетках и со специальными органами размножения, оогониями и антеридиями.
Циановые таким образом проходят, как и бактерии, через всю эволюцию,
сохранив до настоящего времени примитивный тип строения и примитивный
образ жизни;
новое у них — это приспособление отдельных родов и видов к специальным
условиям существования
и различные защитные приспособления. В дальнейшем своем развитии
они упираются в класс водорослей багрянок (Rhodophyceae).
2. Диатомовые, иначе кремнеземки или бацилларии (Diatomeae или Bacillariophyta),
одноклетные или слабо связанные в колонии водоросли. Внутренние слои
клеточной оболочки состоят из пектиновых веществ. Снаружи она покрыта
панцирем, состоящим
из кремнезема. Наличие панциря у диатомей обеспечивает их остаткам
хорошую сохранность. Действительно,
отложения ископаемых диатомей известны во многих странах и эксплуатируются
между прочим как строительный материал. Однако Пиа* утверждает, что
диатомовые впервые появляются только с нижней юры (лейас). Тем не
менее существует
пока никем не опровергнутое утверждение Грюсса, что в девоне Медвежьего
острова
им найдены первые достоверные палеозойские диатомей (Bacillites,
Discoites, Nitschiopsidea и др.). В настоящее время диатомей широко
распространены
и в пресных и в морских водах, изредка встречаются и диатомей, живущие
на сырой
земле. В морях они осаждают местами даже специальный диатомовый ил.
| * J. Р i a. Palaontologisches Zeitschrift, 1931. |
Диатомовые происходят от жгутиковых организмов, окрашенных в желтый
цвет и способных отлагать в своих оболочках небольшие количества
кремнезема. Ряд развития
диатомей предположительно таков. Хризомонады, одноклетные жгутиковые
организмы с желто-бурыми хроматофорами, от них перидинеи (Peridineae
или Dinoflagellatae),
массами развивающиеся в планктоне морей, а иногда и в пресных водах
(сюда
относится рогатик или Ceratium), от перидинеи более редкие силикофлагеллаты,
а от последних
диатомеи.
Диатомовые как бы израсходовали всю свойственную им в начале их истории
(пластичность на разнообразие клеточных форм, скульптуру или рисунок
оболочки и приспособления
к свободно плавающему образу жизни или к образу жизни организма,
прикрепляющегося под водой к скалам или другим твердым предметам.
Они не переходят ни
в какой высший тип. являясь как бы законченным в своем развитии рядом.
3. Зеленые водоросли, или Сhlorophyceae — основная очень пластичная
группа, распадающаяся на 5 классов: протококковые, улотриховыс или
конфервы (зеленые
нитчатки), сифонокладиевые, сифонные и хары, или лучицы. Происходят
от зеленых жгутиковых организмов, причем имеются между двумя этими
группами
прямые переходные
формы, как пирамидомонас и хламидомонас, подвижные одноклетные организмы
наших вод. Среди зеленых, особенно среди дазикладиевых и сифонных,
немало организмов,
отлагающих в своих оболочках кальций и потому прекрасно сохраняющихся
в морских и пресноводных отложениях. Ранее французский палеонтолог
Сапорта приводил целый
ряд окамепелостей из силурийского периода, как остатки сифонных зеленых
водорослей крупных размеров, населявших моря и игравших в те времена
выдающуюся
роль в
деле накопления органического вещества. Однако его билобиты, эофитоны
и пр. не были признаны позднейшими палеонтологами за остатки растений,
а
взамен были
открыты
многочисленные виды дазикладиевых с очень сложным строением из отдельных
камер, особенно многочисленных в период триас-юра-мел. И в настоящее
время сифонные
и дазикладиевые зеленые водоросли поражают своими оригинальными формами.
Живут они в теплых водах южных морей, в их прибрежной полосе, едва
прикрытые водой
во время отлива, на песчаном, илистом или каменистом грунте, особенно
на коралловых отмелях. Особенно много их в Карибском море Америки
и близ островов
Малайского
архипелага.
Строение этих водорослей замечательно тем, что их клеточные ядра
не вызывают, как у других растений, образования клеточных оболочек;
клетчатка
отлагается
у них частью в виде общего для всего организма мешковидного покрова,
частью (например, у различных видов каулерпы — род Caulerpa) в виде
отростков, как бы балочек, внутри; по этим отросткам и ползет протоплазма
сообразно
силе освещения,
то уходя вниз, прочь от чересчур сильных солнечных лучей, то подымаясь
вверх. У дазикладиевых внутренние перегородки делят организмы на
отдельные камеры,
но в каждой камере много клеточных ядер. Такое же строение из камер
имеют и харовые водоросли, твердые плодики которых попадаются, начиная
с девона,
а
веточки с плодиками известны для третичных отложений.
4. Бурые водоросли Phaeophyceae, самые крупные из всех, со сложной
окраской, в которой, кроме хлорофилла и каротина, свойственных всем
аутотрофным
растениям, принимает участие еще желто-бурый пигмент фикофеин, усиливающий
их ассимиляционную
способность. Бурые водоросли и теперь образуют у скалистых берегов
умеренно холодных стран целые подводные леса. Общепризнано присутствие
остатков
бурых водорослей в отложениях юрского, мелового и третичного периодов.
У этих водорослей
не наблюдается отложений кальция или кремния, и потому у них мало
шансов на хорошую палеонтологическую сохранность. Поэтому обычно
думают, что
и в палеозойские
времена они были представлены высоко развитыми формами, обособление
же их в характерную самостоятельную ветвь столь разнородного мира
водорослей произошло
еще ранее.
| * J. Pia. Palaontologisches Zeitschrift, 19S1. ** Gruss. Palaobiologica, 1928. |
И в последнее время такой авторитет полеонтологии растений,
как Пиа*, подтверждает большое значение бурых водорослей в девонский
период, а
Грюсс** описал из девона бурую Nematophora fascigera.
Во всяком случае к началу третичного периода они уже совершенно
сформировались. Находки
в эоцене саргассов
(Sargassum globiferum Steinberg) и
цистозейр (Cystoseira filiformis Stern и С. helvetica Heer) ясно
указывают на то, что разнообразие эоценовых бурых водорослей не
уступало современному.
Если в океанах и происходили крупные катастрофы, связанные с опусканием
так называемых абиссальных областей и другими геологическими изменениями,
то все
же часть океана всегда сохраняла условия, необходимые для жизни
водорослей, и они могли развиваться беспрерывно с девона до нашего
времени.
5. Багрянки, или красные водоросли (Rhodophyceae) с набором пигментов,
в котором, кроме хлорофилла и каротина, участвует еще красный флуоресцирующий
пигмент
фикоэритрин, благодаря чему они способны к ассимиляции при сильно
ослабленном освещении и могут жить в более глубоких частях моря,
опускаясь до границ
проникания солнечных лучей. Группа эта, связанная своим клеточным
строением с высшими
представителями циановых водорослей, отличается чрезвычайным разнообразием
и внешних форм, я цикла развития. Древнейшие из них жили уже в
морях ордовицкого периода (Delesserites salicifolia Ruedemann).
Среди багрянок
выделяется
замечательное семейство кораллиновых, у которых оболочки их в общем
мелких клеточек пропитываются
углекислым кальцием, что придает им внешнее сходство с кораллами.
Они широко распространены в современных морях и образуют местами
обширные
нуллипоровые
рифы, похожие на коралловые, но лишенные характерных для последних
дырочек, в которых у кораллов живут строящие их гидроидные полипы.
Подобные нуллипоры
известны и в ископаемом состоянии, начиная с ордовицких и силурийских
слоев, и были очень обыкновенны в морях мелового периода.
Красные водоросли никаких переходов к выше стоящим группам растений
не дают. Подобные переходы намечаются только у зеленых и особенно
у бурых
водорослей.
3. ПРОИСХОЖДЕНИЕ ГРИБОВ
Грибы лишены хлорофилла и питаются уже готовым органическим веществом.
Они живут или на отбросах животных, на мертвых ветках и других
частях растений, или же паразитируют на различных животных
и растениях. Простейшие трибы
сильно
отличаются от бактерий как строением, так и химизмом, и мы
не можем думать, что эти два класса родственны. Наоборот, с жгутиковыми
организмами
они
сближены во многих своих особенностях. Высшие грибы еще ближе
к некоторым группам водорослей.
Наука рассматривает в настоящее время большинство грибов, как
происходящее от водорослей, через изменение образа жизни. Потеря самостоятельного
питания с помощью солнеч-
ных лучей и замена его паразитизмом или использованием мертвых
частей животных и растений привела некоторые водоросли к потере
зеленой
окраски и соответствующему
изменению химизма. Водоросли превратились в грибы. Конечно,
это не исключает того, что многие грибные организмы произошли
от
грибов же. Палеонтология
обнаружила остатки паразитных грибов в различных растительных
органах лепидодендронов или чешуедревов и других вымерших ныне
растений.
Понятно, что остатки грибов
начинают встречаться только там, где растительность суши достигла
уже высокого развития. Короче, мы не знаем пока грибов древнее
каменноугольных.
Высшие,
особенно шляпные грибы, например трутовики, появляются только
в третичное время;
грибы, близкие к опенкам, обнаружены в миоценовых его слоях,
как и
грибы дождевики.
Известно, что грибы поселяются иногда в тканях корней различных
растений, причем не причиняют им никакого вреда, а наоборот,
даже усиливают
всасывание корнями
почвенных растворов. Явление это, совместная жизнь гриба с
корнем, получило название микоризы, т. е. грибокорня. Микоризы
были найдены
в тканях корней
из верхнекаменноугольных отложений Англии. Считают, что в подземных
частях стебля девонских псилофитов жила микориза. Не ее ли
остатки были описаны
из девонских отложений под именем палеомицеса, т. е. древнего
гриба?
Конечно, полного своего развития достигли грибы только в третичное
время, когда развились те бесчисленные цветковые растения —
деревья, кустарники
и травы,
— на листьях, стеблях, ветвях, стволах и пр. которых грибы
обычно паразитируют. Наши любимые шляпные грибы, идущие в пищу
человека,
большею частью
связаны с корнями тех деревьев, под которыми они растут, и
понятно, не могли
появиться ранее этих деревьев. Близкие к грибам лишайники появились,
по-видимому,
с каменноугольного времени. Грибы не имеют скелета, и сохранять
их сколько-нибудь долгое время
нелегко, но зато паразитные формы грибов, грибница или мицелий
которых разрастается в межклетных ходах высших растений, и
в ископаемом состоянии
сохраняются прекрасно,
защищенные тканями, среди которых растут.
Грибы разделяются на три основные класса: фикомицеты, аскомицеты
и базидиомицеты. Первые, т. е. фикомицеты, не имеют типичного
клеточного строения; нити,
из которых слагается их тело, не разделены перегородками, но
на всем своем протяжении
имеют общую полость, иначе, как и указанные выше водоросли,
они имеют сифонное строение. Такой мицелий легко узнается под
микроскопом
даже
и в ископаемом
состоянии. Многие фикомицеты паразитировали на папоротниках,
лепидодендровых и других растениях каменноугольной эпохи. Особенно
интересен пероноспоровый грибок, Peronosporites antiquarius Worth. Sm., сохранившийся вместе со своими характерными органами
размножения
оогониями.
Обыкновенная
белая плесень (Mucor) также представлена в карбоне (Mucorites).
Сумчатые грибы, или аскомицеты, дающие нередко твердые плодовые
тела, представлены
в ископаемом
состоянии значительно богаче, но только из меловых и третичных
отложений. Очень хороши остатки плесеней с перегородчатыми
нитями грибницы и
характерными цепочками
конидиальных спор, найденные в янтарях (Penicillites curtipes Berk el.).
Базидиальные грибы, у которых материнские клетки спор дают
споры не внутри клетки, как у сумчатых грибов, а снаружи, на
особых
ножках стеригмах,
достигают часто высших форм развития. Высшие, или шляпные грибы,
напр, трутовики,
известны начиная с карбона, но многие из них при более тщательном
исследовании оказались
состоящими из железистого песка и являются так называемыми
конкрециями. Вполне достоверны только позднейшие находки, начиная
с верхнего
мела. Из того же верхнего
мела известны и остатки паразитных ржавчинных грибов.
В вопросе о происхождении грибов существуют две точки зрения.
Одна за то, что все грибы присходят от низших амебообразных
организмов с жгутиками
и представляют
собою самостоятельно развивавшуюся ветвь растительного мира.
Между
амебоидными организмами и грибами существует целый ряд средних
форм, объединяемых
микологами
в особый отдел первичных грибов, или архимицетов. Принадлежащий
к этой группе грибок урофликтис (Urophlyctis Oliverianus Magnus)
жил
некогда
в листьях семенного
папоротника Alethopteris в верхнем карбоне. И сейчас наиболее
примитивные представители этой группы живут или в воде, или
в сырой почве,
и лишь немногие в тканях сухопутных
растений.
Другая точка зрения, сейчас более распространенная, — это та,
что грибы происходят от водорослей, через приспособление к
паразитическому или
сапрофитическому существованию и явившуюся результатом этого
потерю
хлорофилла. Одни из
них
происходят от сифонных зеленых водорослей (фикомицеты), другие
от красных водорослей. Иногда даже точно указывается, какой
род грибов
происходит
от какого рода водорослей
(напр., ржавчинник Puccinia от водоросли багрянки Polysiphonia).
С этой точки зрения грибы — не единая группа, а только придаток
к различным
группам водорослей.
Однако есть одно возражение против такого уничтожения грибов
как класса: это их химическая характеристика.
Дело в том, что грибы отличаются от бесхлорофильных водорослей
и бактерий целым рядом химических особенностей. Так, если приготовить
в высоком
цилиндре питательный
настой
(напр., отвар капустных листьев) и подкрасить его нейтральным
лакмусом в слабо-фиолетовый цвет, то при заражении настоя смесью
грибов
и бактерий мы быстро увидим, что
жидкость разделится на слои, окрашенные различно. Наверху мы
увидим ярко-синий слой с развивающимися в нем грибными организмами
(плесенями),
а внизу
ярко-красный, с кишащими в нем бактериями. Клеточный обмен
бактерий требует мало кислорода
из воздуха и сопровождается выделением кислот, а клеточный
обмен грибов сопровождается обычно довольно сильным поглощением
кислорода
и выделением
щелочей. Таким образом
легко отличить неокрашенный, лишенный хлорофилла и его спутников,
организм водорослевого ряда от организма грибного ряда, даже
в том случае, если
они морфологически сходны.
Возможно, что два основных ряда низших или споровых растений
развивались параллельно, и некоторые особенности в развитии
тех и других проявлялись
по мере их эволюции
сходню ч обоих рядах, независимо от происхождения отдельных
групп.
Последняя группа грибных организмов — это лишайники. Их рассматривают,
начиная с Шимпера, как грибы, выработавшие особый симбиотический
тип питания за счет
заключенных в их тканях водорослей. Большинство лишайниковых
грибов принадлежит к сумчатым грибам, или аскомицетам, ничтожное
меньшинство
тропических
их представителей — к базидиомицетам, сходным с трутовиками.
В ископаемом состоянии лишайники
крайне редки. Достоверные их остатки известны только с верхнего
мела (род
Opegrapha), но особенно многочисленны они в янтарях олигоцена,
а также в пластах бурых
углей того же периода.
Грибы являются законченной в своем развитии группой и не дают
переходов к каким-либо вышестоящим организмам. Их развитие
вылилось в приспособление
к разнообразнейшим
жизненным условиям применительно к строению и химическому составу
питающих их организмов. Среди грибов много так называемых облигатных
паразитов,
которые чрезвычайно тонко реагируют на определенные протеиновые
соединения. Попытки
заразить спорами облигатного паразита какое-либо цветковое
растение, кроме того, на котором он обычно живет, всегда оканчиваются
неудачей.
Благодаря
паразитным грибам иной раз удается точно установить систематическое
положение цветкового
растения, не поддававшегося морфологическому анализу.
Высшие, или шляпные грибы часто вступают в симбиоз с корнями
определенных деревьев (напр., подосиновик на корнях осины),
образуя вместе с
ними так называемую
микоризу. Все это, вызывая массу частных приспособлений, удерживает
грибные организмы на определенной ступени развития, мешая им
развить высшие формы.
4. ВЫХОД РАСТЕНИЙ НА СУШУ
И ПРОИСХОЖДЕНИЕ НАЗЕМНОЙ РАСТИТЕЛЬНОСТИ
В истории развития растительного мира одним из важнейших моментов
является возникновение сухопутной флоры. В то время как
бактерии и водоросли
живут или в водоемах, или, если они обитают в почве, на
сырых склонах или на
голой земле,
в очень тонком слое воды, достаточном для свободного их
передвижения, так как сами они очень малы, высшие растения — мхи, папоротникообразные,
голосеменные
и цветковые, образующие ныне главную массу земной растительности
— приводят в соприкосновение с водой только свои корни,
корневые
волоски
и ризоиды,
все
же остальные их органы находятся в сухой атмосфере и испаряют
воду всей поверхностью.
Если предположить, что родоначальниками растений суши были
водоросли, то трудно понять, каким образом эти легко гибнущие
на воздухе
организмы могли
приспособиться
к резкой потере воды и ее возмещению.
Познакомимся вкратце с теорией Бауера, взявшегося выяснить
этот трудный вопрос. Основой всякого размножения является
деление клеточного ядра,
влекущее за собой
и деление клетки. Если организм одноклеточный, то после
деления клетки получается два организма. Если организм
сложный, то
в
результате деления получается рост
ткани, а рост ткани влечет за собой и рост органа, в состав
которого входит данная ткань. Само собой разумеется, что
каждая клетка
при делении дает
себе подобную.
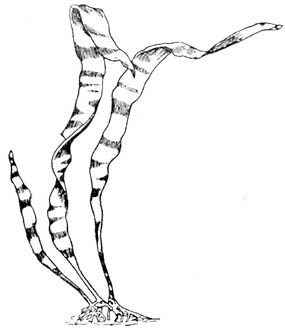
Рис. 22. Гаметофит, или заросток (гаплоид) водоросли ламинария. Сильно увеличено |
Обратим теперь внимание на строение ядра из нитей особого ядерного вещества, или хроматина. В период деления хроматин ядра распределяется между определенным числом отрезков, называемых хромосомами. Согласно данным науки о наследственности, каждая хромосома имеет особое значение как носитель определенных наследственных свойств организма. При передаче этих свойств от одного поколения другому, передача совершается именно через хромосомы ядра. Если одна из хромосом погибнет, то и свойства, носителем которых она является, не передадутся по наследству. Так вот по отношению к хромосомам ядра одного и того же организма неодинаковы. Как правило можно принять, что число хромосом у каждого обособленного организма свое, постоянное. Так, капуста имеет в своих клетках 18 хромосом, рапс 20, у табака и махорки их 24, у липы 41, у осины 8, у одних сортов хлопка 26, у других 52, у кукурузы 20, у пшеницы 42, у ржи 14 и так далее. При делении ядра, в стадии экваториальной пластинки, хромосомы делятся каждая продольно на две, после чего начинают расходиться к полюсам старого ядра, образуя дочерние ядра. Поэтому-то в каждом дочернем ядре будет то же число хромосом, что было в материнском ядре, а самое деление называется уравнительным или эквационным. Однако после неопределенно большого числа эквационных делений клетки начинают делиться иначе. Именно, перед образованием ядерного веретена хромосомы попарно сливаются, и число их уменьшается вдвое. В стадии ядерной пластинки они снова делятся продольно и расходятся, образуя два дочерних ядра, но так как слияние уменьшило их число вдвое, то и дочерние ядра будут обладать лишь половинным числом хромосом. Полное число хромосом называется диплоидным, или двойным, а половинное — гаплоидным, или простым. Из гаплоидной клетки не может развиться такой же организм, как из диплоидной, а будет развиваться организм более слабый, в котором число хромосом все время будет поддерживаться эквационным делением все в том же гаплоидном числе. Только слияние двух ядер в процессе оплодотворения удваивает число хромосом, и возникающий в результате молодой организм снова становится диплоидным. Диплоидный организм размножается обычно почкованием. Если почки одноклеточные, то они называются спорами, если состоят из однородной ткани, то таллидиями, если из органов, каковы стебель и листья, то просто почками. Гаплоид может размножаться подобно диплоиду, оставаясь при этом подобным себе самому, но может и дать так называемые гаметы, т. е. клетки, не способные к самостоятельному развитию, но способные к слиянию друг с другом, после чего число хромосом удваивается и получается так называемая зигота, т. е. двойная клетка, первая клетка следующего поколения. У многих растений, как, например, у мхов и папоротников, у многих водорослей поколение диплоидных клеток неизбежно сменяется поколением гаплоидных. У мхов сам мох гаплоидный, диплоидным же является развивающийся на нем спорогон, состоящий из пяты, ножки, апофиза и коробочки; у папоротников сам папоротник, приносящий во множестве споры, является диплоидным, а вырастающие из спор маленькие заростки гаплоидны.
Рис. 23. Спорофит (диплоид) водоросли ламинария. Сильно уменьшено |
Как правило можно вывести из этих и многих других примеров,
что диплоид приносит после наступления в некоторых его
клетках редукционного
деления споры, а из
спор вырастают гаплоидные организмы, заканчивающие свою
жизнь развитием гамет. Гаметы сливаются, удваивая это число
хромосом
и образуя
зиготу,
из которой
вырастает диплоидный организм. Два типа размножения: почкование
и слияние гамет соответствуют как бы двум различным по
своей внешности и своим
свойствам организмам,
следующим в обязательном порядке один за другим. Это явление
носит в науке наименование смены поколений, или смены спорофита
гаметофитом.
Поразительна смена гаметофита спорофитом у морской водоросли
ламинарии.
Несмотря на широкое географическое распространение, крупные
размеры и большое промысловое значение этой водоросли,
вплоть до работ
Соважо (1915)
и Килина
(1916), ботаники считали ламинарию, или морскую капусту,
за организм, не имеющий полового размножения, так как известны
были только
ее подвижные споры, гаметы
же никогда не наблюдались. Оказалось, однако, и притом
совершенно неожиданно, что споры ламинарии прикрепляются
к камням и
разрастаются
в маленькие
растеньица,
которые после продолжительного периода развития дают: одни
— яйцевые клеточки, другие — подвижные
сперматозоиды. Оплодотворенная яйцеклетка разрастается
в крупное растение с дифференцированными тканями, приносящее
затем периодически
массу
спор.
Для спорофита-диплоида ламинарии характерны быстрый рост,
мощность развития и физиологическое расчленение тканей
тела на щокровные,
ассимиляционные, проводящие, механические и выделительные;
для гаметофита гаплоида (20
хромосом) — малый
рост, медленное развитие, простота строения.
За исключением мхов у всех растений, у которых ясно выражена
смена поколений, мы находим то же взаимоотношение, т. е.
слабый и просто
организованный,
медленно растущий гаметофит, хорошо развивающийся только
в воде или при ее изобилии,
и быстро растущий, сложно построенный и способный сопротивляться
высыханию спорофит. Ясно, что обладание двойным числом
хромосом, короче, двойным
ядром, чрезвычайно выгодно для организма. Ясно также, что
естественный отбор должен
давать в борьбе за существование преобладание тем растениям,
диплоидный аппарат (которых развит сильнее. Если в водной
среде гаплоидные
организмы и могут сравнительно
счастливо заканчивать свой жизненный цикл, то на суше солнце
и ветер быстро сокращают их жизнь. Но в воде существование
гаплоидных организмов
поддерживается
только напряженностью их размножения, при полной нестойкости
особи.
Диплоид не таков—надо видеть, как рвет и треплет океанский
прибой гибкое тело
ламинарии, растущей близ берега на подводных скалах, чтобы
оценить всю силу ее сопротивляемости.
Выше мы видели, что первый расцвет растительной жизни на
суше был приурочен к приморским местностям, особенно к
берегам лагунных
озер и болот,
лежащих между песчаными береговыми валами и подгорной террасой.
Здесь впервые
выработался тип растения, нижней своей частью находящегося
в воде,
а верхушкой в воздушной
среде под прямыми лучами солнца. Споры, развивавшиеся в
мелкой нагретой воде лагун, при понижении уровня воды,
вследствие
испарения, образовывали
заростки
в виде плоских зеленых лопастных пластинок. Отдельные участки
их, выдвигаясь из воды и испаряя воду, вызывали возникновение
токов
воды, возмещавших
испаряемую, и этим способствовали росту своих тканей. Образовывалось
подобие первичного
стебля. Если на таком стебле возникали споры или гаметы,
то они падали в воду и там прорастали, образуя новые заростки,
или же,
сливаясь
в воде, облегчавшей
их движения, давали зиготы, полагавшие начало диплоидному
поколению
— спорофиту.
Позднее, проникновением на незатопляемую сушу, первобытные
растения развили корень (часть спорофита) и получили способность
использовать
грунтовые
воды, что дает возможность
переживать промежутки между дождями. Осталось только выработать
такой способ образования гамет и их слияния, который за
неимением у корней
данного растения
свободной воды позволял бы гаметам двигаться во внутренней
среде самого растения. Теперь почти все клетки сложного
растительного организма становятся диплоидными,
почему оно приобретает и быстроту развития, и способность
к расчленению
клеток на ткани, способность к выработке защитных приспособлений,
каких
не знали его
жившие у воды предки. Гаплоидными остаются только споры
и вырастающие из них микроскопические заростки, защищенные
тканями тела растения
матери. Таким образом,
выход растений на сушу сопровождался полным подчинением
гаметофита
спорофиту и сильным разрастанием последнего.
5. ПРОИСХОЖДЕНИЕ МХОВ
Мхи, несомненно, происходят от водорослей. Их молодая стадия,
так называемые предростки, очень сходна с некоторыми
зелеными водорослями.
В последнее
время, на основании исследования органов размножения
мхов, стали доказывать, что они
произошли от более простых представителей бурых водорослей,
применившихся к жизни на сырых скалах или вообще на
земле.
Древнейшие из несомненных остатков мхов относятся к
верхним каменноугольным слоям и принадлежат каким-то
листостебельным
мхам. Более обильные
их остатки найдены в миоценовых слоях третичной группы.
Мхи очень нежны,
плохо минерализуются,
и вероятность их сохранения очень мала. Однако сравнение
ныне живущих мхов между собою заставляет думать, что
мхи вообще
и, в частности,
та их группа,
которую мы называем печеночниками, существовали задолго
дo каменноугольного времени. Общие соображения заставляют
приурочивать
появление
мхов к тому моменту в истории Земли, когда впервые
стало возможным для
водорослей
поселиться на
влажных частях суши. Здесь-то и должны были выработаться
под влиянием жизни на суше те более сложные отношения
между всасыванием
воды
нижней частью
растения и испарением ее верхушкой, которые привели
к более сложному, чем у водорослей,
строению мха.
Напомним жизненную историю мхов: из споры, в воде,
развивается небольшой предросток, напоминающий по своему
строению
или зеленую нитчатку,
более или менее ветвистую,
или зеленую лопастную пластиночку. Это предросток мхов.
По мере усыхания воды путем образования почек и их
разрастания на предростке
образуется
взрослый
мох, как и предросток, весь гаплоидный, хотя бы он
и состоял из стебля и листьев. В странах с очень влажным
климатом,
лишенным вовсе сухих
периодов, особенно
в горных лесах, полосы туманов и облаков, мхи могут
достигать пышного развития и сложного морфологического
расчленения при длине до 30 см. Чем суше климат, тем
мельче мхи и тем
их меньше.
Развитие мха заканчивается обычно образованием органов,
в которых развиваются гаметы, именно архегониев и антеридиев.
Последние
помещаются в пазухах
листьев или на верхушке стебля между листьями. Если
дождь или роса зальют эти органы
водой, то освобождение сперматозоидов и (передвижение
их к яйцевым клеточкам становится возможным, происходит
оплодотворение
(опять-таки
в воде) и
получается диплоидная зигота, совмещающая в себе сумму
хромосом
обеих гамет. Зигота
дает спорогон, паразитирующий на мхе; спорогон состоит
из диплоидных
клеток, он
врастает в ткань стебля гаплоидного мха особым сосущим
органом — пятой, имеет ножку, заканчивающуюся зеленым
расширением — апофизом и сложнопостроенной
коробочкой.
В тканях коробочки возникает редукционное деление,
как результат
вызываемой усиленным испарением дисгармонии, и образуются
споры.
Предросток мха соответствует жизни в воде, сам мох
— амфибия, то мокнущая, то сохнущая, а спорогон — растение
засухоустойчивое.
Можно сказать,
что и самое строение мхов соответствует переходному
моменту
выхода растений
на сушу. Превращение
водорослевидного предростка в засухоустойчивый спорогон
как бы является эмблемой того процесса, который из
водорослей выработал растения
суши.
В новейших работах фитопалеонтологов указания на ископаемые мхи очень редки. В общем, можно оказать, что в четвертичных отложениях встречаются мхи, не отличающиеся от современных, в верхнеплиоценовых отложениях близ Франкфурта найдены остатки мхов, которые хотя легко определяются только до рода (роды: Neckera, Leskea, Heterocladium, Eurhynchium, Thamnium и Anomodon), но роды эти легко распознаются. Для миоценовых мхов такой возможности нет, и они описаны как особый род Muscites, причем уже самое название обозначает только принадлежность к мхам, не претендуя на большую точность. В олигоцене остатки, включенные в куски янтаря, опять-таки только Muscites. В верхнем эоцене Техаса Беллу* удалось выяснить мох Jungermannites bryopteroides до степени рода и вида, но Харрису** для триасовых отложений Гренландии не осталось ничего другого, как назвать исследованные им остатки мхов неопределенными обозначениями Thallites и Hepaticites, обозначая этим возможную их принадлежность к печеночным мхам. Наиболее древней находкой мха остаются мусцитес, Muscites polytrichoides Ren. et Zeill. из верхнего карбона.
| * О. М. Bell. 1931. ** Т. М. Harris. The fossil flora of Scoresby Sund Fast Greenland, Medd. om Creenl., 1031, 85. *** D. H. Campbell. The origin of land plants. Science, 1930, 72, 177. **** К. И. Мейер. Происхождение наземной растительности, изд. 2. М, 1929. |
Ю. Пиа говорит, как и многие другие, что сосудистые
растения произошли в девонский период от бурых водорослей,
но
ведь мхи не имеют сосудов,
построены гораздо
примитивнее сосудистых растений и должны предшествовать
им в своем появлении на Земле. Пиа не решает этого
вопроса, ограничиваясь утверждением, что
мхи нельзя противопоставлять сосудистым растениям,
как равнозначную
группу. Кемпбель***
говорит, что мхи примитивнее папоротникообразных и
что среди них
мы и можем проследить развитие спорофита и приспособление
его к условиям суши,
тем
более, что мхи легко сопоставить в ряд, начиная от
рода рикчия (Riccia) и кончая родом
антоцерас (Anthoceras): ряд этот и приводит к позднейшему
образованию папоротникообразных. В советской литературе
также есть замечательное
исследование проф. К. И.
Мейера****, в котором автор приходит к той же мысли;
только он считает простейшим и наиболее
близким к водорослям организмом печеночный полуводный
мох сферокарп
(Sphaerocarpus). Он не имеет листьев и весь состоит
из ветвящихся узких
пластиночек, клетки
которых равномерно наполнены хлорофилльными зернами.
К почве он прикрепляется бесцветными нитевидными клетками
— ризоидами.
Спорогон
его также
очень примитивен и защищен от испарения особым покровом.
Легко представить себе, говорит К.
И. Мейер, еще более простой организм, который можно
назвать условно первичным сферокарпом (Protosphaerocarpus),
а
его
уже очень легко
вывести
из водоросли,
покинувшей родную стихию для жизни на отмелом берегу.
Влияние ветра и солнца заставило все более уменьшать
развитие тканей
спорофита
и переход
его к
полупаразитическому питанию, усиливая в то же время
выработку защитных оболочек на спорах.
У мха антоцерас есть еще особенность, унаследованная,
несомненно, от водорослей; когда он затоплен, то содержимое
отдельных
его клеток выходит
через лопающуюся
оболочку наружу в виде зеленого комочка и, увлекаемое
водой, попадает на берег, после чего вырастает в новое
растение.
Такие, как их
называют, «апланоспоры»
довольно обыкновенны у водорослей и повторяются у одного
только этого мха антоцерас.
Если сравнивать мхи с папоротниками, то можно сказать,
что у мхов сильное развитие гаплоида (он же гаметофит
или заросток)
преобладает
над слабым
развитием диплоида
(он
же спорофит или спорогон). У папоротников, наоборот,
развитие диплоида преобладает над развитием гаплоида.
Листья мха,
состоящие из гаплоидных
клеток, соответствуют
не листьям папоротника, а лишь лопастям по краям его
заростка. Листья папоротника, состоящие из диплоидных
клеток, соответствуют
не листьям
мха, а апофизу
на ножке его спорогона. Все это с морфологической точки
зрения, с точки зрения
истории развития. Физиологически, поскольку лист есть
орган фотосинтеза и транспирации (газовый обмен), лист
всегда
соответствует листу,
независимо от происхождения
последнего. Впрочем, апофиз мохового спорогона часто
также выполняет функции листа.
Современная ботаника делит всю совокупность мхов на
два класса: печеночники (Hepaticae) и листостебельные
мхи
(Musci frondosi),
причем спорогон
первых примитивнее, чем спорогон вторых, а у гаметофита
слабее развиты жилки,
проводящие воду от всасывающих ее клеток ризоидов к
испаряющим частям растения. Печеночники
делятся затем на три порядка, причем в одном из них
имеется семейство юнгерманиевых с архегониями, находящимися
на
концах веточек,
и густо сидящими листьями.
Листья эти, однако, лишены жилок, чем и отличаются
от листьев большинства листостебельных
мхов. Среди последних различают три подкласса: Sphagnales,
Andraeales и Bryales. Первые, сфагны или торфяные мхи,
стоят ближе остальных
к печеночникам; при
очень незначительном морфологическом разнообразии сфагны
сыграли в четвертичном периоде выдающуюся роль торфообразователей,
так как, будучи
малочисленны
в тропическом поясе, они получают массовое развитие
в северных странах, покрывают
значительные площади и накопляют массу органического
вещества. Благодаря присутствию в их клеточных оболочках
антисептических
соединений массы
мертвого
сфагна не
гниют, а накопляются из года в год, создавая для торфяников
значительный ежегодный прирост.
Наиболее значительной, вероятно, позднее развившейся
группой мхов является подкласс Bryales, обнимающий
громадное число
форм, развившихся
главным
образом благодаря применяемости мохового организма
к разнообразнейшим условиям жизни.
Громадное число родов и видов настоящих листостебельных
мхов различается между собой строением коробочек, особенно
зубчатого
внутреннего
их края, регулирующего
рассеяние созревших спор; крайне разнообразно и строение
их листьев. Масса мхов живет на ветвях или на коре
деревьев в
сырых лесах,
на скалах, на
тундрах севера, на песках, среди дерновых злаков на
лугах и степях. Будучи, за исключением
сфагнов, слабее других растений суши,
занимают в природе подчиненное место, довольствуясь
клочками почвы, не занятыми более мощными конкурентами.
Зато они
мало зависят от
плодородия почвы, довольствуясь
минимальными количествами минеральных солей, растворенных
в дождевой воде, и потому хорошо уживаются на самых
истощенных почвах.
Несомненно, происходя от водорослей, с которыми их
связывают многие особенности строения и развития, мхи
не дают никаких
переходов к вышестоящим сосудистым
растениям. Они образуют замкнутую ветвь растительной
эволюции. Можно сказать, что построить жизнь на суше,
развивая органы
борьбы за существование
из
гаплоида-гаметофита, не способного к полной дифференцировке
тканей, — роковая ошибка. Задачу эту
могут решить только растения, строящие свою жизнь на
развитии спорофита-диплоида,
с его почти неограниченными возможностями отвечать
на воздействия окружающей среды изменением тканей и
органов
и, прежде
всего, способность построить
корневую систему, что обнаруживается уже у папоротникообразных.
6. ПРОИСХОЖДЕНИЕ ПАПОРОТНИКООБРАЗНЫХ РАСТЕНИИ
Папоротникообразные, по мысли американского ботаника
Джеффрея, можно разделить на папоротникообразные с
крупными, часто
расчлененными на доли и дольки,
листьями, имеющими развитые черешки, и такие, листья
которых мелки, цельны и часто заканчиваются
шиловидно или щетинисто. Вторые — с одной средней жилкою,
первые — с перистым или сетчатым жилкованием.
Мелколистные папоротникообразные составляют более древнюю
и, вместе с тем, основную группу. Они дожили до наших
дней в виде
плаунов
и хвощей. Расцвет
их относится к каменноугольной группе пластов, где
их остатки наиболее обильны и свидетельствуют о времени,
когда эти
мелколиственные, или ликопсиды,
по Джеффрею,
составляли главную массу лесных деревьев.
Первой относящейся сюда группой являются псилофиты
верхнего силура и девона. Самым древним из них является
риния
из древнего красного
песчаника
Шотландии.
Она достигала всего до трети метра высоты и состояла
из ползучего корневища и прямых вильчатоветвистых стеблей.
Некоторые ветви
заканчивались прямостоящим
спорангием, имевшим сложноустроенные стенки и наполненным
мелкими спорами. Корней и листьев не было. Растение
это занимает как
бы середину между
мхами и плаунами, ближе, однако, стоит к последним.
Более поздние представители псилофитов, именно астероксилоны, были крупнее,
и стебли их несли многочисленные
листочки,
имевшие вид твердых острых
чешуи. Они напоминали несколько листья некоторых современных
плаунов. По-видимому, в начале периода псилофитов других
папоротникообразных не было.
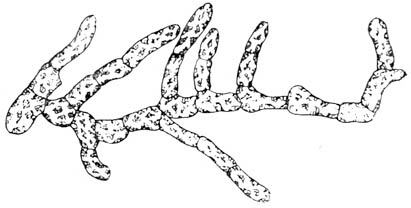
Псилофиты дали начало плаунам, которые с середины девона
начинают быстро развиваться и достигают наиболее пышного
расцвета к
середине каменноугольных
слоев. Особенно
обильны здесь большие деревья лепидодендронов или чешуедревов,
наряду с ними росли и мощные сигиллярии, или печатные
деревья, названные
так потому,
что
следы сброшенных уже листьев оставляли на коре их рубцы,
напоминающие печати.
Корневые системы лепидодендронов и сигиллярий, правильно
разветвленные, мощные, известны под названием стигмарий.
Третьим типом плауновых
деревьев был не менее
мощный ботродендрон, т. е. «деревья с кистями», и,
действительно, их ветви были украшены многочисленными
кистями плодущих
облиственных веточек
со
спорангиями. В каменноугольных пластах найдены и небольшие
плауны, более напоминающие современные.
Другую группу мелколистных папоротникообразных составляли
хвощи и клинолисты, также достигавшие максимального
расцвета в каменноугольное
время. Эти
растения отличались тем, что листья и боковые ветви
сидели у них на стеблях всегда
кольцами, отчего растения имели членистый вид. Появились
они в среднем
девоне, но в начале
мезозойской группы начали вымирать, и к третичному
времени остался только один род хвоща, представители
которого
дожили и до нашего
времени.
Крупнолистные папоротникообразные, или птеропсиды Джеффрея,
начали развиваться с верхнего девона. Они занимали
и в каменноугольном лесу сравнительно
скромное место, хотя и достигали нередко древовидных
форм. Вначале
преобладали группы
папоротников с солидными толстостенными спорангиями,
потомки которых в виде группы мараттиевых папоротников
и теперь
встречаются в жарких
странах.
Позднее
появились и стали вытеснять древние формы наши обыкновенные
папоротники, достигающие в теплых влажных странах и
теперь чрезвычайно пышного
развития. Ледниковый
период выморозил их в северных широтах, и в наших лесах
мало папоротников. Они избегают также пустынь и степей
и ютятся
преимущественно в
более влажных лесах юга.
Основные группы папоротникообразных растений — это
папоротники, хвощи и плауны. Первые два имеют то общее,
что их сперматозоиды
имеют многоресничковый
двигательный
аппарат, плауны имеют сперматозоиды о двух ресничках.
Число ресничек не
зависит ни от каких современных условий; нельзя также
сказать, что функция движения
в том и другом случае различна — энергия и направленность
движения сперматозоидов всех трех групп одинаковы.
Таким образом, различие
в числе ресничек является
признаком, который сложился исторически. Такие признаки
справедливо считаются надежными показателями родства
и происхождения
растительных групп. Гаметофиты
папоротников и хвощей довольно сходны и к остальных
своих признаках, в то время как гаметофиты плауновых
растений
совершенно с ними
не сходны. Следовательно,
происхождение плаунов надо рассматривать независимо
от происхождения остальных
папоротникообразных растений.
Современные плауны — растения суши, их вильчато-ветвящиеся
стебли густо одеты небольшими цельными листьями, спорангии
реже одиночные
на верхней
стороне листьев
близ их основания, спороносные листья чаще собраны
в колосья на концах ветвей. Выпадающие из спорангиев
споры
прорастают
только зарытыми
или полузарытыми в почву, и развивающийся из них гаметофит
представляет собою клубенек или
шнуровидное
образование, причем зеленеет только часть его, выставляющаяся
из
земли
на свет, или все оно бесцветное. Обычно в толще ткани
заростка виден слой клеток, наполненных
грибными нитями. Симбиоз грибов с подземными частями
растений, как известно, способствует переводу запасных
веществ в
растворимое состояние
и тем
повышает всасывающую способность этих подземных частей.
Растения с микоризой часто
называются микотрофами или питающимися с помощью грибов.
На гаметофите развиваются половые железы, архегонии
с яйцевыми клеточками и антеридии, дающие сперматозоиды.
Из яйцевой
клеточки после удвоения
числа ее
хромосом за счет ядра сперматозоида развивается спорофит,
который быстро пробивает себе путь к свету, развивает
в нижней своей
части корни,
а выше — листостебельный
твердый побег с развитыми проводящими пучками. Корень
и проводящие воду пучки из водоносных клеток и волокон
вокруг
них — необходимое
орудие
жизненной борьбы
на суше.
Наиболее примитивным из плаунов является филлоглоссум
из Тасмании и Новой Зеландии. Это — растеньице в 2
— 3 см
вышиной с одним
пучком игловидных
листьев и одним
спороносным колоском, сидящим на прямой ножке. Подземные
его части имеют форму клубеньков, и когда наступает
засуха, растение
переживает
ее, сохраняя
только
эти клубеньки.
Прямой противоположностью этому карлику были великаны
палеозойского мира — лепидодендроны, или чешуедревы.
Лепидодендроны образовали
прямые крепкие
стволы
до 2 м в поперечнике и до 30 м вышины, их многочисленные
вильчато-разветвлявшиеся ветви образовывали густую
крону и были плотно одеты длинными
узколанцетными листьями. С возрастом листья опадали,
оставляя после себя ромбовидные
рубцы, сплошь одевавшие поверхность старых ветвей и
даже главного ствола.
Органы плодоношения состояли из похожих на шишки плотных
колосков, на листьях которых сидели в нижней части
колоса макроспорангии,
а в верхней
— микроспорангии.
Из спорангиев сыпались макроспоры и микроспоры, в которых
внизу на почве происходило развитие маленьких, почти
не выдававшихся наружу,
заростков-гаметофитов.
В
макроспорах на заростках развивались архегонии с яйцеклеточками,
в микроспорах — антеридии с сперматозоидами. Если их
заливала дождевая или болотная
вода, то сперматозоиды начинали двигаться, проникали
в архегонии и
вызывали образование
зигот, прораставших с течением времени и дававших снова
спорофит — дерево лепидодендрона. Пока отдельные части
лепидодендронов
попадались изолированно,
их считали за
самостоятельные растения и давали им родовые назначения.
Так,
например, лишенные коры окаменелые стволы назывались
кнорриями, ветви — галониями
и лепидофлойос
(Halonia и Lepidophloios), а колоски — лепидостробусами.
В течение всего девонского и всего каменноугольного
периодов лепидодендроны
были
украшением
приморских
лесов Земли на пространстве нескольких миллионов лет,
а затем вымерли, не перейдя ни в какую высшую форму.
Контраст между простейшим по строению карликом филлоглоссум
и великаном лепидодендроном нисколько не объясняет
происхождения плауновых
растений. Загадка может быть
несколько расшифрована лишь сближением их с силурийскими
псилофитами (класс Psilophytales, см. рис. 5), не имевшими-корней
и имевшими
лишь слабо развитые
листья. Спорангии псилофитов сидели не на листьях,
а на концах веточек. Их простейшие представители, напр,
риния,
очень
напоминают некоторые
бурые водоросли,
а высшие, как, напр., астероксилон. сходны с плаунами.
Относящаяся сюда же хорнеа (Hornea Lignieri Kidston
et
Lang) обладает
лопастными клубнями,
напоминающими
не то заросток, не то клубень филлоглоссума. Псилофиты
— первые достоверные растения суши, и возможность выделения
типа плауновых
из форм, принадлежавших
к этой группе, весьма велика.
Высшие представители плауновых с их шишковидными колосьями
примыкают к первичным формам голосеменных растений.
Замечательно, что в современном мире растений ближайшими
к лепидодендронам растениями являются представители
рода селагинелла
(Selaginella),
имеющего до 700 видов. Большинство их — небольшие травянистые
растения, вечнозеленые,
с макро- и микроспорангиями.
В лесах Малайского архипелага есть более крупные виды
до 0,5 м вышины, а в Южной Америке даже и в несколько
метров
длины,
но это
не прямостоячие,
а цепляющиеся
растения. Наконец, близок к плауновым, но резко отличается
от них ныне живущий псилотум (Psilotum), широко распространенный
во влажных
лесах
тропического и в некоторых местностях субтропического
пояса вплоть
до Японии. Растет
он
обычно на коре деревьев и состоит из коралловидного
корневища, ветви которого одеты всасывающими волосками,
и прямостоячих
зеленых вильчато-ветвистых
стеблей. Листья очень маленькие в виде острых чешуи,
спорангии округлые трехгнездные,
довольно крупные. Корневище дает многочисленные почки,
похожие на почки мхов, быстро разрастающиеся в новые
растения. Образующиеся
из спор
заростки всего
от 2 до 20 мм длины, цилиндрической формы и без хлорофилла.
Казалось бы, все это сближает псилотум с плаунами и
может выяснить родство последних с псилофитами, так
же, как
и он, не имевшими
корней, но беда
в том, что сперматозоиды его оказались многоресничковыми,
что резко отличает псилотум
от настоящих плауновых.
Мы уделили так много внимания плауновым растениям потому,
что они представляют собою основную ветвь папоротникообразных
растений.
Несомненно происхождение их от водорослей через псилофитов
или другую вымершую группу, и от них, скорее чем от
какой-либо другой
группы,
возникли голосеменные,
это высшее звено эволюционного ряда плаунообразных
растений.
Кроме основного ряда, развитие растительного мира вылилось
в образование боковых рядов, не переходящих в высшие
формы, но
застывших на определенном
уровне.
Таков в первую очередь ряд Articulatae или «членистые».
Эти растения, стебель которых состоял и состоит теперь
из резко
отчлененных
члеников, или междоузлий,
заканчивающихся узлами, на которых сидят мелкие по
отношению к размерам междоузлий листочки, образуя кольца
или мутовки.
Листья, несущие
спорангии и называемые
спорофиллами, у них резко отличаются от
обычных и собраны в колоски на верхушке стебля или
его ветвей.
Сюда относятся 4 подкласса и 8 порядков; три вымерших
подкласса обнимают 5 порядков, а в единственном, имеющем
живых представителей,
2 порядка
из трех
также вымерли. Сохранился только один: хвощи Equisetacea.
Наиболее примитивной группой среди членистых растений
являются Calamophyton primaevum Krausel et Weyland
из среднего девона
Германии и гиения
(Hyenia elegans тех же авторов). Несмотря на членистость
стеблевых органов
и строение колосков,
в этих ископаемых есть что-то, напоминающее псилофиты.
Это что-то, вероятно, относится к сравнительной близости
между
псилофитами
и каламофитами, с одной стороны, и бурыми водорослями
— с другой.
Второй подкласс — псевдоборнии из верхнего девона Медвежьего
острова и третий — сфенофиллы, распространенный в осадочных
пластах от
верхнего девона
до гондванских
слоев нижнего триаса, были травянистыми растениями
с тонкими сравнительно стеблями и сложно построенными
колосками.
Листья сфенофиллов, узкие
у основания, расширялись
кверху, почему их называют часто клинолистами. Образ
жизни
этих растений с их тонкими стеблями недостаточно известен.
Немецкие
палеонтологи
считают их
растениями водными, английские же видят в них растения
наземные, может быть, вьющиеся или лазящие.
Четвертый подкласс — хейростробы (Cheirostrobineae)
гораздо моложе трех первых, ибо окаменелые остатки
его спороносных
колосьев
или шишек известны
из верхнего
карбона.
Наконец, пятый подкласс — хвощевые с вымершими порядками
астерокаламитов и каламитов. Астерокаламиты жили в
период от верхнего девона
до нижнего карбона
и отличались узкими вильчато разделенными в верхней
части листьями. Каламиты пышно развились в каменноугольное
время и дожили до
триаса, их главное
отличие — разноспоровость (деление спор на макроспоры
и микроспоры). Они образовывали
большие заросли в мелководьях и достигали крупных размеров.
От современных хвощей они отличались еще тем, что в
их
спороносных колосьях часть
листьев оставалась бесплодною и, загибаясь концами
кверху, одевала
шишку или
колос своими окончаниями, придавая ей большую защищенность.
Современные хвощи — остаток типа, когда-то несравненно
более богато и разнообразно представленного, построены
значительно
проще каламитов.
В конце палеозоя
хвощи (Equisetales) были представлены несколькими семействами
и многими родами, из
которых сохранился только один род Equisetum, так как
все остальные исчезли
уже до начала третичного периода.
В настоящее время сохранилось всего до 30 видов хвощей,
распространенных почти по всему свету, кроме Австралии,
где их нет вовсе.
Настоящие папоротники ведут свое начало от группы первичных
папоротников Primofilices, живших в период времени
от девона до перми. Это
были относительно мелкие растения
с лазящими корневищами или с прямыми короткими стеблями.
Листья этих примитивных папоротников по большей части
были лишены
плоских пластинок,
и их конечные
разветвления имели цилиндрическую форму. У рода стауроптерис
нет еще полного разделения на стебель и листья, так
как листья почти
не отличаются
от ветвей.
Спорангии со стенкой из одного слоя клеток и лишены
какого-либо механизма для выбрасывания спор, которые
освобождаются
из спорангия через маленькое
отверстие
на его верхушке.
Все более возрастающая сложность в строении листовых
пластинок, или приспособление их к специальным условиям
жизни, а также
усложнение в строении спорангиев
и особенно в строении раскрывающего спорангий аппарата,
у большинства папоротников выраженного в так называемом
кольце,
— таковы
основные направления в истории
настоящих папоротников.
Главнейшее деление папоротников на группы дает возможность
различить между ними три основных подкласса.
Первый подкласс — это папоротники с массивными спорангиями
Eusporangiatae, у которых стенка спорангия состоит
из нескольких слоев клеток
(в толщину), с выделением покровных тканей, наружной
(эпидермис) и
внутренней (гиподермис).
В настоящее время они представлены двумя семействами:
мараттиевых папоротников в лесах жарких стран и мелких
ужовниковых
в умеренном поясе и в странах
севера. Первые развились уже в каменноугольную эпоху,
были очень многочисленны в пермскую
и достаточно богато представлены в более поздних отложениях.
Роды псарониус (Psaronius) и пекоптерис (Pecopteris)
с древовидными стволами и троякоперистыми
крупными листьями напоминают современные нам древовидные
папоротники Бразилии и других влажных тропических стран.
По свидетельству А. Н. Криштофовича, ни один из ныне
существующих видов мараттиевых папоротников не найден
с достоверностью
в ископаемом состоянии,
хотя из мезозоя
и известны остатки, очень напоминающие современные
роды ангиоптерис, данеа и мараттия, сложившиеся не
ранее расцвета
третичных
флор.
Рис. 24. Современный папоротниковый лес в Бразилии 1 — циатея 2 — гемителия |

Спорангии ужовниковых папоротников одиночные, тогда
как спорангии мараттиевых представляют то усложнение,
что
они соединены группами в так называемые синангии, что
лучше защищает развивающиеся споры.
Второй подкласс папоротников — Protoleptosporangiatae
— состоит из одного семейства чистоустовых Osmundaceae.
Стенка
спорангия
построена
у них
из одного слоя клеток,
но раскрывающее спорангий колечко примитивное — в форме
бобовидной пластинки сбоку от верхушки его. Сейчас
это небольшая группа,
обнимающая всего
3 рода и до 20 видов умеренного жаркого пояса. Замечателен
род тодеа (Todea)
с одним
всего видом Т. barbara, растущим в горных ущельях южной
Африки, юго-восточной Австралии и Новой Зеландии; многие
видят в
нем потомка гондванской
флоры, расселившегося некогда на всем протяжении гондванского
материка и уцелевшего
на его обломках.
Ближе к нам чистоуст Osmunda regalis, растущий в Южной
Европе и достигающий крупных размеров в лесных болотах
Черноморского
побережья
Абхазии.
Ископаемые родственники этих папоротников известны
из верхнепермских слоев Белебея,
из юрских отложений Новой Зеландии, меловых Британской
Колумбии и Западной Европы.
Наиболее древними являются дископтерисы среднего карбона,
наиболее молодыми осмундитес, жившая в период от нижней
юры до миоцена.
В общем к группе
осмундовых, или чистоустовых папоротников относится не
менее 15 ископаемых родов и до 40 видов. Можно думать,
что эта группа в настоящее время
угнетена общим изменением условий жизни на Земле.
Третий подкласс папоротников — это лептоспорангиаты
(Leptosporangiatae), т. е. папоротники, спорангии которых
имеют тонкую однослойную
стенку и хорошо
развитое колечко из клеток с сильно утолщенными оболочками,
обеспечивающее рассеяние спор при раскрывании зрелого
спорангия. Сейчас эта
группа распространена широко по всей Земле, достигает
редкого разнообразия
форм и массового
развития особей и лишь на севере, в областях, испытавших
непосредственное действие
последнего послетретичного великого оледенения, сведена
к десятку-другому видов. В нее
входят папоротники с разнообразнейшими формами листьев,
с разнообразнейшим анатомическим их строением сообразно
тене-
или световыносливости
данных видов, а также с весьма разнообразным строением
споровых кучек и даже
спорангиев. Разнообразие это таково, что мысленно легко
представить себе, как выглядела
бы Земля, если бы на ней не было других растений, кроме
папоротников.
Папоротниковые леса, папоротниковые заросли в низинах
и по склонам гор и холмов, папоротники скал, папоротники
водоемов
и болот
дают достаточно полный растительный
покров. Только степи, пустыни и тундры лишены папоротников.
Лептоспорангиатные папоротники обнимают 12 семейств,
более 120 родов и около 300 видов. Их ископаемые остатки
менее
разнообразны, начинают
встречаться
только
с каменноугольной эпохи, гораздо более обильны в юре
и в мелу и еще лучше представлены в отложениях третичного
периода.
Знакомство
с
этими ископаемыми
папоротниками
приводит к мысли, что развитие их не давало переходов
к
вышестоящим типам, а шло путем дифференцирования основного
типа и выработки
приспособлений к внешней постоянно изменявшейся среде.
Из 12 семейств лептоспорангиатных папоротников в каменноугольных
отложениях представлены только три (Schizeaceae, Gleicheniceae
и Hymenophyllaceae),
в юре уже 10, одно (Salviniacea) известно только с
верхнего мела и одно (Parkeriacea)
не известно в ископаемом состоянии вовсе. Несомненно,
что наиболее богатые видами современные семейства диксониевых,
циатовых
(Cyatheaceae) и полиподиевых
папоротников были даже в третичную эпоху менее богаты
видами,
чем теперь. Словом, их развитие продолжается и в наши
дни.
В этом обзоре папоротникообразных совершенно не использованы
данные по анатомии стеблей, а между тем метод тонких
шлифов ископаемых образцов со времен французского
ученого Марселя Рено дал такой богатый материал для
познания строения
и расположения сосудисто-волокнистых пучков папоротников,
хвощей и плаунов,
что постепенное
совершенствование проводящей системы этих растений
в настоящее время выяснено
великолепно и весьма помогает установлению их родства
и места в системе.
7. ПРОИСХОЖДЕНИЕ ШИШКОНОСНЫХ
И ДРУГИХ ГОЛОСЕМЕННЫХ
РАСТЕНИЙ
У папоротникообразных семян не было. Они зарождаются
а в виде спор, очень мелких и слабых носителей жизни,
а развиваются
на почве без
какого бы
то ни было участия
материнского организма.
Семя — большой шаг вперед. Семя образуется и вызревает
медленно, все время получая питание от материнского
организма, пока
в нем не образуется
зародыш,
состоящий из корешка, почечки и семядолей, т. е. зародышевых
листьев. Кроме того, у большинства семенных растений
вокруг или около зародыша
развивается
эндосперм (иначе внутрисеменник), орган, представляющий
собою настоящий магазин питательных веществ, который
помогает прорастающему
зародышу
в его
питании и росте*. Семенные растения имеют перед споровыми
большое преимущество в быстроте и силе развития и всюду,
где их много,
вытесняют споровые.
| * У многих растений эндосперм составляет главную по объему часть семени, например, у пшеницы. У других растений, как у гороха, эндосперм не развивается, и место его в семени занимают семядоли с их запасом питательных веществ. |
Уже в среднедевонских пластах, когда растительный мир
только начал слагаться, появились крупные растения
с листьями
и стволами папоротников,
но на
листьях у них не было спор. Вместо спор на краях листовых
долей помещались несомненные
семена, в которых и происходило заложение и развитие
зародыша. Растения, построенные таким образом, названы
семенными
папоротниками, или птеридоспермами.
Продержались
эти первые на Земле семенные растения вплоть до юрского
периода, но наиболее многочисленными были они в каменноугольное
время.
Несколько позднее семенных папоротников возникли кордаиты,
имевшие уже анатомическое строение хвойных деревьев.
Семена их были уже
не на листьях,
а на ножках и
сидели в пазухах прицветных листьев по одному, образуя
своеобразные «соцветия». Кордаиты имели общих предков
с семенными папоротниками,
время расцвета
обеих этих групп совпадает, как и время их угасания.
Третья группа голосеменных растений — это саговники,
небольшие деревья, не то папоротник, не то пальма,
часто с перистыми
жесткими длинными
листьями. Семена их сидят или по краям коротких шерстистых
перистых листьев, или
на чешуях,
собранных в шишки. Они не ветвятся или, вернее, ветвятся
лишь в исключительных случаях, и листья их собраны
короной у верхушки.
Саговники происходят
от семенных папоротников и особенно многочисленны и
разнообразны были в юрское
время. Теперь
их осталось немного, и живут они в тропиках и вблизи
тропиков, доходя на север до южной части Японии. К
саговникам были
близки так называемые
беннеттиты,
плодоношение которых напоминает цветок. Их остатки
чаще всего находятся
в нижних
пластах меловой системы. Одно время видели в беннеттитах
прародителей цветковых растений, но затем это предположение
было оставлено.
Четвертая основная группа голосеменных растений — это
группа гинкговых. Гинкго — название японское, так как
это чудесное
дерево было впервые
описано по культурным
японским экземплярам. Позднее его нашли и в Китае.
В живых сохранился только один вид, но в ископаемом
состоянии
известно
несколько
родов и много видов.
Возникли они от кордаитов еще в девонское время, когда
появился род псигмофилл, несомненно
принадлежащий к этому классу. Больше всего их было
в юрское время, а в мелу наступил упадок, вызвавший
сильное
сокращение
числа
видов. В третичное
время
жил только современный вид гинкго.
Современные нам хвойные деревья возникли уже позднее
каменноугольного времени. Первое достоверное хвойное
дерево «вальхия» впервые
появляется в красноцветных
толщах пермского времени. Многие исследователи считают,
что первые хвойные произошли от древовидных девонских
или каменноугольных
плауновых деревьев.
Первый расцвет хвойных относится к юрскому времени,
когда уже существовали араукарии; настоящие сосны появляются
только в
верхнем
мелу; ель
старше сосны и появилась уже с нижнего мела.
Рис 25. Семенной папоротник Lyginodendrum oldhamium и его семя, сидящее
в чаше, края |
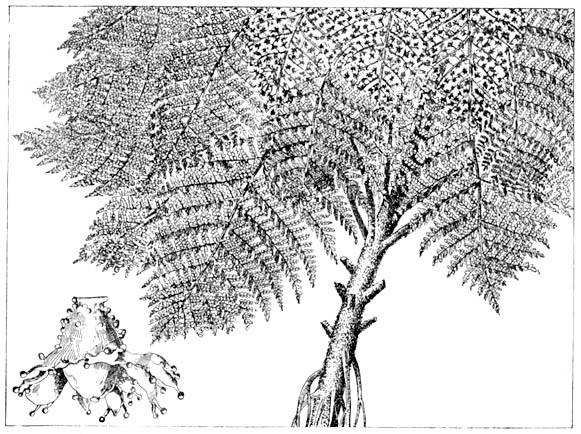
В настоящее время наиболее важный для нас класс голосеменных
— это шишконосные Coniferae, распадается на 5 семейств:
тиссовые, араукариевые,
еловые,
таксодиевые и кипарисовые. Попробуем определить время
и порядок возникновения каждого
из них.
В настоящее время семейство араукариевых представлено
двумя родами, именно даммарой в Новой Зеландии и араукарией
в
Бразилии, Чили,
на островах
Полинезии, в восточной части Австралии и Новой Гвинее
— словом, исключительно в Южном
полушарии. Семейство это является древнейшим из шишконосных,
причем род араукария имел несомненных представителей
в юре в Англии, Китае,
Франции
и др., в мелу
в Северной Америке, на Сахалине и в Чехии. Креузель
приводит один вид (Araucaria Delafondi Zeller) даже
из перми,
сохранившийся в
виде односеменных
чешуи
из разрушенных шишек. В эоценовое время араукарии были
широко распространены в
лесах Северного полушария. В Южном полушарии они ранее
были распространены также шире, чем в наше время, и
ближе подходили
к полюсу. Так,
одна из них, Araucaria imponens Dusen, найдена в третичных
отложениях ныне оледенелого
острова
Сеймур под 64° ю. ш., a A. Nathorsti — в олигоцене
близ Пунта Аренас, на самом юге Южной Америки в безлесной
ныне местности.
Даммара
также
известна
в ископаемом
состоянии из эоценовых отложений Волыни и из более
поздних
отложений с Новосибирских островов.
Ясно, что в семействе араукариевых мы имеем не развивающуюся,
а скорее угасающую группу, о чем нельзя не пожалеть,
так как прекрасная
древесина
и крупные съедобные
семена араукарий, древесина и ценная смола даммары
далеко не лишние и в наше время.
Если теперь начать поиски возникновения этой группы,
то по анатомическим особенностям ископаемых древесин
распространенный
в пермских
и каменноугольных отложениях
род
дадоксилон приводит нас к классу кордаитов. тогда как
последние упираются в некоторые роды плауновых.
Семейство тиссовых отличается от всех остальных шишконосных
тем, что его семяпочки сидят по одной на концах веток,
а тычинки имеют
щитовидную
форму.
В настоящее
время тиссы распространены в горных лесах Европы, Азии
(Гималаи, Китай, Дальний Восток, Япония) и Северной
Америки, другой
род, торрея, — в
лесах Флориды,
Калифорнии, Японии и Китая, третий, аустротаксус, —
во влажных лесах острова Новая Каледония. Все это —
отдельными
изолированными
один
от другого участками,
указывающими на более сплошное и более широкое распространение
в прошлом.
Ископаемые остатки тиссовых были найдены в юрских отложениях
Швеции и Англии, в мелу Гренландии, Северной Америки
и Франции, в третичных
отложениях
Аляски,
Италии, Германии. В четвертичных отложениях семена
и древесина тисса встречаются и в таких местностях,
где
теперь его
нет и в помине.
Словом, тиссовые —
исчезающая группа. Никаких указаний на происхождение
их из папоротникообразных в нашем
распоряжении не имеется. Возможно, что придется искать
родственных отношений между тиссовыми и гинкговыми.
Семейство подокарповых, или многоплодниковых, состоит
из семи родов и 101 вида, распространенных, главным
образом, в Южном
полушарии.
Лишь некоторые
виды рода
подокарпус растут на островах Малайского архипелага
и даже
в южной Японии.
В ископаемом состоянии многоплодниковые попадаются
в юрских отложениях Франции и Швеции, в меловых и третичных
отложениях
(эоцен) Северной
Америки, Гренландии,
Огненной Земли и средней Европы. Возникновение этого
семейства, очень обособленного по своей морфологии,
вероятно,
то же,
что и семейства
тиссовых.
Семейство цефалотиссовых (Cephalotaxaceae) свойственно
исключительно Гималаям, горам Китая и Японии. В ископаемом
состоянии они
известны для меловых отложений
Северной Америки, для миоцена Шпицбергена и Западной
Европы, для плиоцена Германии. Вопрос о происхождении
и этой группы
однороден с вопросом
о происхождении тиссовых.
Семейство таксодиевых — Taxodiaceae, куда относятся
зонтичная сосна Китая, мамонтово дерево Калифорнии
и болотный кипарис
Северной Америки, хорошо
представлено было уже в юрскую эпоху, когда зонтичная
сосна (Sciadopitys) росла в Швеции
и Норвегии, мамонтово дерево (Sequoia) во Франции и
продолжало развиваться в меловую и третичную эпохи,
достигнув в
эоцене своего наиболее полного
процветания. По своей морфологии это семейство может
быть сближено с семейством еловых и
имеет, вероятно, общее с ним происхождение.
Семейство еловых, или сосновых (Pinaceae), обнимает
9 родов и более 210 видов, распространенных частью
в равнинах
севера,
где
некоторые
из них
образуют особую
зону хвойных деревьев, главным же образом в горных
лесах умеренного пояса, заходя в тропики лишь немногими
представителями
рода
Pinus — сосна. В
ископаемом состоянии пихты достоверны только для плиоцена
Европы, ели для нижнего
мела Бельгии и позднейших отложений, лиственницы достоверны
для плиоцена Франции,
настоящие кедры попадаются уже в нижнем мелу и, конечно,
позднее: сосны, которых сейчас насчитывается около
ста видов, начали
развиваться с
начала мелового
периода, причем в прошлом известны и для таких стран,
как Гренландия. Сосны и сейчас богаты разновидностями
и формами
недавнего происхождения,
что
приводит к заключению, что они продолжают развиваться,
вырабатывая новые приспособления
к среде и новые виды.
Семейство кипарисовых обнимает 16 родов и более 130
видов, рассеянных почти по всему свету. Самым крупным
и наиболее
жизненным является
род Juniperus,
или можжевельник, продолжающий и теперь свое развитие.
Начало развития всего семейства кипарисовых относится
к началу меловой
эпохи. Древесина
можжевельника
указана для миоцена Европы, пыльниковые шишки его же
— для янтарей Прибалтики.
Таким образом, развитие голосеменных шло далеко не
преемственно. В палеозое господствовали замечательные
семенные папоротники
и кордаиты. Первые
дали начало саговникам и беннеттитам мезозоя, вторые
гинкговым и, может
быть,
араукариям.
Мезозойские напластования, нередко дающие повод к установлению
особой эры голосеменных параллельно эре пресмыкающихся,
особенно динозавров,
действительно
отличаются
обилием остатков гинкговых и саговниковых с примесью
араукарий. Кайнозойские группы, возникшие с началом
мела, конечно,
не возникли из ничего,
надо полагать, что в это время разнообразие голосеменных
было уже так велико,
что дальнейший
процесс развития базировался уже не на папоротникообразных,
а на гинкговых и араукариевых. Во всяком случае наиболее
вероятной является
следующая
филогения голосеменных растений:
| Первичные папоротники |
Кордаиты |
| Семенные ------»------ |
Гинкговые и араукарии |
| Цикадовые или саговники | Тиссовые. Еловые |
| Беннеттиты . | Кипарисовые |
| Лепидодендроновые |
Остается еще один, седьмой класс голосеменных — это
гнетовые (Gnetales). Он выделен на основании того,
что у
всех его представителей внутренний покров семян
вытянут в трубку, представляющую отдаленное сходство
со столбиком
цветковых
растений.
Но столбик есть
вытянутая часть завязи и развивается за счет плодущих
листьев, составляющих завязь,
трубка же гнетовых — часть семени. Функция и того
и другого органа сходная — регулировка
прорастания пылинок цветени и роста пыльцевой трубки.
Кроме того, гнетовые отличаются еще тем, что имеют
настоящие сосуды во вторичной
древесине,
т. е. в древесине, развивающейся за счет камбиального
кольца,
лежащего на границе
между корой и заболонью, чего нет у других голосеменных,
вторичная древесина которых состоит только из замкнутых
водоносных клеток,
называемых трахеидами.
Этот признак приближает гнетовые к цветковым и
дал повод некоторым авторам утверждать, будто гнетовые
происходят
от цветковых
путем упрощения морфологического
и анатомического строения, с чем согласиться никак
нельзя.
Три рода гнетовых: тропические деревья и лианы
рода гне-тум, безлистные кустарники рода хвойник,
или
эфедра, и совершенно
ни на какое другое
растение не похожая
африканская вельвитчия достоверных ископаемых остатков
не оставили, морфология их совершенно своеобразна,
и только белковая преципитиновая
реакция сближает
их с настоящими шишконосными. Высказывавшаяся в
литературе мысль о происхождении цветковых растений
из гнетовых
не подтвердилась
позднейшими исследованиями.
В лице гнетовых мы имеем глухую ветвь развития
или даже три ветви,
объединенные общностью фазы развития.
8. ПРОИСХОЖДЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ
Цветковые растения называются также покрытосеменными.
Это значит, что их семена скрыты внутри плодов.
Их зародыши, кроме той
защиты, которую
им
дают покровы
семени, защищены еще и стенкой плода, что еще
более обеспечивает их вызревание. Кроме того, они наиболее
совершенны и
в
анатомическом отношении,
что
обеспечивает им наилучшее использование солнечного
света, газов воздуха и воды с почвенным
раствором. Они гораздо совершеннее и быстрее
приспособляются к их среде, чем голосеменные или папоротникообразные,
почему и составляют
в настоящее
время
главную массу растений суши.
Цветок, привлекая своим ароматом и нектаром
насекомых или выставляя навстречу ветру свои
пылинки, много
способствует скрещиванию
растений и образованию
путем скрещивания новых форм, которые потом
благодаря действию
естественного отбора
становятся основным населением какой-либо
соответствующей по своим климатическим и почвенным
свойствам территории.
Иначе говоря, цветок усиливает шансы на скрещивание
между различными особями какого-либо вида,
вызывает так называемую
амфимиксию,
что усиливает пластичность
потомства, являющегося наследственно неоднородным.
Неоднородность потомства обусловливает действие
естественного отбора,
а при этом местные условия,
как фактор отбора, вызывают соответствующие
отклонения, которые и позволяют потомству
данных растений выживать при изменившихся условиях.
Расселяясь по всему лику З'емли и приспособляясь
к местным условиям,
цветковые растения
достигли своего
нынешнего разнообразия, выражающегося цифрою
до 500000 видов.
Само собой разумеется, что такого разнообразия
в первый период после появления на Земле цветковых
растений
быть не могло.
А. Энглер, величайший
знаток
систематики цветковых, начавший свою деятельность
с исторического исследования о растительности
наших дней, как наследнице третичного периода*,
к
концу жизни пришел к убеждению, что появлению
цветковых на
Земле предшествовало
развитие
группы
первичных покрытосеменных
растений Protoangiospermae, возникших в мезозойскую
эпоху ранее верхней юры и отличавшихся более
простым цветком.
| * А. Еngler. Versuch einer Entwicklungsgeschichte der extratropischen Florengebiete der Nordlichen Hemisphare, I, 1879; II, 1882. |
Цветковые растения появляются на Земле в значительном
разнообразии видов только с нижнего мела. Происхождение
их от какой-либо
группы голосеменных
растений
несомненно, но от какой именно группы, пока
наверно сказать нельзя.
Какие из цветковых растений старше других,
какие моложе? Одни авторы признают за древнейшие
цветковые
растения
так называемые
сережчатые
(береза, ива,
тополь, орех и пр.), с мелкими почти лишенными
околоцветника цветами, опыляемыми ветром;
другие — пышные цветы магнолий с массою плодников
и тычинок, находя, что они ближе всего походят
на шишки
голосеменных
деревьев. За
первых долго
высказывалась
и палеонтология, так как остатки тополя, найденные
в Гренландии в слоях нижнего мела, одно время
считались даже древнейшими
остатками цветкового
растения,
какие только известны. Остатки магнолий найдены
лишь в среднемеловых отложениях. На очереди
самые тщательные
поиски
на рубеже
юрских и
меловых отложений,
так как окончательно решить поставленный вопрос
вряд ли можно иным способом.
К новейшим по происхождению цветковым растениям
принадлежат растения, цветки которых собраны
в плотные
обильные соцветия. Особенно выделяется в этом
отношении семейство сложноцветных, к которому
принадлежат
василек, одуванчик
и подсолнечник. Их остатки
появляются только с середины третичной эпохи,
а пышное развитие принадлежит уже настоящему
времени.
9. ПРОИСХОЖДЕНИЕ КУЛЬТУРНЫХ РАСТЕНИЙ
Спутники человека, культурные растения, возникли,
разумеется, лишь после того, как появился
и окреп в своей хозяйственной
жизни сам
человек. Первой
стадией
этой жизни был охотничий быт — человек
убивал животных и питался их мясом. Однако потребность
в растительной
пище сказывалась
и в этом
периоде.
Мужчины охотились, женщины собирали и хранили
ягоды и другие плоды, мясистые корни,
клубни, луковицы и другие части растений,
богатые крахмалом или другими питательными веществами.
Первые путешественники,
посетившие
Камчатку,
Стеллер и Крашенинников,
застали еще в этой отдаленной стране подобный
быт, причем единственными орудиями, известными
людям,
оказались орудия
из камня и рыбьих
костей; железа они вовсе
не знали.
Позднее начинаются робкие попытки земледелия.
Расчищали лес, делали заостренным шестом
ямки и в них бросали
зерна. Начало
земледелия
установить теперь
так же трудно, как и появление на земле
определенных групп растений или животных.
Какие именно растения были впервые выбраны
человеком из дикой природы и послужили
предметом
культуры,
сказать трудно.
Так
же трудно в
настоящее время точно установить,
какое дикорастущее растение стало родоначальником
того или другого культурного.
Достоверно, однако, что земледельческая
культура возникла самостоятельно в нескольких
отдаленных
одна от другой
странах, которые и стали
первичными центрами
земледелия.
Так, юго-восточная Азия, Индокитай и Малайский
архипелаг дали человечеству культуру риса
и, вероятно, сахарного
тростника и банана. Средняя
Америка стала родиной культуры кукурузы
и шоколадного дерева. Южная Америка
разработала культуру
картофеля; в северной Мексике возникла
культура подсолнечника, на островах Вест-Индии
— культура
табака.
В Европе мы находим свой важнейший центр
земледелия. Это побережье Средиземного
моря, где возникла
самостоятельно культура твердых
пшениц, льна, оливки,
свеклы, капусты и многие другие. В Иране,
Туркестане и
Афганистане развилась первоначальная
культура мягких пшениц, ячменя, тыквенных
растений п пр. Отсюда же вышли и северные
культуры ржи
и овса.
Пытаясь узнать, откуда произошли культурные
растения, мы легко находим их родину, но
редко когда нам
удается найти
их дикого
родоначальника. Мы уже нашли дикую
свекловицу, дикую морковь, дикую капусту
и дикую коноплю, но найти дикий картофель
не
удалось.
Еще труднее обстоит
дело
с хлебными
злаками.
Исследуя
породы этих
необходимейших спутников человека, мы убеждаемся
почти для каждого из них, что в его возникновении
участвовало
не одно
дикое растение,
а два
или несколько.
Так как помеси от скрещивания двух разных
видов растений обыкновенно отличаются крупным
ростом
и пышностью
развития, то естественно,
что уже первобытные
земледельцы каменного века обратили внимание
именно на них и размножили помеси хлебных
злаков. Подвергая имевшийся у них посевной
материал отбору, люди постепенно выработали
нужные им
хлебные растения,
никогда не
существовавшие в
природе, но созданные ими самими.
Таким образом создалась новейшая растительность
Земли — культурная. Наше поколение является
свидетелем того, как
исчезают леса
и степи, как всюду
раскидывается
золотой покров зреющих полей, чередующихся
с зеленью огородов и садов.
Растительность, созданная историей Земли
и проявившаяся так пышно в развитии третичных
лесов, все более
и более оттесняется
на
задний план.
Ее прибежище
в наше время — главным образом горы, пустыни,
скалистые острова и другие не удобные для
земледелия
места.
Правда, на Земле еще много незанятых земель,
но быстрый рост человечества за последние
десятилетия показывает,
что в скором
времени все пригодные
для земледелия
территории будут заселены и заняты полями
и садами.
Остается еще вопрос о заповедниках. Почти
во всех странах правительства выделяют
более или
менее
крупные участки
с сохранившейся еще
природой и делают их частью
предметом изучения, частью территорией
для отдыха. Охраняют горные леса, так как
безлесные
склоны
легко размываются
дождевыми водами,
а массы
камней и мелкозема,
снесенные в долину, уничтожают или портят
пашни.
Недалек тот день, когда заповедники и заказники
различного типа одни только сохранят разнообразную
природную
растительность наравне
с
неприступными для культуры местами, все
же остальное будет занято растениями культурными,
выращенными
и созданными человеком.
ОГЛАВЛЕНИЕ
Предисловие............................................................................................................................................................
3
Глава I. Место зеленых растений в
космосе и значение их для человека ........................................................
7
Глава II. Земля до возникновения на
ней растений...........................................................................................
12
Глава III. Как зародилась жизнь на
Земле...........................................................................................................
21
Глава IV. Круговорот жизни и круговорот
вещества в природе......................................................................
34
1. Углерод .............................................................................................................................................................
35
2. Кислород............................................................................................................................................................
37
3. Водород..............................................................................................................................................................
39
4. Азот.....................................................................................................................................................................
40
5. Зольные составные части.................................................................................................................................
42
Глава V. Понятие об эволюции. Палеонтологическая
летопись .....................................................................
46
Глава VI. Архейская группа слоев.......................................................................................................................
57
Глава VII. Палеогеография и условия
жизни прошлых геологических периодов...........................................
60
Глава VIII. Палеозойская группа
слоев...............................................................................................................
64
1. Кембрийская система слоев.............................................................................................................................
64
2. Силурийская система слоев.............................................................................................................................
66
3. Девонская система слоев..................................................................................................................................
69
4. Каменноугольная система слоев......................................................................................................................
72
5. Пермская система слоев....................................................................................................................................
84
Глава IX. Мезозойская группа систем
осадочных образований........................................................................
88
1. Триасовая система слоев...................................................................................................................................
88
2. Юрская система слоев........................................................................................................................................
90
3. Меловая система слоев......................................................................................................................................
96
Глава X. Кайнозойская группа слоев.................................................................................................................
102
1. Третичная система слоев.................................................................................................................................
103
2. Четвертичная система слоев и ледниковый период ....................................................................................
118
Глава XI. Как произошли отдельные
группы растений ............................................................................
129
1. Происхождение бактерий..............................................................................................................................
129
2. Происхождение водорослей..........................................................................................................................
134
3. Происхождение грибов..................................................................................................................................
138
4. Выход растения на сушу и происхождение наземной растительности ...............................................
142
5. Происхождение мхов......................................................................................................................................
146
6. Происхождение папоротникообразных растений .....................................................................................
150
7. Происхождение шишконосных и других голосеменных растений........................................................
159
8. Происхождение цветковых растений.........................................................................................................
165
9. Происхождение культурных растений ......................................................................................................
167
Глава XII. Как произошли главнейшие
органы высших растений...................................................................
169
1. Как произошел корень.......................................................................................................................................
169
2. Происхождение листа.......................................................................................................................................
172
3. Происхождение стебля......................................................................................................................................
173
4. Происхождение женского гаметофита цветковых растений ........................................................................
175
5. Происхождение мужского гаметофита цветковых растений ........................................................................
179
6. Происхождение цветка......................................................................................................................................
181
7. Происхождение семян.......................................................................................................................................
183
Заключение.............................................................................................................................................................
186