Г л а в а XII
КАК ПРОИЗОШЛИ ГЛАВНЕЙШИЕ
ОРГАНЫ ВЫСШИХ РАСТЕНИЙ
Многие из тех заключений, к
которым мы пришли в предшествующих главах, все еще недостаточно доказаны.
Неполнота палеонтологической летописи, которая в
свое время так беспокоила Ч. Дарвина, все еще недостаточно пополнена и
не позволяет довольствоваться ею одной, необходима поддержка со стороны других
отделов ботаники.
Каждый орган растений прошел, пока не стал таким, каким мы его видим, длинную
цепь изменений, и нам интересно проследить хотя бы главнейшие этапы его
развития.
1. КАК ПРОИЗОШЕЛ КОРЕНЬ
Водоросли, как живущие в морях солоноводные, так и живущие в пресных водах
озер, рек и пр., впитывают воду всей поверхностью. Каждая их клетка,
граничащая с внешней средой, представляет собою осмотический аппарат,
всасывающий
растворы минеральных солей и газов и отдающий растворы органических
соединений, напр.,
органических кислот и их солей. Даже те водоросли, которые живут на
коре деревьев или на скалах, одевая их зелеными (плеурококки, празиола) или
красными (трентеполия)
пятнами, поглощают воды росы или дождя всей поверхностью. Если же росы
и дождя нет, то они на время замирают, чтобы снова стать деятельными,
лишь только дождь
коснется их снова. Ясно, что у них и не может быть специальных органов
всасывания воды.
Мхи начинают свою жизнь стадией предростка, построенного наподобие
водоросли и погруженного в воду. В редких случаях, когда предросток
мха развивается
на обсыхающей почве, отдельные его ветви оказываются в земле, и на
них развиваются
длинные трубчатые клетки, называемые ризоидами, которые всасывают воду,
в то время как верхняя, находящаяся на свету часть предростка ее испаряет.
В виде исключения имеется несколько водорослей, имеющих как бы два
конца: всасывающий и испаряющий; иначе говоря, им свойственны явления
полярности.
Нижний полюс
погружен в почву и поглощает воду, верхний обращен к солнцу и ее теряет.
Такова, например, маленькая водоросль ботридий (Botrydium granulatum),
растущая на отмелях и на сырых пашнях. Вся она имеет грушевидную форму,
а нижняя ее
часть вытянута в узкую ветвистую у основания трубочку. Внутренних перегородок
у ботридия нет, строение его сифонное, и вся протоплазма, если освещение
слишком сильное, перекочевывает свободно вниз и так же свободно подымается
наверх,
если небо пасмурно. Воду ботридий поглощает нижней своей частью, разветвления
которой называются ризоидами.
Полярность, т. е. противоположение испаряющей части растения и части,
всасывающей воду,— вот первый импульс к образованию первых корнеподобных
органов.
Гаметофиты мхов, даже наиболее крупных (исключение — сфагновые мхи
и мхи подводные), поглощают воду также ризоидами, т. е. отдельными
клетками,
покрывающими нижнюю
часть стебля. Никакого подобия настоящего корня у мхов нет, как вообще
его нет ни у одного гаметофита гаплоида. Зародыш мха, развивающийся
из оплодотворенной
яйцеклетки и поэтому диплоидный, имеет палочкообразную или овальную
форму, он или вовсе не дифференцирован, или же имеет слабо отличающуюся
пяту,
с помощью которой присасывается к тканям стебля растения матери и из
них
всасывает водные
растворы.
У древнейшей группы наземных растений, у псилофитов, корней не было
вовсе, воду они вбирали, так же как и мхи, с помощью ризоидов. Только
у рода
астероксилон, побеги которого слабо напоминают плауновые растения,
есть вильчато разветвленные,
отходящие от корневища отростки, напоминающие корни.
У плаунов, хвощей и у всех папоротников, как у растений диплоидных,
корни уже вполне развиты. Они всегда невелики, отходят от корневища
или от
коры стебля
и неглубоко погружаются в почву. Ввиду того, что все эти растения жили
и живут во влажных местностях, они могут довольствоваться поверхностными
корнями.
Чем крупнее растение, чем более развита его испаряющая крона, тем более
повышается потребность в глубоко зарывающейся в почву мощной корневой
системе.
У голосеменных растений впервые появляется стержневой корень, закладывающийся
уже в период развития зародыша в семени. Такой корень дает возможность
молодому растению быстро углубиться в почву и распределить боковые
мочки в таком слое
почвы, который наилучше обеспечивает водоснабжение растения.
Однако у корня есть и еще функция — это функция прикрепления к почве,
функция механическая. Уже у крупных морских водорослей типа ламинарии
от нижней
части стебля отходят мощные ризоиды, впивающиеся в камни и удерживающие
водоросль
на определенной глубине, как бы ни били ее волны. Здесь функция всасывания
не имеет значения, все дело в механическом сопротивлении среде.
Корни наших деревьев должны отвечать огромному сопротивлению, так как
ветер очень сильно давит на обширную площадь кроны; дерево гнется,
иногда ломается,
а корни не поддаются, сохраняя связь менаду деревом и почвою. Зато
при условиях, неблагоприятных развитию корневой системы, ветровал неизбежен.
У растений, живущих на очень сухих почвах, поражает длина корней, часто
во много раз превышающая в вышину наземные части растения.
Напомним, что анатомически корень состоит из основного цилиндра, коры,
конуса нарастания и защищающего этот конус чехлика. Кора на некотором
расстоянии от
конца корня несет зону корневых волосков, усиливающих всасывание. Осевой
цилиндр содержит в себе проводящие воду сосуды, через которые она поступает
в стебель.
Клетки коры развивают колоссальное осмотическое давление, обеспечивающее
движение воды в сосудах. Если корень находится в симбиозе с грибками,
образуя микоризу,
то корневые волоски, как правило, отсутствуют.
Наконец, не надо забывать, что корень постоянно растет и своими движениями
при этом роет землю. Подвергаясь действию внешних факторов, как то:
силе тяжести, свету, теплу, влажности, присутствию кислорода воздуха
и химизму
почвенного
раствора,— корень растет неравномерно, искривляется и проникает в наиболее
благоприятные для него слои почвы. Таков результат длительной, постоянно
поддерживающейся борьбы за существование. Разумеется, и эта способность
корня ориентироваться
в почве далась не сразу, а выработалась постепенно.
2. ПРОИСХОЖДЕНИЕ ЛИСТА
У морских и пресноводных водорослей окрашено более или менее равномерно
все растение. Изредка лишены хлорофилла ризоиды, да при большой
толщине самой водоросли
внутренние ткани могут быть очень бедны хромопластами. У мхов дело
обстоит уже иначе, гаметофит мха может иметь зеленый сте-бель,
часто плоский
или крылатый, или же крыло стебля превращено в листья, морфологически
вполне
обособленные,
а у листостебельных мхов даже и с водопроводящей жилкой.
Спорофит мха содержит мало хлорофилльных зерен, которые находятся
у него частью в клетках тканей коробочки, частью в подпирающем
коробочку апофизе.
Настоящих
листьев у него нет вовсе.
У мелколиственных папоротникообразных (Lycopsida), т. е. у хвощей
и плаунов, листья принадлежат уже спорофиту и имеют нормальное
строение, но они
очень малы и работа их невелика. У хвощей ассимиляция углерода
производится главным
образом стеблями и веточками, листья в ней почти не участвуют,
их значение
иное, они охраняют от света растущую зону каждого междоузлия, чем
задерживают в ней дифференцировку тканей, а отсутствие дифференцировки
сохраняет
неизменною зародышевую ткань, главная функция которой — деление
клеток и рост всего
органа.
Первые листья, которые вполне оправдывают такое их обозначение,
— это листья некоторых псилофитов (род Asteroxylon), обладавшие,
по-видимому,
уже вполне
развитым устьичным аппаратом. Листья ископаемых и современных плаунов
следуют за ними. Эти листья все еще примитивные: у них нет завершенного
разделения
на черешок и пластинку, нет развитой сети жилок; анатомически оба
основных
аппарата листа, фотосинтетический — палисадная паренхима — и транспирационный
— паренхима губчатая, выражены несовершенно, как несовершенно и
различие между верхней и нижней сторонами листа. Правда, подобные
листья встречаются
и у растений,
принадлежащих к высоко организованным семействам, но там это результат
опрощения, вызываемого внешними условиями, здесь же — первичное
строение.
У папоротникообразных, названных Джеффреем Pteropsida, листья крупные,
с хорошей дифференцировкой на черешок и пластинку. Их примитивность
сказывается главным
образом в том, что они растут верхушкою, а не основанием, как листья
цветковых. Такой способ роста позволяет им развивать нередко добавочный
рост отдельных
частей листа, курчавость краев и другие уродливости.
У цветковых мы имеем колоссальное разнообразие листовых форм и
строений. Сеть жилок в своей более простой форме дает
очень мало анастомозов. Главные жилки располагаются параллельно
одна другой, и при перерезке любой из них примыкающий к верхней
части
перерезанной жилки
участок ткани засыхает. Более прогрессивный тип жилкования — сетчатый.
Если перерезать одну из главных жилок у листа этого типа, то ткань
кругом остается
жива, так как получит воду обходным путем, через боковую сеть.
Наиболее развитыми являются листья растений из семейства бобовых,
появившегося сравнительно поздно. Лист бобовых состоит из листового
основания, отходящих
от него прилистников, общего черешка и перисто или дланевидно расположенных
пластинок — листочков. Благодаря находящимся на черешочках сочленениям
листовые пластинки бобовых могут перемещаться под влиянием большей
или меньшей напряженности
солнечных лучей и становиться под определенным углом к плоскости
падения последних, регулируя таким образом интенсивность освещения.
Для ассимиляции
особенно благоприятны
лучи определенной напряженности, при более сильном освещении хлорофилл
разрушается быстрее, чем восстанавливается, при более слабом не
хватает энергии.
Такие сложные листья, как листья гороха, акации и массы других
бобовых, с свободным движением отдельных частей, с организованным
отводом
продуктов ассимиляции
из тканей (мякоти) в ситовидные трубки и пр., являются наиболее
совершенным выражением эволюции листа. От листа плауновых до листа
гороха пройден
сложный и долгий путь, приведший к выработке прекрасного пластичного,
сообразно
условиям среды, аппарата фотосинтеза; фотосинтез же, как известно,
— это главный физиологический
процесс зеленых растений.
3. ПРОИСХОЖДЕНИЕ СТЕБЛЯ
Стебель — существенная часть растений, их тело, так как он легко
воспроизводит и корни, и листья. При размножении растений черенками
мы практически
используем это важное свойство стебля регенерировать недостающие
органы. Главные функции
стебля — механическая поддержка кроны и проводящая. По коре
его идет ток питательных веществ из листьев в корни, чем обусловливается
рост
и корневой
системы. По
древесине, точнее по заболони, идет водный ток из корней в
листья. Основная форма стебля — форма колонны — так проста, что в этом
отношении никакой
особой эволюции от крупной морской водоросли лессонии через
стволы плауновых деревьев
каменноугольной эпохи к стволам крупных хвойных и лиственных
деревьев не замечается. Наоборот, анатомия стебля дает очень сложную
картину постепенного усложнения и усовершенствования проводящего
аппарата.
У бурых водорослей в центральной части их стебля мы находим
наличие длинных трубчатых клеток, имеющих анастомозы в сообщающихся
одна
с другой помощью
ситовидных пластинок. Последнее — не что иное, как поперечные
перегородки, отделяющие
одну клетку от другой. Протоплазмы же их проходят через поры
сит, неся с собой питательные вещества. Ствол массивен, ассимиляция
происходит в поверхностных
слоях ткани, внутренние части, затененные наружными, будут
голодать, если их не будет пронизывать система трубочек с движущимися
растворами
внутри.
Даже у грибов в их более массивных стеблевидных образованиях,
каковы, например, шнуровидные ризоморфы опенка, разрастающиеся
под корою
старых пней, внутренние
клетки принимают на себя роль проводящей ткани и функционируют
соответственно, представляя собою пучок длинных нитей, окруженных
мелкоклеточной
плотной корой.
У мхов, в их тонких стеблях, мы всегда находим в центре проводящий
цилиндр из узких тонких трубчатых клеток, образующих точно
пригнанные вертикальные
ряды. Поперечные разрезы шлифов стеблей псилофитов показывают
сразу, что эти первые растения суши были построены сложнее
мхов, хотя
и сходны с ними.
У ринии
в центре стебля находился участок толстостенных клеток, проводивших
воду от корневища к верхушкам побегов. Участок этот был окружен
кольцом многочисленных
тонкостенных трубчатых клеток, проводивших питательные вещества
от зеленых верхних частей растения в корневище. У астероксилона
на продольных
шлифах
видны
водоносные клетки — трахеиды с характерными кольчатыми или
сетчатыми утолщениями на стенках. Совокупность всех таких проводящих
клеток,
имеющая вид внутреннего
шнура, пролегающего между более рыхлой мякотью коры стебля,
получила наименование стеле, или столба.
Эволюция стеле папоротников, приведшая к большому разнообразию
и строения, и расположения, в тех случаях, когда их много,
привела в конце концов
к выработке наиболее рационального построения древесины и коры
у
цветковых растений.
Если у папоротников древесина всегда состоит из водоносных
клеток или трахеид, то у хвойных в их молодых тканях уже есть
небольшие
спиральные
сосуды (в
протоксилеме), у гнетовых их значительно больше и они принимают
некоторое участие также и
в строении вторичной древесины. У однодольных не развит еще
камбий, т. е. специальная зародышевая или образовательная ткань
проводящих
лучков,
что мешает им срастаться
вместе и расти в толщину. Поэтому у них строение стволов менее
плановое, менее совершенное, чем у двудольных растении, дающих
благодаря камбию полное разделение ствола на кору и древесину.
Таким образом, процесс эволюции охватывает все детали строения
стебля, делая его достаточно совершенным орудием как механической
крепости
растения, так
и физиологической связи между двумя активными системами органов
растительной жизни, именно между листовой и корневой системами.
4. ПРОИСХОЖДЕНИЕ ЖЕНСКОГО ГАМЕТОФИТА
ЦВЕТКОВЫХ РАСТЕНИЙ
У простейших размножение состоит в делении клеток; бактерии
и циановые водоросли в короткое время могут образовать
значительные массы
исключительно путем простого
деления. Бактерии клёка за 12 часов нередко превращали
на сахарных заводах содержимое чанов с 500 кг патоки в студенистые комья.
Клетки их обычно
все одинаковы; там, однако, где холод или засуха постепенно
прекращают
рост организмов,
отдельные клетки превращаются в цисты, или покоящиеся споры.
При этом оболочка клеток утолщается, происходит накопление
в протоплазме
запасных
питательных
веществ и клетка несколько разрастается; она переходит
затем
в покоящееся состояние и лишь при наступлении благоприятных
условий снова начинает
ассимилировать, поглощать растворы и делиться.
У жгутиковых организмов, причисляемых к растениям, в некоторых
случаях образуются одинаковые между собой гаметы или изогаметы.
Это клеточки,
снабженные по большей
части двумя ресничками, которые, двигаясь в воде, сталкиваются
между собой и, теряя реснички, сливаются вместе попарно.
Образовавшаяся двухъядерная клетка зигота также превращается
в цисту и переживает
период покоя,
после которого
в ней происходит редукционное деление и образуются молодые
одноядерные клетки следующего поколения.
У водорослей мы имеем уже правильное наличие гамет в цикле
развития. На низшей стадии эволюции стоят водоросли, у
которых гаметы совершено
одинаковы,
как,
например, у обыкновенной на севере зеленой нитчатки улотрикс,
на средней же между гаметами есть уже некоторое различие
в размерах — более крупная
имеет
ресницы и двигается, как это имеет место у простейших водорослей.
Крайним случаем такой анизогамии и будет оогамия, так как
яйцевая клеточка
отличается от изогаметы
крупными размерами, обилием запасных питательных веществ
и полным отсутствием органов движения — ресниц. Дальнейшим
усложнением
является
все растущая
защищенность яйцеклетки водорослей нитчаток, прикрытой
лишь оболочкой материнской клетки. У несколько более совершенных форм
эта оболочка является в виде специального органа оогония.
У многих
бурых водорослей
оогонии
находятся в особых полузамкнутых ямках скафидиях, что значительно
увеличивает их
шансы на спокойное развитие.
Рис. 26. Развитие гаплоидных элементов от изогамет до
зародышевого ядра цветковых растений |
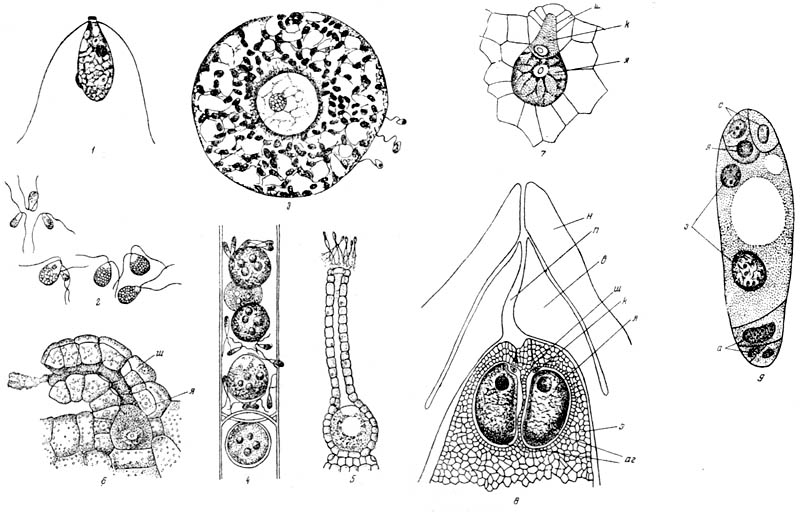
Мы не знаем, как развивались псилофиты, по у современных
мхов мы находим в пазухах листьев, часто собранными помногу
на концах
веточек,
еще
более сложные
органы архегонии. Архегоний состоит из шейки и брюшка,
стенка его многоклетная, полость брюшка заполнена крупной
мощной
яйцеклеткой или иначе макрогаметой;
канал шейки выполнен несколькими легко расплывающимися
клетками. Проникающий сквозь канал сперматозоид вызывает
оплодотворение,
связанное, как и
всегда, с удвоением числа хромосом. Яйцеклетка превращается
в зиготу,
последняя
делится и образует своим дальнейшим ростом палочкообразный
зародыш, из которого позднее
образуется спорогон мха.
У мхов, хвощей, настоящих папоротников и некоторых других
высших споровых все споры одинаковы, и угадать, вырастет
ли из нее
гаметофит, способный
дать и
макро- и микрогаметангии, или же только макрогаметангии
— архегонии, или же только микрогаметангии — антеридии,
невозможно.
Между тем уже у крупных деревьев девонского и каменноугольного
периодов, именно у лепидодендронов, было полное разделение
на макро- и микроспоры.
Из макроспор
вырастали макрогаметофиты, т. е. заростки, дававшие архегонии
и в них яйцеклетки. Английскому палеонтологу Макину (Maclean)
удалось
получить
тонкие шлифы
через макроспоры, на которых хорошо видно, как образовавшийся
уже в макроспоре заросток
своей верхней частью выдается из лопнувшей макроспоры,
образуя тонкую,
вероятно, зеленую волнистую пластинку, на которой сидят
архегонии. Большая часть этого
заростка, вероятно бесцветная, оставалась внутри споры
и состояла из ткани, богатой запасными питательными веществами.
У современных нам плауновых, называемых селагинеллами,
у маленькой группы шильников (Isoetaceae), у некоторых
водяных
папоротников
(Salviniaceae и Marsiliaceae)
деление на макро- и микроспоры выражено очень хорошо и
позволяет во всех
деталях проследить развитие мужских и женских заростков.
Яйцеклетка у них находится
в архегонии, который погружен в ткань заростка. Если бы
заросток не выступал из макроспоры, а развивался бы целиком
внутри
ее, то защита
первых стадий
зародышевой жизни растения была бы еще более обеспечена,
что мы и видим у цветковых растений.
У голосеменных растений редукция (упрощение) гаметофита
идет значительно далее. Архегонии состоят из крупной
яйцеклетки, на верхушке которой сидит несколько маленьких
клеточек, сохранившихся как рудимент шейки архегония.
У голосеменных и цветковых растений, как возникших в процессе
борьбы за овладение засушливыми территориями юрских материков,
гаметофит
и яйцевая клетка глубоко
запрятаны под защитой тканей спорофита. Макроспоры возникают
путем редукционного деления в глубине тканей семяпочки,
в полости завязи,
окруженной покровами
цветка или соцветия. Из четырех макроспор зрелости достигает
только одна, она сильно разрастается и превращается в так
называемый зародышевый
мешок,
крупную
клетку с двумя гаплоидными ядрами в центре. Прорастание
этой макроспоры и развитие в ней заростка состоят в том,
что у
верхнего ее конца
развиваются три крупных
ядра — зародышевое и два его спутника (две синэргиды),
а у нижнего три клетки, называемые антиподами. Последние
можно
рассматривать
как вегетативную
ткань
заростка. Считая и центральные ядра, всего в зародышевом
мешке
развивается 8 ядер. В отдельных случаях число это значительно
больше, так как
антипод может быть много.
Заросток с одним архегонием или, как думают другие ученые,
с тремя архегониями и тремя яйцеклетками готов. Он совершенно
изолирован
от влияния подсыхания,
тесно связан физиологически с материнским растением, и
развитие из яйцеклетки зародыша поставлено в особо благоприятные
условия.
5. ПРОИСХОЖДЕНИЕ МУЖСКОГО ГАМЕТОФИТА
ЦВЕТКОВЫХ РАСТЕНИЙ
Уже у многих водорослей имеются микрогаметангии, в клетках
которых развиваются сперматозоиды. Таковы из зеленых
свободно движущийся
волчок (Volvox),
из нитчаток эдогоний и сфероплея. из сифонных вошерия,
из наиболее сложных все харовые
водоросли. И здесь можно проследить эволюцию в постройке
самого гаметангия от отдельной клепки до сложно построенного
антеридия
с массивной
внешней стенкой.
У мхов антеридий состоит из массы клеточек, у папоротников
число их сильно уменьшается. У тех папоротников, которые
имеют макро-
и микроспоры,
последние
при прорастании дают уже (семейство сальвиний) всего
одну вегетативную клетку и два сильно опрощенных антеридия.
Поэтому неудивительно,
что у голосеменных
микроспоры (называемые пылинками цветени) дают при
прорастании от одной до нескольких вегетативных клеточек,
заменяющих
собою заросток
и упрощенный
антеридий
из двух клеточек. Затем содержимое микроспоры растет
и
вытягивается в трубочку, которая носит название пыльцевой.
Суть этого
процесса в том.
что он превращает
самостоятельно живущий
гаметофит, имеющийся у водорослей и мхов, в орган оплодотворения,
совершенно подчиненный функциям спорофита. Растения,
приобретшие в борьбе за существование
пыльцевую трубку, часто называются сифоногамами. У
водорослей и мхов сперматозоиды попадают по выходе
из антеридиев
в воду и, свободно
двигаясь, направляются
к отверстиям оогониев и архегониев, или к вышедшим
из оогониев яйцеклеткам.
У сосен, елей и других голосеменных ветер разносит
пылинки цветени, причем некоторые из них попадают в
пыльцевые
камеры на семяпочках;
здесь они
попадают в капли нектара, где развивается пыльцевая
трубка. В последней развиваются
из ядра антеридия микрогаметы (сперматозоиды), которые
вызывают растворение оболочек на участке, где стенка
пыльцевой трубочки
соприкасается
со стенкой зародышевого мешка, и проникают к яйцевым
клеточкам архегониев. Все это
далеко нецелесообразно, так как если водоросли и мхи
теряют большую часть сперматозоидов
на путях к яйцеклеткам, то голосеменные, у которых
сперматозоиды имеют
определенное направление и защиту вплоть до момента
проникания внутрь макроспоры и не теряются
зря, теряют зато большую часть микроспор, которые пропадают,
не достигнув пыльцевых камер и не дав развиться сперматозоидам.
Цветковые
растения
в этом отношении
построены целесообразнее. Их пыльца улавливается специально
развитыми рыльцами цветков, их пыльцевая трубочка растет
гораздо сильнее,
число не функционирующих
клеток еще не уменьшено. При переносе пыльцы насекомыми
часто прорастает на рыльцах большая часть пыльцы.
Мужской гаметофит может быть развит у некоторых водорослей
(например, у Ectocarpus из бурых) в целое вполне развитое
растение. У двудомных
мхов
он также представляет
собою все растение мха, отличающееся на глаз от женского
только тем, что на нем не развиваются спорогоны. У
папоротникообразных, при двудомных
заростках,
мужской заросток мельче женского, но все еще у многих
из них
ведет самостоятельное существование. У голосеменных
и покрытосеменных или цветковых растений
мужской
заросток — уже остаточный орган, не имеющий никакой
функции, в
некоторых случаях исчезающий вовсе. На его место становится
пыльцевая трубка,
своеобразное мужское
растеньице, также ведущее самостоятельное существование
от момента прорастания микроспоры-пылинки до момента
оплодотворения.
При водном или земноводном существовании уместен был
гаметофит, дававший сперматозоиды. При чисто сухопутной
жизни на ветру
и на солнце, часто
высоко над землей, у
деревьев место его заняла несравненно более скромная,
но равнозначная ему по функции пыльцевая трубочка.
6. ПРОИСХОЖДЕНИЕ ЦВЕТКА
Гаметофиты водорослей, грибов и мхов дают в некоторых
случаях целые группы гаметангиев: оогониев, антеридиев
или архегониев,
окруженных
иногда защитными
клетками или веточками. Несмотря на это, к водорослям
и грибам никто еще термина «цветок» не применял.
У мхов антеридии располагаются часто группами на
концах стеблей в пазухах защищающих их листочков,
причем последние
принимают
красную или желтую
окраску (например,
у кукушкина льна Polytrichum). Такие скопления
антеридиев некоторые авторы пробовали называть
цветком (большинство
же считает термин
«цветок» в
применении к гаметофиту совершенно неприемлемым,
приурочивая его исключительно к спорофиту)
.
Никаких половых органов спорофит не дает, он дает
только споры. Половые органы дает гаметофит. У
мхов споры
развиваются внутри
коробочки,
которую цветком
никто никогда не называл. У псилофитов, древнейшего
растительного населения суши, спорангий занимал
такое же положение
на конце стебля, как и коробочка
на верхушке спорогона мхов. У всех папоротникообразных
спорангии расположены уже на листьях, причем там,
где они рассеяны
по всей поверхности обыкновенных
зеленых листьев или питающих листьев трофофиллов,
цветка нет. Но вот у нашего черного папоротника
страусопера (Struthiopteris flicastrum All.) к концу лета
развивается в центре его кроны пучок листьев, специализированных
как
носители
спорангиев — спорофиллы. Спорофиллы эти окружены
короною
крупных
зеленых листьев трофофилов, а все вместе можно,
пожалуй, уподобить цветку.
| * A. Engler. Die Naturliche Pflanzenfamilien, Band 14a. Angiospermae. 1926, 140. |
Какое в самом деле различие между строением черного
папоротника и строением пыльникового цветка конопли
или хмеля, состоящего
из пяти
микроспорофиллов
— тычинок и пяти зеленых листочков околоцветника?
И вот А. Энглер*, сводя различия между главнейшими
классами
высших растений, прямо
говорит о
папоротниках, что
их спорофиллы не образуют цветков, за исключением
Struthiopteris и Blechnum. Следовательно,
последние их образуют. На
стр. 1 той же книги
А. Энглер
прямо говорит, что хвощи, плауны, селагинеллы и
все голосеменные
образуют цветы.
Иначе А. Энглер называет цветком всякое организованное
скопление
спорофиллов, т. е. листьев, приносящих споры. Спороносные
колоски хвощей и плаунов,
очень молодые шишки и пыльниковые шишечки хвойных
— все это подходит
под это новое понятие цветка. Если собрание микроспорофиллов
— тычинок вяза или ясеня есть цветок, то и собрание
спорофиллов хвоща
тоже
цветок.
Такие примитивные цветы возникли, по-видимому,
очень рано. У плауновых и кордаитов, приспособившихся
в
девонскую эпоху
к тому,
чтобы раскидывать
свою крону в сравнительно
сухих слоях воздуха, спорофиллы уже собраны помногу
в специальные органы спороношения. Ветер и солнце,
вызывавшие
превращение
зачатков обыкновенных
листьев в спорофиллы,
обусловливали данное явление не у отдельных разбросанных
по кроне листьев, а одновременно на целых веточках,
которые и
превращались
в спороносные
органы — колоски, шишки или цветы.
Цветок, как специфический орган цветковых или покрытосеменных
растений (Angiospermae), отличается от спороносных
колосков папоротникообразных растений тем, что
он не просто орган, дающий споры, а орган перекрестного
опыления. Дело в том, что естественный отбор всего
ярче проявляет
свое действие
в тех
случаях,
когда налицо имеется потомство, полученное путем
скрещивания близких форм. Получаются
особи с различными комбинациями наследственных
признаков,
особи неодинаковые
по существу, и эти особи либо сохраняются действием
борьбы за существование и естественного отбора,
либо истребляются.
Этим
путем из старых
видов образуются новые, более приспособленные к
среде, окружающей растения
в данный исторический
момент. Орудием этого-то чрезвычайно важного процесса
и является цветок.
Большинство ботаников думает, что толчком к развитию
сложного цветка было появление в юрскую эпоху большого
числа сосущих
мед насекомых.
Посещая семяпочки хвойных,
в пыльцевых каморах которых выделяется нектар,
и принося при этом микроспоры — пылинки цветени,
насекомые
дали
толчок к
тому, что
опылявшиеся ранее
исключительно ветром плодущие колоски предков цветковых
растений преобразовались постепенно,
путем стерилизации наружных или нижних спорофиллов
и превращения их в доли
околоцветника, а также совмещения макро- и микроспорофиллов,
тычинок и пестиков в одну систему, в настоящие
цветы. У последних появились
также специальные
нектарники, выделяющие нектар, что еще усилило
привычную связь их с насекомыми.
Наиболее совершенным является цветок с ярким околоцветником,
выделяющим ароматические вещества и нектар, с тычинками
и пестиками, скрытыми
в трубке венчика и тем
защищенными от непогоды; цветок, заметный издали
и систематически . привлекающий насекомых к скрытым
в
нем пищевым веществам
— углеводам и протеинам.
Такой цветок наилучшим образом обеспечивает скрещивание,
образование здоровых семян и действие естественного
отбора, поддерживающее
растение на уровне
требований, предъявляемых к нему внешней средой.
И вот такой-то цветок совершенно отсутствовал в
юрскую эпоху и появился сразу у многих растений
в начале
меловой.
Впоследствии цветковым растениям пришлось во многих
странах, где мало насекомых, например в степях,
снова приспособляться
к опылению
ветром
и упрощать строение
цветка. Так, в семействе злаков цветок заменился
колоском.
Громадное разнообразие форм, строений, окраски,
запахов современных нам цветков объясняется тем,
что процесс
эволюции растительного
мира все
время приводит
орган перекрестного опыления, цветок, к приспособлению,
в соответствии с условиями опыления и внешней средой.
Там, где процесс скрещивания почему-либо выпадает,
редуцируется и цветок. Так у видов рода манжетки
(Alchimilla) зародыш
образуется в семенах
бесполым путем
или апогамно. Скрещивание теряет всякое значение,
и цветок манжетки — маленький, зеленый, с недоразвитыми
органами
— стал неузнаваем.
Его родичи
из семейства
розоцветных блещут прекрасными цветами, а манжетка
стала растением, на цветы которого никто и внимания
не обратит.
Так объясняются и возникновение цветка и бесчисленные
его видоизменения.
7. ПРОИСХОЖДЕНИЕ СЕМЯН
Впервые орган, похожий на семя, мог появиться у
разноспоровых папоротникообразных растений.
Семя состоит из интегументов,
или покровов, под защитой
которых развивается макроспора; в макроспоре
мы должны найти заросток и архегонии.
После оплодотворения
яйцеклетка архегония разрастается в зародыш;
зрелое семя есть хранилище молодого растения с корешком,
почечкой и специальными зародышевыми
листьями — семядолями.
У многих растений, кроме того, развивается
еще специальная питающая зародыш при прорастании
ткань, эндосперм,
иногда
называемый
белком,
так как он
по своей функции напоминает белок куриного
яйца.
Семена голосеменных растений принципиально
отличаются от семян покрытосеменных тем, что
у более древних
голосеменных эндосперм
есть не что иное,
как заросток, т. е. гаметофит. он формируется
очень рано и к моменту оплодотворения
уже
совершенно готов. У более молодых покрытосеменных
растений эндосперм до момента оплодотворения
отсутствует вовсе, здесь он заменен антиподами.
Мы уже упоминали выше, что
число сперматозоидов в пыльцевой трубке у покрытосеменных
два. Один из них оподотворяет яйцеклетку и
тем вызывает образование зародыша,
другой
же сливается
с центральным ядром зародышевого мешка и через
последующие повторные деления образованной
таким образом второй
зиготы дает эндосперм.
Таким образом
эндосперм голосеменных и эндосперм покрытосеменных
не эквивалентны между собой. Один
соответствует заростку, а другой близнецу зародыша.
| * F. W. Oliver and D. H. Scott. On the structure of the palaeozoic
seeds Lagenostoma Lomaxi. Phil. Trans. Roy. Soc., vol. 197, B. 193, 1904. ** E. Dix. Seeds associated with Linopteris Munsteri Eichn. Annals of Botany, vol. 42, 1019-1023, 1928. |
Листья папоротникообразных растений силурийской,
девонской и каменноугольной эпох сначала представлялись
все как
листья настоящих
папоротников.
В 1904 г. Оливер и Скотт в Англии* выяснили
принадлежность находимых в
каменноугольных
пластах семян к листьям древовидного папоротника
Lyginodendron oldamium Will.,
который таким образом оказался не папоротником,
а голосеменным растением и стал первым представителем
особого класса
семенных папоротников
или Pteridospermae. В последнее время доказано**,
что большинство папоротниковых
листьев, находимых
в каменноугольных отложениях, принадлежит птеридоспермам,
но все же семена редко сидят на листьях, чаще
их находят отдельно.
Известны,
однако, и
прямые находки листьев, по краям которых сидят
семена.
Таковы находки некоторых
Pecopteris,
Linopteris и пр. Даже оригинальный глоссоптерис,
этот наиболее типичный представитель гондванской
флоры,
оказался семенным
растением.
Семена папоротников птеридосперм несколько
доходили на семена современных саговников с
крупной пыльцевой
камерой,
часто
они сидели в особой
«чаше», т. е. были окружены
небольшими листочками. Странную на наш взгляд
картину представляли собой эти многочисленные
в свое время
папоротники, часто
с кружевной листвой,
по краям
которой свешивались массивные семена.
В ряду плауновых известен род лепидокарпон
(Lepidocarpon), у которого односеменные макроспорангии
были окружены
специальным покровом,
или интегументом, и
сверх того еще несколькими спорофиллами. Отсюда
легко себе представить
и возникновение
семян у кордаитов и гинкговых.
Таким образом, возникновение семян старше возникновения
цветка. Древнейшие семена одиночны, соответствуют
отдельным макроспорам,
которые были
окружены защитными выростами и листочками.
Значение их то, что заростки, развившиеся
внутри макроспор, получили от материнского
растения больше защиты и питания, чем у тех
папоротникообразных,
макроспоры
которых падали с дерева в болото или на сырую
почву и там уже развивали заростки, архегонии
и зародыши,
как
это было,
например,
у лепидодендронов.
Семя, как совершенный орган зародышевой жизни,
как возможность развивать и сохранять зародыши
от разрушительных
климатических
влияний, дало
растениям возможность проникнуть в глубь обширных
материковых пространств, оторваться
от морских побережий и, используя кратковременные
влажные периоды, в связи с временами года,
— развить обширные
заросли. А там,
где растений много,
они
сами увлажняют воздух, испаряя воду за счет
грунтовых вод, и дают возможность ютиться в
их тени также
и представителям древних
типов,
самостоятельно
живших лишь у воды.
Заканчивая наш обзор, мы видим, что многое
еще не выяснено, не установлены точные факты
прошлого,
нет
возможности
проследить всю цепь изменений,
предшествовавших явлениям современности. Но
это еще придет.
ЗАКЛЮЧЕНИЕ
Перед нами прошла в сжатом, очень кратком изложении
история земной растительности. Мы видели,
что сначала появились
и стали развиваться
простейшие обитатели
водоемов, включая и море. Затем в течение
долгого периода жизнь особенно пышно развивалась
в море, тогда как суша оставалась сравнительно
пустынной. Это был первый период мироздания,
поскольку дело
касается растений,
это
был период
бактерий и водорослей.
Затем жизнь стала возможной и на суше.
На каменной поверхности материков появились
не только пленки
водорослей, но и
подушки растений, близких
к современным мхам.
Они не оставили после себя следов, и мы
еще
не имеем растительных остатков этого периода.
Кембрийские
слои, поскольку дело
касается пресноводных
отложений, по-прежнему немы для нас. Мы
только догадываемся
об этом втором периоде,
когда, сохранив бактерий, жгутиковые организмы
и водоросли, Земля обогатилась еще
и мхами. Но все же мы предполагаем, что
наступил и этот второй период мохообразных
растений.
Верхние, т. е. самые молодые из силурийских
отложений, дали нам доказательство, что
развитие жизни на
суше привело к
образованию сплошного растительного
ковра, низкого, но плотного, вырабатывавшего
такую массу органических веществ, что
это повело не только к развитию более пышной
животной жизни, но и к образованию почв.
Эти растения, более совершенные, чем мхи,
по еще менее сложные, чем плауны, дали
третий период, или период
мохообразных псилофитов.
Девонские осадочные слои дают к концу периода
остатки древесной флоры папоротникообразных
и хвойных —
кордаитов.
Растительность эта развилась много пышнее
в следующий за девонским каменноугольный
период.
Как мы видели
ранее, предполагают, что массовое развитие
этой каменноугольной флоры, создав крупные
запасы
пищевых веществ,
мощные слои почвы и нормальный
процентный состав газов воздуха, было поворотной
точкой в истории
всего живого населения Земли. После этого
развитие как растительного, так
и животного мира
пошло несравненно быстрее, и разнообразие
земного населения чрезвычайно увеличилось.
Так наступил
четвертый период,
или период папортникообразных.
Затем изменение климата, возникновение
больших материковых массивов и крупных
горных хребтов
настолько изменили
условия жизни растений
суши,
что победу
в борьбе за существование могли одержать
только растения с жесткими вечнозелеными
листьями,
хорошо выносящими
и палящее солнце, и
суховеи. Такими растениями
являются голосеменные растения. И действительно,
в осадочных пластах триаса и юры мы находим
ясные доказательства
тому, что наступил
пятый период голосеменных
растений.
К этому периоду относится пышное развитие
таких растений, как саговники, араукарии,
ошкговые
деревья, тиссовые,
родственники наших елей
и сосен, и, наконец, родственники
кипарисов и можжевельников.
В последующую эпоху, когда в морях отлагались
толщи пишущего мела, на суше, наряду с
развитием мира
птиц, млекопитающих
и насекомых,
появились и начали
множиться и в количестве п в разнообразии
форм цветковые растения, деревья,
кустарники и травы. Благодаря своей листве,
быстрому росту и способности быстро распространяться,
они понемногу захватили
большую часть
суши, оставив голосеменным
и папоротникообразным сравнительно ничтожное
место. Только в
морях продолжали господствовать водоросли.
Таким образом, уже с меловой эпохи начинается
господство цветковых растений, образующих
шестой период, или
период цветковых растений.
Период этот достиг своего расцвета в третичное
время. Затем наступление более холодного
ледникового периода
уничтожило
на севере большую
часть живших там
крупных растений, а когда за ледниковым
временем последовало более теплое послеледниковое
время, север снова заселился
растениями, переселившимися с гор, расположенных
южнее.
Так появились на земле предки растений,
так постепенно выработалась п создалась
вся цепь
растительных
организмов, благодаря тем
последовательным изменениям
на поверхности Земли, которые влияли на
организм растений, вызывая в
нем необходимость таких усложнений и усовершенствований,
без
которых растения погибли бы вовсе или ограничились
бы одними лишь водорослями.
Наконец, в близком будущем мы предвидим
решительное господство культурных растений,
созданных
уже не слепыми силами
природы, а работою человека.
Наступает седьмой
период, период господства культурных растений.
Волнующиеся поля пшеницы и других хлебных
растений, стройные
ряды хлопчатника, подсолнечника и пр.,
пышная зелень фруктовых садов, овощные
культуры,
наконец, культуры
декоративных растений (парковые деревья
и кустарники, цветочные культуры)
— все
это сменит
непроходимые чащи первобытных лесов, обманчивую
зелень болот и прочие исторически создавшиеся
формы растительного покрова Земли. Заповедники
сохранят резерв дикой растительности, из
которого еще долгое
время культиваторы
будут черпать
свежий материал
для пополнения введенных в культуру растений,
и только крутые гребни гор сохранят
местами древние типы растительности, в
поучение нашим потомкам. Несомненно, что
организация
земледелия на
научных основах
и связанная
с этим
полная победа человека над природой обещают
человечеству возможность развить
большое и прочное
благосостояние.
ОГЛАВЛЕНИЕ
Предисловие............................................................................................................................................................
3
Глава I. Место зеленых растений в
космосе и значение их для человека ........................................................
7
Глава II. Земля до возникновения на
ней растений...........................................................................................
12
Глава III. Как зародилась жизнь на
Земле...........................................................................................................
21
Глава IV. Круговорот жизни и круговорот
вещества в природе......................................................................
34
1. Углерод .............................................................................................................................................................
35
2. Кислород............................................................................................................................................................
37
3. Водород..............................................................................................................................................................
39
4. Азот.....................................................................................................................................................................
40
5. Зольные составные части.................................................................................................................................
42
Глава V. Понятие об эволюции. Палеонтологическая
летопись .....................................................................
46
Глава VI. Архейская группа слоев.......................................................................................................................
57
Глава VII. Палеогеография и условия
жизни прошлых геологических периодов...........................................
60
Глава VIII. Палеозойская группа
слоев...............................................................................................................
64
1. Кембрийская система слоев.............................................................................................................................
64
2. Силурийская система слоев.............................................................................................................................
66
3. Девонская система слоев..................................................................................................................................
69
4. Каменноугольная система слоев......................................................................................................................
72
5. Пермская система слоев....................................................................................................................................
84
Глава IX. Мезозойская группа систем
осадочных образований........................................................................
88
1. Триасовая система слоев...................................................................................................................................
88
2. Юрская система слоев........................................................................................................................................
90
3. Меловая система слоев......................................................................................................................................
96
Глава X. Кайнозойская группа слоев.................................................................................................................
102
1. Третичная система слоев.................................................................................................................................
103
2. Четвертичная система слоев и ледниковый период ....................................................................................
118
Глава XI. Как произошли отдельные
группы растений ...................................................................................
129
1. Происхождение бактерий................................................................................................................................
129
2. Происхождение водорослей............................................................................................................................
134
3. Происхождение грибов....................................................................................................................................
138
4. Выход растения на сушу и происхождение наземной растительности ......................................................
142
5. Происхождение мхов........................................................................................................................................
146
6. Происхождение папоротникообразных растений ........................................................................................
150
7. Происхождение шишконосных и других голосеменных растений..............................................................
159
8. Происхождение цветковых растений.............................................................................................................
165
9. Происхождение культурных растений ...........................................................................................................
167
Глава XII. Как произошли главнейшие
органы высших растений...........................................................
169
1. Как произошел корень....................................................................................................................................
169
2. Происхождение листа.....................................................................................................................................
172
3. Происхождение стебля...................................................................................................................................
173
4. Происхождение женского гаметофита цветковых растений ................................................................
175
5. Происхождение мужского гаметофита цветковых растений ................................................................
179
6. Происхождение цветка...................................................................................................................................
181
7. Происхождение семян.....................................................................................................................................
183
Заключение...........................................................................................................................................................
186