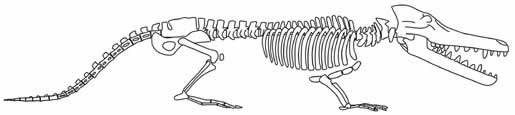
| Главная |
Первый кит, бегущий краем моря.
Ненадёжные киты креационизма.
Индохиус - «меньший брат» китов
Пакицетиды: первые в ряду
Небольшая интермедия: кит с хоботом
Родословная «ходячих китов»
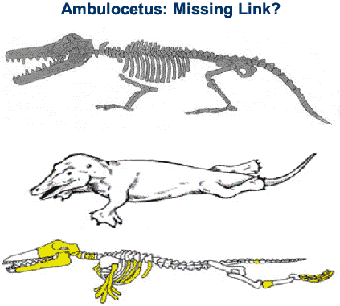
Амбулоцетус – «кинозвезда» из «Прогулок с чудовищами»
Скованные одной цепью: последовательность форм ранних китообразных
Ещё одна интермедия: есть и подревнее киты
Путь к современным китам
Небольшая интермедия: о развитии китового уса
Ушки... не на макушке.
... На задних лапках ходят.
Держи нос по ветру!
Интермедия о правде и полуправде: О том, как НЕ надо говорить о переходных
формах.
Дань прошлому в организме китообразных.
В эпоху становления эволюционной теории Дарвина происхождение китообразных
оставалось загадкой для учёных. Эти животные сильно отличаются от остальных
современных млекопитающих особенностями анатомии и физиологии, и среди ныне
живущих млекопитающих нет видов китообразных (или иных родственных форм), которые
бы сохраняли в большей или меньшей степени явные особенности строения предковых
форм китообразных.
Естественно, это не могло пройти незамеченным для креационистов. Существование
китообразных как чётко оформленной группы животных (1) и отсутствие у них близких
родственников в современной фауне (2) в интерпретации сторонников этого учения
являются несомненными доказательствами сотворения этих животных.
В отечественных источниках по «креационной биологии» проблема происхождения
китообразных освещена крайне скудно. С одной стороны, это не может не радовать
– сторонникам теории эволюции придётся выслушивать меньше нападок со стороны
креационистов. Но с другой стороны, аудитория окажется менее подготовленной
к возможному массированному «вторжению» западных переводных статей. На Западе
всё обстоит иначе – креационисты активно полемизируют с эволюционистами, в
том
числе по
поводу
происхождения
китов. Поэтому «упреждающий удар» в этом вопросе просто необходим. С этой целью
я составил обзор работ, посвящённых эволюции китообразных и изменению отдельных
органов в процессе исторического развития этих животных. Я постарался выделить
основные моменты, по которым происходят разногласия у сторонников и противников
теории эволюции в отношении анатимии, физиологии и эволюции китообразных. Многие
статьи пришлось переводить самостоятельно, поэтому я буду очень благодарен,
если мне укажут на ошибки перевода, и, соответственно, дадут верное значение
переводя с пояснением.
В ходе рассказа я буду цитировать мнения креационистов, выделяя их жёлтой рамкой:
Цитаты из работ эволюционистов выделены голубой рамкой:
Цитаты из источников, не связанных непосредственно с эволюцией китообразных, будут указаны в бесцветной рамке:
Что ж, начнём, пожалуй... Перефразируя поэта, мы говорим «Дарвин», подразумеваем «эволюция», мы говорим «эволюция», подразумеваем «Дарвин». Во всяком случае, креационисты обычно начинают с Дарвина. В русском Интернете желающие могут отыскать статью Малкома Боудена «Киты: никаких признаков эволюции» (в формате PDF). На первой же странице этой небольшой и, на мой взгляд, довольно слабенькой работы есть такие строки:
| Дарвин в первом издании книги «Происхождение видов» (1859) высказывает предположение, что киты эволюционировали от наземных животных. Вероятно, даже ему было сложно предположить возможность эволюции рыбы в млекопитающее. |
Что называется, «приехали», и причём сразу мордой в столб.
Это как же, вашу мать, извиняюсь, понимать? Я просто напомню, что ещё в 1693
году Джон Рэй доказал, что кит – это не рыба, а млекопитающее. Поэтому со
стороны Дарвина логичнее было предположить, что киты эволюционировали не сразу
от водных
животных, а от наземных млекопитающих, хотя бы в свете работы Джона Рэя. А
вот что бывает на уме у антидарвинистов – могу только догадываться. И случайные
«прорывы»
типа
приведённого
пассажа
наводят на совсем скверные мысли.
В русском Интернете на разных креационистских
сайтах также можно отыскать перевод книги Джонатана Сарфати «Несостоятельность
теории эволюции», расплодившийся на разных сайтах до безобразия. В книге
есть глава, посвящённая
эволюции
китообразных – ещё
одна из немногих переведённых на русский язык статей по данной тематике.
Сарфати критикует книгу «Учение об эволюции и природа науки», а также статью
в журнале “Discover” (С. Zimmer, "Back to the Sea," Discover, January
1995, p. 83), в которых изложены факты и теории в пользу эволюции
китов от наземных предков.
Эволюционисты – специалисты по китам, в частности,
покойный Э. Дж. Слайпер (Е. J. Slijper), осознавали отсутствие переходных
форм в летописи окаменелостей: "В
нашем распоряжении нет ни единой ископаемой промежуточной формы между вышеупомянутыми
наземными животными [т. е. хищными и копытными] и китами". (089) Древнейшие ископаемые останки китов в летописи окаменелостей показывают, что киты изначально были полностью водными формами. … 089 - Е. J. Slijper, Dolphins and Whales (Ann Arbor, MI: University of Michigan Press, 1962), p. 17. |
Обратим внимание, что Сарфати цитирует и использует в рассуждениях
текст книги
1962 года издания. Иногда креационисты обижаются на такие
замечания – мол-де, выставляют их на
посмешище как неких ретроградов. А зачем специально выставлять, когда вот оно
– всё перед глазами? Не сомневаюсь, что в наше время за рубежом вышло много
новых
монографий
как о современных, так и об ископаемых китообразных. Но Сарфати в данном случае
виртуозно разделался с суждением, которое до него уже опровергли новые находки.
Это называется «после драки кулаками махать».
Британский зоолог, специалист по наземным позвоночным Даррен Найш рассказывает
об исследованиях ранних китообразных в своей статье с длинным и цветистым названием
“When whales walked the land…
and looked
like antelopes…
and
mimicked crocodiles…. and evolved trunks. What?” («Когда киты ходили по земле...
и выглядели, как антилопы... и подражали крокодилам... и отращивали хоботы.
Что?»), размещённой
в его старом блоге:
… Для меня ранняя эволюция китов стала особенно
интересной по двум причинам. Во-первых, она является предметом полемики,
особенно в течение последних лет. Ранее культивируемая идея относительно
родства китообразных – а именно, представление о том, что киты происходят
от мезонихид – была оттеснена на второй план первоначально непопулярным
и сумасшедшим представлением о том, что киты могли бы быть близкими родственниками
гиппопотамов. ... Далёкие от того, чтобы
быть тюленеобразными прото-китами, пакицетиды – наиболее примитивные
члены радиации китообразных
– были больше похожи на небольших плотоядных антилоп, хотя, возможно,
они проводили по крайней мере некоторое время на мелководье. ... Открытие Ambulocetus и других новых предковых китов показало, что
они не могут все быть отнесены к всеобъемлющим Protocetidae.
Скорее, они
должны
бы классифицироваться как несколько отличающихся друг от друга
групп. Pakicetus и его близкие родственники были помещены в свою
собственную
группу, Pakicetinae,
Gingerich и Russell (1981), и позже она была поднята до уровня
«семейства». Помимо Pakicetus, туда были включены Ichthyolestes
pinfoldi и Nalacetus
ratimitus. Thewissen at al. (1996) создали Ambulocetidae для
Ambulocetus, и кажется, что Gandakasia и Himalayacetus – также члены
этой группы. |
Эта статья появилась в старом блоге Даррена Найша 16 февраля 2007 года, и потому в ней не упомянут Indohyus, одна из интереснейших находок, связанных с ранней эволюцией китообразных. О ней пойдёт речь далее.
Особенно богатыми на находки самых ранних китообразных
были 90-е годы ХХ века. Именно тогда на территории Индии и Пакистана были
открыты многочисленные представители
млекопитающих, демонстрирующих сочетание черт, характерных для наземных
зверей с одной стороны, и несомненных китов, с другой стороны. Позволю
себе привести
список этих форм, приведённый в книге «Происхождение китов» (“The Emergence
of Whales: Evolutionary Patterns in the Origins of the Cetacea” (Advances
in Vertebrate Paleontology) под ред. J. G. M. Thewissen. Plenum Press,
ISBN 0306458535.). Текст обзора
этой книги (сделанного Джеймсом Аккером) на английском языке легко
можно найти в Интернете хотя бы по названию.
Итак, «парад» ранних китообразных (в скобках указан год описания):
- Pakicetidae - - Ambulocetidae - - Remingtoncetidae - |
В этом кратком обзоре не указаны некоторые другие виды, открытые, очевидно, позже написания данной главы книги:
В семействе Pakicetidae:
Himalayacetus subathuensis - 1998
В семействе Remingtonocetidae (названо в честь американского исследователя китообразных Ремингтона Келлогга (Remington Kellogg):
Remingtonocetus domandaensis - 2001
Kutchicetus minimus - 2000
Странные длиннорылые кутхицетусы напоминают помесь выдры с крокодилом – коротколапые млекопитающие с очень длинным заостренным черепом и длинным хвостом. Реконструкция Карла Буэлла.

Скелет кутхицетуса
По семейству Protocetidae есть более новая и подробная сводка в описании одного из новооткрытых китов, откуда я взял лишь информацию по названию и дате открытия видов, входящих в его состав:
Protocetinae Makaracetinae Georgiacetinae |
А продолжают «парад» находки более прогрессивных китов – ныне вымерших базилозаврид. Сведения приводятся по тому же обзору Джеймса Акера; указаны семейство, вид, год открытия, время существования (подробнее о геохронологической шкале будет сказано ниже).
Базилозавриды Дорудонтины: |
Эволюционисты встретили все эти находки с энтузиазмом. Сложно
сказать, какова была первоначальная реакция креационистов, поскольку всё новые
и
новые находки
означали крушение ещё одного мифа, усиленно внедряемого ими в сознание
обывателей. Но, судя по появлению в соответствующих источниках многочисленных
(хотя и
однотипных) статей, креационисты в очередной раз «вышли на тропу войны». Каждый
новый вид ископаемых китообразных, проясняющий родословную этой группы млекопитающих,
сразу же становится объектом в лучшем случае ожесточённых споров, а в худшем
– объектом приложения самых странных умопостроений.
Рассказ о родословной китообразных был бы совершенно неполным без упоминания
об одной из самых новых находок Ханса Тевиссена – загадочном существе под
названием индохиус (Indohyus).
Недавнее открытие Хансом Тевиссеном и его командой исследователей небольшого животного индохиуса (Indohyus) является прекрасным примером предсказательной силы теории эволюции. Если бы это животное было открыто, допустим, около ста лет назад, его вряд ли оценили бы должным образом. Возможно, его сочли бы неким заурядным представителем ранних копытных, составили бы описание, и лишь сто лет спустя, при более подробном изучении открыли бы истинное значение этого открытия. Но индохиус – это находка, которую ждали. Его появлению предшествовал ряд исследований, результаты которых буквально нарисовали портрет этого существа ещё до того, как оно стало известно науке.

Реконструкция индохиуса, сделанная Карлом Буэллом
|
Долгое время предковые формы китообразных не были известны, и родство этих зверей с другими отрядами млекопитающих было предметом дискуссий. Современные китообразные слишком отличаются по строению скелета от прочих млекопитающих, и потому сравнительно-анатомический способ установления родственных связей малопригоден. В предки китам записывали примитивных хищников креодонтов, и даже насекомоядных, а отделение китообразных от прочих млекопитающих умозрительно относили к концу мелового периода (около 70 миллионов лет назад). Гипотеза о родстве китообразных и копытных также существовала: наряду с указанными выше, она упоминается в книге «Фауна мира. Млекопитающие». Но при отсутствии фактического материала все эти гипотезы оказывались предметом критики со стороны креационистов, которые считали отсутствие находок переходных форм от наземных животных к древнейшим китам лучшим показателем сотворённости этих животных. В связи с этим можно припомнить, например, ехидные насмешки креационистов над предположением Дарвина о том, что киты могли произойти от медведей, которые добывали корм в воде, плавая с открытым ртом. Кстати, первоосновой этого предположения послужило наблюдение исследователя Сэмюэля Хирна (Samuel Hearne) в центральной части нынешней провинции Манитоба в Канаде в 1775 году. Он наблюдал американского чёрного медведя, который плавал с открытым ртом в реке, собирая с поверхности воды насекомых. Знал бы он, к чему это приведёт... Шутка. А вот нечто помимо шуток, найденное в блоге Брайана Свитека Laelaps от 13 февраля 2008 года:
Гипотеза Дарвина не полностью необдуманна (см. очерк Стивена Джей Гоулда «Левиафан на крючке его прошлого», 1997), но его широко критиковали за такое предположение в то время, когда наука становилась более систематической, и «теоретики в креслах» презирались. Тем не менее, любопытно, что Ричард Оуэн сделал некоторые интересные комментарии об [этом] параграфе. Как написано Дарвином в письме Чарльзу Лайеллю, у Дарвина была довольно странная встреча с «резким и едким» Оуэном;
Что полагал Дарвин, так это то, что естественный отбор мог воздействовать на медведя, изменяя его признаки при наличии достаточного времени и давления отбора; но его всё ещё высмеивали за этот эпизод, поскольку некоторые истолковывали пример как означающий, что медведи действительно были предками китов. Дарвин продолжал придерживаться этого мнения частным образом, даже когда убрал этот эпизод, как показывает письмо 1860 года к У. Харвею:
|
Более поздние исследования в бласти биохимии позволили определять
родство групп живых организмов по степени сходства и различия в строении белков
и ДНК. Они
позволили, например, установить, что человек и шимпанзе генетически сходны
больше, чем на 99% (и это лишний раз показывает, как легко потерять человеческий
облик. Шутка.). В отношении китов результат подобных исследований был
несколько необычным – медведи окончательно и бесповоротно ушли из кандидатов
в предки/родственники китов, а сами киты оказались более близкими к парнокопытным.
Ископаемые остатки ранних китообразных при сравнении с остатками других
древних млекопитающих вначале показали некоторую степень сходства с мезонихидами
– плотоядными млекопитающими, которые ведут своё происхождение от всеядных
форм
ранних копытных. Мезонихиды – полностью вымершая группа плотоядных млекопитающих,
к которой принадлежали эндрюсарх (Andrewsarchus) и монголоникс
(Mongolonyx),
самые крупные наземные хищные звери. Вроде бы, родство мезонихид и китообразных
было установлено. Но исследования в строении костей ног ранних китообразных
выявило у них несомненные признаки родства с парнокопытными. И на фоне
мозаики
фактов из небытия появился индохиус – существо, для которого словно специально
было оставлено место в этой мозаике. По странному стечению обстоятельств,
его окаменелости были извлечены из земли около 30 лет назад индийским
геологом Ранга Рао, который нашёл зубы и часть челюсти этого животного.
Многие образцы,
собранные Рао, не были отпрепарированы при его жизни, и после его смерти
вдова
Рао отдала их доктору Тевиссену. Во время препарирования один из черепов
был случайно повреждён, и взорам учёных открылась очень интересная особенность
этого животного – строение внутреннего уха, которое позволило сделать
далеко идущие выводы…
Ханс Тевиссен со скелетом индохиуса (Indohyus) |
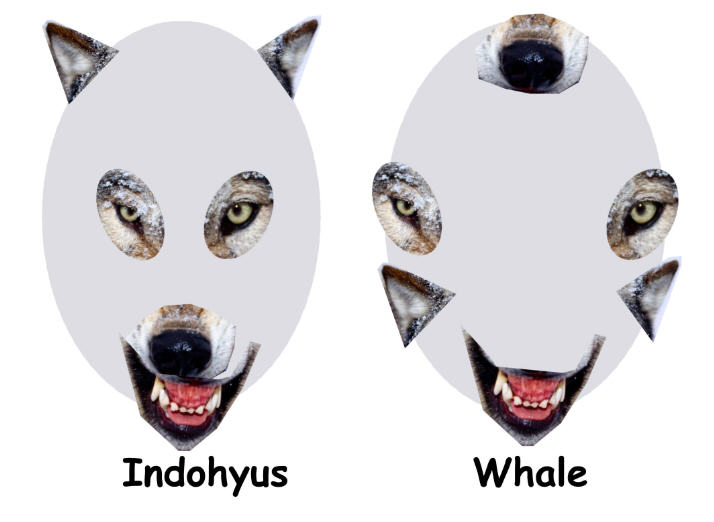
Индохиус (буквально «свинья Индии») – маленькое (размером с кошку) существо
хрупкого сложения, остатки которого найдены на территории Кашмира (Индия).
Чаще всего его сравнивают с современными африканскими водяными оленьками;
сходство нарушает только длинный хвост – общая черта различных групп примитивных
млекопитающих
раннего кайнозоя. Возраст этого существа оценивается в 48 миллионов лет.
Индохиус классифицируется как представитель семейства раоеллид (Raoellidae)
– примитивных парнокопытных. Его считают представителем сестринской группы
по отношению к ранним китообразным на основании особенностей строения ушной
области. Слуховая булла, образованная из барабанной
кости (ectotympanum),
очень необычна по форме и демонстрирует особенности строения, характерные
для открытых
незадолго до этого древнейших китов, и, в частности, для пакицетуса.
У индохиуса есть некоторые адаптации к жизни в воде – наружный слой костной
ткани утолщённый и тяжёлый, как у современных полуводных млекопитающих
типа гиппопотама. Есть и более тонкие признаки, указывающие на водный образ
жизни
индохиуса. Так, в его останках повышено содержание изотопа кислорода 18О,
что указывает на водный образ жизни. Однако, содержание изотопа углерода 13С
говорит о том, что он редко кормился в воде. Впрочем, столь же вероятно,
что его пища могла состоять из высших водных растений (цветковых). В любом
случае, судя по изотопному составу эмали зубов, Indohyus наверняка
не входил в пищевые цепочки, в основании которых лежит пресноводный
фитопланктон, образованный водорослями, а не высшими растениями.
Вполне естественно, что такое открытие не осталось незамеченным креационистами.
Одна из статей, появившаяся в креационном блоге RKBentley
on Creation, носит громкое название «Наука, или рассказывание сказок?»
(Science or Story-telling?). Если уж автор предлагает выбрать между двумя
указанными состояниями, стоит
разобраться в этом вопросе. Автор заметки прочитал статью “Whales May Be
Related to Deer-Like Beast” («Киты могут быть родственны зверю, похожему
на оленя»)
от 19 декабря 2007 года на сайте LiveScience, и утверждает, что встретил
в этой статье немало умозрительных суждений. Судя по всему, сверх этой
статьи он не читал ничего.
R. K. Bentley приводит в своей заметке целый список цитат из статьи, снабжая
их своим комментарием (цветное выделение сохранено). А я, со своей стороны, прокомментирую
комментарии… Люблю я это дело.
«Киты могут быть родственны зверю, похожему на
оленя». Забавно, не правда
ли, они называют его похожим на оленя? Не может ли это само по себе наводить
на мысль о том, что он больше похож на оленя, чем на кита? |
Собственно, Ханс Тевиссен прекрасно ответил на это возражение прямо в критикуемой статье:
«Как животное из зоопарка, он совершенно не выглядит похожим на кита»,
сказал Тевиссен. Но, добавляет он, когда дело доходит до особенностей анатомии,
индохиус
«совершенно потрясающе сходен [с ним]». |
В биологии нельзя делать выводы только на основе сходства внешнего строения сравниваемых живых организмов, поскольку существуют два явления:
Конвергенция – внешнее сходство неродственных форм, обусловленное сходством
в образе жизни.
Параллелизм – появление у организмов, относящихся к родственным группам, сходных
черт строения гомологичных органов, происходящее независимо.
Пример конвергенции – наличие крота обыкновенного в Евразии и крота сумчатого
в Австралии. Их сходство чисто внешнее, но оба этих животных совершенно не
родственны друг другу: обыкновенный крот принадлежит к плацентарным млекопитающим,
а сумчатый крот, соответственно, к сумчатым. Параллелизм демонстрируют ушастые
тюлени (потомки примитивных медвежьих) и настоящие тюлени
(потомки
куньих),
независимо
друг
от друга развившие
черты приспособления к водному образу жизни.
Поэтому вполне очевидно, что строить поспешные выводы на основе сугубо внешнего
сходства сравниваемых видов – попросту ошибочно. Так, дикобразы и ежи не являются
родственниками, хотя и те, и другие очень колючи. И индохиус может быть сколь
угодно похож на оленей некоторыми чертами строения. Но, если у него есть признаки,
которых нет у оленей, и которые не могут в процессе эволюции преобразоваться
в состояние, характерное для оленей, он не будет оленем. И зубки-то у индохиуса
точно не оленьи. А ушки – и подавно. И жил он до того, как
на Земле появились первые олени семейства Cervidae.
Сами по себе первые киты тоже были мало похожи на современных, о чём упоминал
и Тевиссен, причём в той же самой статье:
«Самые ранние киты были вовсе не похожи на китов», сказал Тевиссен. «Они
напоминали помесь между свиньёй и собакой». Они потеряли свои ноги и способность
ходить
по земле около 40 миллионов лет назад, сказал он. |
Так что злополучный индохиус, не имея сходства с современными китами, вполне может быть похожим на архаичных китов. Поэтому я просто не понимаю, над чем так зубоскалит иностранный товарищ. Над собственным непониманием теории эволюции?
«Долгие годы бегемот был главным
кандидатом на роль ближайшего наземного родственника из-за сходства
его ДНК и черт сходства с китами» И снова мы
видим предубеждения в эволюционных исследованиях. Учёные уже решили, что
киты эволюционировали от наземных млекопитающих. Они просто ищут кандидата,
чтобы подогнать под ответ: может быть, это бегемот, может быть, Indohyus,
а может быть, кто-то ещё. Среди ископаемых нет чёткой «последовательности».
Они пытаются её создать. |
Вот здесь наш креационно настроенный оппонент малость путает
разные вещи. Бегемотов считают ближайшими ныне живущими родственниками китообразных,
а индохиус – самый близкий древний родственник древнейших китообразных. В общем-то,
оба утверждения верны и не взаимоисключающи, и я попробую объяснить, почему.
Близость бегемотов к китообразным утверждается на основе исследований их ДНК.
Естественно, что такие результаты можно получить лишь на основе изучения натурального,
не окаменевшего органического материала. Поэтому круг исследованных форм в
данном случае ограничивается только современными и (теоретически) сравнительно
недавно вымершими формами, от которых можно
получить ДНК. Вполне естественно, что из-за этого в исследованную совокупность
объектов не войдут ни индохиусы, ни мезониксы, хотя было бы очень заманчиво
исследовать и их ДНК на предмет родственных отношений с китообразными. Но эти
животные вымерли слишком давно, и их ископаемые остатки давно минерализовались
– замещены
неорганическим
материалом.
Так что
ДНК из них не выбить ни крупицы. Так что близость бегемотов к китам – величина
сугубо относительная, и не предполагающая их прямого
родства. Тем более, что
первые бегемоты появились на Земле только в олигоцене, когда в океане уже вовсю
плавали разнообразные киты нескольких семейств из обоих современных подотрядов.
Если объяснять «на пальцах», можно привести такой пример. Берём на исследование
ДНК таких организмов, как дрожжевые грибки, муха дрозофила, стрекоза, карась,
человек и кит. При сравнении ДНК этих видов на предмет установления близости
к китообразным окажется, что ДНК кита покажет наибольшее сходство в
данной совокупности объектов исследования с ДНК человека. Карась окажется менее сходным
с китом, нежели человек, а ДНК дрозофилы и стрекозы окажется примерно равноудалённой
от ДНК кита по своим отличиям, которые будут больше, чем между ДНК кита и карася.
И совсем «на отшибе» по отношению к ДНК кита окажется ДНК дрожжей. Если же
мы изменим совокупность объектов исследования (при условии сохранения равной
доступности всех объектов для данного вида исследования), добавив к ней современных
верблюда, слона и свинью, то это также повлияет на результат исследования.
Человек уже не будет казаться таким близким родственником кита – между китом
и человеком встанут (по убыванию степени сходства) свинья, верблюд и слон.
Если же добавить к ней ещё и амбулоцетуса в том виде, в каком мы его знаем
(в виде минерализованных древних костей), это никак не повлияет на результаты
исследований, поскольку ДНК амбулоцетуса нам неизвестна, и потому для биохимических
исследований он будет недоступен. Если только на машине времени в прошлое сгонять…
Но это уже фантазия почти по Рэю Брэдбери.
Близость индохиуса к предкам китообразных постулируется на основе изучения
особенностей строения скелетов этих животных. В этом исследовании была задействована
более широкая совокупность объектов – не только современные («ДНК-содержащие»)
организмы, но и многочисленные ископаемые формы. И вполне естественно, что
в этом случае нашлись формы, анатомически более близкие к самым древним китам,
чем бегемоты. Если сравнивать саму методику биохимических и сравнительно-анатомических
исследований, то вполне очевидно, что сравнение прозводилось на разных основаниях
– в одном случае оценивалось сходство биополимеров (более точное, но круг исследуемых
объектов более ограниченный), в другом – чисто внешнее сходсто анатомии (менее
точное, но охватывающее значительно более широкий круг объектов). Соответственно,
при сравнительно-анатомических исследованиях скелета китообразных и их родственников
особенности строения ДНК менее важны, поскольку основное внимание уделяется
особенностям анатомии. Анализ ДНК в этом случае служит как бы для контроля
и коррекции выводов – сходство в строении скелета может быть обусловлено такими
эволюционными явлениями, как параллелизм и конвергенция. Их я имел в виду,
когда называл сравнительно-анатомический метод «менее точным» по отношению
к анализу ДНК. Анализ ДНК помогает отделить результаты параллельной эволюции
в сходных условиях от
результатов
происхождения от общего предка. Вероятность же независимого появления совершенно
идентичных участков в молекулах ДНК пусть сами креационисты рассчитают – они
любят высчитывать такие сверхмалые величины, особенно, когда говорят о зарождении
жизни на Земле. И вообще, биохимический метод неплохо зарекомендовал себя при
идентификации останков разного рода царей-королей… Ведь не может такого быть,
чтобы на царях метод действовал, а на животных – нет.
«Тем не менее, некоторые исследователи
обеспокоены тем, что бегемоты, как кажется, жили не в той части света
и появились слишком поздно, чтобы
быть предками китов». И снова предубеждения. Как видите, учёные уже решили,
где и когда эволюционировали киты – теперь мы лишь должны найти их предков
там! |
Нет, милейший! Учёные просто очертили место и время, где и когда НЕ стоит искать предков китов. Прямого указания на то, кого искать, где и когда, в приведённом отрывке нет. Помните, как Шерлок Холмс расследовал дело об убийстве Чёрного Питера? Того самого, которого прикололи гарпуном к стене? Он просто попробовал пробить этим гарпуном свиную тушу в мясной лавке, и на основании результатов такого эксперимента сразу отсеял из числа подозреваемых людей хрупкого телосложения. Но имя убийцы даже он не смог назвать сразу, и потребовалась дополнительная работа. Так же и здесь – отсеять бегемотов из кандидатов на предков китов (на основании того, что они жили слишком поздно для этого) ещё не означает назвать предков китов с точностью. Об отношении бегемотов к происхождению китов я говорил выше, и потому не буду повторяться.
«Животное является «недостающим
звеном» к сестринским видам древних китов». Вы когда-нибудь интересовались тем, что подразумевается под термином «недостающее
звено»? Он используется, чтобы описать животных, которых учёные рассчитывают
найти, но не находят (вот, почему они названы «недостающими»). Весьма странно
наблюдать учёных, которые бегают вокруг, заявляя, что эволюция – это факт
(!), хотя они до сих пор пытаются найти её свидетельства. «Киты произошли
от наземных животных; как только мы найдём недостающее звено, я это вам
докажу!» Не может ли так оказаться, что они «недостающие» потому, что их
нет? |
Вся беда этого рассуждения в том, что переходных форм находят
достаточно много, чтобы всякий, кто хочет, мог убедиться в том, что теория
эволюции верна.
И
только креационисты отбрасывают их подряд одну за другой, объявляя «мозаичными»
и требуя, чтобы им показали «более другие». А когда начинаешь выяснять, чего
именно они хотят, бравые голоса разом смолкают и оказывается, что господа совсем
даже не определились в своих желаниях.
Вполне естественно, что некоторые из форм, которые официальная наука объявляла
«переходными», также оказывались представителями уклонившихся родословных линий,
и их исключали из числа «переходных». Но это делалось на основе анализа особенностей
строения, причём не только их самих, но и их ближайших родственников, вымерших
или современных.
Выражение «недостающее звено» (в английском языке фраза “missing link” имеет
ещё один смысловой оттенок – «потерянное звено») объяснено не совсем правильно.
Это не просто некие неизвестные существа, а именно члены определённой эволюционной
последовательности от предка к потомку, реконструированной на основе уже имеющихся
палеонтологических
находок и свидетельств смежных биологических дисциплин. Вполне естественно,
что в первых схемах, иллюстрирующих родословные различных групп живых организмов,
время и порядок отделения дочерних групп от предковой группы могли быть выведены
на основе неполных или вообще косвенных данных – например, на основе сравнительно-анатомических
исследований. По мере появления новых находок родственные отношения внутри
изучаемой группы уточняются и пересматриваются. А находки, сделанные позже
и подтверждающие своим существованием ранее сделанные теоретические
выкладки,
как раз и носят название «недостающего
звена».
Иными словами, это формы, которые уже есть, и чьё существование было предсказано,
а не те, которых пока ещё нет в
наличии, как пытается доказать R. K. Bentley.
«Исследование его зубов показало,
что наземное животное проводило много времени в воде и могло кормиться
там, подобно бегемотам и китам». И подобно
выдрам. Вы забыли упомянуть, что выдры проводят много времени в воде. Я
думаю, это животное выглядит больше как выдра, и потому я больше склоняюсь
к тому, чтобы считать его скорее предком современных выдр, чем китов. |
А это уже совершенно глупое предположение, сделанное по типу «лишь бы ляпнуть абы что». Когда на ногах индохиуса растут копытца, представить себе превращение их в когти выдры (то есть, переход из специализированного в более примитивное состояние) довольно сложно. И ещё сложнее представить, как выдра может классифицироваться в отряде хищных, произойди она от индохиуса. Данные биохимии сразу указали бы на её родство с парнокопытными, и это было бы замечено уже в первом приближении. И в анатимии выдры также прослеживались бы особенности, характерные для копытных. Тем не менее, сейчас выдра исправно классифицируется как представитель отряда хищных, и её анатомия не вызывает подозрений в её «чужеродности» по отношению к остальным куньим. В биологии, увы, одного только «выглядит больше как…» совершенно недостаточно для классификации. Было такое раньше – слонов, носорогов и бегемотов объединяли в одну группу «толстокожих» (Pachydermata). И где они, «толстокожие», теперь? Слоны – в отряде хоботных, носороги – среди непарнокопытных, а бегемоты – нежвачные парнокопытные. Так же зайцы, исправно классифицировавшиеся среди грызунов, стали отдельным отрядом. Искусственное объединение по произвольно взятому признаку не выдержало проверки данными сравнительной анатомии и палеонтологии. С тех пор, я полагаю, наука всё же шагнула вперёд достаточно, чтобы не повторять таких явных ошибок. Или у креационистов иное мнение?
«Из набора костей он создал
составной скелет существа возрастом 48 миллионов лет. … Характерное
расположение и форма некоторых коренных зубов сближает
индохиуса с самыми ранними китами возрастом около 50 миллионов лет». Это
любопытно. Поверю ли я, что существо возрастом 48 миллионов лет является
предком существа возрастом 50 миллионов лет. |
И я не поверю. Тем более, что не ранее, чем пять фраз назад
в излияниях R. K. Bentley проскочил термин «сестринские виды», который
вовсе не предполагает непосредственного происхождения одного такого вида от
другого,
зато предполагает их совместное происхождение от общего предка. Конечно, есть
и другое непротиворечивое объяснение этому факту. Учёные не могут быть уверенными,
что откопали имено самый древний экземпляр данного вида. Любой вид существует
на протяжении какого-то времени в истории Земли. В начале своего становления
как самостоятельной группы живых организмов он похож на родственные виды и
отличается от них весьма незначительно. Его популяция на ранних стадиях обособления
очень невелика и вероятность сохранения в ископаемом состоянии совершенно незначительна.
Далее, если вид не вымирает, он переживает определённую фазу расцвета. Тогда
он хорошо отличим от родственных видов, его численность увеличивается, а ареал
может расширяться. И далее вид или дробится на новые виды, или вымирает. В
случае вымирания его ареал разбивается на ряд очагов, в которых существуют
малочисленные популяции, численность которых снижается под действием факторов
окружающей среды. В таком состоянии вид также становится «невидимкой» в палеонтологической
летописи. Поэтому находка, чей возраст оценен в 48 милионов лет, вовсе не ограничивает
фактическое время существования вида кратким промежутком времени, близким к
этому возрасту. Поэтому для Indohyus всё же существует некоторая теоретическая вероятность
того, что он мог быть предком ранних китов.
Кроме того, в приводимом R. K. Bentley отрывке сказано, в общем-то, о том,
что Indohyus был близок к ранним китам, что, в общем-то,
не обязательно подразумевает отношений типа «предок – потомок». Поэтому я могу
утверждать, что наш собеседник ищет в словах исходной статьи то, чего там нет.
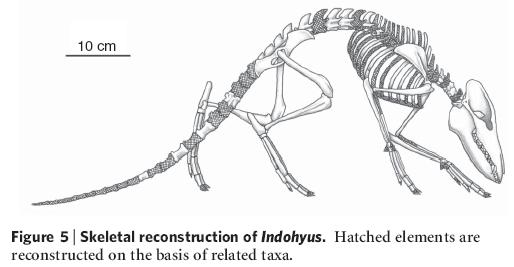
Реконструкция скелета индохиуса. Заштрихованные элементы реконструированы на основе сведений о строении близких видов.
«Ключевая находка, связывающая
индохиуса с китом – это его утолщённая ушная кость, нечто, наблюдаемое
только у китообразных». Когда я это прочитал,
я громко смеялся. Неужели это не может быть истолковано как свидетельство
того, что утолщённая ушная кость также встречается у других животных
помимо китообразных? Разумеется, если это животное не признано родственником китов,
нельзя дальше говорить, что утолщённая ушная кость «встречается только
у китообразных». |
Что ж, это неизбежно в процессе познания. И это говорит о
том, что рамки, в которые человек загоняет природу, очень условны. Например,
раньше выражение «белый, как лебедь» было верным. Но потом в Австралии открыли
чёрного лебедя, а в Южной Америке – черношейного. Или, ближе к теме, ранее
считалось, что для усатых китов характерно наличие китового уса. Затем в 2006
году в Австралии открыли ископаемого кита Janjucetus hunderi, у которого при
особенностях строения, характерных для усатых китов, оказался полон рот зубов,
да таких, что иная акула позавидует. Тем не менее, его всё равно относят к
подотряду Mysticeti, к которому несомненно и без всяких исключений относятся
все современные усатые киты.
Я просто напомню про то, что индохиуса относят к «сестринской группе» по отношению
к древнейшим китам. Поэтому он просто мог унаследовать особенности строения
уха от общего предка. Где здесь противоречия?
Вполне естественно, что в биологии нельзя отрицать версии появления у представителей
разных групп живых организмов сходных признаков в ходе параллелизма или конвергенции.
Параллельное формирование сходного
признака, однако, происходит у близкородственных форм на одной основе – это
главный признак явления. Поэтому, если предположить независимое, но параллельное возникновение
сходного признака у раоэллид и ранних китов, это всё равно автоматически подразумевает
близкое родство данных групп с расхождением незадолго до формирования
характерного строения слухового аппарата). Если же предположить конвергентное происхождение
сходства в строении уха у этих групп (то есть, ранние киты и раоэллиды по условию
представляют собой группы, не родственные друг другу), то
строение органа слуха
у них заведомо должно отличаться, показывая лишь поверхностное сходство. Этого,
однако, не наблюдается.
«При появлении опасности он прыгает в воду и прячется». Как это возможно
узнать нам? Неужели кто-то откопал видеозапись индохиуса возрастом 48 миллионов
лет наряду с его костями? Это 100% домыслов и 0% науки. |
Между прочим, в оригинальной статье сказано:
«А индохиус? «Совсем крошечный олень размером с енота и без рогов», сказал
Тевиссен. Он сказал, что это больше похоже на ныне живущего африканского
оленька, у которого нос похож на крысиный, и «при появлении опасности он
прыгает в воду и прячется». |
Интересное кино получается, не так ли? Это не про самого индохиуса Тевиссен
говорит, а про современного водяного оленька, с которым индохиус сравнивается.
И киносъёмки плывущего под водой оленька не представляют особой редкости. Так
что желчь слита не по адресу.
Честно говоря, я тоже хочу тут шутки юмора пошутить по этому поводу. Например,
мне крайне желательно было бы увидеть видеозапись событий, происходивших, если
архиепископ Ашер в своё время не соврал, в 4004 году до н. э., в период примерно
с 17-го по 23-е октября. Догадываетесь, о чём я? Правильно, о сотворении мира.
Есть
такие видеозаписи? Нету? Тогда и нечего огрызаться на эволюционистов. А то
креационисты с таким апломбом утверждают, что им известна «несомненная истина»,
что я просто начал думать, что в подвалах Ватикана или в тайной кладовке под
Стеной Плача хранится едва ли не глиняная табличка, подписанная самим Яхве…
Но, если отложить шутки в сторону, отдельные элементы поведения доисторических
животных всё же можно реконструировать, основываясь на особенностях строения
скелета и косвенных данных, получаемых при изучении ископаемых остатков. Хотя
бы упомянутое содержание изотопов в костях с опорой на знания о современных
животных уже даёт основание делать какие-то выводы о поведении животного. А форма
самих костей поможет реконструировать развитие мускулатуры и места прикрепления
сухожилий, что, опять-таки, даёт ключ к исследованию физических возможностей
животного: помогает оценить подвижность его суставов и усилие, котрое могли
развивать его мускулы. Соответственно, это позволяет оценить способность исследуемого
существа к тем или иным способам передвижения.
Это прекрасный пример того, как создались многие эволюционные теории.
Вот их новый «научный метод»: 1) Выдвинуть идею о том, как проходила эволюция.
2) Найти окаменелость. 3) Написать сказку о том, как окаменелость подходит
к вашей идее из шага №1. 4) Затем учёные связывают различные сказки в одну
большую сказку под названием Теория Эволюции. |
Ну, в части рассказывания сказок и выдавания их за реальность
эволюционисты вчистую проигрывают своим религиозно настроенным оппонентам.
Мне самому приходилось
сталкиваться с элементами креационистского мифотворчества. То у креационистов
скелет злосчастного австралопитека Люси оказывается разделённым пополам слоем
базальта, то
тот же самый скелет оказывается собранным из скелетов 30 разных особей австралопитека.
Рядом с эогиппусом, якобы, находят останки лошадей современного рода Equus.
Трилобит
оказывается раздавленным современным ботинком (интересно, что вид трилобита
нигде не указан, и торговая марка обуви тоже). Следы динозавров оказываются
перекрытыми следами «древнего человека». И неясно, кто именно пускает таких
«уток»…
Если же говорить о рождении самой теории
эволюции, то его сценарий очень далёк от того, о чём говорит R. K. Bentley.
Начать стоит хотя бы с того, что мысли о постепенном развитии живой природы
от простого к сложному высказывались и до Ч. Дарвина. Далее, если всё же почитать
саму книгу Дарвина, а не её название на корешке, можно увидеть, что Дарвин пришёл
к выводу об эволюции органического мира не спонтанно (типа «а ну-ка, сяду-ка,
да щаз такую чучу отчебучу»...), а после кропотливого сбора и анализа материала.
Он много пишет о различных опытах, собирает факты, касающиеся того или иного
явления, анализирует их. В более поздних изданиях «Происхождения видов...»
он полемизирует с противниками теории эволюции (например, с Майвартом) и анализирует
их возражения. Теория эволюции, к сожалению для R. K. Bentley и других креационистов,
строится не только на окаменелостях. Но они служат весомым доказательством
в её пользу, дополняя данные сравнительной анатомии, эмбриологии и других биологических
дисциплин. Вполне естественно, что палеонтологические свидетельства могут отпровергать
первоначально построенные модели эволюции той или иной группы живых организмов.
Так произошло, например, с предположениями о путях ранней эволюции китов (об
этом подробнее сказано ниже), поэтому находки не обязательно должны «подходить»
к ранее выстроенным логическим цепочкам: здесь R. K. Bentley лукавит по-чёрному.
Тем не менее, практика показывает, что, служа поводом для опровержения некорректных
моделей эволюции той или иной группы
организмов,
или дополняя
и уточняя существующие
модели, палеонтологические свидетельства никогда не опровергают саму теорию
эволюции в целом – вот, с чем не смирятся креационисты.
R. K. Bentley в свете анализа его комментариев выглядит просто бравирующим
неучем, воинственно стучащим себя пяткой в грудь. Вряд ли он имеет какие-то
представления о теории эволюции на более высоком уровне, чем их даёт зарубежный
школьный учебник…
Вполне естественно, что мнение креационистов не ограничивается
этой заметкой. Вот
ещё одна статья:
Post
details: Has the missing link of whale evolution been found? (Последующие
подробности: Найдено ли недостающее звено в эволюции китов?)
21.12.07
Давид Тайлер (David Tyler)
В своей заметке в Nature Тевиссен и др. поясняют, что в истории эволюции китов недостаёт важной ключевой точки: «звено к предку китообразных утрачено». Это не просто случай незнания, а точка основополагающих различий между исследователями. Некоторые делают упор на морфологии, другие на молекулярных свидетельствах – и вас не должно удивлять, если вы услышите о недостатке согласованности в понимании этих данных. Сейчас Тевиссен и его коллеги думают, что у них есть ответ:
Они использовали кладистическую методику, чтобы показать близость раоэллид и китообразных (хотя мы должны заметить, что китообразные представлены Pakicetus, Ambulocetus, Rodhocetus и Artiocetus – все предположительно переходные ходячие киты). Кладистика – не самый честный инструмент, чтобы использовать его для целей отслеживания родословных. Это так, поскольку существует много общих [в оригинале сказано “shared”, что также имеет смысловой оттенок «находящийся в совместном пользовании у кого-либо» - В. П.] черт, которые следует принимать во внимание, и исследователям необходимо выбирать совокупность особенностей, которые они используют в кладограммах. Наибольшее внимание уделяется поиску наиболее экономных родословных древ: с минимальным количеством эволюционных изменений, необходимых для истолкования данных (критерий, который поражает меня как нереалистичный применительно к эволюции китообразных, процессу, который обязательно требует экстраординарного количества изменений). Эти общие черты не всегда чётко подходят к кладистическим рамкам анализа: многие животные (современные и вымершие) демонстрируют загадочную мозаику особенностей. |
Но исследователи, тем не менее, вполне в состоянии выделять
черты, общие для представителей того или иного отряда или иной таксономической
единицы. Вначале признаки, по которым производилась классификация,
действительно могли быть достаточно произвольно выбранными. Чтобы убедиться
в этом, можно полистать старые и репринтные (без современных поправок) издания
А. Э. Брема. Среди птиц выделяются «боевые птицы» (включающие, в частности,
соколообразных и всех водных птиц), к куриным относят киви. Отряды рыб, выделяемые
во времена Брема, вообще не совпадают по объёму ни с одним отрядом рыб в современном
понимании их систематики. Я уже упоминал группу «толстокожих» млекопитающих,
которая также «растворилась» в свете более поздних исследований.
Современная систематика тех же самых млекопитающих обычно не вызывает затруднений.
Среди современных млекопитающих нет групп, положение которых вызывало бы сомнение
(среди птиц, например, фламинго могут включаться в отряд Аистообразных, либо
не включаться, а гоацин у разных специалистов «болтается» между отрядами Курообразных
и Кукушкообразных). Палеонтологические и анатомические исследования помогли
«развести» грызунов и зайцеобразных, а бывших ластоногих сейчас полностью включают
в отряд Хищных на правах даже не подотряда, а просто трёх отдельных семейств.
Неполнозубые, выделенные по состоянию на конец XIX века, уже давно разделены
на собственно неполнозубых, панголинов и трубкозубов. Современные методы исследований
(например, биохимические) помогают установить родственные связи между отрядами,
и их результаты помогают уточнить систематику. Например, бывшие насекомоядные,
которые выглядели как «мусорная корзина», куда «сбрасывали» всех мелких животных,
не вошедших в остальные отряды, сейчас «трещат по всем швам». Так, прыгунчики
выделены в особый отряд Macroscelidea с намёком на их близость к приматам,
златокроты и тенреки выделены в отряд Afrosoricida, а среди бывших насекомоядных
(отряд Eulipotyphla) остались ежи, землеройки, щелезубы и кроты.
На фоне этих перестановок и пересмотров объём отряда китообразных выглядит
неизменным – современные представители этой группы чётко отделены от прочих
млекопитающих по особенностям строения. Но, кроме современных представителей,
в любом отряде есть древние, и китообразные тут не исключение. Пока древние
киты представляют собой, судя по строению скелета, постоянноводные формы, близкие
к современным, проблем с классификацией не возникает. Но, если двигаться
к истокам отряда,
ситуация становится сложнее. Действительно, чем примитивнее представители линии,
ведущей от современных китов в прошлое, к древним представителям группы, тем
меньше у них черт строения, общих с современными китами. И здесь уже возникают
вопросы. У современных китов только одна пара плавников – передняя. У архаичных
китов типа базилозавра и дорудона – две пары. Это киты, или не киты? У современных
китов ноздри находятся в верхней части головы, чаще всего почти на темени (у
кашалота всё же на конце морды). У древних китов они находятся «на полпути»
между кончиком морды и теменем. Это
киты,
или не
киты? У современных китов зубная система гомодонтная (то есть, все зубы одинаковы,
словно у рептилий), а у древних – прослеживается чёткая дифференциация зубов,
как у типичных млекопитающих. Это киты, или не киты? Так по каким же признакам
определить, является ли животное китообразным? Здесь напрашивается параллель
с античной загадкой про Тесеев корабль. Суть её в том, что во время морского
путешествия Тесею и его спутникам приходилось постоянно чинить свой корабль,
пострадавший во время очередной бури, заменяя повреждёные
части новыми. В конце концов, от старого корабля ничего не осталось, поскольку
все его части были одна за другой заменены новыми. И возникает вопрос:
корабль Тесея в конце путешествия – это тот же корабль, что и в начале путешествия,
или уже другой?
Случай с китообразными немного напоминает загадку про Тесеев корабль, но как
бы наоборот: известно конечное состояние объекта (строение современных представителей
отряда китообразных), и от него прослеживается переход к начальной стадии их
эволюции. Китообразные действительно сильно изменились в течение времени их
существования как самостоятельной группы млекопитающих. Поэтому выделить
признаки,
общие для
абсолютно
всех когда-либо
живших
представителей
отряда,
очень сложно. Тем не менее, это вполне можно сделать, хотя таких признаков
становится очень мало. Если двигаться по ряду китообразных, который выстроен
на основе палеонтологических данных, из настоящего в прошлое,
черты современных китов начинают постепенно сглаживаться, и одновременно появляются
черты, совершенно
чуждые современным китам. Дыхало превращается в ноздри и «съезжает» вначале
на переносицу, а затем и вовсе на кончик морды. «Прорезаются» задние конечности,
которые увеличиваются. Позвонки из почти одинаковых дифференцируются на грудные,
поясничные, крестцовые и хвостовые, к ним «прирастает» таз. Конечности постепенно
теряют ластообразную форму, для них становятся характерными подвижные суставы,
локти и колени. Зубы из одинаковых превращаются в дифференцированные. Что же
остаётся от китов-то?
Лукавит Давид Тайлер, когда говорит о какой-то «экономии» и «минимальном количестве
эволюционных изменений». Задача науки, к сожалению для его построений – не
создание красивых картинок, а максимальное приближение к истине, в данном случае
– максимально точная и непротиворечивая реконструкция пути эволюции китообразных.
Поэтому ни о каком «минимальном» или «максимальном» количестве каких-либо признаков
не может идти речь. И признаки для классификации, соответственно, учёные не
«с потолка» берут, а выявляют в ходе анализа строения современных и ископаемых
представителей отряда. Наиболее характерным признаком, который прослеживается
у всех китообразных, от древних до современных, является строение внутреннего
уха. Прочие признаки строения скелета, характерные для древних китообразных,
могут быть не характерны для китов в их нынешнем виде. Тем не менее, их наличие
характеризует древнейших китов как единую группу, и отличает их от прочих млекопитающих
того времени. К таким признакам относятся, в частности, зубы. Строение зубов
отличает китообразных от прочих древних потомков кондилартров, в частности,
от мезонихид (хотя с ними имеется определённое сходство), но зубы ископаемых
китов, в свою очередь, отличаются от зубов современных китов.
Далее, «сходен» ещё не означает «является родственником». Плотный наружный
слой костей конечностей и соотношение изотопов – это не систематические признаки,
а признаки, дающие ключ к пониманию особенностей образа жизни. Например, рацион
ископаемых лошадей также определяется по соотношению изотопов, а кости конечностей
отличаются повышенной плотностью и у современных водных млекопитающих типа
гиппопотама, о чём говорилось выше. Так что это не желаемые признаки, лукаво
и избирательно подобранные для создания в сознании обывателя нужной картины
событий, а результат изучения объективно существующих признаков строения скелета,
которые так просто не отбросить.
Судя по тексту, Давида Тайлера также смущает количество изменений, произошедших
в ходе эволюции китов. Что ж, если сравнивать скорость ранней эволюции китов
со скоростью эволюции китов во второй половине неогена, различие будет заметно.
В эоцене менее чем за десять миллионов лет киты превратились из существ со
скелетом наземного четвероногого в полноценных обитателей океана. Этому нетрудно
дать объяснение, если оценить обстановку в морской фауне прошлого. В эоцене
у первых китообразных не было конкурентов в море. Акулы в то время были сравнительно
мелкими, а ламноидные акулы (те самые, которых мы представляем себе при слове
«акула») появились несколько позже – в позднем эоцене. Ластоногие (несколько
ветвей водных хищников) появились как таковые лишь в олигоцене. Поэтому ранние
киты
могли
относительно свободно занимать новые места обитания, и скорость накопления
новых признаков могла быть гораздо больше, чем в условиях жёсткой конкуренции
с уже занимающими «китовые» экологические ниши видами, когда признак, недостаточно
повышающий конкурентоспособность, отсеивается отбором уже на ранних стадиях.
В мезозое, когда моря кишели морскими
рептилиями разных групп, млекопитающие не слишком
охотно
шли
в воду – ничего крупнее многобугорчатого млекопитающего Castorocauda lutrasimilis величиной
с выдру в отложениях того возраста пока не найдено.
Ещё одним проблемным вопросом Давид Тайлер считает слишком большие изменения,
которые произошли в строении китообразных в ходе их эволюции. Роберт Кэрролл
в книге «Палеонтология и эволюция позвоночных» (глава 22 «Эволюция») приводит
факты, свидетельствующие о том, что потенциал у процесса эволюции просто огромен:
Холдейн (Haldane, 1949) предложил особую единицу для их (скоростей эволюции)
измерения – дарвин. По определению 1 дарвин (d)
– изменение величины признака в e раз за один
миллион лет (e –
основание натурального логарифма). Логарифмическая шкала здесь удобна,
поскольку интерес представляют не абсолютные, а относительные изменения
параметров. ... Во многих случаях распространено изменение размеров особей, происходящее иногда очень быстро за короткое время, но если брать усредненные цифры, учитывающие периодические увеличения и уменьшения размеров тела, эти изменения покажутся куда более медленными. Отдельный скелетный элемент, например, таранная кость непарнокопытных, может заметно измениться за период времени менее 5 млн. лет, а в течение следующих 55 млн. лет оставаться практически неизменным. Скорость преобразований окажется намного ниже, если измерять ее для всего срока существования отряда. ... Следует особо подчеркнуть, что почти все опубликованные результаты расчетов скорости эволюции относятся к ограниченному числу признаков и, возможно, не характеризуют организм как целое. Молекулярные данные говорят о связи с анатомическими изменениями весьма незначительной части генома. Как показали Кинг и Уилсон (King, Wilson, 1975), различия в аминокислотном составе 12 различных белков человека и шимпанзе составляют всего 1%. |
Так что ранние китообразные на заре истории отряда вполне могли развиваться в значительно более быстром темпе, нежели после того, как перешли к полностью водному образу жизни.
Исследователи выделили четыре особенности, которые они считают имеющими значение: строение ушей и коренных зубов, плотность костей конечностей и соотношение стабильных изотопов кислорода в их зубах. Из этого они делают заключение:
До тех пор, пока эти специфические особенности могут использоваться, чтобы отстаивать мысль о том, что животные были водными [в оригинале сказано “water waders”, буквально «бродящими в воде» - В. П.], заявление о том, что они представляют собой переход от парнокопытных к китам, принадлежит к числу предположений. Гипотезы должны быть проверены, а исследователи должны бы давать оценку своим собственным гипотезам по сравнению с альтернативными. В качестве предложения обдумайте гипотезу о том, что Indohyus был специализированным парнокопытным, ищущим свой корм в обводнённых местообитаниях. Более того, исследователи должны оградить себя от вероятности того, что это специально подобранные особенности, которые позволяют им добиться желательного вывода. |
Давид Тайлер забыл упомянуть, что мысль о родстве китообразных и парнокопытных возникла не сразу после находки индохиуса. Если вникать в хронологию событий, то окажется, что идея о происхождении китообразных от парнокопытных появилась приблизительно в 2001 году (тогда же появились критические заметки креационистов на этот счёт), заведомо ранее декабря 2007 года, при изучении строения лодыжки ранних китообразных. Именно тогда оказалось, что таранная кость китообразных имеет одинаковые особенности строения с костями парнокопытных. Индохиус здесь – пример, подтверждающий верность выводов и предсказательную силу теории эволюции.
Сообщение Stokstad ставит под вопрос заявление о том, что найден давно
исчезнувший родственник китов. Он заключает: «Не все, однако, убеждены в том, что Indohyus является ближайшим родственником китообразных. Другой анализ, опубликованный в Cladistics, говорит о том, что вымершая группа плотоядных млекопитающих, называемых мезонихидами, была более близкородственной китообразным». Ссылка на эту заметку дана ниже. [доступ к статье платный, потому прочитать саму статью не удалось – В. П.] |
Хорошо. На секунду представим себе, что ситуация изменилась.
Завтра Ханс Тевиссен или кто-нибудь из его коллег напишет: «Первоначально сложившееся
впечатление о происхождении китообразных от парнокопытных оказалось ошибочным.
На самом деле признаки A, B, C, … X, Y, Z прямо свидетельствуют о происхождении
китообразных от мезонихид». И что тогда? Креационисты выйдут на улицы плясать
и распивать кагор? Ничего подобного. Вывод о происхождении китообразных от
наземных четвероногих млекопитающих останется, и они будут сражаться теми же
методами уже против «мезонихидной» версии.
Возможно, кто-то из креационистов в очередной раз скажет: «Не поймёшь вас –
то от мезонихид, то от парнокопытных. Где же истина, о которой вы говорите?»
Такие доводы в явной или скрытой форме всегда будут. Дело в том, что учёные
заведомо не располагают полной информацией о вымерших животных. Нам неизвестно
строение их мягких тканей, биохимические особенности, особенности поведения.
Для уточнения родословной китообразных не худо было бы заполучить образцы мягких
тканей мезониксов, раоэллид и первых китов – тогда выводы были бы изумительно
точными и однозначными. А нету. Поэтому приходится интерпретировать косвенные
данные – особенности строении скелетов – и спорить, является ли тот или иной
признак результатом конвергенции, параллелизма, или непосредственного наследования
от общего предка. Поэтому, при всей привлекательности и доказанности на данный
момент «парнокопытной» версии происхождения китов для «мезонихидной» версии
всё равно останется хотя бы какое-то место. Работу палеонтологов можно сравнить
с работой криминалистов – появление новой информации заставляет пересматривать
старые
взгляды и порой
отказываться от привлекательных и стройных гипотез, непротиворечиво объяснявших
известную в какой-то момент развития науки совокупность фактов.
Тевиссен и др. предлагают адаптационистский сценарий для пути эволюции к китообразным. Ближайшей аналогией видится африканский оленёк, который живёт вблизи рек, кормится на суше, но при возникновении опасностей бросается в воду.
Сообщение в одной из газет продолжает сказку: «Тем не менее, никто не знает, почему некоторые млекопитающие вернулись в воду. Некоторые из них могли вернуться в реки и озёра, чтобы избежать хищников или в поисках лучшего корма. Поскольку они проводили всё больше времени в воде, их ноги эволюцировали в ласты – тогда как их носы превращались в дыхало, расположенное на вершине китовых голов». Это звучит как хорошее начало для сказки Рэдьярда Киплинга! |
Нет, про слонёнка у него лучше написано. И про то, откуда взялись броненосцы.
Хотя я до сих пор не пойму, откуда в Южной Америке взялись ежи, которые и превратились,
по сказке, в броненосцев. Киплинга-то можно простить – он не зоолог, и «сказка
– ложь, да в ней намёк: добрым молодцам урок». А вот современным сказочникам
из числа креационистов хочется рекомендовать то же самое, что советует в своей
статье Давид Тайлер – проверять свои гипотезы, сравнивая их не только с Книгой
Бытия, но и с более близкими к биологии первоисточниками. Иначе умопостроения
получаются просто термоядерные.
Из сообщения Тевиссена ясно видно, что при выдвижении предположения он опирается
на факты, полученные в ходе исследований (на глаз содержание изотопов не определишь,
не так ли? Тут тонкий анализ нужен) и сравнивает их с особенностями современных
животных, имеющими место быть. Почему Тевииссен, например, не предположил,
что индохиус был лазающим? Правильно, строение его пальцев не предполагает
хватательной функции. А почему не сказано, что он был, допустим, летающим?
Верно, крыльев нет. Все выводы делаются на основе строения реальных ископаемых
остатков, и домыслам, не основанным на реальных особенностях изучаемых объектов,
в науке места нет.
И вот совсем свеженькая, от января 2008 года, статья “Indian whales” («Индийские
киты») за авторством Do-While Johnes, размещённая на сайте Science
against Evolution,
обрушивается с разрушительной и уничтожающей критикой на статью в журнале Nature от 20 декабря
2007 года. Посмотрим, что нам предложит этот товарищ... В цитатах я сохранил
выделение
цветом, использованное в данной работе.
Давайте переведём этот параграф на простой английский. ДНК китов наиболее сходна с ДНК гиппопотамов. Поэтому молекулярные биологи говорят, что киты должны были эволюционировать от раннего бегемота. Палеонтологи не признают этот аргумент, поскольку они думают, что самые старые ископаемые остатки китов имеют возраст 50 миллионов лет, а самые старые ископаемые гиппопотамы – только около 15 миллионов лет. Если киты предшествовали гиппопотамам на 35 миллионов лет, они не могли эволюционировать от них. Но летопись окаменелостей «не разрешила вопроса о родственных связях китообразных». Итак, есть несколько различных предполагаемых предков китов. |
Пример необоснованной «популяризации»
(точнее, «вульгаризации») идей о родстве китообразных и парнокопытных:
в книге А. А. Гангнуса «Технопарк юрского периода» довольно поздний
представитель китов, Rodhocetus, назван «свинокитом», хотя
его непосредственные предки были не свиньями, а китообразными.
Первые китообразные, несомненно, являются
родственниками парнокопытным, но это ещё не означает, что именно свиньям,
так как свинья – это лишь частный случай парнокопытных. Кроме того,
семейство свиней появилось на Земле в олигоцене, а бегемоты в раннем
миоцене – заведомо позже первых китообразных. Поэтому идею о существовании
«свинокитов» лучше оставить для очередного римейка фильма «Остров доктора
Моро». |
Эх, переводчик вы наш, господин хороший… Таких бы «переводчиков»,
да к переводу библейских текстов привлечь, и тогда, боюсь, сторонники одного
перевода Библии порвали бы зубами глотку сторонникам иного перевода, и сожрали
бы своих противников «без соли и без лука». Во всяком случае,
вместо
«перевода» у нас имеет место совершенно отвлечённый комментарий, не имеющий
почти ничего общего с самим текстом, кроме повторения некоторых слов.
Итак, русским по белому… В смысле, русским голосом… Короче, не знаю, впитает
ли Do-While Johnes этот комментарий, но для его единомышленников, пишущих,
говорящих, и (надеюсь!) понимающих по-русски, попробую объяснить, что к чему.
Начнём с того, что в данной цитате НЕ сказано ничего типа «бегемоты были предками
китов». Зато Do-While Johnes старательно пытается создать именно такое впечатление.
В тексте сказано следующее: «Филогенетический анализ молекулярных данных современных
животных надёжно поддерживает утверждение, что гиппопотамиды – ближайшие
родственники китообразных (киты, дельфины и морские свиньи)». Это
не моё желание «обелить» тех, чьё мнение я поддерживаю в силу своих убеждений,
а адекватный перевод:
в английском тексте сказано: “closest relatives”, а не, допустим, “ancestors”,
“ascendants” или “parental group”. «Ближайшие родственники» – это ещё не
означает «предки», верно? В научной литературе, в том числе в работах, посвящённых
родственным
связям ранних китообразных, говорится о том, что бегемоты являются «сестринской
группой» для китов, то есть, среди современных животных они
наиболее близки к китам. Но это ещё не означает, что среди ископаемых животных
не было ещё более близких родственников для китообразных. Другой вопрос, что
биохимические исследования
в этом случае недоступны, т. к. ДНК ископаемых животных в большинстве случаев
просто разрушается до отдельных крохотных фрагментов. Впрочем, о кажущемся противоречии
результатов я уже говорил немного выше.
Итак, чем дальше кормим читателя?
Это новое исследование, предположительно, решило все проблемы. Мы были в какой-то степени сконфужены этим резюме к статье, написанным редактором Nature.
Предположительно, первые 10 миллионов лет эволюции китов должны были
заключаться в эволюции с суши в воду. Если
эти 10 миллионов лет столь хорошо документированы, почему их происхождение
всё ещё неясно? По-видимому,
фраза «хорошо документированы» означает разные вещи для разных людей.
Indohyus похож на китов по форме своих премоляров (зубов), но его рацион
(о котором делаются выводы, главным образом, на основании формы зубов)
был «очень непохожий на китовый». Процесс мышления редактора Nature,
скорее всего, идёт примерно таким образом: |
Итак, что у нас здесь? Do-While Johnes снова что-то путает…
Или опять не путает, а злонамеренно искажает? Во всяком случае, в цитате сказано
про первые 10 миллионов
лет эволюции китов. Не поняли? Хорошо, скажем так: тех
животных, которых уже можно считать именно китами, и никем другим. Но животное, которое
было непосредственным
предшественником самого раннего кита вроде Pakicetus, пока не найдено.
Фигурально выражаясь, у палеонтологов пока есть «рамка без портрета»: они обрисовали
приблизительный
облик предка китообразных из числа не-китообразных (то есть,
того, кого ещё нельзя считать китом), определили, кому он может приходиться
родственником,
и обнаружили Indohyus, ещё одного из его потомков, близкого родственника
архаичных китов.
Относительно причин перехода раоэллид к водному образу жизни можно лишь строить
предположения. Учёным заведомо неизвестны все обстоятельства этого процесса,
которые не могут быть изучены по причине исчезновения экосистем прошлого и
сохранения в ископаемом виде лишь малой части остатков организмов, образовавших
эти экосистемы. В их распоряжении есть только косвенные свидетельства в виде
анатомических особенностей этих животных, которые приходится интерпретировать
в свете сравнения с современными животными, имеющими такие же особенности.
Что же такого есть в черепе Indohyus, что делает его похожим на кита? Ну, есть некоторые черты сходства.
Мы должны отметить, что существуют некоторые различия, которые мы считаем важными. У Indohyus нос расположен в передней части черепа, около его рта. У китов носы расположены на верхней стороне их черепов. У Indohyus глаза расположены близко друг к другу в центре морды. У китов глаза расположены на боковых сторонах их голов. У Indohyus уши находятся на вершине черепа. У китов уши на боковых сторонах черепов. Различия ясно видны ниже.
Но что означают эти различия по сравнению с относительной толщиной медиальной и латеральной стенок барабанной кости? Насколько же это по-дурацки выглядит с нашей стороны не видеть сходства между Indohyus и китом! |
Рисунки, надо сказать, очень наглядны и показательны. Хороший
дидактический приём – облегчает познание анатомии китообразных. Но не всех,
а только современных – и в этом заключается лукавство
Do-While Johnes. Древние китообразные, однако
же, были менее специализированы к водному образу жизни, и черты сходства
между ними и индохиусом были более явственными. Но, если честно и
правдиво сравнивать с ними, пропадёт весь эффект от такого сравнения, поскольку
вместо желаемых
различий
бросится в глаза
поразительное сходство. Сравнение индохиуса и современных китов некорректно
хотя бы потому, что после разделения родословных линий китов и парнокопытных
и те, и другие приобрели большое количество новых признаков, которые заведомо
отсутствовали у общего предка (причём киты приобрели намного больше специализированных
черт, что очевидно). Ясно, что, если сравнить индохиуса (жившего приблизительно
через 2 – 4 миллиона лет после расхождения линий китов и парнокопытных) и современного
кита (живущего в наше время, то есть, спустя где-то 50 – 53 млн. лет после
того же расхождения), различий будет очень много. А сравните-ка индохиуса с
самыми ранними китообразными,
которые
были
его современниками! Не хотите? Боевой задор угас? Не хотите по-дурацки выглядеть?
То-то же… Тем не менее, вам это удалось, как видим.
Далее, некоторые из признаков, указанных как «типично китовые» для большего
эффекта от сравнения, могут проявляться не у всех китов. Так, ноздри китов
сдвинулись на темя сравнительно поздно – позже, чем, например, появился китовый
ус. То есть, на Земле существовали вполне сформировавшиеся киты (иначе их не
назвать)
с ноздрями «на полпути» между рылом и теменем. Об этом я планирую более подробно
рассказать дальше, в отдельной главе.
Глаза ранних китов не всегда смотрели только в стороны. Ambulocetus и Pakicetus имеют черепа с «нормальным» положением глазниц, характерным для типичных млекопитающих.
Ах, ну да, это же были «просто наземные животные ещё одного сотворённого рода,
имеющие лишь некоторые черты сходства с китами»…
Если внимательно рассмотреть череп индохиуса, то видно, что кости уха у него
располагаются там же, где у всех прочих млекопитающих, включая типичных китов
– на боковых частях черепа, ниже «ватерлинии», позади челюстного сустава. Просто
иллюзию высоко расположенных ушей создаёт наличие у индохиуса ушных раковин
(на рисунке, понятное дело, а не на самом черепе, где они просто не сохранились).
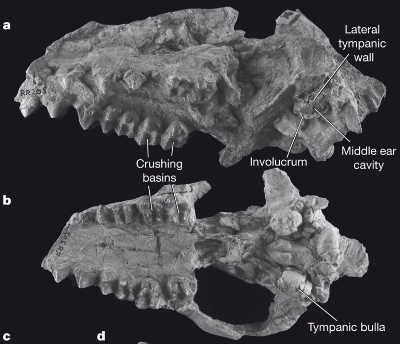
Фото черепа индохиуса. Видно, что кости среднего уха находятся примерно там же, где и у остальных млекопитающих
Ископаемые остатки состоят из черепа и нескольких кусочков костей ног. Что может заставить кого-либо поверить, что в этих костях есть что-то сходное с китом? Хорошо, вот их аргумент.
Их выводы основаны на зубах. Они говорят, что все ныне живущие и вымершие киты имеют небольшие жевательные углубления на своих зубах. Следовательно, разумно было бы предположить, что Indohyus также имел маленькие жевательные углубления, что и заставило бы считать, что Indohyus был предком китов. Но они говорят, что у него были большие жевательные углубления! Indohyus имел зубы, значительно отличающиеся от зубов, что были у китов. Но, чем заключить, что Indohyus не был китом, они говорят, что «это означает, что в процессе происхождения китообразных произошло крупное изменение в функциях зубов, возможно, связанное с изменением рациона при их возникновении». Что они в общих чертах говорят, так это то, что Indohyus должен быть предком китов, поскольку его зубы НЕ похожи на китовые, что является доказательством того, что форма китовых зубов эволюционировала! Но это намного лучше! |
Тпруууууу! Тпруууууууу! Господин хороший, остановите своего
Пегаса, или я буду вынужден его пристрелить. Никто из учёных НЕ ГОВОРИЛ, что
индохиус – предок китов. Его считают представителем
сестринской группы, то есть, группы, произошедшей
от одного с китами предка. Вы ведь не будете утверждать, что ваша сестра –
это ваша мама? Ведь она, являясь близким родственником, тем не менее, не является
предком. Индохиус не может быть предком китов хотя бы из-за своего слишком
молодого возраста (48 миллионов лет против актуальной на момент написания обзора
оценки возраста в 50 – 52 миллионов лет для Pakicetus), это отметил
ещё коллега Do-While Johnes, товарищ R. K. Bentley (см. выше). Так что налицо
явная попытка
подмены
понятий
– выдачи
«родственника»
за «предка». Зачем? Предполагаю, что для того, чтобы в сочных и цветистых выражениях
опровергнуть это умопостроение (такого фиктивного противника ласково называют
«соломенным чучелом»). Если помнить, что раоэллиды (включающие индохиуса) являются
не предками, но
лишь
очень близкими
родственниками
китообразных,
то
факт указанных различий в их зубной системе не вызывает удивления.
Возникает закономерный вопрос: может ли предположительно травоядный Indohyus,
родственник несомненно травоядных парнокопытных, быть также близким родственником
плотоядных китов? Возможна ли чисто физически такая смена рациона? Ископаемые
и современные животные дают чёткий ответ. Начать стоит, пожалуй, с того, что
зубы кондилартров (самый древний отряд копытных млекопитающих, предковый для
всех современных копытных) менее специализированы, чем зубы их потомков, перешедших
на растительный корм. Кондилартры были не только (и даже не столько) травоядными,
но также всеядными с уклоном в плотоядность (например, Arctocyon –
см. ниже рисунок его черепа с огромными клыками и заостренными предкоренными
зубами). При расхождении потомков кондилартров
от общего «корня» разные эволюционные линии
демонстрировали
разные
направления специализации. Думаю, никто не станет отрицать, что мезонихиды
были плотоядными. В отличие от них, большинство современных
копытных (лошадь или слон, например) и в рот не возьмёт мясного. Но есть и
здесь одна тонкость. В Африке и Азии и в наши дни водятся мелкие парнокопытные,
сохранившие множество примитивных черт предковой формы этого отряда – водяные
оленьки (Tragulidae). Эти зверюшки, несмотря на миловидную внешность, охотно
поедают падаль и мелких беспозвоночных, а также рыбу. В неволе африканский
водяной оленёк атакует довольно крупных птиц, если попытаться держать его совместно
с ними. И напомню, кстати, что этот зверь – ближайший экологический аналог
(не родственник!) Indohyus. Это не единственный случай плотоядности
у «настоящих», эволюционно развитых копытных. Второй пример такого рода – ныне
вымершие энтелодоны
и современные свиньи. Свинья мало специализирована к поеданию мяса, но не упустит
случая закусить мелкими животными (вплоть до случаев нападения на новорождённых
детей в эпоху Средневековья) или падалью. А вот энтелодоны с островершинными
зубами и огромными клыками – несомненно плотоядные существа.
Да
что
тут говорить,
если даже северный
олень не упустит возможности съесть яйца или птенцов белой куропатки!
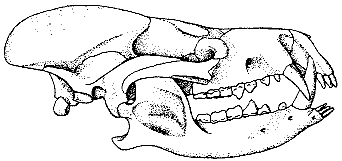
Череп кондилартра Arctocyon.
Итак, для потомка кондилартров, исходно неспециализированных
животных, отсутствует принципиальное ограничение на эволюцию в сторону плотоядности.
У одних потомков
кондилартров, эволюционировавших по пути усиления приспособления к хищному
образу жизни (мезонихиды, киты), развивались особенности строения зубов,
облегчающие умерщвление добычи и потребление мягкого мяса. У других
потомков тех же самых кондилартров,
которые специализировались на питании растениями (современные и многие
ископаемые отряды копытных), развивались особенности зубов, облегчающие обработку
неподвижного и не сопротивляющегося,
но волокнистого, жёсткого и абразивного растительного корма. Произойдя
от одного «универсального» предка, индохиус и ранние китообразные специализировались
в разных направлениях. И где тут противоречия? Всё вполне объяснимо.
Мне жаль, господин Do-While Johnes, но вашего Пегаса всё же пришлось пристрелить.
Ранка от комариного укуса на его заднице могла воспалиться и причинить
ему невероятные страдания.
Пусть для вас будет утешением то, что он не мучался.
На ясном английском, анализ содержания углерода-13 в зубах
говорит о том, что он питался скорее на суше, чем в воде. Итак, это ещё одно
свидетельство того, что это не кит. |
Оп-па, оп-па, оп-па… И куда мы так резво на крыльях своей мечты полетели? На землю не угодно ли, господин хороший? «Питался скорее на суше, чем в воде»… О водных макрофитах вроде как забыли? Или вспоминать не хотите? Для неспециалистов в области ботаники я поясню, что большинство крупных пресноводных растений, которые в обиходе называют «водорослями», на самом деле принадлежит к различным семействам цветковых растений, папоротников и мхов. Достаточно полистать хороший атлас аквариумных растений, чтобы понять это. Водные растения отличаются быстрым ростом, а по ряду особенностей физиологии и биохимии сходны не с водорослями*, а со своими наземными родственниками.
| * «Водорослями» часто называют все водные растения без разбора, но это неверно. В истинном смысле этого слова водоросли – первичноводные растения, не произошедшие от наземных предков. Они, собственно, и образуют фитопланктон. Известные со школы хлорелла и хламидомонада – это из их числа. |
Отсюда
и предположение о питании водными макрофитами. Вполне естественно, эти рассуждения
не отрицают возможности кормления Indohyus на суше – одно другому
не мешает. Ведь и современные гиппопотамы по ночам пасутся на берегу и поедают
наземную растительность, что не мешает им иметь приспособления
к водному образу жизни. Так что сценарий с животным, которое питается на
суше, но проводит значительную часть времени в воде, имеет солидную, в буквальном
смысле «стопудовую» поддержку!
Последнее замечание я целиком оставляю на совести Do-While Johnes. Как говорится,
«кто о чём, а вшивый всё о бане». Как видно из приведённой цитаты, настоящих
учёных интересует получение как можно большего количества информации из имеющихся
находок. О деньгах тут и речи не было. Впрочем, «у кого чего болит…». Наверное,
папаша Ратцингер, взойдя на папский престол, сократил финансирование разного
рода «институтов креационных исследований», или же озолотил, но просто ещё
очень хочется.
Что ж, «на
волке только
слава, а ест овец-то Савва».
Индохиус может служить хорошей моделью предка китов, но он жил слишком поздно, чтобы сам мог быть предком китообразных. Учёным давно известно животное, с полным правом претендующее на роль предка китов – это пакицетус, «кит из Пакистана».
Этот номер журнала Science до сих пор не даёт спокойно спать креационистам из-за первой реконструкции кита Pakicetus, которая, как показали более поздние находки, оказалась неправильной. Дата выхода журнала - 22 апреля 1983 года. Возмущениям противников теории эволюции нет предела до сих пор. |
До недавнего времени учёные могли противопоставить точке
зрения креационистов очень небольшое количество фактического материала. В «Палеонтологии
и эволюции
позвоночных» Р. Кэрролла как пример самой ранней формы отряда китообразных
упоминается Pakicetus («кит из Пакистана»), и в систематическом списке,
завершающем книгу, приводятся названия ещё нескольких родов ранних китообразных,
не включённых
в основной текст. На момент написания той книги никаких элементов скелета
туловища и конечностей (посткраниального, «послечерепного») скелета Pakicetus не было известно. Одна из ранних реконструкций пакицетуса, сделанная Филипом
Джинджеричем почти сразу после открытия этого животного и опубликованная
в журнале Science, изображает это животное таким, каким его ОЖИДАЛИ УВИДЕТЬ
учёные на том уровне знаний об ископаемых китообразных – тюленеобразным
существом с короткими задними лапами и мощным уплощенным хвостом. На тот момент
учёные
не располагали останками иных видов ископаемых китообразных того же времени,
поэтому реконструкция пакицетуса вполне могла оказаться неточной.
Кэрролл
(см. ниже) упоминает, что никаких остатков посткраниального скелета пакицетуса
не найдено, и в его книге приведена лишь реконструкция черепа этого животного.
Реконструкция, основанная на слишком малом количестве ископаемого материала,
стала объектом атаки креационистов, особенно после того, как был найден
достаточно полный скелет этого животного. Дон Баттен в дополнении к своей
статье “A
Whale of a Tale?” пишет о реконструкции Джинджерича:
… кажется, к этим заметкам [палеонтологической
тематики – В. П.], по сравнению с экспериментальной (операционной)
наукой, применяются различные
стандарты. Многим ложным утверждениям оказали таким образом доверие в
престижных обзорных журналах, поэтому я стал довольно скептически относиться
к этим
утверждениям. Например, рассказ Джинджерича о Pakicetus, опубликованный
в престижном журнале Science в 1983 г., был основан на некоторых фрагментах
черепа. Science даже опубликовал на первой странице обложки художественную
реконструкцию целого существа с ногами, преобразующимися в плавники,
плавающего в море и преследующего рыбу на обед. Очень показательно
сравнить его с
более современной реконструкцией, основанной на гораздо большем количестве
полного скелета – теперь ясно, что это наземное существо. |

Красивая реконструкция пакицетуса, сделанная с помощью трёхмерной графики – увы, уже устаревшая.
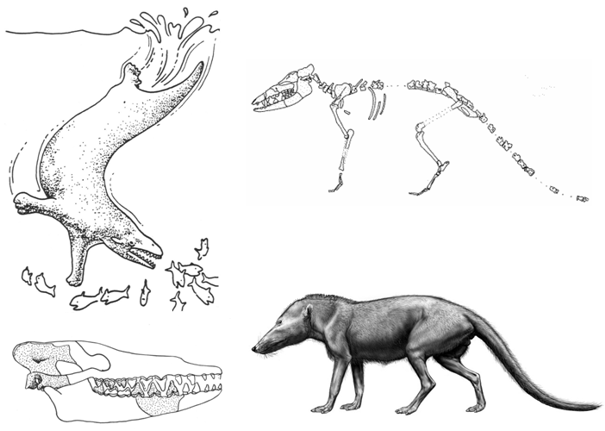
«Компромат» креационистов: первая реконструкция пакицетуса из журнала Science 1983 года; первая находка пакицетуса – реконструированный фрагментарный череп; скелет пакицетуса, найденный командой Ханса Тевиссена; реконструкция пакицетуса, выполненная Карлом Буэллом.
Вот ещё одно эссе, связанное с открытием пакицетуса: Refuting Evolution 2 за авторством Джонатана Сарфати и Майкла Мэттьюса:
Джинджерич обнаружил в Пакистане несколько фрагментов
черепа волкообразного существа, которое, предположительно, имело внутреннее
ухо, подобное китовому. Но окончательные выводы делать рано. Не было
найдено никакого посткраниального скелета, так что у нас нет никаких
идей относительно того, как он передвигался. Однако это не помешало Джинджеричу
написать статью для школьных учителей с иллюстрацией животного, которое
плескалось в море, плавало и ловило рыбу, и убедительно походило на промежуточное
звено между наземными животными и китами.
|
Оп-паньки, господа хорошие! А как вы узнали, что он «волкообразный»?
Это было выяснено уже позже, при раскопках, проведённых… Нет, не Институтом
Муди
(клянусь,
есть
и такой!), и не мормонами, и не свидетелями Иеговы, а эволюционистом Хансом Тевиссеном.
И между открытием черепа пакицетуса и его описанием в 1983 году с одной стороны,
и открытием фрагментов посткраниального скелета пакицетуса в 2001-м году с
другой стороны (тогда, собственно, и признали,
что он был отдалённо похож на волка) прошло в общей сложности больше полутора
десятилетий. Так что Джинджерича вполне можно понять: его построения не вышли
за рамки материала,
известного на тот момент. А вот Сарфати и Мэттьюсу, написавшим свою работу
сравнительно недавно, стоит ответить перед читателями за попытку откровенной
подтасовки фактов.
Новые находки, несомненно, подтвердили ошибочность
первоначальной реконструкции пакицетуса. По современным представлениям,
основанным на реальных находках, это животное
выглядит совсем не так, как представлялось раньше. Конечно, после того, как
палеонтологи сами признали и исправили свою ошибку, основанную на неполноте
исходного материала, критиковать их особенно легко.
Особенности пакицетид в свете недавних находок должны быть рассмотрены особо
из-за того места, которые они занимают в систематике млекопитающих.
Даррен Найш в статье “When whales walked the land…
and looked
like antelopes…
and
mimicked crocodiles…. and evolved trunks. What?” («Когда киты ходили по земле...
и выглядели, как антилопы... и подражали крокодилам... и отращивали хоботы.
Что?») описывает особенности пакицетид:
... Обладая множеством уникальных для китообразных
особенностей ушных костей, Pakicetus был истолкован как вероятно земноводный
хищник,
который был переходной формой между наземными, похожими на волка мезонихиями,
вероятными предками китов (или как это считалось в то время), и больее
крупными и молодыми археоцетами типа базилозаврид. Кости уха Pakicetus демонстрируют уникальное промежуточное состояние между таковыми у наземных
млекопитающих и продвинутых китообразных (Gingerich et al. 1983) и, в
сочетании с его присутствием в мелководных пресноводных местообитаниях,
позволяют предположить, что это было еще не полностью морское животное.
На основании только имеющегося фрагментарного черепа Pakicetus предполагался
как тюленеобразное существо, и соответствующие реконструкции прижизненного
облика изображают его таким образом. Мы теперь знаем, что эти реконструкции
чудовищно неточны, как сочленённые скелеты пакицетид показывают, что
эти млекопитающие внешне не отличались от ранних парнокопытных-«заморышей»
типа оленьков, маленьких оленей и эоценовых примитивных форм (Thewissen
et al. 2001). Фактически мы также теперь знаем, что у пакицетид была
таранная кость с двойным блоком, считавшаяся уникальной для парнокопытных;
так, теперь кажется, что парнокопытные и китообразные были ближе друг
к другу, чем китообразные
к мезонихидам.
|
В журнале “Nature” от 20 сентября 2001 года, в той самой статье “Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls” палеонтологи Ханс Тевиссен и коллеги сообщают об изучении анатомии нескольких видов примитивных животных, относящихся к семейству пакицетид (Pakicetidae):
Есть три рода пакицетидных китообразных: Pakicetus,
Nalacetus и Ichthyolestes. Pakicetus самый крупный, за ним следует Nalacetus (приблизительно на 5% меньше по линейным размерам), и Ichthyolestes (приблизительно
на 29% мельче). До сих пор для пакицетид были описаны только зубы, челюсти
и один череп. Мы откопали четыре частичных черепа, два из которых сохраняют
орбитальную область, несколько фрагментов морды, и приблизительно 150
изолированных посткраниальных костей пакицетид от многих особей. Они
были найдены на одном участке раннеэоценовой формации Кулдана в Пакистане. |
К слову, эта статья широко цитируется критиками теории происхождения китообразных
от наземных животных, и к ней придётся обращаться ещё не раз.
В книге Кэрролла «Палеонтология и эволюция позвоночных» (том 3 русского издания)
приводятся следующие данные о пакицетусе:
Древнейшие киты описаны из самого конца раннего
эоцена Пакистана (Gingerich, Russell, 1981; Gingerich et al., 1983).
Сохранившиеся остатки черепа говорят о строении, промежуточном между
хорошо изученными позднеэоценовыми китами и мезонихидами. Часть фрагментарного
материала сначала относили именно к последней группе, но найденные впоследствии
части показывают, что основание черепа, несомненно, принадлежит китообразному.
Наиболее полные остатки относятся к роду Pakicetus (…). Череп сохранился
не целиком, но длина его составляла, должно быть, 30 – 35 см. Мозговая
коробка очень узкая, с высоким сагиттальным гребнем и ламбдовидным гребнем,
очень напоминающим аналогичное образование у позднеэоценовых китов.
Зубной аппарат и вытянутые челюсти почти такие же, как у хорошо изученных верхнеэоценовых форм. Как и у последних, слуховая булла массивная и образована наружной барабанной костью. У более продвинутых китов ушная область существенно преобразована в связи с развитием дирекционального подводного слуха и для защиты при нырянии. У Pakicetus эти изменения еще не отмечены. Слуховые буллы у современных китов изолированы пазухами, так что звук может восприниматься каждой стороной черепа независимо. У Pakicetus булла все еще соединена с чешуйчатой, основной затылочной и боковой затылочной костями, тогда как у ныне живущих китов – только с околоушной костью. Наличие у Pakicetus хорошо развитой ямки для напрягающей мышцы барабанной перепонки (tensor tympani) говорит о том, что у него сохранялась функция этой перепонки, утраченной продвинутыми китами. Ничто не свидетельствует о васкуляризации среднего уха, позволяющей поддерживать в нем давление при нырянии. Остатки Pakicetus были найдены вместе с костями наземных млекопитающих, т. е. эта форма по крайней мере часть своей жизни проводила на суше. К сожалению, пока ни один из известных фрагментов посткраниального скелета не удается отнести к этому роду. Джинджерич с соавторами предполагают, что он вел земноводный образ жизни и проводил много времени в воде, охотясь за рыбой, но для отдыха выходил на сушу. Премоляры Pakicetus представляют собой пильчатые треугольные пластинки, внешне напоминающие зубы крупных акул. Моляры сохраняют основное мезонихидное расположение бугров. У верхних моляров три корня, хорошо выраженные протокон, паракон и метакон; у нижних, как у мезонихид, сохраняются отчетливые протоконид и гипоконид. |
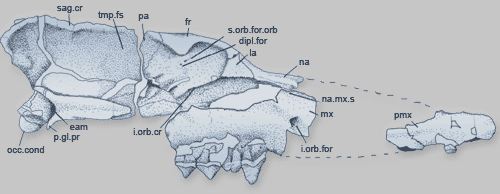
Фрагментарный череп Pakicetus (c сайта Э. Бабински).
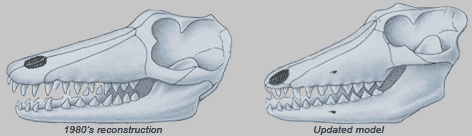
Уточнённая форма черепа Pakicetus (c сайта Э. Бабински).
Благодаря новейшим находкам мнение Джинджерича, высказанное
в 80-х гг. ХХ века, оказалось неверным, а пакицетус обрёл своё «истинное
лицо»: он был длинноногим и длиннохвостым
млекопитающим размером
с волка. У него была довольно крупная и массивная голова с суженным
рылом и остроконечными зубами, немного похожими на зубы акулы. Ставшая классикой
реконструкция этого животного, выполненная Карлом Буэллом, приведена выше.
Скелет, найденный Тевиссеном и коллегами, относительно неполный, но позволяет
сделать
выводы
о том, как выглядело это животное. Вот их мнение из статьи в журнале “Nature”:
Пакицетиды не демонстрируют ни одной из этих [характерных
для водных китообразных – В. П.] особенностей. Шейные позвонки пакицетид
более длинные, чем у позднеэоценовых китов (…), и позвонки туловища увеличиваются
в размере от предыдущего к последующему (…), как у наземных млекопитающих.
Поясничные и хвостовые позвонки (…) длинные по сравнению с таковыми у
современных китообразных, имеющих хвостовой плавник, но не такие длинные,
как у вымерших китообразных, которые плавали, волнообразно изгибаясь
всем позвоночником (например, ремингтоноцетид Kutchicetus).
У Ambulocetus и Kutchicetus мускулистый и гибкий
поясничный отдел позвоночника, тогда как движения у пакицетид ограничены
как результат их отогнутых зигапофизов
(…), черты, общей с бегунами, имеющими жёсткую спину, типа мезонихид
и многих вымерших и современных парнокопытных. Крестец пакицетид состоит
из четырех жёстко соединённых позвонков, и имеется прочное крестцово-подвздошное
соединение, как у наземных млекопитающих и земноводных китов типа Ambulocetus и Kutchicetus,
но не как у более поздних китообразных». (ссылки, приведённые в статье,
пропущены – В. П.)
|

Скелет Pakicetus attocki. Обратите внимание на череп обтекаемой формы, остроконечные зубы и широкие ступни.
Реконструкция Pakicetus, сделанная Карлом Буэллом на основе найденного Тевиссеном
скелета, заставила креационистов обрушиться на сторонников теории эволюции
со шквалом критики.
Пожалуй, наиболее активно проявил себя на этом поприще турецкий креационист
Харун Яхья, чьи работы сейчас активно продвигаются на российский книжный рынок.
В своей статье “Whale Fantasy of National Geographic” (ниже цитируются выдержки
именно из неё) он заявил следующее:
Ископаемые остатки вымершего млекопитающего Pakicetus
inachus, по которым ему дали это название, оказались в центре внимания
в 1983 г. P. D. Gingerich и его помощники, нашедшие это ископаемое, без
всяких колебаний заявили, что это был «примитивный кит», хотя они фактически
нашли только череп.
Другими словами, основываясь на некоторых деталях в его зубах и ушных костях, ‘National Geographic’ посчитал возможным описать этого четвероногого, похожего на волка наземного жителя как «ходячего кита». Только один взгляд на реконструкцию Pakicetus эволюциониста-иллюстратора Карла Буэлла показывает нелепость в наименовании этого животного «ходячим китом». |
В тексте ясно виден излюбленный приём креационистов: сказать,
что, мол, той или этой особенности мало, чтобы считать животное китом (лошадью,
обезьяной,
человеком, и т. д.). На самом деле, строение зубов – это один из самых информативных
признаков для палеонтолога и зоолога, и ошибки здесь достаточно редки. По строению
зуба ёж легко может быть отличим от суслика или летучей мыши, а человекообразную
обезьяну
сложно
спутать
с низшей узконосой или широконосой. Главное – чтобы этот зуб попал в руки опытного
и знающего человека. Именно по зубам кит Basilosaurus был в своё время опознан
как млекопитающее – вначале его описали как гигантскую ящерицу, и память об
этом казусе навсегда осталась в его названии, означающем «царственная ящерица».
Чтобы развеять сомнения в пригодности «зубного» метода определения родства
животных, достаточно просто заглянуть в хороший и подробный определитель млекопитающих.
В нём описано большое количество признаков, относящихся к строению зубов, и
позволяющих точнейшим образом определить вид животного. В случае с пакицетидами
к особенностям строения зубов добавляются ещё особенности черепа, что делает
идентификацию животного более точной. Рассмотрение скелетных признаков наряду
со строением зубов необходимо, поскольку всякое животное само по себе – целостный
организм. И, кстати,
именно
для
случая
с
определением
родственных отношений китообразных
известен
пример явного параллелизма в эволюции зубов между ранними китами и мезонихидами.
Если подумать над тем, что написано в журнале, и как это подал Харун Яхья, можно
сильно удивиться. Ещё раз внимательно читаем выдержку из ‘National Geographic’:
«комбинация мельчайших особенностей ... отсутствующая у других
наземных млекопитающих, но имеющаяся в наличии у более поздних
эоценовых китов». То есть,
за основу «возмутительной» классификации пакицетуса взят, фактически, общий,
характерный
для
древних китов
признак! Ну? Чего тут странного и непонятного?
Харун Яхья продолжает:
Палеонтологи полагают, что Pakicetus был четвероногим
млекопитающим. Строение скелета, показанное слева, опубликованное в журнале
Nature, ясно показывает это. Поэтому реконструкция Pakicetus (…), сделанная
Карлом Буэллом, который основывался на этом строении, является реалистичной. ‘National Geographic’, однако, выбрал для использования картину «плывущего» Pakicetus (…), чтобы изобразить животное как «ходячего кита» и закрепить этот образ у своих читателей. Несостоятельность в этой картине, предназначенной, чтобы заставить Pakicetus казаться более «китообразным», совершенно очевидна: животное изображено в «плавающем» положении. Его задние ноги показаны вытянутыми назад, и им придано подобие плавников. |

«Та самая» иллюстрация плывущего пакицетуса из ‘National Geographic’
Не будем скатываться на обсуждение художественных достоинств
того или иного рисунка. В противном случае придётся обсудить также коллаж с
главной страницы
сайта “Answers in Genesis”, где на привольных зелёных лугах пасутся рядом антилопы
и диплодоки. Нравится Харуну Яхье рисунок пакицетуса, или не нравится – это
его личное дело. Но отказать пакицетусу в способности плавать только на основании
его «волкообразной» формы тела он не вправе: лошади, например, тоже живут на
открытых ландшафтах с твёрдой почвой, а не в болоте, однако же, при необходимости
умеют неплохо плавать. Кроме того, есть в наше время на Земле удивительная
троица
животных.
Это парнокопытное
млекопитающее водяной оленёк (Hyemoschus), обитающий в Африке, певчая птица
оляпка (Cinclus) из Северного полушария, и евразийский водяной паук, или серебрянка
(Argyroneta). У этих животных есть общий признак: они живут в воде, умеют плавать
и нырять, будучи практически лишёнными специфических анатомических признаков
специализации к водному образу жизни. Иными словами, если рассматривать их
анатомию в отрыве от знания об их среде обитания, можно просто не догадаться,
что это водяные животные – об этом в отношении оляпки упоминал ещё Чарльз Дарвин
в «Происхождении видов…». Поэтому вполне возможно, что пакицетус был таким
же существом – водным по поведению, но пока не по строению. По крайней мере,
его остатки приурочены к речным отложениям, о чём говорится выше и далее.
В статье Ленни Фланка «Амбулоцетус как переходная форма китов» (Ambulocetus
as Whale Transitional) рассматривается мнение ещё одного «титана» современного
креационизма,
Дуэйна Гиша:
Первоначальная критика Гишем Pakicetus направлена на среду его обитания. После замечания, что киты являются морскими существами (нет никаких современных пресноводных китов), Гиш продолжает:
Оставив в стороне вопрос, как отложения, в которых находятся «улитки,
рыбы (в частности, сомы), черепахи и крокодилы», могут рассматриваться
как «наземные»
и «континентальные» (все эти животные преимущественно водные), здесь
мы можем отметить, что Гиш прибегает к типичной креационистской тактике
создания
«соломенного чучела» и затем нападения на эволюционную теорию, потому
что она не соответствует его «соломенному чучелу». Аргумент, что любой
предок
китообразных «как ожидалось бы», будет найден в морских отложениях
– целиком собственное построение Гиша. Вообще, нет никакой причины
ожидать, что самые
ранние предки морских китообразных должны обязательно быть морскими
существами, и есть множество причин предположить иначе. Начиная с самых
ранних ископаемых
остатков, которые мы знаем, китообразные не демонстрируют ни одну из
специализированных адаптаций, которыми обладают современные киты для
жизни в открытом море
– васкуляризированные уши, чтобы выравнивать давление при нырянии,
направленный подводный слух, горизонтальные хвостовые лопасти для лучшего
плавания –
и нет никакой причины предполагать, что предки китов должны были жить
в открытом море, пока они развивали эти адаптации. |
Как переводчик этого отрывка, добавлю от себя: пресноводные
китообразные существуют, и список их видов довольно внушителен. Естественно,
в него не попали огромные
«истинные» киты, которых люди представляют себе при слове «кит». Но дельфин
– это тоже китообразное, и речных видов дельфинов существует достаточно много
– часть морских дельфинов регулярно заходит в пресную воду, и даже может встречаться
преимущественно в реках. Это особенно часто происходит в тропиках. Настоящие
дельфины (семейства Delphinidae) встречаются в крупнейших реках мира – Амазонке,
Заире (Конго), Янцзы. Белуха регулярно заходит в реки бассейна Северного Ледовитого
океана, а также в реку Святого Лаврентия в Северной Америке, и высоко поднимается
по ним. Кроме того, примитивнейшие современные китообразные – это особое семейство
речных
дельфинов
(Platanistidae),
включающее,
по разным взглядам, 4 – 6 видов (в настоящее время исследователи
делят их на 4 семейства, о чём я подробнее скажу ниже), и обитающее почти исключительно
в пресной воде. Только один
вид
этого семейства,
лаплатский
дельфин (Pontoporia blainvillei), выходит в море.
Сомы – верный признак того, что отложения были пресноводными: в настоящее время
только немногие виды сомов из двух семейств (ариевые и угрехвостые) живут в
морях. Прочие сомы обитают в пресных водах, и даже в указанных «морских» семействах
сомов немало пресноводных видов.
Кроме того, в оригинальном высказывании Гиша черепахи названы словом “turtles”,
которое в английском языке чаще всего обозначает водных черепах (противопоставляется
слову “tortoises”, обозначающему наземных черепах).
И ещё, упоминание о районе, богатом железной рудой, наводит на воспоминание
о возможности образования железных руд в болотистой среде (так называемые «болотные
руды», известные едва ли не со Средневековья). А это также косвенно указывает
на характер местообитаний пакицетуса. Другое дело, что пресноводные отложения
по определению не могут быть океаническими, и действительно формируются на
континентах, либо на шельфе (например, выносы рек). Интересное дополнение к
информации о среде обитания Pakicetus можно найти на сайте Evidence
for Evolution:
... Хотя фауна млекопитающих, найденная с Pakicetus, состоит из грызунов, летучих мышей, различных парнокопытных, непарнокопытных и хоботных, и даже примата (Gingerich и др., 1983), там также есть водные животные, такие, как улитки, рыбы, водяные черепахи и крокодилы. Более того, осадки, ассоциированные с Pakicetus, демонстрируют свидетельство наличия течения, обычно ассоциируемое с почвами, которые уносит вода. Таким образом, свидетельства палео-окружающей среды ясно показывают, что Pakicetus жил в низинных увлажнённых наземных местообитаниях, совершая нерегулярные заходы в пресную воду. Интересно, что и молочные, и постоянные зубы животного находят в этих отложениях примерно с одинаковой частотой, что поддерживает идею о том, что Pakicetus производил потомство на суше. |
Пакицетус задал нелёгкую задачу не только креационистам, но и эволюционистам: классифицировать это животное и определить его ближайших родственников оказалось довольно сложно. Этим затруднением не преминули воспользоваться креационисты. Харун Яхья продолжает:
Особенности деталей, обсужденных в ‘National Geographic’, «расположение ямок на коренных зубах, складка в костях среднего уха, и расположение ушных костей в пределах черепа» не являются никаким неопровержимым свидетельством, чтобы строить связь между Pakicetus и китом: Как неявно упомянули в статье ‘National Geographic’, описывая «комбинацию
мельчайших особенностей», некоторые из этих особенностей фактически найдены
также у наземных животных. |
Либо Харун Яхья преувеличивает собственную эрудицию, либо
вообще отказывает палеонтологам в профессионализме. Во всяком случае, сумчатых
от плацентарных
отличает не только сумка на животе, но и ряд иных признаков в строении скелета
и мягких тканей тела. Например, южноамериканский Thylacosmilus отнесён
специалистами к числу сумчатых млекопитающих, а
северо- и южноамериканский
Smilodon – к плацентарным (кошачьим), несмотря на их внешнее сходство, явно
хищнический образ жизни и наличие похожих сабельных клыков. Тем более, оба
они вымерли задолго до появления «хитрых и ушлых» эволюционистов, и, если судить
по логике Харуна Яхьи, унесли с собой в могилу ключи от своего происхождения.
Но всё же и их родственные связи «раскусили», а животных классифицировали как
представителей разных отрядов. Точно так же гигантский Diprotodon не стал родственником
ни бегемоту (из-за размеров), ни грызунам (из-за формы резцов), и Zygomaturus,
внешне похожий на носорога, также остался среди сумчатых. Вообще, отношение
западных креационистов к сумчатым млекопитающим заслуживает отдельного
рассказа, и рассуждениям про них тут не место.
Трудно сказать, в каком контексте Гоулд высказал приведённую фразу. Креационисты
очень любят выдёргивать отдельные фразы из контекста, и «козырять» ими. Та
же фраза Дарвина про глаз и его происхождение стала «классикой жанра» в этом
плане. Но, находясь на своём месте, в его книге, она не выглядит «разрушительной»
для теории эволюции. Про другое китообразное с ногами, амбулоцетуса (о нём
будет рассказано далее), Гоулд выразился настолько восторженно и эмоционально,
что слова Харуна Яхьи выглядят
на этом фоне просто грязным пасквилем:
Как заключает Стивен Гоулд, «если бы вы дали мне
чистый лист бумаги и чек без обозначения суммы, я бы никак не смог нарисовать
вам теоретическое промежуточное звено лучшее или более убедительное,
чем Ambulocetus. Те догматики, кто словесным обманом может делать
белое черным, и черное белым, никогда не будут убеждены ни в чем, но
Ambulocetus
– то самое животное, которое они объявили теоретически невозможным» (Gould,
Natural History, May 1994, p. 14) (Цитируется по статье Ленни Фланка
«Амбулоцетус как переходная форма» - В. П.)
|
Короче говоря, описание Pakicetus, который со всей очевидностью является обитателем суши, как «ходячего кита» просто по структурным особенностям его ушных костей и коренных зубов, является ничем иным, как ещё одним примером традиции ‘National Geographic’ «сенсационного, необоснованного бульварного журнализма» В своей статье «Распродажа эволюции китообразных» креационный автор Ashby L. Camp показывает полную неверность утверждения, что класс мезонихид [так в оригинале – Mesonychid class – В. П.], который должен включать наземных млекопитающих вроде Pakicetus, мог быть предками археоцетов, или вымерших китов, такими словами: «Причина того, что эволюционисты уверены в том, что мезонихиды породили археоцетов, несмотря на неспособность идентифицировать любой вид по фактическому происхождению, состоит в том, что мезонихиды и археоцеты имеют некоторое сходство. Эти черты подобия, однако, не достаточны, чтобы представить это как случай преемственности, особенно в свете обширных различий. Субъективный характер таких сравнений очевиден в силу того факта, что так много групп млекопитающих, и даже рептилии были предложены как предки китов». |
В упомянутой статье «Распродажа эволюции китообразных» сказано следующее:
Первая претензия в эволюционистском сценарии – то, что археоцеты произошли от вида мезонихид. Предковый статус Mesonychidae впервые предложил Лей Ван Вален в 1966 году на основании некоторого сходства зубов между мезонихидом Dissacus navajovius (который известен как Dissacus carnifex у Коупа) и некоторыми образцами археоцетов. Его весьма осторожное предположение этой претензии стоит напомнить:
В более обширном анализе, опубликованном три года спустя, Фредерик Салаи предположил, что и гапалодектины (которые тогда считались подсемейством мезонихид), и археоцеты, возможно, «произошли или от ранне-, или от среднепалеоценовых мезонихид, более примитивного вида, чем известные мезонихины» [выделено мной]. Другими словами, Салаи заключил, что и Dissacus, и Ankalagon, единственные среднепалеоценовые мезонихиды, известные в то время, были слишком продвинутыми (эволюционно развитыми), чтобы находиться в ряду археоцетов. Он видел в них «сестринскую группу» археоцетов, но не действительных предков. |
Многочисленные гипотезы относительно происхождения китообразных
– пример вполне обычной ситуации в науке. У креационистов правильный ответ
уже заранее имеется.
Все усилия «креационной науки» сводятся к поиску решения, удовлетворяющего
этому ответу. Приём вполне знакомый со школы, верно? В отличие от креационизма,
в науке окончательный ответ неизвестен, и его нужно найти. Выводы зависят
от полноты информации, которая имеется в наличии, и считаются верными до появления
объективной информации, которая противоречит данным выводам. Поэтому вполне
естественно, что при появлении новой информации происходит пересмотр старых
взглядов. Например, та же информация о происхождении китов от мезонихид,
актуальная
в момент написания статьи Эшби Л. Кампа (1998 год), уже устарела в свете
новых находок китообразных и проведённых анатомических исследований. А в свете
данных
о родстве китообразных, парнокопытных и мезонихид, установленных в ходе исследований
Ханса Тевиссена и коллег, оказывается, что Фредерик Салаи был прав, придавая
мезонихидам указанный статус.
Единственное, что остаётся туманным – «назначение» рептилий на роль предков
китов. Мне, автору этих строк, такая гипотеза ни разу не встречалась ни в
отечественных, ни в переводных источниках. Интересно, в каком году она была
высказана?
В статье Даррена Найша не было рассказано о китах с хоботом,
хотя упоминалось, что они известны. Информацию об этом животном можно найти
в Интернете: вот
ссылка на статью в формате PDF. Это существо получило очень примечательное
название Makaracetus bidens. В названии этого кита символически сочетаются
названия «Makara» – имя сказочного существа, наполовину млекопитающего,
наполовину рыбы (да, да, козерог - это та самая макара и есть!) и «cetus»
– кит (лат.). Одна из сказочных разновидностей макары как раз изображалась
как рыба с головой слона. Остатки макароцетуса были найдены в Восточном Балухистане
(Пакистан) в 2004 году, а описание сделали в 2005 году Филип Джинджерич, Ийяд
С. Залмут, Мунир Уль-Хак, М. Акрам Бхатти.
Остатки макарацетуса найдены в верхних слоях формации Доманда, в отложениях,
сформировавшихся в прибрежной зоне моря Тетис.
От этого кита (образец получил номер GSP-UM 3570) найдены фрагменты черепа,
части двух шейных позвонков, пять грудных позвонков, два крестцовых позвонка.
Все найденные фрагменты – части скелета одной особи.
Череп макарацетуса отличается сравнительно коротким рылом, сильно сжатым с
боков и слегка загнутым вниз. В каждой предчелюстной кости находится по два
резца (всего четыре), что меньше, чем у родственных видов. Отсюда видовой эпитет bidens –
«двузубый». Отверстия ноздрей очень большие (у найденного образца их ширина
составила 48 мм), сдвинуты на верхнюю поверхность рыла; носовое отверстие одно,
но несомненно, что ноздри всё же разделялись хрящевой перегородкой. На боковых
поверхностях черепа имеются ямки для прикрепления очень сильных лицевых мускулов.
В оригинальном описании сказано: «Makaracetus, несомненно, имел короткий
мускулистый хоботок, возможно, для специализированного питания моллюсками или
иной бентосной фауной».
При обсуждении находки упоминается, что остатки моллюсков очень обычны в формации
Доманда.
Череп Makaracetus. Серой штриховкой показаны сохранившиеся части.
A – вид сверху. Обратите внимание на большое отверстие ноздрей.
B – вид сбоку справа. Заметен характерный профиль и обширные углубления (на рисунке обозначены как LFP и LFM), к которым прикреплялись лицевые мышцы .
C – вид снизу. Заметно, насколько сильно сжато с боков рыло этого животного. Pal (palate) – нёбо.
D – срез через носовые кости, показывающий глубину ямок для прикрепления мышц.
Макарацетус пока не вовлечён в ожесточённые баталии креационистов и эволюционистов. Надолго ли?
Прежде, чем рассматривать следующего представителя архаичных китообразных, стоит сделать небольшое отступление и поговорить о различных взглядах на происхождение китов. Эта тема – предмет яростных нападок со стороны креационистов, поскольку дать послабление в таком вопросе равнозначна сдаче позиций ещё в одном ключевом вопросе.
В начале
я должен сделать одну оговорку. В приводимых отрывках из различных статей можно
заметить
разделение взглядов
авторов на две группы. Одни из них поддерживают теорию происхождения китов
от мезонихид, другие сближают их с парнокопытными. Это разногласие связано
с недавними исследованиями Ханса Тевиссена и его коллег. До их открытия
и исследования скелетов примитивных пакицетид, пока был плохо известен посткраниальный
(«послечерепный») скелет ранних китов, господствовала теория о происхождении
китов от мезонихид (Mesonychia) – особой группы плотоядных примитивных
копытных.
В пользу этого говорило сходство строения зубов примитивных китообразных
и мезонихид. Исследования Тевиссена показали, что в строении конечностей
примитивных китообразных и столь же примитивных парнокопытных присутствовали
общие уникальные черты (в частности, строение таранной кости). Своеобразным
подтверждением его исследований служит сходство биохимии современных китов
и парнокопытных. Насколько «парнокопытная» гипотеза противоречит «мезонихидной»,
пока неясно: у давно вымерших мезонихид нельзя взять образец ткани для
биохимических исследований.
Естественно, что открытие пакицетуса и множества других примитивных «китов
с ногами» несколько охладило восторженный пыл креационистов, преподносивших
китов как «очевидный пример сотворённых животных» из-за сильного отличия
их строения от строения любых наземных млекопитающих. Поэтому в ход пошла
более
тонкая политика: необходимо любыми силами «вывести» пакицетид из числа китообразных.
У креационистов есть один такой способ «низвести» пакицетуса. Это популярный
приём игры с объёмом систематических групп, на которые делятся живые организмы.
Когда вид обладает сочетанием черт, характеризующих ранее выделенные отдельные
систематические группы, его точное отнесение к одной из них является затруднительным.
Так происходит, например, в случае с ранними копытными кайнозоя.
Роберт Кэрролл в «Палеонтологии и эволюции позвоночных» (гл. 22) пишет:
«Наши взгляды на эволюцию очень сильно зависят от
таксономических подходов. В основе линнеевской систематики лежит иерархическая
система. Виды объединяются в роды, роды в семейства, семейства – в отряды
и т. д. В результате создается впечатление, что различия видов, входящих
в разные таксономические группы, пропорциональны рангу этих групп. Другими
словами, виды разных семейств должны различаться сильнее, чем виды разных
родов одного семейства, виды разных отрядов – сильнее, чем виды разных
семейств одного отряда, и т.д. Подобное впечатление может оказаться и
верным, если рассматривать только современные формы, но когда речь идет
о начальных этапах радиации крупных таксонов, это явно не так. Чем
больше внимания уделяется вертикальной, или кладистической, классификации
и
чем полнее становится палеонтологическая летопись, тем яснее возможность
отнесения отдельных популяций внутри одного вида к разным родам и даже
семействам или отрядам производных форм. Среди кайнозойских млекопитающих
уже известны случаи, когда подобное разделение предкового вида вполне
оправдано. Уилсон (Wilson, 1971) описал прогрессивную эволюцию примитивных
селенодонтных парнокопытных, у которых популяции одного раннего вида
дали представителей разных родов, а затем и семейств.
Относительно полно документирована крупномасштабная радиация копытных в самом конце мела и начале палеоцена. Тогда из единственного семейства древних кондилартр всего за 2 – 3 млн. лет дифференцировались 16 самостоятельных отрядов (Van Valen, 1978). До сих пор об этих животных, если не считать их зубных систем, известно мало, однако предложенная схема радиации не противоречит гораздо более полным остаткам скелетов из позднейших кайнозойских отложений. С точки зрения изменчивости и дифференциации картина, наблюдаемая на ископаемом материале этих предковых групп, почти не отличается от схемы радиации в кайнозое представителей множества других семейств. Наиболее существенно здесь то, что многие виды и роды на следующих этапах радиации признавались представителями разных семейств и отрядов. На уровне особей и видов эволюция ранних кондилартр выглядит не особенно отличающейся от типичного микроэволюционного процесса. Различия заключаются не в самих видах, а возможностях, реализованных их потомками. Ван Вален обратил внимание на то, что скорость возникновения новых таксонов была здесь существенно выше, чем в среднем для всего кайнозоя, однако эта особенность может быть обусловлена очень малым числом зарождающихся филетических линий и практически полным отсутствием конкуренции. Последующая радиация перспективных отрядов копытных, например парнокопытных и непарнокопытных Северной Америки и нотоунгулат Южной Америки, протекала в общих чертах по одной и той же схеме (Krishtalka, Stucky, 1986; Radinsky, 1966; Simpson, 1948, 1967). Признаки, ранее сохранявшие внутривидовую изменчивость, стабилизировались и усиливались, признаваясь в итоге отличительными чертами семейств и отрядов. Если рассматривать эволюционные события на популяционном и видовом уровне, особых феноменов, с которыми можно было бы связать возникновение родов или других таксонов высшего ранга, не обнаружится. Процессы, ведущие к дифференциации, одинаковы независимо от того, обречен дочерний вид на вымирание или даст начало новому отряду. Таксономические группы более высокого ранга можно выделить лишь по прошествии времени, когда ряд последующих этапов дифференциации приведет к появлению множества производных таксонов. Поиск особых факторов эволюции следовало бы сосредоточить на условиях, облегчающих последующую успешную радиацию группы, а не на моменте образования нового вида.» [выделено мной – В. П.] |
Креационисты пытаются представить систематику как нечто
жёсткое, неделимое и несливаемое. Возможно, это связано с понятием «барамина»,
которое они любят
использовать
в своих работах. «Барамин» - это термин, означающий буквально «сотворённый
род». «Барамином» считается группа живых организмов, произошедшая от одного
ранее «сотворённого»
предка и по определению не имеющая переходных форм к другому такому же «барамину».
Деление на «барамины» больше напоминает упомянутую Кэрроллом линнеевскую
систематику – это как изучение поперечного среза целой кроны дерева. Но кладистическая
классификация
привносит в этот «плоский мир» новое измерение – время, а точнее сказать,
динамику изменения отношений между разными систематическими группами во времени.
Для
большинства креационистов мир остаётся «плоским», уложенным в несколько тысяч
лет, которые
отпущены ему библейской версией творения (хорошо хоть, что они признают шарообразность
самой Земли). И в этом состоит тупик в рассуждениях креационистов: когда они
сталкиваются с формой,
«не влезающей»
в привычные рамки «бараминов» (чаще всего, с ископаемым существом), в их
стане начинаются судорожные метания, граничащие с попытками выдать желаемое
за действительное.
Таким существом, в частности, для них оказался пакицетус – странное существо
с головой, несущей черты, очень характерные для кита, и телом, типичным для
потомка кондилартров.
Вот, что говорится в статье Ленни Фланка «Амбулоцетус как переходная форма»:
«… Гиш играет словами, подобно тому, как он делал
это в аргументах типа «Археоптерикс – это всего лишь птица». Система
классификации, которую используют биологи, которая помещает все организмы
в категории, состоящие из царств, типов, классов, отрядов, семейств,
родов и видов, делает затруднительным подтверждение переходного характера
многих видов, которые обладают характеристиками двух весьма различных
категорий, так как каждый организм должен быть помещен в ту или иную
ранее выделенную категорию, и никакие категории не существуют для чего-то
«полу-того и полу-этого». Как поясняет Л. Беверли Холстед, «у многих
групп мы наблюдаем то, что мы называем мозаичной эволюцией; начинаясь
с животных, которые являются полноценными рептилиями, она заканчивается
формами, которые являются полностью птицами или млекопитающими. Целостная
картина постепенно изменяется, когда развиваются более птицеподобные
или звероподобные черты. Что мы должны делать, поскольку переход столь
постепенный, так это обозначить границу разделения: если некто имеет
признак X, мы назовем его A, если нет, мы назовем его B. Следовательно,
по определению никогда не может быть промежуточного звена, потому что
мы проводили разделительные линии таким образом, что животное вынуждено
быть или одним объектом, или другим». (Montagu, 1984, p. 253)
Гиш взял на себя ответственность утверждать, что киты (даже древние киты) не имеют ног, и поэтому некто, имеющий ноги, должен быть назван как-то иначе, нежели китом, независимо от того, сколь много других признаков кита он может иметь. В действительности не имеет никакого значения, классифицируем ли мы Ambulocetus или Pakicetus как мезонихида или археоцета, так как они – часть непрерывного ряда, который протягивается от одной группы к другой. Можно так же точно изображать Ambulocetus как продвинутого мезонихида со многими особенностями китообразных, или как примитивное китообразное с многими особенностями мезонихид (так же можно одинаково хорошо классифицировать Archaeopteryx как динозавра с птицеподобными чертами или птицу с динозавроподобными чертами – никакая классификация не уничтожит её переходный характер)». [выделено мной – В. П.] |
Иными словами, «перебрасывание» пакицетуса из одного отряда в другой никаким
образом не сможет повлиять на комплекс признаков, которые связывают его
с поздними китообразными. Кто сказал «не связывают»? Кто хочет это утверждать,
пусть докажет,
что «китообразные» черты у «настоящих» китообразных развились отдельно
от
пакицетуса, независимо повторив особенности, характерные для его строения.
Особо одарённых
прошу даже посчитать вероятность этого события, и, возможно, даже сравнить
её с вероятностью самосборки живой клетки из неживого. Или с самосборкой
«Боинга» при урагане на свалке – тоже неплохой пример, которым часто «козыряют»
креационисты.
Остаётся главный вопрос: кому из современных животных китообразные приходятся
наиболее близкими родственниками?
Крупный отечественный знаток китообразных А. Г. Томилин в книге «В мире
китов и дельфинов» (вышла в свет в 1974 году) вообще не упоминает конкретных
предков
китообразных, говоря об их эволюции в самых общих чертах. Пакицетус, напомню,
не был открыт в то время (его череп был найден только в 1980-м году).
Книга «Фауна мира. Звери» под редакцией В. Е. Соколова, вышедшая в 1990
году, также демонстрирует старую точку зрения на этот вопрос, полную неопределённости
(стр. 124):
Предполагают, что представители этого отряда произошли от креодонтов или меловых насекомоядных, или копытных. Некоторые учёные предполагают дифилетическое происхождение отряда; эволюция усатых и зубатых китов, по их мнению, шла путём конвергенции, а не дивергенции, и, следовательно, правильнее считать эти подотряды отрядами. |
От этих слов до открытия амбулоцетуса (о котором речь пойдёт
дальше) оставалось четыре года…
Цитата из работы Ленни Фланка возвращает нас к господствовавшей ранее
гипотезе о происхождении китообразных от особой группы потомков кондилартров
– копытных
хищников мезонихид. Но эта гипотеза долгое время противоречила данным
биохимии, свидетельствовавшим о близости китообразных к парнокопытным.
Естественно,
что проверить биохимическое родство китообразных и мезонихид при нынешнем
уровне
развития техники и технологии вряд ли удастся. Но это даже не нужно.
В настоящее время, благодаря открытию Хансом Тевиссеном более полных
скелетов
пакицетид
появилась ещё одна теория, подтверждённая ископаемыми остатками – о
родстве китообразных
и парнокопытных. Анализ морфологии конечностей ранних пакицетидных
китообразных, сделанный Тевиссеном (результаты изложены в статье в
журнале “Nature”
от 20 сентября 2001 года) говорит о наличии общих признаков в строении
китов
и парнокопытных.
Тевиссен предполагает, что сходство в зубной системе китообразных и
мезонихид, на основе которого ранее поддерживалась мезонихидная гипотеза
(см. у
Кэрролла) появилось независимо.
Естественно, вывод Тевиссена также был взят креационистами
на вооружение, но только в им одним присущей манере. На сайте http://www.scienceagainstevolution.org (громкое
имя у этого сайта: «Наука против эволюции») есть статья Do-While
Johnes
от ноября 2001 года. Он следующим
образом высказывается о родословной китообразных в свете исследований
Тевиссена:
Нам кажется разумным заключением то, что Pakicetus никогда не был предком кита. Это был просто некий вид вымершего наземного животного, у которого только случайно была пара ушных костей, которые имели некоторое подобие ушным костям кита. Но это заключение недоступно эволюционному сообществу, потому что это подразумевало бы, что креационисты были всё это время правы. Pakicetus должен быть сохранен как предок кита, несмотря ни на что. Итак, вот как они пробовали получить некую эволюционную пользу от разрушительного открытия остальной части скелета Pakicetus.
Вы уследили за этой логикой? Не огорчайтесь, если нет. Это было не очень логично. У Pakicetus кости лодыжки точно такие же, как у парнокопытных, которые являются наземными млекопитающими, которые очень хорошо бегают. Это очень удивляет нас, потому что Pakicetus совершенно очевидно был наземным животным, которое могло очень хорошо бегать. Pakicetus не был китом, или даже отдаленным подобием кита. Но название “Pakicetus” означает «кит, найденный в Пакистане». Поэтому по определению он – китообразное. С тех пор Pakicetus произвольно назывался китом, и так как он связан с парнокопытными, следовательно, все киты эволюционировали от парнокопытных. В любом случае, вот их логика. |
Странная, однако же, логика… Нет, не у эволюционистов. Я
просто напомню автору статьи, и всем, кто забыл: пакицетус был исходно описан
по черепу (и, соответственно, поименован исходя из признаков черепа), и много
лет от него не было известно остатков посткраниального скелета. Его череп
как раз имел признаки, характерные для китообразных; потому пакицетус
и был причислен к китам. А вывод о том, что он телом, вроде как, и на кита-то
не похож, был сделан позже, когда были найдены элементы его скелета. Так что
выводы
креационистов о
«неправомерном» причислении волкообразного пакицетуса к китам, и о «кажущемся»
родстве китообразных и парнокопытных сделаны, фактически, «задним числом».
Кроме того, Do-While Johnes должен бы определить, какие признаки уже делают
доистрическое
животное китом, а какие ещё не делают: границу, так сказать, провести. Спорить,
не раскрывая полностью своей позиции, очень легко – тебе дают какой-то пример,
а ты в ответ
надуешь
губёнки
и так с апломбом выдашь: «Не-а, вы не угадали. Давайте другой пример». А что
именно тебе надо, и почему не угадали – то никому неведомо.
Но вернёмся, однако же, к китам. Кроме парнокопытных, в природе есть много
разнообразных групп наземных млекопитающих. У них в задней конечности тоже
есть таранная
кость.
Даже у человека
она есть.
Но
у
китообразных таранная кость почему-то сходна с таранной костью не хищников,
не приматов, не грызунов, и даже не непарнокопытных... И не имеет какой-то
своей специфической формы, отличающей прото-китов от всех прочих отрядов (или
«бараминов») наземных млекопитающих. Ведь не может такого быть, чтобы у Творца
вдруг фантазии не хватило при сотворении каких-то странных существ с остроконечными
зубами и специфическим строением костей уха? Есть, над чем задуматься, верно,
господин
Do-While Johnes? Может, посчитаете вероятность
независимого
появления у археоцетов особенностей строения лодыжки, идентичных таковым у
парнокопытных? Сколько ноликов нарисуете после запятой?
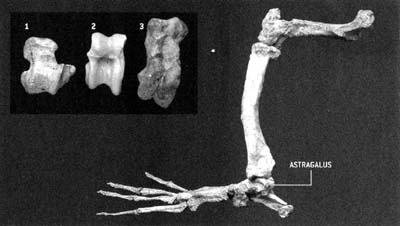
Задняя нога кита Rodhocetus. На врезке: таранные кости (слева направо) мезонихида, парнокопытного и Rodhocetus демонстируют характерную для археоцетов и парнокопытных форму таранной кости с двойным блоком.
Относительно «случайного» сходства костей уха пакицетуса и с таковыми у китообразных
могу лишь ещё раз предложить посчитать вероятность этого события.
Обвинение типа «назвался груздем – полезай в кузов» нельзя считать обоснованным.
По крайней мере, пакицетус был назван пакицетусом на основании того, что его
признаки в результате последующих шагов эволюции превратились в те, что характерны
для китов (естественно, такова точка зрения эволюционистов). Но сам пакицетус,
пока бродил по приречным лесам и мелким ручьям, «не знал», что от него произойдут
киты: ничья эволюция не предопределена и он вполне мог оказаться одной из многочисленных
«тупиковых ветвей», если бы события в природе развивались иначе. Это люди смогли
«постфактум» идентифицировать его как самого раннего кита, опираясь на знание
о более поздних животных, обитавших на Земле миллионы лет спустя, что называется,
«от обратного». Нелепость обвинения Do-While Johnes ещё более подчёркивает
родич пакицетуса с «политкорректным» названием Ichthyolestes – «рыбный разбойник»,
открытый в 1958 году и также идентифицированный позднее как раннее китообразное
(см., в частности, фрагменты из статьи Даррена Найша в начале обзора). По крайней
мере, к нему у товарища претензий нет. «Истинные» пакицетусы были открыты уже
после
него:
Pakicetus
inachus в
1981
году, а Pakicetus
attocki в 1980-м (но признанный как Pakicetus позднее, чем был открыт P.
inachus).
До последнего времени они все причислялись к семейству Protocetidae, но в наше
время на основании
новейших
находок из
Азии
выделяют
до четырёх
семейств примитивных китообразных.
Вообще, если слепо и тупо отталкиваться от названий, то огромный базилозавр,
которого даже такие столпы креационизма, как Джонатан Сарфати с Дуэйном Гишем
признают
китообразным, должен в таком случае
классифицироваться
как рептилия, а не как кит, поскольку его название означает «царственная
ящерица». Но в случае с базилозавром анализ его анатомии (в частности, строения
зубов)
позволил исправить первоначальную ошибку.
Do-While Johnes продолжает, комментируя отрывок из статьи Тевиссена в журнале
“Nature”:
Перевод: новые окаменелости включают кости тела и черепа, так что мы теперь знаем, на кого действительно был похож Pakicetus. Он выглядел точно так же, как наземное млекопитающее. И более того, он выглядел как наземное млекопитающее, умеющее хорошо бегать. Если бы он был найден без черепа, никто не подумал бы, что это был предок кита. Единственная черта этого существа, которое даже близко не напоминает кита – часть черепа, включающая среднее ухо. |
…А если бы тело оленя найти и классифицировать отдельно от его рогатой
головы, оно было бы явно родственно телу антилопы или козы. И никто бы
не подумал,
что это олень, животное из другого семейства. Выходит, так? Возможно, если
бы пакицетус был первоначально описан по остаткам посткраниального скелета,
его могли бы счесть особым видом примитивных копытных. В этом случае даже
название его не имело бы раздражающей креационистов приставки «-цетус».
Но находка образца
с черепом поставила бы всё на своё место. Конечно, название животного от
этого не изменилось бы, но систематическое положение – вполне. Наука не
стоит на
месте и исправляет свои ошибки. Пересмотр устоявшихся взглядов в свете
новых находок – это нормально в научной среде. Уж и не сосчитать, сколько
теорий
стало в связи с этим достоянием истории…
Одна ласточка весны не делает. Один признак чаще всего не определяет чёткой
принадлежности к отряду, особенно когда признаки группы в целом мало дифференцированы
по сравнению с современными родственниками и потомками древнего животного.
Оценка родства проводится по комплексу признаков, что сделал Тевиссен,
обработав большой массив данных и сравнив пакицетид не только с китами,
но и с большой
выборкой ископаемых млекопитающих. В качестве форм для сравнения он взял
несколько форм ранних парнокопытных, мезонихид и кондилартров. Результаты
исследований приводятся в его статье и свидетельствуют о родстве китообразных
и парнокопытных,
а мезонихиды выглядят по этим данным родственной, но отдельной группой. Анализа
этой статьи со стороны креационистов в природе пока не существует. Боятся,
что ли?
Классификация современных отрядов млекопитающих и отнесение
к ним некоего вновь открытого современного зверя не составляет труда, поскольку
мы имеем
дело с уже сформированными комплексами признаков, в наши дни практически не
имеющими промежуточных состояний. У древних представителей ещё «не сформированных»
отрядов
на заре
эры млекопитающих характерные черты были «размыты» - были плохо дифференцированы
и едва прослеживались. Зачастую древнейших представителей современных отрядов относят
к ним лишь на основе строения потомков, реализовавших и усиливших едва
заметные черты предков, прослеживаемые в филогенетических линиях. Об этом прекрасно
сказал Роберт Кэрролл, его цитата из книги «Палеонтология и эволюция позвоночных»
приведена выше.
В относительно недавно вышедшей книга Н. Ю. Клюге «Современная систематика
насекомых» («Лань», СПб, 2000) одна из глав как раз посвящена проблемам классификации.
Поверьте на слово, применительно к насекомым (обширный класс, включающий множество
отрядов разного уровня развития) эта проблема выглядит намного более запутанной,
чем при классификации китообразных и их предков (один отряд), неважно даже,
креационистами, или эволюционистами. Разбираясь с проблемой классификации китообразных
и их четвероногих предков, не лишним будет немного углубиться в терминологию
и полуить представление о подходе к построению родословных древ. Вот некоторые
существенные моменты, изложенные в книге Н. Ю. Клюге (стр. 15 - 16):
1.1.2.1. Термины апоморфия и плезиоморфия
Термин апоморфия (от греческого аро – от) или апоморфный признак означает прогрессивный (в исходном значении этого слова, от латинского pro – вперед и gressus – движение), т. е. продвинутый (или производный, вторичный) признак. Термин плезиоморфия (от греческого plesios – близкий) или плезиоморфный признак означает примитивный (в исходном значении этого слова, от латинского primus, primitivus – первый, первичный) признак (называемый также исходным, анцестральным, предковым или генерализованным признаком). Плезиоморфия и апоморфия – это два альтернативных состояния признака, соответственно до и после произошедшего с признаком эволюционного изменения. Эти понятия являются относительными: если эволюция идет от состояния А к состоянию В и далее к С, то В является апоморфией по отношению к А, но плезиоморфией по отношению к С. Термины «прогрессивный» и «примитивный» нередко связывают с понятиями «более совершенный» и «менее совершенный», или «более сложный» и «более простой», что может внести путаницу. Термины «апоморфный» и «плезиоморфный» имеют только одно значение – направление эволюционного изменения данного признака в данном случае, что делает эти термины более строгими. Апоморфия, отличающая данный таксон от всех прочих, называется его аутапоморфией (или автапоморфией), а апоморфия, общая для нескольких таксонов, называется их синапоморфией. Плезиоморфия, общая для нескольких таксонов, называется их симплезиоморфией. |
Рисунок из книги Н. Ю. Клюге, иллюстрирующий рассказ о принципах классификации.
1.1.2.2. Анализ филогении на основе синапоморфий
Кладистический анализ базируется на предположении о неповторимости и необратимости эволюционных преобразований (известном как закон Долло). Это предположение, в свою очередь, основывается на соображениях вероятности: вероятность того, что одно и то же изменение в генотипе может произойти независимо несколько раз, настолько мала, что ее можно считать равной нулю. Таким образом, предполагается, что каждая апоморфия возникла один раз и унаследована всеми потомками того вида, у которого она возникла. Следовательно, если у разных видов имеется один и тот же апоморфный признак (т. е. синапоморфия), это свидетельствует о том, что все эти виды унаследовали этот признак от своего общего предка и, следовательно, эти виды образуют единую филогенетическую ветвь. Для того чтобы доказать, что несколько таксонов образуют единую филогенетическую ветвь, необходимо выявить у них хотя бы одну синапоморфию, которая при этом являлась бы аутапоморфией для всей этой ветви. Так, на рис. 1 апоморфия 3 является синапоморфией таксонов с и d и аутапоморфией ветви c-d. Наличие этой апоморфии доказывает реальное существование ветви c-d и ее происхождение от общего гипотетического предка e, у которого и возникла эта апоморфия 3. в то же время наличие у нескольких таксонов симплезиоморфий не свидетельствует о том, что эти таксоны имеют общего предка, отличного от предков других таксонов. Так, на том же рисунке плезиоморфия 3 не позволяет ничего сказать о филогенетических отношениях таксонов a и b, кроме того, что эти таксоны не относятся к ветви c-d. Симплезиоморфия 2 относится к филогенетической ветви b-c-d, но наличия одной только этой симплезиоморфии было бы недостаточно для доказательства реального существования этой филогенетической ветви (реальность этой ветви доказывается апоморфией 1). Филогенетическое древо, построенное только на основе кладистического анализа, называется кладограммой; в отличие от некоторых других видов филогенетических древес, кладограмма содержит только информацию о последовательности ветвления и не содержит информации об абсолютном времени ветвления, степени различий между ветвями и числе видов в каждой ветви. Здесь важно отметить, что для реконструирования древа, ветви которого являются надвидовыми таксонами, необходимо не только выявить синапоморфии между ветвями, но и найти аутапоморфии каждой ветви. Если для каждого таксона, состоящего более чем из одного вида, не выявлено ни одной аутапоморфии, мы не имеем права изображать этот такон в виде филогенетической ветви. Так, в примере на рис. 1 для реконструирования филогенетических отношений четырех таксонов (a, b, c и d) необходимо не менее шести апоморфий. (выделено мной - В. П.) |
Что ж, источник не обвинишь в некоей «предвзятости», поскольку он посвящён совсем
иной группе живых организмов, а толкование данных терминов не зависит от того,
к какой группе живых организмов их применять. В свете комментариев к рисунку
утверждение креационистов о том, что признаки, по которым пакицетуса и амбулоцетуса
причислили к китам, мол-де, были взяты произвольно, чтобы дать желаемый результат.
Джинджерич сделал вывод о родстве пакицетуса и ранних китообразных на основе
вполне доступных для изучения признаков строения его черепа. Из статьи Ленни
Фланка «Амбулоцетус как переходная форма»:
…исследователь Фил Джинджерич нашел 52-миллионолетний
череп в мелководных отложениях в Пакистане. Хотя и фрагментарный, череп
имел зубы, которые были почти идентичны с таковыми у мезонихид и археоцетов.
Конфигурация костей в затылочной части черепа, однако, отличалась от
таковых у мезонихид, и была идентична таковой у археоцетов. На этом основании
Джинджерич заключил, что животное, которое он назвал Pakicetus,
было очень примитивным китом. «По времени и его морфологии, – сообщает
Джинджерич,
– Pakicetus совершенно промежуточная форма, недостающее звено
между более ранними наземными млекопитающими и поздними, оформившимися
китами. (Gingerich, "The
Whales of Tethys", Natural History, April 1994, p. 86)
|
Иными словами, пакицетус демонстрирует, с одной стороны, типичные черты предковых
форм (копытных), с другой стороны – раннее проявление черт, характерных для
потомков (китообразных). Таким образом, он соответствует признаками требуемой
креационистами переходной формы.
Данные строки были написаны до находки (точнее, до изучения и описания особенностей)
индохиуса. Очевидно, именно поэтому важный признак строения ушной области пакицетуса
не рассматривался Джинджеричем – он мог появиться у пакицетуса или его непосредственного
предка уже после разделения с мезонихидами, которое признавалось верным на
тот момент развития знаний об ископаемых китообразных. Но и из этого фрагмента
видно, что пакицетуса объединяет с более поздними китами больше, чем только
строение уха и зубов. Но следует обратить внимание, что Джинджерич сравнивает
пакицетуса с археоцетами, а не с зубатыми или усатыми китами. Если сравнить
пакицетуса с современными китами (что делают креационисты), то общих признаков
с ними окажется, естественно, намного меньше, нежели у пакицетуса и археоцетов,
поскольку в процессе эволюции на изначальный план строения наложилось большое
количество признаков, появившихся позже, и потому черты строения ранних китов
были утрачены.
Пакицетус не был типичным представителем парнокопытных, как его преподносят
креационисты вроде Харуна Яхьи. Он демонстрировал признаки, свидетельствующие
о раннем отделении от общего ствола группы. В обзоре Даны Дезони “From Where
the Whale?” от октября 2001 приводятся сведения о строении пакицетуса:
… Pakicetus – это первое китообразное, от которого
хорошо сохранились кости лодыжки. Кости лодыжек являются важными для
идентификации парнокопытных, которые особенно приспособлены к бегу. В
этом смысле пакицетус очень напоминает парнокопытных; таким образом,
раннее китообразное было также и парнокопытным. Скелеты пакицетуса также
демонстрируют некоторые примитивные черты плацентарных, которые были
утрачены у всех современных и большинства более примитивных ископаемых
парнокопытных. Это указывает на то, что линия, ведущая к Pakicetus, отделилась
от прочих парнокопытных до эволюции пакицетуса.
|
Иными словами, эволюция пакицетуса уходит корнями ещё дальше, когда парнокопытные, возможно, были даже не собственно парнокопытными, а специализированной ветвью кондилартров. Но границу в этом случае флажками не пометишь… Тем не менее, такие «флажки» всё же нашлись. В статье А. А. Банниковой «Молекулярные маркеры и современная филогенетика млекопитающих» (2004 год) приводятся любопытные данные о родстве китов и копытных:
Наличие в геноме семейства SINEs является «совершенной
синапоморфией» — надежным маркером клады эволюционного древа. Причина
в том, что, однажды возникнув в стволовой линии, семейство SINEs передается
всем последующим ветвям этого древа (Крамеров и др., 1980; Крамеров,
1987; Shimamura et al., 1997; Serdobova, Kramerov, 1998; Kramerov et
al., 1999; Nikaido et al., 1999; Shedlock , Okada, 2000).
Не менее надежный синапоморфный признак – это наличие отдельных копий SINEs в определенных локусах генома (инсерции). Предполагается, что инсерции SINEs свободны от конвергенций и реверсий, т.е. вероятность возникновения точно такой же вставки дважды в одном и том же месте генома ничтожно мала. Во всяком случае, не известны механизмы таких процессов (Shedlock and Okada, 2000), поэтому считается, что филогенетический сигнал этих маркеров не теряется в шуме гомоплазий. Таким образом, филогенетическая ценность SINEs определяется тем, что 1) присутствие в ДНК разных видов одного и того же семейства SINEs может быть результатом только их общего происхождения, т.е. является апорфным признаком; 2) встраивание этих генетических элементов в геном необратимо; 3) в отличие от LINEs, они распространяются в геномах путем только вертикальной передачи. Все это определяет ценность SINEs как кладистических маркеров видообразования (Miyamoto, 1999; Shedlock, Okada, 2000). Многие исследователи указывают теперь на необходимость тестирования любых филогенетических гипотез помощью SINEs (Waddell et al., 2001; Lin et al., 2002). … Одно из первых удачных применений инсерций SINEs для изучении филогении млекопитающих – обнаружение специфичного короткого ретропозона, общего для китообразных, бегемотов и жвачных (Shimamura et al., 1997; Shedlock , Okada, 2000). Кладу, объединяющую китообразных с бегемотами, поддерживают 4 синапоморфии SINEs из семейства CHR-1; другие 4 инсерции этого SINE и одна инсерция LINE семейства ARE также свидетельствуют в пользу парафилии парнокопытных (Nikaido et al., 1999). Возраст повтора ARE составляет не менее 25 млн. лет, т.е. появление его совпадает с дивергенцией парнокопытных и китообразных (Alexander et al., 1995). Семейства SINEs Bov-A2 и Bov-B, характеризующие подотряд жвачных парнокопытных (Ruminantia), и повтор SSPRE, маркирующий семейство свиных (Suidae), значительно моложе (Yasue, Wada, 1996). |
Со стороны креационистов попытку отделить китов от не-китов предпринял Дон Баттен в статье “A Whale of a Tale?”, опубликованной на сайте “Answers in Genesis”:
Протеро и др. предлагают пять особенностей, объединяющих
китов:
Все резцы параллельны зубному ряду – не сохранилось у Ambulocetus Медиальный ламбдовидный гребень полукруглый – не сохранилось у Ambulocetus Носовые отверстия сдвинуты назад – рострум (морда) не сохранился у Ambulocetus Протоконы маленькие (особенности зубов) Дополнительные выступы зубов большие (особенности зубов) |
Несложно заметить, что эти признаки вовсе не объединяют китов. Так, у достаточно большого количества вымерших китообразных ноздри всё ещё находятся «на полпути» между кончиком рыла и теменем. Таковы они, например, у Aetiocetus – самого раннего известного усатого кита (являющегося, таким образом, несомненным китообразным). У базилозавра ноздри также далеко не достигли темени, но в его принадлежности к китам никто не сомневается, даже креационисты. Далее, у современных зубатых китов зубы вторично упрощены – они конические и слегка изогнутые, похожие на клыки хищных зверей. Между бывшими резцами, клыками и коренными зубами современных китов нет явного различия. Ни о каких дополнительных буграх и вершинах, разумеется, речи не идёт. Выходит, что по классификации Протеро современные киты – это не киты? Вовсе нет. Палеонтолог с мировым именем не может так ошибаться. Позволю себе привести ещё одну цитату, но уже из статьи Ленни Фланка «Амбулоцетус как переходная форма»:
Что касается скелетных особенностей, киты,
тем не менее, могут быть отличены от похожих на них
мезонихид с
использованием пяти основных анатомических характеристик: (1) все резцы
параллельны
зубному ряду, (2), медиальный ламбдовидный гребень полукруглый, (3),
носовые отверстия сдвинуты назад, (4), протоконы маленькие, и (5), дополнительные
выступы зубов большие. Pakicetus, Ambulocetus и Indocetus все обладают
одной или большим количество этих характеристик, и таким образом классифицируются
как «археоцеты» (киты), нежели как мезонихиды. (у скелетов Ambulocetus не хватает значительной части черепа, чтобы определить первые три из
этих характеристик, и потому Тевиссен также добавил несколько других
характеристик черепа в выводе о том, что Ambulocetus был скорее археоцетом,
нежели мезонихидом, вызвав обвинения креационистов, что он «сменил определение
того, что есть кит».) (выделено мною – В. П.)
|
Как говорится, «почувствовали разницу»? Естественно, Протеро не мог так
глупо ошибаться. Просто его слова были в очередной раз выдернуты из контекста
и
использованы в простом словесном трюкачестве. Палеонтолог говорил об отличиях
ранних китов
от близких к ним мезонихид, и из слов Ленни Фланка видно, что пакицетус
обладал некоторыми из них (какими – в первоисточнике не уточняется). Что
ещё важно
в этих словах? То, что все перечисленные особенности не относятся к строению
уха. Поэтому можно сделать вывод, что пакицетус был причислен к китообразным
не только из-за особенностей строения уха.
Оригинальная точка зрения на особенности строения пакицетуса высказана креационистом
Джоном Вудмораппом в статье “Walking whales, nested hierarchies, and chimeras:
do they exist?” («Ходячие киты, вложенные иерархии и химеры: существуют ли они?»,
опубликована
на
сайте
Answers
in
Genesis).
Он
считает, что подобные живые существа – химеры наподобие русалок или «классической»
античной Химеры:
Недавние заявления о «наземных» китах исследованы
и опровергнуты. Тенденции, упоминаемые в эволюции китов, довольно поверхностны
по своей природе, и немного отличаются от тех, которые стали очевидными,
если сравнивать колесные транспортные средства в пределах кладограммы
[Вудморапп сравнивает эволюцию китов с «эволюцией» транспортных средств,
и приводит в статье соответствующие схему и таблицу – В. П.]. Тщательное
изучение эволюции китов вообще, и эволюции уха кита в частности, демонстрирует,
что большая часть анатомических черт не изменяются в последовательном
направлении к китам. Недавно обнаруженные пакицетиды состоят из китовых
«модулей» в составе иного «не-китового» тела. Эти вымершие существа –
примеры химерических существ. Китообразные, мезонихиды и парнокопытные
разделяют множество анатомических черт мозаично, что противоречит любому
типу эволюционной вложенной иерархии [в оригинале сказано “nested
hierarchy”, соотнести это понятие с какими-либо терминами в литературе
по теории
эволюции мне не удалось – В. П.], и это является сильным аргументом в
пользу специального сотворения всех этих существ.
… Эволюционисты говорят нам, что химерических существ не бывает, потому что для того, чтобы их существование стало реальностью, необходим совершенно невероятный набор обстоятельств. Поэтому, организмы, предположительно, эволюционируют как слегка измененные варианты их предков, и таким образом достигают высшей точки во вложенной иерархии всех живых существ. Именно в этой точке некоторые эволюционисты делают большой скачок. Они рассуждают, что Разумный Проектировщик, неоднократно используя один и тот же bauplan (план строительства) при сотворении различных форм жизни, должен создать живые существа, подбирая по крайней мере некоторые из модульных единиц. Результатом этого могли бы стать настоящие химерические животные, препятствующие, таким образом, любому виду классификации живых существ согласно вложенной иерархии. Несуществование химерических существ, как предполагается, поддерживает органическую эволюцию вне Специального Сотворения. |
Для того, чтобы доказать эту точку зрения, Джону Вудмораппу необходимо, как минимум, пересмотреть гораздо больше признаков живых существ, чем он сделал в статье, и доказать полную идентичность внешне сходного признака у различных животных (чтобы этот признак можно было считать образцом «готового модуля», использованного Творцом). Но внешне сходные признаки различных животных или растений чаще всего несут лишь поверхностные черты сходства, обнаруживая коренные различия при тщательном изучении. Так, внешне сходные двупалый и трёхпалый ленивцы оказываются представителями разных семейств, ведущих своё происхождение от разных наземных ленивцев. Ластоногие теперь считаются искусственной группой, объединяющей потомков куньих (настоящие тюлени) и медвежьих (моржи и ушастые тюлени). К слову, вот снова цитата из Банниковой:
Интерес представляют данные о стДНК ластоногих
(Pennipedia) [sic - В. П.]: некоторые сателлиты, найденные у тюленей,
гибридизуются с сателлитами куньих (Mustelidae), но не других Carnivora,
т.е. подтверждается
предположение о филогенетической связи ластоногих с мустелоидной ветвью
наземных хищных (Arnason, Wiedegren, 1986).
|
Так что внешнее сходство в биологии ничего не доказывает, поскольку может быть результатом конвергенции или параллелизма. А более глубокое сходство оказывается результатом общности происхождения хотя бы уже потому, что вероятность появления абсолютно идентичных признаков на одной и той же анатомической основе у видов, которые не родственны друг другу, исчезающее мала. Такое сходство может быть незаметно на первый взгляд, или может быть скрыто признаками, образовавшимися позднее (как у китообразных и парнокопытных, например), но современные методы изучения позволяют его выявить. Особенно показательно в этом плане сходство в строении ДНК.
Джон Вудморапп, “Walking whales, nested hierarchies, and chimeras: do they exist?”:
Ходячие киты, или ходячие химеры? Пакицетиды – интересный набор химерических существ, состоящих из лодыжки, похожей на лодыжку парнокопытных, и внутреннего уха, несколько похожего на таковое у современных китов, соединённых в теле, которое с трудом отличимо от тела типичного вымершего наземного парнокопытного!:
Эволюционный «преемник» Pakicetus едва ли лучше в этом отношении:
Конечно, не столь эффектное, как возможное открытие настоящей русалки, химерическое строение пакицетид и амбулоцетид едва ли могло быть описано в более ясной манере. Любым усилием воображения трудно представить, каким образом пакицетиды предложены как группа, заполняющая промежуток между наземными парнокопытными и водными истинными китообразными. Тот факт, что серьезные ученые-эволюционисты делают такие заявления, лишь продолжает показывать бедность свидетельств эволюции и сопутствующие этому доводящие до отчаяния дистанции, к заполнению которых эволюционисты будут привлекать некоторых вымерших существ в качестве переходных форм. |
А знаете, господин Вудморапп, почему это явление выглядит не так эффектно,
как вам, может быть, того хотелось бы? Потому что китообразные, мезонихиды
и парнокопытные суть производные от одного предка. Поэтому они вполне могут
унаследовать от этого самого предка начальные стадии признаков, которые впоследствии
станут характерными признаками отрядов млекопитающих, в течение миллионов лет
развивавшихся независимо. Если волшебным образом перенестись в тот же ранний
эоцен и изловить тогдашних зверей, парнокопытного, примитивного мезонихида
и пакицетуса, то при сравнении они окажутся весьма сходными и по современной
классификации, основанной на морфологии, могли бы быть признаны в лучшем случае
представителями разных родов в пределах одного – двух семейств одного отряда.
А поскольку воображаемый зоолог-путешественник во времени находится в прошлом,
он должен учитывать, что будущее каждой группы животных не предопределено (ах,
как сложно это сознавать с позиции человека, уже заглянувшего в последнюю главу
детективного романа!), и признаки, которые будут найдены у трёх вышеуказанных
зверей в живом виде, фактически будут лишь признаками, отличающими их только
друг от друга. Ровно так же мы можем лишь фантазировать на тему того, какие
признаки современных животных и в каком направлении разовьются через миллионы
лет эволюции. Тогда ведь некоторых современных животных, которым суждено стать
предками новых таксонов высокого ранга, придётся выделять в самостоятельные
отряды и семейства уже сейчас. А основание для этого есть?
От фантазии всё же перейдём к реальным существам. В свете этих рассуждений признаки,
которые с нынешних позиций характеризуют пакицетуса и амбулоцетуса как представителей
китообразных, будут лишь отличительными признаками некоторой группы потомков
примитивных копытных. Именно как «китовые» эти признаки будут признаны лишь «задним
числом», с позиций знания о том, к чему привело развитие их потомков. Поэтому
данных существ нельзя признать «химерическими», так как их отличительные черты
развились на основе признаков строения, унаследованных от предков. Если посмотреть
в подзорную трубу с другой стороны, можно и современных китов считать химерами,
которые разделяют специфические черты строения уха с ныне вымершими примитивными
копытными. Так ведь? Ведь г-н Вудморапп не станет отрицать того, что характерные
особенности строения уха примитивного копытного млекопитающего Pakicetus,
близкородственного
ранним парнокопытным, удивительным образом повторяются в ухе китов, которых
при всём желании трудно спутать с парнокопытными? Не так ли? Если
даже теперь
продолжается
«исследовательский зуд»,
и
хочется
искать
новых
и
новых
«химер»,
я
могу прямо сейчас,
не
сходя
со
своего места,
отыскать в природе много таких существ. Не верите? Вот одна из них. У неё тело
и голова кролика, а лапы в какой-то степени собачьи. Называется «заяц». Есть
существо с передними лапами обезьяны, головой оленя (только без рогов), задними
лапами зайца и хвостом рептилии. Причём покрыто это животное шерстью антилопы.
Называется
«кенгуру». Но
есть ещё одна, ещё более типичная «химера». У неё заячьи уши, лошадиные ноги,
львиный
хвост и серая козья шерсть на теле. Эта химера называется «осёл». Продолжать
поиск в том же духе? Y/N
Русалка, кентавр и единорог, не говоря уже о мантикоре и грифоне, кажутся нам
необычно-сказочными потому, что их тела состоят из частей, резко отличающихся
друг от друга по строению и физиологии. Зверь + птица, человек + рыба, зверь
+ скорпион… В облике этих существ сочетаются (и в то же время резко разграничены!)
признаки, характерные для представителей разных зоологических классов и даже
типов.
Ну,
ладно,
кентавр
и единорог, вроде, составлены из частей представителей класса млекопитающих.
Но млекопитающих, различающихся на уровне отрядов, причём как поздние представители
отрядов, не связанных общностью происхождения (примат + непарнокопытное и непарнокопытное
+ парнокопытное с рогом непонятной природы, соответственно). Если предположить
естественную эволюцию таких химерических существ, то столь резкой грани между
разными частями их тел просто не должно быть, поскольку любой орган эволюционирует
в составе организма и организм меняется как единое целое – изменение одного признака
неизбежно оказывает влияние на остальные признаки организма. И, если в составе
кентавра трёхлетний ребёнок ещё не отличается взрослым умом, то трёхлетний жеребчик
в его составе уже
вовсю
на
кобыл, э-э-э-э...
реагирует...
Развитие рыбьего хвоста у русалки будет «тянуть» за собой изменение передней
части её
тела в сторону улучшения гидродинамических характеристик, изменения физиологии
дыхания и соответствующих преобразований органов чувств. И личико такой тварины
будет
далеко не столь миловидным, как у человека. Можно
лишь посоветовать
господину
Вудмораппу и иже с ними подумать на досуге, как русалка должна разговаривать
под водой, не имея возможности использовать голосовые связки, и как это отразилось
бы на её анатомии.
В биологии существует понятие «химер», с этим я не спорю. Но это существа, состоящие
из генетически разнородных тканей. Например, растение сансевьера («щучий хвост»)
с листьями, окаймлёнными жёлтым, и пеларгония с белоокаймлёнными листьями – это
типичнейшие химеры. Может ли кто-то поручиться, что клетки «китового» уха пакицетуса
генетически
отличалось клеток от его же собственной «парнокопытной» ноги?
Джон Вудморапп, “Walking whales, nested hierarchies, and chimeras: do they exist?”:
Недавнее обнаружение неких китообразных (фактически
тюленеобразных) протоцетид не даёт ничего существенного для того, чтобы
закрыть огромную пропасть между пакицетидами и истинным китообразными.
За исключением обладания «двублочной» таранной костью (пяткой) «парнокопытного»
типа, эти недавно описанные протоцетиды являются высоко специализированными,
полностью водными существами, которые не показывают никаких чётких наследственных
связей с пакицетидами или амбулоцетидами.
|
Чтобы это стало правдой, Вудмораппу надо доказать, как минимум, что представитель протоцетид Rodhocetus был «высоко специализированным, полностью водным существом». Однако его позвоночник, чётко разделяющийся на отделы, его длиннопалые конечности с подвижными суставами, тазовые кости, сохранившие соединение с позвоночником – все эти признаки указывают на то, что Rodhocetus ещё сохранял способность передвигаться по суше. Подробности смотрите ниже, и, самое главное, обратите внимание на мнение Creation Wiki, где однозначно указывается на способность этого животного ходить по суше. Люблю я креационистов друг на друга натравливать...
Джон Вудморапп, “Walking whales, nested hierarchies, and chimeras: do they exist?”:
Давайте теперь возьмём «китовые» особенности в более широком контексте. Неудивительно, что, поскольку обнаружено большее количество окаменелостей, наша концепция анатомического разнообразия некоторых групп должна расшириться: некоторые анатомические черты, которые считали уникальными для определённых отрядов млекопитающих, проявляются как химерические наборы в других отрядах. Но вместо того, чтобы демонстрировать эволюцию, одновременное химерическое возникновение китовых и не-китовых особенностей у вымерших млекопитающих только показывает, что некоторые особенности, считавшиеся китовыми по своей сущности (потому что они имеют место только у современных китообразных, и ни у каких иных современных млекопитающих), в конце концов, оказались не исключительно китовыми. Это не оправдывает переопределения китообразных до абсурдных крайностей, охватывая все эти химеры, как в настоящее время сделано эволюционистами. |
Что ж, описанное Вудмораппом явление –
появление в разных
группах живых организмов сходных признаков –
реально существует. Доступные
и наглядные примеры этого
есть. Например, саблевидные клыки, торчащие изо рта, впервые появились у звероподобных
рептилий –
достаточно вспомнить плотоядных пермского периода Inostrancevia и Ivantosaurus,
а также травоядных пермских дейноцефалов и триасовых дицинодонтов. Затем они
появились у... нет, не у саблезубых кошек. У палеоцен-эоценовых травоядных
отрядов
Dinocerata
и Pantodonta. Далее у эоценового креодонта Machaeroides, потом у олигоценового
нимравида Hoplophoneus и миоценового нимравида Barbourofelis,
миоцен-плиоценового сумчатого Thylacosmilus, и с миоцена по плейстоцен
у разных родов кошачьих многократно и независимо друг от друга. А ещё с эоцена
по настоящее время сабельными клыками щеголяют...
олени. Кабарга и мелкие олени Юго-Восточной Азии весьма клыкасты, и клыки являются
характерной чертой внешности уже у самых примитивных оленей кайнозоя.
Заговорив об оленях, хочется вспомнить о рогах. Если из дидактических соображений
ограничиться рассмотрением только млекопитающих, то среди них можно увидеть
независимое появление рогов у совершенно разных групп, происходивших от заведомо
безрогих предков. В современной фауне рога есть у оленей, жирафов, полорогих
и вилорогов среди парнокопытных, и у носорогов среди непарнокопытных. В прошлом
рогатых зверей было гораздо больше. Рога отращивали близкие родичи лошадей
бронтотерии, дальние родичи хоботных арсиноитерии, а также уникальные вымершие
травоядные диноцераты.
Современные жирафы рогами не очень-то блещут, но среди ископаемых жирафов можно
отметить Prolybitherium с
рогами, похожими на костяную «бабочку» с раскрытыми крыльями. Некоторые
другие формы жирафов щеголяли ветвистыми,
как у оленей, рогами, которые, однако, не сбрасывались. Для больше остроты
ощущений я бы добавил к «параду рогоносцев» родственников верблюдов из семейства
Protoceratidae с их рогами уникальной формы, вплоть до вилообразной, а также
рогатых грызунов
Epigaulus и рогатую свинью Kubanochoerus. И капелька перца:
шестирогое парнокопытное
Hexameryx и пятирогое Hoplitomeryx –
оба из миоцена острова
Гаргано (Восточная Италия).
И ещё пример («Остапа несло...» © И. Ильф, Е. Петров) –
с «парусными» животными.
«Парус»-терморегулятор на спине ископаемых зверообразных рептилий (подкласс
Synapsida) пермского периода Dimetrodon и Edaphosaurus известен
многим. Более осведомлённые читатели знают про похожий «парус» на спине раннемелового
растительноядного динозавра Ouranosaurus (подкласс
Archosauria, отряд Ornithischia). Кинозрители припомнят фильм «Парк юрского
периода-3», где одним из «действующих морд»
был «парусный»
хищный динозавр
Spinosaurus (отряд Saurischia, тот же подкласс) из мелового периода.
Чтобы завершить (или, сообразно хронологии, практически начать) это представление,
стоит упомянуть
Platyhystrix. Он выделяется в этом ряду уже тем, что вообще не является
рептилией: это земноводное пермского периода.
Просмотрев всё написанное сейчас, я задаюсь вопросом: почему я, вообще, выполняю
работу за господина Вудмораппа? Ему надо убедить читателя –
он пускай и приводит
примеры.
Вудморапп же, вместо того, чтобы доказать типичность явления, ограничился общей
фразой.
Если уж мне выпала честь искать среди животных «вудморапповых химер», то я поищу,
пожалуй... Уж поищу... Так поищу, что не впрок ему это пойдёт.
Мораль
сей
басни такова... Наличие некоторого частного признака, увы, не
позволяет признать
его
обладателя
«химерой»,
если
данный
признак
развивается
на
основе особенностей строения, имеющихся у предка данного вида. То есть, если
у непарнокопытных
не
развиваются зачатки рогов, позже прирастающие к черепу (как у полорогих), то
и рога непарнокопытных не будут полными аналогами рогов полорогих:
у бронтотериев это выросты носовых костей черепа, а у носорогов вообще кожное
образование,
своего рода «мозоль» на носу, не имеющая костной основы. Удлинение клыков или
верхних
остистых отростков
позвонков вообще является сугубо количественным изменением, хотя и влечёт за
собой качественные изменения в анатомии (например, в строении черепа,
если
клыки
используются для нанесения удара жертве) и физиологии (когда «парус» используется
как терморегулятор).
А
принципиальных
препятствий
для независимого
повторения
в разных обстоятельствах единичного частного признака я не вижу. Из-за сабельного
клыка
диноцерат
или
олень
саблезубой
кошкой
не станет –
прочие его признаки будут соответствовать таковым у остальных представителей
таксона,
к которому он принадлежит. Подводя итог, можно сказать так: если особенности
строения существа, рассматриваемого на предмет «химеричности», выводятся из особенностей
строения его предков в соответствии с изученными закономерностями эволюционных
процессов (подробнее об этом рассказано здесь),
это существо никаким образом
нельзя
считать
«химерой».
Вот,
если
без
чуда божественного происхождения не обойтись –
тогда перед нами несомненная химера.
По возможности хотелось бы получить от креационистов примеры этих самых химер,
но
с доказательствами в виде детального рассмотрения особенностей строения представленной
химеры и видов, которые эволюционисты несправедливо полагают родственными ей.
Без этого объяснения слова Вудмораппа пока что выглядят пустословием.
Родственники китообразных? Химерическая трихотомия. Парнокопытные или мезонихиды являются самыми близкими
родственниками китообразных? До недавнего времени вымерший отряд Mesonychia
был в значительной степени принят эволюционистами как сестринская группа
отряда Cetacea (…). Недавний анализ продемонстрировал, что соответствующие
зубные комплексы мезонихид и китообразных стоят в основе объединения
этих двух групп в ветвь. Это поддерживается разнообразием других мезонихидно-китообразных
синапоморфий.
Рисунок 4.
Рисунок 5.
Так как рационалистические предвзятые мнения, конечно,
не будут позволять эволюционисту рассматривать последнюю возможность,
он вынужден будет спотыкаться в своём воображении и моделях организации.
Для некоторых эволюционистов вторичная «обратная эволюция» специализированной
пятки парнокопытного рассматривается как возможная (рисунок 4, слева).
Другие утверждают, что пятка с «двойным блоком»
– гомоплазия. Согласно
этому рассуждению, «двублочная» таранная кость должна возникнуть дважды
независимо (конвергентно) у парнокопытных и мезонихид (рисунок 5, справа).
[Возможно, здесь явная ошибка автора. В оригинале сказано: "According
to this thinking, the ‘double-pulleyed’ astralagus must have arisen
twice independently (convergently) in artiodactyls and mesonychians
(Figure
5, right)". Однако, на схеме ясно видно, что таранная кость с
двойным блоком не возникает у мезонихид (и фактически такого признака
у них
нет), поскольку они отделяются от предкового ствола до возникновения
этого
признака. Очевидно, имелось в виду возникновение сходных признаков
строения зубов – В.
П.] |
Разрешить ситуацию, обрисованную Вудмораппом, помогает, как ни странно, другой креационист, Do-While Johnes. В его работе «Сказка о ките – часть вторая» от ноября 2001 года, фактически, содержится ответ на вопросы Вудмораппа, хотя и в своём, креационистском стиле:
Что заставляет учёных объявлять это существо [Pakicetus]
китом? Комбинация тонких особенностей – расположение бугров на коренных
зубах, складка в кости среднего уха, и расположение ушных костей в пределах
черепа – отсутствует у других неземных млекопитающих, но является характерным
признаком более поздних эоценовых китов. Глубокие, почти вертикальные выемки составляют большую часть следов
износа зубов у пакицетид. Эволюционные аргументы использовались, чтобы
связать этот тип износа с водным хищничеством, охотой на рыбу, но никакой
функциональный модели или современного аналога неизвестно. Кроме того,
этот вид износа зубов также имеет место у парнокопытных Raoellidae.
Хотя этот тип износа зубов, вероятно, представляет особый способ обработки
пищи, он не обязательно подразумевает водный образ жизни. Хотя есть общее сходство у зубов археоцетов [древние
киты] и таковых у мезонихид, такое подобие иногда преувеличено и явно
представляет эволюционную конвергенцию. |
Может, «лживые» и «вредные» эволюционисты всё-таки дадут определение конвергенции?
Пожалуй, стоит это сделать.
«Конвергенция (от лат. convergo – приближаюсь, схожусь), независимое развитие
сходных признаков у разных групп организмов к сходным условиям внешней среды.
Сходство признаков, возникающее в результате конвергенции, называется аналогией
в отличие от гомологии – сходства, основанного на происхождении разных групп
от общего предка путём дивергенции». (определение взято из «Биологического
энциклопедического словаря», М., «Советская энциклопедия», 1989, стр. 276).
Результат конвергенции – аналогия, то есть, появление органов, сходных внешне,
но происходящих из разных зачатков. Классический пример – млекопитающее крот
и насекомое медведка. К сожалению, явление конвергенции не было придумано
для оправдания некоего несходства картины желаемой и картины, которая получается
в результате исследований. Оно было выделено как результат изучения анатомии
современных видов живых существ, не вдаваясь в особенности их происхождения.
Поэтому ехидный комментарий относительно «неувязок» желаемого и действительного,
и придуманного специально для этого «явления» тут не просто неуместный, но
даже откровенно ложный.
В случае со сходством/различием зубов китообразных и мезонихид имеет место
несколько иное эволюционное явление: не конвергенция, а параллелизм. Его
иногда называют «конвергенцией близкородственных форм», поскольку при параллелизме
разные группы живых существ независимо друг от друга формируют в процессе
эволюции сходные признаки на одной основе, унаследованной от общего предка.
В случае с ранними китами и мезонихидами как раз имеет место именно такое
явление. И китообразные, и мезонихиды эволюционировали от одних и тех же
предков, ранних копытных (архаичных парнокопытных или их непосредственных
предков). Поэтому некоторое изначальное сходство в строении их зубов вполне
объяснимо с позиции общего происхождения. Дальнейшее сходство усилилось в
ходе приспособления к хищничеству, которое произошло после разделения этих
групп. Возможность независимого появления зубов мезонихидного типа демонстрирует
совершенно неожиданная находка 2007 года в провинции Саскачеван в Канаде.
В отложениях маастрихтского века мелового периода (самая последняя эпоха
мела) в формации Френчмен была найдена челюсть небольшого млекопитающего,
получившего название Nanocuris improvida. Коренные зубы этого животного как
раз имеют сходство с зубами мезонихид, хотя под сомнением находится даже
принадлежность Nanocuris к плацентарным млекопитающим.
Что ж, большое спасибо Do-While Jones за разрешение вопроса, который Джон
Вудморапп считал совершенно неразрешимым с точки зрения рационального мышления.
Подводя итоги, можно сделать вывод о том, что сходство в строении зубов мезонихид
и китообразных представляет собой хороший пример параллелизма, а вывод о
родстве ранних китообразных и парнокопытных, выведенное на основе строения
костей ног (в частности, таранной кости) является справедливым. Он также
подкрепляется сходством строения ушных костей ранних китов и Indohyus,
несомненного парнокопытного из семейства Raoellidae, которое считается сестринской
группой
для ранних китообразных.
Можно ли считать, что зубы являются ненадёжным признаком
для определения млекопитающих различных групп? Думаю, не стоит делать сразу широкие
обобщения, поскольку биология – это область знания, в которой правила очень
часто имеют исключения. Вне
всяких
сомнений,
зубы
представителей разных групп млекопитающих
несут характерные признаки и могут быть использованы для определения систематической
принадлежности вновь найденных ископаемых существ. Однако, эволюционные явления
типа конвергенции и параллелизма могут определённым образом накладываться на
облик зубов, что приходится учитывать в работе с образцами. Природа – это
область
знания, в которой редко можно найти 100%-но надёжные и однозначные признаки.
Что касается мнения Вудмораппа о «мозаичности»
и «химеричности» ранних парнокопытных, мезонихид и примитивных археоцетов, то
я бы предложил
всем желающим просто
обратиться к мнению специалистов. Тем более, что анализ наборов этих
самых признаков уже сделан, и картина вырисовывается вполне упорядоченная.
Сделал
этот анализ не кто иной, как Ханс Тевиссен, от упоминания имени которого
креационисты содрогаются трусливой дрожью по тёмным углам. Причём результаты
анализа представлены в первоисточнике, который должен быть очень хорошо
знаком сторонникам «креационной науки»: в журнале ‘Nature’ от 20 сентября
2001 года,
в статье “Skeletons of terrestrial cetaceans and the relationship of
whales to artiodactyls”. Ага, в той самой статье, откуда креационисты бездумно
передирают рисунок скелета пакицетуса.
Тевиссен и коллеги пишут о результатах своих исследований следующее:
Современные представители млекопитающих отряда Cetacea
(киты, дельфины, и морские свиньи) – это постоянно-водные плавающие животные,
легко отличимые по морфологии, отсутствию волос и задних конечностей,
имеющие плавники, хвостовые лопасти и обтекаемое тело. Эоценовые ископаемые
документируют многое из перехода китообразных с суши в воду, но до сих
пор наиболее примитивный представитель, для которого был известен скелет,
был явно земноводной формой и жил в прибрежных биотопах. Здесь мы сообщаем
о скелетах двух ранних эоценовых пакицетидных китообразных, Ichthyolestes
pinfoldi размером с лису, и Pakicetus attocki размером с волка. Их скелеты
также объясняют отношения китообразных с другими млекопитающими. Морфологический
кладистический анализ показал, что китообразные более близки к мезонихиям,
группе вымерших архаичных копытных, но молекулярные исследования указали,
что они – сестринская группа гиппопотамид. Наш кладистический анализ
указывает, что китообразные более близкородственны парнокопытным, чем
любому из мезонихий. Китообразные – не сестринская группа никаким мезонихиям,
но гиппопотамам. Наш анализ отвергает идентификацию любого отдельно
взятого семейства парнокопытных как сестринской группы китообразных,
и поддерживает монофилию парнокопытных. (выделено мной - В. П.)
|
Здесь полезно вспомнить метания креационистов при словах, что ранние киты были родственны зверю, похожему на оленя. При этих словах у них начинаются ужимки и прыжки, которые, впрочем, лишь подтверждают верность научных воззрений на антропогенез. Тут и патетическое вопрошение «а может быть, им стоило сказать, что это и был олень?», и шутка Дуэйна Гиша с превращением коровы в кита, которую он ласково называл «сбоем вымени» (в оригинале “udder failure”)... Кто сказал «всё ты придумал»? Вот же оно, здесь можно найти: The Origin of Mammals by Duane Gish, Ph.D.
В летописи окаменелостей попросту не существует переходных форм между морскими млекопитающими и их предполагаемыми наземными млекопитающими предками... Крайне занимательно, начав с коров, свиней или буйволов, попытаться изобразить, на что могли бы быть похожими промежуточные формы. Начав с коровы, можно было бы даже вообразить одну родословную линию, которая безвременно вымерла, из-за чего это могло бы быть названо «сбоем вымени». |
Но Тевиссен однозначно пишет о том, что ни одно
современное семейство парнокопытных не может считаться сестринской
группой для китов. И тем более, добавлю от себя, предком китов.
Поэтому фантазия со «сбоем вымени» пусть используется на креационистских
утренниках в креационистских
же детских садах в злополучном Солт-Лейк Сити (Солёнозёрске, по-нашему) или
ещё где подальше.
Далее в статье из журнала ‘Nature’ сказано:
В противоположность дебатам о сестринской группе
для китообразных, отношения между эоценовыми китообразными и Cetacea
как таковыми – не противоречивы. Все филогенетические исследования указывают,
что пакицетиды более близкородственны ныне живущим китообразным, чем
парнокопытным и мезонихиям, и что пакицетиды разделяет синапоморфии уха
китообразных. В кладограмме за пакицетидами следуют амбулоцетиды, а современные
китообразные (зубатые и усатые киты) близкородственны позднеэоценовым
базилозавридам и дорудонтидам. Наиболее архаичное китообразное, для которого
известен скелет – земноводный Ambulocetus natans. Это было сильное животное
размером с моржа, которое жило в прибрежных местообитаниях и напоминало
крокодила, за исключением длинных задних ног, которые использовались
в плавании. Хотя Ambulocetus не похож на современных китов, он также
значительно отличается от своих родственников из числа наземных млекопитающих,
будь они парнокопытными или мезонихидами. Адаптация к жизни в воде у
Ambulocetus и более поздних китов усложняет установление их самых близких
родственников среди наземных млекопитающих. Данные об ископаемых китах,
которые ближе к основанию филогенетического древа китообразных и имеют
меньшее количество адаптаций к водному образу жизни, могло, возможно
дать новые идеи о филогенетике. Пакицетиды находятся в этом месте и могут
использоваться, чтобы проверить мезонихидную и гиппопотамидную гипотезы.
Есть три рода пакицетидных китообразных: Pakicetus, Nalacetus и Ichthyolestes. Pakicetus самый крупный, за ним следует Nalacetus (приблизительно на 5% меньше по линейным размерам), и Ichthyolestes (приблизительно на 29% меньше). До сих пор для пакицетид были описаны только зубы, челюсти и один череп. Мы откопали четыре частичных черепа, два из которых сохраняют орбитальную область, несколько фрагментов морды, и приблизительно 150 изолированных посткраниальных костей пакицетид от многих особей. Они были найдены на одном участке раннеэоценовой формации Кулдана в Пакистане. Мы используем эти окаменелости, чтобы показать (1), что эти архаичные китообразные были наземными млекопитающими; и (2), что китообразные более близко связаны с парнокопытными, чем к мезонихиями. |
Филогенетический анализ
Есть две текущих гипотезы о самых близких родственниках китообразных,
подкреплённые, соответственно, морфологической и молекулярной систематикой.
Морфологические
филогенетические исследования указывают, что сестринская группа для китообразных
– это (одна из) мезонихиды, вымершая группа плотоядных копытных. С другой
стороны, молекулярные исследования указывают, что китообразные принадлежат
к парафилетической группе парнокопытных, и что гиппопотамы являются их
ныне живущей сестринской группой. |
Вот схема родственных отношений между ранними китами, мезонихидами, кондилартрами и парнокопытными, построенная на основе результатов исследований Тевиссена и коллег (в статье указана под №4):
Схема из статьи Тевиссена в журнале ‘Nature’, иллюстрирующая отношения между ранними китообразными и различными группами копытных.
D. - Diacodexis, род примитивных парнокопытных. Тевиссен и коллеги брали американский и азиатский виды рода отдельно, поскольку исходили из информации о полифилетичности этого рода.
Схема построена на анализе тех самых признаков, которые «мозаично» «разбросаны» у этих животных. Если кому-то интересно, вот комментарии по проведению исследований из оригинальной статьи на английском языке (гиперссылки из оригинальной статьи в цитате не сохранены - В. П.):
Cladistic analysisIn order to test the mesonychian and hippopotamid hypotheses, we chose in-group taxa that both sample ungulate diversity and remain pertinent to the question of sister group relations to Cetacea. We chose four archaic ungulates to root the tree in the ungulate radiation with Arctocyon as outgroup, a practice followed by many other studies4, 44, 46. We limited Cetacea to two taxa (pakicetids, Ambulocetus) because the identity and basal status of these taxa are generally accepted1, 4, 5 and the inclusion of more taxa would make the analysis unwieldy. We included a large sample of mesonychians, following a previous study4. We also included 13 artiodactyls, sampling across the breadth of the modern suborders (Suina, Tylopoda and Ruminantia), but focusing on hippopotamids, their relatives, and Eocene artiodactyls. Following a previous study42 we suspected that the genus Diacodexis was paraphyletic. Therefore, we treated its North American D. secans and Asian D. pakistanensis as separate clades. We also added three families of Eocene Old World artiodactyls: cebochoerids, haplobunodontids, and raoellids. An explicit phylogenetic analysis of artidodactyls42 found raoellids near the base of a radiation of bunodont artiodactyls (including hippopotamids), and cebochoerids to be in an unresolved trichotomy with hippopotamids as part of the second clade, and anthracotheriids as the third. Anthracotheriids (represented in this analysis by Elomeryx4) are commonly considered to be closely related to hippopotamids, and haplobunodontids are traditionally included in the stem group of anthracotheriids. European haplobunodontids are thought to be middle Eocene migrants from Asia, making them possible candidates for the cetacean sister group under the hippopotamid hypothesis. Only three families of artiodactyls are known from Eocene Indo-Pakistan, the probable birthplace of cetaceans. We include all three in our analysis: raoellids, dichobunids (D. pakistanensis) and anthracotheriids. Our matrix contains 29 taxa and 105 characters, 95 of which are parsimony informative and eight of which are ordered. Although gaps and missing characters represent different types of data, they are scored the same. We analysed the matrix using heuristic search algorithms in PAUP 4.0b8 software47. ACCTRAN and DELTRAN optimizations were performed to investigate character transformation and estimated bootstrap values were calculated. Our maximum parsimony analysis produced 38 trees with a length of 281, consistency index of 0.39, and retention index of 0.61. A strict consensus cladogram of these trees is provided in Fig. 4. Character descriptions and scores, their sources, our matrix and our analyses are presented as Supplementary Information. Our heuristic search recovered a second island of trees at 282 steps. There are 38 trees with this tree length and both the Cetartiodactyla and Artiodactyla nodes were maintained in all trees. Our results are robust against addition of Hyopsodus as an outgroup, to exclusion of Meniscotherium and Phenacodus (which are dentally derived), and to the exclusion of Eoconodon and Andrewsarchus (which are in important positions in our cladograms, but are poorly known). The latter two taxa are the primary reason why character support for the Cetartiodactyla node varies with ACCTRAN and DELTRAN optimizations. ACCTRAN optimization shifts several important character changes from the Cetartiodactyla node to the node of Cetartiodactyla + Andrewsarchus. ... 1. Hulbert, R. C.Jr, Petkewich, R. M., Bishop, G. A., Bukry, D. & Aleshire, D. P. A new middle Eocene protocetid whale (Mammalia: Cetacea: Archaeoceti) and associated biota from Georgia. J. Paleontol. 72, 907-927 (1998). | ISI | ... 4. O'Leary, M. A. & Geisler, J. H. The position of Cetacea within Mammalia: phylogenetic analysis of morphological data from extinct and extant taxa. Syst. Biol. 48, 455-490 (1999). | Article | ISI | 5. Uhen, M. D. New species of protocetid archaeocete whale, Eocetus wardii (Mammalia, Cetacea) from the middle Eocene of North Carolina. J. Paleont. 73, 512-528 (1999). | ISI | ... 42. Gentry, A. W. & Hooker, J. J. in The Phylogeny and Classification of the Tetrapods, Vol. 2: Mammals (ed. Benton, M. J.) 235-272 (Clarendon, Oxford, 1988). ... 44. Thewissen, J. G. M. Phylogenetic aspects of cetacean origins: a morphological perspective. J. Mammal. Evol. 2, 157-184 (1994). ... 46. Naylor, G. J. P. & Adams, D. C. Are the fossil data really at odds with the molecular data? Morphological evidence for Cetartiodactyla phylogeny reexamined. Syst. Biol. 50, 444-453 (2001). | ISI | 47. Swofford, D. L. PAUP 4.0b8 Phylogenetic Analysis Using Parsimony (and Other Methods) (Sinauer, Sunderland, Massachusetts, 1998). |
На схеме видно, что, исходя из проанализированных признаков,
животные, взятые для исследований, располагаются на схеме не «мозаично», как
следовало бы ожидать, если Вудморапп был бы прав. Они образуют вполне чётко
оформленные группы кондилартров, мезонихид, китов и парнокопытных. Причём заметно,
что линия, ведущая к мезонихидам, отделяется от ствола парнокопытных + китообразных
раньше, чем разделяются киты и парнокопытные. И сами мезонихиды образуют довольно
целостную группу, из которой несколько выделяются Andrewsarchus и Eoconodon.
Иными словами, анатомические данные вполне поддерживают версию о происхождении
китообразных от парнокопытных, согласуясь, таким образом, с биохимическими
данными. Версию о происхождении китов от бегемотов можно оставить на совести
невежественных газетчиков, мыслящих заголовками.
Амбулоцетус триумфально ворвался в массовое сознание российского обывателя на волне сериала ВВС «Прогулки с чудовищами». Это необычное существо, в котором на первый взгляд сложно признать млекопитающее, привлекло внимание не только учёных-эволюционистов, но и креационистов. Удивительное строение этого зверя, сочетающего китовые признаки в строении черепа и хорошо развитые пятипалые конечности, унаследованные от наземных зверей, не даёт покоя сторонникам теории сотворения мира. Если волкообразного пакицетуса можно каким-то образом обвинить в «некитовости» и причислить к иной группе позвоночных, выбросив из числа китов, то коротконогий амбулоцетус, похожий одновременно на выдру и крокодила, стал более крепким «орешком» для креационистов. Это существо, словно нарочно, является как раз той самой земноводной формой млекопитающих, образцом переходного звена, наличие которого упорно отрицали креационисты. И теперь, когда этот зверь вышел из небытия, им приходится разными способами истолковывать его строение, чтобы оно не портило картину «сотворения мира».

Ханс Тевиссен изучает скелет амбулоцетуса

Реконструкция облика амбулоцетуса.
Самой интересной особенностью амбулоцетуса является наличие
хорошо развитых задних конечностей и таза. У современных китов пояс задних
конечностей редуцирован
до маленьких костей, не связанных с позвоночником и лежащих в толще мышц
на границе хвоста и туловища. Подробнее об этом рассказывается далее.
Джонатан Сарфати пишет в заметке на сайте Answers in Genesis, посвящённой
критике фильма PBS «Эволюция»:
«… Миллер [Кеннет Миллер, автор антикреационной книги «В поисках бога Дарвина» - В. П.] утверждает: «… животное могло свободно двигаться как по суше, так и в воде», и приводит рисунок полного скелета и реконструированного животного. Но это попытка ввести в заблуждение на грани с ложью, указывающая на недостоверность мнения Миллера, потому что нет указаний на тот факт, что далеко не все части были действительно найдены, как показано на схеме. Имеющий критически важное значение тазовый пояс не был найден (…). Без него Миллеру было бы слишком опрометчиво выдвигать такое заявление. Его коллега-эволюционист Аннализа Берта указывает, что:
|
Джонатан Сарфати приводит рисунок скелета амбулоцетуса из самого первого описания этого зверя, сделанного Хансом Тевиссеном в 1994 году. На тот момент известный скелет амбулоцетуса был действительно неполным, и некоторые части, в том числе тазовые кости, отсутствовали.
Вот он, тот самый рисунок, которым часто любят «козырять» креационисты. Сверху - первоначальная реконструкция скелета амбулоцетуса, внизу - первые окаменелости, ставшие известными науке.
Модификация предыдущего рисунка, которая размещена, в том числе, на сайте Answers in Genesis. Жёлтым выделены первоначально найденные кости.
Эдвард Бабински, много дискутировавший с Джонатаном Сарфати по поводу эволюции китообразных, пишет на своём сайте:
«Та же самая история рассказана на сайте AiG 17
августа 2001 в радио-странице «Ответы с Кеном Хэмом» с истеричным названием
«Эволюционисты – после наших детей!». С того времени, как этот рассказ
впервые был размещён в Сети, AiG разместил ещё одну заметку с этой историей
Кена Хэма и Карла Виланда, названную “A Whale of a Tale” [в русском
переводе это произведение называют «Сказка про белого… кита» - В. П.] (на AiG
есть две заметки под одним названием).
… «Ambulocetus может быть чётко соотнесён с археоцетами и другими китообразными на основе особенностей среднего уха, морды, крыши черепа и зубов [ссылка опущена]. Позади черепа, однако, образец показывает экстраординарную комбинацию черт, которые предвосхищают, но не полностью воплощают адаптации китообразных к водному образу жизни. Передние и задние ноги хорошо развиты, с гибкими суставами локтей, запястий, коленей, и пальцами. Кисть крупная и продолговатая, а ступня большая. Пальцы ног завершаются копытцами, как у мезонихид и других копытных, а хвост исключительно длинный». …
Давайте посмотрим ещё немного из того, что писал Тевиссен: |
Эдвард Бабински обратился непосредственно к Хансу Тевиссену, чтобы выяснить подробности находки Ambulocetus, и получил от него по электронной почте такой ответ:
«От: J. G. M. Thewissen
Послано: четверг, 02 декабря 2004 На: Вопрос об открытии Ambulocetus Экземпляр был раскопан в две стадии; результаты первой были опубликованы в 1994 г., итоги второй – в 1996-м. В 1994 г. мы описали несколько позвонков, большую часть костей ног, но никакого таза. В 1996 г. мы описали намного больше позвонков, а также таз. Так что выводы о позвоночнике в 1994 г. были основаны на позвонках, который у нас были тогда. Рисунок, который мы опубликовали в 1994 г., показывает раскраской, что было известно, и что не известно для образца в то время. Поэтому в действительности нет никакой причины, по которой кто-либо должен быть введён в заблуждение (пока он имеет неосторожность обращаться к первоначальной публикации). Причина задержки между этими двумя публикациями выглядит почти как в полицейском фильме. Мы пробовали возвратиться и собрать остальную часть экземпляра перед публикацией в 1994-м. Однако та область превратилась в пристанище для преступников. В тот день, когда мы собирались начинать там работать, был похищен один человек, и на дорогах там было выставлено много полицейских, чтобы противостоять похитителям. Они велели нам продолжать ехать и не останавливаться на той дороге, где это случилось. В силу этого я решил, что не было никакого смысла ожидать возможности собрать больше материала, потому что было неясно, позволят ли нам когда-либо возвратиться к участку. Ханс Тевиссен» |
Иными словами, практически все особенности строения амбулоцетуса известны
достаточно хорошо, чтобы учёные могли сделать далеко идущие выводы о его
родстве с типичными
китами. Но этот вывод абсолютно неприемлем для креационистов, поэтому для
них стало делом чести принизить значение открытия Ambulocetus.
Харун Яхья в статье “Whale fantasy from National Geographic” просто
не
мог обойти этого зверя стороной:
Второе ископаемое существо после Pakicetus в воображаемой
последовательности ‘National Geographic’ – Ambulocetus natans. Это ископаемое
первый раз представлено вниманию мировой общественности в 1984 в статье
в журнале Science. Фактически это наземное существо, которое эволюционисты
заставляют «превратиться в кита».
Название Ambulocetus natans происходит от латинских слов ‘ambulare’ (идти), ‘cetus’ (кит) и ‘natans’ (плавающий), и означает «ходящий и плавающий кит». Очевидно, что животное могло ходить, потому что у него были четыре ноги, подобно всем другим млекопитающим, и даже широкие когти на передних, и копыта на задних ногах. Кроме предубеждения эволюционистов, однако, нет абсолютно никакого основания утверждать, что он плавал в воде, или что он жил на земле и в воде (подобно амфибии). |
Далее он обрушивается с критикой на то, что хитрые эволюционисты, дабы сильнее воздействовать на ум обывателя, злокозненно подрисовали амбулоцетусу плавательные перепонки:
Чтобы увидеть границу между наукой и вожделеющим [в оригинальном тексте сказано: wishful – В. П.] воображением на этом объекте, давайте взглянем на реконструкцию Ambulocetus из ‘National Geographic’. Вот, как он изображён в журнале: |
Как говорится, «кто про что, а вшивый – про баню». Харуну
Яхье, в отличие от учёных, важен пропагандистский эффект, поскольку сейчас
религия – это
большие
деньги. Но, пока деньги не отвели всем глаза, лишний раз сходим в зоопарк
и посмотрим на выдру, а потом попробуем переосмыслить слова Харуна Яхьи.
Выдра
– практически точный аналог древнего амбулоцетуса, только маленький. Если
быть более точным, учёные считают лучшим аналогом амбулоцетуса в плане способа
плавания не европейскую речную выдру, а её гигантскую амазонскую
кузину рода
Pteronura. У неё более длинный хвост, чем у европейской выдры, и
она движется под водой преимущественно за счёт волнообразных изгибов всего
тела.
Ещё одна особенность выдр – то, что на их пальцах есть перепонки. Естественно,
это не мешает этим зверям ходить по земле (или кто-то сомневается в том,
что выдра это умеет?). А утконос с перепончатыми передними лапами умеет даже
норы
рыть. Выдра, кстати, тоже обладает выраженным строительным поведением: например,
гигантская выдра Pteronura строит довольно обширные норы по берегам
рек бассейна Амазонки. Я уже не говорю о способности ходить по земле у уток,
гусей и лебедей,
а также у пеликанов, альбатросов и других птиц с перепончатыми лапами. То
есть, перепонки между пальцами не являются непреодолимым препятствием для
движения
по земле
и прочих жизненных отправлений, связанных с частично наземным образом жизни.
Естественно, поведение животного в ископаемом виде не сохраняется.
Мягкие ткани тоже. Поэтому о некоторых обстоятельствах можно лишь догадываться,
делая выводы
на основе строения сохранившихся частей тела. А их, как было сказано
выше, у амбулоцетуса предостаточно – замечательный по своей полноте скелет
и
несколько фрагментарных остатков. Поэтому возражение Харуна Яхьи отклоняется.
Но успокоить этим эмоционального
турецкого собеседника трудно:
С тем же самым способом эволюционистского подрисовывания,
которое применили к рисунку Ambulocetus, можно сделать любое животное
напоминающим любое другое. Вы смогли даже взять скелет обезьяны, подрисовать
плавники на её спине и перепонки между пальцами, и представить её как
«примата – предка китов».
|
В этом случае палеонтологу придётся очень сильно попотеть,
доказывая, как минимум, её адаптации к водному образу жизни (не говоря уже
о родственных отношениях с китообразными). Ведь признаки приспособленности
к такой среде обитания
должны остаться в скелете животного хотя бы в виде соотношения изотопов
в зубной эмали (об этом уже говорилось выше в обзоре, посвящённом Indohyus).
И характер отложений, в которых найдены остатки этого животного, тоже
должен подсказать некоторые особенности его среды обитания. Впрочем, изредка
ошибки в определении «места жительства» ископаемых животных имели место.
Даже птеродактиль когда-то считался морским животным, и лишь позже люди
догадались,
что он умел летать! Но такие явные ошибки долго не живут, и их быстро исправляют
на основании новой информации, полученной после более подробного изучения.
Что касается изотопного анализа, остатки китообразных чётко идентифицируются
с его помощью. В статье Тевиссена в журнале Nature от 20 сентября 2001
года указывается, что остатки пакицетид чётко отличались по изотопному
составу
(содержание стабильного изотопа углерода 13С) от образцов парнокопытных
из того же местонахождения.
Тевиссен указывает на то, что исследователям удалось таким способом определить
элементы посткраниального скелета пакицетид. Если проводить параллели с
ископаемыми лошадями (тоже одна из любимых тем креационистов), исследования
изотопного
состава костей чётко указывают на характер рациона животных, и согласуются
с особенностями строения зубов. Этот метод креационисты признают, поскольку
используют его как аргумент в своих работах. Поэтому гипотетическая лже-«водяная
обезьяна» Харуна Яхьи была бы очень легко изобличена при современном уровне
развития науки. Иными словами, аргумент тоже отклоняется.
В той же статье Харун Яхья продолжает гневное изобличение нехороших эволюционистов:
Недействительность обмана, выполненного на основе окаменелостей Ambulocetus, может быть видна на рисунке ниже, основанном на реальных палеонтологических данных:
|
Что ж, не всё так плохо… Особенно, если учесть, что эта
реконструкция скелета взята из вполне эволюционистского источника,
и есть даже на
сайте Ханса Тевиссена (только там скелет зеркально развёрнут
в другую сторону, но разве это имеет принципиальное значение?). Это означает,
что при всём мракобесии и звериной ненависти Харуна Яхьи к эволюционному
учению
он
сам всё-таки
заглядывает
в источники по теории эволюции. Только жаль, что видит он там… э-э-э-э… съедобный
плод некоторых фикусов.
Здесь, на реконструкции скелета, действительно не
будет перепонок между пальцами задних лап амбулоцетуса, поскольку реконструкция
скелета не предполагает подрисовывания к нему мягких тканей. Любых. Даже тканей
сердца, которое у амбулоцетуса точно было. А уж в плавательных перепонках позвоночных
никаких скелетных образований и быть не может. Именно потому они и не нарисованы,
а вовсе не по причине какого-то «чудесного прозрения». Зато во всех реконструкциях
прижизненного облика амбулоцетуса пресловутые плавательные перепонки
присутствуют...
Харун Яхья любит в своих рассуждениях ставить всякие мысленные эксперименты:
например, подключать мозг к компьютеру, и т. д. Давайте последуем его примеру,
и представим на секунду, что Харуну Яхье предложили для реконструкции скелеты
ног утки и гуся (современных), но не сказали, что это за птицы. Естественно,
косточки этих ног хорошо препарированы и очищены от мягких тканей. Что мы в
результате получим от Харуна Яхьи при применении им такой же логики? Полагаю,
что обычную трёхпалую ногу некоей усреднённой птицы. А как иначе? Мягкие ткани-то
не сохраняются в ископаемом виде, и препарирование костей было приблизительной
имитацией процесса разложения. Соответственно, ни о какой перепонке в реконструкции
и
речи быть
не могло.
Ладно, это была современная утка. Харун Яхья мог из посторонних источников
вызнать, что именно ему подсунули, и справиться с задачей правильно. А если
реконструировать ископаемую утку? Ладно, это птица из одного «барамина» с современной
уткой, и ей положено иметь перепонки на лапах. А если не утку, а иную птицу
семейства утиных? Хорошо, гусь, лебедь и мускусная утка тоже перепончатолапые
птицы. А если это будет гусеобразная птица не из семейства утиных? Вот здесь
железная логика Харуна Яхьи может дать сбой: к гусеобразным птицам принадлежат
паламедеи, у которых на ногах нет плавательных перепонок.
Задача с реконструированием ископаемых китов похожа на упомянутую реконструкцию
уток и их родственников: от «точно-кита» и «всё-ещё-узнаваемого-кита» плавно
переходим к «ещё-вроде-бы-киту», далее к «не-совсем-но-всё-же-киту-с-натяжкой»,
и далее к «крайне-сомнительному-киту».
С каждым шагом в прошлое китов сомнений и неоднозначных толкований
будет всё больше. Не всегда можно найти свидетельства строения мягких тканей
животных
(например, отпечатки), поэтому приходится искать аналоги строения среди современных
животных.
Я понимаю, что я не Шерлок Холмс, но давайте попробуем предположить, где мог
бы жить амбулоцетус. Его длинные пальцы могли бы использоваться, чтобы лазить
по деревьям. Но это животное размером с очень большого тюленя, и его лапы довольно
короткие. Вряд ли он смог бы лазить по деревьям, даже если бы захотел – он
для этого слишком тяжёл и неуклюж. Его зубы многочисленные и острые – это указывает
на то, что животное явно питалось мясом. Он мог бы быть падальщиком или хищником.
Но его ноги короткие, и животное явно стопоходящее – это совершенно очевидно
не быстрый бегун, и вряд ли он догнал бы другое животное на земле. У него также
нет характерных
для падальщиков дробящих зубов, позволяющих разгрызать кости, поэтому нельзя
предположить, что он питался мертвечиной. Он слишком велик, чтобы жить и охотиться
в норах, как горностай. Хищник, не
умеющий
быстро
бегать,
хищник с длинным телом – вот, кто это. А его находки на берегах древнего моря
Тетис явно указывают на связь амбулоцетуса с водными местообитаниями.
Ханс Тевиссен и его коллеги, описывая облик Pakicetus и его родственников в
статье
“Skeletons
of
terrestrial cetaceans and the relationship of whales to artiodactyls” (Nature
от 20 сентября 2001 года),
сравнивают
его с более
изученными на тот момент китами, в том числе с Ambulocetus. И для Ambulocetus приводится
интересная скелетная особенность, указывающая на его приспособления к водному
образу жизни:
Локтевой отросток (…) составляет меньше 12 % длины
локтевой кости у Ichthyolestes, тогда как он крупнее у Ambulocetus (24
%). У плавающих форм, таких, как базилозавриды и ластоногие, локтевой
отросток удлинён в передне-заднем и проксимально-дистальном направлении,
и служит сильным рычагом для вытягивания локтя и сгибания запястья. Короткие
локтевые отростки характерны для бегающих
форм.
|
Так что даже такие незаметные на первый взгляд особенности указывают на водный образ жизни амбулоцетуса. Но что такое факты, когда вывод уже сделан! Харун Яхья продолжает:
Опубликовав рисунок скелета животного, ‘National
Geographic’ сдал назад от ретуширования реконструкции, которое
сделало её кажущейся больше похожей на кита. Как ясно показывает скелет,
ноги животного были спроектированы, чтобы перемещать его по суше. Не
было никакого признака воображаемых перепонок.
|
Реконструкция головы Ambulocetus с сайта Ханса Тевиссена, выполненная Marion Lipka. Заметны верхнее положение глаз на голове животного, придающее ему отдалённое сходство с крокодилом, и остроконечные зубы плотоядного животного. |
Это гениальный вывод, господин Харун Яхья! Не забудем также
отметить, что по суше прекрасно перемещаются утки, пингвины, пеликаны, выдры
и крокодилы. Причём крокодилы даже умеют бегать галопом. Их ноги уверенно держат
этих животных на твёрдой земле, но это не мешает всем означенным животным иметь
перепонки между пальцами. Даже у лягушек есть перепончатые лапы, а они, поди
же
ты, ещё и прыгать умеют, и лазают по деревьям, и землю роют!
Иными словами, если
ноги амбулоцетуса выдерживают его вес на суше, это ещё не повод отрицать возможность
наличия у него плавательных перепонок. Длинные пальцы
этого животного (заметим, стопоходящего) являются весомым аргументом в пользу
наличия плавательных перепонок. Но это не единственный признак приспособления
амбулоцетуса к жизни в воде. В своей статье в ‘Nature’ Тевиссен упоминает,
к слову, что глазницы амбулоцетуса были
сдвинуты
вверх,
что характерно
для
водных животных типа бегемота.
Почему-то креационисты ни в одной из прочитанных мною работ (а их, поверьте,
было немало) не прокомментировали эту особенность амбулоцетуса, делая упор
на его конечностях, хвосте и строении
уха (последнее, правда, рассматривалось ими крайне поверхностно).
Воинственно настроенный против эволюционистов Малком Боуден приводит в своей
статье «Киты: никаких признаков
эволюции» совершенно
убийственный аргумент против водного образа жизни амбулоцетуса:
Должно быть, этому существу было весьма сложно удержать
добычу маленькими
копытцами, вместо острых когтей!
|
Должно быть, на суше это коротконогое существо охотилось гораздо
успешнее, чем в воде. Я не представляю себе, знает ли господин Боуден о существовании
таких животных, как крокодилы, змеи и тюлени. Все эти животные плотоядны, но
хватают добычу, вообще не пользуясь конечностями. У крокодилов
и тюленей на передних конечностях растут когти, но сами конечности очень коротки
и часто не достают до рта (или ими не пользуются во время еды), а у змей вообще
нет никаких передних конечностей. Тем не менее, это не мешает им всем успешно
хватать
живую
добычу
пастью и удерживать её челюстями. У
амбулоцетуса зубы своей формой указывают на несомненное питание живой добычей,
которую нужно удержать, умертвить и расчленить. Взгляните хотя бы на реконструкцию,
приведёную выше.
В статье Даррена
Найша “When whales walked the land… and looked
like antelopes…
and
mimicked crocodiles…. and evolved trunks. What?” рассказывается об особенностях
строения амбулоцетуса и его возможном образе жизни:
... Обладая спиной, подвижной в дорсовентральной
плоскости, и огромными длиннопалыми ногами, Ambulocetus кажется
сложенным для уверенного
передвижения по земле, но, вероятно, плавал с помощью сильных движений
спины вверх и вниз и гребли с помощью задних лап. Его удлинённый хвост
не имел мощных мускулов и особых позвонков квадратной формы, необходимых,
чтобы давать опору лопастям хвоста.
В отличие от пакицетид, у Ambulocetus была узкая морду и глазницы, направленные в стороны, относительно короткие ноги и в целом мощный скелет. Интригующая возможность, касающаяся палеобиологии Ambulocetus, состоит в том, что это был крокодилообразный хищник-засадчик, который искал добычу, скрываясь на мелководье (Thewissen et al. 1996). Это заключение основано на черепе Ambulocetus, похожим на крокодилий: обе группы имеют длинные, но мощные морды, заострённые зубы, сильные мускулы, закрывающие челюсти, и глаза, расположенные высоко на голове. Ambulocetus также показывает необычный и обширный износ зубов, который указывает, что он питался добычей с костями. Если аналогия с крокодилом жизнеспособна, на кого мог бы охотиться Ambulocetus? Антракобуниды, группа травоядных копытных, вероятно связанных со слонами, и сирены; представители обеих групп жили в тех же самых местообитаниях, что и Ambulocetus. Thewissen и др. (1996) предположили, что Ambulocetus мог бы охотиться на этих животных, а также на других животных, которые, возможно, могли приблизиться к береговой линии. Однако, учитывая, что большие крокодилы уже были обычны в морских и пресноводных местообитаниях, часто посещаемых Ambulocetus, эта теория имеет недостаток. Действительно, было указано, что примитивные китообразные, возможно, страдали от обширной конкуренции и хищничества со стороны крокодилов. Возможно, поэтому, Ambulocetus выбирал местообитания, где крокодилы были очень редки или отсутствовали. |
Джон Вудморапп (статья “Walking whales, nested hierarchies, and chimeras: do they exist?” на сайте Answers in Genesis) критикует реконструкцию Ambulocetus:
Редакторы журнала ‘National Geographic’ сделали бы очень неплохо, если бы, заботясь о своей малоподготовленной аудитории, относились [с вниманием] к следующим отрезвляющим фактам, касающимся реконструкции мягких частей тела, являющимся неэмпирическим (с очень немногими исключениями) и, фактически, направляемыми верой: «Традиционно Ambulocetus, раннее китообразное, изображается с шерстью (…). Как обсуждалось Джоном Гейтси и Морин О’Лири на стр. 562 – 570, гипотезы о филогении, однако, определяют, как мягкие ткани типа кожи, восстанавливаются у ископаемых организмов. Недавние кладистические изучения предполагают, что Ambulocetus был почти безволосым (…) [44]». ... 44. Anonymous, Cover description, Trends in Ecology and Evolution 16(10):inside cover, 2001. |
Харун Яхья Whale fantasy from ‘National Geographic’:
Другой сценарий, который ‘National Geographic’ пробует
навязать без слишком долгих обсуждений, касается поверхности тела рассматриваемых
животных. Подобно другим млекопитающим, Pakicetus и Ambulocetus, которые
считаются наземными млекопитающими, по всеобщему согласию, имеют тела,
покрытые шерстью. И их обоих изображают в ‘National Geographic’ покрытыми
густым мехом. Когда же мы двигаемся к более поздним животным (истинным
морским млекопитающим), весь мех исчезает. Эволюционистское объяснение
этого не ничем не отличается от фантастических сценариев ламаркистского
типа, которые мы видели выше. Истина этого вопроса – это то, что все
рассматриваемые животные были спроектированы в наибольшем соответствии
с окружающей их средой. Нерационально пробовать объяснить этот проект
через мутации или поверхностные истории ламаркистского типа. Подобно
всему замыслу в природе, конструкция этих существ – свидетельство сотворения.
|
Насколько важен вопрос, был ли амбулоцетус безволосым, или
имел густую шерсть? Не думаю, что он имел критическое значение для признания
этого
млекопитающего
китом, или напротив, не относящимся к китообразным. Среди других водных
млекопитающих, наших современников, существуют как мохнатые выдры, каланы
и котики (мех
всех этих животных высоко ценится), так и практически безволосые моржи
и морские
слоны. Тем не менее, котик, морж и морской слон относятся к ластоногим.
Конечно, сейчас ластоногие признаны полифилетической группой: под этим
названием объединяют
потомков древних медведей (моржи и ушастые тюлени) и куньих (настоящие
тюлени). Но всё равно стоит принимать во внимание, что среди водных
млекопитающих встречаются как волосатые, так и безволосые
виды. Поэтому не всегда можно точно
предсказать, когда с каким состоянием признака столкнёшься. Хотя, какая, к
чертям, разница, были ли пакицетус и амбулоцетус голыми, или же покрытыми шерстью?
Представим
себе такую ситуацию: завтра в штаб-квартиру Харуна Яхьи приведут на собачьем
поводке амбулоцетуса, покрытого шерстью. А теперь вопрос: станут ли от этого
особенности строения его уха и зубов менее «китовыми»?
Если амбулоцетусы обитали в тропическом климате исчезавшего тогда моря
Тетис, они, скорее всего, имели волосяной покров подобный шерсти у современного
тюленя-монаха, жителя тропических вод. Но на анатомическое сходство амбулоцетуса
с более поздними китообразными это бы не повлияло. Но хорошо, пусть амбулоцетус
будет
не волосатым,
а голокожим.
Но тогда он будет иметь ещё более «китовый» облик, что опять-таки не понравится
сторонникам «некитовости» амбулоцетуса типа Харуна Яхьи, но уже по другой
причине: мол-де, нарисовав это животное с безволосой кожей, авторы преувеличили
количество черт китообразного у этого вида. Так что, как ни крути, а при таком
подходе всегда найдётся, к чему прицепиться.
Дон Баттен в статье «Кит из сказки?» (A Whale of a Tale?), как и Харун
Яхья, сомневается в том, что
амбулоцетус был водным животным:
Важная черта китов – горизонтальные хвостовые лопасти. Вовлечение хвоста в процесс плавания требует прочных хвостовых позвонков с крупными отростками для прикрепления сильных мускулов. Тевиссен и др. демонстрируют один «хвостовой» позвонок, на котором почти нет отростков для прикрепления мускулов. Более того, этот один хвостовой позвонок даже не был найден с остальным скелетом, будучи «приписанным материалом», найденным пятью метрами выше. Другими словами, поясничная, тазовая и хвостовая части Ambulocetus были «сконструированы» всего лишь из одного поясничного позвонка, одной бедренной кости, небольшого куска большой берцовой кости (малой берцовой кости нет, таза нет), маленького кусочка головки лодыжечного сочленения и небольшого количества костей ступни и пальцев задних лап. И какое же подробное описание дано тому, как животное двигалось в воде и на суше! Утолщённое бедро и наличие копыт подтверждают, что Ambulocetus был наземным животным. Статья была получена журналом Science 28 октября 1993 г. и принята 3 декабря 1993, что указывает на то, что статья прошла процесс рецензирования без, или с минимальными изменениями, необходимыми перед публикацией, и все же статья полна в высшей степени предположительного материала. Реконструкция скелета предполагает, что это «кит». Авторы говорят: «Немногое известно о хвосте, у примитивных китообразных и их родственников всегда много хвостовых позвонков», так что они делали набросок длинного хвоста для Ambulocetus! Есть несколько параграфов догадок о способах передвижения на земле и в воде, и всё же нет даже таза или никаких связанных с ним позвонков! Движение передних конечностей также представлено подробно, и тем не менее, нет никакой плечевой кости или лопатки! Если бы статья такого качества была представлена для публикации в эмпирической области науки типа молекулярной генетики, она была бы отклонена напрямую. Почему тогда она была принята с такой готовностью? Это, вероятно, указание на статус палеонтологии как «науки», а также отчаянное желание неодарвинистов-эволюционистов найти какое-то ископаемое свидетельство «промежуточной» формы, чтобы укрепить веру в градуализм или, несомненно, непосредственно в эволюцию, поскольку светские газеты любезно и некритически сообщают о «находке». Остатки Ambulocetus были найдены в слоях «от нижнего до среднего эоцена».
Ископаемые остатки китов подотряда Archeoceti [орфография сохранена
– В. П.] были найдены в более древних эоценовых слоях, так что Ambulocetus вряд ли был предком современных китов, как требуется Тевиссену и другим. |
Эдвард Бабински отвечает на его аргументы:
Автор утверждает: «утолщённое бедро и наличие копыт
подтверждают, что это существо было наземным животным». «Наземное животное»?
Взгляните на рисунок, и оцените размер существа относительно длины его
передних и задних лап, особенно его укороченные передние лапы, и, кроме
того, его бедро относительно короткое по сравнению с таковым у наземных
животных. И обратите внимание, насколько длинными были его пальцы передних
и задних лап! Он выглядит более приспособленным к плаванию, чем к ходьбе/бегу,
хотя, вероятно,
проводил часть времени на суше. Даже крокодилы с их короткими передними
и задними лапами проводят часть времени на суше, но я не назвал бы их
строго «наземными животными». Что касается «копыт» амбулоцетуса, это
правда,
что он действительно
имел длинные пальцы, которые закончились крошечными копытцами, но это
всё же подтверждает другую связь между самыми ранними китообразными млекопитающими
и семейством Mesonychidae, которое было тем же самым семейством, к которому
принадлежал Pakicetus.
|
Вполне естественно ожидать наличия маленьких копыт у потомков
кондилартров. Если посмотреть на кисти и ступни амбулоцетуса, можно увидеть,
что они
относительно длинные, с пятью тонкими пальцами. Следовательно, ожидать
от амбулоцетуса
типичных копыт, какие есть у лошади или коровы, не стоит. Очевидно, копытца
амбулоцетуса
были небольшими, и напоминали скорее ногти. Наличие копыт не препятствует
универсальности животного, и не слишком сильно ограничивает пути его адаптации
– разве что,
специализированных лазающих форм среди специализированных копытных нет.
Так, копытные мезонихиды были довольно успешными раннекайнозойскими
хищниками (и
среди них эндрюсарх – самый крупный наземный хищный зверь). У других копытных,
халикотериев, копыта превратились в подобие когтей, у некоторых видов даже
втяжных. Неспециализированные копытные, даманы, умеют лазить по деревьям,
а при необходимости это делает даже домашняя коза. Для жизни в воде копыта,
даже
более развитые, чем у амбулоцетуса, тоже не помеха – азиатские буйволы,
бегемоты, десмостилюсы, сирены и водяной оленёк тоже являются копытными, но
они прекрасные
пловцы
и даже ныряльщики. Естественно, это не означает происхождения китов именно
от тех
или других, но показывает, что точка зрения Дона Баттена исключительно
умозрительна.
Читая ответ Эдварда Бабински, следует помнить, что более поздние исследования
Тевиссена и коллег выявили родство китообразных не с мезонихидами, а с
примитивными парнокопытными. Этот вывод можно считать более достоверным,
чем «мезонихидную»
гипотезу, поскольку она подтверждается биохимическими исследованиями. А
сходство молекул ДНК, в отличие от анатомического сходства, не подвержено
конвергенции и параллелизму, хотя и в этом случае приходится делать определённые
поправки,
связанные с особенностями изменения хромосом.
Справедливости ради нельзя не сказать о том, что Дон Баттен «не эволюционирует».
В добавлении к своей статье от 4 января 2002 года он пишет следующее:
Даже если дополнительный материал по Ambulocetus на сайте Тевиссена является приемлемым, он ничем не подтверждает, что это переходная форма между китами и наземными животными. Например, нет свидетельств развития горизонтальных хвостовых лопастей, столь характерных для китов, или уникального слухового аппарата китов (т. е. не имеющей наружного отверстия), или дыхала, и т. д., и т. п. Несомненно, нет ничего уникально «китового», что определяет Ambulocetus как родственника китов. Более того, утолщённое бедро и наличие копыт подтверждают, что это существо было наземным животным. |
«... И ещё я считаю, что Карфаген должен быть разрушен».
Только это и можно припомнить, читая дополнение, сделанное Доном Баттеном.
Сам смысл процитированного абзаца звучит как отчаянный вопль: мол, что
бы вы там нам не показывали, наше мнение всё равно остаётся неизменным. У бегемота,
между прочим, копыта гораздо крупнее, и они также есть на ластах ламантина
и дюгоня
(а у вымершей морской коровы очевидцы вообще описывали растущее на ластах
образование, похожее на лошадиное копыто). Сирены, между прочим, гораздо
более
специализированы
к
водному
образу
жизни, чем был в своё время амбулоцетус. И эти копыта не мешают их обладателям
жить в воде, в чём всяк сам может убедиться, наведя справки о биологии этих
животных.
По поводу уникального слухового аппарата китов Дон Баттен неправ дважды. Во-первых,
следует сказать, что даже у современных китов наружный слуховой проход всё
же есть,
хотя он очень тонкий. У дельфина, например, это крохотное отверстие недалеко
от заднего края глаза. А во-вторых, ушные кости амбулоцетуса исправно демонстрируют
признаки, характерные для китообразных. Подробнее о слуховом аппарате китов планируется
рассказать ниже.
Горизонтальные хвостовые лопасти характерны для китов, слов нет. Но лишь для
современных, составляющих лишь часть общего разнообразия этих животных на протяжении
времени существования всего отряда.
И
не
факт,
что
лопасти
должны
быть
на
хвостах
ископаемых
китов.
А дыхало вообще появилось у зубатых и усатых китов независимо друг от друга и
довольно поздно. А Basilosaurus, несомненный кит (даже Гиш и Сарфати это признают,
и Харун Яхья тоже (см. ниже)),
всё-таки
не
имел
дыхала.
Так
что же он – не кит? Или всё проще и дыхало – не обязательный признак кита?
Отложения, в которых находят ископаемые остатки амбулоцетуса, дают прямые указания
на местообитания этого животного (информация с сайта Evidence
for Evolution):
Отложения, в которых был найден Ambulocetus, содержат отпечатки листьев, а также окаменелости улитки Turritella и других морских моллюсков. Очевидно, что присутствие таких ископаемых должно означать, что окаменелости Ambulocetus были найдены в месте, которое было когда-то мелководным морем – хотя листья могут быть снесены в море водой и быть захороненными там, морские моллюски не могли бы встречаться на суше. |
Как видно, ископаемое окружение чётко указывает на связь амбулоцетуса с морскими прибрежными местообитаниями. Это ещё не пелагическое существо, но уже не строго сухопутный житель. Он искал пищу в воде уже не случайно, а целенаправленно. И на его обеденном столе, судя по крупным зубам амбулоцетуса, была не только рыба, но и довольно крупные существа.
Можно сделать вывод, что, несмотря на все лучшие усилия ‘National Geographic’, факт, что никаких переходных форм между наземными и морскими млекопитающими не было, и что те и другие появились со своими собственными специфическими особенностями, не изменился. Нет никакой эволюционной связи. Роберт Кэрролл принимает это, хотя неохотно и на языке эволюционистов: «невозможно идентифицировать последовательность мезонихид, ведущую непосредственно к китам»[7]. ... 7. Robert L. Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge University Press, 1998, 329 |
Здесь следует сделать ряд существенных оговорок. Кэрролл писал
приведённую строку в период, когда родственные связи между китообразными и
парнокопытными
не были ещё доказаны (напомню, что сходство индохиуса с китообразными
было выявлено лишь в 2007 году), и мезонихидная версия происхождения
китообразных считалась
основной
из-за некоторого
сходства
в строении
зубов
тех и
других. С позиций современного знания о происхождении китообразных
отсутствие сведений о линии мезонихид, ведущей к китообразным, становится вполне
объяснимым: мезонихиды были не предками, а родственниками ранних китов.
В свете рассмотренных особенностей ранних китообразных утверждения Харуна Яхьи
об отсутствии переходных форм между наземными животными и китообразными выглядят
каким-то шаманским заклинанием. Отрицать факты и не принимать их под копеечными
предлогами – не лучшая тактика в свете появления новой информации. Что же,
улитке удобнее прятаться в своей раковине. Но это не спасёт её от пожара,
который испечёт её прямо в собственной ракушке.
Другие ученые принимают точку зрения, что животные,
которых эволюционистские публикации вроде ‘National Geographic’ пытаются
изображать как «ходячих китов», фактически не имеют никакого отношения
к истинным китам, но являются отдельной группой живых существ. Хотя и
будучи эволюционистом, известный российский эксперт по китообразным Г.
A. Мчедлидзе не поддерживает описание Pakicetus, Ambulocetus
natans и
им подобных четвероногих существ как «возможных предков китов», и вместо
этого описывает их как полностью изолированную группу.
|
«О, брат, это жулики!» – так, кажется, говорил Карлсон,
который живёт на крыше, в знакомом с детства мультфильме. Ладно, Харун Яхья
никого
не ограбил
(поскольку,
по законам шариата, за это укорачивают руки с помощью топора). Тем
не менее, жульничать он пытается в полной мере… При этом утверждении
он
ссылается
на следующий источник: G. A. Mchedlidze, General Features of the
Paleobiological Evolution of Cetacea, trans. from Russian (Rotterdam:
A. A. Balkema,
1986), 91.
Ссылка на первоисточник – это похвально. А ещё, если что, можно прикрыться
ею, заявив: это, мол, не я, это вон тот товарищ с умным лицом понаписал
такую фигню, а я, как идиот, за ним её повторяю.
Я не считаю Г. А. Мчедлидзе невежественным человеком или человеком,
злонамеренно искажающим истину и ставящим палки в колёса науке. И
вот, почему. Книга
Мчедлидзе была переведена на иностранный язык и издана в 1986 году,
как можно увидеть
в выходных данных. На русском языке, стало быть, она заведомо издана
ещё раньше (а именно в 1976 году). Амбулоцетус, напомню, был открыт
через восемь лет после выхода иностранного издания
книги, в 1994 году! Я уже не говорю о находке сравнительно полного
скелета пакицетуса (2001 год)… Поэтому я философски отношусь к информации
из
книги
Мчедлидзе. А вот Харуна Яхью не прощу за откровенную попытку обмана
читателей. Кто сказал «это не обман»? Как же объяснить такую «нестыковку», когда
в книге,
написанной
в 1976 году, якобы приводится мнение, касающееся животных, одно из которых открыто
в 1983 году, а другое в 1994-м?
Далее, при рассказе о происхождении современных групп китов придётся
ещё раз обратиться к работе Мчедлидзе.
Пожалуй, индохиус, пакицетус и амбулоцетус – это три животных в родословной китообразных, которые подверглись наибольшему числу нападок со стороны креационистов. В следующей части этого обзора я бы хотел остановиться на более общих вопросах, связанных с эволюцией китообразных. В свете креационного «учения», соответственно.
Выражаю глубокую благодарность за помощь в работе над этим обзором: Brian Switek, Darren Naish, Edward T. Babinski.
Индохиус - «меньший брат» китов
Пакицетиды: первые в ряду
Небольшая интермедия: кит с хоботом
Родословная «ходячих китов»
Амбулоцетус – «кинозвезда»
из «Прогулок с чудовищами»
Скованные одной цепью:
последовательность форм ранних китообразных
Ещё одна интермедия: есть и подревнее киты
Путь к современным китам
Небольшая интермедия: о развитии китового уса
Ушки... не на макушке.
... На задних лапках ходят.
Держи нос по ветру!
Интермедия о правде и полуправде:
О том, как НЕ надо говорить о переходных
формах.
Дань прошлому в организме китообразных.