| Главная |
Первый кит, бегущий краем моря.
Ненадёжные киты креационизма.
Индохиус - «меньший брат» китов
Пакицетиды: первые в ряду
Небольшая интермедия: кит с хоботом
Родословная «ходячих китов»
Амбулоцетус – «кинозвезда» из «Прогулок с чудовищами»
Скованные одной цепью: последовательность форм ранних китообразных
Ещё одна интермедия: есть и подревнее киты
Путь к современным китам
Небольшая интермедия: о развитии китового уса
Ушки... не на макушке.
... На задних лапках ходят.
Держи нос по ветру!
Интермедия о правде и полуправде: О том, как НЕ надо говорить о переходных
формах.
Дань прошлому в организме китообразных.
Ещё один аспект, в котором креационисты пытаются оспорить
эволюционистов – это время существования разных родов ископаемых китообразных.
Если бы данные
о преемственности разных родов китообразных удалось оспорить, это дало
бы креационистам большой козырь в борьбе за свои идеи.
Перед рассмотрением утверждений
креационистов следует дать несколько пояснений по геохронологической шкале.
Кайнозойская эра делится на палеоген (включающий палеоцен, эоцен и олигоцен),
неоген (подразделяется на миоцен и плиоцен) и антропоген (плейстоцен и современная
эпоха – голоцен). Палеоген и неоген образуют третичный период, а антропоген
– четвертичный. Эоценовый отдел кайнозойской эры делится на три подотдела:
верхний, средний и нижний.
Нижний
эоцен (около
54 – 47 млн. лет назад) включает один ярус – ипр. Средний
эоцен делится на два яруса – более древний лютет (около 47
– 42 млн. лет назад) и следующий за ним бартон (42 – 40 млн.
лет назад). Верхний эоцен (40 – 35 млн. лет назад) включает единственный ярус
– приабон.
Харун Яхья в работе A Whale fantasy from ‘National Geographic’ уделяет
внимание этой проблеме:
«Для парада реконструкций на страницах 66 – 69 в номере “National Geographic” от ноября 2001 подразумевалась цель суммировать утверждения журнала относительно происхождения китов. Целая цепочка существ была выставлена одно за другим и описана как переходные формы в развитии кита. Согласно журналу, порядок этих существ, согласно геологическим периодам, когда они жили, был следующим: Pakicetus (50 миллионов лет назад) Список “National Geographic” продолжался, но включал известные категории дельфинов и китов.» |
Ископаемые китообразные, которых находят учёные, образуют достаточно чёткую последовательность, в которой логично связаны время существования, степень эволюционного развития животного и его экология (интерпретированная по характеру отложений, в которых были найдены остатки животного). В обзоре книги “The Emergence of Whales: Evolutionary Patterns in the Origins of the Cetacea” под редакцией Тевиссена приводятся следующие сведения о формациях, в которых были найдены ранние киты:
1. Формация Кулдана, северный Пакистан, ранний эоцен
(ипр) и средний эоцен (лютет)
2. Формация Субату, С.-В. Индия, ранний эоцен (ипр) 3. Формация Доманда, центральный Пакистан (лютет) 4. Формация Гаруди, С.-В. Индия (лютет) 5. Формация Мокаттам, северный Египет (лютет) 6. Формация Амеки, южная Нигерия (лютет) 7. Блю Блафф, известняки Санти, Джорджия и Ю. Каролина, США, средний эоцен (бартон). |
Обратим внимание, что эти формации расставлены в ряд по уменьшению
степени их древности – от самых ранних к более молодым. Пусть это будет своеобразной
«осью» в дальнейших рассуждениях.
Далее в этой же главе приводятся данные о распространении разных групп древних
китообразных по характеру отложений.
Пакицетиды: красноцветные пласты низов формации
Кулдана. Нижняя Кулдана состоит из красных аргиллитов, известняков, песчаников
и конгломератов. Литофации истолкованы как включения почв речной долины,
переработанные в речных руслах во время эпизодических разделений русла.
=> пакицетиды населяли речные местообитания.
Амбулоцетиды: верхи формации Кулдана. Верхняя Кулдана состоит из перемежающихся сланцев, мергелей и известняков со следами эвапоритов. Переход от нижней к верхней отмечен пёстрыми сланцами, интерпретируемыми как пресноводные отложения. Обширная последовательность приливных отложений в засушливых местообитаниях. => Амбулоцетиды населяли приливные области с сильным влиянием пресных вод. Ремингтоноцетиды: формации Доманда и Гаруди (самые ранние – в формации Кулдана – Кохат, переслоенной тонкими аргиллитными известняками и известковыми сланцами, содержащими нуммулитов, двустворчатых и брюхоногих моллюсков. Похоже на заливаемое приливами прибрежное местообитание амбулоцетид). Доманда и Гаруди переслоены красно-коричневыми и зелёными сланцами, иловыми мергелями, тонкими известняками, богатыми устрицами. Интерпретируется как удалённые от берега морские мелководья, переходящие в дальние богатые карбонатами среды обитания. => дифференциация местообитаний между ремингтоноцетидами и протоцетидами??? Более вероятно, ремингтоноцетиды и протоцетиды населяли прибрежные морские воды с некоторым влиянием пресных вод. Протоцетиды: формация Мокаттам (также Гиши, Маади) и Блю Блафф – часть Мокаттам: ровное мелководное карбонатное местообитание (нуммулитовый известняк с небольшим количеством моллюсков, мшанок и морских ежей), регрессивное. Типичнейшая ассоциация с мелководными морскими шельфовыми отложениями, удалёнными от берега. Блю Блафф представляет собой песчанистые аргиллиты, богатые устрицами мергели; известняки Санти – это мергели и более чистые карбонатные отложения. Они представляют собой мелководные морские местообитания недалеко от берега, возможно, с приливным влиянием. => протоцетиды также населяли мелководные прибрежные и удалённые от берегов местообитания. |
В течение эоцена китообразные совершили резкий скачок в морфологических
изменениях: из четвероногих наземных животных они превратились в полностью
водные формы, совершенно не похожие внешне на свои наземных предков и родственников.
Возможная причина этого явления – отсутствие конкурентов в новой среде обитания.
На границе мелового периода и палеогена произошло массовое вымирание крупных
животных на суше (знаменитое исчезновение динозавров). Но последствия массового
вымирания были ещё более страшными для морских организмов. Большинство крупных
животных морей мезозоя, как позвоночных, так и беспозвоночных, вымерло. Для
палеоцена неизвестны ископаемые остатки несомненно морских млекопитающих, хотя
точно известно о существовании специализированных морских птиц – первых пингвинов.
Зато благодаря недавним исследованиям более поздние эоценовые отложения явили
научному сообществу прекрасно документированную историю покорения моря первыми
китообразными.
Креационисты не хотят смириться с таким положением дел, и всеми силами стараются
низвести значение обильного ископаемого материала, ставшего известным в наши
дни. Относительно взглядов на возраст этих животных Харун Яхья в работе A
Whale fantasy from ‘National Geographic’ пишет
следующее:
Пока мы довольно долго рассматривали небольшие, но вводящие в заблуждение добавления к реконструкциям животных в схеме. Наряду с этим даты, приписанные им ‘National Geographic’ были отобраны в соответствии с предубеждениями дарвинистов. Животные демонстрируются как следующие одно за другим на геологической линии, тогда как это под вопросом. Ashby L. Camp разъясняет ситуацию, основываясь на палеонтологических данных: «Согласно стандартной схеме Pakicetus inachus датируется поздним ипром, но некоторые эксперты признают, что он может датироваться ранним лютетом. Если принимается более поздняя датировка (ранний лютет), тогда Pakicetus будет почти, если не полностью, современником Rodhocetus, ранне-лютетского ископаемого из иной формации Пакистана. Более того, возраст Ambulocetus, который найден в той же самой формации, что и Pakicetus, но на 120 метров выше, должен быть пересмотрен в сторону уменьшения на ту же величину, что и у Pakicetus. Это сделало бы Ambulocetus моложе, чем Rodhocetus и, возможно, моложе, чем Indocetus и даже Protocetus». |
Собственно, вот отрывок из статьи Эшби Л. Кампа The Overselling of Whale Evolution («Распродажа эволюции китов»), написанной в 1998 году, который более подробно раскрывает претензии креационистов к возрасту образцов ископаемых китообразных:
Образуют ли ранние археоцеты последовательность
или ряд переходных форм, зависит, конечно, от их морфологии и их стратиграфичского
положения. Претензии состоят в том, что для каждого из этих ископаемых
степень эволюционной продвинутости соответствует стратиграфическому положению.
Иными словами, чем старше окаменелость, тем менее продвинутые её признаки;
чем моложе окаменелость, тем более продвинуты её признаки. Это и есть
то соответствие формы и положения (возраста), которое создаёт впечатление
направленного изменения с течением времени.
Общепринятый порядок видов археоцетов, в понятиях как морфологического (от примитивных к продвинутым), так и стратиграфического (от ниже/старше к выше/моложе) критериев, следующий: Pakicetus, Ambulocetus, Rodhocetus, Indocetus, Protocetus, и Basilosaurus (...) Одна из проблем этой аккуратной картинки состоит в том, что стратиграфические отношения большинства из этих ископаемых неоднозначны. Согласно стандартной схеме, Pakicetus inachus датируется поздним ипром, но некоторые эксперты признают, что он может датироваться ранним лютетом. Если принимается более поздняя датировка (ранний лютет), тогда Pakicetus будет почти, если не полностью, современником Rodhocetus, ранне-лютетского ископаемого из иной формации Пакистана. Более того, возраст Ambulocetus, который найден в той же самой формации, что и Pakicetus, но на 120 метров выше, должен быть пересмотрен в сторону уменьшения на ту же величину, что и у Pakicetus. Это сделало бы Ambulocetus моложе, чем Rodhocetus и, возможно, моложе, чем Indocetus и даже Protocetus. По стандартной схеме Protocetus датируется средним лютетом, но некоторые эксперты датировали его ранним лютетом. Если принимать более старый возраст (ранний лютет), то Protocetus станет современником Rodhocetus и Indocetus. В этом случае то существо, которое, как считается, было полностью морским археоцетом, уже будет на сцене в то же время, или около того времени, когда археоцеты впервые появляются в летописи окаменелостей. Та важность, которой эволюционисты наделили эти окаменелости в своём сражении с креационистами, не сможет помочь, но вызовет удивление: неужели их стратиграфическое положение в стандартной схеме вызвано влиянием их морфологии? Тот, кто склонен признавать эволюцию, должен бы менее критично относиться к возрасту, который выстраивает эти окаменелости в морфологическую последовательность, но более критично – к возрасту, который нарушает эту последовательность. Как показывает разнообразие и чередование мнений экспертов, стратиграфическое соответствие – это искусство в гораздо большей степени, чем обычно оценивается. Опираясь на вышесказанное, будет оправданно считать, даже внутри рамок, очерченных эволюционистами, что все ранние археоцеты были по большей части современниками. |
Для пакицетуса указывался гораздо больший возраст: череп, найденный Джинджеричем, изначально оценивался даже в 53 миллиона лет. Понятно, что цифры возраста отложений берутся не «с потолка», а являются результатом кропотливого анализа различных показателей. Филип Джинджерич занимался, в том числе, изучением возраста формаций, в которых найдены ранние китообразные, и в настоящее время придерживается взглядов о более молодом возрасте пакицетуса. Вот выдержка из резюме к его работе «Стратиграфические и микропалеонтологические ограничения среднеэоценового возраста формации Кулдана в Пакистане, содержащей ископаемые остатки млекопитающих» (2003 год):
Формация Кулдана из Пакистана наиболее известна своими ископаемыми млекопитающими, включая примитивных китообразных, таких, как Pakicetus и его близкие родственники. Млекопитающие формации Кулдана оценивались по возрасту в различных исследованиях как раннелютетские (нижний средний эоцен), позднеипрские (верхний ранний эоцен), или, в настоящее время, промежутком времени от большей части ипра до раннелютетского времени (от нижнего раннего эоцена до нижнего среднего эоцена). Формация Кулдана – это сравнительно тонкая, 20 – 120 метров толщиной, прослойка континентальных красноцветных отложений, сформировавшаяся в период низкого уровня моря и лежащая внутри намного более мощной последовательности морских формаций, богатых фораминиферами. Планктонные и мелководные бентосные фораминиферы ограничивают возраст формации Кулдана верхним ранним эоценом или нижним средним эоценом, и текущая интерпретация стратиграфии мирового уровня океана поддерживает последнюю. Малая продолжительность времени существования низкого уровня моря, в котором найдены млекопитающие Кулданы, означает, что различия между образцами, возраст которых оценивается, возможно, представляют собой скорее различия в местных условиях обитания, участках осадконакопления и выборках, чем какое-то существенное различие в возрасте. |
Полный текст работы (с платным доступом) находится здесь. В описании ещё одного вида ранних китообразных, Himalayacetus subathuensis, авторы, Сунил Баджпаи и Филип Джинджерич уточняют возраст Pakicetus:
По возрасту Pakicetus считался поздне-раннеэоценовым, когда
был впервые описан (...), поскольку красноцветные отложения, в которых
он найден, интерпретировались как представляющие период низкого уровня
моря (...). В то время признавался один период понижения уровня моря
при переходе от раннего к среднему эоцену (...). Он был в конце раннего
эоцена; красноцветные отложения с костями Pakicetus, как считалось, представляли
это время снижения уровня моря, и Pakicetus считался раннеэоценовым
по возрасту. |
Нарушает ли «омоложение» Pakicetus картину, построенную
палеонтологами? Естественно, можно ожидать того, что время существования разных
родов китообразных
может
перекрываться.
Этому
есть
очень простое и непротиворечивое
объяснение:
после отделения от предковой группы живых организмов какой-либо дочерней группы,
предковая группа не прекращает своего существования в полном составе. Отделившаяся
от неё дочерняя группа существует сама по себе, осваивая иные местообитания
(собственно, различия в образе жизни как раз и обусловили разделение некогда
единой предковой группы на несколько потомков), и до определённого момента
её существование не влияет
на выживаемость
представителей предковой группы. Поэтому после отделения от пакицетид ветви,
ведущей к животным, ещё более приспособленным к водному образу жизни, пакицетиды
продолжали существовать в привычных местообитаниях, не конкурируя с ними. Отсюда
и возможный более молодой возраст пакицетуса – эволюционное событие на тот
момент могло уже свершиться, а пакицетусы спокойно доживали свой век, занимая
иную экологическую нишу и не конкурируя со своими потомками. Эволюционисты
формулируют «проблему», придуманную креационистами, просто: «если вы живёте
сейчас, почему живы ваши родители?».
Нельзя исключать и того, что Ambulocetus также был найден в
более поздних отложениях, чем он фактически появился на Земле
как самостоятельный таксон: эти два события, как понимаете, не одно и то
же, и появление ископаемых остатков того или иного вида в летописи окаменелостей
всегда «запаздывает» по отношению к фактическому появлению в природе
вида, который пока не достиг
своего расцвета
и максимальной
численности. Тогда при оценках времени дивергенции придётся оценивать по целому
комплексу признаков
строения
скелета,
когда от
него мог
дивергировать иной потомок, и какие черты специализации Ambulocetus приобрёл
уже после этого разделения. Пока найдено сравнительно немного остатков амбулоцетуса:
имеется
в виду количество экземпляров, а не полнота образца, которая для известного
вида очень хорошая и демонстрирует ключевые особенности этого животного. Они
относятся к отложениям примерно одного и того же возраста, поэтому нет уверенности,
что найденные экземпляры представляют собой именно животных,
самых древних в своём роду, и вообще нельзя сказать, насколько долго Ambulocetus существовал
в истории Земли как самостоятельный таксон. Ведь по единственной находке или
по
находкам из одного и того же слоя отложений (фактически, современников) весьма
сложно предположить,
насколько
долго
мог существовать
вид до его появления в летописи окаменелостей.
И ещё один вопрос возникает... Джинджерич утверждает, что формация Кулдана образовалась
в течение сравнительно короткого отрезка времени. Разница между возрастами Pakicetus и Ambulocetus составляет
один миллион лет (см. цитату из Харуна Яхьи). Если Pakicetus из
формации
Кулдана стал «моложе» на два миллиона лет, это означает, что возраст
Ambulocetus из той
же формации должен составлять около 47 миллионов лет, если следовать
логике Дона
Баттена. Но означает ли это, что и в других местах находки указанных животных
будут иметь тот же самый возраст? Кроме того, целесообразно ли «разводить» на
целый миллион лет животных, одно из которых найдено в верхах, а другое в нижней
части формации, если специалисты считают, что она сама по себе сформировалась
в
течение
очень
короткого
времени?
Эдвард Т. Бабински, обсуждая проблему сосуществования и перекрывания в пространстве
и времени предковых
и
потомковых
разновидностей китообразных,
говорит
следующее:
То, что все вышеуказанные существа собраны вместе
в геологическом времени, с черепами сходной формы и ушными костями переходной
формы, не похожими на кости современных китов, и что все они были млекопитающими,
приспособленными в различной степени к водной среде обитания, и что все
они предшествовали современным китообразным, говорит громче, чем акцентирование
автора на разногласиях в отношении датировки, создающее прецедент для
креационизма. Карл Зиммер добавляет: |
Креационисты слишком прямо сравнивают родословное древо живых организмов с настоящим деревом. Они сетуют: мол-де, находим только «листья», «боковые ветви», но почему-то не видим ствола. И не понимают, что «ствол» и «скелетные ветви» родословного древа (в отличие от дерева, растущего на улице) – это и есть самые тонкие его части. А где тонко – там и рвётся. Переход от одной группы животных к другой – это практически единичный акт, совершавшийся немногими популяциями внутри вида-родоначальника. А ископаемые остатки животных – это свидетельство существования широко распространённых форм, добившихся определённого успеха в борьбе за существование и обладавших какое-то время достаточно высокой численностью, что позволило с некоторой степенью вероятности произойти даже такому редкому и случайному явлению, как захоронение. Такие распространённые формы, как правило, отличаются некоторыми специализациями, уклоняются от «главной дороги» и не могут в силу этого быть признанными предками более поздних форм. Ископаемые остатки показывают время, когда вид живых организмов добился значительного эволюционного успеха, и стал особенно многочисленным. Но они ни в коем случае не показывают точной даты его появления или вымирания. Например, уже появляются отдельные свидетельства о существовании динозавров в палеоцене (не повсеместно, но лишь в нескольких местонахождениях), а в летописи кистепёрых рыб зияет колоссальный пробел между современным целакантом и меловыми представителями этой группы.
Работа палеонтолога по установлению родственных связей и построению
родословного древа какой-либо группы живых организмов больше похожа на работу
следователя: конечный результат неизвестен, и при построении целостной картины
событий приходится приводить в соответствие множество фактов, зачастую полученных
косвенным путём. И каждый новый факт, получаемый позже, может либо подтвердить
построенную картину, и даже заполнить оставленную для него «рамку без портрета»,
или же, напротив, частично или полностью разрушить предыдущие построения,
заставляя пересматривать ранее сделанные выводы и повторять анализы и пр. Наука
не застрахована от этого, и это вполне нормальный процесс познания мира. Для
сторонников креационизма, имеющих дерзость примазываться к науке, такой способ
познания мира не годится. У них уже имеется конечный вывод, представляющий
собой незыблемую религиозную догму, и «научное познание мира» сводится у них
к поиску и отбору фактов, хотя бы частично соответствующих желаемому результату.
При этом груда фактов, противоречащих таковому, просто отбрасывается.
Держа в памяти слова Карла Зиммера, приведённые выше, следует признать, что палеонтология
преподносит порой крайне «неудобные» находки. Одной из таких находок был новый
вид примитивнейших китообразных из Индии, получивший название Himalayacetus
subathuensis – «гималайский кит из формации Субату». Находка представляет
собой обломок левой ветви нижней челюсти с двумя коренными
зубами
– 2-м и 3-м. Описание нового рода и вида археоцетов было сделано в 1998 году.
Возраст этого китообразного определён по присутствию бентосного простейшего Nummulites
atacicus, которое указывает на середину раннего эоцена, около 53 миллионов
лет
назад. Pakicetus, род китообразных, прежде признававшийся самым древним,
отошёл на второй план. Но это было сделано не только потому, что его возраст
оценивался лишь в 50 миллионов лет. Это же самое исследование возраста
отложений, которое дало столь древнюю оценку для Himalayacetus, и одновременно
«омолодило» Pakicetus.
Описание Himalayacetus сделано Сунилом Ваджпаи и Филипом Джинджеричем:
Himalayacetus subathuensis – новый пакицетидный
археоцет из формации Субату на севере Индии. Типовой образец зубов имеет
маленький канал нижней челюсти, указывающий на отсутствие специализаций
органов слуха, отмеченных у более продвинутых китообразных, и у него Pakicetus-подобные
коренные зубы, предполагающие, что он питался рыбой. Himalayacetus важен
тем, что это самый древний известный археоцет, и тем, что он был найден
в морских отложениях, связанных с морской фауной. Himalayacetus продлевает
летопись окаменелости китов приблизительно на 3,5 миллиона лет назад
в геологическом времени, к средней части раннего эоцена [53,5 миллионов
лет назад (млн. л. н.)]. Кислород в фосфате зубной эмали имеет соотношение
изотопов, промежуточное между значениями, указанными для пресноводных
и морских археоцетов, показывая, что Himalayacetus, вероятно,
проводил некоторое время в обеих средах. Когда временной диапазон Archaeoceti
калиброван радиометрически, сравнение вероятностей ограничивает время
происхождения Archaeoceti и, следовательно, Cetacea приблизительно 54
– 55 млн. л. н. (начало эоцена), откуда следует, что их расхождение с
ныне живущими Artiodactyla, возможно, произошло уже 64 – 65 млн. л.
н. (начало кайнозоя). … Новый пакицетид был найден приблизительно на 100 м ниже стратиграфически и на 3,5 миллиона лет старше геологически, чем эквивалент Кулдана – Калакот, интервал красноцветных отложений верхней Субату, содержащий остатки Pakicetus в других местах. Это не только продлевает летопись окаменелостей Cetacea назад во времени, но также поддерживает идею о том, что киты возникли на берегах Тетиса и подкрепляет интерпретацию пакицетид как начальной земноводной стадии эволюции китообразных, заселивших Тетис, чтобы питаться рыбой. |
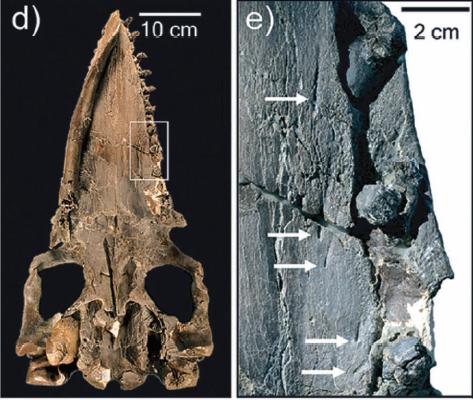
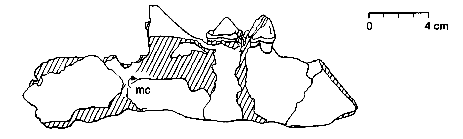
Образец челюсти Himalayacetus. MC - mandibular channel (нижнечелюстной канал).
Что ж, в принципе, находка более раннего возраста, нежели Pakicetus, вполне ожидаема. В статье Ханса Тевиссена и др. в журнале Nature (в той самой, где приводится реконструкция скелета этого животного как волкообразного существа с длинным хвостом) упоминается, что, помимо Pakicetus, экспедиция обнаружила остатки скелетов Nalacetus и Ichthyolestes, которые были его современниками. А это уже само по себе предполагает, с эволюционной точки зрения, что у них должен быть некий общий предок, живший ранее.
Himalayacetus имеет общую форму коренных зубов, характерную для Archaeoceti. Он несомненно является пакицетидом, потому что у него отсутствует увеличенный нижнечелюстной канал, заметный у всех более продвинутых археоцетов (рис. 1). Himalayacetus похож размерами на более позднего Pakicetus (…) но отличается наличием более низких коренных зубов с большим и более округленным главным бугром (протоконидом) на тригониде и меньшим и менее приподнятым малым бугром (гипоконидом) на талониде (рис. 2). Himalayacetus отличается от Pakicetus, Gandakasia (…), и Ambulocetus (…) наличием антеромедиального бугра в основании тригонида, проецирующегося в более переднем направлении, чем антеролатеральный бугор. |
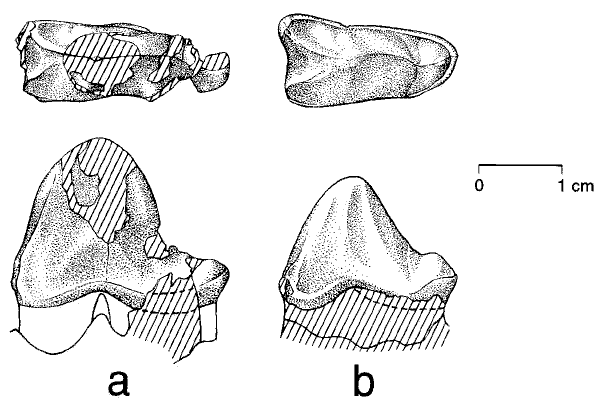
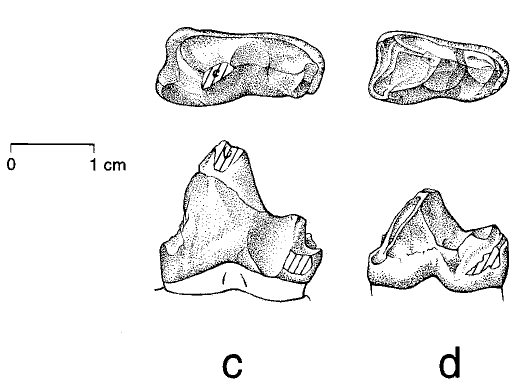
Зубы Himalayacetus в сравнении с зубами Pakicetus inachus и P. attocki:
A) Коронка второго левого коренного зуба Himalayacetus
B) Коронка третьего левого коренного зуба Himalayacetus
С) Коронка второго левого коренного зуба Pakicetus attocki
D) Коронка третьего левого коренного зуба Pakicetus inachus
Пока от Himalayacetus известна только фрагментарная нижняя челюсть, и о его облике можно сказать только то, что это было животное размером приблизительно с пакицетуса. Поэтому о строении ушной области животного, к сожалению, ничего нельзя сказать. Зубы животного демонстрируют сходство с зубами более поздних китообразных.
Himalayacetus также важен тем, что показывает,
что некоторые киты, вероятно, были отчасти морскими очень рано в ходе
эволюции китообразных. Himalayacetus происходит из мелководных
морских отложений, содержащих устриц, тогда как Pakicetus и
другие ранее известные древнейшие пакицетиды происходят из континентальных
красноцветных отложений и были найдены совместно с наземными млекопитающими
(…). Изотопы кислорода отличают ныне живущих пресноводных Cetacea от
морских (…). Кислород в пресноводной обстановке разделяется на фракции
– обогащён лёгким 16O и обеднён тяжёлым 18O – по
сравнению с кислородом в морском окружении, в котором сохраняется более
тяжёлый 18O (рисунок 4). Относительно изотопов кислорода
у эоценовых Pakicetus и Nalacetus сообщалось,
что их соотношение было характерно скорее для пресноводной, чем для морской
среды значение, что привело Тевиссена и др. (…) к заключению, что пакицетиды,
геологически древнейшие киты, не были морскими. Интерпретация усложняется
из-за того, что изотопный состав кислорода в фосфате костей и зубов наземных
млекопитающих часто тяжёлый (…), отражая фракционирование в течение дыхания
и других метаболических процессов. Таким образом, значения содержания
тяжёлого изотопа, наблюдаемые у Himalayacetus и по крайней мере
некоторых [образцов] Pakicetus можно объяснять дыханием на суше
животных, которые почти наверняка всё ещё были земноводными, получением
с пищей некоторого количества тяжёлого кислорода из морской воды во время
питания, или обоими способами. Жизнь на суше соответствует находкам всех Pakicetus совместно
с наземными млекопитающими, а получение с пищей тяжёлого кислорода в
морской воде совместимо с открытием Himalayacetus совместно
с морскими устрицами и другими моллюсками. Интерпретация всё ещё неоднозначна,
но две линии подтверждений – находка в морских отложениях и соотношение
изотопов кислорода в пользу тяжёлого – говорят в пользу вторжения в
морские воды на ранних этапах эволюции китообразных. Происхождение китов
обычно
объясняется доступностью рыбы как пищи, на высокопродуктивных морских
мелководьях восточного Тетиса (…), и ранние пакицетиды, вероятно, питались
рыбой в пресноводных реках, солоноватоводных эстуариях, и на морских
мелководьях.
|
Вот схема из того же описания, демонстрирующая результаты исследования содержания изотопов кислорода в эмали ископаемых и ныне живущих китообразных.
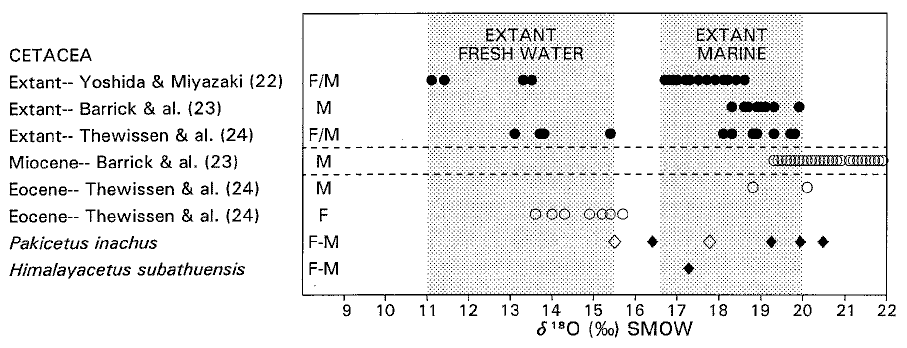
Рисунок 4. Соотношение изотопов в фосфате
костей и зубов ныне живущих и ископаемых китов. Современные киты, живущие
в пресной воде, имеют значение содержания лёгкого кислорода в пределах
11 – 15,5 (частиц на тысячу δ 18O СЗОВ), тогда как
для китообразных, живущих в морской воде, значение содержания тяжёлого
изотопа находится в пределах 16,5 – 20 ([чёрный кружок] и
заштрихованные области). У миоценовых китообразных значение содержания
тяжёлого кислорода в пределах 19 – 33 ([светлый кружок]).
Сообщается о немногих эоценовых китообразных, и они делятся на пресноводные
и морские формы ([светлый кружок]). Значения для среднеэоценового P.
inachus варьируют от 15,5 для молодых особей ([светлый ромб])
до 20,5 для взрослых ([чёрный ромб]; А. Госвами и Ф. Д. Джиндж.,
неопубликованные сведения). Для фосфата зубной эмали H. subathuensis из
середины раннего эоцена это значение составляет 17‰ δ 18O
Стандартного Значения для Океанской Воды (СЗОВ), и потому тяжелее,
чем кислород,
найденный в фосфате ныне живущих и эоценовых пресноводных китобразных. |
На рисунке английские буквы обозначают: F – пресноводные (freshwater), M – морские (marine). Источники информации, на которые ссылаются авторы описания:
22. Yoshida, N. & Miyazaki, N. (1991) J. Geophys.
Res. 96, 815–820.
23. Barrick, R. E., Fischer, A. G., Kolodny, Y., Luz, B. & Bohaska, D.
J. (1992) Palaios 7, 521–531.
24. Thewissen, J. G. M., Roe, L. J., O’Neil, J. R., Hussain, S. T., Sahni,
A. & Bajpai, S. (1996) Nature (London) 381, 379–380
Разрушает ли морской образ жизни примитивного Himalayacetus логичную
цепочку форм, ведущую от пресноводных и солоноватоводных животных к полностью
морским формам? В некотором роде, да – среди ожидаемых ранних
пресноводных форм неожиданно объявляется морская. Но это лишь дополняет наше
знание о ранней эволюции этих животных. К тому же, переход к жизни в море возможен
без радикальных изменений строения скелета. Так, среди множества выдр, распространённых
на всех материках, кроме Австралии и Антарктиды, есть настоящий морской
житель, калан, отличающийся от прочих выдр не больше, чем сами пресноводные
выдры друг от друга – это представитель лишь ещё одного
рода внутри разнообразного семейства куньих. А другой морской житель, белый
медведь, вообще принадлежит
к тому же роду Ursus,
что и вполне наземные медведи. Пока посткраниальный скелет (и даже сам по себе
череп) Himalayacetus неизвестен, преждевременно делать какие-то выводы
относительно степени его приспособленности к водному образу жизни. Возможно,
это такой же «белый медведь» среди пакицетид, морской скорее по поведению,
нежели по морфологии. Но так это, или нет – покажут
находки в будущем.
Здесь хочется лишний раз попенять креационистам. Учёные не
скрывают своих методик изучения ископаемых образцов, и результаты исследований
можно найти
как в открытом доступе, так и в статьях, распространяемых за некоторую плату
(это, ИМХО, политика не столько учёных, сколько издателей). И в свете этого
утверждать, что результаты, мол, подгоняются под предубеждение – это как в
русской народной сказке лиса перемазывала мёд со своего живота на живот медведю.
Только она-то убедила медведя, что он сам свой мёд и съел, а в случае с креационистами
я успехов в таком убеждении не вижу. Хотя... Отдельные товарищи на форумах
пытаются оперировать сведениями, заимствованными из креационных источников.
Но у специалистов такие аргументы неизменно вызывают лишь здоровый смех, а
креационистское умопостроение легко разрушается при помощи твёрдых фактов. В
этом случае возражений, как правило, больше не поступает.
Оригинальное название статьи с описанием Himalayacetus subathuensis:
A new Eocene archaeocete (Mammalia, Cetacea) from India and
the time of origin of whales
SUNIL BAJPAI AND PHILIP D. GINGERICH
Джон Вудморапп, “Walking whales, nested hierarchies, and chimeras: do they exist?”:
Образуют ли ископаемые киты цепь переходных форм?
Ранее требовалась находка «переходных форм ископаемых китов». Недавний
номер ‘National Geographic’ привлекает внимание читателя
к множеству предполагаемых тенденций в эволюции китообразных. Это:
1) Всё большая
специализация к водному образу жизни; 2) Подводный слух, 3) Сокращение
размера задних конечностей, и 4) перемещение nares (ноздрей) на
заднюю часть верхней (дорсальной) поверхности черепа. Предполагаемая
тенденция
к подводному слуху заслуживает некоторого внимания, и более детально
обсуждена ниже. |
Обвинение серьёзное, особенно в свете того, что именно креационисты
любят непросвещённую аудиторию, и излагают свои аргументы в надежде на то,
что их собственные подтасовки не будут замечены. Тем не менее, давайте взглянем,
насколько хорошо (или плохо?) китообразные демонстрируют изменения, связанные
с переходом от наземных животных к полностью водным формам. Я должен заметить,
что ни в одной статье креационистов ложность тенденций, выделяемых эволюционистами,
почему-то не разбирается подробно, прямо по видам животных. Впрочем, это их
проблемы.
На останках китообразных хорошо прослеживаются определённые тенденции в изменении
формы тела. Об этом хорошо сказано в обзоре книги “The
Emergence of Whales: Evolutionary Patterns in the Origins of the Cetacea” (глава
11), откуда я далее буду брать сведения об особенностях строения
ископаемых китов. Автор данной главы - Emily A Buchholtz, Dept. of Biological
Sciences, Wellesley College, Wellesley, MA. Итак:
... «Посткраниальные скелеты ранних китообразных, хотя и являются почти исключительно фрагментарными, на удивление информативны и могут дополнить морфологические свидетельства рациона, специализации органов чувств и изменения в строении черепа (!!), заполняя некоторые пробелы в нашем понимании драматичного перехода (!!) ранних китообразных с суши в воду». ... «Китообразные обладают большим набором приобретённых признаков, связанных с морским образом жизни». [в обзоре Джеймса Акера приведены цитаты из оригинальной работы - В. П.] Сравним с мезонихидами. По всей видимости, Pachyaena ossifraga (...) является здесь лучшим выбором - в статье Джинджерича о Rodhocetus в Nature есть схема скелета. Сведения о позвоночном столбе P. ossifraga: 7 шейных , 12 грудных, 7 поясничных, 3 тазовых и 15+ хвостовых позвонков. Особенности передних шейных позвонков указывают на поддержание крупной головы. Далее следует много деталей, включая указание на широкие пределы подвижности хвоста. Китообразные сильно отличаются! Строение позвоночного столба Delphinus delphis (дельфина, конечно): 7 шейных, 13 грудных, 22 поясничных и 31 хвостовой позвонок. (И креационисты говорят, что эволюция не добавляет ничего нового! Как насчёт 15 новых поясничных позвонков?) Шейные позвонки очень короткие (у P. ossifraga длинные шейные позвонки). |
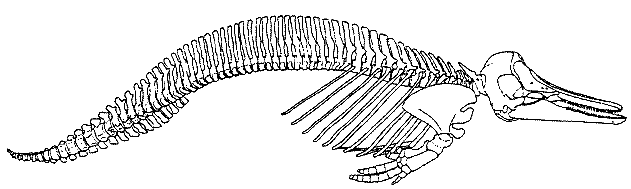
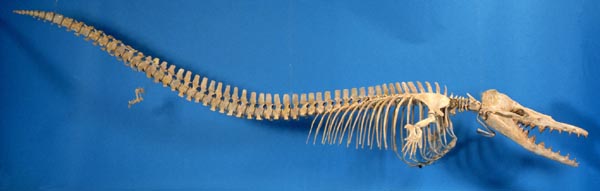
Скелет современного тусклого дельфина Lagenorhynchus obscurus демонстрирует существенные изменения скелета китообразных по сравнению с другими отрядами млекопитающих. Обратите внимание, насколько плавно происходит изменение формы позвонков от шейных к хвостовым. Границы между отделами позвоночника практически не выражены. Рудименты задних конечностей не были показаны в источнике иллюстрации.
“The Emergence of Whales: Evolutionary Patterns in the Origins of the Cetacea” (глава 11):
У китообразных больше позвонков, чем у их наземных предков; увеличение [их числа] распределено по разным функциональным отделам позвоночника неравномерно. Интересно, что Слайпер (Slijper) (1936), рассматривая только Protocetus, наземных млекопитающих и китообразных, которых он считал «примитивными», предсказал, что у «предкового кита» должно быть 7 шейных, 17-19 нешейных предтазовых позвонков (12-13 грудных, 5-6 поясничных) и 2 крестцовых позвонка. Цитата: «Шестьдесят лет и много открытий ископаемых форм спустя, это предсказание было в значительной степени подтверждено». |
Набросившись на Слайпера (см. часть 1), Джонатан Сарфати, наверное, упустил из виду это дивное предсказание специалиста. А может быть, и не упустил, но просто не захотел об этом говорить своим читателям. Я не знаю. Чужая душа - потёмки, а я не телепат, чтобы наверняка знать, что именно думал Сарфати, читая (надеюсь!) работы Слайпера при подготовке своих статей.
3. Устройство позвонков и локомоция археоцетов
… 3.1 Ambulocetus natans От скелета имеется только несколько разрозненных позвонков. Потому их общее количество неизвестно; грудных больше, чем поясничных, крестцовые сочленены с сильным тазом. Длина тел шейных позвонков относительно большая. Тело одного измеренного поясничного позвонка более широкое и заметно более длинное, нежели высокое. Предположение Тевиссена о «плавании с помощью ступней» (…) поддерживается строением известных позвонков. Вероятно, только частично водный образ жизни; меньше водных и больше наземных навыков, чем у ныне живущих тюленей». |
Немного прервусь с цитатой из обзора, и приведу цитату с сайта Эдварда Бабински, разбирающего статью Дона Баттена «Кит из сказки?», опубликованную на сайте “Answers in Genesis”:
«Автор утверждает: «… нет никакого свидетельства
развития горизонтальных хвостовых лопастей [у Ambulocetus], столь характерных
для китов.»
Эволюционисты не ожидают, что у Ambulocetus были хвостовые лопасти: «В отличие от современных китообразных, Ambulocetus имел длинный хвост и потому, вероятно, не имел хвостовых лопастей». (Thewissen, Science, январь 1994, стр. 211) (Ни один хрящ, который образует хвостовые лопасти китов, не сохраняется подобно костям) Поскольку мы говорим о хвосте Ambulocetus, позвольте мне добавить «обзор в обзоре» другой креационистской статьи, автор которой заявил, что «движения хвоста [самых ранних китов] начнут давить его репродуктивные органы об тазовые кости. Это привело бы к некоторому снижению сексуального влечения животного, и оно быстро потеряло бы интерес к воспроизводству – не очень положительный эволюционный шаг. Доведённое до крайности, это новое движение хвостом могло бы просто разрушить весь таз. Селективное давление окружающей среды или естественного отбора работало бы против любого такого изменения хвоста у наземных млекопитающих». Это «виляние хвостом» идёт с 1984 года, когда Гиш имел обыкновение показывать свои слайды коровы, эволюционирующей в кита, называя это «сбоем вымени» (Gish, 1985). Но даже до 1984 г. биологи знали, что «... гигантская речная выдра Pteronura) из Южной Америки обладает горизонтально расширенным хвостом, который производит толчки, даже при том, что хвостовые позвонки имеет типичную для наземных животных форму (Fish, 1998)». И речная выдра всё ещё не вымерла. «Кроме того, способ плавания этих выдр (дорсовентральные волнообразные изгибы позвоночного столба) дают очень хорошую аналогию метода плавания Ambulocetus. (Thewissen, 1998)».» [ссылки опущены – В. П.] |
Нет, люди... Хвосты китов решительно не дают спать креационистам... Во ещё одна цитата, из статьи Creationist mindblocks to Whale evolution («Креационистские препятствия для разума и эволюция китообразных», критика статьи Scientific Roadblocks to Whale Evolution by Frank Sherwin):
А теперь от плохо изученного к совершенно смешному…
«Ни у кого из предполагаемых наземных предков китов (копытные или хищники) не было вертикального движения хвоста. Тем не менее, китообразные (и предполагаемое связующее звено, Ambulocetus) способны к волнообразным движениям позвоночника вверх и вниз. Когда это случилось? Где окаменелости, документально подтверждающие, как движение из стороны в сторону хвоста наземных млекопитающих сменилось движением вниз и вверх у Ambulocetus и китов)?» (Шервин, 1998). Выглядит странным, что он хочет поговорить о движении позвоночника, и затем сравнивает его с движением хвоста. Для начала, позвоночный столб млекопитающих изгибается во время передвижения вверх и вниз, в противоположность изгибам из стороны в сторону у рептилий. Я должен согласиться с тем, что виляющий хвост млекопитающего представляет гораздо большую проблему. Мы все знаем, как больно должно быть кошке ходить с хвостом, поднятым трубой, или собаке, поджимающей хвост между задними лапами, не говоря уже об обитателях скотного двора, сгоняющих мух со своих спин. … Заявление Шервина о том, что «Совершенно случайное событие должно было бы одновременно создать эти горизонтальные хвостовые лопасти независимо, уменьшить тазовые кости и позволить деформированному наземному существу продолжать жить и даже процветать в море.» (Шервин, 1998) – это в лучшем случае «соломенное чучело», а в худшем – или следствие отсутствия знаний о предмете обсуждения или заранее намеченная попытка скрыть изобилие информации, известной об эволюции, палеобиологии и физиологии китообразных. |

Бразильская речная выдра Pteronura (фото с сайта Palaeos Vertebrates)
Можно добавить ещё немного креационных шуток юмора. Вот байка, которую приводит в своей статье «Киты: никаких признаков эволюции» Малком Боуден:
Промежуточное звено Есть одно забавное дополнение к этой проблеме. Много лет назад Дж. Хэлдейну (J.B.S. Haldane) было предложено составить ряд из морских животных, которые могли быть предками китов. Он представил следующую последовательность: морж – тюлень – дюгонь – кит. На первый взгляд это кажется правдоподобным, т.к. прослеживается возрастающая тенденция к жизни в воде. Однако все портит тот факт, что первые два млекопитающих передвигаются в воде, производя горизонтальные движения хвостом, тогда как два последних производят вертикальное движение. Так что, несчастному переходному животному между тюленем и дюгонем приходилось двигать хвостом под довольно странным углом[4]. ... [4] Dewar, D. and Merson Davies, L., Is Evolution a Myth? Evolution Protest Movement, 1957, p.22 |
А ещё наш зоркий друг Малком Боуден не разглядел того факта,
что у моржей и тюленей хвостишко совсем коротенький и не играет никакой
роли в передвижении под водой, а роль основного двигателя тела в
толще воды взяли на себя ластообразные задние лапы, работающие
попеременно (левой-правой, левой-правой...). Кроме
того, в ряд плотоядных животных как-то по-странному вклинивается вполне растительноядный
дюгонь, что,
сами
понимаете, не поддерживает такой картины. Я не могу сказать, в каком контексте
Холдейну было предложено построить такой ряд, поэтому не могу сказать, где
и насколько приврал Боуден. Однако, в ряду форм, предложенных Холдейном, есть
определённая закономерность. Морж – это бентофаг, проводящий много времени
на берегу (или на льду) и не удаляющийся на большое расстояние от твёрдой поверхности.
Он не удаляется с мелководья и не встречается над глубинами. Тюлень также связан
с берегом, но способен удаляться от берега надолго и охотится в толще воды.
Дюгонь – полностью водное животное,
не
выходящее
на сушу,
но
также
не удаляющееся
далеко от берега. И, наконец, киты полностью порвали с сушей и освоили пелагический
образ жизни. Они водятся и на мелководьях, и над большими глубинами, могут
кормиться на глубине или потреблять специфические виды корма, которые можно
найти лишь вдалеке от берегов (планктон, пелагические стайные рыбы). Иными
словами, для них нет препятствий для расселения в океане, кроме сугубо географических
и климатических. Так что в плане изменения экологических требований в
ряду предков китообразных схема Холдейна вполне правдоподобна, если опустить
тот
факт, что ранние киты были не бентофагами, как моржи, а хищниками: прослеживается
постепенная утрата связи с сушей и освоение пелагических местообитаний. Более
точно я ничего не могу утверждать, поскольку располагаю только той информацией,
которую нашёл у Боудена, а придумывать и додумывать за другого человека –
одно из самых неблагодарных занятий.
Говоря о способах передвижения Ambulocetus, целесообразно
добавить мнение Ханса Тевиссена о возможностях
передвижения амбулоцетуса по суше (с сайта Эдварда Т. Бабински):
В номере National Geographic от мая 1994 года рассказывается
о находке Ambulocetus в бывшем внутреннем море Пакистана. Тевиссен сомневается,
что древний ископаемый кит Ambulocetus natans был «ходячим плавающим
китом». Он объясняет, что на суше он двигался бы неуклюже, как морской
лев, и «он бы выглядел неповоротливым, но он всё ещё мог передвигаться».
Его команда обнаружила много окаменелостей, включая череп, который идентифицирует
ископаемое животное как китообразное. Позвоночный столб указывает, что
существо двигалось сходным с современными китами образом, используя заднюю
часть тела для движений вверх и вниз, тогда как задние конечности использовались
для толчков. Считается, что его передние конечности использовались, чтобы
рулить.
|
Поведение не сохраняется в ископаемом состоянии, мягкие ткани
тоже. Но о них можно судить по особенностям строения ископаемых остатков животного.
Главное чтобы каждый вывод учёных был подтверждён и обоснован реальными особенностями
и закономерностями. И не беда, если некоторые ранние гипотезы оказываются на
«свалке науки» (например, усиленно смакуемая креационистами гипотеза Ф. Джинджерича
о тюленеобразном облике пакицетуса) – в отличие от религии, в науке нет догм,
и всякие утверждения,
ранее считавшиеся
верными, могут быть опровергнуты. Но опровергнуты только одним способом – фактами,
а не личной прихотью.
Вернусь, однако же, к обзору Джеймса Акера. Итак, после амбулоцетуса тело китообразных
менялось
следующим
образом:
О родоцетусе стоит сказать немного больше. Претензии креационистов связаны с тем, что первоначально был описан неполный скелет этого кита. Вот выдержки из Интернет-ресурса Creation Wiki (далее CW), где обсуждаются аргументы с сайта TalkOrigins (далее TO):
CW: |
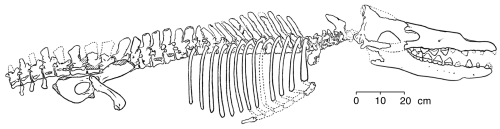
Исходный образец Rodhocetus; рисунок с сайта TalkOrigins. Честно говоря, в черепе животного мало дельфиньего, да и в позвоночнике тоже.

Реконструкция Rodhocetus, взятая с
сайта Palaeos Vertebrates.
Обратите внимание на год, указанный на рисунке.
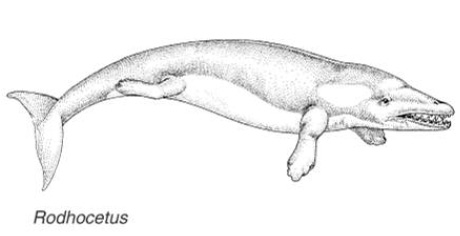
Эта «дельфинообразная» реконструкция Rodhocetus часто встречается на разных сайтах.
CW: |
А теперь вопрос на засыпку: что потеряла наука, когда выяснилось, что реконструкция родоцетуса, основанная на первоначальной находке, оказалась неточной? Думаю, ничего, кроме ошибочных воззрений и реконструкций. А что приобрела? Более точные сведения о строении данного животного, и к тому же дополнительное свидетельство о родстве китообразных и парнокопытных, заключающееся в «двухблочном» строении таранной кости. Плюс уточнённые представления о роли конечностей в передвижении животного. Если сравнить более старую «дельфиноподобную» реконструкцию, представленную выше, с более новой, «крокодилообразной», заметны различия в стиле плавания, предполагающемся для этих реконструкций. «Дельфиноподобный» родоцетус изображён с широким хвостовым плавником, что предполагает движение почти исключительно с помощью изгибов позвоночника в вертикальной плоскости. «Крокодилообразный» родоцетус передвигается не только с помощью изгибов позвоночника, но и с помощью движения конечностей, что и показано на рисунках, представленных ниже. Поскольку кончик хвоста для Rodhocetus неизвестен, сложно оценить вероятность появления у него хвостового плавника.
|
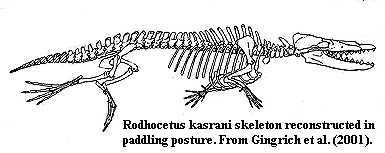
Реконструкция Rodhocetus kasrani в плавающем положении, созданная после открытия Джинджеричем костей конечностей этого китообразного. Напомню, что вывод о родстве китобразных и парнокопытных был сделан, в том числе, на основании изучения строения костей конечностей Rodhocetus. |
Современные представления об облике Rodhocetus. Не исключено, что они также могут быть неточными. Обратите внимание, что на реконструкции скелета этого животного (см. выше) хвостовые позвонки заштрихованы: точная длина хвоста Rodhocetus неизвестна, и потому сложно оценить, в какой степени хвост у этого животного был вовлечён в процесс плавания, и были ли у него хвостовые лопасти. Но крупные отростки позвонков свидетельствуют о мощной мускулатуре, которая приводила в движение хвост животного.
|
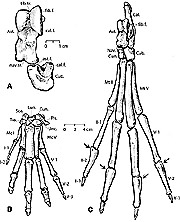
Исходя из пропорций скелета Rodhocetus, Джинджерич
с соавторами предположил, что это китообразное плавало с помощью движений задних
конечностей, как выхухоль.
В черепе родоцетуса заметны первые стадии изменений, которые наблюдаются в
черепе более развитых китообразных, в том числе современных. Предчелюстные
кости и нижняя челюсть животного удлиняются, и череп животного становится более
длинным, чем у ранних китообразных. Зубы также начинают постепенно упрощаться,
а коренные зубы имеют более высокую коронку, чем у более ранних видов китообразных.
Дифференциация зубов, характерная для млекопитающих, у Rodhocetus постепенно
стирается.
Примечательно положение ноздрей этого животного: из-за удлинения предчелюстных
костей положение ноздрей меняется по сравнению с ранними китами. Ноздри животного
располагаются над клыками – и это с учётом того, что резцы животного расположены
в передней части черепа с довольно большими промежутками.
В строении уха Rodhocetus также происходят изменения в сторону характерных
для современных китообразных: ушные кости состоят из характерной очень плотной
костной ткани. Однако в среднем ухе животного отсутствуют пазухи, характерные
для современных китов и связанные с приспособлением к направленному восприятию
звука под водой.
3.4 Protocetus atavus |

Реконструкция Protocetus
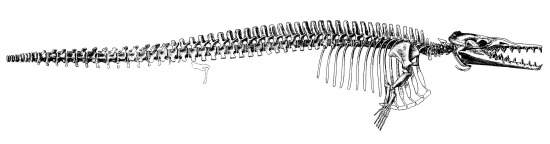
Скелет Zygorhiza. Не все виды семейства Basilosauridae были длиннотелыми змеевидными существами.
Снова позволю себе «припасть» к первоисточнику в лице Creation
Wiki. Вполне очевидно, что ряд китообразных, выстроенный там, не так полон,
как в книге “The
Emergence of Whales: Evolutionary Patterns in the Origins of the Cetacea”.
Товарищи могли бы очень красиво попенять составителям эволюционного ряда китообразных
с сайта TalkOrigins, что, мол-де, те скрывают правду от несведущих читателей,
и в свете этого замечания предстать практически положительными героями-просветителями.
Ан, нет. Цель не та: ряд форм китообразных получился бы гораздо более длинным,
плавным и красивым, что вредно повлияло бы на достижение поставленной цели
формирования в сознании читателя образа «эволюционного ряда китообразных» как
разорванной цепочки совершенно не относящихся друг к другу форм.
Но это лишь мои предположения. Истинный смысл «молчаливости» креационистов
в этом отношении остаётся тайной, скрытой в чужом сознании. А я не телепат,
чтобы утверждать что-либо за другого человека наверняка. Судя по логике
построения ряда китообразных, очередной фрагмент из анализа Creation Wiki надо
«втиснуть» в обзор Джеймса Акера где-то здесь:
CW: |
Для того, чтобы так утверждать, необходимо доказать, что половые органы (непосредственно отвечающие за воспроизводство у любого вида животных) прикреплялись не к тазовым костям, а к костям ног. Здесь видится довольно простое логическое построение: половые органы прикрепляются к тазовым костям (в чём всяк может убедиться), и задние конечности (то есть, ноги) прикрепляются к тазовым костям, но это ещё не говорит о том, что половые органы прикрепляются именно к ногам. Кроме того, строение органа в рассуждениях автора из Creation Wiki плавно подменяется его функцией. И от того, что ноги Prozeuglodon утратили функцию передвижения по суше, ещё не означает того, что они перестали быть задними конечностями. Впрочем, тема задних конечностей китообразных будет подробно рассмотрена далее.
3.7 Basilosaurus cetoides Насколько мы знаем, B. cetoides был полностью морской формой. Короткие шейные позвонки, отсутствие размерной неоднородности в тазовых позвонках, отсутствие крестцово-тазового сочленения указывает на отсутствие способностей к наземному движению. Характер длин позвонков иной, нежели у других археоцетов, с серией однородных по размерам позвонков. Показывает угреобразную форму плавания (подобно безногим ящерицам) с дорсовентральной ориентацией [изгибы тела не в горизонтальной, как характерно для рептилий, а в вертикальной плоскости – В. П.]. Это означает, что маневренность возобладала над скоростью. Маленькие хвостовой стебель и хвостовой плавник показывают, что волнообразные движения туловища обеспечивали основное усилие». |
Подводя итоги рассмотрения этих тенденций, автор обзора делает заключение:
4. ОБСУЖДЕНИЕ
… я собираюсь пропустить большую часть вспомогательного текста ради основных точек морфологических тенденций. 1. Сокращение числа крестцовых позвонков (из-за потери слиянения, что улучшает осевые волнообразные движения). 2. Потеря крестцово-тазового сочленения. Сохранение одного позвонка с тазовым сочленением (Rodhocetus) предполагает, что слияние крестцовых позвонков было утрачено до [потери] тазового сочленения. У животных с некоторыми способностями к наземной жизни возможно волнообразное движение [тела]. 3. Сокращение относительной длины шейных позвонков. Улучшает гидродинамику. 4. Удлинение туловища. Увеличивает диапазон волнообразного движения для большего усилия. 5. Смещение в заднем направлении точки максимальной амплитуды дорсо-вентрального движения. 6. Модификация хвоста для формирования хвостового стебля и хвостовых лопастей. Прежде всего обратите внимание, что Rodhocetus и Protocetus значительно различаются, нежели Georgiacetus, Zygorhiza, и Basilosaurus. Функциональные изменения в стиле плавания могут быть дополнительно добавлены к условной филогении археоцетов. Заметны две тенденции: разрастание позвоночного столба, представляющего туловище, совершающее волнообразные движения, и сдвиг назад точки максимума волнообразного движения со сменой поверхности, придающей толчок. Второй «набор» изменений заключается в пропорциях тела и имеет тенденцию (как Вы можете догадаться) к гладкой и обтекаемой форме тела (с сужением к хвосту). |
Подводя итог, замечу, что последовательность, в которой располагаются обсуждаемые
виды китообразных, полностью соотносится со временем их существования, а также
логически связана с характером отложений, в которых были найдены те или иные
виды китов. Иными словами, последовательность времени существования, усиление
анатомической специализации к водному образу жизни и смена мест обитания (которая
выводится из характера отложений, в которых находят различные виды ранних китообразных)
идут параллельно друг другу. Это более чем явное указание на эволюцию данных
животных.
Единственное, с чем эти данные не согласуются, так это с желанием креационистов
представить всё не как чёткую и закономерную последовательность, а как кучу отдельных
событий, никак не связанных друг с другом.
В свете современных открытий, значительно прояснивших раннюю
историю китообразных, стало очевидно, что креационизм как мировоззрение потерял
ещё один некогда сильный аргумент. Найденная учёными последовательность видов
древнейших китообразных отличается достаточной полнотой, чтобы можно было проследить
их эволюцию. Но у креационистов есть свой взгляд на подобные выкладки. Они
считают каждую из форм в ряду некоей отдельно взятой единицей, не имеющей ничего
общего ни с той, что стоит в ряду до неё, ни с той, что находится после.
На эволюционном древе китов есть точка, на которую направлены атаки креационистов.
Это точка перехода от археоцетов к современным группам китов. По мнению креационистов,
все разнообразные древние «киты с ногами» и киты без ног, относящиеся к группе
археоцетов, не имеют ничего общего с современными китами. То есть, современные
усатые и зубатые киты сотворены.
Эшби Л. Камп
«Распродажа эволюции китообразных» (The Overselling of Whale Evolution by
Ashby L. Camp), 1998:
От археоцетов к современным китам Третье громкое заявление в эволюционистской цепочке событий – это
то, что археоцеты дали начало современным китам. Это иногда преподносится
как факт, но родственные отношения между этими подотрядами долгое
время
были объектом обсуждения.
|
В работе Ashby L. Camp дана ссылка на книгу: George Gaylord
Simpson, “The Principles of Classification and a Classification of Mammals,”
Bulletin of
the American
Museum
of Natural
History 85 (1945). Свежачок-с... Эта работа особенно актуальна
в свете находок последних десятилетий. Нет, я не отрицаю её ценности в части,
не противоречащей современным взглядам, и я признаю её важной в аспекте истории
науки. Я понимаю, что Джордж Гейлорд Симпсон был высокообразованным специалистом,
причём такого уровня, до которого большинству креационистов просто по определению
не добраться. К сожалению, он уже умер, и потому неизвестно, как он оценил
бы современные находки архаичных китообразных. Так что, ребята-акробаты,
не обижайтесь, если вас снова обвинят в том, что вы под видом
новейшей научной
информации
пичкаете
читателя
откровенным старьём,
которое
опровержено самим развитием науки. В данном случае идёт «война цитат», а
не оперирование реальными фактами: из первоисточника просто вы(б, д)ирается
удобная фраза, звучащая
красиво и «убойно», вне учёта уровня знаний, имевшего место на момент
написания данной фразы, и вываливается на несведущего читателя как «официальная
точка зрения науки на настоящий момент».
Я не могу говорить за Дж. Г. Симпсона,
но могу лишь предположить, что он говорил о происхождении современных
групп китообразных непосредственно от археоцетов.
Пусть меня поправят более сведущие в этом вопросе люди, и я никогда не отказываюсь
вносить дополнения и изменения в свою работу, если их необходимость будет обоснована.
Далее
Эшби Л. Камп сообщает следующее:
Эта точка зрения была повторена снова двумя десятилетиями
позже А. В. Яблоковым, который писал: «Сейчас многим исследователям ясно,
что Archaeoceti не могут быть признаны прямыми предками современных китообразных».
До недавнего времени это было единодушное мнение.
|
Похоже, я всё-таки был прав... Эшби Л. Камп буквально в следующем после мнения Симпсона абзаце уже говорит немного по-другому. Просто «предки» превращаются уже в «прямых предков», что, согласитесь, не одно и то же. Между археоцетами и современными семействами китообразных укладывается целый ряд семейств китообразных, не доживших до наших дней, и вряд ли Яблоков об этом не знал.
Современные ведущие специалисты в этой области считают,
что археоцеты были предковыми для современных китов, но нет согласия
в том, какое семейство археоцетов участвовало в этом. Фактически, предполагались
все три семейства (Protocetidae, Remingtonocetidae и Basilosauridae).
Это особенно показательно, если знать, насколько радикально отличаются
ремингтоноцетиды от остальных археоцетов.
|
Вполне нормальное явление в науке, занимающейся поиском правильного ответа, а не подгонкой под него решения. Нельзя исключать того, что те же ремингтоноцетиды, своеобразные «гавиалы среди млекопитающих», были на каком-то этапе развития науки известны по неполным остаткам, и комплекс известных особенностей этих животных не противоречил мнению о происхождении от них более поздних китообразных. А затем новые находки ремингтоноцетид выявили специфические особенности их строения, исключающие возможность их перехода в состояние, характерное для более современных китообразных. Всегда надо понимать, что палеонтологи имеют дело не с цельными свежими и натуральными тушками животных, а с остатками остатков в буквальном смысле этого слова. Поэтому первоначальные выводы могут быть ошибочными. Ведь, как известно, «не ошибается тот, кто ничего не делает».
В дополнение к сказанному, не было выявлено ни одной
цепочки, ведущей от археоцетов к современным китам. Филогенетические
отношения между главными родословными линиями внутри отряда Cetacea продолжают
оставаться «очень плохо понятными», и вот почему в современных филогенетических
древах преобладают тупики, пунктирные линии и
знаки вопроса. Что касается Basilosaurus isis, то в общем случае установлено,
что Basilosaurinae были изолированным подсемейством, не имеющим отношения к происхождению
современных китов.
|
Естественно, а вы, господа, чего хотели? Летопись окаменелостей – это не банковский счёт, созданный именно с целью сохранения и преумножения. Никто из богов, включая Яхве, не станет гарантировать того, что до наших дней целиком и полностью сохранятся остатки всех когда-либо существовавших видов, в частности, китообразных. Выводы же о родстве делаются на основании имеющихся данных, и в разные моменты времени объёмы информации, на основании которой делаются выводы о родственных связях китообразных, различаются. А часть информации заведомо может быть утрачена навсегда. Поэтому в построениях родословных древ всегда будут присутствовать знаки вопроса, отражающие пробелы... нет, скорее, не в знаниях, а в летописи окаменелостей. Косвенные данные помогут восстановить родственные связи хотя бы в том плане, что какие-то из древних групп можно однозначно исключить из числа предков той или иной группы современных китов. И любые выводы, воплощённые в виде схем, графиков и родословных древ, обязательно строятся на реальных фактах. В этом нетрудно убедиться, скачав хотя бы несколько научных статей с описаниями тех или иных видов доисторических животных. В них, кроме короткой вводной части и чисто «созерцательного» описания имеющейся находки, достаточно большое место отводится части с названием «Обсуждение». Обсуждая находку, специалист обосновывает её систематическое положение и какие-то свои выводы путём всестороннего сравнения реально имеющих место особенностей строения находки с соответствующими особенностями более или менее родственных форм. В креационистской литературе я ни разу не наблюдал обсуждения креационистами действительно научных статей, а все нападки направлены преимущественно на популярные статьи и сайты с заведомо более упрощенным уровнем подачи материала, который рассчитан на малоподготовленного читателя.
Basilosaurus isis, ..., был гигантским морским археоцетом, датируемым ранним бартоном. Эволюционисты полагают, что базилозавриды произошли от более ранних Protocetidae (которые включают археоцетов, обсуждавшихся выше), но специалисты соглашаются с тем, что «ясные отношения предка – потомка отсутствуют». Конечно, огромная разница в размерах между Basilosaurinae и протоцетидами бросает тень сомнения на эту гипотезу. Все протоцетиды были менее десяти футов длиной, тогда как Basilosaurus cetoides был свыше 80 футов длиной а Basilosaurus isis превышал 50 футов. Было подсчитано, что даже в бурно эволюционирующей линии изменения размера обычно составляют порядка 1 – 10% за миллион лет [28]. ... [28] Ernst Mayr, Animal Species and Evolution (Cambridge, MA: Harvard University Press, 1963), 238 (цитируется работа J. B. S. Haldane). Также см. Robert Wesson, Beyond Natural Selection (Cambridge, MA: MIT Press, 1991), 53. В этом плане стоит обратить внимание на то, что череп Pakicetus составлял в длину примерно половину размера черепов Ambulocetus, Rodhocetus, и Indocetus. Gingerich, et al., (1994): 844-845. |
«Было подсчитано»... На каком материале и насколько эти расчёты
соотносимы с эволюцией китообразных? Просто ранее, в первой части данной работы,
вот здесь, была процитирована немного другая
информация, причём со ссылкой на того же Холдейна.
Ради интереса я попросил свою жену (учителя математики) посчитать время, которое
требуется для превращения существа длиной в 8 футов (2,4 метра) в существо
длиной 80 футов при указанных темпах прироста (нижний и верхний пределы). Результаты
оказались следующими: при приросте в 1% за 1 миллион лет на такой переход потребуется
примерно 231,41 миллиона лет (в три с небольшим раза дольше, чем вся кайнозойская
эра), а при максимальном 10%-ном приросте – примерно 24,159 миллиона лет
(где-то треть кайнозоя). И тот, и
другой
показатели,
естественно,
не
вписываются
в палеонтологическую летопись, где первый полностью морской протоцетид Rodhocetus датируется
средним эоценом, а Basilosaurus известен
из позднего эоцена. Однако, процесс увеличения размеров тела может проходить
гораздо быстрее. Поскольку Эшби Л. Камп высказывает сомнение, связанное сугубо
с линейными размерами, я специально не стану обсуждать качественные различия
между исходным видом и его возможным потомком. Я лишь приведу сколько-то примеров,
показывающих, что в соответствующих условиях увеличение размеров тела может
происходить намного быстрее.
Я думаю, всем известна домашняя птица курица. Так вот, дикий петух весит порядка
700 граммов, а домашние петухи породы кохинхин достигают веса 5 – 5,5 кг.
Петух породы брама весит до 7 кг, а гибридные петухи-бройлеры иной раз бывали
и до
8 кг живым весом. Нетрудно посчитать, что уже при увеличении веса в 8 раз (то
есть, при весе чуть более 5,5 кг) линейные размеры птицы сходных пропорций
увеличились бы в два раза, что фактически имеет место, и в чём всяк может убедиться,
сравнив дикого петуха (при наличии такового) с домашним. А сколько времени
насчитывает история домашних кур? Не миллионы лет, а
лишь несколько
тысячелетий.
Волнистые попугайчики разводятся в неволе недавно: с момента первого их разведения
в неволе не прошло и двухсот лет. Тем не менее, среди них уже есть крупные
породы (так называемые «выставочные»), которые примерно в полтора раза больше
своего дикого собрата.
В последнее столетие зайцы-русаки стали активно расселяться на север. И в их
популяциях уже замечено статистически достоверное увеличение размера тела по
сравнению с популяциями из прежних мест обитания.
А в Австралии уже подстреливали кошек весом около 30 кг, которые по результатам
генетической экспертизы
оказывались
потомками домашних кошек (в частности, Даррен Найш в своём блоге несколько раз
обращался
к
этому вопросу).
Давно
ли
домашние кошки
появились
в
Австралии? Уж точно не раньше белого человека.
Эти примеры показывают, что линейные размеры – не такое уж большое препятствие
для эволюции более мелкого вида в более крупный. При соответствующем направлении
естественного отбора наращивание массы тела (не путать с ожирением!) может
проходить достаточно быстро. Ах, ну да, в этих примерах «совсем нет никакой
эволюции, поскольку куры остались курами, попугаи попугаями, кошки кошками,
а зайцы зайцами; это всё лишь разновидности в пределах одного изначально сотворённого
вида».
Понимаю, что и так можно сказать: много раз видел это возражение у креационистов,
практически слово в слово. Но я заранее оговорился, что не касался качественных
изменений,
которые как раз и превращают один вид в другой, и специально свёл обсуждение только
к количественной стороне вопроса, поскольку в данном случае Эшби
Л. Камп видит препятствие именно в этом.
Не имея обоснованных аргументов, что Basilosaurus
isis действительно произошёл от протоцетид, эволюционисты заявляют,
что он был переходной формой в том смысле, что он демонстрирует особенности,
[промежуточные] между более ранними протоцетидами и поздними китообразными.
Если Protocetus был полностью морским, как сейчас считают
некоторые эксперты, возникает вопрос, могут ли, и в какой степени,
особенности Basilosaurus считаться более «продвинутыми». Но
что более важно, так это если Basilosaurus не произошёл от
протоцетид и не был предком для китообразных [очевидно, имеется
в виду «для более поздних», поскольку он уже был настоящим китообразным
– В. П.] (смотри
далее), то что доказывает наличие промежуточных особенностей у Basilosaurus?
Кажется, самое большее, что можно сказать, это то, что он косвенно
подтверждает
утверждение
о происхождении с изменением, когда демонстрируется существо, похожее
на существо, подразумеваемое гипотетически в реальной цепочке. Креационисты
находят это слишком слабым звеном, чтобы оно несло бремя экстраординарного
заявления об эволюции китообразных .
|
| * Полишинель - французский «коллега» нашего Петрушки. «Секрет Полишинеля» - тайна, известная всем. |
Вам открыть один большой секрет, который для палеонтологов
является «секретом Полишинеля»*? На самом деле виды, которые находят палеонтологи,
большей частью являются не собственно переходными, а их близкими родственниками,
уклонившимися в большей или меньшей степени от направления, ведущего к очередному
звену в цепочке. Ведь в ископаемом состоянии с гораздо большей вероятностью
сохранится не образец из той самой единичной популяции, которая дала
начало новому виду-потомку, а его более многочисленные соседи, которые добились
сиюминутного успеха в борьбе за существование, заняв удобные места обитания,
приспособившись к ним и приобретя в это время некие черты строения, отличающиеся
от таковых у популяции-предка следующего в цепочке вида. Такие виды называют
сейчас «фоновыми». Так что находки ископаемых животных скорее «обрисовывают»
очертания древа жизни, как ребёнок обводит карандашом свою ладонь, положенную
на лист бумаги.
Вопрос о том, «что доказывает наличие промежуточных особенностей
у Basilosaurus?», можно рассматривать с разных сторон. Я не
стану доказывать, что, базилозавр, мол-де, был крайне важным промежуточным звеном
в эволюции китообразных. Я лишь приму мнение специалистов в этой области, и
считаю, что поступить так будет гораздо правильнее, чем доказывать что-то
несуществующее просто «в пику» оппоненту. Также лишний раз напомню, что я
не телепат, и потому не могу со 100%-ной точностью утверждать, о чём думал
тот
или иной
человек, совершая то или иное действие. В частности,
я прямо говорю, что не знаю, какими соображениями руководствуется в каждом
конкретном примере автор того
или иного обзора, посвящённого эволюции китообразных, когда помещает в список
обсуждаемых видов Basilosaurus. Если препятствие для мышения состоит
только в этом, то можно, например, спросить у самого автора – они часто указывают
свой адрес электронной почты.
Мои собственные соображения (ИМХО) по этому вопросу таковы.
Базилозавр – один из наиболее известных видов ископаемых китообразных, к тому
же первый из них, достигший истинно «китовых» размеров, сравнимых с размерами
ныне живущих китов. Поэтому могу предположить, что в обзоры его вводят сугубо
для наглядности, как пример одного из первых и хорошо известных полностью морских
видов китов. Если сказать больше, то в научных статьях он появляется ровно
с такой частотой, какая требуется. Например, если он никаким боком не приходится
родственником тому или иному киту из современных групп, то в статьях про этих
животных нет даже слова "Basilosaurus". Поэтому в данном
случае нападка Эшби Л. Кампа направлена на популярные статьи с заведомо упрощенным
изложением. Если читателя не интересует систематическое положение базилозавра
и степень его родства с современными китами, он и не станет копать глубже.
А кому надо, тот легко найдёт в соответствующей литературе интересующую информацию.
Учёные не делают секрета из того, что базилозавр – не предок современных
китов. Хотя бы специализация его позвонков не позволяет этого сделать: у базилозавра
эволюция стиля плавания привела к появлению длинных, похожих на брёвна позвонков,
тогда как в линии, ведущей к современным китам, увеличивалось количество самих
позвонков при сглаживании перехода между формой позвонков из разных отделов
позвоночника.
Анатомические особенности Basilosaurus могут считаться продвинутыми
не только по отношению к особенностям строения протоцетид, но и по отношению
к особенностям более ранних представителей китообразных. А понятие «переходный»
может применяться в двух смыслах: переходный фактически и переходный анатомически.
Фактически (с филогенетической точки зрения) базилозавр не является переходной
формой между самыми ранними и современными китам. Это учёные признают вполне
официально и говорят об этом прямым текстом. Но с точки зрения анатомии это
животное демонстрирует переходное состояние признаков. У базилозавра конечности
уже превратились в плавники, но сохраняют подвижность в локтевом суставе (кстати,
у дорудона тоже есть такая особенность). У него явно заметна редукция задних
конечностей по сравнению с ранними китами, но конечности ещё достаточно дифференцированы
по строению. И ещё в его черепе наблюдается заметный сдвиг ноздрей
назад: они находятся «на полпути» между кончиком морды (как у самых ранних китов)
и теменем (как у поздних китов). К слову, дорудоны демонстрируют те же особенности,
но учёные считают, что они как раз и представляют собой линию,
ведущую к современным китам.
Джонатан Сарфати «Несостоятельность теории эволюции» ГЛАВА 5 «ЭВОЛЮЦИЯ КИТОВ?»
Однако Basilosaurus был полностью водным существом, поэтому он никак не мог представлять собой переходную форму от наземных млекопитающих к китам. Кроме того, Барбара Сталь (Barbara Stahl), специалист по палеонтологии позвоночных, подчеркивает:
|
Хорошо, базилозавры с их змеевидными телами составляли боковую ветвь эволюции китообразных. А как насчёт представителей второго подсемейства в семействе Basilosauridae? Тех, которые Dorudontinae? Креационисты предпочитают не вдаваться в лишние подробности, но шила в мешке не утаишь. В статье «Креационистские препятствия для разума и эволюция китообразных» критикуется статья Frank Shervin “Scientific Roadblocks to Whale Evolution”. В частности, там указывается именно на этот пример лукавства креационистов:
Да, в отношении Basilosaurus не делалось
заключений, что он является предком для современных китов; тем не менее,
цитируя
Сталь [Барбару – В. П.], Шервин совершает ошибку, смешивая их «[т.
е. Basilosaurus и
родственные виды животных]» со «всеми археоцетами». Есть два семейства
(или, что
спорно, подсемейства) поздних археоцетов; Basilosauridae и Dorudontidae.
Дорудонтиды обладали телами, типичными для более поздних китов, и очень
немногое в их посткраниальных скелетах сильно отличается от [скелетов]
поздних зубатых и усатых китов. Итак, Dorudon также известен
тем, что обладал небольшими задними конечностями, похожими на конечности Basilosaurus,
и его локтевые суставы были подвижными (Uhen, 1998). Многие палеонтологи
считают группу дорудонтид предковой для современных подотрядов китообразных.
|
 |
|
 |
Сверху: скелет Dorudon с сайта
Ф. Джинджерича. Позвоночник животного лишён характерных признаков,
отличающих Basilosaurus: позвонки короткие и их форма достаточно плавно
меняется от грудных к поясничным, крестцовым и хвостовым. Слева: реконструкция Dorudon. Обратите внимание
на положение ноздрей «на полпути» между кончиком морды и теменем. |
Также вспоминается скелет Zygorhiza, короткотелого представителя базилозаврид (или дорудонтид). Его строение ясно показывает, что даже не все базилозавриды были специализированными длиннотелыми существами, не говоря уже обо всех археоцетах. В обзоре книги “The Emergence of Whales” (глава 16) Джеймс Акер приводит мнение учёных относительно родства базилозаврид и дорудонтид, основанное на особенностях их морфологии:
Базилозавриды и дорудонтиды, вероятно, произошли от протоцетид. Он [Hulbert – В. П.] считает их различающимися на уровне семейств; другие считают их подсемействами, Джинджерич не считает. Стоит отметить, что значительная длина тела у Basilosaurus уникальна. «Это различие между базилозавридами и дорудонтинами значительно больше, чем различия между многими родословными линиями современных китов, которые считаются различающимися на уровне семейств». |
В геологической летописи известно много видов и родов ископаемых китообразных. Часть их принадлежит к современным группам, а часть – к давно вымершим. Наличие в природе вымерших групп китообразных чем-то ненравится креационистам. Во всяком случае, когда речь заходит о переходных формах между ископаемыми и современными китами, некоторые креационисты буквально по-кавалерийски «рубят» классификацию. Примером этого может служить статья в Creation Wiki (далее по тексту CW), где обсуждается информация с сайта TalkOrigins (далее ТО):
CW: |
Мог быть «разновидностью Dorudon», а мог и не быть.
И скорее всего, что не был. Wikipedia – это свободная энциклопедия, заметки
для неё пишет кто угодно,
и не всегда им можно доверять. Для не назвавшего себя креациониста, составившего
этот обзор, очень характерно вынесение скоропалительных, но ничем не обоснованных
«диагнозов»: это – дельфин, это – тоже дельфин, а это – обратно дельфин (или
кто-то другой). Вряд ли Agorophius был разновидностью дорудонов хотя
бы потому, что Agorophiidae – это группа китообразных, отличная как от
базилозаврид,
так и от дорудонтид (или от подсемейств базилозаврин и дорудонтин; это уж в
зависимости от того, как понимаются родственные отношения дорудонов и «типовых»
базилозаврид). На основании анатомии семейство Agorophiidae однозначно относят
не к археоцетам, а к зубатым китам. Некоторые авторы-эволюционисты вообще
считают, что Agorophiidae – это не самостоятельное семейство примитивных зубатых
китообразных,
а своеобразная «ступень», уровень развития, которого одновременно и независимо
друг от друга достигли неродственные друг другу ранние китообразные. В свете
исследований, отделяющих Agorophiidae от поздних археоцетов, утверждение, что,
мол, «информации недостаточно», выглядит крайне лицемерным. Одно логичное объяснение
для этого явления есть: для креациониста уже очевиден конечный результат, и
логические построения, ведущие к нему, просто не важны.
Agorophius в данном случае датируется поздним олигоценом, но иногда
для него указывается более ранний возраст. В базе
данных Н. Н. Каландадзе и А. С. Раутиана позднеэоценовый возраст указывается
для родов Agorophius, Archaeodelphis и Xenorophus,
хотя другие авторы указывают для них более поздний, олигоценовый возраст. Собственно
дорудоны и базилозавры вымерли в конце эоцена, задолго до этого времени, но
несколько родов археоцетов также указывается для олигоцена. К тому же среди
специалистов общепризнано, что собственно базилозавриды (не дорудонтиды!) вымерли,
не оставив потомков. Некоторые креационисты даже пеняют этим эволюционистам,
если замечают базилозавра, поставленного эволюционистами в середину ряда форм
китообразных: об этом я уже говорил выше.
Для олигоцена отмечается (в частности, у Р. Кэрролла в «Палеонтологии и эволюции
позвоночных», т. 3, стр. 93) относительная бедность материала
по китообразным:
Это объясняют экологическими факторами, сократившими
численность китов (Lipps, Mitchell, 1976), или геологическими процессами,
сказавшимися на сохранности их остатков (Dawson, Krishtalka, 1984). Возможно,
эволюцию зубатых и усатых китов подтолкнули изменения в циркуляции воды
южных океанов в раннем олигоцене с последующим формированием циркумантарктического
течения (Fordyce, 1980).
|
В работе Mark D. Uhen и Nicholas D. Pyenson «Оценки разнообразия, тенденции и историографические эффекты: обзор разнообразия китообразных в третичный период» (DIVERSITY ESTIMATES, BIASES, AND HISTORIOGRAPHIC EFFECTS: RESOLVING CETACEAN DIVERSITY IN THE TERTIARY) приводится диаграмма многообразия родов китообразных, описанных для каждой эпохи кайнозоя, начиная со времени их появления:
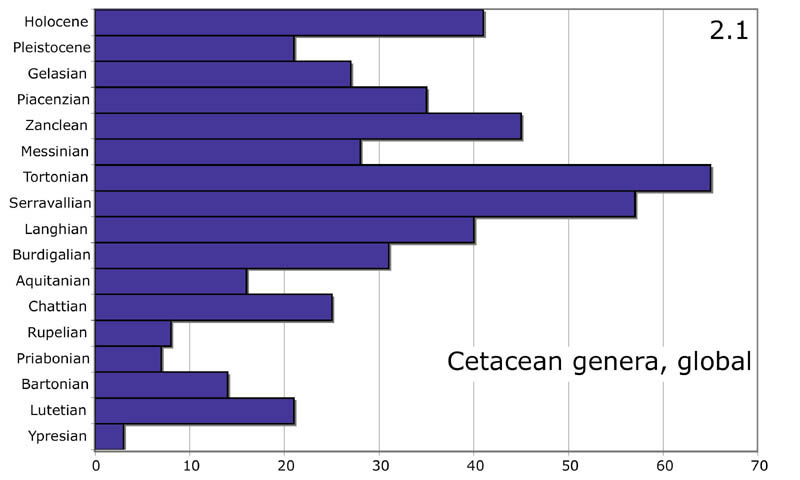
Рисунок 2.1 Многообразие родов китообразных для всей Земли в соответствующие геологические эпохи:
Holocene – голоцен (современная эпоха)
Pleistocene – плейстоцен
Gelasian – геласский век (поздний плиоцен)
Piacenzian – пьяченский век (средний плиоцен)
Zanclean – занклийский век (ранний плиоцен)
Messinian – мессиний (вторая половина позднего миоцена)
Tortonian – тортоний (первая половина позднего миоцена)
Serravallian – серраваллий (вторая половина среднего миоцена)
Langhian – лангий (первая половина среднего миоцена)
Burdigalian – бурдигалий (вторая половина раннего миоцена)
Aquitanian – аквитаний (первая половина раннего миоцена)
Chattian – хатт (поздний олигоцен)
Rupelian – рюпель (ранний олигоцен)
Priabonian – приабон (поздний эоцен)
Bartonian – бартон (вторая половина среднего эоцена)
Lutetian – лютет (первая половина среднего эоцена)
Ypresian – ипр (ранний эоцен)
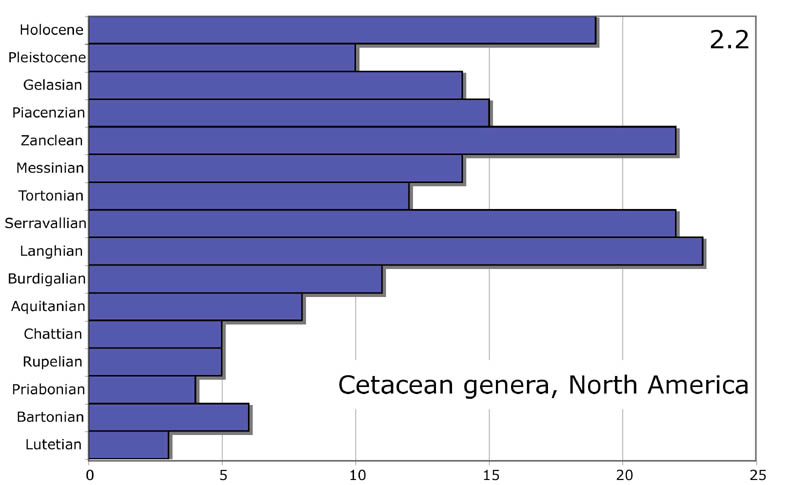
Рисунок 2.2 Те же данные для Северной Америки. Обратите внимание на изменения масштаба линеек, показывающих количество родов. Однако авторы отмечают сходство динамики колебаний родового многообразия китообразных на обеих схемах.
Из этих схем видно, что в позднем эоцене и раннем олигоцене
разнообразие китообразных резко сократилось, но уже в позднем олигоцене превзошло
разнообразие архаичных китов эоцена. Однако, для позднего олигоцена уже отмечаются
представители совершенно иных семейств китообразных, относящихся к ныне существующим
подотрядам.
А можно ли найти более конкретный материал? Вполне, нужно лишь немного поискать,
если информация действительно нужна. На сайте http://paleodb.org есть
возможность делать выборки по группам живых организмов и времени их существования.
Введя
название отряда "Cetacea" и отрезок времени, соответствующий раннему
олигоцену (от 34 до 28 миллионов лет назад), я буквально в течение нескольких
секунд
получил список из видов, найденных в отложениях того времени. И ещё немного
времени ушло на то, чтобы уточнить по той же базе данных их систематическое
положение. Итак, вот некоторые раннеолигоценовые китообразные:
Археоцеты:
Семейство Basilosauridae
Phococetus vasconum
Зубатые киты:
Семейство Agorophiidae
Agorophius pygmaeus
Семейство Squalodelphinidae
Medocinia tetragorhina
Семейство Simocetidae
Simocetus rayi
Семейство Squalodontidae
Squalodon sp.
Не определённые до рода и вида представители сквалодонтид:
Squalodontidae,"Genus X"
Squalodontidae,"Genus Y"
Squalodontidae,"Genus Z"
Odontoceti incertae sedis (зубатые киты неясного
систематического положения)
Saurocetus gibbesii
Xenorophus sloanii
Xenorophus sp.
Не определённые до рода и вида зубатые киты:
Odontoceti,"Genus X"
Odontoceti,"Genus Y"
Odontoceti,"Genus Z"
Усатые киты:
Семейство Aetiocetidae
Willungacetus aldingensis
Семейство Cetotheriopsidae
Micromysticetus rothauseni
Вполне возможно, что база данных на указанном сайте не исчерпывающе
полна. Например, в on-line базе данных Дж. Сепкоски для нижнего олигоцена упоминаются также
киты родов Manicetus, Oligodelphis, Platyosphys.
Из всей этой информации видно,
что в раннем олигоцене фауна китообразных была всё же достаточно разнообразной,
и даже включала некоторых
представителей
обоих
современных
подотрядов. Даже представитель рода Agorophius найден среди раннеолигоценовых
китообразных, что уже само по себе интересно при обсуждении родословной китообразных.
Так что претензии составителя из Creation Wiki уходят не по адресу. Кому надо,
тот
легко может
получить информацию
хотя
бы
на уровне
перечисления
родового и видового разнообразия (а в указанной электронной базе данных можно
сделать также выборки по странам и континентам). А кому это совсем не надо,
тот будет строить обиженное личико и водить корявым пальчиком по
строчкам,
безосновательно
упрекая
учёных
в разных смертных грехах.
Что у нас там дальше?
CW: |
Prosqualodon не относится ни к одному из современных семейств дельфинов. Нужно внимательнее текст читать, господа креационисты. Вам ничего не говорит упоминание такой особенности, как бугры на коренных зубах? Просто у современных китообразных зубы гомодонтные – все практически одинаковой формы (вот и пример возврата к предковому, рептильному состоянию; эволюция – далеко не всегда морфофизиологический прогресс). А наличие дополнительных бугров на коренных зубах – характерный признак примитивных семейств зубатых китов типа семейства Squalodontidae. Из статьи «Креационистские препятствия для разума и эволюция китообразных»:
Зубная система ранних археоцетов гетеродонтная (подразумевается типичная для остальных млекопитающих, так как в наличии резцы, клыки и коренные зубы), а у зубатых китов имеет тенденцию к гомодонтности (похожие друг на друга зубы во всей челюсти, колышкообразные у дельфинов и кашалотов, лопатообразные у морских свиней), а также тенденцию к проявлению полиодонтии (многочисленные зубы). Здесь есть исключения. У многих китов зубная система редуцирована. «Примечательные зазубренные щёчные зубы», упоминаемые Stahl (Шервин, ссылка 11) – не совсем точное высказывание. Зубы археоцетов имеют не зазубрины, а серию зубчиков (бугорков), которые сохранялись, хотя прогрессивно уменьшались с течением времени, во многих линиях архаичных китов, как у зубатых, так и у усатых китов (Fordyce, et al, 1994), а гетеродонтная зубная система сохранялась у сквалодонтид до их вымирания в позднем миоцене. Некоторые ныне живущие речные дельфины имеют моляроподобные задние зубы, а зубы усатых китов (рассасывающиеся в утробе) гетеродонтны: типичные колышкообразные – передние, и несущие дополнительные бугры – задние. [Шервин ссылается на книгу Барбары Сталь B. J. Stahl, Vertebrate History: Problems in Evolution (Dover Publications, Inc., 1985), p. 489. - В. П.] |
Сквалодонтид сближают с ныне живущими речными дельфинами семейства Platasnistidae. В статье “Shark-toothed dolphins (Family Squalodontidae)” R. Ewan Fordyce пишет:
Французский учёный J-P-S Grateloup предложил название
Squalodon для фрагмента челюсти в 1840 году. Grateloup считал, что окаменелость
может представлять собой рептилию. Более поздние находки в районе Северной
Атлантики показали, что эти животные были млекопитающими, с черепами,
чем-то похожими на черепа ныне живущих одонтоцетов – зубатых китов, дельфинов
и морских свиней. Однако, зубы были непохожи на зубы любого из современных
видов. Ныне живущие зубатые киты обычно имеют простые конические зубы,
которые не сильно меняются в челюстях от передних к задним, но зубы сквалодонтид
дифференцированы: на кончике челюсти растут резцы с коническими вершинами,
которые спереди назад сменяются зубами с более низкими коронками и многочисленными
мелкими зубчиками, или буграми. Сходным образом дифференцированные зубы
присутствуют у архаических, или предковых китов, Archaeoceti.
… В течение многих десятилетий считалось, что сквалодонтиды представляют собой исходную группу, от которой эволюционировало много, или большая часть современных зубатых китов. В конце 1980-х гг. доктор Christian de Muizon заключил, что сквалодонтиды более родственны ныне живущему гангскому дельфину Platanista gangetica, находящемуся под угрозой исчезновения, нежели любому другому ныне живущему виду. De Muizon основал свою оценку родства главным образом на [строении] лопатки, а также ушной области. Похоже, сквалодонтиды имеют мало отношения к происхождению большей части ныне живущих дельфинов. |

Образец Squalodon был одной из первых окаменелостей, описанных в научной литературе. Рисунок Ангустино Сцилла 1670 года показывает обломок челюсти сквалодона с тремя коренными зубами, имеющими два корня и характерные пильчатые края. Этот образец принадлежит виду Squalodon melitensis. Сама окаменелость находится ныне в Седжвикском музее, Кембридж, Соединённое Королевство.

Реконструкция Squalodon

Реконструкция Squalodon, на врезке – фото черепа. С сайта Palaeos Vertebrates.
Одновременное обитание видов в какой-то момент истории ещё не означает, что они появились одновременно. Например, на современной Земле сосуществуют как относительно молодой человек разумный (возраст нашего вида менее 1 миллиона лет), так и гораздо более древние африканские носороги (возраст 3 – 4 миллиона лет), и даже щитень (самый долгоживущий вид ныне живущих животных: его возраст около 230 миллионов лет). Кроме того, нельзя быть уверенным, что оба этих рода китообразных всегда сосуществовали одновременно. Agorophius может быть реликтом более ранней стадии эволюции китообразных и расценивается как переходная форма с позиций анатомии. Кроме того, поиск по сетевым палеонтологическим базам данных, как было показано выше, указывает на находки Agorophius в раннем олигоцене.
CW: |
Насчёт раннего миоцена всё хорошо, спасибо за беспокойство. Я посмотрел базу данных Дж. Сепкоски на предмет китообразных, существовавших в раннем миоцене, и обнаружил следующие роды китообразных, существовавшие в раннем миоцене (c добавлениями и распределением по семействам согласно списку в книге Р. Кэрролла «Палеонтология и эволюция позвоночных», т. 3, стр. 227 - 228, а также базам данных по палеонтологии on-line):
Зубатые киты:
Семейство Squalodontidae: Neosqualodon, Phoberodon, Prosqualodon,
Sachalinocetus, Squalodon, Tangaroasaurus
Семейство Eoplatanistidae: Eoplatanista
Семейство Platanistidae (речные
дельфины): Proinia
Семейство Ziphiidae (клюворылы): Notocetus, Squaloziphius
Семейство Delphinidae (дельфиновые):
Delphinavus, Iniopsis, Protodelphinus, Stereodelphis
Семейство Rhabdosteidae
(Eurhinodelphidae): Argyrocetus (в т. ч. Doliodelphis), Macrodelphinus, Ziphodelphis
Семейство Acrodelphidae:
Acrodelphis, Cyrtodelphis, Schizodelphis
Семейство Phocaenidae (морские свиньи):
Phocaenopsis
Семейство Physeteridae (кашалоты): Apenophyseter, Diaphorocetus,
Idiorophus,
Orycterocetus, Physeter, Physeterula
Семейство Kogiidae (карликовые кашалоты): Miokogia
Семейство Squalodelphidae:
Squalodelphis
Odontoceti incertae sedis (зубатые киты неясного систематического положения): Miodelphis, Champsodelphis, Microsqualodon, Rhytisodon (интересно, что под этим названием описан ещё и крокодил!), Trirhizodon.
Усатые киты:
Семейство Cetotheriidae: Cetotheriopsis, Mauicetus
Семейство
Balaenopteridae (полосатики): Plesiocetus
Семейство Balaenidae (гладкие киты):
Aglaocetus, Morenocetus
Ну вот, я же говорил,
что у китообразных в нижнем миоцене всё хорошо. Я сознательно опускал виды,
для которых временем существования указан просто «миоцен». Работа
заняла в общей сложности не более двух часов, включая некоторое время
на уточнение систематического положения некоторых родов китообразных, не включённых
в книгу
Кэрролла, но
имеющихся
в базе данных Сепкоски, с помощью баз данных в Интернете (в частности, с сайта
The
Paleobiology Database).
А вот с Kentriodon у автора из Creation Wiki явно не всё получается
так, как задумано. Все современные зубатые киты, в том числе дельфины как
их частный случай, отличаются очень
характерным признаком: асимметрией черепа. Такой признак, как симметричность
черепа, выводит Kentriodon не только из числа дельфинов, но и вообще не позволяет
причислить его ни к одной современной группе зубатых китов.
Относительно эволюции зубатых китов, и, в частности, приобретения асимметричности
черепов современными и ископаемыми зубатыми китами Р. Кэрролл в книге «Палеонтология
и эволюция позвоночных» (т. 3, стр. 94) пишет следующее:
Можно сгруппировать зубатых китов в пять надсемейств,
соответствующих крупным филогенетическим радиациям (…). Каждое надсемейство
характеризуется особым типом асимметрии [черепа – В. П.]. Squalodontoidea
включают четыре семейства из олигоцена – раннего миоцена и, вероятно,
предков всех современных групп зубатых китов. Наиболее примитивно семейство
Agorophiidae, хронологически и морфологически занимающее промежуточное
положение между археоцетами и более продвинутыми одонтоцетами. «Телескопизация»
черепа еще не особенно заметна, щечные зубы по-прежнему многокоренные
и несут дополнительные зубчики. В семействе Squalodontidae череп полностью
«телескопический» и ноздри находятся между глазницами. Теменные кости
с крыши черепа вытесняются и верхнечелюстные приходят в соприкосновение
с верхнезатылочной. Щечные зубы, хотя и увеличиваются в числе, сохраняют
примитивную треугольную конфигурацию. Возможно, сквалодонтиды - предки
большинства позднейших зубатых китов. Вымершие Rhabdosteidae (Eurhinodelphidae)
– длиннорылые формы, у которых все зубы редуцировались до простых заостренных
колышков. У Squalodelphidae уже в раннем миоцене был асимметричный череп.
К надсемейству Delphinoidea относится большинство ныне живущих зубатых китов. Kentriodontidae, известные с нижнего миоцена до раннего плиоцена, связывают их с первой (сквалодонтоидной) радиацией зубатых китов. Длина кентриодонтид не превышала 2 м, сохранялся примитивный симметричный череп. Их потомки, современные Delphinidae (дельфиновые), Phocaenidae (морские свиньи) и Monodontidae (белуховые, или нарваловые) появляются в позднем миоцене примерно 10-11 млн. лет назад. Асимметрия черепа у каждого из этих семейств развивалась, по-видимому, самостоятельно. |
Не всякий относительно небольшой кит относится к дельфинам,
и не всякое более крупное китообразное можно отнести к одному из семейств крупных
китов. Как
пример можно привести косатку – самого крупного дельфина (до 10 метров длиной).
Несмотря на своё название «кит-убийца» (англ. “killer whale”), особенно закрепившееся
в англоязычной литературе, это всего лишь очень крупный вид семейства дельфинов.
То же самое можно сказать о гринде (по-английски “pilot whale”, то есть,
«кит-пилот») – это тоже дельфин, хотя его длина – около 6,5 метров. А вот ремнезубы
(Mesoplodon),
уступающие по размерам указанным разновидностям дельфинов (длина разных видов
от 4,4 до 6 метров), относятся не к ним, а к особому семейству клюворылых китов.
В семействе кашалотовых карликовый кашалот, или когия (Kogia) имеет длину до
4 метров.
Тем не менее, это не дельфин.
Подробности об эволюции зубатых китов можно найти на этом сайте:
В течение раннего миоцена лобная жировая линза
развилась в свою современную форму, и усовершенствовалась акустическая
система. Процветали различные дельфинообразные семейства, ныне вымершие.
Squalodelphinidae были самыми ранними известными зубатыми китами с асимметричными
черепами. Было определено два рода: Squalodelphis из Северной Италии,
и Diochoticus из Аргентины. Эти существа были мелкими, возможно, менее
трёх метров в длину, и имели многочисленные гомодонтные зубы. Черепа
были телескопическими характерным для Rhabdosteidae и многих производных
видов Squalodonotidae способом. Некоторые считают, что клюворылые киты
эволюционировали от этого вида, но свидетельства в пользу этого очень
ограниченные.
В наше время надсемейство Delphinoidea включает большинство ныне живущих видов китообразных, включая семейства Delphinidae, Phocoenidae и Monodontidae, равно как вымершие семейства Albireonidae и Kentriodontidae, которые считаются их предками. Большинство представителей семейства Kentriodontidae было очень мелкими, длиной только до двух метров. У них были короткие клювы и много гомодонтных зубов. Хотя они были разнообразны в течение среднего и позднего миоцена и в Атлантике, и в Пацифике, образцов моложе 10 миллионов лет нет. Семейства Delphinidae, Phocoenidae и Monodontidae генетически очень близки, имеют около 10 – 15% вариаций в гетерохроматине С. Почти у всех одно и то же число хромосом, 2n = 44. Единственное исключение – косатка, с только одной парой t-хромосом, имеющей короткие плечи и сателлитные структуры. m-хромосомы у косатки также очень малы. |
Вот, заодно, ещё один ответ на упрёк не назвавшего себя креациониста из Creation Wiki – эволюция китообразных в раннем миоцене расписана довольно подробно. Мало того, если посмотреть сетевые источники, сведений по морфологии, эволюции и родственным связям ранних китообразных может быть найдено более, чем достаточно. Вот, например, сведения из обзора эволюции зубатых китов с сайта М. Милинковича:
Odontoceti Особый случай так называемых «речных дельфинов» В то время как большинство из 83 признных ныне живущих видов китообразных
исключительно морские, несколько видов живёт периодически или
исключительно в пресной воде. Некоторые из них (напр., белуха, амазонский
дельфин
(тукаши), иравадийский дельфин, беспёрая морская свинья) чётко
характеризованы филогенетически, т. е. они принадлежат к надсемейству
Delphinoidea
(см., напр., Heyning, 1989; LeDuc и др., 1999). С другой стороны,
таксономические отношения так называемых «речных дельфинов» были
в неупорядоченном состоянии в течение более чем столетия. Они
включают три исключительно речных вида:
Филогенетический анализ нуклеотидных последовательностей
(митохондриальных и ядерных генов) подтверждает, что ныне живущие речные
дельфины – не
монофилетичны (Cassens и др. 2000; Hamilton и др. 2001)
и предполагает, что они - реликтовые виды, чья адаптация
к речным местообитаниям случайно гарантировала их выживание при крупных
изменениях окружающей среды в
морской экосистеме или при появлении Delphinidae (Cassens и др.
2000). […] Обсуждение некоторых проблематичных вымерших групп Несколько ископаемых групп зубатых китов (напр., Agorophiidae, Squalodontidae, Eurhinodelphinidae и Kentriodontidae) всё ещё таксономически проблематичны; вероятно, что некоторые из них образуют парафилетические группы.
|
 |
 |
Череп Simocetus rayi (вид
открыт и описан в 2002-м году) демонстрирует смесь примитивных и высокоспециализированных
черт. Частично беззубые челюсти указывают на то, что это китообразное питалось, засасывая ртом кормовые объекты. Их форма предполагает, что животное питалось в придонных слоях воды. Форма черепа указывает на способность к эхолокации. |
|
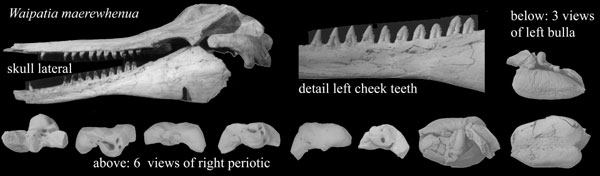
Waipatia maerewhenua открыта в 1994 году в известняках Отекаике, Новая Зеландия. Длина черепа около 60 см.
|
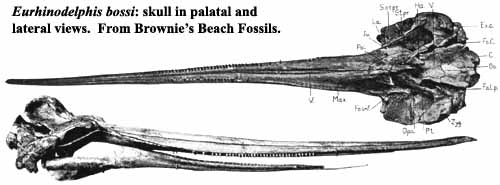
Череп Eurhinodelphis bossi со стороны нёба и сбоку. Рисунок с сайта Palaeos vertebrates.

Реконструкция Eurhinodelphis.
|
Если далее говорить о сотворении/эволюции у китообразных, возникает ещё один
закономерный вопрос: почему признаки, характерные для современных зубатых китообразных,
появляются в истории китообразных не одновременно? Вначале появляются китообразные
с уменьшенными задними конечностями, разделённым на отделы позвоночником, дифференцированными
зубами и
симметричным черепом с ноздрями вблизи кончика. Затем появляются китообразные
со сдвинутыми
на переносицу ноздрями и тазом, не связанным с позвоночником, но задними конечностями,
сохраняющими типичное строение. Далее их облик становится ещё более «китообразным»,
позвонки становятся более-менее одинаковыми, их количество увеличивается, а
границы между отделами позвоночника сглаживаются. Видимые
снаружи
задние
конечности
исчезают,
череп
становится
«телескопическим»
и
ноздри сдвигаются практически на темя. Но симметричность черепа указывает на
то, что эхолокация всё
ещё не развита. И зубы пока ещё остаются дифференцированными, хотя различия
постепенно начинают сглаживаться. Потом появляются китообразные с гомодонтной
зубной системой.
А
затем
разные
киты
независимо
друг
от
друга
развивают
несимметричные
черепа, которые указывают на появление способности к эхолокации.
Я, конечно, понимаю, что прочитать мысли бога-творца
никто не может. Во всяком случае, сторонники религиозного подхода к познанию
мира наделяют богов особым свойством непознаваемости их мышления людьми. Тем
не менее, по сравнению с современными китообразными древние виды выглядят «неполноценными»:
при внешнем подобии, подразумевающем сходный образ жизни, одни китообразные
имели способность к эхолокации, а другие не имели. Для того же семейства кентриодонтид
указывается разнообразие видов, сравнимое с таковым у современных дельфинов.
Но их симметричный череп говорит об отсутствии способностей к эхолокации. Поэтому
со стороны всё выглядит похожим не на мудрое творение изначально совершенных
существ, а на метод проб и ошибок, при котором каждое новое приспособление
возникает
«в порядке общей очереди», и его обладатели, имея преимущества в выживании
перед представителями более древних групп китообразных, вытесняют их, а затем
сами
оказываются вытесненными иными представителями китобразных, обладающими новым
приспособлением. Археоцеты с обтекаемой формой тела вытеснили древних земноводных
«полу-китов», ранние киты современных подотрядов с более совершенным строением
позвоночника вытеснили
археоцетов и образовали ряд самостоятельных семейств (Squalodontidae, Waipatiidae
и др.),
киты
с
дыхалом и гомодонтной зубной системой (или китовым усом) вытеснили более ранних
представителей зубатых и усатых китов. И затем среди зубатых китов виды с совершенной
системой
эхолокации сменили ранних зубатых китов с симметричными черепами.
Вторая ветвь современных китообразных представлена усатыми китами, животными, которых большинство обывателей представляет себе в первую очередь при слове «кит». У этих животных в их современном виде зубы исчезли, и функционально их заместил китовый ус, роговое образование кожного происхождения. Но у древних усатых китов следы происхождения от зубастых предков значительно более выраженные: ранние усатые киты имели хорошо развитые зубы. И спор Creation Wiki и TalkOrigins продолжается...
CW: |
 |
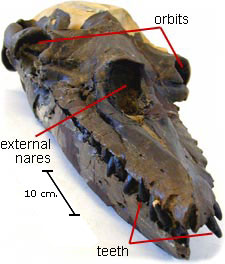 |
Сверху: череп Aetiocetus.
Подписи на рисунке: Слева: великолепная иллюстрация Aetiocetus, выполненная Карлом Буэллом. |
Комментарии Creation Wiki к информации с TalkOrigins можно практически считать законченными: в списке сайта TalkOrigins указан ещё один вид ископаемых китообразных, Mesocetus:
Mesocetus был разновидностью усатых китов, и у него не было зубов. Что у нас есть, так это зубы, а затем отсутствие зубов, без свидетельства постепенной утраты, что соотносится с вышеизложенной гипотезой. |
Комментатор из Creation Wiki
с ходу относит его к «разновидности
усатых
китов», даже не задумываясь над тем, что современные (я не беру в расчёт ископаемых!)
усатые киты образуют несколько хорошо различающихся семейств, которые даже
с большой
натяжкой
не
втиснуть
в один
«барамин». Я оставлю это утверждение на его совести (при наличии таковой) и
на совести тех, кто будет тиражировать в своих статьях это утверждение (при
том же условии). Кстати, в настоящее время учёные склоняются к мнению, что
род Mesocetus является сомнительным, а ископаемые остатки, объединённые
под этим названием, принадлежат разным видам примитивных усатых китов. К слову,
часть их относят к роду Piscobalaena из семейства Cetotheriidae (тоже
считающегося сборным). Понятно, что при отнесении Mesocetus (в его
прежнем понимании) к «разновидности
усатых
китов» (в «бараминном» смысле) придётся крепко переосмыслить объём самого «барамина»
усатых китов и поднять его до ранга подотряда. И тогда придётся «сбараминить»
кошек с гиенами и виверровыми, а собак с медвежьими, енотами, пандами, куньими
и тремя семействами водных «ластоногих» в нагрузку. Панда, лиса и морской слон
в одном «барамине» – не слишком ли круто? Есть и другой выход: признать искусственность
самого понятия «барамин» и более осторожно объединять разные виды в
«разновидности» каких-то неопределённых групп.
Aetiocetus –
очень интересное китообразное. Он сочетает в строении признаки усатых
китов (особое
строение
челюстей, характерное
для
этой группы) и зубы.
Для эволюционистов это животное представляет собой интерес как форма, переходная
между примитивными ранними китами, имеющими зубы, и более специализированными
усатыми китами. Стоит заметить, что в последние годы сделано много находок
доисторических усатых китов (во всяком случае, эти животные классифицируются
в подотряде Mysticeti, а не Odontoceti), и Aetiocetus уже не является
«наиболее примитивным известным усатым
китом».
Креационисты,
как видно из приведённого отрывка, имеют свой особый взгляд на это существо.
По
их мнению,
это «просто
усатый
кит с
зубами».
Такая точка зрения очень напоминает толкование креационистами археоптерикса:
были, мол, птицы с зубами, и птицы без зубов, с пальцами на крыльях, и без
пальцев. В таком случае киноплёнка – это просто набор картинок на длинной прозрачной
ленте, и картинки не имеют ничего общего между собой, поскольку невооружённым
глазом ясно видно, что они разделены чёрными полосочками.
Относительно ранней эволюции усатых китов хорошо написал Эрих Фитцжеральд, автор
описания одного из удивительных усатых китов, о котором я расскажу ниже. В описании
этого вида он пишет следующее:
Свидетельство того, как эволюционировали усатые
киты и их замечательные приспособления дли питания, представлено плохо.
Фильтрование пищи en masse, обладание китовым усом, отсутствие зубов
во взрослом состоянии и большой размер тела олицетворяют ныне живущих
и ископаемых усатых китов ветви Chaeomysticeti (Pivorunas 1979; Sanderson
и Wassersug 1993; Fordyce и Muizon 2001; Marshall 2002; Geisler и Sanders
2003; …). Геологически самый старый из известных усатых китов, Llanocetus
denticrenatus, известен с рубежа эоцена/олигоцена (приблизительно 34
млн. лет назад) Антарктиды; он имел сильно разделённые промежутками зубы
с широкими пальчатыми зубчиками и, возможно, «прото-китовый ус», который
дополнял вероятную фильтрационную функцию его зубов (Fordyce 2003; …).
Другие зубастые усатые киты включают разнообразных Aetiocetidae, которые,
как думают, были на начальной стадии питания путём фильтрации и обладали
китовым усом, и Mammalodon colliveri, у которого не было китового уса,
но экология его питания остаётся загадочной (Barnes и другие. 1995; Fordyce
и Muizon 2001; Berta и Deme.re. 2005; Deme.re. 2005; Ichishima 2005;
Sawamura и др. 2005; Berta и
др. 2006).
|
Весьма любопытно объяснение автора из Creation Wiki по поводу
утраты зубов усатыми китами. Мнение о том, что допотопные библейские патриархи
жили несколько
веков, ничем, кроме самой Библии, не подтверждается. Поэтому доверять Библии
в этом плане нельзя. Тем более, что в священных книгах других религий, имеющих
такие же сверхъестественные (или наоборот, вполне земные) источники вдохновения,
ничего не говорится о том, какими были первые люди и сколько они жили. Впрочем,
иногда говорится, но не совсем то, что в Библии. И даже количество и порядок
природных катастроф, уничтожавших разные варианты человечества, различается
по разным книгам, которые в одинаковой степени священны, только для разных
народов. Поэтому ссылаться на Библию и прямо переносить описанные в ней события
на
людей и животных нельзя – она не выдерживает критики даже в свете сравнения
с иным
религиозным текстом.
Но можно подключить воображение, и представить себе Землю до и после Всемирного
Потопа по библейской версии в представлении «научных креационистов». Можно
представить себе, как пролился многодневным дождём «водно-паровой экран»,
защищавший Землю от губительного ультрафиолетового излучения. Можно посочувствовать
людям
и животным, чья продолжительность жизни стала снижаться из-за последовавшего
за этим облучения. Например, динозавры у креационистов в результате такого
катаклизьма «выродились» в ящериц, змей, крокодилов и даже чуть
ли не в саламандр (и это при том, что ящерицы древнее динозавров!). Так что
катастрофа должна быть та ещё. Применительно к теме рассказа
одно остаётся непонятным: киты живут в воде, которая защищает их от ультрафиолетового
излучения
подобно
озоновому
слою.
Почему
тогда
на них
повлияло
космическое
излучение? Или дело не в потопе было?
Ещё один способ создать иллюзию «сотворённости» китообразных – это показать, что все их группы (археоцеты, усатые и зубатые киты) появились независимо друг от друга и не имеют никаких родственных связей. В этом преуспел в своё время Дуэйн Гиш, и у него есть последователи. Мнение креациониста Тимоти Уоллеса (TW) из работы “Five Major Evolutionist Misconceptions of Evolution” комменирует Джон Хоппнер (JH):
TW:
В любом случае, весьма примечательно (и явно отсутствует в документах Хант),
что эти Archeoceti [sic] (или предполагаемые «примитивные киты») также не
общеприняты. Г. A. Мчедлидзе, российский эксперт по китам, выразил серьезные
сомнения относительно того, могут ли существа, подобные Pakicetus, Ambulocetus
natans, и другие – даже если считать их водными млекопитающими – должным
образом рассматриваться как предки современных китов. Вместо этого он видит
их как полностью изолированную группу. […]
«проблема филогенетических отношений между Archaeoceti и современными Cetacea – весьма спорная проблема, которая породила много противоречивых мнений. Некоторые авторы (Абель, 1913; Спасский, 1954; Ромер, 1966) подтверждают возможность таких отношений с усатыми китами. Другие (Ван Вален, 1966, 1968) допускают возможность филогенетических отношений между Archaeoceti и усатыми и зубатыми китами. Наконец, множество авторов (Келлогг, 1928, 1936; Миллер, 1923; Яблоков, 1964; и другие) противоречат этим и другим подобным гипотезам, рассматривая Archaeoceti как полностью изолированную группу, не имеющую ничего общего с типичным Cetacea». [Г. А. Мчедлидзе, Общие Особенности Палеобиологической Эволюции китообразных, Серия Русских Переводов, A.A.Balkema/Rotterdam, 1984, p. 91] |
JH:
Мчедлидзе – это его единственная цитата для построения «соломенного кита».
Это – прекрасный пример проблем, которые возникают у креационистов, когда
они только повторяют, как попугаи, ложные заявления других креационистов.
Искажение от этого только усугубляется. Все, что он сделал, это перефразировал
страницу 205 из книги Гиша. Часть, на которую, вероятно, ссылается г-н
Уоллис (или Гиш), находится в начале главы 5 «Филогенетических отношений
Cetacea». Мы видим, что Мчедлидзе только отмечает противоречие в последовательности,
и на следующих страницах он обсуждает более подробно каждую точку зрения,
сосредотачиваясь главным образом на родах Mirocetus, Aetocetus, и Ferecetotherium из семейства Basilosauridae. Это находится в следующей главе – той, где
он приводит нам своё собственное заключение:
«Последние палеоценовые [в оригинале Paleocene; очевидно, имеются в виду палеогеновые – В. П.] формы ранних Cetacea (Mirocetus, Aetocetus, Ferecetotherium) характеризуются специфической комбинацией особенностей Archaeoceti и беззубых китов. Они, очевидно, занимают промежуточное положение между Archaeoceti и Mysticeti. Усатые киты [Mysticeti] близко связаны филогенетически с Archaeoceti, в то время как зубатые киты, вероятно, представляют собой независимую ветвь общей совокупности архаичных млекопитающих. Усатые киты эволюционировали от некоторых форм позднеэоценовых или раннеолигоценовых Archaeoceti; Ferecetotherium, который жил в позднем олигоцене, может рассматриваться как одна из форм, не слишком отличающихся от группы Archaeoceti, которая дала начало усатым китам». [Мчедлидзе, (1984), стр. 118 – 119] |
JH:
Если бы г-н Уоллис читал книгу, он увидел бы, что Мчедлидзе очень ясно говорит,
что он думает, что Archaeoceti – переходные формы от наземных млекопитающих
к Mysticeti. Это родословная линия от Archaeoceti к Odontoceti (зубатые киты)
о которых идёт речь. Также следует отметить, что Pakicetus и Ambulocetus
natans – члены семейства Protocetidae подотряда Archaeoceti, но они даже
не были упомянуты Мчедлидзе. К тому же, эта книга была только переведена
с русского на английский в 1984 году, книга была фактически издана в 1976
году, на восемь лет раньше, чем был обнаружен Pakicetus, и на восемнадцать
лет раньше, чем Ambulocetus natans! Итак, как он мог выражать серьезные сомнения
и рассматривать их как совершенно иную группу, как говорит г-н Уоллис, когда
они были ещё неизвестны? Может быть, именно поэтому Мчедлидзе отсутствовал
в документе мисс Хант? (имеется в виду обзор Кейтлин Хант на сайте TalkOrigins,
посвящённый переходным формам в различных
группах животных – В. П.).
|
TW:
В 1988 г. Р. Л. Кэрролл, ведущий палеонтологический авторитет среди эволюционистов,
высказал в печати предположение, что киты эволюционировали от наземного млекопитающего,
подобного мезонихидам. С тех пор кажется, что была яростная попытка приписать
происхождение китов кому-нибудь напоминающему этих волкообразных существ,
создавая поведенческие сценарии водного образа жизни, чтобы таким образом
помочь воображению в «заполнении» одного из многих неприятных промежутков
в летописи окаменелостей.
|
JH:
Г-н Уоллис сообщает нам что книга [Роберт Л. Кэрролл, Палеонтология и эволюция
позвоночных, W. H. Freeman и Co., Нью-Йорк, 1988] вдохновила «яростную попытку
приписать происхождение китов кому-нибудь напоминающему волкообразных существ»,
но он будет не в состоянии процитировать никаких статей, где это происходит,
или сообщать нам, какие «волкообразные» существа ошибочно классифицируются
как мезонихиды. Если бы Уоллис читал книгу, он увидел бы, что это – тщательно
детализированное описание того, какие ископаемые и как интерпретируются.
На страницах 520 и 521 он обнаружил бы, какие реальные морфологические характеристики
и как используется для отнесения образцов к Mesonychia. Его серьезное невежество
в палеонтологии делает непростительным для него высказывание таких заявлений,
если он не может обеспечить хоть какие-то свидетельства, чтобы доказать свои
сваленные в кучу обобщенные обвинения. Изучение свидетельств, найденных во
всех его источниках,
оказывается более неприятным для Уоллиса с его воображением.
|
Иными словами, источник громких заявлений креационистов здесь такой же, как
обычно – выхваченные из контекста «удобные» цитаты, замалчивание фактов, вольное
толкование объективной информации, а также банальное невежество и попытки выдать
желаемое за действительное.
Новые находки доисторических китообразных сильно затрудняют жизнь креационистов.
Возможно, именно поэтому креационисты предпочитают пользоваться «проторенными
дорожками». Новые и старые, но пока малоизвестные находки не успевают попасть
под
«каток» креационистских умопостроений. Я могу с полной уверенностью утверждать
это,
поскольку
просмотрел
достаточно
много креационистских статей, и не заметил особого разнообразия упоминаемых в
них видов ископаемых китообразных: их не больше, чем в соответствующих статьях,
написанных с эволюционной точки зрения. Особенно «крепко» достаётся первым китообразным
– пакицетусу, амбулоцетусу и базилозавру. Упоминания о других разновидностях
китообразных скудны и единичны, не несут никакой конкретной информации, и ограничиваются
лишь туманными аргументами. Пример этого – попытка «опровержения» информации
с сайта TalkOrigins со стороны не назвавшего себя автора из Creation Wiki (см.
выше). У меня сложилось ощущение, что креационисты не наступают, воинственно
размахивая предметами культа, а как-то злобненько «огрызаются», боясь переходить
в наступление.
Что говорят об эволюции усатых китов эволюционисты? Вернусь к статье
Эриха Фитцжеральда:
Филогения Mysticeti, представленная здесь [схема
эволюции усатых китов будет приведена далее - В. П.], согласуется с концепцией
взрывообразной радиации высших китообразных в течение олигоцена,
особенно в раннем олигоцене (28,5 – 34,0 млн. лет назад; Fordyce 1992;
Nikaido и другие. 2001; Sasaki и другие. 2005). Эта адаптивная радиация
совпадала по времени с глобальными изменениями климата, и увеличила продуктивность
и разнородность океанских местообитаний, связанных с установлением антарктического
кругополярного течения после заключительного распада Гондваны и изоляции
Антарктиды (Fordyce 1980, 1992; Nikaido и другие. 2001). Предыдущие исследования
делали упор на первостепенное значение фильтрующего типа питания как
толчка для начальной радиации усатых китов (Fordyce 1980; Fordyce 1992;
Fordyce и Barnes 1994; Barnes и др. 1995; Fordyce и Muizon 2001; Nikaido
и др. 2001; Берта и др. 2006); но нестандартная морфология и базальное
положение Janjucetus (и Mammalodon) в филогении усатых
китов, которые оба, предположительно, не питались путём фильтрации, предполагают,
что
начальная радиация усатых китов не была связана с эволюцией фильтрующего
типа питания.
Эволюции фильтрующего типа питания предшествовала умеренная радиация исходных Mysticeti, которая включала эволюцию мелких китообразных, конвергентно сходных с некоторыми мезозойскими морскими рептилиями и ныне живущими настоящими тюленями. Потому олигоценовая эволюция усатых китов выглядит характеризующейся значительно большими разнообразием и разнонаправленностью, и, таким образом, сложностью, чем считалось ранее. Усатые киты, несомненно, заняли широкий диапазон экологических ролей в ходе своей ранней эволюционной истории, являясь значительно более многообразными, чем могли бы показаться их представители от миоцена до современности. |
В конце XX и начале XXI века появилось множество новых находок
доисторических китообразных. Вполне естественно, что не все они являются предками
современных
китов, но их изучение позволяет более полно представить направления эволюции
этих млекопитающих. Иногда открытия оказываются и вовсе неожиданными. Так,
в начале 2006 года в Австралии были обнаружены ископаемые остатки совершенно
неожиданного вида китообразных. Общая длина этого животного оценивается приблизительно
в 3,5 метра на основании размеров черепа. Череп животного сохранился очень
хорошо, а от туловища известны лишь разрозненные кости: шейные позвонки с 1-го
по 3-й, лопатки, два ребра и лучевая кость. Это китообразное отличается от
ранее известных
представителей
группы огромными глазами, значительно
превышающими
глаза любого
другого известного
до того момента вида (для сравнения: глаз дельфина афалины, имеющего примерно
такую же длину, не крупнее глаза собаки). Лишь у странного, похожего на моржа
с бивнями разной длины, кита Odobenocetops глаза имеют приблизительно
такой же относительный размер.
По месту находки и фамилии подростка, обнаружившего остатки животного, новый
род и вид ископаемых китов был назван Janjucetus hunderi. При внимательном
изучении этого животного было установлено, что это представитель усатых китов.

Реконструкция Janjucetus, выполненная Bryan Choo
«Улыбочку!» На черепе Janjucetus (рисунок из описания Э. Фитцжеральда) хорошо заметны передние зубы, предназначенные для вырывания кусков мяса из добычи. Масштабная линия 100 мм. Обозначения: nuc – затылочный гребень, sac – теменной гребень, zmx – скуловой отросток верхнечелюстной кости, la – слёзная кость, vo – сошник.
В описании вида, которое составил Эрих Фитцжеральд, приводятся такие данные об особенностях этого животного:
Janjucetus разделяет с остальными усатыми
китами два ключевых синапоморфных признака: (i) направленный латерально
[в боковом
направлении – В. П.] зигоматический отросток верхнечелюстной кости с
наклонной передней поверхностью, ясно отделяющей его от ростральной части
верхнечелюстной кости; и (ii) широкие луковицеобразные нижне-затылочные гребни. Janjucetus представляет
собой исходную ветвь в пределах подотряда Mysticeti, но не является наиболее
базальным представителем усатых китов
(…).
|
В свете вывода о систематическом положении Janjucetus совершенно неожиданной стала выглядеть другая особенность этого кита, заметная буквально невооружённым глазом: Janjucetus имел полный рот острых зубов с зазубренными краями, да таких, что иная акула позавидует. Эта особенность резко контрастировала с предыдущими предположениями палеонтологов о том, что определяющим моментом в разделении усатых и зубатых китов было появление фильтрующего аппарата – китового уса.
Череп и зубы Janjucetus производят устрашающее впечатление. Масштабная линия на рисунке (a) – 100 мм, на рисунках (b – e) – 20 мм. Сокращения на рисунке: sq – чешуйчатая кость, ju – скуловая кость, pal – нёбо; зубы (заглавными обозначены зубы верхней челюсти, строчными – нижней, цифра указывает номер зуба, считая от симфиза – места соединения правой и левой половин челюсти): I – резцы, C – клыки, P – предкоренные, M – коренные.
Был ли Janjucetus hunderi фильтратором? Этот фундаментальный
вопрос, в контексте филогенетического положения Janjucetus, имеет широкие
последствия для интерпретации эволюции усатых китов. С питанием путём
фильтрования функционально соотносятся несколько морфологических особенностей
усатых китов: (i) удлинённые и расширенные рострум и верхнечелюстные
кости по отношению к общей длине черепа (увеличивает объём ротовой полости
и площадь фильтрующих образований); (ii) уменьшенный размер глазниц и
глаз, а глазницы направлены вбок (зрение не столь критично для выбора
и поимки добычи); (iii) крупный, наклоненный в переднем направлении надглазничный
щиток (увеличивает площадь прикрепления надосевой мускулатуры, поднимающей
голову вверх, и увеличивает сопротивление наклону головы вниз, когда
рот открыт); (iv) редуцированные/отсутствующие зубы, и (v) рострум и
верхнечелюстные кости не утолщённые и не сильно окостеневшие (регуляторы
течения воды в ротовую полость, но не активные элементы захвата добычи
(…).У Janjucetus нет ни одной из перечисленных особенностей, функционально
соотносящихся с методом ловли добычи путём фильтрования. Нёбная поверхность
верхней челюсти Janjucetus лишена питающих отверстий и борозд, ассоциирующихся
с питающими китовый ус сосудами (…). Это сильное свидетельство против
того, что у Janjucetus был китовый ус, даже в начальной форме.
|
|
Пасть Janjucetus отличается множеством особенностей, свидетельствующих о том,
что это был хищник-макрофаг, то есть, существо, питающееся, подобно современным
зубатым китам, крупной одиночной добычей – большими рыбами, в том числе акулами,
головоногими моллюсками, а также, возможно, пингвинами и мелкими китообразными.
Остатки Janjucetus датированы поздним олигоценом, а первые пингвины существовали
на Земле уже в эоцене, и, возможно, ранее. Поэтому хищный усатый (но фактически
«безусый») кит вполне мог отыскать себе достаточно добычи. Совместно с черепом
Janjucetus найдены остатки акул, скатов, костных рыб, неидентифицированных
морских птиц, мелких зубатых китов Prosqualodon sp. и неидентифицированный
вид, возможно, из семейства Waipatiidae, а также «безусого» усатого кита Mammalodon
colliveri.
Исследователи сравнивают Janjucetus hunderi с ныне живущим морским
леопардом (Hydrurga leptonyx) – плотоядным видом тюленей, нападающим
на другие виды тюленей, а также на пингвинов. Во всяком случае, пропорции черепов
этих двух видов обнаруживают
внешнее сходство. Разумеется, это лишь поверхностное сходство, результат
конвергенции: китообразных и ластоногих (по сути – три семейства хищников,
ведущих водный образ жизни) связывает весьма отдалённое родство.
Зубы нового
китообразного покрыты многочисленными бороздками, края зубчиков на коренных
зубах были лезвиеподобными, а форма костей черепа свидетельствует
о том, что челюстная мускулатура этого кита позволяла ему сжимать челюсти,
развивая значительное усилие. Резцы и клыки этого китообразного (его зубная
система не гомодонтная; зубы чётко различаются по форме) собраны в своеобразную
«розетку» на концах челюстей.
Находка этого вида китов не осталась незамеченной креационистами, что лишний
раз говорит о том, что они понимают глобальную роль и биологический смысл процесса
эволюции (фигурально, разумеется): если не приспосабливаться к новым условиям,
можно «вымереть» для читателя, оказавшись на обочине научного прогресса.
Австралийский креационист Стивен Джонс, анализируя в своём блоге статьи про
своего знаменитого ископаемого «земляка», вывел ряд вопросов, которые возникли
у него в связи с этим примечательным и неожиданным открытием. Вполне естественно
для него, что в исполнении креациониста эти вопросы звучат в упрёк сторонникам
теории эволюции. Но насколько справедливы эти упрёки?
Стивен Джонс «Происхождение китового уса: вот ещё одна проблема для дарвинизма»:
[Обратите внимание: 1) Прежде «считали,
что усатые киты быстро приобрели в процессе эволюции китовый ус …
после отделения от общего с зубатыми китами предка почти 40 миллионов
лет назад;» хотя
2) на ~15 миллионов лет позже этот «25-миллионолетний ископаемый» «усатый
кит даже не имел китового уса» (выделено мною)]
|
[Обратите внимание: 3) то, что это был «древний
усатый кит, настолько примитивный, настолько первичный, что он имел
зубы» приведёт к необходимости «перерисовать эволюционные схемы во
всём мире», поскольку 4) «фактически этот древний хищник был усатым
китом.» (выделено мною)]
|
И чего здесь удивительного? В очередной раз креационисты строят
удивлённо-негодующее лицо, пеняя эволюционистам, что, мол, их точка зрения
обратно поменялась. Но
точка зрения меняется не просто так, а на основании вновь появляющейся информации.
Прежние построения могли соответствовать тому объёму информации, который был
известен до момента, когда они были сделаны. Вполне естественно, что новые
данные могут или подтвердить построения, или опровергнуть их. Честно говоря,
иногда мне очень хочется, чтобы такого рода креационисты попали к какому-нибудь
следователю, который на основании мелочного совпадения или чего-то подобного
выдвинул бы против них обвинение в чём-нибудь малоприятном и грозящем отсидкой
в местах, не столь отдалённых, на солидный срок. А затем, по мере появления
новых фактов, свидетельствующих в пользу невиновности этих
товарищей,
просто
произвольно отметал бы эти факты, не принимая их во внимание и сохраняя фанатично-тупую
верность первоначально выдвинутой версии. Думаю, тогда бы креационно настроенные
товарищи, попав в такой «переплёт», по-другому посмотрели бы на необходимость
пересмотра прежних версий в свете новых фактов, причём не только у эволюционистов,
но и у себя, любимых. Правда, боюсь, что в данном случае «горбатого могила
исправит».
Возращаясь к «безусым» усатым китам, можно предположить, что изначально была
выдвинута простейшая гипотеза: усатые киты отделились от зубатых, когда приобрели
китовый ус и стали фильтраторами. Такая гипотеза могла быть выдвинута, исходя
из знаний о современных китах, принадлежащих к отряду Mysticeti: у них у всех
есть китовый ус. Она не противоречила знаниям об усатых китах, имевшимся у
учёных где-то в первой половине ХХ века: все известные на тот момент китообразные,
современные и ископаемые, имели китовый ус, или были анатомически сходны с
современными усатыми китами. Вполне естественно, что у данной группы китообразных,
помимо наличия китового уса, были скелетные признаки, отличающие их от прочих
групп древних и современных китов, причём не всегда связанные именно с наличием
китового уса и способностью к фильтрованию пищи из воды. Но далее были открыты
киты, у которых скелетные признаки соответствовали таковым у усатых китов,
однако скелет не нёс особенностей, связанных с наличием китового уса. Возникает
закономерный вопрос: как классифицировать таких китов? На зубатых китов они
«не тянут» строением скелета, а на усатых – «отсутствием присутствия» китового
уса. Если учитывать, что они оказываются древнее несомненных усатых китов современного
типа, можно прийти к заключению, что усатые киты как самостоятельная группа
животных, объединённая общими анатомическими особенностями, отделились от общего
с зубатыми китами предка до того, как у них появился китовый ус сам по себе.
И само название «усатые киты» они получили, фактически, на основании неполных
знаний об этой группе животных: на момент именования усатых китов брались во
внимание лишь особенности современных представителей этой группы, у которых
у всех был китовый ус. Получается, как с выражением «белый, как лебедь»: оно
было верным до открытия чёрных лебедей.
Ситуация с усатыми китами – не единственная в зоологии. Так, современные носороги
были названы «носорогами» на основании того, что у всех современных представителей
этой группы есть рог. Даже подотряд непарнокопытных, к которому относятся носороги,
называется Ceratomorpha – «подобные рогатым». Но древние носороги далеко не
все были рогатыми, и потому в палеонтологической и научно-популярной литературе
встречается выражение «безрогий носорог». Ничего, не режет слух? Во всяком
случае, претензий к названию «носорог» со ссылкой на строение ископаемых видов
я в креационистской литературе не встречал.
Итак, китовый ус в свете данных палеонтологии выглядит лишь частным признаком
одной из групп животных, относящихся к подотряду Mysticeti. И его наличие в
одной группе этих животных не выглядит настоятельной обязанностью для прочих
животных, входящих в этот подотряд, немедленно эволюционировать в направлении
развития
китового
уса. Разделение древних Mysticeti (да и любых других живых организмов) на самостоятельные
семейства означает, что разные группы этих животных вели заведомо различный
образ жизни, отличаясь, в том числе, используемыми источниками пищи. Поэтому
наличие зубастого Janjucetus наряду с «истинно усатыми» китами-фильтраторами
не вызывает удивления с точки зрения теории эволюции.
[Обратите внимание: 5) что «исходные особенности усатых китов не включали фильтровальный аппарат.» (выделено мною)] |
И опять, чего тут непонятного? «Лыко да мочало, начинай сначала».
И приходится начинать.
Являются ли усатые киты отдельной, хорошо отличающейся особенностями анатомии,
группой в пределах отряда китообразных? Несомненно. Справедлив ли вывод о
том, что «усатый кит = зубатый кит – зубы + китовый ус»? («Коммунизм
есть советская власть плюс…») Вряд ли. Достаточно взглянуть на признаки,
по которым
Janjucetus был причислен к усатым китам. Эти признаки – особенности
черепа, не связанные непосредственно с наличием китового уса. Сам по себе
китовый ус не сохраняется в ископаемом состоянии, поэтому прямых свидетельств
его наличия у тех или иных китообразных можно и не получить, особенно если
скелет фрагментарный. Тем не менее, у усатых китов, как видно, есть иные
признаки строения, объединяющие их и противопоставляющие усатых китов зубатым
китам и археоцетам.
[Обратите внимание: 6) заявление о том, что «Этот новый вид – не предок современных усатых китов», потому что «Самые древние ископаемые остатки усатого кита … на 10 миллионов лет древнее» (выделено мною)] |
… И вся сложность ситуации, в которую попадает Стивен Джонс с таким объяснением, заключается в том, что на момент описания Janjucetus уже был известен кит Eomysticetus с китовым усом, возрастом около 30 миллионов лет. То есть, кит с китовым усом уже появился до того, как на арену жизни выплыл зубастый Janjucetus. Иными словами, в эпоху существования Janjucetus в океанах уже плавали киты, обладавшие китовым усом. Если бы Стивен Джонс поднялся на уровень выше нападок на заведомо упрощённые популярные статьи, и обратился к собственно описанию Janjucetus, имеющемуся в свободном доступе в Интернете. Там есть интереснейшая схема, включающая, в том числе, распределение во времени некоторых видов усатых китов. Вот она, с моим переводом обозначений на русский язык (в оригинале англоязычная, разумеется):
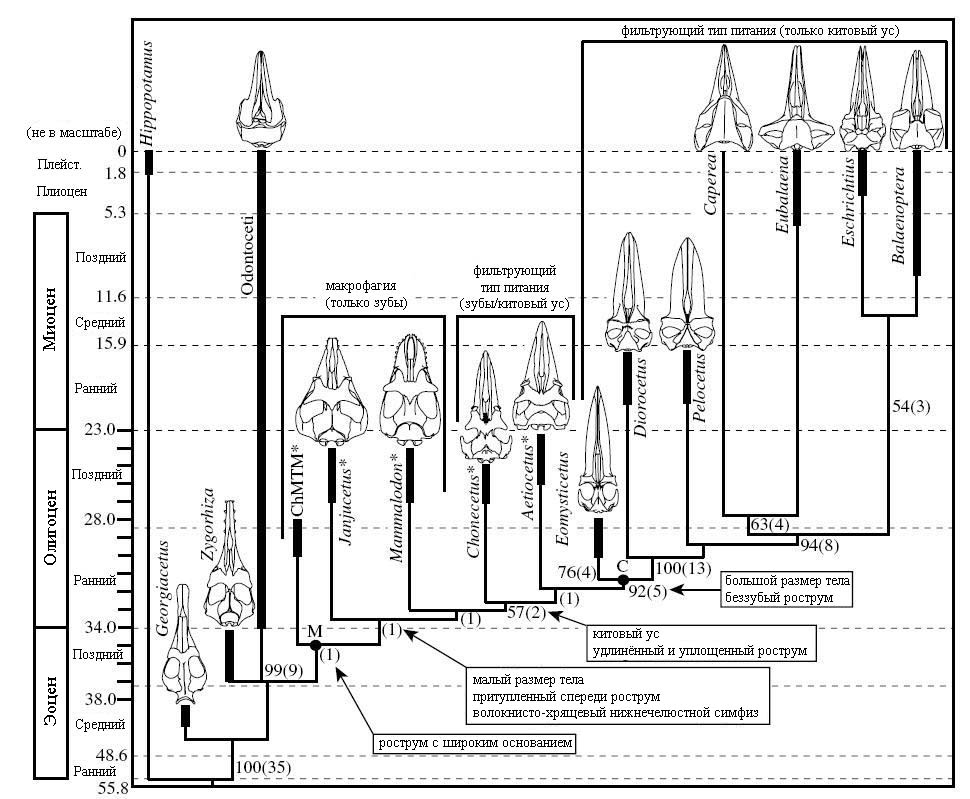
Схема,
отражающая этапы ранней эволюции усатых китов, взята из описания Janjucetus.
Подпись к ней гласит:
«Филогения и стратиграфическая летопись Mysticeti, включая Janjucetus,
и эволюция экологии питания у усатых китов. Филогенетические отношения Janjucetus основаны
на строгом соответствии трёх филогенетических древ, построенных на основании
анализа парсимонии [сходства признаков - В. П.] 266 признаков
в 26 родах (некоторые таксоны удалены из древа). Реконструкции черепов китообразных
показаны как вид сверху. Признаки, существенные для эволюции питания усатых
китов, размещены
в узлах, где они появляются. Таксоны, помеченные *, представляют зубастых усатых
китов. Числа и числа в скобках, указанные в узлах, представляют собой значения bootstrap и
поддержки ветвей, соответственно. Закрашенные круги обозначают названные ветви.
Жирные чёрные
линии на ветвях представляют собой линии пределов
стратиграфической ошибки для их соответствующей родословной
линии. Возрасты даны в миллионах лет, шкала времени в одном масштабе только
для позднего эоцена и олигоцена. Шкала времени дана по Gradstein и др.
(2004). Сокращения: ChM TM – зубастые усатые киты Чарльстонского Музея (Charleston
Museum toothed mysticetes); C – Chaeomysticeti; M – Mysticeti.»
[Заметьте, что это 8) «опровергает представления
о том, что все исходные усатые киты питались путём фильтрования», и
9) «что происхождение и исходная радиация усатых китов были связаны
с эволюцией фильтрующего типа питания» (выделено мною).
|
И чего в этом катастрофического? Я уже упоминал не раз о том,
что выводы науки строятся на основе знаний, имеющихся на момент формулирования
такого вывода,
и считаются истинными, пока не противоречат имеющимся на тот или иной момент
времени фактам. И они могут быть опровергнуты каким-либо новым фактом, который
заставит пересматривать построенные связи и сформулированные умозаключения.
Поскольку окончательный ответ при научном познании мира неизвестен, наука легко
расстаётся с выводами, которые признаны неверными. У религии другая позиция:
правильный ответ уже известен, и надо лишь подогнать под него решение.
Честно, я рад и за столь интересную находку Janjucetus, и за выводы,
которые были сделаны на её основании, даже если они заставили отвергнуть какие-то
более
ранние заключения.
Я не считаю отдельным вопросом заявление Фитцжеральда о том, что это ископаемое является свидетельством того, что «эволюция признаков, связанных с питанием путём фильтрования, была постепенной», поскольку оно демонстрирует полную противоположность, по крайней мере, в дарвиновском смысле, «медленному градуализму»: ... Если он был свидетельством последовательности «слегка различающихся шагов» к «фильтрующему типу питания» от самых ранних усатых китов, по путям, реальность которых отстаивал Дарвин (...), то в этом случае он должен бы поддерживать это заявление. Но до тех пор, пока этот наиболее морфологически «древний усатый кит, настолько примитивный, настолько первичный, что он имел зубы» остаётся существовавшим ~25 млн. лет назад, т. е. через ~15 млн. лет после предположительного разделения линий зубатых (Odontoceti) – усатых (Mysticeti) китов, которое я принимаю как имевшее место, до этих пор это будет свидетельством того, что появление китового уса произошло в более короткие временные сроки, и потому не было, или, по крайней мере, было менее «постепенным» в вышеназванном дарвиновском смысле. Как говорилось в первой заметке [в блоге обсуждаемого автора - В. П.], до этой находки «Многие учёные считали, что усатые киты быстро приобрели в процессе эволюции китовый ус для питания мелкой рыбой и планктоном после отделения от общего с зубатыми китами предка почти 40 миллионов лет назад» (выделено мною)!] |
А теперь остынем после столь громкого «разоблачения нехороших
эволюционистов», и немного подумаем. В первом приближении смешным выглядит
обвинение в том,
что некий процесс идёт «не по Дарвину». Я лишь напомню о том, что Дарвин работал
над своим «Происхождением видов...» лет эдак сто пятьдесят тому назад, и, вполне
естественно, не мог знать о каких-то более поздних научных открытиях. Поэтому
обвинение в адрес сторонников теории эволюции в «отступничестве» от Дарвина
можно пропустить мимо ушей. В противном случае придётся обвинять средневековых
математиков Европы в том, что на одном из этапов развития математики они не
знали числа «ноль».
Кстати, с математикой у товарища всё в порядке. Не нужно большого ума, чтобы
вычесть из сорока или пятнадцать, или двадцать пять, а затем потрясать воображение
читателя полученными результатами. Здесь напрашивается совершенно другой вопрос:
а нужна ли эта «высшая математика» применительно к находке Janjucetus?
Если внимательно посмотреть на текст заметки, предваряющий замечание 6, можно
увидеть две интересных детали. В статье говорится:
... Тем не менее, этот новый вид – не предок современных усатых китов. Самые древние ископаемые остатки усатого кита, найденные в Антарктике, на 10 миллионов лет древнее. Вместо этого J. hunderi принадлежит к неизвестной боковой линии эволюции, существования которой никто не ожидал. |
Не надо особенно напрягать логику, чтобы выделить три момента:
1) Janjucetus - не предок современных китов.
2) Первые усатые киты древнее его.
3) Janjucetus - представитель боковой линии (в оригинальном
тексте сказано "sidetrack") усатых китов.
Что сделал Стивен Джонс? Он сам соединил первое и второе утверждение логической
связью («не предок, потому что есть древнее»), которая фактически отсутствует
в оригинальном тексте, и сознательно или случайно не обратил внимания на
следующее утверждение, которое
поясняло,
почему Janjucetus не
предок современных китов. Так и родился ещё один миф.
Теперь самое время вернуться к его последнему утверждению. Знание математики
на уровне первого класса начальной школы – это очень важно и нужно. Но начинать
складывать
и вычитать разного рода циферки можно только в одном случае: если Janjucetus находится
на линии, ведущей непосредственно к современным китам. Тогда его анатомия
будет
важной «узловой точкой» при расчёте темпов появления китового уса. Но у нас есть
упоминание о том, что Janjucetus представляет собой боковую линию в
эволюции усатых китов. То есть, он НЕ находится на линии, ведущей от самых ранних
«безусых» усатых китов к современным усатым китам. И тогда какого дьявола надо
складывать и вычитать циферки?
Если бы Стивен Джонс был чуть-чуть любопытнее и действительно хотел бы оценить
место и значение Janjucetus в эволюции усатых китов,
он
должен бы
по хорошему счёту поискать
составленное
Эрихом Фитцжеральдом описание
Janjucetus. Оно вполне доступно и его можно прочитать бесплатно. Тем
более, оно сделано на родном Стивену Джонсу английском языке. И в нём можно найти
немало интересного. Например, вышеприведённую схему родственных отношений ископаемых
китов. И из неё можно узнать крайне любопытные вещи. Например, ниже (стало быть,
древнее) Janjucetusна схеме находится другое животное, Eomysticetus.
Пояснительные подписи позволяют установить, какие признаки были характерны для
этого животного. Даже на схематичном
рисунке видно, что череп этого животного обладал формой, характерной для обладателей
китового уса. То есть, к моменту появления Janjucetus в океане уже плавали
киты,
обладающие китовым усом! И это низводит математические построения Стивена Джонса
к нулю.
Вообще, этот пример сильно перекликается с эволюцией лошади, о которой так любят
говорить креационисты. Только подход здесь прямо противоположный. В случае
с серией ископаемых лошадей креационисты
«изобличают» нехороших эволюционистов, демонстрируя наличие нескольких родословных
линий лошадей, из чего, впрочем, сами эволюционисты не делают секрета: в книгах
эволюционного направления родословное древо лошадей исправно ветвится. В случае
с усатыми китами виден прямо противоположный
подход: все ископаемые киты, принадлежащие к одной группе высокого систематического
ранга (в данном случае подотряду),
волевым решением спихиваются
в
одну цепочку, даже если они принадлежат к разным семействам, чётко отличающимся
друг от друга. Всё. Можно делать далеко идущие выводы.
Китовый ус уникален для усатых китов, но зубатые киты, не имея его, тем не менее, также являются китами. Поэтому данный признак нельзя считать каким-то основополагающим: это частная адаптация одной группы китообразных. В природе существует много аналогов этого приспособления: например, тонкие волосовидные зубы птерозавра Pterodaustro, росшие на нижней челюсти. С помощью этих зубов птерозавр отфильтровывал мелких животных, находясь на берегу и черпая воду челюстями, или же «чиркая» клювом по поверхности воды в полёте. Среди современных животных самый близкий аналог китового уса представляет собой фильтровальный аппарат в клюве гусеобразных птиц - уток и гусей. Именно с цедильным аппаратом гусей и уток сравнивал китовый ус Чарльз Дарвин, рассуждая о возможности его развития в книге «Происхождение видов...» (6-е издание, глава VII «Различные возражения против теории естественного отбора»):
Гренландский кит — одно из самых замечательных животных
в свете, и так называемый китовый ус представляет одну из его наиболее
выдающихся особенностей. Китовый ус состоит из ряда пластинок, которые,
находясь приблизительно в числе 300 на каждой стороне верхней челюсти,
расположены очень тесно друг к другу поперек продольной оси ротовой полости.
С внутренней стороны главного ряда находится несколько добавочных. Концы
и внутренние края всех пластинок расщеплены в крепкие щетины, которые
покрывают все огромное нёбо и служат для просачивания или процеживания
воды и для задержания мелкой добычи, которой питается это огромное животное.
Относительно китового уса м-р Майварт замечает, что, «однажды достигнув такой величины и такого развития, при котором он вообще стал полезен, он мог сохраняться и увеличиваться в нужных размерах уже только под влиянием одного естественного отбора. Но как могло быть получено самое начало такого полезного образования!» В ответ на это можно спросить, почему бы отдаленные предки [современных] китов, обладающих китовым усом, не могли иметь рта, построенного более или менее сходно с клювом утки, снабженным [роговыми] пластинками? Утки, подобно китам, кормятся, процеживая грязь и воду, и это семейство иногда называли Criblatores, т. е. цедильщики. Я надеюсь, что не буду ложно понят в том смысле, будто я говорю, что предки китов имели в действительности рот с пластинками подобно клюву утки. Я хочу только сказать, что это нельзя считать невероятным и что огромные пластины китового уса гренландского кита могли постепенно развиться из таких пластинок, которые на каждой стадии развития были полезны для их владельца. (Выделено мною - В. П.) |
Во времена Дарвина ископаемые китообразные были плохо известны, и он рассуждал о возможном развитии фильтрационного аппарата на гораздо более доступном примере – птицах семейства утиных. Дарвин приводит примеры разной степени развития фильтрующего аппарата в клювах утиных птиц – от примитивного и малоэффективного в роли фильтрующего приспособления до сложно устроенного и позволяющего эффективно фильтровать воду, демонстрируя прямую связь между его развитием и образом жизни. Не проводя прямых аналогий между строением клюва утки и пасти кита, Дарвин показывает, что каждая стадия развития цедильного аппарата эффективна при том образе жизни, который ведёт его обладатель. Далее Дарвин пишет:
Вернемся к китам. Hyperoodon bidens лишен настоящих,
пригодных для работы, зубов, но его нёбо затвердело и снабжено, по Ласепеду,
небольшими, неравными, жесткими роговыми выступами. Поэтому нет ничего
невероятного в предположении, что какая-нибудь из ранних форм Cetacea
имела нёбо, покрытое подобными же ороговевшими выступами, несколько более
правильно расположенными, которые, подобно буграм на клюве гуся, помогли
схватывать и размельчать пищу. Но если так, то едва ли можно отрицать,
что указанные выступы, посредством изменений и естественного отбора,
могли превратиться в пластинки. Постепенное развитие их, наблюдаемое
у ныне существующих Cetacea, приводит нас к огромным пластинам китового
уса гренландского кита. У нас нет ни малейшего основания сомневаться
в том, что каждая ступень в этом ряде могла быть настолько же полезна
некоторым древним Cetacea при постепенном изменении функций в процессе
развития медленно изменяющихся частей, насколько полезны последовательные
изменения клюва у разных существующих ныне членов семейства утиных. Мы
должны помнить, что каждый вид утиных подвержен жестокой борьбе за существование
и что строение каждой части тела птицы должно быть хорошо приспособлено
к условиям ее существования.
|
Стивен Джонс "The origin of whale baleen: Yet another problem for Darwinism #2" :
что 1) в типичном «скользком» дарвинском стиле «который
не может быть согласован даже с обычной интеллектуальной честностью»
(Darlington, C.D., «Место Дарвина в истории», Basil Blackwell: Oxford
UK, 1959, стр. 60), Дарвин риторически считал спорный вопрос решенным,
утверждая, «едва ли можно отрицать, что указанные выступы, посредством
изменений и естественного отбора, могли превратиться в пластинки, столь
же развитые, как таковые у Египетского гуся» (выделено мною); и 2) возможно
надеялся, что никто из читателей не заметит, что выбранный им в качестве
примера Hyperoodon bidens не развил китового уса!
|
Насчёт этого самого кита (в современной систематике это карликовый кашалот,
или когия) вывод Стивена Джонса поражает глубиной и интеллектуальной насыщенностью.
Сто с лишком лет пелена закрывала глаза дарвинистов, и они слепо заучивали
этот пример, не пытаясь сунуть руку в пасть этого кита, чтобы убедиться, что
он не развил китового уса…
На самом деле всё намного проще: Дарвин привёл пример этого китообразного как
аналог возможного предка усатых китов в том понимании эволюции этих животных,
которое существовало в его времена. Он и не ждал развития истинного китового
уса у этого кита!
Дарвина, который подкреплял свои утверждения реальными фактами,
наблюдаемыми в природе, креационисты пытаются обвинять в «скользкости» и «нечестности».
Хорошо, но что в таком случае можно сказать о креационистах, которые лепят
свои обвинения в адрес сторонников
теории
эволюции едва ли не на пустом месте, злостно искажают и замалчивают факты, и
не поднимаются при обсуждении выше статей в популярных газетах и журналах???
Не лучше ли заняться собой в первую очередь, чем лить помои на человека, который
к тому же много лет как умер?
Суть состоит в том, что это ископаемое показывает,
что Дарвин был неправ в своём утверждении, что «отдаленные
предки [современных] китов, обладающих китовым усом, … имели рот, построенный
более или менее сходно с клювом утки, снабженным [роговыми] пластинками».
Оно показывает, что фактически «отдаленные предки [современных] китов,
обладающих китовым усом ... имели рот» с «большим набором зубов без
всякого свидетельства наличия гребневидной бахромы, используемой другими
усатыми китами, чтобы фильтровать свою пищу из морской воды» (…).
|
Нет, народ… Что-то мне не нравится такая дискуссия. Я понимаю Майварта, я понимаю Дарвина. Они жили и творили в 19 веке. И они оперировали совокупностью сведений, которая была известна науке в то время. Но на дворе, напомню, начало XXI века, новые технологии, новые данные, новые методы исследований. Поэтому я решительно не понимаю Стивена Джонса, который в заметке от августа 2006 года продолжает «доказывать» «несостоятельность теории Дарвина» данными сомнительной свежести. По хорошему счёту, это нужно делать на основании информации, которая актуальна, по меньшей мере, на 2004 – 2005-й годы.
Более того, пока естественный отбор не снабдил этого
усатого кита китовым усом, то что-то иное, нежели естественный отбор должно быть истинным объяснением того, почему одна родословная линия
усатых китов развила китовый ус, а другая линия не сделала этого.
|
Что ж, пока не было найдено ископаемых остатков древнейших
усатых китов, считалось, что отделение ветви усатых китов от общих с зубатыми
китами предков произошло именно при формировании китового уса. Выше это признавал
и сам Стивен Джонс, и не стоит отрицать того, что именно так обстояли дела
в науке. Почему? Потому что данные, имевшиеся на момент формулирования данного
утверждения, не противоречили ему, и не было основания предполагать что-либо
другое. Однако, наука тем отличается от религии, что при появлении нового фактического
материала наука легко пересматривает прежние выводы, и при признании их недостоверности
легко отбрасывает их, делая частью истории. Может ли религия похвастать этим?
Не думаю. Хотя, похоже, гелиоцентрическая система и шарообразность Земли всё
же признаны.
Находки ископаемых китообразных в настоящее время указывают на то, что ближайший
аналог самых ранних усатых китов в плане способа питания – не гусеобразная
птица (фильтратор массовой микроскопической добычи), а тюлень-крабоед (хватальщик
относительно крупной стайной добычи). Часть китообразных, которые по строению
черепа относились к усатым китам, продолжили приспособление к питанию крупной
добычей. Так на арене жизни появились Mammalodon и Janjucetus. Другие киты
стали приспосабливаться к питанию мелкой стайной добычей – мелкой рыбой, ракообразными,
возможно, мелкими кальмарами. У них развился китовый ус.
Осуждать Дарвина и Майварта в том, что они не располагали информацией о «зубастых
усатых китах», которая стала известной лишь недавно, в конце XX и начале XXI
века, бессмысленно и смешно. Их выводы могут быть хоть тысячу раз неверными
в свете новейших находок, но в свете той совокупности информации, которая была
известна во 2-й половине XIX века, их аргументы выглядят вполне обоснованными,
и на это надо обязательно делать скидку. А вот какого дьявола господин Стивен
Джонс не упомянул в своих рассуждениях (от августа 2006 года) кита Llanocetus
denticrenatus,
об открытии которого было сообщено в декабре 1989 года – это уже другой вопрос...
Здесь
попахивает
чем-то одним: либо желанием скрыть информацию и составить ложное представление
о положении вещей, либо банальным невежеством самого автора. Второй случай
не предосудителен: в свете полного объёма информации по данной проблеме Стивен
Джонс покажется просто громкоголосым дурачком, который не до конца разобрался
в вопросе, и всё. Но в первом случае всё гораздо хуже, поскольку такой нечестный
приём ведения
дискуссии
явно рассчитан
на неосведомлённого читателя, не владеющего полной информацией по данному довольно
узкому и специфическому вопросу. Конечно, тогда такой читатель поверит в наличие
«чего-то иного, нежели естественный отбор», что и требовалось этому товарищу.
А можно ли обойтись без всяких «потусторонних» объяснений? Думаю, вполне. Для
начала стоит перечитать ещё раз слова Дарвина, и уяснить одну простую вещь:
«У нас нет ни малейшего основания сомневаться в том, что каждая ступень в этом
ряде могла быть настолько же полезна некоторым древним Cetacea при постепенном
изменении функций в процессе развития медленно изменяющихся частей, насколько
полезны последовательные изменения клюва у разных существующих ныне членов
семейства утиных». Дарвин специально подчёркивает главное условие развития
какого-либо приспособления: на любом этапе своего развития оно должно быть
полезным и давать преимущества его обладателю.
GATESY, J. **; DEMERE, T.; MCGOWEN, M.; BERTA, A.; U. C. Riverside; San Diego Natural History Museum; U. C. Riverside; San Diego State U.: Пошаговая эволюция фильтрующего типа питания у усатых китов Синтез молекулярных и палеонтологических данных может предложить уникальные взгляды на историю комплексной адаптации. Здесь мы используем объединенный филогенетический анализ морфологических признаков и данных о последовательностях митохондриального и ядерного геномов, чтобы отследить крупное эволюционное преобразование у Mysticeti (усатые киты). Происхождение китового уса и потеря зубов коррелируют с эволюцией исключительно фильтрующего способа питания у усатых китов. Переход от несущих зубы челюстей к беззубым, покрытым китовым усом верхним челюстям ныне живущих усатых китов, однако, не был выполнен за один эволюционный шаг. Новое свидетельство от позднеолигоценового усатого кита, имеющего зубы, Aetiocetus weltoni документирует присутствие нёбных питающих отверстий у этого архаичного таксона; сходным образом располагающиеся отверстия на нёбе содержат сосуды, которые питают китовый ус у ныне живущих усатых китов. В контексте нашего объединенного филогенетического анализа эти наблюдения наводят на мысль, что у Aetiocetus и, вероятно, других исходных усатых китов китовый ус существовал совместно с функциональными зубами. Чтобы дополнить данные, основанные на ископаемых, мы секвенировали три гена, имеющих ключевое значение для выработки дентина/эмали у ныне живущих усатых китов и внешнего таксона. Enamelin и ameloblastin, экспрессия которых происходит в процессе развития ткани эмали у млекопитающих, имеют нонсенсные мутации в нескольких линиях усатых китов и, очевидно, нефункциональны. Одновременный филогенетический анализ палеонтологических и молекулярных данных подразумевает, что ранние зубастые усатые киты приобрели в процессе эволюции китовый ус, и что минерализованные зубы взрослых особей были впоследствии утрачены за промежуток времени в несколько миллионов лет в одной родословной линии Mysticeti, ведущей к ныне живущим усатым китам, у которых остались дегенерировавшие гены эмали и рудиментарные эмбриональные зачатки зубов. |
Честно говоря, в литературе я встретил очень мало ответов
на вопрос: «каким образом прошёл переход к фильтрующему типу питания?». Поэтому
попробую высказать свою точку зрения на этот вопрос.
Может ли крупный кит питаться маленькой добычей? Среди современных крупных
китов кашалот даёт, как ни странно, возможность ответить на этот вопрос. Обычно
в популярной литературе этого кита изображают закусывающим гигантским кальмаром.
Но это, хотя и известная, но далеко не столь часто встречающаяся добыча. Естественно,
на китобойных станциях при разделке кашалотов из их желудков извлекали таких
моллюсков. Но также известно, что часто в желудках кашалотов находили большое
количество сравнительно мелких кальмаров и рыб. Даже если рыба достигала порой
метровой длины, она всё равно мала по сравнению с многотонным кашалотом. Современные
зубатые киты часто питаются мелкими рыбами и кальмарами, просто засасывая их
ртом. У клюворылых китов значительно редуцированы зубы – иногда всего лишь
до двух крупных зубов, явно не приспособленных для схватывания такой добычи,
как рыба и кальмары.
У самых ранних усатых китов («усатых» скорее по анатомии скелета, а не по наличию
китового уса) зубы достаточно редко сидят на челюстях. Можно предположить,
что они постепенно утрачивали функцию захвата и непосредственного удержания
добычи, и служили скорее для «запирания» рта. Кстати, у современной китовой
акулы мелкие зубы выполняют именно такую роль. Первоначальная добыча прото-усатых
китов, скорее всего, была достаточно крупной, а мелкая рыба имела возможность
проскользнуть между зубами хищника. Не стоит улыбаться: современная сельдь
тоже имеет реальный шанс выскользнуть даже из гораздо более совершенного фильтровального
аппарата кита-горбача. Собственно говоря, так ловит добычу, мелких плавающих
ракообразных, современный тюлень-крабоед (Lobodon), у которого зубы
приобрели специфическую форму, став
многовершинными и плоскими. На острове Сеймур близ Антарктического полуострова
в 1989 году был обнаружен интересный кит, названный Llanocetus denticrenatus,
о котором я уже упоминал выше. Канадского исследователя Эдварда
Д. Митчелла удивило необычное строение зубов этого животного: они сидели в
челюстях с большими промежутками,
имели
неглубокие корни и глубокие выемки на коронке, делящие зуб на лопасти. Популярная
статья,
в которой рассказывалось про открытие этого животного,
называлась "Ancient
whale smiled like a sieve" («Улыбка древнего кита похожа на решето»). Это
наглядное подтверждение того, что первые усатые киты использовали зубы, чтобы
закрывать выход из своего рта мелким кормовым объектам, которых схватывали
по несколько штук.
В процессе эволюции у китов развивалось приспособление,
которое одновременно позволяло воде свободно выходить из пасти животного, но
более эффективно задерживало рыбу, ракообразных или кальмаров, оказавшихся
во рту кита. На приведённой выше реконструкции Aetiocetus,
выполненной Карлом Буэллом, видна следующая стадия этого изменения: роговые
образования перекрывают промежутки между зубами, функционально замещая зубы,
которые в какой-то степени ещё сохраняют свою функцию, мешая добыче покинуть
пасть кита.
Ключ к познанию дальнейшего изменения ловчего аппарата китообразных также даёт
знание об «испорченных» генах, отвечающих за развитие зубов, найденных в геноме
современных
китов. При дальнейшем развитии китового уса наличие зубов постепенно стало
нейтральным признаком, и мутанты с «испорченными» генами, отвечающими за развитие
зубов, не отличались по успешности выживания от китов с неповреждёнными генами,
у которых отрастали анатомически полноценные зубы. В дальнейшем исчезновение
зубов сняло анатомические ограничения на развитие более совершенного цедильного
аппарата.
В блоге Брайана Свитека Laelaps 13 февраля 2008 года была опубликована статья «Древние усатые киты могли иметь зубы». В ней приводится, в частности, признаки, которые указывают на наличие китового уса у ископаемых китов. Фотографии также взяты оттуда:
... Вокруг края верхней челюсти ныне живущих китов есть некоторое количество отверстий с бороздами, тянущимися вперёд (к передней части рта) и в буккальном направлении (в сторону «щеки», начинающейся в области задней четверти черепа), и эти особенности обозначают присутствие китового уса. (Отверстия – не гнёзда для китового уса, а полости, которые когда-то вмещали кровеносные сосуды, которые питали китовый ус.) Удивительно, но вымерший зубастый кит Aetiocetus weltoni также обладает этими особенностями (см. иллюстрацию выше), и таким образом, теперь есть конкретное свидетельство [существования] зубастого кита, который также имел китовый ус. Дальнейший анализ Aetiocetus cotylalveus и Chonecetus goedertorum также обнаружил подобные полости, но они сохранились далеко не так хорошо, как изображенный череп A. weltoni. Аналогично, неописанный вид рода Morawanocetus мог бы иметь подобные особенности, но подробности того, было это, либо не было, ещё не опубликованы. |
Теперь можно попытаться ответить на вопрос, сформулированный
Стивеном Джонсом: почему одна родословная линия усатых китов развила китовый
ус, а другая не сделала этого? В принципе, этот вопрос сводится к детскому
вопросу: почему леопард пятнистый, а тигр полосатый?
Итак, ранее строго считалось, что «усатый кит = кит, имеющий во рту китовый
ус». Более поздние находки установили, что это не всегда так, особенно на ранних
стадиях эволюции этой группы. Если взглянуть на многообразие усатых китов на
ранних стадиях эволюции отряда, то видно, что эти животные изначально приспосабливались
к различным источникам питания, вплоть до появления свирепых хищников типа Janjucetus,
которые явно охотились на крупную одиночную добычу, которая не всегда могла быть
рыбой.
То
есть,
их
приспособление к питанию путём фильтрования воды выглядит лишь одной частной
адаптацией внутри
группы,
призванной
снизить конкуренцию с прочими китообразными. Здесь виден обычный процесс адаптивной
радиации, при котором от одного предка происходит широкий спектр потомков, уклоняющихся
в разных направлениях эволюции для снижения конкуренции. «Безусые усатые киты»
вымерли по разным причинам, а одна ветвь былого многообразия китов подотряда
Mysticeti выжила и дошла до нашего времени. Из-за того, что они оказались единственными
современными представителями подотряда, их можно считать просто
«не
репрезентативной
выборкой» многообразия представителей подотряда: отсюда и удивление при открытии иных усатых
китов,
приводящее
к странным умопостроениям.
Примеров такой «не
репрезентативной
выборки»
много: те же современные жирафы. Ранее считалось, что «жираф = парнокопытное
с длинной шеей». Затем всё впечатление от этого вывода испортили ископаемые жирафы,
среди которых явно преобладают виды с короткими шеями, мало отличавшиеся пропорциями
от антилоп, и даже виды с ветвистыми рогами, являющиеся практически двойниками
оленей. И очень некстати в лесах Конго обнаружился окапи с не слишком длинной
шеей. И длинношеие жирафы стали выглядеть своего рода исключением среди множества
древних короткошеих родственников, хотя при слове «жираф» традиционно представляется
именно
парнокопытное
с длинной шеей. Сила стереотипа, если угодно.
Информация по эволюции усатых и зубатых китов после отделения этих групп от предковых археоцетов освещена в популярной литературе (в том числе на доступных рядовому пользователю страницах Интернета) крайне скупо и разрозненно. А на русском языке таких первоисточников практически нет. Очевидно, с этим напрямую связана скудность (стремящаяся к полному отсутствию) статей креационистов, посвящённых «разбору полётов» эволюционистов. Это лишний раз говорит о том, что так называемый «научный креационизм» без эволюционизма просто мёртв, и может существовать лишь как паразитическое явление. Впрочем, здесь уже я начинаю позволять себе некоторое злословие в отношении оппонентов (в свете изученного материала, думаю, вполне заслуженное). А потому вернусь к теме. Возможно, дальнейшее изложение покажется несколько похожим на «лоскутное одеяло»: я постарался выбрать информацию по эволюции усатых китов из различных статей, в том числе посвящённых описанию отдельных видов этих животных.
Mysticeti
Усатые киты
Michel C. Milinkovitch и Olivier Lambert
Родственные отношения среди вымерших и ныне живущих
групп усатых китов были изучены не столь широко, чем родственные отношения
среди зубатых китов. В настоящее время признаны четыре семейства: Balaenidae,
Balaenopteridae, Neobalaenidae (включающее единственный ныне живущий
вид, карликового гладкого кита Caperea marginata), и Eschrichtiidae (включающее
единственный ныне живущий вид, серого кита Eschrichtius robustus).
Некоторые молекулярные исследования определили место гладких китов (настоящих китов) как исходную родословную линию ныне живущих усатых китов (например, Arnason и Gulberg, 1994, 1996; Gatesy 1998). Неуверенность остается в отношении филогенетического положения Neobalaenidae: Caperea мог быть сестринской группой по отношению к полосатикам и Eschrichtius (серый кит) (Arnason и Gulberg 1994), или к гладким китам (Gatesy 1998). Последняя гипотеза также поддерживается морфологическими исследованиями McLeod и др. (1993) и Bisconti (2000). Обсуждение некоторых проблематичных вымерших групп Несколько групп ископаемых усатых китов (например, Aetiocetidae и Cetotheriidae) всё ещё таксономически проблематичны; вероятно, что некоторые из них образуют парафилетические группы. До формулирования разумных гипотез относительно филогении этих групп будут необходимы систематические ревизии, дополнительные анатомические данные и кладистические анализы.
|

Скелет кита Cetotherium из отложений Сарматского моря.
Сами по себе Cetotheriidae выглядят искусственной группой, совмещающей животных с различными признаками, разделяющими мало общих черт. В резюме к статье A NEW CETOTHERE (CETACEA: MYSTICETI) FROM THE EARLY MIOCENE OF JAPAN Toshiyuki Kimura and Tomowo Ozawa, Journal of Vertebrate Paleontology, 2002, 22(3):684–702 об описании нового китообразного Isanacetus laticephalus, про признаки этой группы китообразных сказано следующее: при анализе 77 признаков у 16 видов китообразных не выявлено ни одного общего признака (синапоморфии) для Cetotheriidae.
Анализ поддерживает следующие гипотезы: "Cetotheriidae" в
обычном понимании являются парафилетическим уровнем развития; «цетотерии»
более близкородственны Balaenopteridae Eschrichtiidae, чем Balaenidae;
и «цетотерии» образуют две подгруппы; одна, включающая
Cetotherium, и другая, которая включает Isanacetus,
Parietobalaena и Aglaocetus.
Последняя подгруппа более близкородственна Balaenopteridae
Eschrichtiidae, но также является парафилетической.
|
Интересно, что по данным, полученным на основе изучения биохимических признаков, зубатые киты также елятся на две группы. В резюме к статье Molecular phylogeny of cetaceans prompts revision of morphological transformations (автор Michel C. Milinkovitch) приводятся интересные сведения о родстве кашалота и усатых китов, которое было установлено молекулярными исследованиями:
Способные к эхолокации зубатые киты и питающиеся
путём фильтрования воды усатые киты традиционно рассматриваются как две
монофилетических родословных линии, которые произошли от вымерших китообразных
подотряда Archaeoceti. Тогда, как текущая интерпретация совокупности
морфологических и поведенческих данных поддерживает монофилию зубатых
китов, молекулярные
филогении противоречат этому таксономическому делению, принимавшемуся
в течение долгого времени. Молекулярные данные указывают, что одна группа
зубатых китов, кашалоты, более близкородственна сильно разнообразным
морфологически усатым китам, чем другим зубатым китам. Более того, эти
молекулярные исследования экспериментально предполагают менее давнее
происхождение усатых китов чем было принято считать. Хотя всё ещё необходим
полный кладистический анализ всех существенных морфологических данных,
переоценка некоторых наиболее важных из этих признаков помогает согласовать
морфологический и молекулярный подходы.
|
В статьях креационистов мне приходилось встречать эту информацию,
подаваемую с характерной ехидцей: мол, эволюционисты снова накопали что-то,
что противоречит ранее сделанным выводам. Что ж, вывод о причислении кашалота
к зубатым китам и противопоставление зубатых китов усатым были сделаны без
учёта молекулярных данных, поскольку в 19 веке, когда формировалась систематика
современных китообразных, такой метод установления родства не был известен,
то есть, знания об особенностях этих животных были неполными. Что изменится,
если разделят кашалотов и мелких зубатых китов? Старые отряды рыб (для наглядности
сравните современный справочник по ихтиологии с «Жизнью животных» А. Э. Брема)
покромсали так, что мало не покажется. Но со стороны креационистов протестов
по этому поводу не наблюдается. Думаю, всё дело в слабом знании фактического
материала...
Сближение кашалотов с усатыми китами, с другой стороны, небесспорно. Причиной
этого считается противоречие между данными, которые получены с помощью митохондриальной
ДНК с одной стороны, и ядерной ДНК с другой стороны. Об этом упоминает, в частности,
и сам Милинкович. Он же, говоря об этом противоречии, замечает, что зубатых
китов объединяет ряд признаков строения черепа и среднего уха.
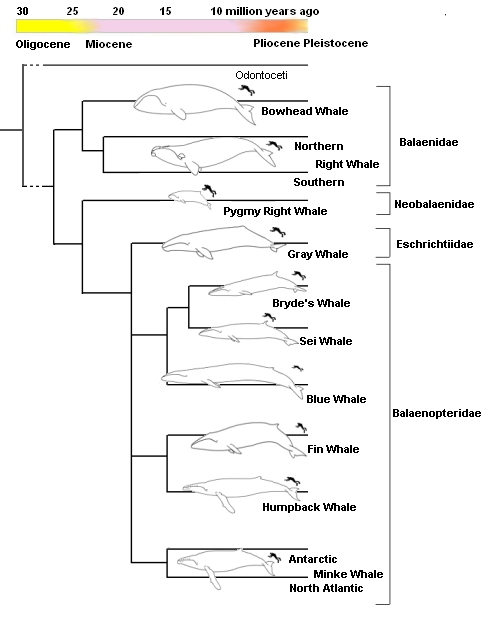
Филогенетическое древо усатых китов, составленное Peter Tomiak и Felix G. Marx, Бристольский университет. Виды современных китообразных, указанные на схеме (сверху вниз):
Гренландский кит (Balaena mysticetus)
Южные киты (Eubalaena glacialis/australis) – бискайский и австралийский. В более старых отечественных источниках они признаются одним видом.
Карликовый кит (Caperea marginata)
Серый кит (Eschrichtius robustus)
Полосатик Брайда (Balaenoptera brydei)
Сейвал (Balaenoptera borealis)
Голубой кит (Balaenoptera musculus)
Финвал (Balaenoptera physalus)
Кит-горбач (Megaptera novaeangliae)
Малый полосатик (Balaenoptera acutorostrata/bonaerensis) – в данном случае выделено два вида: соответственно, антарктический и североатлантический.
Хочется закончить обзор эволюции основных групп отряда китобразных на тёплой мажорной ноте, поэтому я предоставлю слово любезному Харуну Яхье и его работе Whale fantasy from ‘National Geographic’:
Вопреки притязаниям палеонтолога Ханса Тевиссена, который играет главную
роль в эволюционистской пропаганде на предмете происхождения морских
млекопитающих, и является одним из наиболее важных источников информации
для ‘National Geographic’, мы имеем дело не с эволюционным процессом,
подкреплённым эмпирическими свидетельствами, а со свидетельствами, притянутыми
к предполагаемому эволюционному генеалогическому дереву, несмотря на
многие противоречия между ними. Громкая эволюционистская пропаганда о морских млекопитающих, напоминает однако, «лошадиную серию», которая была однажды выдвинута таким же образом, но которую эволюционисты затем признали недействительной. Множество вымерших млекопитающих, которые жили в разное время, было выставлено в ряд одно за другим, и эволюционисты временами пробовали представить их как «твёрдое доказательство». И всё же правда всплывает через некоторое время, наступает понимание, что эти животные не могли быть предками одно другому, что они появились в различные периоды, и что они были фактически отдельными вымершими видами. |
А как же красивая легенда о том, что все виды современных и ископаемых лошадей жили до Потопа в одно и то же время, и чуть ли не бок о бок? Или Всемирного Потопа не было, и время существования мира всё же подразделялось на геологические периоды?!!! Я думаю, младоземельным креационистам стоит «поставить на вид» Харуну Яхье такую вольность и откровенную крамолу. Ведь это нарушает стройность и единство их рядов, а Харун Яхья в данном случае рассуждает, фактически, на пользу эволюционистам.
Найлз Элдредж, хорошо известный палеонтолог Американского Музея Национальной Истории, где были выставлены, и где все ещё в основном сохраняются схемы развития лошади, сказал о них однажды это: «Было ужасное количество историй, одни более воображаемые, чем другие, о том, чем в действительности является природа этой истории [жизни]. Наиболее знаменитый пример, всё ещё выставленный в подвальном этаже, это выставка эволюции лошади, подготовленная, возможно, пятьдесят лет назад. Тогда это было представлено как истинная правда в одном учебнике за другим. Теперь я думаю, это довольно прискорбно, особенно когда люди, которые предложили некоторые из этих историй, могут сами сознавать предположительный характер части того материала». |
Трудно было не заметить в рассуждениях креационистов о китах такого поразительного сходства с «лошадиной серией». И те же самые логические приёмы в креационистской дискуссии легко распознаются. Как ни прискорбно, креационисты почти не эволюционируют в меняющемся мире палеонтологических находок. Что ж, это их проблемы. Другое дело – что они пытаются навязывать свою точку зрения, пользуясь невежеством или излишней доверчивостью слушателей. Но их истории тают, как снег на солнце, в свете фактов.
Волшебная история об эволюции китов, так отчаянно
защищавшаяся ‘National Geographic’, является одной из этих фантазий естествознания.
Подобно её предшественникам, она также скоро окажется в мусорном ведре
науки.
|
Ох, уж эти креационисты... Если надо, наука легко выбрасывает в «мусорное ведро» построения, оказавшиеся неправильными в свете вновь полученной информации. Но вся беда для креационистов заключается в том, что на смену отброшенным построениям эволюционистов появляются новые, более близкие к истине. Так что очередной ритуальный танец победителя над «мусорным ведром» науки – это лишь временный самообман креационистов. Тем хуже для них будет потом.
Выражаю глубокую благодарность за помощь в работе над этим обзором: Brian Switek, Darren Naish, Edward T. Babinski.
Индохиус - «меньший брат» китов
Пакицетиды: первые в ряду
Небольшая интермедия: кит с хоботом
Родословная «ходячих китов»
Амбулоцетус – «кинозвезда» из «Прогулок с чудовищами»
Скованные одной цепью: последовательность форм ранних китообразных
Ещё одна интермедия: есть и подревнее киты
Путь к современным китам
Небольшая интермедия: о развитии китового уса
Ушки... не на макушке.
... На задних лапках ходят.
Держи нос по ветру!
Интермедия о правде и полуправде: О том, как НЕ надо говорить о переходных
формах.
Дань прошлому в организме китообразных.