Глава 3. Рыбы глубоководной пелагиали
Кончилась очередная океанологическая станция, опустела палуба. Судно на малом ходу неторопливо «переползает» с одной волны на другую, но забортные работы еще не завершены. С блока крамбола косо протянулся к корме и режет воду, уходя вглубь, напряженный, как струна, трос: идет траление разноглубинным тралом Айзекса-Кидда... Полчаса – час лова на выбранном горизонте (скажем, на глубине 1000 м), и вот трал поднят на борт грузовой стрелой — в свежую погоду эта операция не так уж проста, и требуется приложить немало мускульных усилий, чтобы усмирить летающий над палубой 100-килограммовый депрессор-заглубитель. Наконец, сетной мешок развязан и улов высыпан в таз. Чего там только нет. Ярко-красные креветки с длинными усами-антеннами, полупрозрачные миниатюрные кальмарчики, сальпы, пиросомы, медузы.... Но сейчас нас интересуют только рыбы. Их в трале больше, чем беспозвоночных, но какие же они странные, непохожие на тех, к которым мы привыкли с детских лет. Светящиеся анчоусы сверкают сине-зелеными или голубыми пуговичками фотофоров. У змеевидно удлиненного бархатно-черного идиаканта, кроме золотых фотофоров, светятся еще и фиолетовые пятна железистой ткани на теле. Еще более длинные нитехвостые угри удивляют своими клювовидными тонкими челюстями. Зубастый хаулиод, раскрыв пасть, демонстрирует устрашающие непропорционально большие клыки. Рыбы-топорики действительно очень напоминают хорошо известный плотницкий инструмент, в миниатюре отлитый из никеля или серебра да еще и украшенный изумрудами фотофоров.
Среди глубоководных пелагических рыб наибольшим разнообразием выделяется семейство светящихся анчоусов, или миктофовых (Myctophidae), детально изученное в последние годы трудами В. Э. Беккера (Институт океанологии АН СССР), Б. Нафпактитиса, Дж. Пэкстона, Б. Халли и других ихтиологов. Оно насчитывает более 210 видов, 92 из них входят в состав двух больших родов — Diaphus и Lampanyctus. Миктофиды - некрупные рыбки длиной от 2,5 до 25 см, большеглазые и большеротые (ротовая щель далеко заходит за вертикаль заднего края глаза), но в целом имеющие вполне стандартный «рыбообразный» облик (рис. 17). Их своеобразие состоит, однако, в необычайно сильном развитии светящихся органов — фотофоров и разнообразных желез на голове и туловище.

Рис. 17. Светящиеся анчоусы, или миктофиды: миктофум (1), протомиктофум (3), диафус (3), лампаниктус (4)
Особенно сложное строение имеют фотофоры
— одинаковые по форме круглые бляшки, число которых у разных видов варьирует
от 50 до 80 (их нет только
у одного,
самого глубоководного вида из рода Taaningichthys). Эти бляшки располагаются,
как правило, в нижней части тела и сгруппированы в серии, генеральная
топография которых характерна для всех представителей семейства (только
у Scopelopsis
multipunctatus мелкие фотофоры усеивают всю поверхность головы и туловища).
В то же время число фотофоров в сериях и их взаимоположение специфичны
для каждого вида. Основой отдельного фотофора является погруженная
в кожу чешуя,
которая видоизменена в чашечку, выстланную пигментным и отражающим
слоями и содержащую внутри нежные светопродуцирующие клетки, окруженные студенистой
тканью. Чешуйка, прикрывающая все эти образования снаружи, модифицирована
в
совершенно прозрачную двояковыпуклую линзу. Каждый фотофор имеет независимое
кровоснабжение и иннервируется особым нервным ответвлением. Его голубоватый
свет испускается в результате химической реакции, которая сводится
к
окислению специфического вещества — люциферина, выделяемого железистыми
клетками,
в присутствии особого фермента люциферазы.
Светящиеся железы миктофовых рыб группируются в несколько типов. Так,
у представителей большинства родов (за исключением Diaphus и Gymnoscopelus)
развиты хвостовые
железы, расположенные на верхнем и нижнем краях стебля. Они могут быть
маленькими или большими, одиночными или составленными серией чешуевидных
пластинок, простыми
по строению или сложно организованными — с пигментной выстилкой, рефлектором
и линзой. В некоторых родах (Lampanyctus и др.) верхне- и нижнехвостовые
железы одинаково развиты у особей обоих полов, в других (Myctophum
и др.)
самцы имеют
только верхние железы, а самки только нижние. Иногда такие железы присутствуют
только у самцов или развиты у них гораздо лучше, чем у самок (род Electrona).
Кроме хвостовых, у всех миктофид есть одна или несколько пар окологлазничных
желез, которые лучше всего развиты у видов рода Diaphus. У некоторых
из них эти железы очень велики и занимают почти всю предглазничную
часть головы.
В
их строении и особенно размерах также нередко наблюдается половой диморфизм.
Набор желез еще одного, последнего типа — чешуйковидных или аморфных
скоплений светящейся ткани у оснований плавников, на верхней поверхности
головы и
в иных местах сильно варьирует у представителей разных родов.
Функция системы светящихся органов, по-видимому, многозначна, но главное
ее назначение состоит, без сомнения, в обеспечении распознания особей
своего вида,
а в некоторых случаях и их пола. Сосредоточение светопродуцирующих
источников преимущественно на нижней части тела позволяет предполагать,
что здесь
имеет место и так называемый противотеневой эффект, способствующий
лучшей маскировке
рыбы на фоне рассеянного верхнего освещения. Есть также предположения,
что вспышки хвостовых желез могут дезориентировать преследующего хищника.
Систематика миктофид, которую нередко вполне серьезно именуют
«миктофидологией», - очень своеобразный раздел ихтиологической науки. Миктофидологи
(в нашей стране эта специфическая профессия полноправно представлена В.
Э. Беккером)
определяют
роды и виды, рассматривают их родственные отношения, строят систему
семейств, основываясь главным образом на расположении фотофоров,
каждую группу
которых принято обозначать определенной буквенной аббревиатурой,
а отдельные органы
в ее составе — порядковыми номерами. Поэтому научные рисунки светящихся
анчоусов часто напоминают технические чертежи, а видовые описания
типа «SAO1 под VO4 или впереди его вертикали; SАО2 примерно
на вертикали AOa2; SAO3 над АOа2» и т. д. понятны
«без перевода» только тем, кто хорошо знаком
с этой группой
рыб.
Такая «геометричность» миктофид, казалось бы, очень облегчает их
опознание: на схематических рисунках различия между видами очень
наглядны и очевидны.
Это, однако, вовсе не так. На самом деле в уловах глубоководных тралов
крайне редко можно встретить идеально сохранившийся экземпляр с полным
набором светящихся
органов, а установление видовой принадлежности по сохранившимся на
своих местах остаткам полуободранных фотофоров — процедура, которая
не терпит
стандартного
подхода. Определение поврежденных миктофид, на мой взгляд, имеет
нечто общее с разгадыванием кроссворда, и поиск верного решения сам
по себе
очень увлекателен.
Я люблю возиться с этими симпатичными рыбками, знаю многих из них
«в лицо» и узнаю некоторых даже при полном отсутствии кожи, но тем
не
менее очень
часто пасую перед непреодолимыми трудностями, доводя идентификацию
только до родового
уровня.
Светящиеся анчоусы очень широко распространены во всех широтах Мирового океана (их нет только в Центральном полярном бассейне), но достигают наибольшего видового разнообразия в тропических и субтропических водах. Все они принадлежат к числу океанических рыб, причем большинство видов постоянно обитают в толще воды и лишь немногие ведут бенто-пелагический образ жизни у материковых склонов и подводных поднятий. Среди пелагических миктофид резко преобладают среднеглубинные виды, которые держатся в светлое время суток на глубине порядка 300 — 1500 м. Для них очень характерны суточные вертикальные миграции с ночным подъемом на несколько сот метров в составе так называемых мигрирующих звуко-рассеивающих слоев* (ЗРС) — комплекса макропланктонных и микронектонных животных, перемещающихся по вертикали в течение суток. В тропиках эти миграции, как правило, полностью укладываются в пределы основного термоклина (в высоких широтах верхняя граница сезонного слоя скачка также лимитирует выход миктофид к поверхности в летне-осеннее время). В то же время представители тепловодных родов Myctophum (кроме М. selenops), Symbolophorus, Gonichthys, Centrobranchus и Loweina (всего около 40 видов), относящихся к никтоэпипелагической группе, проникают ночью к самой поверхности воды. Батипелагические миктофиды (Taaningichthys, некоторые виды Lampanyctus и Lampadena) не меняют существенно горизонта своего обитания в течение суток.
| * Звукорассеивающие слои — регистрируемые гидроакустическими приборами (эхолотами) слои, образованные скоплениями макропланктона. |
Пелагические миктофиды — типичные макропланктонные рыбы, постоянно
дрейфующие вместе с перемещающимися водами. Поэтому почти все видовые
ареалы этих
рыб находят объяснение в приуроченности их основ к тем или иным
крупномасштабным круговоротам (из этого правила существуют, однако,
отдельные не очень
понятные исключения). Пассивным концентрированием особей определяются
по-видимому,
и
скопления миктофид в некоторых районах схождения течений, например
вдоль Южного полярного фронта.
Биология светящихся анчоусов изучена явно недостаточно. Известно,
однако, что они почти безвыборочно питаются разнообразным мезопланктоном
и
принадлежат к числу консументов второго — третьего уровня. Продолжительность
их жизни
невелика:
большинство тропических и мелкие умеренноводные виды живут всего
1 — 2 года, а более крупные умеренноводные — 4 — 5 (по другим данным
— 7
— 8)
лет. В тропической
зоне миктофиды размножаются круглогодично, а за ее пределами преимущественно
в теплое время года, причем их личинки доминируют в зпипелагическом
ихтиопланктоне всех районов открытого океана (это является косвенным
доказательством
их высокой численности). Некоторые тропические формы (в частности,
никтоэпипелагические виды из родов Myctophum и Symbolophorus, а
также Notolychnus valdiviae
и др.)
относятся к моноцикличным рыбам: они созревают уже в возрасте нескольких
месяцев и после довольно продолжительного многопорционного нереста
погибают примерно
в годовалом возрасте. Индивидуальная плодовитость разных видов
сильно варьирует,
достигая нескольких тысяч икринок, и находится в прямой зависимости
от размеров производителей.
Как правило, миктофиды держатся разреженными стайками, хотя в некоторых
участках океана (например, в зоне антарктической конвергенции)
они образуют более или
менее значительные скопления. Такие скопления более характерны,
однако, для отдельных участков над материковым склоном, и образуют
их в основном
специфические
«присклоновые» виды. Для одного такого вида — Ceratoscopelus maderensis
— у восточного побережья Северной Америки отмечены на глубине 600
м плотные линзовидные
стаи диаметром до 100 м и толщиной до 10 м, в которых плотность
рыб достигает 10 — 15 экз./м3 (в одной стае, следовательно,
может насчитываться
300
— 400 тыс. особей средней длиной около 6 см). В целом светящиеся
анчоусы, несомненно,
являются наиболее многочисленной группой мезопелагических рыб,
играющей огромную роль в пищевых цепях открытого океана.
К числу массовых рыб глубоководной пелагиали принадлежат также
некоторые представители близкородственных семейств Photichthyidae
(фотихтиевые)
и Gonostomatidae (гоностомовые),
которые, как и большинство других обитателей мезопелагиали, обладают
хорошо развитыми органами свечения (рис. 18). Среди фотихтиевых
особенно высокой
численности достигают мезопелагические винцигуерии (род Vinciguerria),
широко распространенные
в тропических и субтропических водах. Впрочем, за исключением отдельных
участков восточной части Тихого океана, где один из видов (V. lucetia)
может доминировать
в ихтиоцене, они нигде не конкурируют с миктофидами по биомассе.
Пищу винцигуерии, максимальная длина которых не превышает 5,5 см,
составляет
мелкий планктон.
Откорм происходит главным образом в верхних слоях океана (у V.
nimbaria и V. lucetia иногда даже у поверхности), куда рыбы поднимаются
ночью
в составе ЗРС;
горизонт их дневного обитания находится на глубине 300 — 600 м.
Продолжительность жизни не превышает 2 лет, причем начинающийся
с наступлением половой
зрелости примерно в годовалом возрасте порционный нерест продолжается
до естественной
гибели рыб.
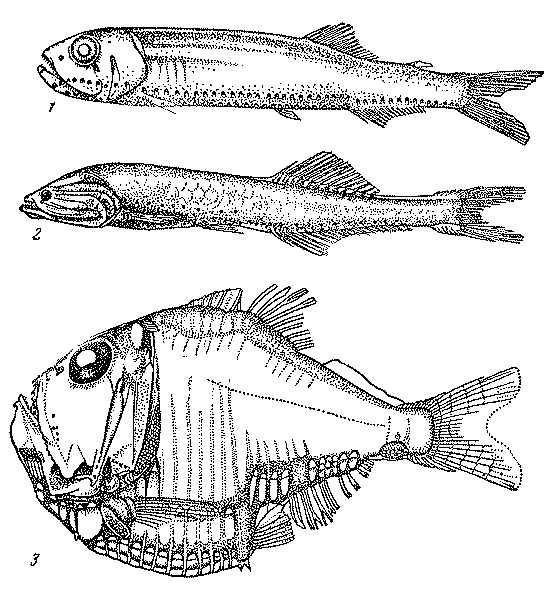
Рис. 18. Винцигерия (1), циклотона (2) и топорик аргиропелекус (3)
Из гоностомовых рыб, в изучение которых большой вклад
внесла В. А. Мухачева, необходимо в первую очередь отметить род циклотон
(Cyclothone),
который
считается, если судить не по биомассе, а по числу особей, самым
высокочисленным среди
всех нынеживущих родов рыб. Сюда относятся 11 видов мелких рыбок
(длина взрослых особей от 3 до 6 — 7 см, масса — от 60 до 600
мг - нижнее значение
близко к
минимальному для рыб), из из которых 2 вида мезопелагические,
6 — мезобатипелагические, 2 — батипелагические и 1 — батиабиссопелагический.
Вертикальные миграции
для циклотон не характерны.
С увеличением глубины обитания окраска тела видов меняется от
прозрачной (например, у С. alba) до черной (С. microdon и др.),
а размеры
фотофоров уменьшаются и
у самого глубинного вида (С. obscura) отсутствуют. «Светлые»
циклотоны, как правило, гораздо меньше «темных» по длине и биологически
отличаются
от них.
У всех видов самцы в 1,2 — 1,6 раза мельче самок и имеют сильно
развитые обонятельные органы, служащие для отыскания особей противоположного
пола по запаху. Циклотоны
питаются в основном мелкими планктонными ракообразными, причем
батипелагические
виды, живущие в бедных пищей водах, питаются от случая к случаю,
даже не каждый день. Их рост и возраст почти не изучены (на отолитах
С.
acclinidens из севере-восточной
Пацифики насчитывали не более 3 годовых колец). Плодовитость
варьирует от 100 до 10 тыс. икринок. Для двух батипелагических
видов характерен
протандрический
гермафродитизм: все особи первоначально финкционируют как самцы,
а затем большая
их часть становится самками.
Такая же реверсия пола отмечена у тех видов близкородственного
рода Gonostorna, которые обитают на большой глубине (G. bathyphilum,
G.
gracile). Гоностомы,
как и циклотоны, принадлежат к числу плаиктофагов, но имеют более
крупные размеры (от 7 до 27 см), и некоторые из них входят в
состав мигрирующих
ЗРС.
Очень своеобразным обликом отличается семейство рыб-топориков
(Sternoptychidae) (в последние годы его изучали Р. Бэрд и О.
Д. Бородулина), в котором,
как и в двух рассматриваемых выше, объединяются как пелагические,
так и бенто-пелагические
виды (их длина варьирует от 4 до 10 см). Наиболее массовые мезопелагические
роды (Argyropelecus и Sternoptyx) широко распространены в теплых
водах. Топорики
не совершают значительных суточных миграций: аргиропелекусы,
характерную особенность которых составляют телескопические глаза,
направленные
вверх, держатся обычно
в верхней мезопелагиали, на глубине 200 — 500 м, стерноптиксы
— в нижней на глубине 500 — 1500 м. Возраст крупных особей A.
pacificus
длиной
6 — 7,5 см
составляет 3 — 4 года. Пищей топориков служат в основном мелкие
рачки,
а сами они являются объектом питания многих хищников, включая
тунцов, заходящих в
пограничные с эпипелагиалью слои основного термоклина для откорма.
Еще одно семейство относительно некрупных (длина до 20 см) мирных
рыб — батилаговые (Bathylagidae) представлено во всех зонах Мирового
океана,
но особенно характерно
для умеренных и высоких широт. Достаточно сказать, например,
что дальневосточная серебрянка (Leuroglossus schmidti) является
самой
массовой среднеглубинной
рыбой Охотского моря. Батилагиды (в семействе, по С. Г. Кобылянскому,
около
20 видов), подобно циклотонам, разделяются на две группы — «светлых»
и «темных», преимущественно приуроченных в своем распространении
соответственно к верхней
и нижней мезопелагиали, причем некоторые светлые виды (тропический
Bathylagoides nigrigenys) совершают вертикальные миграции в составе
ЗРС. Батилаги питаются
зоопланктоном, потребляя в основном не ракообразных, а животных
с мягкими покровами — крылоногих моллюсков, щетинкочелюстных
(хетогнат), аппендикулярий.
По возрасту
и росту данных очень мало. Известно, однако, что особи одного
из темных северотихоокеанских видов при длине 11 см имеют 3,
а при
15
см — 5
годовых колец на отолитах. Антарктический
Bathylagus antarcticus нерестится 3 — 4 раза в году, выметывая
до 5 тыс. икринок.
К семейству меламфаевых (Melamphaidae) относятся рыбы, населяющие
толщу вод открытого океана и наиболее обычные на глубине 500
— 2000 м, т.
е. в мезо-
и батипелагиали. Они имеют однотонную темную окраску, большой
рот с очень мелкими зубами и сильно развитую систему чувствительных
к колебаниям
воды сейсмосенсорных
органов и слизевых каналов на голове. В состав семейства входит,
по А.
Эбелингу, около 35 видов, многие из которых имеют ограниченное
распространение и приурочены
к определенным водным массам. Интересно, что виды, населяющие
наиболее
кормные районы с высокой биологической продуктивностью, достигают
больших размеров,
чем те, ареалы которых приурочены к бедным пищевыми ресурсами
водам. Так, в малокормных центральных круговоротах преобладают
карликовые
виды рода
Melamphaes,
длина которых во взрослом состоянии не превышает 2,5 — 3 см,
тогда как размеры наиболее крупных представителей того же рода
достигают
10 — 12
см. Пищу меламфаев
составляют различные планктонные беспозвоночные, в основном мелкие
ракообразные, а их самих поедают более крупные глубоководные
хищники, в частности рыбы-удильщики.
Несмотря на малые размеры тела, представители семейства имеют,
по-видимому, довольно большую продолжительность жизни: поромитры
(род Poromitra),
например, достигают 10-летнего возраста при максимальной длине
15 см. В отличие от
многих других глубоководных рыб меламфаевые не совершают протяженных
вертикальных миграций, хотя некоторые виды ночью несколько расширяют
зону своего обитания.
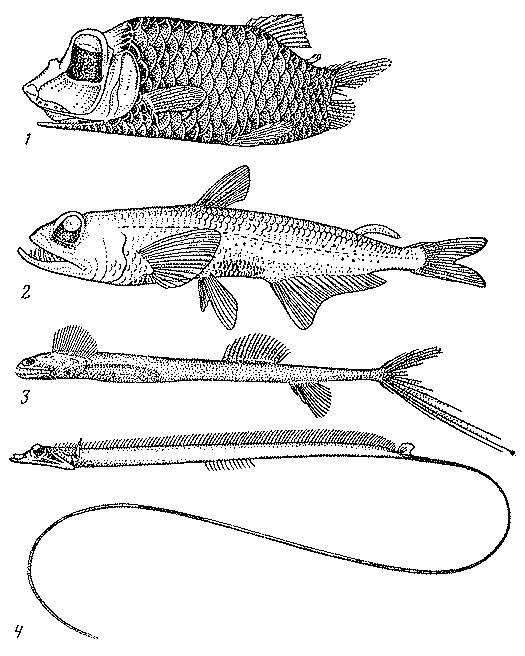
Рис. 19. Глубоководные рыбы с телескопическими глазами: опистопрокт, или рыба-утюг (1), жемчужноглазка (2), розаура (3), палочкохвост (4)
Из других рыб, обитающих в глубоководной
пелагнали, мелким планктоном питаются немногие и к тому же
малочисленные преимущественно среднеглубинные
виды,
но некоторые из них несомненно заслуживают специального упоминания,
хотя бы в
связи со своим необычайным внешним обликом. Обладающий длинным,
почти цилиндрическим в передней половине телом ксенофтальмихт
(Xenophthalmichthys danae) имеет
сильно увеличенные трубчатые глаза, направленные прямо вперед.
Такие
же телескопические
глаза, но ориентированные вверх, развиты
у видов семейства опистопроктовых (Opisthoproctidae), среди
которых крайним своеобразием выделяются рыбы-утюги (два вида
рода Opisthoproctus)
(рис.
19). У этих небольших (до 10 см) рыбок брюшная поверхность
тела образует совершенно
плоскую «подошву», покрытую серебристыми чешуями и отражающую
свет. У близкого рода Bathylychnops с каждой стороны головы
расположено по два
глаза, из
которых нормально развитый основной направлен косо вверх, а
маленький дополнительный, находящийся под передней частью основного,—
вперед
и вниз. Этот дополнительный
зрительный орган, развивающийся как вырост (дивертикул) стенки
главного глазного
яблока, имеет все структурные части глаза, включая хрусталик,
и
снабжен мускулатурой, позволяющей менять направление взгляда.
Батилихнопс — уникальная четырехглазая
рыба, которая пользуется преимуществами почти полного кругового
обзора.
Хорошо развитые трубчатые глаза, смотрящие вперед, имеют и
совсем не родственные всем этим рыбам гигантуровые (семейство
Giganturidae
с
родами Gigantura
и Rosaura), которые обитают на глубине 500 — 1500 м. Они обладают
мощным) зубным вооружением
и, несмотря на относительно мелкие размеры (до 17 см), питаются
главным образом рыбами, заглатывая даже такую добычу, которая
превышает их
по величине (в желудке
экземпляра Gigantura chuni длиной 8 см был однажды обнаружен
хаулиод длиной 14 см). Личинки гигантуровых, подобно личинкам
других глубоководных
рыб,
живут в эпипелагиали. По внешнему виду они не имеют ничего
общего с взрослыми особями
и до последнего времени даже выделялись в особое семейство.
Еще один вид с телескопическими глазами — причудливый палочкохвост
(Stylephorus chordatus) (длина до 28 см), живущий на глубине
300 — 800 м, имеет в
нижней части хвостового плавника пару утолщенных и очень длинных
лучей, образующих
упругий прут, длина которого вдвое превышает длину тела; между
этими лучами проходит канал боковой линии. Нормальное положение
рыбы в
толще воды —
вертикальное, головой вверх (т. е. такое же, как у его родичей
— других вогмеровидных рыб)
— позволяет палочкохвосту пассивно дрейфовать, выжидая появления
добычи — мелких копепод. Когда рачок оказывается поблизости,
рыба резко выдвигает
вперед свой
трубчатый рот (ротовая полость при этом увеличивает объем почти
в 40 раз) и втягивает в него жертву.
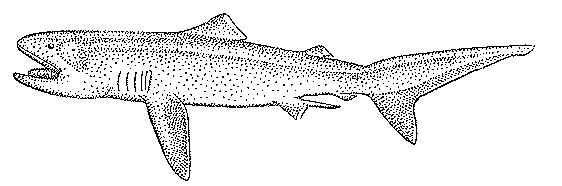
Рис. 20. Большеротая акула, или мегахазма
По-видимому, к числу мезопелагических рыб принадлежит и большеротая акула мегахазма (Megachasma pelagios) (рис. 20), известная пока лишь по двум экземплярам длиной 4,5 м. Первый из них был случайно пойман в 1976 г. у Гавайских островов — он запутался в плавучем якоре-парашюте научно-исследовательского судна на глубине 165 м, второй добыт сетями в 1984 г. в районе Калифорнии. В желудке этой акулы были найдены только эвфаузиевые раки тизаноподы, обитающие на глубине 150 — 1100 м. О глубоководном образе жизни этого вида свидетельствует также слабая мускулатура, мягкие покровы, малообызвествленные позвонки и другие особенности строения. Огромная пасть и хорошо развитые жаберные тычинки позволяют считать, что большеротая акула, подобно китовой и гигантской, является облигатным планктофагом-фильтратором, а серебристая выстилка нёба дала основание для предположения о том, что она может привлекать добычу, подсвечивая изнутри открытую пасть.

Рис. 21. Глубоководные пелагические угри: немихтис (1), циема (2), большерот, или пеликаний угорь (3)
Очень своеобразны
и крайне редко встречающиеся мирапинновидные рыбы (подотряд Mirapinnoidei),
но даже среди них особенно
выделяется сенсационно
знаменитая «волосатая», или «мохнатая», рыба Mirapinna
esau, 30 лет назад часто упоминавшаяся в газетах разных стран. Этот
вид до сих
пор остается
известным по единственному
экземпляру длиной 4 см, пойманному у поверхности воды близ
Азорских островов (по-видимому, случайный вынос с глубины)
. Тело рыбки,
включая голову
и плавники, действительно покрыто мягкими волосовидными
выростами длиной до 1,5 мм (они
несут железистые клетки, функция которых остается невыясненной).
Представители близкого семейства Eutaeniophoridae, известные
с глубины 300 — 1400
м, характеризуются наличием длинного лентообразного придатка
на хвостовом плавнике.
Остальные мезо- и батипелагические рыбы питаются более
крупными — макропланктонными организмами, в большинстве
случаев не
отдавая полного
предпочтения
каким-либо специфическим животным. Из этого правила есть,
однако, несколько исключений,
и все они связаны с выборочным потреблением ракообразных,
главным образом эвфаузиид и креветок.
Среди таких стенофагов нужно отметить прежде всего нитехвостых
угрей (Nemichthyidae), представленных как в верхней мезопелагиали
(род
Nemichthys), так и на больших
глубинах — до 2000 м и глубже (род Avocettina) (рис. 21).
Нитехвостые угри, достигающие 75 — 150 см в длину, характеризуются,
как
недавно выяснили И. Нильсен, Д. Смит и Э. С. Кармовская,
очень интересными
особенностями
жизненного цикла.
В период между ювенильным метаморфозом (эти рыбы, как и
все угреобразные, имеют лептоцефальную личинку) и наступлением
половой зрелости
они обладают длинными
тонкими челюстями, усаженными чрезвычайно мелкими частыми
зубами, но созревшие
самцы теряют и зубы, и челюсти, резко изменяя свой внешний
облик (в связи с этим их даже выделяли в особые роды и
виды).
Есть
предположения, что
аналогичное
превращение может происходить и с самками. Эти дегенеративные
изменения, совпадающие с созреванием, и редкость поимок
готовых к размножению
особей позволяют думать,
что нитехвостые угри гибнут сразу же после нереста. Продолжительность
их жизни достигает 10 лет, а может быть, и большая. Из
подводных аппаратов
немихтиды
наблюдались неподвижно зависшими в толще воды: по-видимому,
таким способом они подкарауливают своих жертв — эвфаузиид
и креветок
сергистид, хватая
их, когда те прикоснутся антеннами к щетке челюстных зубов
и зацепятся за нее.
Для глубоководной пелагиали характерно еще несколько семейств
угреобразных. Пилосошниковые угри (Serrivomeridae), длина
которых достигает 75
см, довольно
обычны в батипелагиали. Там же на глубине 1400—3000 м обитают
и гораздо более редкие большероты, или пеликаньи угри (Eurypharynx
pelecanoides),
длиной до
1 м и мешкороты (9 видов рода Saccopharynx) длиной до 170
см, выделяющиеся
непропорционально большой по сравнению с черепом пастью
и сильно растяжимой глоткой (см. рис. 21). Еще глубже (до
5800
м) живут
циемы (два вида
родов Cyema и Neocyema) — короткотелые плоские угри, экология
которых почти
не изучена.
Как ни удивительно, в составе мезопелагической фауны рыб находит себе законное место и обыкновенный европейский речной угорь, знакомый многим «сухопутным» рыболовам нашей страны, прежде всего, конечно, тем, кто ловит в реках Балтийского бассейна. Как-то и мне довелось присутствовать при поимке этой рыбы в одном из заливов Московского моря (оно же Иваньковское водохранилище). Местный парнишка с гордостью показал мне свою добычу: на дне лодки среди самых банальных окуньков и подлещиков извивался змеевидный полуметровый гость Подмосковья, явившийся сюда из весьма экзотических краев. Я сказал молодому рыбаку, что скользкая грязно-зеленая рыбина принесла ему привет из далекого тропического океана, но не удивил его этим: будучи вполне современным человеком, он уже знал историю угря. Предложение моей сердобольной спутницы дать рыбе , шанс вновь вернуться на свою родину — в теплое Саргассово море также было с ходу отвергнуто на основании полной нереальности такого возврата. Юный рационалист, бесспорно, был прав: вероятность такого события, действительно, бесконечно мала. Даже среди ученых есть скептики, которые сомневаются в возможности европейских угрей пересечь Атлантику. Но ведь угриное племя (а это как-никак особый вид!) все-таки существует, и, следовательно, кто-то все-таки возвращается на нерест, преодолевая все опасности на своем трудном пути и реализуя, казалось бы, совсем нереализуемые шансы...
Действительно, европейский угорь (Anguilla anguilla),
как это было показано И. Шмидтом уже около 60 лет назад,
размножается
в глубинах
центральной
части Саргассова моря, воды которого отличаются наиболее
высокой
соленостью в пределах
Северной Атлантики. Отсюда начинается дрейф икры, а
затем личинок — лептоцефалов, пассивно переносимых на северо-восток
поверхностными
течениями. На это
путешествие, которое заканчивается заходом прозрачных
мальков (их называют
стеклянными
угрями) в европейские реки, уходит 2,5 — 3 года. В
пресной воде самцы угря обычно проводят 6 — 12 лет,
достигая
длины 30 — 40
см, а самки
— не менее
10 — 12 лет, вырастая за это время до 55 — 65 см. В
океане взрослые угри мигрируют
на запад, придерживаясь течений западного направления
на глубине 1000 — 1200 м, достигают в конце концов
места назначения
и,
отнерестившись там,
погибают,
дав начало новому поколению. Такая дальняя миграция
требует, конечно, огромных энергетических затрат. На
этом основании
английский ихтиолог
Д. Таккер
предположил, что все взрослые особи, выросшие в реках
Европы, гибнут в океане, не достигнув
нерестилищ, а воспроизводство осуществляется за счет
американской популяции
(обычно она рассматривается в качестве особого вида
A. rostrata). Различия между европейским и американским
угрями этот автор
относил на счет
прямого влияния температуры в местах вымета икринок
(в
воды Европы попадают лептоцефалы,
выклюнувшиеся в восточной части Саргассова моря) и
отрицал, таким образом, само существование вида A.
anguilla.
| * В ихтиологии под хищниками традиционно понимают рыб, питающихся другими рыбами, и противопоставляют их мирным рыбам, поедающим беспозвоночных животных и растения. В гидробиологии принято другое содержание терминал хищником считается любое животное, потребляющее в пищу животный корм. |
Перейдем теперь к хищным рыбам, пищу которых составляют объекты, сопоставимые с ними по размерам (не менее 15 — 20% длины хищника), — крупные ракообразные, головоногие моллюски, другие рыбы*. В мезопелагиали наибольшего разнообразия среди них достигают представители «усатых стомиеобразных», или стомиевидных (сюда относятся 6 семейств), прежде всего меланостомиевые (Melanostomiidae), которых насчитывается, по современным представлениям, не менее 180 видов (рис. 22). Это — некрупные рыбы длиной от 10 до 40 см. Они имеют в той или иной степени удлиненное, лишенное чешуи тело интенсивного черного цвета, иногда отливающего металлическим блеском. У всех видов есть подбородочный усик, длина которого может у некоторых из них превышать длину тела. Система светящихся органов хорошо развита и включает обычно мелкие фотофоры, беспорядочно рассеянные по телу, крупные туловищные фотофоры, расположенные в продольных рядах вдоль нижнего профиля, большие заглазничные фотофоры, а также светящиеся органы на усике и пятна люминесцентной ткани на голове и туловище. Аквариальные наблюдения над эхиостомой (Echiostoma barbatum) показали, что свет, испускаемый заглазничным органом, может меняться от ярко-розового до зеленовато-белого. Цвет крупных фотофоров у особей этого вида варьирует от красноватого до темно-алого, мелких фотофоров — от розового до пурпурного, а светящаяся ткань на теле имеет зеленоватый цвет. Исключительного разнообразия (как по цвету, так и по числу, форме и расположению) достигают светящиеся железы на подбородочном усике, строение которого у многих видов рода Eustomias (всего их более 100) служит, как установлено Р. Гиббсом, единственным строгим признаком для их идентификации. Меланостомиевые довольно обычны в тропических и субтропических водах, многие живут в умеренных широтах. В дневное время они концентрируются на глубине 500 — 1500 м, а ночью поднимаются к верхней границе основного термоклина (молодые экземпляры Photonectes albipennis даже к поверхности воды). Питаются преимущественно рыбами (светящимися анчоусами, циклотонами), но некоторые виды (например, северотихоокеанская Tactostoma macropus) отдают предпочтение крупным ракообразным. Предельный возраст — 8 лет, а возможно, и больше.
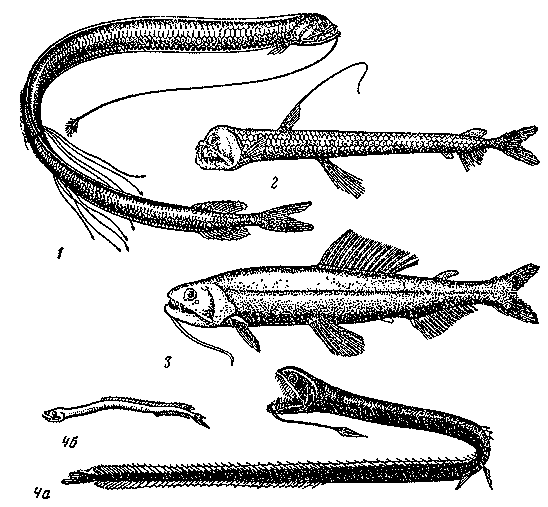
Рис. 22. Стомиеобразные рыбы: стомия (1), хаулиод (3), астронестес (3), идиакант — самка (4а) и самец (46)
В
течение нескольких лет — это было в середине 70-х годов — меланостомиевые
рыбы (на профессиональном жаргоне
их
называют «черненькими»)
находились в самом центре моих научных интересов.
Представлялось поразительным,
что в этой исключительно
многообразной группе виды многих родов (например,
эустомиасы и меланостомиасы) различаются между собой практически
только по строению
чрезвычайно
сложно устроенного подбородочного усика, и хотелось
разобраться в изменчивости этого аппарата.
Надо сказать, что изучение этих усиков, несущих
миниатюрные светящиеся луковицы и разного рода придатки,— исключительно
тонкое дело,
и мне очень
повезло в
том отношении, что я смог кооперироваться в этой
работе с художницей
нашей лаборатории Глафирой Похильской. Ведь в данном
случае никакие словесные описания не могут заменить
в высшей степени
информативного
научного рисунка.
Обработка собранной в Тихом и Индийском океанах
коллекций дала возможность описать несколько новых
форм и высказать
предположения
об эволюционных
тенденциях в пределах группы. Наш материал оказался,
однако, не очень представительным, и это исследование
не получило
дальнейшего развития.
Более обширные коллекции
эустомиасов были изучены американскими коллегами,
которые пришли к заключению об исключительно высокой
видоспецифичности
строения
усика
и, основываясь
главным
образом на этой особенности, чуть ли не удвоили
число видов в роде.
Их результаты значительно продвинули вперед изучение
этих своеобразных рыб,
однако я думаю,
что внутривидовая изменчивость усиков в ряде случаев
осталась все-таки недооцененной, и именно этим
объясняются странные
узколокальные ареалы некоторых видов в Атлантическом
океане. В общем, до полной ясности в познании видового
состава «черненьких» еще далеко.
Близкие к мелоностомиевым семейства астронестовых
(Astronesthidae) с 5 родами (35 видов) и монотипические
хаулиодовые
(Chauliodontidae, 8
видов),
стомиевые
(Stomiatidae, 14 видов), идиакантовые (Idiacanthidae,
3 вида) также имеют приспособления к питанию
относительно крупной
добычей — очень
большой
рот, острые клыковидные
зубы, откидывающуюся вверх при глотании голову,
огромный желудок (см. рис. 22). Все они используют
при охоте
сходную тактику
— выжидание и приманивание
жертв, — и все способны заглатывать добычу, лишь
в 2 —
3 раза уступающую им по размеру. Пассивный образ
жизни дает
полное
основание относить
стомиевидных рыб к макропланктонной группировке.
Пожалуй, наиболее соответствует представлениям
о страшных глубоководных хищниках — недаром они
так
часто изображаются
в популярных
книгах — хаулиоды с их гигантскими
кривыми клыками, которые не умещаются в пасти
и устрашающе торчат наружу даже при закрытом
рте
(эти страшилища,
правда, невелики
по размеру:
самый крупный
вид Chauliodus sloani не превышает 30 см в длину).
Подбородочный усик у всех хаулиодов очень маленький,
но первый луч
спинного плавника вытянут
в длинную
нить со светящимся кончиком, которая, как показывают
подводные наблюдения, свободно свисает перед
ртом и служит для привлечения
доверчивых жертв.
Идиаканты представляют интерес прежде всего с
точки зрения их биологии. Самки, имеющие длинное
угревидное
тело,
достигают длины
40 см и
по образу жизни не
отличаются от других стомиевидных рыб. Этого
никак нельзя сказать о самцах — они гораздо мельче
(длина
всего лишь
6 — 7 см) и
отличаются от самок
отсутствием зубов, усика, брюшных плавников и,
главное, полной дегенерацией пищеварительного
тракта. По окончании личиночной стадии (личинки
идиакантов, кстати сказать, очень своеобразны
— их глаза сидят
на концах длиннейших,
тонких,
как
нитка, стебельков, что облегчает парение в воде)
самцы не питаются и вряд ли живут
дольше нескольких месяцев. В то же время самка
северотихоокеанского вида Idiacanthus antrostomus
длиной 32,5 см имела
6 лет от роду. Плодовитость крупной самки
тропического вида I. fasciola определена в 14
тыс. икринок.
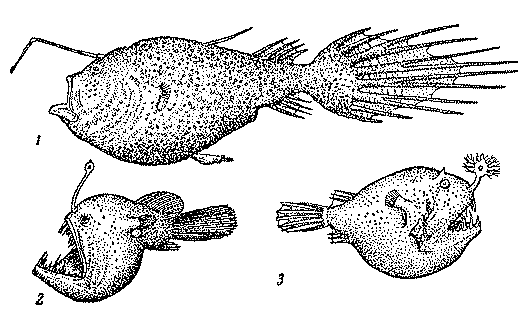
Рис. 28. Глубоководные удильщики: церация — самка с карликовым самцом (1), меланоцет (2), борофрина (3)
Еще одна весьма примечательная группа
макропланктонных рыб — церациевидные или глубоководные удильщики
(подотряд Ceratioidei),
очень большой
вклад в изучение которых внесен Э. Бертельсоном
и Т. Питчем, весьма характерна
для бати- и абиссопелагиали.
Сюда относятся 11 семейств и около 120 видов,
довольно разнообразных по размерам и внешнему
облику (рис.
23), но обладающих одной
общей особенностью — видоизмененным
в «удилище» (иллиций) первым лучом спинного
плавника. На конце иллиция находится «приманка» (эска),
которая у разных
видов
сильно
различается
по строению, величине
и форме. Все удильщики имеют довольно короткое
тело (исключение составляет семейство Gigantactinidae)
с непропорционально
большой головой, огромный, как правило, зубастый
рот и сильно растяжимый желудок. Окраска однотонная
черная,
реже
темно-коричневая.
Органов
свечения на туловище
и голове нет, и у большинства видов единственной
светоизлучающей структурой служит железа, входящая
в состав эски. Исключение
составляют только
линофрины (род Linophryne), у которых в дополнение
к удочке имеются подбородочные
усики со светящимися придатками. В заполненном
слизью просвете эскальной трубчатой
железы живут светящиеся симбиотические бактерии,
которых до сих пор не удалось размножить в
культуре и изучить
детально. Эскальная
железа
и
ее придатки (иногда
сложно устроенные) изолированы от наружной
среды отражающим и пигментным слоями, прободаемыми
«окнами» и «щелями»,
которые обеспечивают
видоспецифичность иллюминации
эски. Регуляция интенсивности свечения обеспечивается
только изменением интенсивности кровоснабжения,
т. е. скорости
подачи в железу кислорода,
необходимого для
жизнедеятельности бактерий. Функциональная
роль
эски как приманки,
привлекающей кормовые объекты удильщиков, совершенно
очевидна, однако серьезные межвидовые
различия в строении органа заставляют предполагать
и его коммуникационное значение.
Взрослые удильщики наиболее обычны на глубине
1500 — 2500 м. Они известны, впрочем, и из
более глубоких
горизонтов,
хотя
их личинки
проходят развитие
в приповерхностных горизонтах (туда всплывает
имеющая положительную
плавучесть икра) и лишь по мере роста опускаются
все глубже и глубже. Возрастные
изменения горизонта обитания удильщиков представляют
собой один из классических примеров
вертикальных онтогенетических миграций. В глубинах
океана образ жизни разнополых удильщиков имеет
очень мало общего.
Половозрелые
самки,
к которым, собственно
говоря, и относятся все приведенные выше сведения
об удильном аппарате, как правило, имеют длину
от 5 —
8 до 20 см, но
у гигантактиса (Gigantactis
gargantua)
она достигает 41 см, криптопсара (Cryptopsaras
couesi) — 44 см, гимантолофа (Himantolophus
groenlandicus) — 60 см,
а у
церации
(Ceratias holboelli)
— даже 145 см. Их пищу составляют различные
глубоководные рыбы (миктофиды, топорики,
меламфаи), иногда настолько крупные, что попытка
проглотить их приводит к гибели самого хищника.
Совсем
недавно,
например, на
поверхности
океана была обнаружена
самка дицерации (Diceratias bispinosus) длиной
11 см, подавившаяся 37-сантиметровым долгохвостом
(по
заглоченному
ею экземпляру
был, кстати говоря, описан
новый вид Ventrifossa johnboborum).
Весной 1975 г. «Витязь» проводил глубоководные биологические исследования в западной тропической Пацифике и внутренних морях Индонезии. В том памятном рейсе, которым завершались работы судна в бассейне Тихого океана, было выполнено рекордное число ловов разноглубинным тралом (своя рука - владыка: я был тогда начальником экспедиции) и поймано множество интереснейших рыб, включая относящихся к новым видам. На фоне ставших обычными редкостей, естественно, осталась почти незамеченной поимка на станции 7316 сравнительно небольшого (60 мм в длину) удильщика, принадлежавшего к весьма банальному виду (Melanocetus johnsoni). На следующий день, разбирая улов в лаборатории, я обратил внимание на туго набитое брюхо этой рыбы. При вскрытии из желудка было извлечено семь свежезаглоченных жертв, в том числе четыре рыбы – фотонект (Photonectes) длиной 165 мм и массой 16,1 г, поромитра длиной 68 мм, пара светящихся анчоусов длиной 57 и 31 мм, один кальмар и две креветки. Все кормовые объекты весили лишь немногим меньше, чем сам хищник после освобождения его желудка (28,9 г против 37,2 г). Правда, остается неясным, удалось ли ему удовлетворить свой аппетит еще на воле, или он поспешил воспользоваться теми возможностями, которые ему дал кратковременный, по тесный контакт с другими пленниками трала (последний вариант я считаю значительно более вероятным).
Самцы всех видов удильщиков отличаются от
самок отсутствием иллициума, намного меньшими
размерами,
более прогонистым
телом, сильным
развитием обонятельных
органов, наличием своеобразных вторичных
зубов, сидящих на внешней стороне челюстей,
направленных
вперед
и действующих как щипчики.
В большинстве
семейств они, несмотря на свою карликовость
(длина 15 — 25
мм), ведут вполне активную
самостоятельную жизнь, питаясь мелкими
планктонными рачками и отыскивая самок по запаху. В то
же время представители
четырех семейств —
Caulophrynidae, Ceratiidae,
Linophrynidae и Neoceratiidae — демонстрируют
совершенно уникальные отношения между полами,
не известные
ни у каких других позвоночных
животных. Встретив
самку своего вида, самцы этих удильщиков
сначала накрепко вцепляются в нее зубами,
а затем полностью
срастаются
с ее телом, теряя
ненужные теперь
органы
(челюсти, глаза, кишечник), подключаясь
к ее кровеносной системе и превращаясь в результате
этих преобразований
в самых настоящих
паразитов,
выполняющих
только одну функцию — продуцирование спермы.
Как правило, различия в размерах самок
и паразитических самцов очень впечатляющи:
так, на крупной церации
длиной 119 см и массой 7 кг был обнаружен
самец
длиной 16 мм и массой всего
14 мг (по-видимому,
он лишь недавно обрел свою «хозяйку», так
как у других особей того же вида самцы-паразиты
вырастают за счет
самки до длины
16 см).
У криптопсара известен,
однако, случай прикрепления 10-миллиметрового
самца
к совсем юной, еще
не половозрелой самочке длиной 15,5 мм,
находившейся, по примерной оценке, в возрасте между
6 и 12 месяцами. Наибольший возраст крупной
самки удильщика гигантактиса определен
в 18
лет.
К макропланктону есть основания относить
и саблезуба (Anoplogaster cornuta), отдаленно
напоминающего
удильщиков коротким телом,
угольно-черной окраской
и огромными клыками, но принадлежащего
к отряду
бериксообразных рыб. Мальки саблезуба по
внешнему виду сильно отличаются
от крупных особей:
они имеют
довольно светлую коричневатую окраску,
почти треугольное в поперечном сечении
туловище
и «рогатую» голову, украшенную длинными
заостренными шипами. Долгое время молодых
и взрослых саблезубов
относили к разным
видам и даже
родам, и
лишь недавно
было установлено, что индивидуальное развитие
этой рыбы сопровождается столь существенными
изменениями
внешнего
облика. Этот вид
широко распространен в
мезопелагиали тропической зоны, но размножается
только в центральных круговоротах. Он достигает
примерно
15 ем в длину
(при возрасте
около 4 лет) и питается
ракообразными, мелкой рыбой и кальмарами.
Молодь, обитающую на глубине порядка 50
— 200 м,
нередко поедают тунцы и другие хищные рыбы.
В мезо- и батипелагиали довольно обычны
также представители семейства живоглотовых
(Chiasmodontidae)
— черные
или коричневые бесчешуйные
рыбы длиной до 30
см со слабой мускулатурой и огромным ртом,
вооруженным многочисленными клыковидными
зубами. Наиболее характерная особенность
вошедшего во все учебники черного живоглота
(Chiasmodon
niger) — способность
заглатывать
крупную добычу,
превышающую по размерам самого хищника
(рис. 24). Челюсти, стенки тела и желудок
у этого
и близких видов могут сильно растягиваться.
Светящиеся живоглоты (виды рода Pseudoscopelus),
возможно,
достойные обособления
в отдельное семейство,
отличаются
наличием мелких фотофоров, расположенных
отдельными группами на нижней поверхности
тела — на горле,
у брюшных и грудных
плавников и у начала
анального плавника.
Живоглоты долгое время считались весьма
редкими рыбами, и лишь сравнительно недавно
было
установлено, что
виды этого
семейства
(в основном псевдоскопелы)
играют важную роль в питании тех тунцов
и марлинов, которые опускаются для откорма
к границам основного
термоклина.
При изучении состава
пищи большеглазого
тунца в западной части Тихого океана, например,
живоглоты были в том или ином количестве
найдены больше, чем
в половине
всех
исследованных
желудков.
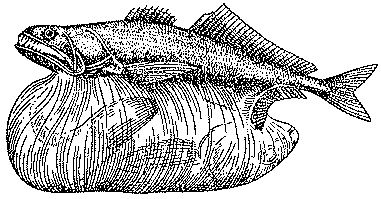
Рис. 24. Хиазмод, или чepный живоглот, заглотивший крупную добычу
Еще одна группа пассивных макропланктонных
рыб, изучаемых сейчас Дж. Пэкстоном,
— отряд китовидообразные,
по
общему абрису тела
действительно несколько напоминающие
миниатюрных настоящих китов. Они особенно
характерны для батипелагиали, но редко
попадают в руки
исследователей; в мезопелагиали живут
только ронделетии (род Rondeletia с 2
видами). Эти темно-красные, коричневые
или черные рыбы,
имеющие голое тело, большую голову с
огромным ртом, маленькие или рудиментарные
глаза и прекрасно развитую сейсмосенсорную
систему, ведут, по всей вероятности,
хищный образ жизни.
Некоторые из
них
(например, виды
рода Cetomimus),
по-видимому, обладают способностью к
люминесценции (светящимися органами считаются
скопления
губчатой ткани у анального
отверстия и вдоль
оснований плавников).
Размеры большинства видов не превышают
10 — 15 см, но самые крупные особи барбуризии
(Barbourisia
rufa)
достигают
30
— 32 см в длину.
Переходное положение между макропланктонной
и микронектонной группировками занимают
эверманнелловые (семейство
Evermannellidae) и жемчужноглазковые
(Scopelarchidae) рыбы, недавно ревизованные
Р.
Джонсоном.
Эверманнеллиды — мезопелагические рыбы,
длина которых варьирует от 7 до 18,5
см, с мощным
зубным вооружением
и довольно
слабой туловищной
мускулатурой.
В состав семейства входят три рода, хорошо
различающиеся строением глаз
— у
одонтостомопса (Odontostomops normalops)
они имеют нормальное строение, у коккорелл
(род
Coccorella) — полутелескопические,
а у эверманнелл
(Evermannella) — настоящие
трубчатые, ориентированные вверх со слабым
наклоном вперед. Обитающие в том же диапазоне
глубин (300
— 1000 м, редко
глубже) и тоже
пользующиеся преимуществом
бинокулярного зрения жемчужноглазки имеют
более упругое тело и, по-видимому, обладают
лучшими
возможностями
для активного
плавания.
Длина самых
мелких видов, ареалы которых приурочены
к тропической зоне, составляет лишь
6
— 10 см (род
Scopelarchus и др.), тогда как крупные
представители рода Benthalbella, живущие
в холодных водах
Северной Пацифики
и Антарктики, достигают
22— 24 см в длину.
Характерной особенностью этих рыб, как
и других, рассматриваемых ниже представителей
подотряда
алепизавровидных — веретенников,
кинжалозубов, алепизавров,—
является способ их размножения. Все они
принадлежат
к числу синхронных гермафродитов,
гонады которых одновременно продуцируют
как яйца, так и сперму. Таким образом,
потенциально
они
способны к самооплодотворению,
но никаких
доказательств в пользу его реального
существования нет, и
при нересте партнеры скорее
всего
ведут себя так же, как и раздельнополые
рыбы: один из них функционирует в качестве
самца,
другой —
самки.
Семейство веретенниковых (Paralepididae)
содержит, по А. Посту, 12 родов и около
50 видов, распространенных
на глубине
200—1500
м (обычно
300
— 600 м)
во всех широтах Мирового океана, но наиболее
разнообразных в теплых водах. Особенно
большой ареал, простирающийся
от 40° с.
ш. до 40°
ю. ш. (отдельные
нахождения и за пределами этой зоны,
в Атлантике, например, вплоть до западной
части Баренцева
моря), у ложного
нотолеписа
(Notolepis
rissoi),
достигающего
30 см в длину. Другие виды семейства
имеют
длину от 15 см до 1 м (Sudis hyalina),
и все они относятся
к
нектонной
жизненной
форме.
Удлиненное
тело веретенников
прекрасно приспособлено для быстрого
плавания и преследования добычи «в угон»
(по этой
причине они почти не попадают
в тралы
исследовательских
судов), однако
из подводных аппаратов наблюдали, как
они медленно передвигаются в вертикальном
положении
головой
вниз или вверх. Представители
тропических
родов Lestidium
и Lestrolepis имеют светящиеся органы
в виде одного
или двух трубчатых каналов, расположенных
в брюшной мускулатуре
и,
по-видимому, содержащих
светящиеся бактерии.
Веретенники питаются мелкими рыбами и
различными беспозвоночными, но
один
вид (Notolepis coatsi), живущий в антарктических
водах, специализирован на питании
крилем. Сами они составляют очень существенную
часть диеты тунцов и других крупных хищников.
Выделяемый в отдельное семейство, но
близкий к веретенникам кинжалозуб (Anotopterus
pharao) длиной
до 90 см (этого
размера он достигает
в возрасте 6 лет) особенно
характерен для умеренных вод северного
и южного полушарий (крупные особи могут
быть
встречены
даже у Гренландии
и в Антарктике),
хотя размножение
идет, судя
по ловам молоди, только в субтропической
зоне. Кинжалозуб хорошо отличается от
всех
родственных
ему рыб отсутствием
спинного
плавника, особенно
резко контрастируя с алепизаврами (семейство
Alepisauridae), обладающими огромным
флаговидным
плавником на спине.
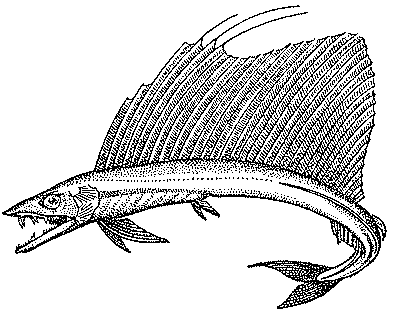
Рис. 25. Большой алепизавр
Большой алепизавр (Alepisaurus ferox)
(рис. 25) имеет очень широкое распространение.
Он встречается
как
в тропических, так и в умеренных
водах в очень растянутом
диапазоне глубин — от подповерхностных
горизонтов до 1000
м, будучи особенно обычным в слое от
150
до 350 м, и часто ловится
на крючки
тунцеловных
ярусов. Максимальная зарегистрированная
длина этой рыбы 210 см (второй вид рода
— короткорылый алепизавр А. brevirostris
не превышает 90 см). Экземпляры длиной
1,5
м и
массой
около 5 кг (по соотношению этих величин
видно, насколько прогонисто их тело)
вполне обычны
в уловах. Алепизавры
— прожорливые хищники,
не очень
разборчивые
в выборе добычи. В списках их жертв значатся
десятки видов рыб, головоногих моллюсков,
ракообразных, а также сальпы,
гребневики, медузы, пелагические
полихеты и другие объекты. Следует отметить,
что
переваривание пищи у алепизавров (как
и у родственных им рыб) происходит только
в кишечнике, а желудок используется лишь
в качестве
своего
рода «провизионной кладовой»
и всегда содержит
абсолютно целые объекты питания (из этого
«орудия лова» происходит
немало голотипов
мезопелагических животных). Общее число
жертв в желудке может быть очень велико:
у одного
алепизавра, например, там были найдены
41 рыба, 3 кальмара,
1
амфипода и
1 полихета, а у другого сразу 83 экземпляра
рыб-топориков. Трофологические исследования
показывают,
что именно алепизавры являются главными
пищевыми конкурентами «ярусных»
тунцов, но они же принадлежат к числу
первостепенных кормовых объектов этих
тунцов.
Среди мезопелагических хищников нельзя
не упомянуть небольших акул из преимущественно
бентопелагического
семейства
катрановых (Squalidae),
представляющих довольно
специфический элемент океанической ихтиофауны.
Их насчитывается около 10 видов. Акула-карлик
(Euprotomicrus bispinatus)
встречается во
всех океанах
и приурочена
преимущественно к центральным водным
массам (в других
районах открытого океана ее замещает
близкий и также карликовый
вид — акула-пигмей
Squaliolus laticaudus).
Акула-карлик принадлежит к числу никтоэпипелагических
рыб, мигрирующих ночью к поверхности
(акула-пигмей не поднимается,
как правило,
выше 200
м), а в дневное
время опускается, как предполагают, на
глубину
порядка 1000 — 1500 м. Самцы этого вида
созревают при длине
17 — 19 см
и достигают
длины
22 см, самки немногим крупнее — самая
большая из них имела размеры 27 см. Размножается
путем яйцеживорождения,
принося
на свет 8
детенышей длиной
около 6 см. Характерной
особенностью акулы-карлика является способность
к произвольному
свечению. При возбуждении вся брюшная
поверхность
ее тела и нижние края боков
светятся
ровным
бледно-зеленым светом, ярко вспыхивая
при резких движениях рыбы и затухая,
когда
она успокаивается.
Органами свечения
служат
мелкие фотофоры диаметром
0,3 — 0,8 мм, густо усеивающие всю нижнюю
часть туловища. Люминесценция может быть
очень интенсивной:
известен
случай, когда такую
акулу заметили ночью на
расстоянии 15 м от борта.
Близко к поверхности океана изредка встречается
ночью и сигарная акула Isistius brasiliensis,
широко распространенная
в тропиках,
но она,
как правило, не поднимается
выше верхней границы основного термоклина.
Впечатляющий размах ее вертикальных миграций
(с 3500 до 85
м) облегчается практически
нейтральной
плавучестью
тела, обеспечиваемой большим количеством
жидкого жира в
полости тела и огромной печени.
Эта акула, достигающая 50 см в длину
(самцы несколько мельче — до 40 см),
— прекрасный
пловец. Наряду
с потреблением в пищу относительно
небольших рыб
и головоногих она, как и второй вид рода
— большезубая акула I. plutodus,
способна к чрезвычайно своеобразному
способу питания, который,
по сути
дела, может быть
определен как факультативный паразитизм
(уникальное явление в классе хрящевых
рыб). Оба вида могут нападать на самых
крупных рыб и китообразных. В этом случае
акулы сначала присасываются толстыми губами
к боку своей жертвы и вонзают в нее свои
непропорционально
большие
нижнечелюстные
зубы (рис.
26), имеющие
форму
треугольных
лезвий и образующие в своей совокупности
подобие острой пилы. Затем они производят
вращательные
движения, позволяющие
выпилить
из кожи
и мышц
жертвы конический
или цилиндрический кусок, который извлекается
из раны шиловидными верхнечелюстными
зубами
и препровождается
в широкую глотку.
Следы таких атак — характерные
кратеровидные шрамы — отмечаются не так
уж редко. Они
были обнаружены на телах марлинов,
тунцов, ваху, корифен, большеротой акулы,
дельфинов и китов. Интересно, что такие
же отметины были
обнаружены при осмотре
внешнего резинового
корпуса американских
атомных подводных лодок. Оба вида обладают
способностью к люминесценции.
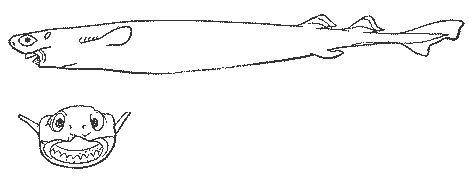
Рис. 26. Большезубая акула — факультативный паразит крупных рыб и китообразных
После лова
в мезопелагиали сети и тралы иной раз
приходят на борт с характерными
дырами,
происхождение которых
долго оставалось
непонятным
— ведь в толще
воды нет, как известно, предметов,
за которые можно
зацепиться. Оказалось, что эти
разрезы — своеобразные автографы сигарных
акул, побывавших в трале и благополучно
ушедших на
свободу. Лишь в
крайне редких случаях,
видимо,
будучи пойманными
в самых верхних горизонтах, они разрешают
извлечь себя из воды, но и
в этом случае чаще всего не ждут, пока
их освободят из сети, а сами прорезают
ее острыми,
как бритвы, зубами, снова возвращаясь
в море или, гораздо реже, падая на
палубу.
Живые акулы (брать их в руки, как показывает
мой опыт, надо с большой осторожностью),
помещенные в аквариум,
интенсивно светятся
в темноте
таинственным зеленоватым
светом, и я видел эту иллюминацию несколько
раз. Впервые такое свечение наблюдалось
в начале прошлого
века натуралистом
старого
закала Ф.
Беннеттом, который оставил
его прекрасное описание, приводимое
здесь с некоторыми купюрами.
«В темном помещении экземпляр представлял
собой необычное зрелище, Вся
нижняя поверхность
тела и головы ярко излучала зеленоватый
фосфоресцирующий свет, что поистине
придавало животному страшный
и пугающий облик.
Люминесценция была постоянной
и заметно не усиливалась при раздражении...
Когда акула погибла (после
трех часов наблюдений) люминесценция
полностью исчезла... Моим первым впечатлением
было,
что рыба
случайно
получила некоторое
количество
светящегося вещества
из моря, но более тщательное изучение
не подтвердило этого подозрения, потому
что постоянство, с которым свечение
занимало определенные части тела и
плавников,
его
неизменность при жизни
и прекращение после смерти
не оставляют
сомнения
в том, что это был жизненно необходимый
процесс...»
Самыми крупными мезопелагическими хищниками
являются, по-видимому, гемпиловые
рыбы — змеиная макрель
максимальной длиной около
1 м (как интерзональный
мигрант, она была рассмотрена в предыдущей
главе), эсколяр (Lepidocybium flavobrunneum)
длиной до 2 м (обычно 1 — 1,2 м),
достигающая иногда еще больших размеров руветта
(Ruvettus pretiosus).
Оба последних
вида более
обычны, впрочем,
над материковым
склоном и подводными поднятиями и,
может быть, должны считаться бентопелагическими.
В своем
вертикальном распространении они
тяготеют к верхней
части
основного термоклина. Истинно мезопелагические
гемпилиды
из родов Diplospinus,
Paradiplospinus и Nealotus не превышают
25 — 45 см в длину.
В целом рыбы, входящие в состав глубоководных
пелагических сообществ, характеризуются
значительно меньшими
размерами, чем рыбы приповерхностных
слоев. Как показывают
ориентировочные расчеты (в качестве
примера проанализированы данные по
максимальной
длине приблизительно 400
видов, известных, согласно
новейшей
сводке, из вод
Южной Африки), длина «среднестатического»
глубоководного вида равна примерно
12 см, что на целый порядок
меньше длины «среднего»
эпипелагического
вида, которая достигает 102 см (рис.
27). При этом наибольшая длина двух
третей
(64%) мезобатипелагических видов
не превышает 20 см, а преобладающей
их
части (85%)
—50 см, тогда как абсолютное большинство
(93%) эпипелагических видов может
достигать длины
более 20 см, а почти
три четверти (72%) — и
более 50 см.
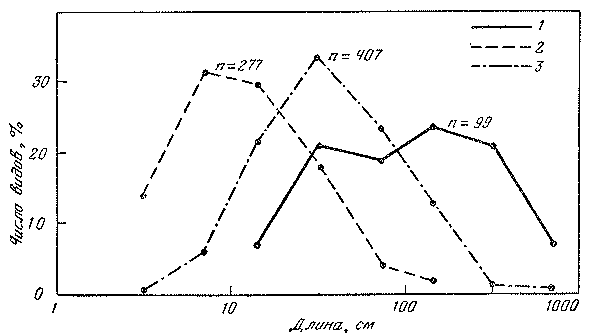
Рис. 27. Распределение видов эпипелагических (1), глубоководных пелагических (3) и глубоководных донно-придонных (3) рыб по максимальной длине особей
Среднегеометрическая длина вида составляет 102 см в эпипелагиали, 12 см в глубинной толще воды и 37 см у дна и над ним
Неудивительно поэтому, что подавляющее
большинство глубинных видов относится
к пассивной жизненной форме (т.
е. к планктону в широком понимании термина). К ней принадлежат
и многие
из крупных рыб (например,
полутораметровый удильщик церация).
Настоящие
нектонные виды характерны только
для самых верхних горизонтов мезопелагиали,
а в холодных
глубинах
с ничтожными кормовыми
ресурсами отсутствуют
даже микронектонные рыбы: по-видимому,
активный поиск и преследование
добычи
энергетически не оправдывают
себя в этих специфических условиях.
Рассматриваемые в этой главе глубоководные
пелагические рыбы входят в состав
сообществ четырех крупномасштабных
биотопов
— мезо-, бати-,
абиссо-
и хадопелагиали,
однако комплексы видов, населяющие
эти биотопы, далеко не равнозначны
по объему
и разнообразию
(табл. 3).
Наиболее богатый видами
(около 950) мезопелагический
ихтиоцен достаточно хорошо обособлен
от батипелагического (в нем около
200
видов), но с дальнейшим увеличением
глубины происходит лишь
общее обеднение фауны. Специфических
абиссопелагических
и тем более
хадопелагических рыб, насколько
можно судить по имеющимся сейчас
данным,
не существует,
и поэтому
ихтиоцены
трех нижних зон водной толщи можно
рассматривать совместно. Общее
число видов в глубоководной пелагиали, по-видимому, близко к 1000 (все цифры
в табл. 3 следует рассматривать впрочем, как приблизительные).
Таблица 3
Систематический состав глубоководных пелагических ихтиоценов
(ЭМ — эпимезо-, М — мезо-, МБ — мезобати-, Б — бати-, БА — батиабиссопелагиаль)
| Отряд, семейство |
Всего видов |
Видов в ихтиоценах |
Встречаемость в других ихтиоценах | ||||
| ЭМ | М | МБ | Б | БА | |||
Катранообразные (Squaliformes) Катрановые акулы (Squalidae) |
68 |
1(9) |
— |
— |
— |
— |
Эпимезобатибентопелагиаль |
Ламнообразные (Lamniformes) Лисьи акулы (Alopiidae) |
3 |
(1) |
— |
— |
— |
— |
Неритическая эпипелагиаль |
| Большеротые акулы (Megachasmidae) | 1 |
(1) |
— |
— |
— |
— |
Нет |
| Ложнопесчаные акулы (Pseudocarchariidae) | 1 |
(1) |
— |
— |
— |
— |
» |
| Сельдевые акулы (Lamnidae) | 5 |
(5) |
— |
— |
— |
— |
Эпипелагиаль |
Кархаринообразные (Carcharhiniformes) Серые акулы (Carcharhinidae) |
48 |
(1) |
— |
— |
— |
— |
» |
Угреобразные (Anguilliformes) Нитехвостые угри (Nemichthyidae) |
11 |
— |
2 |
9 |
— |
— |
Нет |
| Циемовые (Cyemidae) | 2 |
— |
— |
— |
1 |
1 |
» |
| Острохвостые угри (Ophichthidae) | 240 |
(1) |
— |
— |
— |
— |
Эпибенталь |
| Дерихтовые (Derichthyidae) | 3 |
— |
2 |
1 |
— |
— |
Нет |
| Пилосошниковые угри (Serrivomeridae) | 10 |
— |
1 |
9 |
— |
— |
» |
| Мешкоротовые (Saccopharyngidae) | 9 |
— |
— |
— |
9 |
+ |
» |
| Большеротовые (Eurypharyngidae) | 1 |
— |
— |
— |
1 |
— |
» |
Лососеобразные (Salmoniformes) Серебрянковые (Argentinidae) |
35 |
— |
12 |
— |
— |
— |
Мезобентопелагиаль |
| Батилаговые (Bathylagidae) | 19 |
— |
19 |
— |
— |
— |
Нет |
| Опистопроктовые (Opisthoproctidae) | 11 |
— |
11 |
— |
— |
— |
Нет |
Стомиеобразные (Stomiiformes) Гоностомовые (Gonostomatidae) |
27 |
— |
16 |
5 |
1 |
1 |
Мезобентопелагиаль |
| Топориковые (Sternoptychidae) | 40 |
— |
15 |
— |
— |
— |
» |
| Фотихтовые (Photichthyidae) | 21 |
2 |
14 |
— |
— |
— |
» |
| Хаулиодовые (Chauliodontidae) | 8 |
— |
8 |
— |
— |
— |
Нет |
| Стомиевые (Stomiidae) | 10 |
— |
10 |
— |
— |
— |
» |
| Астронестовые (Astronesthidae) | 35 |
2 |
28 |
— |
— |
— |
Мезобентопелагиаль |
| Меланостомиевые (Melanostomiidae) | 180 |
— |
178 |
— |
— |
— |
» |
| Малакостеевые (Malacosteidae) | 15
|
—
|
13
|
2
|
—
|
—
|
Нет |
| Идиакантовые (Idiacanthidae) | 3
|
—
|
3
|
—
|
—
|
—
|
» |
Ayлопообразные (Aulopiformes) Жемчужноглазковые (Scopelarchidae) |
17 |
— |
17 |
— |
—
|
—
|
» |
| Нотосудиевые (Notosudidae) | 19
|
—
|
2
|
—
|
—
|
—
|
Мезобентопелагиаль |
| Гигантуровые (Giganturidae) | 2
|
—
|
1
|
1
|
—
|
—
|
Нет |
| Веретенниковые (Paralepididae) | 50
|
—
|
50
|
—
|
—
|
—
|
» |
| Кинжалозубовые (Anotopteridae) | 1
|
—
|
1
|
—
|
—
|
—
|
» |
| Эверманнелловые (Evermarmellidae) | 7
|
—
|
7
|
—
|
—
|
—
|
» |
| Омосудовые (Omosudidae) | 1
|
—
|
1
|
—
|
—
|
—
|
» |
| Алепизавровые (Alepisauridae) | 2
|
(2)
|
—
|
—
|
—
|
—
|
» |
Миктофообразные (Myctophiformes) Неоскопеловые (Neoscopelidae) |
5 |
— |
1 |
— |
— |
— |
Мезобентопелагиаль |
| Миктофовые (Myctophidae) | 220
|
40
|
150
|
10
|
—
|
—
|
» |
Трескообразные (Gadiformes) Меланоновые (Melanonidae) |
2 |
— |
1 |
— |
— |
— |
» |
| Брегмацеровые (Bregmacerotidae) | 7
|
—
|
2
|
—
|
—
|
—
|
» |
| Долгохвостовые (Macrouridae) | 250
|
—
|
—
|
5
|
—
|
—
|
Мезобатибенталь и бентопелагиаль |
Ошибнеобразные (Ophidiiformes) Ошибневые (Ophidiidae) |
135 |
— |
— |
4 |
— |
— |
» |
Удильщикообразные (Lophiiformes) Каулофриновые (Caulophrynidae) |
3 |
— |
— |
3 |
— |
— |
Нет |
| Церациевые (Ceratiidae) | 3
|
—
|
—
|
3
|
—
|
—
|
» |
| Гигантактовые (Gigantactinidae) | 17
|
—
|
—
|
17
|
—
|
+
|
» |
| Неоцерациевые (Neoceraciidae) | 1
|
—
|
—
|
1
|
—
|
—
|
» |
| Линофриновые (Linophrynidae) | 25
|
—
|
—
|
25
|
—
|
+
|
» |
| Онейродовые (Oneirodidae) | 30
|
—
|
—
|
30
|
—
|
+
|
» |
| Тауматихтовые (Thaumatichthyidae) | 5
|
—
|
—
|
3
|
—
|
—
|
Абиссобенталь |
| Центрофриновые (Centrophrynidae) | 1
|
—
|
—
|
1
|
—
|
—
|
Нет |
| Гимантолофовые (Himantolophidae) | 4
|
—
|
—
|
4
|
—
|
—
|
» |
| Меланоцетовые (Melanocetidae) | 5
|
—
|
—
|
5
|
—
|
+
|
» |
Опахообразные (Lampridiformes) Опаховые (Lampridae) |
2 |
(2) |
— |
— |
— |
— |
» |
| Лофотовые (Lophotidae) | 2 |
(2) |
—
|
—
|
—
|
—
|
» |
| Радицефаловые (Radiicephalidae) | 1
|
(1)
|
—
|
—
|
—
|
—
|
» |
| Вогмеровые (Trachipteridae) | 9
|
(9)
|
—
|
—
|
—
|
—
|
» |
| Ремень-рыба (Regalecidae) | 2
|
(2)
|
—
|
—
|
—
|
—
|
» |
| Палочкохвостовые (Stylephoridae) | 1
|
—
|
1
|
—
|
—
|
—
|
» |
| Мирапинновые (Mirapinnidae) | 1
|
—
|
1
|
—
|
—
|
—
|
» |
| Лентохвостовые (Eutaeniophoridae) | 3
|
—
|
3
|
—
|
—
|
—
|
» |
| Мегаломиктеровые (Megalomycteridae) | 5
|
—
|
5
|
—
|
—
|
—
|
» |
Бериксообразные (Bericiformes) Диретмовые (Diretmidae) |
5 |
— |
1 |
— |
— |
— |
Мезобентопелагиаль |
| Саблезубовые (Anoplogasteridae) | 2
|
—
|
2
|
—
|
—
|
—
|
Нет |
| Меламфаевые (Melamphaidae) | 33
|
—
|
20
|
10
|
1
|
—
|
Мезобентопелагиаль |
Китовидкообразные (Cetomimiformes) Ронделетиевые (Rondeletiidae) |
2 |
— |
2 |
— |
— |
— |
Нет |
| Барбурисиевые (Barbourisiidae) | 1
|
—
|
1
|
—
|
—
|
—
|
» |
| Китовидковые (Cetomimidae) | 23
|
—
|
—
|
23
|
—
|
+
|
» |
Скорпенообразные (Scorpaeniformes) Пинагоровые (Cyclopteridae) |
27 |
(1) |
— |
— |
— |
— |
Эпибенталь |
Липаровые (Liparididae) |
150
|
—
|
1
|
—
|
—
|
—
|
Эпи — абиссобенталь |
Окунеобразные (Perciformes) Эпигоновые (Epigonidae) |
27 |
— |
— |
1 |
— |
— |
Мезобентопелагиаль |
| Морские лещи (Bramidae) | 17
|
(15)
|
—
|
—
|
—
|
—
|
» |
| Гривачевые (Caristiidae) | 4
|
—
|
4
|
—
|
—
|
—
|
Нет |
| Живоглотовые (Chiasmodontidae) | 17
|
—
|
6
|
11
|
—
|
—
|
» |
| Икостеевые (Icosteidae) | 1
|
(1)
|
—
|
—
|
—
|
—
|
» |
| Луваровые (Luvaridae) | 1
|
(1)
|
—
|
—
|
—
|
—
|
» |
| Скомбролабровые (Scombrolabracidae) | 2
|
—
|
1
|
—
|
—
|
—
|
» |
| Гемпиловые (Gempylidae) | 22
|
1(2)
|
3
|
—
|
—
|
—
|
Мезобентопелагиаль |
| Скумбриевые (Scombridae) | 49
|
(7)
|
—
|
—
|
—
|
—
|
Эпипелагиаль |
| Меч-рыбы (Xiphiidae) | 1
|
(1)
|
—
|
—
|
—
|
—
|
» |
| Парусниковые (Istiophoridae) | 8
|
(5)
|
—
|
—
|
—
|
—
|
» |
| Амарсиповые (Amarsipidae) | 1
|
(1)
|
—
|
—
|
—
|
—
|
Нет |
| Центролофовые (Centrolophidae) | 22
|
(9)
|
—
|
—
|
—
|
—
|
Мезобентопелагиаль |
| Номеевые (Nomeidae) | 16
|
(16)
|
—
|
—
|
—
|
—
|
Нет |
| Алетовые (Tetragonuridae) | 3
|
(3)
|
—
|
—
|
—
|
—
|
» |
Скалозубообразные (Tetraodontiformes) Рыбы-луны (Molidae) |
4 |
(4) |
— |
— |
— |
— |
» |
—46
|
(103)
|
611
|
182
|
13
|
2+
|
||
Примечание. В графу ЭМ (эпимезопелагиаль) включены 149 видов, общих для эпи- и мезопелагиали: цифрами без скобок обозначены никтоэпипелагические виды, мигрирующие к поверхности ночью; в скобках показаны виды, встречающиеся в пограничной зоне. В графе БА знаком «+» отмечена вероятная встречаемость в абиссопелагической зоне.
Мезопелагический биотоп занимает, как уже указывалось, воды основного пикноклина,
как правило, совпадающего с основным термоклином. Положение и четкость его
верхней границы определяются в первую очередь температурным фактором. Именно
поэтому в высоких широтах, где в зимне-весеннее время температура поверхностных
и промежуточных вод становится практически одинаковой, нередко наблюдаются
выходы глубоководных рыб даже к самой поверхности. В тропиках граница между
мезо- и эпипелагиалью полностью проницаема только для никтоэпипелагических
мигрантов. Эта граница, или, точнее говоря, пограничная зона, приуроченная
к наиболее градиентным по температуре слоям и имеющая толщину порядка 100
— 150 м, должна рассматриваться как экотон — зона смешения видов из обоих
контактирующих сообществ. Одни из этих видов проводят в нем определенную
часть жизненного цикла, другие встречаются там постоянно или почти постоянно.
Для эпимезопелагического экотона особенно характерны крупные нектопланктонные
и нектонные рыбы, находящие в этой зоне оптимальные условия существования.
Некоторые из них — алепизавры, ряд видов веретенников и гемпилид — более
тяготеют к верхней мезопелагиали, другие — упоминавшиеся в предыдущей
главе сельдяные короли и их родичи, морские лещи, центролофы — к нижней эпипелагиали,
но это разделение представляется весьма условным. Здесь же нагуливается во
взрослом состоянии большинство крупных тунцов, мечерылых и акул, служащих
объектами ярусного рыболовства. Основной источник пищи всех таких хищников
— макропланктонные организмы (рыбы, кальмары, ракообразные), мигрирующие
к верхней границе термоклина ночью, однако те хищники, которые питаются преимущественно
на утренних и вечерних зорях (алепизавры, тунцы и др.), могут использовать
этот источник пищи только опосредованно — через постоянно живущих в мезопелагиали
менее крупных хищных нектеров (это паралепидиды, гемпилиды ц хиазмодонтиды
из рыб и, возможно, некоторые кальмары).
Преобладающие в мезопелагиали макропланктонные и микронектонные рыбы делятся
на две основные экологические группы. Одну из них (большую) образуют виды,
совершающие регулярные суточные вертикальные миграции с ночным подъемом и
дневным опусканием, другую — виды, сохраняющие во взрослом состоянии более
или менее постоянный горизонт обитания.
Следует отметить, однако, что большинству представителей обеих групп (а также
рыбам бати- и абиссопелагиали) свойственны онтогенетические миграции: их
икринки, личинки и мальки проходят развитие в слоях воды, расположенных ближе
к поверхности, чем горизонты обитания подрастающих и взрослых особей. Проявлением
одномоментного состояния онтогенетической миграции многих видов служит прямая
связь размера рыб с глубиной обитания — в пределах вертикального диапазона,
характерного для данного вида, более крупные особи распределены на большей
глубине, чем мелкие. По мнению М. Е. Виноградова, основной смысл онтогенетических
перемещений, которые хорошо выражены и у многих беспозвоночных, состоит в
том, что растущие личинки и молодь могут откармливаться в более кормных верхних
слоях воды.
Суточные миграции рыб (как и других пелагических животных) в самой общей
форме обычно объясняют с трофических позиций, указывая, что вертикальные
мигранты, поднимаясь в ночное время на меньшие глубины (вплоть до поверхности
океана), используют в качестве пищи относительно более богатые ресурсы мезопланктона
этой зоны или поедают животных, перемещающихся вместе с ними. Такое объяснение
ничего не дает, впрочем, для понимания обратного процесса — перемещения мигрантов
на глубину после откорма. Для обоснования этого опускания было предложено
несколько гипотез.
Так, в 50-е годы советский ихтиолог Б. П. Мантейфель, который был в нашей
стране одним из основоположников изучения поведения рыб, высказал предположение,
что сельди (а также другие рыбы из числа зрительных планктофагов) активно
уходят вниз, спасаясь от «пресса хищников» поверхностного слоя, и придавал
этому уходу прямое адаптивное значение. Такое объяснение не может быть принято:
как ни трудно количественно оценить упомянутый «пресс», он выражен в мезопелагиали
никак не меньше, чем в эпипелагиали (это особенно очевидно в тепловодной
зоне, где наблюдаются самые масштабные вертикальные миграции миктофид и других
макропланктонных рыб). Другая теория, также исходящая из адаптивности миграций,
предполагает активное опускание вниз с целью уменьшения энергетических затрат
мигрантов в том случае, если они питаются в более теплой воде, чем та, в
которой происходит переваривание их пищи. Эта гипотеза, приложимая в отличие
от предыдущей только к тропическим водам, не проверялась балансовыми расчетами
и вряд ли выдержит такую проверку в подавляющем большинстве случаев.
По мнению Ю. А. Рудякова, ритмические вертикальные перемещения пелагических
животных могут быть следствием изменения их двигательной активности в течение
суток, причем опускание на глубину происходит в те часы, когда уровень этой
активности оказывается недостаточным для компенсации пассивного погружения
под действием силы тяжести. Для вертикально мигрирующих рыб наиболее правдоподобно
выглядит следующая схема суточных перемещений: активный подъем вечером с
целью встречи с главными кормовыми объектами, ночной откорм в слое обитания
этих объектов (иногда с перерывом в полуночные часы), более или менее пассивное
утреннее опускание на глубину (возможно, соответствующую переходу мигрантов
в состояние нейтральной плавучести) и дневной пассивный отдых («сон») на
этой глубине. В связи с этим нужно отметить, что большая часть мигрирующих
рыб имеет плавательный пузырь — орган, довольно эффективно, хотя и не быстро
регулирующий изменение удельной массы тела. У многих рыб вертикальные миграции
начинают формироваться еще на личиночной стадии развития. Известно, например,
что личинки многих массовых миктофид встречаются у поверхности воды только
днем (это пример так называемой инвертированной, или обращенной, миграции),
что прямо связывается с существованием у них суточного хода двигательной
активности, обусловленной ритмом питания.
Общепризнано, что миграции контролируются суточными изменениями освещенности.
Этот фактор может действовать как ориентирующий, сигнальный, определяющий
ритм питания или синхронизирующий эндогенные (циркадные) ритмы отдельных
особей.
Размах вертикальных перемещений может сильно варьировать. Наибольшей протяженности
— с глубины 400 — 800 (1000) м до поверхности — они достигают у никтоэпипелагических
рыб, уже упоминавшихся выше. К ним относятся около 40 видов светящихся анчоусов
из 5 родов, акула-карлик, змеиная макрель, которые, по-видимому, поднимаются
к поверхности каждую ночь, а также отдельные виды винцигуерий и астронестов
(например, Astronesthes niger), у которых такая миграция не имеет массового
характера, и значительная часть популяции остается ночью в подповерхностных
слоях.
У преобладающего большинства тропических мезопелагических рыб вертикальные
миграции полностью укладываются в пределы своего биотопа — основного термоклина
(т. е, между нижней границей поверхностного изотермического слоя и верхней
границей глубинных однородных вод). При этом положение верхней границы ночного
подъема разных видов довольно сильно варьирует: по-видимому, оно контролируется
в основном температурой воды (может быть, также положением слоя концентрации
пищевых объектов). Известную роль в размахе вертикальной миграции играет
и освещенность (многие миктофиды, например, поднимаются в новолуние на 30
— 50 м выше, чем в полнолуние). Имеются различия и в батиметрическом распределении
разных размерно-возрастных групп одного вида. У некоторых видов (из родов
Diaphus, Ceratoscopelus и др.) в нижние слои эпипелагиали проникают, как
правило, только неполовозрелые особи.
Мигрирующие по вертикали рыбы перемещаются в толще воды в многовидовых ассоциациях
— звукорассеивающих слоях, образующих особые сообщества неполного состава
(без автотрофного звена), которые можно называть схизоценами. Эти слои объединяют
рыб, креветок, эвфаузиид, кальмаров, киленогих и крылоногих моллюсков, сифонофор,
сальп и других животных. Суточные миграции ЗРС (как и другие виды вертикальных
перемещений) связывают мезопелагических консументов с пищевыми ресурсами
вышележащих горизонтов и имеют существенное значение в перераспределении
органического вещества в океане. Эти слои имеют очень широкое распространение
и наблюдаются почти повсеместно. В продуктивных районах часто формируются
сразу несколько звукорассеивающих слоев, перемещающихся по-разному, хотя
иногда и сливающихся вместе ночью или днем. Основную роль в мигрирующих ЗРС
играют макропланктонные рыбы-планктофаги, среди которых резко преобладают
миктофиды из родов Diaphus, Ceratoscopelus, Lampanyctus, Hygophum и др.,
составляющие до 80 — 90% общей численности рыб в схизоцене. В некоторых участках
океана к доминирующим формам относятся также винцигуерии и батилагиды. Хищные
рыбы представлены в сообществах ЗРС хаулиодами, стомиями, гоностомами, астронестами,
идиакантами и другими относительно крупными видами отряда Stomiiformes, питающимися
только в пределах самого слоя. Глубины дневного и ночного положения звукорассеивающих
слоев колеблются в довольно значительных пределах — у светящихся анчоусов,
например, амплитуда суточных перемещений варьирует от 200 м (Diaphus regani)
до 700 — 800 м (Ceratoscopelus warmingi), — но почти всегда остаются в границах
мезопелагиали, т. е. зоны, занятой промежуточными водами. В светлое время
суток мигрирующие рыбы живут на глубине 300 — 1000 м (обычно глубже 400 —
500 м), а ночью концентрируются в слое от 50 — 100 до 300 м.
В мезопелагиали, как уже отмечалось, имеются и слабо мигрирующие рыбы, у
которых размах вертикальных перемещений не превышает 50 — 100 м, т. е. настолько
ничтожен, что им можно пренебречь (у некоторых, например у топориков рода
Argyropelecus, ночью поднимается только верхняя граница). Есть там и рыбы,
вовсе не мигрирующие на протяжении суток. Глубина обитания этих постоянно
живущих на одном горизонте видов (их еще называют «стационарными») также
не одинакова. Грубо говоря, на средних глубинах можно выделить два комплекса
видов, разделение которых особенно четко выражено в тропических водах с развитым
главным термоклином. Там, на глубине 400 — 500 м, температурный градиент
сильно уменьшается и понижение температуры становится гораздо более медленным,
чем в самом слое скачка, и это приводит к обособлению (конечно, не абсолютному)
двух среднеглубинных группировок — верхне- и нижнемезопелагической. К первой
относятся, в частности, рыбы-топорики рода Argyropelecus, эверманнеллиды,
жемчужноглазки, светящиеся живоглоты и др. Многие из них, можно повторить
еще раз, входят в число основных кормовых объектов тунцов рода Thunnus и
других крупных хищников. В нижней мезопелагиали постоянно живут «светлые»
циклотоны (например, Cyclothone alba), представители родов Melamphaes, Valenciennellus,
Sternoptyx и др., и там же сосредоточиваются днем все рыбы, предпринимающие
вертикальные миграции. На глубине порядка 300 — 400 м находится и нижняя
граница ночного подъема наиболее слабо мигрирующих ЗРС. В результате верхние
горизонты мезопелагиали — выше 300 — 500 м — в светлое время имеют очень
скудное макропланктонное население, а макропланктонные рыбы (если не считать
личиночно-мальковых стадий развития) в этом слое днем отсутствуют.
Мезопелагический ихтиоцен, как уже говорилось, исключительно разнообразен
по видовому составу. Он содержит около 960 видов, из которых две трети специфичны
для среднеглубинных горизонтов. Наибольшее число мезопелагических видов содержат
семейства Myctophidae {почти 200) и Melanostomiidae (примерно 180), а также
Paralepididae (около 50), Melamphaidae (около 35), Astronesthidae (около
35), Gonostomatidae (около 30) и др. Всего в этой зоне океанской пелагиали
представлено более 75 семейств, из которых около 20 являются голомезопелагическими,
т. е. не представлены ни в прибрежной зоне, ни в других биотопах океанской
пелагиали.
Видовое разнообразие батиабиссопелагического ихтиоцена значительно меньше,
чем мезопелагического. Здесь отмечено, впрочем, не так уж мало видов — почти
200, которые, как уже указывалось, все относятся к макропланктону. Для этой
зоны особенно характерны рыбы-удильщики подотряда Ceratioidei (около 100
видов, т. е. половина видового состава ихтиоцена). Типичными обитателями
глубинных однородных вод являются также «черные» циклотоны, миктофиды рода
Taaningichthys, китовидки, глубоководные угреобразные.
На всех горизонтах океанской пелагиали видовой состав рыб в тропических и
субтропических водах значительно разнообразнее, чем в субполярных. В антарктической
мезопелагиали, например, обитает чуть больше десятка видов (все эндемичные),
причем более чем одним видом представлено только четыре рода — Bathylagus,
Protomyctophum, Electrona, Gymnoscopelus. Это различие отчасти объясняется
отсутствием термосферы в высоких широтах, однако такое объяснение неприложимо
к бати- и абиссопелагиали в связи с полнейшим однообразием глубинных вод.
Общая причина видовой бедности всех высокоширотных пелагических ихтиоценов
состоит в том, что ранний онтогенез всех глубоководных рыб проходит в верхних
горизонтах пелагиали — там, где условия среды в разных широтах различаются
коренным образом. Поэтому-то адаптации большинства видов к развитию икры
и личинок при высокой температуре препятствуют освоению ими субполярных районов.
Количественное распределение рыб в глубоководной пелагиали зависит от продуктивности
поверхностных вод, так как первичное продуцирование органического вещества
идет только в эпипелагиали и количество пищи, попадающей на глубины, определяется
в конечном итоге
именно этим переносом. Как и в эпипелагиали, наименьшая биомасса рыб наблюдается
в центральных частях субтропических круговоротов, а наибольшая — в экваториальных
и умеренно теплых водах, а также в нерито-океанических участках. Иногда наблюдаются
некоторые отклонения от этого правила, которые всегда объясняются подтоком
глубинных вод из районов, отличающихся иной продуктивностью поверхностного
планктона. Биомасса пелагических рыб в целом уменьшается с глубиной, но в
тропической зоне пик концентрации макропланктона и нектона находится не в
эпипелагиали, а в верхней мезопелагиали.
Количественно оценить обилие рыб в пелагиали совсем не просто, даже если
ограничить эту задачу только некрупными — макропланктонными и микронектонными
объектами. К этой цели ведут разные пути (ловы разноглубинными тралами, гидроакустические
съемки, прямые наблюдения из подводных аппаратов, телеучеты), но в любом
случае очень важно, чтобы способы оценки были единообразными во всех странах
и учреждениях, так как только в этом случае обеспечивается полная сравнимость
данных. Для обсуждения этой проблемы и выработки соответствующих рекомендаций
Научный совет по океаническим исследованиям (сокращенно СКОР) при ЮНЕСКО
образовал в 1977 г. международную рабочую группу, в состав которой входил
и я. Результатом нашей деятельности были предложения по унификации сетных
орудий лова и акустических методов, однако для полного внедрения подготовленных
рекомендаций в широкую практику нужно еще преодолеть немало препятствий.
С участием в этой рабочей группе у меня связано курьезное воспоминание об одной поездке в США. Я прилетел в Сан-Диего, не имея представления, как добираться оттуда до небольшого городка Айдилвилл (именно там мы должны были заседать), но будучи твердо уверенным, что меня, как было обещано, встретят в аэропорту. Однако в небольшой группе ожидавших у входа в здание я не увидел никого из известных мне ихтиологов. Не было там и плакатика с моей фамилией – так коллеги обычно встречают незнакомого человека, и я с некоторым недоумением прошел в почти пустой (дело было поздним вечером) зал. Что же делать? Вернулся назад, потоптался у входа, пока не иссяк поток пассажиров. Опять ушел в зал, отыскал в справочной книге около телефона-автомата номер знакомого ученого, но, прежде чем позвонить, решил еще раз показаться на входе. Теперь там стоял только один представительный мужчина с седой бородкой клинышком, и вид у него был довольно растерянный. Может быть, этот человек встречал меня? Я демонстративно встал перед ним, поймал его отрешенный взгляд... Несколько секунд мы в упор глядели друг на друга, и вдруг его глаза ожили. «Вы не доктор Парин?» – спросил он без большой, впрочем, надежды. «Да». Не могу сказать, кто из нас — Пол Смит (так его звали) или я — был больше рад благополучному исходу дела. Потом он признался, что, действительно, был очень растерян, так как считал, что узнает русского с первого взгляда, а среди пассажиров не оказалось никого, отличающегося от обычных людей. Я не стал уточнять, какими именно Пол представлял себе наши отличительные особенности, но догадываюсь, что специфический образ «русского» сложился в его представлении не без влияния местных средств информации.
Экология рыб глубоководной пелагиали изучена крайне слабо. Сведения об
их питании и трофических связях, о размножении и росте, об образе жизни
и поведении
явно недостаточны, хотя результаты отдельных исследований уже начали
публиковаться в последние годы. Это дает возможность сделать некоторые предварительные
обобщения.
Размножение. Икра и ранние личинки самых разных рыб глубоководной пелагиали
— светящихся анчоусов, циклотон, удильщиков — проходят развитие в поверхностных
горизонтах, но размножающихся особей в эпипелагиали иикогда не наблюдали.
Логично заключить, что нерест происходит на глубине обычного обитания
взрослых рыб, откуда оплодотворенные икринки, обладающие положительной
плавучестью,
быстро всплывают. Вылупившиеся личинки по мере развития и роста постепенно
опускаются все глубже и глубже.
Насколько сейчас известно, почти все глубоководные пелагические рыбы
относятся к порционно нерестующим. Число икринок в порции подвержено
довольно сильной
изменчивости. В районе Гавайских островов оно варьирует, по данным Т.
Кларка, у миктофид от 30 — 150 (крохотный Notolychnus valdiviae длиной
не более
2,5 см) до 2,6 — 9,5 тыс., а у других мезопелагических рыб от 70 — 250
до 27
— 61 тыс. ооцитов (относительно крупная Gonostoma elongatum длиной до
27,5 см), причем общее число порций составляет 10 — 20. Антарктический
батилаг
(Bathylagus antarcticus) выметывает от 250 до 1300 икринок в 3 — 4 порциях,
а североатлантический светящийся анчоус (Benthosema glaciale) — 190 —
470 икринок, возможно, при разовом нересте. В яичниках «светлых» циклотон
насчитывали
0,1 — 0,9 тыс. ооцитов, «черных» циклотон — около 2 — 10 тыс., угря Derichthys
serpentinus — 4,1 тыс., рыб-удильщиков, как правило, 7,5 — 10,8 тыс.,
но у очень крупной (65 см) самки цератии (Ceratias holboelli) — почти
5 млн.
ооцитов. Суммарная абсолютная плодовитость у рыб мезо- и батипелагиали
в общем невелика и заметно уступает плодовитости эпипелагических рыб,
которые, впрочем, имеют гораздо более крупные средние размеры. Относительная
плодовитость
(число икринок на 1 г массы тела) у глубоководных видов также достоверно
меньше, чем у приповерхностных.
Размножение глубоководных пелагических рыб в тропических водах, по всей
вероятности, продолжается в течение всего года, но у краев тепловодной
зоны либо имеет
пик в летне-осеннее время, либо вообще ограничено этим сезоном. В то
же время в субполярных водах существуют как зимненерестующие, так и летненерестующие
виды (в Северной Пацифике, например, к числу первых принадлежит миктофида
Stenobrachius leucopsarus).
Некоторые из упоминавшихся специфических особенностей батипелагических
рыб, связанные с их размножением, в частности, половой диморфизм по размерам
(и
крайнее его проявление — самцовый паразитизм) и по строению органов обоняния,
а также синхронный гермафродитизм, обеспечивающий возможность нереста
любой пары случайно встретившихся производителей, несомненно, объясняются
трудностями
отыскания партнеров в условиях разреженного существования.
Возраст. Данных по возрасту глубоководных рыб пока совсем
немного, и почти все цифры, приводимые здесь, получены Дж. Фитчем и Р.
Лавенбергом
при исследовании
отолитов отдельных экземпляров из вод, прилегающих к берегам Калифорнии.
Продолжительность жизни этих рыб в общем невелика. Скорее всего созревают
(еще до наступления годовалого возраста) и меньше всего живут (1,5— 2
года) мигрирующие в эпипелагиаль тропические миктофиды и винцигуерии:
до наступления
половой зрелости они быстро растут, а затем почти беспрерывно нерестятся
вплоть до самой смерти. Так, Benthosema pterotum, наиболее массовый вид
миктофид Аравийского моря, становится половозрелой к 6 — 7 месяцам жизни
при длине
порядка 40 мм (максимальная длина этого вида 50 мм) и гибнет после окончания
нерестового периода в возрасте чуть больше 1 года. Созревание слабо мигрирующих
и постоянно остающихся на глубинах, а также обитающих в высоких
и умеренных широтах рыб наступает позже (у батипелагического угря Serrivomer
sector, например, в возрасте около 4, у северного светящегося анчоуса
Benthosema glaciale — в 2 — 3 года). Продолжительность их жизни составляет
от 3 —
5 (циклотоны, рыбы-топорики, батилаги, умеренно-холодноводные и глубинные
миктофиды,
веретенники рода Lestidium и др.) до 6 — 8 лет (хаулиоды, идиаканты,
кинжалозуб, китовидки, живоглоты) и даже 10 — 18 лет (представители родов
Poromitra,
Nemichthys, Gigantactis). Это свидетельствует о более высоком уровне
энергетических процессов у мигрирующих по вертикали видов по сравнению
с ограниченно мигрирующими
и немигрирующими.
Питание. В каждом биотопе глубоководной пелагиали есть и планктоядные
и хищные рыбы. Среди мезопелагических планктофагов доминируют, как уже
отмечалось,
мигрирующие по вертикали миктофиды и винцигуерии. Исследования, выполненные
в последние годы Т. А. Гореловой, Т. Кларком и другими специалистами,
показывают, что те из них, которые достигают ночью поверхности, питаются
в основном
приповерхностными
беспозвоночными (копеподы понтеллиды и др.), а перемещающиеся в составе
ЗРС без выхода в верхний однородный слой — мигрирующим вместе с ними
интерзональным мезопланктоном (главным образом также ракообразными —
копеподами, остракодами,
амфиподами, эвфаузиидами) и являются основными потребителями этого планктона.
Рыбы-мигранты питаются по преимуществу ночью или в вечерние и утренние
часы
(т. е. на закате и восходе солнца) и, подобно всем прочим зоофагам океанской
толщи, захватывают обнаруженную ими (как правило, визуально) добычу,
какой бы размер она ни имела, поодиночке (единственным фильтратором среди
глубоководных
рыб является, насколько сейчас известно, большеротая акула мегахазма).
Об этом свидетельствует, в частности, явное преобладание в их желудках
непрозрачных,
более или менее контрастных по окраске объектов. Несмотря на очень широкие
пищевые спектры и их большое сходство у разных видов тропических миктофид,
между ними, по меньшей мере в некоторых районах, существуют определенные
различия в составе пищи, которые, по всей вероятности, связаны с различиями
в длине рыб (корреляция между размерами потребителя и жертвы часто выражена
довольно явно) и в видовом составе планктона на горизонтах их ночного
подъема. Это, по-видимому, способствует снижению межвидовой конкуренции
за пищу
в условиях ограниченных трофических ресурсов, поскольку у миктофид, живущих
в более продуктивных по планктону умеренных и холодных водах, различий
в
составе пищи фактически не обнаружено. В олиготрофных участках океана
миктофиды демонстрируют исключительно высокую кормовую пластичность:
Ceratoscopelus
warmingi, например, может использовать в пищу не только излюбленных копепод,
но также хетогнат, оболочников, сифонофор, личинок рыб и даже фитопланктон.
Немигрирующие планктофаги — циклотоны, гоностомы, рыбы-топорики, меламфаи,
батилаги (кроме совершающего суточные миграции Bathylagoides nigrigenys)
— питаются независимо от времени суток или, что представляется характерным
только для мезопелагических форм — жителей сумеречной зоны (Sternoptychidae
и др.), в дневные часы. Основной пищей топориков, гоностомовых и меламфаев
служат ракообразные (у более мелких — копеподы, у крупных — можно напомнить,
что длина Gonostoma elongatum превышает 25 см, — эвфаузииды). Батилаги
предпочитают планктеров с мягкими покровами — крылоногих моллюсков, личинок
головоногих,
аппендикулярий и самых мелких рачков. Средние индексы наполнения желудков
у пелагических рыб уменьшаются с глубипой в соответствии с разреженным
распределением глубинного плактона, и батипелагические циклотоны (в частности,
Cyclothone
pallida) влачат, казалось бы, самое жалкое существование (для них, впрочем,
совершенно нормальное), поглощая в среднем только одного маленького рачка
раз в 10 дней. Рассчитано, что суточные рационы иммигрирующих рыб составляют
примерно 1% сырой массы их тела и заметно уступают рационам мигрирующих
видов, у которых они достигают 3 — 5, а у быстро растущей молоди даже
10 — 15% сырой
массы тела.
Среди рыб, питающихся макропланктонными объектами, особое место занимают
специализированные потребители относительно крупных ракообразных — больших
глубоководных эвфаузиид и креветок. Это все виды бекасорылых, или нитехвостых,
угрей (семейство Nemichthyidae), желудки которых никогда не содержат
другой пищи, а также, возможно, еще два семейства угрей — пилосошниковые
(в питании
Stemonidium hypomelas отмечены только ракообразные, но виды рода Serrivomer
наряду с ними поедают рыб и кальмаров) и нессарамфовые (для представителей
обоих родов — Nessarhamphus и Derichthys — ракообразные указываются в
качестве основной пищи). Предположительно такая специализация связана
с какой-то
особенностью поведения: можно думать, что все эти угри «знают секрет»
приманивания крупных
ракообразных, так как активное выборочное преследование именно этих жертв
хищникам явно не по силам. Интересно, что узкоспециализированные ракоеды
(карцинофаги) есть и среди стомиеобразных рыб (Tactostoma macropus, виды
рода Photostomias), которые не имеют ничего общего с глубоководными угрями.
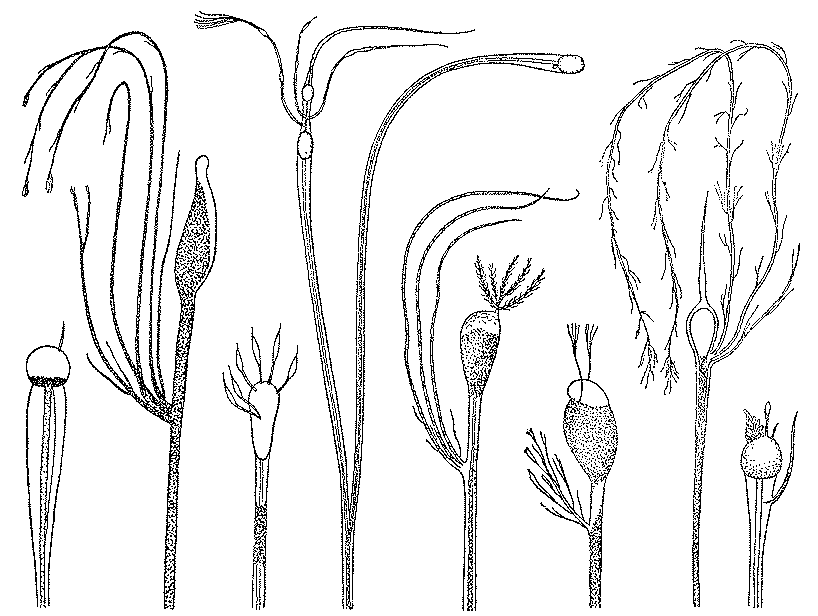
Рис. 28. Концевые части подбородочных усиков у разных видов эвстомий (род Eustomias)
Обычно считается, что остальные хищные рыбы не имеют особых пищевых предпочтений,
но это, по-видимому, не совсем правильно. Так, стомиеобразные хищники
— хаулиоды, астронестовые, меланостомиевые и другие, входящие в сообщества
ЗРС, питаются
только рыбами и
беспозвоночными, также мигрирующими в составе этих слоев. Мало того,
недавно выяснилось (эти данные получены Т. Кларком), что хищники, обладающие
длинными
подбородочными усиками со светящимися органами на конце (представители
родов Stomias, Eustomias, Melanostomias и др.) или удлиненным лучом спинного
плавника,
также оснащенным люминесцентной железкой у конца (виды рода Ghauliodus),
питаются главным образом рыбами — миктофидами и др., которые привлекаются
светом этих желез. При этом в тех случаях, когда представители разных
родов или разных видов одного рода (например, рода Eustomias) сильно
различаются
по строению светящихся приманок (рис. 28), между ними есть и более или
менее существенные различия в составе кормовых объектов — у некоторых
в желудках
преобладают такие жертвы, доля которых в ихтиоцене, судя по траловым
ловам, совсем незначительна. Преимущественными ихтиофагами являются и
батиабиссопелагические
удильщики, которые также подманивают добычу светящимися эскальными приманками
и в некоторых случаях светящимися придатками усиков.
Виды стомиевидных, у которых усик отсутствует или развит слабо (как упомянутые
выше тактостома и фотостомия), питаются только ракообразными или преимущественно
ими и, по всей вероятности, активно ловят свою добычу. В некоторых случаях
хищник (например, Thysanactis dentex) может использовать оба способа
питания — подманивать рыб с помощью светящейся «удочки» и визуально выискивать
зоопланктон (включая и довольно крупных раков). Самый глубоководный представитель
этой
группы — Malacosteus niger, немигрирующий обитатель батипелагиали, имеет
широкий спектр питания — от мелких копепод до рыб.
Немигрирующие мезопелагические хищники сильно различаются по размерам
— мелкие эверманнеллиды, жемчужноглазки и светящиеся живоглоты (Pseudoscopelus)
имеют
всего 6 — 7 см в длину, тогда как крупные веретенники (Sudis hyalina),
кинжалозуб, алепизавры могут достигать 1 — 2 м. Известно, что их пища
достаточно
разнообразна
и состоит в основном из рыб, головоногих и крупных ракообразных, однако
специальные исследования касались только питания двух видов алепизавров.
Исключительно
широкий кормовой спектр этих рыб имеет, пожалуй, лишь одно ограничение
— в нем практически отсутствуют все организмы, ассоциированные с ЗРС,
в частности
основной их компонент — миктофиды. Мигрирующие формы (диафусы, винцигуерии,
креветки сергестиды, кальмары кранхииды) входят вместе со стационарными
обитателями нижней мезопелагиали (светлые циклотоны и др.) в рацион эверманнелловых,
жемчужноглазковых и веретенниковых рыб. Это различие можно объяснить
тем, что алепизавры (как и тунцы) добывают пищу на утренней и вечерней
заре,
когда
ЗРС находятся выше горизонта их охоты, а другие хищники — преимущественно
днем, т. е. в период нахождения ЗРС на глубине. Получается, таким образом,
что очень мощная трофическая система мигрирующих слоев, которая сохраняет
автономию по меньшей мере на трех уровнях консументов и включает преобладающую
часть биомассы мелких среднеглубинных рыб (миктофиды и пр.), кормит на
выходе только стационарных нижнемезопелагических хищников. Их численность
не поддается
пока прямой оценке (особенно численность нектонных видов — веретенников,
гемпилид и др.), но не может быть малой, что явствует также из данных
о значении этих объектов в питании тунцов и других хищников, кормящих
в переходной
между
эпи- и мезопелагиалыо зоне.
Распространение. Пространственное распределение макропланктонных видов
океанических рыб более или менее соответствует распределению естественных
участков пелагической
среды — замкнутых или частично замкнутых круговоротов течений, в которых
циркулируют водные массы. Крупномасштабные круговороты (тропические,
субтропические и субполярные), простирающиеся от берега до берега каждого
океана, и зоны
их соприкосновения населены «океаническими» (в понимании К. В. Беклемишева)
видами. В участках схождения и расхождения течений у материков живут
дальнеиеритические виды.
К этим большим круговоротам и к среднемасштабным круговоротам, существующим
в их пределах в качестве особых структурных элементов, приурочены основы
видовых ареалов макропланктонных рыб, т. е. такие участки, в которых
вид может существовать неопределенно долго вне зависимости от его наличия
в
других частях океана. Ареал, кроме его основы, включает также нестерильную
и стерильную
области выселения в выносящих течениях, и они могут быть очень протяженными.
Правда, определить границу между ними в большинстве случаев непросто:
это требует специальных исследований физиологического состояния рыб,
вроде
осуществленных У. О'Деем и Б. Нафпактитисом на атлантических миктофидах
рода Lobianchia.
Таким образом, основой ареалов планктонных видов рыб могут служить как
крупномасштабные круговороты океанских течений, так и круговороты, расположенные
в нейтральных
областях деформационного поля. Основа ареала многих широко распространенных
рыб (в частности, видов с циркумтропическим распространением) может охватывать
несколько круговоротов. Существенно отметить, что во вторичных водных
массах (например, между субтропическими и субполярными круговоротами)
смешанное
население состоит из зависимых популяций видов, имеющих основы ареалов
в соседствующих водных массах или в дальненеритических круговоротах.
Изложенные взгляды вполне соответствуют широко распространенным представлениям
(так называемая «гипотеза водных масс») о том, что стациями популяций
планктонных видов являются водные массы, понимаемые как совокупности
однородных по
своим характеристикам (температура, соленость и др.) объемов воды. В
то же время
надо отметить, что выделение этих однородных объемов по одним только
температурно-соленостным показателям недостаточно дробно для выявления
всех пелагических биотопов.
Некоторые круговороты (особенно среднемасштабные), практически не различающиеся
по температурно-соленостным характеристикам, служат основами ареалов
разных видов, что, по-видимому, определяется их обособленностью по другим
факторам
среды, важным для жизни этих видов.
Рассматривая географическое распространение конкретных видов, как правило,
почти всегда удается с той или иной степенью достоверности, зависящей
прежде всего от изученности объекта, определить, хотя бы в предположительной
форме,
положение основы (или нескольких основ) их ареалов. Почти все затруднительные
случаи могут быть объяснены неполнотой данных или недостаточностью сведений
о жизненных циклах вида, но имеются и такие факты, которые не поддаются
объяснению в рамках изложенной концепции. В качестве примера можно упомянуть,
например,
ареалы двух централь-нотихоокеанских миктофид — Protomyctophum beckeri
(к югу от Гавайских островов — между 8 — 20° с. ш. и 144 — 166° з. д.)
и Lampanyctus
hubbsi (вдоль экватора — между 10° с. ш. — 5° ю. ш. и 120 — 160° з. д.),
в пределах которых нет, судя по имеющимся данным, никаких замкнутых горизонтальных
циркуляции (возможно, правда, что основы их ареалов приурочены к вертикальным
круговоротам).
 |
 |
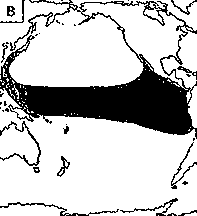 |
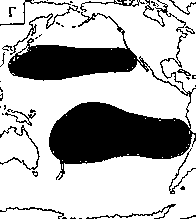 |
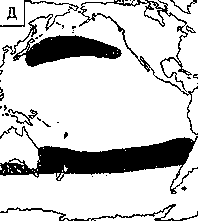 |
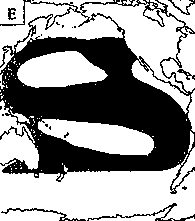 |
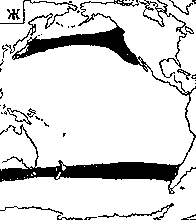 |
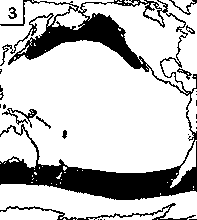 |
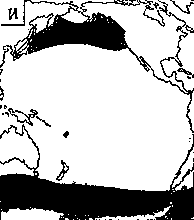 |
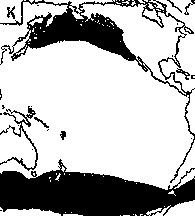 |
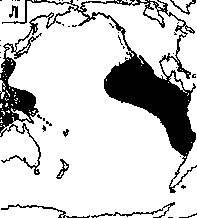 |
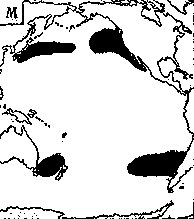 |
Рис. 29. Типы океанических (А – К) и дальненеритических (Л, М) ареалов макропланктонных
мезопелагических рыб (на примере тихоокеанских миктофид)
Обозначения в тексте
Основные типы ареалов (их называют также «географическими элементами
фауны») макропланктонных рыб группируются следующим образом (рис. 29,
А — М):
I. «Океанические ареалы (ареалы океанических круговоротов).
A. Широкотропический тип. Широкотропические виды живут, грубо
говоря, во всей зоне между 40° с. ш. и 40° ю. ш. Основы их ареалов
находятся
во всех
тропических и субтропических круговоротах, зависимые популяции есть
во вторичных водных массах на периферии тропиков, а некоторые виды
распространены
и в
тропических дальненеритических участках. В то же время следует отметить,
что многие широкотропические рыбы имеют более или менее обширные разрывы
в географическом распространении, отсутствуя, например, либо в ядрах
центральных круговоротов, либо в переходных, либо в дальненеритических
районах.
Б. Экваториально-центральный тип. У этих видов основы ареалов
расположены в экваториальных и центральных водах (часто с пробелами
в тех или иных
их участках), но области их распространения не столь обширны, как у
широкотропических рыб, так как они отсутствуют во вторичных водных
массах у границ тропической
зоны, а обычно и в дальненеритических районах.
B. Экваториальный тип. Основы ареалов лежат
в тропических круговоротах у экватора, но отдельные виды имеют дополнительные
основы в некоторых дальненеритических участках, так что области их
распространения расширяются у берегов (особенно в западных частях океанов).
Очень характерный
подтип этого географического элемента составляют ареалы видов, отсутствующих
в восточноэкваториальных дальненеритических районах.
Г. Центральный тип. Все ареалы этого типа имеют основы в олиготрофных
субтропических круговоротах. Некоторые из центральных видов живут во
всех пяти этих круговоротах
и имеют бицентральное (антиэкваториальное) распространение, другие
встречаются только в одной-двух центральных водных массах.
Д, Е. Периферический тип. Эти виды живут на периферии центральных водных
масс, главным образом на севере северных и (или) на юге южных субтропических
круговоротов (Д), но в относительно немногих случаях также у восточных
(реже у западных) берегов материков и на экваторе (Е).
Ж. Переходный тип. Эти ареалы приурочены .либо к северной части Субантарктики
(«зона субтропической конвергенции»), либо Северотихоокеанскому и (или)
Североатлантическому течениям, т. е. ко вторичным водным массам, образованным
в результате смешения
субполярных и центральных вод. Основы ареалов, по-видимому, находятся
в круговоротах, прилегающих к материкам. Особенно многочисленны такие
рыбы
в южном полушарии.
3. Нотально-южнобореальный тип. Ареалы этого типа располагаются в южных
частях Субантарктики (между субтропической конвергенцией и южным полярным
фронтом)
или Субарктической Пацифики, где примесь центральных вод незначительна.
В Атлантическом океане этот географический элемент ихтиофауны отсутствует.
И. Антарктическо-севернобореальный тип. Эти ареалы имеют своей основой
в северном полушарии западные круговороты субарктических вод Тихого
и Атлантического океанов, а в южном — циркуляции, образованные течениями
Восточных и Западных
ветров.
К. Широкий антарктическо-бореальный тип. Виды, принадлежащие к этому
типу, имеют основы ареалов в субполярных круговоротах либо северного,
либо южного
полушария.
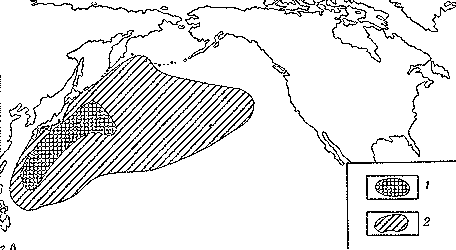
Рис. 30. Ареал северотихоокеанской гоностомы
1 — область размножения (основа ареала); 2 — область распространения
взрослых особей
II. Дальненеритические ареалы (ареалы нейтральных областей).
Л. Тропический тип. Основы ареалов тропических дальненеритических видов
расположены в нейтральных областях между тропическими и субтропическими
круговоротами,
но видов, общих для всех этих областей, нет. Особенно много эндемичных
рыб обитает в восточной тропической Пацифике.
М. Переходный тип. Эти дальненеритические ареалы находятся в нейтральных
областях между субтропическими и субполярными круговоротами.
Н. Широкий дальненеритический тип. Виды с ареалами этого типа живут
в тропических и в переходных нейтральных областях.
Ареалы батипелагических рыб изучены пока менее детально, чем ареалы
мезопелагических видов. Некоторые из них имеют очень широкое распространение
в Мировом
океане. Крайний пример представляет в этом отношении одна из циклотон
(Cyclothone
pseudopallida), обладающая почти космополитическим ареалом (этого
вида нет только в Северном Ледовитом океане, восточной тропической
Пацифике
и Аравийском
море), но, к сожалению, положение основы ареала этого вида еще не
установлено. Между тем, как показывает пример Gonostoma gracile (рис.
30), область
размножения батипелагического вида может занимать очень малую часть
акватории, на которой
этот вид распространен.
Известно, что многие батипелагические рыбы имеют широкотропическое
распространение. Из более ограниченных ареалов следует отметить такие
географические элементы,
как экваториальный, восточнотихоокеанский и восточноатлантический
тропические, антарктическо-нотальный и бореальный. Наблюдающееся
сходство в типологии
ареалов мезо- и батипелагических рыб объясняется, по всей вероятности,
тем, как уже говорилось, что раннее развитие всех глубоководных видов,
независимо
от глубины, на которой они живут во взрослом состоянии, происходит
в верхних горизонтах пелагиали.
Что касается нектонных глубоководно-пелагических рыб, которые, следует
напомнить, характерны главным образом для переходного слоя между
эпи- и мезопелагиалью
и для верхних горизонтов последней, то их географическое распространение
пока еще не изучено в деталях. Можно думать, что оно определяется,
как и распределение эпипелагического нектона, температурой воды и
кормовыми условиями.
Гемпилиды и веретенник судис (Sudis hyalina) имеют циркумтропические
ареалы, а алепизавры встречаются как в тропической зоне (там происходит
их нерест),
так и за ее пределами. Для антитропического кинжалозуба характерно
разобщение
репродуктивной части ареала, приуроченной к субтропическим водам,
и области нагула, охватывающей субполярные районы.
Оглавление
Предисловие ..................................................................................................................................................................
3
От автора .......................................................................................................................................................................
7
Введение ......................................................................................................................................................................
11
Приложение: Иерархия высших таксонов рыбообразных
и рыб, представленных в морской среде
обитания ......................................................................................................................................................................
22
Глава 1. Открытый океан как среда обитания рыб
..................................................................................................
23
Глава 2. Рыбы приповерхностных вод .....................................................................................................................
50
Глава 3. Рыбы глубоководной пелагиали ..........................................................................................................114
Глава 4. Донные, придонные и придонно-пелагические
рыбы ..........................................................................
169
Глава 5. Происхождение океанических рыб ..........................................................................................................
236
Глава 6. Современное состояние и перспективы
океанического рыболовства .................................................
254
Заключение ................................................................................................................................................................
270