Как жирафе седло
или
Нужен ли бог эволюции?
Ответ на работу А. К. Москвитина "Теория эволюции в
теологическом аспекте"
Текст оригинальной работы А. Москвитина, авторская орфография сохранена.
Комментарий П. Волкова дан синим цветом.
С момента появления на свет трудов, заложивших основу того, что ныне называется
теорией эволюции, а вернее с того момента, когда в обиход вошел сам термин "теория
эволюции" считалась и считается, что этой теории принадлежит фундаментальная
роль в вопросе формирования атеистического мировоззрения. И это понятно, поскольку,
отменив акт Божественного творения, теория эволюции вместо Творца вывела на
арену возникновения живого изменчивость и естественный отбор - факторы сугубо
материальные и наблюдаемые.
Тем не менее я рискну задаться вопросом: а чиста ли теория эволюции от Божественного
вмешательства? - и попробую ответить на этот вопрос.
Ну, для начала здесь стоит, вообще-то, доказать несомненное существование
некоей силы, которая в культурах разных народов именуется «бог» или «боги»
(в зависимости от того, как её понимала каждая из больших и малых цивилизаций
по всему миру). Как доказать? Элементарно. Предложить серию опытов, которые
в контролируемых и чётко определённых условиях дадут устойчиво воспроизводимый
результат. Этим результатом должно быть зафиксированное приборами (то есть,
независимое от ЛИЧНОСТИ экспериментатора) явление присутствия некоего существа
(ладно, применим для описания этого феномена термин «сущность»), которое должно
отвечать признакам, которые в культуре приписываются богу (богам): уметь сотворять
всё, что само пожелает, принимать жертвы, карать, быть милостивым, и т. д.
Пока такой эксперимент не будет проведён, пока он не докажет наличия данной
сущности, приплетать её к каким-либо рассуждениям, и искать следы её вмешательства
во что-либо просто бессмысленно.
Конечно, разные народы по-своему пытались познать богов. Но результаты этого
процесса ни коим образом нельзя считать научно достоверными. Употребление порошка
сушёного мухомора или каких-то неведомых белому человеку тропических трав,
разного рода психотехники, практикуемые в ряде религий (например, «крутящиеся
дервиши» ислама) – это не научные методы. Естественно, что они не проводятся
в контролируемых условиях; совершенно очевидно, что их результат полностью
субъективен; наконец, результат применения этих приёмов попросту невоспроизводим.
Сравнить результаты познания богов разными народами вполне возможно – для этого
достаточно почитать их мифы и легенды. Можно увидеть, что общность божеств
и их функций связана с общностью происхождения самих народов, либо с последующим
взаимовлиянием разных культур. Даже количество богов, извините, сильно варьирует:
от всего лишь одного в иудаизме и производных религиях до 330 миллионов в Непале
(автору этого комментария доводилось читать в журнале «Вокруг света» заметку
про Непал, и такая цифра приводилась в ней со ссылкой на подсчёты специалистов
по религиям). Плюс функции каждого из богов крайне отличаются: у политеистических
народов главным богом мог быть бог любого из явлений природы, а боги-«многостаночники»
имели различные наборы областей ведения, даже если одна из этих областей была
общей у богов разных народов. Ещё одна особенность – каждый народ считал (и
поныне считает) собственные результаты познания богов единственно правильными,
а прочие – заведомо неверными. К людям, исповедующим «не такую» религию, относятся,
в лучшем случае, с жалостью, а в худшем – с презрением и отвращением, считая
допустимым обманывать их, облагать налогами иную религию, ограничивать в возможностях
отправления религиозных культов, и даже просто убивать.
Сами видите, разброс результатов познания богов у разных народов говорит о
том, что объект познания отличается крайней эфемерностью признаков и их изменчивостью
в крайне широких пределах. Это заставляет подумать о том, что в действительности
какого-либо единого или многоликого объекта может попросту не быть.
Чувствую возражения оппонента: мол-де, религия давно уже принимает во внимание
наличие бога, и показала множество его проявлений в окружающем мире. Но религия
– это, извините, заинтересованная сторона (причём в свете реалий сегодняшнего
дня – материально заинтересованная в государственных масштабах). И ещё – все
доказательства подобного рода являются интерпретацией явлений окружающего мира
с «задней мыслью» о том, что бог всё-таки есть.
Понятие "экологической ниши"
Экологическая ниша - совокупность всех факторов среды в пределах которых возможно
существование вида в природе.
Ладно, к определению из «Биологического энциклопедического словаря» (М., «Советская
энциклопедия», 1989, стр. 730) я лишь добавлю для ясности ещё немного из соответствующей
словарной статьи:
«Совр. концепция Э. н. сформировалась на основе модели Э. н., предложенной
Дж. Хатчинсоном (1957, 1965). Согласно этой модели, Э. н. можно представить
как часть воображаемого многомерного пространства (гиперобъёма), отд. измерения
к-рого соответствуют факторам, необходимым для нормального существования вида.
Расхождение Э. н. разных видов путём дивергенции происходит б. ч. за счёт приуроченности
к разным местообитаниям, разной пище и разному времени использования одного
и того же местообитания».
Начиная изложение предмета настоящей работы с рассмотрения термина "экологическая
ниша" я имею целью показать какие последствия влечет за собой необдуманное
употребление даже безупречно сформулированных понятий. А к такому употреблению
я отношу выражение вид "занял экологическую нишу".
В основе тезиса о необдуманном употреблении этого понятия лежит тот очевидный
факт, что в любом установившемся биоценозе уже по определению связи, установившиеся
между входящими в него видами не имеют разрывов, ибо в противном случае он
не может считаться установившимся. То есть в стабильном биоценозе нет незанятых
экологических ниш.
Учёные считают, что это не есть факт.
Например, в Австралии к моменту колонизации её плацентарными грызунами были
свободные экологические ниши, не занятые местными сумчатыми. Далее, ограничения
сумчатых в области освоения водных биотопов (детёнышам, которые находятся в
сумке в течение нескольких месяцев, сами понимаете, водные процедуры противопоказаны)
привели к тому, что в Австралии не появилось своих местных «выдр». Утконос –
это, скорее, аналог выхухоли: он питается беспозвоночными. А вот хищного млекопитающего-рыбоеда
в реках Австралии так и не нашлось. Крокодилы, вараны и черепахи? Ну, да, отчасти
они «поделили» не востребованное млекопитающими место в экосистемах Австралии.
Но своим существованием они не исключили возможность существования рыбоядных
млекопитающих в тропических условиях, ведь в это же самое время в тропиках Азии,
Африки и Америки благополучно (если человек не вмешивается) обитают выдры разных
видов и родов.

"Моа-нало" - ныне вымерший гигантский гавайский гусь. Рост около полутора
метров - для гуся много, для "жирафа" маловато. |

Австралийский дроморнис,
птица, сближаемая с гусеобразными - одна из самых крупных птиц мира,
современник первых аборигенов. Ими же он и был истреблён. |
Хотим знать дальше? Поехали. Место действия: Гавайи
– рай для птиц, поросший тропическими лесами. Время действия – до колонизации
человеком. Действующие
лица – нало, гигантские гавайские гуси нескольких родов и видов, впоследствии
истреблённые человеком и известные по останкам из лавовых тоннелей и кухонных
куч аборигенов. Рост нало не превышал полутора метров, хотя аппетитная растительность
Гавайских островов процветала и на большей высоте. Почему бы не быть пернатому
«жирафу»? Рост новозеландских моа, разделивших позже судьбу нало, достигал
3,5 – 4 метров. Очевидно, почти таким же был рост нелетающих австралийских
дроморнисов и гениорнисов, сближаемых ныне с гусеобразными. Это говорит о
том, что нет принципиальных ограничений для того, чтобы гусеобразная птица
выросла
до
таких размеров. Ан,
нет на Гавайях гусей-жирафов, ни живых, ни мёртвых.
Ещё один вопрос: а есть ли вообще стабильные биоценозы? Если исходить из представлений
А. Москвитина, стабильным можно назвать лишь полностью закрытый от других
биоценоз, либо биоценоз, который наблюдается в течение очень короткого времени.
А если
брать большие отрезки времени и рассматривать судьбу биоценоза на их протяжении,
может выясниться, что стабильных биоценозов практически не существует! Точно
так же, бросив один взгляд на часы, можно утверждать, что часовая стрелка не
движется. Но, глядя на часы много раз в течение дня, наблюдатель убеждается,
что стрелки движутся вполне нормально.
Вернёмся для примера обратно на Гавайи.
Это превосходный пример островной изоляции –
располагаясь
в центре
Тихого океана
и никогда не
имев
сухопутной
связи с
материком,
Гавайские острова заселялись очень медленно. Так, до первых поселенцев на
Гавайях пополнение флоры островов, по расчётам учёных, шло крайне низкими темпами:
один вид за несколько десятков тысяч лет. В эпоху «флота каноэ», первых поселенцев
на островах, новые растения стали появляться на Гавайях с частотой нескольких
видов за век. А в наши дни на Гавайях регистрируется несколько новых видов
растений в год. Но, даже если оставить наше время и эпоху первых поселенцев,
видно, что Гавайи всё равно чисто физически не были закрытым сообществом.
На
них регулярно прилетали зимовать птицы, приносившие с собой семена, а ветры
и морские течения также способствовали обогащению флоры. Иными словами, «стабильный
биоценоз» чем-то подобен средневековому «идеальному рыцарю» - он существует
только на бумаге и в представлении А. Москвитина. В действительности между
биоценозами происходит постоянный обмен видами, более или менее интенсивный.
Это не значит, что в состав образующих данный биоценоз видов не могут войти
новые виды. Это значит только то, что теплого местечка для нового вида никто
не приготовил. Появление нового вида ведет к переформированию связей между
видами в обновленном биоценозе и после того, как эти связи приобретут устойчивый
характер, о новом виде можно будет сказать, что он "занимает определенную
экологическую нишу", которую он сам и создал, сформировав определенные
цепочки взаимоотношений с "аборигенами".
Не совсем точно. Для Австралии, как я уже сказал выше, допускалось наличие
свободных экологических ниш. Но это скорее исключение, чем правило. Обычно
же новых поселенцев в устоявшемся сообществе действительно никто не ждёт. И
всё же они приходят и даже натурализуются в уже имеющемся сообществе. Благодаря
своим особенностям, дающим преимущество в выживании, вид-поселенец может просто
«потеснить» уже имеющиеся виды, «вклиниться» между ними и получить свой «кусок»
общего «пирога» - этот процесс как раз можно назвать выражением «занял экологическую
нишу». Если у него нет таких преимуществ (сформировавшихся в иных условиях,
в которых он обитал раньше), то судьба этого вида в роли переселенца печальна
– он не сможет конкурировать с аборигенами, и в течение считанных поколений
вымрет (а то и вовсе не даст потомства).
Мало того, даже если нового вида в экосистеме не появилось, связи могут «пересматриваться»:
например, один из прежних видов травоядных в силу мутации получил возможность
эффективно нейтрализовать яд в листьях одного из растений, которое ранее занимало
незначительную долю в его рационе. Итог? Это растение займёт большее место
в его рационе, численность травоядного животного будет расти, а бывшего ядовитого
для него растения – сокращаться. Соответственно, видам живых организмов, зависящим
от этих двух видов, придётся либо хорошо, либо плохо – в зависимости от того,
на чьей они стороне. И неизвестно, чьим опылителем (может быть, очень узко
специализированным!) окажется бабочка, личинки которой кормились только на
листьях растения, ранее ядовитого для антилопы или жирафа…
Выражение же "вид занял экологическую нишу" не может не подразумевать,
что этот самый вид осознал факт несуществующего наличия такой ниши и мало того
- принял активные действия с тем, чтобы эту несуществующую нишу занять. Оставив
в стороне вопрос о существовании ниши зададимся вопросом, а как вид может осознать
ее наличие? Ответ очевиден - только надвидовым сознанием или же видовым подсознанием.
В общем, З. Фрейд со своим "коллективным бессознательным" может отдыхать.
Это самое "коллективное бессознательное" при изложенном подходе к
делу оказывается присущим уже колонии тараканов.
А вот тут уже видно, как совершенно необдуманно (я бы даже сказал, «насильственно»)
вводится некий «разумный компонент». Естественно, что я прошу А. Москвитина
привести факты, свидетельствующие о его наличии. В качестве этих фактов я принимаю
общепризнанные результаты опытов, реально проведённых компетентными в области
этологии учёными, проделанных с соблюдением чистоты научного эксперимента,
не вызывающих сомнения в своей объективности, результаты которых признаны мировым
научным сообществом. Не откажусь увидеть ссылки на публикации в научных журналах
и Интернете. А до этого знаменательного события оставим понятия «надвидовое
сознание» и «видовое подсознание» на откуп писателям-фантастам, а также специалистам
по психической энергии и торсионным полям, которых сейчас в России развелось
больше, чем легендарных «русских медведей».
Надо ли вообще вводить какое-то понятие «осознания» в процесс поселения какого-либо
вида в новом биоценозе? Особенно, если речь идёт о растениях? Или у растений
тоже есть некое «надвидовое сознание» и «видовое подсознание»? Тогда каким
местом… Извините, какой орган каждого отдельно взятого растения отвечает за
формирование этих самых квазимыслительных штуковин?
В трудах по теории эволюции никаких понятий подобного рода просто не существует.
Я, конечно, не исключаю, что А. Москвитин выводит наличие подобных процессов,
исходя из текстов школьных учебников и научно-популярной литературы, где автор
может для упрощения понимания (то есть, из дидактических соображений) применить
образное выражение. И атака, как я понимаю, идёт на это самое выражение. Помните
сказку и мультфильм «Бобик в гостях у Барбоса»? В ней как раз и есть пример
подобного рода: Бобик не понимал, как часы «ходят» без ног, и очень удивился,
что они умеют «бить»: «Они ещё и дерутся!?» Косой взгляд А. Москвитина на выражение
«вид занял экологическую нишу» - это пример того же рода.
Так что же, собственно говоря, означает выражение «вид занял экологическую
нишу»? Это же элементарно. Чаще всего он просто «расталкивает локтями» уже
существующие виды в том самом многомерном пространстве Хатчинсона, о котором
я упомянул выше. Он просто отбирает у «старожилов» большую или меньшую часть
этого самого пространства в тех пределах, где он сам может нормально существовать,
и где ему позволит это сделать успешная конкуренция с прежними обитателями.
Конкуренция происходит на границах жизненных интересов разных видов, где каждый
вид выживает и противопоставляет прессингу со стороны вида-конкурента какие-то
свои особенности и преимущества. И то, что для одного вида уже может быть пределом
выраженности какого-либо фактора, ограничивающего его существование, для другого
вида является фактором благоприятным или же очень далёким от лимитирующего.
Пример? Пожалуйста. В опреснённой лагуне (или в том же Азовском море) рядом
друг с другом плавает три вида рыб – карповая (лещ), бычок и камбала. Для леща
солёность Азовского моря – это предел, большей он просто не вынесет. Для камбалы
тоже предел – её икра уже не развивается в воде с такой низкой солёностью,
и для нереста она мигрирует в Чёрное море. И лишь бычок сможет нормально жить
и нереститься в воде с такой же, большей или меньшей солёностью, плюс переносит
колебания этой самой солёности. В этом его сильная сторона. Но в иных местах
обитания он будет поигрывать другим видам по другим показателям (способность
защищаться, добывать разные виды корма, и т. д.). Пример, конечно, приблизительный,
поскольку жизненные интересы этих рыб лишь немного соприкасаются.
Но здесь я, похоже, немного отвлёкся. Главный залог успеха в освоении новой
экологической ниши у вида-вселенца – это особенности анатомии и физиологии,
которые у него уже есть (сформировались в прежних условиях обитания).
Этот пример, надеюсь достаточно показателен. Впрочем, к употреблению термина "экологическая
ниша" я еще вернусь ниже.
Показательным в примере оказалось лишь то, что человек, явно не знакомый с
основами теории эволюции, пытается критиковать не саму теорию, а собственное
представление о ней. Либо пытается вводить в объяснения термины и понятия,
существование которых само по себе уже под большим вопросом.
Изменение условий обитания, как фактор, способствующий возникновению нового
вида
Парадоксы, неразрывно сопровождающие возникновение нового вида в свете теории
эволюции я предлагаю рассмотреть на примере возникновения такого вида, как
жирафа (Giraffa camelopardalis).
Как известно "…в желтой жаркой Африке не видать идиллий…" (В. Высоцкий).
Вот так случилось и с появлением на свет жирафы. Все было хорошо, но почему-то
расплодились (есть такое предположение) злые антилопы, которые начали активно
поедать растительность не только на высоте своего роста, но, вставая на задние
ноги, и много выше. Тем самым были нарушены сложившиеся трофические связи и
бедным предкам жирафы пришлось туго. То ли они категорически отказались вставать
на задние ноги, сочтя это крайне неприличным, то ли такое вставание оказалось
бесполезным вследствие малости их роста, но предки жирафы решили в полном соответствии
со словами еще не родившегося В.И. Ульянова пойти другим путем. А именно путем
эволюции, отрастив на этом пути длинные шеи. Перспективы это сулило блестящие
- ведь никакая антилопа не заберется на дерево, чтобы объесть листья, а жирафе
с длинной шеей и забираться никуда не надо.
Увы, это не сказка. Это вольное изложение предыстории появления вида Giraffa
camelopardalis рассказываемой адептами теории эволюции.

Палеотрагус (картина К.
К. Флерова) |

Самотерий (картина К. К.
Флерова) |
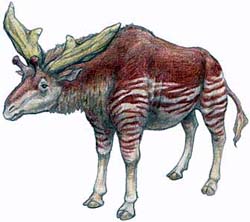
Сиватерий (рисунок с одного
японского сайта)
Трудно узнать "классических"
жирафов в этих ископаемых зверях, но с точки зрения систематики, палеонтологии
и анатомии все они - самые настоящие жирафы, пусть даже у них нет
легендарной длинной шеи.
|
Изложение крайне вольное, поскольку крупные антилопы
типа современной канны или нильгау на задние ноги вставать не могут. Антилопы
в Африке питаются преимущественно
двудольными растениями (зебры – более жёсткими однодольными (злаками)), растущими
на земле, и вставать на задние ноги могут лишь немногие виды. Примитивные жирафы,
внешне похожие на современного окапи (гелладотерий, палеотрагус), лося (сиватерий)
или того самого лубочного короткошеего жирафчика, над которым любят потешаться
креационисты
(самотерий), были достаточно высокими, чтобы любая тогдашняя антилопа, даже
вздумав корячиться на задних ногах, не дотянулась бы до той высоты, где кормились
они. Даже рост современного окапи достигает двух с лишним метров, чего любой
современной антилопе можно только желать. Таким образом, конкуренция жирафов
с антилопами получается не столь жёсткой, как пытается показать А. Москвитин.
Тем более, что жирафы не кормятся ни травой на земле, как большинство антилоп,
ни болотными растениями, как антилопы водяной козёл и ситатунга. И в горах,
на каменистых склонах, жирафы не обитают, а антилопы могут, и живут там. Иными
словами, у жирафов и антилоп точек соприкосновения жизненных интересов изначально
было немного. Только антилопы типа нильгау, похожие телосложением на примитивных
жирафов, либо тонкие и изящные существа типа геренука могли отчасти конкурировать
с первыми жирафами. Но это ведь не вся куча видов африканских антилоп, а единичные
виды!
А что до «неприличия» или «приличия» вставать на задние ноги – оставим это
суждение на совести А. Москвитина. Хотя в условиях жёсткой конкуренции о всякого
рода приличиях и прочих предубеждениях логичнее всего забыть, либо придётся
вымереть, но зато таким красивым и приличным.
А с кем же конкурировали-то жирафы, если не с антилопами? Не забудем про слонов,
которые всегда отличались более крупными размерами, чем антилопы, и объедали
растительность выше, чем они. Судя по всему, жирафы конкурировали даже не столько
с антилопами, сколько со слонами. А уж слонов-то разных видов и родов в Африке
в доисторические времена было много, причём приспособленных к самым различным
условиям обитания – от лесов и саванн до болот. И в наши дни у жирафов в Африке
единственный реальный конкурент в питании ветками с вершин деревьев – слон.
Поэтому жирафы, отрастив длинную шею, совсем не избавились от конкурентов –
они лишь уменьшили конкуренцию до достаточно низкого уровня.
Я готов допустить истинность этой предыстории при условии, что не мешает рассмотреть,
как же процесс пошел дальше. Необходимым условием процесса, между прочим, является
условие, что предки жирафы не встали перед проблемой изменения кормовой базы
вдруг. Понятно, что тогда бы они просто вымерли. Итак, процессы изменения кормовой
базы и отращивания шеи у жирафы были синхронизированы. И вот тут звенит первый
звоночек: какой фактор синхронизировал два независимых процесса - беззастенчивое
истребление растительности антилопами и медленное от поколения к поколению
отращивание шей жирафами? Такое отращивание шей это не изменение окраски
у бабочек в промышленных районах - не та плодовитость у жирафы и не та численность.
Антилопы к тому же не настолько гуманны, чтобы посматривать на предков жирафы
и вести себя в соответствии с тем насколько у тех удлинились шеи. Предки
жирафы
в свою очередь не могли ускорить процесс воспроизводства, чтобы угнаться
за антилопами. Значит синхронизирующий фактор был внешним, способным к оценке
ситуации (читай: разумным) и способным претворить в жизнь некоторые действия
в обеспечение требуемой синхронизации. Кто или что могло быть этим фактором?
- задам я риторический вопрос.
Что ж, действительно, доисторические жирафы не «вдруг» столкнулись с проблемой
конкуренции за корма. Наряду с ними растительность пожирали десятки видов антилоп
и множество видов совершенно незнакомых нам травоядных, например, халикотериев.
Семейство жирафовых известно с нижнего миоцена. Наряду с ними в это же время
существовали относительно рослые травоядные из семейства полорогих (как раз
к нему относятся злополучные антилопы), а также представители ещё нескольких
ныне вымерших семейств жвачных. Плюс из непарнокопытных лошади, носороги и
халикотерии (я перечисляю только группы из Старого Света, которые реально могли
конкурировать с первыми жирафами). Добавим сюда разнообразных динотериев и
мастодонтов из числа хоботных, а также тот факт, что ранние жирафы были не
столь рослыми, как сейчас, а это значит, что их конкуренция со всем этим доисторическим
зверинцем была крайне острая. Вот и получается, что жирафам пришлось серьёзно
постараться, чтобы «прогрызть» себе дорогу к кронам деревьев.
Замечу, к слову, что плодовитость тут никакой роли не играет. У хоботных (посмотрите
хотя бы нынешних слонов) плодовитость ещё ниже, чем у жирафов (один детёныш
раз в четыре года), но эволюционировали они столь же исправно, и видовое разнообразие
слонов на протяжении их истории было вполне сравнимо с жирафьим, и даже превосходило
его. Весь темп эволюции определяет скорость изменения среды обитания. Если
среда относительно стабильна, то поддерживается наиболее оптимальный тип представителя
вида (по-научному, действует стабилизирующий отбор). Если условия меняются,
получает преимущество одна из отклоняющихся от нормы форм, случайно оказавшаяся
адекватной этому изменению среды. Прочие вымирают, и бывшая «норма» постепенно
«съезжает» в число не очень хороших признаков, и становится неблагоприятной
для выживания. Это называется «движущий отбор».
В популяции условных пражирафов (их истинное систематическое положение пока
неважно, речь идёт о механизме) существовали как чуть более длинношеие особи,
так и чуть более низкорослые особи. Пока полорогие и прочие звери позволяли
им жить, как есть (то есть, не вторгались в их экологическую нишу), пражирафам
для нормального кормления достаточно было их обычных шей. Почему раньше не
лезли к ним? Возможно, ели разные виды растений (как сейчас антилопы в Африке
– куча видов на одной территории, но друг другу не мешают), или действительно
были совсем коротышками рядом с дюжими, выше оленя, первыми жирафами. Внезапности
вторжения не было – просто какой-то сосед условных пражирафов освоил новый
вид растительной пищи, ранее доступный лишь пражирафам, либо кто-то новый пришёл
из Евразии в эти экосистемы и слегка «растолкал локтями» установившееся сообщество.
Естественно, он не вышиб полностью пражирафов с их исконных кормушек, но просто
дал понять, что тоже имеет на это дело виды. Что стало со средой? Она изменилась!
И теперь понятие «нормы» также поменялось – оптимально выживать теперь начали
не пражирафьи коротышки, которых объедали наглые копытные пришельцы (или бывшие
добрые соседи), а чуть более долговязые существа. Времена изменились, и теперь
коротышкам не стало житья. Они потихоньку стали более редкими, а потом и вовсе
сгинули. Зато «нормой» стали те, кто раньше даже мог испытывать некие неудобства
из-за высокого роста. И естественный отбор в новых условиях ПОМЕНЯЛ направление
– он стал не приводить всех пражирафов к некоей умеренно-рослой разновидности,
но стал способствовать тому, чтобы жирафы приспосабливались не только анатомически,
но и поведенчески к поеданию высокорослых растений, на которые за миллион лет
до этого они даже не смотрели.
Можно предположить также, что из-за деятельности наземных травоядных часть
растений стала более рослой, чтобы «проскочить мимо рта» всевозможных прожорливых
коротышек, и пышно разрастись на высоте, недоступной носорогу, мелкой лошади
или антилопе. И единственные, кто смог воспользоваться таким источником пищи,
были слоны и жирафы. Так что возможно даже, что деятельность антилоп и их соседей
не только не помешала жирафам кормиться, но даже наоборот, сделала их стол
изобильным.
Одновременно в популяциях пражирафов шло накопление разных признаков, позволявших
оптимизировать существование более крупному и долговязому существу. Какие-то
признаки были бесперспективными, и число их носителей в каждом поколении снижалось.
А другие признаки, наоборот, становились достоянием всей популяции.
Подводя промежуточные итоги, скажу так: жирафы всегда жили в условиях конкуренции,
поэтому новая беда «свалилась» на них не сразу и не вдруг на всю популяцию
в полной мере. Среди пражирафов были и такие, кто в силу определённого рода
полиморфизма в популяции мог избегать действия нового фактора. Изменение условий
обитания вызвало изменение направления отбора, и бывшая «ненорма» стала оптимумом.
А что же некий «синхронизирующий фактор», на который возлагает надежды А. Москвитин?
Тут стоит спросить: а был бы процесс удлинения шеи жирафами, не появись у них
конкурент? У жирафов, как я понимаю, нет врождённого стремления вытягивать
шею. Понимаю, что «если бы, да кабы…», но представим себе ситуацию, в которой
не было бы у тех самых пражирафов наглого и жизнелюбивого конкурента. Тогда
и остались бы жирафы короткошеими крепко сложенными молодцами. Ведь среди всего
множества жирафов нынешние долговязые существа, которых мы представляем себе
при слове «жирафа», представляют собой скорее исключение из правила. Было много
вполне привычных по облику, не долговязых жирафов – гелладотерий, либитерий,
самотерий и сиватерий, например. Несмотря на отсутствие длинной шеи, с точки
зрения систематики это тоже жирафы. Но конкуренция с другими копытными была
столь велика, что короткошеим видам не суждено было выжить в саванне – лишь
окапи уцелел в лесах, где самые рослые антилопы всего около метра ростом. Так
что жирафы сознательно или бессознательно не тянули шею вверх лишь потому,
что они жирафы. Появление конкурентов было лишь толчком к изменению в этом
направлении. Без него слишком длинношеие пражирафы чувствовали себя хуже из-за
разного рода физиологических проблем.
Я не исключаю того, что, «опередив» каких-то полорогих или непарнокопытных
конкурентов, «уже-довольно-рослые» жирафы прошлого столкнулись с конкурентами
в лице мастодонтов и слонов, что подстегнуло новый виток эволюции. А никакой
синхронизации и в заветах не было – была лишь причинно-следственная связь:
одно событие послужило толчком для другого. События-то как раз были зависимы,
но не с точки зрения анатомии, а с точки зрения экологии. А уж в какой форме
проявились последствия – это дело десятое. Пражирафы могли, например, научиться
есть такой кустарник, от которого любой их конкурент сразу копыта отбросит.
А что? Коалы, например, пошли таким путём – кто кроме них сможет глодать малопитательные
и ядовитые листья тех эвкалиптов, что они едят?
Я должен упомянуть, что сам процесс изменения длины шеи у предков жирафы не
невозможен. Ничто не мешает произвести нечто подобное путем выведения длинношеей
породы, скажем, коров. Практическая ценность будет равна нулю, но результат
может впечатлить.
Возникает лишь вопрос «зачем оно надо?»… Когда это действительно необходимо,
длинношеие животные появляются в природе исправно. Те же маменьчизавры и индрикотерии,
например.
То есть некий постоянно действующий и действующий достаточно мягко фактор
в принципе мог стать фактором естественного отбора, способствующего увеличению
длины шеи у предков жирафы. Что это за фактор - не берусь судить. Могу только
сказать что к его определению нужно подходить с большой осторожностью, иначе
можно поиметь то, что было сказано выше, в том числе и в части употребления
термина "экологическая ниша".
Но отвлекусь на время от процесса отращивания шеи предками жирафы
Репродуктивно изолированные популяции
Перейду от жирафы к мышам. Мыши - моя слабость. Довелось мне быть в экспедиции
на озере Чаны (Новосибирская область). Лавливали мы там мышей как на островах,
так и на материке. Так что кто-то крутил хвосты коровам, а я проделывал то
же самое с мышами.
Возьму для примера далеко не фантастическую ситуацию. Жила на острове популяция
каких-то там полевок. Жила и горя не знала. До той поры пока не случился катаклизм,
разделивший остров пополам и изменивший условия проживания полевок на этих
двух частях острова, причем изменил в разных направлениях. Многие полевки повымерли,
но нашлись и выжившие, поскольку оказались они носителями аллелей, контролировавшими
наличие признаков, обеспечивших выживание в новых условиях. Подобные процессы
идут постоянно. Катаклизм же придуман мной просто для ускорения процесса. Таким
образом в итоге возникли две разные популяции полевок, да к тому же из-за перераспределения
состава аллелей в их геноме стали они репродуктивно изолированными. То есть
и тут полевки и там полевки. И предки у них общие, родня одним словом, а вот
потомства поиметь друг от друга не могут. Что поделать, дело житейское.
И все бы было ничего, только вот в дело вмешивается понятие "биологический
вид".
Очень коротко вид определяется как репродуктивное сообщество. А это определение
имеет далеко идущие последствия. Одним из этих последствий является то, что
те самые две популяции полевок на несчастном острове взаимно представляют два
новых вида. Пусть даже отыщется предок этих полевок, счастливо живущий на соседнем
острове. Пусть этот предок может иметь плодовитое потомство как с представителями
одной так и другой из вновь возникших популяций, пусть представителей этих
двух популяций с первого взгляда друг от друга не отличить, все равно это два
разных вида и все тут. На этот счет теория эволюции сурова. Если сказано, что
бурундук - птичка, значит, никаких зверьков!
Что же, ситуация, указанная А. Москвитиным, повторялась в природе без всяких
«допустим» много раз. И один из итогов такого явления, описанный А. Москвитиным,
имеет место быть в реальном мире. В книге В. Гранта «Эволюция организмов» указано
формирование подобных популяций на примере насекомых и растений. Среди них:
североамериканская пчела Hoplitis producta, подвиды которой ведут себя по отношению
друг к другу как хорошо сформированные виды. Также подобные полувиды известны
для мухи Drosophila palustrosum из тропиков Нового Света. Для понятия вида
существует очень много определений, учитывающих разные аспекты этого вопроса.
В «Биологическом энциклопедическом словаре», которым (надеюсь!) пользовался
А. Москвитин, сказано:
«Строгое общепринятое определение В[ида] до сих пор не разработано, обычно
под В. понимается совокупность популяций особей, способных к скрещиванию с
образованием плодовитого потомства, населяющих определ. ареал, обладающих рядом
общих морфо-физиол. признаков и типов взаимоотношений с абиотич. и биотич.
средой и отделённых от других таких же групп особей практически полным отсутствием
гибридных форм».
Таково определение вида, учитывающее большинство критериев. Под данное А. Москвитиным
определение «репродуктивное сообщество» вполне подойдёт простая популяция –
внутривидовое образование. Характерным признаком популяции как раз является
самовоспроизводимость в течение неограниченно долгого времени. Исходя из данного
А. Москвитиным понятия, я могу скромно предположить, что товарищ просто не
хочет принимать во внимание множество остальных аспектов существования вида.
А ведь они очень важны. Поэтому «далеко идущие последствия» приведённого куцего
определения ограничатся лишь тем, что под него можно будет просто подсунуть
что-то иное, и плавно сменить тему обсуждения. Но последуем авторской логике,
и ограничимся критерием репродукции.
Естественно, видовая самостоятельность некоей группы живых организмов определяется
далеко не так, как представляет неискушённому читателю А. Москвитин. Разумеется,
авторитет бородатых или лысоватых дядюшек в строгих очках, живших десятки и
сотни лет назад, а ныне взирающих на нас с портретов в аудиториях и кабинетах,
так просто не отбросишь (особенно, если они всё же правы!), но ведь сказал
же в своё время старик Аристотель, что у насекомых восемь ног! Конечно, человеку
свойственно ошибаться. Но, кроме того, в наше время наука шагнула далеко вперёд,
и на её вооружении стоят передовые методы исследования.
Помните, в каком восторге был Шерлок Холмс, когда открыл реактив, позволяющий
отличить кровь от красной краски? А что бы он сказал, если бы узнал, что по
информации, «вытянутой» из мазка крови, современные криминалисты смогут определить
множество личных генетических параметров человека? То же самое можно сказать
про биологию. Сейчас на вооружении биологов стоят новейшие технологии, а новые
виды описывают не только на основе ломких гербариев и блеклых заспиртованных
образцов! Так известные сто лет назад виды рыб геофагусов (Geophagus (Satanoperca))
семейства цихлид (Cichlidae) поделили ещё на несколько новых на основе особенностей
поведения. Генетика и изучение белков позволили выделить из одного вида озёрных
лягушек (Rana ridibunda Pallas, 1771, Что может быть более знакомым? Больше
двухсот лет известна!) десяток новых видов с перспективой дальнейшего деления!
А ведь если их выложить в баночках перед любым самым искушённым герпетологом,
он не различит ни одного из этих видов – их можно различить лишь путём электрофореза
белков. Тут их не то, что «с первого взгляда», а даже со «второго», «третьего»
и «десятого» взгляда так просто не различить. А вы говорите – мышам хвосты
крутить...
Кстати, сходное явление наличия видов-дублёров (так это явление называется)
обнаружено у серых крыс (выделен вид крыса туркестанская, внешне крайне незначительно
отличающийся от обычной серой крысы), а также у малярийных комаров Anopheles.
Причём одни виды-дублёры комаров способны переносить малярию, а другие – нет.
Внешние различия у них почти не выражены (например, цвет оболочки яиц). Поэтому
в наше время совершенно не лишними являются генетические исследования, которые
проводят, изучая голотип (образец, по которому проводится описание) вновь описываемого
вида. Поэтому, прежде чем выдать диагноз «бурундук – это птичка», отдельно
взятый специалист перелопатит гору литературы, и ещё с десяток специалистов
«поставит на уши». И то дальнейшие открытия и исследования могут поставить
его диагноз под сомнение. Не бойтесь, уважаемый А. Москвитин, если уж бурундук
и впрямь зверёк, то рано или поздно найдётся специалист, который признает вашего
бурундука
зверьком. Может, не на вашем веку, но будет. А если он и взаправду птичка,
то никто его ни за какие коврижки зверьком не признает. Но всё будет проведено
аргументированно, со ссылкой на специалистов.
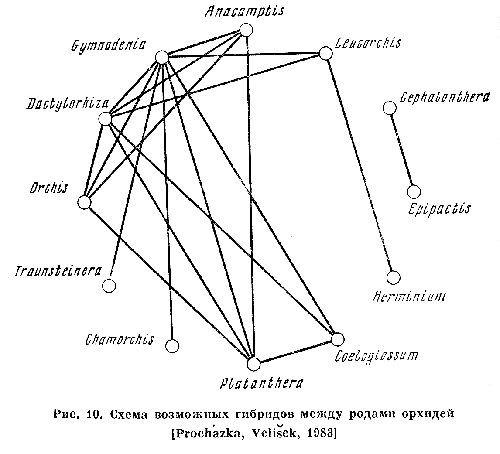 Схема
из книги М. Г. Вахрамеевой и др. "Орхидеи нашей страны" (М., "Наука",
1991, стр. 56) показывает возможность естественной гибридизации европейских
орхидей.
Схема
из книги М. Г. Вахрамеевой и др. "Орхидеи нашей страны" (М., "Наука",
1991, стр. 56) показывает возможность естественной гибридизации европейских
орхидей.
|
|
|
Рукотворные гибриды
орхидей, результат межродового скрещивания этих растений (сверху вниз):
Одонтиода
(Одонтониум х Кохлиода)
Вуйлстекеара (Мильтония х Кохлиода х Одонтоглоссум)
Брассолелиокаттлея (Брассавола х Лелия х Каттлея)
Потинара (Брассавола х Лелия х Каттлея х Софронитис)
|
Вернёмся к нашим полёвкам. Судя по тексту,
А. Москвитина очень удивляет теоретическая возможность их давать потомство с
родительской популяцией, и не скрещиваться друг с другом. Что же, в природе
это наблюдается, и учёные как-то не склонны считать это чем-то чудесным и из
ряда вон выходящим. Я уже упоминал примеры с пчелой Hoplitis producta и мухой
Drosophila palustrosum. У этих видов (правильнее их считать уже «надвидами»)
как раз проявляется подобное явление. В опытах по совместному разведению разных
полувидов дрозофил гибриды между симпатрическими (обитающими в природе вместе)
формами наблюдались гораздо реже, чем между аллопатрическими (раздельно обитающими)
формами – коэффициент изоляции в среднем 0,85 против 0,67 соответственно. Здесь
уже видна изоляция на уровне поведения – размножающиеся особи избегают «чужаков»,
обитающих совместно, хотя легче дают гибриды с «незнакомцами».
Репродуктивная изоляция не всегда проявляется на
генетическом уровне. Иногда виды, вполне способные давать гибриды, не образуют
их по иным причинам. Золотой (Chrisolophus pictus) и алмазный (Ch. amherstiae)
фазаны легко образуют неограниченно плодовитые гибриды в фазанниках. В конце
19 века любители-птицеводы даже сетовали, что чистокровного алмазного фазана
уже не найдёшь – одни гибриды. Но в природе они даже не встретятся – алмазный
фазан обитает в горах Юго-Восточной Азии, а золотой – в тёплых долинах. Цихловые
рыбы озёр Танганьика и Малави (Ньяса) представлены множеством видов. В природе
в каждом озере разные виды цихлид живут совместно, но гибриды между ними крайне
редки. В аквариумах же они дают настолько много гибридов, что в клубах любителей
этих рыб специально проводят сохранение «чистых» природных видов. Дело в том,
что любитель, стремясь развести редкий вид, отбирает и инкубирует икру искусственно.
В результате мальки-«сироты» не запечатлевают родителей как особей своего вида,
и более склонны к образованию гибридов. Ещё один мой любимый пример – орхидеи.
В природе гибриды орхидей встречаются, хотя крайне редко: орхидеи крайне разборчивы
в опылителях, и иной раз «доверяют» услуги опыления лишь одному – двум видам
насекомых. Но в теплицах, где насекомых нет, а мягкая кисточка в руках цветовода
присутствует, выведены разнообразные гибриды, в том числе межродовые. Так, к
роду вуйлстекеара (x Vuylstekeara) отностятся гибриды мильтонии (Miltonia),
кохлиоды (Cochlioda) и одонтоглоссума (Odontoglossum), к роду софролелиокаттлея
(x Sophrolaeliocattleya) гибриды софронитиса (Sophronitis), лелии (Laelia) и
каттлеи (Cattleya). Вильсонара (Wilsonara) – также тройной межродовой гибрид,
а потинара (x Potinara) – вообще четверной.
Можно предположить (в том числе на основе результатов опытов с полувидами дрозофил),
что генетический барьер возникает чаще всего между симпатрическими видами. Но
пример с цихлидами озера Танганьика говорит, что это не абсолютная истина, и
иные барьеры (поведенческий, например) также могут обеспечивать нескрещиваемость
симпатрических видов.
Здесь необходимо сделать важное замечание. Теория эволюции прекрасно объясняет
процессы подобные удлинению шеи у предков жирафы или возникновению репродуктивно
изолированных популяций у мышей. Объясняет убедительно и обоснованно. Забывая,
однако, зачастую упомянуть, что происходят эти процессы за счет рекомбинации
уже содержавшегося в геноме предков генетического материала.
Ага, спинным мозгом чувствую, что мы возвращаемся к старой-престарой сказке
о том, что полезных мутаций не бывает. К чему я это говорю? Да всё к тому же
– что разнообразие генетического материала пополняется как раз за счёт мутаций.
Но об этом – немного ниже.
Что следует из этого замечания? Только то, что теория эволюции непогрешима
в части микроэволюции - эволюции внутри вида. Но как ни выдавай репродуктивно
изолированную популяцию мышей за новый вид, они останутся теми же мышами. А
значит и предки их были мышами, и предки предков. Продолжая же эту ретроспективу
мы упремся в изначального предка мышей, который был той же самой мышью, то
есть (звучит второй звонок) в барамин (изначальную популяцию), который возникновением
своим обязан Создателю. Это называется - приплыли…
Хе-хе-хе, прямо «теми же мышами»? Или всё же новым видом? Продолжим цепочку
рассуждений… Мыши двух разных биологических родов – это «те же мыши», или уже
не «те же»? Где она, эта самая грань между «ещё теми» и «уже не теми»?
В процессе эволюции происходит переход, если угодно, количественных изменений
в качественные. Когда у популяции какого-либо вида накапливается слишком большое
количество специфических признаков, отличающих её от прочих группировок такого
уровня, учёные вправе задуматься над тем, представляет ли она собой тот же
самый вид, или уже новый. То же самое происходит на более высоком уровне –
рода, семейства, отряда.
Если «продолжить ретроспективу» и, в сущности, отбросить все признаки специализации,
«наслоившиеся» на «первообразец» – это будет «та же» мышь, или нечто иное?
Если отбросить признаки специализации хомяков, то «изначальный хомяк» будет
«тем же» хомяком, или нет? А если поставить рядом «первомышь» и «первохомяка»,
будут ли они отличаться друг от друга как представители двух разных родов (не
в библейском, а в биологическом смысле!), или как два вида одного рода? Беда
А. Москвитина в том, что он просто не представляет себе многообразия той или
иной группы живых существ, приводя её как пример. Он рассуждает на «бытовом»
уровне, в то время, как истинное многообразие тех же мышей, крыс или хомяков
фантастически велико. Если собрать всех их вместе, А. Москвитин, пожалуй, затруднится
сказать, где будет крыса, а где крысовидный хомяк рода ихтиомисов, либо не
различит мышь (одного из многих родов!) и какого-нибудь мышевидного хомяка.
И возможно, что он спутает мышовку из семейства тушканчиков с какими-то экзотическими,
но всё же мышами. Так что креационизм непогрешим лишь на бытовом уровне, достаточном
для обывателя. При более внимательном рассмотрении и обоснованных фактическим
материалом «придирках» он съёживается и испаряется, как снег под весенним солнцем.
Ещё один вопрос – введение в рассуждение термина «барамин». Что он означает?
Вот цитата из работы У. Дж. Гладсхауэра «Как возник наш мир»:
«Для отличия библейского понятия вида от принятого
в биологии профессор Фрэнк Марч использует здесь термин "барамин" (сотворенная группа; евр. "бара" –
творить, "мин" – род). Мы хотим использовать этот термин для обозначения
отдельной группы живых организмов. Эта группа включает все родственные разновидности,
происходящие от созданного в дни творения одного представителя библейского
вида. Напрашивается вопрос: насколько велика такая сотворенная группа? Насколько
она соответствует биологическому виду? Или она ближе к роду или семейству?
В любом случае бесспорно, что библейское понятие вида "барамин" почти
всегда шире биологического.»
Понятие «барамин» крайне расплывчато по объёму. В креационной литературе
упоминается о возможном соотношении «барамина» с таксономическими единицами.
Вот отрывок
из замечательной работы И. И. Дзеврина и П. В. Пучкова ««Сад» креационизма
или «дерево» эволюции?» (её
можно найти здесь),
где дана ссылка на книгу Г. Морриса «Библейские основания современной науки»:
«Таксону какого ранга соответствует baramin? По
Моррису "это иногда вид,
иногда род, иногда семейство", но чаще "по крайней мере среди высших
животных ... по-видимому, семейство: медвежьи, волчьи и т. п. ... может приблизительно
представлять изначальный род"»
С точки зрения науки такая классификация не может считаться естественной. В
данном случае в основу её заложено не естественное родство живых существ (возможное
возражение креациониста: «… выдвинутое предположительно в свете теории эволюции»),
а итог принципиально недоказуемого действия некоей сущности, чьё существование
само по себе нуждается в более чем серьёзных доказательствах. Точно так же,
привлекая к доказательству непроверяемое и невоспроизводимое действие высших
сил, я могу утверждать, что, хоть в большинстве случаев дважды два равняется
четырём, в моём отдельно взятом случае в результате чудесного вмешательства
оно стало равно пяти. Естественно, такое доказательство «не потянет» даже у
доски в первом классе. Тем не менее, им пользуются мужи, считающие себя учёными.
«Барамин» невозможно соотнести ни с одним таксоном живых существ. Попытки «привязать»
его к семейству (или даже отряду – «ступеньке более высокого ранга) обязательно
натыкаются на бытующие в среде учёных тенденции деления и слияния таксонов
на основании данных анатомии, физиологии, генетики, паразитологии, палеонтологии
и прочих дисциплин. Чтобы мои рассуждения стали более понятными, приведу простой
пример. Курица и кукушка – птицы разных «родов» (библейских), или одного? Несомненно,
разных. Тогда курица и фазан – это птицы разных родов, или одного? Возможно,
кто-то скажет, что разных, а кто-то вспомнит многочисленных азиатских фазанов
с гребешками (хотя бы того же серебристого фазана (Lophura nycthemera)), и,
скрипя зубами, сведёт их в один «род». Возможно, методом таких поэтапных рассуждений
все куриные птицы смогут быть сведёнными в один «барамин», чётко отделимый
от прочих. Сюда же войдут примитивные гокко, или краксы – куриные птицы из
Южной Америки. И тут придётся обратить внимание ещё на одну птицу, которая
точно известна креационистам. Это гоацин – излюбленный ими пример «птицы, сохраняющей
в наши дни черты примитивного археоптерикса». В систематике гоацин «сидит на
двух стульях» - он близок как к низшим куриным, так и к кукушкообразным. И
эта птица становится связующим звеном между существами, которых, на первый
взгляд, можно отнести к разным «бараминам»: одни орнитологи считают его примитивным
куриным, другие – кукушкообразной птицей. В свете этого как-то даже бледно
выглядит тот факт, что цесарки и индейки могут как помещаться среди фазановых,
так и выводиться в самостоятельные семейства. Если этого примера мало, приведу
ещё один: систематическое положение семейства барракуд (Sphyraenidae). Одни
учёные относят их к отряду атеринообразных, а другие – к кефалеобразным. И
всей этой ситуации придаёт пикантность тот факт, что указанные отряды рыб принадлежат
даже к разным надотрядам – таксонам ещё более высокого ранга, чем отряд. Похоже,
границы «барамина» тут расплылись до безобразия. Если «копать» дальше, то на
арене обязательно появятся одноклеточные жгутиконосцы – связующее звено между
растениями и животными, с одинаковым успехом помещаемые и среди царства растений,
и среди царства животных. Здесь где прикажете проводить границу между «бараминами»?
Иными словами, расплывчатость и субъективность понятия «барамин» делают его
совершенно неприменимым к реальной систематике.
Новый вид и ароморфозы
Ароморфоз - … эволюционное преобразование строения и функций организмов, имеющее
общее значение для организма в целом и ведущее к морфофизиологическому прогрессу…
Взглянув на собаку и кошку, каждый вправе воскликнуть: "Могём мы ямбов
от хореев, как вы ни бейтесь, отличить!" То есть собака есть собака, а
кошка есть, соответственно, кошка. Это два разных вида и как представители
этих разных видов, они имеют разные, но общие для кошек и общие для собак признаки,
которые обоснованно можно назвать видовыми.
Предположив, чисто умозрительно, конечно, что и те и другие произошли от общего
предка, нельзя отрицать того, что тут дело без ароморфозов не обошлось.
Почему? В данном примере это вполне возможно отрицать. Именно тут как раз
обошлось. Указанные различия между кошкой и собакой – это идиоадаптации, приспособительные
изменения частного характера, не влияющие на общую организацию рассматриваемой
группы организмов. И кошка, и собака остались млекопитающими со всеми вытекающими
отсюда признаками. Они по-прежнему принадлежат к одному отряду внутри данного
класса, разве что, семейства разные. Потому говорить, что кошка по уровню организации
стоит выше собаки (или наоборот) – это бессмысленно. А вот то, что А. Москвитин
«не могёт» ароморфозов от идиоадаптаций отличить, говорит о многом. Возможно,
именно в этом неумении кроется причина написания им этой самой работы.
Каков же механизм возникновения ароморфозов. Согласно теории эволюции ароморфозы
возникают вследствие мутаций и естественного отбора.
Возьмем этот факт на заметку и вернемся к предкам жирафы, неустанно отращивающим
шеи.
А при чём тут эта идиоадаптация? Увы, увы, увы. Второй раз на те же грабли,
причём подряд. Жирафы при своей шее остались млекопитающими, разве что стали
очень специализированными. А так наряду с длинношеими существами, которые представляются
обывателю при слове «жираф», в семейство жирафов входит множество ископаемых
родов этих зверей, плюс современный окапи, также не отличающийся длинной шеей.
Для специалиста по систематике млекопитающих «жираф» - это, в первую очередь,
животное с определённым строением зубов, черепа, рогов, и т. д. И лишь как
частная специализация это существо длинношЕЕЕ. Так что в удлинении шеи ароморфозов
не наблюдается.
Мобилизовав все ресурсы имевшегося у них исходно генома, они приходят к выводу,
что процесс нуждается в продолжении, тогда как ресурсов уже нет. Остается одна
надежда - мутации.
И вот тут-то звенит третий звонок.
Мутации - вещь сугубо случайная, результаты их непредсказуемы и происходят
они далеко не каждый день. А предкам жирафы необходимо, чтобы они были синхронизированы
все с тем же фактором, ухудшающим кормовую базу, с возможностями репродуктивного
воспроизводства самих пражирафов и имели сугубо определенную направленность.
Удлинение шеи ставит, кроме всего, вопрос о кровоснабжении мозга. Проблема.
Решена она у жирафы наличием артериальных клапанов. Но как она решалась, вот
в чем вопрос! Возникали ли эти клапаны один за одним, пока (кем-то) не было
сказано: "Хватит!" Или они возникли все сразу, а шея потом сама доросла
до заданного этой клапанной системой ограничения? Вопросов масса, но решены
они могут быть только при наличии все того же внешнего управляющего фактора
- разумного и деятельного. И тем более разумного и деятельного, поскольку требуемый
ароморфоз возможен лишь при синхронизации уже трех процессов - к названным
выше двум прибавляется третий - целенаправленный отбор мутации. Целенаправленный
потому что требуемым итогом этих мутаций является возможность создания для
себя жирафами, которым предстоит возникнуть, экологической ниши, частью которой
является корм на вершинах деревьев. Сбой означает вымирание вновь возникающего
вида. И тут уже естественный отбор нам не помощник, поскольку он может отбраковать
единичные вредные мутации но не в силах определить потенциальную полезность
каждой мутации из той серии конечной и неизвестной ему целью которой является
возникновение нового вида.
Ни одного ароморфоза тут не было указано, поэтому рассуждения А. Москвитина
о жирафах кажутся мне совершенно неуместными. Вот, кабы рассказал он о происхождении
зверей от тероморфов, либо о широком «потоке» птицеобразных форм мезозоя, параллельно
развивавших птичьи признаки – это было бы в самое «яблочко». А так – я остаюсь,
прямо-таки, в состоянии полного непонимания. И к чему тут жирафы?
В представлениях креационистов мутации или на 100% вредны, или «практически
всегда» негативны и лишь «в лучшем случае» нейтральны. Отталкиваясь от этого
допущения, они объясняют появление новых признаков именно таким способом –
перекомбинацией уже существующих. В свою очередь я могу обратиться к первоисточникам
креационизма, а именно – к Библии, и спросить креационистов: а как объяснить
тот факт, что в настоящее время в популяциях разных животных с диплоидным набором
хромосом существует больше, чем по 4 аллеля для одного гена? Как они появились,
если Ной взял на ковчег только по паре животных разных видов? Ведь тогда они
должны нести максимум по 2 аллеля одного гена каждое при условии, что ген не
дублируется (такое бывает - например, оттенок кожи человека кодируется не двумя,
а четырьмя генами). Можно, конечно, придраться к этому, и сказать, что взято
было по две пары «нечистых»
и по семи
пар «чистых»,
но так говорится в одном месте Библии, а в другом сказано, что взято было лишь
по паре. На мой взгляд, это лишний раз говорит о том, что Библию нельзя использовать
как буквальный и достоверный источник информации.
Немного отвлёкся я от основной темы. Конечно, я не телепат и не психоаналитик,
но то, что А. Москвитин не упомянул о мутациях, рассуждая о причинах генетического
многообразия, заставляет думать о том, что его точка зрения аналогична таковой
у большинства креационистов. Если я не прав, то я просто прошу автора данной
работы уточнить свою точку зрения, указав источник УВЕЛИЧЕНИЯ генетического
многообразия в популяции.
Почему-то креационистами деликатно опускается такой вопрос, как относительность
пользы и вреда мутаций. Про многие мутации нельзя точно и однозначно сказать,
«полезны» они, или «вредны». Например, при исчезновении или ослаблении действия
какого-либо фактора окружающей среды «вредная» мутация может стать «нейтральной».
А если, например, «вредный» при атаке хищника признак приносит пользу в брачный
сезон, помогая очаровывать особей противоположного пола, он автоматически переходит
в разряд «полезных» для вида, хотя носителям этого признака рано или поздно
приходится нести «персональную ответственность» за его наличие, причём собственной
головой. Кроме того, в природе существуют разного рода нейтральные мутации,
которые могут обнаружиться, разве что, в лабораторных условиях. Даже у человека
такие есть: например, чувствительность к фенилтиокарбамату (описанная в старой
книге Шарлотты Ауэрбах «Генетика»). Одни воспринимают его на вкус как нечто
горькое, а другие совершенно не ощущают его вкуса. Я, например, не знаю, какова
моя чувствительность к этому веществу. Полагаю, что большинство читающих эти
строки тоже не скажет о себе в этом плане ничего определённого.
Ещё одна традиционная ошибка креационистов, которой можно избежать, изучив
школьный курс общей биологии. Они упорно удивляются тому, как «для появления
такого рода изменений у особи должен произойти целый ряд мутаций одна за другой»
(приблизительно такими словами выражается их недоумение). В работе А. Москвитина
это чётко не прописано, но ощущение присутствия такой мысли текст его работы
навевает. Во всяком случае, идёт чёткий намёк на то, что мутаций, мол-де, маловато
будет, и редкие они. Но выход из этого явно надуманного «тупика» прост: единица
эволюции – не особь, а популяция. Ждать цепочки мутаций в потомстве одной конкретной
пары можно очень долго. А в многочисленной популяции мутации происходят в большем
количестве, и их носители имеют шансы передать их потомству. Какие-то из мутаций
отсеются, какие-то распространятся. Между популяциями одного вида идёт постоянный
обмен особями, который приводит к перемешиванию генов и возникновению новых
сочетаний, иной раз с неожиданным эффектом (это явление сочетанного действия
генов). Поэтому в «родословную» конкретной пары особей (если целесообразно
будет в порядке тысячелетнего эксперимента отслеживать её в поколениях) нужный
ген может прийти просто в готовом виде «со стороны».
Обратим внимание, как плавно вводится в рассуждение некий разумный фактор.
Вначале вопрошается: «так, или не так?» А затем ответствуется: «только так,
и не иначе!» Причём анализ невозможности иного исхода событий остаётся где-то
«за бортом» рассуждений. Я не согласен с А. Москвитиным, и мне как читателю
Также не совсем понятно рассуждение А. Москвитина о возможности «создания для
себя жирафами, которым предстоит возникнуть, экологической ниши, частью которой
является корм на вершинах деревьев». Боюсь, что, поучая читателей в начале
своей работы, А. Москвитин сам забыл, о чём идёт речь. Ведь он сам определил
экологическую нишу как «совокупность всех факторов среды в пределах которых
возможно существование вида в природе». Я, конечно, мог бы представить себе,
как жирафы влияют на климат в местах своего обитания, разгоняя тучи рожками…
Но моего воображения на это не хватит. Жирафы, равно как и другие виды, не
«создают для себя экологическую нишу», а «по одёжке протягивают ножки». Формируясь
как вид, они приспосабливаются к ресурсам и условиям, но в отдельности от множества
других видов не в состоянии существенно повлиять на них. Я не хочу сказать,
что живые существа совершенно не влияют на среду своего обитания. Достаточно
вспомнить, что весь кислород в атмосфере Земли имеет биогенное происхождение,
а известняки, на которых формируется особый комплекс флоры и фауны, являются
чаще всего останками животных. Но применительно к единственному виду такая
возможность выгладит сомнительной. Исключение здесь составляет только человек.
Ну, а что жирафы иной раз вымирали – это не новость для палеонтологов. Некоторые
жирафы, столь же длинношеие и специализированные, как современная саванновая
жирафа, всё же вымерли. Это говорит о том, что в природе процесс «ожирафливания»
древних короткошеих «пражирафов» шёл не столь прямолинейно и гладко, как хочется
представить. Что именно было первичным – нарастание клапанов в шее, или удлинение
самой шеи – вопрос, который можно однозначно разрешить лишь путём анатомирования
натуральных экземпляров представителей «магистральной линии», ведущей к современной
жирафе, которые жили в разное время и были связаны отношениями преемственности.
Но они безвозвратно вымерли, и решение вопроса откладывается до постройки машины
времени.
Я лично могу предположить (ИМХО), что количественные изменения «на полголовы»
опережали качественные. Так часто бывает в эволюции: вначале появляются «грубо
сделанные» представители некоей группы живых организмов, а затем их сменяют
«улучшенные варианты». Достаточно рассмотреть раннюю эволюцию млекопитающих,
чтобы понять, о чём я говорю. Так что для первых длинношеих жирафов я предполагаю
существование вначале просто примитивных носителей специализации, «улучшенных»
ровно настолько, насколько дают им возможности особенности строения предков.
Это получаются этакие «полудлинношеие» формы. Они быстро достигают предела
физиологических возможностей, но одновременно идёт накопление разного рода
мутаций, позволяющих расширить пределы этих возможностей. Затем наступает качественное
улучшение – появляются формы, сумевшие разными путями преодолеть «застой».
А после этого следует новая волна количественных изменений – «опробование»
возможностей полученных признаков. Эти «волны», в принципе, могут накладываться
друг на друга. И никакого внешнего разумного фактора тут не требуется. Если
бы разум действительно оказывал стороннее давление на процесс эволюции, не
пришлось бы наблюдать ни превращения одних форм в другие «широким фронтом»
(от рыбы к земноводному, от тероморфа к млекопитающему, и т. д.), появления
и существования в течение миллионов лет групп, которые всё равно затем вымирают
(те же моллюски аммониты – с девона до мела!). Если разум всё же направляет
процесс эволюции, то делает он это как-то хаотично, как если бы нету его.
Вот за что стоит похвалить А. Москвитина – это за то, что он совершенно справедливо
признаёт, что у естественного отбора цель крайне неопределённая в плане конечного
результата. Всё то, что мы называем «видами» сейчас – это как бы срез непрерывного
процесса. Живи мы на миллион лет раньше или позже, мы бы назвали «видом» то
состояние неких представителей живых организмов, что существовало тогда. А
нынешнее состояние было бы относительно того воображаемого «вида» всего лишь
«подвидом», «расой», «переходной формой», «формой неясного таксономического
статуса». Естественный отбор действует не так, как человек. Человек задался
целью вывести собак, похожих на льва – и появились леонбергер и пекинес (ну
да, китайцы именно так льва и представляли, только не маленьким, конечно).
Захотели вывести золотых рыбок с «глазами дракона» - появились рыбки телескопы.
Порода совершенствуется – для этого есть стандарт с определёнными предписаниями
и бонитировка (оценка производителей на соответствие стандарту). А в природе
нет конечной цели – есть лишь цель сиюминутная. И процесс естественного отбора
действительно осуществляется без разумного начала, поскольку не может сохранить
мутации, которые могут стать (а могут в равной степени и не стать) полезными
когда-нибудь в Светлом Будущем. В природе надо выжить и достичь процветания
не через миллионы лет, когда (мамой клянусь!) будет намного лучше, чем сейчас,
а именно сейчас. Именно такая тактика приводит к появлению многочисленных «тупиковых
линий» и «боковых ветвей» на древе жизни. Уклонение от неспециализированной
линии – это, фактически, принесённое в жертву будущее эволюционирующей совокупности
во имя процветания в настоящем. Будущее не предопределено – поэтому неизвестно,
что будет сопутствовать линии живых организмов, которая начала эволюционировать
в том или ином направлении. Палеонтология говорит о том, что успешные эволюционные
ходы – это процесс, в котором было много участников, но мало выигравших. Переход
от рыб к четвероногим шёл не одним стволом – разные лопастепёрые рыбы параллельно
развивали сходные признаки. Пандерихтисы – одна из таких групп. От амфибий
к рептилиям тянется тоже не один ствол.

Синорнис - существо,
представляющее одну из ветвей "многоствольного" скачка от
рептилий к птицам в мезозое.
|
Полагают, что современные черепахи
развили рептильные признаки независимо от прочих рептилий: советский палеонтолог
Ивахненко выделяет их вместе с частью бывших «котилозавров» в класс парарептилий.
«Многоствольная» эволюция тероморфов в млекопитающих известна довольно широко,
а в последние годы появилось множество свидетельств того, что эволюция птиц
также шла «широким фронтом».
Следует отметить, что в той или иной степени описанная картина (изменение
условий обитания, "осознание" наличия новой экологической ниши и
целенаправленные мутации, приводящие к требуемому для занятия этой новой ниши
ароморфозу) характерна для любого случая, когда мы встречаемся с попытками
объяснить с позиций эволюционного учения процесс возникновения нового вида.
Именно вида, а не репродуктивно изолированной популяции.
А что же такое то самое «осознание» наличия новой экологической ниши? Это,
увы, лишь кажущийся процесс. Как сказал профессор Селезнёв в мультфильме «Тайна
третьей планеты», «только звери об этом не знают – они неграмотные». Процесс
«осознания» и последующего занятия «места под солнцем» - это результат успеха
в постоянной «толкотне локтями», испытании на прочность границ экологической
ниши соседних видов. Где есть отпор – туда не лезут, а где «нащупали слабину»
- там и начинается расширение границ собственных жизненных интересов за чужой
счёт. У людей всё аккурат то же самое, только осознанное и облечённое в «цивилизованную
форму» - например, «миротворческие» войска в других странах и авианосцы у чужих
берегов.
В отличие от кратковременной истории цивилизации, в природе есть ещё и изменение
условий среды, медленное и неотвратимое. Это дополнительный фактор, испытывающий
мутации на полезность. Изменения среды (в том числе появление новых видов в
прежней среде обитания) приводят к сдвигу «нормы» в строении представителей
вида. При наличии в достаточно большой популяции множества мутаций в ответ
на такое изменение всегда найдётся адекватная, и её носители становятся в НОВЫХ
УСЛОВИЯХ «нормой». А если нет адекватной мутации? Что ж, бывает и такое. Причём
бывает часто. Разрешением такой ситуации бывает самое элементарное вымирание
вида, которое случалось на Земле миллионы раз.
Что же мы имеем с гуся? С одной стороны барамины, возникшие благодаря Создателю,
с другой - сам Создатель, направляющий процесс эволюции. И это в теории эволюции
- фундаменте атеизма!
Слишком много мы с этого самого вашего «гуся» имеем. Если бы это была корова,
у неё бы уже мозги выдоили такими темпами. Мы «имеем» совсем не то, что хотелось
бы: совершенно искусственное, произвольно раздуваемое понятие «барамин», которое
придётся переосмысливать применительно к каждому конкретному случаю. Из-за
неконкретности и крайней субъективности это понятие не подходит для использования,
и может служить только как инструмент воздействия на не особенно искушённую
аудиторию. И второй вопрос: при чём тут боги? Где они в теории эволюции? Теория
эволюции никогда не пользовалась моррисианским понятием «барамин» ввиду его
искусственности и ориентации на сверхъестественное существо как на первопричину
появления такого понятия и движущую силу процесса исторического развития жизни.
И никакие божественные силы не берутся в расчёт при исследовании процесса эволюции.
Теория, не могущая обойтись без Создателя, но всячески это скрывающая, отрицающая
свое родство с религиозными воззрениями может быть названа только ересью в
изначальном смысле этого слова. Таким образом теория эволюции в той ее части,
которая описывает возникновение новых видов, а не репродуктивно изолированных
популяций есть еретическое религиозное учение.
Странная позиция у А. Москвитина. Сначала «пришил» к теории эволюции белыми
нитками какие-то искусственные «барамины», затем присовокупил не доказанную
и принципиально недоказуемую сущность, называемую «бог» («Создатель»), и теперь
пытается доказать, что оно тут к месту, и всегда было, только трусливо замалчивалось.
Не знаю, кто как, а я не увидел в его рассуждениях никакой надобности в существовании
богов. Если окинуть взором хотя бы более-менее изученные отрезки истории развития
живой природы, то окажется, что никаким участием разума, ни внутреннего, ни
внешнего, в этом процессе даже «не пахнет». Наблюдается процесс хаотического
«искания и испытания», появление большого количества недолго живущих групп
(иногда даже в высоком ранге типа – вспомните ископаемых животных, участников
«Кембрийского взрыва»), сменяемое качественным улучшением, а также вымирание
групп, существовавших иной раз дольше, чем некоторые из современных таксонов.
 Иллюстрация
с сайта Martin Jehle о палеоценовых
млекопитающих. Примеры ранней эволюции млекопитающих, представители очень недолго
существовавших вымерших отрядов, не оставивших потомков в нашем мире: гигантские
пантодонты Pantolambda и представитель тиллодонтов Ectoganus. В чём смысл "творения"
таких "пробных моделей"? Неужели бог действует методом "проб
и ошибок"?
Иллюстрация
с сайта Martin Jehle о палеоценовых
млекопитающих. Примеры ранней эволюции млекопитающих, представители очень недолго
существовавших вымерших отрядов, не оставивших потомков в нашем мире: гигантские
пантодонты Pantolambda и представитель тиллодонтов Ectoganus. В чём смысл "творения"
таких "пробных моделей"? Неужели бог действует методом "проб
и ошибок"?
Странно выглядит здесь словечко «ересь». В обывательском смысле оно может означать
просто чепуху, в более возвышенном – смелую идею, идущую вразрез с общепринятым.
Но здесь явно прослеживается религиозный подтекст.
В «Словаре живого великорусского
языка» Даля читаем: «Ересь ж. вообще, различие
в мнениях веры; раскол или отщепенчество, отступничество. … уродливые толки,
более или менее отвергающие сущность христианского
учения…» Всякая ветвь христианства считает «ересью» все остальные ветви.
Так, для православия все западные христианские течения – типичная ересь.
Так что
моррисианство, взращенное на западной неправославной почве – это как раз
самая настоящая ересь. И идущее оттуда понятие «барамин», которым оперирует
А. Москвитин – как
раз типично
еретическое. Где костёр? Или предпочитаем дыбу?
А как же насчёт теории эволюции? Очень просто. Она находится вне религии и
для объяснения процессов, происходящих в природе, не нуждается в богах. Поэтому
понятие
«ересь» в религиозном смысле этого слова к ней неприменимо.
Кажется, добрались до конца статьи. Итак, «три звоночка» прозвенели. Обычно
после этого начинается спектакль. Но начинать-то, на мой взгляд, просто нечего.
Окинем взглядом всё, что было написано ранее.
Первый звоночек: синхронизация двух процессов – изменения среды и изменения
анатомии. Анатомия меняется не независимо, а вслед за средой. Колебания, отклонения
от усреднённой «нормы» есть всегда. Но лишь изменения среды сдвигают понятие
«норма» в каком-либо направлении. Без изменений среды шея жирафа сама расти
не будет.
Второй звоночек: «изначальный» предок какой-либо группы живых организмов вряд
ли был «рафинированным» представителем этой группы. Он, несомненно, нёс основополагающие
черты, общие со своими будущими потомками, но также обладал рядом признаков,
которые были утрачены его потомками. Таким образом, путём простой «редукции»
специализированных признаков потомков «изначальный» вид (соответствующий библейскому
«барамину») не получишь. Редукция частных признаков приведёт лишь к тому, что
получающиеся таким способом «предковые формы» окажутся близкими друг к другу.
Фактически это демонстрируют ископаемые копытные кондилартры, среди которых
предки нескольких разных отрядов копытных (явных «бараминов») могут быть сгруппированы
с точки зрения систематики в два – три семейства, если не ближе.
Третий звоночек: мутационный процесс идёт в популяции, а не в «роду» какой-то
одной пары особей. И ничто не мешает носителям разных мутаций свободно скрещиваться
и передавать мутации новым поколениям (кроме 100%-ной летальности мутации,
разумеется, но и тут возможна передача в рецессивной форме).
Заключение
Я включаю телевизор и смотрю "В мире животных". В желтой жаркой Африке
бредут стада антилоп - и больших и маленьких. И деревья стоят и листвой они
покрыты снизу доверху. Хватает антилопам и большим и маленьким и даже средним.
Что-то случилось, листва вон вновь почему-то отросла, а шея у жирафов не уменьшилась…
А может быть эта листва и не исчезала? Зачем же тогда возникли жирафы? Я смотрю
и думаю: "Чудны дела твои, Великая Ересь под названием Теория Эволюции!"
Стоит, конечно, принять во внимание тот факт, что стволы
баобабов и акаций не очень-то и ветвисты, и съедобной для копытных листвы
на них не хватит, чтобы
прокормить даже самую необжористую антилопу. Стоит также подумать над тем,
что антилопа ситатунга живёт на болоте, куда не полезут ни тяжеловесная коровья
антилопа канна, ни куду, ни гну. А антилопа-прыгун с помощью маленьких твёрдых
копыт лазает по каменистым склонам, куда не запрыгнет ситатунга с её длинными
копытами. Одни антилопы объедают кусты, другие предпочитают траву. Одни живут
на болотах, другие в горах, третьи в сухой пустыне. Борьба за существование
не заканчивается. Просто в процессе эволюции виды «разбредаются» по своим
экологическим нишам, по своим многомерным пространствам. Они могут кормиться
рядом, но поедать
разные травы. Они могут есть примерно одинаковые травы, но жить в разных местах.
И лишь жирафы посматривают на антилопью круговерть у себя под ногами свысока:
с ними могут соперничать только слоны, и то не в полной мере.
Эволюция продолжается, господа присяжные заседатели!
Волков П. И.
г. Владимир, 2006 год.



