
Even a completely degraded environment can be successfully exploited by
certain species Ś but others are sure to perish.
62
FOUR
REUNITING GONDWANALAND

| |
Be fertile and multiply, fill the Earth and instill fear
and terror into all the animals of the Earth and birds of the sky.
Ś GOD, in a conversation with Noah |
In the now far-off decade of the 1960s a famous bumper sticker graced many
a Volkswagen bus: Reunite Gondwanaland. In those days the theory of plate
tectonics (also known as continental drift) was still in its infancy, and
the waggish slogan was a cry to bring back all of the southern continents
into the single continental landmass that existed at the end of the Paleozoic
era, some 250 million years ago. In a strange way, that call has been heeded:
Gondwanaland has been reunited. Not in any physical way Ś Africa is not measurably
closer to Australia or South America than it was thirty years ago. But functionally
they have been brought together as barriers to biotic exchange between them
have been eliminated. The common travel of boats and ships across the oceans
has shrunk those oceans by giving the animals and plants of the now separated
continents access to their age-old corridors of dispersal. When Gondwanaland
existed, it was a time of greater global homogeneity, of fewer ecological
niches, of fewer and lower mountain ranges, of more uniform global climate
Ś and, because of these factors, it was characterized by far lower planetary
biodiversity than the present era. The functional reuniting of Gondwanaland
may take us back to a lower global biodiversity reminiscent of that bygone
age. This renewed homogenization of the worldÆs biota may set the current
mass extinction
63
apart from all such previous events, for after this event there may not
be a subsequent diversity increase. The planet may well stay at the low levels
typical of a single landmass, rather than the higher diversity of numerous
separated continents.
Biological diversity is so commonplace to us that it is taken for granted.
Yet the factors leading to diversity are still great biological enigmas.
Since the Cambrian Period more than 500 million years ago, the diversity
of species on Earth has been fluctuating, but increasing overall. Will it
continue to do so? Here I will argue that the new mass extinction, which
is causing a dramatic decrease of diversity on Earth, will not be followed
by a renewed burst of diversification, or even a return to pre-extinction
diversity values until, perhaps, many millions of years have gone by.
Trends in Diversity
What controls the diversity of a given region? How can a coral reef be so
rich in life and a sand bed beside it so poor? And if we change scale, how
can a large region be species-rich and a neighboring province species-poor?
If we define diversity as the number of species present in any given area,
can we arrive at some rough mathematical rule governing diversity? There
are no simple answers to these questions. Many factors enter into the equations,
such as nutrient availability, habitat type, and amount of water; there are
also numerous factors affecting the formation of species, such as rates of
barrier formation, rates of genetic change, and, especially, rates of extinction.
Biologists have long recognized that diversity appears to be roughly related
to habitat size, and this makes good sense: the larger the area of habitat
available, the more animals and plants, and at the same time, the more different
kinds of animals and plants, can be accommodated. But is extinction rate
also related to habitat size, in some inverse way? Do larger habitats or
only larger population sizes protect individual species from extinction?
Some rough rules of thumb about this relationship were first formulated in
the 1960s by two famous ecologists, Robert MacArthur and E. O. Wilson, who
proposed a new theory relating diversity to habitat area. MacArthur and Wilson
called their idea the equilibrium theory of island biogeography. In essence,
it relates the area of habitat to the number of species present: as habitat
area increases, so too do species numbers, and they do so in a predictable
way. Similarly, as habitat area decreases, species numbers fall. Because
the number of species bears such a predictable relationship to the area available,
we can analyze the way in which defor-
64
estation, for example, leads to the shrinking of habitat and thus to the
loss of species. This influential model was one of the seminal theories about
the regulation of biodiversity through time. While it was originally designed
to examine diversity on islands, models patterned on the theory of island
biogeography have now been scaled upward to encompass continental and even
global scales of community and evolutionary diversity.
MacArthur and WilsonÆs equations can be used to predict rates of extinction.
They found, for example, that an island always has fewer species than a mainland
or continental habitat area of the same size, even if the habitats are otherwise
exactly identical. The implications of this finding are that parks and reserves,
which essentially become islands of habitat surrounded by disturbed areas,
will always suffer a loss of species. It also means that subdividing any
sort of larger habitat into smaller patches of disturbed and undisturbed
regions will increase the rate of extinction.
With these implications in mind, paleontologist Michael McKinney of the University
of Tennessee has recently summarized the general traits of global diversity:
The causes of change in diversity have been debated for over a century.
Generally, the proposed causes fall into two categories: abiotic factors
(those brought about by nonliving aspects of the environment, such as climate
change) and biotic
65
factors (those brought about by life itself, such as competition, predation,
and disease). Not surprisingly, ecologists have stressed the importance of
biotic factors, viewing the world in short time spans and at the limited
geographic scales of individual habitats and ecosystems. Those who examine
global biodiversity from the perspective of a larger and longer framework
(such as paleontologists) have long believed that abiotic changes are the
most important factors determining diversity. According to this view, the
two most important mechanisms regulating diversity are the rate of origination
of new species and the rate of species extinction. These two competing factors
are affected by abiotic factors and by each other.
In terms of the shorter ecological time scale, speciation is always a relatively
slow process, while extinction can be either fast or slow. In the present-day
Age of Humanity, it appears that the large-scale environmental changes causing
the observed rise in extinction are abiotic Ś climate change and changes
in landscape and vegetation Ś yet their ultimate cause is biotic Ś the actions
of humans. These circumstances have no precedent on Earth.
Our understanding of the rate of diversification relies on the concept of
niche saturation. For many decades ecologists have used the concept of a
niche to describe how a particular species lives and interacts in its ecosystem.
The niche is somewhat analogous to the profession of a species: what it eats,
where it lives, what it does in its community. As more and more species either
evolve in or invade a given community with finite energy resources, more
and more of the available niches are filled. It may be that the overall carrying
capacity of a given habitat, community, continent, or even the Earth, limits
the number of available niches, and that these niches can become saturated
with species, thus limiting the potential for new speciation. Human activities
appear to be reducing the number of niches available, at least in terrestrial
habitats. The replacement of a forest with a field, or a field with a city,
reduces niche availability. Suddenly the world is a less heterogeneous place
Ś just as it was during the time of Gondwanaland, 200-300 million years ago.
Disturbance and Diversity
Since the actual number of species on Earth today is so important, knowing
what controls that number is also important. Why are there not twice as many,
or half as many, plants and animals? Why are there more now than during the
time of Gondwanaland? Although there is an enormous scientific literature
on diversity, this is a question that has perplexed biologists for nearly
two centuries, and it appears that
66
there is no easy answer. The most famous book on the topic Ś Charles DarwinÆs
On the Origin of Species Ś does not even address the issue. Darwin was concerned
with the transformation of individuals, rather than how and why new species
form. Most of the more recent treatises on diversity, such as Yale ecologist
Evelyn HutchisonÆs famous paper ōHomage to Santa Rosalia, or Why Are There
So Many Kinds of Animals?ö do not examine the mechanisms leading to the origin
of species, but simply describe the maintenance of species once they have
evolved.
Nevertheless, this problem was reexamined recently in a thoughtful essay
by paleontologists Warren Allmon, Paul Morris, and Michael McKinney, who
attacked it in a different way. They asked how short-term environmental changes,
or perturbations, as well as more severe and longer-lasting changes, which
they called disturbances, affect evolution and diversity. Because humans
are producing both perturbations and disturbances in copious quantities around
the globe, this particular question is highly relevant to understanding and
predicting possible future trends in diversity.
All organisms encounter perturbations in their daily lives. Fluctuations
in temperature, food availability, rates of predatory attacks Ś these and
a thousand other environmental changes are part of the everyday lives of
all organisms. Sometimes, however, one or several of these changes are severe
enough to kill off or otherwise remove a species or group of species from
a given geographic area, creating a patch in space from which these organisms
are now absent. Of course, what constitutes a perturbation or disturbance
varies from species to species Ś a disturbance for a protozoan may not even
be noticeable to a fish. Disturbances are thus species-specific. They can
also be thought of as acting at many environmental scales, as well as many
scales of time. Perhaps the most interesting for our purposes are time scales
ranging from a thousand to a hundred thousand years Ś the intervals of time
necessary for the speciation of large animals and plants.
Ecologists have long understood that there is a relationship between the
degree of disturbance and the ability of nature to maintain diversity. Many
studies of marine intertidal zones have shown that in areas of either too
little or too much disturbance, few species occur. The disturbances can be
both abiotic Ś such as a violent storm Ś and biotic Ś such as the incursion
of a new predator. Both types of disturbance create patches of open space
or habitat. By reducing the numbers of abundant species, they allow rare
species to maintain their existence or allow new species to gain a foothold
in the environment. In environments with little disturbance diversity drops
as a few species outcompete all the others and dominate the
67
environment. In high-disturbance areas, diversity also stays low, since
only a few species can maintain viable populations in the face of high mortality.
Maximum diversity is found in areas that can be considered to have intermediate
levels of disturbance. Such conditions allow many species to survive, but
do not allow any particular species to take over through predation or competition.
On the other hand, there have been virtually no studies trying to link disturbance
with speciation, or the creation of diversity. Allmon and his colleagues
have suggested that, like the maintenance of diversity, the creation of diversity
through the formation of new species may occur in regions of intermediate
disturbance. Paleontologist Steven Stanley has postulated a similar model,
noting that ōhigh rates of speciation are actually promoted by less severe
environmental deterioration Ś deterioration severe enough to elevate extinction
rates to a moderate degree but not so severe as to cause wholesale extinctionö.
This idea has some interesting implications. It predicts that endemic species
Ś those restricted to specific and hence small geographic regions Ś will
encounter relatively higher levels of disturbance than more broadly adapted
species, and therefore experience both higher levels of origination and extinction.
These species Ś the specialists and types found in restricted ranges Ś are
those that produce the largest number of new species. Yet they also have
the highest extinction rates.
Global diversification remains a simple equation: origination minus extinction.
The highest net rates of diversification seem to occur in animals with small
body sizes, short generation times, wide distributions, and high abundances
Ś beetles and rodents, for example. Although two of these traits Ś wide distribution
and high abundance Ś seem to negate new speciation, they retard extinction
even more. The net result is higher diversification than extinction.
Compounded Disturbance and Ecological Surprise
All species have evolved in the presence of disturbance. Thus, disturbances
that happen within a particular range of intensity Ś not too extreme Ś result
in little long-term change in the nature, composition, and energy flow of
a population, or even an ecosystem. But what of ōcompoundedö disturbances,
when major disturbances occur repeatedly at higher than normal frequencies?
This was the question posed by ecologist Robert Paine and his colleagues
in a 1998 article. Paine has spent his entire research career studying intertidal
organisms and has contributed fundamental discoveries about the architecture
of ecosystems and diversity. According to Paine, disturbances, ranging from
small-scale and
68

Deforestation and fragmentation are the future Ś and bane Ś of post-industrial
ecosystems.
69
frequent perturbations to large and infrequent catastrophes, occur from
time to time in any habitat. It is these cycles of disturbance that led to
ecologyÆs first paradigm, that of ecological succession. Disturbances often
cause widespread mortality, leaving a residual assemblage of flora and fauna,
which provides a legacy that subsequent processes and populations use to
rebuild. Even large, infrequent disturbances do not appear to override the
biotic mechanisms that structure the eventual recovery. Paine and his colleagues
used the example of the catastrophic 1988 Yellowstone National Park fire,
which burned nearly 40% of the park and was an order of magnitude larger
than previous fires in the park region. Even a decade after this major event,
there have been no ecological ōsurprisesö; the ecosystems returning are similar
to those present prior to the fire. But what if the park underwent another
such fire ten years after the first, and then another a year after that?
If such major catastrophes are compounded, will ecosystems return to their
previous state? Paine and his colleagues argue that they will not.
Compounded disturbance can be portrayed in two ways. First, it can occur
in the manner proposed in the Yellowstone fire example, in which a normal
community undergoes a second (or multiple) disturbance before recovery from
the first is completed. Second, a major stress can be superimposed on a community
altered by some significant disturbance. Examples of the this second type
of compound disturbance can be seen when fish stocks are depleted by overfishing
and then subjected to some other type of large-scale disturbance. In such
a case their recovery is markedly delayed, if it occurs at all. Climate change
may produce the same effect: a series of major storms one after another may
markedly alter ecosystems that have evolved under lower storm frequency regimes.
Paine and his colleagues note that the prime cause of compounded disturbance
is human activity. The prime result is lowered diversity Ś a return to Gondwanaland.
Plate Tectonics and Diversity
The studies above (and many others as well) suggest that compounded disturbance
produced by humanity may have caused the equilibrium level of world biodiversity
to drop. Yet there is a second and equally important factor that is taking
us back to Gondwanaland: the functional removal of barriers to migration.
In a way, to borrow from another hoary bumper sticker, we have indeed stopped
continental drift.
One of the major influences on the equilibrium value of global biodiversity
is continental configuration. When the various continents were united, there
was obviously easy faunal interchange around the globe. When the continents
are
70
widely separated, however (as they are today), there is greater heterogeneity
in environments, less faunal interchange, and many more species. Two hundred
fifty million years ago, all the major continents were merged, and biodiversity
was far lower than it is today. But by introducing non-native species across
ecological boundaries and continents, humanity has found a way to functionally
reunite the various continents, as least as far as gene flow is concerned.
Since the majority of the EarthÆs biodiversity today is found on continents
(and there is no reason to believe that this relationship has changed over
the last 300 million years), the processes of plate tectonics are especially
important for life and its ecosystems. As continents have shifted their positions
through time, they have affected global climate, including the overall albedo
(the planetÆs reflectivity to sunlight), the occurrence of glaciation events,
the pattern of oceanic circulation, and the amounts of nutrients reaching
the sea. All of these factors have biological consequences that affect global
biodiversity. Moreover, continental drift can help augment diversity by increasing
the number and degree of separation of habitats (which promotes speciation).
Plate tectonics also promotes environmental complexity Ś and thus increased
biotic diversity Ś on a global scale. A world with mountainous continents,
oceans, and myriad islands is far more complex, and offers more evolutionary
challenges, than a planet dominated by either land or ocean. James Valentine
and Eldredge Moores first pointed out this relationship in a series of classic
papers written in the 1970s. They showed that changing the positions and
configuration of the continents and oceans would have far-reaching effects
on organisms, causing increases in both diversification and extinction. Changes
in continental position would affect ocean currents, temperatures, seasonal
rainfall patterns and fluctuations, the distribution of nutrients, and patterns
of biological productivity. Such changing conditions would cause organisms
to migrate out of the new environments, and would promote speciation. The
deep sea would be affected least by such changes, but the deep sea is the
area on Earth today with the fewest species: over two-thirds of all animal
species live on land, and the majority of marine species live in the shallow-water
regions that would be most affected by plate tectonic movements.
The most diverse marine faunas on Earth today are found in the tropics, where
communities are packed with vast numbers of highly specialized species. Not
only are there fewer species at higher latitudes, but species composition
is different from that in the tropics as well. Most species have fairly narrow
temperature limits imposed by physiological adaptation, and since temperature
conditions change rapidly with latitude, itÆs not surprising that the north-south
coastlines of continents show a continuously changing mix of species.
71
In 1996, biologist P. Vitousek and three colleagues used mathematical modeling
to project the number of mammalian species that would be expected on Earth
if all of the continents were reunited into the configuration present at
the end of the Paleozoic Era, some 250 million years ago. They concluded
that the world would contain about half of the nearly 4,000 mammalian species
present if we reunited Gondwanaland. These same authors speculated that
the current transport of mammals from continent to continent is leading
to an extinction rate of mammals that will yield approximately this same
global biodiversity: 2,000 mammalian species.
Aliens Among Us
Whenever a species arrives in an area where it was not previously found,
there is a potential for biological change. Such invasions of new species
have occurred throughout geologic time, yet the rate of invasion has vastly
increased during the Age of Humanity. Today, no area on Earth is immune to
such biological invasions. It is estimated that about 11% of all species
now living in France have been introduced; in Australia the proportion is
10%, in Hawaii 18%, and in New Zealand more than 40%. These biological invasions
are particularly marked in floral communities. There are records of about
1,200 native plant species in New Zealand, but there are now over 1,700 non-native
plants living there as well. Although it could be argued that the introduction
of so many non-native plant species has more than doubled the diversity of
plant life in New Zealand, this is only a transitory result. Over time, many
non-natives will inevitably drive the natives to extinction, causing world
biodiversity to decline.
Biological invasions aided by humans have come in three major pulses. Over
a period starting several thousand years ago until about 1500 A.D., human
movement and migration caused the transport of plant and animals mainly in
the Old World. Beginning in about 1500, however, a second phase of invasions
began with the increasing contact between the Old World and the New due to
European exploration and conquest, during which many Old World species were
transported to the New World. The final phase began about 150 years ago with
the globalization of species movement due to the vastly increased efficiency
of human transport.
There have been many reasons for these species introductions. In some cases
the introduced species were purposely brought to a new location to become
animal or plant crops. Some were brought to serve as ornamentals or pets,
while others were introduced for sport or hunting. Still others were introduced
to control
72
ōpestsö, only to have the introduced species become even more destructive
than the species they were brought in to control. Ironically, some introductions
have occurred for purposes of either biotechnology or scientific research.
Finally, there have been many accidental introductions from ship ballast
and airplane holds, of ōhitchhikingö seeds escaped from either wild or agricultural
areas, and simply as side effects of habitat alteration.
The majority of introduced species do not survive. It is estimated that of
a hundred introduced species, approximately ten will successfully colonize
or naturalize, and only two or three will become pests. But those that do
often become major problems, especially in fragile, endangered, or rare ecosystems,
such as early successional habitats, ecosystems with few species, and ecosystems
that traditionally have a low number of predators or grazers. The pest invaders
often show a suite of similar characteristics: they have a high reproductive
potential, many offspring, and generalized habits and food requirements.
They can thus be characterized as ōpioneerö species, in that they can colonize
and flourish in a wide variety of ecosystems. They are often human ōcommensalsö
Ś species that thrive in the presence of humanity.
While the greatest consequences of these invasions are biological, their
economic impact is not trivial. In the United States alone, it is estimated
that the Russian wheat aphid causes as much as $130 million in crop damage
each year, the Mediterranean fruit fly as much as $900 million, and the gypsy
moth about $750 million. The boll weevil may have caused as much as $50 billion
in damage to cotton crops during the twentieth century.
The ultimate effect of many invasions is extinction of native species, and
examples of such extinctions abound. In 1959, in the Rift Valley of Africa,
British colonials introduced a northern African fish called the Nile perch
into Lake Victoria for sport fishing. The Nile perch is a voracious predator
on smaller fishes. Prior to its introduction, the lake was home to over 300
species of endemic cichlid fishes. Yet by the early 1980s, when the problem
was finally recognized, over half of the cichlid species in Lake Victoria
had gone extinct because of the Nile perch.
Of all the factors causing the translocation of species, the exchange of
ballast water may be among the most important and the most difficult to stop.
Thousands of species are transported around the globe in shipsÆ ballast water.
When a ship takes on ballast water, it takes up the plankton of a given region,
which often contains the juvenile stages of marine animals and plants. These
organisms are
73

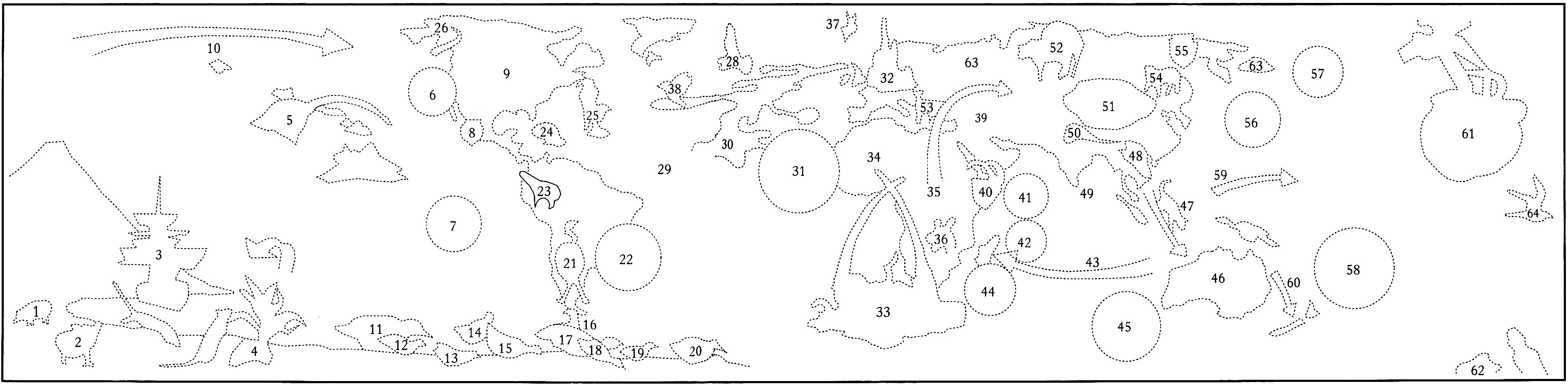
|
1. Feral pig
2. Norway rats
3. European exploration
of remote oceanic
islands
4. Coconut palm
5. Hawaiian honeycreeper
and mosquito
6. California condor
7. Easter Island
8. Mexican grizzly bear
9. North America |
10. Human migration
through Beringia
11. Barredwing rail
12. Aukland Island slate-
breasted rail
13. Laysan rail
14. Ponape crake
15. Samoan wood rail
16. Chatham Island banded rail
17. Lord Howe wood rail
18. Wake Island rail
19. Hawaiian rail |
20. Tahitian rail
21. Rhea
22. Rio de Janeiro
23. Tapir
24. Haitian solenodon
25. Piping plover
26. Alaskan pipeline
27. House sparrow
28. Feral pigeon
29. Caiman hunting
30. North Sea oil industry
31. Gorilla |
32. EuroDisney
33. Big game hunting
34. The Sahara
35. Origin of modern human
36. Quagga
37. Research whaling
38. Eurasian starling
39. Ostrich
40. Walia ibix
42. Brookesia peyrieresi
43. Human migration to
Madagascar
44. Dodo and dodo tree |
45. Thylacine
46. Feral rabbits
47. Human migration brings dingo to
Australia
48. Giant land snail
49. Ceylon elephant
50. Snow leopard
51. Traditional Chinese medicine
(bear gall bladder, tiger paw, and
rhinoceros
horn)
52. Eurasian bison
53. Cretan deer
54. Tiger |
55. Kamchatkan bear
56. Shistosoma
57. Hunting woolly mammoth
58. Stephen Island wren
59. Human migration from Asia into the Pacific
60. Human migration to New Zealand
61. Brown tree snake
62. Ocean-borne garbage
63. StellarÆs seacow
64. Sandwich tern |
A recent history of the world, from an ecological perspective.
then discharged when the ship reaches its destination port. One such invader
was the infamous zebra mussel, which made its way into the Great Lakes of
North America. The zebra mussel, which originated in Europe, is an extremely
efficient filter-feeding organism, straining plankton from the surrounding
water so efficiently that it outcompetes native species, which then starve
to death. It multiplies rapidly, attaching itself to pipes, boats, and the
shells of other mollusks. Zebra mussels clog water intake pipes, thus affecting
public water supplies, irrigation, sewage treatment plants, shipping lanes,
and recreational activities.
Governments around the world are trying to monitor the alien species arriving
in ballast water. A recent study conducted on Japanese ships entering Oregon
ports discovered the presence of over 350 alien species being discharged
into Oregon waters. Among the most undesirable of such invaders are predatory
crabs, which are capable of wreaking havoc on shellfish beds. Such an invasion
began in the 1990s with the appearance of the green crab in Washington State.
The green crab feeds on small clams, and is capable of decimating local populations
of clams and snails.
Plants also suffer a great deal from biological invasions. Because plant
seeds are usually small, they are easily transported long distances, and
often can colonize and take over new ecosystems quickly. In different areas
of the United States, introduced plants make up between 7% and 48% of the
total plant diversity. Many of these non-natives, such as kudzu, were deliberately
introduced to control soil erosion. Others were introduced as agricultural
crops. On rangelands, invasive plants such as cheatgrass crowd out more nutritious
native plants, cause soil erosion, and pose threats to native wildlife.
Even more deleterious than these plant invasions has been the transport of
plant pathogens from one part of the world to another. Dutch elm disease
decimated elm trees in both England and the United States after it was accidentally
imported. The introduction of the chestnut blight from Asia to America in
1890 drove the American chestnut tree to the verge of extinction in less
than fifty years. In Australia, native Jarrah forests have been destroyed
due to the introduction of a root fungus imported from Eastern Australia.
Winners and Losers
Predicting winners and losers in the future can be as perilous for biologists
as it is for stockbrokers. In both cases, however, there are some clear signs
of what may prosper (and even diversify) and what may die out.
76
One clear insight into predicting whether a species will flourish or not
comes from the size of its geographic range. In the late 1980s, biologists
J. Brown and M. Maurer showed that species of North American birds with small
geographic ranges almost always had low population densities within those
ranges. In other words, there are virtually no species of birds in North
America that are both narrowly distributed and abundant within their small
geographic ranges. On the other hand, birds with widespread geographic ranges
are usually abundant in most regions within their ranges. Although we see
this all around us, it is not an obviously intuitive finding; but it is a
generalization that has the utmost importance for picking winners as well
as losers among species in the coming years.
The correlation between range size and abundance can be understood by looking
at the geometry of species ranges. Geographic ranges exist because they encapsulate
all the areas where a species can exist. Thus the outer limits of a geographic
range tend to be less favorable areas for the species than the interior of
the range. If the size of the less favorable perimeter is large relative
to total area of the range, it becomes limiting to the overall population,
and small geographic ranges have higher perimeter-to-area ratios than large
ones. Not surprisingly, then, when a large geographic range is suddenly broken
up into many smaller ranges, the abundance of the species will drop. Geographic
range fragmentation can thus influence the likelihood of extinction by affecting
the rate of extinction of local populations that find themselves confined
to ōhabitat islandsö.
This correlation between range size and abundance is the single greatest
nightmare of conservation planners. The urbanization and ōagriculturalizationö
of the world has been fragmenting the ranges of most wild species while greatly
expanding the geographic ranges of agricultural species. This effect will
in essence spell doom for a majority of the worldÆs rare species, many of
which are ōmegamammalsö. Once again, the conclusion of the Age of Megamammals
is the functional reuniting of Gondwanaland.
77
CONTENTS
IMAGES
|
|
vii |
FOREWORD
|
Biological Futures
Niles Eldredge |
ix |
| PREFACE |
|
xiii |
| INTRODUCTION |
The Chronic Argonauts |
1 |
| ONE |
The Deep Past: A Tale of Two Extinctions
|
13 |
| TWO |
The Near Past: The Beginning of the End of the Age of Megamammals |
37 |
| THREE |
Into the Present |
47 |
| FOUR |
Reuniting Gondwanaland
|
63 |
| FIVE |
The Near Future: A New World
|
79 |
| SIX |
The First Ten Million Years: The Recovery Fauna
|
103 |
| SEVEN |
After the Recovery: A New Age?
|
119 |
| EIGHT |
The Future Evolution of Humans
|
139 |
| NINE |
Scenarios of Human Extinction: Will There Be an ōAfter Manö?
|
155 |
| TEN |
Deep Time, Far Future
|
169 |
| BIBLIOGRAPHY |
|
177 |
| INDEX |
|
183 |