
| Главная | Библиотека | Форум | Гостевая книга |
|
Глава
10
«Акустические
глаза»
дельфинов
Впервые явление ориентации по звуку было обнаружено у летучих мышей итальянцем
Лаццаро Спалланцани в 1793 году, а эхолокация у китообразных открыта лишь
в пятидесятых годах XX века.
Насколько точен эхолокационный аппарат дельфинов и какова роль эхолокации
в жизни китообразных? По эхолокационным сигналам дельфины могут с большой
точностью ориентироваться в пространстве, плавать с наглазниками, определять
глубину дна и близость берега, разный характер грунта, величину и качество
погруженных предметов, находить вещи в мутной воде. Действие гидролокатора
у дельфинов хорошо изучено в океанариумах.
На каком расстоянии дельфины могут обнаруживать пищу с помощью своего гидролокатора?
Ответ получили В. Шевилл и Б. Лоренс в следующем опыте: от лодки перпендикулярно
ее борту протягивали разделительную сеть длиной в несколько метров, и рыбу
бесшумно опускали в воду в условиях темноты то с носа, то с кормы. Подопытный
самец должен был заблаговременно решать, по какую сторону сети ему плыть,
чтобы найти ожидаемую рыбу. Эту задачу он правильно решал с расстояния до
4,5 м. Дельфин оказался способным не только обнаруживать отдельные эхо своих
скрипов от рыбки длиной 15 см, но мог распознавать именно это эхо среди прочих
звуков, отраженных от берега, дна, камней, лодки и других предметов в пруду.
Точность исследования Шевилла и Лоренс была подтверждена специальной проверкой
физиком Джоном Скоттом на афалине Сальти.
Профессор Флоридского университета психолог Уинтроп Келлог с сотрудниками
в морской лаборатории близ гавани Аллигатор несколько лет изучал эхолокацию
афалин. Опыты он проводил над двумя обученными дельфинами Альбертом и Бетти
в бассейне площадью 350 м2 и глубиной до 2 м. Илистое дно и стенки
бассейна хорошо поглощали звуки, не давая эха. Дельфины, плавая, взмучивали
воду так,
что видимость во время опыта не превышала 35 — 85 см. Чтобы дельфины не могли
видеть действий человека ни в воде, ни в воздухе, эксперименты проводились
ночью. Звуки их записывали специальной аппаратурой. Предметы погружали в
воду за фанерным экраном. Если в водоеме было спокойно, дельфины лишь изредка
издавали скрипы. При холостом шуме (например, когда капали жидкость на поверхность
воды), животные издавали короткий скрип и замолкали. Если в воду бросали
погружающийся несъедобный предмет, то вслед за первым издавалось еще несколько
скрипов с промежутками в 1 — 2 секунды. Но когда кидали рыбу, следовал целый
залп скрипов, частота импульсов возрастала до нескольких сотен в секунду,
и дельфин безошибочно направлялся к цели. При этом он покачивал головой из
стороны в сторону на 10°, как бы прицеливаясь на объект посредством отраженного
звука, и увеличивав частоту излучаемых щелчков по мере приближения к рыбе.
Темнота не мешала ему различать рыбу по размерам. Крупной кефали (30 см)
Альберт и Бетти явно предпочитали вдвое меньшего пятнистого горбыля. Если
погружали обеих рыб вместе, афалины устремлялись к предпочитаемой рыбе: в
первых 16 испытаниях самец ошибся только 4 раза, а в 140 последующих — ни
разу! Когда горбыля подвешивали позади стеклянного экрана (он был виден,
но недоступен для эхолокации), а кефаль — впереди экрана (она была доступна
для ультразвукового распознавания), то дельфин не пытался ловить горбыля
и довольствовался лишь кефалью.
В последующих опытах в Калифорнийском океанариуме экспериментаторы стали
«ослеплять» подопытных дельфинов, надевая им наглазники (рис. 39), и обучили
их принимать пищу по сигналу. Оказалось, что дельфины с выключенным зрением
могут свободно плавать в лабиринте из подвешенных металлических стержней,
не касаясь их, и точно различать заграждения из тонкой проволоки и нейлоновых
нитей. Такие способности помогают дельфинам тонко отличать разные виды кормовых
объектов (рыб, моллюсков, рачков) и выбирать предпочитаемую пищу в условиях
любой освещенности среды, безопасно плавать в щелях между выступами дна.
В бассейне лаборатории Пойнт-Магу Кеннет Норрис и физиолог Рональд Тернер
обучили взрослую самку-афалину Элис с надетыми наглазниками распознавать
два металлических шарика размером 3,7 и 6,2 см. Шарики располагались в 60
— 120 см от животного и могли взаимно переставляться. Если самка выбирала
больший шарик (6,2 см), она получала в награду рыбу. Затем меньший шарик
стали заменять все более и более крупным, уменьшая разницу с большим шариком.
Когда меньший шарик стал диаметром 5 см, дельфин продолжал безошибочно выбирать
больший шарик (6,2 см). А когда разница в диаметрах шариков уменьшилась до
0,8 см, афалина все же в большинстве случаев делала правильный выбор. Этот
опыт продемонстрировал поразительную точность распознавания величины лоцируемых
объектов. В других опытах К. Норриса и его коллег «ослепленная» афалина с
расстояния 6 — 7 м отличала капсулу из желатина от куска рыбы такого же размера.
Акустические лучи, проникая внутрь предметов, несут в виде эха информацию
о размере, форме и материале этих предметов.
Советские исследователи Н. А. Дубровский, А. А. Титов, В. П. Бабкин и другие,
пользуясь той же методикой разделительной сети, показали, что афалина в темноте
надежно различает шары диаметром 5 см, сделанные из акустически жестких и
мягких материалов, с расстояния 11 м.
Эхолокационные импульсы дельфинов человек слышит как разного рода скрипы.
Эти сигналы животные посылают направленно с помощью координированной работы
воздушных мешков, носового канала, лобно-носовой (жировой) подушки и сложной
системы мышц, расположенных в выпуклой части головы. Лобно-носовая подушка,
лежащая на челюстных и межчелюстных костях, и вогнутая передняя поверхность
черепа действуют как звуковая линза и рефлектор: они концентрируют (фокусируют)
сигналы, излученные гидролокатором, и в виде звукового пучка направляют на
лоцируемый объект. Доказательства действия такого ультразвукового прожектора
получены с помощью экспериментов как за рубежом (В. Эванс, Д. Прескотт, В.
Сатерланд, Р. Бейл, 1964), так и у нас (Е. В. Романенко, А. Г. Томилин и
Б. А. Артеменко, 1963 — 1965).
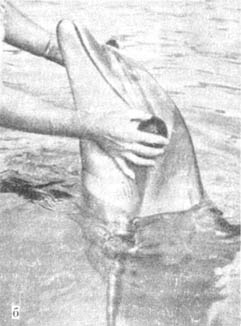
Рис. 39.
 |
 |
 |
|
Афалине закрывают глаза латексовыми наглазниками (а, б). «Ослепленное» животное свободно плавает между подвешенными трубами, не касаясь их (в), а, б — фото сотрудников океанариума «Морская студия»; в — фото Рихарда Хьюэтта.
Особенно высока разрешающая способность к локации у дельфинов в горизонтальном
плане и хуже — в вертикальном. Они могут, находясь на поверхности воды, лоцировать
объекты и на глубине. Это для них очень важно: ведь им приходится постоянно
то выныривать (чтобы дышать), то погружаться вглубь. Но куски рыбы, находящиеся
под нижней челюстью, они лоцировать не могут. Может быть, потому, что нижняя
челюсть служит экраном, отражающим звуковые волны, издаваемые системой воздушных
мешков. Возможно, в этом имеет значение и то, что лобно-носовая подушка расположена
наклонно к продольной оси черепа.
Американские исследователи Эванс и Пауэлл в 1967 году доказали способность
ослепленного дельфина различать как одинаковые по площади пластины — медную,
алюминиевую и латунную, так и разные по толщине (2,2 и 3,2 мм) медные пластины.
Советский исследователь А. Е. Резников экспериментально показал, что верхний
порог слуха афалины достигает 180 кгц, и это животное в условиях опыта различает
одинаковые по внешнему виду стеклянные и пенопластовые предметы. В опытах
В. М. Бельковича и его сотрудников дельфины отличали пенопласт от эбонита,
алюминий от плексиглаза, а в опытах А. П. Абрамова — латунь от текстолита,
фторопласта и стали.
Оказалось, что дельфины могут различать геометрические фигуры, помещенные
внутри холщевых мешочков (А. Г. Голубков, Ю. В. Иваненко), или даже различные
положения поршня внутри металлического цилиндра. Этого нельзя достичь с помощью
зрения, так как свет отражается от поверхности предметов; ультразвуковые
же лучи с их большей длиной волны по сравнению с оптическими проникают внутрь
предметов. Это позволяет дельфинам с помощью акустических волн исследовать
не только размер и объем, но и качество (материал) предметов и отлично «видеть»
ушами при выключенном зрении.
За последние годы появились интересные, внешне привлекательные, гипотезы
о действии приемной системы дельфина по голографическому принципу (Джон Дреер),
либо по принципу звуковидения (Л. Е. Резников, В. М. Белькович, Б. Г. Хоменко,
Г. Б. Агарков, Б. В. Солуха и др.). При разработке этих гипотез обращалось
внимание на структуры надчерепной части головы дельфина — на лобно-жировую
подушку, способную фокусировать звуки, на характер взаимного расположения
воздушных мешков, жировой подушки и черепа, на гистологию эпителия воздушных
мешков. Эти гипотезы исходят из признания, что в звуковидении дельфинов имеют
значение кожные рецепторы на голове и в надчерепных тканях, якобы способные
воспринимать акустическую информацию в воде.
Американский исследователь Д. Дреер в 1969 году предположил, что дельфины
при звуковидении используют «голографические изображения», которые как бы
отпечатываются на коже лобно-носовой подушки головы.
Москвич А. Е. Резников придает особое значение внутренней поверхности воздушных
мешков, представляющей собой пигментированный кожный эпителий и играющей
якобы роль сетчатки «акустических глаз». Он считает, что «пространственные
акустические изображения лоцируемых объектов», возникающие в теле жировой
подушки, могут восприниматься как образы «пространственного осязания». По
мнению этого исследователя, звуковидение дельфина столь же острое, как и
разглядывание окружающих предметов с помощью зрения у людей.
Ученые Г. Б. Агарков, Б. В. Солуха и Б. Г. Хоменко связывают ориентацию дельфинов
в окружающей обстановке также с голографическим механизмом приема эхо-сигналов
и получения объемного представления об окружающем пространстве на основе
отраженных звуков. Они полагают, что отраженный акустический сигнал воспринимается
многочисленными механорецепторами, размещенными в коже и мягких тканях головы,
особенно в лобном выступе. Каждый механорецептор отдельным нервным каналом
связан с мозгом, где и создается образ на базе звуковой информации.
Во всех выше изложенных гипотезах оказались уязвимые места. Их обнаружил
крупный советский биоакустик Н. А. Дубровский. Он считает, что в этих концепциях
не уделено должного внимания следующим акустическим и нейрофизиологическим
аспектам:
1. Чтобы приемная система, работающая по принципу звуковидения и голографии,
могла функционировать столь же эффективно, как слух и зрение, необходимо
чтобы число рецепторных элементов достигало величины порядка 105 или более
(30 тысяч внутренних и 20 тысяч внешних волосковых клеток в улитке млекопитающих).
Между тем нейрогистологические исследования показали, что плотность и общее
число рецепторных окончаний в структурах надчерепной части головы дельфина,
в том числе вблизи воздушных мешков, достигает лишь 102 на 1 см2, т. е. близко
к числу, характерному для кожи человека (например, в области предплечья).
Известно, что кожный анализатор имеет крайне ограниченный частотный (не выше
200 — 300 гц) и динамический диапазон. При такой плотности рецепторных элементов
можно воспринимать лишь самую грубую (интегральную) картину сфокусированного
звукового поля.
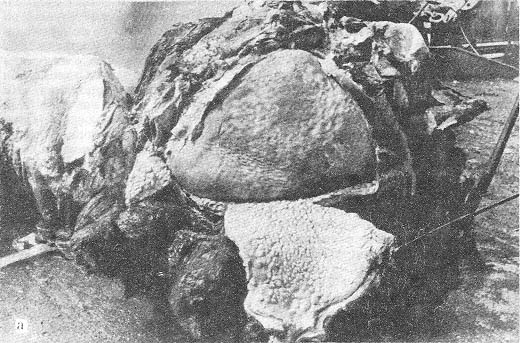
Рис. 40.
 |
 |
Акустический глаз кашалота: а — пузырчатая ткань, выполняющая роль сетчатки; б — верхний спермацетовый мешок-звукопровод, проводящий акустические лучи на пузырчатую ткань-сетчатку. Фото В. А. Козака.
2. В ходе эволюции органов чувств (зрения, слуха) происходила
их постепенная защита от грубого непосредственного внешнего воздействия.
Например, улитка
органа слуха млекопитающих оказалась спрятанной в толще черепа, а сетчатка
глаза — на дне глазного яблока в углублении костных глазниц. В условиях,
такой защиты органов чувств и развились очень чувствительные сенсорные системы.
Авторы же обсуждаемых гипотез поместили воспринимающие структуры приблизительно
в те же самые места, где происходит излучение эхолокационных импульсов дельфина,
звуковое давление в которых может достигать огромных значений — до 4 • 104 дин/см2.
Какой невероятной скоростью релаксации должны обладать рецепторные и нейронные
элементы, чтобы, спустя считанные миллисекунды после воздействия
мощного импульса они были в состоянии воспринимать тончайшее эхо с ничтожно
малым звуковым давлением (сотые и даже тысячные доли бара, т. е. в миллион
раз меньшие сигналы). В природе неизвестны случаи пространственного совмещения
органа звукоизлучения и звукоприема. Напротив, следует ожидать у дельфинов
существования специальных дополнительных механизмов защиты слуховой системы
от воздействия собственных зондирующих сигналов.
3. Детальность звукового изображения зависит от отношения длины волны звукового
сигнала к размеру входной аппертуры, в данном случае к диаметру звуковой
линзы. Так как максимум спектра эхолокационного сигнала дельфина приходится
на 75 кгц (длина волны 2 см), а диаметр линзы не превышает 10 см, то отношение
этих величин составляет 0,2. В этих условиях звуковое изображение, если бы
оно существовало, было бы весьма грубым и не позволило бы дельфинам иметь
высокую разрешающую способность.
4. Трудно себе представить способность рецепторов надчерепных структур дифференцировать
частоты звуковых сигналов. В органе слуха для этой цели служит специальная
система — улитка с базилярной и текториальной мембраной и органом Корти.
Непонятно, как могло бы происходить различие частоты звука, если бы не функционировала
слуховая система дельфина.
5. Трудно с эволюционной точки зрения объяснить необходимость нового органа
звуковой рецепции при наличии у млекопитающих великолепно развитого слуха,
в частности бинаурального слуха, позволяющего объяснить способность человека
и животных осуществлять пространственный слуховой анализ. Это тем более верно,
что именно слуховая система дельфина развита исключительно хорошо.
Все это вместе взятое позволяет считать указанные гипотезы, основанные на
принципе голографии, искусственными.
В связи с вопросом о голографическом восприятий у дельфинов рассмотрим очень
интересную концепцию биолога В. А. Козака (Институт физиологии АН УССР) о
звуковидении у кашалота. Этот автор исполинскую голову крупнейшего зубатого
кита уподобляет гигантскому плавающему «акустическому глазу», в котором есть
и «сетчатка», и «линза», и «стекловидное тело», и сложный нервный аппарат.
К вогнутой передней стенке черепа кашалота примыкает полость — фронтальный
воздушный мешок. Внутри этого мешка на его задней стенке В. А. Козак обнаружил
множество (3 — 4 тысячи) пузырьков размером от горошины до голубиного яйца
(рис. 40, с). Пузырьки заполнены жидкостью, сильно иннервированы и снабжены
огромным количеством рецепторных клеток. По мнению исследователя, это пузырчатое
рецепторное поле выполняет роль сетчатки, позволяющей воспринимать звуковые
образы, аналогично зрительным образам, воспринимаемым глазом. Акустическую
линзу в голове кашалота создает уплотненная ткань, расположенная впереди
верхнего спермацетового мешка. Эта «линза» концентрирует и направляет звуковые
лучи на пузырчатую выстилку. Верхний спермацетовый мешок (рис. 40, б), прозрачный
для звуковых лучей, играет роль стекловидного тела и служит звукопроводом.
Предполагается, что ход звуковых лучей при эхолокации кашалота таков: отраженный
от объектов звук падает на переднюю стенку головы животного, проходит, концентрируясь,
через плотную линзу в вытянутый спермацетовый мешок и попадает на пузырчатую
выстилку — рецепторное поле фронтального мешка. Отсюда раздражение поступает
в соответствующий центр головного мозга, где и создается звуковой образ.
Звуковидение для кашалота, питающегося в зоне вечного мрака, имеет первостепенное
значение и позволяет даже слепым особям иметь нормальную упитанность. В ходе
эволюции глаза этого кита все больше и больше сдвигались на бока головы,
а вперед выдвигалась звукопреломляющая акустическая линза и спермацетовый
мешок — акустический канал — волновод. Орган акустического зрения кашалота
формировался для работы на глубинах, где протекала большая часть его жизни.
Как полагает В. А. Козак, огромный «акустический глаз» позволяет кашалоту
получать стереоскопическую картину окружающей обстановки в трехмерном измерении.
Свою концепцию об акустическом, зрении кашалота этот ученый построил на основании
им же открытого рецепторного пузырчатого поля, которое не обнаружено у дельфинов.
Поэтому, в какой мере критические замечания Н. А. Дубровского о голографическом
восприятии дельфинов затрагивают и теорию В. А. Козака о звуковидении кашалота,
предстоит выяснить в будущем.
В естественных условиях локационный аппарат китообразными используется на
каждом шагу. Дельфины применяют его, например, для разведки препятствий на
их пути. При этом время между произведенным щелчком-сигналом и возвратом
его эха животные, видимо, используют для определения расстояния до объекта.
Калифорнийские исследователи Вильям Эванс и Джон Дреер с судна «Си-Квест»
наблюдали поведение стайки афалин в лагуне Скаммона. На пути плывущей группы
из пяти афалин оказался звуковой барьер из вертикальных алюминиевых труб.
Животные, эхолоцируя, заметили барьер в 360 м от себя и замедлили ход. Затем
стайка повернула к берегу, а от нее отделился разведчик. Он, лоцируя, подплыл
к препятствию и вернулся к группе; тогда вся стайка, издавая эхо-сигналы,
направилась к преграде, проплыла между алюминиевыми трубами и продолжила
путь в первоначальном направлении.
Подобное наблюдение повторили биологи США Давид и Мельба Колдуэллы и Сибеналер
во время лова атлантической афалины в узком заливчике во Флориде. Как только
одна стайка дельфинов вошла в залив, выход из него перекрыли сетями. Тогда
от стайки отделился вожак, разведал преграду, а затем в сопровождении крупной
самки атаковал сеть. Через разрыв сети и через места, где верхний край ее
погрузился, вся стайка ушла в море.
На основании опытов в океанариумах и наблюдений в природе был сделан общий
вывод, что эхолокация у китообразных — основной способ распознавания объектов,
погруженных в воду. Скрипучими сериями звуков высокой частоты они «ощупывают»
окружающую обстановку и получают необходимую информацию.
В условиях хорошей освещенности дельфины ориентируются и зрением. Французские
исследователи Р. Бюснель и А. Дзидзик наблюдали в Средиземном море, что белобочки
в ясную и тихую погоду осуществляют «молчаливую навигацию» и пользуются своим
гидролокатором гораздо реже, чем при недостающем свете.
Эхолокация разъясняет многие таинственные явления в жизни, мелких китообразных.
Например, почему они в мутных водах и ночью не ловятся в мелкоячеистые сети,
но попадают в крупноячеистые, «прозрачные» для отраженных звуков, не касаются
мин, когда резвятся на минных полях; почему речные дельфины — инии и соталии,
— не повреждая себя, свободно плавают в сумеречных лесах, заливаемых во время
половодья реки Амазонки, а обитатель мутного Ганга — слепой дельфин платаниста,
возможно, обходится вообще без зрения.
Принцип действия эхолокационного аппарата дельфинов ныне широко применяется
в технике в виде сонаров. Термин «сонар» происходит от начальных букв трех
английских слов: «sound» — звук, «navigation» — навигация и «range» — дальность.
Звукоизлучающие приборы посылают импульсы в воду, а их эхо доставляет информацию
о направлении и расстоянии до подводных лодок противника, до невидимых препятствий,
опасных для навигации (айсберги, мели, рифы и т. п.). На этом же принципе
построена работа эхолотов при измерении глубин и поисках косяков рыб.
Такие приборы — верные помощники человека в службе безопасности кораблевождения
и в рыболовстве. Однако эффективность природных локаторов оказалась гораздо
выше технических характеристик существующих сонаров и эхолотов. Гидролокатор
дельфина поражает ученых своей универсальностью, миниатюрными размерами и
точностью лоцирования. Он выгодно отличается от применяемых в морской практике
гидролокационных аппаратов более высокой устойчивостью к помехам, большей
шириной спектра излучаемых колебаний, богатством модуляций по интенсивности
и частоте повторения. Гидролокатор дельфина является ныне важнейшим прототипом
для моделирования в технике.
Естественные свойства дельфинового сонара объединяют оба основных принципа
современной технической гидроакустики: активный поиск, когда посылается пакет
волн и улавливается от него эхо, и пассивное подслушивание шумов и звуков,
присущих самим лоцируемым объектам.
Гидробиоников привлекает феноменальная точность локации дельфинов и замечательная
портативность их локатора, превосходящая все образцы аппаратов того же назначения,
созданных человеком. Рыбопоисковый эхолот «Дельфин» весит столько, сколько
и афалина, тогда как гидролокатор афалины составляет ничтожную часть ее веса.
Современные подводные лодки снабжены мощным гидролокационным оборудованием.
Природа гидролокатора идентична дельфиновой, но этот сложный прибор занимает
огромное место в лодке, и его обслуживает целый штат техников.
Удивление вызывает способность дельфинов выделять из хаоса звуков только
необходимые. В бассейн, где жили афалины, с целью их дезориентации излучались
их же гидролокационные сигналы, записанные ранее. Однако эти сигналы не могли
сбить животных и помешать точности их гидролокации.
1 При такой точности лоцирования преград нельзя считать столкновения кашалотов с кораблями случайными, а не преднамеренными. Вот недавний пример: советский китобоец «Циклон» вел промысел в океане. После выстрела из пушки по кашалоту тот ринулся на китобоец и ударил в борт массивной головой так, что судно сильно накренилось, а двигатели вышли из строя. Поврежденное судно пришлось отбуксировать во Владивосток. |
В голове дельфинов имеется сложный вычислительный механизм, с помощью которого
нужные эхо-сигналы, отраженные от цели, отделяются от ненужных, отраженных
от посторонних предметов, дна и поверхности1. Столь замечательный
сонар позволяет дельфинам «видеть» ушами, «слушать» челюстями, а «разговаривать»
ноздрей
и макушкой.
О чем же могут «разговаривать» дельфины?

Глава 1. Нашествие сенсаций 8
Глава 2. «Помощники» и друзья человека в море 12
Глава 3. С суши в воду 18
Глава 4. Человек и дельфины 34
Глава 5. Поведение узников 46
Глава 6. «Тираны» китов — любимцы зрителей 62
Глава 7. Скороходы морей 70
Глава 8. Покорители морских глубин 84
Глава 9. Судьба анализаторов 94
Глава 10. «Акустические глаза» дельфинов 110
Глава 11. Поющие киты и «морские попугаи» 122
Глава 12. Обладают ли дельфины речью? 140
Глава 13. Мышление дельфинов конкретно, а действия
трафаретны 148
Глава 14. Рефлексы выныривания и инстинкт сохранения
вида 152
Глава 15. Сознательно ли дельфины спасают людей
160
Глава 16. «Самоубийства» среди китообразных или
жертвы инстинкта 164
Глава 17. Странные попутчики или бездумные лоцманы
172
Глава 18. Дельфины и наука 176
Глава 19. Сигналы дальнего действия и звуковые
маяки в океане 184
Глава 20. «Звезды моря» служат человеку 188
Глава 21. Будущее дельфинов 198
Систематический список видов отряда китообразных 202
Алфавитный список видов китообразных 205