| Главная | Библиотека | Форум | Гостевая книга |
|
Глава
7
Скороходы морей
Бионика — новая наука, возникшая на стыке биологии и техники. Она использует
биологические процессы, живые системы как модели для создания новых технических
устройств и решения инженерных задач. Поскольку живые биологические прототипы
для целей моделирования в технике могут относиться к самым различным группам
живой природы, возможность применения бионики очень широка.
В мире существует почти 1,5 млн. разных видов животных, и все они отлично
приспособлены к окружающей среде. Такая приспособленность — результат естественного
отбора, в котором выживают наиболее приспособленные и гибнут неприспособленные.
Естественный отбор — непрерывно действующий механизм, который оттачивает
до совершенства любые признаки всякого вида, наиболее соответствующие условиям
жизни в данной среде. Для водных млекопитающих важное преимущество в борьбе
за существование дает быстрота передвижения. Завоеванию дельфинами Мирового
океана способствовали их быстроходность и отличная звуковая ориентация, обеспечиваемая
совершенно устроенным гидролокатором. Изучение приспособлений быстро плавающих
водных животных может подсказать ценные идеи для повышения скоростных качеств
судов. Особо достойны изучения дельфины (рис. 23, 24), способные стремительно
двигаться и совершать гигантские прыжки при сравнительно слабом расходе энергии:
ведь в течение многих тысячелетий они приспособлялись догонять быстроходных
рыб, чтобы питаться1.
1 Остается загадкой, как в желудок тихоокеанского кашалота
попали три полных меча — по 80 см длиной — самой быстроходной меч-рыбы,
извлеченных В. А. Козаком при разделке кита на китобазе «Дальний Восток»
в 1969 г. |
Их скорость зависит, конечно, от хорошей обтекаемости как всего тела, так
и его отдельных частей. Главный локомоторный орган китообразных — хвост,
т. е. сжатый с боков хвостовой стебель и горизонтально расположенные хвостовые
лопасти. Хвостовой стебель совершает удары вверх-вниз, а лопасти работающего
хвоста принимают разные углы наклона к продольной оси стебля: когда стебель
идет вниз, лопасти поворачиваются вверх и наоборот. Вращательных движений
при плавании хвост не производит. Частота и размах ударов хвоста и степень
наклона хвостовых лопастей влияют на быстроту плавания. Наблюдения показывают,
что дельфины, плывущие возле носа эсминцев и быстроходных катеров, могут
двигаться со скоростью до 65 км/ч. Однако в этих случаях животные используют
толкающую силу «оседланной» корабельной волны, и такая скорость не свойственна
им при свободном движении в море. В экспериментах, проведенных на Гавайских
островах с малайскими продельфинами, специально выпускаемыми для скоростного
пробега, Т. Ланг и К. Прайор установили максимальную скорость не более 40,6
км/ч (11,3 м/сек). Близкую цифру (36 км/ч, или 10 м/сек) нашли советские
исследователи В. Шулейкин, В. Лукьянова и И. Стась в 1937 году для дельфина-белобочки.
У гринды максимальная скорость в Калифорнийском океанариуме была 40 — 48
км/ч, а у косаток, по наблюдениям с судна «Монтерей», варьировала в течение
20 минут от 38 до 55 км/ч. Даже самые быстроходные дельфины способны двигаться
с большой скоростью недолго, лишь несколько минут.
Удивляло то, что по расчетам гидродинамиков дельфин не может развивать скорость
более 20 км/ч, а в действительности он превышает эту цифру в 2 — 3 раза.
Каким образом обеспечивается высокая скорость передвижения китообразных?
Еще в 1936 году американец Джемс Грэй, изучавший движение водных животных,
заинтересовался необычайной легкостью плавания дельфинов. Грэй буксировал
по бассейну жесткую модель дельфина, замеряя сопротивление воды. В результате
он пришел к поразительному выводу: дельфины развивают такую высокую скорость,
которая по расчетам выходит за пределы их мускульной силы. Получалось, что
дельфины либо должны иметь неестественно высокую мощность мышц, в 7 раз большую,
чем у любого млекопитающего (что невозможно), либо им удается каким-то способом
уменьшать сопротивление воды. Но каким? Это необъясненное в то время явление
— двигаться быстрее своих возможностей — получило название парадокса Грэя.
Гидродинамические свойства дельфина оказались совершеннее, чем в конструкциях
торпед или подводных лодок, созданных человеком. Этот факт заинтересовал
конструкторов подлодок и скоростных самолетов. Гидродинамик инженер Макс
Крамер в США в 1958 — 1960 годах показал, что сопротивление воды, испытываемое
дельфином при движении, в 10 раз меньше, чем сопротивление при движении модели
того же размера и формы с обычной обшивкой. Стало очевидно, что быстроходность
китообразных зависит не только от формы тела и сильной мускулатуры хвоста,
но и от их «волшебной» кожи.
Гладкая, упругая кожа дельфинов привлекла внимание судостроителей, и американская
фирма «Раббер компани» смоделировала гладкую обшивку для судовых корпусов.
Заинтересовались дельфиньей кожей и военно-морские силы США, чтобы имитировать
аналогичное каучуковое покрытие и тем самым повысить скорость подводных лодок.
1 Существует у судостроителей ошибочное мнение, что кожа
дельфинов выделяет полимерную смазку, которая уменьшает сопротивление
трения жидкости и гасит турбулентные пульсации благодаря длинно-цепочной
структуре молекулы-полимера. В действительности же у дельфина нет ни
кожных желез, ни их выделений и «рыбный» (за счет кожной слизи) способ
уменьшения сопротивления воды при плавании к нему неприменим. Однако
в коже дельфина все время отшелушивается тонкий наружный слой. Это позволяет
ему постоянно очищаться от водорослей и других морских организмов, которые
снижали бы его скорость. У трупов китов, лежащих на воздухе, верхний
слой эпидермиса затвердевает и легко отделяется вроде целлофановой пленки. |
Какие же приспособления в коже позволяют дельфину быть «скороходом морей»
и отличным прыгуном?
Кожа дельфина очень упругая, эластичная, несмачиваемая (гидрофобная), противостоит
гидравлическим ударам при большой скорости его движения1. Для
объяснения парадокса Грэя возникло несколько гипотез. Одна из них объясняла,
«что стремительно
плывущий дельфин может уменьшать вихри в потоке, обтекающем его тело, за
счет передачи тепла вдоль тела от одного участка кожи к другому». По другой
гипотезе, дельфин «уменьшает сопротивление воды поверхностью своей кожи и
способен при увеличении скорости гасить завихрения, возникающие в пограничном
слое жидкости».
Рис. 23
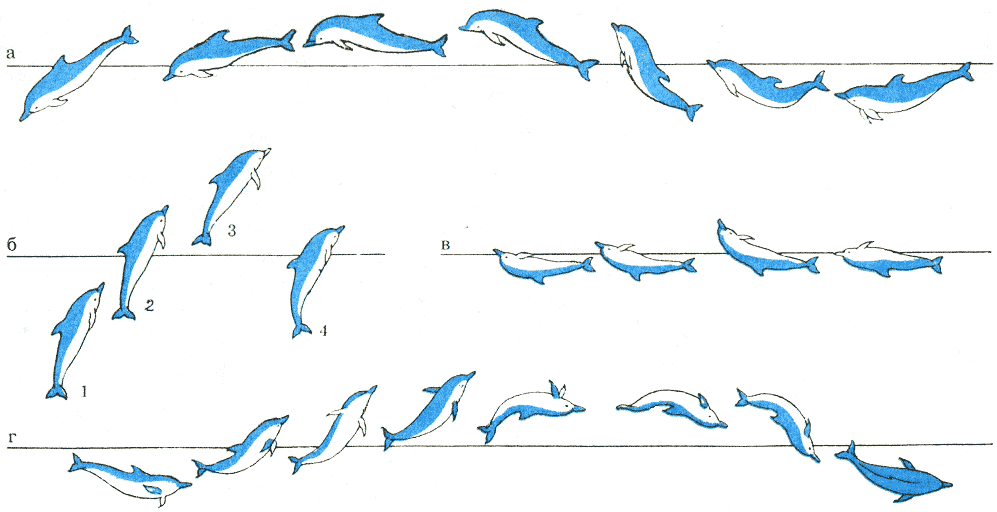
Различные прыжки обыкновенных дельфинов: а — пологий; б — высотный (по вертикали);
в — с плаванием на спине; г — с поворотом вокруг оси тела. По Г. Пиллери.
При движении в воде всякое тело испытывает сопротивление. Сила сопротивления
зависит от строения потока около тела. У хорошо обтекаемых тел можно различать
несколько областей. У носа или головного конца в пограничном слое (вблизи
тела) обтекание упорядоченное (ламинарное) и сила трения минимальная. Затем
на некотором расстоянии ламинарное обтекание нарушается и становится турбулентным:
в пограничном слое возникают вихри (поперечные движения частиц жидкости),
и сопротивление на этом участке резко возрастает. Еще дальше — к корме или
хвосту — может произойти срыв обтекания с образованием еще больших вихрей.
При конструировании обтекаемого тела основная задача — не допустить перехода
ламинарного течения в турбулентное и срыва потока. Турбулизация пограничного
слоя всегда связана с первоначальным возникновением колебаний в этом слое.
Гибкая кожа при известных условиях будет задерживать возникновение вихревого
потока в пограничном слое.
М. Крамер пытался найти в коже дельфина ту структуру, которая способствует
уменьшению сопротивления воды. В коже дельфина есть два основных слоя — эластичный
наружный (эпидермис) и лежащий под ним упругий внутренний (дерма с высокими
сосочками и жировым отложением). По мнению ученого, наружный слой выгибается
и пружинит под давлением воды, подобно поролоновому матрацу, который прогибается
в определенных местах под тяжестью тела. Бионик В. Мартека так комментирует
концепцию Макса Крамера: «Там, где вода, обтекающая быстро плывущего дельфина,
должна была бы образовывать маленькие вихри турбулентного течения, кожа прогибается
внутрь и как бы вбирает в образовавшееся углубление потенциально опасный
(в смысле турбулентности) участок водной среды. Зарождающееся в этом месте
завихрение оказывается словно изолированным от других частиц воды, проносящихся
мимо. Таким образом обеспечивается ламинарное обтекание дельфина».
Рис. 24

Вертящийся продельфин. Славится вращением тела в воздухе: при высотном прыжке успевает повернуться по оси тела 2,5 раза. Фото Лоу Джекобса.
М. Крамер учел строение кожи дельфина в 1958 году при создании им искусственного
покрытия «ламинфло» (от слов «ламинар флоу», т. е. ламинарное течение), имитирующего
кожу дельфина. Модель торпеды, одетая в оболочку «ламинфло» и испытанная
в потоке воды при скорости 70 км/ч, встретила сопротивление жидкости на 60%
меньшее, чем контрольная модель.
Оболочка «ламинфло» сделана из трех резиновых слоев общей толщиной 3,5 мм:
гладкого верхнего (0,5 мм), среднего эластичного с гибкими столбиковидными
выступами (2,5 мм) и нижнего слоя (0,5 мм), примыкающего к корпусу модели
(рис. 25). В пространство между выступами вводилась демпфирующая, т. е. погашающая
колебания, кремнийорганическая жидкость, заключенная между нижним слоем и
крышей среднего слоя. Гладкий верхний слой имитировал эпидермис кожи дельфинов,
средний подражал дерме с ее коллагеновой и жировой тканями, а нижний играл
роль опорной пластины. Позже Крамер предложил более тонкое покрытие (2,5
мм) с удлиненными гибкими гребешками вместо столбиков.
Рис. 25.
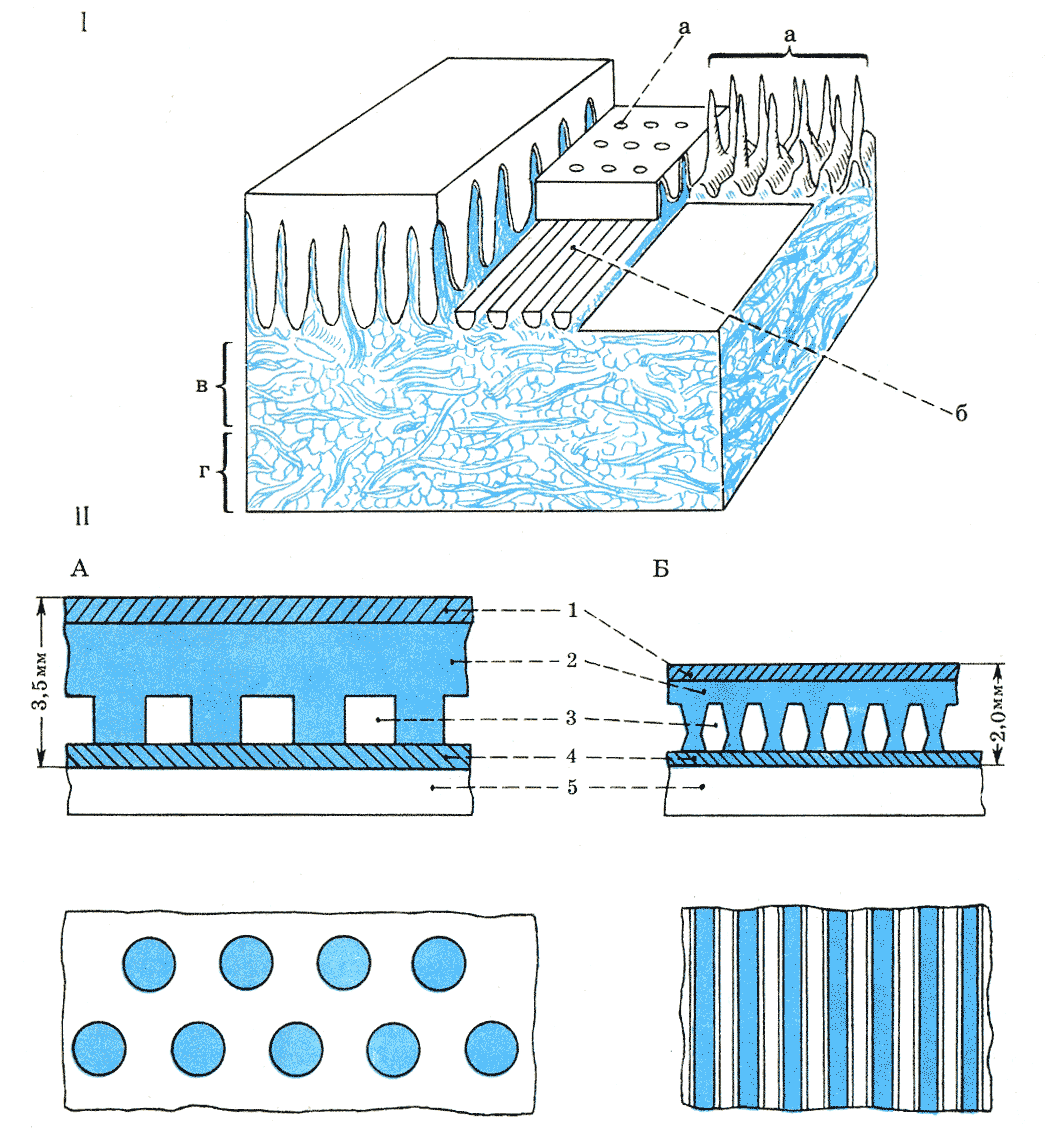
Схема строения кожи дельфина (I) и модель обшивки «ламинфло» (II): а — дермальные сосочки; в — продольные эпидермальные перегородки, между ними каналы; в — подсосочковый слой дермы; г — подкожная жировая клетчатка с пучками коллагеновых волокон; А — толстая обшивка со столбиковыми опорами; В — тонкая обшивка с ребристыми опорами: 1, 4 — верхняя и нижняя бесшовные резиновые оболочки; 2 — резиновая диафрагма; 3 — вязкая жидкость; 5 — стенка жесткой модели. По С. В. Першину и В. Е. Соколову.
Рис. 26.
Подкожная мускулатура морской свиньи: а — вид сбоку; б — вид снизу. По Р. М. Суркиной. |
Однако искусственное покрытие «ламинфло» не может до конца соперничать с
кожей дельфина, в которой демпфирование осуществляется гораздо совершеннее.
Главная причина гидродинамического совершенства дельфинов заключается в том,
что они активно управляют своей кожей, к которой подходит большое количество
нервов. Тонко улавливая силу сопротивления трения и меняющееся давление в
тех или иных участках кожи, дельфин может активно предотвратить начальные
фазы процесса турбулизации потока. От рецепторов кожи плывущего дельфина
поступает точная информация в головной мозг, и отсюда в виде биотоков по
двигательным нервам подается команда в тот ответственный участок кожи и кожной
мускулатуры, где в данный момент ощущается возросшее сопротивление воды.
Немедленно следует соответствующая двигательная реакция кожной мускулатуры,
и опасность турбулентных пульсаций ликвидируется.
Кожную мускулатуру, расположенную между дермой и подкожной жировой клетчаткой
на груди, брюхе, спине и шее дельфинов, тщательно исследовал киевский морфолог
Р. М. Суркина (рис. 26). Оказалось, что при сокращении кожной мышцы весь
слой кожи может сдвигаться и образовывать складки на поверхности тела дельфинов.
Такие складки еще в 1955 году установил с помощью подводной киносъемки во
Флоридском океанариуме Франк Эсапян: при стремительных движениях дельфинов
он заметил бегущую по телу животных мышечную волну (рис. 27). Бегущая волна,
возможно, уменьшает гидродинамическое сопротивление и предотвращает возникновение
вихревого потока в пограничном слое: работа подкожной мускулатуры как бы
настраивает кожу на оптимальный для быстрого плавания режим и задерживает
появление турбулентных пульсаций в пограничном слое.
Наконец, еще одна особенность структуры кожи имеет значение при движении
дельфинов — направленные вдоль струй потока дермальные валики. Впервые на
них обратил внимание английский биолог П. Пурвес, давший схему их расположения
на теле быстроходного и тихоходного дельфинов. Более точную схему расположения
этих валиков на теле дельфина-белобочки дала киевлянка Р. М. Суркина (рис.
28). Валики были в основном направлены вдоль тела, а на каждой хвостовой
лопасти следовали полукругами назад и сходились у выемки хвостового плавника.
Предполагается, что дермальные валики отображают направление линии потоков
вокруг плывущего китообразного. Сформировались же они в ходе эволюции в соответствии
с направлением потоков воды на коже животного при его движении. Главная адаптация
кожных покровов китообразных к быстрому плаванию, как установили советские
исследователи, заключается в самонастройке кожи путем демпфирования (см.
главу 18). Осуществляется это сосудистой системой и нервным аппаратом мельчайших
сосочков кожи, основание которых составляют дермальные валики.
Рис. 27.
Складки вокруг тела у афалины появились в момент движения с максимальной скоростью. Фото Ф. Эсапяна. |
В настоящее время кораблестроители уже заимствовали некоторые черты живых моделей — китообразных, что позволило получить выигрыш в скорости судов. За рубежом появляются суда с китовидной формой и дельфинообразными обводами носовой части. Такая конструкция позволила заметно повысить скорость хода судов при той же мощности двигателей. Известно, что в судостроении в настоящее время применяются и другие варианты судов рыбовидной формы. В частности, имеется ряд проектов полупогруженных судов со скрытым подводным корпусом и специальными надстройками, выступающими над водой. При определенных условиях такие суда могут быть более выгодными. Таким образом, в судостроении применяются и китовидные и рыбообразные формы судов. Японский профессор Тако Инуи на модели корабля «Куренаи Мару» показал, что китовидная форма корпуса корабля более пригодна для движения в море, чем острокилевая, и кораблестроители построили океанский корабль с китовидным корпусом. Новое судно оказалось более экономичным: оно обеспечивало ту же скорость и грузоподъемность, хотя мощность его двигателей была на 25% меньше, чем у судов того же назначения.
Рис. 28.
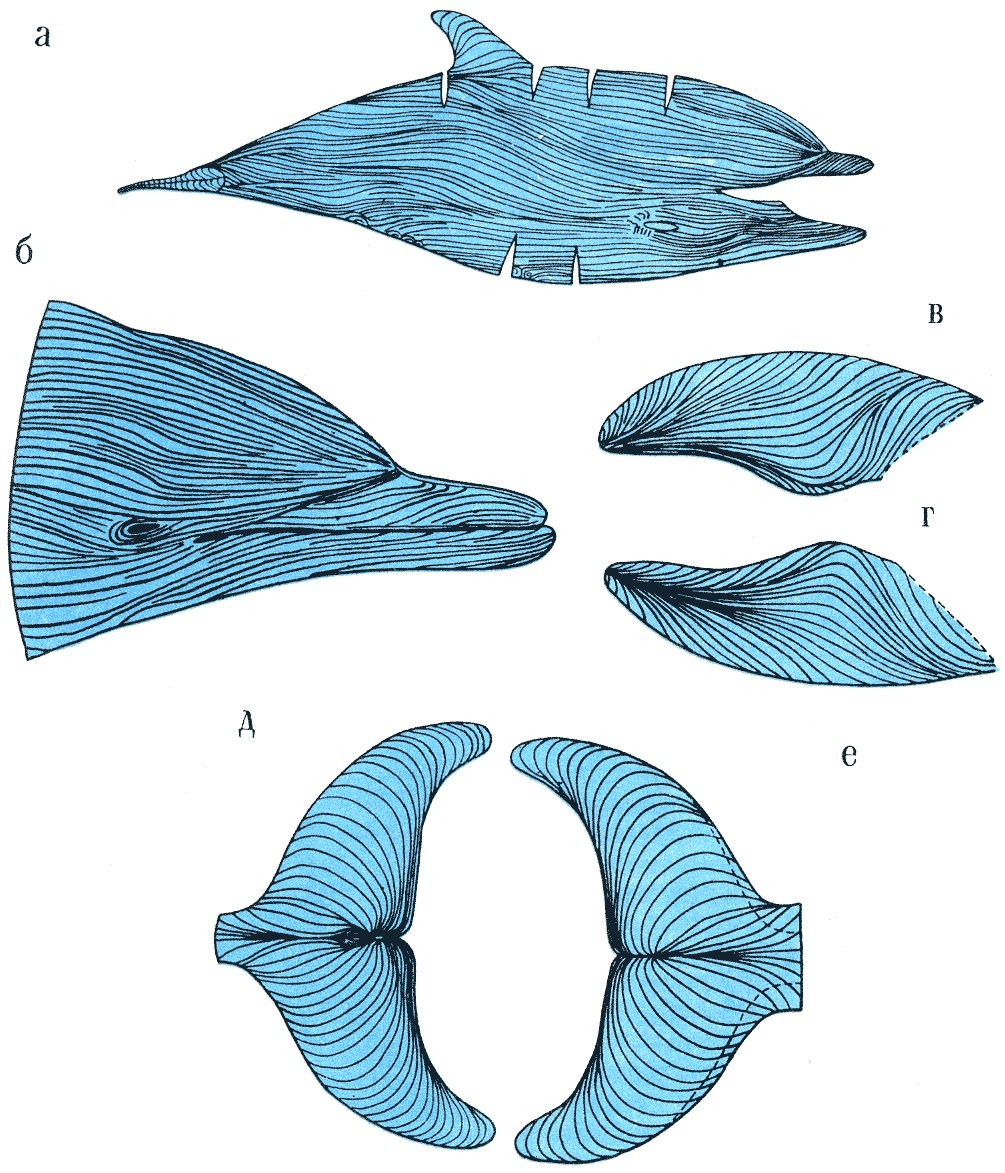
Расположение и направление дермальных валиков кожи обыкновенного дельфина: на теле (а), на голове (б), на грудных плавниках сверху (в) и снизу (г), на хвостовых плавниках сверху (д) и снизу (е). По. Р. М. Суркиной.
Дельфины замечательно используют законы гидродинамики, когда «оседлывают» волны, т. е. пассивно скользят возле носа идущего судна. Хвостовые плавники их при этом не работают, а тело движется (иногда на сотни метров) вместе с волной, возникающей у носа корабля. Поскольку корабль следует вперед, он создает давление гидродинамического поля впереди носа, которое мы не видим, но которое дельфины, тонко обнаруживают своим кожно-мускульным чувством. Сила, толкающая дельфина, находящегося в таком поле, образно сравнивается с сильным ветром, толкающим человека на улице. Эту теорию, предложенную учеными США Э. Феджером и Р. Баккусом, подтверждают подводные фотографии: позы пристроившихся к кораблю дельфинов свидетельствуют, что животные используют давление поля в корабельной волне всем телом, а не только нижней поверхностью хвостовых лопастей. Лопастями хвоста они контролируют гидродинамическую толкающую силу (рис. 29). Действие поля дельфины могут ощущать на расстоянии до 1,5 м впереди и до 1 м в сторону от носа корабля, притом не только близ поверхности, но и на всю глубину погружения носовой часа судна. Стайки этих юрких животных держатся возле носа кораблей обычно от нескольких минут до 1 часа и выстраиваются временами в два или три этажа. Пока не найдено нужное положение, дельфины крутятся в воде, стремительно ныряют, энергично работают плавниками, а потом плывут какую-то часть времени пассивно, затем снова чередуют активное поведение с пассивным скольжением и т. д.
Рис. 29.

Пятнистые продельфины оседлали корабельную волну. Фото сотрудников Флоридского океанариума.
Рис. 30.

Эшелонированное плавание матери и детеныша афалины во Флоридском океанариуме «Морская студия». Фото сотрудников океанариума.
Для оседлывания корабельных волн дельфинам нужна высокая скорость кораблей.
Стайки быстроходных белокрылых морских свиней редко следуют возле судов,
идущих со скоростью 17 км/ч, и то лишь на 1 — 2 минуты, но очень часто и
на более продолжительный срок пристраиваются к носу судов, если те идут со
скоростью 20 — 30 км/ч.
Недавно немецкий исследователь Генрих Гертель установил, что пассивно движущийся
дельфин, оседлавший корабельную волну, испытывает разное сопротивление воды
в зависимости от глубины его погружения: в 50 см от поверхности сопротивление
воды бывает максимальным, а на глубине 1,5 — 2 м — становится минимальным.
Если дельфин способен так тонко улавливать разницу в сопротивлении воды на
разной глубине в пределах двухметрового слоя, то он, вероятно, широко использует
эту способность и в жизни, уменьшая сопротивление воды путем регулирования
глубины своего хода в поверхностной толще моря. Этим патентом природы человек
еще, несомненно, воспользуется в скором будущем.
Явление «оседлывания волны» объясняет, почему дельфины, двигаясь близ носа
быстроходных судов, имеют увеличенную скорость в сравнении с их естественной.
В природе аналогичное явление известно под названием «лоцманирования» у акул:
маленькая, сравнительно тихоходная рыбка-лоцман плывет впереди быстроходной
акулы как «поводырь», держась почти вплотную к телу хищника в пограничном
слое; она словно приклеивается к этому слою воды и плывет, почти не затрачивая
усилий.
В 1961 году у китообразных было открыто другое интересное для гидробиоников
явление, которое калифорнийские зоологи К. Норрис и Д. Прескотт назвали «эшелонированным
плаванием». Оказалось, что детеныш, плывущий рядом с кормящей маткой, выбирает
наиболее выгодное положение относительно ее тела. Это экономит силы молодого
животного: сосунок, используя давление поля, создающегося вокруг стремительно
идущей самки, может плыть пассивно, как дельфины при оседлывании корабельных
волн, и не отстает от родителя, несмотря на свои еще слабые удары хвоста.
Таким образом, с действием гидродинамического поля дельфины знакомятся очень
рано. Взрослый дельфин может «буксировать» молодого, если тот придерживается
строго определенного положения. Это место легко себе представить, если поместить
детеныша рядом и параллельно боку тела матери, притом так, чтобы конец морды
детеныша не выдавался вперед за грудной плавник родителя (рис. 30).
Вначале сосунок редко отплывает от матери и тут же снова пристраивается в
излюбленном месте. Иногда он занимает положение между спинными плавниками
двух взрослых дельфинов. В океанариумах наблюдали, как детеныш, спасаясь
от преследования крупного самца афалины, пристраивался к материнскому боку,
чтобы таким способом поддержать высокую скорость.
Эшелонированное плавание свойственно и китам. Нам не раз приходилось видеть
сосунков-полосатиков и всегда только очень близко возле родителя. При выныривании
юркое тело детеныша появлялось на поверхности одновременно с массивным телом
кита.
Случалось, что мелкие дельфины «оседлывали» более крупных китообразных другого
вида. В океанариумах замечали, как афалины, полосатые и обыкновенные дельфины
пристраивались к плывущим взрослым гриндам. Вероятно, по этой же причине
в стадах гринд в море изредка присутствуют небольшие дельфины другого вида.
Поскольку такая «буксировка» задерживает скорость хода крупного животного,
то последнее нередко прогоняет от себя «наездника».
Английский ученый Бриан утверждает, что стадо косаток, выстроенное фронтом,
плывет быстрее, чем одиночки. Изучение всех этих закономерностей при движении
китообразных строем, шеренгой, тесным стадом представляет определенный практический
интерес. Это могло бы иметь значение, например, при разработке конструкций
катамаранов (двухкорпусных судов), при поисках более выгодного взаиморасположения
судов и катеров при их групповом передвижении или наилучшего размещения лодок
во время гонок, соревнований и т. п.
Большой интерес к гидродинамическим исследованиям дельфинов проявляет военно-морское
ведомство США, исходя из девиза «живые прообразы — ключ к новой технике».
Человек повторил форму китообразных в контурах подводных кораблей. Дельфин
освободил человека от необходимости искать ее и выдал ему один из секретов
своей быстроходности: люди построили подводную лодку с дельфинообразными
обводами. Таковы, например, формы таинственно погибшей в 1963 году американской
атомной подводной лодки «Трешер» и экспериментальной подводной лодки США
«Дольфин», которые развивали скорость до 50 км/ч. Таким образом, дельфин
становится важным объектом для моделирования в гидробионике.
Мысли гидробиоников и океанологов направлены сейчас на изучение загадочных
океанских глубин. В каком направлении следует вести поиски с целью увеличения
глубины и продолжительности свободного ныряния человека без технического
снаряжения? Не поможет ли в решении этой проблемы изучение приспособлений
лучших ныряльщиков мира — китов и дельфинов?

Глава 1. Нашествие сенсаций 8
Глава 2. «Помощники» и друзья человека в море 12
Глава 3. С суши в воду 18
Глава 4. Человек и дельфины 34
Глава 5. Поведение узников 46
Глава 6. «Тираны» китов — любимцы зрителей 62
Глава 7. Скороходы морей 70
Глава 8. Покорители морских глубин 84
Глава 9. Судьба анализаторов 94
Глава 10. «Акустические глаза» дельфинов 110
Глава 11. Поющие киты и «морские попугаи» 122
Глава 12. Обладают ли дельфины речью? 140
Глава 13. Мышление дельфинов конкретно, а действия
трафаретны 148
Глава 14. Рефлексы выныривания и инстинкт сохранения
вида 152
Глава 15. Сознательно ли дельфины спасают людей
160
Глава 16. «Самоубийства» среди китообразных или
жертвы инстинкта 164
Глава 17. Странные попутчики или бездумные лоцманы
172
Глава 18. Дельфины и наука 176
Глава 19. Сигналы дальнего действия и звуковые
маяки в океане 184
Глава 20. «Звезды моря» служат человеку 188
Глава 21. Будущее дельфинов 198
Систематический список видов отряда китообразных 202
Алфавитный список видов китообразных 205