Эдуард Израилевич Колчинский
Эволюция биосферы

АКАДЕМИЯ НАУК СССР
ИНСТИТУТ ИСТОРИИ, ЕСТЕСТВОЗНАНИЯ
И ТЕХНИКИ
Э. И. КОЛЧИНСКИЙ
ИСТОРИКО-КРИТИЧЕСКИЕ ОЧЕРКИ
ИССЛЕДОВАНИЙ В СССР
Ответственные редакторы:
акад. А. М. УГОЛЕВ, А. В. ЛАПО
Ленинград
„НАУКА”
Ленинградское отделение
1990
УДК 576.82
Колчинский Э. И. Эволюция биосферы: Историко-критические очерки
исследований в СССР. – Л.: Наука, 1990. – 236 с.
В книге анализируются основные направления в разработке проблемы эволюции
биосферы, сложившиеся в нашей стране под влиянием работ В. И. Вернадского.
Особое внимание уделяется проблеме геохимических факторов эволюции биосферы,
ее биогеохимических функций, преобразующего влияния жизни на атмосферу,
литосферу и гидросферу, а также главным закономерностям в преобразовании
биосферы. Анализируются пути управления эволюцией в современной биосфере.
Ставится задача синтеза эволюционной теории и учения о биосфере. Библ.
663 назв. Ил. 16. Табл. 7.
Рецензенты:
Д. В. Лебедев, Ю. И. Полянский
| К |
1903040000-595 |
494-90, l полугодие |
|
© Э. И. Колчинский, 1990 |
| 042(02)-90 |
|
|
ISBN 5-02-026646-9
ПРЕДИСЛОВИЕ
Уже в период формирования современного дарвинизма некоторые биологи предвидели,
что в канву создаваемого синтеза необходимо включить „данные биохимии,
исторической геологии и геохимии” (Завадский, Шапоренко, 1934, с. 159).
Несколько позже М.М. Камшилов (1947) подчеркнул, что биогеохимические
исследования В.И. Вернадского и А.П. Виноградова относятся к важнейшим
достижениям советской науки в познании закономерностей эволюции органического
мира, так как прокладывают путь к раскрытию закономерностей эволюции
биосферы. Однако в те годы эти идеи не только не получили всеобщего
признания, но даже казались странными для большинства биологов-эволюционистов,
которые
еще совсем недавно считали организм первичной и основной формой существования
жизни, а эволюцию живого сводили преимущественно к формированию адаптации
организмов к окружающей среде.
Синтетическая теория эволюции покончила с подобной „монополией” организма
в эволюции, доказав, что последняя может совершаться только в статистически
насыщенных ансамблях родственных организмов — популяциях. Отныне преобразования
генетических и экологических структур популяций и способы видообразования
оказались в центре эволюционно-биологических исследований. Эволюцию же
надвидовых уровней организации жизни практически не изучали. Даже главные
этапы эволюции биогеоценозов и биосферы характеризовали не специфическими
признаками, присущими каждой из этих форм организации живого как целостной
системы, а описанием организмов и видов, входящих в их состав на правах
элементарных компонентов. Биогеоценоз рассматривали только как арену
эволюционных преобразований, а развитие биосферы, по словам П. Дювиньо
(1979), не признавалось
эволюционистами за достоверный процесс.
Глобальные изменения в поверхностных оболочках Земли, происшедшие за
последние десятилетия под влиянием промышленности, расширения сельскохозяйственных
угодий, урбанизации и т. д., заставили по-новому взглянуть на процессы
развития природы. Изменилось и отношение к проблеме эволюции надвидовых
форм организации живого. Стало общепризнанным, что эволюционный процесс
захватывает не только отдельные организмы, виды, биогеоценозы, но и высший
уровень организации живого – биосферу. В различных отраслях естествознания,
связанных с изучением биосферы, накапливается все больше данных о том,
что эволюция органического мира изменяет геохимическое строение и энергетику
биосферы, ее биогеохимические функции и биогеохимический круговорот веществ.
В свою очередь эти преобразования биосферы в целом становятся мощными
факторами
дальнейшей эволюции ее важнейшего компонента — органического мира.
3
Не случайно многие биологи-эволюционисты пришли к выводу, что „органическая
эволюция — это прежде всего эволюция биосферы, в которую каждый вид вносит
свой специфический вклад” (Wahlert, 1978, S. 116). Изучение преобразований
таких параметров биосферы, как продуктивность, „энерговооруженность”, доступность
минеральных веществ и их оборачиваемость в биогеохимических циклах, по
мнению Г. Валерта, позволяет объяснить прогрессивную эволюцию одних организмов
и сохранение на более низком уровне других. Он даже считает, что попытки
рассматривать филогенез какой-либо таксономической группы без учета ее
роли в осуществлении биогеохимических и энергетических функций биосферы
неизбежно ведут к финализму и телеологии. По мнению А. В. Яблокова и А.
Г. Юсуфова, учение об эволюции биосферы и составляющих ее биогеоценозов
в будущей эволюционной теории должно „занимать не меньший объем, чем сегодня
занимает учение о микроэволюции” (1981, с. 313). В последние годы все чаще
появляются статьи и книги, в которых с позиций эволюционной теории делаются
попытки проанализировать связи эволюции органического мира и всей биосферы
(Камшилов, 1974; Шварц, 1976; Цветков, 1981; The evolving biosphere, 1981;
Колчинский, 1981, 1983а, 1983б, 1987; Палеонтология и эволюция биосферы,
1983; Левченко, Старобогатов, 1986; Соколов, Барсков, 1988, и др.).
Вместе с тем существуют объективные трудности в познании эволюции биосферы.
Для такого познания требуется анализ следующих вопросов: в чем заключается
сущность эволюции биосферы; как осуществляется взаимная детерминация эволюционных
преобразований на уровне видов и биосферы; каковы основные тенденции в
эволюции биосферы как целостной системы; каковы основные принципы выделения
главных этапов эволюции биосферы; в чем заключаются особенности эволюции
биосферы в условиях научно-технической революции и каково их влияние на
эволюцию органического мира в настоящее время? Историко-критический анализ
этих вопросов является задачей данной книги.
Ответы на эти вопросы не могут быть даны в рамках какой-то отдельной науки
или даже какого-либо отдельного цикла наук, например биологического, геологического,
географического и т. д. Поистине эта проблема является комплексной и глобальной.
Биосфера представляет собой сложную систему, объединяющую компоненты различной
природы, каждый из которых отличается относительно самостоятельной историей
развития и собственными движущими силами этого развития. Так, эволюция
недр Земли и химического состава мантии шла под влиянием гравитационной
дифференциации веществ различной массы и энергии распада радиоактивных
элементов. Образование первичной атмосферы и гидросферы связано с дегазацией
лав, выплавившихся из верхней мантии и создавших земную кору. Вероятно,
чисто абиогенный характер носят процессы тектогенеза, орогенеза, вулканизма,
дрейф материков и т.д. Между тем в последние годы нередко выходят работы,
где развитие именно этих процессов именуется эволюцией биосферы. Нисколько
не умаляя роли органической эволюции в детерминации многих геологических,
физико-географических и геохимических процессов на поверхности
4
нашей планеты, все же необходимо признать, что вряд ли ее образование,
движения континентов и тектонико-магматические события как-то связаны с
жизнедеятельностью организмов. Следовательно, нет оснований их называть
эволюцией биосферы.
Следует подчеркнуть, что и без изучения эволюции вулканизма и дрейфа континентов
необычайно широк круг вопросов, связанных с изучением эволюции биосферы.
Сюда относятся поиски прямых и косвенных данных о кардинальных перестройках
органического мира, об эволюции биогеохимических и энергетических функций
биосферы, о перестройках ее биотического круговорота, о количестве биомассы
и биопродуктивности в биосферах геологического прошлого, о роли живого
вещества в преобразовании лито-, атмо- и гидросферы.
Такие данные, естественно, могут быть получены лишь в результате усилий
ученых различных специальностей. Соответственно и история получения этих
данных и развития представлений об эволюции биосферы может быть написана
лишь в результате совместной работы историков соответствующих отраслей
знания. В данной работе сделана лишь попытка проанализировать этапы и направления
в изучении эволюции биосферы, результаты которого могут иметь значение
для эволюционной теории и прежде всего для ее нового раздела о движущих
силах и закономерностях эволюции надвидовых форм организации жизни. Естественно,
при этом пришлось касаться вопросов, обсуждение которых не связано прямо
с многолетней специализацией автора в области истории эволюционной теории,
а иногда даже вторгаться в области естествознания, довольно далекие от
биологии. Автор испытывал известные трудности при анализе работ по геохимии,
геологии, почвоведению и т. д. Вероятно, при этом не удалось избежать просчетов
и упущений, которые могут вызвать возражения со стороны соответствующих
специалистов. Единственное оправдание автор находит в том, что эти историко-критические
очерки, возможно, пробудят интерес к истории познания закономерностей эволюции
биосферы.
Следует отметить также, что, хотя книга задумана как историко-критическое
исследование проблемы эволюции биосферы, состояние данной проблемы нередко
заставляло отступать от выбранного жанра. Дело в том, что большинство исследователей
не шло дальше отдельных высказываний, в лучшем случае проспектов будущих
исследований. Сейчас данная проблема представляет собой скорее совокупность
заявок и постановок вопроса, чем систему фактов и теоретических обобщений.
В этих условиях задача историка науки заключается не только в анализе развития
проблемы, но и в попытке на основе такого анализа систематизировать хотя
бы в первом приближении имеющиеся разрозненные данные, для того чтобы лучше
представить современное состояние этой проблемы и наметить пути ее дальнейшего
исследования.
В заключение я хочу поблагодарить своих коллег, которые на разных этапах
обдумывания замысла книги и работы по его реализации приняли участие в
обсуждении различных аспектов проблемы эволюции биосферы.
5
Глава 1
ИСХОДНЫЕ ПОЛОЖЕНИЯ
1.1. О структуре и функциях биосферы
В одной из своих последних статей В. И. Вернадский (1942) отметил расхождения
в трактовке разрабатываемых им представлений о поверхностных оболочках
Земли, особенно в трактовке понятия биосферы. Расхождения не преодолены
и по сей день. В многочисленных публикациях о биосфере бытуют резко отличающиеся
друг от друга представления об ее структуре и функциях, пространственных
границах и основных компонентах (Ефремов, 1959; Наумов, 1964; Сукачев,
1964а; Ковда, Тюрюканов, 1970; Перельман, 1973, 1977; Камшилов, 1974, 1975;
Арманд, 1975; Вассоевич, 1976; Вассоевич, Иванов, 1977, 1983а, 1983б, 1984;
Иванов, 1978, 1979; Гегамян, 1980, 1981; Голубец, 1982; Лапо, 1982, 1987;
Соколов, 1983, 1986; Голубец, Одинак, 1983, и др.).
Даже в Большой Советской Энциклопедии (БСЭ, третье издание) в различных
статьях приводятся несходные определения биосферы и соответственно по-разному
трактуются ее границы, функции и место среди других геологических оболочек
Земли. Так, авторы статьи „Биосфера” В. А. Ковда и А. Н. Тюрюканов характеризуют
биосферу как оболочку „Земли, состав, структура и энергетика которой в
существенных чертах обусловлены прошлой или современной деятельностью живых
организмов” (БСЭ, т. 3, с. 364). В состав биосферы ими включается часть
атмосферы, вся гидросфера и верхняя часть литосферы, связанные между собой
сложными циклами миграции химических элементов и обмена энергией. Аналогичное
определение биосферы в статье „Земля” приводит К. П. Флоренский, подчеркивая
специально, что в биосферу входит область существования не только современных
организмов, но и их „былого обитания” (БСЭ, т. 9, с. 484).
В статьях же „Геосферы” и „Геохимия” (последняя написана А. П. Виноградовым)
биосферой называется „область на границе твердой, жидкой и газовой оболочек
Земли, занятая живым веществом – совокупностью организмов” (БСЭ, т. 6,
с. 335). Помимо этого еще встречаются предложения понимать под биосферой
лишь совокупность живых организмов (Бернал, 1969).
Такая неопределенность в трактовке биосферы побуждает нас охарактеризовать
основные исходные положения, на которые мы будем опираться в данной книге.
Но вначале несколько слов об основных
6
моментах формирования и развития представлений о биосфере, что позволит
лучше понять причины современных разногласий.1
Истоки этих представлений уходят в естествознание XVII и XVIII веков. В
работах Ж. Бюффона, Б. Варрениуса, Вик д’Азира, Д. Вудворта, Дж. Геттона,
К. Гоффа, X. Гюйгенса, К. Линнея, М. В. Ломоносова не раз высказывались
идеи о взаимосвязанности всех процессов неорганической и органической природы.
Основой этих идей были данные о зависимости распространения животных от
климатических условий, о газовом обмене у организмов, об участии животных
и растений в формировании почв, известняков и горючих ископаемых, а также
первые гипотезы о существовании биогенного круговорота элементов (А. Лавуазье,
А. Сьнядецкий).
Само понятие „биосфера”, по мнению Вернадского, сформулировано Ж.-Б. Ламарком.
В лекциях 1800-го года Ламарк отметил, что у „живых тел обнаруживаются
все сложные неорганические вещества, наблюдаемые в природе” (1955, с. 10),
и что в местах, не заселенных организмами, минералы весьма однородны. Это
объясняется тем, что животные и растения активно участвуют в формировании
поверхности Земли.
Несколько позже в книге „Гидрогеология” Ламарк (Lamarck, 1802) подробно
остановился на этих идеях. Поставив вопрос о влиянии живых организмов на
„вещества, находящиеся на поверхности земного шара и образующие его кору”,
он пришел к однозначному выводу о том, что все минералы внешней коры Земли,
а следовательно, и состоящие из „них низменности, холмы, долины и горы
являются исключительно продуктами животных и растений, которые существовали
на этих участках поверхности земного шара” (цит. по: Вассоевич, Иванов,
1977, с. 59—60). Даже граниты Земли, по его мнению, возникли при активном
участии организмов. Тем самым Ламарк „дал яркое представление значения
биосферы в истории нашей планеты” (Вернадский, 1965, с. 64).
Для распространения подобных идей в естествознании XIX в. особое значение
имели труды А. Гумбольдта, который начиная с 1793 г. не раз высказывал
мысль об органическом мире как неотъемлемой части земной коры и отмечал
связь химического состава организмов с неорганическим веществом среды их
обитания (см., например: Гумбольдт, 1936, с. 189—198). Во втором издании
своих знаменитых „Картин природы”, опубликованных в 1826 г., Гумбольдт
(1959) ввел понятие „жизненная среда” (die Lebensphare), под которой понималась
специфическая оболочка Земли, где в единую целостную систему объединены
атмосферные, морские, континентальные процессы, а также явления органического
мира. Позже Ф. Ратцель (Ratzel, 1869) называл поверхность Земли „пространством
жизни» (der Lebensraum), местом обитания организмов. В эти же годы Э. Реклю
в специальном разделе многотомного труда
1 История учения о биосфере исследована в трудах самого
В. И. Вернадского, который очень внимательно относился к идеям своих предшественников.
В последние
годы развитию идей о биосфере были посвящены работы Д. И. Гордеева (1974а),
Н. Б. Вассоевича и А. Н. Иванова (1977), Б. П. Высоцкого (1977), А. Н.
Иванова (1979).
7

Эдуард Зюсс (1831-1914).
„Земля” (1914) в ярких красках описывал роль живого вещества в преобразовании
Земли.
Таким образом, начиная с работ Ламарка в науке укоренилось представление
о существовании специального пространства на нашей планете, охваченного
и организованного жизнедеятельностью организмов. Но из всех терминов, предложенных
для обозначения этого пространства, международное признание получил лишь
термин „биосфера”,1 введенный Э. Зюссом в 1875 г. при рассмотрении
основных геологических оболочек Земли: лито-, атмо- и гидросферы. Он полагал,
что
„в области взаимодействия верхних сфер и литосферы и на поверхности материков
можно выделять самостоятельную биосферу. Она простирается теперь как над
сухой, так и над влажной поверхностью, но ясно, что раньше она была ограничена
только гидросферой” (Suess, 1875, S. 159-160). Вводя понятие „биосфера”,
Зюсс не дал его определения, что позволяло употреблять это понятие в различных
смыслах.
Позже в книге „Лик Земли” Зюсс называл биосферой „совокупность организмов,
ограниченную в пространстве и во времени и обитающую
1 В немецкой литературе в качестве синонима „биосферы”
используют иногда термин Ратцеля „пространство жизни”, а в англоязычной
литературе
- термин
„экосфера”, предложенный К. Ламонтом (La Mont, 1958).
8
на поверхности Земли” (Suess, 1909, S. 779). Эта совокупность представлялась
ему гармоническим единством. К сожалению, и здесь Зюсс не пошел дальше
общих замечаний о геологическом месте жизни и ее зависимости от планетарных
параметров Земли.
Выделение биосферы в качестве специфической геологической оболочки стало
важным теоретическим обобщением, позволяющим синтезировать данные разных
наук о роли живого в геологических и геохимических процессах. Однако Зюсс
не подчеркнул особого места биосферы среди других геосфер. Между тем биосфера
„состоит из совокупности трех геосфер – тропосферы (нижней воздушной геосферы),
гидросферы и коры выветривания (верхней твердой геосферы) ” (Вернадский,
1926а, с. 51). Характеристика же Зюссом биосферы как „совокупности организмов,
ограниченной в пространстве и во времени” дала повод использовать данное
определение для обозначения „слоя живой материи” (Вальтер, 1911, с.6).
Такая трактовка биосферы стала широко употребляться в конце XIX в. в работах
географов, почвоведов и геологов (Д. Н. Анучин, П. И. Броунов, Г. Вагнер,
И.Ф. Мушкетов, Ф. Ратцель, Н.М. Сибирцев и др.). Оно получило впоследствии
название биологического, хотя, как отмечают Вассоевич и Иванов, биологи
того времени „термином «биосфера» вообще не пользовались” (1977, с. 65).
Таким образом, задолго до работ Вернадского сложилась традиция использовать
термин „биосфера” в двух смыслах. Одни авторы под биосферой понимали совокупность
организмов, или слой живого вещества. Другие биосферой называли пространственную
оболочку Земли, в формировании и функционировании которой активное участие
приняла жизнь.
В результате трудов В. В. Докучаева (1883, 1892, 1899), его учеников и
последователей (Г.Н.Высоцкий, Г. Ф. Морозов, Н. М. Сибирцев и др.) на рубеже
XIX и XX веков стала четко осознаваться необходимость создания целостного
учения о взаимозависимых изменениях органического мира, рельефа, вод, почв,
осадочных пород и климата. Это учение было разработано В.И.Вернадским,
который, следуя методологическим установкам своего учителя Докучаева, сумел
связать воедино процессы развития органического мира с геологической историей
других оболочек Земли.
Биосферой Вернадский называл оболочку Земли, строение и организованность
которой обусловлены жизнедеятельностью организмов. Для Вернадского биосфера
была планетарной биокосной системой, в которую жизнь входила на правах
хотя и главного, но далеко не единственного компонента. Тем самым биологическое
определение биосферы теряло всякий смысл. С введением И. Д. Лукашевичем
(19096) термина „живое вещество” для обозначения „совокупности организмов”
становится недопустимым применение в этом смысле термина „биосфера”.1
1 Для обозначения совокупности организмов в биологии используют
чаще всего такие термины, как биомасса, биос, биота, биострома, биома и
др.
9
Высказывания Вернадского о пространственных границах биосферы далеко не
всегда согласовываются друг с другом (см. подробнее: Гегамян, 1980; Вассоевич,
Иванов, 1983а, 1983б). В 20-х годах он границы биосферы сужал обычно до
„поля существования жизни”, верхние пределы поля ограничивались высотой
9 — 13 км, а нижние — 2.5 — 2.7 км на суше и 5.5 км в океане. В книге „Биосфера”
он писал: „По существу биосфера может быть рассматриваема как область земной
коры, занятая трансформаторами, переводящими космические излучения в фиксированную
земную энергию — электрическую, химическую, механическую, тепловую и т.
д. ” (1926а, с. 14). В более поздних работах Вернадский (1937, 1942) склонялся
к расширению и „поля существования жизни”, и самих границ биосферы. В биосферу
уже включалась не только вся тропосфера, но и часть стратосферы (до высоты
22 км), гидросфера, в том числе и подземная, частично (а в некоторых высказываниях
и вся) стратосфера (наибольшая глубина 8.5 км). В книге „Химическое строение
биосферы Земли и ее окружения”, опубликованной посмертно в 1965 г., есть
высказывания, где к зоне биосферы относятся фактически все горизонты земной
коры, несущие на себе печать воздействия прошлой или современной жизни.
К расширению границ биосферы побуждала Вернадского необходимость учесть
связь между геохимическими и энергетическими процессами, протекающими в
„поле существования жизни” и в других геосферах, преобразованных прошлой
жизнью и содержащих громадное количество аккумулированной ею раньше энергии.
Различающиеся, а иногда и взаимоисключающие друг друга высказывания Вернадского
о границах биосферы и вызвали современные разногласия. Одни авторы, считая,
что „дух” биогеохимических идей Вернадского точнее выражен в его ранних
работах, требуют понимать под биосферой только область обитания современных
организмов (Вассоевич, 1976; Вассоевич, Иванов, 1977, 1983а, 1983б; Шипунов,
1980; Лапо, 1982, 1987; Соколов, 1983, и др.). Другие же, отдавая явное
предпочтение более поздним высказываниям Вернадского, пишут о биосфере
как мощной геологической оболочке, испытавшей когда-либо влияние жизни,
т. е. включают в нее и зоны былых биосфер (Тимофеев-Ресовский и др., 1969;
Ковда, Тюрюканов, 1970; Гегамян, 1980; Шилова, 1983, и др.).
Путь к преодолению этих разногласий был предложен Вассоевичем и Ивановым.
Для этого достаточно различать „биосферу как область активной жизни и мегабиосферу,
включающую как биосферу (эубиосферу), так и прилегающие к ней сверху и
снизу оболочки, лишенные жизни, но испытавшие то или иное прямое или косвенное
ее влияние” (1983б, с. 291 — 292). Структура мегабиосферы в понимании
Вассоевича (1976) выглядит следующим образом. Верхний слой ее составляет
апобиосфера,
т. е. безжизненное пространство верхней части атмосферы, еще содержащей
кислород. Затем следует парабиосфера — область временного проникновения
организмов, спор, пыльцы. Названные области расположены над биосферой (эубиосфера
— арена жизни). Снизу к ней примыкает метабиосфера, или былые биосферы,
составленные из измененных остатков прошлой жизни. Но не все былые биосферы
входят в состав
10
метабиосферы. Часть из них содержится в осадочных породах современной
биосферы.
Представления Вассоевича о биосфере как части более мощной геологической
оболочки используются в нашей работе. При изучении эволюции биосферы учитываются
и данные о строении всей мегабиосферы, вплоть до гнейсов и гранитов, ибо
в осадочных и метаморфизированных породах содержатся „следы былых биосфер”
(Лапо, 1987).
Подчеркивая целостность, организованность биосферы и „всюдность” ее живого
вещества, Вернадский (1965) вместе с тем указывал на наличие в ней четких
дискретных подразделений. Особо он выделял охваченный жизнью слой биосферы,
где сосредоточена основная масса организмов. В этом слое различались три
„пленки жизни” (геохоры): наземная, планктонная, донная. Мощность наземной
пленки колеблется от нескольких десятков сантиметров до десятков метров,
а в гидросфере зона максимального сосредоточения жизни достигает мощности
100 м и более.
В 1949 г. Е.М. Лавренко ввел понятие „фитогеосферы” как основной структурной
единицы биосферы, в пределах которой и происходят геохимические и энергетические
процессы, связанные с жизнедеятельностью организмов. При этом называлось
несколько типов фитогеосфер: поверхностная, подводная и наземная. Под фитогеосферой
понимался не только растительный покров, но и связанные с ним консументы
и редуценты вместе с населенной ими средой.
Предложение Лавренко вызвало затяжную дискуссию, носящую преимущественно
терминологический характер (см.: Голубец, Одинак, 1983). Соглашаясь с необходимостью
выделения в биосфере слоя максимальной концентрации живого вещества, участники
дискуссии, как правило, предлагали собственные термины для его обозначения.
Так появились аналогичные по содержанию понятию „фитогеосфера” термины:
„биогеосфера” (Ефремов, 1959), „геомерида” (Беклемишев, 1964), „биогеоценотический
покров” (Сукачев, 1964а, 19646), „витасфера” (Тюрюканов, Александрова,
1969), „биокаллима” (Вассоевич, 1976) и мн. др. Но поскольку каждый из
терминов при буквальном переводе не совсем адекватно отражал суть заключенного
в нем понятия, то в литературе сейчас используется около десятка терминов-синонимов.
Не вдаваясь в лингвистические споры о том, какой термин лучше отражает
суть данного понятия, заметим, что в данной работе предпочтение отдается
термину „биогеоценотический покров”, так как он тесно связан с учением
В. Н. Сукачева (1942, 1944) о биогеоценозах как элементарных хорологических
подразделениях биосферы (Тимофеев-Ресовский, Тюрюканов, 1966). „Биогеоценоз
- это те блоки, из которых состоит вся биосфера и в которых происходят
вещественно-энергетические круговороты, вызванные жизнедеятельностью организмов
и в сумме составляющие большой биотический круговорот” (Тимофеев-Ресовский
и др., 1969, с. 330). В понятии „биогеоценоз” наиболее полно отражен дискретный,
мозаичный характер распределения живого вещества в биосфере. Для обозначения
дискретных единиц вещественно-энергетического круговорота предлагались
и другие термины: „голоцен”, „микрокосм”,
11
„эпиморфа”, „элементарный ландшафт”, „микроландшафт”, „биосистема”, „биохора”,
„фация”, „эпифация”, „биоэкос” и мн. др. Однако в литературе на германских
и славянских языках использовались преимущественно термины „биогеоценоз”
или „геобиоценоз”. Правда, сейчас и здесь наметилась тенденция к вытеснению
его термином А. Тенсли (Tansley, 1935) „экосистема”. Но последний все-таки
менее удобен при описании структурных единиц биосферы, так как экосистемой
является и вся биосфера, и любая ее подсистема, объединяющая группы популяций
разных видов и их биотоп.
Вернадский (1965, с. 58—60) предлагал следующую классификацию входящих
в биосферу веществ: 1) живое вещество или совокупность организмов; 2) биогенное
вещество, созданное и переработанное жизнью; 3) косное вещество, в образовании
которого живое вещество не участвует; 4) биокосное вещество, представляющее
собой динамические равновесные системы образующих его живого и косного
веществ; 5) радиоактивные элементы; 6) рассеянные атомы, возникшие в результате
воздействия космических излучений; 7) вещество космического происхождения.
Эта классификация, построенная на различных критериях (характер самого
вещества, причины его возникновения, природа исходного вещества, степень
дисперсии), оказалась далеко не идеальной. Из-за обилия критериев классификации
выделенные типы или частично перекрывали друг друга, или один из выделенных
типов полностью входил в состав другого.
Более последовательна классификация М. М. Камшилова (1975), который называет
четыре типа вещества: живое, биогенное, косное и биокосное. Однако биокосное
вещество – не какой-то самостоятельный тип, существующий наравне с живым
и косным. Если говорить о биокосном веществе в смысле Вернадского, то это
означает некую динамическую систему, образуемую организмами, продуктами
их жизнедеятельности и материальными условиями обитания. Поэтому прав А.
И. Перельман (1977), предлагая всю биосферу рассматривать как некую гигантскую
биокосную систему, подсистемами которой являются биокосные тела более низкого
уровня организации: почвы, илы, кора выветривания, поверхностные воды,
водоносные горизонты, ландшафты и т. д.
Особое место живого вещества в биосфере определяется его энергетическими
и геохимическими функциями. Первая из них связана с ассимиляцией солнечной
энергии и обогащением ею биогенного и косного вещества. Геохимические функции
выражаются в участии организмов в перемещениях и концентрациях химических
элементов, а также в преобразованиях окружающей среды и в создании новых
минералов. Вернадский выделял пять видов таких функций: „I) газовые функции;
II) концентрационные функции; III) окислительно-восстановительные функции;
IV) биохимические функции; V) биохимические функции человека – Homo sapiens.
Вместе взятые, они определяют основные химические проявления живого вещества
в биосфере” (1965, с. 237).
Накопленные к настоящему времени знания позволили А. В. Лапо (1987, с.
114) разработать следующую классификацию функции живого вещества: 1) энергетическая
— поглощение солнечной энергии при фото-
12
синтезе и химической энергии при разложении веществ; 2) концентрационная
— избирательное накопление определенных элементов; 3) деструктивная — минерализация
органического вещества и разложение неорганического вещества; 4) средообразующая
– преобразование физико-химических параметров среды; 5) транспортная –
перенос организмами элементов при миграциях.
Учитывая, что биосфера как целостная система играет ведущую роль в детерминации
геохимических свойств и энергетики других геосфер, видимо, можно говорить
об энергетической, средообразующей и концентрационной функциях самой биосферы.
Таким образом, под биосферой будет пониматься поверхностная оболочка Земли,
организованная живым веществом. Важнейшим структурным компонентом биосферы
является биогеоценотический покров, состоящий из биогеоценозов разного
уровня сложности и эволюционной продвинутости. В биогеоценозы входит косное,
живое и биогенное вещество. Как гигантская биокосная система биосфера выполняет
энергетические, геохимические и средообразующие функции на поверхности
Земли.
Эта совокупность представлений и используется в нашем дальнейшем изложении.
1.2. Эволюция биосферы (общие представления)
Несмотря на то что в последние годы были опубликованы десятки книг и статей,
в заголовках которых имеется словосочетание „эволюция биосферы”, а также
проведен ряд конференций и симпозиумов, посвященных данной проблеме, в
самой ее постановке остается много неясного.
Справедливо писал К. М. Завадский, характеризуя уровень знаний об эволюции
надвидовых форм существования жизни: „Ступени эволюции биоценотической
и биостроматической форм организации живого исследованы еще совсем мало.
До сих пор преобладают характеристики эволюции типов биоценозов по ведущим
типам организмов (например, псилофитовые биоценозы девона и т. п.), а не
по специфическим структурно-функциональным закономерностям (конструктивные
особенности, особые черты биотических связей, особые циклы круговоротов
веществ)” (1968, с. 193).
Такая оценка относится и к изученности эволюции биосферы, при исследовании
закономерностей которой мы сталкиваемся с принципиальными трудностями.
Эти трудности обусловлены тем, что биосфера как планетарная оболочка сложилась
более трех с половиной миллиардов лет назад и в течение этого громадного
промежутка времени оставалась постоянной в основных чертах своего строения,
организации круговорота веществ и потоков энергии и выполняемых биогеохимических
функциях. Формирование принципиальных особенностей биосферы восходит ко
времени, о котором мы не имеем прямых палеонтологических данных (Гиляров,
1973; Чернов, 1983; Медников, 1985; Левченко,
13
Старобогатов, 1986). И поэтому реконструкции начальных этапов эволюции
биосферы всегда гипотетичны. Практически нам известен лишь один этап в
развитии биосферы — это современная, сложно дифференцированная биосфера,
включающая несколько миллионов видов, каждый из которых выполняет в ней
различные функции: средообразующие, деструктивные, концентрационные, энергетические
и транспортные.
Такая биосфера могла сформироваться лишь в результате длительного развития.
Поэтому кажется странным, что сама правомерность постановки вопроса об
эволюции биосферы еще недавно вызывала дискуссии. Многие авторы, ссылаясь
на устойчивость биогеохимических функций биосферы, стабильность ее основных
энерго-вещественных параметров, отрицали эволюцию биосферы как целостной
системы. Исключение делалось только для современного этапа в ее истории,
когда под влиянием производственной деятельности человека в ней произошли
грандиозные изменения. Другие же ученые, отождествив биосферу с одним из
ее компонентов — органическим миром, даже филогенетические преобразования
отдельных таксономических групп пытались представить как показатели эволюции
биосферы, а коренные преобразования в органическом населении Земли в отдельные
геологические периоды рассматривали как время крупных перестроек самой
биосферы. В результате выделялось несколько критических периодов в истории
биосферы, когда происходило вымирание одних крупных таксонов и их замена
другими.
Сейчас становится очевидной ограниченность обеих точек зрения. Данные,
накопленные в различных областях геологии, географии и биологии, свидетельствуют
о том, что в ходе эволюции органических форм происходили и определенные
изменения в биосфере: изменялись ее биогеохимические функции, расширялась
зона распространения жизни, усложнялся биотический круговорот и т. д. В
то же время эти изменения не автоматически следовали за любым изменением
в органическом мире. Биосфера в целом оказывалась более устойчивой, чем
входящие в нее отдельные виды и ценозы.
Еще в 1928 г. В. И. Вернадский отметил противоречие между „эмпирическим
фактом” материально-энергетической устойчивости биосферы и установленным
Ч. Дарвином фактом исторических преобразований живого. Обдумывая это противоречие,
он сделал ряд важных выводов, имеющих большое значение для понимания эволюции
биосферы (см. подробнее гл. 2). Как подчеркивал Б. С. Соколов, вывод Вернадского
о живом веществе как мощной геологической силе, растущей с ходом времени,
не оставляет сомнения в том, что „речь идет об эволюции биосферы в самом
полном и точном смысле слова” (1983, с. 11).
Будучи в некоторых своих фундаментальных чертах строения геологически вечной,
биосфера претерпевает подлинные качественные и необратимые изменения во
времени. Опираясь на данные различных наук, изучающих историю биосферы,
можно сказать, что эволюция биосферы есть точно такой же „эмпирический
факт”, как и факт ее высокой устойчивости и стабильности. Своеобразие этой
эволюции заключается в том, что она происходит в пределах уже сложившегося
уровня организации
14
живого. К ее изменениям трудно применить критерии прогрессивного или регрессивного
развития. Вместе с тем это не были и флуктуации отдельных параметров биосферы
вокруг каких-то средних величин. Напротив, в преобразовании многих параметров
биосферы можно установить четко выраженные тенденции.
Высокая устойчивость биосферы делает понятным ту познавательную ситуацию
в ее изучении, когда до сих пор нет разработанного представления о сущности
эволюции этой формы организации жизни. Многокомпонентное строение биосферы,
разнообразие происходящих в ней абиотических и биотических процессов, непрерывные
преобразования ее отдельных компонентов и связей затрудняют выделение и
характеристику существенных черт в эволюции биосферы. Концентрируя внимание
на изменении самых общих параметров биосферы (общая биомасса, биогеохимические
и энергетические функции, информационная „емкость” биосферы, область распространения
жизни, структура потоков вещества и энергии и т.д.), можно упустить из
виду субстрат этих преобразований, и в первую очередь — организменный и
видовой уровни организации живого, а также преобразования, совершающиеся
на уровне экосистем (биогеоценозов) как основных структурно-функциональных
единиц биосферы. С другой стороны, существует опасность свести эволюцию
биосферы к сумме преобразований ее отдельных компонентов. В результате
многие работы, в названиях которых есть словосочетание „эволюция биосферы”,
посвящены фактически не развитию этой специфической формы организации жизни,
а необратимым изменениям климата, тектогенеза, вулканизма, осадкообразования,
почвообразования, флоро-фаунистических комплексов и даже филогенезу отдельных
таксономических групп. В этом отношении характерны многие статьи в книге
„Палеонтология и эволюция биосферы” (1983), содержащей труды XXV сессии
Всесоюзного палеонтологического общества.
Биосфера — это планетарная система, материально-энергетическая устойчивость
которой определяется всей совокупностью органических форм. Как всякая система,
она обладает собственными закономерностями эволюции, которые нельзя свести
к изменениям, вызванным эволюцией той или иной таксономической группы.
Источником развития биосферы выступают противоречивые взаимодействия между
всем живым и косным веществом в поверхностной оболочке Земли. „Разрешение
этого противоречия в ходе обменных процессов между организмами и окружающей
их средой обеспечивает процесс развития биосферы как целостной материальной
системы” (Гирусов, 1976, с. 120). Ведущая роль в этом взаимодействии принадлежит
жизни. Органический мир в целом, а не отдельные группы растений и животных,
детерминирует основные параметры биосферы. Поэтому необходимы не частные,
а интегральные характеристики для оценки результатов взаимодействия эволюирующей
биостромы с косным веществом биосферы.
Такой подход был намечен в трудах В. И. Вернадского, который стремился
найти характеристики, связывающие эволюцию органического мира с общим строением
и энергетикой биосферы. Эта связь наиболее ярко проявилась в изменении
геохимического строения биосферы, ее
15
общей биомассы и биопродуктивности, энергетики, концентрационных биогеохимических
функций, информационной „емкости” и, наконец, общепланетарного биотического
круговорота.
Жизнь, возникнув в „доактуалистическом” периоде истории Земли, быстро в
геологическом масштабе времени „растеклась” по поверхности нашей планеты.
В ходе эволюции биосферы постепенно все геологические и геохимические процессы
превратились в биогеологические и биогеохимические. Это объяснялось все
увеличивающимся давлением жизни на окружающую среду, усложнением взаимодействий
между живым и косным веществом. Следы средопреобразующего воздействия жизни
зафиксированы в геохимическом строении биосферы: в газовом составе атмосферы,
в химическом и минералогическом составе литосферы, в структуре и текстуре
образующих ее пород. Решающая роль живого сказывается в детерминации химической
активности природных вод и общего термодинамического баланса биосферы.
Переход от восстановительной атмосферы к кислородной был центральным событием
в геохимической эволюции биосферы, вызвавшим грандиозные изменения в гидросфере
и литосфере. Эволюция живого привела к постепенному уменьшению кислотности
вод и превращению их в щелочные.
Продукты жизнедеятельности организмов в результате литогенеза превратились
в залежи полезных ископаемых и в осадочные породы. Возможно, даже гранитная
оболочка Земли представляет собой метаморфизированные остатки „былых биосфер”.
Преобразование геохимической структуры биосферы под влиянием эволюции живого
вещества отразилось и в эволюции минералообразования, формировании залежей
полезных ископаемых, эволюции почв. Таким образом, образование четырех
планетарных оболочек Земли — атмосферы, гидросферы, литосферы и педосферы
— является итогом жизнедеятельности организмов и их эволюции.
Рост биомассы и продуктивности биосферы вел к прогрессивному накоплению
запаса превращаемой энергии в поверхностных оболочках Земли и тем самым
к уменьшению „производства” непревращаемых форм энергии в земных условиях.
В результате накопления продуктов жизнедеятельности организмов увеличивалась
негэнтропия биосферы. По мере развития органического мира происходило обогащение
живого, биогенного и косного вещества биосферы аккумулированной солнечной
энергией. Ее аккумуляторами стали не только горючие ископаемые, но и все
„вадозные” минералы, в которых находится большая часть энергии, накопленной
в земной коре.
В ходе развития биосферы непрерывно нарастала дифференцированность ее строения.
Это проявлялось в увеличении видового разнообразия биосферы, формировании
ее физико-географической зональности, в возникновении геохимических барьеров
и т.д. Иными словами, происходило увеличение неорганической и биологической
информации. Рост информации в биосфере был связан прежде всего с тем, что
эволюция органического мира, как отмечал еще Ч.Дарвин (1859), в целом идет
в сторону достижения наибольшей „суммы жизни”, которая возможна лишь при
16
наибольшем разнообразии видов биологического сообщества. Возрастание информационной
„емкости” биосферы, т. е. усложнение ее структуры и взаимодействий между
ее компонентами, резко повышало целостность биосферы, степень ее автономности
от космоса и от других геологических оболочек Земли.
Одним из показателей эволюции биосферы служат также изменения в концентрационных
функциях живого вещества. Среди них – усиление одних концентрационных функций
и ослабление других, появление качественно новых функций, перенос отдельных
концентрационных функций с одной группы организмов на другую и т. д. Эволюция
концентрационных функций проявилась как в увеличении содержания в организмах
разных химических элементов, так и в изменении соотношений между ними.
Изменения концентрационных функций живого вещества отразились в формировании
месторождений полезных ископаемых. Время образования многих рудных месторождений
совпадает с периодом доминирования в биосфере организмов с повышенным содержанием
этих
металлов.
Однако общая значимость названных показателей эволюции биосферы становится
понятной только в рамках постановки проблемы эволюции биосферы с точки
зрения преобразования биологического круговорота как основной формы существования
жизни. Эволюция биосферы представляет собой прежде всего процесс „расширения,
углубления и интенсификации биотического круговорота веществ”, и поэтому
„особенности биологических явлений на разных уровнях, различия отдельных
особей, видов и групп организмов прежде всего следует рассматривать как
приспособление, обеспечивающее этот круговорот” (Наумов, 1964, с. 138).
Уже развитие примитивных форм жизни вело к возникновению особых биогенных
циклов в сложных круговоротах вещества и потоках энергии на нашей планете.
По мере захвата жизнью все новых и новых регионов эти циклы приобретали
общепланетарный характер. В процессе эволюции органического мира происходила
взаимная „пригонка» видов таким образом, чтобы подольше сохранить в круговороте
вещества, используемые в метаболизме животных и растений, и повысить скорость
их круговорота. Это обеспечивалось за счет интенсификации окислительных
процессов и ускорения процессов обновления живого вещества. Эволюция живого
была „запрограммирована” на то, чтобы при сохранении биотического круговорота
химических элементов „выделялось больше кислорода, фиксировалось больше
энергии и могло образовываться все больше живого вещества” (Вудвелл, 1972,
с.41).
Важнейшими событиями в эволюции биосферы, по-видимому, были возникновение
и эволюция основных способов питания (хемотрофного, автотрофного и гетеротрофного),
типов экологических взаимодействий (хищничество, паразитизм, конкуренция,
кооперация) и становление биотического круговорота, осуществляемого продуцентами,
консументами и редуцентами. Возникший на заре жизни биотический круговорот
продуцентов и редуцентов из числа бактерий, низших грибов и других древних
групп постоянно усложнялся за счет встраивания в него
17
все новых и новых звеньев, расширения арсенала используемых химических
элементов, минеральных веществ и энергетических источников. Таким образом,
эволюция биосферы проявляется прежде всего в расширении сферы действия
биотического круговорота и в усложнении его структуры. Это обеспечивается
выработкой наиболее устойчивой макроструктуры жизни, возрастанием многообразия
органических форм и усиления их преобразующего воздействия на окружающую
среду. Эволюция геохимического строения биосферы, ее биомассы и биопродуктивности,
биогеохимических функций, информационной „емкости” и „энерговооруженности”
служит показателем эволюции биотического круговорота. Благодаря последней
возрастает целостность и организованность биосферы, совершенствуются механизмы
ее саморегуляции.
1.3. Взаимосвязь эволюции видов и развития биосферы
Эволюция биотического круговорота является наиболее общим и специфическим
показателем исторических преобразований жизни на биосферном уровне ее организации.
Однако нельзя понять эволюцию биосферы без изучения количественной и качественной
роли отдельных видов в ее геохимических и энергетических процессах, без
исследований исторических преобразований биогеоценозов. Именно усложнение
трофических связей внутри экосистем, возрастание зависимости между продуцентами,
консументами и редуцентами приводят к тому, что минеральные соединения,
выхваченные из геологических циклов организмами, обнаруживают тенденцию
к сохранению в биотических циклах.
В свою очередь эволюция биогеоценозов не может быть понята вне эволюции
составляющих ее популяций и видов. Образование биоценозов, подчеркивал
М. С. Гиляров, зависит прежде всего от межвидовой конкуренции, так как
сохраняются прежде всего те виды, которые лучше всего адаптированы к данному
биотическому сообществу и его абиотическим факторам. „Вид и биоценоз состоят
из одних и тех же структурных единиц: из популяций» (Гиляров, 1983а, с.
83). Энергетическая эффективность планеты и роль живого вещества в аккумуляции
солнечной энергии и в планетарном круговороте веществ возрастает благодаря
дифференциации сообществ и популяций, специализации жизненных форм и увеличению
числа экологических ниш.
Преобразования на организменном и популяционно-видовом уровнях организации
живого, детерминируясь сложными внутрипопуляционными и внутрибиоценотическими
взаимодействиями, в свою очередь выступают в качестве важнейших факторов
эволюции, протекающих на надвидовых уровнях организации живого. Так, повышение
в ходе мегоарогенеза общей активности животных (и прежде всего способность
к разумному поведению) сыграло важную роль в интенсификации биотического
круговорота, что обеспечивалось развитием нового принципа преодоления неблагоприятных
условий внешней среды. Там, где раньше выживание было возможно только с
помощью инактивации или же развития пассивных приспособлений, теперь создается
способность к поиску
18
оптимальных условий или к активному преодолению вредных условий. При дисперсии
видов по различным экосистемам особенно важными были активные миграции,
осуществляемые животными с высокоразвитой нервной системой и способностью
к сложным локомоторным движениям.
Активность отдельных организмов оказывается важнейшим регулятором функционирования
и эволюции биосферы. Как отмечал С. С. Шварц (1976), эволюция организмов
приводит к изменению структуры биогеоценозов, а в конечном итоге и биосферы,
так как свойства последней в значительной степени определяются слагающими
ее биогеоценозами. Освоение новых зон жизни и овладение новыми ресурсами
вещества и энергии всегда были следствием эрогенных преобразований организмов.
Появление полиареальных видов и широкие миграции млекопитающих, птиц, насекомых,
рыб, головоногих обеспечивали усиление обмена веществом и энергией между
различными биогеоценозами. Тем самым первооснова этого глобального по своей
сути процесса эволюции биосферы заключается в прогрессивных преобразованиях
организмов.
Об этом свидетельствуют результаты сравнительных исследований геохимических
функций у беспозвоночных, выступающих в качестве инициаторов освоения экологических
зон, и у их функциональных последователей - позвоночных (Симкин, 1969).
Беспозвоночные, как правило, представлены более специализированными формами
в осуществлении этих функций. Их средообразующая роль осуществляется благодаря
большему разнообразию видов и огромной численности популяций. При этом
интенсивность геохимической функции отдельной особи у беспозвоночных животных
оказывается значительно ниже. Филогенетическая дивергенция ведет к формированию
новых экологических комплексов и эволюционных взаимодействий (Wahlert,
1978).
Методологической установкой при изучении взаимосвязи эволюции видов с эволюцией
биосферы служит положение о том, что вид - не только элементарная единица
эволюционного процесса, но и элементарная геохимическая и энергетическая
единица биосферы (Камшилов, 1961; Наумов, 1964, 1967, 1972). Более того,
каждый вид является фактически уникальным по своим экологическим функциям.
Вот почему не удается, как правило, найти равноценную замену виду, вымершему
в результате антропогенной деятельности. Особенно велика биогеохимическая
и энергетическая роль в экосистемах ключевых видов, вымирание которых ведет
к коренной перестройке биотических сообществ.
В связи с этим преобразование химического состава и геохимической энергии
популяции представляет собой и элементарное явление в эволюции биосферы.
Образование каждого вида, писал Н. В. Тимофеев-Ресовский (1958), есть шаг
в освоении организмами биосферы. Согласно закону Г. Ф. Гаузе (1934а), становление
нового вида невозможно без формирования специфической экологической ниши,
без овладения ресурсами, которые ранее не использовались. Диверсикационные
процессы видообразования повышают общую адаптивную пластичность данной
таксономической группы и тем самым увеличивают „общую интенсивность использования
организмами поверхности Земли, повышая степень дифференциации биомассы
в биосфере” (Тимофеев-Ресовский, 1958, с. 333).
19
Для понимания эволюции биосферы важно получить максимальное количество
сведений об общей биомассе отдельных видов, их химическом составе и геохимической
энергии. Эти видовые признаки характеризуют участие того или иного вида
в энерготрофических процессах биосферы. Поэтому важно процессы видообразования
изучать не только с точки зрения изменения морфологических и физиологических
признаков организмов, но и с учетом их веса, элементного состава и геохимической
энергии. Уже первые исследования такого рода, проведенные в 20—30-х годах
под руководством В. И. Вернадского и А. П. Виноградова, позволили получить
точные данные об участии отдельных видов в осуществлении биогеохимических
функций биосферы. Установление геохимической определенности видов не только
дополняет традиционные представления о процессах видообразования и их критериях,
но и позволяет изучать эти процессы как элементарные явления в эволюции
биосферы. Не случайно широкое распространение среди советских и зарубежных
биологов получили представления о том, что появление практически каждой
новой группы растений означает усовершенствование способности использовать
энергию солнца, а появление новых групп животных связано с формированием
механизмов дальнейшей утилизации этой энергии (Бей-Биенко, 1972; Ничипорович,
1973; Шварц, 1973; Родендорф, Жерехин, 1974; Paulus, 1978; Wahlert, 1978;
Вольские, 1986, ими. др.). Ниже будет показано (гл. 3), что в некоторых
работах удалось довести результаты изучения микроэволюционных преобразований
геохимических признаков до уровня биосферных исследований. В частности,
было доказано, что адаптированность организмов к геохимической среде предопределяет
и интенсивность выполнения ими биогеохимических функций. От степени приспособленности
организмов зависит скорость роста их биомассы, а следовательно, и объем
химических элементов, вовлекаемых ими в биотический круговорот. Приспособленность
организмов сказывается и на их воздействии на окружающую среду. Высокоадаптированные
организмы, как правило, обладают наибольшей геохимической энергией и играют
более важную роль в биогеохимических процессах биосферы по сравнению с
менее адаптированными. Исследования по геохимической экологии подошли близко
к исследованию взаимосвязи процессов микроэволюции и видообразования с
эволюцией биосферы. Изучение причинных зависимостей между микроэволюцией
и условиями геохимической среды — важный путь к правильному решению этой
задачи.
Взаимосвязь преобразований на организменном уровне и на уровне экосистем
можно видеть и на примере перехода свободноживущего вида к паразитизму.
Упрощение организма, потеря им ряда органов и функций связаны с перераспределением
этих функций между другими членами экосистем, т. е. с усложнением экосистемных
взаимодействий. Применительно к биосфере даже регрессивная эволюция отдельных
форм и вымирание организмов могут быть важными факторами ее прогрессивной
эволюции. Исчезновение биологических сообществ нередко является необходимой
предпосылкой для появления новых форм, которые более интенсивно используют
ресурсы окружающей среды.
20
Наряду с влиянием эволюции видов на процессы преобразования биосферы необходимо
учитывать и обратное воздействие структуры биосферы на эволюцию каждого
вида. Как подчеркивалось не раз на 20-м Международном филогенетическом
симпозиуме, посвященном специально проблемам коэволюции, экологические
взаимодействия всецело определяют коэволюционные процессы, доминирующие
в биосфере (см.: Co-Evolution, 1978). Практически каждый вид, каждая более
крупная таксономическая группа эволюируют в тесной связи с другими таксонами.
Примером могут служить коэволюция обитателей коралловых рифов, фитофагов
и высших растений, цветковых растений и их опылителей, наземных позвоночных
и растительности и т.д. Фактически эволюция любого таксона, любой жизненной
формы растений или животных может быть понята только с точки зрения биосферы,
куда они входят на правах элементарных компонентов. Виды связаны между
собой не только энерго-трофическими отношениями, но и общностью становления
и коэволюции в рамках единой глобальной экосистемы — биосферы.
О зависимости процессов видообразования от структуры биосферы свидетельствует
и тот факт, что после периодических массовых вымираний организмов биоценотический
покров восстанавливается в сходных жизненных формах, хотя и в ином видовом
составе. Это объясняется, по-видимому, наличием в биосфере определенных
экологических лицензий (в смысле Я. И. Старобогатова, 1988), побуждающих
конвергентно эволюировать представителей различных филумов. В пользу утверждения
о детерминирующем влиянии структуры биосферы на процессы видообразования
говорят не только факты параллельной эволюции на разных континентах, но
и результаты моделирования на ЭВМ мегафилогенеза позвоночных (Меншуткин,
1977).
Некоторые филогенетические закономерности эволюции становятся более понятными
с учетом принципов строения и функционирования биосферы. Так, например,
увеличение размеров тела в филогенезе животных может быть оценено как результат
эволюции живого в сторону все более эффективного выполнения биогеохимических
функций. Увеличение размеров тела, как правило, ведет к повышению активности
животных в захвате пищи, в борьбе с конкурентами, а следовательно, и к
повышению скорости переноса вещества в биосфере. Усиление же цефализации,
т. е. разумного способа поведения животных, не только обеспечивало быстрое
приспособление к меняющимся условиям существования, но и совпадало с общей
направленностью эволюции жизни к усложнению биогенной миграции атомов в
биосфере, согласно биогеохимическим принципам, сформулированным Вернадским.
Эти принципы определяют главные направления в эволюции жизни, которая идет
в сторону создания форм, усиливающих влияние жизни на окружающую среду
и увеличивающих миграцию атомов в биосфере.
Исследование взаимодействий между процессами видообразования и эволюцией
биосферы только начинается. Имеющиеся данные позволяют судить лишь в общих
чертах о закономерностях этих взаимодействий. Однако ясно, что эволюция
видов, будучи детерминирована
21
структурой биосферы, в свою очередь является главным фактором ее преобразования.
И напротив, эволюционные преобразования биосферы предопределяют будущие
изменения отдельных видов, что в целом ведет к ограничению их эволюционных
возможностей. По мере возрастания целостности и автономности биосферы уменьшается
вероятность случайных, независимых преобразований отдельных видов. Видообразование
все больше сливается в единый, целостный процесс преобразования биостромы.
Возрастает и организованность эволюционного процесса, который все больше
контролируется сложившейся в биосфере системой круговорота веществ и потоков
энергии.
В связи с этим неизбежно возникает вопрос о соотношении и взаимодействии
факторов эволюционных преобразований на различных уровнях организации живого.
Этот вопрос крайне дискуссионен. До сих пор нет „глубоких разработок проблемы
механизмов и закономерностей временного преобразования сообществ и их соотношения
с процессами видообразования, микро- и макроэволюции... ” (Чернов, 1983,
с. 472). Одни авторы стремились провести близкую аналогию между процессами
видообразования и эволюции сообществ, утверждая, что последняя протекает
под действием „отбора сообществ”, „естественного отбора второго порядка”,
„межвидового отбора”, „межъярусной и межфитоценозной борьбы за существование”,
„борьбы сообществ”, „группового отбора” и т. д. (Ревердатто, 1935; Соколов,
1948; Ярошенко, 1961; Dunbar, 1968: May, 1976, 1978, и др.). Утверждалось
даже, что геометрическая прогрессия размножения, борьба за существование,
естественный отбор, хорошо известные как факторы видообразования, являются
в то же время факторами формирования биоценозов (Гиляров, 1980).
Другие авторы считают, что процессы видообразования и эволюции экосистем
не только самостоятельны, но и полностью независимы друг от друга и поэтому
необходимо искать специфические факторы и движущие силы эволюции экосистем.
Так, М. А. Голубец пишет: „В биосфере нет борьбы за существование между
биогеоценозами, и биосфера лишена свойства естественного отбора наиболее
устойчивых экосистем. Биогеоценозы не рождаются и не умирают. Эволюцией
этих систем движут не только внутренние (биотические, присущие живым организмам),
но и какие-то другие силы” (1982, с. 62).
В числе специфических факторов эволюции биосферы, как правило, называют
резкие изменения абиотических процессов: колебания активности солнечного
и космического излучений, тектогенез, орогенез, дрейф материков, вулканизм,
оледенения и т. д. К ведущим факторам эволюции биосферы относят воздействие
электромагнитных космических полей, изменение суммарной солнечной активности,
увеличение радиоактивных элементов в поверхностных оболочках Земли, резкие
изменения климата и т. д. (Будыко, 1975: Пресман, 1976; Красилов, 1977:
Alvarez, 1979; Хохряков, 1981, и др.).
Вряд ли кто-то будет отрицать в наше время, что существует необозримое
множество процессов, воздействующих на эволюцию отдельных популяций и видов,
флор и фаун целых областей и даже на эволюцию всего живого на Земле. Они
могут быть региональными, как например
22
засоление или опреснение данного водоема, распространение вулканических
газов после подводного извержения. Но эти факторы могут носить и характер
процессов планетарного масштаба, действие которых развертывается в течение
геологического времени (изменение уровня океана, глобальные изменения климата,
изменение состава атмосферы и т. п.). Независимо от того, каково происхождение
этих факторов — дальнекосмическое, солярное или геологическое, все они
по сути дела являются факторами, внешними по отношению к биосфере, действием
которых вряд ли возможно объяснить глобальный процесс ее преобразований
на протяжении миллиардов лет.
Что же касается периодических колебаний космических излучений, и прежде
всего изменений, вызванных циклическими изменениями солнечной активности
и движениями солнечной системы вокруг центра Галактики, то есть все основания
предполагать, что жизнь еще на начальных этапах своего становления приспособилась
к подобным изменениям, так же как она приспособилась к периодическим сменам
дня и ночи или времен года. Несмотря на то что во всем мире сейчас развернуты
крупномасштабные исследования влияния космических факторов на динамику
численности организмов, их миграцию, заболеваемость и т.д. (см.: Чижевский,
1973), практически нет надежных данных о влиянии этих факторов на эволюцию
организмов. Важно учитывать и то, что с момента образования биосферы космические
факторы могли воздействовать на эволюцию организмов, лишь преломляясь через
сложную цепь внутрипопуляционных и биоценотических взаимодействий. Если
же обратиться к разнообразным гипотезам о якобы решающем воздействии космических
факторов на эволюцию органического мира в определенные периоды его истории,
то подобные гипотезы, как правило, не подкреплены ни астрофизическими,
ни палеонтологическими данными.1 Хотя значению космических факторов
в истории биосферы была посвящена не одна монография и даже конференция
(например,
совещания „Космические факторы и эволюция органического мира”, Москва,
1974; „Космическая антропоэкология”, Ленинград, 1984), в решении этого
вопроса практически нет прогресса, подобно тому, как многочисленные труды
и симпозиумы по проблемам экзобиологии не продвинули нас вперед в доказательстве
реального существования жизни вне земных условий.
Наиболее удачная попытка выделить основные факторы эволюции биосферы с
учетом взаимодействия внешних и внутренних факторов ее развития принадлежит
М. М. Камшилову (1979). Он называл три группы факторов: взаимодействие
живого и неживого в биосфере; своеобразие отношения биосферы к абиотической
среде и особенности взаимоотношений между организмами. При этом он отмечал,
что по мере возрастания упорядоченности, сложности биосферы, приобретения
ею способности к саморегуляции значение внутренних биотических факторов
все более возрастало.
Далее мы еще не раз будем обращаться к вопросу о взаимодействиях между
эволюционными преобразованиями на популяционно-видовом
1 Космическими факторами нельзя объяснить и крупные ароморфозы.
23
и биосферном уровнях организации живого. Здесь отметим только, что, несмотря
на отличия в детерминации эволюции популяций и экосистем, нет оснований
отрывать их друг от друга. Набор каузальных факторов по существу один и
тот же, меняются лишь соотношения между ними. Абиотические агенты и межвидовые
взаимодействия, будучи второстепенными, внешними факторами в адаптивных
преобразованиях популяций, становятся ведущими в эволюции сообществ. В
свою очередь основные факторы микроэволюции на экосистемном уровне действуют
лишь опосредованно, через эволюционные изменения видов и популяций. Пока
нет оснований предполагать существование иного механизма исторических преобразований
живого, кроме факторов, установленных при экспериментальных исследованиях
адаптивных преобразований популяций. „Движущими силами, регулирующими структуру,
а следовательно, и эволюцию целых биоценозов, являются и здесь внутренние
взаимоотношения между элементами биоценоза. В этом эволюционном процессе
широкого масштаба перестраиваются постепенно флора и фауна... В основе
всех этих преобразований лежит, однако, эволюция наиболее интегрированных
систем — организмов как индивидуальных представителей видов” (Шмальгаузен,
1968, с. 175).
Таким образом, познание движущих сил и закономерностей эволюции биосферы
— это прежде всего установление взаимосвязи эволюции органического мира
с эволюцией биосферы, так как основные параметры последней (общая биомасса,
продуктивность, химический состав живого вещества, общий запас аккумулированной
энергии и т.д.) являются интегральным результатом аналогичных признаков
вида. Популяция выступает не только элементарной единицей эволюции, но
и элементарной (в биогеохимическом, энергетическом и трофическом отношениях)
единицей биогеоценозов. Необратимые преобразования популяции, затрагивая
ее биогеохимические и энергетические функции, так или иначе сказываются
на функционировании биогеоценозов. Именно преобразования химического состава
популяции, ее геохимической функции и роли в энерготрофической структуре
экосистемы означают элементарный шаг в эволюции биосферы. И хотя масштаб
одного и того же явления — образования видов — оказывается различным для
популяционно-видового и биосферного уровней организации жизни, причины,
обусловливающие решающий момент в адаптивных преобразованиях популяций
и элементарное явление в эволюции биосферы, по-видимому, одни и те же.
В противоречивых взаимодействиях эволюции организмов и эволюции биосферы
вырабатывались наиболее эффективные механизмы эволюции живого (Колчинский,
1985а, 1985б). Тем самым биосфера создает условия, которые в дальнейшем
определяют основные направления ее развития. И только в рамках этого процесса
становится понятным, что эволюция жизни — это внутренне детерминированный
процесс.
24
1.4. Этапы и направления в изучения эволюции биосферы
Начатые В. И. Вернадским исследования эволюции биосферы проводились в
СССР широким кругом ученых. Среди них – биогеохимики (А. П. Виноградов,
В. В. Ковальский, Я. В. Самойлов), географы (М. И. Будыко, И. П. Герасимов,
А. А. Григорьев, Б. Б. Полынов, В. Б.Сочава), геохимики и геологи (Н. Б.
Вассоевич, Н. Н. Верзилин, Ю. А. Жемчужников, А. В. Лапо, Б. Л. Личков,
К. И. Лукашев, А. И. Перельман, А. Б. Ронов, А. В. и С. А. Сидоренко, Н.
М. Страхов, А. Л. Яншин), геоботаники (М. А. Голубец, Е. М. Лавренко, В.
Н. Сукачев, И. А. Титов), математики (А. А. Александров, В. А. Костицын,
Н. Н. Моисеев, Ю. М. Свирижев, А. М. Тарко), палеонтологи (А. Г. Вологдин,
Л. Ш. Давиташвили, Д. Н. Соболев, Б. С. Соколов), почвоведы (В. Р. Вильямс,
В. А. Ковда, А. А. Роде, Е. И. Шилова), фитофизиологи (А. А. Ничипорович),
философы (Э. В. Гирусов, П. А. Водопьянов), экологи (А. М. и М. С. Гиляровы,
М. М. Камшилов, К. М. Хайлов, Ю. И. Чернов, С. С. Шварц) и др.
Столь широкий фронт исследований выдвигает особые трудности при выделении
направлений в развитии проблемы эволюции биосферы. Трудности усугубляются
еще тем, что, несмотря на участие в этих исследованиях представителей различных
отраслей естествознания, сама проблема эволюции биосферы находится еще
на стадии постановки и первых попыток ее решения. До сих пор нет твердо
очерченных направлений в ее разработке. Данной проблемой, как правило,
занимаются лишь в контексте глобальных задач, связанных с дальнейшей разработкой
учения о биосфере и познанием ее современного состояния. В существующих
сейчас биогеохимических, физико-географических, кибернетических и термодинамических
подходах к изучению биосферы удельный вес эволюционной проблематики крайне
незначителен.
Основными принципами при выделении этапов в развитии любой науки или научной
проблемы служат крупные открытия, создание фундаментальных обобщающих трудов,
внедрение новых методов исследования. При выделении этапов в развитии проблемы
эволюции биосферы наряду с этими общепризнанными критериями периодизации
науки следует учитывать также количество трудов, связанных с ее разработкой,
распространенность идеи эволюции биосферы среди естествоиспытателей различных
специальностей, формирование новых эволюционных подходов к изучению различных
сторон строения и функционирования биосферы. Руководствуясь этими соображениями,
целесообразно выделить три периода в разработке анализируемой проблемы.
Первый период, начавшийся с опубликования „Биосферы” В. И. Вернадского
(1926а), можно оценить как время утверждения этого учения в науке. Этот
период охватывает около двух десятилетий. Он отличался доминированием биогеохимического
подхода к разработке учения о биосфере и закономерностях ее эволюции. В
эти годы проводились историко-сравнительные исследования химических признаков
организмов и химических особенностей ископаемых остатков, а также древних
биогенных пород. В этих исследованиях преимущественно участвовали ученики
В. И. Вернадского. Вместе с тем в эти же годы изучалось влияние
25
биогеохимической деятельности организмов на процессы выветривания, осадкообразования,
почвообразования и образование залежей полезных ископаемых. Полученные
данные впоследствии использовались при разработке проблемы эволюции биосферы
и построении первых гипотез о сопряженной эволюции биологических и неорганических
процессов.
Второй период охватывает 40—60-е годы. Важнейшим событием этих лет стало
создание В. Н. Сукачевым учения о биогеоценозах как основной структурно-функциональной
энергетической и геохимической единице биосферы. Это учение положило начало
экологическому направлению в изучении биосферы, в рамках которого предпринимались
первые попытки поставить вопрос об основных тенденциях и главных этапах
эволюции биогеоценотических систем. В эти же годы был опубликован ряд обобщающих
работ, где эволюция биосферы связывалась с преобразованием геохимических
ландшафтов и биогеохимических провинций, изменением газового состава атмосферы
и химизма природных вод, преобразованием осадочных пород литосферы под
влиянием организмов и продуктов их жизнедеятельности. Среди исследований
этих лет широкое развитие получили работы по геохимическим факторам эволюции
биосферы и эволюции ее биогеохимических функций.
Начало третьего, современного, периода в развитии проблемы эволюции биосферы
может быть датировано первой половиной 70-х годов, когда общенаучная значимость
этой проблемы получила всеобщее признание. Из проблемы, привлекающей внимание
геологов, геохимиков, почвоведов и отчасти географов и экологов, она превратилась
в важнейшую глобальную задачу современности, в решение которой включались
специалисты различных отраслей науки. Широкое развитие получают информационные
и энергетические трактовки эволюции биосферы. Строятся математические модели
эволюции биосферы и ее отдельных составляющих. Становится очевидной необходимость
разработки учения об эволюции биосферы как важнейшего раздела эволюционной
теории. Первые шаги в этом направлении были предприняты в трудах М. И.
Будыко, М. М. Камшилова, Н. В. Тимофеева-Ресовского и С. С. Шварца. Именно
в эти годы стал осуществляться синтез учения о биосфере с современной эволюционной
теорией.
В дальнейшем мы постараемся рассмотреть историю изучения биогеохимических
закономерностей эволюции биосферы и значение полученных результатов для
современной эволюционной теории. В связи с этим мы проанализируем лишь
следующие направления: 1) изучение роли геохимических факторов эволюции
биосферы; 2) исследование химического состава современных организмов, а
также древних биогенных пород с целью выяснения эволюции биогеохимических
функций биосферы; 3) изучение влияния эволюции органического мира на эволюцию
химического состава атмосферы, гидросферы, литосферы, на процессы почвообразования,
литогенеза и т. д.; 4) сравнительные исследования биологической продуктивности,
биомассы и биогеохимических циклов в современных адаптивных зонах и геохимических
ландшафтах для постановки вопроса об эволюции биотического круговорота;
5) изучение энергетических и информационных аспектов эволюции биосферы.
26
Глава 2
В. И. ВЕРНАДСКИЙ
ОБ ЭВОЛЮЦИИ БИОСФЕРЫ
Весь комплекс вопросов, связанных с изучением закономерностей функционирования
и эволюции биосферы, был впервые поставлен в трудах В. И. Вернадского.
Его взгляды претерпели определенную эволюцию в течение почти тридцатилетней
разработки учения о биосфере. В книгах „Биосфера” (1926а) и „Очерки геохимии”
(1927а) он главное внимание уделял исследованию постоянства фундаментальных
черт организованности биосферы и ее биогеохимических функций. Это дало
основание некоторым исследователям научного творчества Вернадского считать,
что в вопросе об эволюции биосферы он занимал отрицательную позицию, считая
биосферу в целом неизменной в течение геологического времени. При этом
игнорируется тот факт, что уже в работах последующих лет (1928, 1931, 1944),
и прежде всего в посмертно изданных трудах „Химическое строение биосферы
Земли и ее окружения” (1965) и „Размышления натуралиста” (1977), Вернадский
неоднократно обращался к изучению существенных преобразований в биогеохимических
круговоротах вещества и потоков энергии в биосфере. „Эволюционный процесс
живых веществ, — подчеркивал Вернадский (1977, с. 18), — непрерывно в течение
всего геологического времени охватывает всю биосферу и различным образом,
менее резко, но сказывается, на ее косных природных телах. Уже по одному
этому мы можем и должны говорить об эволюционном процессе самой биосферы
в целом” (курсив наш. – Э. К.). Он не раз отмечал, что эволюция органического
мира придала биосфере облик, который не свойствен всем другим геологическим
оболочкам Земли. При этом ,,в ходе геологического времени растет мощность
выявления живого вещества в биосфере, увеличивается его в ней значение
и его воздействие на косное вещество биосферы” (там же). Из опубликованных
недавно выдержек из дневников В. И. Вернадского видно, что уже в момент
формирования учения о биосфере он ясно понимал, что „оно представляет другую
сторону, другой аспект – эволюционного учения” (цит. по: Неаполитанская,
1986, с. 18).
В последнее десятилетие учение о биосфере привлекает внимание исследователей
самых различных специальностей, в том числе биологов-эволюционистов и историков
науки. Все большее количество статей и монографий освещает отдельные аспекты
этого учения и его современное состояние (Личков, 1948; Камшилов, 1970,
1974; Перельман, 1973; Вассоевич, Иванов, 1977, 1983а, 1983б, 1984; Шипунов,
1980; Яншин, 1981, 1986; Голубец, 1982; Лапо, 1982, 1987; Будыко, 1984;
В.И.Вернадский и современность, 1986; Сытник и др., 1988; Щербак, 1988;
В. И. Вер-
27
надский и современная наука, 1988; Научное и социальное значение..., 1989,
и др.). В 1982 г. вышла прекрасная книга И. И. Мочалова, в которой на основе
анализа научных трудов Вернадского и архивных материалов прослежен путь
формирования естественнонаучных и философских воззрений ученого, в том
числе и его учений о биосфере и ноосфере. Но, к сожалению, за исключением
статьи, опубликованной нами в 1981 г., до сих пор нет трудов, в которых
бы исследовались эволюционные взгляды Вернадского и его идеи об эволюции
биосферы. Лишь частично этот вопрос затрагивался в ряде работ (Сытник и
др., 1987; Назаров, 1988). В этой главе мы постараемся в какой-то степени
заполнить этот пробел в историко-биологической и эволюционно-биологической
литературе и показать значение работ Вернадского для развития современной
эволюционной теории.
2.1. Идеи о взаимодействии эволюции видов
и эволюции биосферы
Единство процессов видообразования и эволюции биосферы. Вернадский впервые
обосновал идею об эволюции поверхностной оболочки Земли как целостном процессе
взаимодействия живого вещества и косной материи. Ведущая, активная роль
в этом взаимодействии принадлежит жизни, которая признавалась Вернадским
„великим, постоянным и непрерывным нарушителем химической косности поверхности
нашей планеты” (1926а, с. 26).
Вернадский не ограничился констатацией ведущей роли живого в эволюции биосферы,
а стремился найти „кипящий слой” элементарных преобразований биосферы.
Опережая широкое проникновение популяционных представлений в эволюционную
биологию, он уже в середине 20-х годов считал необходимым выделить вид
как главный носитель эволюции. Основная задача в изучении эволюции биосферы,
по мнению Вернадского, заключалась в установлении связи „эволюции видов
с механизмом биосферы, с ходом биогеохимических процессов” (1928, с. 234).
Существование подобной связи не вызывало у него сомнений, так как „основные
числа, характеризующие эти процессы, являются видовыми признаками, меняющимися
в процессе эволюции” (там же). К числу этих признаков он относил общую
биомассу вида, его химический состав и геохимическую энергию.
Позднее, конкретизируя эти мысли, Вернадский характеризовал эволюцию химической
и энергетической структур биосферы как интегральный результат преобразований,
происходящих на видовом уровне организации жизни. Факторы и движущие силы
видообразования, по-видимому, и представляют собой ведущие источники преобразования
всей биосферы. Поэтому необходимо исследовать, как „эволюция видов
переходит в эволюцию биосферы” (1977, с. 18). В связи с этим при постановке
проблемы эволюции биосферы Вернадский использовал основные результаты исследований
эволюции органического мира, в том числе и дарвинистские представления
о причинах видообразования.
28

Владимир Иванович Вернадский (1863 – 1945).
Вместе с тем он считал необходимым учитывать и детерминирующее влияние
структуры биосферы на эволюцию каждого вида. „Жизнь, — писал Вернадский,
- неразрывно связана с биосферой, и ее эволюция в значительной степени
определяется ее строением” (1928, с. 242). Изучение влияния структуры
биосферы на эволюцию видов, по мнению Вернадского, представляет определенный
интерес и для эволюционной теории, так как позволяет объяснить некоторые
закономерности биологической эволюции действием „целой совокупности
физических законов и механизмов функционирования биосферы” (там же, с.
243).
Таким образом, Вернадский впервые выдвинул задачу синтеза основных положений
эволюционной теории с созданным им учением о биосфере. С одной стороны,
для познания механизмов эволюции биосферы следует опираться на данные
эволюционной теории о субстрате эволюции органического мира и ее движущих
силах. С другой
стороны, изучение структуры биосферы должно способствовать углублению
знаний о закономерностях органической эволюции. Эти положения являются
ключевыми
для понимания взглядов Вернадского на сложные
29
взаимодействия эволюционных преобразований, происходящих на разных уровнях
организации жизни.
Об отношении Вернадского к дарвинизму. В той области эволюционной теории,
которая позднее получила название „микроэволюция”, Вернадский в целом придерживался
принципов селекционизма. Он правильно отмечал особенности дарвиновского
понимания субстрата эволюции. В учении об естественном отборе, писал Вернадский
в работе „Живое вещество” еще в начале 20-х годов,1 элементарным
носителем эволюции выступает не особь, а сообщество организмов. „Статистическое
представление
о живом... проникает в дарвинизм и эволюционное мировоззрение. Это то значение,
какое в них получает вид, заменяющий индивид” (Вернадский, 1978, с. 55).
При этом он привел дополнительные аргументы против монотипической концепции
вида, неоднократно подчеркивая, что даже в морфологически однородных видах
имеется множество разновидностей, отличающихся друг от друга по физиологическим,
этологическим и химическим признакам. И эти различия генетически закреплены.
Поэтому он считал невозможным ограничиться исследованиями химического состава
вида в целом. Необходимо „идти дальше вида, — писал Вернадский, — принимать
во внимание меньшие, чем вид или раса, группы организмов”, так как „биохимические,
экологические и элементарные группы играют огромную роль в геохимической
реакции организмов” (1978, с. 26—27). Геохимическое воздействие вида на
биосферу зависит также от его половой, возрастной и сезонной структур.
Вернадский отмечал, что по своему химическому составу, размеру, массе,
геохимическим функциям и скорости размножения внутривидовые группы сильно
различаются, и это „не проходит бесследно в геохимическом отношении” (там
же, с. 286).
Статистический характер дарвиновских законов импонировал Вернадскому тем,
что и в геохимических процессах „жизнь выражается совместным действием
отдельных организмов, их совокупностью. Мы в ней изучаем лишь статистические
законы и обобщения, связанные с жизнью” (Вернадский, 1927а, с. 47). В этих
процессах роль отдельной особи исчезает полностью и заметен лишь общий
геохимический и энергетический эффект их действия.
Вот почему внедрение статистического подхода к явлениям органической эволюции
оценивалось Вернадским как важнейшее достижение дарвинизма, равное по своей
значимости формированию статистически-вероятностных представлений в физике
и общественных науках. „Во второй половине XIX века в учении о борьбе за
существование формы статистических законов природы обрели научную почву
в научном мировоззрении” (Вернадский, 1978, с. 55). Для него были чужды
концепции о строго направленном и предопределенном ходе эволюции, в основе
которых лежали принципы жесткого детерминизма. Эволюция для Вернадского
была незапрограммированным, статистическим процессом.
1 Эта рукопись опубликована только в 1978 г.
30
В связи с этим он высоко оценивал теорию естественного отбора Ч.Дарвина,
называя ее „величайшим обобщением”, оказавшим огромное влияние на все мировоззрение
современного естествознания. В трудах Дарвина его привлекали строгая обоснованность
каждого положения фактами и доказательствами, стремление избегать натурфилософских
схем и спекулятивных рассуждений, от обилия которых так страдала биология
XIX в. Но в дарвинизме Вернадский видел преимущественно учение, объясняющее
процессы видообразования, но не раскрывающее движущие силы и закономерности
макроэволюции. Свои высказывания по основным проблемам макроэволюции Вернадский
формулировал очень осторожно, со многими ограничениями и оговорками, так
как не считал уместным вмешиваться в споры по проблемам, которые не входили
прямо в круг его специальных занятий.
Геохимическая трактовка вида и видообразования. Стремясь
связать эволюцию видов с эволюцией биосферы, Вернадский подчеркивал необходимость
изучать
видообразование не только с точки зрения преобразования морфологических
и физиологических признаков организмов, но и изменения их массы, элементного
состава и геохимической энергии. По его инициативе были начаты широкие
исследования элементного состава организмов, принадлежащих к различным
таксономическим группам (Вернадский, 19226, 1927в, 1929 и др.). Он предложил
оригинальную методику для определения биомассы, химического состава и геохимической
энергии у однолетних цветковых растений и у насекомых (1926б, 1926в),
не потерявшую свое значение и по сей день.
В результате проведенных исследований было получено большое количество
точных данных и сделан ряд обобщений, характеризующих участие отдельных
видов и других таксономических групп в осуществлении концентрационных функций
биосферы. Благодаря этим работам было показано, что вид, по выражению А.
П. Виноградова (1931), перефразировавшего известную формулировку В. Л.
Комарова, является морфологической системой, помноженной на геохимическую
определенность. Столь же видоспецифическими оказались и разнообразные формы
географической, сезонной, суточной и возрастной изменчивости химического
состава организмов.
Все эти исследования не только дополняли традиционные представления о критериях
вида и процессах видообразования, но и позволяли оценить видообразование
как элементарное явление в эволюции биосферы, выясняя, по словам Вернадского,
место каждого вида в общей миграции химических элементов в поверхностных
оболочках Земли.
Вернадский не раз подчеркивал, что хотя носители геохимической энергии
– организмы – дискретны, в совокупности они представляют собой единое целое,
занимающее определенное место в геохимических и энергетических процессах
биосферы.
Вернадский впервые поставил вопрос о геохимических факторах эволюции организмов.
Химическая дифференциация вида в значительной степени обусловлена различием
химической среды в пределах его ареала. В доказательство этого он ссылается
на работы Вебера, показавшие связь экологической химической изменчивости
лиственницы и бука
31
при переходе от низменных районов обитания к горным. Однако геохимические
факторы действовали не прямо, а косвенно, преломляясь через конституцию
каждого вида. Так, у лиственницы по мере перехода к низменным формам увеличивается
количество калия и фосфора, а у бука при снижении калия и фосфора увеличивается
содержание кремния и кальция.
Если влияние химического состава почв на видообразование растений Вернадский
считал в принципе установленным, то еще большего он ожидал от изучения
взаимосвязи между разнообразием химической среды и многочисленностью видов
насекомых. „Становится очень вероятным, - писал В. И. Вернадский в начале
20-х годов, - что значительная часть видов организмов, в частности растений,
является морфологическими видами, созданными благодаря химическим особенностям
той среды, на которой они живут, и того состава, какой они при этом получают”
(1978, с. 28). Несколько позже, опираясь уже на результаты многочисленных
исследований химического состава разных видов, Вернадский утверждал: „Судьба
организмов материально связана и может быть количественно охарактеризована
на фоне изменения биосферы и законов миграции ее атомов. Как химический
состав организмов определяет генетически связанные с ним породы и минералы,
так и обратно общие законы распространения и сочетания атомов в мертвой
природе закономерным образом обусловливают состав живого вещества” (Вернадский,
Виноградов, 1931, с. 149).
Борьба за существование как геохимический процесс биосферы. Борьбу за существование
Вернадский считал твердо установленным фактором эволюции, ведущая роль
в которой принадлежит противоречивым взаимодействиям внутри биологических
сообществ. „В природе, - писал Вернадский, — мы всегда имели биоценозы,
сообщества, качественный состав которых и количество организмов регулируются
борьбой за существование, приспособлением, явлениями солидарности и цепями
жизни” (1965, с. 306—307). Всякие попытки объяснить эволюцию видов или
биологических сообществ прямыми формообразующими действиями абиотических
агентов (например, гипотезу И. Вальтера об эксплозивных преобразованиях
морских организмов в результате изменения химизма вод) он считал ошибочными.
Вернадский попытался дать геохимическую и энергетическую оценку процессов
борьбы за существование. Он ввел специальное понятие „геохимическая энергия
организмов” (1925), которое, по его мнению, предопределяет эффект воздействия
данного вида на окружающую среду. С повышением геохимической энергии ускоряется
осуществляемая данным видом биогенная миграция атомов в биосфере и тем
самым усиливается его участие в биогеохимических циклах биосферы. В связи
с этим количественные определения геохимической энергии отдельных видов
Вернадский считал важнейшей задачей современного естествознания. Суть этой
задачи состоит в том, чтобы в математических формулах выразить связь геометрической
прогрессии размножения с потенциальной скоростью заселения данным видом
всей планеты. Он подчеркивал: „Борьба за существование идет на определенной
территории, которая
32
ограничена. ... Меня интересовал не организм, но геохимическое отражение
его на планете” (1965, с. 293).
Как свидетельствуют рукописные работы Вернадского, опубликованные в 70-х
годах, уже с первых своих раздумий о геохимической роли живого вещества
в биосфере он стремился найти основные математические понятия, отражающие
геохимические и энергетические аспекты борьбы за существование. При этом
значительную помощь ему оказал Б. А. Холодовский. Первые результаты работ
в этом направлении были изложены Вернадским в статье „Ход жизни в биосфере”
(1925) и в докладе „О размножении организмов и его значении в механизме
биосферы”, который был прочитан в январе 1926 г. в Масариковском университете
в Брно и в апреле этого же года на заседании Ленинградского общества естествоиспытателей.
В том же году доклад был опубликован в „Известиях АН СССР”. В 1926 г. вышли
в свет и две брошюры Вернадского, где излагалась методика определения геохимической
энергии у насекомых и однолетних цветковых растений (19266, 1926в). Основные
теоретические взгляды Вернадского на геохимические аспекты борьбы за существование
и роли последней в эволюции биосферы были изложены в серии работ, начиная
с „Биосферы” (1926а) и „Бактериофаг и скорость передачи жизни в биосфере”
(19276) и кончая книгой „Химическое строение биосферы Земли и ее окружения”
(1965).
Основные положения Вернадского по проблеме геохимической энергии сводятся
к следующему. Давление жизни на окружающую среду проявляется в росте и
размножении организмов как результате превращения энергии солнца в биохимическую
энергию. Эти взаимоотношения можно выразить при помощи некоторых математических
формул. Поскольку каждый вид имеет специфический темп размножения, цифровые
значения формул для каждого вида сильно различаются. Особенно велики различия
между бактериями и крупными долгоживущими млекопитающими. Вернадский отметил
обратную пропорциональную зависимость между темпом размножения организмов,
с одной стороны, и размером особи, ее массой, продолжительностью жизни
— с другой.
Связывая темп размножения отдельных видов с параметрами биосферы, Вернадский
в величине экватора Земли находит предельное, одинаковое для всех организмов
расстояние, по которому может распространиться вид и которое может преодолеть
его геохимическая энергия в процессе расселения по планете.
Введя понятие геохимической энергии организмов для характеристики борьбы
за существование, Вернадский указывал на необходимость различать реальный
геохимический эффект вида и максимально возможное проявление его геохимической
энергии при благоприятных условиях.
Таким образом, Вернадский существенно расширил дарвиновское понимание борьбы
за существование как фактора органической эволюции. Введя понятие „геохимической
энергии” и связав его с некоторыми основными константами биосферы, Вернадский
по существу открыл путь к точному количественному изучению геохимического
воздействия отдельных видов на эволюцию биосферы. Как справедливо отмечал
Е. М. Лавренко: „Если Дарвин доказал основное значение размножения
33
организмов для развития (эволюции) органического мира, то В. И. Вернадский
с неменьшей убедительностью выявил общеземное, планетное значение этого
биологического процесса, его роль в механизме земной коры” (1945, с.44).
Само развитие биосферы В. И. Вернадский связывал с постоянным противоречием
между способностью организмов к быстрому размножению и ограниченностью
материальных и энергетических ресурсов жизни в каждый геологический момент
ее истории. Очевидно, что разрешение этого противоречия было возможно только
на пути создания все более прогрессивных форм жизни, способных к завоеванию
новых зон жизни, овладению новыми источниками минеральных веществ и рациональному
использованию уже освоенных ресурсов.
Для понимания взаимосвязи эволюции организмов с эволюцией биосферы особый
интерес представляют составленные Вернадским (1925, 19276, 1965) таблицы
размножения организмов. Из данных табл. 1 видно, что наибольшей геохимической
энергией отличаются бактерии и цианофиты, выживаемость которых обеспечивается
за счет продуцирования и расхода большего количества биомассы. Таксоны
с наиболее высокой геохимической эволюцией подвергаются интенсивной элиминации,
в результате чего они в течение миллиардов лет оставались на относительно
низком уровне морфологической организации. Это объясняется, видимо, тем,
что предел в создании форм, способных к быстрому накоплению биомассы и
к „мгновенному» заселению потенциально доступной зоны жизни, был достигнут
еще на заре жизни. В дальнейшем главным направлением в эволюции стало формирование
организмов, наиболее эффективно использующих солнечную энергию для синтеза
органических соединений (эволюция растительного покрова) и усложняющих
циклы миграции химических элементов за счет активного переноса их в разные
регионы биосферы (эволюция двигательной активности животных).
С большим интересом Вернадский отнесся к начавшимся в нашей стране экспериментальным
исследованиям борьбы за существование (Сукачев, 1927; Гаузе, 1934а, 1939).
Эти исследования, по мнению Вернадского, позволяли приступить к познанию
элементарных преобразований в биосфере, так как в процессах борьбы за существование
„резко изменяется ... биогенная миграция атомов” (Вернадский, 1965, с.
304).
Вместе с тем Вернадский считал, что в дарвинистской трактовке борьбы за
существование как фактора эволюции односторонне выпячены различные формы
конкуренции и конституциональной борьбы и „не учтены два других больших
биологических явления ..., тесно связанных со структурой живого вещества”
(1965, с. 288). Это, во-первых, взаимопомощь и кооперация у организмов,
ведущих групповой образ жизни, а во-вторых, прямая борьба между хищниками
и жертвами, паразитами-хозяевами и т. п. Интересно отметить, что он подчеркивал
также необходимость учитывать значение аллелопатических взаимодействий
между растениями, открытых в конце прошлого века В. Хэдсоном „волн жизни”,
а также своеобразие процессов борьбы за существование в пределах различных
таксономических групп и в разных экосистемах. Все эти высказывания Вернадского
направлены на исправление одностороннего
34
Таблица 1
Скорость размножения организмов (по: В. И. Вернадский, 1965)
Организмы |
Δ |
a |
Бактерии |
|
|
| Bacterium coli comm. |
61.27 |
2.78 • 1018 |
| Vibrio cholerae asiat. |
61.02 – 62.47 |
2.2 •1018 – 6.4 • 1018 |
| Bacillus ramosus |
53.33 |
1.16 • 1016 |
Диатомеи |
|
|
| Nitzschia putrida |
4.8 |
26.8 |
| Среднее для морских диатомей (планктон) |
0.5 |
0.41 |
Piridinae |
|
|
| Среднее для Ceiatium (планктон) |
0.25 |
0.19 |
Phytomonadina |
|
|
| Eudorina elegans |
0.79 |
0.72 |
Heliozoa |
|
|
| Actinophrys |
1.25 |
1.38 |
Инфузории
|
|
|
Paramecium aurelia
|
1.78 |
2.32 |
P. caudata
|
1.09 |
2.12 |
| Leucophrys patula |
7.0 |
127.0 |
Dicotyledonae
|
|
|
Trifolium repens
|
0.016 |
0.0088 |
Solanum nigrum
|
0.04 |
0.029 |
| Пшеница (среднее для Франции) |
0.019 |
0.013 |
Mammalia
|
|
|
Sus scrofa
|
0.9 – 2.25 · 10-3 |
0.5 – 1.5 · 10-3 |
S. domestica
|
9.7 – 10.6 · 10-3 |
6.7 – 7.10-3 |
Mus decumanus
|
9.6 (мин) · 10-3 |
7.7 · 10-3 |
| Elephas indicus |
0.096 – 0.4 · 10-3 |
0.067 – 0.07 · 10-3 |
понимания борьбы за существование не столько самим Дарвином, сколько его
некоторыми последователями. В дальнейшем был выполнен ряд исследований
механизмов кооперации и их роли в эволюции. На многочисленных объектах
изучались разнообразные аспекты (в том числе и эволюционные) взаимодействий
типа хищничества, паразитизма и т. д. Находит свой отклик и призыв Вернадского
учитывать эволюцию самой борьбы за существование (Шмальгаузен, 1940; Галл,
Колчинский, 1972). Однако всестороннее исследование этого процесса остается
еще задачей будущего.
Биосферная детерминация процессов макроэволюции. В трактовке эволюции живого
Вернадский стоял на позициях адаптациогенеза,
35
понимая эволюцию как непрерывный процесс приспособления к сложной констелляции
абиотических и биотических факторов. Высокая эволюционная пластичность
органических форм, их способность адаптироваться к разнообразным по температуре,
давлению, химизму средам обусловливают потенциальную возможность многообразных
путей эволюции. „Создание эволюционным путем новых форм жизни, приспосабливающихся
к новым условиям ее бытия, увеличивает всюдность жизни, расширяет ее область.
Жизнь этим путем вносится в такие места биосферы, в которых она раньше
не существовала” (Вернадский, 1928, с. 240). В качестве примеров он называет
образование пещерной фауны и глубоководного бентоса. Завоевание жизнью
горячих источников, ледниковых и снежных полей, высокогорных областей и
т. д., по мнению Вернадского, свидетельствует о том, что адаптивная экспансия
жизни продолжается и в настоящее время. Максимальное разнообразие органического
мира достигается „способностью живого к приспособлению к среде, способностью,
действующей неуклонно и непрерывно сотни миллионов, по-видимому, миллиарды
лет. Приспособляемость жизни необычайна, и формы ее проявления бесконечны.
Идет, по-видимому, расширение поля жизни” (Вернадский, 19276, с. 446).
Однако существует и ряд ограничений для эволюционных преобразований органического
мира и его экспансии в поверхностной оболочке Земли. Среди них, по мнению
Вернадского, наиболее важной является сложившаяся в биосфере система круговоротов
вещества и потоков трансформации энергии, которая и определяет главные
направления в эволюции живого, канализируя в какой-то мере всю эволюцию
органического мира. Под действием этих ограничений в эволюции создаются,
как правило, „формы, все более усиливающие влияние жизни на окружающую
среду” (Вернадский, 1928, с. 243).
Для понимания взаимосвязи эволюции биосферы и видообразования особое значение
имели три биогеохимических принципа, сформулированных Вернадским. В этих
принципах по существу выражены фундаментальные закономерности, лежащие
в основе эволюции органического мира. Так, в первом принципе, согласно
которому биогенная миграция атомов стремится к максимуму, Вернадский
охарактеризовал общую направленность биогенной миграции химических элементов
в биосфере.
Во втором принципе выражена главная тенденция в биогеохимической эволюции
организмов. „Эволюция видов в ходе геологического времени, приводящая
к созданию форм жизни, устойчивых в биосфере, идет в направлении, увеличивающем
проявление биогенной миграции атомов в биосфере” (там же, с. 242-243).1
Согласно этому принципу, способность организма интенсифицировать биогенную
миграцию атомов была одним из важнейших критериев действия отбора.
Вернадский подчеркивал, что в этих двух принципах отражается „биогенная
миграция эволюционного процесса, в масштабе исторического
1 Этот принцип соответствует сформулированному Э. Бауэром (1935)
принципу возрастания внешней работы организмов, рассматриваемому как важнейшая
закономерность прогрессивной эволюции.
36
времени не меняющаяся, но резко меняющаяся в аспекте геологического времени”
(1965, с.285). Это означало, что отнюдь не каждый вид должен эволюировать
в сторону ускорения миграции атомов. Персистентные формы неизменны в течение
десятков и сотен миллионов лет не только в морфологическом, но и в химическом
отношении. В то же время сложившаяся в биосфере система биогеохимических
круговоротов препятствует возникновению форм с более низкой биогеохимической
энергией. Даже в ходе регрессивной эволюции форм, перешедших к паразитизму,
резко увеличивается скорость размножения, т. е. чрезвычайно увеличивается
биогенная миграция их атомов, что „усиливает их положение в борьбе за существование”
(там же).
В третьем принципе сформулировано положение о том, что в каждый период
геологического времени „заселение планеты должно было быть максимально
возможное для всего живого вещества, которое тогда существовало” (там же,
с. 286). Очевидно, этот принцип не имеет ничего общего с встречающимися
в последние годы утверждениями о том, что с возникновения биосферы область
ее распространения и ее биомасса якобы не менялись.
Вернадский отмечал и изменение детерминирующего влияния отдельных компонентов
биосферы на эволюцию жизни. Пока жизнь, писал он, была представлена только
водными формами, ее биомасса и продуктивность ограничивались физико-химическими
свойствами гидросферы, регулирующими содержание углекислого газа в воде.
Такая регуляция не позволяет достигнуть значительного увеличения биомассы
растительности, так как не дает извлечь из воды весь углекислый газ. Завоевание
растительностью суши означало снятие этих ограничений. „ …Жизнь, — писал
Вернадский, — выходит из того сдерживающего ее количество напряжения,
какое оказывает Всемирный океан” (1927а, с. 208). Проблема факторов-ограничителей
эволюционного процесса активно обсуждается в эволюционной теории (см.,
например: Тимофеев-Ресовский и др., 1969; Сутт, 1977). Ставится задача
изучения различных биохимических, геномных, онтогенетических, видовых,
биоценотических и других „запретов” в эволюции. Поэтому особенно актуальны
идеи Вернадского о существовании ограничений на биосферном уровне.
В своих последних работах Вернадский уделял внимание таким филогенетическим
закономерностям эволюции, как неравномерность темпов эволюции и внезапные
вымирания крупных групп растений и животных на границах геологических
периодов, направленность и необратимость эволюционного процесса, полифилетичность
происхождения крупных таксономических групп. Уровень филогенетических
исследований
того времени не позволял объективно оценить значимость этих закономерностей
макроэволюции, которые изучались преимущественно в недарвиновских концепциях
эволюции. Вернадский же воздерживался от окончательного суждения о природе
этих закономерностей. Он был согласен и с идеями Д. Н. Соболева (1924)
и Б. Л. Личкова (1936) о совпадении крупных преобразований органического
мира с периодически происходящими внезапными усилениями орогенетических,
тектонических, вулканических и других процессов. Допускал он и влияние
на эволюцию
37
органического мира периодических колебаний в интенсивности космических
излучений. Однако Вернадский оставлял в стороне вопрос о конкретном механизме
синхронизации преобразований в органическом и неорганическом мире.
Как следствия адаптации органического мира к выполнению биогеохимических
функций в биосфере рассматривались Вернадским закон Э. Копа – Ч. Депере
об увеличении размеров тела в процессе эволюции филогенетических ветвей
и закон Д. Дана о цефализации, т.е. тенденции к неуклонному усложнению
нервной системы и психической деятельности в ходе прогрессивного развития
живого. Ему не только удалось выйти за рамки традиционно антидарвинистских
трактовок этих филогенетических закономерностей, но и впервые объяснить
их с точки зрения структуры биосферы. Увеличение размеров тела, как правило,
ведет к повышению активности организмов в захвате пищевых ресурсов, борьбе
с конкурентами, хищниками, к более экономичному обмену веществ, к снижению
плодовитости. „Увеличение веса, - писал Вернадский, - показывает большую
энергию организма в добывании пищи, и, очевидно, организмы, имеющие благоприятные
условия для своего развития, будут... стремиться достигнуть максимума проявления
своей энергии, насколько это дозволяют внешние обстоятельства” (1978, с.
92). Поэтому увеличение размеров тела характерно для быстро прогрессирующих
групп организмов. Параллельно это способствовало и росту общего значения
организмов в геохимических процессах.
Усиление же доли „разумности” в поведении животных не только обеспечивало
быстрое приспособление к новым условиям существования, как это отмечал
А. Н. Северцов (1922), но и совпадало с общим направлением эволюции органического
мира в сторону усложнения биогенной миграции атомов в биосфере. Так, например,
с появлением птиц миграция химических элементов в тропосфере достигла максимума,
так как птицы стали играть решающую роль в обмене веществ между сушей и
океаном (Вернадский, 1928). Эта роль противоположна геохимической деятельности
рек. Если реки переносят химические элементы и минеральные вещества в моря
и океаны, то рыбоядные птицы обеспечили их обратное движение. „Создание
этих летающих видов позвоночных не только породило новые формы биогенной
миграции, но и резко отразилось на химическом богатстве моря и суши, оно
внесло изменения в историю отдельных элементов, в частности фосфора” (там
же, с. 246). Столь же значительные изменения в биогеохимических круговоротах
вызывали и другие эволюционные преобразования, связанные с усовершенствованием
поведения животных.1 Именно высокоразвитые животные (насекомые,
рыбы, птицы, китообразные) нередко мигрируют на сотни, тысячи, а иногда
и десятки тысяч
километров.
1 Взгляды Вернадского на цефализацию как способ ускорения биогенной
миграции атомов в биосфере фактически противоречат трактовке П. Тейяр де
Шарденом
цефализации как реализации божественной сущности в эволюции. Для Вернадского
цефализация была следствием эволюции в условиях все возрастающей сложности
взаимодействий организмов, когда возможности биохимической и морфологической
эволюции были уже исчерпаны.
38
Мысль Вернадского о существовании в эволюции органического мира тенденции
к цефализации, т. е. к „разумному способу поведения”, получила дальнейшее
развитие в концепции неограниченного прогресса (Завадский, 1958; Huxley,
1963). Так, К.М.Завадский, выделяя магистральную линию в эволюции живого
вплоть до появления человека, подчеркивал, что она оказалась возможной
„только при совершенно определенном течении процессов эволюции в
других ветвях, и в этом смысле она сама есть результат развития всей живой
природы в целом” (1958, с. 103). Благодаря прогрессивному развитию нервной системы
у групп животных, занимающих, как правило, верхние звенья в цепях питания,
резко повышалась функциональная целостность биосферы и ее способность к
саморегуляции, усовершенствовались процессы трансформации вещества и энергии
(Шмальгаузен, 1972; Одум, 1975; Шварц, 1976, и др.).
По-новому удалось Вернадскому подойти и к традиционной проблеме о моно-
или полифилетическом происхождении основных таксономических групп. Он считал,
что представления о едином или немногих предках современного органического
населения Земли не только не подкрепляются прямыми данными палеонтологии,
но и противоречат тому, что нам известно о „геологическом значении живого
вещества» (Вернадский, 1965, с. 291). Чем дальше мы уходим в глубь веков,
тем отчетливее становится дискретность основных типов животных и растений.
Поэтому для него было удивительным, что на разработку проблемы полифилетического
происхождения органического мира „по недоразумению... направлены усилия
противников дарвинизма” (там же), а в рамках школы А. Н. Северцова по-прежнему
придерживаются монофилетической концепции.
Решение проблемы полифилетического происхождения основных таксонов Вернадский
видел в своей оригинальной гипотезе возникновения жизни на нашей планете.
Далее мы подробнее остановимся на этой гипотезе. Здесь отметим только,
что в противоположность глубоко укоренившимся представлениям о первичности
организма и производности всех остальных форм существования живого, Вернадский
впервые обосновал идею о том, что первичная биосфера с самого начала была
сложной неоднородной системой. В ее состав входили не только первичные
виды, но и первичные биоценозы, так как ни один вид не мог бы самостоятельно
исполнять основные геохимические функции биосферы.
Эта позиция Вернадского принципиально отлична от предшествующих концепций
полифилетического происхождения органического мира (см., например: Бекетов,
1896; Берг, 1922). Новизна развиваемых Вернадским представлений заключалась
в признании одновременного образования нескольких форм существования жизни,
которая изначально могла функционировать лишь в форме биотического круговорота
веществ и потоков трансформации энергии. И только могуществом консервативных
традиций в биологии можно объяснить тот факт, что эти идеи Вернадского
не нашли в свое время должного признания.
Таким образом, Вернадский существенно углубил представления об эволюции
органического мира. Если раньше в эволюционной теории
39
учитывались лишь изменения морфологических и физиологических свойств организмов,
то Вернадский показал путь к изучению геохимических аспектов эволюции.
В области микроэволюции он предложил геохимическую оценку интенсивности
борьбы за существование, в учении о виде — геохимическую трактовку вида
и видообразования. С учетом фундаментальных черт организованности биосферы
становятся понятными и такие закономерности макроэволюции, как неравномерность
темпов эволюции, возрастание многообразия органических форм, повышение
уровня организации животных и растений, а также направленность эволюции
отдельных филетических ветвей и мегоарогенеза в целом. Сформулированная
Вернадским геохимическая трактовка эволюции организмов намечала новые подходы
к изучению процессов видообразования и стала методологической основой для
постановки вопроса о главных тенденциях в эволюции биосферы.
Мысль Вернадского о взаимосвязи процессов видообразования и эволюции биосферы
получила отзвук и у биологов-эволюционистов. Например, М. М. Камшилов подчеркивал:
„Суммарная жизнедеятельность развивающихся видов организмов определяет
особенности биосферы, которая в свою очередь обусловливает возможности
выживания и направления эволюционных преобразований отдельных видов. Короче,
направление эволюционных преобразований отдельных видов представляет собой
функцию их положения в эволюирующей биосфере” (1974, с. 69-70).
2.2. Гипотеза о возникновении биосферы
К своей оригинальной гипотезе происхождения биосферы Вернадский пришел
в ходе длительных раздумий над вопросами о сущности жизни, о пространственных
и временных границах распространения жизни в космосе, о возможности абиогенного
синтеза живого вещества, о причинах появления жизни на Земле и т. д. Постоянно
возвращаясь в течение четверти века к этим вопросам, Вернадский вносил
в свои представления изменения и уточнения. Это дало повод некоторым его
критикам усмотреть противоречивость во взглядах Вернадского и даже приверженность
к агностицизму и витализму (Новогрудский, 1931; Деборин, 1932, 1933). Поверхностное
знакомство с трудами Вернадского по проблемам происхождения биосферы привело
и некоторых современных авторов к аналогичному выводу, что „Вернадский
в учении о биосфере занял иррациональную позицию в вопросе о происхождении
жизни, считая исследование причин происхождения жизни занятием бесполезным,
иллюзорным и опасным” (Dose, Rauchfuss, 1975, S. 2).
Такие оценки сделаны на базе лишь некоторых высказываний Вернадского. В
них не учитывается развитие его представлений по данной проблеме. Игнорируется
тот факт, что гипотезы о вечном существовании жизни в космосе и о появлении
жизни на нашей планете в результате заноса в латентном состоянии микроорганизмов
или спор
40
из космического пространства Вернадский считал вероятными в первые годы
своих биохимических исследований.
Необходимо отметить, что идея вечности жизни была широко распространена
среди естествоиспытателей в конце XIX-начале XX веков. К ее сторонникам
принадлежали К. Бернар, Г. Гельмгольц и др. Гипотезы Г. Рихтера и С. Аррениуса
о заносе зародышей жизни из космоса при помощи световых лучей многими рассматривались
тогда как удовлетворительные, вполне научные объяснения появления жизни
на Земле. После опытов Л. Пастера идея абиогенного возникновения жизни
вызывала скептическое отношение у многих естествоиспытателей как сугубо
спекулятивное натурфилософское построение. Для ученых более приемлемыми
были взгляды, выраженные в яркой форме польским географом и геологом И.
Д. Лукашевичем, который, кстати, оказал большое влияние на формирование
биогеохимических взглядов Вернадского. Вот как описывал Лукашевич пути
распространения жизни в космосе: „Угасающие светила на своих прощальных
лучах рассылают бесчисленные зародыши в бездны Вселенной, и там, где упадут
эти лучи на молодую планету, вспыхивает новая жизнь, начинается новый цикл
развития, вечно однообразный по существу, вечно различный по конкретным
результатам” (19096, с. 32).
В книге „Начало и вечность жизни” (1922а) Вернадский по существу присоединился
к идее вечности жизни. В пользу этой идеи, по мнению Вернадского, свидетельствовала
вся многовековая история неудачных попыток синтезировать органические вещества
абиогенным путем и экспериментальные опровержения возможности „самозарождения”
организмов. И в исторической геологии не находил он фактов о существовании
азойных эпох в геологический период развития Земли. Абиогенез в земных
условиях казался Вернадскому невозможным из-за специфики живого вещества
по сравнению с окружающим его косным веществом. Эти особенности проявлялись
в дисимметричности живого вещества, в однородности его изотопного состава
и т. д. Зависимость жизни от космических излучений Вернадский рассматривал
как подтверждение принципа X. Гюйгенса о всеобщем распространении жизни
во всех частях космоса. Вот почему в статьях и книгах, опубликованных в
начале 20-х годов, он заявлял: „Идея вечности и безначальности жизни, тесно
связанная с ее организованностью, есть то течение научной мысли, последовательное
проявление которого открывает перед научным творчеством широчайшие горизонты”
(1922а, с. 58). Он ссылается при этом на принцип Ф. Реди, согласно которому
„все живое от живого”, т. е. отрицающий идею самозарождения.
Однако, как видно из рукописей Вернадского, написанных в эти же годы, но
опубликованных только в 70-х годах, он далеко не был уверен, что принципы
Гюйгенса и Реди, гипотезы Рихтера и Арррениуса могут объяснить появление
жизни на Земле. Эти принципы указывают лишь на отсутствие абиогенеза
в наши дни, но совершенно не объясняют земной структуры, столь приспособленной
для развития жизни. Эти принципы оставляют нерешенным вопрос о том, почему
на Земле жизнь существует в форме биосферы. Вернадский не мог допустить,
что все многообразие
41
живого вещества возникло из немногих примитивных организмов, „поселившихся
на земной поверхности из космического пространства” (1978, с. 133).
Поэтому он считал необходимым всесторонне исследовать ту комбинацию
космических
и геологических условий, которая создала предпосылки для образования
и развития жизни как единой целостной системы.
К началу 30-х годов Вернадский занял еще более осторожную позицию
в трактовке принципа Реди. Он писал: „В действительности принцип
Реди
не отрицает абиогенеза,
он только указывает пределы, в которых абиогенез отсутствует. Возможны
такие условия в земной истории, когда не было биосферы и существовали
на земной коре физико-химические явления или состояния, которые сейчас
в ней
отсутствуют и которые были необходимы для абиогенеза” (1931, с. 635).
Он даже допускал возможность, что и сейчас существуют физико-химические
условия,
„которые допускают абиогенез, происходящий и ныне на Земле, но по
своей незначительности и недостаточной точности наших обычных методов
исследования
ускользающий от внимания” (там же).
К этим выводам Вернадский, видимо, пришел под влиянием ряда новых
открытий в области химии и биохимии. В частности, было показано,
что нет принципиальных
отличий в изотопическом составе большинства элементов в живом и косном
веществе. Экспериментально была показана возможность создания оптически
активных (дисимметричных) веществ при помощи циркулярно-поляризованного
солнечного света. В лаборатории А. Н. Баха в эти годы шли опыты по
абиогенному синтезу биоорганических соединений. И наконец, в 1924
г. была опубликована
знаменитая гипотеза А. И. Опарина о закономерном характере возникновения
жизни на Земле в условиях восстановительной атмосферы.
Признавая возможность абиогенного возникновения жизни, Вернадский
считал правильнее говорить о возникновении не отдельных организмов,
а биосферы.
В 1931 г. в статье „Об условиях появления жизни на Земле” Вернадский
впервые изложил принципиально новый подход к данной проблеме, основанный
на понимании
жизни как неотъемлемой части определенного механизма Земли — биосферы.
„Необходимо считаться, — подчеркивал Вернадский, — с особенностью
геохимических функций живых организмов и механизма биосферы, вызывающих
сложность
жизни, существование неразрывного комплекса организмов, распадающихся
на многочисленные
морфологически различные формы. В биосфере всегда наблюдалось, говоря
терминами геохимии, разнородное живое вещество и жизнь всегда исполняла
одновременно
разнородные биогеохимические функции” (1931, с. 638). Он считал недопустимым
рассматривать возникновение жизни на Земле как абиогенез однообразных
организмов. Напротив, „должен был одновременно появиться сложный
комплекс живых форм,
развернувшийся затем в современную живую природу” (там же, с. 639).
Среди организмов нет форм, способных одновременно выполнять все функции
биосферы.
По мнению Вернадского, первичная жизнь могла существовать в форме
комплекса „многих видов, морфологически принадлежащих к разным,
резко разделенным
классам организмов” (там же, с. 648). В противном случае,
42
полагал он, пришлось бы допустить существование какой-то „видовой” формы
организмов.
Таким образом, согласно Вернадскому, надиндивидуальные формы организации
жизни оказываются столь же древними, как и сама жизнь. Органический мир
представляет собой исторически сложившуюся форму организации биогенных
потоков вещества и энергии. Его становление включало не только появление
дискретных носителей жизни (вначале молекулярных систем возрастающей сложности,
затем прокариотной и эукариотной клеток и, наконец, многоклеточного организма),
но и образование надорганизменных систем – видов и биогеоценозов.
Почти одновременно с Вернадским сходные идеи были высказаны Тейяр де Шарденом,
который также писал о возникновении жизни на нашей планете в форме биосферы.1 Однако французский палеонтолог исходил не из анализа геохимических функций
биосферы, как это было сделано Вернадским, а из философской концепции
панпсихизма (Ермоленко, 1975). Здесь был не столько новый подход к проблеме
возникновения
жизни, сколько новая попытка оправдать спиритуалистический трансформизм.
Долгое время эти оригинальные идеи Вернадского по существу оставались вне
поля зрения исследователей, занимающихся проблемой происхождения жизни.
Экспериментальные исследования предбиологической эволюции, стимулированные
трудами А. И. Опарина, были нацелены прежде всего на выяснение путей образования
ее дискретных носителей — организмов. Вероятно, только Н. Г. Холодный (1945)
попытался как-то синтезировать гипотезы А.И.Опарина и Вернадского, относя
формирование биосферы к периоду возникновения архебионтов, питавшихся готовыми
летучими органическими веществами из первичной атмосферы.
Постепенно высказанные Вернадским идеи находят все большее число сторонников
среди биогеохимиков, экологов и биологов-эволюционистов (Камшилов, 1961,
1974; Ковальский, 1963а; Наумов, 1964; Завадский, 1968; Шапошников, 1975;
Cloud, 1976; Fiermes, 1976; Заварзин, 1979; Чернов, 1983; Уголев, 1985,
и мн. др.). В свете современных знаний становится ясным, что жизнь — это
свойство, присущее экосистеме в целом, а не отдельным организмам или изолированным
скоплениям молекулярных соединений. Поэтому „центральный вопрос происхождения
жизни — это не вопрос о том, что возникло раньше, ДНК; или белок, а вопрос
о том, какова простейшая экосистема” (Патти, 1970, с. 178). Отсюда следует,
что в раннем археозое на основе разнообразных высокомолекулярных белковых
и нуклеотидных соединений возникали не единицы, а миллионы открытых систем,
способных более или менее продолжительное время находиться в состоянии
динамического равновесия. Но лишь немногие из них достигали той степени
внутренней слаженности и сбалансированности процессов метаболизма, которые
были
1 Книга Тейяр де Шардена, где излагались эти взгляды,
была написана в 30-х годах, но опубликована только после его смерти (1965,
рус. пер.).
43
необходимы для их самосохранения и воспроизведения в условиях зарождающейся
биосферы.1
Современная теоретическая биология в целом приходит к идеям, высказанным
Вернадским несколько десятилетий тому назад. К проблеме возникновения жизни
в форме биосферы все чаще обращаются и зарубежные биохимики и биогеохимики
(Флоркен, 1959; Cloud, 1976; Цветков, 1981, и др.). Предпринимаются первые
попытки реконструировать ландшафтные условия возникновения жизни на Земле
(Панфилов, 1977) и процессы формирования круговорота органических соединений
— „зародыша будущего биотического круговорота” (Камшилов, 1974, с. 43).
Исходя из идей Вернадского, Камшилов пришел к выводу, что биотический круговорот
сформировался на основе абиогенных процессов синтеза и деструкции органического
вещества, происходящих в добиологический период истории Земли. Он предполагал,
что биоценозы возникли раньше отдельных организмов и видов. Биотический
круговорот лишь постепенно дифференцировался, образуя отдельные виды.
Но трудно представить, как биоценоз может функционировать без его составляющих
— гетерогенных компонентов организменного и видового уровней. Жизнь действительно
не может существовать вне биоценозов и биосферы. И в этом Камшилов безусловно
прав. Но нельзя согласиться с тем, что жизнь могла существовать без ее
дискретных носителей — организмов. Поэтому более приемлемыми являются
работы,
в которых обосновывается мысль об одновременном возникновении на Земле
четырех основных форм организации жизни: организмы, виды, биоценозы,
биосфера (Завадский, 1968).2
Несколько слов о других сторонах научных воззрений Вернадского на проблему
возникновения жизни. Все большее признание находят его идеи о неразрывной
связи земной жизни с космосом. Накапливаются и данные о том, что в момент
образования Земля в „наследство” от космической эволюции могла получить
разнородные углеродистые соединения, послужившие в дальнейшем основой для
синтеза органических веществ и протобионтов. Действительно, в межзвездном
пространстве
1 Мы полагаем, что наиболее разработанной на сегодняшний
день является концепция отбора гиперциклов М. Эйгена и П. Шустера (1982).
В ней учтены
такие важные критерии отбора самоорганизующихся систем, как каталитическая
активность макромолекул, совершенствование способов превращения вещества
и энергии, возрастание их скорости, экономичности и согласованности,
совершенствование механизмов наследственной информации и т. д. Отбор
по этим признакам неизбежно
вел к становлению биологических систем, главными критериями которых
стало
соответствие биоценотическим условиям, выражающееся в дифференциальной
размножаемости и выживаемости (Колчинский, 1983б).
2 Вместе с тем нельзя полностью игнорировать возможность
монофилетического происхождения жизни. Об этом говорят не только
традиционные данные
о единстве генетического кода, биохимических процессов и т. д., но
и соображения экосистемного
порядка. Так, на примере термофильной фауны Б. М. Медников (1985)
достаточно убедительно доказывает вероятность монофилетического возникновения
протоорганизма – акцептора электронов в условиях стабильного источника
– донора электронов.
44
обнаружены низкомолекулярные соединения углерода (формальдегид, циан и
его производные), спирты, эфиры, аминокислоты и др. Однако вряд ли в древнейших
слоях Земли удастся найти „следы” космических соединений. Они могли разрушиться,
войти в состав первичной атмосферы или первых органических соединений.
При палеобиохимических исследованиях докембрийских пород имеют дело не
с „пришельцами из космоса”, а с веществом, заново синтезированным в этих
недрах под влиянием биохимических процессов (Опарин, 1979).
Сами идеи о заносе жизни из космоса теперь обычно излагаются в модифицированном
виде. Жизнь на нашей планете рассматривается как результат ее аккреционного
образования из космической пыли (см., например: Robinson, 1966, 1967).
Подобные представления отнюдь не исключают экспериментального подхода к
проблемам биопоэза. Более того, они способствуют сбору данных для „решения
вопроса о земном или неземном происхождении жизни” (Фокс, Дозе, 1975, с.
303).1 Весьма перспективны и исследования связи биосферных
процессов с различными космическими излучениями (Чижевский, 1973; Дубров,
1974), которые
лежат в русле основных идей Вернадского.
Иначе обстоит дело с идеей Вернадского о широком распространении жизни
во Вселенной. Результаты многолетних радиоастрономических исследований
показывают, что Земля вряд ли могла получить готовые формы жизни в период
своего формирования. Не оказалось организмов ни на Венере, ни на Марсе,
которые еще недавно не только писателями-фантастами, но и многими крупными
учеными оценивались как планеты, населенные организмами. Земля — это единственная
планета солнечной системы, пригодная для существования жизни на углеродисто-белковой
основе. Возможно, что биосфера Земли уникальна и в рамках всей нашей Галактики.
Предположение Вернадского о практически одновременном (в масштабе геологического
времени) формировании Земли и жизни завоевывает популярность и среди геологов
(Войткевич, Холодков, 1976; Сидоренко, 1984). На смену взглядам о длительном,
охватывающем несколько миллиардов лет периоде биопоэза приходят идеи о
том, что предбиологическая эволюция шла очень быстро. Ее основные „изобретения”
(абиогенный синтез органических соединений, формирование матричного синтеза
органических молекул, образование жизни вироидного типа, обладающей точным
аппаратом самовоспроизведения макромолекулярных комплексов РНК и ДНК, развитие
прокариотов) заняли гораздо меньше времени, чем вся последующая эволюция
жизни.
1 Мы не рассматриваем гипотезу Ф. Крика и Л. Оргеля о
происхождении жизни на Земле в результате „направленной панспермии” – переноса
зародышей
жизни
с помощью неких разумных человекоподобных существ с молибденовых звезд
(Crick, Orgel, 1973). Совершенно прав Р. Дикерсон, указавший на принципиальную
непроверяемость подобных умозрительных построений (Dickerson, 1978,
р. 70).
45
2.3. Основные тенденции в эволюции биосферы
Возникновение жизни, по мнению Вернадского, положило начало геологическому
периоду в развитии нашей планеты и длительному процессу эволюции биосферы.
Подчеркивая неразрывную связь эволюции видов с эволюцией биосферы, Вернадский
вместе с тем стремился найти интегральные характеристики эволюции биосферы,
ее специфические закономерности, не сводимые к закономерностям эволюции
на нижележащих уровнях организации живого. Эволюция химического состава
организмов и их метаболических процессов рассматривалась им прежде всего
в плане ее влияния на геологическое строение биосферы, ее биогеохимические
функции и общую энергетику. „Благодаря эволюции видов, непрерывно идущей
и никогда не прекращающейся, резко меняется отражение живого вещества на
окружающей среде. Благодаря этому процесс эволюции — изменения — переносится
в природные биокосные и биогенные тела, играющие основную роль в биосфере,
в почвы, в наземные и подземные воды (в моря, в озера, реки и т.д.), в
угли, битумы, известняки, органогенные руды и т.п. ” (Вернадский, 1977,
с. 18).
Рост биомассы биосферы и ее организованности. Вернадский был уверен, что
благодаря способности к размножению в геометрической прогрессии сформировавшаяся
жизнь быстро „растеклась” по поверхности нашей планеты, приспосабливаясь
к разнообразным условиям существования. С самого начала эволюция живого,
считал он, может идти в соответствии с биогеохимическими принципами. Поэтому
на каждом этапе истории органического мира его биомасса и область распространения
достигали возможного предела.
Расширение зоны обитания жизни и связанное с этим увеличение размеров биосферы
Вернадский оценивал как важную тенденцию в ее эволюции. „Биосфера, — писал
он, — не есть нечто неподвижное... Это проявляется в расширении области
жизни, т. е. биосферы, в геологическое время. Вырабатываются организмы,
которые, приспосабливаясь к условиям безжизненных областей, их заселяют:
процесс расширения идет быстро в ходе геологического времени” (1937, с.
12). В то же время он допускал, что процесс освоения жизнью новых областей
продолжается и в наши дни.
В связи с этим неизбежно возникал вопрос о том, менялось ли количество
живого вещества в ходе развития биосферы. Позиция Вернадского здесь была
неоднозначной. С одной стороны, он считал, что общее количество живого
вещества оставалось примерно одинаковым в течение всего геологического
времени. Объем живого вещества казался ему предопределенным количеством
химических элементов, пригодных для использования в биологических процессах.
А это количество, как считал Вернадский, было постоянным. Кроме того,
он опирался на общепринятые тогда представления о том, что биомасса Мирового
океана (область обитания древнейшей жизни) во много раз превышает общую
биомассу более молодых надземных экосистем. С другой стороны, многочисленные
высказывания Вернадского о непрерывной экспансии жизни по поверхности
Земли,
об образовании все более сложных и продуктивных
46
экосистем, об усложнении циклов миграции химических элементов и т. д.
говорят о том, что он допускал возможность увеличения биомассы в эволюции
биосферы.
Такая несогласованность в высказываниях Вернадского обусловливалась прежде
всего знаниями о распределении живого вещества в биосфере. Н. Б. Вассоевич
и А. Н. Иванов (1984) указали, что приводимые Вернадским в разные годы
оценки общей биомассы планеты отличались на несколько порядков величин.
Опираясь на преувеличенные оценки количества живого вещества в океане,
Вернадский ошибался при сравнении биомассы суши и океана по меньшей мере
на три порядка величин. Конечно, и сейчас оценки продуктивности и биомассы
биосферы в целом, возможно, не более близки „к действительности, чем простая
догадка” (Одум, 1975, с. 71). Еще хуже обстояло дело в 20-30-х годах, когда
Вернадский, поставив вопрос о необходимости суммарной оценки всей биомассы,
не смог правильно оценить распределение живого вещества в различных регионах
биосферы. Тем более у него не было никаких количественных данных для решения
вопроса об изменении биомассы биосферы во времени.
В качестве важной тенденции в эволюции биосферы Вернадский выделял возрастание
ее организованности. Непрерывный обмен атомов, отмечал он, в ходе геологического
времени стремится к динамическому равновесию. Но это равновесие постоянно
смещается по мере насыщения биосферы живым веществом, захвата им новых
зон, общего усложнения биотического круговорота. В своем развитии биосфера
„никогда не возвращается по существу к прежнему состоянию; совпадение даже
в основном... с былым состоянием никогда не наблюдается, сходство всегда
приблизительное” (1937, с. 11). В последнем издании „Очерков геохимии”
Вернадский подчеркивал, что циклы химических элементов „обратимы
лишь в главной части атомов, часть же элементов неизбежно и постоянно
выходит
из круговорота. Этот выход закономерен, т. е. круговой процесс не является
вполне обратимым” (1954, с. 40). В целом же биогенные циклы атомов „привели
к выявлению организованности биосферы, приспособленной к жизни и жизнью
воссоздаваемой” (там же, с. 48). Правда, он не дал объяснения причин организованности
биосферы, отмечая лишь усиление ее стабильности, способности к саморегуляции
и независимости от других земных оболочек. Возможно, это вызвано тем, что
в процессе коренных перестроек биосферы сохранялись прежде всего те группы
организмов, которые были устойчивы к воздействию астрономических и геологических
факторов. Возрастание целостности и автономности элементарных компонентов
в биосфере в целом способствовало и увеличению ее организованности.
Роль живого вещества в становлении и стабилизации поверхностных
оболочек Земли. Как уже отмечалось, под влиянием организмов
и продуктов их жизнедеятельности постепенно все геологические и геохимические
процессы превратились в биогеологические
и биогеохимические. „На земной поверхности, — писал Вернадский (1926а,
с. 25), — нет химической силы, более постоянно действующей, а потому
и более могущественной по своим конечным последствиям, чем живые организмы,
взятые
47
в целом”. Решающая роль живого в эволюции биосферы особенно ярко, проявилась
в формировании и стабилизации газового состава атмосферы, в превращении
восстановительной обстановки на нашей планете в окислительную, в преобразовании
химической и минералогической структуры литосферы, в детерминации химической
активности природных вод и общего термодинамического баланса биосферы (1926а,
1965). Избирательно извлекая из окружающей среды химические элементы и
их изотопы и выделяя их обратно в виде органических соединений, живое вещество
создает и сохраняет в охваченной им области общепланетарный биотический
круговорот. „Живое вещество, — писал Вернадский, - охватывает и перестраивает
все химические процессы биосферы, действенная его энергия по сравнению
с энергией косного вещества... огромна. Живое вещество есть самая мощная
геологическая сила, растущая с ходом времени” (1965, с. 127) (курсив наш.
– Э. К.).
Остановимся кратко на идеях Вернадского об эволюции геохимической структуры
биосферы под воздействием жизни.
До работ Вернадского считалось, что состав и химические свойства атмосферы
определяются законом всемирного тяготения и планетарными свойствами Земли.
И хотя уже с конца XVIII в. была известна роль растений в обогащении атмосферы
кислородом, все-таки предполагалось, что они играют второстепенную роль
в детерминации газового состава атмосферы. Органический мир рассматривался
в лучшем случае как придаток к атмосфере (Ж. Буссенго, Ж. Дюма). Уже в
„Очерках геохимии” Вернадский дополнил идеи Буссенго и Дюма, показав, что
само соотношение основных газов атмосферы обусловлено жизнедеятельностью
организмов. Регуляция газового состава, по мнению Вернадского, осуществлялась
двумя путями. Первый из них связан с жизнедеятельностью организмов или
с разложением их остатков, а второй – с процессами метаморфизации и вулканизма.
Вернадский считал, что большинство газов, выделяемых при метаморфизме горных
пород и извержении вулканов, по своему происхождению биогенны, т. е. являются
преобразованными продуктами жизнедеятельности организмов.
Позднее Вернадский не раз возвращался к вопросу о влиянии живого вещества
на атмосферу. „Атмосфера, — писал ученый, — всецело создана жизнью, она
биогенна” (1942, с. 252).
Вывод Вернадского о том, что жизнь, преобразуя атмосферу, как бы создает
себе необходимую среду обитания, в последние годы в нашей стране стал
азбучной истиной. И странно читать, что „по мере более глубокого познания
тропосферы,
а также благодаря догадкам, которыми мы в значительной мере обязаны английскому
ученому и изобретателю Джеймсу Лавелоку, становилось более очевидным,
что нижняя атмосфера Земли была сильно модифицирована жизнью, причем так,
чтобы
поддерживать жизнь в еще больших масштабах» (Маргелис, 1983, с. 298).
Идея о том, что жизнь в значительной степени предопределяет основные свойства
своей среды, благодаря Вернадскому укрепилась в отечественной науке задолго
до гипотезы Геи, выдвинутой Дж. Лавелоком в 70-х годах (Lovelock, 1979,
1983). И именно Вернадскому, а не Лавелоку, принадлежит заслуга во всестороннем
доказательстве этой идеи.
48
Вернадский показал также ведущую роль живого вещества в детерминации химической
активности природных вод. „Состав воды океана, - писал Вернадский, - в
главной своей части регулируется жизнью, живым веществом, его энергией”
(1923, с. 27). Эта регуляция осуществляется путем концентрации элементов
организмами из вод океана. Включенные в состав биохимических соединений,
они получают совсем иную судьбу и дают начало совсем другим продуктам,
чем получались бы при чисто химических процессах испарения морской воды.
На основе анализа роли живого вещества моря в миграции кремния, алюминия,
железа, кальция, магния он показал: „Какой бы элемент мы ни брали, всюду
мы видим одну и ту же картину – всюду живое вещество, жизнь является
основным агентом, создающим химию моря” (Вернадский, 1923, с. 34).
Вернадский существенно углубил представление Ч. Дарвина и В. В. Докучаева
о ведущей роли организмов в генезисе и эволюции почв. Уже в статье „Генезис
минералов” (1892), опубликованной в Энциклопедическом словаре Брокгауза
и Эфрона, он подчеркнул необходимость рассматривать почвообразование как
часть более общего биогеохимического преобразования поверхностных оболочек.
В дальнейшем (1913, 1921а, 1988) он не раз возвращался к этому вопросу,
отмечая перспективность изучения процессов почвообразования с биогеохимической
точки зрения. И здесь мы хотели бы напомнить авторам, доказывающим постулирование
Вернадским принципа неизменности гидросферы и педосферы (см.: Посохов,
1981), лишь одно его высказывание: „Почвы и реки девона, например, иные,
чем почвы третичного времени и нашей эпохи” (Вернадский, 1977, с. 18).
Не случайно ведущие советские почвоведы (Полынов, 1956; Ковда,1973) не
раз подчеркивали, что идеи Вернадского и учение В. Р. Вильямса стали основами
современных представлений об эволюции почв.
И до Вернадского было известно о геологической деятельности организмов.
Были выделены органогенные породы (известняки, горючие/ископаемые и т.
д.). Большое внимание геологической роли организмов уделяли в своих работах
Н. И. Андрусов, М. А. Егунов, Г. А. Надсон, Н. А. Соколов и, особенно,
ученик Вернадского Я. В. Самойлов. Но в целом жизнедеятельность организмов
и разложение их остатков оценивались как второстепенный фактор в образовании
осадочных пород, полезных ископаемых, минералов и т. д. Например, наиболее
распространенные горные породы — пески и глины — считались абиогенными.
Совсем иначе смотрел на эту проблему Вернадский. „Все минералы верхних
частей земной коры – свободные алюминокремниевые кислоты (глины), карбонаты
(известняки и доломиты), гидраты окиси железа и алюминия (бурые железняки
и бокситы) и многие сотни других непрерывно создаются в ней только под
влиянием жизни... С исчезновением жизни не оказалось бы на земной поверхности
силы, которая могла бы давать непрерывно начало новым химическим соединениям”
(1926а, с. 25). Этот вывод Вернадский подкреплял и экспериментальными
исследованиями. Он доказал, что диатомовые водоросли способны разлагать
49
каолиновые глины, разрушая связи алюминия и кремния. Без организмов эта
реакция идет только при высокой температуре.
В книге „Очерки геохимии” (1927а) Вернадский подробно проанализировал роль
организмов в геохимических преобразованиях соединений алюминия, железа,
магния, углерода, кальция, кремния. Так, разбирая циклы кремния в земной
коре, он показал, как организмами захватываются атомы кремния из водных
золей кремнезема. Одни организмы разрушают каолиновые ядра алюмосиликатов
и извлекают из них атомы кремния. Под влиянием продуктов жизнедеятельности
организмов кремний концентрируется в твердые кремнеземистые коллоиды —
опалы, а затем и выкристаллизовывается в кварц, собирается в водных магниевых
силикатах и в глинах. Образовавшиеся „вадозные” минералы со временем проникают
в метаморфизированные оболочки, где образуют серпентины, тальки, хлоритоиды,
хлориты, или возвращаются к исходному состоянию — слюдам, полевым шпатам
и т. д. Их ядра, разбиваясь, дают начало силлиманиту и кварцу. В более
глубоких метаморфических оболочках вновь синтезируются различные каолиновые
алюмосиликаты. Особенно в больших масштабах этот синтез идет при застывании
расплавленной магмы.
Вернадский доказывал, что литифицированными продуктами жизнедеятельности
организмов являются не только громадные запасы горючих ископаемых, но и
все осадочные породы и даже, возможно, гранитная оболочка представляет
собой метаморфизованные остатки былых биосфер.
Разбирая вопрос об эволюции каустобиолитов, Вернадский пришел к выводу,
что она обусловлена изменениями в химическом составе организмов, остатки
которых служат основой для их образования. Он высоко ценил пионерские исследования
В. О. Таусона (1936), который показал, что каустобиолиты „являют индивидуальность
и неповторяемость в геологическом времени и представляют собой историческое
явление, что выражается в их чрезвычайном химическом разнообразии и просто
объясняется тем, что для каждого геологического момента организмы, тела
которых они представляют, были неповторяемы, резко химически различны,
так же как различны те микробы, которые вызвали их образование и тела которых
входят в их состав” (Вернадский, 1965, с. 269). Идеи Вернадского о ведущей
роли живого вещества в становлении и стабилизации геохимических особенностей
поверхностных оболочек Земли и приводимый им фактический материал в их
подтверждение требуют специального анализа, который далеко выходит за рамки
задач нашей книги. Сказанного достаточно, чтобы подчеркнуть приоритет Вернадского
в выдвижении и доказательстве этих идей.
Некоторое время они с трудом воспринимались геологами и географами. Приведем
только два примера.
Полемизируя с Вернадским, крупный советский геолог Л. В. Пустовалов писал,
что организмы, сами зависящие в своем развитии от абиотических факторов,
не могли и не могут „играть той будто бы ведущей и определяющей роли,
которую ошибочно нередко приписывают им” (1940, с. 375). При этом явно
не учитывалось, что многие из так называемых
50
абиотических агентов образования горных пород на нашей планете являются
продуктами жизнедеятельности организмов.
Сравнительно недавно Д. Л. Арманд (1975) уверял, что географ не может принять
идеи о ведущей роли жизни в создании современных геологических оболочек
Земли. Жизнь, отмечает он, - вторичное явление на Земле. Основные физико-географические
процессы (тектогенез, выветривание, эрозия, сток, оледенение и т. д.) протекают
без всякого влияния жизни. Любопытна и форма аргументации автора. Жизнь,
пишет он, преобразует лик Земли в той же мере, в какой стенная роспись
— интерьер здания. Ни один архитектор не станет утверждать, что фрески
несут на себе здание. Поэтому он считал правильнее говорить не об эволюции
биосферы, а об эволюции ландшафтосферы или географической оболочки в смысле
А. А. Григорьева (1966).
Подобная критика идей Вернадского о ведущей роли жизни в эволюции атмо-,
гидро-, педо- и литосферы – достояние прошлого. Как подчеркивал А.В.Сидоренко
(1984),с самого начала образования биосферы (3.5—4 млрд лет тому назад)
органическое вещество стало одной из главных действующих сил всех поверхностных
и приповерхностных процессов, вызвавших глубокую и мощную дифференциацию
и трансформацию первичного земного вещества наружной оболочки нашей планеты.
Идеи о громадном преобразующем влиянии живого вещества на геологические,
в том числе и геохимические процессы постепенно занимают все большее место
и в трудах зарубежных ученых. В 1940 г. Тейяр де Шарден и Леруа основали
впервые институт по проблемам геобиологии и журнал Geobiologia. В настоящее
время эти идеи разрабатываются в Геобиологической лаборатории в Австралии,
в Биогеологической лаборатории в США и других лабораториях. Поставленная
Вернадским проблема эволюции химического состава Земли, ее оболочек, минералов
и горных пород интенсивно исследовалась в трудах Дж. Пуля (Poole, 1951),
В. Руби (Rubey, 1955), Б. Мейсона (1971), Г. Голланда (Holland, 1972),
Д. Аллена и М.Нельсона (Allen, Nelson, 1986).
К настоящему времени затихла дискуссия между сторонниками биогеохимических
объяснений литогенеза и приверженцами физико-химических трактовок этого
процесса. „Не может быть сомнения, — пишет голландский ученый П. Вестброк
(Westbrock, 1983, р. 91), — что жизнь оказала сильное влияние на развитие
нашей планеты. История жизни и история Земли являются коэволюцией”. Вместе
с тем стало ясно, что роль живого вещества в геологических процессах различна,
„начиная от процессов огромного биогеохимического значения и кончая лишь
небольшой дополнительной ролью” (Ферсман, 1934, с. 296). Нередко живое
вещество является лишь катализатором геологических процессов.
Накопление энергии в биосфере. Вернадский дополнил энергетический
подход к эволюции организмов, намеченный еще в трудах А. Больцмана, Г.
Гельмгольца,
С. А. Подолинского, В. Томсона и К. А. Тимирязева. Ему глубоко импонировала
мысль В. Анри о том, что жизнь с космической точки зрения „есть не что
иное, как постоянное задержание и накопление химической и лучистой энергии,
замедляющие превращение
51
полезной энергии в теплоту и препятствующие рассеянию последней в мировом
пространстве” (1917, с. 455). Опираясь на высказывания своих предшественников
о направленности эволюции в сторону уменьшения энтропии, Вернадский независимо
от В. Л. Комарова (1921) и А. Лотки (Lotka, 1925) предложил рассматривать
находящуюся в биосфере энергию как результат органической эволюции.
В „Очерках геохимии” Вернадский охарактеризовал основные способы увеличения
энергии в биосфере: во-первых, это фотосинтез и выделение организмами кислорода,
обладающего высокой химической активностью; во-вторых, захват растениями
новых областей Земли, превращение их в области аккумуляции солнечной энергии
при фотосинтезе; в-третьих, аккумуляция солнечной энергии в горючих ископаемых
и биогенных минералах. Эволюция биосферы, отмечал Вернадский, ведет к прогрессивному
накоплению запаса превращаемой энергии в поверхностных оболочках Земли,
прежде всего в литосфере, и тем самым к уменьшению „производства” непревращаемых
форм энергии в земных условиях. Препятствуя деградации энергии, жизнь обусловливает
рост негэнтропии в биосфере; „происходит увеличение деятельной энергии”
(Вернадский, 1927а, с. 228). По мере развития растительности и усложнения
трофических связей в биосфере идет обогащение ее живого и биокосного вещества
аккумулированной энергией.
Вернадский впервые показал, что аккумуляторами солнечной энергии в литосфере
являются не только горючие ископаемые, как было общепринято после работ
Ю.Майера, но и все „вадозные” минералы, причем в них содержится большая
часть накопленной в земной коре энергии. И хотя вначале эта мысль вызвала
немало возражений со стороны геологов и геохимиков, впоследствии благодаря
работам А. Е. Ферсмана о геоэнергетике минералов, Б. Б. Полынова о биогенном
генезисе минералов глин и особенно Н. В. Белова о „геохимических аккумуляторах”
она получила широкое признание.
О возникновении новой формы миграции химических элементов. В
специальной главе, посвященной изучению эволюции концентрационных функций
биосферы,
мы подробнее рассмотрим взгляды Вернадского на проблему изменения форм
миграции химических элементов.1 Здесь отметим только, что,
выделяя биогеохимические функции живого вещества, он полагал, что функции,
связанные с питанием,
размножением и дыханием организмов, с разложением их остатков, издавна
существовали в биосфере. Функция же, связанная с перемещением атомов
в процессах преобразования внешней среды в соответствии с потребностями
организмов,
стала возможной только с появлением животных со сложным способом поведения.
Деятельность роющих организмов, постройка муравейников, ульев, плотин,
гнезд и т. д. — все это было для Вернадского примерами возникновения
в ходе эволюции биосферы качественно новой формы биогенной миграции.
Она
не была связана с прохождением химических элементов через тело организмов.
Такая форма биогенной
1 Это вызвано тем, что работы Вернадского в этом направлении
были опубликованы позже исследований его ученика Я. В. Самойлова (см.
гл. 4).
52
миграции и биогеохимической функции живого вещества достигла наибольшего
развития в наши дни, когда значительная часть мигрирующих атомов в биосфере
перемещается в соответствии с потребностями человека.
Биосферные адаптации. Вернадский впервые поставил вопрос о возможности
адаптивных преобразований на биосферном уровне. Важнейшей адаптацией биосферы
он считал возникновение озонового экрана в результате фотосинтезирующей
деятельности растений, защитившей жизнь от губительного воздействия ультрафиолетового
излучения (Вернадский, 1942). Способность растений улавливать солнечную
энергию и переводить ее в формы, пригодные для обеспечения самых разнообразных
биосферных процессов, он вслед за Тимирязевым считал другой важнейшей адаптацией
биосферы.
Идея Вернадского о биосферных адаптациях получила поддержку в трудах С.
С. Шварца (1973, 1976) и Г. Л. Шкорбатова (1982). Такие адаптации Шварц
называл „линиями обороны” биосферы. Первая линия обеспечивается за счет
организованной разнородности трофических уровней, каждый из которых включает
сотни и тысячи видов, состоящих в свою очередь из миллионов и миллиардов
особей, уникальных в генетическом отношении. Это многообразие жизни повышает
вероятность сохранения главных геохимических и энергетических функций биосферы
в случае каких-то катастроф планетарного масштаба. Иерархичность структурных
уровней биосферы (см. подробнее гл. 6) оценивалась Шварцем как вторая „линия
обороны” биосферы. К числу биосферных адаптации Шкорбатов относит „регуляцию
потоков энергии в экосистемах и соотношение основных трофических групп
в биомах, регуляцию круговоротов биогенных элементов и газового состава
атмосферы” (1982, с. 785). И у Шварца и у Шкорбатова организованность биосферы
по существу отождествляется с ее адаптивностью.
В целом же вопрос о возникновении адаптации на уровне биосферы до сих пор
остается совершенно неразработанным, что в значительной степени затрудняет
понимание сущности преобразований биосферы.
* * *
Как видно из вышеизложенного, в эволюции биосферы Вернадский выделял
несколько основных тенденций. Это расширение области миграции элементов
в результате
захвата жизнью все новых зон обитания; усиление преобразующего влияния
жизни на среду, на косные компоненты биосферы; общее ускорение темпов
миграции элементов; появление качественно новых форм миграции элементов,
непосредственно
не связанных с внутриорганизменным обменом (например, деятельность
животных); резкое усиление отдельных биогеохимических функций биосферы;
концентрация
громадного количества солнечной энергии в поверхностной оболочке
Земли и, наконец, общее усложнение структуры биогеохимического круговорота,
связанное с появлением новых потоков миграции элементов. Все эти
преобразования
биосферы
шли в соответствии с биогеохимическими принципами, сформулированными
Вернадским.
53
В распоряжении Вернадского не было палеобиогеохимических и палеонтологических
данных о влиянии преобразований отдельных видов на структуру и функции
биосферы. Впрочем, даже в будущем не приходится рассчитывать на получение
таких данных. В лучшем случае возможным станет лишь изучение изменений
в химическом строении биосферы и в биотических круговоротах в результате
возникновения крупных таксонов. Познание главных тенденций в эволюции биосферы
может идти „путем изучения общего эффекта жизни в истории химических элементов
Земли — биогенной миграции химических элементов и тех правильностей, которые
с этой миграцией связаны” (Вернадский, 1928, с. 234).
Разрабатывая проблему эволюции биосферы, Вернадский в то же время полагал,
что основные физико-географические параметры нашей планеты были неизменны
в течение геологического времени. Он скептически относился к различным
геологическим гипотезам о пульсирующих изменениях размеров Земли, о постепенном
остывании ее поверхности и т. д. Это дало основание некоторым историкам
геологии характеризовать взгляды Вернадского как антиэволюционные (Guntau,
1967; Urbanek, 1973). Приведенный материал показывает, насколько ошибочны
подобные оценки. Именно Вернадский наметил пути для изучения связи эволюции
видов с геохимическими и энергетическими преобразованиями биосферы. Этот
путь давал возможность выделять главные тенденции в эволюции биосферы.
2.4. Учение о ноосфере как заключительном этапе
эволюции биосферы
Разработанные Вернадским биогеохимические и энергетические подходы к изучению
эволюции биосферы требовали дополнить современную периодизацию истории
жизни, которая строится преимущественно на изучении остатков ископаемых
организмов, новыми более существенными критериями. Не раз возвращаясь к
этому вопросу, Вернадский обычно отмечал четыре крупных этапа в истории
биосферы: появление первичных автотрофов, возникновение животных с кальциевым
скелетом, формирование лесных биогеоценозов и, наконец, создание ноосферы.
Как видно, для выделения этих этапов он использовал разные критерии. Так,
первый этап связан с появлением нового способа питания и важнейшего компонента
биосферы, второй — с усилением одной из биогеохимических функций, третий
— с возникновением биотического сообщества, наиболее эффективно осуществляющего
аккумуляцию солнечной энергии, а четвертый — с резким изменением всех биогеохимических
и энергетических процессов в биосфере. Начатый Вернадским поиск критериев
периодизации истории органического мира, отличных от принятых в геохронологии,
впоследствии был продолжен в ряде работ (см. гл. 6).
Особое внимание уделял Вернадский (1916, 1927а, 1937, 1944, 1965, 1977
и др.) заключительному этапу в истории биосферы, когда человеческая деятельность
стала решающим фактором преобразования природы.
54
К концу своей жизни он сформулировал основы учения о ноосфере, которое
сейчас оказалось в центре внимания ученых самых различных специальностей
как у нас в стране, так и за рубежом.
Сама мысль о громадном влиянии деятельности человека на эволюцию природы
часто высказывалась в трудах крупнейших ученых XIX в. — создателей эволюционных
концепций (Ж.-Б. Ламарк, Ч. Лайель, Ч. Дарвин, В. В. Докучаев и др.). Еще
в начале XIX в. Ламарк отмечал, что силы человечества стали столь велики,
что существует опасность уничтожения им самого себя и окружающей среды
(Ламарк, 1935). Автор концепции классического униформизма Лайель (Lyell,
1830—1832, 1863) готов был признать, что появление человека и его влияние
на геологические процессы нарушают даже основополагающий принцип униформизма,
т. е. принцип неизменности факторов и законов. Хорошо известно, какое внимание
уделял Дарвин анализу селекционной деятельности человека и ее роли в преобразовании
органических форм. И наконец, В. В.Докучаев (1883, 1892) показал, что сельскохозяйственная
и производственная деятельность — важнейшие факторы почвообразования.
В середине XIX в. вышла книга Г. Марша „Человек и природа или о влиянии
человека на изменение физико-географических условий природы”. Цель книги
заключалась в том, чтобы указать на „размеры изменений, произведенных человеком
в физических условиях обитаемой им планеты, — раскрыть опасность неблагоразумия
и необходимость осторожности, когда речь идет о вмешательстве в широких
размерах в непосредственный порядок органического или неорганического мира”
(Марш, 1866, с. VII). Автор обобщил громадный материал о последствиях производственной
деятельности, которая вела к уменьшению числа диких животных и растений,
к изменению численных соотношений между видами, к сокращению площади лесов,
изменению местного климата и т. д. Марш подчеркивал, что только человек
,,имеет разрушительную силу, только он один обладает такой энергией, против
которой совершенно бессильна сама природа” (1866, с. 39). Поэтому необдуманная
эксплуатация природных ресурсов, предупреждал английский ученый сто лет
тому назад, грозит существованию самого человека.
Важность перехода к планомерному взаимодействию человека со средой стала
осознаваться широкими кругами научной общественности в начале XX в. Ясно
эта мысль была сформулирована А. Е. Ферсманом в брошюре „Химические проблемы
промышленности”. Он подчеркивал, что человек должен согласовывать свою
деятельность с законами окружающего мира. В приспособлении к нему заключается
„весь закон эволюции, борьба за существование, естественный отбор, вся
сложность химических превращений и физических процессов, наконец, вся жизнь
человека с ее постоянной борьбой и постоянными исканиями” (1924, с. 50).
Не раз предпринимались попытки выделить современную эпоху как начало новой
эры в истории Земли. Эту эру американские геологи Ч. Шухерт (Schuhert,
1923) и Д. Леконт (Le Cont, 1904) предложили назвать психозойной, а академик
А. П. Павлов (1922) – антропогенной. А. Е. Ферсман (1934) называл совокупность
геологических и геохимических процессов, вызванных деятельностью человека,
техногенезом.
55
Существовали и философские предпосылки учению о ноосфере. Истоки рачительного
отношения к природе восходят к мифам различных народов. У философов Древней
Греции можно найти немало призывов следовать за природой в своих действиях.
Идеями о закономерном характере превращения человека в решающую силу развития
биосферы и даже космоса пронизаны труды многих мыслителей на грани XIX
и XX веков, и прежде всего сторонников русского космизма – Н. Ф. Федорова,
Н. А. Умова, К. Э. Циолковского и др.
Таким образом, сама идея о превращении человека в решающий фактор биосферных
преобразований еще до работ Вернадского получила общенаучное признание
и философское обоснование. Заслуга Вернадского заключается в том, что он
впервые показал закономерный характер этого процесса, связав его с предшествующей
эволюцией биосферы. Он создал основу учения о ноосфере как заключительном
этапе в истории биосферы.
Это учение Вернадский разрабатывал практически на протяжении всей жизни.
В 1989 г. впервые были опубликованы фрагменты из писем и дневников Вернадского,
отражающие зарождение и развитие идей о переходе биосферы в ноосферу. Оказывается,
уже в студенческом докладе „О предсказании погоды”, прочитанном в 1882
г. на заседании студенческого научно-литературного общества Санкт-Петербургского
университета, Вернадский писал о превращении человека в геологическую силу
в связи с возрастанием мощи воздействия его на природу. „Человек настоящего
времени, - говорил юный Вернадский, - представляет из себя геологическую
силу и сила эта сильна именно тем, что она возрастает и предела ее возрастанию
нам не видно” (там же, с. 152). К этим мыслям он не раз возвращался в своих
дневниках и письмах. Но к систематической разработке учения о ноосфере
Вернадский приступил практически в последние 30 лет. Еще до революции,
работая в Комиссии по изучению естественных производительных сил России,
в докладе на собрании Петроградского общества естествоиспытателей в 1915
г. он подчеркивал, что своеобразие геологической деятельности человека
по сравнению с другими организмами заключается в том, что „он один коснулся
в своей работе почти всех химических элементов и, вероятно, в конечном
итоге коснется всех элементов” (1916, с. 80). Умение человека использовать
потенциальную энергию, сконцентрированную в полезных ископаемых, а также
энергию рек, ветра, приливов и отливов и т. д., по мнению Вернадского,
свидетельствует о том, что в биосфере действует качественно новый фактор
ее преобразования.
Глава „Человечество как часть однородного живого вещества” из книги Вернадского
„Живое вещество” показывает, насколько он был увлечен проблемой человека
как фактора развития биосферы в период своих первых биогеохимических
исследований. Он выдвинул тогда мысль о необходимости рассматривать историческое
развитие
человеческой культуры „как естественноисторическое проявление жизни на
нашей планете, ... как одно из проявлений геологической истории Земли”
(1978, с. 46). Краткий анализ влияния человека на живую природу со времен
неолитической революции привел Вернадского к выводу, что,
56
несмотря на незначительное число доместицированных растений и животных
по сравнению с дикими формами, влияние их на геохимические процессы биосферы
огромно. Занимая громадные и все увеличивающиеся пространства биосферы,
культурные сообщества резко меняют биогеохимические процессы биосферы.
„При создании рас, — подчеркивал Вернадский, - человек производит огромную
геохимическую работу, но она остается для нас совершенно неизвестной...
” (1978, с. 49).
Уже в первом издании „Очерков геохимии” (1927а) Вернадский ввел специальный
раздел „Геохимическая деятельность человечества”. В нем он отмечал, что
появление человека означало принципиально новую форму воздействия жизни
на геохимические циклы. В процессы обмена вовлекались элементы, которые
не нужны ему для поддержания жизни, а использовались лишь для создания
орудий труда. Человек резко „изменяет геохимическую историю всех металлов,
он образует новые соединения, воспроизводит их в количествах того же порядка,
какой создался для минералов, продуктов природных реакций” (1927а, с. 231).
Вызванные человеком изменения в геохимических циклах подготовлены всей
предшествующей эволюцией биосферы. Они способствуют ускорению миграции
атомов и увеличению свободной энергии в биосфере.
В те же годы на французском языке публикуется статья Вернадского „Автотрофность
человечества”, в которой автор обосновывает неизбежность перехода к тому
этапу в истории биосферы, когда не только взаимоотношения природы и человека,
но и само общество будут перестроены в соответствии с требованиями научного
знания. Разум человека, по выражению Вернадского, предстает в истории биосферы
как „устремленная и организованная воля его” (1980, с. 228). Его могущество
росло в геометрической прогрессии. Отмечая необходимость превращения человека
из существа социально-гетеротрофного в социально-автотрофное, т.е. перехода
к искусственному синтезу пищи, Вернадский считал, что это станет возможным
только тогда, когда разум человека, наука станут действительно ведущей
силой на Земле. Уже в этой статье по существу была сформулирована идея
о неизбежности перехода биосферы в ноосферу. Однако в эти годы Вернадский
еще не использовал термин „ноосфера”, который в 1927 г. был предложен приверженцем
философии А. Бергсона Э. Леруа (Le Roy, 1927). Рассмотрев закономерности
эволюции жизни, он пришел к выводу, что в человеке биологическая эволюция
исчерпала себя. Дальнейшая эволюция „живого на нашей планете будет совершаться
только духовными средствами: индукция, общество, язык, разум и т. д. И
это будет ноосфера, которая последует за биосферой” (ibid., p. 195-196).
Под ноосферой автор понимал не преобразованную деятельностью человека геологическую
оболочку Земли, а закономерный этап в развитии органического мира, когда
доминирующая роль в биосфере принадлежит духовному творчеству человека
и продуктам его труда. И в этом взгляды Леруа отличались от представлений
Тейяр де Шардена (1965), который ноосферу понимал прежде всего как некий
„мыслящий пласт”, постепенно разворачивавшийся из мира животных и растений
и приходящий на заключительном этапе психогенеза к осознанию тождества
всего существующего с точкой Омега – божеством.
57
В статье „Несколько слов о ноосфере” Вернадский использовал понятие „ноосфера”,
вложив в него содержание, отличающееся от трактовок и Леруа, и Тейяр де
Шардена. Для Вернадского ноосфера была закономерным этапом в развитии биосферы,
в пределах которого разум человека станет общепланетной силой, преобразующей
лик Земли не только в соответствии с потребностями человека, но и с учетом
законов биосферы. Ноосферу Вернадский понимал как естественное тело, компонентами
которого будут литосфера, гидросфера, атмосфера и органический мир, преобразованные
разумной деятельностью человека. Со временем в ноосферу будет включено
и околоземное космическое пространство.
Вернадский был уверен в закономерном характере возникновения ноосферы.
„Взрыв научной мысли в XX веке подготовлен всем прошлым биосферы
и имеет глубочайшие корни в ее строении. Он не может остановиться и пойти назад.
Он может только замедлиться в своем темпе... Биосфера неизбежно перейдет
так или иначе, рано или поздно, в ноосферу... ” (1977, с. 33). Такая уверенность
позволяла Вернадскому с оптимизмом смотреть в будущее. В разгар самой разрушительной
в истории войны Вернадский не сомневался, что будущее в руках прогрессивного
человечества, которое обязательно обуздает враждебные человеку социальные
и природные силы. Идеализируя участников антигитлеровской коалиции и надеясь
на их благотворное сотрудничество после войны, Вернадский утверждал: „Идеалы
нашей демократии идут в унисон со стихийным геологическим процессом, с
законами природы, отвечают ноосфере” (1944, с. 120). Это высказывание явно
находилось в противоречии с реальностями сталинского режима.
Вернадский осознавал, что человечество находится лишь на подступах к ноосфере.
На страницах, посвященных особенностям органической эволюции в биосфере,
Вернадский с горечью признавал, что усиливающееся воздействие человека
ведет к исчезновению многих видов, к изменению численных соотношений между
другими видами. Природные биогеоценозы заменяются культурными, а в оставшихся
природных экосистемах коренным образом перестраиваются биотические связи,
упрощается структура трофических цепей. При этом факторы органической эволюции
не переставали действовать, а проявляются „на каждом клочке территории,
захваченной человеком” (1965, с. 306).
Вместе с тем процессы борьбы за существование и естественного отбора приобретают
специфические формы в культурных биоценозах, так как „вопрос идет о заселении
на неопределенное время занятых раньше пространств” (там же). Прошедшие
длительный путь эволюции природные биоценозы, как правило, отличаются
более высокой биологической продуктивностью и стабильностью биогеохимических
круговоротов, чем культурные биоценозы. Качественный состав природных
биоценозов
отрегулирован процессами конкуренции, взаимопомощи, паразитизма, хищничества
и т. д. и обеспечивает обычно максимальное использование ресурсов биотопа.
С биогеохимической и энергетической точек зрения культурные биоценозы
„отличаются большей простотой состава” (там же, с. 297) и меньшей геохимической
энергией,
так как они могут
58
сохраняться только при условии дополнительной затраты энергии со стороны
человека (Вернадский, 1929).
Опираясь на данные А. Н. Лебедянцева о химическом составе, массе, коэффициенте
размножения, скорости распространения и геохимической энергии культурных
растений, Вернадский утверждал, что создаваемые человеком виды обладают
меньшей геохимической энергией, чем их дикие предки. Не случайно конкуренция
между культурными растениями и сорняками, как правило, заканчивается в
пользу последних. Поэтому повсеместная замена диких форм культурными растениями
таит угрозу дестабилизировать биотический круговорот.
Мысль Вернадского о закономерном характере перехода к эволюции, управляемой
человеком, стала особенно актуальна в наши дни. Влияние человечества на
процессы эволюции на глазах последних поколений стало глобальным и таким
интенсивным, что требования Вернадского о всесторонней планомерности воздействия
и учете его отдаленных последствий признается всеми (см.: Яншин, 1981,
1986). Как справедливо отмечалось М.М. Камшиловым, „на наших глазах совершается
революционный переход от эволюции, управляемой стихийными биологическими
факторами (период биогенеза), к эволюции, управляемой сознанием, к периоду
ноогенеза” (1974, с. 232). Для успешного осуществления этого перехода необходимо
приступить к созданию частной теории эволюции, учитывающей своеобразие
общих и частных факторов органической эволюции в современных условиях,
определяемых могучим воздействием антропогенных факторов (Завадский, Колчинский,
1977). Создание такой теории является задачей первостепенной важности как
по практическим, так и теоретическим соображениям.
К учению о ноосфере обращаются сейчас ученые самых различных специальностей.
Среди них геологи, геохимики, биогеохимики, географы, биологи, философы,
социологи, экономисты и др. Учение о ноосфере, возникшее как важное философское
обобщение на базе геологии, биогеохимии, эволюционной теории и истории
науки, должно стать основополагающим при планировании взаимодействия между
природой и человеком. Термин „ноосфера”, как справедливо отмечал Дж. Бернал,
позволяет „увидеть место умственной и общественной деятельности человека
в общей системе эволюции жизни” (1969, с. 226). Это учение побуждает человечество
скорее встать на путь подлинного управления биосферой. В противном случае
под угрозой оказывается само будущее человечества. Вот почему ежегодно
выходят сотни книг и статей, посвященных ноосфере, а на многих международных
конгрессах не раз звучали доклады под названием, аналогичным докладу П.
Дювиньо (1979) „Ноосфера и будущее растительности земного шара” на XII
Международном ботаническом конгрессе в Ленинграде в 1975 г.
* * *
Более сорока лет тому назад А. Е. Ферсман сказал о Вернадском: „Десятилетиями,
целыми столетиями будут изучаться и углубляться его гениальные идеи,
а в трудах его будут открываться новые страницы, служащие источником новых
исканий: многим исследователям при-
59
дется учиться его острой и отчеканенной, всегда гениальной, но трудно
понимаемой творческой мысли” (1946, с. 5). Эти слова оказались пророческими.
Чем больше времени уходит со дня смерти Вернадского, тем очевиднее величие
и значимость его научного подвига. Все большее число представителей самых
различных специальностей включаются в разработку его творческого наследия.
Поражаясь разносторонности научных изысканий Вернадского, академик Л. С.
Берг, А. П. Виноградов, Н. Д. Зелинский, С. А. Зернов, А. Н. Крылов, И.
И. Шмальгаузен и другие назвали его ученым, представляющим в своем лице
всю Академию наук. Но если для своих современников Вернадский был прежде
всего минералогом, геохимиком, радио-геологом, то сейчас мы знаем Вернадского
преимущественно как основателя биогеохимии, создателя учения о биосфере
и ноосфере, глубокого философа и блестящего историка науки. Человечество
ценит убежденность Вернадского в том, что закономерным итогом развития
биосферы будет победа разума и гуманизма.
Многие идеи и концепции Вернадского не были должным образом оценены при
его жизни. Многие работы оставались неопубликованными, и лишь в последнее
десятилетие научная общественность могла ознакомиться с ними. Некоторые
из основных идей Вернадского остались фактически незамеченными и по сей
день. К ним относятся и высказывания Вернадского по проблемам эволюции
биосферы.
Эволюционной теории еще предстоит включить основные положения учения Вернадского
о биосфере, которое также нуждается в осмыслении с позиций современного
дарвинизма. В первую очередь это относится к гипотезе об изначальном существовании
четырех форм организации жизни, к идее о взаимозависимости процессов видообразования
и эволюции биосферы, о ведущей роли антропогенных факторов в эволюции современной
биосферы.
С учетом фундаментальных черт организованности биосферы становятся более
понятными такие закономерности макроэволюции, как неравномерность темпов
эволюции, возрастание сложности и разнообразия экосистем, существование
определенных направлений в эволюции отдельных филогенетических ветвей,
а также мегоарогенез. Для изучения процессов микроэволюции особый интерес
представляют геохимическая оценка интенсивности борьбы за существование
и геохимическая трактовка вида и видообразования.
Подводя итоги развития эволюционной теории в СССР за 30 лет, М. М. Камшилов
писал, что для „разработки действенной эволюционной теории мысли и факты,
приводимые в работах В. И. Вернадского, имеют исключительное значение”
(1947, с. 310). Он особенно выделял идеи Вернадского об эволюции биосферы
в сторону усиления преобразующего влияния жизни на среду, увеличения
ее энерговооруженности и завоевания новых минеральных и энергетических
ресурсов,
экспансии жизни на поверхности Земли, об усилении взаимодействия живого
с неживым. Однако эти идеи Вернадского долгое время оставались вне поля
зрения эволюционистов. Не случайно некоторые биогеохимики считают,
60
что эволюцию биосферы, которая происходит одновременно с эволюцией организмов
и видов, „эволюционисты не признали достаточно очевидной” (Дювиньо, 1979,
с. 77). В последнее время, однако, необходимость синтеза эволюционной теории
с учением Вернадского не раз подчеркивали М. С. Гиляров, Ф. Г. Добржанский,
К. М. Завадский, Н. В. Тимофеев-Ресовский, С. С. Шварц, А. Г. Юсуфов, А.
В. Яблоков и многие другие крупные эволюционисты.
Основополагающее значение трудов Вернадского для познания биосферы и закономерностей
ее эволюции признают и представители других наук. Ограничимся двумя примерами.
Американский эколог и биогеохимик Дж. Хатчинсон отмечал в предисловии
к сборнику статей о биосфере: „Концепция биосферы, которую мы принимаем
сейчас,
в основе опирается на идеи Вернадского” (1972, с. 9). С ним согласен
французский эколог Г. Гегамян, который подчеркивал, что до Вернадского
не изучалось
„значение биосферы в общей энергетике и геохимической работе Земли, и
только В. И. Вернадский в 1924 г. в своей книге «La geochimie» впервые
глубоко
проанализировал и выявил значение «живого вещества»... в геологической
эволюции Земли» (1980, с. 581). Идеи Вернадского используются в Международной
научно-исследовательской программе „Человек и биосфера”, проводимой под
эгидой ЮНЕСКО, и учитываются практически во всех новейших теориях, посвященных
эволюции экосистем. И если проблемой эволюции биосферы по-прежнему занимаются
главным образом геохимики, палеобиогеохимики, палеогеографы, палеонтологи
и экологи, а не биологи-эволюционисты, то объясняется это тем, что все
еще остается вне поля зрения мысль Вернадского о неразрывной связи эволюции
видов с биогеохимическими процессами биосферы.
61
Глава 3
ИЗУЧЕНИЕ ГЕОХИМИЧЕСКИХ ФАКТОРОВ
ЭВОЛЮЦИИ БИОСФЕРЫ
Влияние химических факторов на эволюцию организмов интересовало биологов
еще со второй половины XIX в. Были проведены опыты с целью выяснить возможность
наследования морфологических и физиологических изменений, возникающих у
организмов при выращивании их в условиях с повышенным содержанием солей,
извести, цинка, меди, кальция. Предполагалось, что химические соединения
и элементы, воздействуя на внутриорганизменный обмен веществ, способны
прямо вызвать адекватные адаптивные наследственные изменения. Эти эксперименты
окончились безрезультатно. Адаптация к химической среде по механизму своего
возникновения в сущности ничем не отличается от адаптации к другим абиотическим
факторам (температуре, свету, влажности). В эволюционной теории прочно
укоренилось представление о том, что вопрос о роли химической среды в эволюции
живого является лишь частью более общей проблемы об абиотических факторах
эволюции.
Подобное представление справедливо при изучении адаптивных преобразований
на организменном и популяционно-видовом уровнях организации живого. Однако
при переходе к исследованию закономерностей эволюции биосферы требуется
его существенная корректировка. Функциональную основу биосферы составляет
прежде всего биогенная миграция элементов, а химический состав живого вещества
– его важнейшая характеристика. Единство процессов жизни и геохимической
среды является важнейшим условием, определяющим развитие, строение и функции
биосферы (Ковальский, 1979). Возникает необходимость объяснить установившийся
химический состав живого вещества, а также причины его своеобразия в различных
регионах биосферы.
Такая задача была поставлена в трудах В. И. Вернадского (19226, 1926а,
1965), который отмечал закономерный характер распределения химических элементов
в живом веществе биосферы и его зависимость от геохимического строения
поверхностных оболочек Земли. Впоследствии эти вопросы затрагивались в
работах ряда советских и зарубежных биогеохимиков.
В начале 30-х годов была сделана попытка установить связь между распространенностью
химических элементов в биосфере и их положением в периодической таблице
Д. И. Менделеева (А. П. Виноградов). Впоследствии к этой проблеме обращались
А. Е. Ферсман, В. Гольдшмидт, А. И. Войнар, В. Шоу, Дж. Уолд, С. А. Шукарев,
А. А. Кист, В. В. Ковальский и др.
62
Несколько позже (с конца 30-х годов) стали исследовать закономерности
химического состава организмов и его особенности в различных биогеохимических
провинциях (Я. М. Берзиня, Г. Бертранд, Р. И. Блохина, Т. Ф. Боровик-Романова,
А. П. Виноградов, М. И. Густун, Э. Кидсон, B. В. Ковальский, С. В. Летунова,
Дж. Макхарт, Д. П. Малюга, В. Ноддак, Н. С. Петрунина, Ю. И. Раецкая, X.
Ремедже, И. А. Самарина, А. М. Симорин, М. Фокс, В. С. Чебаевская и мн.
др.).
С середины 50-х годов стала формироваться геохимическая экология, в задачи
которой вошло изучение геохимических факторов микроэволюции, причины внутрипопуляционного
и внутривидового полиморфизма по пороговой чувствительности организмов
к резко повышенным или пониженным концентрациям тех или иных химических
элементов, механизмы формирования адаптации к геохимической среде (В. В.
Ковальский, С. В. Летунова, Н. С. Петрунина, М. А. Риш, А. Брэдшоу, C.
С. Шварц, М. Д. Скарлыгина-Уфимцева, М. Я. Школьник, В. М. Емец, А. В.
Жулидов, С. А. Шарыгин и др.).
Главные исследования концентрировались вокруг проблемы: зависимость химического
состава живого вещества в биосфере от биологических свойств элементов и
их положения в периодической таблице Менделеева (проблема „предопределенности”
химического состава организмов); общие закономерности химической дифференциации
организмов в биосфере; биогеохимические провинции и их роль в эволюции;
геохимические факторы микроэволюции. Далее мы остановимся на этих исследованиях,
уделяя особое внимание работам А. П. Виноградова и В. В. Ковальского, в
которых были сформулированы задачи проводимых в данных направлениях исследований
и сделаны важнейшие обобщения, имеющие большое значение для изучения геохимических
факторов эволюции биосферы.
3.1. „Химическая предопределенность” живого вещества
Вопрос о закономерном характере химического состава организмов был поставлен
еще в конце XVIII – начале XIX веков в трудах Дж. Пристли, А. Гумбольдта,
Ф. Джона, Г. Фехнера, К. Шпренгеля и Ю. Либиха. Однако в рамках господствовавшей
естественной телеологии казалось очевидным, что организмы используют именно
те химические элементы, которые лучше всего служат для отправления их важнейших
функций и для строения протоплазмы.
Следует учитывать и широко распространившиеся позже представления об единой
протоплазме всех организмов, состоящей якобы из ограниченного числа элементов:
четырех органогенов (водорода, углерода, азота, кислорода) и семи зольных
элементов (фосфора, серы, кальция, калия, натрия, железа, магния). Совокупность
этих элементов составляет более 99% общей массы у всех организмов. Другие
же элементы, встречавшиеся, как правило, в микроколичествах (от 10-3
до 10 -5 % от общей массы), расценивались как случайные примеси.
Открытие физиологической роли таких элементов, как медь, йод, марганец,
бор, кремний
и др.,
63
заставило по-новому взглянуть на химический состав организмов с учетом
всей его сложности и многообразия.
Во второй половине XIX в. внимание исследователей привлекла определенная
закономерность в использовании организмами химических элементов. Это побуждало
к изучению связи между биологическими свойствами химических элементов и
их положением в периодической таблице Д. И. Менделеева. Распространение
элементов в органическом мире связывали прежде всего с их атомной массой.
Считалось, что именно легкая масса основных биогенных элементов, расположенных
главным образом в двух первых периодах таблицы Менделеева, и предопределяет
химический состав живого вещества. Однако данное объяснение оказалось явно
неудовлетворительным, поскольку в этих же периодах расположены и элементы,
содержащиеся в организмах в незначительных количествах. В то же время относительно
тяжелые элементы из четвертого периода (калий, кальций, железо) иногда
встречаются в организмах в весьма значительных количествах. Вот почему
все больше внимания стали уделять месту элементов в ряду электрических
положительных и отрицательных зарядов, валентности атомов и их способности
образовывать многочисленные соединения с углеродом (Sestini, 1885).
Особое внимание этой проблеме было уделено в книге американского физиолога
Л. Гендерсона „Среда жизни” (1924), опубликованной впервые в 1912 г. В
этой работе развивались идеи о приспособленности физико-химических свойств
неорганического мира к потребностям жизни. Правильнее было бы, конечно,
сказать, что жизнь в своем химическом составе предопределена той геохимической
обстановкой, которая существовала на нашей планете в период биогенеза.
Тем не менее, разбирая свойства водорода, углерода и кислорода, Гендерсон
показал, что физико-химические особенности этих элементов делают их удивительно
подходящими не только для образования органических соединений, но и для
создания той неорганической среды, в рамках которой живое вещество может
существовать и эволюировать. Особую роль при этом, по мнению Гендерсона,
играют элементы, участвующие в образовании воды и углекислоты.
Исключительные термические свойства воды (большая теплоемкость, низкая
теплопроводность по сравнению с металлами и высокая теплопроводность по
сравнению с другими жидкостями, расширение перед замерзанием, диэлектрическое
постоянство, способность к ионизации, исключительная подвижность и способность
образовывать растворы, высокое поверхностное напряжение и др.) превратили
воду в необходимый компонент среды для возникновения жизни. Вода играет
особую роль и в жизнедеятельности организма. Второе место после воды по
своей пригодности для жизни занимает углекислота, которая способна растворяться
в воде, поддерживать постоянство концентрации водородных ионов, что оказалось
очень важным для поддержания реакции крови.
Работа Гендерсона сыграла большую роль в изучении биологических свойств
химических элементов, поскольку доказала, что „свойства материи и явления
космического развития... тесно связаны со строением живых организмов
и их приспособлением: поэтому эти свойства являются
64
более важными для биологии, чем это предполагали раньше” (Гендерсон, 1924,
с. 197). Но из-за слабой изученности химического состава организмов Гендерсон
был вынужден ограничиться анализом причин распространения в биосфере лишь
нескольких биофильных химических элементов и их соединений.
Новый период в разработке данного вопроса был стимулирован биогеохимическими
трудами В. И. Вернадского, которые были продолжены А. П. Виноградовыми
В. Гольдшмидтом. Вернадский (19226) попытался свести в единую таблицу данные
о распространении химических элементов в земной коре, гидросфере, атмосфере
и живом веществе. Впоследствии Вернадский не раз обращался к этому вопросу,
отмечая, правда, недостаточность точных количественных данных для его решения.
Так, в книге „Химическое строение биосферы Земли и ее окружения” (1965)
он писал, что нам известны только 14 элементов, которые содержатся во всех
организмах. Почти все они, за исключением железа, принадлежат к числу легких.
Кроме того, во многих организмах встречаются такие элементы, как бор, фтор,
марганец, кобальт, никель, медь, цинк, бром и др. В связи с этим Вернадский
считал необходимым количественно исследовать содержание элементов в живом
веществе с целью выяснить закономерности их распространения в биосфере,
подобно тому как такие закономерности были установлены ранее для распространения
элементов в космосе и литосфере.1
Такая работа применительно к химическому составу живого вещества была выполнена
А. П. Виноградовым (1933,1935б). Объясняя закономерности распространения
химических элементов в организмах, он полагал, что химический состав
живого вещества есть периодическая функция атомного номера. Содержание
тех или
иных элементов находится в обратной пропорции к их атомным массам и в
конечном счете определяется зарядом ядер атома. Подавляющую часть живого
вещества
составляют элементы, расположенные в первых четырех периодах таблицы
Менделеева.
Однако зависимость между количественным содержанием атомов в живом веществе
и их атомным номером не может быть выражена в виде прямой линии (рис.
1). Существует определенная периодичность в убывании числа атомов, которая
точно наблюдается в пределах первых 50 – 60 элементов. Кривая их распределения
имеет довольно правильные периоды с максимумами и минимумами. В целом
вся
кривая может
1 В. Гольдшмидт сформулировал геохимические законы распределения
элементов в космосе и на Земле в многотомном издании „Geochemische Verteilungegesetzeder
Elemente”, Bd 1 – 8 (1923 – 1928). В его геохимической классификации
химических элементов учитывались размеры атомов и ионов, величина зарядов
ионов, положение
элемента на кривой атомных объектов, валентность, сродство с углеродом,
кислородом, серой и т. д. Гольдшмидт доказал, что распределение элементов
в поверхностных оболочках Земли определяется их физико-химическими
свойствами и их поведением в кристаллических решетках. А. Е. Ферсман (1932,
1937)
существенно углубил и развил эти идеи Гольдшмидта, сформулировав ряд
закономерностей, устанавливающих связь между содержанием элементов в
космосе и литосфере
и их положением в таблице Менделеева.
65
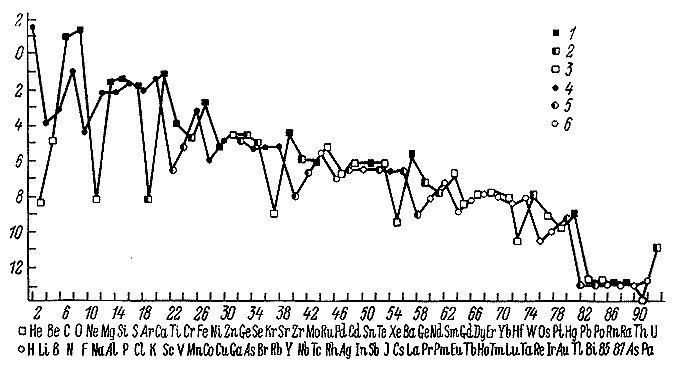
Рис. 1. Периодичность химического элементарного состава организмов (по:
Виноградов, 1935б).
1 – 3 — четные элементы, 4 – 6 — нечетные. 1, 4 – химические элементы,
количественно определенные; 2, 5 – вероятное положение элементов, количественно
не определенных; 3, 6 – вероятное положение элементов, еще не найденных
в организмах. По оси абсцисс – порядковый номер атома, по
оси ординат – lg процентного содержания атомов.
быть разделена на пять больших периодов по 18 элементов (без водорода
и гелия), т. е. от лития до кальция, от скандия до стронция и т.д.
Виноградов впервые отметил, что периодичность для кривых четных и нечетных
элементов различна, хотя формулы у них совпадают. У четного ряда периодичность
выражается формулой 2 + 6 + 6, а у нечетного 1 + 6 + 6... Элементы, отвечающие
этим числам, находятся в максимумах кривой. Максимумы кривой четного ряда
находятся постоянно на более высоком уровне, хотя постепенно сама кривая
сближается с кривой нечетного ряда. В максимумах четного ряда находится
кислород, кремний, кальций, железо, стронций, олово, барий, ртуть, радон.
Правда, здесь пришлось пропустить германий и рутений, так как тогда еще
не было данных о количественных их содержаниях в живом веществе. В нечетном
ряду в максимумах оказались водород, азот, алюминий, калий, марганец, рубидий.
Здесь Виноградов вынужден был пропустить галлий по тем же соображениям,
по которым в четном ряду были пропущены рутений и германий.
Виноградов показал, что химические элементы, находящиеся в максимумах кривых,
являются „главным субстратом (связанных с их геохимической функцией)
наиболее древних организмов и их современных потомков” (1935б, с. 23).
Для всех
этих элементов имеются организмы – мощные их концентраторы, у которых
содержание кремния, калия, железа, алюминия и т. д. достигает целых процентов
от общей
66

Рис. 2. Химический элементарный состав организмов и периодическая система
Менделеева (по: Виноградов, 1935б).
I – элементы 1-го уровня, II – элементы
2-го уровня.
массы (рис. 2). Нередко эти организмы выступают в качестве породообразователей.1
Существует и определенная связь между биологическими и геохимическими функциями
элементов. Атомы железа и марганца выступают в качестве переносчиков
кислорода и в биохимических, и в геохимических процессах, а физиологический
антагонизм ряда элементов, находящихся в максимумах четного и нечетного
рядов (К/Са, Fe/Mn; Pb/Sr, Cs/Ba), соответствует их геохимическому антагонизму.
В итоге организмы, богатые Са (Corallinoideae) бедны К, и наоборот (Laminareae).
При этом „химический элементарный состав различных видов развивается
закономерным образом путем изменения в содержании сразу ряда отдельных
химических элементов” (там же, с. 25).
Сравнивая результаты исследования закономерностей распределения элементов
в живом веществе с выявленными закономерностями их распространения в
других оболочках Земли, Виноградов отмечал, что в принципе везде действуют
одни
и те же закономерности. Но в каждой
1 Суждения Виноградова о концентрационных
функциях организмов и их эволюциях подробно анализируются в следующей
главе.
67
из оболочек они проявляются по-разному. Кривая распределения атомов в
биосфере будет отличаться в деталях от подобных кривых, построенных для
литосферы, гидросферы и почвы.
Таким образом, А. П. Виноградов показал, что распространение элементов
в биосфере зависит не только от их массы и заряда ядра, что отмечалось
и до него, но от целого комплекса физико-химических свойств элементов.
Жизнь использует прежде всего те атомы, которые наиболее пригодны для осуществления
биохимических и физиологических функций. В заключение статьи Виноградов
писал: „Мы должны ожидать, что новая экспериментальная проверка может многое
исправить в наших построениях, но основная идея о периодичности в содержании
химических элементов в живом веществе несомненно сохранится” (1933, с.
35). И это предположение Виноградова подтвердилось.
Почти одновременно с работами Виноградова появился ряд статей, в которых
делались попытки классифицировать химические элементы с учетом их биологических
свойств и положений в таблице Менделеева (Thatcher, 1934; Frey-Wyssling,
1935; Steinberg, 1938, и др.). В этих работах предлагались различные классификации,
построенные на основе представлений об уже установленных или предполагаемых
функциях тех или иных элементов в биохимических, физиологических и энергетических
процессах организмов. Критический анализ этих классификаций сделал В. Стайлс
(1949).
Внимание ученых привлекла и поставленная Виноградовым проблема периодичности
в распространении химических элементов в биосфере (Webb, Fearon, 1937;
Mason, 1958). Вскоре после публикации статей Виноградова А. Е. Ферсман
(1937) сравнивал результаты своих исследований о зависимости распространения
элементов в космосе и литосфере от их положения в таблице Менделеева с
выводами Виноградова. Ферсман подчеркивал, что химические элементы, составляющие
живое вещество, в целом подчиняются основным закономерностям распределения
этих элементов в неорганическом мире. Живое вещество продолжает тенденцию
геохимических процессов, заключающуюся в концентрации наиболее подвижных
атомов.
Ферсман отмечал, что 23 элемента верхней части таблицы, составляющие большую
часть живого вещества, группируются около оси благородных газов так, что
ближе всего к этой оси расположены основные биогенные элементы. „С точки
зрения таблицы Менделеева, - писал Ферсман, — элементы биогенеза, стремясь
к наиболее верхним периодам Менделеевской системы, вместе с тем очень определенно
группируются в поле ионов сферических, наибольших радиусов, наименьшей
валентности и потому наименьших энергетических показателей, отвечая способности
к наибольшей активности и миграции” (там же, с. 430).
В структуре клетки преимущественно участвуют легкие элементы большой валентности,
которые, однако, в условиях избытка свободного кислорода не остаются
катионами, а превращаются в подвижные комплексные анионы больших радиусов
и малых
величин энергии решетки и энергии иона в решетке. В то же время жизнь
использует и элементы, способные образовывать устойчивые решетки с большими
величинами
68
энергетических связей внутри нее. Подобные соединения нерастворимы, прочны
механически, термически, гидро- и электростатически. Они используются для
образования защитных и опорных органов (например, [SiO4]-4, Са+2[СО3]-2,
Са+2[РО2]-2). Жизнь, способствуя накоплению свободного кислорода в атмосфере,
ведет к образованию легких подвижных анионов, а в итоге к усилению процессов
миграции элементов в биосфере, „к накоплению в ней элементов, подвижных,
малостойких в своих соединениях, тогда как более стойкие и менее подвижные
комплексы выпадают в отдельных более устойчивых зонах (таков SiO2 в подзолах)
” (там же, с. 430).
Позднее идеи Виноградова и Ферсмана были подтверждены и существенно дополнены
в работах А. И. Войнара (1960), В. Шоу (Shaw, 1960), Дж. Уолда (1964),
С. А. Шукарева (1971), А. А. Киста (1973). Так, Войнар дал обзор данных
о распространении 60 микроэлементов в организме животных и человека. На
основе этих данных он обратился к вопросу о зависимости биологический роли
микроэлементов от их расположения в таблице Менделеева. Сопоставляя данные
Виноградова о химическом составе почв и литосферы с данными Ф. Левтгардта
(Leuthardt, 1941) о содержании микроэлементов в живом веществе, он пришел
к выводу, что „содержание элементов в живом веществе пропорционально составу
среды, с поправкой на растворимость соединений, куда входят эти элементы”
(Войнар, 1960, с. 12). Поэтому главную массу живого вещества составляют
отнюдь не самые распространенные в поверхностных оболочках Земли элементы,
а те из элементов, которые легко образуют газы и водорастворимые соединения.
Несмотря на принадлежность алюминия и титана к числу наиболее распространенных
элементов в земной коре, в организмах они встречаются в незначительных
количествах. Другая картина у углерода, азота, фосфора и йода. В сумме
микроэлементов в живом веществе гораздо меньше, чем в земной коре. При
этом близкие по химическим свойствам элементы, как правило, встречаются
в организмах примерно в тех же соотношениях, в которых они содержатся в
окружающей среде. Войнар подчеркивал, что по мере расширения знаний о взаимосвязи
между строением элементов и их биологическими функциями все реальнее становилась
задача по построению периодической системы биологических элементов.
Наиболее обоснованной на сегодняшний день является биогеохимическая периодическая
таблица В. Шоу (Shaw, 1960), в которой автор суммировал все известные
к началу 50-х годов знания о распространенности элементов в живом веществе
с вьщелением необходимых, а иногда и незаменимых для жизни атомов. В
этой
таблице приведены основные физико-химическое свойства, обусловливающие
биологическую значимость атомов. В таблице электронная структура атомов,
их число и атомная масса сопоставлены с такими свойствами элементов,
как электроотрицательность, ионный и квантовый радиусы, ионный потенциал
и
потенциал ионизации и т. д. Все эти свойства элементов обусловливают
их химическую активность и способность образовывать элементы различной
степени
устойчивости и пригодности для использования в физиологических и биохимических
процессах. Из данных таблицы видно,
69
что распространение атомов в живом веществе определяется соотношением
их различных свойств, а также их содержанием в среде и способностью организмов
их избирательно концентрировать.
Шоу предложил разделить химические элементы на три типа в зависимости от
того, как их использование в организмах соответствует принимаемому многими
биогеохимиками правилу, что „более важные для организмов элементы относятся
к числу наиболее распространенных в морской воде” (р. 200).
Классификация Шоу позволяет лучше понять закономерность в эволюции химического
состава живого вещества. Суть этой закономерности заключается в том, что
в ходе приспособления первичных организмов к геохимическим факторам среды
был отобран тот комплекс элементов, который создает структурную и функциональную
основу жизни.
Предложенная Шоу таблица не только дает возможность в графическом виде
суммировать современные знания о биологических свойствах элементов, но
и позволяет в принципе предсказывать элементы, биологические функции которых
пока еще неизвестны. Так, например, Шоу предсказывал участие лития и бериллия
в образовании ферментативных систем.
Дальнейшее усовершенствование и уточнение биогеохимической периодической
таблицы предполагает углубленное изучение химико-физических свойств отдельных
элементов, оказавшихся жизненно необходимыми. В связи с этим огромное значение
имеет получившая широкую известность работа Дж. Уолда (1964) о причинах
преимущественного использования жизнью элементов первых трех периодов системы
Менделеева. Атомы этих периодов, будучи наименьшего размера, „приобретают
стабильные электронные конфигурации после присоединения соответственно
1, 2, 3, 4 электронов” (там же, с. 103). Они образуют устойчивые связи,
имеющие кратный характер.
В заключение этого раздела уместно вспомнить, что интенсивные экспериментальные
исследования по синтезу низкомолекулярных органических веществ показали,
что все они являются термодинамически и энергетически наиболее вероятными
продуктами, возникающими под контролем химического отбора (Буханан, 1966,
с. 109). Случайные смеси атомов углерода, водорода, кислорода, азота,
фосфора, серы и их простейших молекул с необходимостью приводят к синтезу
соединений,
составляющих основу белков, углеводов, нуклеотидов и т. д. (см. обзоры:
Кальвин, 1971; Руттен, 1973; Фокс, Дозе, 1975; Поннамперума, 1977, и
др.). Было доказано, что химические свойства этих элементов определяют
их высокую
активность и способность избирательно вступать во взаимодействия, ведущие
к возникновению более сложных соединений различной степени устойчивости
и подвижности. Характерная для них делокализация электронов повышает
стабильность молекул и дает выигрыш в энергии („энергия резонанса”). Образовавшиеся
из них молекулы нуклеотидов, флавинов, аминокислот характеризуются в
свою
очередь рядом специфических свойств: низкая энергия электронного
70
возбуждения, низкий потенциал ионизации, высокое сродство к электрону,
высокая электронная поляризуемость и т. д. Ясно, что и в этом „подборе
свойств” кроется „нечто существенное для функций живых систем” (Блюменфельд,
1977, с. 14).
3.2. Общие закономерности химической дифференциации
живого вещества в биосфере
Химический состав живого вещества в своих основных чертах сложился еще
в процессах предбиологической эволюции, когда шел отбор химических элементов
и соединений, необходимых для поддержания целостности метаболических процессов
и передачи наследственной информации (А. И. Опарин, Н. Пири, Е. М. Крепе,
А. Львов и др.). Но происходящая затем экспансия жизни, ее приспособление
к различным геохимическим средам, разделение организмов на автотрофы и
гетеротрофы сопровождались нарастающей дифференциацией живого вещества.
На этот аспект в изучении закономерностей химического состава живого вещества
не раз указывал В.И.Вернадский. Он подчеркивал, что „химический состав
не является одинаковым на всем лике Земли, но резко меняется, как функция
литологического состава и климатических зон, в разных частях биосферы в
связи с литологией верхних частей планеты” (1965, с. 63). Среди факторов,
обусловливающих изменение химического состава живого вещества в пределах
климатических зон, Вернадский называл высоту над уровнем моря, близость
океана, химический состав почв и горных пород. Вместе с тем он отмечал,
что химический состав живого вещества изменяется не случайно, а „различен
закономерно” (там же). Вернадский по существу сформулировал положение о
зависимости биохимических процессов, химического состава организмов и их
эволюционных преобразований от геохимических факторов среды.
Эти идеи Вернадского получили обоснование и развитие в трудах А. П. Виноградова
(1933, 1935а, 1944 и др.), который считал, что химическая дифференциация
живого вещества связана прежде всего с завоеванием жизнью принципиально
новых в геохимическом отношении сред обитания. Переход морских форм в
пресноводные бассейны, а тем более их выход на сушу он считал наиболее
крупными событиями
в эволюции химического состава организмов. Поэтому существенные различия
химического состава в пределах царств наблюдаются между таксонами, обитающими
в морских и наземных сообществах. Морские организмы содержат гораздо
больше химических соединений нечетного ряда, чем имеется этих элементов
у наземных
организмов. Эти отличия особенно ярко проявляются при сравнении водорослей
и ксерофитов. Водоросли содержат гораздо больше водорода, бора, фтора,
натрия, алюминия, чем любое ксерофильное растение. Эти различия в значительной
степени обусловлены тем, что многие элементы нечетного ряда относятся
по классификации Гольдшмидта к числу талласофильных элементов, т. е. их
содержание
в океанской воде выше, чем в почвах (например, натрий, хлор, бор и т.д.).
71
В целом этот вывод хорошо согласуется со сводками по сравнительному содержанию
химических элементов в организмах (см., например: Bowen, 1966). При всей
приблизительности средних характеристик, выводимых для всех наземных или
всех морских растений или животных, из составленных Г. Бовеном таблиц (р.
68—73) сравнительного содержания химических элементов видно, что различия
между морскими и наземными растениями или морскими или наземными животными
очень значительны. Морские водоросли содержат углерода 34500 мг/100 г сухого
вещества, азота – 1500, водорода – 4100, кальция – 1150, магния – 520,
натрия – 3300, калия – 5200, серы – 1200. В наземных растениях, например
покрытосеменных, эти показатели выглядят следующим образом: углерода –
45 400, азота – 3000, водорода – 5500, кальция – 1800, магния – 320, натрия
– 120, калия – 1400, серы – 340. У морских и наземных животных отличия
также достаточно отчетливы, хотя и значительно меньше, чем между растениями.
В заключительных строках своей книги „Химический элементарный состав организмов
моря” (1944) Виноградов подчеркивал, что среди обитателей гидросферы наиболее
сильное изменение претерпевали организмы, переселяющиеся в филогенезе из
моря в солоноватые воды и затем в пресные (или наоборот). В этом случае
химические изменения были значительнее, чем у организмов, все развитие
которых в течение геологического времени проходит в море.
Собранный Виноградовым материал о географической изменчивости химического
состава организмов свидетельствовал о том, что химические особенности
отдельных регионов биосферы накладывают глубокий отпечаток на их обитателей,
вызывая
далеко идущую конвергенцию химического состава у филогенетически далеких
друг от друга таксонов. Он считал необходимым выяснить механизм глубокого
и тонкого влияния, которое оказывает нивелирующее действие среды на организмы,
имеющие общие местообитания. „Это влияние распространяется на целые области,
образуя зоны, в которых морские организмы имеют сходные черты в химическом
составе” (Виноградов, 1937, с. 40). Так, изменение в широтном направлении
концентраций карбоната кальция, окиси кремния, йода, магния в морских
водах обусловливает и географическую изменчивость у морских организмов
по концентрационным
способностям этих элементов и соединений. В результате водоросли теплых
и экваториальных морей, воды которых насыщены углекислым газом, характеризуются
сильно выраженной способностью концентрировать карбонат кальция. Такой
же способностью обладают и обитающие здесь беспозвоночные, результаты
геохимической деятельности которых хорошо известны в виде рифового известняка.
Планктон
теплых морей содержит многочисленные организмы с известковым скелетом.
С перемещением в более холодные воды такие организмы встречаются все
реже, а главную роль в планктоне играют диатомовые с кремниевым скелетом,
рачки
с хитиновым скелетом и т. д. Для обитателей умеренных и северных бассейнов
характерно более высокое содержание йода, в то же время количество магния
в скелетах иглокожих увеличивается по мере
72
продвижения их от полюса к экватору. И подобных примеров в книге Виноградова
очень много.
Значительные изменения происходят и при перемещении организмов в глубоководные
области моря. По мере опускания в абиссальные области исчезают организмы
с известковым скелетом, замещаясь формами с органическим или кремниевым
скелетом. Это объясняется прежде всего недостаточностью кальция в среде.
На основе этих данных Виноградов имел полное основание для вывода о том,
что химические факторы внешней среды оказывают огромное влияние на химический
состав живого вещества и на географическое распределение организмов в биосфере.
В то же время он был далек от распространенных в те годы механоламаркистских
представлений, нашедших свое отражение и в некоторых исследованиях о влиянии
геохимических факторов среды на эволюцию организмов. Так, Ф. Нопча (Nopcsa,
1923) полагал, что сильное уплотнение и утолщение костей у некоторых ископаемых
четвероногих, вернувшихся к морскому образу жизни, возникают в результате
наследственного закрепления патологических явлений (пахиостиоз). Эти явления
он считал результатом реакции крови на изменения химических факторов среды.
Упрощенные механоламаркистские рассуждения свойственны и палеобиогеохимическим
работам Ю. Вильзера.
Виноградов не пошел по заманчиво легкому пути объяснения химической эволюции
организмов как результата прямого формообразующего воздействия среды с
последующим наследственным закреплением приобретаемых признаков. Им в 40—50-х
годах опубликованы таблицы среднего химического состава почв, гидросферы,
литосферы, атмосферы и живого вещества.1 Из таблиц видно,
что средний состав растений по отдельным элементам в десятки и сотни
раз отличается от среднего
состава этих элементов в геосферах. Аналогичная картина и у животных,
хотя и отличающаяся по отдельным элементам.
Вопреки гипотезам о наследовании приобретенных признаков. Виноградов подчеркивал,
что „химический элементарный состав, как показывают нам тысячи анализов,
не является простым отражением, повторением химического состава среды”
(Виноградов, 1944, с. 188), а складывается в длительных эволюционных преобразованиях,
обусловленных взаимодействиями организмов с внешними факторами. Он не раз
отмечал, что естественный отбор является главной причиной приспособления
организмов к химическим особенностям среды. Тем самым возникала возможность
для синтеза биогеохимии с учением об естественном отборе, которая позднее
была реализована в ряде работ по геохимической экологии.
Влияние геохимических факторов на эволюцию химического состава организмов,
по мнению Виноградова (1944), всегда было опосредовано совокупностью регуляторных
механизмов, контролирующих и направляющих химические взаимодействия организмов
с окружающей средой.
1 Эти таблицы приведены в комментариях Виноградова к
книге Вернадского „Химическое строение биосферы Земли и ее окружения» (с.
341—346).
73
Эти механизмы, выработанные в ходе филогенеза, обеспечивают стабильность
химического состава организмов, необходимую для нормального протекания
всех биохимических и физиологических процессов. Возрастание стабильности
химического состава и независимости его от резких колебаний в геохимической
среде является одной из важнейших закономерностей арогенной эволюции. Особенно
ярко она проявилась в филогенезе животных, где создание кровеносной системы,
а также механизмов нервной и гуморальной регуляции обеспечило высокую стабильность
химического состава организмов. Противоположная картина наблюдается у прокариот
и одноклеточных эукариот, где „внутри каждого класса исключительное разнообразие
типов своего химического состава, отличающихся легкой изменчивостью состава”
(1944, с. 194).
Важно отметить, что одновременно с Виноградовым сходную закономерность,
т. е. возрастание независимости от внешней среды, и в эволюции морфологических
признаков и органов установили А. А. Машковцев (1936) и И. И. Шмальгаузен
(1938). Вероятно, стабилизация химического состава является одним из следствий
общей тенденции прогрессивной эволюции в сторону выработки защитных механизмов
индивидуального развития и усовершенствования аппарата его регуляции.
Как одно из подтверждений установленной Виноградовым закономерности, по-видимому,
можно рассматривать результаты исследований различий в эволюционных преобразованиях
у позвоночных животных разных классов (Шварц и др., 1965). Если в эволюции
млекопитающих наблюдается значительная устойчивость биохимически-тканевого
уровня организации, то у рыб уже при микроэволюционных изменениях затрагиваются
биохимический и тканевой уровни организации. Поэтому у млекопитающих виды
одного семейства в биохимическом отношении дифференцированы слабо, а у
рыб внутривидовые группы отличаются целым комплексом биохимических особенностей.
Высокая изменчивость биохимического уровня организации рыб обеспечивает
их существование в различных водоемах при отсутствии устойчивости внутренней
среды организмов.
Интересная попытка оценить роль геохимических факторов в эволюции организмов
сделана Б. Б. Шлыковым (1948). Он исходил из укоренившейся в геохимии классификации
зольных элементов, которые были разделены В. Гольдшмидтом на пять групп:
сидерофильные, халькофильные, литофильные, атмофильные и биофильные. Поскольку
каждая из этих групп концентрируется преимущественно в разных геосферах,
то, как полагал Полынов, данная классификация в какой-то степени отражает
геохимическую историю нашей планеты. Биофильная группа, насчитывающая 17
элементов, по мнению Гольдшмидта, объединяет элементы, которые лишь в позднейшие
фазы эволюции Земли стали концентрироваться в организмах. Вскоре Вернадский
добавил к этим элементам еще шесть. Виноградов же считал, что обязательное
присутствие в организмах установлено по крайней мере для 40 элементов.
Полынов посчитал возможным применить исторический подход и при классификации
биофильных элементов. Он предложил встречающиеся в организмах элементы
разделить по их биологической значимости
74

Борис Борисович Полынов (1877-1952).
на две группы: органогены и примеси. В свою очередь органогены включают
как элементы, без которых невозможно существование любых организмов (абсолютные
органогены: Н, С, О, N, Mg, К, Р, S), так и элементы, необходимые только
для некоторых крупных таксонов (специфические органогены: Си, Na, Ca, Fe,
Cl, F, Mn, Sr, В, Zn, Be, I).
Полынов специально подчеркивал, что предлагаемая им классификация отражает
современные отношения элементов биосферы к живому веществу, которые неизбежно
менялись в ходе исторического развития жизни. Поскольку в состав биофилов
входят типичные литофилы, сидерофилы и атмофилы, то такое сочетание не
могла обеспечить ни одна из первоначальных геосфер Земли в отдельности.
Такое сочетание элементов могло появиться на поверхности „первозданной»
литосферы, орошаемой водой и соприкасаемой с атмосферой. Иначе говоря,
эти условия аналогичны тем, в которых на магматических породах формируются
аллювиальные почвы. В связи с этим Полынов предложил
75
оригинальную гипотезу происхождения жизни и первых этапов почвообразовательного
процесса, на которой мы остановимся ниже (гл. 5). Здесь же отметим только,
что, согласно гипотезе Полынова, в условиях прискального образования почв
формирующиеся предбиологические системы могли получить все необходимые
для жизни абсолютные органогены. Специфические же органогены, по мнению
Полынова, не играли существенной роли на начальных этапах эволюции живого
и встречались лишь в качестве примесей у некоторых организмов, живущих
в специфических в химическом отношении регионах биосферы. Постоянно поглощая
эти элементы и приспосабливаясь к их участию в метаболических процессах,
организмы постепенно превратили их в специфические органогены, необходимые
для нормального протекания биохимических физиологических процессов у данной
группы.
Как видно, Полынов сильно сузил число абсолютных органогенов. Уже вскоре
В. А. Ковда (1956), анализируя классификацию Полынова, показал, что в число
абсолютных органогенов следует включить также йод, бор, кальций, железо,
медь, кобальт. Их отсутствие ведет к резким нарушениям в эмбриональном
развитии и в итоге к летальному исходу. Согласно современным данным (Войнар,
1960; Bowen, 1966; Ковальский, 1974), в составе организмов определено 67
элементов. Из них по меньшей мере 46 являются органогенами в смысле Полынова.
Но следует учитывать, что при современном уровне знаний нет возможности
окончательно сказать, какие из них являются абсолютными органогенами, а
какие - специфическими. Количественные определения для многих микроэлементов
очень скудны и требуют дополнительных исследований и проверок. Возможно
в принципе и открытие новых органогенов. Поэтому окончательные классификации
элементов на абсолютные и специфические органогены — дело будущего. Но
сам предложенный Шлыковым эволюционный подход к оценке роли химических
элементов совпадает с принципами последующих классификаций атомов по выполняемым
ими биогеохимическим, биохимическим и физиологическим функциям (В. А. Ковда,
А. И. Перельман, В. Андервуд и др.).
Фудзио Эгами (1983) предлагает разделить химические элементы на пять групп
в соответствии с их поведением и концентрацией в морской воде — предполагаемой
„колыбели” жизни. В первую группу он включает элементы, наиболее распространенные
в морской воде. В нее входят те биологически значимые элементы, за исключением
фосфора, которые составляют основную массу живого вещества. В последнюю
же пятую группу Эгами помещает элементы типа свинца и ртути, токсичные
для организмов.
Прежде чем закончить этот раздел, остановимся на интересных работах А.
И. Перельмана (1981, 1982) о влиянии геохимических факторов на эволюцию
органического мира. В центре внимания Перельмана оказался вопрос о влиянии
геохимических особенностей природных вод на обитающие в них организмы.
Он попытался кратко рассмотреть, как воздействуют различные классы природных
вод (сильнокислые, кислые, нейтральные, слабощелочные, сильнощелочные
и т.д.) на процессы видообразования. Он отмечает, что многие элементы оказывают
особое
76
воздействие на некоторые системы органов животных и растений: железо,
кобальт, медь — на кровеносные системы; йод — на щитовидную железу; кальций,
фтор, стронций и кремний — на скелет и т.д. Поскольку морские животные
содержат брома больше по сравнению с наземными (соответственно 4 · 10-3
и 2.5 · 10-5 на грамм живого веса), то Перельман считает, что высокое
содержание брома в морской воде „могло замедлять цефализацию” (1982,
с. 111). Перельман
назвал это явление „бромным прессом на нервную систему”, который был
снят лишь с освоением животными пресноводных водоемов и суши. Автор предлагает
целую программу комплексных геохимических, сравнительно-физиологических
и эволюционно-биологических исследований в современных морских, пресноводных
и наземных экосистемах с целью установить роль геохимических факторов
в
эволюции органического мира. Реализация этой исследовательской программы
остается делом будущего.
Работы по химическому составу организмов и факторах его эволюции, по мнению
Камшилова, представляют „большой интерес для эволюциониста, указывая новый
аспект в изучении эволюции и значительно углубляя наши знания о закономерностях
этого процесса” (1947, с. 310). Тем не менее результаты исследования Виноградова,
Перельмана, Полынова и других биогеохимиков очень редко упоминаются в работах
по эволюционной теории, вышедших за последние годы как у нас в стране,
так и за рубежом. Так, уникальные данные Виноградова о закономерностях
географической изменчивости химического состава морских организмов даже
не упомянуты в обстоятельной работе Е. И. Лукина (1983), посвященной специально
истории изучения этих закономерностей в нашей стране и опубликованной в
книге „Развитие эволюционной теории в СССР”. Исключение, увы, подтверждающее
правило, составляет только книга Н. В. Тимофеева-Ресовского, Н. Н. Воронцова,
А. В. Яблокова (1969) „Краткий очерк теории эволюции”. Здесь в разделе
об эволюции биосферы сказано буквально несколько слов о биогеохимических
исследованиях Полынова и Перельмана. Ссылок же на работы Виноградова нет.
Подобное положение, видимо, не случайно. Дело в том, что в рассмотренных
работах основное внимание уделялось среднему химическому составу или всего
живого вещества, или крупных таксонов. Иначе говоря, речь шла о макроэволюции.
Такой подход, имеющий решающее значение для восстановления картины эволюции
и обоснования закономерностей филогенеза, мало что дает для понимания каузальных
основ эволюции. Естественно, что в современный синтез эволюционных знаний,
нацеленный прежде всего на выяснение факторов и движущих сил микроэволюции,
интересные выводы Виноградова о филогенетических закономерностях эволюции
живого оказались не включенными.
3.3. Исследование биогеохимических провинций
и их роли в эволюции биосферы
Важный шаг к преодолению разрыва между изучением геохимических факторов
макроэволюции и эволюционных преобразований, происходящих внутри популяций
и видов, был сделан Виноградовым еще
77

Александр Павлович Виноградов (1895-1975).
в конце 30-х годов в работе
о биогеохимических провинциях. Так, он называл районы биосферы, в которых
повышенная или пониженная концентрация каких-либо
химических элементов предопределяет специфику видового состава животных
и растений. В условиях избыточности или недостаточности одного или
нескольких элементов (Li, В, F, Mg, Si, P, S, Ca, Cr, Co, Ni, Cu, Zn,
Se, I, Mo,
Pb и др.) организмы или гибнут, или претерпевают резкие фенотипические
изменения.1 У них возникают различного рода эндемические
реакции, носящие чаще всего характер морфозов, дисфункций, эндемических
заболеваний
1 Такие провинции были впервые выделены А. П. Виноградовым
и А. М. Симориным в 30-х годах при изучении районов уровской эндемии в
Читинской
обл. и провинций
эндемичного зоба в Дагестане. Для разработки представлений о биогеохимических
провинциях исходными были, по-видимому, идеи Вернадского о „химических
областях биосферы” (1978), в которых количество какого-либо элемента
резко отличается от его среднего содержания в земной коре или коре
выветривания.
А. Е. Ферсман (1934) предложил такие районы называть геохимическими
провинциями. Это название было использовано и в совместном докладе
В. И. Вернадского
и А. П. Виноградова 5 июня 1936 г. на заседании Московского терапевтического
общества.
78
(остеодистрофия, рахит, зоб, атаксия, анемия, гипо- и авитаминозы, хлорозы
и т. д.). Подобные отклонения нередко ведут к резкому снижению жизнеспособности
и плодовитости организмов, и их носители подвергаются сильному негативному
отбору. Специфичность флоры и фауны выражается, однако, не только в возрастании
числа вредных модификаций, но и в формировании эндемичных, наследственно
закрепленных форм. Их возникновение, по мнению Виноградова, обусловлено
действием естественного отбора, направленного в сторону выработки адаптации
к специфической геохимической среде. Виноградов полагал, что „через существующие
и вновь возникающие биогеохимические провинции распространяются разнообразные
флоры и связанные с ними фауны. Биогеохимическая провинция является этапом
на их пути, поэтому рассмотрение и изучение влияния этих провинций на эволюцию
организмов вследствие влияния их на отбор представляет глубоко интересную
задачу” (1938, с. 286).
С эволюционной точки зрения особенно важно, что адаптивные изменения в
этих провинциях одновременно захватывают разные группы организмов. Такие
формы оказываются устойчивыми к избытку или недостатку тех или иных элементов
в почвах, в почвенных, подпочвенных растворах и т. д. по сравнению с их
общими кларками в биосфере. Впоследствии Виноградов не раз возвращался
к вопросу о роли биогеохимических провинций в эволюции органического мира.
В статье, посвященной задачам химического исследования биосферы (1945),
он отметил особое значение изучения влияния среды на химический состав
организмов при их расселении в пределах той или иной биогеохимической провинции
или при резком изменении какого-либо элемента в данном районе. При этом
возникают разные типы морфологических, наследственно обусловленных реакций
у организмов одного и того же вида на изменения в концентрациях элементов.
Одни организмы, перенесенные в области, отличающиеся от исходной по уровню
концентрации отдельных элементов, нормально развиваются и размножаются.
У них незаметно морфологических отклонений, хотя химический состав и претерпевает
некоторые изменения. В то же время популяции, взятые из других мест, страдали
от морфозов и погибали. Все это свидетельствовало о том, что изменения
геохимических факторов могут становиться ведущими агентами в формировании
адаптивной нормы популяции. Наследственно закрепленной становится и потребность
организмов в тех или иных концентрациях химических элементов. „Флора и
фауна, — писал Виноградов, — шествуя в геологическом времени через ряд
биогеохимических провинций, должны были подвергнуться отбору и измениться,
и в частности изменить свой химический состав” (1945, с. 353). Виноградов
(1963) выделил два типа биогеохимических провинций в современной биосфере.
Первый тип характерен для определенных почвенно-климатических зон. Например,
в лесной зоне часто встречаются области с недостатком йода, кальция,
кобальта, меди и т. д. Такие провинции приурочены к подзолистым и дерново-подзолистым
почвам и никогда не встречаются на черноземных почвах. Второй тип биогеохимических
провинций не связан с природными зонами. Они обычно
79
возникают в местах повышенного содержания какого-либо элемента, вызванного
геологическими причинами.
Ко времени публикации работ Виноградова в начале 60-х годов было известно
более 30 элементов, колебания концентраций которых вели к возникновению
биогеохимических провинций. Накопился и новый материал об изменчивости
организмов в этих регионах. Опираясь на эти данные, Виноградов (1963) выделил
три типа реакции организмов на условия биогеохимических провинций: 1) естественный
отбор форм, оказавшихся в условиях новой для них биогеохимической провинции;
2) появление ненаследственных изменений; 3) формирование новых рас, подвидов
и видов. В целом же он не касался вопроса о механизмах перехода ненаследственных
изменений, вызванных геохимическими факторами, в наследственно закрепленный
признак. И это, возможно, объясняется его намерением не вмешиваться в споры
сторонников и противников идеи наследования приобретаемых признаков.
Виноградов был уверен, что „изучение эволюции органического мира на геохимической
основе с привлечением обширных сведений из палеонтологии, систематики,
химической экологии и сравнительной биохимии привело бы к новым открытиям
в области эволюции органического мира” (1963, с. 212). Он считал возможным
геохимическими факторами объяснять смену кровяного пигмента гемоцианина
на гемоглобин, появление теплокровности и т. д. Но все эти сюжеты о геохимических
факторах макроэволюционных событий затрагивались им вскользь, лишь в плане
общей постановки вопроса.
Виноградов (1960) считал полезным выделять и палеобиогеохимические провинции,
образующиеся в результате действия геологических процессов. Вызывая резкую
геохимическую дифференциацию биосферы, процессы орогенеза, трансгрессии
и регрессии, породообразования и вулканизма обусловливают различные направления
биогеохимической эволюции целых биологических сообществ, а тем самым и
эволюцию всей биогеохимической и энергетической обстановки в биосфере.
Особенно велика роль горообразовательных процессов, вызывающих дифференциацию
горных пород в соседних долинах, быструю изоляцию биогеохимических провинций
и т. д. Не случайно центры происхождения многих групп растений приурочены
к горам, где изоляция биогеохимических провинций создавала благоприятные
условия для быстрого видообразования. „В геологическом прошлом, — писал
Виноградов, — биогеохимические провинции ... должны были играть значительную
роль в преобразовании флоры и фауны” (1963, с. 210).
Подчеркивая, что эволюция органического мира является и геологическим процессом,
Виноградов считал необходимым изучать ее с точки зрения влияния геохимических
факторов, и прежде всего избыточности или недостаточности концентраций
химических элементов и соединений, на процессы видообразования. В частности,
он отмечал, что виды растений и животных с высоким содержанием натрия
и хлора, вероятнее всего, возникли в бессточных областях или на морских
побережьях,
хотя в настоящее время они обитают не только на солончаках и в соленых
водоемах. Возникновение растений с высоким содержанием алюминия
80
шло, по мнению Виноградова, на латеритах, а пески и вулканические почвы
являются первичными областями распространения злаков. Эти и другие примеры
Виноградов не раз приводил в своих работах с целью показать воздействие
геохимических факторов на формирование специфической флоры и фауны в отдельных
регионах биосферы, В то же время он вынужден был признать, что современный
уровень знаний позволяет только в самых общих чертах объяснить роль геохимических
факторов в появлении эволюционных новшеств. В связи с этим предполагалось
развивать химическую экологию, которая, устанавливая корреляции незначительных
изменений органических форм с геохимическими условиями среды, должна выяснить
роль последних в обшей эволюции флоры и фауны.
Сформулированные Виноградовым представления о биогеохимических провинциях
получили дальнейшее фактическое обоснование и конкретизацию в многочисленных
исследованиях. Так, с 1944 г. проводились исследования биогеохимических
провинций таежных и лесных нечерноземных областей биосферы. Были выявлены
провинции, характеризующиеся недостатком кобальта, что вело к возникновению
у животных эндемичных гипо- и авитаминозов В12 (В. В. Ковальский, В. С.
Чебаевская, Ю. И. Раецкая, Я. М. Берзиня), недостатком меди, обусловливающим
анемию у животных, недостатком йода, вызывающим эндемичный зоб, бесплодие
(В. В. Ковальский, М. И. Густун, Р. И. Блохина и др.). После 1954 г. объем
исследований в этом направлении резко увеличился. Были открыты провинции,
характеризующиеся недостатком двух и более элементов, а также провинции,
в которых недостаток одного какого-либо элемента (например, кальция) сочетается
с избытком другого (стронция). В биогеохимическое районирование биосферы
и изучение отдельных биогеохимических провинций были втянуты в дальнейшем
и другие регионы биосферы. Затрагивались и вопросы о механизмах воздействия
геохимических факторов на формирование структуры биогеоценозов и на их
функционирование, об изменчивости различных звеньев биогеохимических пищевых
цепей, об изменении биогенных циклов элементов, участвующих в метаболических
процессах и органических соединениях. Было показано, что органический мир,
вовлекая химические элементы в биогенную миграцию, не только преобразует
среду своего обитания, но и эволюционирует вместе с ней.
Мы не будем подробно останавливаться на этих работах, так как обзор проведенных
исследований можно найти в обобщающих сводках по биогеохимии (Лукашев,
Банковская, 1973; Ковальский, 19746, 1982). Отметим только, что они позволили
изучить различные биогеохимические провинции нашей страны, особенности
существующих в них трофических структур и биогенных циклов элементов,
своеобразие населяющих их организмов. Тем самым они сыграли важную роль
в разработке
биогеохимической концепции биосферы, подняв изучение особенностей отдельных
биологических сообществ до биосферного уровня. Было выяснено, что „химический
состав элементов биосферы — почв, природных вод, организмов - обнаруживает
географическую неоднородность, носит мозаичный характер” (Ковальский,
1963б, с. 830).
81
но мнению В. Н. Сукачева, подобная „мозаичность биосферы и есть следствие
того, что химизм природных явлений, связанных с ней, отражает ее сложение
из различных биогеоценозов” (1964а, с. 30). О размахе химической гетерогенности
лито-, педо- и гидросфер красноречиво свидетельствуют лишь некоторые
данные, приведенные в последней книге В. В. Ковальского „Геохимическая среда
и
жизнь” (1982). Так, содержание меди в почве колеблется от 0.1 · 10-4
до 154 · 10-4 %, т.е. в 1500 раз, цинка — от 0.1 · 10-3 до
99 · 10-4, т.
е. в
1000 раз. Аналогичны данные о колебаниях в почвах содержания кобальта,
марганца, стронция и др. Наземные пресные воды также могут различаться
между собой по содержанию кобальта – почти в 330 раз, бора – в 33, меди
– в 40, стронция – в 100, цинка – в 220 раз. В десятки и сотни раз отличаются
концентрации этих элементов и в тропосфере.
Эти различия являются результатом эволюции, происходящей на биоценотическом
уровне. Биогеохимические изменения биосферы в пространстве в какой-то степени
отражают ее изменения во времени, а ее отдельные биогеохимические провинции
являются этапами этих изменений. Сравнительные методы изучения ныне существующего
разнообразия органического мира, как известно, дают богатую информацию
о прошедших эволюционных событиях. Изучение особенностей факторов и движущих
сил эволюции у отдельных таксономических групп по существу является единственным
надежным способом для разработки проблемы „эволюция эволюции” (Завадский,
Колчинский, 1977). Подобно этому и развитие сравнительно-исторических исследований
различных биогеохимических провинций должно стать важным источником знаний
об основных этапах и закономерностях биогеохимической эволюции биосферы.
Биогеохимическое районирование биосферы несет в себе важную экологическую
информацию, позволяющую понять не только механизм функционирования биосферы,
но и ее эволюцию. Ведущее значение сравнительных исследований современных
биогеохимических провинций объясняется тем, что данные о палеобиогеохимических
провинциях или полностью отсутствуют, или крайне скудны. Их познание оказывается
возможным только на основе сравнительных исследований современных провинций,
дополненных некоторыми, очень отрывочными сведениями о химическом составе
ископаемых почв и его предполагаемом влиянии на обитающих в тех провинциях
животных и растений. Примером такого анализа палеобиогеохимических провинций
может быть работа А. И. Перельмана и Е. Н. Борисенко (1965) о пермской
палеобиогеохимической провинции в Приуралье.
Необходимо отметить также, что изучение особенностей эволюционного процесса
в отдельных биогеохимических провинциях позволяет не только объединить
основные положения эволюционной теории и биогеохимии, но и через изучение
адаптивных преобразований популяций приступить к изучению эволюции биотического
круговорота веществ. Популяция является не только элементарной единицей
эволюционного процесса, но и элементарной биогеохимической и энергетической
единицей биогеоценоза (Шварц, 1973). Существует тесная связь между
82
процессами видообразования и эволюцией экосистем (биогеоценозов) (Пианка,
1981). Так, например, усовершенствование защиты жертвы ведет к уменьшению
переноса энергии с одного трофического уровня на другой, в то время как
эволюция хищника ведет, напротив, к увеличению переносимой энергии. Полифагность
хищников, их способность перестраивать свою диету в зависимости от доступности
той или иной жертвы обеспечивает дополнительный механизм для сохранения
популяций жертв, а в итоге для стабилизации экосистемы.
Необратимые изменения популяции по биохимическим или энергетическим признакам
так или иначе скажутся на функционировании биогеоценоза (Наумов, 1967).
Поэтому преобразование химического состава популяции и ее геохимической
энергии по сути дела представляет элементарный эволюционный акт в биогеохимической
эволюции биосферы, В биогеохимической провинции специфическое направление
естественного отбора захватывает не отдельные популяции, а целые флоры
и фауны данного региона. Это неизбежно сказывается на всей сложившейся
биогеохимической и энергетической структуре сообщества, на различного рода
трофических, конкурентных, кооперативных взаимодействиях.
В связи с этим особую актуальность приобретает проблема геохимических факторов
микроэволюции, которая в последние десятилетия стала разрабатываться в
рамках геохимической экологии. Ее общетеоретические и эволюционные положения
были сформулированы в серии работ В. В. Ковальского (1941, 1943, 1957,
1963б, 1974 и др.).
3.4. Геохимические факторы микроэволюции и ее влияние
на биогенные циклы элементов
Ковальский исходил из того, что геохимические факторы воздействуют на
все формы организации живого. Эти воздействия проявляются как в изменениях
активности ферментов, так и в преобразовании форм миграции химических элементов
в экосистемах. Поэтому при их изучении внутрипопуляционные и внутриорганизменные
процессы можно рассматривать как составные элементы единой целостной системы
жизни.
Если Вернадский и Виноградов основное внимание уделяли устойчивости химического
состава вида, то Ковальский в центр своих работ поставил задачу выяснить
причины внутривидовой изменчивости организмов. Уже в одной из первых статей
по проблемам биогеохимии он сформулировал цель будущих исследований: „Изучение
явлений химической изменчивости организмов представляет основную предпосылку
для установления в фило- и онтогенетическом развитии химических особенностей
организмов и для исследования химических сторон эволюционного процесса (курсив
наш, — Э. К.), т. е. для построения всей сравнительной биохимии” (1941,
с. 381). При изучении химического состава организмов он считал необходимым
учитывать их пол, возраст, норму реакции и т. д.
83

Виктор Владиславович Ковальский (1899 – 1984).
Ковальский полагал, что диапазон изменчивости химического состава организма
обусловлен его „видовой” историей и связан с его приспособлением к естественным
ритмическим колебаниям среды. Особенно важно исследовать сезонные колебания
химического состава среды, так как в зависимости от сезона меняется направление
и интенсивность действия борьбы за существование и отбор. Сезонная изменчивость
химического состава организмов, как полагал Ковальский, имела большое эволюционное
значение. Она связана с космической сезонной ритмикой, являющейся одним
из главных факторов эволюции биосферы. Сезонная ритмика способствует выработке
адаптации широкого значения, позволяющих организмам выжить в колеблющихся
условиях среды.
Позднее он сформулировал главные задачи изучения геохимических факторов
микроэволюции следующим образом. „Установление связей между химическими
элементами среды и приспособительной изменчивостью организмов, а также
перестройки обмена веществ в условиях различных биогеохимических провинций
позволяют создавать геохимическую экологию растений и животных, которая
выясняет закономер-
84
ности приспособляемости организмов к различному содержанию химических
элементов в среде, химической и морфологической их изменчивости, расселения
организмов. Изучение этих вопросов служит важной основой развития эволюционной
физиологии и биогеохимии, задачами которых являются не только познание
закономерностей химических и физиологических изменений организмов, но и
выяснение химических и физиологических закономерностей эволюционного процесса
— изменчивости, приспособляемости, видообразования и естественного отбора”
(Ковальский, 1960, с. 32).
Реализуя эту программу, Ковальский показал, что на популяционно-видовом
и организменном уровнях организации живого воздействие геохимических факторов
проявляется в изменении динамики численности и воспроизводства популяции,
в ее фенотипической и генетической структуре, в преобразовании процессов
морфогенеза, метаболизма, химического состава и т. д. Организмы сохраняют
нормальную жизнеспособность и сбалансированность всех физиологических и
биохимических процессов в пределах определенного диапазона в изменении
геохимических условий. Эта сбалансированность обеспечивается регуляционными
механизмами, закрепленными генетически, нарушение которых означает выход
концентрации тех или иных химических элементов и соединений за пределы
пороговой чувствительности организмов.
Внутри каждой популяции существует генотипическая гетерогенность по чувствительности
организмов к геохимическим условиям. Эта гетерогенность проявляется особенно
ярко при воздействии экстремальными дозами, когда у части организмов возникают
различного рода эндемичные заболевания, нарушения морфогенеза, дисфункции,
подавление репродуктивной способности и т. д. В этих условиях и выявляются
группы особей с повышенной толерантностью к необычным концентрациям химических
элементов. В условиях значительных колебаний геохимических факторов, по
мнению Ковальского, возрастает темп эволюционных преобразований.
Особенно важным для понимания роли геохимических факторов эволюции, как
справедливо отмечал В. Н.Сукачев (1964а), был разработанный Ковальским
комплексный подход к изучению этих факторов. Он показал, что „никогда в
природе не действует изолированно один химический элемент, значение имеет
соотношение между элементами. Поэтому при изменении концентрации какого-либо
элемента в организмах наблюдается не только усиление или ослабление отдельных
процессов, но и дисфункции процессов обмена веществ” (1960, с. 17).
Эти сформулированные Ковальским положения стали теоретической базой для
многочисленных исследований причинных зависимостей между геохимическими
факторами среды и разнообразными биологическими реакциями организмов
и популяций. Следует отметить необычайно большой размах проводимых исследований.
Их объектами стали представители практически всех крупных таксономических
групп от бактерий и актиномицетов до цветковых растений и высших позвоночных.
Своеобразие морфологических, физиологических и биохимических адаптации
организмов изучалось в различных биогеохимических провинциях
85
и зонах СССР. Проводились и экспериментальные исследования реакции организмов
на изменения концентраций того или иного химического элемента, моделировались
селективные процессы выработки адаптации к необычным геохимическим условиям.
Рассмотрим кратко основные результаты этих исследований.
Экспериментальное изучение адаптации микроорганизмов получило широкое развитие
еще в начале 30-х годов. Во многих работах были получены интересные данные
о „приручении” микроорганизмов к повышенным содержаниям лития, меди, кобальта,
селена (см.: Имшенецкий, 1956). Новизна геохимико-экологического подхода
заключалась в том, что изучались адаптации микроорганизмов к геохимическим
условиям, воспроизводящим природную обстановку, а также ставились эксперименты
с целью изменить эти адаптации. На базе широко используемого в экологии
микроорганизмов метода сравнительного изучения поведения отдельных видов
микроорганизмов (С. Н. Виноградский, С. П. Костычев, Е. Н. Мишустин) Ковальский
и Летунова (1959, 1974; Летунова, 1960; Летунова и др., 1968) разработали
методику, построенную на сравнении биологических реакций микроорганизмов
одного и того же вида, но выделенных из биотопов с различными концентрациями
испытуемых элементов. В рамках эколого-геохимического направления изучались
адаптивные реакции микроорганизмов на различные концентрации химических
элементов, изменение закономерностей их роста, влияние микроэлементов на
процессы метаболизма, а также на осуществление микроорганизмами накопительных
функций в биосфере.
Результаты изучения более чем тысячи штаммов бактерий и актиномицетов,
выделенных из иловых отложений с различными концентрациями Со, В, Se, Mo,
Ba, Cu, U, показали, что организмы способны к росту лишь в определенных
пределах концентрации этих элементов. При этом штаммы, взятые из районов
с высоким содержанием химических элементов, сохраняют нормальную жизнеспособность
и рост в более широком диапазоне колебаний их концентраций, чем штаммы
из районов с пониженными их концентрациями. Организмы, способные гибко
реагировать на колебания концентраций химических элементов и одновременно
удовлетворять свои потребности в довольно широких диапазонах их колебаний,
регулируют процессы поступления из среды химических элементов и их накопление
в организмах. Такая регуляция, по мнению Летуновой (1979), является одной
из важнейших адаптации микроорганизмов к окружающей среде. Другой путь
адаптации основывается на способности микроорганизмов превращать растворимые
соединения в нерастворимые.
Были поставлены и эксперименты с целью изменить адаптацию штаммов Act.
clivaceus, Вас. megaterium на обедненных кобальтом илов путем внесения
кобалысодержащих удобрений (Летунова, 1960) (табл. 2). Выделенные через
три месяца формы оказались способны к нормальному росту и синтезу витамина
В12 при значительно больших концентрациях кобальта, чем линии этих же
штаммов, но выделенные до внесения удобрений и культивируемые при обычных
для них
концентрациях кобальта. Данный эксперимент позволяет изучать скорость
86
Таблица 2
Рост иловой микрофлоры при различном содержании хлористого кобальта
в естественной
среде обитания (по: Летунова, 1960)
№ группы
микроорганизмов |
Характеристика
ростовых групп микроорганизмов |
% от общего числа штаммов, выделенных из илов с содержанием
кобальта (% на сухое вещество) |
| 5.0 · 10-4 (Ярославская обл.) |
7.7 ·10-4 (Курская
обл.) |
3.0 · 10-3 (Аз.ССР) |
1 |
Рост возможен без частичного или полного подавления при концентрациях
2-240 мг/л |
20.0 |
31.6 |
88.2 |
2 |
Рост возможен только в пределах концентраций 2-100 мг/л; при 240 мг/л
он отсутствует |
30.0 |
25.4 |
11.8 |
3 |
Рост возможен без частичного или полного подавления лишь в узких
пределах концентраций – 2 – 65 мг/л |
50.0 |
43.0 |
0 |
адаптации организмов к измененным геохимическим условиям. Но при этом
оставалось неизвестным, носила ли адаптация характер длительных модификаций
или же была результатом отбора мутантов. По мнению Ковальского и Летуновой
(1963), во многих случаях адаптации микроорганизмов по своему характеру
являются длительными модификациями, а их носители представляют собой экологические
штаммы. В то же время были выделены штаммы бактерий и актиномицетов, которые
в течение многих месяцев и даже лет сохраняют специфику реакций на избыток
или недостаток химических элементов, т. е. экологические реакции являются
наследственными (Мишустин, 1947). Так, безрезультатно окончились попытки
изменить адаптации штамма 8 Вас. megaterium к высоким содержаниям селена
и штаммов 1100 и 1029 Act. indocolor к высоким содержаниям бора (Летунова
и др., 1968). Все это свидетельствует о том, что способность к оптимальному
росту при определенных концентрациях химических элементов может быть и
наследственно закрепленным признаком. Об этом же говорят и результаты экспериментов
по генетической трансформации некоторых адаптивных реакций микроорганизмов
(Ковальский, Летунова, 1974). Так, признак устойчивости к высоким концентрациям
селена был перенесен от штамма 8 Вас. megaterium с конституционной селенредуктазой
к штамму 10 того же вида, который ранее не встречался с высокими концентрациями
этого элемента. Устойчивость к селену и способность к синтезу селенредуктазы
сохранялась у реципиентного штамма в течение долгого времени культивирования
его в среде без селена.
Была выявлена также внутрипопуляционная изменчивость у бактерий и грибов
по пороговой чувствительности к концентрациям бора, селена, меди, цинка
(Ковальский, Летунова, 1974). Эта изменчивость
87
установлена у организмов, обитающих в одних и тех же геохимических условиях,
что свидетельствует о генетическом характере этого полиморфизма. Естественно,
что подобная гетерогенность способствует селективным процессам в случае
резких изменений геохимических условий. Экспериментальные исследования
в этом направлении были бы весьма полезными в плане изучения взаимодействий
между геохимическими факторами среды, генетической и экологической структурой
популяций и отбором.
Не меньшее значение они имеют и для познания эволюции биосферы. Ковальскому
и Летуновой удалось довести результаты изучения микроэволюционных процессов
у микроорганизмов до уровня биосферных исследований (Ковальский, 1974;
Ковальский, Летунова, 1974). Было показано, что адаптированность микроорганизмов
к геохимической среде предопределяет характер и скорость выполняемых ими
биогеохимических функций. От степени приспособленности зависит скорость
роста биомассы, а следовательно, и объем вовлекаемых в биотический круговорот
химических элементов, а также воздействие организмов на окружающую среду.
Так, у иссыккульского штамма Вас. mesentericus, приспособленного к высоким
концентрациям урана в почвах, происходит максимальный рост биомассы и соответственно
максимальное накопление этого элемента. У штаммов же, не адаптированных
к этому элементу, рост биомассы подавлен. Подобно этому интенсивная азотфиксация,
рост биомассы и накопление молибдена наблюдаются у узбекского штамма Az.
chroococum, приспособленного к высоким концентрациям молибдена. „Гетерогенность
популяций, по мнению авторов, создает благоприятную обстановку для естественного
отбора (особенно в экстремальных условиях) и эволюции организмов в направлении
усиления их биогеохимической роли” (Ковальский, Летунова, 1974, с. 5).
Таким образом, высокоадаптированные организмы играют более важную роль
в биогеохимических процессах биосферы по сравнению с менее адаптированными.
Это доказывает связь эволюционных преобразований, происходящих на популяционно-видовом
и биосферном уровнях организации живого. В силу высокой скорости размножения,
быстрой смены поколений и громадной численности популяций микроорганизмы
являются хорошей моделью для изучения роли геохимических факторов в эволюции
биосферы. Поэтому продолжение и расширение исследований по геохимической
экологии микроорганизмов является чрезвычайно важным и перспективным для
познания биогеохимических закономерностей эволюции биосферы.
В последние десятилетия благодаря эколого-геохимическим исследованиям Г.
Гриффита, В. В. Ковальского, Э. Кэннона, П. Митчелла, Я. В. Пейве стало
ясно, что геохимическая среда является важным экологическим фактором
эволюции растений и животных (Уэбб, 1965). Ежегодно публикуется много работ,
в которых
сообщаются новые данные о влиянии геохимических факторов на жизнедеятельность
и размножение организмов, о возникновении различного рода морфологических
и физиологических отклонений у организмов в случае резких изменений концентраций
того или иного химического элемента. Однако в целом
88
проблема геохимических факторов эволюции до сих пор остается на уровне
самых общих решений. В распоряжении исследователей имеются лишь факты адаптированности
организмов к специфическим условиям среды (например, квасцовая, галмейная,
серпентиновая и т.д. флоры). Для объяснения же механизмов формирования
этих адаптации неизбежно обращение к эволюционной теории.
Причины адаптации растений к геохимической среде обсуждались в работах
Е. А. Беловой, Т. Ф. Боровик-Романовой, В. В. Ковальского, Д. П. Малюги,
Н. С. Петруниной, М. Д. Скарлыгиной-Уфимцевой, М. Я. Школьника и др. Было
показано, что растения сохраняют нормальную жизнеспособность лишь в пределах
определенных колебаний тех или иных химических элементов. Выход за эти
пределы вызывает резкие отклонения от нормального развития и приводит к
биогеохимическим эндемиям (Малюга, 1963; Ковальский, Петрунина, 1965; Петрунина,
1974, и др.). В числе таких отклонений – хлорозы и некрозы, укорачивание
междоузлий, карликовость, недоразвитие генеративных органов, нарушения
плодо- и семяобразования и т.д. Как правило, такие отклонения захватывают
подавляющее большинство растений, что, по мнению Ковальского (19636), обусловлено
их непосредственной связью с геохимическими условиями. Вместе с тем незначительная
часть популяции, оказавшейся в необычных геохимических условиях, сохраняет
нормальную жизнеспособность. Усиленный отбор таких форм, по мнению Ковальского,
приводит к возникновению эндемичных видов. В связи с этим он высказал предположение,
что в районах с резкими колебаниями геохимических условий происходит интенсивное
видообразование. Как доказательство этого он рассматривал приуроченность
центров происхождения культурных растений, выделенных Н. И. Вавиловым (1926),
к горным районам, где геохимическая среда наиболее разнообразна. Ковальский
и Петрунина (1965) предложили схему, в которой различные реакции растений
на необычные концентрации химических элементов представлены как определенные
стадии видообразования. Среди адаптивных форм растений они выделяли три
группы: 1) растения, не концентрирующие химические элементы; 2) непривычные
концентраторы; 3) привычные концентраторы. Для изменения биогеохимических
процессов в биосфере особое значение имеют привычные концентраторы, содержащие
элементы в значительно больших количествах, чем их среднее содержание в
почвах ареала. Классическим примером мощных концентраторов хлора является
ряска (Вернадский, Виноградов, 1931), молибдена – все представители семейства
бобовых (Ковальский, Петрунина, 1965), лития – семейства лютиковых, магнолиевых,
пасленовых (Боровик-Романова, Белова, 1974), алюминия – плауны (Виноградов,
1933),и т. д.
Изучение популяций Rumex acetosa, Pyrethrum parthenifolium, Scrophularia
variegata, S. grossheimii, Hypericum perforatum в молибденовых провинциях,
Alhagi kirghisorum, Cynodon dactylon, Glycyrrhiza glabra в кальциево-стронциевых
провинциях, а также ряда других растений, произрастающих на почвах с
повышенным содержанием никеля, меди, кобальта, показало, что даже в популяциях
растений
— концентраторов
89
химических элементов существует значительная физиологическая изменчивость
по этому признаку. Среди них встречаются как привычные, так и непривычные
концентраторы. Для каждого вида характерен свой размах изменчивости, т.е.
соотношение привычных и непривычных концентраторов, а также форм с пониженной
или повышенной способностью к концентрации химических элементов. Так, в
популяциях тимьяна кочи (Thymus kotschyanus) и ромашки девичьей (Pyrethrum
parthenifolium), произрастающих в молибденовых провинциях, установлено,
что в первой популяции 58 % особей содержали молибдена больше, чем его
было в почвах, 33 % особей — меньше, чем в почвах, и у 9 % установилось
такое же содержание молибдена, как и в почвах. Во второй популяции соответствующие
цифры равнялись 31, 61 и 8 % (Ковальский, 1982).
Все это позволяет рассматривать внутрипопуляционную изменчивость в качестве
исходного материала для действия отбора в сторону формирования видов —
мощных концентраторов. Экстремальные геохимические условия среды способствуют
выявлению внутрипопуляционного полиморфизма по устойчивости к различным
концентрациям химических элементов и резко обостряют процессы борьбы за
существование и естественного отбора. Очевидно, что возникновение мощных
концентраторов так или иначе должно отразиться на процессах биогенной миграции
химических элементов. Примером группы, где интенсивно идет процесс формирования
концентраторов стронция, согласно Ковальскому, может служить верблюжья
колючка (Alhagi kirghisorum). В ее популяциях, произрастающих в стронциевых
биогеохимических провинциях Таджикистана, положительным градиентом концентрации
стронция обладают 97.6 % особей, а отрицательным — 2.4 %.
Однако до сих пор нет прямых данных о том, как возникают виды — мощные
концентраторы и как их возникновение сказывается на биогеохимических процессах
биосферы. В экспериментальном отношении этот вопрос практически не изучен.
Для получения таких данных перспективны исследования микроэволюционных
преобразований у растений в районах горных разработок, где почвы содержат
резко повышенные концентрации химических элементов. Такие исследования
в последние годы стали проводиться как у нас в стране, так и за рубежом.
Из-за интенсивного развития горнодобывающих предприятий существуют громадные
территории, где резко повышено содержание тяжелых металлов (свинец, цинк,
медь), способных в короткий срок остановить рост растений и вызвать их
гибель. Важно, что в этих местах складываются свободные ниши с экстремальными
геохимическими условиями. Вместе с тем нельзя сказать, что на этих территориях
вообще нет растений. Иногда там встречаются виды, которые обычно произрастают
только в районах с нормальным геохимическим фоном. Еще в 1934 г. С. Прат
(Prat, 1934) впервые показал на примере Silene vulgaris, что популяции,
растущие на территории с повышенным содержанием тяжелых металлов, генетически
отличаются от популяций того же вида, но произрастающих на нормальных
почвах. Они толерантны к таким концентрациям металлов, которые для обычных
популяций
являются токсичными. Теперь
90
благодаря многочисленным исследованиям установлено, что это справедливо
для всех видов растений, встречающихся в индустриальных пустынях”. А. Брэдшоу
(Bradshaw, 1971, 1984) доказал, что Agrostis tenuis, Antoxanthum odoratum,
Plantago lanceolata, Rumex acetosella образуют микрогеографические расы,
произрастающие в районах бывших шахт, где почва содержит соли меди, свинца
и цинка в дозах, летальных для других рас тех же самых видов. Устойчивость
оказалась наследственно закрепленной. Она не теряется при выращивании растений
в обычных условиях. Устойчивость чаще всего специфична к отдельным металлам
и обусловлена несколькими генами.
Были получены и доказательства быстрого селективного ответа нормальной
популяции на экстремальные условия. Так, из 1000 особей Agrostis tenuis
выжили только три растения, которые и положили начало новой расе. Это показывает,
что адаптация к необычным геохимическим факторам может идти под воздействием
катастрофического отбора в течение одного – двух поколений.
Определенную роль в адаптации к геохимическим факторам может играть миграция
особей из толерантных популяций, если ареалы обычных и толерантных популяций
близко расположены. Как показали исследования Брэдшоу, между толерантными
и нетолерантными расами нет строгой изоляции, между ними сохраняется определенный
поток генов. В результате взрослые растения микрогеографических рас были
хорошо устойчивы к высоким концентрациям солей, но некоторые из их семян
давали менее жизнеспособные формы с резкими морфологическими и физиологическими
отклонениями. Таким образом, помимо фено- и генотипической изменчивости
и отбора изоляция является важнейшим фактором адаптации растений к геохимической
среде.
Толерантные расы растений в районах бывших горных разработок чаще всего
отличаются замедленным ростом. Однако благодаря им в биогенные круговороты
включаются все же значительные количества тяжелых металлов, которые в противном
случае оставались бы в почвах. Таким образом, формирование популяций, устойчивых
к высоким концентрациям тяжелых металлов, способствует усложнению биотического
круговорота веществ. Этот процесс, протекающий у нас на глазах, позволяет
экспериментально изучать связь процессов микроэволюции с эволюцией биосферы.
В отличие от микроорганизмов и растений животные более независимы от геохимических
условий среды, так как последние действуют на них не прямо, а опосредованно,
преломляясь через сложную цепь трофических отношений. Кроме того, у животных
имеются сложные механизмы регуляции взаимодействий организмов с окружающей
средой. Среди них системы, обеспечивающие депонирующую, барьерную, выделительную
функции, распределение микроэлементов между тканями и органами. И хотя
резкие отклонения в концентрациях химических элементов вызывают срыв
регулирующих систем, однако вредящему воздействию геохимических факторов
подвергается,
как правило, незначительная часть популяции. Такое соотношение нормальных
и с пониженной жизнеспособностью организмов является, по-видимому, результатом
91
длительного отбора на выработку защитных механизмов от вредящего воздействия
геохимической среды. Это отмечалось в ряде работ В.В.Ковальского, М. А.
Риша, С. С. Шварца по геохимической экологии животных. Тем не менее в работе
Шварца (1954) было показано, что повышенное содержание никеля в тех водоемах,
где проходит метаморфоз зеленая жаба, ускоряет онтогенез ее особей и резко
тормозит их рост. У зеленых же лягушек в этих условиях происходят столь
значительные повреждения внутренних органов (кишечника, почек, легких),
что их пребывание в таких водоемах становится невозможным. В то же время
у многих наземных позвоночных повышенное содержание никеля в почвах сказывается
лишь косвенно, через пищевые цепи. Здесь отклонений от нормального развития
значительно меньше. В частности, таких отклонений практически нет у насекомоядных
птиц. Эти исследования Шварца демонстрируют, сколь сложно воздействие геохимических
факторов.
В связи с этим большой интерес представляют работы по изучению зависимости
химического состава 17 видов амфибий и 9 видов рептилий от геохимических
условий среды (Шарыгин, 1984; Шарыгин, Коржневский, 1982). Генетически
закрепленная способность организмов извлекать и накапливать в различных
тканях и органах металлы (алюминий, марганец, железо, хром и т. д.) формируется
в зависимости от геохимических условий среды, биотопа и характера пищевых
цепей. Существуют и достоверные различия по элементному составу близких
подвидов. Например, у четырех подвидов скальной ящерицы (Lacerta saxicola)
различия по некоторым элементам больше видовых и даже родовых. Крымский
подвид (L. saxicola lindcholmi) и нормальный подвид (L. saxicola saxicola)
различаются по 11 элементам, крымский и западно-северокавказский (L. saxicola
darevskii) — по 9 элементам. Сопоставление химического состава двух внутривидовых
форм остромордой лягушки (Rana arvalis) — полосатой и пятнистой — показало,
что первая форма содержит марганца в 3.5, хрома — в 5, никеля — в 4, цинка
— в 6, стронция — в 2 раза меньше, чем вторая. Железа, однако, в 2 раза
меньше у последней формы. По мнению Шарыгина, повышенное содержание микроэлементов
у пятнистой формы обеспечивает ее лучшую приспособленность к горным условиям.
Дифференциальная выживаемость двух форм в разных ареалах объясняется автором
сложным взаимодействием геохимических факторов внешней среды и особенностью
химического состава каждого из подвидов.
Работы по изучению воздействия геохимических факторов на популяции позвоночных
(Криволуцкий, 1983; Покаржевский, 1985) в целом показали, что, как и у
растений, основой формирования адаптации организмов к геохимической среде
выступает генетическая гетерогенность популяций животных по их пороговой
чувствительности к концентрациям химических элементов. Это свидетельствует
о том, что геохимические процессы, интегрирующие биосферу в единую систему,
непосредственно вплетаются и в ход микроэволюционных преобразований.
Сравнивая итоги изучения геохимических факторов эволюции животных с результатами
эволюционных исследований по геохимической экологии микроорганизмов и
растений, мы должны отметить,
92
что в работах с животными главное внимание уделялось биохимическим и физиологическим
механизмам адаптации организмов к повышенным содержаниям тех или иных химических
элементов, а эволюционные аспекты этой проблемы практически не исследовались,
хотя в некоторых зарубежных работах был показан наследственный характер
различий (Д. Фолкнер, Г. Вине, Ф. Люкс и др.). Остается неясным значение
этих исследований для разработки проблемы эволюции биосферы. В целом можно
сделать вывод, что исследования по геохимической экологии микроорганизмов
наиболее близко подошли к реализации поставленной Вернадским задачи изучения
взаимосвязей процессов микроэволюции и видообразования с эволюционными
преобразованиями биосферы. Было показано, что степень адаптации организмов
к геохимической среде непосредственно сказывается на процессах миграции
химических элементов, на биологической продуктивности и общей биомассе
экосистем, на количестве аккумулированной в ее биокосных компонентах энергии.
Следует согласиться с Ковальским, что „без установления причинной зависимости
между развитием живых организмов и формированием живого вещества, с одной
стороны, и геохимическими условиями — с другой, не может быть понята эволюция
биосферы” (1972, с. 515). В заключение несколько слов о взаимоотношении
между геохимической экологией и эволюционной теорией.
В своих последних работах С. С. Шварц не раз подчеркивал, что изучение
воздействий микро- и макроэволюции на исторические преобразования биосферы,
на ее биогеохимическую и энергетическую эволюции является важнейшей задачей
современной эволюционной теории. Правда, он вынужден был признать, что
еще нет возможности решить проблему взаимодействия эволюции организмов
и эволюции биосферы. „Время для этого еще не пришло, — писал С. С. Шварц,
— но оно придет завтра. Целесообразно поэтому выделить те вопросы, решение
которых будет способствовать решению проблемы в целом” (1973, с. 213).
Вместе с тем он считал установленным тот факт, что адаптивное преобразование
видов в геохимическом и энергетическом отношении имеет не локальное, а
общепланетарное значение.
Изучение геохимических факторов эволюции организмов является одним из важнейших
путей для решения вопросов и об эволюции биосферы. В то же время это один
из наиболее перспективных путей синтеза биогеохимических представлений
с эволюционной теорией.
При обсуждении вопроса о геохимических факторах микроэволюции необходимо
учитывать также, что „отдельные организмы не только приспособлены к физической
среде, но и своим совместным действием в рамках экосистемы приспосабливают
геохимическую среду к своим биологическим потребностям” (Одум, 1975,
с. 34). Благодаря жизнедеятельности организмов идет непрерывное изменение
окружающей среды, в нее поступают все новые вещества, источники энергии,
изменяются концентрации и распределения отдельных элементов. Таким образом,
при анализе воздействия геохимических факторов на эволюцию живого вещества
нельзя забывать, что многие из этих факторов порождены самой жизнью.
93
В ходе эволюции сложилось единство геохимической среды и жизни, выражающееся
в постоянстве зависимости жизни от геохимических условий, водного режима,
температуры, инсоляции и т.д. В то же время многие из этих условий — результат
предшествующей эволюции органического мира. В ходе эволюции шло усложнение
взаимосвязей организмов с геохимическими факторами. Влияние геохимических
факторов становилось все более косвенным и опосредованным.
* * *
Проанализированные направления отечественной эволюционно-биологической
школы немало сделали для реализации теоретических установок Вернадского
о взаимодействиях между процессами видообразования и эволюцией биосферы.
Было показано, что эти процессы находятся в сложных каузальных отношениях,
где причины и следствия постоянно меняются местами. Эволюция видов, контролируемая
действующими в биосфере биогеохимическими принципами, в свою очередь является
ведущим фактором ее эволюции. Изменение химического состава и геохимической
энергии видов — это элементарное явление в эволюции биосферы. От степени
адаптированности организмов к среде зависит характер и интенсивность выполняемых
ими биогеохимических функций и их участие в биотическом круговороте.
Сформулированные советскими учеными положения о зависимости распространения
химических элементов в живом веществе биосферы от их места в таблице Менделеева,
о геохимических факторах географической изменчивости организмов, о возрастании
автономности организмов от геохимических факторов внешней среды в ходе
прогрессивного развития жизни, о внутривидовой дифференциации организмов
по приспособленности к экстремальным геохимическим факторам среды и по
их участию в концентрации химических элементов имеют большое значение для
понимания факторов, движущих сил и закономерностей эволюции видов. Принципы
современной эволюционной теории исключительно важны в качестве исследовательских
установок для изучения самыми различными методами эволюционных преобразований
на уровне биосферы. Следует умножить усилия биологов-эволюционистов по
интеграции данных и обобщений, полученных в ходе биогеохимических исследований,
с основными положениями современной эволюционной теории. Такая работа будет
полезна и для изучения факторов, движущих сил и закономерностей эволюции
самой биосферы.
94
Глава 4
ЭВОЛЮЦИЯ БИОГЕОХИМИЧЕСКИХ КОНЦЕНТРАЦИОННЫХ
ФУНКЦИЙ БИОСФЕРЫ
Само понятие „биогеохимическая функция биосферы” было сформулировано Вернадским
в работах 1928 – 1931 годов. Под биогеохимической функцией он понимал способность
организмов вызывать движение вещества в биосфере (Вернадский, 1980, с.
38). Этой способностью и обусловлены те грандиозные изменения в химическом
составе атмосферы, гидросферы, литосферы и педосферы, которые являются
наиболее яркими и очевидными показателями исторических преобразований геохимического
строения биосферы. Среди разнообразных биогеохимических функций биосферы
(газовой, кислородной, окислительно-восстановительной, деструктивной и
т. д.), Вернадский особенно выделял концентрационные функции. Суть этих
функций заключается в способности организмов аккумулировать химические
элементы в количествах, превышающих среднее их содержание в окружающей
среде в десятки, сотни и тысячи раз.
Представления Вернадского о концентрационных функциях биосферы позволили
по-новому взглянуть на известные еще с середины прошлого века факты о таксономических
различиях в химическом составе организмов (Rochleder, 1854; Bezold, 1857).
Так, появилась проблема корреляции между элементами химического состава
организмов и осуществляемыми ими биогеохимическими концентрационными функциями,
а следовательно, и вопрос о взаимосвязи их эволюционных преобразований.
Можно выделить три этапа в разработке проблемы эволюции концентрационных
функций биосферы в нашей стране.
Первый этап связан с палеобиогеохимическими работами Я. В. Самойлова, опубликованными
в период с 1906 по 1923 г. Самойлов был инициатором систематических исследований
минерального и химического состава скелетов и тканей современных и ископаемых
организмов с целью реконструкции биогеохимических особенностей прошлых
биосфер. При этом изучение химических особенностей органогенных пород,
образовавшихся в тот или иной период геологического времени, рассматривалось
как главный источник знаний для реконструкции особенностей былых биосфер.
Второй период (с середины 20-х до конца 40-х годов) проходил под определяющим
воздействием идей Вернадского о биогеохимических концентрационных функциях
биосферы. В эти годы под руководством В. И. Вернадского и А. П. Виноградова
шли интенсивные исследования
95
геохимических констант видов и более крупных таксономических единиц. Исследования
позволяли оценить участие того или иного таксона в осуществлении биогеохимических
функций биосферы, а также проследить изменения этих функций в связи с
преобразованием таксономического состава органического мира. Итоги изучения
закономерностей
изменения концентрационных функций биосферы были подведены в книге А.
П. Виноградова „Химический элементарный состав организмов моря” (1935а,
1937,
1944) и в посмертно изданной книге В. И. Вернадского „Химическое строение
биосферы Земли и ее окружения” (1965).
Третий период (с 50-х годов до настоящего времени) характеризуется первыми
попытками количественно оценить эволюцию концентрационных функций не только
в связи с филогенезом отдельных таксономических групп, но и в связи с эволюцией
всей биосферы в целом. Работы в этом направлении вела Е. А.Бойченко с сотрудниками
и учениками. Здесь изучалась взаимосвязь эволюционных преобразований окислительно-восстановительных
и концентрационных функций биосферы и сопоставлялись полученные данные
со сведениями о мировых запасах полезных ископаемых.
Мы остановимся главным образом на работах Я. В. Самойлова, В. И. Вернадского,
А. П. Виноградова и Е. А. Бойченко, в которых были не только сформулированы
основные задачи названных направлений исследований, но и сделаны обобщения,
имеющие значение для понимания закономерностей в эволюции биосферы.
4.1. Роль палеобиогеохимических исследований
в изучении эволюции концентрационных функций биосферы
Общепризнано значение Я. В. Самойлова в создании и развитии палеобиогеохимических
исследований (Архангельский, 1929; Давиташвили, 1940: Вольфкович, 1974;
Гордеев, 19746; Дроздова, Соколов, 1979; Мирзоян, 1984, и др.). Оценивая
его вклад в разработку проблем палеобиогеохимии, Вернадский писал в 1940
г., что Самойлов первым пошел по пути изучения геологической роли органического
мира и выдвинул в связи с этим ряд новых проблем, которые были „поставлены
и сведены к мере и числу”. Вместе с Вернадским Самойлов по существу заложил
основы учения о биолитах.
Уже в 1917 г. Самойлов доказал необходимость создания новой науки „палеофизиологии”,
т. е. палеобиогеохимии. В ее задачу должно было войти изучение особенностей
биогеохимической деятельности организмов в прошедшие геологические эпохи.
К идее о создании такой науки Самойлов пришел в ходе исследований месторождений
барита, целестина, фосфоритов, кремнистых пород, железо-марганцевых и
медных руд, которые, по его мнению, могли образоваться только при активном
участии
организмов. Вопреки доминирующим тогда представлениям об образовании
этих минералов и горных пород абиогенным путем Самойлов считал, что их
месторождения
созданы в результате
96
жизнедеятельности организмов. Генезис морских осадочных отложений, по
его мнению, можно понять лишь при учете биохимических и физиологических
особенностей ископаемых организмов, которые были способны концентрировать
их в больших количествах.
Самойлов развивал биогеохимические представления не на уровне биосферы,
а применительно к генезису минералов, горных пород и месторождений полезных
ископаемых. Для объяснения их происхождения он и начал изучать роль „организмов
в геохимических и геологических процессах земной коры” (Вернадский, Ферсман,
1929, с. 3). Но объективно значение его работ оказалось гораздо шире. В
них содержится ряд обобщений, имеющих прямое отношение к анализируемой
нами проблеме.
Остановимся кратко на идеях Самойлова о концентрационных функциях биосферы
и их исторических преобразованиях.
К проблеме участия организмов в образовании минералов и горных пород Самойлов
обратился еще в 1906 г. в своей докторской диссертации.1 Ссылаясь
на работы С. Н. Виноградского (1888), Н. И. Андрусова (1897) и Г. А.
Надсона (1903),
он полагал, что „путем исследования свойств минералов можно определять
не только все физико-химические свойства той среды, в которой происходило
осадкообразование, но в известных пределах и живые организмы фауны и
флоры среды” (1906, с. 12—13), Согласно Самойлову, концентрация в осадочных
породах
редких металлов скорее всего обусловлена жизнедеятельностью морских организмов,
способных накапливать эти металлы. Эта идея получила дальнейшее обоснование
и развитие в последующих публикациях Самойлова.2
Самойлов полагал, что для понимания концентрационных функций биосферы необходимо
знать точно химический состав организмов, и прежде всего их скелетных образований.
К сожалению, систематические исследования этих признаков еще только начинались.
Это побудило Самойлова „не просто использовать для решения интересовавших
его вопросов накопленный другими материал, но и поставить самому работу
в этом направлении, перекинув, таким образом, мост от минералогии к физиологии”
(Архангельский, 1929, с. 8). Он старался выявить механизм превращения остатков
организмов в минеральные и горные породы.
В 1910 г. Самойлов обратил внимание на то, что крупные запасы барита, как
правило, приурочены к ярусам верхней юры. Это навело его на мысль, что
в отдельных регионах биосферы того периода существовали организмы, содержащие
в своих раковинах барий и способствовавшие тем самым аккумуляции стойких
соединений бария (его сульфатов)
1 В эти же годы проблема геологического значения живого вещества
привлекает все большее внимание Вернадского. В письме к Самойлову от 9
июля 1908
г. он писал: „Много в последние годы обдумано в связи с вопросом о количестве
живого вещества, о чем я говорил Вам раньше” (цит. по: Тр. Биогеохим.
лабор.
1980. Т. 16. С. 6).
2 Мы цитируем статьи Самойлова по посмертному сборнику его
биогеохимических работ „Биолиты» (1929).
97
в виде конкреций. В случае справедливости этого предположения изучение
месторождений барита могло привести к открытию специфической ископаемой
группы, содержащей барий в значительных количествах.
К сожалению, в литературе того времени имелись только отдельные указания
на наличие бария в организмах (Р. Квентон, Ф. Кларк и Дж. Меррей). Такие
данные не могли быть доказательством того, что образование месторождений
барита связано с доминировавшей в верхней юре группой организмов с высоким
содержанием бария. Лишь данные О. Бютчли о сульфате стронция (т. е. металла
той же группы) в скелетах радиолярий Acantharia в какой-то степени могли
служить косвенным подтверждением гипотезы Самойлова. Суть гипотезы заключалась
в том, что „связь между отложениями некоторых минеральных тел и определенными
группами растительного или животного мира окажется, быть может, много теснее,
чем это представляется в настоящее время” (Самойлов, 1929, с. 34).
Но в том же 1910 г. в книге А. Щепотьева „Исследования над низшими организмами”
гипотеза Самойлова получила прямое подтверждение. В теле корненожки Xenophyophora
были найдены образования сернокислого бария (BaSO4). Это явно свидетельствовало
о возможности биогеохимического происхождения барита.
К аналогичным выводам пришел Самойлов в 1912 г. при изучении генезиса месторождений
целестита (сульфат стронция), приуроченных к верхнемеловым и нижнетретичным
отложениям. Сопоставляя сведения об этих месторождениях с данными О. Бютчли
о нахождении сульфата стронция у радиолярий, Самойлов предположил, что
„существованию таких стронцийсодержащих организмов обязаны меловые отложения
своим относительным богатством рассматриваемым (щелочноземельным) металлом”
(там же, с. 49).
В 1912 г. Самойлов четко формулирует гипотезу, согласно которой крупные
месторождения металлов, содержащихся в морской воде в ничтожных количествах,
могли создаваться только в результате осуществления концентрационных функций.
Он считал неправильным при изучении ископаемых ограничиваться лишь морфологическими
признаками. Необходимо выявлять и особенности физиологических процессов,
а следовательно, и эволюцию химических соединений, участвующих в этих процессах.
Он ставил вопрос о том, всегда ли в биосфере существовали те же соотношения
между животными, содержащими в крови гемоглобин или гемоцианин, как и в
современной биосфере. Если в прошлом эти соотношения были иными, то не
в этом ли кроются причины образования мощных месторождений меди в пермских
отложениях? Подобные месторождения могли быть созданы доминирующими в перми
организмами, у которых функцию переноса кислорода в крови выполняли молекулы
гемоцианина, содержащего медь вместо железа. И наличие ванадия в крови
у асцидий Самойлов также использовал для объяснения повышенных содержаний
этого элемента в ряде осадочных пород.
Опираясь на эти данные и предположения, Самойлов уже в 1912 г. формулирует
вывод, важный для изучения биогеохимических функций палеобиосфер. Он
писал: „Если мы примем, что различные организмы
98
с указанными характерными своеобразными особенностями, являющиеся в настоящее
время относительно более редкими, в отдаленные периоды истории Земли количественно
преобладали, представляли собой обычные, распространенные группы животных,
тогда будет ясно, какое значение может иметь детальное изучение всех этих
соотношений в деле правильного объяснения генезиса различных минералов,
заключенных в осадочных породах” (там же, с. 51).
В последующие годы Самойлов публикует ряд статей, в которых далее углубляет
этот вывод и подкрепляет его новыми доказательствами. Так, широкое распространение
плавикового шпата в известняках московского яруса карбона он объяснял наличием
в это время фторсодержащих организмов, подобных некоторым современным видам
плеченогих, пресноводных и морских моллюсков, у которых К. Андрее, И. Андерсон,
А. Куффер, А. Мюнтц, А. Хатин и др. установили наличие фтора в раковинах.
Приуроченность фтора к фосфоритам метасоматического типа, по мнению Самойлова,
могла быть вызвана сильным развитием в соответствующий геологический период
животных с повышенным содержанием фтора, как это установлено у роговых
губок (Aplysinedae, Spongidae) и кораллов (Gorgonia carolinii). Высокое
содержание в известняках сульфата кальция связано с распространением тех
групп беспозвоночных, у которых Ф. Кларк установил следующее процентное
содержание CaSO4: у брахиопод – Lingula anatina – 4.18% и Discinisea lamellosa
– 8.35%; у кораллов (альционарии) – Gorgonia acerosa – 4.08%; у Paramuricea
borealis – 4.69 и Gorgonia suffruticosa – 5.43%; у иглокожих – Mellita
sexiesperferatus – 2.53 и Ohiomyxa flaccida – 4.17%. Среди брахиопод, сходных
по морфологическим признакам, существуют две группы, резко отличающиеся
по химическому составу. У замковых брахиопод скелет составлен из СаСО3
(карбонатный), а у беззамковых – из Са3(РO4)2 (фосфатный). Значительные
колебания наблюдаются в содержании карбоната магния (MgCO3) у брахиопод
– (от 1 до 8.63%, у альционарии – от 6.18 до 15.73%, у иглокожих – от 11.56
до 14.95 %. Иногда в пределах одного семейства или даже рода животные образовывали
раковины совершенно разного химического состава. Все это побуждало искать
связь преобразований морфологических и биохимических признаков и выяснять
адаптивное значение различий в химическом составе скелетов близкородственных
организмов.
В целом было ясно, что „если бы качественно и количественно химический
состав скелетов был бы во все геологические времена одинаков, то и первоначальный
химический состав осадочных пород — биолитов, откладывающихся в одинаковых
физико-географических условиях в течение всей истории Земли, был бы неизменным”
(там же, с. 128). В первоначальном составе биолитов неизбежно отражалось
количественное соотношение организмов со скелетами различного химического
состава.
Вместе с тем нельзя было забывать и о тех неизбежных изменениях, которые
претерпевал первоначальный химический состав ископаемых остатков в процессах
диагенеза. Поэтому особенно были важны комплексные исследования биологов
и геологов. При переходе элементов из биосферы в литосферу происходят
химические преобразования
99
ископаемых остатков. Здесь возможен вторичный процесс концентрации таких
редких элементов, как бериллий, ванадий, ртуть (Гольдшмидт, 1938; Султанов,
Исаев, 1967). Подобные вторичные концентрации элементов естественно затрудняют
познание биогеохимических функций прошлых биосфер.
Подлинной программой исследования эволюции биогеохимических функций биосферы
стала классическая статья Самойлова „Биолиты”, опубликованная в 1921 г.
В этой статье систематизированы все ранее приводимые данные и гипотезы
об изменении концентрационных функций биосферы в связи с эволюцией химического
состава организмов. Для изучения этого процесса было необходимо: во-первых,
искать ископаемые организмы со специфическим химическим составом и специфическими
биогеохимическими функциями, не встречающиеся в современной биосфере; во-вторых,
выяснить количественные соотношения в прошлых биосферах групп организмов
с различным химическим составом и изучать изменения этих соотношений в
ходе органической эволюции.
Самойлов обосновал важную мысль о том, что при отсутствии фоссилизированных
остатков изучение химического состава биолитов и закономерностей их образования
является надежным способом для реконструкции геохимических функций вымерших
организмов. Различия в химическом составе биолитов, образовавшиеся в разные
геологические эпохи, служат показателями „количественных соотношений организмов
со скелетами различного химического состава” (там же, с. 66). В этой статье
Самойлов вновь ссылается на ранее высказанные предположения о биогенном
происхождении целестина, барита, фосфоритов, кремнистых пород.
Годом позже в статье, опубликованной на немецком языке, Самойлов приводит
данные исследований железо-марганцевых конкреций со дна Черного, Балтийского
и Баренцевого морей. Образования этих пород, по мнению Самойлова, могли
происходить в результате деятельности бактерий. Разбирая работы Р. Ланге,
И. Помпеску и других авторов о содержании меди в германском медистом сланце,
Самойлов вновь возвращается к гипотезе о доминирующем положении в морских
сообществах перми организмов (например, моллюсков), у которых функцию переноса
кислорода выполняют молекулы гемоцианина.
В своих последних выступлениях и статьях Самойлов старался далее развить
высказанные им теоретические положения и наметить пути их проверки. В докладе,
прочитанном 19 февраля 1922 г. на торжественном заседании в честь 75-летия
со дня рождения А. П. Карпинского, Самойлов остановился на вопросе об эволюции
минерального состава скелетов.
Кремниевая функция, по мнению Самойлова, более древняя, чем кальциевая.
Организмы, обладающие кремниевым скелетом, не продвинулись дальше губок
среди животных и диатомовых водорослей среди растений. „На древе развития
организмов, — писал Самойлов, — древнейшая кремниевая группа представляет
отдельную самостоятельную ветвь, скоро остановившуюся в своем дальнейшем
развитии” (там же, с. 98). У более сложных организмов ведущая роль в
образовании скелетов
100
принадлежит кальцию. Но и здесь есть определенная закономерность в эволюции
скелета. Если у беспозвоночных он преимущественно сложен из углекислого
кальция, то у позвоночных в скелете доминирует фосфат кальция и лишь отдельные
части скелета формируются из карбонатов.
Самойлов попытался объяснить причины более раннего появления организмов
с сильно развитой кремниевой функцией, положив начало тем самым идущим
и по сей день спорам о причинах внезапного усиления кальциевой функции
биосферы на границе протерозоя и палеозоя.1 Самойлов высказал
гипотезу, не получившую, правда, признания. Согласно этой гипотезе, подобная
последовательность
в эволюции кремниевых и кальциевых скелетов объясняется химическими свойствами
Si и Ca и их распространением в земной коре. Преимущественное использование
вначале кремния связано с тем, что этот элемент занимает второе место
после кислорода по распространенности в биосфере. Кальций — только пятое.
Натрий
и калий, находящиеся между кремнием и кальцием в этом ряду,2 образуют
растворимые соли и поэтому не могли быть промежуточным звеном при переходе
от кремниевых
скелетов к кальциевым. Таким образом, по мнению Самойлова, жизнь вначале
использовала более доступный элемент, а затем перешла к утилизации элемента,
способного образовывать более эффективные в структурном и функциональном
отношениях скелеты. Аналогичную закономерность он усматривал и в эволюции
кальциевого скелета. Здесь древнейшие группы использовали более распространенный
элемент углерод, а более прогрессивные животные — менее распространенный
фосфор.
Опираясь на эти данные и соображения, Самойлов допускал даже возможность
существования древней жизни на кремниевой основе. И хотя гипотеза о „высокоустойчивой
к температурным колебаниям протоплазме”, содержащей вместо углерода кремний,
не раз излагалась в литературе как до Самойлова (Д. Рейндольс, Н. Н. Морозов),
так и после него (А. Азимов, М. Г. Воронков, В. Фирсов), нет никаких геологических
данных о том, что в земных условиях могли существовать организмы, в основе
которых лежали кремнийсодержащие высокомолекулярные соединения. Весьма
убедительны и соображения Дж. Уолда (1964) о том, что кремний в силу ряда
своих химических свойств в принципе не может быть носителем жизни.
Вместе с тем подмеченный Самойловым факт более мощного развития кремниевой
функции в древних биосферах не раз впоследствии использовался в работах,
в которых рассматривались вопросы о геохимической эволюции поверхностных
оболочек Земли (Вильямс, 1926; Виноградов, 1932, 1944, и др.). Современные
данные в целом подтверждают вывод Самойлова о том, что в ходе прогрессивной
эволюции доля кремния в составе организмов уменьшается. Если у одноклеточных
водорослей (диатомовые и силикофлагелляты) кремний по массе среди других
элементов занимает третье место, то уже в папоротниках,
1 В дальнейшем мы подробнее остановимся на этих дискуссиях.
2 По современным данным, на 3-м и 4-м местах по доле в литосфере
находятся алюминий и железо (Наука и Вселенная. 1983. С. 130).
101
содержащих кремния больше всего среди высших растений, он перемещается
на седьмое место, и, наконец, у покрытосеменных оказывается на четырнадцатом
месте. Если в планктоне соотношение углерода и кремния составляют 1:1,
в папоротниках – 1:100, то у млекопитающих – 1:5000.
Были предложены и другие объяснения столь широкого распространения кремниевых
организмов в более древних биосферах. Так, М. Г. Воронков и И. Г. Кузнецов
(1984) считают, что отложившиеся в наружных частях организма соединения
кремния могли выполнять роль теплоизолирующего механизма, защищающего
бактерии и примитивные животные и растения от переохлаждения или перегрева
в условиях
резких колебаний температур в первичной биосфере. Ссылаясь на исследования
В. Г. Александрова над силикатными бактериями, они отмечают их удивительную
способность сохраняться при температуре –196 °С и +160 °С. Такая устойчивость
к экстремальным температурам, вероятно, позволяла первичным обитателям
Земли осваивать ту среду, в которой фактически еще не было органических
источников питания и озонового экрана, защищающего жизнь от губительного
воздействия космического излучения. Вполне возможно, что именно организмы,
способные разрушать силикаты и алюминосиликаты, переработали силикатный
покров Земли в почву и тем самым создали условия для организмов, у которых
сильное развитие получила кальциевая и фосфорная функции.
В свете современных знаний о трофических цепях интересна гипотеза Самойлова
о том, что химический элементный состав животных в значительной степени
предопределяется потребляемыми ими растениями. Растения являются в этом
отношении промежуточным звеном между животными и внешней средой. Осуществление
концентрационных функций биосферы распадается на два этапа. Вначале рассеянные
в биосфере элементы поглощаются растениями из почвы и водных растворов,
а затем уже аккумулируются в теле животных. Эта гипотеза Самойлова вполне
согласуется с представлениями о повышении концентрации металлов при переходе
от низших звеньев в трофических цепях к высшим. Учитывая двухступенчатый
характер аккумуляции организмами химических элементов, становится понятным
тот факт, что у гетеротрофов концентрация таких элементов, как натрий,
кальций, фосфор, хлор, цинк и т. д., значительно выше, чем у автотрофов.
Вполне возможно, что усложнение трофической структуры биосферы способствовало
усилению концентрационных функций.
Предложенный Самойловым палеобиогеохимический подход к изучению концентрационных
функций биосферы открывал и новый путь в познании эволюции жизни. Если
до сих пор ископаемые организмы реконструировались лишь по морфологическим
признакам, то отныне изучение химических признаков современных организмов
давало возможность познать „прошедшую жизнь через изучение химического
состава ныне живущих организмов” (Виноградов, 1932, с. 6). Исследование
химического состава биолитов позволяло в принципе открывать группы организмов,
которые в силу различных причин не оставили никаких следов в палеонтологической
летописи. В связи с этим особое
102
значение приобретает совместное изучение морфологической и химической
эволюции, построенное на комплексном применении актуалистических и историко-сравнительных
методов. На их основе вырабатывалась более полная характеристика той жизни,
которая отвечает каждой геологической эпохе.
Имеющиеся тогда данные заставляли ограничиваться только самой постановкой
вопроса. Суть поднятой Самойловым проблемы заключалась в том, что не только
известняки и горючие ископаемые, но и другие осадочные породы образуются
на границе литосферы и биосферы при непосредственном, а иногда и решающем
участии организмов и продуктов их жизнедеятельности. Многие из этих организмов
сейчас или исчезли с лица Земли, или занимают в экономике биосферы незначительное
место.
Но выяснение конкретного механизма образования органогенных минералов оставалось
задачей будущего. На долю Самойлова выпала миссия лишь обосновать значение
палеобиогеохимии для познания биосфер прошлого. Он не успел полностью обработать
даже собранный им и его сотрудниками материал и завершить начатые в этом
направлении исследования (Вольфкович, 1974).
Исследования Самойлова были впоследствии продолжены в многочисленных работах
отечественных и зарубежных ученых. Изучался химический состав ископаемых
организмов и различных продуктов их жизнедеятельности, анализировались
закономерности преобразования органического вещества в различных геологических
процессах (седиментогенез, образование полезных ископаемых и т. д.). Особое
внимание уделялось роли органического вещества в концентрации и миграции
химических элементов. В частности, было установлено участие лигнина растений
в образовании углей, торфов, выяснено значение ароматических структур лигнина
в накоплении урана, меди, германия и т. д. в ископаемом органическом веществе.
Применение новых методов исследования дало возможность изучать геохимическую
роль не отдельных элементов, а органических соединений (порфирины, аминокислоты,
липиды и т. д.) и продуктов их распада в процессах образования ископаемого
органического вещества. Итоги многолетних палеобиогеохимических исследований
были подведены во многих статьях и книгах (Abelson, 1963; Манская, Дроздова,
1964; Манская, Кодина, 1975; Schopf, 1975; Lowenstam, 1975, 1984; Degens,
1976; Cloud, 1976; Вологдин, 1976; Дроздова, 1977; Дроздова, Соколов, 1979,
и др.).
Мы не намерены вмешиваться в споры геологов о том, какие из гипотез Самойлова
о биогенном происхождении того или иного минерала подтвердились, а какие
нет. Данный вопрос выходит за пределы нашей компетентности. Но необходимо
отметить главное. К настоящему времени полностью подтверждено предположение
Самойлова о способности живого вещества концентрировать в больших количествах
элементы из разбавленного раствора. „Известно, что в современной биосфере
организмами массами извлекаются из недонасыщенных растворов углекислые
соли кальция, магния и стронция, кремнезем, фосфаты, йод, фтор и другие
компоненты” (Лапо, 1982, с. 90). Благодаря интенсивному
103
белковому обмену животные в окружающую среду выделяют огромные количества
веществ, содержащих фосфор и азот. Значительно расширено и число минералов,
в образовании которых участвуют организмы. Так, согласно X. Ловенштаму
(Lowenstam, 1975; Lowenstam, Margulis, 1980), организмы образуют следующие
минералы: карбонаты (кальцит, арагонит, фатерит, моногидрокалыщт, аморфный
карбонат кальция); фосфаты (даллит, франколит, аморфные гидрогели фосфата
кальция и фосфата железа); кремнезем (опал); окислы и гидроокислы железа
(магнетит, гетит, лепидокрокит, аморфные гидраты железа); сульфаты (целестин,
барит, гипс); галиды (флюорит).
Вместе с тем палеобиогеохимические исследования подтвердили вывод Самойлова
о том, что ископаемые остатки не всегда отражают химический состав древних
организмов, а следовательно, базируясь только на них, нельзя делать однозначные
выводы о выполняемых ими биогеохимических функциях в биосфере. Так, ископаемые
аминокислоты редко соответствуют аминокислотному составу организмов, образовавших
осадочную породу. Содержание и состав ее аминокислот предопределяются в
значительной степени процессами диагенеза и катагенеза. Поэтому сравнительные
исследования химического состава различных групп современных организмов,
каждая из которых может рассматриваться как представитель определенного
этапа в истории биосферы, остается по-прежнему главным методом изучения
эволюции биогеохимических функций биосферы. Именно этот способ стал доминирующим
в последующем изучении эволюции концентрационных функций биосферы в трудах
В. И. Вернадского, А. П. Виноградова, Е. М. Бойченко.
4.2. Проблема эволюции биогеохимических функций биосферы
в трудах В. И.
Вернадского
Проблема биогеохимических функций биосферы привлекла внимание Вернадского
в самом начале разработки им учения о биосфере. В двух лекциях о химическом
составе живого вещества и его связи с химией земной коры, прочитанных
в 1922 г. в Петрограде и в Праге, Вернадский (1922б) подчеркнул, сколь
грандиозна
роль организмов в геохимической истории отдельных химических элементов.
Опираясь на скудные и очень разрозненные данные того времени о химическом
составе органического мира, он составил таблицу распределения 28 химических
элементов в биосфере. Даже при самом грубом сравнении среднего химического
состава живого вещества с содержанием этих элементов в гидросфере, атмосфере
и литосфере было ясно, как велико значение организмов в распределении
и миграции элементов в биосфере.
Вместе с тем Вернадский подчеркнул, что по среднему содержанию того или
иного элемента нельзя судить о концентрационных возможностях организмов
и их участии в геологической истории данного элемента. Так, при среднем
процентном содержании марганца в живом веществе 10-4 – 10-2%
встречаются виды, содержащие марганец до 1 % (лишайники, грибы) и даже
6 – 7% (бактерии
– Bacterium marganicum, Crenotrix
104
leptotrix) (Вернадский, 1927а, с. 74). Подобные различия в концентрационных
способностях особенно ярко проявляются между гетеротрофами и автотрофами,
но нередко они встречаются и внутри царств. Дело в том, что морфологическая
эволюция организмов, которая уже более полувека изучалась биологами, была
тесно связана с историческими преобразованиями химического состава организмов.
Как видно из составленной Вернадским таблицы распределения химических элементов
в отдельных таксономических группах, каждая из них отличалась химической
специфичностью.
Базируясь на этой таблице, Вернадский в 1922 г. сформулировал представление
о геохимических концентрационных функциях отдельных таксономических групп.
Он выделял четыре группы организмов по их способности концентрировать те
или иные химические элементы. В первую группу он включил организмы, накапливающие
элементы в процентном отношении намного больше, чем их имеется в среде.
Такие организмы он называл кремниевыми, серными, железными, кальциевыми
и т.д. Во вторую группу попали организмы, просто „богатые данным элементом”,
т. е. имеющие его больше, чем кларк данного элемента в биосфере. К первой
группе он относил организмы, содержащие определенный элемент от 10% и выше,
ко второй – от 1 до 10%. Далее по классификации Вернадского следовали „обычные”
и „бедные” организмы.
Казалось бы, что такая классификация организмов, взятая в филогенетическом
аспекте, должна была навести Вернадского на мысль об эволюции концентрационных
функций всей биосферы. Но в начале 20-х годов главная задача Вернадского
заключалась не в этом. Основное внимание он уделял доказательству закономерного
характера связи живого вещества с земной корой. Поэтому основной упор тогда
был сделан на положении, что в целом геохимические функции живого вещества
остаются одинаковыми в течение геологического времени. В те годы Вернадский
писал, что лишь интенсивное развитие промышленности вызвало значительные
изменения в биогеохимических функциях биосферы.
Для познания участия организмов в миграции были необходимы количественные
данные о химическом составе живого вещества. В связи с этим большое значение
имели исследования по определению химического состава отдельных видов,
начатые Вернадским в 1919 г. Впоследствии в эти исследования включились
В. С. Садиков, предложивший методику химического анализа живого вещества,
и А. П. Виноградов (Садиков, Виноградов, 1924; Садиков, 1923,1926; Виноградов,
1929). По этой методике под руководством Вернадского проводились исследования
химического состава организмов, принадлежащих к различным группам. С
образованием в 1928 г. Биогеохимической лаборатории эти исследования приобрели
особый
размах и масштабность. В работах сотрудников лаборатории, а также ученых
из других научных учреждений и высших учебных заведений (М. П. Белая,
Г. Бергман, Б. К. Бруновский, X. Г. Виноградова, Т. И. Горшкова, В. М.
Дирш,
В. В. Ковальский, К. Г. Кунашева, А. Н. Лебедянцев, Д. П. Малюга, Л.
С. Селиванов, М. Р. Сканави-Григорьева, К. Ф. Терентьева, Б. А. Холодовский)
изучалась роль организмов
105
в концентрации брома, йода, фтора, титана, радия, никеля, меди, марганца,
кобальта, молибдена и др. В результате было получено большое количество
точных данных и сделан ряд теоретических обобщений, касающихся участия
организмов в осуществлении биогеохимических концентрационных функций.1 Так, исследуя влияние беспозвоночных на образование некоторых горных
пород, К. Ф. Терентьева (1932) показала высокое содержание углекислого
магния
в скелете у современных иглокожих. На основании этого она предположила,
что если у древних иглокожих содержание магния было столь же велико,
то эти организмы могли способствовать образованию доломитов.
Было начато изучение изменений концентрационных функций организмов в процессах
видообразования. Обитающие в одних и тех же водоемах близкородственные
виды рясок (Lemna minor и L. polyrhiza) отличались между собой по содержанию
радия (Бруновский, 1932). Причем сезонные колебания радия в воде находились
в обратной зависимости от содержания этого элемента у организмов. Весеннее
обеднение воды радием вызывалось интенсивным ростом рясок, концентрирующих
радий. Осенью же накопленный радий вновь возвращался в воду. Бруновский
показал, что в предельных случаях содержание радия у рясок в 56.5 раза
больше его содержания в пруду.
С 1937 г. к биогеохимическим исследованиям подключился Отдел сравнительной
биохимии (заведующий В. В. Ковальский) Института биохимии АН УССР. В проведенных
ими исследованиях суточных и сезонных колебаний химического состава животных
было показано, что каждая таксономическая группа отличается своеобразным
характером подобной ритмики (Ковальский, 1941). Тем самым представления
о видоспецифичности концентрационных функций дополнялись данными и о динамическом
характере этой специфичности.
Аналогичные исследования в те годы проводили и зарубежные биогеохимики,
биохимики, геохимики (Э. Кидсон, Дж. Макхарт, В. Ноддак, X. Ремедже, М.
Фокс и др.). Упомянем только некоторых из них. Это работы В. Р. Дж. Аткинса
о концентрации соединений фосфора и азота морскими организмами. Г. Лунде
и К. Клосс доказали способность обитателей моря концентрировать йод в количествах,
превышающих содержание его в морской воде в сотни тысяч раз. Об участии
организмов в концентрации ванадия в асфальтитах писал Г. Фестер.
Среди зарубежных работ, выполненных в 30-е годы, особенно выделяются классические
исследования норвежского геохимика В. Гольдшмидта (1938), который, изучая
угли из округа Гартли в Англии, установил, что почти все редкие элементы
концентрируются в золе углей.2
1 Эти данные публиковались преимущественно
в „Трудах Биогеохимической лаборатории”, которые выходят с 1930 г.
2 Работы Гольдшмидта получили такую популярность,
что даже идея Вернадского о концентрации живым веществом рассеянных элементов
за рубежом
стала
приписываться Гольцшмидту под названием „принципа биогенной концентрации
элементов”.
Однако сам Гольдшмидт во всех своих работах подчеркивал, что приоритет
в биогеохимических исследованиях принадлежит Вернадскому, под влиянием
которого он и обратился к изучению роли биогенной концентрации
106
в образовании горючих ископаемых. Поэтому А. Е. Ферсман имел все основания
писать в 1934 г.: „Классические исследования акад. Вернадского первые поставили
глубоко научный и точный анализ биогеохимических процессов, и по его пути
(курсив наш, - Э. К.) пошли американцы (Кларк), немцы, скандинавы (В. М.
Гольдшмидт) и французы” (с. 51).
Коэффициент обогащения золы этих углей редкими элементами по сравнению
с их содержанием в земной коре достигал для германия и мышьяка – 1600,
бора и висмута – 1000, цинка – 250, бериллия – 180, платины – 120 и
т. д. Как видно из этого перечня, в одинаковой мере концентрировались элементы,
принадлежащие к самым разным группам периодической таблицы и отличающиеся
химическими свойствами, радиусом атомов, валентностью, зарядами ионов
и
т. п. Здесь типичные литофилы концентрировались вместе с сидерофильными
и халькофильными элементами. Совпадение во многих случаях коэффициента
обогащения у элементов, резко различающихся по химическим свойствам,
по мнению Гольдшмидта, свидетельствовало о том, что их концентрации объяснялись
процессами, протекающими в растениях при поглощении ими почвенных растворов
и испарении. Растения выполняли преимущественно роль фильтрирующего
и испаряющего
аппарата. Элементы накапливались при испарении в листьях, остатки которых
служили вначале для образования гумуса, а позднее и углей „при окаменении
богатых гумусом почв” (там же, с. 315). Признавал Гольдшмидт и роль
чисто биологических процессов при концентрации таких, например, элементов,
как
бор. В любом случае (физические или биологические процессы лежали в
основе концентрирования редких элементов в растениях) было ясно, что обогащение
золы углей редкими элементами обусловлено развитием специфических концентрационных
функций у растений, остатки которых и превратились в горючие ископаемые.
Опираясь на многочисленные результаты отечественных и зарубежных исследований,
Вернадский пришел к выводу, что химический состав организмов не только
не повторяет геохимическое строение биосферы, но, напротив, в значительной
степени оказывает активное влияние на среду. Этот состав является результатом
эволюционно сложившегося обмена веществ, под воздействием которого
и идет изменение концентраций элементов в биосфере. Этот вывод составил
ядро представлений
Вернадского о биогеохимических функциях биосферы, которые он изложил
в книге „Химическое строение биосферы Земли и ее окружения”. Вернадский
выделял
пять основных типов биогеохимических функций: 1) газовые, проявляющиеся
в участии организмов в создании и поддержании газового состава атмосферы;
2) концентрационные; 3) окислительно-восстановительные; 4) биогеохимические
функции, связанные с биогенной миграцией атомов в процессах питания,
дыхания, размножения и разрушения организмов; 5) биогеохимические функции
человечества.
В ходе осуществления концентрационных функций биосферы, по мнению Вернадского,
образуется множество соединений, которые не возникают абиогенным путем.
Среди них не только белки, аминокислоты,
107
но и металлоорганические соединения. Вернадский полагал, что число соединений,
созданных организмами, значительно превышает количество известных к тому
времени 3000 минералов, возникающих абиогенным путем.
Вернадский в 30-40-х годах выделял две группы концентрационных функций
(1940, 1965). К первой группе он относил функции, в ходе осуществления
которых происходит захват элементов, встречающихся у всех организмов. Вторую
же группу составляют концентрационные функции, которые осуществляются только
отдельными группами организмов. Уже из этого деления видно, что Вернадский
в эти годы четко ставил вопрос об эволюции концентрационных функций биосферы.
Если в своих первых работах по биосфере он основной упор делал на общих
характеристиках влияния организмов на среду, то позднее не раз подчеркивал,
что в ходе геологического времени не только существенно менялись в количественном
отношении отдельные функции, но и появлялись качественно новые функции.
Допускал он также, что отдельные частные функции становились всеобщими.
При этом он чаще всего в качестве примера приводил кальциевую функцию.
Уже давно было установлено, что организмы, будучи мощными концентраторами
кальция, создают громадные месторождения углекислого кальция. Между тем
в докембрийских отложениях не было организмов с кальциевыми скелетами.
Внезапное появление организмов с известковым скелетом в позднем докембрии
оценивалось Р. Дэли (Daly, 1909) и Р. Лаллом (Lull, 1923) как возникновение
качественно новой кальциевой функции. В четвертом издании „Очерков геохимии”
Вернадский (1934) по существу присоединился к этой точке зрения. Он подчеркивал,
что на грани протерозоя и палеозоя, возможно, впервые в биосфере появились
организмы с кальциевым скелетом. Эти организмы, по его мнению, коренным
образом изменили историю кальция в земной коре. Он считал возможным, что
именно в этот период истории биосферы кальций „впервые вошел в состав всех
организмов” (1965, с.249). Этот перелом в истории кальция он связывал с
возникновением моллюсков, брахиопод, иглокожих, кораллов, криноидей и других
беспозвоночных. Правда, Вернадский сомневался, „явился ли в это время кальций
впервые в концентрационной функции 1-го рода, или, что вероятнее, его значение
усилилось и изменилось для концентрационной функции 2-го рода” (там же).
Вернадский попытался свести в рамках единой таблицы имеющиеся тогда данные
о среднем химическом составе всего живого вещества, а также об изменениях
его концентрационных функций в связи с морфологической эволюцией. В таблице
было показано участие отдельных таксономических групп в концентрации
уже 39 (а не 28, как это было в начале 20-х годов) элементов. Из таблицы
видно,
как смена доминирующих групп в биосфере могла сказаться на осуществлении
концентрационных функций живого вещества (там же, с. 251—263). Вернадский
показал, что „биогенная миграция атомов также меняется в течение всего
эволюционного процесса, так как меняются организмы” (там же, с. 265).
Но, к сожалению, эта эволюция чаще всего отражается в газообразных
108
соединениях, что затрудняет исследование количественных и качественных
параметров этих преобразований.
Немногие из биологов-эволюционистов, современников Вернадского, смогли
в полной мере оценить значение этих идей Вернадского для понимания сложных
причинно-следственных связей в преобразованиях организмов и окружающей
их среды. К числу этих немногих принадлежит один из создателей эволюционной
экологии С. А. Северцов. Ссылаясь на идеи Вернадского о постоянстве химического
состава видов, Северцов (1951, с. 48) отмечал, что подобное постоянство
„определяет однородность химического воздействия на среду и однородность
восприятия вещества из внешней среды при метаболизме” и в конечном счете
„определяет энергию экологической жизнедеятельности вида как целого”. Под
влиянием продуктов жизнедеятельности организмов происходят направленные
изменения самой среды. В свою очередь эти изменения выступают в качестве
факторов видообразования.
Таким образом, в трудах Вернадского впервые были сформулированы представления
о биогеохимических концентрационных функциях биосферы и предложена их классификация.1 Выделение общих и частных концентрационных функций открывало путь к изучению
их эволюции. Вернадский впервые поставил также вопрос не только о количественных
преобразованиях общих концентрационных функций, но и о возможности появления
качественно новых частных функций в ходе эволюции биосферы. Разработка
этих идей Вернадского была осуществлена в трудах А. П. Виноградова.
4.3. Учение А.П.Виноградова об эволюции химического состава
организмов и выполняемых ими геохимических функций
В разработке представления об эволюции биогеохимических функций биосферы
особая роль принадлежит исследованиям А.П.Виноградова, выполненным с
конца 20-х до середины 40-х годов. Крупным событием в разработке данной
проблемы
стала публикация его книги „Химический элементарный состав организмов
моря” (1935 – 1944, ч. 1 – 3), в которой был обобщен материал о химическом
составе
не только современных и ископаемых организмов, но и некоторых биогенных
пород. В начале 50-х годов эта книга была издана в США, и по сей день
она является наиболее полной сводкой по химическому составу живого вещества.2
1 В настоящее время отмеченная Вернадским концентрационная
функция организмов широко используется как у нас в стране, так и за рубежом
при поиске рудных
месторождений на основе биогеохимического анализа растений (см.: Малюга,
1963).
2 В примечаниях к таблицам о содержании химических элементов
в разных группах организмов, составленных Г. Бовеном (Bowen, 1966, р. 73),
автор
прямо
пишет, что подавляющее большинство данных он взял из американского издания
книги
Виноградова.
109
Впервые к интересующей нас теме Виноградов обратился в докладе на заседании
Ленинградского общества естествоиспытателей, прочитанном 28 февраля 1931
г. Вскоре этот доклад был опубликован в журнале „Природа”. Здесь Виноградов
попытался рассмотреть химический состав организмов в связи с вопросами
их систематики и морфологической эволюции. Он полагал, что обсуждение данных
о содержании тех или иных химических элементов в различных таксономических
группах будет способствовать не только выяснению закономерностей распределения
этих элементов в биосфере, но и установлению связи между процессами видообразования
и биогеохимической эволюции всей биосферы. Как и Вернадский, Виноградов
был убежден, что изучение химического состава организмов „связывает явления
жизни с историей химических элементов в земной коре” (1931, с. 254). При
разработке этой проблемы Виноградов столкнулся с рядом трудностей как объективного,
так и субъективного порядка. Дело в том, что большинство ископаемых организмов,
лишенных скелета, практически не оставили следов в палеонтологической летописи.
Лишь современное живое вещество было „доступно в целом нашему непосредственному
изучению”, — писал Виноградов в 1932 г. в небольшой научно-популярной книге
„Геохимия живого вещества” (с. 7).1 Поэтому для выяснения
закономерностей эволюции концентрационных функций биосферы прежде всего
было важно установить
точный химический состав современных организмов. Сравнительное изучение
его позволяло заглянуть в прошлое биосферы, так как многие из современных
организмов являются „химическими реликтами” и содержат информацию об
особенностях химического состава живого вещества в прошлых биосферах,
а следовательно,
и информацию об особенностях выполняемых тогда биогеохимических функций.
Имевшийся к тому времени материал не раскрывал подлинной картины химического
состава отдельных типов, классов, отрядов, а тем более родов и видов.
Несмотря на 130-летнюю историю изучения химического состава организмов,
практически
не было работ, в которых был бы проведен анализ всего организма. Исследовались
чаще всего отдельные органы: скелет у животных, семена, листья, корни
и т.д. у растений. К тому же обычно учитывалось количественное содержание
5 – 10 так называемых биогенных элементов (Н, С, О, Р, N, К и др.), составляющих
более 99% живой массы организмов. На остальные элементы долгое время
не
обращалось внимание, так как они оценивались как результат случайного
„химического загрязнения”. Только к началу 20-го в. было доказано, что
многие из этих
элементов (например, Си, Mn, Zn, Si, As) необходимы для нормального протекания
физиологических процессов. Однако количественные данные о содержании
этих элементов у разных организмов практически еще отсутствовали. К тому
же
методика
1 Эту книгу мы называем сейчас научно-популярной ретроспективно,
так как за ней последовали фундаментальные труды Виноградова (1935а,
1937, 1944)
по данному вопросу. Но именно эту книгу А. Е. Ферсман (1934) назвал
прекрасной сводкой о содержании элементов в живом веществе, к которой по
состоянию
знаний на тот день нельзя было ничего прибавить.
110
исследования химического состава организмов по процентному содержанию
элемента к живому весу, к сухому весу и к золе была столь различна, что
мешала сведению воедино данных о его содержании во всем веществе биосферы.
И наконец, только 0.5 % всех современных видов были изучены в химическом
отношении.
Отсутствие крупных обобщающих работ о химическом составе разных таксономических
групп послужило, по-видимому, причиной того, что в начале 30-х годов Виноградов
стоял фактически на антиисторических позициях в трактовке биогеохимических
функций биосферы. Он утверждал, что на протяжении всей истории биосферы
действовали одни и те же концентрационные функции (кальциевая, магниевая,
кремниевая, карбонатная и т. д.), которые лишь переходили от одной группы
организмов к другой. В качестве примера он называл апатитовую функцию,
которая в палеозое выполнялась брахиподами, трилобитами, а впоследствии
— позвоночными животными. В те же годы Виноградов и кальциевую функцию
относил к числу вечно действующих в биосфере, называя в качестве ее возможных
носителей в докембрийской биосфере бактерии, водоросли и простейшие.
Вместе с тем даже тот весьма фрагментарный материал, которым располагал
Виноградов при написании книги „Геохимия живого вещества» по существу
мог бы трактоваться и с эволюционных позиций. Прежде всего это данные об
эволюции
геохимических функций в филогенезе отдельных таксономических групп. Гомологичные
ряды химических признаков, т. е. возрастающие или убывающие ряды содержаний
брома, йода, марганца, ванадия, магния и т. д. в разных группах организмов,
явно свидетельствовали о количественных изменениях геологических функций
в ходе эволюции. Имелись и факты появления качественно новых концентрационных
функций и угасания некоторых более древних. Так, концентрация серы, железа,
магния осуществляется главным образом бактериями и низшими растениями.
Высокое содержание ванадия (10-2 % живого веса или 1% золы)
характерно только для аспидий [sic – В. П.], а меди (1.10-2 –
5.10-3 %) - у моллюсков
и членистоногих. Мощными концентраторами алюминия были и остались хвощи
и плауны, роль которых, однако, в современной биосфере намного ниже,
чем в биосфере позднего палеозоя.
Даже столь беглый перечень некоторых данных, приведенных в названной книге,
о специфике геохимических функций у отдельных организмов, явно свидетельствует
о том, что в ходе эволюции биосферы происходило и преобразование геохимических
функций живого вещества. Тем более это стало очевидным после выхода в
свет книги „Химический элементарный состав организмов моря”. В ней систематизированы
данные о химическом составе более 4000 видов. Диапазон приводимых сведений
простирается от бактерий до высших растений и позвоночных. В качестве
литературных
источников использовано более 2000 работ. Результаты собственных аналитических
исследований и обработка данных других авторов были суммированы в 325
таблицах, многие из которых содержат более сотни показателей, а одна таблица
— даже
2400 показателей.
111
Виноградов выполнил многосторонний анализ химического состава организмов
моря в зависимости от их систематического положения, области обитания,
особенностей физико-географических и химических условий среды, сезонной
ритмики и т. д. Были выявлены также некоторые тенденции изменений в содержании
тех или иных элементов в пределах отдельных таксономических групп. В основу
представлений о химическом составе организмов прошлых биосфер Виноградов
положил данные о химическом составе реликтовых организмов и биогенных пород.
Использовались и знания о процессах преобразования остатков ископаемых
организмов. Таким образом, при реконструкции химического состава вымерших
групп учитывались как прямые палеонтологические, так и косвенные (минералогические
и неонтологические) данные.
Опираясь на сравнительно-исторические и актуалистические приемы исследования,
Виноградов сформулировал представления о главных направлениях в эволюции
химического состава органического мира. Эти представления имеют непосредственное
отношение к исследуемой нами проблеме. По сути дела был выявлен ряд закономерностей
в эволюции концентрационных функций. Среди них — уменьшение числа биогеохимических
функций у высших организмов по сравнению с низшими, нарастающая специализация
организмов по их функциям, параллелизмы в эволюции функций в генетически
несвязанных филумах, усиление одних биогеохимических функций и ослабление
других, приобретение организмами новых функций, перенос отдельных функций
с одной группы организмов на другие. Остановимся кратко на этих закономерностях,
оставляя в стороне подробные сведения об эволюции химического состава организмов.
Прежде всего поражает удивительное разнообразие геохимических функций прокариот
и эукариот (одноклеточных), доминирующих в докембрии. Как правило, в роли
концентраторов различных элементов выступают бактерии и цианофеи. Среди
них существуют мощные концентраторы железа, меди, серы и др. Интересен
тот факт, что прокариоты по количеству азота резко отличались от всех других
растений, к царству которых их относили в те годы. Опираясь на сугубо геохимические
соображения, Виноградов предложил эту группу организмов выделить из царства
растений. Это предложение полностью соответствует современным представлениям
о макросистеме органического мира. Бактерии и синезеленые водоросли теперь
выделены в самостоятельное царство дробянок, Mychotalia (Тахтаджян, 1973;
Whittaker, Margulis, 1978). В самостоятельное царство протистов выделяют
и одноклеточных эукариот, геохимические функции которых столь же разнообразны,
как и функции дробянок (Серавин, 1984). Минеральные скелеты одноклеточных
могут содержать арагонит, кальцит с примесями МаСO3 [sic –
В. П.], целестин, сульфат бария, опал, гидроокиси и карбонаты Mn, Fe,
Al, кремнезем. У них
имеются все типы минеральных скелетов, за исключением фосфатовых. Значительная
часть этих скелетов, по Виноградову, не встречается
112
у многоклеточных, а другие — лишь у наиболее примитивных их представителей.1
Это исключительное разнообразие сочетается с высокой модификационной изменчивостью
химического состава низших организмов. Можно предположить, что специфический
фактор эволюции простейших — способность к длительным модификациям —
оказывается и фактором стабилизации биосферы.
Виноградов не раз отмечал, что в ходе прогрессивной эволюции происходила
интенсификация концентрационных функций. Весьма характерны в этом отношении
красные водоросли, у которых впервые появляется способность концентрировать
практически все рассеянные элементы в десятки, сотни и даже тысячи раз
больше, чем их содержится в среде. Важным этапом в эволюции концентрационных
функций стал переход от неспециализированного накопления элементов во всех
тканях примитивных организмов к накоплению их в виде сложных комплексных
соединений в специализированных тканях, где они играют важную физиологическую
роль (например, йод в щитовидной железе, медь в дыхательных пигментах).
„В течение эволюции, - писал Виноградов, - та или другая геохимическая
функция для известных форм была ими потеряна, но сохранилась вызванная
ею физиологическая функция того или иного химического элемента” (19356,
с. 20). В связи с этим уместно вспомнить оригинальное объяснение закономерностей
распространения элементов в живом веществе, предложенное Виноградовым.
Он полагал, что химический состав живого вещества есть периодическая функция
атомного номера. Из составленного им графика встречаемости элементов в
живом веществе видно, что „все химические элементы, находящиеся по распространению
в житом веществе в максимумах кривой, являются главным субстратом (связанных
с их геохимической функцией) жизнедеятельности древних форм организмов
и их современных потомков” (там же, с. 23).
Виноградов показал, что совершенствование способа концентрации того или
иного элемента протекает зачастую параллельно в различных филумах. Так,
среди бурых и красных водорослей, а также губок наряду с семействами,
умеренно концентрирующими йод, встречаются и сильные его концентраторы,
а скрепи
последних — и исключительно мощные концентраторы йода. Аналогичная картина
наблюдается с изменениями концентраций меди и цинка у моллюсков, ванадия
у асцидий, магния, марганца, железа, серы и азота в других группах животных.
Это явление Виноградов (1944, с. 199) назвал параллельными рядами накопления.
Как правило, существует прямая зависимость интенсивности той или
1 Последний вывод не соответствует новейшим данным. Как следует
из составленной крупным американским биогеохимиком X. Ловенштамом (Lowenstam,
1975,
1984) таблицы распределения минеральной составляющей живого вещества океана,
сложноорганизованные животные (хордовые и моллюски) образуют наибольшее
количество минеральных скелетов, в то время как бактерии и некоторые
водоросли синтезируют только по одному минералу. Фактически лишь простейшие
- это
вершина эволюции на клеточном уровне – могут сравниться с высокоорганизованными
животными по количеству выделяемых минералов.
113
иной концентрационной функции от общей эволюционной продвинутости группы.
В роли мощных концентраторов в каждом филуме выступают обычно более прогрессивные
формы. „Наиболее богатые (в смысле накопления того или иного химического
элемента) организмы — крайние члены этих параллельных рядов накопления
— одновременно являются в эволюционном ряду наиболее высокостоящими по
сравнению с другими” (там же, с. 202). Правда, „точный смысл, генетическое
значение этих рядов, - вынужден был признать Виноградов, - можно выяснить
лишь путем дальнейших исследований” (там же). К сожалению, ни сам Виноградов,
ни его ученики к этой проблеме больше не обращались.
Оценивая выделенные Виноградовым закономерности в эволюции концентрационных
функций биосферы, следует признать, что их эволюция в пределах отдельных
филумов идет в соответствии со вторым биогеохимическим принципом Вернадского,
согласно которому жизнь эволюирует в сторону создания форм, усиливающих
биогенную миграцию атомов в биосфере. Виноградов показал также, что некоторые
биогеохимические функции биосферы получают наибольшее развитие лишь на
отдельных этапах ее эволюции, а затем резко снижают свою интенсивность.
Плауны, доминирующие в карбоне и перми, активно осуществляли концентрацию
алюминия, в значительной степени сейчас подавленную. К числу угасающих
функций по накоплению элементов он относил и кремниевую, которая встречается
лишь среди наиболее низкоорганизованных представителей животных и растений.
У высших животных кремний сохранил лишь физиологическое значение. Здесь
он встречается в незначительных количествах (Tacke, Wannagat, 1979; Воронков,
Кузнецов, 1984).
Вслед за Вернадским Виноградов считал, что в ходе эволюции биосферы возможен
перенос тех или иных химических функций с одной группы организмов на другую.
Он показал, что такие действующие с раннего кембрия концентрационные функции,
как апатитовая, магнезиально-кальциевая, железная и т. д., в разные периоды
осуществлялись различными таксонами.
Виноградов вполне справедливо считал, что химический состав организмов
характеризует выполняемую им геохимическую функцию в биосфере. В связи
с этим он ставил задачу установить связь повышенного содержания в организме
того или иного элемента с выполняемой им физиологической функцией. Ее решение,
по мнению Виноградова (1944), доказало бы, что вовлечение данного химического
элемента в обмен веществ организма означало приобретение им новой геохимической
функции. Тем самым открывалась возможность для изучения взаимодействий
биохимической и физиологической эволюции организмов с биогеохимической
эволюцией биосферы.
В начале 60-х годов Виноградов вновь вернулся к этому вопросу, подчеркивая,
что выполнение той или иной геохимической функции зависит от систематического
положения организма. Таким образом, он впервые доказал, что геохимические
концентрационные функции действительно „получают различное развитие в
ходе эволюции организмов” (1935а, с. 247). Выдвинутые Самойловым и Вернадским
теоретические
114
положения он дополнил богатым фактическим материалом, а также сделал целый
ряд новых обобщений, раскрывающих закономерность эволюции биогеохимических
функций биосферы. Следует согласиться с Л. А. Зенкевичем, что именно А.
П. Виноградову удалось на богатом фактическом материале показать, что преобладание
в разные геологические периоды групп организмов с различным химическим
составом обусловило изменения в химическом составе всей биосферы (Зенкевич,
1951).
4.4. Основные направления современных исследований
Суть современных представлений об эволюции биогеохимических функций биосферы
выразил В. В. Ковальский в статье, посвященной возникновению и эволюции
биосферы. „Вместе с филогенетическими изменениями организмов... менялась
роль организмов в миграции атомов в биосфере” (1963а, с. 45). Это положение
в той или иной форме встречается в самых разнообразных работах геологов,
геохимиков, почвоведов и биогеохимиков, в которых так или иначе затрагивалась
обсуждаемая нами проблема. При этом исследования велись по двум основным
направлениям. Первое из них связано с попыткой выяснить основные тенденции
в эволюции концентрационных функций в пределах отдельных таксономических
групп. Для второго направления характерно стремление исследовать эволюцию
концентрационных функций не отдельных таксонов, а всей биосферы в целом.
Нет возможности рассмотреть все работы, в которых продолжались начатые
В. И. Вернадским и А. П. Виноградовым исследования концентраций тех или
иных атомов разными видами одного таксона. Ограничимся некоторыми наиболее
характерными из них. Так, изучение содержания кальция, магния, стронция,
марганца, железа, кремния, титана, алюминия в скелетных остатках современных
и ископаемых кораллов позволило выделить некоторые тенденции в изменении
концентраций этих элементов в пределах данной группы (Козлова и др., 1980).
Кораллы раннего палеозоя отличались повышенной способностью концентрировать
магний, марганец, железо, кремний по сравнению с кораллами из позднего
палеозоя и мезозоя. Соотношения Mg/Sr и Ca/Sr, достигнув максимального
значения в ордовике и силуре, в дальнейшем существенно снизились. Соотношение
Ca/Mg, несмотря на ряд колебаний, в целом оказалось более высоким у современных
форм, чем у их палеозойских и мезозойских предков.
Подобные комплексные исследования, базирующиеся на сравнении химического
состава предковых форм и их современных потомков, имеют большое значение
для изучения исторических преобразований концентрационных функций, выполняемых
той или иной группой организмов. Но при этом приходится постоянно сталкиваться
с искаженной картиной этой эволюции в результате вторичного изменения
химического состава в фоссилизированных остатках (перекристаллизация, диагенетические
и катагенетические изменения ископаемого органического вещества).
115
Тем не менее этот подход интенсивно развивается в работах геологов и геохимиков,
которые на основе данных об эволюции органогенных пород судят об особенностях
развития концентрационных функций у организмов, из остатков которых сложены
эти породы. Приведем в качестве примера работы В. Г. Кузнецова (1983) и
Ю. П. Казанского (1983). Эти авторы эволюцию рифостроителей считают одной
из главных причин эволюции химического и минерального состава рифов и общего
морского карбонатонакопления. Этим объясняется высокое содержание магния
в рифейских и нижнекембрийских рифах, их преимущественное сложение из кальцита
в нижнем и среднем палеозое и появление рифов с преобладанием арагонита
во второй половине палеозоя. Состав рифостроителей в мезозое и кайнозое
предопределил первичное сложение рифов из арагонита и высокомолекулярного
кальцита. Возрастание интенсивности фотосинтеза в районах формирующихся
рифов обусловило рост концентраций стронция. Этому способствовали и организмы
с арагонитовым скелетом, где стронция всегда больше, чем в кальциевых скелетах.
Для понимания механизмов эволюции концентрационных функций биосферы интересны
попытки выяснить связь процессов видообразования с изменением способностей
организмов накапливать те или иные элементы (Ковальский, 1974; Ковальский,
Боровик-Романова, 1978; Жулидов, Емец, 1981; Емец, Жулидов, 1983; Криволуцкий,
1983; Шарыгин, 1984; Покаржевский, 1985, и др.). Так, В. М. Емец и А. В.
Жулидов исследовали содержание Zn, Co, Си, Мо трех видов жуков-мертвоедов
из рода Silpha в районах их совместного обитания в Восточной Сибири. По
мере увеличения концентрации этих элементов был установлен следующий ряд:
S. obscura, S. carinata, S. perforata. Оказалось, что и филогенез, построенный
по морфологическим признакам, совпадает с этим рядом. Наиболее продвинутый
по морфе вид S. perforata содержит наибольшее количество микроэлементов
и отличается резкими половыми различиями по этому признаку.
Авторы попытались выяснить выявленные ими тенденции в эволюции химического
состава жуков-мертвоедов с точки зрения учения о биосфере. Они пришли
к выводу, что полученные данные согласуются со вторым биогеохимическим
принципом
Вернадского. „Повышение роли видов насекомых в биологических круговоротах
химических элементов, - по мнению авторов (1983, с. 555), - может достигаться
за счет увеличения содержания в их теле химических элементов в эволюционном
ряду близких форм и за счет формирования у эволюционно продвинутых видов
половых различий в аккумулировании химических элементов”. К сожалению,
такие работы по изучению видообразования крайне редки. И к изучению эволюции
биогеохимических функций по-прежнему обращаются гораздо чаще геологи,
минералоги, геохимики, чем биологи-эволюционисты. Правда, в последние годы
интерес
к элементному составу отдельных таксонов все больше усиливается в связи
с попытками использовать организмы как индикаторы степени загрязнения
среды (Криволуцкий, 1983; Покаржевский, 1985, и др.).
116
Особенности развития концентрационных функций у отдельных таксонов все
больше исследуются и зарубежными авторами. Их результаты не раз излагались
в обзорах современных знаний в области биогеохимии, которые с 1946 г. по
инициативе Дж. Хатчинсона выпускает Американский Музей естественной истории
(Kamen, 1946; Bertrand, 1950; Hutchinson, 1950). Здесь особенно необходимо
отметить фундаментальный труд Дж. Хатчинсона (Hutchinson, 1950), который
дал исчерпывающую сводку данных о специфической геохимической деятельности
морских птиц. Ее результатом стали мощные месторождения гуано, содержащие
в значительных количествах азот, фосфорную кислоту, калий и кальций. В.
Крумбейн (Krumbein, 1978) показал, что в продуктах жизнедеятельности некоторых
микроорганизмов концентрация марганца в 1 200 000, железа в 650 000, серебра
в 24 000, ванадия в 420 000 раз больше по сравнению с концентрациями этих
элементов в окружающей среде. Очевидно, что столь высоких концентрационных
способностей не достигла никакая другая группа организмов, будь то растения,
грибы или животные. В связи с этим нужно признать, что второй биогеохимический
принцип Вернадского применим лишь в пределах царства. За пределами царств
этот принцип теряет свое значение, так как бактерии, синезеленые и простейшие
– нередко более мощные концентраторы отдельных элементов, чем многоклеточные
организмы.
Но, превосходя многоклеточные организмы по развитию отдельных концентрационных
функций, они в целом уступают им по разнообразию синтезируемых органических
соединений и даже минералов. В таблице, составленной X. Ловенштамом (Lowenstam,
1975) и дополненной Л. Маргелис (1983), показано, как усложнялись в целом
концентрационные функции биосферы с появлением высокоорганизованных животных
и растений. Бактерии выделяют всего три минерала (даллит, арагонит, кальцит).
Ярче выражена концентрационная функция у простейших, где помимо названных
минералов образуются также аморфные гидраты железа, целестин, барит. Но
подлинными чемпионами по выработке минералов стали моллюски, которые выделяют
еще и моногидрокалыгит, аморфный карбонат кальция, фатерит, аморфный карбонатный
апатит, франколит, магнетит, гетит, лепидокрокит и флюорит. По числу синтезируемых
минералов моллюски превосходят даже позвоночных животных. К тому же скелет
у них нередко строится из нескольких минералов.
В последние десятилетия внимание исследователей нередко было направлено
на поиск механизмов эволюции конкретной концентрационной функции. Особенно
часто объектом подобных исследований становилась кальциевая функция, так
как внезапное появление организмов с кальциевым скелетом, как отмечалось
выше, издавна рассматривалось в качестве примера эволюции биогеохимических
функций биосферы. Это событие получило название „скелетной революции”.
К настоящему времени накопилось немало гипотез, призванных объяснить эту
„революцию”. Одни авторы, вслед за А. П. Виноградовым (1940), дают этому
явлению чисто геохимическое объяснение, утверждая, что в условиях перенасыщения
первичных морских вод углекислым газом, хлором и аммиаком был раствор,
в котором раковины не могли
117
возникнуть, а если даже и возникали, то должны были вскоре раствориться.
Другие авторы приобретение скелетов объясняли необходимостью защиты от
хищников, возникновение которых якобы приурочено к концу докембрия (Hutchinson,
1961; Stanley, 1976; Заварзин, 1979; Медников, 1985). При этом Г. А. Заварзин
отмечал, что образование у эукариот скелетов означало вовлечение в биологический
круговорот таких химических элементов, как кремний, стронций, калий и т.
д. До этого прокариоты лишь косвенно влияли на миграцию элементов путем
изменения физико-химической обстановки. В неокатастрофических гипотезах
причины данного события усматривались в каких-то космических процессах,
вызвавших резкое ускорение мутагенеза в разных группах животных. В результате
могли одновременно появиться мутанты — обладатели кальциевого скелета (Schindewolf,
1963).
Были и попытки как-то синтезировать эти геохимические, адаптационистские
и неокатастрофические гипотезы. Такая попытка была сделана И. Н. Крыловым
(1972), который считал целесообразным учитывать широкий набор факторов,
обусловливающих возникновение и закрепление в биосфере животных преимущественно
с кальциевым скелетом. Среди геохимических факторов он называет не только
изменение химического состава морских вод, но и изменение газового состава
атмосферы, образование озонового экрана и т. д. Такое комплексное изменение
геохимических параметров среды, по мнению Крылова, вызвало направленный
мутагенез, в ходе которого возникали формы, адаптированные к новым условиям.
Каждая из этих гипотез уязвима для критики, и споры вокруг них не утихают
по сей день. При обсуждении гипотезы Виноградова указывалось, что высокое
содержание углекислого газа в морской воде не помешало почему-то образованию
мощных отложений карбоната кальция под воздействием денитрифицирующих и
декальцинирующих бактерий, а также сохранению остатков породообразующих
цианофитов в виде строматолитов, онколитов и т.д. (Вологдин, 1976). А между
тем возраст этих пород достигает 2 – 2.3 млрд лет. Следует учитывать также,
что практически одновременно с образованием карбонатных скелетов появились
фосфатные и кремниевые. И этот факт не объяснить снижением концентрации
углекислого газа в морских водах.
Трудно объяснить повсеместное „одевание” организмов в скелеты и только
необходимостью защиты от хищников. Ни одна адаптация не является абсолютной.
Не гарантирует защиту от хищников и твердый скелет. Обладатели твердого
скелета – диатомовые водоросли – лежат в основе трофической структуры в
морских экосистемах. Давая выигрыш в пассивной защите, скелет нередко закрывал
для животных путь к более активной борьбе с хищниками.
Недостаток неокатастрофических гипотез эволюции заключается в том, что
в них постулируется некий механизм эволюционных преобразований, реальность
которого еще должна быть доказана. Что же касается модифицированного
варианта этой гипотезы в работах Крылова о направленном химическом мутагенезе,
то
в ней по сути дела постулируется возможность возникновения наследственных
изменений,
118
Таблица 3
Эволюция восстановительных функций
(по: Бойченко, Удельнова, Юферова, 1969)
| Восстановление организмами E'o, В, при рН 7 |
Усвоение СО2 клетками, % от сухой массы/ч |
Участие соединений металлов |
Время появления в биосфере |
При окислениях органических веществ абиогенного происхождения E'o >
до +0.05 |
Гетеротрофное усвоение первичными организмами,
n · 10-3 |
K, Mg |
Архей |
| При окислениях неорганических веществ в окружающей среде E'o до
+0.30 |
Хемосинтезы у автотрофных бактерий и синезеленых водорослей,
n · 10-2 |
Fe |
Архей,
протерозой |
При окислениях веществ
на свету с образованием
перекисей E'o, до +0.55 |
Фоторедукция у разных
представителей водорослей,
n · 10-1 |
Fe
Cu, Ti
Mn, Ni |
Протерозой,
палеозой |
При окислении воды на
свету с выделением кислорода E'o, до +0.82 |
Фотосинтез у разных типов зеленых растений,
n · 100 - 101 |
Fe, Mn |
Протерозой,
палеозой,
мезозой |
адаптированных к факторам, вызвавшим эти мутации. Таким образом, проблема
„скелетной революции” до сих пор не имеет удовлетворительного решения.
В серии работ Е. А. Бойченко, Л. Н. Захаровой, М. А. Пущевой, Г. Н. Саенко,
Т. М. Удельновой, С. М. Черногоровой, опубликованных в конце 60-х – начале
80-х годов, были подведены итоги многолетних исследований по эволюции окислительно-восстановительных
и концентрационных функций биосферы под влиянием эволюции растительного
мира. Эти исследования были начаты еще в 1935 г. по инициативе Вернадского.
Авторы систематизировали и обобщили данные физиологии, биохимии, а также
исторической геохимии об участии отдельных групп микроорганизмов и растений
в эволюции концентрационных функций. Им удалось показать, что вовлечение
все большего числа металлов в качестве катализаторов в окислительно-восстановительные
реакции автотрофов означало и эволюцию концентрационных функций биосферы,
так как автотрофы составляли более 99 % всей ее биомассы.
В ряде работ (Бойченко, 1967, 1968, 1984; Бойченко и др., 1968, 1969, 1972,
1977, 1979) были намечены основные четыре этапа эволюции восстановительных
функций, связанные с изменениями источников энергии: переход от брожения
при низких значениях окислительно-восстановительного потенциала (E'o)
к более эффективному восстановлению в процессах фотосинтеза (табл.3).
В целом эта эволюция завершилась еще в докембрии. Осуществлялась она путем
использования все большего количества веществ с высокими значениями E'o,
благодаря которым и осуществлялись восстановительные функции. При гетеротрофном
усвоении первичными
119
организмами CО2 в качестве катализаторов используются ионы калия
и магния. Переход к хемосинтезу был связан с использованием железа с низким
значением
E'o. При фоторедукции наряду с железом в восстановлении углекислоты
участвуют также липидные соединения, включающие медь, титан, марганец,
никель. И
наконец, в процессах фотосинтеза восстановление CО2 осуществляется
соединениями двухвалентного железа с участием марганца. Рост окислительно-восстановительного
потенциала от +0.05 В до +0.82 В привел соответственно и к ускорению
прироста органического вещества от 10-3% за 1 ч от сухой
массы клетки при гетеротрофном
усвоении CО2 до 101% при фотосинтезе. В итоге
произошло резкое увеличение общей биомассы биосферы.
В эволюции наземной растительности неуклонно повышалось содержание марганца,
достигшее особенно высокого уровня в специализированных органах фотосинтеза
у семенных растений. Авторы пришли к выводу, что „наблюдающееся в эволюции
биосферы возрастание биохимических функций находится в тесной связи с изменением
содержания и форм нахождения в организмах поливалентных металлов... которые
шире и чаще используются в обмене, и они наложили яркий отпечаток на эволюцию
всех биогеохимических функций” (Удельнова и др., 1979, с. 186—187). Нарастающее
разнообразие обмена веществ у растений и усиление их биогеохимических функций
проявилось как в увеличении содержания разных металлов в ферментах, так
и в уменьшении доли железа по сравнению с другими металлами. Так, отношение
Fe/Mn с 15 у цианофитов падает до 1 у покрытосеменных. Уменьшается и отношение
Fe/Cu и Fe/Ti, достигая минимума у моховидных; минимум Fe/Cr наблюдается
у зеленых водорослей.
В совместных работах Е. А. Бойченко, Г. Н. Саенко и Т. М. Удельновой (1968,
1972) было доказано также, что вовлечение все большего числа металлов в
качестве катализаторов в окислительно-восстановительные реакции означало
и эволюцию концентрационных функций биосферы. Она выражалась в увеличении
числа концентрируемых металлов, в изменении количественных соотношений
между ними, в переходе от „прямого поглощения” металлов к созданию ферментов,
включающих металлы. В этих ферментах концентрация металлов в сотни и тысячи
раз превышает среднее содержание в клетке. Само возникновение разнообразных
концентрационных функций, по мнению авторов, связано со становлением автотрофного
питания.
Собранный материал свидетельствовал, что основные этапы эволюции концентрационных
функций биосферы связаны со сменой доминирующих групп автотрофов (Бойченко
и др., 1968; Удельнова и др., 1979). Авторы показали, что наиболее высокое
содержание железа у синезеленых водорослей (табл. 4). У красных водорослей
при некотором уменьшении железа повышается содержание кобальта и меди.
Для зеленых и бурых водорослей характерно повышенное содержание меди,
ванадия, титана и хрома, у мхов и папоротников высокие значения алюминия,
у покрытосеменных
— марганца. Сопоставление данных об элементном составе различных групп
растений с результатами исследований палеобиогеохимиков, геохимиков и
геологов (Г. Бовен, Дж. Шопф,
120
Таблица 4
Эволюция концентрационных функций растений
(по: Бойченко, Саенко, Удельнова,
1968)
| Элемент |
Растения с более высоким % элементов |
Содержание Mn в клетках % · n |
Время появления в биосфере |
Mg |
Бактерии (Bacteria) гетеротрофные |
10-6 - 10-5 |
Архей |
Fe |
Синезеленые водоросли (Суаnophyta) |
10-4 |
Конец архея, протерозой |
Cu |
Зеленые водоросли (Chlorophyta) |
10-3 |
Конец протерозоя, начало палеозоя |
Zn |
Бурые водоросли (Phaceophyta); папоротникообразные (Pteridophyta) |
10-3
10-2
|
Палеозой |
Mn |
Покрытосеменные (Anthophyta), их листья |
10-2 - 10-1 |
Конец мезозоя, кайнозой |
Дж. Ламсден, Н. М.Страхов, А.Б.Ронов и др.) показало, что наибольший
расцвет автотрофов с повышенным содержанием того или иного элемента синхронен
с
образованием их мощных месторождений и увеличением содержания в осадочных
породах. Так, наиболее крупные месторождения никеля (72 % запасов капиталистических
стран), железа (68 %) и кобальта (93 %) относятся к периоду доминирования
в биосфере синезеленых водорослей, которые содержат эти элементы в больших
количествах (Быховер, 1963). В осадочных породах верхнего протерозоя
и нижнего палеозоя зафиксировано высокое содержание ванадия, что, возможно,
связано с доминирующим положением в биосфере зеленых и бурых водорослей
(Перельман, 1961; Сауков, 1961). Деятельность бурых водорослей и папоротников,
по мнению авторов, обусловила формирование верхнепалеозойских месторождений
цинка. И наконец, широкое распространение покрытосеменных скоррелировано
во времени с образованием многочисленных рудных месторождений марганца
(73% разведанных запасов).
В связи с этим большой интерес представляют собранные Е. А. Бойченко с
соавт. данные об ежегодной концентрации современными автотрофами элементов
и о разведанных запасах их месторождения (в т):
Элемент
|
Концентрируется ежегодно при фотосинтезе |
Мировые запасы сырья |
Углерод
|
1011 |
1012 |
Фосфор
|
109 |
1010 |
Хром
|
105 |
108 |
Марганец
|
107 |
108 |
Железо
|
108 |
1011 |
| Кобальт |
105 |
106 |
121
| Элемент |
Концентрируется ежегодно
при фотосинтезе |
Мировые запасы сырья |
| Никель |
106 |
107 |
| Медь |
107 |
108 |
| Цинк |
107 |
107 |
| Молибден |
105 |
106 |
Если допустить, что накопление металлов растениями является единственным
способом образования рудных месторождений, то тогда известные к настоящему
времени месторождения различных геологических времен могли бы образоваться:
железа – за 1400 млн лет; меди – 38 млн лет; кобальта – 19 млн лет; никеля
– 14 млн лет; марганца и молибдена – 6 млн лет (Удельнова и др., 1979).
Таким образом, вполне допустимо предположение о том, что эволюция рудных
месторождений связана с эволюцией концентрационных функций биосферы.
Хотя исследования в этих направлениях только начинаются, уже ясно, что крупные
изменения в биогеохимических функциях биосферы в значительной степени
связаны
с преобразованием состава металлов и их соединений в ферментативных системах
фотосинтеза. Об этом свидетельствуют данные о химическом составе всех
автотрофных организмов и данные о содержании металлов в фоссилизированных
остатках
древних организмов и биогенных пород.
* * *
Таким образом, за 75-летнюю историю исследований концентрационных функций
биосферы достигнут значительный прирост знаний о закономерностях
их эволюции. Начатые Самойловым исследования изменений химического состава
организмов
как факторов эволюции горных пород и минералов были в дальнейшем
подняты
на уровень биосферных исследований. Было показано, что в ходе прогрессивной
эволюции биосферы возрастает участие организмов в миграции элементов
и увеличивается число биогеохимических функций. Дальнейшее изучение
взаимообусловленной эволюции биохимических и физиологических процессов организмов
с осуществляемыми
ими биогеохимическими функциями представляется нам чрезвычайно перспективным
для познания закономерностей в эволюции биосферы.
122
ОГЛАВЛЕНИЕ
Стр.
Предисловие ...............................................................................................................................................................
3
Глава 1. Исходные положения ...............................................................................................................................
6
1.1. О структуре и функциях биосферы ................................................................................................................
6
1.2. Эволюция биосферы (общие представления) .............................................................................................
13
1.3. Взаимосвязь эволюции видов и развития биосферы .................................................................................
18
1.4. Этапы и направления в изучении эволюции биосферы ...........................................................................
25
Глава 2. В. И. Вернадский об эволюции биосферы ...........................................................................................
27
2.1. Идеи о взаимодействии эволюции видов и эволюции биосферы ............................................................
28
2.2. Гипотеза о возникновении биосферы ...........................................................................................................
40
2.3. Основные тенденции в эволюции биосферы ..............................................................................................
46
2.4. Учение о ноосфере как заключительном этапе эволюции биосферы ...................................................
54
Глава 3. Изучение геохимических факторов эволюции биосферы ...............................................................
62
3.1. „Химическая предопределенность” живого вещества .............................................................................
63
3.2. Общие закономерности химической дифференциации живого вещества в биосфере
...................... 71
3.3. Исследование биогеохимических провинций и их роли в эволюции биосферы
................................. 77
3.4. Геохимические факторы микроэволюции и ее влияние на биогенные циклы элементов...............
83
Глава 4. Эволюция биогеохимических концентрационных функций биосферы .......................................
95
4.1. Роль палеобиогеохимических исследований в изучении эволюции
концентрационных функций биосферы ..............................................................................................................
96
4.2. Проблема эволюции биогеохимических функций биосферы в трудах
В. И. Вернадского ...................................................................................................................................................
104
4.3. Учение А. П. Виноградова об эволюции химического состава организмов
и выполняемых ими геохимических функций ................................................................................................
109
4.4. Основные направления современных исследований ..............................................................................
115
Глава 5. О преобразующем влиянии биологической эволюции
на геохимическое строение биосферы ....................................................................................................................
123
5.1. Влияние эволюции живого на газовый состав атмосферы ...........................................................................
123
5.2. Жизнь и эволюция гидросферы .......................................................................................................................
129
5.3. Биогенные факторы эволюции литогенеза .....................................................................................................
134
5.4. Роль биотических агентов в эволюции почв ..................................................................................................
141
Глава 6. Первые шаги к изучению главных тенденций
в эволюции биосферы .................................................
147
6.1. Эволюция общей биомассы биосферы и ее биологической продуктивности .............................................
147
6.2. Изменения энергетики биосферы .....................................................................................................................
152
6.3. Изменение информационного „фонда” биосферы ..........................................................................................
162
6.4. Эволюция биотического круговорота ..............................................................................................................
166
6.5. К вопросу об основных этапах развития биосферы .......................................................................................
179
Глава 7. Особенности органической эволюции в современной биосфере
(вместо заключения) .................................................................................................................................................
192
Послесловие ..............................................................................................................................................................
204
Литература .................................................................................................................................................................
206
Именной указатель ....................................................................................................................................................
228