| Главная | Библиотека | Форум | Гостевая книга |
Глава 5
О ПРЕОБРАЗУЮЩЕМ ВЛИЯНИИ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
НА ГЕОХИМИЧЕСКОЕ
СТРОЕНИЕ
БИОСФЕРЫ
Начиная с 40-х годов в СССР публикуется все больше работ, в которых эволюция биосферы трактуется прежде всего как возникновение под влиянием биологической эволюции необратимых изменений в химическом составе ее биокосных и биогенных компонентов. При этом обычно имеют в виду преобразование газового состава атмосферы, химических свойств гидросферы и осадочных пород литосферы, эволюцию процессов образования минералов и формирования месторождений полезных ископаемых, почвообразования и т. д. Эти исследования проводятся геологами, геохимиками, географами, почвоведами. Их тематика выходит за рамки целей и задач нашей монографии. Мы остановимся лишь кратко на главных результатах работ, связанных с изучением влияния организмов на исторические преобразования атмосферы, гидросферы, верхних слоев литосферы и почв.
5.1. Влияние эволюции живого на газовый состав атмосферы1
В. И. Вернадский предполагал, что все газы современной атмосферы по своему
происхождению связаны с живым веществом. Но внимание последующих исследователей
сконцентрировалось преимущественно на выяснении роли живого вещества в изменениях
концентраций кислорода и углекислого газа. Стало общепринятым, что „современное
состояние биосферы и ее эволюция на протяжении последних двух миллиардов
лет в известной мере определялись взаимодействиями организмов с атмосферными
газами, в основном с углекислым газом и кислородом” (Будыко,1984, с. 214).
Сейчас в атмосфере содержится 1.2 · 1021 г кислорода,
или примерно 21 % ее объема, и 0.0002 · 1021 г углекислого
газа, что составляет 0.03 %. Если учесть,
что в процессах фотосинтеза ежегодно выделяется около 2 · 1017 г кислорода (Шипунов, 1980), то весь имеющийся в атмосфере кислород мог образоваться
за несколько тысяч лет. Это период полного
1 Мы не затрагиваем здесь пионерские работы В. А. Костицына (1984), начавшего еще в 30-х гг. во Франции разработку математических моделей эволюции атмосферы, биосферы и ее органического вещества. В дальнейшем это направление в изучении эволюции биосферы получило развитие в трудах Н. Н. Моисеева и его сотрудников (1980, 1985).
123
круговорота кислорода. Разность между его приходом в биосферу и расходом
на окислительные процессы — решающий фактор в эволюции газового состава атмосферы.
С этим согласны почти все занимающиеся проблемой исторических преобразований
атмосферы. Но конкретные выводы каждого из них резко отличаются друг от друга.
Не имея возможности останавливаться на анализе расчетов и методов, положенных
в основу той или иной гипотезы, мы попытаемся охарактеризовать только некоторые
итоги развития проблемы.
Долгое время наиболее признанной была гипотеза, в которой центральным событием
в истории атмосферы считался переход от атмосферы, состоящей из аммиака,
метана, углекислого газа и паров воды, к кислородной атмосфере. Этот переход
коренным образом изменил геохимическую обстановку в биосфере, преобразовав
ее геологические, физико-географические, биогеохимические и эволюционно-биологические
процессы. Гипотеза о восстановительном характере первичной атмосферы получила
широкую известность благодаря работам А. И. Опарина и Дж. Холдейна, в которых
принималось, что только в условиях такой атмосферы могла зародиться жизнь.
Зачастую обсуждался вопрос не о том, какой была первичная атмосфера, а какой
она должна была быть, чтобы в ней могла появиться жизнь (Заварзин, 1984).
Начиная с 20-х годов стали накапливаться геологические доказательства об
отсутствии кислорода в докембрийской атмосфере (см. обзоры: Виноградов, 1947;
Urey, 1952, 1959; Rubey, 1955; Rankama, 1955; Руттен, 1973, и др.). В сочетании
с традиционными исследованиями физиологических и биохимических аспектов эволюции
фотосинтеза (К.А.Тимирязев, В. Н. Любименко, Д. И. Сапожников и др.) эти
доказательства использовались для реконструкций эволюции атмосферы.
Одним из первых в нашей стране к этой проблеме обратился А. А. Григорьев
(1936), который поставил вопрос о зависимости изменений концентраций кислорода
и углекислого газа в атмосфере и эволюции организмов. Он предположил, что
объем кислорода, выделяемого при фотосинтезе, превышал в среднем его расход
на дыхание организмов и на окисление химических соединений. В итоге „количество
кислорода в атмосфере постепенно возрастало” (там же, с. 15). Обсуждая механизмы
воздействия организмов на расход углекислого газа в разные периоды истории
биосферы, Григорьев допускал, что в дотретичных биосферах, где доминировали
пойкилотермные животные, при потеплении климата увеличивалось количество
углекислого газа.
Для понимания механизма влияния фотосинтеза на состав атмосферы большое значение
имели работы А. П. Виноградова. В 1941 г. он совместно с Р. В. Тейс экспериментально
доказал, что основная масса выделяемого растениями кислорода образуется не
путем восстановления углерода из углекислого газа, как это было тогда общепринято,
а в процессе дегидрогенизации воды.1 Несколько позже (1947) он
показал, что ни один из абиогенных способов образования кислорода по своим
1 В том же году аналогичные результаты были получены американскими учеными С. Рубеном, М. Каменом и др.
124
масштабам не может сравниться с фотосинтезирующей деятельностью растений
и обеспечить накопление свободного кислорода в атмосфере. В эти годы он вслед
за Вернадским полагал, что переход от восстановительной атмосферы к кислородной
произошел в геологически короткое время, буквально за несколько тысяч лет.
Впоследствии Виноградов отводил этому переходу сотни миллионов и даже миллиардов
лет. В работе о воздействии фотосинтеза на эволюцию биосферы Виноградов (1973)
обратил особое внимание на образование кислорода как важнейшего биогеохимического
процесса. Он подчеркивал, что создание азотно-кислородной атмосферы и озонового
экрана коренным образом изменило радиационный пояс Земли и в значительной
степени предопределило всю дальнейшую эволюцию жизни. Возрастающая концентрация
кислорода резко расширила пространственные границы биосферы и обеспечила
захват жизнью новых экологических зон. Превращение же дыхания в главный способ
окисления органических соединений повысило эффективность энергетических процессов
в организмах, общую активность их жизнедеятельности, а тем самым и общую
интенсивность биогеохимических и энергетических процессов в биосфере.
Возросший интерес к проблемам биопоэза стимулировал появление ряда работ
по эволюции атмосферы (Poole, 1941, 1951; Urey, 1952, 1959; Rubey, 1955;
Rankama, 1955; Соколов, 1959; Олейников, 1962, и др.). Среди них наибольшую
известность в 50-е годы получили работы Дж. Пула и Г. Юри. Их основное положение
заключалось в том, что метан, аммиак, углекислый газ и водяные пары были
главными компонентами атмосферы на первых этапах ее развития. Кислород лишь
в незначительных количествах мог образовываться при фотодиссоциации водяного
пара. Обогащение атмосферы кислородом началось только после возникновения
фотосинтеза. Как полагал Юри, с достижением концентрации кислорода 0.001
(точка Юри) от его современного уровня стало возможным развитие аэробных
организмов и постепенное становление кислородной атмосферы. Предполагалось,
что точка Юри была достигнута примерно 2 млрд лет тому назад. И с этого времени
фотосинтезирующая деятельность растений превратилась в основной источник
кислорода.
Несмотря на популярность взглядов Пула и Юри в 50-е годы, высказывались сомнения
в существовании восстановительной атмосферы на древней Земле. Так, В. Руби
(Rubey, 1955) допускал, что наиболее вероятными компонентами древней атмосферы
были углекислый газ и азот, а не метан и аммиак. Процессы фотодиссоциации
воды, по его мнению, могли быть мощными источниками свободного кислорода
и сами по себе могли привести к значительному накоплению свободного кислорода
в атмосфере.
Вопрос о составе древней атмосферы в настоящее время оживленно дискутируется
(Квасов и др., 1980; Gribben, 1982; Holland, 1983; Крылов, 1983, Будыко,
1984; Заварзин, 1984, и др.). Большинство ученых склоняется к мысли, что
без учета паров воды основными компонентами докислородной атмосферы были
не метан и аммиак, а углекислый газ (примерно 81 % от общего объема) и азот
(6.4%). Эти выводы
125
базируются на изучении состава вулканических газов и атмосфер Венеры и Марса
(Оуэн, 1986; Принн, 1986). Вместе с тем в последние десятилетия становятся
вновь популярными и гипотезы об относительно больших количествах свободного
кислорода в первичной атмосфере (Соколов, Барсков,1988).
Долгое время все рассуждения об эволюции газового состава атмосферы носили
чисто качественный характер. Вот почему большой интерес вызвали работы Л.
Беркнера и Л. Маршалла (Berkner, Marshall, 1964, 1965), в которых предлагались
количественные оценки содержания кислорода в атмосфере разных геологических
периодов. Авторы выделили ряд переломных моментов в истории атмосферы, оказавших
огромное воздействие на эволюцию жизни. Они исходили из того, что в течение
долгого времени существования биосферы насыщение ее атмосферы кислородом
шло очень медленно, и лишь к началу фанерозоя была достигнута критическая
точка, равная 1 % кислорода от его современного содержания. В конце силура
в накоплении кислорода выделяют вторую критическую точку (10%), которая,
по мнению авторов, дала возможность жизни выйти на сушу. В карбоне содержание
кислорода было максимальным и в 2-3 раза превышало современное. Затем произошло
резкое его снижение. И лишь в мезозое устанавливаются концентрации, близкие
к современным. Опираясь на данные эволюционной палеонтологии, Беркнер и Маршалл
(Berkner, Marshall, 1964,1965) попытались показать, как рост концентрации
свободного кислорода в атмосфере вел к освоению новых регионов биосферы и
к бурной адаптивной радиации жизни. Высказывалась ими и мысль о биогенной
регуляции климата путем воздействия растительности на газовый состав атмосферы.
Гипотеза Беркнера и Маршалла получила дальнейшее развитие и обоснование в
трудах Г. В. Баринова (1972), М. И. Будыко (1975, 1977а, 1984 и др.) и О.
П.Добродеева (1975).
Вопрос о взаимодействии изменений газового состава атмосферы и эволюции органического
мира особенно подробно обсуждался в работах Будыко, опубликованных за последние
15 лет. Он показал, что уменьшение содержания углекислого газа в атмосфере
привело в конце палеозоя к падению биопродуктивности растительности и к снижению
общей биомассы биосферы. Заслуживает внимания и его гипотеза о неслучайном
характере временных совпадений появления животных с высоким уровнем метаболизма
с периодами максимального содержания кислорода в атмосфере (млекопитающие
– в конце триаса, птицы – в середине юрского периода). Будыко полагает, что
в эпоху повышения содержания кислорода в атмосфере увеличивалось разнообразие
организмов, а в эпохи снижения этих концентраций происходил обратный процесс.
Происходящее сейчас увеличение углекислого газа в атмосфере он оценивает
как „омоложение биосферы”, как ее возврат к тем периодам в истории биосферы,
когда ее продуктивность была выше.
В работах О. П. Добродеева и Г. В. Баринова развивалась гипотеза об исторических
преобразованиях растительности, атмосферы и климата как едином саморегулируемом
процессе. Ведущая роль в этом процессе отводилась изменениям в интенсивности
фотосинтезирующей
126
деятельности растений. Этими изменениями объяснялись периодические смены
эпох оледенения и потепления. Баринов полагал, что прямые и обратные связи
между растительностью и климатом осуществляются путем саморегуляции и самоускорения
биологической эволюции. Периодические смены климатических эпох сопровождаются
интенсивными процессами видообразования. Вновь возникающие виды растений,
по мнению автора, обладали более мощными и совершенными механизмами фотосинтеза
и метаболизма, что ведет к сокращению периодов биосферных ритмов и к ускорению
эволюции в органическом мире.
Работы Баринова, Будыко и Добродеева чрезвычайно важны для решения традиционной
проблемы о роли климатических и других физико-географических факторов в эволюции,
так как доказывают биогенное происхождение многих факторов, ранее считавшихся
сугубо абиотическими.
Отмеченные Беркнером и Маршаллом тенденции в эволюции атмосферы не вызывали
принципиальных возражений. Но их количественные расчеты уже вскоре были подвергнуты
сомнению. Так, Ю. П. Казанский с соавторами (1969) полагали, что в конце
докембрия концентрация кислорода в атмосфере была довольно высока. М. Руттен
(1973) утверждал, что точка Пастера (1 % современного содержания кислорода)
была достигнута уже в середине архея, 10%-ный уровень - в позднем рифее.
По мнению Б. С. Соколова (1976), точка Пастера также была достигнута в протерозое,
примерно 1 млрд лет тому назад. Еще дальше это событие отодвигает С. Г. Неручев
(1977), согласно расчетам которого 1 %-ное содержание свободного кислорода
могло быть превышено в период от 1.6 до 1.0 млрд лет тому назад. Как общепринятую
цифру 1.5 млрд лет называют Дж. Браун и А. Массет (Brown, Mussett, 1981).
И наконец, некоторые авторы вообще считают, что кислородная атмосфера существует
по меньшей мере 2.5 млрд лет и что концентрации кислорода, близкие к современным,
установились в довольно короткий период времени (Виноградов и др., 1969:
Крылов, 1983).
В последние годы внимание советских ученых сконцентрировалось на проблеме
изменения углекислого газа и кислорода в фанерозое (Будыко, Ронов, 1979;
Верзилин, 1980; Посохов, 1981; Ушаков, Ясаманов, 1984; Будыко и др., 1985;
Ясаманов, 1985). Этот интерес стимулирован исследованиями М. И. Будыко и
А. Б. Ронова (1979), предложившими оригинальную методику для расчета изменений
концентраций углекислого газа и кислорода на основе данных о содержании органического
углерода в осадочных и вулканогенных породах континентов. Из составленной
таблицы динамики газового состава атмосферы (рис. 3, 4) видно, что уже к
началу фанерозоя масса атмосферного кислорода составила приблизительно треть
его современного содержания. В дальнейшем рост концентраций кислорода был
неравномерным. Его резкое увеличение произошло в девоне и карбоне, когда
был достигнут уровень, равный современному. В триасе содержание кислорода
вновь снизилось, и лишь в середине мезозоя произошло новое резкое увеличение,
сменившееся затем длительным периодом убывания. Концентрации же углекислого
газа колебались в течение палеозоя и мезозоя в пределах 0.1 – 0.4%
127
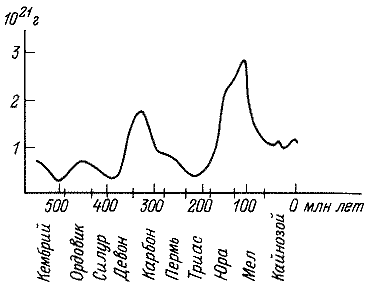
Рис. 3. Изменение массы кислорода в фанерозое (по: Будыко и др., 1985).
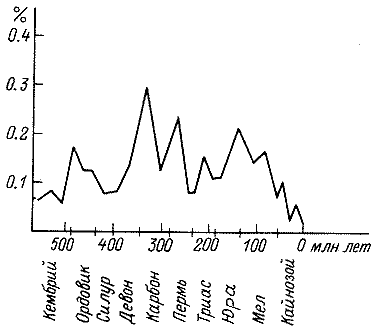
Рис. 4. Изменение концентрации углекислого газа в фанерозое (по: Будыко и др., 1985).
от общего объема атмосферы. И только в конце мезозоя наметилась четкая тенденция
снижения, приведшего в плиоцене к беспрецедентно низким значениям, которые
в несколько раз меньше среднего содержания углекислого газа в фанерозойских
атмосферах.
Н. Н. Верзилин (1980) попытался дополнить исследования Будыко и Ронова. Он
старался учитывать не только данные об органическом углероде в осадочных
породах фанерозоя, но и то количество кислорода, которое было потрачено на
окисление органического и неорганического вещества, содержащегося в толщах
более древних пород. Хотя Верзилин и основывался на тех же данных, что Будыко
и Ронов, использованная им методика расчетов привела к иным результатам.
Это, как отмечает сам автор, ясно показывает, что при реконструкции древних
атмосфер принятая система расчетов имеет большее значение, чем ,,первичный
исходный материал” (Верзилин, с. 15).
Из приведенного выше видно, сколь существенно различаются взгляды советских
географов и геологов о конкретных путях эволюции
128
атмосферы. Аналогичная картина и в зарубежной литературе (Carrels et al.,
1976, Cloud, 1968, 1976; Hart, 1978; Gribbin, 1982; Holland, 1983; Уокер
1984; Оуэн, 1986; Принн, 1986, и мн. др.). Даже популярная сейчас гипотеза
Геи Лавелока (Lovelock, 1979,1983), в которой немалое место занимает вопрос
о биогенном происхождении атмосферы и ее регуляции органическим миром, нередко
подвергается критике. Существуют и гипотезы о сугубо абиогенном происхождении
свободного кислорода из железной сердцевины ядра Земли (Сорохтин, 1974).
Таким образом, результаты многолетних исследовании эволюции атмосферы подтвердили
в целом идеи Вернадского о биогенном происхождении основных компонентов биосферы
и о регуляции ее газового состава жизнедеятельностью организмов. Но если
Вернадский считал, что газовый состав атмосферы, близкий к современному,
установился где-то в раннем архее, то сейчас большинство ученых полагает,
что только в позднем фанерозое произошла подобная стабилизация. Советские
ученые внесли существенный вклад в изучение основных тенденции в преобразовании
газового состава атмосферы. Было подтверждено, что многие абиотические факторы
эволюции контролируются и регулируются живым веществом. Сформулирована мысль
о тесной взаимосвязи процессов преобразования газового состава атмосферы,
климата и эволюции органического мира. Однако и по сей день существуют серьезные
трудности в выборе данных и методов для количественных оценок изменений атмосферы
на разных этапах развития биосферы. Особенно проблематична история докембрийской
атмосферы. Обилие гипотез не компенсирует отсутствие надежных данных о скорости
изменении в древних атмосферах углекислого газа, о времени появления или,
наоборот, исчезновения из атмосферы кислорода, азота, метана, аммиака. Сравнительные
исследования происхождения и эволюции атмосфер планет солнечной системы,
развернувшиеся буквально в последние десятилетия, до сих пор дают скорее
больше вопросов, чем ответов, о механизмах образования атмосферы Земли и
составе первичной атмосферы. Остается неясным также количественное соотношение
биогенного и абиогенного кислорода в современной атмосфере.
5.2. Жизнь и эволюция гидросферы
Благодаря высокой химической активности кислорода увеличение его парциального давления в атмосфере отразилось и на других геосферах. В основе современных взглядов на эволюцию гидросферы лежат идеи Вернадского о магматическом происхождении океанических вод и ведущей роли живого в детерминации их химических свойств. При этом Вернадский подчеркивал, что океан, как продукт эндогенных процессов, заложивших основу его химического состава и концентрации солеи, мало менялся с архейских времен. „С самых древних геологических отложений, с архейской эры, не только в общем состав и формы нахождения воды должны быть аналогичны современному” (Вернадский, 1933, с. 18).
129
Подобное высказывание часто оценивалось как отрицание Вернадским эволюции
гидросферы (Пустовалов, 1940; Посохов, 1981). При этом проходят мимо не менее
характерных для Вернадского высказываний о том, что „в истории природных
вод мы имеем несомненные указания на то, что геологически преходящими могут
быть минеральные виды природных вод, которые не повторяются и которые раньше
не существовали” (1933, с. 12). Не обошел Вернадский и вопрос о факторах
исторических изменений в гидросфере. В живом веществе он видел фактор как
стабилизации основных параметров гидросферы, так и последующих ее изменений
в определенных пределах. Живое вещество, по мнению Вернадского, „целиком
создает все те химические процессы, законы которых мы можем изучать в химической
океанографии” (1923, с. 34).
Взгляды Вернадского на проблему происхождения и эволюции гидросферы океана
заключаются в следующем. В целом океан – продукт магматических процессов,
но в определении его основных свойств немалую роль играла жизнь. Главные
параметры океана стабилизировались еще в раннем архее, а дальнейшие преобразования
гидросферы шли в пределах, в которых могли существовать и эволюировать организмы.
В целом эти взгляды Вернадского принимаются как советскими, так и зарубежными
учеными. Различие коренилось не в том, менялась или не менялась в масштабе
геологического времени гидросфера, а в оценках времени стабилизации ее главных
химических свойств и пределов их изменений. Здесь сложились три точки зрения.
Одни авторы полагали, что эволюция гидросферы прекратилась к началу фанерозоя
(Виноградов, 1959б, 1967; Валяшко, 1971; Гавриленко, 1975; Богданов и др.,
1978). Другие считают, что эволюция гидросферы продолжалась в течение всего
фанерозоя (Пустовалов, 1940; Ронов, 1959; Страхов, 1962; Посохов, 1981).
Наконец, в последние десятилетия стала распространяться точка зрения, согласно
которой с раннего докембрия океан практически не менялся (Garrels, MacKenzie,
1971; Виноградов, 1975; А. и С. Сидоренко, 1971, 1975). Остановимся кратко
на работах названных авторов.
Ведущим фактором на первых этапах эволюции гидросферы была дегазация вещества
мантии. Эта идея, высказанная в трудах Дж. Калпа (Kulp, 1951) и В. Руби (Rubey,
1951), получила всестороннее обоснование в теории происхождения океана А.
П. Виноградова (1967). Отводя дегазации мантии ведущую роль в образовании
океана, Виноградов признавал вместе с тем и влияние жизни на стабилизацию
его современных химических свойств и солевого состава. Об этом свидетельствует
тот факт, что они сформировались одновременно с завершением формирования
главных типов организмов (3 млрд – 600 млн лет назад) . Виноградов выделял
три стадии в истории океана: эволюция океана в отсутствие биосферы (архей)
: эволюция океана под влиянием жизни (конец архея – начало палеозоя); стабильный
состав океана (палеозой – современность). Особенности второго этапа Виноградов
описал в специальной главе „Образование биосферы и эволюция океана”. Уже
появление органических соединений в период зарождения жизни, по мнению Виноградова,
стабилизировало растворимость многих неорганических соединений,
130
изменило характер транспорта взвесей и солей в донные отложения. Постоянной
стала и стратификация вод океана. Особое значение для эволюции гидросферы
имело возникновение фотосинтеза, а следовательно, и кислородной атмосферы.
Окисление СН4 в СO2 в конечном счете вело к стабилизации карбонатно-бикарбонатной
системы океана, для которой характерны активные реакции океанических вод
и устойчивость показателей рН.
Эволюция жизни, согласно Виноградову, привела к смене первичной восстановительно-глеевой
обстановки на окислительную, к постепенному уменьшению кислотности вод и
соответствующему возрастанию их щелочности. Первичный океан постепенно терял
часть элементов катиогенного происхождения. Менялись миграционные формы таких
элементов, как сера, азот и железо. Главными формами миграции для серы стал
ион SO2-4, для железа – Fe2+, а для азота – N2 и NO3. Повысилась подвижность
кальция и магния. Происходило осаждение труднорастворимых солей. Стали отлагаться
мощные биогенные карбонаты кальция и магния. Окисление сульфидов до сульфатов
обусловило переход океанических вод к современному хлоридо-сульфатному типу.
Таким образом, Виноградов доказал существенную роль жизни в формировании
химического состава гидросферы в докембрии. Она стала и важным фактором сохранения
сложившегося состава в фанерозое.
Теорию Виноградова поддержали в своих работах М. Г. Валяшко (1971), Е. С.
Гавриленко (1975) и др. Так, Валяшко писал, что „стабилизация современного
состава произошла где-то между 1500 õ 106 и 500 õ 106 лет
назад. С того времени на Земле не было таких сил и факторов, которые могли
бы изменить
состав вод”
(1971, с. 102). В целом соглашаясь с Виноградовым и Валяшко о времени завершения
эволюции океана, Гавриленко вынуждена признать, что „результаты тектонического
и палеогеографического изучения не дают однозначного решения вопроса о времени
стабилизации объема Мирового океана и его границ” (1975, с.37).
Не случайно многие советские геологи отдавали предпочтение представлениям
У. Твенхофера и И. Вальтера о существенных изменениях как химического состава
солей океана, так и их концентраций на протяжении всей геологической истории
Земли. Критикуя Вернадского, Л. В. Пустовалов писал: „Самая элементарная
логика подсказывает, что состав морских солей и концентрация их меняются
во времени в сторону все возрастающего содержания по крайней мере тех солей,
которыми в настоящее время океан насыщен” (1940, с. 98). Но это утверждение
не подкреплялось фактическими доказательствами. Оставались неизвестными и
причины эволюции гидросферы, так как Пустовалов резко выступал против идеи
о какой-либо существенной роли живого вещества в геологических процессах.
Н. М. Страхов (1962) в основном был согласен с гипотезой Виноградова, но
полагал, что эволюция гидросферы захватила и весь фанерозой. Он выделял четыре
этапа в эволюции океана: азойский, археозойский, протерозойско-рифейский
и исторический (фанерозойский). Основные
131
события этой эволюции, по его мнению, были „связаны с живым веществом” (там,
же, с. 15).
Увеличение биомассы в фанерозойских биосферах, по мнению Страхова, вело к
использованию целого ряда минеральных компонентов для внутриклеточного метаболизма
организмов, построения их металлоорганических соединений, образования различных
внутренних и внешних скелетов. Это особенно справедливым казалось ему относительно
меди, ванадия, железа, кремнезема, карбонатов кальция и магния, фосфатов,
барита и целестина, подвижность и концентрация которых в гидросфере менялись
в течение всего фанерозоя. Также непрерывно увеличивалась щелочность океанских
вод. Как достоверный факт приводил он изменение общей солености океана и
особенно содержания в нем сульфатов. Без всяких оговорок он писал: „В течение
исторического этапа не только колебалась минерализация океанской воды, но
и изменялись количественные соотношения между отдельными солевыми компонентами”
(там же, с. 17).
В результате анализа соленакопления в разные геологические эпохи А. Л. Яншин
пришел к выводу, подтверждающему точку зрения Н. М. Страхова. „За последние
600 млн лет, - пишет Яншин, - состав вод Мирового океана... подвергался существенным
изменениям как в отношении общей величины солености, так и в отношении состава
растворенных солей” (1977, с. 13). Базируясь на количественных соотношениях
между преобладающими ионами в разные геологические эпохи, Ю. П. Казанский
(1977) выделил пять основных этапов в эволюции океана: „1) аммоний-магний-кальциевый
хлоридо-карбонатно-бикарбонатный (архей – ранний протерозой); 2) натрий-кальций-магниевый
бикарбонатно-хлоридный (средний – поздний протерозой); 3) кальций-магний-натриевый
бикарбонатно-хлоридный (кембрий – ордовик); 4) магний-натриевый бикарбонатно-хлоридный
(силур – карбон) и 5) магний-натриевый сульфатно-хлоридный (пермь – кайнозой)”
(с. 64). Как видно из этой периодизации, в пределах фанерозоя было по меньшей
мере три крупных изменения гидросферы. В фанерозое сокращалось количество
кальция, карбоната и бикарбоната, а содержание магния, натрия, сульфат-иона,
напротив, увеличилось.
В начале 80-х годов вышли две монографии: Е. В. Посохова „Химическая эволюция
гидросферы” (1981) и А. И. Перельмана „Геохимия природных вод” (1982). В
них также отстаивалась мысль об эволюции гидросферы в течение всего геологического
времени. Но в отличие от работ Страхова, Яншина и Казанского, Посохов и Перельман
поднимали вопрос не только об эволюции океана, но и других природных вод,
в том числе подземных. Посохов (1981) полагал, что развитие гидросферы шло
постоянно в сторону создания условий, наиболее благоприятных для жизни. Подобное
„улучшение” химизма водной среды проходило при активном участии самих организмов
и продуктов их жизнедеятельности. Особо значительной для гидросферы фанерозоя
оказалась эволюция наземной растительности. С ее появлением увеличилось разнообразие
ионного состава континентальных вод, их обогащение органическим
132
веществом и соединениями CО2, Н2S, NН3,
NO3. Растения оказались также важным фактором формирования гидрохимической
зональности
грунтовых
вод.
Перельман (1982) основное внимание уделил дифференциации природных вод в
связи с возникновением геохимических барьеров. Так, в местах окисления сульфидов
формировались сернокислые воды. Единая слабовосстановительная среда разделилась
на две противоположные: резко окислительную и резко восстановительную. Последняя
стала характерной для болот, придонных горизонтов озер и водоносных слоев
литосферы.
Казалось бы, биогенная эволюция вод океана и гидросферы в целом – общепризнанный
факт. Но в последние годы вновь звучат высказывания о необходимости вернуться
к взглядам Вернадского о стабилизации основных черт гидросферы еще в раннем
архее. Так, в статье „Сколько лет океану? ” В. И. Виноградов пишет, что „основная
масса газоводной оболочки Земли сформировалась еще на начальной догеологической
жизни нашей планеты” и с тех пор „химической состав океанической воды и атмосферы
почти не менялся” (1975, с. 50). Этот вывод автор строит на данных о распределении
изотопов серы в ее окисленных и восстановленных соединениях из архейских
отложений. Он полагал, что биогенный круговорот серы был неизменным 2.5 –
0.2 млрд лет, а достижение современного химического состава вод заняло небольшой
в геологическом масштабе отрезок времени. Подобные выводы В. Виноградов строил
только на результатах собственных исследований, не пытаясь как-то сопоставить
их со всеми геологическими и геохимическими доводами других геологов в пользу
эволюции гидросферы.
И за рубежом есть сторонники идеи о стабилизации химического состава гидросферы
еще в раннем архее. В качестве примера сошлемся на книгу Р. Гаррелса и Ф.
Маккензи (Garrels, MacKenzie, 1971). Правда, авторы признают ненадежность
доводов как сторонников идеи об эволюции гидросферы в рамках биосферы, так
и ее противников. Но сами они присоединяются к постулатам последних. Относя
формирование основных особенностей современной гидросферы на 1.5 – 2.0 млрд
лет в прошлое, они сомневаются в возможности ее значительных изменений в
более поздние периоды развития биосферы.
Таким образом, в отечественной литературе в целом укоренились взгляды о магматическом
происхождении вод океана и о влиянии жизни на становление химического состава
современной гидросферы и концентрации в ней солей различных органических
и неорганических соединений. Гидросфера как под влиянием самих организмов
и продуктов их жизнедеятельности, так и в результате обмена газами с атмосферой,
преобразованной живым веществом, претерпела определенные изменения в ходе
развития биосферы. Однако масштабы изменений гидросферы, основные этапы и
тенденции ее развития остаются все еще не выясненными. Имеющиеся геологические
факты допускают разные интерпретации этих процессов.
133
5.3. Биогенные факторы эволюции литогенеза
Важным направлением в изучении закономерностей эволюции биосферы стали исследования,
посвященные эволюции процессов литогенеза и общих путей и способов миграции
элементов в литосфере. Мысль об эволюции биогенного осадкообразования была
высказана в 1893 г. И. Вальтером, который считал, что „между морфологической
формой и ее литогенетической деятельностью, между формой и образом жизни
организмов существуют постоянные связи, что каждое изменение видового признака
влечет за собой эволюцию литогенетических процессов” (Walther, 1893 – 1894,
S. 1004). Эта мысль далеко не сразу получила должное развитие за рубежом.
За исключением работ Р. Дэли и Е. Штейдманн, исследовавших роль организмов
в эволюции кремниевых и карбонатных пород, а также работ К. Бойрлена об эволюции
углеобразования, идея Вальтера практически не использовалась иностранными
геологами.1
Непростой оказался ее путь и в отечественной науке, хотя периодически она
становилась теоретической основой большинства отечественных исследований
по эволюции литогенеза (см.: Страхов, 1971). Это объяснялось укоренившейся
в нашей стране еще с конца прошлого века биохимической концепцией осадкообразования
(Н. И. Андрусов, М. А. Егунов, Г. А. Надсон, Н. А. Соколов), а также революционизирующим
влиянием биогеохимических работ В. И. Вернадского и палеофизиологических
исследований Я. В. Самойлова. Одна из задач палеофизиологии, по мнению Самойлова,
заключалась в познании закономерностей эволюции породо- и минералообразования
в связи с эволюцией живого (см. гл. 4).
Апелляция к жизни в работах советских геологов в 20 – 30-х годах порою стала
принимать скорее форму словесной отмычки, чем продуктивного метода анализа
геохимических и геологических процессов. Это порождало у некоторых ученых
скептическое отношение к биогеохимической концепции литогенеза и минералообразования.
Бурное внедрение новых физико-химических методов в литологию привело к увлечению
геологов многочисленными физико-химическими концепциями литогенеза, в том
числе и сформулированной Л. В. Пустоваловым (1940) общей физико-химической
теории осадочного образования.
Но уже вскоре стала очевидной неоправданность подобных увлечений. В трудах
А. Д. Архангельского, Л. С. Берга, А. Г. Бетехтина, Г. И. Бушинского, А.
П. Виноградова, А. Г. Вологдина, М. Н. Годлевского, Ю. К. Горецкого, Ю. А.
Жемчужникова, М. Д. Залесского, Б. Л. Исаченко, М. И. Кантора, С. И. Кузнецова,
Б. Л. Личкова, В. П. Маслова, Е. П. Молдаванцева, Д. В. Наливкина, Б. В.
Перфильева, Б. Б. Шлыкова, А. Б. Ронова, А. А. Саукова, Ю. Н. Сеньковского,
Н. М. Страхова, В. О. Таусона, А. И. Тугаринова, А. В.Хабакова, М. С. Швецова
и других исследователей,
1 В то же время в работах палеоботаников Ш. Бертрана, Г. Потонье, Б. Рено, выполненных в конце XIX-начале XX в., было доказано, что различия каустобиолитов зависят от химического состава образующих их древесных растений, водорослей и микроорганизмов животных.
134
опубликованных в 20 – 60-х годах, приводились все новые и новые доказательства
роли организмов и продуктов их жизнедеятельности в процессах осадконакопления,
породообразования и их эволюции (см. подробнее: Лапо, 1982, 1987). Было продемонстрировано
влияние живого на изменение темпов литогенеза, минералогического и химического
состава осадочных пород, закономерностей их географического распространения
и т. д. Благодаря детальному биогеохимическому и геохимическому изучению
ископаемых остатков, проводимому на уровне микрокомпонентов, молекул или
даже атомов, делались выводы о палеогеографической обстановке фоссилизации
этого вещества, о характере его диагенеза, катагенеза, об особенностях живого
вещества в ту или иную эпоху. Среди различных подходов к изучению эволюции
осадкообразования советские ученые сконцентрировали главное внимание на разработке
вопроса об изменении главных факторов осадкообразования в геологической истории
Земли и о влиянии этих изменений на коренные преобразования условий седиментации
и эволюции типов осадочных пород и формаций (Яншин и др., 1983). Эволюция
органического мира, как правило, рассматривалась в тесной связи с изменениями
других факторов осадкообразования (термический режим, газовый состав атмосферы,
окислительно-восстановительный потенциал, солевой состав гидросферы, тектонический
режим Земли и т. д.). Это создавало возможности для синтетических концепций
эволюции литогенеза.
Важным событием в разработке проблемы воздействия эволюции органического
мира на геологические процессы стала V сессия Всесоюзного палеонтологического
общества, состоявшаяся в 1959 г. Ее материалы опубликованы в книге „Значение
биосферы в геологических процессах. Вопросы взаимосвязи палеонтологии и тектоники”
(1962). На этой сессии наряду с палеонтологами, зоологами, ботаниками (Р.
Ф. Геккер, И. И. Горский, К. Б. Корде, Б. С. Соколов, А. И. Толмачев, Д.
М. Федотов) участвовали также и геологи – ведущие специалисты по полезным
ископаемым осадочного происхождения (В. А. Гроссгейм, Е. П. Левандо, Б. Л.
Личков, В. Д. Наливкин, A. M. Обут, В. А. Успенский, В. Е. Хаин). В многочисленных
докладах была рассмотрена роль живого вещества в формировании земной коры
в целом, в образовании карбонатных пород, каустобиолитов, железных и марганцевых
руд, фосфоритов, бокситов, аллитов, в подготовке исходного вещества на стадии
гипергенеза для образования глинистых пород. Как подчеркивалось в программном
докладе сессии, сделанном Б. П. Марковским, необходимо „рассматривать развитие
жизни на Земле как одну из сторон развития Земли” (1962, с. 10). Проблемы
эволюции биосферы стали центральными и в 1979 т. на XXV сессии Всесоюзного
палеонтологического общества (Палеонтология и эволюция биосферы, 1983).
С середины 40-х годов Н. М. Страхов, опираясь на исследования советских ученых
и обобщая данные мировой литературы, попытался создать первую общую теорию
литогенеза и эволюции литогенетических процессов. В наиболее полной форме
она была изложена в начале 60-х годов (Страхов, 1960, 1962, 1963). Автор
показал, что наиболее достоверные следы эволюции осадкообразования связаны
с эволюцией биоса
135
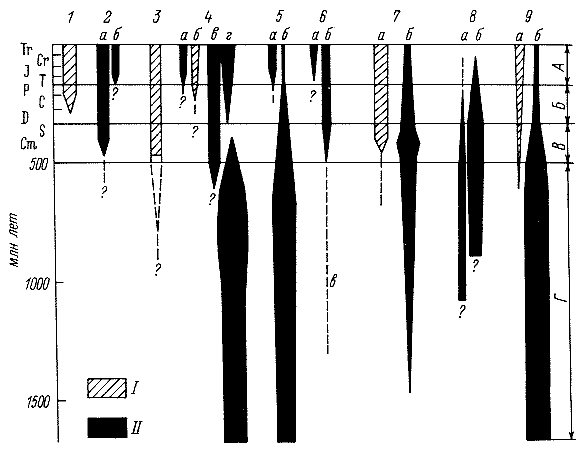
Рис. 5. Стратиграфическое размещение биогенных (I) и хемогенных (II) пород
(по: Страхов, 1963).
Этапы: А – альпийский, Б – герцинский, В – каледонский, Г – докембрийский. 1 – угли; 2 – галогенные породы: а – каменная соль и ангидриты (гипсы), б – калийные соли; 3 – фосфориты; 4 – железные руды: а – коры выветривания, б – озерно-болотные, в – морские оолитовые шалюзит-гидрогетитовые, г – глауконитовые породы, джеспилиты; 5 – марганцевые руды: а – коры выветривания, б – морские; 6 – бокситовые накопления: а – коры выветривания, б – морские и озерные, в – метаморфизованные в наждаки; 7 – известняки: а – органогенные, б – хемогенные; 8 – доломиты первичные: а – нормально морские, б – лагунные; 9 – кремнистые породы: а – органогенные, б – хемогенные.
и его влиянием на литогенез. Начиная с середины протерозоя хемогенный литогенез
постепенно сменился биогенным. Факторами этой замены были все усиливающаяся
фотосинтезирующая деятельность растений, рост общей биомассы в связи с освоением
жизнью пелагических зон моря, выходом ее на сушу и включением все большего
числа химических соединений и элементов в процессы метаболизма. Страхов разработал
оригинальную схему эволюции внешних геосфер и осадочного породообразования
в истории Земли (см. рис. 5).
Вначале влияние биоса, согласно Страхову, было косвенным и проявлялось главным
образом в изменении набора и минерало-петрографических признаков пород. В
результате повышения окислительно-восстановительного потенциала и приближения
рН к нейтральным значениям снизилась подвижность элементов с переменной валентностью
(Fe, Mg, Аl), которые стали перемещаться лишь в составе комплексных органических
соединений, а их осаждение стало происходить преимущественно в форме окисей
в прибрежных районах. Появляются новые типы минералов (глаукониты, фосфориты,
органогенные известняки и биогенные
136
кремнистые породы). К концу протерозоя резко сокращается хемогенное образование
доломитов, которое в дальнейшем стало преимущественно биогенным за счет вноса
некоторыми организмами в осадок MgCО3. Все более вытесняется
и хемогенное образование CaCО3. Если в архее осадкообразование
было одностадийным, то позднее в результате дифференциации биосферы на горизонты
с различными
окислительно-восстановительными
и кислотными условиями диагенез становится важной стадией осадочного породообразования.
В отличие от одностадийного закисно-доломито-джеспилитового литогенеза в
архее; в протерозое осадкообразование приняло форму двустадийного окисно-закисного
доломито-джеспилитового. На следующем этапе оно превратилось в двухстадийное
углисто-карбонатно-галогенное. Отныне жизнь стала оказывать прямое влияние
на процессы литогенеза, а эволюция состава породообразующих остатков организмов
стала отражаться на изменениях химических свойств осадков, минеральных и
петрографических особенностях пород.
Работы Страхова — наиболее всесторонняя и хорошо обоснованная попытка создать
общую концепцию эволюции литогенеза и геохимических процессов в осадочной
оболочке Земли под влиянием исторических преобразований живого. Дальнейшее
обоснование и развитие эта концепция получила в работах А. А. Саукова (1961),
С. И. Кузнецова и др. (1962), А. Б. Ронова (1964, 1972, 1980, 1983), В. П.
Петрова (1967), А. Б. Ронова и др. (1972), А. И. Анатольевой (1972, 1983),
Т. В. Аристовской (1972), А. П. Лисицына (1974, 1978), Н. Н. Верзилина (1977а,
19776), Н. В. Лопатина (1979), М. А. Жаркова (1981), В. И. Бгатова (1983),
А. И. Пака (1983) и др. В работах этих авторов приведен новый фактический
материал о воздействии органического мира на эволюцию континентального породообразования
и океанического осадконакопления.
Основные итоги этих исследований подведены в книгах: А. Б. Ронов „Осадочные
оболочки Земли” (1980), А. В. Лапо „Следы былых биосфер” (1987) и А. Л. Яншин
„Эволюция геологических процессов в истории Земли” (1988), а также в сборниках:
„Эволюция литогенеза в истории Земли” (1981), „Проблемы эволюции геологических
процессов” (1981) и „Биологические аспекты формирования осадочных пород и
руд” (1983). Анализ итогов исследований советских геологов о биогенном происхождении
осадочных пород Земли привел А. В. Лапо (1982, 1987) к выводу, что горные
породы, составляющие метабиосферу, своим происхождением так или иначе связаны
с жизнедеятельностью организмов. Некоторые из них (карбонаты, кремнистые
породы, каустобиолиты и частично фосфатные породы) образованы из остатков
организмов. Другие породы (железистые и марганцевые породы и аллиты) формируются
под влиянием факторов, преобразованных их жизнедеятельностью. Наконец, для
солей, обломочных и глинистых пород роль живого вещества сводилась преимущественно
к подготовке исходного вещества на стадии гипергенеза.
Вместе с тем в ряде работ, сыгравших важную роль в разработке проблемы эволюции
осадочного породообразования и рудообразования
137
(Яншин, 1977, 1988; Яншин и др., 1977; Боголепов, Жарков, 1981), было показано,
что хотя наиболее яркие примеры этой эволюции связаны с развитием жизни,
качественные изменения в вещественном составе пород могут происходить и без
заметного участия организмов. Такие изменения проявляются и в тех формациях,
где биогенный материал не был основным породообразующим фактором.
Начиная со второй половины XX в. в нашей стране развертываются систематические
исследования органического вещества в ископаемых осадках. Здесь особо важно
отметить работы советских ученых о влиянии эволюции растений и микроорганизмов
на характер и тип накопления горючих ископаемых. Задача установить связь
между качественными и количественными составами флор и специфическими особенностями
каустобиолитов почти одновременно была сформулирована в 30-х годах в работах
К. Бойрлена, М. Д. Залесского, Г. Потонье, В. О. Таусона. Особенно плодотворной
оказалась мысль В. О. Таусона (1936, 1941) о необходимости учитывать эволюцию
гетеротрофных организмов при обсуждении генезиса „всяких органогенных пород”
(1936, с. 279). Задача выяснить зависимость особенностей угленакопления от
эволюции растений и микроорганизмов успешно решалась в трудах А. И. Гинзбург,
И. И. Горского, А. Н. Гусевой, Ю. А. Жемчужникова, А. А. Карцева, Н. С. Коржневской,
И. Е. Лейфмана, В. В. Мокринского, Д. В. Наливкина, Л. Н. Ржанниковой, П.
И. Степанова, С. Г. Неручева и др. Было показано, что состав углей существенно
менялся от девона до наших дней. Например, нижнекарбоновые угли стеблевого
происхождения сменились углями, образованными из стволов. В мезозое создавались
уникальные угли, состоящие из спресованных листьев хвойных, птеридоспермовых,
гинкго и т. д. Для кайнозоя характерны лигниты.
Благодаря трудам Н. Б. Вассоевича (1984) и его школы были не только рассеяны
сомнения в происхождении нефти из биогенного органического вещества, но и
сформулирована задача исследовать связи эволюции нефтеобразования с эволюцией
живого вещества. Как было показано Н. В. Лопатиным (1979), современные данные
о „филогенетическом развертывании” горючих ископаемых явно свидетельствуют
о тесной зависимости эволюции типов угленакопления и образования нефтей фанерозоя
от исторических преобразований растительного мира. Лопатин предложил выделить
ряд этапов в эволюции биосферы, каждый из которых связан с крупными ароморфными
преобразованиями в органическом мире и возникающими под их влиянием изменениями
в образовании углеродистых горючих ископаемых.
Большое влияние на проведение исследований ископаемого органического вещества
оказала книга В. А. Успенского „Баланс углерода в биосфере в связи с вопросами
о распространении углерода в земной коре” (1956) и серия работ А. В. Сидоренко
об органическом веществе в докембрийских осадочно-метаморфических породах
(Сидоренко, 1976а, 1976б, 1984; А. и Св. Сидоренко, 1971, 1975). Установлено,
что органическое вещество присуще за редким исключением всем современным
и ископаемым осадкам. Оно играет исключительно важную роль во всех процессах
литогенеза, начиная с гипергенеза, седиментогенеза,
138
диагенеза и кончая катагенезом. Его влияние сказывается в образовании многих
сингенетических и эпигенетических рудных месторождений. В концентрированном
виде органическое вещество создает угли, горючие сланцы, а жидкие и газообразные
продукты его фоссилизации лежат в основе образования нефти и углеводородных
газов.
Сопоставление данных о периодичности накопления высокоуглеродистых формаций
в докембрийском осадконакоплении с основными этапами развития живого в докембрии
привело ряд авторов к выводу, что „все крупные эпохи повышенного накопления
в осадках органического вещества не просто отвечают... возрастанию биомассы
в эти эпохи, но... являются следствием принципиальных «биологических обновлений»
живых организмов” (Сидоренко и др., 1978, с. 925). Среди важнейших событий
органической эволюции, обусловивших крупные эпохи осадкообразования, регионального
метаморфизма и магматической активности, называют возникновение фотосинтеза
у прокариот (3.7 – 3.5 млрд лет тому назад), появление эукариот (1.9 – 1.6
млрд лет тому назад), массовое развитие колониальных форм (1.6 – 1.3 млрд
лет), возникновение митоза, мейоза, метафитов и метазоев (1.0 – 0.9 млрд
лет), экспансия многоклеточных (0.6 – 0.5 млрд лет) (Сидоренко и др., 1978;
Св. Сидоренко, Теняков, 1978). Другие же авторы считают, что нет однозначного
решения вопроса о причинах периодических глобальных поступлений в литосферу
органического вещества (Соколов, Барсков, 1988). По их мнению, эпохи угленакопления,
образования горючих сланцев, биогенного карбонатообразования и т. д. могли
быть связаны с резким увеличением биомассы некоторых групп организмов, с
утилизацией которой не справлялись редуценты, с распадом ранее существовавшей
трофической пирамиды, с особыми условиями захоронения и т. д.
Открытие П. Эйбельсоном (Abelson, 1954, 1963) аминокислот в остатках ископаемых
организмов и некоторых органогенных пород стимулировало новые направления
в изучении влияний химического состава организмов и их остатков на эволюцию
литосферы. Изучалась геохимическая роль отдельных компонентов органического
вещества (аминокислот, углеводов, лигнина, липидов). В нашей стране это направление
представлено работами С. М. Майской и ее сотрудников Т. В.Дроздовой, В. Н.
Генераловой, Л. А. Кодиной.
Мы смогли упомянуть только о некоторых работах советских геологов и геохимиков,
связанных с проблемой эволюции биогенного осадкообразования. Число этих исследований
поистине огромно. Следует заметить, что не все из упомянутых теоретических
положений получили всеобщее признание. Иногда подвергается сомнению мысль
о решающем влиянии жизни на процессы изменения даже таких пород, как карбонатные
и кремнистые. В других же работах, хотя и признается подобное влияние, ему
отводится второстепенное место в процессах литогенеза по сравнению с различными
тектоническими явлениями. Мы не намерены, да и не имеем права вмешиваться
в споры геологов, какие же факторы и в какой степени воздействовали на эволюцию
процессов выветривания, осадкообразования и рудогенеза.
139
Но, несмотря на дискуссионность многих проблем, связанных с изучением влияния
живого вещества на эволюцию литогенеза, а следовательно, и развитие верхних
слоев литосферы, мы полагаем, что уже сейчас достаточно твердо установлено
следующее. Во-первых, живые организмы и продукты их жизнедеятельности с возникновения
биосферы активно участвовали во всех процессах гипергенеза и литогенеза,
нередко изменяя ход и характер осадконакопления и породообразования. Во-вторых,
преобразующее влияние живого вещества на ход геологических процессов неуклонно
возрастало в ходе развития биосферы. В-третьих, эволюция угленакопления,
образования карбонатных и кремнистых пород оказалась наиболее тесно связанной
с историческими преобразованиями органического мира.
Важно подчеркнуть также, что теория эволюции литогенетических процессов Н.
М. Страхова была первой попыткой в систематизированной форме отразить место
и роль эволюции жизни в преобразовании процессов литогенеза и рудообразования.
В этой теории были собраны важные аргументы в пользу теории биогенного осадкообразования.
И в работах других советских геологов были подробно проанализированы различные
типы горных пород с целью установить место и роль биологических агентов на
разных стадиях преобразования вещества в верхних частях земной коры и на
ее поверхности.
Судя по высказываниям некоторых зарубежных геологов и геохимиков, они лишь
недавно стали обращаться к вопросу о „коэволюции жизни и неорганического
мира”. В практику геологических исследований все шире входят работы, посвященные
геологическому влиянию жизни (Cloud,1976; Lowenstam, 1975, 1984; Monty, 1979).
Но, по мнению П. Вестброка, в настоящее время такие работы представляют лишь
теоретический каркас для сбора материала, который призван их подтвердить
или опровергнуть (Westbrock, 1983). До недавнего времени оставались наиболее
типичными работы, подобные книге Р. Гаррелса и Ф. Маккензи (Garrels, Mackenzie,
1971). Хотя книга названа „Эволюция осадочных пород”, в ней проводится мысль,
что геологические процессы в общем не менялись во времени, типы осадков были
аналогичны современным, а изменения состава древних осадочных пород были
вызваны не различиями в их исходном материале, а действием вторичных постседиментационных
процессов.
В дальнейшем потребуются новые факты о воздействии органической эволюции
на процессы гипергенеза, седиментации и диагенеза, об изменениях состава
горных пород в соответствии со сменой таксонов в биосфере. Необходимы также
реконструкции микробных палеоценозов с целью установить их участие в процессах
минералообразования. Все это позволит глубже понять закономерности биологической
детерминации осадочных пород литосферы.
140
5.4. Роль биотических агентов в эволюции почв
Среди биокосных компонентов биосферы особое место занимает почва, которая,
по определению В. А. Ковды (1973), выполняет функцию общепланетарного аккумулятора
и распределителя космической энергии в биосфере и является универсальным
экраном, удерживающим в биосфере биофильные элементы. Ее особая роль в биосфере
не раз отмечалась натуралистами XIX в., но основателями биологического направления
в генетическом почвоведении стали Ч. Дарвин и В. В. Докучаев (Гиляров, 1960,
1983б). Классический труд Дарвина (1881) о дождевых червях и их роли в образовании
почв вышел незадолго до работ Докучаева, который традиционно считается основателем
современной концепции почв как четвертого царства природы, в добавление к
трем ранее выделенным К. Линнеем – минеральному, растительному и животному.
Дарвин впервые дал количественную оценку роли биотических агентов в формировании
почв. Докучаев (1883) существенно углубил этот подход в учении о происхождении
русского чернозема.
Для развития идеи эволюции биосферы в почвоведении, видимо, небезразличным
был тот факт, что у истоков генетического почвоведения стояли создатель эволюционной
теории в биологии – Дарвин и учитель Вернадского – Докучаев. Докучаевское
учение оказало огромное влияние на развитие почвоведения во всех странах
(см. подробнее: Крупеников, 1981). Но поставленная им проблема эволюции почв
стала предметом исследования преимущественно отечественных ученых: Д. Г.
Виленский, В. Р. Вильяме, В. Р. Волобуев, Г. Н. Высоцкий, К. К. Гедройц,
К. Д. Глинка, В. А. Ковда, С. И. Коржинский, П. С. Коссович, С. П. Кравков,
И. М. Крашенинников, С. С. Неуструев, И. К. Пачоский, Б. Б. Полынов, Л. И.
Прасолов, Н. П. Ремезов, А. А. Роде, А. Н. Розанов, Н. М. Сибирцев, И. И.
Синягин, Г. И. Танфильев и др.
Особый интерес для нас представляет подчеркнуто биологический подход к эволюции
почв, предложенный одновременно в работах Пачоского и Вильямса. Наиболее
развернутое обоснование он получил в учении Вильямса о малом биологическом
круговороте как форме развития почв. Это учение в законченной форме было
опубликовано в 1926 г. Вильяме попытался синтезировать накопленные к тому
времени теоретические соображения о биотических агентах почвообразовательного
процесса и их роли в эволюции. Сущность почвообразовательного процесса, по
Вильямсу, заключалась в циклическом создании и разложении органических соединений.
Особое значение он придавал накоплению их в почвах, что изменяло режимы минерального
и водного питания растений и вызывало в итоге смену растительных формаций.
Вильямс подчеркивал, что почва должна обладать способностью удерживать и
концентрировать элементы зольного питания. Особое значение он придавал малому
биологическому круговороту зольной и азотной пищи растений, в который включены
процессы синтеза и деструкции органического вещества.
Вильямс рассматривал эволюцию почв как составную часть единого почвообразовательного
процесса, как проявление одного общего
141
динамического процесса воздействия элементов биосферы на окружающую среду.
В ходе этого процесса происходят взаимосвязанные и взаимообусловленные эволюционные
преобразования не только органического мира, но и рельефа, водных бассейнов,
климата и т. д.
Эволюция почв, по мнению Вильямса, подчинялась строгим закономерностям. Она
шла от тундровых почв через подзолистые, болотные ко все более зрелым черноземам,
а затем к сухостепным и, наконец, к пустынным солончакам. Однако подобная
схема могла быть применена лишь к почвообразовательному процессу, развертывавшемуся
на освобождавшихся от ледников территориях. Она по существу охватывала лишь
заключительные стадии почвообразовательного процесса, примерно 1 – 2 млн
лет. Основная же часть истории почв, насчитывающая 300 – 400 млн лет, оставалась
вне ее рамок. Кроме того, в ней не учитывалось влияние различных геологических
факторов на эволюцию почв, а между тем тектогенез, орогенез, вулканизм, эрозия,
денудация, трансгрессии, регрессии и т. д. нередко вмешивались в этот процесс,
уничтожая на громадных территориях почвы. При абсолютизации биотических факторов
эволюции почв по существу оставались в стороне такие важные факторы, как
глобальные изменения климата, рельефа, уровня грунтовых вод и т. д.
В ходе развернувшихся дискуссий вокруг работ Вильямса, в которых немалую
роль играли его фантастические рекомендации мер по повышению плодородия почв,
неоднозначную оценку получила и эта гипотеза. Одни авторы не только ее всецело
поддержали, но и старались далее развить и дополнить (Титов, 1934; Герасимов,
1951). Другие же, принимая исходные положения Вильямса, критиковали его за
упрощение реальных биогеоценотических взаимодействий, за стремление построить
общую схему эволюции почв, базируясь лишь на немногих относительно хорошо
известных ему примерах смен почв, за игнорирование роли абиотических факторов
и т. д. (Роде, 1947; Шилова, 1957, 1959; Сукачев, 1964б; Глазовская, 1972,
и др.). Попытки Вильямса и особенно его последователей максимально идеологизировать
и политизировать научные дискуссии привели к тому, что его труды наряду с
работами Лепешинской и Лысенко провозглашались краеугольным камнем некой
особой советской биологии.
Отметая философскую аргументацию Вильямса, В. Н. Сукачев (1964б) считал неверным
принимать биогеоценотические процессы как единые и однообразные, хотя каждый
из компонентов биогеоценоза составляет часть единого природного явления.
Каждый из этих компонентов, по мнению Сукачева, развивается по собственным
внутренним закономерностям и их нельзя свести к общим закономерностям эволюции
биогеоценоза. Эволюция биогеоценоза представляет собой, по справедливому
замечанию Сукачева, интегральный результат сложных и внутренне противоречивых
процессов развития таких компонентов биогеоценоза, как атмосфера, литосфера,
кора выветривания, фитоценоз, микробоценоз и т. д. В связи с этим представления
об едином почвообразовательном процессе Сукачев оценивал не только как неудачные,
но и как глубоко ошибочные по существу. В то же время П. Д. Ярошенко
142
(1961) использовал термин „почвообразовательный процесс”, рассматривая,
правда, последний как составную часть процесса эволюции биосферы.
Были предложены и другие, иногда принципиально отличные схемы эволюции почв
(Коссович, 1911; Ремезов, 1932; Полынов, 1933). Так, Коссович исходным пунктом
почвообразования считал пустынные, а не тундровые почвы, рассматривая последние
вместе с болотными почвами как заключительные стадии эволюции. Отсутствие
данных об ископаемых почвах и их связи с живущими в те эпохи организмами
стало причиной серьезных расхождений при выделении главных этапов и тенденций
в эволюции почв. Эти расхождения резко проявились во время дискуссий на Всесоюзной
почвенной конференции в 1933 г.
Мы не будем останавливаться на остальных точках зрения, высказанных в ходе
этой и последующих дискуссий. Сошлемся на мнение А. А. Роде, который в результате
анализа истории данной проблемы пришел к выводу, что „наши представления
об эволюции почв в большинстве случаев имеют лишь весьма общий характер.
Отдельные стадии этой эволюции, движущие факторы ее в большинстве случаев
представляются еще в той или иной мере гипотетическими и требующими проверки
с приложением более надежных, преимущественно экспериментальных методов”
(1947, с. 50). Аналогичные оценки можно встретить в работах Глазовской, Полынова,
Ковды. Вместе с тем важно подчеркнуть, что в концепции Вильямса было четко
сформулировано представление об эволюции органического мира как главной движущей
силе эволюции почв. Как отмечалось в работах ведущих отечественных почвоведов
и геохимиков (Полынов, 1956; Ковда, 1973; Перельман, 1973), основная идея
биологической концепции В. Р. Вильямса вместе с биогеохимическими идеями
В. И. Вернадского составляют основы современных знаний об эволюции почв.
Для разработки проблемы эволюции почв большое значение имело внедрение геохимических
идей Вернадского в практику почвоведения. Здесь особую роль сыграли разработанный
Б. Б. Шлыковым (1934) метод сопряженного анализа почв, грунтовых вод, зоны
гипергенеза и растительности и осуществленные им на основе этого метода классические
исследования начальных этапов почвообразовательного процесса под влиянием
литофильной растительности (Полынов, 1945). Было показано, что литофилы,
захватывая химические элементы пропорционально их содержанию в породе, обусловливают
изменение форм их миграции и накопления в коре выветривания, а также образование
вторичных минералов. В пользу этой гипотезы говорит тот факт, что современные
скалы в первую очередь заселяются бактериями и синезелеными водорослями,
обладающими способностью извлекать различные соединения литофильных элементов,
являющиеся, согласно Полынову, абсолютными органогенами. Вслед за бактериями
и цианофитами на скалах размещаются диатомеи и низшие грибы. Позже появляются
простейшие и лишайники, которые обладают способностью не только химически,
но и механически разрушать скальные породы. Наконец, мхи окончательно готовят
почву для заселения ее высшими растениями. Таким образом,
143
в формировании современных почв и биогеоценозов на скальных породах наблюдается
строгая последовательность. Низшие формы организмов готовят условия существования
для более высокоорганизованных растений. Это позволяло Полынову рассматривать
процесс скального почвообразования „как некоторую схему общей эволюции” (1956,
с. 443).
Эта работа Полынова в общих чертах описывает развитие биосферы при завоевании
растительностью суши. Наряду с исследованиями зольного состава лесной (Н.
П. Ремезов, С. В. Зонн), степной и галофитной растительности (В. А. Ковда,
Н. И. Базилевич) работы Полынова подтвердили, что биогеохимическое почвообразовательное
действие растительности осуществляется прежде всего через малый биотический
круговорот. При этом особая роль в эволюции почв принадлежит процессам разложения,
а не создания органических веществ, так как при разложении освобождается
громадное количество энергии в тепловой или химически работоспособной форме.
В настоящее время биогеохимические аспекты эволюции почв разрабатываются
в трудах В. А. Ковды, М. А. Глазовской, К. И. Лукашева и многих других. Тесная
взаимосвязь жизнедеятельности организмов и почвообразовательных процессов
была доказана в работах по почвенной микробиологии (Н. А. Красильников, Е.
Н. Мишустин, Б. В.Перфильев, Н. Г. Холодный) и почвенной зоологии (К. А.
Арнольди, М. С. Гиляров и др.). Были установлены корреляции типа почв с находящимися
в них зооценозами и микробоценозами и последовательность распада остатков
организмов под воздействием почвенных грибов, простейших, червей, насекомых,
их личинок и продуктов их жизнедеятельности. Воздействие корневых систем
растений, микроорганизмов и их ферментов на процессы разрушения и синтеза
биогенных почвенных соединений обсуждалось также в работах М. М. Кононова,
Я. В. Пейве, И. В. Тюрина и др.
В заключение остановимся кратко на представлениях об эволюции почв, изложенных
в фундаментальном труде В. А. Ковды „Основы учения о почвах” (1973). Историческое
развитие почв автор связывает с преобразованиями биотического круговорота
веществ, возникавшими в результате эволюции автотрофов и их экспансии на
поверхности Земли. Автор опирается как на палеоботанические данные об эволюции
растительного покрова, так и на факты почвенной геохимии о почвообразовательной
роли отдельных групп бактерий, грибов и растений. Ковда выделяет пять этапов
в эволюции почв: скальный почвообразовательный процесс, или первичное биогеохимическое
выветривание; первичный почвообразовательный процесс; лесное и болотное тропическое
почвообразование; возникновение луговых, черноземных почв; формирование современных
почв.
Начало первичного почвообразовательного периода Ковда связывает с выходом
на сушу цианофитов и некоторых групп водорослей. В этот период стал осуществляться
синтез вторичных материалов, а также формировались болотные почвы. Лишь с
широким расселением растительности кустарникового и древовидного типа (псилофиты,
хвощи, папоротники) возникает подлинно почвообразовательный процесс, хотя
144
и носящий довольно примитивный характер. Позднее с появлением археоптерисовой
флоры стали формироваться кислые каолинитовые, аллитные, бокситовые, гидроморфные,
обогащенные железом почвы. Развитый лесогумусовый перегноиный горизонт окончательно
сложился в эпоху доминирования антрокофитовой флоры. Лесная растительность
давала обильный материал для формирования мощной подстилки, значительного
количества перегноя, а микробо- и микоценозы обеспечивали его разложение.
В условиях доминирования голосеменных преимущественно формировались кислые
подзолистые почвы, желтоземы, красноземы, бокситы, а также латериты, торфы.
Наконец, с появлением покрытосеменных, и прежде всего трав, существенно ослаб
подзолообразующий процесс. Возникает дерновый процесс, ведущий к образованию
гумусированных луговых и черноземных почв. Складываются все основные типы
современного почвообразования: бокситовый, ферралитный, красноземный, желтоземный,
подзолообразование, буроземный, лесостепной (сероземный), черноземный, пустынно-солончаковый
и лугово-дерновый. Под антропогенным влиянием резко ускорился процесс смены
подзолообразования дерновым процессом. Предлагаемая Ковдой схема, по признанию
автора, является гипотезой.
Трудности в познании эволюции почв связаны прежде всего с тем, что маломощные
почвенные покровы геологического прошлого были уничтожены денудационными
процессами. Ископаемые почвы древних геологических эпох – явление довольно
редкое. Из доплейстоценовых почв, т. е. с возрастом приблизительно более
1 млн лет, сохранились только локальные участки. Еще хуже обстоит дело с
сохранением более древних почв.
И хотя мы еще весьма схематично представляем себе основные этапы становления
и эволюции этого важнейшего биокосного компонента в биосфере, необходимо
подчеркнуть, что отечественная школа генетического почвоведения, сложившаяся
в результате синтеза идей В. В. Докучаева и В. И. Вернадского, стала инициатором
в развертывании исследований о биотических агентах почвообразовательного
процесса и основных этапах его эволюции.
* * *
Даже краткий обзор основных результатов исследований, непосредственно относящийся к проблеме биогеохимических закономерностей эволюции, позволяет сделать вывод, что идея Вернадского о переходе эволюции органического мира (эволюции видов) в эволюцию биосферы в целом была подтверждена. Следы этого перехода запечатлены в газовом составе атмосферы, в химических свойствах и солевом составе гидросферы, в химическом и минералогическом составе литосферы, в структуре и текстуре образующих ее пород. Была доказана и биогенная природа многих факторов, издавна считающихся абиотическими (озоновый экран), но наличие которых необходимо для существования современной биосферы. Не случайно мысль о необходимости создания
145
новой науки – биогеологии, изучающей воздействие жизнедеятельности организмов
и их эволюции на развитие геологических процессов и на формирование в земной
коре месторождений полезных ископаемых, завоевывает все большее число сторонников
(Walther, 1936; Teihard de Charden, 1943; А. и Св. Сидоренко, 1971; А.Сидоренко
и др., 1978; Соколов, 1983). По точному выражению Соколова, стало очевидным
единство стратисферы Земли как результат развития былых биосфер.
Проведенные в СССР исследования геологов, географов, геохимиков и почвоведов
показали, что воздействие жизни на эволюцию биосферных компонентов было весьма
различно. Наибольшее влияние жизнь оказала на формирование почв и атмосферы.
Биогенное преобразование последней в свою очередь оказывало громадное воздействие
на эволюцию органического мира. Эволюционные преобразования гидросферы и
осадочных пород литосферы в меньшей степени зависели от жизни. Влияние жизни
сказалось лишь на установлении современного солевого состава и химических
свойств океанов, а также на эволюции процессов литогенеза.
В то же время необходимо учитывать, что в трактовке геохимической эволюции
поверхностных оболочек Земли до сегодняшнего дня переплетаются „факты различной
степени достоверности и различные предположения и гипотезы” (Ронов и др.,
1972, с. 88). Имеющихся данных явно недостаточно, для того чтобы разработать
единую, учитывающую достижения различных наук концепцию эволюции геохимического
строения биосферы и ее движущих сил. Предстоят еще длительные исследования
в плане поиска дополнительных доказательств этой эволюции и с целью выработки
методов изучения взаимосвязи эволюционных процессов на популяционно-видовом
и биосферном уровнях организации живого. Только после открытия таких методов
может быть успешно решена задача синтеза имеющихся данных о геохимической
эволюции биосферы с эволюционной теорией.
146
Глава 6
ПЕРВЫЕ ШАГИ К ИЗУЧЕНИЮ
ГЛАВНЫХ ТЕНДЕНЦИЙ
В ЭВОЛЮЦИИ БИОСФЕРЫ
Поиск интегральных характеристик эволюции биосферы является одним из важнейших, но, к сожалению, и наименее разработанных подходов к изучению исторических преобразований этой формы организации жизни. К числу параметров, характеризующих биосферу как целостную систему, следует отнести ее общую биомассу и биологическую продуктивность, энергетику, информационный „фонд” и, наконец, общепланетарный биотический круговорот. Как уже отмечалось, задача изучения этих параметров биосферы была поставлена еще в трудах В. И. Вернадского, но из-за ее громадной сложности и отсутствия надежных данных она еще очень далека от своего решения. Здесь по сути дела еще нет сложившихся направлений исследований, а существуют лишь отдельные попытки постановки и решения отдельных проблем. В данной главе мы постараемся изложить основные результаты подобных попыток. В заключительном разделе главы ставится вопрос о необходимости введения понятия „эволюционно-биосферная формация”, которое должно отражать существование крупных этапов в истории биосферы. При этом имеется в виду, что каждая эволюционно-биосферная формация характеризовалась особыми структурно-функциональными и эволюционными закономерностями.
6.1. Эволюция общей биомассы биосферы
и ее биологической продуктивности
Напомним, что позиция самого Вернадского в вопросе об изменении в ходе геологической
истории общей биомассы (общего количества живого вещества) биосферы была
весьма противоречива. С одной стороны, он считал, что общее количество живого
вещества оставалось примерно одинаковым в течение всего геологического времени.
С другой стороны, его многочисленные высказывания о непрерывной экспансии
жизни на поверхности Земли, об образовании все более сложных и продуктивных
сообществ, об усложнении потоков вещества в биосфере и т. д. свидетельствуют
о том, что он допускал возможность увеличения биомассы в ходе эволюции биосферы.
Подобная противоречивость в оценке биомассы прошлых и современных биосфер
до сих пор сохранилась в литературе. Вывод Вернадского о постоянстве биомассы,
как правило, не находил поддержки
147
у биологов и палеонтологов. Исключение составляют, пожалуй, только взгляды
А. Мейер-Абиха, который считал, что „правило Вернадского” о постоянстве биомассы
биосферы должно стать основополагающим постулатом современных эволюционных
представлений (Meyer-Abich, 1964). Применяя это „правило” к анализу проблемы
вымирания организмов, он утверждал, что процессы вымирания предопределены
необходимостью сохранить общую биомассу живого вещества планеты в периоды
появления более высокоорганизованных групп животных и растений. Большинство
же биологов считало, что в ходе истории органического мира значительно возрастало
общее количество живого вещества. Об этом свидетельствовали нарастающее разнообразие
органического мира, резкое расширение областей, населенных организмами, и
завоевание все новых и новых экологических зон и нарастающая дифференциация
экологических ниш. Так, Л. Ш. Давиташвили писал: „Биомасса всего органического
мира с течением времени возрастала, потому что биос захватывал все новые
и новые области, необитаемые пространства становились обитаемыми, заселялись
разнообразными организмами” (1971, с. 42).
В работах геологов, биогеохимиков и философов встречаются скептические высказывания
о возможностях решить этот вопрос однозначно (Виноградов, 1975; Водопьянов,
1981; Лапо, 1982). А. Г. Вологдин утверждал: „В настоящее время нет точных
данных об изменениях количества живого вещества во времени, и поэтому решение
этого вопроса пока оказывается невозможным” (1976, с. 91 – 92). Тем не менее
нередки утверждения о неизменности общего количества биомассы биосферы в
ходе геологического времени. Например, ссылаясь на отдельные высказывания
Вернадского и на результаты собственных исследований по истории геохимических
циклов серы, В. И. Виноградов пришел к выводу, что ни видовой состав биосферы,
ни ее пространственное строение не определяют количество живого вещества.
Биомасса биосферы, по мнению автора, всецело обусловлена „наличием тех элементов
и определенных условий в пределах земной коры, которые необходимы для построения
органического вещества” (1975, с. 57). Он полагает, что уже первичные организмы
в кратчайший (в геологическом масштабе времени) срок сумели использовать
все потенциальные возможности для увеличения общей биомассы биосферы. „В
дальнейшем эволюция живого шла в пределах выдержанной примерно на одном и
том же уровне биомассы” (там же, с. 57). По мнению П. А. Водопьянова, „устойчивость
биосферы есть результат постоянства ее общей массы, массы живого вещества,
энергии, связанной с живым веществом и среднего химического состава всего
живого” (1981, с. 188).
Как видно из приведенных высказываний, в пользу и той и другой точек зрения
существуют веские теоретические аргументы. Но решение этой проблемы, по-видимому,
зависит не столько от них, сколько от объективных возможностей более или
менее точно оценить количество биомассы в современной биосфере и в биосферах
прошлого. Один из таких методов был предложен В. А. Успенским в книге „Баланс
углерода в биосфере в связи с вопросом о распространении углерода в земной
148
коре” (1956). Исходя из постоянства коэффициента фоссилизации органического
углерода (0.8 %) и данных об его содержании в осадочных породах, Успенский
попытался определить суммарную биомассу биосферы за все время ее существования,
взятое условно за 600 млн лет. Даже при столь заниженном возрасте биосферы
оказалось, что годовая продукция органического углерода в современной биосфере
превышает среднюю годовую продукцию в биосферах геологического прошлого в
29.5 раз. Полученный результат, по мнению автора, дает однозначный ответ
о возрастании общей биомассы в историческом развитии биосферы и хорошо согласуется
с данными о среднем содержании углерода в осадочных породах палеозоя, мезозоя
и кайнозоя, которые показывают „постепенное возрастание органического углерода
в суммарной биомассе фоссилизирующего углерода” (там же, с. 85). Так, среднее
содержание органического углерода в палеозойских отложениях 0.62 %, в мезозойских
– 0.68 % и в кайнозойских – 1.32 %. Автор утверждает, что содержание органического
углерода буквально тает по мере увеличения катагенетической изменчивости
горных пород.1
Оценивая возрастание содержания углеродистых соединений в породах вверх по
стратиграфическому разрезу, А. Б. Ронов (1964) пришел к выводу, что это объясняется
прежде всего неуклонным увеличением биомассы, биологической активностью растений
и усилением перехода их остатков в ископаемое состояние, а также потерями
летучих соединений древними отложениями. Этот вывод был поддержан Н. М. Страховым
(1971) и Н. Б. Вассоевичем (1973). Предпринимались попытки как-то количественно
оценить данный процесс. Так, например, Н. М. Страхов считал, что переломным
моментом в возрастании общей биомассы стало завоевание жизнью суши. „Завоевание
континентов, - писал он, - сопровождалось резким возрастанием общей биомассы
живого вещества: она по крайней мере удваивается. В океане также продолжается
рост биомассы и расползание живой материи по дну – бентос захватывает новые
участки глубокого дна, продвигается в пелагическом направлении и планктон,
что несомненно также увеличивает биомассу морских организмов” (1971, с. 549).
Подобные расчеты базировались на ранее существовавших представлениях о том,
что общие биомассы морских и наземных биологических сообществ примерно равны.
Сейчас же, однако, установлено, что живое вещество океана в 700 – 1000 раз
меньше живого вещества суши (Суетова, 1976), и, следовательно, выход растений
на сушу означал более значительные увеличения биомассы, чем это допускалось
в расчетах В. А. Успенского и Н. М. Страхова.
В последние десятилетия были опубликованы работы А. В. и Св. А. Сидоренко
(1971, 1975), В. М. Швеца (1973), Н. Б. Вассоевича (Вассоевич и др., 1976),
в которых приводились данные о содержании ископаемого
1 Этот вывод подтверждается данными Ф. Викмана, К. Марса, К. Ранкамы и А. В. Сидоренко о содержании рассеянного органического вещества в докембрийских осадочных породах. Согласно Св. А. и А. В. Сидоренко (1975), его среднее содержание в отложениях докембрия равняется 0.2 – 0.3 %.
149
органического вещества в породах различных геологических фаций. На их основе
предпринимались попытки оценить увеличение первичной продуктивности. Так,
например, С. Г. Неручев (1979) считает, что в фанерозое количество накапливаемого
органического вещества возросло от 0.004 · 1015 т С орг/млн
лет в кембрии до 0.03 · 1015 т С орг/млн лет в неогене, т. е.
в 7 раз. С учетом
более высоких
потерь органического углерода в древних осадках он полагает, что это означает
увеличение биопродуктивности биосферы за этот период в 3.5 – 7 раз.
Однако только по этим данным нельзя сделать вывод о количественных показателях
изменений биомассы, так как скорость и характер фоссилизации биогенного вещества
также изменялись в ходе геологического времени. Фактически мы не знаем, какая
часть отмершей биомассы переходит в ископаемое состояние. Если принять вывод
В. О. Таусона (1947) о непрерывном возрастании интенсивности и объема деструктивной
деятельности микроорганизмов, тогда неизбежно признание, что в ходе прогрессивной
эволюции биосферы все большая часть ее биомассы поступала вновь в биотический
круговорот вскоре после захоронения. К аналогичному выводу приходит А. П.
Хохряков (1981), который считает, что в эволюционном ряду (хвойные леса –
лиственные леса – травянистые сообщества) шло уменьшение процента органики,
поступающей в захоронение. Фитоценозы, возникшие на последних этапах эволюции
биосферы, отличаются все более замкнутым биотическим круговоротом, почвами
со все большим запасом быстро мигрирующей органики и со все меньшим запасом
органических веществ, выключенных из биотического круговорота. В таком случае
данные о содержании ископаемого органического вещества в горных породах немного
говорят о количестве общей биомассы биосферы в тот или иной период ее истории.
Данные геологии и геохимии о содержании органического вещества в литосфере
должны дополняться результатами сравнительных исследований биомассы в различных
современных экосистемах, каждая из которых может быть рассмотрена как модельная
система какого-либо этапа в эволюции биосферы. Особенно перспективными оказываются
сравнительные исследования биомассы и биотической продуктивности океана и
суши, так как долгое время область распространения жизни, по-видимому, была
ограничена гидросферой, где сложились все царства органического мира и подавляющее
большинство типов и классов животных (Зенкевич, 1971). По мнению многих авторов,
только в фанерозое содержание кислорода в атмосфере достигло такого уровня,
который защищал бы органические формы на континентах от губительного воздействия
ультрафиолетовых излучений (Berkner, Marshall, 1964).
Следует заметить, что подобные сравнительные исследования стали возможны
лишь в последние десятилетия благодаря значительному прогрессу знаний о биомассе
растений и животных в отдельных экосистемах. К числу наиболее надежных и
обоснованных сводок о распределении живого вещества в биосфере относятся
и работы советских биологов (Винберг, 1960, 1983; Кобленц-Мишке и др., 1968;
Богоров, 1969; Базилевичи др., 1970).
150
В 1951 г. Е. Рабинович, базируясь на данных Дж. А. Райли о продуктивности
планктона, оценил среднюю годовую фиксацию органического углерода в Мировом
океане в 155 млрд т, но уже вскоре после работ Е. Штимана, Е. Йенсена и Г.
Г. Винберга сложилось убеждение, что эти показатели сильно завышены. В книге
„Первичная продукция водоемов”, опубликованной в 1960 г., Г. Г. Винберг пришел
к выводу, что наиболее вероятной величиной будет 35 – 70 млрд т в год. Уточнение
этих расчетов на основе сопоставления данных различных авторов позволило
О. И. Кобленц-Мишке с соавт. (1968) оценить валовую биологическую продуктивность
океана в 25 – 30 млрд т в год, а чистую продукцию — в 15 – 18 млрд т. В большинстве
же работ, опубликованных позднее, приведенные цифры колеблются в пределах
45 – 60 млрд т (см., например: Богоров, 1969, 1970; Whittaker, 1970; Винберг,
1983, и др.). При сравнении первичной продукции океана с первичной продукцией
континентов за год, которая, по оценке Н. И. Базилевич, Л. Е. Родина, Н.
Н. Розова (1970), равна 172.5 млрд т, становится очевидным, что продуктивность
биосферы, а следовательно, и количество вовлекаемых за год в биотический
круговорот химических элементов увеличилось по крайней мере в три – четыре
раза. К началу 60-х годов большинство ученых было согласно с тем, что валовая
продукция живого вещества континентов значительно превышает валовую продукцию
океана. Лишь отдельные исследователи продолжали настаивать на цифрах, близких
к оценкам Райли – (126 ± 82) · 109 т углерода в год. В.
Б. Цейтлин (1983) оценивал первичную продуктивность океана в 50 – 110 млрд
т в год,
а Э. А. Шушкина
с соавт. (1984) – даже 190 млрд т.
Советско-французская экспедиция, работавшая в 1983 г. по Международной программе
„Plancton rate processes in the oligotrophic oceans”, получила данные, которые,
возможно, вынудят вновь пересмотреть устоявшееся мнение о первичной продукции
океана (Крупаткина и др., 1985). Было показано, что первичная продуктивность
олиготрофных вод, оцениваемая недавно цифрой 50 – 150 мг углерода на 1 м2,
оказалась значительно заниженной. Вклад в первичную продуктивность этих областей
океана пикопланктона, который раньше не учитывался, составляет 30 – 70 %
первичной продукции вод в акватории тропических и субтропических широт. Сейчас
еще только начинаются исследования продуктивности пикопланктона. Но, как
отмечает Д. К. Крупаткина с соавт. (1985, с. 62), „если эти величины подтвердятся
в дальнейшем, то суммарная продукция Мирового океана повысится примерно на
порядок величин и составит около 187 õ 109 т углерода в
год”. И тогда вновь окажется, что она по крайней мере равна биопродуктивности
наземных
экосистем.
А это значит, что выход жизни на сушу привел к удвоению биологической продуктивности
биосферы. Этот вывод можно сделать лишь с учетом того, что оценки продуктивности
всей биосферы в целом носят очень приблизительный характер.
Более убедительно сравнение биомассы в экосистемах, различающихся по геологическому
возрасту. В океане, биологическая структура которого сложилась в конце палеозоя
(Зенкевич, 1948), биомасса равняется 3.42 млрд т сухой массы (Богоров, 1969,
1970). В более молодых
151
 Рис. 6. Соотношения масс продуцентов (I) и консументов с редуцентами (II)
в океане (1) и на континентах (2) (по данным Базилевич и др., 1971).
Рис. 6. Соотношения масс продуцентов (I) и консументов с редуцентами (II)
в океане (1) и на континентах (2) (по данным Базилевич и др., 1971).
наземных биогеоценозах биомасса составляет 1.8 · 1012 т
(Рябчиков, 1971), 2.42 · 1012 т (Базилевич и др.,
1970), 2.63 · 1012 т (Суетова, 1976)
или даже
3.1 · 1012 - 1.0 · 1013 т (Ковда,
1973). Иначе говоря, даже при столь значительных различиях в оценке биомассы
континентов, данной
названными авторами, ясно,
что „общая биомасса Мирового океана в общем балансе планеты по весу играет
ничтожную роль” (Базилевич, 1979, с. 56). Это является доказательством значительного
возрастания биомассы в ходе эволюции биосферы и увеличения количества вещества,
включенного в биотический круговорот.
Для понимания закономерностей в эволюции биомассы и биологической продуктивности
биосферы (сухое вещество, т/%) очень важны данные, суммированные Базилевич
с соавторами (1970,1971):
| Континенты: | |
| зеленые растения | 2.4 · 1012/99.2 |
| животные и микроорганизмы | 0.02 · 1012/0.8 |
| Итого: | 2.42 · 1012/100 |
| Океан: | |
| зеленые растения | 0.0002 · 1012/6.3 |
| животные и микроорганизмы | 0.003 · 1012/93.7 |
| Итого: | 0.0032 · 1012/100 |
Как можно видеть, в ходе эволюции биосферы коренным образом менялось и соотношение
между биомассой продуцентов, с одной стороны, и консументов и редуцентов
— с другой. Если в океане, древнейшей части биосферы, биомасса продуцентов
составляет всего 1/16 часть от общей биомассы организмов океана, то в наземных
экосистемах соотношение совершенно другое (рис. 6). Здесь преобладает биомасса
растений, которая более чем в 99 раз превышает общую биомассу животных и
микроорганизмов. Это является одним из косвенных доказательств изменения
биомассы нашей планеты.
Точные количественные характеристики этих процессов являются задачами будущих
исследований, в которых, по-видимому, должны участвовать биологи и геологи
различных специальностей.
6.2. Изменения энергетики биосферы
Увеличение биомассы и накопление ископаемого органического вещества изменяют энергетику биосферы. По самым грубым подсчетам, только в месторождениях горючих ископаемых сконцентрировано более
152
1 · 1013 т органического вещества (Перельман, 1973),
а всего в осадочных породах - около 3.48 · 1015 т
(Вассоевич, 1976; Швец, 1973), 1.5 · 1016 т (Базилевич,
1979). Энергия, накопленная в осадках, равняется примерно 1 · 1025
Дж (Вольскис, 1982). Эти цифры позволяют представить, какое колоссальное
количество энергии
накопилось в земной коре и в почве за всю историю биосферы. Кроме того, значительная
часть энергии, аккумулированная ранее живым веществом, теперь представлена
в форме поверхностной энергии дисперсных частиц в зоне выветривания, а также
расходуется во многих геологических процессах (Полынов, 1956; Синицын, 1972;
Тимофеев, Щербаков, 1983). Живое вещество, участвуя в раздроблении горных
пород на тонко дисперсные коллоидные вещества, передает им дополнительную
энергию. И чем больше суммарная поверхность вещества, тем выше такая энергия.
Наиболее высокой энергией отличаются глины, илы, ископаемые угли, чернозем,
которые обладают способностью сорбировать из окружающей среды различные соединения.
Следует напомнить, что энергетическая трактовка органической эволюции не
раз предлагалась еще во второй половине XIX в. в работах Ю. Майера, В. Томсона,
С. А. Подолинского, Л. Больцмана. Так, Больцман еще в 1886 г. отмечал, что
борьба за существование – это прежде всего борьба за энергию. Особая термодинамическая
роль органического мира в функционировании нашей планеты подчеркивалась К.
А. Тимирязевым и Н. А. Умовым. В лекции „О космической роли растений”, прочитанной
в 1903 г., Тимирязев (1904) рассматривал хлорофилл как посредник между рассеянной
солнечной энергией и энергией синтезируемых в растениях органических веществ.
Впоследствии эта энергия используется в самых различных процессах, протекающих
на поверхности нашей планеты. Умов (1916) считал даже целесообразным ввести
дополнительный закон в термодинамике, который бы отражал тот факт, что отбор
направлен на преодоление энтропии. Были предложены и такие понятия, как „эктропия”
и „негэнтропия” для обозначения общепланетарной функции живого вещества,
препятствующей возрастанию энтропии в биосфере.
В статье „Энергетика жизни”, оказавшей огромное влияние на развитие представлений
об энергетике живого вещества, В. Анри (1917) подчеркивал, что жизнь препятствует
превращению полезной энергии в теплоту и тем самым препятствует ее рассеиванию
в мировом пространстве. Он был уверен, что в ходе прогрессивной эволюции
в биосфере увеличивалось количество энергии, аккумулированной организмами.
Этому процессу, полагал Анри, нет предела, так как жизнь еще не скоро достигнет
предела в своем распространении в поверхностных оболочках Земли. Поэтому
количество свободной энергии в биосфере возрастает постоянно. Это было обусловлено
увеличением количества органического вещества, накапливаемого в осадочных
породах и месторождениях горючих ископаемых, а также в форме поверхностной
энергии дисперсных частиц.
Попытки использовать энергетический критерий для оценки основных тенденций
в эволюции биосферы были сделаны в работах многих
153
советских ученых. Одним из первых, кто попытался применить эти представления
для понимания биосферных процессов, был В. Л. Комаров (1921). Он считал,
что „смысл” органической эволюции заключается в выработке форм, замедляющих
энтропию солнечной энергии. По его мнению, все многообразие органических
форм есть не что иное, как своеобразный способ усложнения циклов трансформации
энергии на Земле и повышения энергетической эффективности живого, проявляющейся
в образовании „стойких систем энергии” (с. 45). „При этом задерживается,
обесценивается и удлиняется весь цикл превращений, испытываемых данным количеством
энергии в момент появления ее на Земле в виде солнечного луча до исчезновения
с Земли” (там же). Благодаря эволюции живого стала возможной концентрация
солнечной энергии на сотни миллионов лет. Аналогичные идеи высказывали в
30 – 40-е гг. Э. Бауэр, А. Е. Ферсман, Э. Шредингер, но они не получили тогда
должного развития.
Только в последние десятилетия в связи с возрастанием интереса к проблеме
эволюции биосферы они привлекают внимание все большего числа ученых различных
специальностей. Так, Г. Ф. Хильми подчеркивает: „С энергетической точки зрения
появление и развитие биосферы на Земле... следует рассматривать как возникновение
грандиозного процесса постоянного накопления запаса превратимой энергии в
поверхностной оболочке Земли и тем самым направленного процесса уменьшения
«производства» непревратимых форм энергии в земной природе” (1966, с. 271).
Это обеспечивается, по мнению Хильми, прежде всего выработкой у растительного
покрова адаптации к максимальному захвату солнечной энергии. На континентах
оно достигается путем увеличения фотосинтезирующей поверхности (образование
листовой мозаики, формирование непрерывного растительного покрова, его ярусной
структуры, а также дифференциация растений по сезонам вегетации).
В числе основных путей замедления энтропии в биосфере Е. М. Лавренко (1972)
называет морфологическое усложнение растений в сторону увеличения фотосинтезирующей
поверхности и формирования разнообразных жизненных форм, захват новых зон
жизни, возникновение гетеротрофных организмов, способных максимально использовать
аккумулированную автотрофами энергию. Качественное многообразие органического
мира, по мнению Лавренко, играет особо важную роль в устойчивости биосферы
и в выполнении ею „негэнтропийной роли”.
В связи с этим интересно привести некоторые цифры, характеризующие эффективность
использования солнечной энергии в биосфере в целом и в различных экосистемах.
В среднем в биосфере используется лишь 0.1 – 0.2% годовой величины солнечной
радиации (Дювиньо, Танг, 1973; Моисеев и др., 1980). Характерно, однако,
различие между эффективностью использования энергии в морских и наземных
экосистемах. Если на суше она колеблется в пределах 0.3 – 0.46 %, а в некоторых
оптимальных условиях может достигать даже 1 – 2% (Будыко, 1984), то эффективность
использования энергии фитопланктоном гораздо ниже. Согласно расчетам Богорова
(1970), только 0.04% солнечной энергии, достигающей поверхности океана, аккумулируется
в процессах фотосинтеза.
154
Таблица 5
Значение К (степени прогрессивности ландшафта)
для основных типов ландшафта
(по: А. И. Перельман, 1973)
Тип ландшафта K = lg П / lg Б |
Тип ландшафта K = lg П / lg Б |
I. Эволюционный ряд ландшафтов: голосеменные – покрытосеменные 1. Таежные ландшафты 0.54 – 0.55
3. Травяные ландшафты
II. Прочие типы современных ландшафтов 4. Тундра 0.59
|
6. Примитивные пустыни - такыры водорослевые 1.0 III. Вымершие ландшафты (предположительно) 7. Лесные ландшафты с папоротникообразными среднего палеозоя < 0.55 IV. Культурные ландшафты 9. Культурные ландшафты с высокой агротехникой на почвах > 1 (П>Б)
|
Для познания энергетических закономерностей эволюции биосферы перспективным
является сравнение энергетической эффективности экосистем различного эволюционного
возраста. Базируясь на уникальной сводке Л. Е. Родина и Н. И. Базилевич „Динамика
органического вещества и биологический круговорот зольных элементов и азота
в основных типах растительности”, А. И. Перельман (1973, с. 57) составил
таблицу отношений ежегодной продукции к биомассе, точнее, отношений их логарифмов
— коэффициент К в основных типах растительности (табл. 5).
В плане обсуждаемой нами проблемы особенно интересны данные о современных
ландшафтах, отражающие эволюцию сообществ в связи со сменой доминирующих
продуцентов (голосеменных деревьев покрытосеменными деревьями и далее вытеснение
их травянистыми покрытосеменными). Оказалось, что в таежных ландшафтах, возникших
в середине пермского периода, коэффициент равен 0.54 – 0.55, в ландшафтах
с покрытосеменными деревьями, возникших в середине мела, коэффициент колеблется
от 0.59 до 0.68 и, наконец, в ландшафтах с травянистыми покрытосеменными
(степи, саванны), возникших в неогене, он достигает 0.95. По мнению Перельмана,
возрастание ежегодной продукции на единицу биомассы следует рассматривать
как показатель прогрессивного развития ландшафта, выражающегося в повышении
„экономичности”
155
его функционирования. Экстраполяция установленных закономерностей на вымершие
формации (влажные леса папоротникообразных среднего палеозоя и псилофитовые
формации нижнего палеозоя) дали соответственно коэффициенты 0.4 и 0.3.
Значительно более высокие цифры приводят В. Ф. Левченко и Я. М. Старобогатов
(1986), которые считают, что эффективность наземных экосистем по использованию
поступающей солнечной энергии возросла за последние 540 млн лет примерно
в 100 раз.
Энергетическая эволюция биосферы тесно связана с возрастанием целесообразности
живых систем, которую можно измерить повышением таких показателей, как КПД
системы, „энергетическая доходность” и „степень элевации” по В. Францу (Franz,
1951). Она проявляется в росте общей экономичности обмена веществ и круговорота
биомассы, уменьшении непроизводительных потерь при большей работоспособности
структур, в более высокой их надежности при большей активности организмов,
нарастании тенденции к автоматизации (по Шмальгаузену), интенсификации и
к усилению гомеостатических процессов и т. д. Высокий консерватизм биоэнергетики
клетки у высших растений обусловлен, по-видимому, тем, что уже на начальных
стадиях развития жизни была выработана весьма совершенная структура внутриклеточных
энергетических процессов. Вместе с тем наличие тонких различий в структурных
и функциональных компонентах, связанных с производством энергии, позволяет
предположить, что некоторая дошлифовка процессов брожения, фосфорилирования
и фотофосфорилирования продолжалась и после того, как в целом было завершено
формирование этих биоэнергетических процессов (Брода, 1978). Однако до сих
пор еще трудно установить корреляцию этих различий в биоэнергетических процессах
с различиями в образе жизни отдельных групп организмов и их различной ролью
в энергетике всей биосферы.
Одним из наиболее общих и наглядных способов характеристики энергетической
эффективности биосферы, по-видимому, может стать критерий средней выживаемости
особи у доминирующих в биосфере таксонов. Этот критерий был одновременно
предложен К. М. Завадским (1958) и Дж. Худеем (Thoday, 1958). Если у низших
организмов, представителей древнейших биосфер, средняя выживаемость особи
очень мала и приближается к величинам 1/109 или 1/1010 из
числа родившихся, то у высших животных она достигает 10 – 30%, т. е. в среднем
каждая из десяти
или даже из трех-четырех родившихся особей принимает участие в воспроизведении
вида. Колоссальные количества особей в периоды пиков численности у низших
организмов оказываются нормальной избыточностью, страхующей вид от вымираний
в периоды депрессий. Как неоднократно подчеркивал Завадский, вероятность
выполнения видовой функции – воспроизведения – у каждой средней особи таких
организмов крайне мала. Существование любого вида обеспечивается в основном
статистическими механизмами надежности — высокой плодовитостью и огромной
численностью в экологически оптимальных условиях. Естественно, такой способ
воспроизведения видов был неэкономичным и неэффективным. Из миллионов народившихся
особей в среднем
156
только одна выполняет видовую функцию, а остальные идут в качестве „дани”
за несовершенство своей организации. Большое количество живого вещества затрачивается
при этом на преодоление факторов „сопротивления среды”. У высших организмов,
занимающих доминирующее положение в современных экосистемах, вид является
более экономичной системой. Ослабляются механизмы надежности вида, построенные
лишь на воспроизводстве громадного числа особей в популяциях. Возросшая надежность
особи и увеличение числа групповых адаптации, основанных на ней, позволили
резко сократить непроизводительные расходы биомассы. Популяции высших животных
затрачивают гораздо меньше энергии на поддержание стабильной средней численности,
чем это происходит в популяциях низших организмов.
Повышение средней выживаемости особи стало главной стратегией в прогрессивной
эволюции живого. В то же время повышалась энергетическая эффективность живого
вещества в биосфере.
Зависимость энергетики биосферы от адаптации организмов не раз отмечал в
своих последних работах С. С. Шварц (1973, 1976), в том числе и в посмертно
опубликованной книге „Экологические закономерности эволюции” (1980). Он особо
выделял исследования Н. И. Калабухова (1946), который впервые показал энергетическую
значимость всякой адаптации. В эволюции биосферы особо эффективным оказался
адаптациогенез, связанный с увеличением потребления энергии не популяцией,
а особью, активно преодолевающей экстремальные воздействия внешней среды.
Это обеспечивает захват новых экологических зон, интенсификацию энергетических
процессов в биосфере, повышение скорости миграции в ней элементов и т. д.
Появление гомойотермных животных Шварц оценивал как подлинную революцию в
энергетике биосферы, так как расходование ими громадного количества энергии
для поддержания физиологического оптимума потребовало дополнительных запасов
биомассы на низших уровнях трофических цепей. По предположениям Шварца в
древних ценозах биомасса растений в 4 – 5 раз превышала биомассу животных
и не менее 15% энергии, накопленной в низших звеньях пищевой цепи, переходило
в верхние звенья. В третичных же ценозах биомасса автотрофов уже в сотни
раз превышает биомассу консументов и лишь 2 – 3 % ее идет на построение их
новых органов и на размножение. Это служит еще одним доказательством связи
энергетической эволюции биосферы с общим повышением активности животных,
увеличением разнообразия их двигательных реакций и расширением ареала миграции
(Северцов, 1934; Зенкевич, 1944; Ливанов, 1955).
Здесь уместно напомнить, что повышение (интенсификацию) общей энергии жизнедеятельности
А. Н. Северцов рассматривал как главный критерий прогрессивной эволюции.
И хотя сам Северцов не дал точного определения термина „энергия жизнедеятельности»,
но из приведенных им примеров ароморфных преобразований (превращение трубчатого
сердца в двухкамерное, мешковидной печени в трубчатую сложную железу, увеличение
дыхательной поверхности, усовершенствование теплообмена, усиление окислительных
процессов и т. д.) ясно, что автор
157
обращал внимание на два момента: 1) „все эти изменения вызывали, само собой
разумеется, резкое повышение обмена веществ” (1934, с. 121); 2) они усиливали
двигательную активность животных (там же, с. 76). Активность организмов,
проявляясь в актах добычи пищи, поиска полового партнера, миграции, заботы
о потомстве и т. д., способствует появлению все новых и новых потоков энергии
в биосфере и общему повышению его интенсивности.
В работах М. И. Будыко (1975, 1977а, 1984 и др.) не раз подчеркивалось исключительное
значение учения Северцова об ароморфозах для понимания энергетических аспектов
эволюции биосферы. „Вся история эволюции организмов, — пишет Будыко, — связана
с глубокими изменениями в механизмах их энергоснабжения, причем у прогрессивных
организмов эффективность указанных механизмов постепенно возрастала” (1984,
с. 284). Особенно подробно он разбирает увеличение расходов энергии у наземных
позвоночных, обусловленных необходимостью преодолевать силы тяготения, переходом
от ходьбы к бегу и к летанию, общей тенденцией к увеличению массы организмов,
становлением эффективного механизма терморегуляции и т. д. Будыко предполагает,
что повышение энергии жизнедеятельности происходило и у растений при увеличении
продуктивности фотосинтеза, так как это „делало возможным усложнение структуры
растений и повышение массы их непродуктивных органов” (там же, с. 289).
Характерно, что общая продукция в пределах одного трофического уровня консументов
находится в обратной зависимости от интенсивности использования ассимилированной
энергии (Риклефс, 1979). У птиц и крупных млекопитающих эффективность использования
ассимилированной энергии меньше 1 %, а у мелких млекопитающих она достигает
6%, у насекомых колеблется в пределах 5 – 13% и, наконец, у некоторых водных
животных превышает 30%. Наименее активные пойкилотермные животные, обитающие
в воде, затрачивают значительную часть ассимилированной энергии на рост и
размножение, достигая тем самым максимума в эффективности использования энергии
на воспроизводство биомассы данной особи. Однако особи со столь высоким коэффициентом
использования ассимилированной энергии относятся к видам, отличающимся низким
коэффициентом выживаемости.
В ходе эволюции биосферы была выработана оптимальная организация, связанная
с особенностями использования ассимилированной энергии на различных уровнях
жизни (Шварц, 1976). В предложенной Шварцем схеме показано, что эффективность
ее использования падает по мере повышения уровня биологической интеграции.
Эта эффективность составляет (%): элементарные физиологические функции –
70 – 80; работа организма в целом и комплексные физиологические функции –
15 – 50; рост, размножение и развитие индивида – 1.5 – 15; размножение и
развитие популяций – 0.5 – 7; использование энергии сообществом фотосинтезирующих
растений – 0.1 – 2; использование энергии солнечного излучения высшими трофическими
звеньями – 0.01 – 1; использование солнечной энергии для продуцирования новых
тканей животных – 0.0002 – 0.05.
158

Станислав Семенович Шварц (1919 – 1976).
Высокая эффективность использования энергии на уровне элементарных физиологических
функций обеспечивает выживание организмов, а относительно высокий процент
использования энергии в процессах роста, размножения и развития обеспечивает
воспроизводство выживших особей. Шварц подчеркивал: „Что бы ни случилось
на верхних этажах природы, какие бы катаклизмы ни потрясли биосферу и составляющие
ее биогеоценозы, высшая эффективность использования энергии на уровне клеток
и тканей гарантирует жизнь организмам, которые и восстановят структуру жизни
на всех этажах ее проявления в форме, наиболее соответствующей новым условиям
среды” (1976, с. 65).
Таким образом, сложившаяся структура использования энергии на различных уровнях
организации живого является фактором, гарантирующим стабильность биосферы.
В ходе ее эволюции была создана такая структура жизни, при которой распределение
энергии по разным уровням и эффективность ее использования на каждом уровне
обеспечивают целостность и устойчивость биосферы.
159
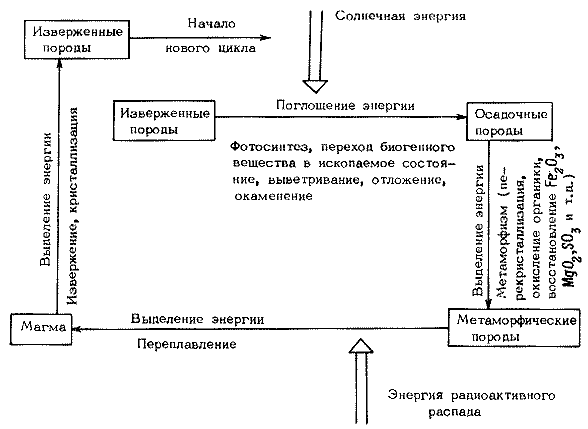
Рис. 7. Схема большого круговорота и взаимодействий солнечной энергии с глубинной энергией Земли (по: Белов, Лебедев, 1964 с изменениями).
Интересная гипотеза об эволюции энергетики Земли под влиянием эволюции живого
была предложена Н. В. Беловым и В. И. Лебедевым (1964). Согласно этой гипотезе,
аккумулированная солнечная энергия, погружаясь в глубинные слои Земли, становится
источником мощных тектонических и магматических процессов (рис. 7). Авторы
допускали, что минералы, заряженные энергией, образуются практически во всех
процессах, в которых участвуют или живое веществ, или его продукты. Среди
этих процессов они называли гипергенез, образование глин, кремниевых пород,
бокситов, известняков, солей, каустобиолитов и т. д. Погружаясь в зонах геосинклиналей
в недра Земли, минералы „разряжаются”. Освободившаяся энергия способствует
разогреванию земных недр и используется в процессах гранитизации, образования
месторождений рудных ископаемых, а также орогенетической и интрузивной деятельности
Земли (Лебедев, 1957). Механизм подобного запасания энергии минералами был
предложен в геоэнергегической гипотезе А. Е. Ферсмана (1934) и теории геохимических
аккумуляторов Н.В.Белова (1952).
Гипотеза Белова и Лебедева вызвала оживленную дискуссию среди геологов и
геохимиков. Опираясь на некоторые термодинамические представления и данные
кристаллографии, Д. С. Коржинский (1955) утверждал, что нет основания отводить
солнечной энергии какую-либо роль в глубинных процессах Земли. Богатые энергией
глинистые минералы могут образовываться не только биогенным путем (установленным
Шлыковым), но и в процессах абиогенных изменений полевых шпатов. Ссылаясь
на выполненные В. Г. Хлопиным и Г. В. Войткевичем расчеты об уменьшении радиоактивной
энергии, многие геологи и геохимики утверждали, что этой энергии вполне достаточно
для объяснения
160
всех геологических процессов. Поэтому апелляция к энергии, аккумулированной
жизнью, казалась им излишней.
Вместе с тем многие авторы поддержали гипотезу Белова и Лебедева. Выступившие
в ее защиту Н. Н. Верзилин, В. А.Ковда, А. С. Монин, А. И. Перельман, А.
А. Сауков, П. П. Тимофеев, А. В. Щербаков и другие полагали, что представления
о „большом круговороте”, связывающем эндогенные и экзогенные геологические
и геохимические процессы, позволяют лучше понять роль органической эволюции
в геологическом развитии Земли. Так, например, Н. Н. Верзилин (1977а, 19776)
считает, что возрастание интенсивности геологических процессов на протяжении
истории Земли явно свидетельствует о том, что их энергетическим источником
служит не столько радиоактивный распад, сколько эволюция органического мира.
Ускорение эволюции геологических процессов происходит параллельно с „усилением
интенсивности биологических круговоротов, скорости миграции элементов в биосфере,
скорости накопления в минеральном веществе потенциальной химической энергии”
(1977а, с. 12). В результате автор пришел к выводу, что „основной движущей
силой развития земной коры является жизнедеятельность живого вещества во
всем многообразии ее проявления” (там же). По мнению некоторых авторов, анализируемая
гипотеза Белова и Лебедева может быть использована для объяснения энергетических
источников и механизмов в процессах глобальной тектоники литосферных плит
(Монин, 1977).
Представления о большом геологическом круговороте вещества, который связывает
воедино процессы органической эволюции с тектогенезом и магматизмом и обеспечивается
солнечной энергией, аккумулированной организмами, в настоящее время находятся
на стадии гипотезы. Однако сама попытка связать процессы эволюции живого
с геологическими процессами осадкообразования, тектогенеза, орогенеза и магматизма
может оказаться перспективной для познания основных тенденций в эволюции
биосферы и прилегающих к ней других поверхностных оболочек Земли. „Возможно,
что в связях между поверхностными и магматическими процессами заключается
один из основных законов геологии, а эти процессы – лишь различные проявления
единого, грандиозного по длительности и сложности процесса развития земной
коры, преобразования вещества, энергии и информации” (Перельман,
1973, с. 50).
В заключение этого раздела надо отметить, что мы находимся только в начале
пути познания энергетических закономерностей эволюции биосферы. Как и 60
лет назад, еще нет достаточных данных для выделения основных тенденций в
энергетической эволюции биосферы как целостной системы (Lotka, 1925). По
сути дела приходится ограничиваться лишь констатацией положения о том, что
по мере прогрессивного развития жизни усиливается накопление в биосфере превращаемой
энергии и обогащение этой энергией других поверхностных оболочек Земли. В
результате постепенно возрастает негэнтропийная роль биосферы. Однако только
исследования энергетической роли отдельных групп организмов и отдельных видов
могут дать материал для понимания
161
энергетических закономерностей в эволюции биосферы, обусловленной появлением новых более прогрессивных форм животных и растений и формированием новых экосистем.
6.3. Изменение информационного „фонда” биосферы
Широкое использование понятия „информация” затронуло и учение о биосфере.
В работах, посвященных эволюции биосферы, не раз отмечалось, что в ходе истории
органического мира происходило и изменение ее информационной „емкости”. Об
изменении информации биосферы как одной из главных тенденций в ее эволюции
почти одновременно стали писать биологи-эволюционисты (Камшилов, 1970), почвоведы
и биогеохимики (Ковда, Тюрюканов, 1970), геохимики (Перельман, 1973).
Изменение информации биосферы выражалось в возрастании разнообразия и структурированности
биосферы. Происходило увеличение числа органических форм, геохимических барьеров,
нарастала дифференцированность физико-географического строения биосферы и
т. д. И хотя в отдельные периоды широкого распространения зон с теплым и
влажным климатом (карбон, мезозой) объем живого вещества был, возможно, и
больше, чем в современной биосфере, разнообразие современных физико-географических
условий (наличие аридных и гумидных зон, высокогорий и т. д.), в которых
обитают организмы, обусловливает большее разнообразие видов животных и растений
или, иными словами, большее количество биологической информации.
Один из первых попытался применить кибернетические представления об информации
к эволюции органических форм и к эволюции биосферы М. М. Камшилов в 1961
г. В книге „Значение взаимных отношений между организмами в эволюции” он
отмечал, что с точки зрения кибернетика „морфофизиологический прогресс отдельных
видов является способом увеличения запаса информации в результате установления
многочисленных связей с внешней средой и в особенности с биотическим окружением”
(с. 99). И далее он подчеркивал неотделимость эволюции отдельных видов от
прогрессивного развития всего живого, от „роста запаса его информированности”
(там же). Это обеспечивалось за счет усложнения и расширения взаимодействий
между живым и неживым веществом в биосфере, вовлечения в биосферные процессы
все новых и новых материально-энергетических ресурсов.
Впоследствии Камшилов не раз возвращался к вопросу об увеличении запаса информации
в биосфере (1970, 1974). Ссылаясь на отмеченные Н. Винером (1958) закономерности,
характеризующие процессы концентрации информации, он пришел к выводу о неизбежности
ускорения роста информации в биосфере по мере возрастания ее сложности. Для
количественной оценки роста информации в биосфере весьма показателен подсчет
числа водных и наземных организмов, сделанный Камшиловым (1974) на основе
данных Ф. Г. Добржанского (Dobzhansky, 1951) о видовом разнообразии органического
мира. Число наземных
162

Михаил Михайлович Камшилов (1910 – 1979)
животных составляет 93 %, а водных — только 7 % от общего числа видов в царстве животных. Сходные соотношения существуют и в царстве растений, где наземных видов – 92%, а водных – 8%. Их этих цифр ясно видно, что с освоением регионов биосферы, отличающихся разнообразием условий, возрастали и возможности видообразования. При этом лишь немногие формы оказались способными эволюировать в сложных условиях наземных экосистем. Примерно 10% из числа классов животных смогли приспособиться к наземной жизни, а остальные остались в гидросфере. Но именно эти 10% дивергировали в удивительное разнообразие видов, дав подавляющее большинство современных видов животных. То же самое и в царстве растений. Подобное преобладание наземных видов объясняется тем, что „эволюция жизни на суше пошла явно ускоренными темпами” (Камшилов, 1974, с. 76).
163
В книге В. А. Кордюма „Эволюция и биосфера” (1983) подчеркивается особое
значение информационного подхода в познании эволюции биосферы. Правда, Кордюм
исходит из совершенно иных, чем Камшилов, представлений. Он полагает, что
в биосфере постоянно находится громадное количество генетической информации,
которая благодаря различным способам внеполовой „горизонтальной” (трансформация,
трансдукция, симбиоз и т. д.) передачи постоянно обрушивается на организмы.
Это позволяет организмам быстро обрести новые признаки, на „приобретение
которых обычным путем ушли бы миллионы и даже миллиарды лет” (там же, с.
105). Предположив наличие широкого обмена генетической информации между столь
отдаленными таксонами, как бактерии и млекопитающие, Кордюм признает реальной
единицей эволюции не вид, а всю биосферу в целом. Он пишет: „Основным источником
информации в эволюции является привносимая извне информация. Она собирается,
перетасовывается, обновляется, перераспределяется и т. д. за счет всего генофонда
биосферы, претерпевает интенсивные изменения и внешней является только по
отношению к отдельным организмам (да и то до поры до времени). По отношению
же ко всей биосфере переносимая информация является неотъемлемой частью”
(с. 119).
Мы не будем останавливаться на этих более чем дискуссионных воззрениях автора,
так как их анализ выходит за пределы нашего исследования. Отметим только,
что наши знания о роли горизонтального переноса наследственной информации
у высших эукариот столь малы (Lewin, 1984; Хесин, 1984), что нет научных
оснований сейчас для гипотез об его исключительном значении в эволюции многоклеточных
организмов. Возвращаясь же к предмету нашего исследования, важно подчеркнуть,
что хотя автор и пишет об эволюции биосферы, но фактически у него никакой
новой информации на уровне биосферы не создается. Все преобразования по существу
сведены лишь к перекомбинированию уже существующей информации. Эволюируют
лишь отдельные таксоны, и то только за счет привносимой в их геномы уже готовой
информации. Откуда последняя берется, остается неизвестным. Невольно вспоминается
гипотеза Я. Лотси о перекомбинировании неизменных генов, как единственном
источнике новообразований. Гипотеза Кордюма вряд ли может быть признана информационной
теорией эволюции биосферы.
Информационный подход с позиций геохимии наиболее подробно рассмотрен в книге
А. И. Перельмана „Геохимия биосферы” (1973), который отмечал, что разделение
веществ в ходе истории биосферы означало рост количества неорганической информации.
Он проявлялся в прогрессивной дифференциации химических элементов, в образовании
месторождений полезных ископаемых, в возникновении геохимических барьеров,
в изменениях „кларков концентрации” отдельных химических элементов и т. д.
Перельман предлагает назвать увеличение неорганической и биологической информации
„законом прогрессивного развития биосферы” (с. 35), или законом В. И. Вернадского.
Информационный подход к эволюции биосферы позволяет выделить одну из основных
тенденций этого процесса — повышение ее надежности и устойчивости. Устойчивость
экосистем и их возможности
164
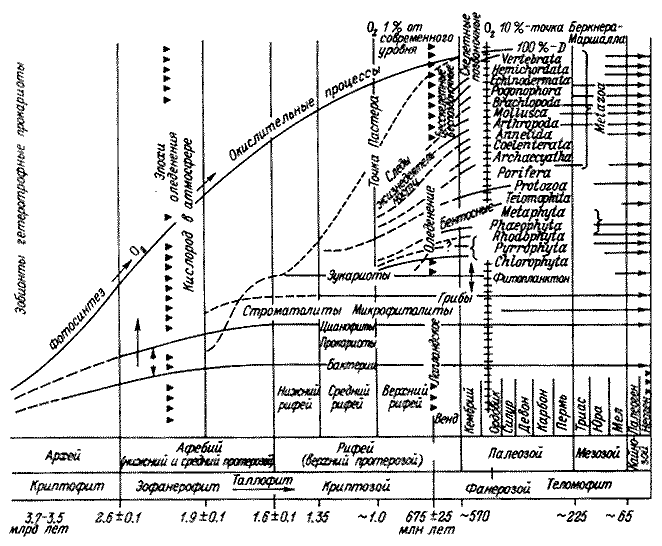
Рис. 8. Нарастающее разнообразие органического мира (по: Соколов, 1986).
приспособиться к изменяющимся абиотическим факторам в значительной степени
зависят от их сложности, т. е. от числа составляющих их видов животных, растений,
микроорганизмов и взаимодействий между ними. Это отмечал еще Ч. Дарвин. Органическая
эволюция в целом идет в сторону достижения наибольшей суммы жизни, которая
возможна лишь при наибольшем разнообразии видов в экосистемах (рис. 8). Возрастание
структурной сложности биосферы и усложнение взаимодействий между ее компонентами
вели к повышению ее целостности и независимости от космоса и от других геологических
оболочек Земли (Забелин, 1963; Сукачев, 1964а, 19646).
Повышение количества информации в экосистемах обусловливало не только количественные,
но и качественные преобразования биосферы. Повышение индекса таксономического
разнообразия обеспечивало устойчивость и надежность функционирования биосферы
и в то же время было источником ее дальнейшего развития в связи с усложнением
биогеоценотических связей.
Следует учитывать, что рост информации не был каким-то непрерывным и монотонным
процессом. Были периоды резкого сокращения биологической (Красилов, 1986),
а возможно, и неорганической информации. Часть информации постоянно переходила
в ископаемое состояние в виде однородных горных пород. Так, все многообразие
современных месторождений горючих ископаемых не может хоть в какой-то степени
165
отразить многообразие организмов, из которых образовались эти месторождения.
С точки зрения информационного подхода интересен тот факт, что рост разнообразия
органических форм на уровне семейств, родов и видов происходил только тогда,
когда был достигнут максимум разнообразия классов. Так, наибольшее число
классов в мире животных приходится на начало позднего палеозоя. В начале
мезозоя число классов сократилось и с тех пор придерживается на более или
менее одинаковом уровне. Последние геологические и палеоклиматологические
исследования заставляют внести определенные коррективы в традиционные представления
о нарастающей природной зональности Земли. Изучение основных типов метаморфических
пород докембрия, их структурных и текстурных особенностей показало, что уже
в раннем докембрии имелись различные климатические зоны (гумидные, аридные,
ледовые щиты и т.д.) (Сидоренко, 19766).
Информационный критерий позволяет выразить такие тенденции в эволюции биосферы,
как возрастание дифференциации ее живого, биокосного и косного вещества.
Однако он не отражает такие составляющие этой эволюции, как расширение зоны
распространения жизни, перестройка энергетики биосферы, преобразование биогеохимических
функций, усиление роли живого вещества в процессах выветривания, почвообразования,
осадкообразования, в преобразовании химических свойств атмосферы и гидросферы.
Интегральная характеристика всех этих процессов может быть выработана на
основе изучения исторических преобразований биотического круговорота веществ
— главного механизма биосферы, обеспечивающего его функционирование.
6.4. Эволюция биотического круговорота
Увеличение биомассы живого вещества, возрастание энерговооруженности биосферы
и ее информационной „емкости” по сути дела являются сторонами единого процесса
– эволюции биотического круговорота. Впервые эта мысль в самой общей форме
была высказана Вернадским. Он отмечал, что живое вещество, избирательно извлекая
из окружающей среды элементы и их изотопы, создает и сохраняет в охваченной
им области общепланетарный биотический круговорот (Вернадский, 1926а, 1927а).
Тем самым эволюция органического мира обусловливает развитие сложной общепланетарной
оболочки, включающей организмы и косное вещество, охваченное и организованное
деятельностью живого вещества.
Эти идеи Вернадского получили поддержку прежде всего в работах почвоведов,
географов и геохимиков (Вильямс, 1926, 1950, 1951; Герасимов, 1951; Шилова,
1959, 1960; Виноградов, 1959а, 19596, 1967; Перельман, 1961, 1973; Камшилов,
1961, 1970, 1974; Ковда, 1973; Лапо, 1982, 1987; Морозов, 1983, и др.).
Так, эволюцию биосферы В. Р. Вильяме понимал „как общий, грандиозный
по масштабу и продолжительности процесс, зародившийся в то время, когда
термодинамические и электрохимические условия
166
безжизненных процессов развития Земли определили появление жизни” (Вильяме,
1950, с. 51). По мнению Вильямса, „единственный способ придать количественно
ограниченному свойство бесконечного – это придать ему циклическое движение,
заставить его вращаться в круговороте” (там же, с. 50). Именно этим способом
и воспользовалась жизнь, выработав его в процессах длительного отбора и дошлифовки
биотических отношений. Уже развитие примитивных флоро-фаунистических комплексов
и начальная их дифференциация вели к возникновению особых биогенных циклов
в сложных круговоротах вещества и процессах превращения энергии. Эти биологические
циклы развивались на базе большого геологического цикла круговорота веществ,
наиболее ярко проявляющегося в круговороте воды и в циркуляции газов атмосферы.
Формирующиеся трофические отношения в зарождающейся биосфере способствовали
изыманию элементов из большого геологического круговорота и вовлечению их
в новый, меньший по сравнению с исходным биотический круговорот. Эти элементы
начали вращаться „расширяющейся спиралью” в биотическом круговороте. Сложившаяся
в биосфере система отношений продуцентов, консументов и редуцентов, отмечал
Вильямс, обеспечивает удержание зольных элементов в биотическом круговороте.
Постепенно биогенные циклы вещества и энергии приобрели общепланетарный характер,
используя при этом особенности геологического цикла.
Одним из первых, кто попытался применить идею эволюции биотического круговорота
для объяснения эволюции биосферы, был геоботаник И. А. Титов. В своей гипотезе
эволюции „георастительных сообществ”1 автор развивал мысль
о сопряженной эволюции абиотических и биотических компонентов биосферы. „Биотическая
эволюция
свойств среды жизни, — писал Титов, — в георастительных системах воздействует
на генетические превращения и отбор растительных организмов, симбиофизиологических
консорциев и растительных формаций, участвующих в развитии этих систем” (1934,
с. 147). Но сами эти превращения и отбор, подчеркивал Титов, определяют ход
биотической эволюции факторов среды. Новые конструктивные и физиологические
свойства организмов обусловливают способы использования в биотических круговоротах
элементов зольного и азотного питания. Поэтому в ходе эволюции биосферы „шел
отбор более совершенных, в равных условиях местообитаний, биологических трансформаторов
энергии, осуществляющих более совершенно в этих условиях круговорот материального
фактора-минимума, элементов зольного и азотного питания” (там же, с. 23).
Титов предпринял попытку объяснить и эволюцию активности животных с точки
зрения усовершенствования биотического круговорота. Он считал, что рост активности
способствовал, во-первых, расширению используемых животными территорий и
их пищевых ресурсов. Животные, расширяющие арсенал используемых в биосфере
ресурсов, тем самым способствуют интенсификации биотического круговорота.
1 Под георастительным сообществом Титов понимал целостную систему растительного сообщества и условий его развития в водосборном бассейне.
167
Конкретные выводы работ Титова были не лишены недостатков. Он абсолютизировал
роль сообществ водосборного бассейна в развитии растительного покрова Земли
и всей биосферы, оставляя фактически в стороне все другие структурно-функциональные
единицы биосферы. Однако теоретическое значение его работ бесспорно. Они
способствовали выработке методологических установок на необходимость синтеза
учения о биосфере, эволюционной теории и представлений о биотическом круговороте
с целью познания исторических преобразований на уровне биоценозов и биосферы.
Развивая представления Вильямса об эволюции биосферы как сопряженном процессе
преобразования растительного покрова, животного мира, атмосферы, почвы и
т. д., И. П. Герасимов показал, что в процессе приспособления растений к
условиям низкого естественного плодородия на поверхности Земли постепенно
складывался сплошной биогеоценотический покров. Появлялись все более сложные
по своей структуре и видовому составу биогеоценозы. Чрезвычайно усложнялась
пищевая цепь и повышалась интенсивность всех форм биотического круговорота.
В результате в „их орбиту стали вовлекаться все большие массы, все более
разнообразные по своему составу химические соединения, которые раньше на
более ранних этапах геологической истории свободно мигрировали, не захватываясь
живыми организмами в поверхностных и грунтовых водах, и достигали в конечном
счете океана” (1951, с. 14). Возрастающее влияние живого на окружающую среду,
расширение сферы существования жизни и вызванные этим изменения геохимических
соотношений между океаном и сушей в свою очередь оказывали воздействие на
дальнейшую эволюцию живого. Тем самым усложнение и усовершенствование биотического
круговорота представляет собой не только наиболее важный результат эволюции
биосферы, но и источник ее дальнейшего развития.
В связи с этим большое значение имеют различные попытки охарактеризовать
круговорот веществ на разных стадиях эволюции биосферы и очертить главные
тенденции в его эволюции. Эволюция круговорота веществ на начальных этапах
развития Земли обсуждалась в работах А. П. Виноградова (1959а, 1959б, 1967).
Он показал, что развитие поверхностной оболочки Земли в сторону наиболее
устойчивого динамического равновесия приводит к обмену веществ между отдельными
геосферами и внутри них. Вначале существенную роль играли процессы выноса
веществ из недр Земли в результате его дегазации и выплавления. Особенно
интенсивно этот обмен шел в архее, когда происходили крупные колебательные
движения земной коры, обширный орогенез, интенсивный вулканизм, гранитизация.
В этот период громадные количества магматического вещества и вулканических
газов поступали из глубинных слоев Земли на ее поверхность. В дальнейшем
зона обмена веществом между глубинными и поверхностными оболочками резко
сократилась, так как области геосинклиналей сузились. Важное значение приобрел
большой круговорот воды, благодаря которому происходила миграция элементов
и эволюция химического состава геосфер.
168
В. В. Ковальский (1963а), обсуждая вопрос о возникновении биосферы, высказал
предположение, что в протобиосфере существовала очаговая „оболочка”, составленная
из абиогенных органических и активных координационных соединений микроэлементов.
Эти соединения могли выполнять зачаточные биогеохимические функции (концентрационную,
окислительную, восстановительную), которые впоследствии получили свое максимальное
развитие в биосфере. Осуществление этих зачаточных функций, по мнению Ковальского,
и было зарождением биотического круговорота.
Для познания эволюции биогеохимического круговорота веществ большое значение
имеют исследования его особенностей в отдельных фанерозойских палеобиосферах
и в различных современных геохимических ландшафтах. Эти исследования проводились
в рамках геохимического ландшафтоведения, созданного трудами Б. Б. Полынова
(1934, 1953а), который синтезировал геохимические идеи Вернадского и Ферсмана
с учением Докучаева о зонах природы. Разработанный им метод сопряженного
геохимического анализа всех компонентов ландшафта активно использовался в
трудах его многочисленных учеников и последователей (Н. И. Базилевич, М.
А. Глазовская, В. В. Добровольский, К. И. Лукашев, А. И. Перельман и др.),
в которых изучались различные геохимические ландшафты. Эти исследования стали
основой для постановки вопроса об этапах и тенденциях в эволюции биогеохимического
круговорота в фанерозое, так как сохранилось немало ландшафтов, которые в
известном смысле могут рассматриваться как реликты прошлых биосфер: пустыни,
горные тундры, хвойные и тропические леса и т. д. (Перельман, 1961,1973;
Лукашев, Вадковская, 1973; Ковда, 1973). Таким образом, комплексное использование
историко-сравнительных и актуалистических приемов исследований биогеохимического
круговорота позволяет выделить некоторые тенденции в его эволюционных преобразованиях.
Выступая на Втором научном чтении имени А. Е. Ферсмана 21 декабря 1960 г.,
А. А. Сауков остановился на значении органической эволюции в коренных изменениях
геологических и геохимических процессов. Он показал, как в ходе постепенного
завоевания жизнью гидросферы, верхней части литосферы и нижней части тропосферы
жизнь оказывала все возрастающее воздействие на миграцию химических элементов.
И это воздействие увеличивалось в „соответствии как с увеличением массы живого
вещества, так и с качественными изменениями органического мира” (1961, с.
10). Сауков отмечал три главных события, оказавших влияние на эволюцию биотического
круговорота: появление фотосинтезирующих растений; выход растений на сушу;
появление человеческого общества. Так, например, появление наземной флоры
вело к накоплению органического вещества в почвах, в залежах торфа, угля
и т. д. Все это означало формирование неких „органических барьеров”, препятствующих
элементам мигрировать с суши в водоемы. Свою задачу Сауков ограничивал только
доказательством того, что „в ходе эволюции нашей планеты происходят количественные
и качественные изменения факторов миграции химических элементов” (там же,
с. 16). Он не поднимал вопрос
169
о главных тенденциях в этих изменениях. Такой вопрос был поставлен Е. И.
Шиловой (1960а, 1962), А. И. Перельманом (1961) и В. А. Ковдой (1973).
Е. И. Шилова попыталась охарактеризовать механизм обмена веществ и энергии
на различных этапах истории биосферы и связать его как-то с необратимыми
макроэволюционными преобразованиями организмов. Опираясь на некоторые хорошо
известные факты из области палеонтологии, эволюционной теории, почвоведения
и геохимии, Шилова осветила „важнейшие черты развития единого в своей биохимической
сущности, многообразного в формах проявления процесса обмена веществ в биосфере”
(19606, с. 43). Особое внимание при этом уделялось изменениям в круговороте,
связанным с увеличением содержания в атмосфере кислорода и уменьшением содержания
углекислого газа.
Наиболее развернутая характеристика эволюции геохимического круговорота была
дана А. И. Перельманом в книге „Геохимия ландшафта” (1961). Он выделил три
периода в эволюции геохимического ландшафта: абиогенный, биогенный и культурный.
Для обсуждаемой нами проблемы особое значение имеет второй период, разделенный
на ряд этапов (докембрий, ранний и поздний палеозой, мезозой и кайнозой).
Становление биогеохимического круговорота Перельман относит к периоду докембрийских
геохимических ландшафтов, когда жизнь „пробовала различные варианты биологического
круговорота, вовлекая в интенсивную миграцию различные элементы, пока в ходе
эволюции не возник наиболее целесообразный круговорот, свойственный современным
ландшафтам” (1961, с. 385). Среди главных тенденций в эволюции биотического
круговорота Перельман отмечал нарастающую его дифференциацию, связанную с
появлением все новых и новых циклов элементов и изменением их форм миграции,
ускорение темпа круговорота, усовершенствование способности к максимальному
использованию минерального вещества и воды, к повышению биотической продуктивности
ландшафтов. Эти процессы сопровождались нарастающим выщелачиванием и обеднением
ландшафтов элементами, необходимыми для минерального питания растений. Особенно
сильно это сказывалось в периоды спокойного тектонического развития Земли.
В эпоху же интенсивного орогенеза магматические процессы выносили в биосферу
глубинные породы, обогащенные фосфором, калием, кальцием, натрием и т. д.
Несмотря на направленность в эволюции биогеохимического круговорота, обусловленную
прежде всего прогрессивным развитием жизни, в ней наблюдается и некоторая
периодичность.
Высокая зависимость биотического круговорота от эволюции растительного покрова
была показана В. А. Ковдой (1956, 1973, 1985), который посвятил эволюции
биогеохимического круговорота специальный раздел в книге „Основы учения о
почвах”. Он считал, что возрастание числа видов с различными биогеохимическими
и экологическими функциями означало усиление биотического круговорота, вовлечение
в него все большего числа химических элементов, а также возрастание роли
живого вещества в процессах выветривания, осадкообразования, преобразования
атмосферы, гидросферы и литосферы. Вслед за Вильямсом
170
Ковда оценивает возникновение почвообразовательного процесса как важнейшее
событие в эволюции биосферы, подготовленное появлением на суше микроорганизмов
и растений. Постепенно почвенный покров превратился в важнейший компонент
биосферы, представляя собой область концентрации процессов разложения остатков
животных и растений, а следовательно, и освобождения громадного количества
энергии и накопления питательных веществ. Почва стала важнейшим аккумулятором
вещества и энергии, используемых в биотическом круговороте. Из составленной
Ковдой таблицы зольного состава растений (табл. 6) видно, что уже появление
водорослей на Земле расширило роль биотического круговорота минеральных элементов,
так как зольность водорослей в 3 – 4 раза выше, чем бактерий. Особенно возросла
доля натрия, серы, хлора в минеральных веществах, поглощаемых водорослями.
Наметилась тенденция к увеличению доли кремния и фосфора, получившая дальнейшее
развитие у наземных растений.
Ковда в целом считал, что „размеры биологического круговорота минеральных
веществ непрерывно увеличивались как вследствие увеличения числа видов и
массы живого вещества, так и вследствие общего возрастания зольности от хвойных
к лиственным древесным породам и от последних к травянистым” (1973, с. 125).
Ковда указывал на противоречивый характер эволюции биотического круговорота.
Увеличение в тканях растений содержания фосфора, серы, железа, магния, кальция
и особенно калия происходило с колебаниями и отклонениями. Содержание кремния,
достигнув максимума в хвощах, хвойных и злаках, в дальнейшем уменьшается.
Доля хлора в наземных растениях в целом постепенно снижалась.
Проводимые исследования распределения в горных породах содержания различных
элементов позволяют выяснить тенденции в эволюции круговорота веществ в масштабе
геологического времени. Как было показано А. В. Лапо (1982), изучение „следов
былых биосфер” не только является важнейшим способом познания эволюции всей
биосферы, но и ее биотического круговорота.
В целом же проблема эволюции биогеохимического круговорота разрабатывается
еще в самой общей форме, так как до сих пор нет необходимых палеогеографических
и палеобиогеохимических данных для более детального изучения этого процесса.
Изучение минерального и зольного состава современных организмов и исследование
особенностей биогеохимического круговорота в различных современных геохимических
ландшафтах по существу остаются единственными путями его познания. Аналогичная
картина наблюдается и в зарубежных геохимических исследованиях.
Мысль о том, что эволюция биосферы выражается прежде всего в исторических
преобразованиях биотического круговорота, получила распространение и среди
биологов-эволюционистов (Камшилов, 1961, 1970, 1974; Наумов, 1964; Тимофеев-Ресовский
и др., 1969; Шварц, 1973, 1976; Мазинг, 1970; Гиляров, 1973; Уголев, 1980,
1985, 1987; Голубец, 1982; Ситник, Голубец, 1983; Левченко, Старобогатов,
1986, и мн. др.).
171
Таблица 6
Зольный состав растений (в % к золе) (по: Ковда, 1973)
| Группа растений | Зола, % |
К |
Na |
Са |
Mg |
Fe |
Р |
S |
Si |
Cl |
| Бактерии (среднее из 10) | 7.3 |
14.7 |
0.6 |
6.0 |
4.8 |
0.6 |
1.0 |
1.2 |
0.6 |
— |
| Водоросли: | ||||||||||
|
25.3 |
5.0 |
12.2 |
23.1 |
1.7 |
0.5 |
1.5 |
8.3 |
2.6 |
10.3 |
|
27.8 |
18.2 |
12.3 |
8.6 |
3.2 |
0.7 |
1.2 |
6.3 |
1.0 |
20.0 |
|
20.0 |
11.0 |
14.8 |
7.0 |
4.2 |
0.6 |
1.6 |
12.7 |
0.7 |
6.7 |
| Грибы (среднее из 46) | 7.2 |
28.4 |
2.7 |
3.2 |
2.4 |
1.4 |
16.5 |
2.3 |
1.3 |
1.3 |
| Лишайники: | ||||||||||
| накипные (среднее из 3) | 8.7 |
0.98 |
0.5 |
77.6 |
2.1 |
0.9 |
0.09 |
0.8 |
8.2 |
— |
| листоватые (среднее из 1 9) |
4.5 |
10.0 |
1.9 |
16.0 |
2.8 |
3.5 |
2.6 |
4.0 |
6.7 |
— |
| кустистые (среднее из 90) | 2.6 |
9.3 |
3.5 |
12.0 |
2.4 |
3.0 |
2.3 |
2.9 |
16.8 |
0.4 |
| Мхи (среднее из 29) |
4.6 |
8.0 |
2.5 |
16.0 |
4.0 |
5.3 |
2.1 |
2.3 |
12.2 |
4.0 |
| Папоротники (среднее из 9) |
6.9 |
35.4 |
3.3 |
20.0 |
5.0 |
0.6 |
3.6 |
2.4 |
3.7 |
10.2 |
| Хвощи (среднее из 49) |
19.0 |
11.2 |
1.5 |
8.1 |
1.6 |
0.6 |
1.8 |
2.2 |
29.3 |
4.1 |
| Плауны (среднее из 4) | 5.1 |
13.7 |
0.9 |
4.3 |
2.4 |
0.8 |
1.7 |
1.6 |
6.3 |
1.4 |
172
| Голосеменные: | ||||||||||
|
3.8 |
15.4 |
— |
26.4 |
4.5 |
2.0 |
6.2 |
6.2 |
4.2 |
— |
|
4.5 |
6.5 |
— |
21.0 |
1.7 |
0.4 |
2.6 |
6.0 |
16.0 |
— |
| Однодольные: | ||||||||||
| водные (среднее из 12) | 16.3 |
15.5 |
5.1 |
20.5 |
5.2 |
4.2 |
3.5 |
2.3 |
4.0 |
3.4 |
| злаки (среднее из 260) | 6.6 |
23.0 |
3.1 |
4.4 |
1.9 |
2.1 |
2.1 |
2.4 |
19.0 |
6.1 |
| лилейные (среднее из 10) | 8.1 |
30.7 |
4.8 |
11.2 |
3.5 |
1.1 |
4.6 |
2.4 |
2.7 |
7.3 |
| Двудольные: | ||||||||||
|
9.5 |
25.0 |
5.0 |
20.0 |
7.5 |
1.4 |
2.6 |
1.9 |
1.5 |
2.9 |
|
20.5 |
12.4 |
19.5 |
7.3 |
3.8 |
0.9 |
1.6 |
5.0 |
2.4 |
14.5 |
|
9.6 |
23.0 |
7.7 |
17.0 |
2.3 |
1.5 |
4.0 |
4.0 |
3.0 |
7.9 |
|
7.9 |
27.0 |
3.4 |
18.0 |
3.4 |
1.0 |
4.7 |
1.7 |
5.1 |
4.1 |
|
13.0 |
28.4 |
— |
18.6 |
4.1 |
— |
3.1 |
2.0 |
— |
7.8 |
|
2.1 |
16.0 |
4.0 |
16.8 |
6.3 |
1.9 |
3.7 |
2.3 |
5.9 |
2.5 |
|
7.0 |
18.8 |
8.0 |
12.1 |
3.5 |
1.0 |
3.2 |
2.7 |
3.2 |
9.0 |
| Культурные растения: | ||||||||||
|
7.0 |
31.8 |
1.8 |
4.8 |
2.0 |
0.4 |
3.2 |
2.6 |
15.0 |
5.5 |
|
10.4 |
27.0 |
3.2 |
11.8 |
3.2 |
1.7 |
3.4 |
2.4 |
6.6 |
5.8 |
173
Одним из первых среди биологов к такому выводу пришел Н. П. Наумов (1964),
который считал, что всякое адаптивное преобразование на уровне организма
или вида должно оказывать влияние на геохимические и энергетические параметры
биотического круговорота.
Систематизированное развитие данный подход получил в серии работ М. М. Камшилова
„Значение взаимных отношений между организмами в эволюции” (1961), „Биотический
круговорот” (1970) и „Эволюция биосферы” (1974). В этих книгах автор попытался
дать синтез дарвинизма с идеями В. И. Вернадского. Уже в 1961 г. он отмечал,
что необходимо дальнейшее обоснование вывода Вернадского о расширении области
существования жизни и усилении давления живого вещества на окружающую среду.
Подобными примерами, по словам Камшилова, „буквально полна палеонтология”
(с. 109). Он пишет о расширении области обитания и источников питания у морских
ежей, об удивительно широкой адаптивной радиации моллюсков и насекомых в
палеозое и мезозое, а также гомойотермных животных в кайнозое. Возникновение
последних открыло исключительные возможности для распространения жизни на
Земле и создало предпосылки для сознательного регулирования обменом веществом
и энергией между органической и неорганической компонентами биосферы.
Камшилов подчеркивал, что даже организмы, вставшие на путь морфофизиологического
регресса (в смысле А. Н. Северцова), способствовали прогрессивной эволюции
биосферы. Эндопаразит повышает степень эффективности в использовании жизненных
ресурсов, побуждая хозяина поглощать больше пищи, чем это необходимо для
его собственного существования. Возникновение пещерной фауны расширяло область
существования жизни и усложняло взаимодействие между живым и неживым. Вымирающие
формы, как правило, заменялись более прогрессивными и разнообразными группами
организмов, в результате чего возрастала общая интенсивность биологических
процессов, усложнялись связи между организмами, расширялся арсенал материально-энергетических
источников их существования. Важнейшим фактором этой эволюции Камшилов считал
естественный отбор, который формирует все новые и новые группы организмов,
способствующих реализации главных тенденций в развитии биосферы.
Впоследствии Камшилов не раз возвращался к этим вопросам, уточняя высказанные
им ранее положения. Исходя из мысли Вернадского (1931) о появлении жизни
на Земле в форме комплекса организмов, он отмечал, что жизнь изначально существовала
в форме биотического круговорота, основанного на взаимодействии синтетиков
и деструкторов (Камшилов, 1970). Этот круговорот возник на базе абиотического
круговорота, в недрах которого стали формироваться циклы органического вещества
в виде синтеза и распада химических соединений углерода на первых этапах
биопоэза.
Автор намечал некоторые основные этапы в эволюции биотического круговорота
(1974): круговорот, осуществляемый прокариотными формами; возникновение одноклеточных
форм; усложнение циклической структуры жизни в результате надстройки трофических
отношений
174
Рис. 9. Стадии развития биосферы (по: Камшилов, 1974 с изменениями).
А –
большой абиотический круговорот, Б – биосфера, Ч - человечество.
из многоклеточных животных, растений и грибов; превращение биотического
круговорота в общепланетарный процесс в связи с завоеванием жизнью суши;
усложнение и расширение области действия биотического круговорота с завоеванием
доминирующего положения в биосфере гомойотермными животными; возникновение
человеческого общества и, наконец, переход к биотическому круговороту, сознательно
регулируемому действиями людей. В конечном счете человек выходит за пределы
круговорота Земли, проникая в космическое пространство.
Характерной чертой в эволюции биотического круговорота оказалось то, что
жизнь, извлекая из окружающей среды необходимые вещества и обогащая ее продуктами
жизнедеятельности и свободной энергией, неизбежно изменяет свои условия существования.
Поэтому она должна постоянно приспосабливаться ко все новым и новым созданным
ею же условиям существования. И вместе с ней неизбежно должен меняться и
биотический круговорот. „В силу самой природы жизни, - подчеркивал Камшилов,
- биотический круговорот не может быть полностью замкнутым, последующие витки
круговорота не совпадают с предыдущими. Биотический круговорот оказывается
спиралью – великой спиралью жизни” (1974, с. 54-55). В ходе развития биотического
круговорота многие вещества на долгие периоды геологического времени исключались
из биосферы и захоронялись в виде мощных месторождений руд и горючих ископаемых.
Развитие биосферы было связано с усложнением биотического круговорота. Новые
органические формы нередко могли существовать лишь на базе предшествующих.
В основе современного круговорота лежит круговорот продуцентов и редуцентов
из числа бактерий, низших грибов и других древних групп, над которыми надстроены
этажи из многоклеточных растений и животных, связанных трофическими, средообразующими,
конкурентными отношениями, а также отношениями взаимоблагоприятствования
и кооперации. Новые формы жизни, возникнув
175
на базе старых, нередко могут существовать лишь вместе с ними. Эволюция
биотического круговорота предстает прежде всего как прогрессирующая дифференциация
форм живого, где наряду со старыми формами постоянно возникают все новые
и новые. Это не только усложняет структуру биотического круговорота, но и
повышает надежность его функционирования. При этом постоянно ускоряется темп
эволюции в силу усложнения биотических взаимодействий, выступающих в роли
факторов адаптивных преобразований.
Для характеристики сложившегося биотического круговорота представляются важными
соотношения численности различных видов в биосфере, проанализированные Камшиловым,
с точки зрения эволюции биосферы. Он опирался на данные Ф. Г. Добржанского
(Dobzhansky, 1951). Согласно этим данным, численность видов животных (1 млн)
почти в четыре раза превышает численность всех видов растений (265.5 тыс.).
Не случайно и соотношение видов различных групп организмов в пределах отдельных
царств и таксонов. Так, покрытосеменные, представленные в основном сухопутными
формами, насчитывают 150 тыс. видов. Водорослей всего 14 тыс., они уступают
не только покрытосеменным, но и таким сухопутным формам, как грибы (70 тыс.)
и мхи (15 тыс.). Среди животных наиболее дифференцированными оказываются
также наземные формы – насекомые, насчитывающие более 750 тыс. видов. Такое
разнообразие насекомых и покрытосеменных объясняется их тесной взаимосвязью
в процессе эволюции, на что обращал внимание еще Дарвин. Столь значительное
преобладание наземных форм над водными явно свидетельствует о том, что завоевание
жизнью суши привело к усложнению биотического круговорота.
Необходимо отметить также, что, несмотря на меньшее видовое разнообразие
растений, именно на них выпадает 99% всей биомассы Земли. По мнению Камшилова,
это свидетельствует о существовании особого закона в эволюции биотического
круговорота. Согласно этому закону, „более высокий уровень дифференциации
сосредоточен в меньшем объеме, чем уровень менее дифференцированный” (1974,
с. 77).
Ведущее значение эволюции биотического круговорота в ходе исторических преобразований
живого отмечали и многие другие биологи. Так, согласно Н. В. Тимофееву-Ресовскому
с соавт. (1969), эволюция биосферы связана прежде всего с повышением интенсивности
биотического круговорота и с изменением самих биогеоценозов как целостных
систем. По мнению Г. Н. Симкина (1969), жизненные формы растений и животных
выступают в качестве функциональных звеньев в системе потоков вещества и
энергии в биосфере. Поэтому он полагает, что нарастающая функциональная и
структурная сложность биосферы протекает параллельно с увеличением разнообразия
жизненных форм. В результате эволюция их дает возможность более полно использовать
ресурсы биосферы, упорядочивает и интенсифицирует энергообмен в биосфере
и ее биогеохимический круговорот.
А.П.Хохряков (1981) считает, что определенные тенденции в эволюции высших
растений в сторону ускорения смены органов в ходе онтогенеза обусловлены
главным образом действием второго биогеохимиче-
176
ского принципа Вернадского, согласно которому в ходе прогрессивного развития
жизни неизбежно усиливается биогенная миграция атомов в биосфере. М. А. Голубец
(1982) среди важнейших характеристик эволюции экосистем называет „эволюцию
материально-энергетических (прежде всего трофических) и информационных связей
между их компонентами, а также межэкосистемных взаимосвязей, эволюцию их
биотического круговорота” (с. 62). Однако в целом он вынужден признать, что
мы можем лишь только в общих чертах охарактеризовать данный процесс развития.
Под влиянием трудов В. Н. Сукачева (1942, 1952, 1964а) было сформировано
учение о биогеоценозах как основных структурно-функциональных единицах биосферы.
В связи с этим в нашей стране уже с конца 50-х годов были предприняты первые
попытки поставить вопрос об основных тенденциях и главных этапах в эволюции
биогеоценотических систем (Быков, 1957, 1970, 1975; Ярошенко, 1961; Мазинг,
1970; Гиляров, 1973; Плотников, 1979; Старобогатов, 1984, и др.). В качестве
критериев эволюции биогеоценозов обычно назывались усложнение структуры экосистем,
повышение их эффективности в использовании ресурсов биосферы, рост биомассы,
совершенствование механизмов регуляции и т. д. Основные итоги исследований
в этом направлении подведены Ю. И. Черновым (1983). Поэтому мы не будем на
них подробнее останавливаться.
Отметим только исключительную важность учения о биологической структуре океана,
разработанного Л. А. Зенкевичем (1948, 1951, 1967), для понимания эволюции
биотического круговорота. Автор не раз указывал на перспективность сравнительных
исследований биогеоценозов суши и океана, биотическая структура которого
сформировалась задолго до заселения жизнью суши. Сам выход организмов на
сушу мог осуществиться лишь на базе сложившегося в море триединства „микробного,
растительного и животного компонентов с основными характерными особенностями
биологических цикловых связей продуцентов, консументов и редуцентов” (1967,
с. 523). Среди характерных особенностей морских биогеоценозов Зенкевич выделял:
огромный масштаб горизонтальных и вертикальных миграций его обитателей; широкие
межбиоценотические связи; различие в жизненных формах; мощный биокосный слой,
захватывающий всю толщу океана; ведущее значение одноклеточного фитопланктона
в биологической продуктивности; приуроченность растительного покрова к областям
литорали и сублиторали. В работах Зенкевича был дан обстоятельный анализ
основных различий между морскими и наземными биогеоценозами, их систематического
состава, морфологических и физиологических особенностей их обитателей, своеобразия
биогеохимических циклов, первичной продуктивности и трансформации
вещества и энергии.
В работах советских и зарубежных биологов не раз отмечалось, что эволюция
биотического круговорота должна анализироваться прежде всего с точки зрения
пирамиды жизни, т.е.пищевых цепей. Так, В. М. Виллиамсон еще в 1946 г. указывал,
что пирамида жизни Ч. Элтона представляет собой окончательный результат эволюции
в данную геологическую
177
эпоху. Трехслойная цепь (растения – травоядные животные – хищники), возникла
еще у простейших. Автор полагал, что взаимодействие трех уровней обусловливает
эволюцию каждого из них, но особая роль принадлежит верхнему уровню, так
как хищники всегда являются более активными, сообразительными и привередливыми.
По мнению А. М. Гилярова (1973), усложнение трофоценотической структуры экосистем
(появление добавочных трофических уровней, новых групп консументов, увеличение
разнообразия видов в пределах каждого трофического уровня) не может идти
бесконечно. Скорость эволюционных преобразований неизбежно замедляется по
достижении максимально возможного числа трофических уровней, которое, согласно
Р. Линдеману (Lindeman, 1942), равно шести, и максимально возможного видового
разнообразия в пределах каждого трофического уровня.
Оригинальный подход к проблеме становления и эволюции биотического круговорота
развивает А. М. Уголев (1980, 1985, 1987). Он предлагает создать новую науку
– трофологию, изучающую механизмы и закономерности трофических взаимодействий
в биосфере, а также трофологические аспекты эволюции видов, биоценозов и
биосферы. Трофические взаимодействия, по мнению Уголева, представляют собой
единую целостную систему, в которой отдельные виды и организмы функционируют
в виде взаимосвязанных звеньев. Соответственно биотический круговорот должен
рассматриваться прежде всего как трофический процесс. Формирование этого
процесса происходило с конца, т. е. с редуцентов, использующих абиогенные
органические вещества. По мере истощения его запасов возникали различные
организмы (фотосинтетики и хемосинтетики), способные к синтезу органических
соединений. Для нормального существования биотического круговорота необходимы
в равной степени гетеротрофы и автотрофы, так как преобладание последних
привело бы к быстрому исчерпанию необходимых химических элементов и соединений.
Эволюция каждого вида, по мнению Уголева, определяется в первую очередь его
приспособленностью к трофической структуре биосферы. Определяющую роль играют
не только его обеспеченность источниками питания и способы его использования,
но и „поедаемость” каждого члена трофической цепи, т. е. „доступностью для
другого организма в качестве источника пищи и трофичностью, т. е. питательными
свойствами и способностью быть ассимилированными” (1980, с. 57). Эволюция
трофической системы биосферы, подчеркивает Уголев, направлена на обеспечение
высокой эффективности, экономичности, регулируемости и надежности функционирования
биосферы. Тем самым делается попытка установить связи между эволюционными
преобразованиями трофики организмов с закономерностями функционирования и
эволюции биотического круговорота.
В заключение следует подчеркнуть, что проблема эволюции биосферы и ее биотического
круговорота настолько сложна, что требует усилий специалистов самых различных
естественных наук. Только в результате совместных исследований палеонтологов,
геохимиков, литологов, почвоведов, биогеохимиков, палеоэкологов и биологов-эволюцио-
178
нистов можно будет приступить к реконструкции исторических преобразований
биосферы. Само развитие биосферы связано с постоянным разрешением противоречия
между безграничной способностью организмов к размножению и ограниченностью
в каждый конкретный момент геологической истории материальных ресурсов, которые
могут быть использованы в процессах жизнедеятельности организмов. Разрешение
этого противоречия возможно только на пути создания все более прогрессивных
форм, способных к овладению новыми источниками вещества и энергии и к рациональному
их использованию. Все это вело к общему усложнению структуры биосферы, расширению
области ее распространения и увеличения числа различных органических форм.
Ближайшей задачей эволюционной теории является изучение воздействий микро-
и макроэволюции на исторические преобразования биосферы, на ее биогеохимическую
и энергетическую эволюцию. Создание общей теории эволюции биосферы станет
возможным лишь в результате интенсивной разработки биогеохимических, энергетических,
информационных подходов к ее изучению. Необходим также синтез этих подходов
с основными положениями современной эволюционной теории, которые исключительно
важны в качестве исследовательских установок для изучения самыми различными
методами эволюционных преобразований на уровне биосферы (см. подробнее: Колчинский,
1983а).
Требуется усилить труды биологов-эволюционистов по интеграции данных и обобщений,
полученных в ходе биогеохимических исследований, с основными положениями
дарвинизма. Эволюция биосферы, ее химического строения и энергетики определяется
прежде всего суммарными преобразованиями, происходящими на видовом уровне
организации живого. Поэтому призыв Вернадского исследовать, как эволюция
видов переходит в эволюцию биосферы, становится особенно актуальным в наши
дни.
6.5. К вопросу об основных этапах развития биосферы
Наиболее крупные этапы в эволюции биосферы были связаны, как правило, с появлением новых групп организмов, выполняющих специфические геохимические и энергетические функции в экосистемах. Нередко они также отличались специфическими формами действия общих факторов эволюции и наличием частных факторов. Эволюция биосферы и „эволюция эволюции” по существу два тесно связанных друг с другом процесса (Хозацкий, 1979; Татаринов, 1983; Колчинский, 19856), так как доминирующее положение в биосфере всегда занимали формы с наиболее прогрессивным механизмом эволюции. Переломные моменты в развитии биосферы и перестройка механизмов эволюции у доминирующих таксонов в биосфере совпадают во времени. Поэтому совместное изучение этих процессов дает возможность выработать единую адекватную методологию выделения основных этапов в истории органического мира и биосферы в целом.
179
Среди причин, затрудняющих разработку проблемы эволюции биосферы, одной
из главных, надо полагать, является отсутствие целенаправленного сбора знаний
о таких этапах в истории органического мира, которые характеризуются наличием
своеобразных форм действия общих факторов эволюции, а также специфических
факторов, действующих только на данном этапе. Эти этапы мы предложили назвать
историко-биологическими (Колчинский, 1971) или эволюционно-биологическими
формациями (Завадский, Колчинский, 1972, 1977). При их выделении учитывались,
однако, лишь изменения в каузальных факторах и движущих силах эволюции. Но
реальный процесс детерминации органической эволюции более сложен. Необходимо
принимать во внимание и кондициальные факторы, а также общее детерминирующее
воздействие структуры биосферы на процессы видообразования. Биогенная природа
факторов, обусловливающих возможность дальнейшей эволюции, видна хотя бы
из того, что само существование жизни вне толщи воды стало возможным лишь
с образованием озонового экрана, который является результатом фотосинтезирующей
деятельности древних водорослей. Кроме того, важно учитывать и воздействие
исторических преобразований каузальных основ эволюции видов на развитие биосферы.
В связи с этим возникает необходимость уточнить представление об основных
этапах в истории органического мира.
До сих пор предпринимались лишь отдельные попытки продолжить начатую Вернадским
разработку периодизации истории живой природы на иных основаниях, чем в геохронологии,
в которой не учитываются такие признаки, как изменение движущих сил эволюции
у доминирующих в биосфере таксономических групп и существенные преобразования
в структуре и функциях биосферы.
Современная геохронология сыграла важную роль в установлении общей последовательности
развития живого, в выделении основных преобразований флоры и фауны Земли,
в описании смен групп организмов и т. д. Однако ни геохронология, ни палеонтология
сами по себе не давали и не дают ответа на вопрос о причинах эволюции. Ограничены
их возможности и в изучении преобразований биосферы.
Описательный характер современной периодизации (геохронологии) вызвал необходимость
поиска новых, более глубоких критериев классификации и построения на их основе
принципиально новой схемы.
Подразделяя историю жизни на два этапа, Р. Маттей (Matthey, 1954) руководствовался
различием факторов и движущих сил эволюции. Первый этап (в течение первого
миллиарда лет) характеризовался формированием типов и классов. В то время
возникли все основные тины организации, движущей силой эволюции был не отбор,
а захват макромутантами еще не занятых зон жизни и крупных экологических
ниш. Это была эпоха господства „макромутации” и „типогенеза”. Второй этап
начался с того момента, когда плотность пригодных для жизни зон стала достаточно
высокой. С возрастанием количества организмов на нашей планете естественный
отбор приобретал все большее значение. Под его контролем происходят процессы
дифференциации популяций и видообразования.
180
Однако на этом этапе по существу ничего нового уже не возникало, а лишь
реализовались ранее накопленные генетические возможности.
Подобная трактовка содержит ряд рациональных моментов. Особенность докембрийской
биосферы состояла в слабой заселенности нашей планеты, в наличии множества
свободных экологических ниш и даже целых зон жизни. Относительная простота
протобиогеоценозов, в которых также была упрощена и система межвидовых отношений,
возможно, и ослабляла интенсивность естественного отбора. Можно предположить,
что в тот период, как пишет М. М. Камшилов, „проб было много, а критиков
мало” (1974, с. 197). Возможно, что лишь с усложнением структуры биосферы
и плотным заселением пригодных для жизни зон возникли более жесткие условия
для действия естественного отбора и все большую роль стал играть отбор на
конкурентоспособность. Вместе с тем несомненно, что на ранних этапах эволюции
существовала интенсивная конституциональная борьба, а также внутривидовая
конкуренция. Предположение же Маттея о том, что естественный отбор не действовал
на протяжении всего докембрия, ничем не обосновано.
В. В. Ковальский (1963а) предлагал выделить три этапа в истории живой природы:
протобиосферу, биосферу и ноосферу. При этом он правильно учитывал своеобразие
действия факторов эволюции и особенности биотического круговорота на каждом
из выделенных этапов. Однако предложенная Ковальским схема отражает только
наиболее крупные этапы в истории биосферы. Кроме того, первый и последний
этап по существу выходят за рамки чисто биологической истории органического
мира, так как представляют собой периоды перехода от неорганической формы
движения материи к биологической и от последней к социальной. Время же существования
самой биосферы практически не разделяется на этапы.
Д. И. Сапожников (1959) предложил при выделении основных этапов в эволюции
живого базироваться на эволюции типов обмена веществ, которая составляет
следующий ряд: 1) первичные гетеротрофы; 2) гетеротрофные усвоители углекислого
газа; 3) хеморедукторы; 4) фоторедукторы; 5) фотосинтетики. Эволюция типов
питания, по мнению Сапожникова, на первом этапе состояла в постепенном преобразовании
гетеротрофов, способных усваивать соединения, только очень богатые энергией,
в гетеротрофов, способных использовать продукты диссимиляции первичных форм.
На втором этапе возникла автогетеротрофность, т. е. автотрофность в отношении
аминокислот и витаминов при способности усваивать аммиачный азот. С возникновением
хеморедукции усвоение углекислого газа осуществлялось за счет энергии неорганического
вещества в анаэробных условиях. На четвертом этапе появилась способность
к фоторедукции (т.е. использование световой энергии для различных окислительно-восстановительных
реакций). И наконец, на пятом этапе происходило становление фотосинтеза:
повышение окислительно-восстановительного потенциала; появление и совершенствование
железосодержащих ферментных систем; включение в обмен свободного молекулярного
кислорода.
181
По мере становления механизма фотосинтеза анаэробиоз сменялся аэробиозом
и в атмосфере начал накапливаться биогенный кислород, что оказало в дальнейшем
существенное влияние на ход всех геологических, геохимических и биологических
процессов в биосфере (см. гл. 5). Однако в предложенной схеме нет временного
обозначения этапов эволюции типа обмена веществ, связанных с коренными переменами
в биосфере.
Многие ученые (Бернал, 1959; Гаффрон, 1962) выделяют в истории органического
мира шесть этапов, связанных со становлением следующих процессов: абиогенные
химические реакции, фотохимические реакции, ведущие к элементарным формам
накопления энергии; круговорот соединений углерода, включающий реакции консервации
лучистой энергии и ее освобождения, т. е. возникновение биотического круговорота;
усложнение и усовершенствование круговорота, связанное с возникновением организмов;
прогрессивная дифференциация живых существ, представляющая собой морфологический
процесс, ведущий к появлению и прогрессивному развитию многоклеточных организмов;
развитие нервной системы и органов чувств, приведшее к появлению поведенческих
реакций. Эта классификация основных стадий „биогенеза” далеко выходит за
рамки истории биосферы. Значительная часть этих стадий включает в себя процессы
не биологической, а геохимической эволюции.
В качестве основных критериев для выделения этапов в истории биосферы Р.
Фейбридж (1970) предложил использовать биолиты угольного ряда и карбонатных
пород. Он выделял пять этапов в истории биосферы, названных им „биогеохимическими
революциями”. Первая из них связана с возникновением жизни на Земле (3.8±0.3 · 109 лет
тому назад), вторая – с появлением первичных фотосинтезирующих организмов
(2.9±0.2 · 109 лет),
третья – с возникновением организмов с карбонатными раковинами (6.2±0.3 · 108 лет),
четвертая – с великой эпохой
образования
углей (2.5±0.3 · 108 лет) и пятая — с общебиосферным
распространением фораминифер (1.0±0.2 · 108 лет).
Эта периодизация не раз использовалась советскими геохимиками
для изучения геохимических особенностей архейских, протерозойских, палеозойских
и мезозойских биосфер (Лукашев, Вадковская, 1973).
И. М. Забелин (1978) выделяет основные этапы эволюции биосферы, учитывая
пространственную приуроченность основных эволюционных преобразований в тот
или иной период истории биосферы. Он называл это пространственными фазами
эволюции и намечал три главных этапа: существование биосферы в мелководных
прибрежных участках; освоение зоны всего океана; выход жизни на сушу. Очевидно,
что этот критерий только в совокупности с другими может быть использован
при выделении крупных этапов в истории биосферы.
М. А. Голубец называет три крупных этапа в истории биосферы, „знаменующих
переходы к новым качественным состояниям биосферы” (1982, с. 98): этап гетеротрофной
биосферы в условиях восстановительной обстановки на поверхности Земли; этап
существования организмов в условиях окислительной обстановки; этап антропогенного
воздействия
182
на процессы биосферы. Им в данной схеме учтены только наиболее крупные переломные
моменты в эволюции биосферы.
Рассматривая историю кислорода в атмосфере, Г. А. Заварзин (1984) назвал
следующие этапы доминирования в биосфере геохимических процессов от настоящего
времени к прошлому: „циклический механизм, обусловленный высшей наземной
растительностью и грибами, а также животными, ведущий к формированию гумуса
за счет лигноцеллюлозы с нижней границей около 0.4 млрд лет назад; циклический
механизм, обусловленный эукариотными водорослями и беспозвоночными океана
примерно 1.5 – 2 млрд лет назад; цианобактериальные сообщества и их анаэробное
разложение, формирование строматолитов, окисление резервуаров восстановленных
веществ, поставляемых эксгаляциями, – до 3.5 млрд; фотохимические реакции
в примитивной атмосфере у поверхности Земли” (с. 8—9).
Успехи в изучении докембрия побудили некоторых авторов предложить более детальную
датировку первых этапов органической эволюции и формирования биосферы. Так,
например, Б. С. Соколов (1976) считает что простейшие эобионты появились
на Земле, вероятно, ранее 4.25 млрд лет тому назад, возникновение механизмов
фотосинтеза у прокариотических пробионтов произошло 3.7 – 2.5 млрд лет, а
достоверное появление эукариотов – 1.6 – 1.35 млрд лет. Недавно Б. С. Соколов
и И. С. Барсков (1988) уточнили и детализировали представления о глобальных
этапах в развитии биосферы. В качестве начального этапа они выделили биосферу
с „прокариотной энергетикой” (возраст 3760 – 1900 млн лет), когда трофическая
структура биосферы и биогеохимические функции ее обитателей были иные, чем
в наше время. К важнейшим этапам в эволюции биосферы авторы относят также
„появление эукариот (1900 млн лет назад); возникновение многоклеточных организмов
(900 млн лет); развитие биоминерализации у животных (570 млн лет); заселение
суши растениями и животными, появление настоящих почв (400 млн лет), появление
покрытосеменных растений (130 млн лет); появление ноосферы... ” (с. 248),
Как видно, и здесь нет четко сформулированных критериев для выделения этапов.
Согласно М. Руттену, история жизни в докембрии разделяется на три основных
этапа: преджизнь; ранняя жизнь, существовавшая в условиях бескислородной
атмосферы; поздняя жизнь, связанная с кислородной атмосферой (1973, с. 159).
При этом он высказал интересную идею о том, что в течение долгого времени
преджизнь и ранняя жизнь существовали одновременно. Такое сосуществование,
по мнению автора, могло продолжаться от первого появления ранней жизни до
перехода к кислородной биосфере.
Были сделаны некоторые попытки связать воедино принципиальные крупнейшие
ароморфозы с эпохами повышенного накопления в осадках органического вещества,
с изменением хода всех биогеохимических процессов в биосфере и ростом ее
биомассы (Страхов, 1971; Соколов, 1976; Сидоренко, Теняков, Сидоренко, 1978).
Так, А. В.Сидоренко с соавторами среди важнейших событий органической эволюции,
обусловивших крупные эпохи осадкообразования, регионального метаморфизма
183
и магматической активности, назвали возникновение фотосинтеза и прокариот
(3.7—3.5 млрд лет тому назад), появление эукариот (1.9—1.6 млрд лет), массовое
развитие колониальных форм (1.6—1.3 млрд лет), возникновение митоза, мейоза,
метафитов и метазоев (1.0—0.9 млрд лет), экспансия многоклеточных (0.6—0.5
млрд лет). Из этой схемы видно, что существует довольно отчетливая корреляция
между преобразованиями каузальных основ эволюции, сменами доминирующих таксонов
в биосфере и изменениями в ее геохимических и геологических процессах. Даже
столь краткий обзор свидетельствует о том, что среди представителей различных
наук, изучающих биосферу, распространено убеждение о необходимости уточнить
понятие об основном этапе развития органического мира. Вместе с тем проблема
периодизации истории биосферы разработана слабо. Практически во всех рассмотренных
периодизациях учитываются только два этапа: первый, обусловленный возникновением
жизни, и последний, связанный с доминирующим положением человека в биосфере.
Следует согласиться с В. В. Друшицем (Друшиц и др., 1983, с. 80) в том, что
ни объем основных „этапов в развитии биосферы, ни их рубежи до сих пор не
разработаны и не выработаны единые критерии”. Для определения понятия об
основном этапе мы с К. М. Завадским предложили руководствоваться следующими
соображениями. Прежде всего учитывать крупные события в истории органического
мира, связанные с преобразованием каузальных факторов и движущих сил эволюции.
Во-вторых, принимать во внимание эволюцию способов питания.
Кроме того, мы предлагали учитывать особенности механизма эволюции и структуры
вида у доминирующих таксонов, названные нами эволюционно-биологическим укладом,
а также особенности соотношений таких укладов в биосфере. На заре эволюции,
когда структура протобиосферы была относительно простой, когда население
Земли было довольно однородным, а жизнь существовала в относительно простых
условиях первичного океана, возник одноукладный этап эволюции, который можно
назвать эобионтным. Для него было характерно специфическое взаимодействие
формирующихся факторов эволюции. В дальнейшем все основные этапы эволюции
были многоукладными. При переходе к новому этапу эволюции предшествующие
уклады, как правило, сохранялись и заключались в качестве структурной части
нового этапа, выполняя нередко важную роль в биотическом круговороте. Иногда
предыдущий уклад элиминировался как целое, и в новую биотическую систему
включались лишь отдельные его элементы (виды, биогеоценозы, а возможно, даже
целые флористические и фаунистические комплексы). При выделении основных
этапов развития живой природы следует учитывать их многоукладность, а также
структурную сложность каждого уклада.
Крупные преобразования в биосфере происходили обычно постепенно. Характерные
черты нового уклада зарождались в недрах предшествующего этапа, оставаясь
в течение долгого времени на положении второстепенного уклада. Существование
примитивных млекопитающих в течение всего мезозоя является хорошим примером
такого взаимоотношения между старым и новым укладами. Смена этапов никогда
не происходила путем полного уничтожения ранее доминирующих групп
184
организмов. Включение их в состав новых экосистем влекло за собой возрастание
многоукладности органического мира, увеличение видового разнообразия биосферы,
усложнение ее структуры за счет расширения области обитания жизни, дифференциации
экологических ниш и усложнения трофических цепей. Предшествующие формации
нередко не только сохранялись, но и служили базисом для нормального функционирования
и развития нового уклада. Подобная преемственность укладов обеспечила поступательную
эволюцию биосферы в сторону усложнения ее строения и повышения стабильности
и эффективности биотического круговорота.
При выделении основных этапов в эволюции биосферы необходимо учитывать доминирующие
типы географически наиболее распространенных биоценозов. Во время каждого
этапа существовало множество экосистем. Но среди них, во всяком случае в
пределах крупных экологических зон, всегда имелись доминантные биоценотические
отношения, образуемые видами доминирующих классов (Вишаренко, 1975). Их представители
образуют громадное число процветающих видов. У животных эти виды занимают
высшие звенья в цепях питания. В силу доминирующего положения в экологических
системах на данном этапе развития биосферы эти таксоны оказывали существенное
влияние на геохимические и энергетические функции биосферы, а специфические
закономерности эволюции доминирующего типа оказываются важными критериями
для выделения того или иного этапа в истории биосферы.
Возникновение более совершенного механизма эволюционных преобразований позволяло
таксону, занимавшему ранее субдоминантное положение в биосфере, в момент
коренных перетурбаций выходить на широкую эволюционную дорогу, а особенности
механизмов и структуры составляющих его видов, характеризовали новый доминирующий
уклад. Наступал новый период в истории органического мира. Именно появление
качественно новых, более эффективных механизмов эволюционных преобразований,
возможно, и было основной причиной завоевания господствующего положения одними
группами организмов и повсеместного вытеснения ими других.
Ранее мы предлагали основные этапы в истории органического мира называть
историко-биологическими или эволюционно-биологическими формациями, каждая
из которых характеризовалась специфическими формами действия общих факторов
эволюции, а также наличием некоторых частных факторов, действующих только
на данном этапе. При этом мы сознавали, что сейчас еще трудно сказать, сколько
было эволюционно-биологических формаций в течение развития жизни на Земле
и где проходят их временные границы. К тому же состояние наших знаний об
особенностях действия в геологическом прошлом факторов эволюции таково, что
нельзя выделять формации, опираясь только на этот критерий. Необходимо прибегать
и к помощи других критериев, в качестве которых могут быть использованы и
наиболее показательные результаты эволюции. Надо помнить, что такие крупные
эволюционные новшества, как например появление клеточного ядра, полового
процесса, многоклеточности, являясь результатом эволюции,
185
в то же время выступали в качестве ее мощных субстратных, а иногда и каузальных
факторов.
При характеристике и описании этих формаций полезно учитывать также специфичность
абиотических условий, оказывающих влияние на направление, характер и темп
эволюционных преобразований. Хотя физико-географические условия, будучи внешними,
кондициональными факторами видообразования, не оказывают решающего воздействия
на эволюционный процесс, однако колебания климата, процессы трансгрессии
и регрессии, орогенетические процессы, вулканизм и т. д. влияли на интенсивность
мутагенеза и модифицировали действие естественного отбора. В первых же формациях
в силу относительной простоты их структур и примитивной организации заселяющих
их форм такие условия среды, как состав атмосферы, космические и солнечные
излучения и т. д., выступали в качестве важнейших агентов отбора.
Переход от одного этапа эволюции к другому не может быть понят без „учета
всего комплекса причин и факторов, осуществляющих этот процесс” (Хозацкий,
1979, с. 56). Сущность того или иного этапа в эволюции органического мира
может быть раскрыта лишь при учете всех пространственных, геохимических,
энергетических и информационных характеристик биосферы, которая не только
является ареной процессов видообразования, но и обеспечивает его единство
и непрерывность на протяжении всей истории жизни. Эволюция каузальных основ
эволюции не может быть понята без соотнесения ее с общей эволюцией биосферы
как целостной системы, без учета изменений ее геохимического строения, энерговооруженности
и информационного „фонда”.
В связи с этим было бы правильнее основные этапы в развитии органического
мира называть не эволюционно-биологическими, как это предлагалось нами раньше,
а эволюционно-биосферными формациями. Структурность развития органического
мира заключается не в этапности его отдельных филумов, а в этапности эволюции
всей биосферы. При этом арогенезы наиболее крупного масштаба, ведущие к появлению
новых таксонов высокого ранга, обусловливали и эволюцию биосферы, вызывая
смену этапов в ее развитии.
Таким образом, для разработки проблемы эволюции биосферы необходимо дальнейшее
уточнение представлений об эволюционно-биосферных формациях как главных этапах
в истории биосферы, каждый из которых характеризовался особыми структурно-функциональными
и эволюционными закономерностями. Эволюционно-биосферные формации далеко
не всегда совпадали с выделяемыми сейчас геологическими формациями. Однако
это не означает, что при выделении их следует отказаться от использования
геохронологии. Описывая ту или иную эволюционно-биосферную формацию, мы не
можем оставить в стороне субстрат эволюции, т. е. основные свойства эволюирующих
групп (в особенности доминирующих таксонов) и надорганизменных систем. При
описании формаций следует указывать как доминирующую группу, так и время
существования этой формации. Нам представляется, что были бы удачными такие
названия эволюционно-биосферных формаций, как
186
например катархейская эобионтная, архейская протобионтная, протерозойская
прокариотная, протерозойская эукариотная и т.д.
Приведем пример возможной характеристики одной из первых формаций, названной
нами протерозойской прокариотной. При этом мы будем опираться на работы советских
и зарубежных палеонтологов, геологов, геохимиков и палеобиогеохимиков по
докембрию (Глесснер, 1963; Крылов, 1972; Барнхорт, 1972; Schopf, 1975; Вологдин,
1976; Cloud, 1976; Заварзин, Крылов, 1983; Заварзин, 1984) и др. „Состав
и состояние внешних геосфер в докембрии были благоприятными для существования
бактериально-водорослевого биоса, и действительно, имеется немало прямых
и косвенных доказательств развития и деятельности древнейших организмов”
(Вассоевич и др., 1973, с. 153).
Область распространения биосферы тогда была, по-видимому, ограничена мелководьями,
так как разрушительное действие ультрафиолетовых лучей при отсутствии озонового
экрана не позволяло существовать наземным формам. Плотность заселения этой
биосферы определялась прежде всего физико-географическими факторами, прямо
воздействующими на продуктивность формирующихся ценозов. В процессе растекания
жизни по поверхности Земли и насыщения ею пригодных местообитаний происходила
дифференциация живого на скопления, приспособленные к качественно различным
комплексам абиотических факторов. К ним относятся температура и прозрачность
вод, наличие минеральных соединений и т. д. Возникновение прокариотов существенно
изменило взаимоотношения организмов со средой: начали формироваться первичные
микроценозы, аналоги которых и сейчас существуют в некоторых водоемах, не
пригодных для жизни более высокоорганизованных форм. Если предбиологические
формы (протобионты, эубионты) не вступали между собой в устойчивые связи,
то у прокариотов возникают аллелопатические связи, различные симбиотические
и фагоцитозные взаимодействия, послужившие в дальнейшем основой для развития
сложных экологических отношений. Все большее значение приобретают внутренние
факторы развития живого, т. е. взаимодействия живого с живым. Отныне соответствие
только абиотической среде и наличие пищевых ресурсов оказываются недостаточными
для выживания. Успеха добиваются более конкурентоспособные организмы. Сами
агенты отбора становились все более сложными, а специализация - одним из
важнейших условий сохранения вида. Сущность отбора заключалась в переживании
наиболее приспособленного не только к абиотическим, но и к биотическим факторам
среды.
Происходило и освоение новых зон жизни. В это время как бы испытывались разнообразные
способы использования энергии и обмена веществ (Stanier, 1961). Среди бактерий
существуют четыре основные группы: фотолитотрофные, фотоорганотрофные, хемолитотрофные,
хемоорганотрофные. Многие из них способны переходить с дыхания на брожение
и обратно в случае изменения содержания кислорода в окружающей среде. Биохимические
преобразования преобладали на этой стадии эволюции биосферы.
187
В экосистемах данной эволюционно-биосферной формации существовали лишь две
основные группы организмов: цианофеи и бактерии, которые и обеспечивали синтез
и разложение органических веществ. Функция же консументов практически отсутствовала
в древних экосистемах (Заварзин, Крылов, 1983).
Если судить по общей биомассе современных бактерий и синезеленых водорослей,
то биомасса рассматриваемой формации была меньше биомассы последующих биосфер.
Но благодаря высокой геохимической энергии прокариот высокая продуктивность
биосферы была обеспечена на начальных этапах ее эволюции.
Несмотря на относительную простоту экосистем этой эволюционно-биосферной
формации, результаты ее функционирования оказались грандиозными с геохимической
и энергетической точек зрения. Существенные черты геохимического и энергетического
строения биосферы сложились уже в рамках этой формации. Фотосинтезирующие
организмы стали главными источниками энергии всех биологических процессов.
Их жизнедеятельностью сформирована и стабилизирована кислородная атмосфера.
Микроорганизмы „изъяли” из первичной атмосферы громадные количества углекислого
газа, превратив его в мощные залежи карбонатов (Вологдин, 1976). Благодаря
их жизнедеятельности происходило очищение атмосферы от ядовитых газов вулканического
происхождения. Они сыграли важную роль в регулировании солевого состава морей
и океана, в стабилизации основных химических свойств гидросферы Земли (Страхов,
1962; Виноградов, 1967; Валяшко, 1971; Перельман, 1982). С их жизнедеятельностью
связано не только коренное преобразование всей геохимической обстановки на
поверхности нашей планеты от восстановительной в окислительную, но и формирование
громадных месторождений железных и полиметаллических сульфидных пород, фосфоритов
и т. д. (Сауков, 1961; Кузнецов и др., 1962; Ронов, 1964,1980, и мн. др.).
Развитие прокариотной эволюционно-биосферной формации не просто предопределило
всю дальнейшую судьбу органической эволюции. Оно практически решило вопрос:
быть или не быть на нашей планете сложным формам жизни.
Необходимо отметить существенные особенности каузальных основ эволюции у
организмов, господствующих в данной формации. Эти формы отличались от предшествующих
наличием клеточной оболочки, создавшей возможность для возникновения и нормального
развитая внутриклеточных обменных процессов и оградившей генетический аппарат
от непосредственного влияния факторов внешней среды. В повышении устойчивости
наследственной информации важную роль сыграло возникновение хромосом и митотического
аппарата ее редупликации. Характерными факторами этой эволюционно-биосферной
формации были микроскопические размеры тела, ультракороткий срок жизни отдельной
особи, высокая плодовитость, громадная численность популяций, пассивность
организма, развитие химической регуляции (через среду) в биоценотических
отношениях, еще слабая расчлененность изменчивости на генотипическую и фенотипическую.
В результате взаимодействия всех этих факторов эволюционные процессы протерозойской
прокариотной формации
188
были крайне неэкономными. Для одного шага эволюции требовалось сотни миллиардов
особей. Вновь возникающая мутация не могла быть сохранена в рецессиве и подвергалась
немедленной проверке. Вместе с тем широкое развитие получили такие формы
обмена генетической информацией между различными группами организмов, как
конъюгация, трансформация и трансдукция. Широкое распространение механизмов
горизонтального переноса генетической информации в принципе позволяет говорить
о наличии единого генофонда биосферы на данном этапе ее развития.
Роль избирательной элиминации была незначительна, в то время как катастрофический
отбор и генетико-автоматические процессы были широко распространены. Творческая
роль отбора проявлялась лишь в суммировании полезных мутаций, а его основными
(преобладающими) направлениями были отбор на координацию биохимических процессов
в клетке, на большую плодовитость и т. д. Дав начало более высокоорганизованным
формам, прокариоты в дальнейшем изменялись, видимо, очень мало, совершенствуясь
главным образом биохимически и физиологически в пределах уже достигнутого
уровня организации.
В результате крупных эволюционных изобретений (образование клеточного ядра,
образование органелл движения и возрастание активности фенотипа, возникновение
полового процесса и диплоидности и т. д.) существенно изменились движущие
силы эволюции, что послужило основой перехода к новой протерозойской эукариотной
эволюционно-биосферной формации, в которую предшествующая не просто вошла
в качестве эволюционного уклада, но и составила основу ее энергетико-геохимических
процессов. Более того, и в настоящее время „многие процессы жизненного цикла
микроорганизмов составляют основу жизненных функций высших растений и животных”
(Руттен, 1973, с. 145). И если бы исчезли внезапно все прокариоты, то вряд
ли биосфера могла сохраниться.
Мы не будем останавливаться на характеристике других эволюционно-биосферных
формаций, хотя накопленные к настоящему времени данные палеонтологии, палеобиогеохимии,
геохимии, эволюционной теории и некоторых других наук позволяют дать описание
некоторых из них, например протерозойской эукариотной или гомойотермной формаций.
Так, в ряде работ протозоологов обобщен богатый материал об особенностях
эволюции простейших (Полянский, 1971, 1981; Райков, 1978; Серавин, 1984,
и др.), которые занимали доминирующее положение в протерозойской эукариотной
формации. В пределах этой крупной таксономической группы, выделяемой теперь
в самостоятельное царство (Whittaker, 1969), общие факторы и причины эволюции
действуют в специфических формах (особенности аппарата наследственности;
разнообразие способов агамного и полового размножения; своеобразие аппаратов
митоза и мейоза; особенности процессов конкуренции, хищничества, конституциональной
борьбы за существование и естественного отбора). Кроме того, существует ряд
частных факторов эволюции, характерных только для этой группы животных (длительные
модификации, эпигеномные изменения, высокая полиплоидность и ядерный дуализм,
189
симбиогенез и паразитизм как факторы прогрессивной эволюции и т.п.). В ряде
работ были предприняты попытки охарактеризовать специфику факторов и движущих
сил эволюции в формации, где доминирующее положение занимали млекопитающие
и покрытосеменные (Колчинский, 1971, 1983б; Завадский, Колчинский, 1977).
Однако задачей данного параграфа является только постановка вопроса о необходимости
понятия „эволюционно-биосферная формация”. При его разработке, видимо, будет
происходить объединение данных общей и частных теорий эволюции с материалами
палеобиологии, включая палеоэкологию, а также данные исторической геологии,
палеобиогеохимии, палеогеографии, палеоклиматологии и других наук, позволяющих
реконструировать особенности среды обитания прошлых эпох, геохимических и
энергетических функций живого вещества, структуры экосистем, специфические
типы взаимодействия между факторами эволюции и т. д.
Мы полагаем, что методология формационного анализа может стать базой для
синтеза данных различных геологических и биологических наук, связанных с
изучением эволюции биосферы. Формационный анализ позволяет лучше представить
целостность развивающейся биосферы, вскрыть основные противоречия, лежащие
в основе ее развития, более подробно охарактеризовать все многообразие источников
развития и системную сложность его каузальных взаимодействий.
* * *
В начале 50-х годов Л. А. Зенкевич, отмечая исключительную важность изучения эволюции биосферы, вынужден был признать, что мы совершенно не знаем основных тенденций целостного развития жизни и главных параметров биосферы на отдельных этапах ее истории. Он писал: „Мы совсем ничего не можем сказать, как изменялось общее количество организмов в биосфере, как усложнялись системы взаимодействий между основными биологическими группами в биосфере и как сложилась та система циклических процессов в биосфере, которую мы ныне знаем” (1951, с. 140). В настоящее время становятся понятными принципиальные трудности, связанные с изучением эволюции биосферы. Становление основных особенностей биосферы и ее биотического круговорота явно относится к тому времени, о котором мы не можем судить по палеонтологическим данным. В ископаемое состояние переходят лишь остатки организмов и различные следы их жизнедеятельности, а структура и функции биосферы не фоссилизируются. В связи с тем, что метод прямых наблюдений за процессами эволюции биосферы неосуществим, необходимо пользоваться совокупностью косвенных методов. Суть этих методов заключается в том, чтобы на основе сравнительных исследований современных экосистем,
190
каждая из которых может быть рассмотрена как модель определенного этапа
в эволюции биосферы, и на базе синтеза данных палеобиохимии, палеонтологии,
палеоэкологии и других наук, связанных с изучением палеобиосфер, попытаться
выделить главные тенденции в эволюции биосферы.
Выполненный нами анализ первых результатов изучения исторических преобразований
общей биомассы биосферы, ее продуктивности и энергетики, информационного
„фонда” и общепланетарного биотического круговорота показывает, что в целом
начинают вырисовываться некоторые основные тенденции и закономерности в
эволюции биосферы. Но в сущности мы находимся только в начале длительного
пути, ведущего
к их познанию.
191
Глава 7
ОСОБЕННОСТИ ОРГАНИЧЕСКОЙ ЭВОЛЮЦИИ
В СОВРЕМЕННОЙ БИОСФЕРЕ
(ВМЕСТО ЗАКЛЮЧЕНИЯ)
Глобальные изменения в поверхностных оболочках Земли, прошедшие за последние
десятилетия под влиянием промышленности, расширения сельскохозяйственных
угодий, урбанизации и т. д., превратили проблему эволюции биосферы в одну
из важнейших проблем, стоящих перед современным человечеством. Государственные
деятели разных стран, ученые самых различных специальностей, писатели, художники,
кинематографисты и другие деятели культуры — все стараются как-то понять
и оценить происходящие сейчас эволюционные необратимые преобразования биосферы.
Но среди литературы, посвященной проблемам эволюции биосферы в современных
условиях, очень мало работ, написанных биологами-эволюционистами. Внимание
биологов чаще всего концентрируется на эволюционных изменениях какого-либо
конкретного вида или биологического сообщества. Проблема же особенностей
органической эволюции в современной биосфере практически обсуждалась только
в работах 3. И. Бермана, А. А. Парамонова, Ю. И. Полянского (1967), М. М.
Камшилова (1970, 1974), С. С. Шварца (1973, 1976) и А. В. Яблокова (1978,1983).
Между тем выработка способов управления эволюционным процессом в условиях
все возрастающего давления общества на природу возможна лишь на основе знаний
не только общих, но и специфических факторов эволюции видов и всей биосферы
в целом. Изучение особенностей факторов и движущих сил эволюции в современной
биосфере необходимо для планирования будущего и управления эволюционным процессом
во все более широком и, наконец, в глобальном масштабе. Цель этой заключительной
главы состоит в том, чтобы определить особенности эволюции в современной
биосфере и наметить пути к управляемой эволюции.
Заключительные периоды в развитии биосферы связаны со становлением антропозойной
эволюционно-биосферной формации, где человеческая деятельность постепенно
стала ведущей преобразующей силой на поверхности нашей планеты. По новейшим
данным (Урысон, 1972; Бунак, 1980; Leakey, 1981; Джохансон, 1984; Алексеев,
1984), антропогенез начался гораздо раньше, чем это предполагалось в годы
открытия питекантропа яванского и синантропа. Видимо, более 4.5 млн лет тому
назад антропогенез протекал в немногих малочисленных популяциях антропоидов,
занимавших ограниченный ареал и по своему воздействию
192
на окружающую среду еще практически ничем не отличающихся от популяций других
высших приматов. И так же как по мере развития живого все возрастало его
влияние на безжизненное, косное окружение, а геохимические процессы становились
все более биогенными, так и по мере становления и развития человечества все
усиливалось и становилось качественно разнообразным его влияние на органическую
эволюцию. В течение последних тысячелетий человечеству удалось уничтожить
плейстоценовую фауну в крупнейших регионах биосферы (Евразия, Америка и Австралия)
вскоре после проникновения в них.
Определение нижней границы антропозоя как этапа в развитии биосферы вызывает
разногласия, связанные с выбором критериев выделения. Если в качестве такового,
например, избирается доместикация и начало эволюции в условиях культуры,
то эта граница совпадает с периодом неолитической революции в истории человечества
и проходит примерно 8000 – 10 000 лет до н. э. Если же избираются признаки,
характеризующие начало ноосферы в смысле В. И. Вернадского, то и ныне мы
находимся только на подступах к ней. На глазах последних поколений влияние
человечества на процессы эволюции стало глобальным и таким исключительно
большим, что требования всесторонней планомерности воздействия и учета его
отдаленных последствий становятся настоятельно необходимыми. В связи с этим
изучение особенностей эволюционного процесса в современных условиях приобретает
громадное практическое значение.
Следует заметить, что для неспециалистов проблемы исторического развития
жизни кажутся связанными только с процессами далекого геологического прошлого
и ассоциируются с описательными методами палеонтологии. И в этом смысле они
представляются очень далекими от современности. Действительно, крупнейшие
преобразования живого, так называемая мегаэволюция, осуществлялись на протяжении
миллиардов, сотен миллионов лет, лишь в случаях крайне быстрой эволюции —
в течение десятков миллионов лет. Основываясь на том бесспорном факте, что
все крупные таксоны возникли в далеком геологическом прошлом и что по мере
приближения к современности все меньше становился масштаб новообразований,
некоторые биологи сформулировали финалистическую концепцию эволюции (см.
подробнее: Назаров, 1984). Так, Виганд полагал, что запас „образовательной
силы” достигал максимума лишь на начальных этапах эволюции живого, а затем
постепенно уменьшался и полностью исчез к настоящему времени, так как эта
„сила” иссякла. Виды стали неизменными и вновь могут возникать лишь расы
и разновидности. Подобные оценки эволюционных возможностей современных органических
форм часто базируются на филогеронтологических идеях о запрограммированности
срока существования каждой таксономической группы, которая неизбежно должна
проходить те же стадии развития, что и индивид (Д. Роза, К. Бойрлен, А. Вандель,
О. Шиндевольф). В рамках этих финалистических представлений бессмысленна
даже постановка вопроса об эволюции современных организмов.
Исходя из других оснований, к учению о „финале эволюции” пришел Б. М. Козо-Полянский
(1922). В нашу эпоху, полагал он, совокупная
193
мощь воздействия человечества на живую природу чрезвычайно возросла и эволюция
практически прекратилась как стихийный процесс, регулируемый статистическим
механизмом естественного отбора. Рост численности населения, развитие производительных
сил, приручение животных и окультуривание растений, распашка громадных площадей
и создание пастбищ, массовая вырубка лесов, индустриализация добычи морских
животных — все эти и другие прямые и косвенные воздействия человека на дикоживущие
виды и среду их обитания, по мнению Козо-Полянского, привели к тому, что
биологическая эволюция как процесс природы закончилась. Человечество стало
единственным фактором незначительных преобразований органических форм, которые
возможны лишь под разумным контролем со стороны человека. „Процесс эволюции
весь в прошлом, - писал он, - а формообразование сейчас идет в поистине ничтожных
размерах” (там же, с. 8).
Такая постановка вопроса хотя и не содержала философски ошибочных идей, характерных
для настоящих финалистических гипотез, все же оказалась неверной (Хахина,
1977). В ней по существу отрицалась возможность эволюционных преобразований
большинства организмов, не вовлеченных непосредственно в сельскохозяйственную
практику человека. Доместикация животных и окультуривание растений практически
расценивались как единственные пути эволюционных преобразований современных
видов.
Действительно, эволюция, направляемая волей человека, — это реальный процесс,
протекающий в биосфере со времен неолитической революции. Уже тогда в процессы
производства для удовлетворения утилитарных и эстетических потребностей человека
были вовлечены десятки видов животных и сотни видов растений. Впоследствии
путем бессознательного и методического отбора удалось вывести громадное количество
сортов культурных растений и пород домашних животных, не существовавших до
этого в природе. Тем самым человек стал мощным фактором увеличения разнообразия
органических форм в биосфере. Не случайно теоретическое осмысление практики
селекционных работ было одним из важнейших источников для создания учения
Ч. Дарвина, а селекция превратилась в „учение об эволюции, направляемой волей
человека” (Вавилов, 1934).
Но современные взаимоотношения человека с природой характеризуются тем, что
его сознательное воздействие на эволюцию ограничено очень узким кругом видов.
До недавнего времени их число составляло 30 видов животных и около 250 видов
растений из 2 млн видов, существующих в антропозойной биосфере. Если считать
и виды, с которыми только начата селекционная работа, то и сейчас они составляют
лишь сотые доли процентов всех современных форм. По словам П. М. Жуковского,
„феноменальное историческое явление состоит в том, что почти все основные
культурные растения были одомашнены за несколько тысяч лет до нашей эры.
То же случилось и с одомашниванием животных” (1970, с. 3). Если допустить,
что в антропозое эволюция идет только под разумным контролем человека, то
окажется, что подавляющее большинство органических форм обречено
194
на вымирание, будучи не в состоянии адаптироваться к среде, резко измененной
человеком.
На первый взгляд это предположение подтверждается массовыми вымираниями видов.
Ежедневно исчезает один вид животных и еженедельно – один вид растений (Яблоков,
1983). Сейчас под угрозой вымирания находится около 1200 видов животных,
несколько десятков тысяч растений (Большаков, 1983). Только в „Красной книге
СССР” (1978) среда редких и находящихся под угрозой исчезновения видов насчитывается
62 вида млекопитающих, 63 – птиц, 21 – рептилий, 8 – амфибий, 418 видов и
подвидов растений. Вместе с тем органический мир отнюдь не пассивно реагирует
на воздействия человека. Эволюционная пластичность многих форм и в наши дни
достаточно высока. Вопреки воле человека интенсивно формируются новые формы
сорных растений, вредителей сельского хозяйства, возбудителей различных заболеваний
и т. д. На урбанизированных территориях доминирующее положение занимают рудеральные
растения и животные — комменсалы и паразиты человека. Идет интенсивный отбор
на синантропизацию, т. е. на выработку свойств, позволяющих выжить и оставить
потомство в среде, измененной человеком. Трудно представить конечный итог
этих эволюционных преобразований. Возможно, они приведут к созданию форм,
с которыми человек не сможет уже справиться. И вряд ли эту эволюцию следует
считать „находящейся под разумным контролем человека”.
Среди разнообразных форм воздействия человека на окружающий мир (разрушение
привычных местообитаний животных и растений, вырубка лесов, распашка степей
и лесостепей, осушение болот, создание искусственных водоемов, загрязнение
атмосферы и гидросферы, горнодобывающие работы, урбанизация громадных территорий,
сокращение и подрыв кормовой базы животных, чрезмерная добыча хозяйственно
ценных видов) практически нет факторов, которые можно было бы отнести к числу
„разумно контролируемых” и направляющих эволюцию в нужную для нас сторону.
Напротив, они вызывают эволюционные преобразования, близлежащие (не говоря
уже об отдаленных) последствия которых предсказать практически невозможно.
Как правило, мы не можем предвидеть эволюционные последствия даже таких факторов,
которые, вызывая массовую гибель организмов того или иного вида, в то же
время являются агентами катастрофического отбора, способного вызвать быстро
новую адаптацию.
В биологии накопилось много примеров того, что органический мир реагирует
на антропогенные факторы активными микроэволюционными преобразованиями. Примерами
таких эволюционных ответов служат индустриальный меланизм, выработка ядоустойчивых
рас насекомых, приобретение бактериями и вирусами устойчивости к лекарственным
препаратам и т. д. Чаще всего они вызывают дополнительные трудности во взаимодействии
человека с природой. Выработка резистентности у болезнетворных микроорганизмов
к лекарствам заставляет изыскивать все новые и новые сильнодействующие антибиотики.
Устойчивость некоторых рас насекомых и грызунов к ядам возросла в сотни раз,
195
и потери от вредителей по-прежнему составляют десятки процентов урожая,
несмотря на все защитные меры.
Под воздействием человека идут и нежелательные изменения численных соотношений
видов в природных экосистемах. Преобладающими становятся хозяйственно малоценные
виды с высокими коэффициентами размножения, с громадной численностью популяций,
со сложной эколого-генетической структурой. Мышевидные грызуны, „сорные виды”
рыб, малоценные породы древесных растений и сорные травы зачастую занимают
в экосистемах места ранее доминирующих высокоразвитых животных и ценных растений.
Многим микроорганизмам и растениям удалось довольно быстро приспособиться
к самым, на первый взгляд, неблагоприятным условиям. Радиоактивные и химические
мутагены резко ускорили мутационный процесс. В больших масштабах идет отбор
на резистентность организмов к фенолам, к загазованной атмосфере, к нефтяной
пленке в водоемах и т. д. Всеобщая гибель живого отнюдь не единственный ответ
на подобные результаты деятельности человека. Например, эффективность бактерий-нефтедеструкторов,
разлагающих нефтяные отходы, не только не снизилась, но даже стала повышаться
при росте концентраций цинка и ванадия. В течение нескольких поколений полевица,
ценное кормовое растение, приобрела генетически закрепленную адаптацию к
почвам, обогащенным свинцом, медью, никелем и обедненным к тому же кальцием
и фосфором (Шварц, 1976). Некоторые растения оказались способными переносить
высокие концентрации токсических элементов в атмосфере и почвах и создавать
довольно стабильные и продуктивные сообщества в регионах интенсивной промышленной
деятельности (Кулагин, 1971). Наблюдаются и случаи сопряженной эволюции с
растениями насекомых-фитофагов, приспособленных к питанию растениями с большими
концентрациями пестицидов (Ehrlich A., Ehrlich P., 1982).
Уникальный пример выработки новой адаптации на протяжении одного поколения
людей буквально обошел всю научную и популярную литературу: приспособление
калифорнийского кузнечика Romalia microphtera к гербициду 2,4 дихлорфенол.
В его популяции возникли и распространились особи, которые не только нейтрализовали
яд, как это часто происходит и в популяциях других насекомых. Мутантные особи
оказались способными трансформировать 2,4 дихлорфенол в еще более ядовитое
вещество – 2,5 дихлорфенол, которое образуется при соединении 2,4 дихлорфенола
с секретируемыми кузнечиком веществами. Благодаря этому кузнечик стал неуязвим
для своих врагов – хищных насекомых и птиц.
Эволюционные преобразования происходят и в популяциях рыб, птиц и млекопитающих,
подвергающихся постоянному, но не слишком интенсивному истреблению в процессах
лова и охоты. Как правило, они ведут к омолаживанию структуры популяций,
к сокращению продолжительности жизни, к ускорению смены поколений, к уменьшению
как средних, так и абсолютных размеров тела (Попов, 1971; Никольский и др.,
1972, и др.). Особенно резкие изменения проявляются в стереотипах поведения,
связанные с потребляемой пищей, сезонными миграциями,
196
выбором местообитаний и т.д. (Владышевский, 1973; Банников, Флинт, 1973;
Захаров, 1984, и др.). Все больше диких видов животных начинают обитать в
антропогенных ландшафтах в непосредственной близости с человеком (например,
белки, бобры, барсуки, лебеди и т. д.). И хотя эти эволюционные изменения
нельзя оценить как негативные для человека (напротив, они направлены в сторону
адаптации диких животных к антропогенной среде), они не являются и следствиями
эволюции, управляемой человеком. Его деятельность здесь выступает лишь как
один из агентов отбора. Эволюция идет помимо целей и намерений человека,
хотя объективно и совпадает с ними.
Человек вынужден считаться с действием естественных факторов эволюции даже
на территориях, которые, на первый взгляд, всецело находятся под его контролем.
Культурные растения должны устоять в сложных межвидовых отношениях с вредителями
и сорняками. Наряду с хозяйственно ценными признаками культурные растения
должны отличаться высокой конкурентоспособностью, устойчивостью к грибковым
и вирусным заболеваниям и паразитам, высокой приспособленностью к абиотическим
условиям существования.
Даже в процессах селекции естественный отбор не утратил полностью своего
значения, а продолжает действовать наряду с искусственным отбором, приводя
к созданию „локальных и региональных сортов, т. е. экологических форм, которые
отличаются от природных экологических рас лишь привязанностью к искусственным
биогеоценозам” (Шмальгаузен, 1969, с. 258). При этом естественный отбор может
как тормозить, так и усиливать действие искусственного отбора. Игнорирование
этого обстоятельства может привести к вырождению ценных сортов растений и
высокопродуктивных пород животных. Лишь при соблюдении биотехнических и агрономических
приемов может быть обеспечено как выведение, так и поддержание на требуемом
уровне хозяйственно ценных сортов и пород.
Следует учитывать также, что на обрабатываемых полях образуются свободные
экологические ниши, куда устремляется множество сорных растений и вредных
животных. Природные биогеоценозы, как отмечал еще Вернадский, обычно отличаются
большей стабильностью биогеохимического круговорота, чем культурные ценозы.
Качественный состав биогеоценозов обеспечивает максимальное использование
условий биотопа. Создаваемые же человеком виды связаны с меньшей биогенной
миграцией химических элементов, чем их дикие предки, что угрожает дестабилизацией
существующего биотического круговорота, так как противоречит биогеохимическим
принципам, лежащим в основе функционирования и развития биосферы. Все это
оказывает дополнительные воздействия со стороны естественного отбора на популяции
растений и животных, вовлеченные в селекционный процесс и культивируемые
в агроценозах.
И наконец, нельзя забывать и о том, что в наши дни эволюция происходит во
многих группах организмов без видимого участия антропогенных факторов. Здесь
нет возможности перечислить все новые расы, подвиды и даже виды, возникновение
которых почти на наших глазах было
197
зафиксировано. Достаточно сослаться на примеры сверхмолодых видов, перечисленных
К. М. Завадским (1968): рябинокизильник из восточной Якутии, ячмень-клинэлимусовый
гибрид с Памира и др. Многие из этих недавно возникших форм отличаются высокой
конкурентоспособностью и захватывают широкие ареалы.
Таким образом, эволюция в современной биосфере не только не прекратилась,
но идет огромными темпами. И хотя человеческая деятельность и ее последствия
чаще всего являются ведущими агентами селективных процессов, в целом эволюция
идет стихийно, нередко против воли и целей человека. Нет оснований говорить
ни о финале эволюции, ни об эволюции, контролируемой человеком. Переживаемый
сейчас биосферой период — это не период кризиса, а период ее коренной перестройки.
Отмечаемый во многих работах кризис биосферы скорее является кризисом той
биосферы, к которой приспособлен человек. В истории биосферы не раз были
периоды, когда под воздействием каких-то мощных абиотических факторов резко
возрастала интенсивность действия отбора и ускорялись процессы видообразования
(Будыко, 1973; Родендорф, Жерехин, 1974). Образование новых форм нередко
влекло за собой такие изменения в окружающей среде, которые оказывали губительное
воздействие на ранее существовавшие группы организмов. В результате исчезали
многие семейства, отряды и даже классы, а их место в биосфере занимали другие
формы, более приспособленные к новым условиям. Смена анаэробных организмов
аэробными является наиболее ярким примером коренных преобразований биосферы.
С самого своего возникновения организмы меняли биосферу и сами изменялись
под воздействием вызванных ими преобразований. И сейчас органический мир
в целом может приспособиться к той экологической ситуации в биосфере, которая
возникла в результате деятельности человека.
Трудно, конечно, предсказать дальнейший ход эволюции биосферы. „Мы не знаем,
к чему в конце концов приведет новая эволюция. Человечество знает только
самое ее начало” (Вайскопф, 1977, с. 233). Но ему далеко не безразлично,
каков будет ее конец, т. е. какой будет формирующаяся сейчас биосфера. Человек,
выросший в недрах голоценовой биосферы, может поддерживать нормальную жизнедеятельность
только в том случае, если будут сохранены главные параметры именно этой биосферы,
ее основные черты и видовой состав. Крах голоценовой биосферы чреват для
человека многими нежелательными последствиями, о которых нет необходимости
здесь говорить, так как они не раз подробно анализировались во многих работах,
посвященных экологии человека в современных условиях. Необходимо сохранить
и рационально использовать ресурсы современной биосферы. Нельзя согласиться
с высказываниями отдельных ученых о том, что „человек проигнорировал такое
множество открываемых перед ним перспектив, и поэтому для нашей планеты было
бы полезным его исчезновение с последующим появлением какого-то другого более
высокоинтеллектуального вида” (Fiennes, 1976). Нам надо биосферу сохранить
такой, какой она нужна человеку, а не человека приспосабливать к меняющейся
биосфере.
198
В определении стратегии во взаимодействиях человека с природой все больше
надо учитывать ныне идущий стихийный процесс эволюции. Огромный эмпирический
материал как о негативных воздействиях человека на органический мир, так
и о попытках как-то изменить биосферные процессы в сторону улучшения качественного
состава экосистем, увеличения их продуктивности, повышения скорости накопления
биомассы и общего количества ее выхода с единицы площади и т.д. остается
критически не проанализированным и мало обобщенным. Наши неудачи в рациональном
использовании ресурсов биосферы обусловлены и тем, что неизвестны скорости
и направления эволюции громадного числа видов в современных условиях. Не
изучены воздействия на биосферные процессы не только эволюционных преобразований
отдельных видов, но и всего органического мира в целом. Мы не знаем, перестала
ли биосфера быть саморегулируемой системой и какие ее механизмы и в каких
размерах требуют корректирующего влияния со стороны человека.
Необходимо выработать такую систему воздействия человека на окружающую среду,
чтобы удовлетворение его потребностей не только не наносило бы ущерба биосфере,
но, напротив, способствовало бы повышению ее продуктивности и устойчивости
(Камшилов, 1974). При этом невозможен путь, связанный с консервацией биосферы.
Индустриализацию нельзя сдержать не только потому, что она является основой
повышения благосостояния человека, но и потому, что неиндустриальные методы
ведения хозяйства при современной численности населения наносят еще больший
ущерб окружающей среде (Дювиньо,1979). Современное воздействие человека на
органический мир, как отмечалось выше, глубоко противоречиво. С одной стороны,
оно ведет к резкому снижению численности и вымиранию многих видов, к общему
ухудшению природной среды. Природные ценозы, испытавшие воздействие человека,
как правило, упрощаются, их видовое разнообразие уменьшается, а продуктивность
снижается на несколько порядков величин. С другой стороны, как отмечал С.
С. Шварц (1976), некоторые изменения следует рассматривать как факторы, благоприятствующие
развитию биосферы. Так, рост концентрации CО2 в атмосфере является предпосылкой
для повышения продуктивности растительных сообществ. Человек резко увеличивает
энергетический обмен в биосфере, что, по мнению Шварца, также способствует
ее „омоложению” за счет ускорения процесса эволюции и развития прогрессивных
биоценозов. К этим результатам приводят многие мероприятия по обводнению
пустынь, работы по мелиорации, осушению болот и т. д. Агроценозы, созданные
в результате этих мероприятий, иногда отличаются повышенной продуктивностью
по сравнению с биоценозами, ранее существовавшими на этих местах. Тем самым
в отдельных регионах биосферы деятельность человека могла бы способствовать
увеличению ее продуктивности. Однако необдуманность таких мероприятий, в
частности, узковедомственный подход к их проведению, чаще ведет к деградации
экосистем и к резкому падению их продуктивности.
Рост общественного производства в целом соответствует общей тенденции в эволюции
биосферы в сторону усиления преобразующего
199
влияния жизни на косные компоненты биосферы, ускорения биогенной миграции
химических элементов в биосфере и увеличения ее энерговооруженности. Как
писал А. Лотка (Lotka, 1925, р. 245) „общая тенденция достижений человечества
идет вперед к циркуляции все большего количества материи, к увеличению круговорота
жизни. Можно не сомневаться, что эта тенденция будет продолжена и усилена
в будущем, так как она является одним из выражений более общего закона природы”.
К настоящему времени предложено несколько возможных путей управляемой эволюции
(Берман и др., 1967; Дювиньо, Танг, 1973; Камшилов, 1974; Шварц, 1976; Лаптев,
1982, и др.). Наиболее важными и осуществимыми в настоящее время являются
следующие: 1) расширение числа доместицируемых и селектируемых видов; 2)
направленное изменение свободноживущих видов; 3) повышение продуктивности
природных биогеоценозов за счет интродукции видов; 4) создание искусственных
ценозов; 5) охрана основных типов природных биогеоценозов для сохранения
видового разнообразия биосферы и ее суммарного „генофонда” как источника
для будущей селекционной работы.
Расширение числа одомашненных животных и культурных растений является наиболее
показательным примером способности человека контролировать эволюцию. Сейчас
несколько тысяч видов животных, растений и микроорганизмов находятся на различных
стадиях доместикации. Если еще совсем недавно „пекарские” дрожжи были чуть
ли не единственным домашним видом микроорганизмов, то сейчас буквально сотни
и тысячи их штаммов используются в медицинской промышленности для получения
лекарственных препаратов. Весьма широк и спектр мероприятий по вовлечению
животных в процессы частичной и полной доместикации. Он простирается от мер
по регуляции численности некоторых видов в природе и частичной селекции свободноживущих
видов с целью их приспособления к измененным человеком биогеоценозам до создания
специальных хозяйств по разведению и селекции ранее диких форм. Благодаря
регуляции численности многих промысловых видов птиц и млекопитающих удалось
восстановить их численность на уровне, не только не угрожающем вымиранием,
но и позволяющем вести их регулярный отстрел. Это нередко расширяет продовольственную
базу человечества. Оказалось, что в условиях некоторых природных биоценозов
(пустыни, полупустыни, саванны) регулируемая охота на диких животных обеспечивает
более высокий уровень продукции мяса с единицы площади, чем содержание в
этих районах сельскохозяйственного скота (Huxley, 1962).
Охрана и регуляция численности свободноживущих животных нередко идут параллельно
с попытками включить их в процессы непосредственного сельскохозяйственного
производства. Создаются целые производственные комплексы по одомашниванию,
разведению и использованию таких животных, как антилопы, маралы, овцебыки,
лоси, зубры, бегемоты, крокодилы, змеи и т. д. Сотни видов насекомых разводятся
для борьбы с вредителями сельскохозяйственных растений и леса. Эксплуатация
рыбных запасов в водоемах и их восстановление в значи-
200
тельной степени стали зависеть от рыбоводных заводов, где выводятся и выращиваются
до определенного возраста мальки наиболее ценных рыб.
Сильно увеличилось и число животных, вовлеченных в новую отрасль сельского
хозяйства – звероводство. Десятки видов пушных зверей разводятся сейчас на
зверофермах, а их вклад в общую стоимость заготовляемой пушнины во много
раз превышает стоимость пушнины, добываемой путем охоты (Глембоцкий, 1972).
Селекционная работа с вновь доместицируемыми видами ведет не только к усилению
хозяйственно ценных признаков, но и к выработке наследственно закрепленных
новых стереотипов поведения (Беляев, 1972; Трут, 1978, и др.). Это дает основание
надеяться на превращение этих животных в домашние.
Несмотря на успехи в доместикации, следует учитывать, что подавляющее большинство
ныне существующих видов находится вне этого процесса. В ближайшем будущем
не приходится надеяться на фронтальное преобразование органического населения
биосферы путем полной доместикации морских и сухопутных организмов. И хотя
в доместикационных процессах человек ближе всего подошел к управляемой эволюции
как процессу создания „новых форм (штаммов, пород, сортов), обладающих нужным
человеку сочетанием морфологических и физиологических свойств, адаптированных
к экологическим условиям и стойко передающих свои полезные для человека особенности
потомству” (Берман и др., 1967, с. 475), все же приходится искать и другие
пути для сохранения многообразия органических форм.
Направленное изменение свободноживущих видов, по мнению А. В. Яблокова (1978),
является одним из таких путей. В качестве примеров он называет включение
в круг синантропных видов все большего числа позвоночных, сосуществующих
с человеком на полностью урбанизированных территориях. Эти формы прошли отбор
на выживание особей, не только не боящихся соседства с человеком, но и склонных
вступать с ним в контакты (белки в парках). Примерами синантропных растений
могут быть льняной рыжик, клевер ползучий, подорожник и т. д. Сейчас, однако,
трудно сказать, насколько обширным может оказаться круг животных и растений,
способных существовать как свободноживущие виды на урбанизированных территориях.
Другой путь связан с внедрением отдельных видов в новые для них природные
биогеоценозы с целью повышения их продуктивности. За полвека в СССР интродуцировано
около 50 видов промысловых зверей и 8 видов птиц, которые неизбежно испытали
микроэволюционные изменения в новых условиях.
Восстановление разрушенных природных биоценозов и создание искусственных
является также важным способом управления эволюцией. Всякий вид может существовать
только в рамках экосистемы. И эволюция видов в сторону приспособления к человеку
и результатам его деятельности возможна только при определенных преобразованиях
самих экосистем. До сих пор главным направлением в этих преобразованиях было
постепенное превращение природных биоценозов в полуприродные, а последних
– в агроценозы. При этом вновь формирующиеся
201
ценозы должны превосходить природные не только по продуктивности и отношению
продуктивности к биомассе, но и по устойчивости. И если первые показатели
в целом уже достигнуты во многих сельскохозяйственных угодьях, то достижение
последних является задачей будущего. У антропогенных экосистем, как правило,
понижена способность к саморегуляции и циклическому возобновлению основных
компонентов. Однако иногда вновь возникшие ценозы, например на орошаемых
землях бывших пустынь, отличаются от исходных природных ценозов сложностью
видового состава и повышенной продуктивностью. Здесь возможно формирование
экосистем со всеми характерными для них свойствами саморегуляции, циклического
возобновления и т. д.
И наконец, как это ни странно звучит, охрана природных биогеоценозов путем
создания сети заказников и заповедников по-прежнему остается одним из главных
путей управления эволюцией. Каждый вид уникален в генетическом отношении,
и его вымирание означает потерю генетической информации, которая вряд ли
в будущем может быть восстановлена. Поэтому важно принять все меры для сохранения
каждого вида. Человек должен сохранить не только нынешнее видовое разнообразие,
но и экосистемы в том виде, в котором они сложились до антропогенных воздействий.
„Бережное сохранение экосистем и функционирующих в них популяций и видов
абсолютно необходимо” (Ehrlich, Money, 1983, p. 252), так как человечество
сейчас еще не в состоянии предусмотреть последствия для природных экосистем,
вызванные вымиранием того или иного вида.
Не удастся, конечно, полностью изолировать эти заповедные участки биосферы
от происходящих в ней глобальных перемен. Так или иначе человек вынужден
вмешиваться в биоценотические процессы на территории заповедников. Это вмешательство
также ведет к микроэволюции живущих в заповедниках видов, но темп и масштабы
этих изменений значительно меньше адаптивных преобразований видов на урбанизированных
территориях.
В заключение следует еще раз подчеркнуть, что влияние прямых или косвенных
воздействий человека на адаптивные преобразования организмов предопределяют
особенности эволюции в условиях антропозойной формации. Но в целом этот процесс
продолжает происходить стихийно, регулируясь преимущественно действием статистических
сил при направляющем участии естественного отбора. С последним человек вынужден
считаться даже в селекционных процессах. Однако само развитие взаимоотношений
человека со средой требует перехода к эволюции отдельных видов и экосистем,
контролируемой человеком. Несмотря на все „мнимые и реальные” трудности в
управлении эволюцией (Сутт, 1978), человек должен направить эволюцию живого
по пути ноогенеза, т. е. по пути разумного управления обменом веществ между
биосферой и человечеством (Камшилов, 1970). Эволюция организмов, контролируемая
человеком, будет одной из важнейших предпосылок для преобразования биосферы
в ноосферу в смысле Вернадского.
Более сорока лет тому назад Вернадский, отмечая качественный перелом в истории
биосферы, писал: „Мы живем в исключительное
202
время в истории нашей планеты, в психозойскую эру, когда создается новое
ее состояние — ноосфера и когда геологическая роль человека начинает господствовать
в биосфере и открываются горизонты его будущего развития” (1965, с. 270).
От самого человека зависит, удастся ему обеспечить более или менее гармоничный
переход биосферы в ноосферу или же своей неразумной деятельностью он поставит
под угрозу собственное существование.
Одним из главных показателей начала эры ноогенеза станет резкое увеличение
численности видов растений и животных, эволюция которых будет поставлена
под прямой контроль человека. Вероятно, это будут не только окультуренные
и одомашненные виды, но и виды, существующие в составе природных экосистем.
Практические успехи в управлении эволюцией видов и экосистем будут служить
лучшей проверкой достигнутой глубины знаний в изучении особенностей органической
эволюции в современной биосфере.
203
ПОСЛЕСЛОВИЕ
Работая над этой книгой, мы исходили из того, что история науки особенно
полезна в тех случаях, когда исследуется развитие наиболее актуальных проблем.
Их историко-критический анализ может помочь при выборе направлений современных
исследований и позволяет лучше понять тенденцию развития проблемы в ближайшем
будущем. Не вызывает сомнения, что вопрос, ставший предметом этих очерков,
является первоочередным не только для эволюционной теории, но и всего комплекса
естественных наук, так или иначе связанных с изучением биосферы. Успехи в
изучении закономерностей эволюции биосферы небезразличны и для решения целого
ряда глобальных проблем, стоящих перед современным человечеством.
Мы надеемся, что данная книга — это только начало историко-критических исследований
проблемы эволюции биосферы, разработка многообразных аспектов которой возможна
лишь при участии историков науки различной специальности. Цель этой книги
заключалась прежде всего в том, чтобы доказать плодотворность синтеза эволюционной
теории с учением Вернадского, необходимость которого не раз подчеркивали
М. С. Гиляров, К. М. Завадский, М. М. Камшилов, Н. В. Тимофеев-Ресовский,
Дж. Хатчинсон, С. С. Шварц и многие другие крупные эволюционисты. Только
изучение преобразований таких параметров биосферы, как продуктивность, „энерговооруженность”,
доступность минеральных веществ и скорость их оборота в биогеохимических
циклах позволяет объяснить прогрессивную эволюцию одних форм и сохранение
других на более низком уровне. Уместно привести здесь заключительные слова
посмертно изданной книги Шварца: „Имеется достаточно оснований рассматривать
эволюцию как процесс прогрессивной экспансии жизни на нашей планете, совершающийся
на основе создания в ходе филогенеза отдельных групп новых экологических
ниш. Таким путем в круговорот биосферы вовлекаются все новые и новые потенциальные
среды жизни, повышается продуктивность и стабильность отдельных биогеоценозов
и биогеоценотического покрова Земли в целом и создаются предпосылки для создания
все новых экологических ниш. Это гарантирует бесконечность эволюции в пространстве
и во времени. Но первооснова этого глобального по своей сути процесса заключается
в ароморфной специализации отдельных видов и групп животных. Истоки этого
процесса отчетливо просматриваются уже на внутривидовом уровне. Связь микро-
и макроэволюционного процесса делается, таким образом, очевидной” (1980,
с. 253).
204
Ближайшей задачей эволюционной теории является изучение воздействия микро- и макроэволюции на исторические преобразования биосферы, на ее биогеохимическую и энергетическую эволюцию. Создание общей теории эволюции биосферы станет возможным лишь в результате интенсивной разработки биогеохимических, экологических и информационных концепций в эволюции биосферы. Приоритет в создании этих концепций принадлежит отечественной науке. Здесь же предприняты и первые попытки синтеза этих концепций с современной эволюционной теорией. Однако, осуществление этого синтеза является задачей будущего.
205
ЛИТЕРАТУРА
Алексеев А. П. Становление человечества. М., 1984. 462 с.
Анатольева А. И. Домезозойские красноцветные формации. Новосибирск, 1972.
346 с.
Анатольева А. И. Эволюция континентального красноцветного породообразования
в истории Земли // Эволюция осадочного процесса в океанах и на континентах.
М., 1983. С. 121 – 128.
Андрусов Н. И. Бактериология и геология, их взаимные отношения // Учен,
зап. Юрьев, ун-та. 1897. № 1. С. 1 – 20.
Анри В. Энергетика жизни // Природа. 1917. № 4. С. 443 – 456.
Аристовская Т. В. Теоретические проблемы численности и продуктивности почвенных
бактерий // Вопросы численности биомассы и продуктивности почвенных микроорганизмов.
Л., 1972. С. 7 – 20.
Арманд Д. Л. Наука о ландшафтах. М., 1975. 287 с.
Архангельский А. Д. Яков Владимирович Самойлов // Самойлов Я. В. Биолиты.
Л., 1929. С. 5 – 11.
Базилевич Н. И. Биогеохимия Земли и функциональные модели обменных процессов
природных экосистем // Тр. Биогеохим. лаб. 1979. Т. 17. С. 55-73.
Базилевич Я. И., Родин Л. Е., Розов Н. Н. Географические аспекты
изучения биологической продуктивности // Материалы V съезда Геогр. о-ва
СССР. М.,
1970, 28 с.
Базилевич Н. И., Родин Л. Е., Розов Н. Н. Сколько весит вещество планеты?
// Природа. 1971. № 1. С. 46 – 53.
Банников А. Г., Флинт Ф. Е. Проблемы слонов в Восточной Африке
// Природа. 1973. №11. С. 87 – 96.
Баринов Г. В. Биосферные ритмы и проблемы сохранения кислородного равновесия
// Журн. общ. биологии. 1972. Т. 33, № 6. С. 771 – 779.
Барнхорт Э. Древнейшие органические остатки на Земле // Природа. 1972.
№ 5. С. 77 – 87.
Бауэр Э. Теоретическая биология. М.; Л., 1935. 206 с.
Бгатов В. И. Земная атмосфера и кора выветривания // Эволюция осадочного
процесса в океанах и на континентах. М., 1983. С. 103 – 117.
Бей-Биенко Г. Я. Мир насекомых и охрана природы // Природа. 1972. № И.
С. 32 – 39.
Бекетов А. Н. География растений. СПб., 1896. 359 с.
Беклемишев В. Н. Об общих принципах организации жизни // Бюл. МОИП. Отд.
биол. 1964. Т. 69, вып. 2. С. 22 – 38.
Белое Н. В. Геохимические аккумуляторы // Тр. Ин-та кристаллографии. 1952.
№7. С. 73 – 80.
Белов Н. В., Лебедев В. И. Источники энергии геохимических процессов //
Природа. 1964. № 5. С. 22 – 28.
Беляев Д. К. Генетические аспекты доместикации животных // Проблемы доместикации
животных. М., 1972. С. 39 – 45.
Берг Л. С. Номогенез или эволюция на основе закономерностей. Пб., 1922.
VIII+ 306 с.
Берман 3. И., Парамонов А. А., Полянский Ю. И. Проблема управления процессами
органической эволюции // Современные проблемы эволюционной теории. Л.,
1967. С. 442 – 478.
Бернал Дж. Проблема стадий в биогенезе // Возникновение жизни
на Земле. М., 1959. С. 42 – 59.
206
Бернал Дж. Возникновение жизни на Земле. М., 1969. 391 с.
Биогеохимические аспекты формирования осадочных пород и руд. Л., 1983. 126
с.
Биосфера // БСЭ. 3-е изд. 1970. Т. 3. С. 364 – 365.
Блюменфельд Л. А. Проблемы биологической физики. М., 1977. 336 с.
Богданов Ю. А., Каплин П. А., Николаев С. Д. Происхождение и развитие океана.
М., 1978. 160 с.
Боголепов К. В., Жарков М. А. Проблемы эволюции геологических процессов //
Проблемы эволюции геологических процессов. Новосибирск, 1981. С. 7 – 19.
Богоров В. Г. Жизнь океана. М., 1969. 64 с.
Богоров В. Г. Биологическая продуктивность океана и особенности ее географического
распределения // Вопр. географии. 1970. № 84. С. 80 – 102.
Бойченко Е. А. Об эволюции окислительных функций биосферы // Геохимия. 1967.
№8. С. 971 – 976.
Бойченко Е. А. Участие металлов в эволюции окислительно-восстановительных
процессов растений // Изв. АН СССР. Сер. биол. 1968. № 1. С. 24 – 33.
Бойченко Е. А. Факторы взаимодействия соединений металлов в эволюции растений
// Изв. АН СССР. Сер. биол. 1984. № 3. С. 403 – 409.
Бойченко Е. А., Грыжанкова Л. Н., Удельнова Т. М. Значение биогенных
комплексов металлов в эволюции биосферы // Разделение элементов и изотопов
в геохимических
процессах. М., 1979. С. 68 – 74.
Бойченко Е. А., Саенко Г. Н., Удельнова Т. М. Эволюция концентрационных функций
растений в биосфере // Геохимия. 1968. № 10. С. 1260 – 1264.
Бойченко Е. А., Саенко Г. Н., Удельнова Т. М. Изменение соотношений металлов
в эволюции растений в биосфере // Очерки современной геохимии и аналитической
химии. М., 1972. С. 454 – 458.
Бойченко Е. А., Удельнова Т. М. Значение металлов в окислительно-восстановительных
функциях растений // Изв. АН СССР. Сер. биол. 1977. № 2. С. 194 – 200.
Бойченко Е. А., Удельнова Т. М., Юферова С. Г. Эволюция восстановительных
функций биосферы // Геохимия. 1969. № 11. С. 1392 – 1396.
Большаков В. Н. Экологическое прогнозирование. М., 1983. 64 с.
Боровик-Романова Т. Ф., Белова Е. А. К биогеохимии лития//Тр. Биогеохим.
лаб. 1974. Т. 13. С. 118 – 143.
Брода П. Плазмиды. М., 1982. 220 с.
Брода Э. Эволюция биоэнергетических процессов. М., 1978. 304 с.
(Бруновский Б. К.) Brunowsky В. Studien iiber die Konzentiation
des Radiums durch Lebenwesen // Тр. Биогеохим. лаб. 1932. Т. 2. С. 9 – 25.
Будыко М. И. Человек и биосфера // Вопр. философии. 1973. № 1. С. 51 – 70.
Будыко М. И. Экологические факторы эволюции // Журн. общ. биологии. 1975.
Т. 36, №1. С. 36 – 48.
Будыко М. И. Глобальная экология. М., 1977а. 328 с.
Будыко М. И. Прошлое и будущее биосферы // Вести. АН СССР. 19776. № 12. С.
70 – 79.
Будыко М. И. Эволюция биосферы. Л., 1984. 488 с.
Будыко М. И., Ронов А. Б. Эволюция химического состава атмосферы в фанерозое
// Геохимия. 1979. № 5. С. 643 – 653.
Будыко М. И., Ронов А. Б., Яншин А. Л. История атмосферы. Л., 1985. 207 с.
Бунак В. В. Род Homo, его возникновение и последующая эволюция. М., 1980.
321с.
Буханан Дж. Речь председателя // Происхождение предбиологических систем.
М., 1966. С. 109 – 112.
Быков Б. А. Геоботаника. Алма-Ата, 1957. 371 с.
Быков Б. А. Введение в фитоценологию. Алма-Ата, 1970. 234 с.
Быков Б. А. Биоценозы и эволюция // Флора и растительные ресурсы Казахстана.
Алма-Ата, 1975. С. 23 – 35.
Быховер Н. А. Распределение мировых ресурсов минерального сырья по эпохам
рудообразования. М., 1963. 474 с.
Вавилов Н. И. Центры происхождения культурных растений. Л., 1926.
248 с.
207
Вавилов Н. И. Селекция как наука. М.; Л., 1934. 16 с.
Вайскопф В. Физика в двадцатом столетии. М., 1977. 269 с.
Вальтер И. История Земли и жизни. СПб., 1911. 447 с.
Валяшко М. Г. Эволюция химического состава воды океана // История Мирового
океана. М., 1971. С. 97 – 104.
Вассоевич Н. Б. Основные закономерности, характеризующие органическое вещество
современных и ископаемых осадков // Природа органического вещества современных
и ископаемых осадков. М., 1973. С. 11 – 60.
Вассоевич Н. Б. Различное толкование понятия „биосфера” // Исследования
органического вещества современных и ископаемых осадков. М., 1976. С. 381—399.
Вассоевич Н. Б. Учение о биосфере // Изв. АН СССР. Сер. геол. 1977. № 1.
С. 5 – 13.
Вассоевич Н. Б. Представление В. И. Вернадского о происхождении нефти. М.,
1984. 20с.
Вассоевич Н. Б., Гусева А. Н., Тараненко Е. И. К проблеме нефте- и газообразования
// Природа органического вещества современных и ископаемых осадков. М., 1973.
С. 150 – 157.
Вассоевич Н. Б., Иванов А. Н. К истории учения о биосфере // Методология
и история геологических наук. М., 1977. С. 57 – 94.
Вассоевич Н. Б., Иванов А. Н. О различном понимании биосферы // Палеонтология
и эволюция биосферы. Л., 1983а. С. 17 – 33.
Вассоевич Н. Б., Иванов А. Н. О биосфере и мегабиосфере // Журн. общей биологии.1983б.
Т. 44, № 3. С. 291 – 304.
Вассоевич Н. Б., Иванов А. Н. О живом веществе, подразделениях биосферы и
биосферологии // Журн. общ. биологии. 1984. Т. 45, № 4. С. 435 – 448.
Вассоевич Н. Б., Конюхов А. И., Лопатин Н. В. Общее и особенное в образовании
углей, нефти и углеводородных газов // Горючие ископаемые. М., 1976. С. 7
– 19.
Верзилин Н. Н. Методы палеогеографических исследований. Л., 1977а. 129 с.
Верзилин Н. Н. Роль живого вещества в образовании земной коры //
Вести. ЛГУ. Сер. геол. 19776. № 24. С. 5 – 13.
Верзилин Н. Н. Проблема изменения содержания свободного кислорода в атмосфере
Земли в фанерозое // Вестн. ЛГУ. Сер. геол. 1980. № 24. С. 12 – 20.
Верзилин Н. Н., Верзилин Н. Н., Верзилин Н. М. Биосфера, ее настоящее, прошлое,
будущее. М., 1976. 224 с.
Вернадский В. И. Генезис .минералов // Энцикл. словарь / Брокгауз и Ефрон.
1892. Т. 8. С. 306 – 311.
Вернадский В. И. К вопросу о химическом составе почв // Почвоведение. 1913.
№2 – 3. С. 1 – 21.
Вернадский В. И. Об использовании химических элементов в России // Русская
мысль. 1916. № 1. С. 73 – 88.
Вернадский В. И. Записка (о необходимости организации химического изучения
организмов) // Протоколи зсiдання фiз.-мат. виддiлу Укр. АН у Юев.
Юев. 1919. Вып. 1.С. 43 – 46.
Вернадский В. И. О геохимическом анализе почв // Бюл. III Всерос. съезда
почвоведов в Москве. М., 1921а. № 3 – 4. С. 1 – 4.
Вернадский В. И. Записка об изучении живого вещества с геохимической точки
зрения // Изв. Рос. АН. Сер. 6. 19216. Т. 15, № 1 – 18. С. 120 – 123.
Вернадский В. И. Начало и вечность жизни. Пг., 1922а. 58 с.
Вернадский В. И. Химический состав живого вещества в связи с химией земной
коры. Пг., 19226. 48 с.
Вернадский В. И. Живое вещество в химии моря. Пг., 1923. 36 с.
Вернадский В. И. од жизни в биосфере // Природа. 1925. № 10 – 12. С. 26
– 38.
Вернадский В. И. Биосфера. Л., 1926а. 148 с.
Вернадский В. И. Определение геохимической энергии (величины Д, v, s) некоторых
групп насекомых. Л., 19266. 12с.
Вернадский В. И. Определение геохимической энергии (величины Д, v, s) однолетних
цветковых растений. Л., 1926в. 9 с.
208
Вернадский В. И. О размножении организмов и его значении в механизме биосферы
// Изв. АН СССР. Сер. 6. 1926 г. Т. 20, № 10. С. 697 – 726; № 12. С.
1053 – 1060.
Вернадский В. И. Очерки геохимии. М.; Л., 1927а. 368 с.
Вернадский В. И. Бактериофаг и скорость передачи жизни в биосфере //
Природа. 19276. № 6. С. 443 – 446.
Вернадский В. И. О проделанной Биогеохимической лабораторией работе по
анализу живого вещества // Изв. АН СССР. Сер. 6. 1927в. Т. 21, № 18.
С. 1589 – 1591.
Вернадский В. И. Эволюция видов и живое вещество // Природа. 1928. №
3. С. 227 – 250.
Вернадский В. И. О геохимических постоянных культурных растений // Докл.
АН СССР. Сер. А. 1929. № 2. С. 38 – 40.
Вернадский В. И. Об условиях появления жизни на Земле // Изв. АН СССР.
Сер. 7. 1931. №5. С. 633 – 653.
Вернадский В. И. История минералов земной коры. Т. 2. История природных
вод. Ч. 2, вып. 1. Л., 1933. 202 с.
Вернадский В. И. Очерки геохимии. М., 1934. 380 с.
Вернадский В. И. О пределах биосферы // Изв. АН СССР. Сер. геол. 1937.
№ 1. С. 3 – 24.
Вернадский В. И. Биогеохимические очерки. М.; Л., 1940. 250с.
Вернадский В. И. О геологических оболочках Земли как планеты // Изв.
АН СССР. Сэр. геогр. и геофиз. 1942. № 6. С. 251 – 262.
Вернадский В. И. есколько слов о ноосфере // Успехи соврем, биологии.
1944. Т. 18, вып. 2. С. 113 – 120.
Вернадский В. И. Избранные соч.: В 5-ти т. М.; Л., 1954. Т. 1. 696 с.
Вернадский В. И. мическое строение биосферы Земли и ее окружения. М.,
1965. 374 с.
Вернадский В. И. Размышления натуралиста. Кн. 2: Научная мысль как планетное
явление. М., 1977. 191 с.
Вернадский В. И. Живое вещество. М., 1978. 358 с.
Вернадский В. И. Проблемы биогеохимии // Тр. Биогеохим. лаб. 1980. Т.
16. 320с.
Вернадский В. И. Об участии живого вещества в создании почв // Сытник
К. М., Апанович Е. М., Стойко С. М. Вернадский: жизнь и деятельность
на Украине.
Киев, 1988. С. 186 – 193.
Вернадский В. И. Биосфера и ноосфера. М., 1989. 261 с.
Вернадский В. И. , Виноградов А. Я. О химическом элементарном составе
рясок (Lemna) как видовом признаке // Докл. АН СССР. 1931. Сер. А. №
6. С. 148
– 158.
Вернадский В. И. , Ферсман А. Е. Предисловие // Самойлов Я. В. Биолиты.
Л., 1929. С. 3.
В. И. Вернадский и современность. М., 1986. 230 с.
В. И. Вернадский и современная наука. Л., 1988. 112с.
Вильямс В. Р. Почвоведение. М.; Л., 1926. Ч. 1. 323 с.; Ч. 2. 448 с.
Вильямс В. Р. Избранные произведения: В 2-х т. М., 1950. Т. 1. 790 с.
Вильямс В. Р. Почвоведение: Земледелие с основами почвоведения // Собр.
соч.: В 10-ти т. М., 1951. Т. 6. С. 3 – 576.
Винберг Г. Г. Первичная продукция водоемов. Минск, 1960. 329 с.
Винберг Г. Г. Биологическая продуктивность водоемов // Экология. 1983.
№ 3. С. 3 – 12.
Винер Н. Кибернетика или управление и связь в животном и машине. М.,
1958. 281с.
Виноградов А. П. Химический состав планктона Екатерининского пруда в
Детском селе близ Ленинграда // Докл. АН СССР. Сер. А. 1929. № 2. С.
41 – 42.
Виноградов А. П. Химический элементарный состав морских организмов в
связи с вопросами их систематики и морфологии // Природа. 1931. № 6.
С. 230—254.
Виноградов А. П. Геохимия живого вещества. Л., 1932. 67 с.
Виноградов А. П. Химический элементарный состав организмов и периодическая
система Д. И. Менделеева // Природа. 1933. № 8 – 9. С. 28 – 36.
209
Виноградов А. П. Химический элементарный состав организмов моря. Ч. 1 –
3 // Тр. Биогеохим. лаб. 1935а. Т. 3. С. 53 – 278; 1937. Т. 4. С. 5 – 226;
1944. Т. 6. С. 5 – 273.
Виноградов А. П. Химический элементарный состав организмов и периодическая
система Д. И. Менделеева // Тр. Биогеохим. лаб. 19356. Т. 3. С. 5 – 30.
Виноградов А. П. Биогеохимические провинции и эндемии // Докл. АН СССР. 1938.
Т. 18, № 4 – 5. С. 283 – 286.
Виноградов А. П. О причине отсутствия известковых (из СаСО3) скелетов у докембрийских
беспозвоночных // Докл. АН СССР. 1940. Т. 27, № 3. С. 231 – 233.
Виноградов А. П. К химическому познанию биосферы // Почвоведение. 1945. №
3. С. 348 – 354.
Виноградов А. П. Геохимическая история кислорода и фотосинтеза // Изв. АН
СССР. Сер. биол. 1947. № 3. С. 409 – 422.
Виноградов А. П. Возникновение биосферы // Возникновение жизни на Земле.
М., 1959а. С. 28 – 41.
Виноградов А. П. Химическая эволюция Земли. М., 19596. 44 с.
Виноградов А. П. О генезисе биогеохимических провинций // Тр. Биогеохим.
лаб. 1960. Т. 11. С. 3 – 7.
Виноградов А. П. Биогеохимические провинции и их роль в органической эволюции
// Геохимия. 1963. № 3. С. 199 – 213.
Виноградов А. П. Введение в геохимию океана. М., 1967. 215 с.
Виноградов А. П. Геохимия // БСЭ. 3-е изд. 1971. Т. 6. С. 330 – 336.
Виноградов А. П. Фотосинтез и биосфера // Современные проблемы фотосинтеза.
М., 1973. С. 7 – 14.
Виноградов А. П., Тейс Р. В. Изотопный состав кислорода разного происхождения
// Докл. АН СССР. 1941. Т. 33, № 9. С. 497 – 501.
Виноградов В. И. Сколько лет океану? // Природа. 1975. № 12. С. 50 – 57.
Виноградов В. И., Иванов П. В., Мизарев М. А. и др. О возрасте кислородной
атмосферы Земли // Докл. АН СССР. 1969. Т. 188, № 5. С. 1144 – 1147.
(Виноградский С. Н.) Winogradsky S. Ueber Eisenbacterien // Botan. Zeitung,
1888. Bd 46, N17. S. 261 – 270.
Вишаренко В. С. Детерминация в биологических процессах. Л., 1975. 88 с.
Владышевский Д. В. Морфологические реакции птиц на изменение условий существований
в антропогенных ландшафтах // Проблемы эволюции. Новосибирск, 1973. Т. 3.
С. 242 – 248.
Водопьянов П. А. Устойчивость и динамика биосферы. Минск, 1981. 248 с.
Войнар А. И. Биологическая роль микроэлементов в организме животных и человека.
М., 1960. 544 с.
Войткевич Г. В., Холодков Ю. И. Следы древней жизни на Земле. Ростов н/Д,
1976. 100 с.
Вологдин А. Г. Земля и жизнь. М., 1976. 240 с.
Вольскис Р. С. К проблеме вида и эволюции // Материалы II совещания
представителей национальных комитетов МАБ социалистических стран и 12 (20)
заседаний Сов.РГ
по проекту № 8 Б: Вид и его продуктивность в ареале. Вильнюс, 1982. С. 18
– 29.
Вольскис Р. С. Вид как элемент биоты // Система интеграции вида. Вильнюс,
1986. С. 254 – 270.
Вольфкович С. И. Жизнь и деятельность Я. В. Самойлова // Самойлов Я. В. –
минералог и биогеохимик. М., 1974. С. 5 – 12.
Воронков М. Г., Кузнецов И. Г. Кремний в живой природе. Новосибирск, 1984.
158с.
Вудвелл Дж. Круговорот энергии в биосфере // Биосфера. М., 1972. С. 41 –
60.
Высоцкий Б. П. Проблемы истории и методологии геологических наук. М.,
1977. 280с.
Гавриленко Е. С. Гидрогеология тектоносферы. Киев, 1975. 196 с.
Галл Я, М., Колчинский Э. И. К постановке проблемы эволюции закона борьбы
за существование // Закономерности прогрессивной эволюции. Л., 1972. С. 66
– 71.
210
Гаузе Г. Ф. О процессах уничтожения одного вида другим в популяциях инфузорий
// Зоол. журн. 1934а. Т. 13, вып. 1. С. 18 – 26.
(Гаузе Г. Ф.) Gause G. F. The struggle for existence. Baltimore,
1934б. 163 p.
Гаузе Г. Ф. Исследование над борьбой за существование в смешанных популяциях
// Зоол. журн. 1939. Т. 18, вып. 4. С. 557 – 571.
Гаффрон Г. Эволюция фотосинтеза // Тр. V Междунар. биохим. конгр.:
Эволюционная биохимия. Симпоз. 3. М., 1962. С. 116 – 131.
Гегамян Г. В. О биосферологии В. И. Вернадского // Журн. общ. биологии.
1980. Т. 41, №4. С. 581 – 594.
Гегамян Г. В. Ламарк, Вернадский и биосферология // Природа. 1981. № 9.
С. 78 – 81.
Гендерсон Л. Ж. Среда жизни. М.; Л., 1924. 197 с.
Геологический словарь. М., 1973. Т. 1. 486 с.; Т. 2. 456 с.
Геосферы // БСЭ. 3-е изд. М., 1971. Т. 6. С. 323.
Геохимия // БСЭ. 3-е изд. М., 1971. Т. 6. С. 330 – 336.
Герасимов И. П. Палеогеографическое значение учения В. Р. Вильямса о едином
почвообразовательном процессе // Пробл. физ. географии. 1951. № 16. С. 7
– 15.
Гиляров А. М. Эволюция на уровне экосистем // Журн. общ. биологии. 1973.
Т. 34, № 1.С. 13 – 20.
Гиляров М. С. Почвенные беспозвоночные как фактор плодородия почв //
Журн. общ. биологии. 1960. Т. 21, № 2. С. 81 – 88.
Гиляров М. С. Закономерности приспособления членистоногих к жизни на
суше. М„ 1970. 276 с.
Гиляров М. С. Биогеоценология и теория естественного отбора // Журн. общ.
биологии. 1980. Т. 41, № 3. С. 321 – 331.
(Гиляров М. С.) Gilarow М. S. Invited discussion remark // Abh. Akad. Wiss.
DDR. Abt. Math.-Naturwiss.-Techn. 1983a. N 1. S. 83 – 84.
(Гиляров М. C.) Gilarow M. S. The formation of vegetable mould through the
actions forms. Its methodological premices // Abh. Akad. Wiss. DDR. Abt.
Math.-Naturwiss.-Techn. 1983б. N 1. S. 35 – 47.
Гирусов Э. В. Система „общество-природа” (Проблемы социальной экологии). М., 1976. 167 с.
Глазовская М. А. Почвы мира: Основные семейства и типы почв. М., 1972.
Т. 1. 231 с.
Глембоцкий Я. Л. Роль генетики в селекции животных. М., 1972. 48 с.
Глесснер
М. Древнейшие бесскелетные организмы // Природа. 1963/№ И.С. 73 – 78.
Голубец М. А. Актуальные вопросы экологии. Киев, 1982. 158 с.
Голубец М. А., Одинок А. П. Современные представления о биогеоценотическом
покрове // Биогеоценотический покров Бескид и его динамические тенденции.
Киев, 1983. С. 5 – 19.
Гольдшмидт В. Принципы распределения химических элементов в минералах и горных
породах // Успехи химии. 1938. Т. 7, вып. 2. С. 288 – 320.
Гордеев Д. И. История учения о роли биосферы в развитии земной коры.// Тр.
3-го Междунар. конгр. по истории науки. Секция 8. М., 1974а. С. 84 – 87.
Гордеев Д. И. Я. В. Самойлов – один из основоположников биогеохимии // Я.
В. Самойлов – минералог и биогеохимик. М., 19746. С. 17 – 18.
Григорьев А. А. О некоторых взаимоотношениях основных элементов физико-географической
среды и их эволюция // Пробл. физ. географии. М.; Л., 1936.
Вып. 3. С. 3 – 30.
Григорьев А. А. Закономерности строения и развития географической среды.
М., 1966. 382с.
Гумбольдт А. География растений. М., 1936. 230с.
Гумбольдт А. Картины природы. М., 1959. 270 с.
Давиташвили Л. Ш. Развитие идей и методов в палеонтологии после Дарвина.
М.; Л., 1940. 263 с.
Давиташвили Л. Ш. Эволюция условий накопления горючих ископаемых в связи
с развитием органического мира. М., 1971. 296 с.
211
Дарвин Ч. (1881). Образование растительного слоя Земли деятельностью
червей и наблюдения над их образом жизни // Соч.: В 9-ти т. М.; Л., 1936.
Т. 2.
С. 115 – 138.
Дарвин Ч. (1859). Происхождение видов путем естественного отбора
// Соч.: В 9тат. М.; Л., 1939. Т. 3. С. 171 – 832.
Деборин А. М. Проблема времени в освещении акад. Вернадского
// Изв. АН СССР. 1932. Сер. 7. № 4. С. 543 – 569.
Деборин А. М. Критические замечания на критические замечания
акад. В.И.Вернадского // Изв. АН СССР. 1933. Сер. 7. № 3. С. 409 – 419.
Дегенс Э. Геохимия осадочных образовании. М., 1967. 299 с.
Джохансон Д. Люди [sic – П. В.]: Истоки рода человеческого.
М„ 1984. 295 с.
Добродеев О. П. Живое вещество и оледенения Земли // Природа.
1975. № 6. С. 67 – 71.
Докучаев В. В. Русский чернозем. СПб., 1883. 376 с.
Докучаев В. В. Наши степи прежде и теперь. СПб., 1892. 128 с.
Докучаев В. В. К учению о зонах природы: Горизонтальные и вертикальные
почвенные зоны. СПб., 1899. 28 с.
Дроздова Т. В. Геохимия аминокислот. М., 1977. 196 с.
Дроздова Т. В., Соколов Б. С. Палеобиогеохимия // Тр. Биогеохим.
лаб. 1979. Т. 17. С. 37 – 45.
Друшиц В. В., Шиманский В. Н., Соловьев А. Н. Особенности перестроек
биосферы в фанерозое // Палеонтология и эволюция ноосферы. Л., 1983.
С. 78 – 87.
Дубров А. П. Геомагнитное поле и жизнь. Л., 1974. 175 с.
Дыпис Н. В. Учение В. И. Вернадского о биосфере и его значение
для развития биогеоценологии // Бюл. МОИП. Отд. биол. 1978. Т. 83, №
1. С. 62 – 68.
Дювиньо П. Ноосфера и будущее растительности земного шара //
Отчет 12-го Междунар. ботан. конгр. Л., 1979. С. 76 – 96.
Дювиньо П., Танг М. Биосфера и место в ней человека. М., 1973.
270 с.
Емец В. М., Жулидов А. В. Видовая и половая специфичность в накоплении
микроэлементов у жуков-мертвоедов (Coleoptera, Silphadae) рода Silpha
// Журн. общ. биологии.
1983. Т. 44, № 5. С. 553 – 558.
Ермоленко М. Т. К критике теоретико-эволюционных основ тейярдизма //
История и теория эволюционного учения. Л., 1975. Вып. 3. С. 152 – 161.
Ефимов Ю. И. Специфичность закономерностей антропосоциогенеза // Вопр.
философии. 1969. № 7. С. 120 – 130.
Ефремов Ю. К. Ландшафтная сфера Земли // Изв. Всесоюз. геогр. о-ва. 1959.
Т. 91, вып. 6. С. 525 – 528.
Жарков М. А. Классификация соленосных формаций и эволюция соленакопления
в фанерозое // Эволюция литогенеза в истории Земли. Новосибирск, 1981.
С. 4 – 12.
Жуковский П. М. Мировой генофонд растений для селекции. Л., 1970. 87
с.
Жулидов А. В., Емец В. М. О половых различиях в накоплении тяжелых металлов
у водных жуков // Журн. общ. биологии. 1981. Т. 42, № 4. С. 583 – 586.
Забелин И. М. Физическая география и наука будущего. М., 1963. 112 с.
Забелин И. М. Физическая география в современном естествознании: Вопросы
истории и теории. М., 1978. 335 с.
Завадский К. М. К пониманию прогресса в органической природе // Проблема
развития в природе и обществе. М.; Л., 1958. С. 78 – 120.
Завадский К. М. Вид и видообразование. Л., 1968. 404 с.
Завадский К.
М., Колчинский Э. И. Эволюционно-биологические формации // Закономерности
прогрессивной
эволюции. Л., 1972. С. 149 – 156.
Завадский К. М., Колчинский Э. И. Эволюция эволюции: Историко-критические
очерки проблемы. Л., 1977. 236 с.
Завадский К. М., Шапоренко К. К. Реф. кн.: Haldane J. В. S. The causes
of evolution. London, 1932 // Сов. ботаника. 1934. № 2. С. 151 – 159.
Заварзин Г. А. Прокариотные системы в связи с филогенией бактерий //
Журн. общ. биологии. 1979. Т. 40, № 1. С. 5 – 16.
Заварзин Г. А. Бактерии и состав атмосферы. М., 1984. 192 с.
212
Заварзин Г. А., Крылов И. Я. Циано-бактериальные сообщества – колодец в прошлое
// Природа. 1983. № 3. С. 59 – 68.
Захаров И. А. Экологическая генетика и проблемы биосферы. Л., 1984. 32 с.
Земля // БСЭ. 3-е изд. 1972. Т. 9. С. 476 – 494.
Зенкевич Л. А. Очерки по эволюции двигательного аппарата животных // Журн.
общ. биологии. 1944. Т. 5, № 3. С. 129 – 171.
Зенкевич Л. А. Фауна и биологическая продуктивность моря. М., 1947. Т. 2.
588с.; 1951. Т. 1.507с.
Зенкевич Л. А. Биологическая структура океана // Зоол. журн. 1948. Т. 27,
№ 2. С. 113 – 124.
Зенкевич Л. А. Материалы к сравнительной биогеоценологии суши и океана //
Журн. общ. биологии. 1967. Т. 28, № 5. С. 523 – 537.
Зенкевич Л. А. К вопросу о древности океана и его фауны // История мирового
океана. М., 1971. С. 77 – 83.
Значение биосферы в геологических процессах: Вопросы взаимосвязи. М.; Л.,
1962. 248 с.
Иванов А. Н. Об истории и соотношении понятий о географической оболочке и
биосфере // Вопросы истории и теории физической географии. Саратов, 1978.
Вып. 4. С. 5 – 12.
Иванов А. Н. Биосфера в трудах географов и геологов до Вернадского // Вопросы
истории и теории физической географии. Саратов, 1979. Вып. 5. С. 41 – 51.
Имшенецкий А. А. Экспериментальная изменчивость микроорганизмов. М.,
1956. 42 с.
Казанский Ю. П. Об изменении газового и солевого состава океанических
вод // Геология и геофизика. 1977. № 8. С. 56 – 66.
Казанский Ю. П. О развитии морского карбонатонакопления в геологической истории
Земли // Эволюция осадочного процесса в океанах и на континентах. М.,
1983. С. 157 – 162.
Казанский Ю. П., Катаева В. Я., Шугурова Н. А. Опыт изучения газовой
и жидкой фаз как реликтов древних атмосфер и гидросфер // Геология и география.
1969. №11. С. 39 – 43.
Калабухов Я. И. Сохранение энергетического баланса организма как основа процесса
адаптации // Журн. общ. биологии. 1946. Т. 7, № 6. С. 417 – 434.
Кальвин М. Химическая эволюция. М., 1971. 240 с.
Камшилов М. М. Исследования советских биологов в области эволюционной теории
за тридцать лет // Успехи соврем, биологии. 1947. Т. 24, № 2. С. 287 – 320.
Камшилов
М. М. Значение взаимных отношений между организмами в эволюции. М.; Л., 1961.
136с.
Камшилов М. М. Биотический круговорот. М., 1970. 160 с.
Камшилов М. М. Эволюция биосферы. М., 1974. 256 с.
Камшилов М. М. Изучение биосферы и вопросы воспроизводства и охраны животного
и растительного мира // История биологии с начала XX века и до наших
дней. М., 1975. С. 543 – 560.
Камшилов М. М. Факторы эволюции биосферы // Вопр. философии. 1979.
№3. С. 128 – 137.
Квасов Д. Д., Гольцев А. П., Сафрай А. С. Влияние парникового эффекта в
первичных атмосферах на формирование Земли и планет // Астрон. вестн. 1980,
Т. 24, № 2. С. 72 – 79.
Кист А. А. Биологическая роль химических элементов и периодический закон.
Ташкент, 1973. 65 с.
Кобленц-Мишке О. И., Волковинский В. В., Кабанова Ю. Г. Новые данные о
величине первичной продукции Мирового океана // Докл. АН СССР. 1968. Т.
183,
№5. С. 1189 – 1192.
Ковальский В. В. Периодическая изменчивость химических свойств организмов
и ее биологическое значение // Успехи соврем, биологии. 1941. Т. 14, вып.
3.
С. 380 – 423.
Ковальский В. В. Вопросы биохимического изучения факторов эволюции //
Конференция по проблемам органической эволюции. Уфа, 1943. С. 12 – 14.
213
Ковальский В. В. Новые направления и задачи биологической химии сельскохозяйственных
животных в связи с изучением биогеохимических провинций. М., 1957.
35 с.
Ковальский В. В. Биогеохимические провинции и методы их изучения // Тр.
Биогеохим. лаб. 1960. Т. 11. С. 8 – 32.
Ковальский В. В. Возникновение и эволюция биосферы // Успехи соврем,
биологии. 1963а. Т. 55, вып. 1. С. 45 – 67.
Ковальский В. В. Геохимическая экология и ее эволюционное направление
// Изв. АН СССР. Сер. биол. 19636. № 6. С. 830 – 849.
Ковальский В. В. Некоторые задачи геохимической экологии // Очерки
современной геохимии и аналитической химии. М., 1972. С. 514 – 528.
Ковальский В.
В. Геохимическая экология. М., 1974. 282 с.
Ковальский В. В. Современные
задачи
и проблемы биогеохимии // Тр. Биогеохим. лаб. 1979. Т. 17. С. 12 –
29.
Ковальский В. В. Геохимическая среда и жизнь. М., 1982. 78 с.
Ковальский
В. В., Боровик-Романова Т. Ф. Значение бабочек в биогенной миграции
химических элементов и элементарный состав их различных таксономических
групп //
Тр. Биогеохим. лаб. 1978. Т. 15. С. 175 – 186.
Ковальский В. В., Летунова С. В. Значение микрофлоры в миграции кобальта
и приспособление микроорганизмов к среде в биогеохимических провинциях
с различным содержанием кобальта // Докл. АН СССР. 1959. Т. 126, №
1. С. 167
– 170.
Ковальский В. В., Летунова С. В. Адаптация иловой микрофлоры
к искусственному повышению содержания кобальта в естественной среде
обитания
// Микробиология.
1963. Т. 32, № 5. С. 850 – 856.
Ковальский В. В., Летунова С. В. Геохимическая экология микроорганизмов
// Тр. Биогеохим. лаб. 1974. Т. 13. С. 3 – 37.
Ковальский В. В., Летунова С. В. Пятьдесят лет Биогеохимической лаборатории
// Тр. Биогеохим. лаб. 1979. Т. 17. С. 3 – 13.
Ковальский В. В., Петрунина Н. С. Геохимическая экология и эволюционная
изменчивость растений // Проблемы геохимии. М., 1965. С. 565 – 577.
Ковда В. А. Минеральный состав растений и почвообразование // Почвоведение.
1956. № 1. С. 6 – 38.
Ковда В. А. Основы учения о почвах. М., 1973. Кн. 1. 447 с.
Ковда В.
А. Биогеохимия почвенного покрова. М., 1985. 263 с.
Ковда В. А., Тюрюканов
А. Н. Биосфера
// БСЭ. 3-е изд. М., 1970. Т. 3. С. 364 – 365.
Козлова Л. Е. и др. Сравнительное изучение скелетного вещества ископаемых
и современных кораллов // Палеобиогеохимия беспозвоночных. Новосибирск,
1980. С. 3 – 23.
Козо-Полянский Б. М. Финал эволюции. Краснодар, 1922. 24 с. Колчинский
Э. И. Об изменяемости законов эволюции // Методологические проблемы
биологии. Л., 1971. С. 20 – 26.
Колчинский Э. И. Значение трудов В. И. Вернадского для эволюционной
теории // Проблемы новейшей истории эволюционного учения. Л., 1981.
С. 68 –
94.
Колчинский Э. И. Эволюция биосферы // Развитие эволюционной теории
в СССР.
Л., 1983а. С. 479 – 512.
(Колчинский Э. И.) Kolchinsky E. Evolution of natural selection //
General questions of evolution. Praha, 1983б. P. 53 – 60.
Колчинский Э. И. Геохимические факторы эволюции биосферы //Экология
и теория эволюции. Л., 1984. С. 166 – 183.
(Колчинский Э. И.) Kolchinsky E. On the mane tendencies in the evolution
of the biosphere // Evolution and morphogenesis. Praha, 1985a. P. 771
– 778.
Колчинский Э. И. Философско-методологический анализ проблемы
„эволюция эволюции”: Автореф. дис. ... докт. филос. наук. Л., 1985б.
36 с.
Колчинский Э. И. Идеи В. И. Вернадского об эволюции биосферы. Л., 1987.
16 с.
Комаров В. Л. Смысл эволюции // Дневник I Всерос. съезда рус.
ботаников. Пг., 1921. №5. С. 45.
Кордюм В. А. Эволюция и биосфера. Киев, 1983, 212 с.
214
Коржинский Д. С. Преувеличение роли солнечной энергии в энергетике земной
коры // Изв. АН СССР. Сер. геол. 1955. № 1. С. 52 – 64.
Коссович П. С. Основы учения о почвах. СПб., 1911. Вып. 1. 264 с.
Костицын В. А. Эволюция атмосферы, биосферы и климата. М., 1984.
96 с.
Красилов
В. А. Эволюция и биостратиграфия. М., 1977. 256 с.
Красилов В. А. Нерешенные проблемы теории эволюции. Владивосток, 1986.
138с.
Красная книга СССР. М., 1978. 459 с.
Криволуцкий Д. А. Радиоэкология сообществ наземных животных. М.,
1983. 86 с.
Крупаткина Д. К., Берлин Б., Маэстрани С. Лидер первичной продукции
– океан, а не суша // Природа. 1985. № 4. С. 56 – 62.
Крупеников И. А. История почвоведения. М., 1981. 328 с.
Круть И. В. Введение в общую теорию Земли. М., 1978. 368 с.
Крылов И. Н. На заре жизни. М., 1972. 105 с.
Крылов И. Н. Органический мир докембрия и его влияние на
осадочные процессы // Биогеохимические аспекты формирования осадочных
пород
и руд. Л.,
1983. С. 23.
Кузнецов В. Г. Некоторые черты эволюции рифообразования в истории
Земли // Эволюция осадочного процесса в океанах и на континентах.
М., 1983.
С. 162
– 173.
Кузнецов С. И. Геохимическая деятельность микроорганизмов в месторождениях
полезных ископаемых // Изв. АН СССР. Сер. биол. 1972. № 3. С. 301
– 314.
Кузнецов С. И., Иванов М. В., ЛяликоваН. Н. Введение в
геологическую микробиологию. М., 1962. 239 с.
Кулагин Ю. З. Преадаптация растений и антропогенные факторы
// Журн. общ.
биологии. 1971. Т. 32, № 5. С. 593 – 596.
Кунашева К. Г. Определение углерода в некоторых видах Acridiidae
// Тр. Биогеохим. лаб. 1932. Т. 2. С. 27 – 30.
Лавренко E. M. О значении биогеохимических работ акад. Вернадского
для познания растительного покрова Земли // Природа. 1945. № 5. С.
40 – 44.
Лавренко Е. М. О фитогеосфере // Вопр. географии. 1949. № 15. С.
53 – 56.
Лавренко Е. М. Об энергетической трактовке В. Л. Комаровым эволюции
организмов // Сто лет со дня рождения В. Л. Комарова. Л., 1972. С.
82 – 89.
Ламарк Ж.-Б. Философия зоологии. М.; Л., 1935. Т. 1. 330 с.
Ламарк Ж.-Б. Вступительные лекции к курсу зоологии // Избр. произведения:
В 2-х т. М.; Л., 1955. Т. 1. С. 9 – 36.
(Лапо А. В.) Lapo А. V. Traces of bygone biosphere. M., 1982. 221
p.
Лапо А. В. Следы былых биосфер. М., 1987. 206 с.
Лаптев И. Д. Экологические проблемы. М., 1982. 247 с.
Лебедев В. И. Основы энергетического анализа геохимических
процессов. Л.,
1957. 342 с.
Левченко В. Ф. Старобогатов Я. И. Два аспекта эволюции жизни: физический
и биологический // Физика: Проблемы, история науки. Л., 1986. С.
101 – 142.
Летунова С. В. О роли иловой микрофлоры стоячих водоемов в миграции
кобальта // Тр. Биогеохим. лаб. 1960. Т. 11. С. 107 – 119.
Летунова С. В. Проблемы геохимической экологии микроорганизмов //
Тр. Биогеохим. лаб. 1979. Т. 17. С. 101 – 110.
Летунова С. В., Ковальский В. В., Ермаков В. В. Геохимическая экология
почвенной микрофлоры в условиях различного содержания селена в естественной
среде обитания
// Тр. Биогеохим. лаб. 1968. Т. 12. С. 238 – 258.
Летунова С. В., Ковальский В. В., Романова С. Н. Геохимическая экология
микроорганизмов в условиях различного содержания урана в илах //
Журн. общ. биологии. 1970.
Т. 31, № 1. С. 111 – 120.
Ливанов Н. А. Пути эволюции животного мира. М., 1955. 400 с.
Лисицын А. П. Осадкообразование в океанах. М., 1974. 438 с.
Лисицын А. П. Процессы океанской седиментации. М., 1978. 392 с.
Лисицын А. П., Богданов Ю. А. Главные события мезозоя и
кайнозоя океанов // Эволюция осадочного процесса в океанах и на континентах.
М., 1983.
С. 82
– 103.
215
Личков Б. Л. Движение, материков и климаты прошлого Земли. М.; Л.,
1936.
128 с.
Личков Б. Л. Владимир Иванович Вернадский. М., 1948. 103 с. Лопатин
Н. В. Эволюция биосферы и полезные ископаемые // Изв. АН СССР.
Сер. геол.
1979.
№7. С. 5 – 22.
Лукашев К. П., Вадковская И. К. Биосфера и биогеохимические провинции.
Минск, 1973. 239 с.
Лукашевич И. Д. Неорганическая жизнь Земли. СПб., 1909а. Ч. 2.
311 с.
Лукашевич И. Д. Что такое жизнь? Биологический этюд. СПб., 19096.
32 с.
Лукин Е. И. Закономерности географической изменчивости организмов
// Развитие эволюционной теории в СССР. Л., 1983. С. 175 – 188.
Мазинг В. В. К вопросу об эволюции биоценотических систем //
Теоретические проблемы фитоценологии и биогеоценологии. М., 1970.
С. 95 – 107.
Малюга Д. П. Биогеохимический метод поисков рудных месторождений.
М., 1963.264с.
Минская С. М., Дроздова Т. В. Геохимия органического вещества.
М., 1964. 315 с.
Минская С. М., Кодина Л. А. Геохимия лигнина. М., 1975.
230 с.
Маргелис Л. Роль симбиоза в эволюции клетки. М., 1983.
351 с.
Марковский Б. П. Жизнь как геологический фактор // Значение биосферы
в геологических процессах. Вопросы взаимосвязи палеонтологии
и тектоники. М.; Л., 1962. С.
8 – 11.
Марш Г. Человек и природа. СПб., 1866. IX+ 592 с.
Машковцев А. А. Смена эндогенных и экзогенных факторов эмбриологического
развития в онтогенезе и филогенезе // Изв. АН СССР. Сер. биол.
1936. № 5. С. 945 – 997.
Медников Б. М. Монофилия органического мира и эволюция экосистем
// Журн. общ. биологии. 1985. Т. 46. № 4. С. 462 – 470.
Мейсон Б. Основы геохимии. М., 1971. 311 с.
Меншуткин В. В. Опыт имитации эволюционного процесса на вычислительной
машине // Журн. эволюц. биохимии и физиологии. 1977. Т. 13, №
5. С. 545 – 555.
Мирзоян Э. Н. Развитие сравнительно-эволюционной биохимии в России.
М., 1984. 272 с.
Моисеев Н. Н. Коэволюция человека и биосферы: кибернетические
аспекты // Природа. 1984. № 1. С. 59 – 67.
Моисеев Н. Н. Алгоритмы развития. М., 1987. 304 с.
Моисеев Н. Н., Александров А. А., Тарко А. М. Человек
и биосфера. М., 1985.
272 с.
Моисеев Н. Н., Крапивин В. Ф., Свирижев Ю. М., Тарко А. М. Системный
анализ динамических процессов в биосфере // Человек и биосфера.
М., 1980. Вып.
4. С. 228 – 258.
Монин А. С. История Земли. М., 1977. 228 с.
Морозов Н. П. Глобальный биогеохимический круговорот химических
элементов // Биогеохимические аспекты формирования осадочных
пород и руд. Л.,
1983. С. 25 – 26.
Мочалов И. И. В. И. Вернадский – человек и мыслитель. М., 1970.
176 с.
Мочалов И. И. Владимир Иванович Вернадский. М., 1982.
488 с.
Надсон
Г.
А. Микроорганизмы
как геологические деятели. СПб., 1903. 98 с.
Назаров А. Г. Эволюционная
идея в творчестве В. И. Вернадского // Журн. общей биологии.
1988. Т. 49, № 2.
С. 147 – 165.
Назаров В. И. Финализм в современном эволюционном учении. М.,
1984. 285 с. Наука и Вселенная. М., 1983. 296 с.
Наумов Н. П. О методологических проблемах биологии // Филос.
науки. 1964. № 1.С. 136 – 145.
Наумов Н. П. Об эволюции макросистем // Журн. общ. биологии.
1967. Т. 28, № 6. С. 635 – 649.
Наумов Н. П. Проблемы и задачи популяционной биологии
// Развитие концепции структурных уровней в биологии. М., 1972.
С. 322 –
331.
216
Научное и социальное значение деятельности В. И. Вернадского. Л., 1989.
416 с.
Неаполитанская В. С. Вернадский и современность // Вернадский и современность.
М., 1986. С. 11 – 21.
Неручев С. Г. Опыт количественной оценки параметров древних атмосфер Земли
// Изв. АН СССР. Сер. геол. 1977. № 10. С. 9 – 22.
Неручев С. Г. Возможная масса и продуктивность биосферы в фанерозое и эпохи
образования каустобиолитов // Палеонтология и эволюция биосферы: Тез. докл.
21-й сессии Всесоюз. палеонтолог, о-ва. Л., 1979. С. 38 – 40.
Никольский Г. В., Соколов И. И., Цепнин Е. А. Закономерности изменений ихтиофауны
за период антропогена // Журн. общ. биологии. 1972. Т. 33, № 2. С. 274 –
280.
Ничипорович А. А. Фотосинтетическая деятельность и первичная продуктивность
фитоценозов на современном этапе эволюции биосферы // Проблемы биогеоценологии.
М., 1973. С. 157 – 173.
Новогрудский Д. И. Геохимия и витализм // Под знаменем марксизма. 1931. №
7 – 8. С. 168 – 203.
Одум Ю. Основы экологии. М., 1975. 742 с.
Олейников А. Н. О роли живого вещества в эволюции атмосферы Земли // Значение
биосферы в геологических процессах. М.; Л., 1962. С. 117—127.
Опарин А. И. Возникновение и начальное развитие жизни на Земле.
М., 1966. 204 с.
Опарин А. И. О сущности жизни // Вопр. философии. 1979. № 4. С. 35 – 49.
Оуэн Т. Земля как одна из планет солнечной системы (сравнительное изучение
происхождения и эволюции атмосфер планет). М., 1986. 27 с.
Павлов А. П. Ледниковые и межледниковые эпохи Европы в связи с историей ископаемого
человека. Пг., 1922. 52 с.
Пак А. И. Эволюция кор выветривания в истории Земли // Эволюция осадочного
процесса в океанах и на континентах. М., 1983. С. 179 – 191.
Палеонтология и эволюция биосферы. Л., 1983. 152 с.
Панфилов Д. В. Эколого-ландшафтные условия возникновения жизни на Земле //
Бюл. МОИП. Отд. биол. 1977. Т. 82, вып. 6. С. 5 – 15.
Патти Г. Замечание // На пути к теоретической биологии. М., 1970. Т. 1. Пролегомены.
С. 177 – 179.
Перельман А. И. Геохимия ландшафта. М., 1961. 496 с.
Перельман А. И. Геохимия биосферы. М., 1973. 156 с.
Перельман А. И. Биокосные системы Земли. М., 1977. 160 с.
Перельман А. И. Геохимические факторы биологической эволюции // Вопросы палеобиогеохимии.
Баку, 1981. С. 86 – 97.
ПерельманА. И. Геохимия природных вод. М., 1982. 115 с.
Перельман А. И., Борисенко Е. Н. Пермские равнины Приуралья как особая геохимическая
и, возможно, палеобиогеохимическая провинция // Проблемы геохимии. М., 1965.
С. 614 – 619.
Петров В. П. Основы учения о древних корах выветривания. М., 1967. 343 с.
Петрунина Н. С. Геохимическая экология растений в провинциях с избыточным
содержанием микроэлементов (Ni, Со, Си, Mo, Pb, Zn) // Тр. Биогеохим. лаб.
1974. Т. 17. С. 57 – 117.
Пианка Э. Эволюционная экология. М., 1981. 399 с.
Плотников В. В. Эволюция структурных растительных сообществ. М., 1979. 274с.
Покаржевский А. Д. Геохимическая экология наземных животных. М., 1985. 300
с.
Полынов Б. Б. Основы построения генетической классификации почв // Задачи
и методы почвенных исследований. М,; Л., 1933. С. 23 – 33.
Полынов Б. Б. Кора выветривания. Л., 1934. 240 с.
Полынов Б. Б. Первые стадии почвообразования на массивнокристаллических породах
// Почвоведение. 1945. № 7. С. 327 – 339.
Полынов Б. Б. К вопросу о роли элементов биосферы в эволюции организмов
// Почвоведение. 1948. № 10. С. 594 – 607.
217
Полынов Б. Б. Учение о ландшафтах // Вопр. географии. 1953а. Вып. 33. С.
30 – 44.
Полынов Б. Б. О геологической роли организмов // Вопр. географии.
19536. Вып. 33. С. 45 – 64.
Полынов Б. Б. Избранные труды. М., 1956. 751 с.
Полянский Ю. И. О своеобразных чертах прогрессивной эволюции на клеточном
уровне // Журн. общ. биологии. 1971. Т. 32, № 5. С. 541 – 548.
Полянский Ю. И. О специфике эволюционного процесса в крупных таксонах
(на примере эволюции простейших) // Проблемы новейшей истории эволюционного
учения. Л., 1981. С. 156 – 169.
Поннамперума С. Происхождение жизни. М., 1977. 176 с.
Попов В. А. Антропогенные факторы эволюции млекопитающих // Бюл. МОИП.
Отд. биол. 1971. Т. 76, № 2. С. 16 – 21.
Посохов Е. В. Химическая эволюция гидросферы. Л., 1981. 286 с.
Пресман
А. С. Идеи В. И. Вернадского в современной биологии. М., 1976. 64 с.
Принн Г. Химия и химическая эволюция атмосфер Венеры, Сатурна и Титана по данным
новейших космических исследований. М., 1986. 29 с.
Проблемы эволюции геологических процессов. Новосибирск, 1981. 207 с.
Пустовалов Л. В. Петрография осадочных пород. М.; Л., 1940. Ч. 1. 476
с.
Развитие эволюционной теории в СССР: 1917 – 1970 гг. Л., 1983. 614 с.
Райков И. Б. Ядро простейших:
Морфология и эволюция. Л., 1978. 327 с.
Ранняя история Земли. М., 1980. 620
с.
Ревердатто В. В. Введение в фитоценологию. Томск, 1935. Ч. 1. 98 с.
Реклю
Э. Земля: Описание земного шара. М., 1914. Т. 10. 111 с.
Ремезов Н. П. Противоположные процессы как основа генетического развития почв // Почвоведение.
1932. №
2. С. 178 – 211.
Риклефс Р. Основы общей экологии. М., 1979. 424 с.
Роде А. А. Почвообразовательный процесс и эволюция почв. М., 1947. 142
с.
Роде А. А. Докучаевское почвоведение в Академии наук в 20 – 30-е годы
// Природа. 1974. № 5. С. 59 – 67.
Родендорф Б. Б., Жерехин В. В. Палеонтология и охрана природы // Природа.
1974. №5. С. 82 – 91.
Родин Л. Е., Базилевич Н. И. Динамика органического вещества и биологический
круговорот элементов и азота в основных типах растительности земного
шара. М.; Л., 1965. 253 с.
Ронов А. Б. К послекембрийской геохимической истории атмосферы и гидросферы
// Геохимия. 1959. № 5. С. 397 – 409.
Ронов А. Б. Общие тенденции в эволюции состава земной коры, океана и
атмосферы // Геохимия. 1964. № 8. С. 715 – 743.
Ронов А. Б. Эволюция состава пород и геохимических процессов осадочной
оболочки Земли // Геохимия. 1972. № 2. С. 137 – 147.
Ронов А. Б. Осадочная оболочка Земли. М., 1980. 76 с.
Ронов А. Б. Эволюция осадкообразования в истории Земли // Эволюция осадочного
процесса в океанах и на континентах. М., 1983. С. 49 – 74.
Ронов А. Б., Мигдисов А. А. Эволюция химического состава пород щитов
осадочного покрова Русской и Северо-Американской платформы // Геохимия.
1970. №4.
С. 403 – 438.
Ронов А. Б., Мигдисов А. А., Ярошевский А. А. К геохимической истории
внешних оболочек Земли // Очерки современной геохимии и аналитической
химии. М.,
1972. С. 88 – 93.
Руттен М. Происхождение жизни. М., 1973. 413 с.
Рябчиков А. М. Баланс энергии и биомасса Земли // Вестн. МГУ. Сер. 5.
География. 1971. № 1.С. 13 – 19.
Садиков В. С. Химия жизни. Пг., 1923. Вып. 1. 88 с.
Садиков В. С. К методике химического анализа животных организмов // Изв.
АН СССР. Сер. 6. 1926. № 9. С. 649 – 654.
(Садиков В. С., Виноградов А. П.) Sadikow W. S., Winogradow A. P. Untersuchungen
uber die Zussamentsetzung des lebendingen Substrats // Biochem. Ztsch.
1924. Bdl50,H. 5/6. S. 372 – 391.
218
Самойлов Я. В. Минералогия жильных месторождений Нагольного кряжа
(Донецкий бассейн) // Материалы для геологии России. 1906. Т. 23, вып. 1.
С.
1 – 260.
Самойлов Я. В. Биолиты. Л., 1929. 140 с.
Сапожников Д. И. Возникновение и эволюция фототрофного типа питания
// Возникновение жизни на Земле. М., 1959. С. 28 – 41.
Сауков А. А. Эволюция факторов миграции элементов в геологической
истории // Изв. АН СССР. Сер. геол. 1961. № 5. С. 3 – 16.
Северцов А. Н. Эволюция и психика. М., 1922. 54 с.
Северцов А. Н. Главные направления эволюционного процесса. М.; Л.,
1934. 150 с.
Северцов С. А. Проблемы экологии животных. М., 1951. Т. 1. 172 с.
Серавин Л. Н. Простейшие. Что такое? Л., 1984. 174 с.
Сидоренко А. В. Проблемы литологии докембрия и полезные ископаемые
// Вестн. АН СССР. 1976а. № 1. С. 113 – 125.
Сидоренко А. В. Осадочная геология докембрия – ключ к познанию
истории земной коры // Наука и человечество. 1977 (Международный
ежегодник).
М., . 1976б.
С. 101 – 111.
Сидоренко А. В. Новое в учении о биосфере. М., 1984. 15 с.
Сидоренко А. В., Сидоренко Св. А. Органическое вещество в докембрийских
осадочно-метаморфических породах и некоторые геологические проблемы
// Сов. геология. 1971. № 5. С.
3 – 30.
Сидоренко А. В., Теняков В. А., Сидоренко С. А. О едином биогеологическом
процессе созидания и развития сиаля земной коры // Докл. АН СССР.
1978. Т. 238, № 4. С. 924 – 927.
Сидоренко Св. А., Сидоренко А. В. Органическое вещество в осадочных
метаморфических породах докембрия. М., 1975. 116 с.
Сидоренко Св. А., Теняков В. А. О глобальных „эпохах” накопления
в докембрии углеродистых формаций // Докл. АН СССР. 1978. Т. 242,
№ 1.
С. 172 – 175.
Симкин Г. Н. Явление жизни и функциональная организация биологических
макросистем // Бюл. МОИП. Отд. биол. 1969. Т. 74, вып. 3. С. 158
– 160.
Синицын В. М. Сиаль: Историко-генетические аспекты. Л., 1972. 167
с.
Ситник К. М., Голубец М. А. До питания про эволюцию экосистем //
Укр. ботан. журн. 1983. Т. 40, № 1. С. 1 – 8.
Соболев Д. Н. Начала исторической биогенетики. Харьков, 1924. 203
с.
Соколов Б. С. Органический мир Земли на пути к фанерозойской дифференциации
// Вестн. АН СССР. 1976. № 1. С. 126 – 143.
Соколов Б. С. Палеонтология и эволюция биосферы // Палеонтология
и эволюция биосферы. Л., 1983. С. 4 – 17.
Соколов Б. С. Биосфера: понятие, структура, эволюция // В. И. Вернадский
и современность. М., 1986. С. 96 – 123.
Соколов Б. С., Барсков И. С. Палеонтология и эволюция биосферы
// Современная палеонтология. М., 1988. Т. 2. С. 245 – 254.
Соколов В. Эволюция атмосферы Земли // Возникновение жизни на Земле
М., 1959. С. 59 – 69.
Соколов С. Я. Типы борьбы за существование в мире растений // Конференция
по проблемам дарвинизма. М., 1948. С. 27 – 29.
Сорохтин О. Г. Глобальная эволюция Земли. М., 1974. 184 с.
Стайлс В. Микроэлементы в жизни растений и животных. М., 1949. 186
с.
Старобогатов Я. И. Эволюция пресноводных экосистем // Актуальные
вопросы гидробиологии. Л., 1984. С. 24 – 33.
Старобогатов Я. И. О соотношении между микро- и макроэволюцией //
Дарвинизм: история и современность. Л., 1988. С. 138 – 145.
Страхов Н. М. Основы теории литогенеза. М., 1960. Т. 1. 212 с.; 1963.
Т. 3. 550 с.
Страхов Н. М. Этапы развития внешних геосфер и осадочных пороцообразований
// Изв. АН СССР. Сер. геол. и геогр. 1962. № 12. С. 3 – 22.
Страхов Н. М. Развитие литогенетических идей в России и СССР. М.,
1971. 608 с.
219
Суетова И. А. География живого вещества // Природа, 1976. № 2. С. 34 –
37.
Сукачев В. Н. К вопросу о борьбе за существование между биотипами одного
и того же вида // Юбилейный сборник, посвященный И. П. Бородину. Л.,
1927. С. 195 – 219.
Сукачев В. Н. Идея развития в фитоценологии // Сов. ботаника. 1942. №
1 – 3. С. 5 – 17.
Сукачев В. Н. О принципах генетической классификации в биогеоценологии
// Журн. общ. биологии. 1944. Т. 5, № 4. С. 213 – 227.
Сукачев В. Н. К вопросу о развитии растительности // Ботан. журн. 1952.
Т. 37, № 4. С. 496 – 507.
Сукачев В. Н. Основные понятия лесной биогеоценологии // Основы лесной
биогеоценологии. М., 1964а. С. 5 – 49.
Сукачев В. Н. Динамика лесных биогеоценозов // Основы лесной биогеоценологии.
М., 19646. С. 458 – 486.
Султанов К. М., Исаев С. А. К вопросу о сравнительном изучении химического
элементарного состава раковин современных и ископаемых беспозвоночных
// Уч. зап. Азерб. ун-та. Сер. Геология-география. 1967. № 2. С. 3 –
7.
Сутт Т. Я. Проблема направленности органической эволюции. Таллин,
1977. 139с.
Сутт Т. Я. Управление эволюцией – проблемы мнимые и реальные
// Эволюционная теория и проблема „человек-природа”. Тарту, 1978. С.
14 – 17.
Сытник К. М., Апанович Е. М.,. Стойко С. М. В. И. Вернадский: Жизнь и
деятельность на Украине. Киев, 1988. 366 с.
Татаринов Л. П. Палеонтология и теория эволюции // Вестн. АН СССР. 1983.
№ 12. С. 40 – 51.
Таусон В. О. Об эволюции микроорганизмов в течение геологических эпох
// Архив биол. наук. 1936. Т. 43, вып. 2 – 3. С. 267 – 286.
Таусон В. О. Изменения направления и биохимизма некоторых процессов в
растительной клетке в ходе эволюции // Сборник работ по физиологии растений:
Памяти
К. А. Тимирязева. М.; Л., 1941. С. 171 – 215.
Таусон В. О. Наследство микробов. М.; Л., 1947. 148с.
Тахтаджян А. Л. Четыре царства органического мира // Природа. 1973. №
2. С. 22 – 32.
Тейяр де Шарден П. Феномен человека. М., 1965. 296 с.
Теняков В. А. Бокситообразование в геологической истории Земли
и проблемы бокситов докембрия // Докембрийские коры выветривания. М.,
1975. С. 52
– 65.
Терентьева К. Ф. Минеральный состав скелетов некоторых современных иглокожих
// Тр. Биогеохим. лаб. 1932. Т. 2. С. 45 – 63.
Тимирязев К. А. Космическая роль растений // Научное слово. 1904. № 3.
С. 31 – 77.
Тимофеев П. П., Щербаков А. В. Проблема энергетики гипергенных процессов
// Докл. АН СССР. 1983. Т. 269, № 1. С. 184 – 187.
Тимофеев П. П., Щербаков А. В., Ильин В. А. Роль биосферы в энергетике
земной коры // Биогеохимические аспекты формирования осадочных пород
и руд. Л.,
1983. С. 31.
Тимофеев-Ресовский Н. В. Микроэволюция // Ботан. журн. 1958. Т. 43, №
3. С. 317 – 336.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк
теории эволюции. М., 1969. 408 с.
Тимофеев-Ресовский Н. В., Тюрюканов А. Н. Об элементарных биохорологических
подразделениях биосферы // Бюл. МОИП. Отд. биол. 1966. Т. 71, вып. 1.
С. 123 – 132.
Титов И. А. Последствия жизнедеятельности растительных сообществ: Климато-геоботаническое
исследование. М., 1934. 192 с.
Томшин А. Т. Содержание микроэлементов у насекомых отряда чешуекрылых
(Lepidoptera), обитающих в различных экологических условиях: Автореф.
дис. ... канд. биол.
наук. Красноярск, 1974. 23 с.
Трут Л. Н. Очерки по генетике поведения. Новосибирск, 1978.
255 с.
220
Тюрюканов А. Н., Александрова В. Д. Витасфера Земли // Бюл. МОИП. Отд.
биол. 1969. Т. 74, вып. 4. С. 14 – 26.
Уголев А. М. Трофология – новая междисциплинарная наука // Вестн.
АН СССР. 1980. № 1. С. 50 – 61.
Уголев А. М. Эволюция пищеварения и принципы эволюции функций. Л.,
1985. 542 с.
Уголев А. М. Естественные технологии. Л., 1987. 318с.
Удельнова Т. М., Пушева М. А., Черногорова С. М., Бойченко Е.
А. Соотношение поливалентных металлов в эволюции биогеохимических функций
// Тр. Биогеохим.
лаб. 1979. Т. 17. С. 183 – 189.
Умов Н. А. Характерные черты и задачи естественнонаучной мысли. СПб.,
1914. 47 с.
Умов Н. А. Физико-химическая модель живой материи // Собр. соч. М.,
1916. Т. 3. XVI+ 666 с.
Уокер Дж. С. Г. Эволюция химического состава атмосферы // Импакт.
1984. № 1. С. 17 – 25.
Уолд Дж. Почему живое вещество базируется на элементах второго и
третьего периодов периодической системы? Почему фосфор и сера способны
к образованию
макроэргических связей? // Горизонты биохимии. М., 1964. С. 102 –
113.
Урысон М. И. Истоки семейства гоминид и филогенетическая дифференциация
высших приматов // Человек: эволюция и внутривидовая дифференциация.
М., 1972. С.
9 – 22.
Успенский В. А. Баланс углерода в биосфере в связи с вопросом о распространении
углерода в земной коре. Л., 1956. 101 с.
Ушаков С. А., Ясаманов Н. А. Дрейф материков и климат Земли. М.,
1984. 207 с.
Уэбб Дж. Прикладная геохимия и общество // Проблемы геохимии. М.,
1965. С. 578 – 588.
Фейбридж Р. В. Карбонатные породы и палеоклиматология в биогеохимической
истории планеты // Карбонатные породы. М., 1970. Т. 1. С. 357 – 386.
Ферсман А. Е. Химические проблемы промышленности. Л., 1924. 50 с.
Ферсман А. Е. Периодический закон количества элементов // Докл. АН
СССР. 1932. №11. С. 261 – 266.
Ферсман А. Е. Геохимия. Л., 1934. Т. 2. 352 с.; 1937. Т. 3. 503 с.
Ферсман А. Е. Жизненный путь академика В. И. Вернадского (1863 –
1945) // Зап. Всерос. минерал, о-ва. 1946. Сер. 2. Ч. 75, № 1. С.
5 – 24.
Флоренский К. П. Биосфера // БСЭ. 3-е изд. 1972. Т. 9. С. 484 – 485.
Флоркен М. Развитие биосферы и биохимическая эволюция // Возникновение
жизни на Земле. М., 1959. С. 505 – 516.
Фокс С., Дозе К. Молекулярная эволюция и возникновение жизни. М.,
1975. 374с.
Хатчинсон Дж. Биосфера // Биосфера. М., 1972. С. 9 – 25.
Хахина Л. Н. О роли человека в живой природе // Наука и техника.
Л., 1977. Вып. 9. С. 118 – 120.
Хесин Р. В. Непостоянство генома. М., 1984. 472 с.
ХильмиГ. Ф. Основы физики биосферы. Л., 1966. 300 с.
Хозацкий Л. И. Эволюция организмов и эволюция биосферы // Палеонтология
и эволюция биосферы: Материалы 25-й сессии Всесоюз. палеонтол. о-ва.
Л., 1979.
С. 56 – 57.
Холодный Н. Г. К проблеме возникновения и развития жизни на Земле
// Успехи соврем, биологии. 1945. Т. 19, вып. 1. С. 65 – 78.
Хохряков А. П. Эволюция биоморф растений. М., 1981. 168с.
Цветков Л. Произход и еволюция на биосферата. София, 1981. 179 с.
Цейтлин В. Б. Продукция и биомасса фитопланктона в Мировом океане
// Докл. АН СССР. 1983. Т. 69, № 1. С. 247 – 250.
Чернов Ю. И. Проблема эволюции на биоценотическом уровне
организации жизни // Развитие эволюционной теории в СССР. Л., 1983.
С. 464 –
479.
221
Чижевский А. Л. Земное эхо солнечных бурь. М., 1973. 349 с.
Шапошников
Г. X. Живые системы с малой степенью целостности // Журн. общ.
биологии. 1975.
Т. 36, № 3. С. 323 – 335.
Шарыгин С. А. Приложимость данных по химическому элементарному
составу к систематике некоторых амфибий и рептилий // Журн. общ.
биологии.
1984. Т.
45, № 5. С. 624 – 630.
Шарыгин С. А., Коржневский В. В. Микроэлементы в биогеоценозах
Южного берега Крыма // Наземные и водные экосистемы. Горький,
1982. Вып.
5. С. 118 –
122.
Шварц С. С. Влияние микроэлементов на животных в естественных
условиях рудного поля // Тр. Биогеохим. лаб. 1954. Т. 10. С.
76 – 81.
Шварц С. С. Эволюционная экология животных // Тр. Ин-та экологии
животных и растений. 1969. Вып. 65. 199 с.
Шварц С. С. Эволюция и биосфера // Проблемы биогеоценологии.
М., 1973. С. 213 – 228.
Шварц С. С. Эволюция биосферы и экологическое прогнозирование
// Вести. АН СССР. 1976. № 2. С. 61 – 72.
Шварц С. С. Экологические закономерности эволюции. М., 1980.
278 с.
Шварц С. С., Добринская Л. А., Добринский Л. Н. О принципиальных
отличиях
в
характере эволюционных преобразований у рыб и высших позвоночных
животных // Внутривидовая
изменчивость наземных позвоночных животных и микроэволюция. Свердловск,
1965. С. 59 – 76.
Швец В. М. Органическое вещество подземных вод. М.,
1973. 191 с.
Шилова Е. И. Еще раз о необходимости критического
отношения
к учению
акад.
В. Р. Вильямса
о едином почвообразовательном процессе // Вести. ЛГУ. Биология.
1957. № 15, вып. 3. С. 33 – 42.
Шилова Е. И. О современном состоянии проблемы эволюции почв и
методах ее изучения // Вести. ЛГУ. Биология. 1959. № 15, вып.
3. С. 26
– 35.
Шилова Е. И. К вопросу об эволюции обмена веществ в биосфере
// Вести. ЛГУ. Биология. 1960. № 3, вып. 1. С. 41 – 55.
Шилова Е. И. К вопросу об эволюции обмена веществ в биосфере
// Значение биосферы в геологических процессах. М., 1962. С.
122 –
144.
Шилова Е. И. О понятии „биосфера”, его значении и современном
толковании // Вести. ЛГУ. 1983. Биология. № 3, вып. 1. С. 5 –
16.
Шипунов Ф. Я. Организованность биосферы. М., 1980. 291
с.
Шкорбатов Г. Л. К построению общей теории адаптации // Журн.
общ. биологии.
1982. Т. 43,
№ 6. С. 775 – 787.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом
развитии. М.; Л., 1938. 144 с.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса.
М.; Л., 1940. 231 с.
Шмальгаузен И. И. Кибернетические вопросы биологии.
Новосибирск, 1968. 224 с.
Шмальгаузен И. И. Проблемы дарвинизма. Л., 1969. 494 с.
Шмальгаузен И. И. Факторы прогрессивной (ароморфной) эволюции
– снижения энтропии // Закономерности прогрессивной эволюции.
Л.,
1972. С. 5
– 27.
Шукарев С. А. Периодическая система Д. И. Менделеева
и современная химия // Периодический закон и строение атома.
М., 1971. С. 118
– 203.
Шушкина Э. А. Продукция основных экологических групп планктона
в эпипелагиали океана // Океанология. 1985. Т. 25, вып. 5. С.
839 –
845.
Шушкина Э. А., Виноградов М. Е., Востоков С. В. Оценка первичной
продукции и генетической деструкции в эпипелагиали океана //
Океанология. 1984.
Т. 24, №1. С. 130 – 138.
Щепотьев А. Исследования над низшими организмами. СПб.,
1910. 376 с.
Щербак Н. П. В. И. Вернадский. Киев, 1988. 107 с.
Эволюция литогенеза в истории Земли. Новосибирск, 1981, 163 с.
Эволюция осадочного процесса в океанах и на континентах.
М., 1983. 246 с.
222
Эгами Фудзио. Химическая эволюция в первобытном океане и роль
в ней
ионов переходных элементов // Изв. АН СССР. Сер. биол. 1980.
№ 4. С. 519 – 527.
Эйген М., Шустер П. Гиперцикл: Принципы самоорганизации макромолекул.
М., 1982. 270 с.
Яблоков А. В. Многообразие живого – драгоценное наследие
эволюции //
Наука и жизнь, 1978. № 7. С. 26 – 27, 42 – 49.
Яблоков А. В. Управляемая эволюция как путь сохранения разнообразия
жизни // Диалектика в науках о природе и человеке. Кн. 2:
Эволюция материи
и ее структурные уровни. М., 1983. С. 302 – 311.
Яблоков А. В., Юсуфов А. Г. Эволюционное учение. М., 1981.
344 с.
Яншин А. Л. Основные проблемы соленакопления // Проблемы
соленакопления. Новосибирск, 1977. Т. 1. С. 5 – 15.
Яншин А. Л. Методологическое значение учения В. И. Вернадского
о биосфере и преобразовании ее в ноосферу // Методология
науки и научный
прогресс.
Новосибирск, 1981. С. 194 – 204.
Яншин А. Л. В. И. Вернадский и его учение о биосфере и переходе
ее в ноосферу // В. И. Вернадский и современность. М„ 1986.
С. 28 –
40.
Яншин А. Л. Эволюция геологических процессов в истории
Земли. Л.,
1988. 39 с.
Яншин А. Л., Жарков М. А. Об эволюции осадочных формаций
в геологической истории // Эволюция осадочного процесса в
океанах
и на континентах.
М.,
1983. С. 74 – 81.
Яншин А. Л., Жарков М. А., Казанский Ю. П. Эволюция осадочного
породообразования в истории Земли и связанные с ней закономерности
размещения полезных
ископаемых // Геология и геофизика. 1977. № 11. С. 90 – 97.
Яншин А. Л., Казанский Ю. П., Николаева И. В. Проблема
осадочного процесса // Эволюция осадочного процесса в океанах
и на континентах.
М., 1983.
С. 5 – 16.
Ярошенко П. Д. Геоботаника. М.; Л., 1961. 474 с.
Ясаманов
Н. А. Древние климаты Земли. М., 1985. 96 с.
Abelson P. Amino acids in fossils // Science. 1954. Vol. 119,
N 3096. P. 576. Abelson P. Organic geochemistry // J. Wash.
Acad. Sci. 1963.
Vol.
53, N 1.
P. 105 – 115.
Allen J., Nelson M. Space biospheres. Arizona, 1986. 88
p.
Alvarez W. An indium clue to the dinosaurs' demise // New
Sci. 1979. Vol 82 N1158. P. 798.
Berkner L., Marshall L. The history of growth in the earth's
atmosphere // The origin and evolution of atmosphere and
oceans. New York,
1964. P. 102
– 126.
Berkner L., Marshall L. On the origin and rise of oxygen
concentration in the earth's atmosphere // J. Atmos. Sci.
1965. Vol. 22,
N 3. P. 225 – 261.
(Berkner L, Marshall L). Беркнер Л., Маршалл Л. Кислород
и эволюция // Земля и Вселенная. 1966. № 4. С. 32 – 39.
Bertrand D. Survey of existing knowledge of biogeochemistry.
2. The biogeochemistry of vanadium // Bull. Amer. Mus.
Natur. Hist.
1950.
Vol. 94. P. 403 –
455.
Bezold A. Untersuchungen uber die Verteilung von wasserorganischer
Materie und anorganischen Verbindungen in Tierreiche //
Ztsch. Wiss. /ool. 1857.
Bd 8. S. 487 – 517.
Biogeochemical cycling of mineral-forming elements. Amsterdam;
New York, 1979. 612p.
Boimineralization and biological metal accumulation. International
Symposium on Biomineralization(1982, Renesse, Netherlands).
Reidel, 1983. 573 S.
Bowen H. Trace elements in biochemistry. London; New York,
1966. X + 241 p.
Bradshaw A. D. Plant evolution in extreme // Ecological
genetic and evolution Oxford; Edinburgh, 1971. P. 79 –
93.
Bradshaw A. Adaptation of plants to soils containing toxic
metals – a test for conceit // Origins and development
of adaptation. London, 1984.
P.
4 – 14.
Brown G., Mussett A. The inaccessible Earth. London
etc., 1981. 262 p.
223
Cloud P. Atmospheric and hydrospheric evolution on the
primitive earth // Science. 1968. Vol. 160, N 3829.
P. 729 – 736.
Cloud P. A working model primitive Earth // Amer. J.
Sci. 1972. Vol. 272, N 6. P. 537 – 548.
Cloud P. Beginnings of biosphere evolution and their
biogeochemical consequences // Paleobiology. 1976.
Vol. 2, N 4. P. 351
– 387.
Co-Evolution. 20-ste Phylogenet. Simp. (Hamburg, 28
– 30 Nov., 1975). Hamburg; Berlin, 1978.128 S.
Crick F., Orgel L. Directed panspermia // Icarus. 1973.
Vol. 19, N 3. P. 341 – 346.
Daly R. First calcareous
fossils and
evolution of
limestone
// Bull. Geol. Soc. Amer. 1909. Vol. 20, N1. P. 153
– 170.
Degens E. Molecular mechanism of carbonate, phosphate
and silica deposition in the living cell // Top Curr.
Chem.
1976. Vol.
64. P. 3 – 112.
Degens E. Why do organisms calcify? // Chem. Geol.
1979. Vol. 25, N 5. P. 257 – 269.
Dickerson R. Chemical evolution and the origin of life
// Sci. Amer. 1978. Vol. 239. P. 70 – 87.
Dobzhansky Th. Genetics and origin of species. New
York, 1951. 364 p.
Dose K., Rauchfuss H. Chemische
Evolution
und der Ursprung
lebenden
Systeme.
Stuttgart, 1975. 217 S.
Dunbar M. Ecological development in polar regions:
A study in evolution. Prentice-Hall, 1968.119 p.
Ehrlich A., Ehrlich P. Coevolution,drugs and pest control
// Bios. 1982. Vol. 53, N 3. P. 134 – 139.
Ehrlich P., Money A. Extinction, substitution and ecosystem
services // BioScience. 1983. Vol. 33, N 4. P. 248
– 254.
Fiennes R. Ecology and Earth history. New York, 1976.120
p.
Franz V. Zum jetzigen Stand der Theorie von biotechnischen
Fortschritt in der Pflanzen und Tiergeschichte // Biol.
General. 1951. Bd
19, H. 3. S. 368 – 385.
Frey-Wyssling A. Die unentbehrlichen Elemente der Pflanzenahrung//
Naturwissenschaften. 1935. Bd 23, H. 6. S. 767 – 769.
(Garrels R.) Гаррэлс Р. Круговорот углерода, кислорода
и серы в течение геологического времени. М, 1975. 47
с.
Garrets R., Lerman A., MacKenzie F. Controls of atmospheric
O2 and CO2: past, present and future // Amer. Sci.
1976. Vol. 64,
N 3. P.
306 – 315.
(Carrels R., MacKenzie F.) Гаррелс Р., Маккензи Ф. Эволюция
осадочных пород. М., 1971. 272 с.
Gribbin J. Carbon dioxide ammonia and life // New Sci.
1982. Vol. 94, N 1305. P. 413 – 414.
Guntau M. Der Aktualismus in den geologischen Wissenschaften
// Freiberg. Forschungsh. R. D. 1967. Bd 55. S. 1 – 139.
Hart M. The evolution of the atmosphere of the earth
// Icarus. 1978. Vol. 33, N 1. P. 23 – 29.
Holland H. The geologic history of sea water and attemp
to solve problem // Geochim. etcosmocnim.acta. 1972.
Vol. 36,
N 5. P.
637 – 651.
(Holland H.) Холланд Г. Эволюция атмосферы // Энциклопедия:
Океан – атмосфера. Л., 1983. С. 438 – 442.
Hutchinson G. Survey of existing knowledge of
biogeochemistry. 3. The biogeochemistry of vertebrate
excretion // Bull.
Amer. Mus. Nat.
Hist.
1950. Vol. 96. XVII
+ 554 p.
Hutchinson G. The biologist poses some problem // Oceanography.
1961. N 67. P. 85 – 94.
Huxley J. Eastern Africa: the ecological base // Endeavour.
1962. Vol. 21, N 82. P. 98 – 107.
Huxley J. Evolution: The modern synthesis. London, 1963.
652 p.
Kamen M. Survey of contemporary knowledge of biogeochemistry.
1. Isotopic phenomen in biogeochemistry // Bull. Amer.
Mus. Nat. Hist.
1946. Vol.
87, N 2. P. 101
– 138.
Krumbein W. Geomicrobiologische Prozesse bei
der Anreicherung nutzbarer Minerale und sedimentarer
Lagerstatten // Erdol.
u. Kohle, Erdgas,
Petrochemie. 1978.
Bd 31, H. 3. S. 147 – 151.
224
Krumbein W. Calcification by bacteria and algae // Biogeochemical
cycling of mineral-forming elements. Amsterdam; New
York, 1979. P. 180 – 232.
Kulp J. Origin of the hydrosphere // Bull. Geol. Soc.
Amer. 1951. Vol. 62, N 3. P. 326 – 330.
Lamarck J. B. Hydrogeologie. Paris, 1802. 268 p.
La Mont C. The ecosphere // Sci. Amer. 1958. N 4.
P. 83 – 86. Leakey R. Themakingof mankind. London,
1981.
256
p.
Le Come J. Elements of geology. New York. 1904.667
p.
Le Roy E. Lexidence idealiste et le foit de 1'evolution.
Paris, 1927. 270 p.
Leuthardt F. Mineralstoffwechsel und die Spurenelemente
// Ergeb. Physiol. 1941. Bd44. S. 588 – 643.
Lewin B. R. The science as way of knowing – molecular
evolution // Amer. Zool. 1984. Vol. 24, N 2. P. 451
– 464.
Lindeman R. The trophic – dynamic aspect of ecology
// Ecology. 1942. Vol. 23, N 3. P. 399 – 419.
Lotka A. Elements of physical biology. Baltimore,
1925. 460 p.
Lovelock J. Gaia: A new look at life on the earth.
Oxford; New York, 1979. XI + + 157 p.
Lovelock J. Gaia as seen throught the atmosphere
// Biomineralization and biological metal accumulation.
Reidel, 1983. P. 3
– 19.
Lovelock J., Whitfleld M. Life, span of the biosphere
// Nature. 1982. Vol. 296, N5857. P. 561 – 563.
Lowenstam H. A. Impact of life on chemical and physical
progress // The sea. New York, 1975. Vol. 5. P. 715
– 796.
Lowenstam H. A. Biomineralization processes and products
and evolution of biomineralization // 27 Int. Geol.
Congr. Proc.
Utrecht, 1984.
Vol. 2. P.
79 – 95.
Lowenstam H., Margulis L. Evolutionary prerequisites
for Phanerozois skeletens // BioSystems. 1980. N
12. P. 27
– 41.
Lull R. The pulse of life // The evolution of the
earth and its inhabitans. New Haven, 1923.146р.
Lyell Ch. Principles of geology. London, 1830 – 1832.
Vol. 1. XXXII+ 420 p.; Vol. 2. 453 p.
Lyell Ch. The geological evidence of the antiquity
of man. London, 1863. 528 + + XVI p.
Mason B. Principles of geochemistry. New
York, 1958. X + 276 p.
Matthey R. Quelques reflexions sur le probleme
de l'evohition // Scientia. 1954. Vol. 89, N6. P.
200
– 202.
May R. Pattern in multi species communities // Theoretical
ecology. Oxford, 1976. P. 142 – 162.
May R. The ecolution of ecological systems // Sci.
Amer. 1978. Vol. 239, N 3. P. 119 – 133.
Meyer-Abich A. The historico-philosophical background
of the modern evolution biology. Leiden, 1964.170
p.
Monty C. Consequences biosedimentologiques de revolution
des ecosustemes // Petrole et Techneques. 1979. Vol.
260, N 1.
P. 3 – 11.
Nopcsa F. Eidolosaurus und Pachyophis: Zwei neue
Neokomreptilien // Paleontographica. 1923. Bd 64.
S. 99 – 154.
Paulus H. Co-Evolution zwischen Bluten und ihren
tierischen Bestaubern // Co-Evolution. 20-ste Phylogenet.
Symp.
(Hamburg, 28-30 Nov.,
1975). Humburg;
Berlin, 1978.
S. 51 – 81.
Poole J. The evolution of the atmosphere // Proc.
Roy. Dublin Soc. Sci. 1941. Vol. 22. P. 345 – 363.
Poole J. The evolution of the earth's atmosphere
// Proc. Roy. Dublin Soc. Sci. 1951. Vol. 25. P.
201 –
224.
Prat S. Die erblichkeit der Resistenz gegen Kupfer
// Ber. Dtsch. bot. Ges. 1934. Bdl02,H. l.S. 65 –
67.
Rankama K. Geologic evidence of chemical
composition of the precambrian atmosphere // Geol.
Soc. Amer.
Spec. Pap.
1955.
Vol. 62. P. 651
– 664.
225
Ratzel F. Sein und Werden der organischen Welt. Leipzig, 1869. 514 S.
Regenfuss H. Ursachen und Konsequenzen einer parallelen phylogenetischen
Aufspaltung von Parasiten und Wirten // Co-Evolution. 20-ste Phylogenet.
Symp. (Hamburg, 28 – 30 Nov., 1975). Hamburg; Berlin, 1978. S. 83 – 99.
Robinson R. The origins of petroleum // Nature. 1966. Vol. 212, N 5068. P.
1291 – 1294.
Robinson R. Origins of oil, a correction and further comment on the Brunnock
even C-number predominance in certain higher alkanes of african crudes and
on the biogenesis of nonacesane // Nature. 1967. Vol. 214, N 5085. P. 263.
RochlederA. Phytochemie. Leipzig, 1854. 270 S.
Rubey W. Geologic history of water // Bull. Geol. Soc. Amer. 1951. Vol. 62,
N 9. P. 1111 – 1147.
Rubey W. Development of hydrosphere and atmosphere with special reference
to probable composition of the early atmosphere // Geol. Soc. Amer. Spec.
Pap. 1955. Vol. 62. P. 651 – 654.
Schindewolf O. Neokatastrophism? // Zschr. Dtsch. Geol. Gesell. 1963. Bd
114, H. 2. S. 430 – 455.
Schopf J. Precambrian paleobiology: problems and perspectives // Ann. Rev.
Earth Planet. Sci. 1975. N 3. P. 213 – 249.
Schuhert Ch. The earth's changing surface and climate during geologic time
// The evolution of the earth and inhabitans. New Haven, 1923. P. 45 – 81.
Sestini F. Relazione tra il peso atomico e l'afficio fisiologico
degli elementi chimici // Gaz. Chim. Int. 1885. Bd 15. S. 107 – 132.
Shaw W. Studies in biogeochemistry. 1. A biogeochemical periodic table //
Geochim et cosmochim. acta. 1960. Vol. 19. N 3. P. 196 – 207.
Sherlock A. Man as a geological agent. London, 1922. 87 p.
Stanier R. La place des bacteries dans le monde vivant // Ann.
Inst. Pasteur. 1961. Vol. 101, N2. P. 297 – 312. .
Stanley S. M. Possil data and the Precambrian – Cambrian evolutionary transition
// Amer. J. Sci. 1976. Vol. 276. P. 56 – 76.
Steinberg R. Correlations between biological essentiality and atomic structure
of the chemical elements // J. Agric. Res. 1938. Vol. 57, N 6. P. 851 – 858.
Suess E. Die Entstehung der Alpen. Wien, 1875.168 S.
Suess E. Das Antlitz der Erde. Wien; Leipzig, 1909. Bd 3. 789 S.
Tacke R., Wannagat V. Syntheses and properties of bioactive organosilicon
compounds // Top. Curr. Chem. 1979. Vol. 84. P. 7 – 75.
Tansley A. The use and abuse of vegetational concepts and terms // Ecology.
1935. Vol. 16, N2. P. 284 – 307.
Teihard de Charden P. Geobiologie et „Geobiologia” // Geobiologia.
1943. Vol. 1. P. 1 – 5.
Thatcher R. A proposed classification on the chemical elements with respect
to their functions in plant nutrition // Science. 1934. Vol. 79, N 2056.
P. 463 – 466.
The evolving biosphere. New York, 1981. 311 p.
Thoday J. Natural selection and biological progress // A centure of Darwin.
Cambridge (Mass.), 1958. P. 368 – 378.
Urbanek A. Rewolucja naukowa w biologii. Warszawa, 1973. 238 S.
Urey H. On the early chemical history of the earth and the origin of life
// Proc. Nat. Acad. Sci. 1952. Vol. 38. P. 351 – 363.
(Urey H.). Юри Г. Первичные атмосферы планет и происхождение жизни // Возникновение
жизни на Земле. М., 1959. С. 20 – 27.
Wahlert G. Phylogenie als okologischer Prozess// Naturwiss. Rdsch.
1973. Bd 26, H. 6. S. 247 – 250.
Wahlert G. Co-Evolution herrscht uberall // Co-Evolution. 20-ste Phylogenet.
Symp. (Hamburg, 28-30 Nov., 1975). Hamburg; Berlin, 1978. S. 101 – 125.
Walther I. Einleitung in der Geologie als historische Wissenschaft. Iena,
1893 – 1894. XXV+1017S.
Walther I. Mediterranen: Geobiologische Untersuchungen uber Gestaltung
und Besiedlung des mediterranen Lebensraumes. lena, 1936. 587 S.
226
Webb D., Fearon W. Studies of the ultimate composition of biological material
// Proc. Roy. Dublin Soc. Sci. 1937. Vol. 21. N 3. P. 487-504.
Westbrock P. Life as a geological force: New opportunities for paleontology
//Paleobiology. 1983. Vol. 8, N 2. P. 91 – 96.
Whitfleld M., Watson A. The influence of biomineralization on the composition
of seawater // Biomineralization and biological metal accumulition. Reidel,
1983. P. 86 – 93.
Whittaker R. New concepts of kingdoms of organisms // Science. 1969. Vol.
163. P. 150 – 160.
Whittaker R. Communities and ecosystems. London, 1970. 162 p.
Whittaker R., Margulis L. Protist classification and the kingdoms
of organism // Biosystems. 1978. Vol. 10, N1. P. 3 – 18.
227
ИМЕННОЙ УКАЗАТЕЛЬ
Азимов А. 101
Александров А. А. 25
Александров В. Г. 102
Александрова В. Д. 11
Алексеев А. П. 192
Анатольева А. И. 137
Андервуд В. 76
Андерсон И. 99
Андрее К. 99
Андрусов Н. И. 49, 97, 134
Анри В. 51,153
Анучин Д. Н. 9
Аристовская Т. В. 137
Арманд Д. Л. 6, 51
Арнольди К. А. 144
Аррениус С. 41
Архангельский А. Д. 96, 97, 134
Аткинс В. 106
Базилевич Н. И. 144, 150 – 153, 155,169
Банников А. Г. 197
Баринов Г. В. 126, 127
Барнхорт Э. 187
Барсков И. С. 4, 126, 139, 183
Бауэр Э. 36, 154
Бах А. Н. 42
Бгатов В. И. 137
Бей-Биенко Г. Я. 20
Бекетов А. Н. 39
Беклемишев В. Н. 11
Белая М. П. 105
Белов Н. В. 52, 160, 161
Белова Е. А. 89
Беляев Д. К. 201
Берг Л. С. 39, 60, 134
Бергман Г. 105
Бергсон А. 57
Берзиня Я. М. 63, 81
БеркнерЛ. 126, 127
Берман 3. И. 192, 200, 201
БерналДж. 6, 59, 182
Бернар К. 41
Бертран Ш. 134
Бертранд Г. 63
Бетехтин А. Г. 134
БлохинаР. И. 63, 81
Блюменфельд Л. А. 71
Бовен Г. 72, 109, 120
Богданов Ю. А. 130
Боголепов К. В. 138
Богоров В. Г. 150, 151, 154
Бойрлен К. 134, 138, 193
Бойченко Е. А. 96, 119 – 121
Больцман А. 51, 153
Большаков В. Н. 195
Борисенко Е. Н. 82
Боровик-Романова Т. В. 63, 89, 116
Браун Дж. 127
Брода Э. 156
Броунов П. И. 9
Бруновский Б. К. 105, 106
Брэдшоу А. 63, 91
Будыко М. И. 22, 25 – 27, 123, 125 – 128,
154, 158, 198
Бунак В. В. 192
Буссенго Ж. 48
Буханан Дж. 70
Бушинский Г.
И. 134
Быков Б. А. 177
Быховер Н. А. 121
Бютчли О. 98
Бюффон Ж. 7
Вавилов Н. И. 89, 194
Вагнер Г. 9
Вадковская И. К. 81, 169
Вайскопф В. 198
Валерт Г. 4
Вальтер И. 9, 32, 131, 134
Валяшко М. Г. 130, 131, 188
ВандельА. 193
Варрениус Б. 7
Вассоевич Н. Б. 6, 7, 9 – 11, 25, 27, 47,
138, 149, 153, 187
Верзилин Н. Н. 25, 127,128, 137, 161
Вернадский В. И. 3, 6, 7, 9 – 12,
14, 15,
20, 21, 25, 26 – 54, 56 – 62, 65, 71, 73,
74, 78, 83, 89, 93, 95 – 97, 104 – 109, 110, 114 – 116, 123, 125, 129
– 131,
133, 141, 143, 145, 147, 148, 164,
166, 169, 174, 177, 179, 180, 193, 202
Вестброк П. 51, 140
Вик д'Азир 7
Викман Ф. 149
Виленский Д. Г. 141
Виллиамсон В. 177
Вильзер Ю. 73
Вильямc В. Р. 25, 49, 101, 141 – 143, 166,
167, 168, 170
228
Винберг Г. Г. 150, 151
Вине Г. 93
Винер Н. 162
Виноградов А. П. 3, 6, 20, 25, 31, 60,62, 63, 65 – 69, 71, 72 – 74, 77 –
81, 83, 89, 95, 96, 101, 102, 105, 109 – 115, 117, 118, 124, 125, 130, 131,
134, 141, 166, 168, 188
Виноградов В. И. 127, 130, 133, 148
Виноградов X. Г. 105
Виноградский С. Н. 86, 97
Вишаренко В. С. 185
Владышевский Д. В. 197
Водопьянов П. А. 25, 148
Войнар А. И. 62,69, 76
Войткевич Г. В. 45, 160
Волобуев В. Р. 141
Вологдин А. Г. 25, 103, 118, 134, 148, 187,188
Вольскис Р. С. 20, 153
Вольфкович С. И. 96, 103
Воронков М. Г. 101, 102, 114
Воронцов Н. Н. 77
Вудвелл Дж. 17
Вудворт Д. 7
Высоцкий Б. П. 7
Высоцкий Г. Н. 9, 141
Гавриленко Е. С. 130,131
Галл Я. М. 35
Гаррелс Р. 133, 140
Гаузе Г. Ф. 19, 34
Гаффрон Г. 182
Гегамян Г. В. 6,10,61
Гейдройц К. К. 141
Геккер Р. Ф. 135
Гельмгольц Г. 41, 51
Гендерсон Л. Ж. 64, 65
Генералова В. Н. 139
Герасимов И. П. 25, 142, 166, 168
Геттон Дж. 7
Гиляров А. М. 13, 25, 171, 178
Гиляров М. С. 18, 22, 25, 61, 141, 144,
177, 204
Гинзбург А. И. 138
Гирусов Э. В. 15, 25
Глазовская М. А. 142 – 144, 169
Глембоцкий
Я. Л. 201
Глесснер М. 187
Глинка К. Д. 141
Годлевский М. Н. 134
Голланд Г.
51
Голубец М. А. 6,11, 22, 25, 27, 171, 177,
182
Гольдшмидт В. М. 62, 65, 71, 74, 100,
106, 107
Гордеев Д. И. 7, 96
Горецкий Ю. К. 134
Горский И. И. 135, 138
Горшкова Т. И. 105
Гофф К. 7
Григорьев А. А. 25, 51, 124
Гриффит Г. 88
Гроссгейм В. А. 135
Гумбольдт А. 7, 63
Гусева А. Н. 138
Густун М. И. 63, 81
Гюйгенс X. 7, 41
Давиташвили Л. Ш. 25, 96, 148
Дана Д. 38
Дарвин Ч. 14, 16, 31, 35, 49, 55, 141, 165,
176, 194
Деборин А. М. 40
Депере Ч. 38
Джон Ф. 63
Джохансон Д. 192
Дикерсон Р. 45
Дирш В. М. 105
Добржанский Ф. Г. 61, 162, 176
Добровольский В. В. 169
Добродеев О. П. 126,
127
Дозе К. 45., 70
Докучаев В. В. 9, 49, 55, 141, 145,169
Дроздова Т. В. 96, 103, 139
Друшиц В.
В. 184
Дубров А. П. 45
Дювиньо П. 3, 59, 154, 199, 200
ДэлиР. 108, 134
Дюма
Ж. 48
Егунов М. А. 49, 134
Емец В. М. 63, 116
Ермоленко М. Т. 43
Ефремов Ю.
К. 6, 11
Жарков М. А. 137, 138
Жемчужников Ю. А. 25, 134, 138
Жерехин В. В. 20, 198
Жуковский П. М. 194
Жулидов А. В. 63, 116
Забелин И. М. 165,182
Завадский К. М. 3,13,39,43, 59, 61, 82,
156, 180, 184, 190, 198, 204
Заварзин Г. А. 43, 118, 124, 125, 183,
187
Залесский М. Д. 134, 138
Захаров И. А. 197
Захарова Л. Н. 119
Зелинский Н.
Д. 60
Зенкевич Л. А. 115, 150, 151, 157, 177,
190
Зернов С. А. 60
Зонн С. В. 144
Зюсс Э. 8, 9
229
Иванов А. Н. 6, 7, 9, 10,27, 47
Имшенецкий А. А. 86
Исаев С. А. 100
Исаченко Б. Л. 134
Йенсен Е. 151
Казанский Ю. П. 116, 127, 132
Калабухов Н. И. 157
Калп Дж. 130
Кальвин М. 70
Камен М. 124
Камшилов М. М. 3, 4, 6, 12, 19, 23, 25 –
27, 40, 43, 44, 59, 60, 77, 162 – 164,
166, 171, 174 – 176, 181, 192, 199,
200, 202, 204
Кантор М. И. 134
Карпинский А. П. 100
Карцев А. А. 138
Квасов
Д. Д. 125
Квентон Р. 98
Кидсон Э. 63, 106
Кист А. А. 69
Кларк Ф.
98
Клосс К. 106
Кобленц-Мишке О. И. 150, 151
Ковальский В. В. 25, 43, 62, 63, 76,
81 – 90, 92, 93, 105, 106, 115, 116,
169, 181
Ковда В. А. 6, 10, 25, 49, 76, 141, 143 –
145, 152, 161, 162, 166, 170 – 172
Кодина Л. А. 103, 139
Козлова Л.
Е. 115
Козо-Полянский Б. М. 193, 194
Колчинский Э. И. 4, 24, 35,
44, 59,
82,
179, 180, 190
Комаров В. Л. 31, 52, 154
Кононов М. М. 144
Коп Э.
38
Корде К. Б. 135
Кордюм В. А. 164
Коржинский Д. С. 160
Коржинский
С.
И. 141
Коржневская Н. С. 138
Коржневский В. В. 92
Коссович П. С. 141, 143
Костицын В.А. 25, 123
Костычев С. П. 86
Кравков С. П. 141
Красилов В. А. 22, 165
Красильников
Н. А. 144
Крашенинников И. М. 141
Крепс Е. М. 71
Криволуцкий Д. А.
92, 116
Крик
Ф. 45
Крумбейн В. 117
Крупаткина Д. К. 151
Крылов А. Н. 60
Крылов И. Н. 118, 125, 127, 187,
188
Кузнецов В. Г.116
Кузнецов И. Г. 102, 114
Кузнецов С. И. 134, 137,
188
Кулагин Ю. 3. 196
Кунашева К. Г. 105
Куффер А. 99
Кэннон Э. 88
Лавелок Дж. 48, 129
Лавренко Е. М. 11, 25, 33, 154
Лавуазье А. 7
Лайель Ч. 55
Лалл Р. 108
Ламарк Ж.-Б. 7,8, 55
Ламонт К. 8
Ламсден Дж. 121
Ланге Р. 100
Лапо А. В. 6, 10, 11, 12, 25, 27,103, 135,
137,148,171
Лаптев И. Д. 200
Лебедев В. И. 160, 161
Лебедянцев А.
Н. 59, 105
Левандо Е. П. 135
Левтгардт Ф. 69
Левченко В. Ф. 4,
13, 156,
171
Лейфман
И. Е. 138
Леконт Д. 55
Лепешинская О. Б. 142
Леруа Э. 51, 57, 58
Летунова
С. В. 63, 86 – 88
Либих Ю. 63
Ливанов Н. А. 157
Линдеман Р. 178
Линней К. 7,
141
Лисицын А. П. 137
Личков Б. Л. 25, 27, 37, 134, 135
Ловенштам
X. 104,113, 117
Ломоносов М. В. 7
Лопатин Н. В. 137, 138
Лотка
А. 52, 200
Лотси Я. 164
Лукашев К. И. 25, 81, 144, 169
Лукашевич И. Д. 9, 41
Лукин Е. И.
77
Лунде Г. 106
Лысенко Т. Д. 142
Львов А. 71
Любименко В. Н.
124
Люкс
Ф. 93
Мазинг В. В. 171, 177
Маккензи Ф. 133, 140
Макхарт Дж. 63, 106
Малюга Д.
П. 63, 89, 105
Майская С. М. 103, 139
Маргелис Л. 48, 117
230
Марковский Б. П. 135
Марс К. 149
Марш Г. 55
Маршалл Л. 126,
127
Маслов В. П. 134
Массет А. 127
Маттей Р. 180, 181
Машковцев
А.
А. 74
Медников
Б. М.
13, 44, 118
Мейер Ю. 52, 153
Мейер-Абих А. 148
Мейсон Б.
51
Менделеев Д. И. 62, 64, 68
Меншуткин В. В. 21
Меррей Дж.
98
Мирзоян Э. Н. 96
Митчелл П. 88
Мишустин Е. Н. 86, 87, 144
Моисеев
Н. Н.
25, 123
Мокринский
В. В. 138
Молдаванцев Е. П. 134
Монин А. С. 161
Морозов
Г. Ф. 9
Морозов Н.
Н. 101
Морозов Н. П. 166
Мочалов И. И. 28
Мушкетов И. Ф.
9
Мюнтц А. 99
НадсонГ. А. 49, 97, 134
Назаров А. Г. 28
Назаров В. И.
193
Наливкин Д. В. 134, 135, 138
Наумов Н. П. 6, 17,
19,43, 83,
171, 174
Неаполитанская
В. С.
27
Неручев С. Г. 127, 138, 150
Неуструев С. С. 141
Никольский
Г. В. 196
Ничипорович А. А. 20, 25
Новогрудский Д. И. 40
Ноддак
В. 63, 106
Нопча Ф. 73
Обут А. М. 135
Одинак А. П. 6, 11
Одум Ю. 39, 47, 93
Олейников
А. Н. 125
Опарин А. И. 42 – 45, 71, 124
Оргел Л. 45
Оуэн Т. 126, 129
Павлов А. П. 55
Пак А. И. 137
Панфилов Д. В. 44
Парамонов
А. А. 192
Пастер Л. 41, 127
Патти Г. 43
Пачоский И.
К. 141
Пейве Я. В. 88, 144
Перельман А. И. 6, 12, 25, 27, 76, 77, 82, 121, 132,
133, 143, 153, 155, 161, 162, 164, 166, 169, 170,
188
Перфильев Б. В. 134, 144
Петров В. П. 137
Петрунина Н. С. 63, 89
Пианка Э. 83
Пири Н. 71
Плотников В. В. 177
Подолинский С. А. 51, 153
Покаржевский А. Д. 92, 116
Полынов Б. Б. 25, 49, 52, 74 – 77, 134, 141, 143,
144, 153, 160, 169
Полянский Ю. И. 189,192
Помпеску И. 100
Поннамперума С. 70
Попов В. А. 196
Посохов Е. В. 49, 127, 130, 132
Потонье Г. 134, 138
Прасолов Л. И. 141
Прат С. 90
Пресман А. С. 22
Принн Р. 126,129
Пристли Дж. 63
Пул Дж. 51, 125
Пустовалов Л. В. 50, 130, 131, 134
Пушева М. А. 119
Рабинович Е. 151
Раецкая Ю. И. 63,81
Райков И. Б. 189
Райли Дж. 151
Ранкам К. 149
Ратцель Ф. 7, 9
Ревердатто В. 22
Реди Ф. 41, 42
Рейндольс Д. 101
Реклю Э. 7
Ремедже X. 63, 106
Ремезов Н. П. 141, 143, 144
Рено Б. 134
Ржанникова Л. Н. 138
Риклефс Р. 158
Рихтер Г. 41
Риш М. А. 63, 92
Роде А. А. 25, 141 – 143
Родендорф Б. Б. 20, 198
Родин Л. Е. 151, 155
Роза Д. 193
Розанов А. Н. 141
Розов Н. Н. 151
Ронов А. Б. 25, 121, 127,128, 130, 134,
137, 146, 149, 188
Рубен С. 124
Руби В. 51, 125,
130
Руттен М. 70, 124, 127, 183, 189
Рябчиков А.
М. 152
231
Садиков В. С. 105
Саенко Г. Н. 119 – 121
Самарина И. А. 63
Самойлов Я. В. 25, 49, 52, 95 – 104, 114, 134
Сапожников Д. И. 124, 181
Сауков А. А. 121, 134, 137, 161, 169, 188
Свирижев Ю. М. 25
Северцов А. Н. 38, 39, 157, 158, 174
Северцов С. А. 109
Селиванов Л. С. 105
Сеньковский Ю. Н. 134
Серавин Л. Н. 112, 189
Сибирцев Н. М. 9, 141
Сидоренко А. В. 25, 45, 51, 130, 138, 139, 146, 149,
166, 183
Сидоренко С. А. 25, 130, 138, 139, 146,
149, 183 ;
Симкин Г. Н. 19, 176
Симорин А. М. 63, 78
Синицын
В. М. 153
Синягин И. И. 141
Сканави-Григорьева
М. Р. 105
Скарлыгина-Уфимцева
М.
Д. 63, 89
Соболев Д. Н. 25, 37
Соколов Б. С. 4, 6, 10,
14, 25, 96, 103,
126, 127, 135, 139, 146, 165, 183
Соколов В. 125
Соколов Н. А. 49, 134
Соколов С. Я. 22
Сорохтин О.
Г. 129
Сочава В. Б. 25
Стайлс В. 68
Старобогатов Я. И.
4, 14, 21, 156, 171,
177
Степанов П. И. 138
Страхов Н. М. 25, 121, 130
– 132, 134 –
137, 140, 149, 183, 188
Суетова И. А. 149, 152
Сукачев В. Н. 6, 11, 25, 26, 34, 82, 85,
142, 165, 177
Султанов К. М. 100
Сутт Т.Я. 37,
202
Сытник (Ситник) К. М. 28, 171
Сьянедцкий
А. 7
Танг М. 154, 200
Танфильев Г. И. 141
Тарко А. М. 25
Татаринов Л. П. 179
Таусон В. О. 50,134, 138, 150
Тахтаджян А. Л. 112
Твенхофер У. 131
Тейс Р. В. 124
Тейяр де Шарден П. 38, 43, si, 57, 58
Тенсли А. 12
Теняков В. А. 139, 183
Терентьева К. Ф. 105, 106
Тимирязев К. А. 51, 124, 153
Тимофеев П. П. 153,
161
Тимофеев-Ресовский Н. В. 10, 11, 19,
26, 37, 61, 77, 171, 176, 204
Титов И. А. 25, 142,
167, 168
Толмачев А. И. 135
Томсон В. 51, 153
Трут Л.
Н. 201
Тугаринов А.
И. 134
Тудей
Дж.
156
Тюрин И. В. 144
Тюрюканов А. Н. 6, 10, 11, 162
Уголев А. М. 43, 171, 178
Удельнова Т. М. 119
– 122
Умов Н. А. 56, 153
Уокер Дж. Г. 129
Уолд Дж. 62, 69,
70, 101
Урысон М.
И.
192
Успенский В. А. 135, 138, 148, 149
Ушаков С.
А. 127
Уэбб Дж. 88
Федоров Н. Ф. 56
Федотов Д. М. 135
Фейбридж
Р. 182
Ферсман А. Е. 51, 52, 55, 59, 62, 65, 68,
69, 78, 97,107, 110, 154, 160, 169
Фестер
Г. 106
Фехнер Г. 63
Фирсов В. 101
Флинт
Ф. Е.
197
Флоренский К. П. 6
Флоркен
М. 44
Фокс
М. 63, 106
Фокс С.
45, 70
Фолкнер Д. 93
Франц В. 156
Хабаков А. В. 134
Хаин В. Е. 135
Хайлов К. М. 25
Хатин А. 99
Хатчинсон Дж. 61, 117, 204
Хахина Л. Н. 194
Хесин Р. В. 164
Хильми Г. Ф. 154
Хлопин В. Г. 160
Хозацкий Л. И. 179, 186
Холдейн Дж. 124
Холодков Ю. И. 45
Холодный Н. Г. 43, 144
Холодовский Б. А. 33, 105
Хохряков А. П. 22, 150, 176
Хэдсон В. 34
Цветков Л. 4, 44
Цейтлин В. Б. 151
Циолковский
К. Э. 56
232
Чебаевская В. С. 63, 81
Чернов Ю. И.
13, 22, 25, 43, 177
Черногорова С.
М. 119
Чижевский
А. Л. 23, 45
Шапоренко К. К. 3
Шапошников Г. X. 43
Шарыгин С. А. 63, 92, 116
Шварц С. С. 4, 19, 20, 25, 26,
39, 53, 61,
63, 74, 82, 92, 93, 157 – 159,
171,
192, 196, 199, 200, 204
Швец З.
М. 149, 153
Швецов М. С. 134
Шилова
Е.
И. 10, 25,
142, 166, 170
Шиндевольф О. 193
Шипунов
В. Я. 10,
27, 123
Школьник М. Я.
63, 89
Шкорбатов Г. Л. 53
Шмальгаузен
И. И.
24, 39, 60, 74, 156,
197
Шопф Дж. 120
Шоу В. 62, 69, 70
Шпренгель К. 63
Шредингер Э.
154
Штейдманн Е. 134
Штиман Е. 151
Шукарев С. А. 62, 69
Шустер П.
44
Шухерт
Ч. 55
ШушкинаЭ.
А. 151
Щепотьев А. 98
Щербак Н. П. 27
Щербаков А. В. 153, 161
Эгами Фудзио 76
Эйбельсон П. 139
Эйген М. 44
Элтон Ч. 177
Юри Г. 125
Юсуфов А. Г. 4, 61
Юферова С.
Г. 119
Яблоков А. В. 4, 61, 77,
192, 195, 201
Яншин А.
Л. 25, 27, 59, 132, 135,
137, 138
Ярошенко П.
Д. 22,
177
Ясаманов
Н. А. 127
* * *
AbelsonP. 103,139
Allen
J. 51
Alvarez L. W.
22
Berkner L. 126, 150
Bertrand D.
117
Bezold A. 95
Bowen H. 72, 76, 109
Bradschaw A.
91
Brown
G. 127
Cloud P. 43, 44, 103, 129, 140
Crick F. 45
Daly R. 108
Degens
E. 103
Dickerson
R. 45
Dobzhansky
Th. 162, 176
Dose K. 40
Dunbar M. 22
Ehilich A. 196
Ehrlich P.
196, 202
Fearon W. 68
FiermesR.
43, 198
Franz V.
156
Frey-Wisling
A. 68
Garrels R.
129, 130,
133, 140
Gribben J.
125, 129
Guntau
M. 54
Hart M.
129
Holland
H. 51,
125, 129
Hutchinson G.
117,
118
Huxley
J. 39,
200
Kamen M.
117
Krumbein M. 117
KulpJ.
130
Lamarck
J.-B.
7
La
Mont
C.
8
Leakey
R.
192
Le
Conte
J.
55
Le
Roy
E.
57
Leuthardt
F.
69
LewinB.
R.
164
LindemanR.
178
Lotka
A.
52,161,
200
Lovelock
J.
48,129
Lowenstamm
H.
A.
103,
104,
113,
117,
140
LullR.
108
Lyell
Ch.
55
MacKenzie
F. 130, 133, 140
Margulis L. 104,112
Marshall L.
126, 150
Mason
B. 68
Mattey R.
180
233
MayR.
22
Meyer-Abich
A. 148
Money A.
202
Monty
C. 140
Musset A.
127
Nelson
M. 51
Nopfisa F.
73
Orgel L. 45
Paulus
H. 20
Poole J.
51, 125
Prat S.
90
Rankama K.
124, 125
Ratzel F.
7
Rauchfuss
H. 40
Robinson R.
45
Rochleder
A. 95
Rubey W. 51, 124,
125, 130
Schindewolf
O. 118
Schopf J.
103, 187
Schuchert
Ch. 55
Sestini F.
64
Shaw W.
69, 70
Stanier R.
187
Stanley
S. 118
Steinberg
R.
68
Suess
E. 8,
9
Tacke R.
114
Tansley
A. 12
Teihard
de Charden
P. 146
Thatcher
R. 68
Thoday
J. 156
Urbanek
A. 54
Urey H.
124, 125
Wahlert G.
4, 19, 20
Walther I.
134, 146
Wannagat
V. 114
Webb
D.
68
Westbrock P.
51, 140
Whittaker R.
112, 151,
189
234
ОГЛАВЛЕНИЕ
Стр.
Предисловие ................................................................................................................................................................
3
Глава
1.
Исходные
положения ...................................................................................................................................
6
1.1.
О
структуре
и
функциях
биосферы .....................................................................................................................
6
1.2.
Эволюция
биосферы
(общие
представления) ..................................................................................................
13
1.3.
Взаимосвязь
эволюции
видов
и
развития
биосферы ......................................................................................
18
1.4.
Этапы
и
направления
в
изучении
эволюции
биосферы .................................................................................
25
Глава
2.
В.
И.
Вернадский
об
эволюции
биосферы ................................................................................................
27
2.1.
Идеи
о
взаимодействии
эволюции
видов
и
эволюции
биосферы
.................................................................
28
2.2.
Гипотеза
о
возникновении
биосферы ...............................................................................................................
40
2.3.
Основные
тенденции
в
эволюции
биосферы ..................................................................................................
46
2.4.
Учение
о
ноосфере
как
заключительном
этапе
эволюции
биосферы
............................................................ 54
Глава
3.
Изучение
геохимических
факторов
эволюции
биосферы ........................................................................
62
3.1.
„Химическая
предопределенность”
живого
вещества .....................................................................................
63
3.2.
Общие
закономерности
химической
дифференциации
живого
вещества
в
биосфере .................................
71
3.3.
Исследование
биогеохимических
провинций
и
их
роли
в
эволюции
биосферы .........................................
77
3.4.
Геохимические
факторы
микроэволюции
и
ее
влияние
на
биогенные
циклы
элементов...........................
83
Глава
4.
Эволюция
биогеохимических
концентрационных
функций
биосферы .................................................
95
4.1.
Роль
палеобиогеохимических
исследований
в
изучении
эволюции
концентрационных
функций
биосферы ...................................................................................................................
96
4.2.
Проблема
эволюции
биогеохимических
функций
биосферы
в
трудах
В.
И.
Вернадского ......................................................................................................................................................
104
4.3.
Учение
А.
П.
Виноградова
об
эволюции
химического
состава
организмов
и
выполняемых
ими
геохимических
функций .......................................................................................................
109
4.4.
Основные
направления
современных
исследований ....................................................................................
115
Глава
5.
О
преобразующем
влиянии
биологической
эволюции
на
геохимическое
строение
биосферы ...............................................................................................................
123
5.1.
Влияние
эволюции
живого
на
газовый
состав
атмосферы
....................................................................
123
5.2.
Жизнь
и
эволюция
гидросферы ....................................................................................................................
129
5.3.
Биогенные
факторы
эволюции
литогенеза ...............................................................................................
134
5.4.
Роль
биотических
агентов
в
эволюции
почв
.............................................................................................
141
Глава
6.
Первые
шаги
к
изучению
главных
тенденций
в
эволюции
биосферы
.......................................
147
6.1.
Эволюция
общей
биомассы
биосферы
и
ее
биологической
продуктивности
.....................................
147
6.2.
Изменения
энергетики
биосферы ................................................................................................................
152
6.3.
Изменение
информационного
„фонда”
биосферы ....................................................................................
162
6.4.
Эволюция
биотического
круговорота .........................................................................................................
166
6.5.
К
вопросу
об
основных
этапах
развития
биосферы .................................................................................
179
Глава
7.
Особенности
органической
эволюции
в
современной
биосфере
(вместо
заключения) ..............................................................................................................................................
192
Послесловие .............................................................................................................................................................
204
Литература ...............................................................................................................................................................
206
Именной
указатель .................................................................................................................................................
228
Научное издание
Эдуард Израилевич Колчинский
ЭВОЛЮЦИЯ
БИОСФЕРЫ
ИСТОРИКО-КРИТИЧЕСКИЕ
ОЧЕРКИ
ИССЛЕДОВАНИЙ
В
СССР
Утверждено
к
печати
Институтом
истории
естествознания
и
техники
Академии
наук
СССР