
| Главная | Библиотека сайта | Форум | Гостевая книга |
АКАДЕМИЯ НАУК КАЗАХСКОЙ ССР
Научно-популярная серия
Г. А. ТИХОВ
Член-корреспондент Академии наук СССР, действительный член
Академии наук Казахской ССР
АСТРОБОТАНИКА

ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК КАЗАХСКОЙ ССР
Алма-Ата
1949
ВВЕДЕНИЕ
Слово «астроботаника» впервые произнесено мною публично на
заседании Президиума Казахского филиала. Академии наук СССР в конце 1945 года.
В печати это слово впервые появилось б марта 1946 года в газете «Казахстанская
правда≫. Писатель С. Полоцкий, после длительной беседы со мною, написал статью
под заглавием «Путь к светилам», где и изложил сущность моих основных научных
работ. Статья заканчивалась фразой: «Можно сказать, что мы стоим на пороге
новой науки — астроботаники».
* * *
§ 1. КОРНИ АСТРОБОТАНИКИ. Еще в 1909 году, наблюдая в Пулкове планету Марс, я заинтересовался вопросом о растительности на нем, а потому и стал изучать оптические свойства земной растительности, в частности, полосы поглощения хлорофилла у земной зелени. Поясню это несколько подробнее. Если мы посмотрим на зеленый лист в спектроскоп — прибор, разлагающий свет на составляющие его цвета, то увидим в крайних красных лучах темную полосу. Это значит, что зеленый лист поглощает эти лучи. Как показал знаменитый русский ботаник К. А. Тимирязев, именно за счет поглощения этих лучей зеленое вещество листа, так называемый хлорофилл, производит разложение углекислого газа нашей атмосферы на кислород и углерод, а из последнего и имеющейся в листе воды образуются первые органические соединения — крахмал и сахар. Таким образом, можно было ожидать, что и в спектре марсианской растительности должна наблюдаться полоса поглощения хлорофилла в крайних красных лучах. Вот отсюда и появился у меня интерес к изучению оптических свойств земной растительности. Я изучал и спектр поглощения спиртовых вытяжек хлорофилла из зеленых листьев и из игл хвойных растений, и спектр света, отраженного
3
листвой и хвоей, и спектральную прозрачность зеленых листьев.
Таким образом, я занялся вопросами ботаническими, но применял способы
только оптические. Это были, по существу, первые шаги применения ботаники
к астрономии, так что еще в 1909 году появились первые ростки астроботаники.
Прошло девять лет. В 1918 и 1920 годы были опять удобные для наблюдений положения
Марса, называемые «противостояниями»; я снова посвятил некоторое время
наблюдениям этой планеты в Пулкове. При этом много внимания было уделено поискам
полосы поглощения хлорофилла в темных местах Марса, считающихся растительными
покровами, т. е. по существу я опять занимался астроботаникой, не употребляя
этого слова.
§ 2. БОТАНИЧЕСКИЕ ИССЛЕДОВАНИЯ, СВЯЗАННЫЕ С АЭРОФОТОСЪЕМКОЙ. В 1930
году я был приглашен в основанный в Ленинграде Институт аэрофотосъемки для организации
фотометрической лаборатории. Как одну из важных тем этой лаборатории я ввел
изучение спектральной отражательной способности земных образований, в частности,
растительных покровов. Наблюдения состояли в том, что я и мои сотрудники фотографировали
спектр света, рассеянного разными почвами, травой, деревьями в разные периоды
их лиственного убора, ковры цветов и т. п. Полученные спектрограммы измерялись
на разных фотометрах и таким образом определялась яркость в разных лучах спектра
по отношению к яркости стандарта белизны, каковым является барит или магнезия.
Особенно много таких наблюдений сделал мой ученик и друг Е. Л. Кринов, опубликовавший
в 1947 году свои исследования в книге «Спектральная отражательная способность
природных образований».
Эти исследования, в частности, подтвердили давно известный факт, что зелень
очень сильно рассеивает инфракрасные лучи.Таким образом, у меня получился значительный
опыт по исследованию оптических свойств земной растительности.
§ 3. ЛЕКЦИИ В АЛМА-АТА. РОЖДЕНИЕ АСТРОБОТАНИКИ. С первого года пребывания
в Алма-Ата, именно с 1941 года, я неоднократно выступал с публичными лекциями,
причем темой их чаще всего был вопрос о возможности жизни на других планетах,
особенно на Марсе. Я касался исключительно вопроса о растительной жизни. На
своих лекциях я указывал, что мысль о земноподобной растительности на Марсе
встречается с несколькими затруднениями, главнейшее из которых состоит в том,
что места на Марсе, где могут быть растительные покровы, не обнаруживают сколько-нибудь
значительного
4
рассеяния инфракрасных лучей. После одной из лекций (в начале
1945 г.), когда аудитории было предложено задавать вопросы, агрометеоролог А.
П. Кутырева спросила меня: «Не считаете ли Вы возможным, что марсианские
растения путем длительного приспособления к суровому климату этой планеты утеряли
вредное для них свойство сильно рассеивать инфракрасные лучи, приносящие половину
всего солнечного тепла?». Я ответил, что эта мысль представляется мне очень
интересной, и что я над ней подумаю. На следующий день я пришел к выводу, что
проверить высказанное А. П. Кутыревой мнение можно следующим образом: надо сравнить
отражательную способность в инфракрасных лучах растений лиственных и хвойных.
У последних, особенно зимой, эта способность должна быть значительно меньше,
чем у лиственных, несущих свой зеленый наряд только в теплое время года. Чтобы
проверить эту мысль, я воспользовался результатами, тогда еще рукописными, Е.
Л. Кринова и нашел следующее: при сравнении березы и ели оказалось, что при
одинаковых значениях отражательной способности в синих лучах в инфракрасных
лучах эта способность была равна для березы 0,54, а для ели – 0,16, т. е. для
ели в три с лишним раза меньше, чем для березы.
При сравнении овса с тундровым можжевельником было обнаружено, что при одинаковых
значениях отражательной способности в зеленых лучах отражательная способность
в крайних красных лучах для овса равна 0,75, а для можжевельника – 0,24, т.
е. для можжевельника в три слишним раза меньше, чем для овса. Таким образом,
можно было думать, что отражательная способность марсианской растительности
в инфракрасных лучах еще меньше, чем у земных хвойных растений и даже, быть
может, близка к нулю. Итак, одно из важнейших затруднений для допущения земноподобной
растительности на Марсе было снято. Найденное объяснение и послужило толчком
для основания новой науки – астроботаники. Стало ясно, что для понимания
свойств марсианской растительности необходимо изучать оптические свойства земных
растений, живущих в суровых условиях высоких гор и Арктики.
§ 4. НАБЛЮДЕНИЯ НА ВЫСОТАХ ЗАИЛИЙСКОГО АЛАТАУ. Изучение оптических
свойств высокогорных растений было включено в тематический план Института астрономии
и физики на 1946, а затем и на 1947 годы. При исследованиях применялись походный
кварцевый спектрограф (Фиг. 1) с фотопластинками, чувствительными до инфракрасных
лучей включительно, и фотоаппарат с такими же пластинками и со свето-
5
фильтрами, дающими возможность делать снимки в разных лучах,
в том числе и в инфракрасных.
§ 5. НАБЛЮДЕНИЯ 1946 ГОДА. В 1946 году наблюдения производились в условиях
горной местности – в долине Терес-Бутак (2200 м над уровнем моря), в долине
реки Малой Алматинки (до высоты 3000 м над уровнем моря) и на морене Туюк-Су
(3400 м над уровнем моря), фиг. 2.
Большинство спектрограмм было получено научной сотрудницей В. С. Соколовой.
Обработка спектральных негативов 1946 года привела к открытию совершенно нового
явления. Оказалось, что некоторые из исследуемых цветов имеют в инфракрасных
лучах яркость, значительно превосходящую яркость баритового экрана. Вот эти
цветы: журавельник (Geranium grandiflorum) на Сухом хребте (высота 3000 м над
уровнем моря) с яркостью 1,7 – 1,5 по отношению к бариту; пион (Paeonia intermedia)
на высоте 1350 м с яркостью 1,6 и желтая алтайская фиалка (Viola altaica) на
высоте 3000 м над уровнем моря с яркостью 1,3 – 1,2 .
Кроме того, некоторые цветы показали в крайних красных лучах, на длине волны
694 mμ, сильное усиление яркости сравнительно с соседними местами спектра. Такова
была алтайская желтая фиалка и алтайская кортуза (Cortusa altaica) на Сухом
хребте.
Чем же можно было объяснить найденные явления? Это можно было объяснить тем,
что некоторые цветы излучают собственное свечение в инфракрасных и в узком участке
крайних красных лучей. Найденные особенности качественно и количественно различны
у разных растений, и это дало возможность на основе собранного материала опубликовать
статьи «Спектральный анализ растений»1 и «Спектральный анализ зелени
и цветов»2, понимая под этим возможность отличать одни растения от других
по их спектру, и располагать растения в классы по их спектральным свойствам.
В 1946 году были получены также очень интересные снимки лиственных и хвойных
деревьев в инфракрасных лучах, показывающие сильное рассеяние этих лучей растительной
зеленью. На фиг. 3 и 4 воспроизведены сделанные мною на обсерватории в Алма-Ата
снимки яблонь и груш в синих и в инфракрасных лучах, а на фиг. 5 и 6 – сделанные
В. С. Соколовой снимки тяньшанской ели также в синих и в инфракрасных.
1 Доклады Академии наук СССР, 1947, т. 57, № 7.
2 Известия Академии наук КазССР, серия астрономическая и физическая, вып. 3,
1948.
6
лучах. Инфракрасные снимки производят впечатление, что они
сняты зимой после обильного снегопада.
Между тем фотографии 3 и 4 были сняты одна вслед за другой в теплый солнечный
день 12 сентября 1946 г., а фотографии 5 и 6 – в теплый солнечный день 24 сентября
1946 г. близ лагеря «Локомотив» на высоте 2400 м над уровнем моря.
§ 6. НАБЛЮДЕНИЯ 1947 ГОДА. В 1947 году работы по наблюдениям производились
в тех же местах и с той же аппаратурой, что и в 1946 году. Как и в 1946 году
большинство спектрограмм было сделано В. С. Соколовой. Большую часть спектрограмм
обработал М. П. Остяков. Наиболее интересные выводы были таковы: листья ореха
на высоте 1350 м над уровнем моря дали яркость в инфракрасных лучах 1,20, цветок
лапчатки (Potentillai nivea) –1,50. Ель на высоте около 1500 м показала яркость
0,52, а на высоте 2200 м – 0,32. Это подтверждает высказанное уже мнение (в
§ 3), что отражательная способность зеленых растений должна быть тем меньше,
чем суровее климат, в котором живет растение.
В связи с этим очень интересно отметить следующее. В своей книге Е. Л. Кринов1
пишет, что хвойные породы лесных насаждений в зимний период дают в инфракрасных
лучах яркость, равную всего 0,19, тогда как в летний период яркость их в инфракрасных
лучах поднимается до 0,30. Сильное уменьшение отражательной способности хвойных
деревьев в инфракрасных лучах от лета к зиме подтверждается и нашими алмаатинскими
наблюдениями.
§ 7. УЧРЕЖДЕНИЕ СЕКТОРА АСТРОБОТАНИКИ. Быстрое развитие работ по астроботанике
показало на неооходимость организации специального сектора. Об этом мною была
подана докладная записка в Президиум Академии наук Казахской ССР 8 сентября
1946 года. Президиум одобрил мое предложение, и в 1947 году проект организации
специального сектора был направлен на утверждение соответствующих центральных
органов. В результате 11 ноября 1947 года Сектор астроботаники получил
официальное утверждение.
Итак, новая наука астроботаника получила официальное признание. На Земле был
учрежден впервые сектор, имеющий назначением изучать оптические свойства земных
растений для понимания растительных явлений на планете Марс и для суждения о
том, при каких крайних физических условиях могут еще существовать земноподобные
растения.
1 Е. Л. Кринов. Спектральная отражательная способность природных образований, 1947, стр. 96 – 97.
7
§ 8. ВЫСОКОГОРНЫЕ НАБЛЮДЕНИЯ 1948 ГОДА. ОТКРЫТИЕ ФЛУОРЕСЦЕНЦИИ
ЦВЕТОВ. Работы 1948 года проводились уже по смете Сектора астроботаники.
Одна часть научных сотрудников сектора была направлена на высоты Заилийского
Алатау, где и вела наблюдения под руководством З. И. Новиковой.
Другая часть в это время проводила наблюдения под руководством М. П. Остякова
в районе близ устья реки Оби.
Кроме продолжения наблюдений по спектральной отражательной способности растений
в плане работ стояли задачи фотографирования растений в инфракрасных лучах,
пользуясь собственным свечением растений. Иными словами, надо было проверить
и уточнить самосвечение, или флуоресценцию растений в инфракрасных
лучах.
Для этого были построены так называемые «ф л у о р е с ц е н т н ы е» ящики
по типу, показанному на фиг. 7. Ящик не имеет дна. Нижнее отверстие ящика равно
50 × 40 сантиметров. Высота ящика — 50 сантиметров. На одной из сторон его сделан
скос, в который вставляется плоско-параллельный стеклянный сосуд с внутренним
расстоянием между стеклянными стенками в 2,5 сантиметра, наполняемый водным
раствором медного купороса. Этот раствор имеет голубой цвет и при определенной
густоте этого цвета (концентрации раствора) раствор полностью поглощает крайние
красные и ближние инфракрасные лучи и в то же время хорошо пропускает все остальные
лучи спектра до ближних ультрафиолетовых включительно. До такой именно густоты,
и доводится цвет раствора в плоско-параллельном сосуде, который мы будем в дальнейшем
называть «жидким медно-купоросным светофильтром» или просто «жидким
светофильтром».
Сбоку в флуоресцентный ящик вставляется объектив «Биотар», имеющий светосилу
1 : 0,85. Объектив закрыт темнокрасным светофильтром, пропускающим только крайние
красные и инфракрасные лучи, т. е. те, которые поглощает жидкий светофильтр.
Таким образом, жидкий светофильтр на ящике и темнокрасный светофильтр на объективе
являются, как принято говорить, «дополнительными или скрещенными светофильтрами».
Если их сложить вместе, то они не пропустят никаких лучей, и через них не видно
даже Солнца. Объектив питает камеру, кассета которой заряжена пластинкой, чувствительной
к крайним красным и к ближним инфракрасным лучам.
Исследуемое растение покрывается ящиком, повернутым так, чтобы жидкий светофильтр
был обращен в сторону Солнца. Ящик окапывается внизу землею, чтобы в него не
проник
8
никакой свет, помимо того, который проходит через жидкий светофильтр.
Для проверки этого в ящик помещается рядом с растением толстая гипсовая пластинка.
Итак, вследствие «скрещенности» светофильтров на фотопластинку не проникает
ни один солнечный луч. Поэтому, если растение даст свой отпечаток, то только
за счет собственного свечения в крайних красных или инфракрасных лучах или в
тех и других вместе.
Весною 1948 г. были построены два флуоресцентных ящика с полным оборудованием.
Один из них был использован для проведения целой серии наблюдений в условиях
Субарктики, а другой ящик применялся в высокогорных условиях близ г. Алма-Ата.
День 3 августа 1948 г. оказался для Сектора астроботаники историческим. В этот
день на Сухом хребте был получен первый удачный негатив самосвечения цветка.
Это был колокольчик скученный (Campanula glomerata L .). Негатив был снят на
высоте 2800 м над уровнем моря при солнечном свете в 13 час. 41 мнн„ истинного
местного времени с выдержкой в 7 секунд.
Столь большая выдержка при громадной светосиле объектива показывает, что самосвечение
цветка было очень слабое. Увеличенный отпечаток с описанного негатива воспроизведен
на фиг. 8 . Мы видим, что никаких следов гипсовои пластинки на фиг. 8 нет. Следовательно,
цветок дал свое изображение благодаря самосвечению, или флуоресценции. Самосвечение
разных цветов имеет разную силу. Зеленые части растений флуоресцируют особенно
сильно. На фиг. 9 представлен снимок мелколепестника золотистого (Erigeron aurantiacus),
положенного на зеленую траву. Этот снимок сделан также на Сухом хребте 5 августа
1948 г. в 16 час. 4 мин. истинного местного времени с выдержкой в 15 секунд
при солнечном свете. Мы видим, что сравнительно с травой цветок светится очень
слабо, а потому дает лишь свои силуэты на светлом фоне травы. Здесь опять не
видно никаких следов гипсовой пластинки. Если растение флуоресцирует очень сильно,
то оно освещает своим светом гипсовую пластинку достаточно сильно для того,
чтобы от нее получилось изображение; при этом пластинка бывает всего ярче на
ближайшем расстоянии от растения и слабее на более далеких расстояниях. Это
очень хорошо видно на фиг. 10, где снят со значительной передержкой девясил
корнеголовый (Inula rhizocephala Schrenk.). Негатив снят на Джайлау (2200 м
над уровнем моря) 9 августа 1948 г. в 10 час. 47 мин. истинного местного времени
с выдержкой в 15 секунд при ярком солнечном свете. Все снимки вы
9
сокогорных экспедиций сделаны лаборантами М. К. Джунусовым
и Л. Д. Костенко.
Итак, известная уже раньше флуоресценция живой зелени в инфракрасных лучах под
воздействием более коротких волн получила наглядное подтверждение, а флуоресценция
цветов была открыта впервые.
Из этого видно, что астроботаника переросла уже свое первоначальное назначение
и вошла в задачи самой ботаники. Поднимается вопрос, какое физиологическое значение
имеет флуоресценция цветов? Как она связана с климатом, с временами года, с
отдельными этапами жизни растений? Поднимается также вопрос о привлечении насекомых
цветами. Не видят ли некоторые насекомые, например, пчелы, инфракрасных лучей?
Таким образом, здесь уже как бы перекидывается мост к науке о насекомых – энтомологии.
Чувствуется уже возможность связи с агрокультурой, садоводством. В самом деле,
если мы разместим большое число растений в классы по их оптическим свойствам
и сопоставим эти классы с уже известными классами, то, быть может, будет найдена
связь между оптическими свойствами растений и их морозоустойчивостью, засухоустойчивостью
и другими важными свойствами, и таким образом появится возможность выбора растений,
подходящих для того или другого климата по их оптическим свойствам.
Читатель, быть может, спросит: «А где же тут астроботаника, где тут применение
к исследованию растительности на планете Марс?». Ответ очень прост. Рано или
поздно, с развитием наблюдательной техники, можно будет получать спектры тех
мест планеты Марс, на которых предполагается существование растительности. Сличая
эти спектры со спектрами разных, земных растений, живущих в суровом климате
высоких гор или Субарктики, можно будет сделать более определенные заключения
о растительности на Марсе и даже, быть может, сказать, к какому семейству земных
растений всего ближе растения марсианские.
§ 9. СУБАРКТИЧЕСКИЕ НАБЛЮДЕНИЯ 1948 ГОДА. Наблюдения 1948 года в районе
Полярного круга дали возможность получить 34 спектральных негатива и собрать
богатый гербарий субарктический растительности.
По невыясненным еще причинам флуоресцентный ящик не дал положительных результатов.
Является ли это следствием каких-либо инструментальных (технических) ошибок
или таково свойство субарктической растительности – этого нельзя решить до новых
наблюдений.
Во всяком случае этот вопрос чрезвычайно интересен и
10
важен, так как между климатическими условиями в период жизнедеятельности растений, живущих на южных горах и в Арктике, существует очень большое различие. Дело в том, что, например, близ г. Алма-Ата (широта 43°) Солнце ежедневно восходит и заходит, тогда как в Арктике в период жизнедеятельности растений Солнце не заходит вовсе. Заимствую из книги академика А. А. Григорьева «Субарктика» (1946 г.) следующую таблицу:
Северная широта |
Число суток с незаходящим солнцем |
Северная широта |
Число суток с незаходящим солнцем |
75° |
108 |
69° |
62 |
73° |
97 |
68° |
54 |
70° |
73 |
67° |
32 |
В наблюдениях 1948 года принимала участие ботаник А. З. Соломченко.
Заимствую из ее отчета описание некоторых характерных особенностей субарктической
растительности:
«Обследованную территорию следует отнести к подзоне южной тундры – лесотундры.
Крупная древесная растительность заходит далеко в глубь полуострова (Ямал),
чуть дальше Нового порта (широта 67°40'); из хвойных здесь растут ель (Picea
obovata), и лиственница (Larix sibirica) высотой незначительно выше роста человека;
попадаются в большом изобилии кривые, извилистые стволики березы (Betula tortuosa),
а также ивы, разросшиеся по берегам речушек и по болотам.
Изредка можно видеть остатки деревьев, вероятно, погибших от изменившихся почвенных
условий, что доказывает наступление тундры на лес. Подобные массивы выделяются
отдельными островами с темнозеленым фоном, чередуясь со светлозелеными пятнами
более пониженных мест типичной кустарниковой тундры. Среди тех и других элементов
почва покрыта моховым и лишайниковым покровом.
Первые спектры отражательной способности растений были получены на коренном
берегу протока Полуя-Поляпты, включающем элементы лесной и тундровой растительности,
состоящей в основном из кустарников с преобладанием карликовой березки (Betula
nana); она группируется там, где много ягодников, и выделяется темнозеленым
фоном на всей территории; на близком расстоянии вырисовываются блестящие, круглые,
по краю пильчатые листочки ее. Местами бурый и вблизи красноватый тон дают кусты
багульника
11
(Ledum palustre); еще реже выделяются белые пятна ивы, очень
низкой, почти стелющейся по поверхности почвы, имеющей белые пушистые и серебристые
листья.
Во втором (считая сверху) ярусе выделяются полукустарники ягодника, окружающие
первый ярус: сизые листья с ягодами голубики (Vaccinium uliginosum), еще ниже
кожистые, блестящие листья вечнозеленой брусники (Vaccinium vitis idaea); в
виде ползучей елочки листья и стебли водяники (Empetrum nigrum) и толокнянки
(Arctous alpina). Среди такой тундры характерно наличие кочек, покрытых вышеописанным
ярусным покровом. Пространство между кочками занято сизыми пятнами лишайников
– Stereocaulon.
В третьем ярусе растительного покрова кочек обилие мхов: кукушкин мох (Polytrichium
strictum), Hypnum, Hylocomium. Среди кочек выделяются чуть покачивающиеся от
ветра метелки типчака (овсяница овечья, Festuca ovina L .); чрезвычайно обильны
зеленые «свечки» ползучих стеблей плауна; из лишайников обильны, кроме других,
ягели. Процент покрытия 95.
Незначительные пространства заняты луговинами, заселенными травянистыми растениями,
каковы вейник (Calamagrostis sp .), золотая розга (Solidago virga aurea), девясил
(Inula sp ). иван-чай (Epilobium angustifolium L .) и др.
В большом количестве встречается торфяной мох сфагнум (Sphagnum sp .), переплетающийся
с различными видами растений. Здесь встречаемся с клюквой (Oxycoceos palustre
sp .), розеточными листьями морошки (Rubus chamemorus) и грушанкой (Pirola sp
.).
Основной фон в верхнем ярусе создают кусты ивы с пушистыми белыми листьями;
в малом количестве присутствуют также карликовая березка, голубика (Vaccinium
uliginosum L.) и брусника (Vaccinium vitis idaea L .).
С приближением к открытому пространству воды господство занимает торфяной мох
– сфагнум, переплетаясь с коричневыми ниточками клюквы (Vaccinium oxycoccos
L.)».
Заимствую еще из книги академика JI. С. Берга «Географические зоны Советского
Союза» (1947 г.) некоторые места, важные для астроботаники.
«Растительность тундры, в соответствии с окружающей обстановкой, носит своеобразный
отпечаток... Весьма много вечнозеленых растений. Польза от этого та, что растение
может использовать солнечный свет, лишь только он появится, не затрачивая времени
на образование листьев. Многие растения растут дерновинками (например, куропаточья
трава – Dryas octopetala) или стелются по почве, например, полярные
12
ивы и березы, что, как думают, дает им возможность лучше использовать
летнюю теплоту почвы.
Как известно, растения можно разделить на две группы: одни требуют для цветения
и плодоношения, длинного дня (т. е. продолжительного освещения) и способны зацветать
даже при непрерывном освещении; они цветут, главным образом, летом. Другие же,
напротив, нуждаются в коротком дне, требуя перерыва освещения; они цветут у
нас осенью. К первой группе относятся, между прочим, злаки – пшеница, ячмень,
рожь, овес. Но есть и растения, для наступления цветения которых продолжительность
дня не имеет значения, например, однолетний мятлик (Роа annua). Растения тундры,
очевидно, относятся преимущественно к первой группе – это растения длинного
дня.
Для жизни растений (и животных) тундры имеет важное значение большая напряженность
ультрафиолетового излучения на севере, чем в средних широтах. При тех же высотах
Солнца ультрафиолетовых лучей (с длиной волны 290 – 320 mμ) на севере (68° с.
ш.) почти вдвое больше, чем под 47 – 54° с. ш. Это обстоятельство было открыто
сравнительно недавно (в 1927 г.). Как известно, тем же свойством отличаются
и высокогорные зоны. По мнению Л. Иванова (1928 г.), следует принять во внимание
значительную прозрачность воздуха на севере – почти полное отсутствие здесь
пыли.
Восковой налет, опушение листьев, красная окраска клеточного сока и оболочек,
блестящая, отражающая свет листва – все это служит для защиты хлоропластов1
от интенсивного света.
Растения тундры (как и высокогорные) имеют склонность расти подушками (например,
многие камнеломки – Saxifraga), что дает им, как полагают, лучшую защиту от
ветра, а следовательно, и от высыхания, столь губительного в тундре.
Большую роль в тундре играют лишайники. Особенно важное значение в качестве
корма для оленей имеет ягель или олений «мох» (Cladonia). О нем Шренк говорит,
что он для тундр важнее, чем хлеб для более умеренных широт.
Цветковые растения тундры отличаются обилием цветов, крупной их величиной и
яркостью окраски. С. Григорьев (1913 г.) на Канином носу видел цветы (цветочные
корзинки) ромашки диаметром до 8 см; на некоторых экземплярах этого растения
наблюдалось свыше двухсот соцветий. Тут же бросается в глаза обилие крупных,
ярко окрашенных цветов:
1 Белковые тельца, окрашенные в зеленый цвет красящим веществом – хлорофиллом.
13
желтой купальницы, ястребинки, крестовника, синюхи (Polemonium),
мытника, живокости и других.
На Гусиной Земле (Новая Земля) на сланце в щелях можно наблюдать группы ярких
цветов: синих незабудок (Myosotis alpestris), розовых и белых звездчаток (Silene
acaulis), почти совершенно лишенных листьев, с короткими стеблями. Они растут
так тесно, что цветы образуют как бы букеты, ярко выступающие на темном фоне
покрытой сланцем почвы (Житков и Бутурлин, 1901 г.).
Растения тундры удивительно приспособлены к перенесению морозов: на Чукотской
земле ложечная трава к наступлению зимы 1878/79 года была, по наблюдениям Чильмана,
в полном цвету застигнута морозами; тем не менее на следующий год она продолжала
вполне благополучно расти, перенеся морозы до 46°.
Некоторые растения севера лесной зоны перезимовывают в стадии цветения. Так,
у однолетних фиалок (Viola arvensis и Viola tricolor) в Петергофе можно наблюдать
открытые цветы в течение всей зимы. То же бывает у некоторых многолетников:
одуванчика, маргаритки и других».
Следует обратить особенное внимание на следующее. Как сказано выше, «красная
окраска клеточного сока и оболочек служит для защиты хлоропластов от интенсивного
света» и, в частности, вероятно, от интенсивного ультрафиолетового облучения.
Между тем, в книге академика А. А. Григорьева «Субарктика» (1946 г.) говорится,
что одним из приспособлений субарктических растений, защищающих их от избытка
ультрафиолетовых лучей, является усиление синего и фиолетового оттенков. В частности,
листья имеют фиолетовый оттенок, цветы незабудки не голубые, а темноголубые,
цветы тысячелистника не белые или розоватые, как в лесной полосе, а розовые
с фиолетовым оттенком, дельфиниум (шпорник) не синий, а густо темносиний и т.
п.
«В настоящее время можно считать доказанным, – пишет академик Григорьев, – что
изменения окраски являются приспособлением растений к облучению светом, изобилующим
ультрафиолетовыми лучами».
Я понимаю это так: если растение имеет темносиний цвет или фиолетовый оттенок,
то оно сильно отражает синие и фиолетовые лучи и, вероятно, соседние с последними
ультрафиолетовые лучи.
Как же понимать, что для той же цели может служить красная окраска клеточного
сока и оболочек? Я думаю, что клеточный сок и оболочки такой окраски поглощают
ультрафиолетовые лучи и не допускают их до хлоропластов. С другой
14
стороны, красный сок и оболочки свободно пропускают на хлоропласты
длинноволновую часть видимого спектра, особенно
важную для их жизненной деятельности.
Таким образом, две крайних окраски – сине-фиолетовая и красная – достигают
одной и той же цели. Этот вывод очень интересен во всех отношениях, но
он требует еще тщательной и углубленной проверки.
Интересно еще отметить, что исследованный в 1948 году лишайник цетрария (Oetraria
islandica), имеющий буроватый цвет, очень сильно рассеивает ультрафиолетовые
лучи, до 25%.
Мы сделали довольно подробные выписки о жизни растений в Субарктике, так как
суровый субарктический климат имеет хотя бы отдаленное сходство с еще более
суровым климатом Марса, и свойства субарктических растений могут приблизить
нас к пониманию особенностей растительности марсианской.
§ 10. ПРОДОЛЖЕНИЕ РАБОТ ПО ВЫСОКОГОРНЫМ НАБЛЮДЕНИЯМ 1948 ГОДА. В начале
августа 1948 года были получены первые снимки цветов в их собственном инфракрасном
свечении. Однако этот чисто качественный результат необходимо было перевести
в количественный, т. е. узнать, какую долю по отношению к рассеянному цветком
красному и инфракрасному свету составляет свет флуоресценции.
Для этого жидкий светофильтр на флуоресцентном ящике мы заменили красно-инфракрасным
светофильтром, т. е. тем самым, каким был закрыт объектив. Тогда и цветок и
гипсовая пластинка должны были дать свои изображения в отраженном ими красно-инфракрасном
свете, так как, при справедливости закона Стокса, флуоресценция, возбужденная
этими лучами, должна проявиться в лучах с еще-большей длиной волны, к которым
применяемые нами фотопластинки уже не чувствительны.
На фиг. 11 – 14 даны образцы снимков листвы и цветов растений при облучении
их через купоросный и через красно-инфракрасный светофильтры. На фиг. 11 и 12
показаны снимки жимолости (Lonicera Karelinii Bge), сделанные через купоросный
светофильтр и через инфракрасный. Во время съемки на растении были зеленые листья
и красные ягоды. На первом снимке видны только черные силуэты ягод и светлая
точка на одной из них; эта точка происходит от отражения на ягоде света флуоресценции
листьев. На правом снимке ягоды вышли светлыми, так как они были красные.
На фиг. 13 и 14 показаны снимки растения «Борец» (Асоnitum rotundifolium) в
полном цвету. Обстановка была такая.
15
Несколько дней перед съемкой шел снег. 14 сентября было ясно,
и цветы вышли из-под снега. На фиг. 13 видно, что флуоресцируют и листья, и
цветы. На фиг. 14 интересно то, что некоторые части цветов и цветоножек выглядят
гораздо ярче, чем плоская поверхность гипсовой пластинки. Однако правый боковой
край этой пластинки значительно ярче, чем ее плоская поверхность, и уже сравним
по яркости с наиболее яркими частями цветов. По всей вероятности, наиболее яркие
места цветов и гипсовой пластинки происходят от того, что падающий от Солнца
свет был близок к скользящему, почти зеркально-отраженному. Во всяком случае
этот вопрос требует дополнительных исследований.
Каково же соотношение яркостей цветка при облучении его отдельно через красно-инфракрасный
и через купоросный светофильтры?
Для ответа на этот вопрос мы располагаем следующими данными. Изображения большинства
цветов при освещении их через купоросный и через красно-инфракрасный фильтры
получаются приблизительно одинаковой плотности при следующих условиях: при освещении
через купоросный, светофильтр цветок фотографируется при полном отверстии объектива,
равном 65 мм и с выдержкой в 15 секунд, а при освещении через красно-инфракрасный
фильтр объектив диафрагмируется до диаметра 8 мм, и выдержка равна 0,5 секунды.
Плотность изображения на негативе пропорциональна площади объектива и, в первом
приближении, выдержке. Площади отверстий объектива в двух случаях относятся
как (65/8)2 = 66 и отношение выдержек равно 15/0,5 = 30.
Таким образом, для получения того же результата от флуоресценции надо дать экспозицию
в 66 × 30 = 1980 раз большую. Иными словами, флуоресценция цветов в крайних
красных и в инфракрасных лучах для большинства цветов приблизительно в 2000
раз слабее, чем свет этих цветов в тех же лучах при облучении их крайними красными
и инфракрасными лучами.
Нами было исследовано несколько десятков растений, и почти все они оказались
флуоресцирующими в большей или меньшей степени.
§ 11. ИССЛЕДОВАНИЕ СПЕКТРАЛЬНЫХ СВОЙСТВ РАСТЕНИЙ СПОСОБОМ СВЕТОФИЛЬТРОВ.
Для более быстрой разведки основных оптических свойств растений можно применять
фотографирование их через светофильтры, пропускающие тот или другой участок
спектра. При этом употребляются пластинки
16
чувствительные к лучам соответствующей длины волны. Так, например,
при фотографировании в инфракрасных лучах мы пользовались пластинками научно-исследовательского
кинофотоинститута (НИКФИ), называемыми «инфрахром», которые чувствительны
до длины волны 840 mμ. При фотографировании в красных лучах применялись пластинки
НИКФИ «панинфра», при фотографировании в зеленых лучах – пластинки
«ортохром» и для ультрафиолетовых лучей применялись обыкновенные
фотопластинки.
В качестве предмета сравнения (эталона) служит испытательная пластинка из смеси
барита и угля, устанавливаемая между ветками и цветами.
В 1948 году растения снимались нами и на горах, и в опытном цветнике, разведенном
около лабораторий Сектора астроботаники. Растения снимались в четырех участках
спектра: в инфракрасном, красном, зеленом и ультрафиолетовом.
При таких съемках растения непосредственно обнаруживают свои оптические особенности.
Так, мы уже видели на фиг. 3 – 6 , что зеленые деревья, кусты и трава очень
сильно рассеивают инфракрасные лучи.
На фиг. 15 и 16 воспроизведены снимки садового цветка «боярышня», или «принцесса»
(Cosmea), сделанные в красных и в ультрафиолетовых лучах. Рядом помещена круглая
угольно-баритовая пластинка в деревянной шкатулке. При непосредственном рассмотрении
цветка он имеет чисто белый цвет, как гипс. Фиг. 15, дающая снимок в красных
лучах, показывает, что в этих лучах некоторые цветки несколько ярче эталона,
другие – несколько слабее. В ультрафиолетовых лучах картина совершенно иная.
Эталон попрежнему светлый, а цветки более или менее темные. В Секторе астроботаники
имеется большое число негативов растений, полученных описанным способом, но
они еще ждут обработки.
§ 12. ЕЩЕ ДВА ОПТИЧЕСКИХ СВОЙСТВА МАРСИАНСКОЙ РАСТИТЕЛЬНОСТИ, ОТЛИЧАЮЩИХ
ЕЕ ОТ РАСТИТЕЛЬНОСТИ ЗЕМНОЙ. Мы видели (§ 3), как астроботаника объяснила
первое из затруднений, стоявших перед допущением того, что на Марсе существует
земноподобная растительность. Это затруднение состояло в том, что у марсианской
растительности нет сколько-нибудь заметного рассеяния инфракрасных лучей. Наше
объяснение состоит в том, что марсианские растения утеряли это свойство в процессе
длительного приспособления к суровому климату.
Мысль о существовании на Марсе земноподобной растительности наталкивалась еще
на два затруднения: на отсутствие в свете, рассеянном марсианской растительностью,
по-
17
лос поглощения хлорофилла и на преимущественно голубой цвет
этой растительности.
Займемся сначала первым из этих вопросов. Как мы уже кратко говорили (в § 1
), в длинноволновой части видимого света, рассеянного земною зеленью, более
или менее сильно ослаблены лучи некоторых длин волны. Всего сильнее ослаблены
красные лучи в пределах от 690 до 660 mμ, меньше ослаблены оранжевые лучи между
635 и 615 mμ и всего меньше желтые лучи между 600 и 585 mμ. При рассматривании
или при фотографировании спектра растений указанные места имеют вид более или
менее темных полос, называемых полосами поглощения хлорофилла.
У разных 'растений эти полосы занимают несколько иное положение, а слабые полосы
могут и совершенно отсутствовать. Вот, например, что дали мои старые наблюдения,
произведенные в Пулковской обсерватории еще в 1929 и 1930 годах, когда я начал
исследовать спектр рассеяния растений для понимания снимков земной поверхности,
сделанных с самолетов. Молодая листва березы (24 мая 1930 г.) дала середины
полоc поглощения хлорофилла на волнах 680, 630 и 595 mμ, старая листва березы
(24 сентября 1929 г.) – на волнах 678, 618 и 590 mμ. Молодая трава (24 мая 1930
г.) показала только главную полосу на волне 676 mμ.
Поглощение света хлорофиллом, правда, преимущественно в спиртовых его вытяжках,
особенно тщательно изучал еще в прошлом веке наш знаменитый соотечественник
Климент Аркадьевич Тимирязев. Он показал, что поглощение света главной полосой
хлорофилла имеет жизненное значение для растений. Как мы уже говорили
(в § 1), зеленое растение поглощает из воздуха углекислый газ, разлагает его
на кислород и углерод, кислород выделяет в воздух, а из углерода и воды образует
первые органические вещества – крахмал и сахар. Тимирязев и показал своими опытами,
что наиболее быстрое разложение углекислоты происходит именно под влиянием тех
лучей света, которые поглощаются главной полосой хлорофилла.
Уже давно появился интерес к тому, существует ли полоса поглощения хлорофилла
и у марсианской растительности. Еще в конце прошлого века французский астроном
Фламмарион писал, что полосу поглощения хлорофилла на Марсе откроют раньше,
чем линии поглощения кислорода в его атмосфере.
Вопрос о полосе поглощения хлорофилла на Марсе интересовал и Тимирязева. И вот
в 1909 году он пишет знаменитому исследователю Марса американцу Ловеллу письмо
с
18
вопросом, не наблюдается ли главная полоса поглощения хлорофилла
в спектре зеленых областей Марса.
Ловелл ответил Тимирязеву следующим образом: «Два года тому назад мой сотрудник
Слайфер делал снимки при помощи спектрографа с целью доказать присутствие хлорофилла
на Марсе. Он получил хорошие результаты для земных объектов, но для Марса дело
гораздо труднее, частью вследствие малого количества света, частью вследствие
особенного характера поставленной задачи. Растительность в сколько-нибудь значительных
количествах встречается только в некоторых частях диска, и расположить в этих
частях щель спектрографа почти невозможно, хотя со временем, может быть, мы
с этой трудностью справимся.
Вопрос этот занимает нас вот уже четырнадцать лет, и Слайфер уже много работал
с отраженным светом...»
В 1918 и 1920 гг. я неоднократно наблюдал в Пулкове спектр темных областей Марса
в окулярный спектроскоп, привинченный к 15-дюймовому рефрактору, и никаких затруднений
в наведении на щель спектроскопа темных мест Марса не встречал. Однако полосы
поглощения хлорофилла с уверенностью я не видел, но в спектре экваториальных
темных мест заметил особенно сильное поглощение в зеленых лучах. Это должно
давать темным местам Марса голубой цвет, что неоднократно отмечалось разными
наблюдателями.
Чем же объяснить, что до сих пор не получено решительных указаний на присутствие
главной полосы хлорофилла у марсианской растительности? Как может быть, чтобы
в течение целых 14 лет, да, повидимому, и до настоящего времени, на обсерватории
Ловелла, посвященной в основном исследованию планет и, в частности, Марса, не
удалось решить такую сравнительно простую задачу, несмотря на мощное оборудование
этой обсерватории?
Ответ может быть такой: полоса хлорофилла не найдена потому, что у марсианской
растительности ее нет.
Почему же у марсианской растительности нет этой полосы? Вот как я объяснял это
еще в 1946 году:1
«Вторым отличием марсианской растительности от земной является отсутствие в
первой главной полосы поглощения хлорофилла, лежащей в крайних красных лучах.
И это можно объяснить суровым климатом Марса, в котором, для разложения углекислоты,
растениям недостаточно поглощать только сравнительно узкую полосу спектра солнечного
света, а
1 Вестник Академии наук Казахской ССР № 9 (18), сентябрь, 1946, стр. 22.
19
необходимо воспринимать возможно более обширную область лучей.
Поэтому в растительности Марса должны быть ослаблены все наиболее длинноволновые
лучи видимого спектра, что дает этой растительности отмеченную многими наблюдателями
голубоватую окраску».
Необходимо было проверить это объяснение на земных деревьях северного типа,
имеющих голубой цвет. Такова, например, канадская ель (Picea Engelmanni). В
Алма-Ата на одном дворе растет такая ель приблизительно тридцатилетнего возраста.
Я поручил своему аспиранту В. С. Тихомирову снять спектр ели и для сравнения
– спектр сосны в университетском ботаническом саду. Для этих съемок применялся
спектрограф, дающий для главной полосы поглощения хлорофилла ширину в 2,5 миллиметра
на фотопластинке. На каждом из негативов снят также спектр света, рассеянного
гипсовой пластинкой. Снимки производились в ясный день около полудня, так что
условия освещения деревьев и гипса были одинаковы. Выдержка во всех случаях
была одинаковая, но менялись диафрагмы, обусловливающие светосилу спектрографа.
На фиг. 17 и 18 показаны увеличенные снимки спектра сосны и спектра канадской
ели. Мы видим, что у сосны полоса поглощения хлорофилла1 очень отчетливая, а
у канадской ели она едва заметна. Благодаря спектрам гипсовой пластинки свойства
полос хлорофилла у сосны и у канадской ели можно выразить в числах, подвергнув
негативы фотометрическим измерениям. Более определенным стандартом является
спрессованный порошок барита, а потому числа, полученные по отношению к гипсу,
приведены затем к бариту, с которым была сравнена гипсовая пластинка. Полученные
результаты представлены кривыми на фиг. 19.
Мы видим, что у сосны спектр сильно ослабевает в области главной полосы поглощения
хлорофилла, тогда как у канадской ели спектр сильно и равномерно ослаблен в
области красных, оранжевых и зеленых лучей. Таким образом, мое объяснение, высказанное
еще в 1946 году, полностью оправдалось.
Этот результат устраняет второе и третье возражения против существования растительности
на Марсе, и это является большим успехом астроботаники. Она уже вышла на широкий
научный путь, так как может не только объяснять, но и предвидеть неизвестные
явления природы.
§ 13. СВЯЗЬ АСТРОБОТАНИКИ С ДРУГИМИ НАУКАМИ. Мы уже видели, что астроботаника
вошла в тесную связь с
1 Несколько правее середины спектра.
20
ботаникой и открыла флуоресценцию цветов. Теперь перед нами
стоит задача – понять жизненное значение для растений флуоресценции их зелени
и цветов. Для решения этого вопроса у нас еще слишком мало данных, но все-таки
уже вырисовываются некоторые перспективы дальнейших исследований.
Мы спрашиваем себя, какое значение имеет это явление для теплового состояния
растения? Связано ли оно с коротким южным днем, или оно присуще и растениям
длинного субарктического дня? На этот вопрос мы надеемся получить со временем
определенный ответ.
Второй вопрос, интересующий сотрудников Сектора астроботаники, относится уже
к зрению насекомых, т. е. к науке, изучающей насекомых и носящей название энтомологии.
Не видят ли насекомые инфракрасных лучей? Если да, то цветы, сильно флуоресцирующие
в инфракрасных лучах, должны быть особенно привлекательными для насекомых, что
важно для опыления. В частности, не видят ли инфракрасных лучей пчелы, и какова
флуоресценция у особо медоносных цветов? Таким образом, астроботаника связывается
с пчеловодством, и мы проектируем установку в опытном цветнике Сектора астроботаники
нескольких ульев, чтобы следить за поведением пчел по отношению к цветам, изучаемым
нами на флуоресценцию.
Далее, нас интересует связь флуоресценции с морозоустойчивостью и засухоустойчивостью
растений. Если, тут будет обнаружена определенная связь, то это покажет практическую
значимость астроботаники.
Таким образом, перед астроботаникой раскрываются широчайшие перспективы не только
астрономические, но и в приложении к наукам, связанным с практическими целями.
ЗАКЛЮЧЕНИЕ
Мы видели, что корни астроботаники связаны с вопросом о растительности
на Марсе. В связи с этим понадобилось изучение оптических свойств земных растений.
Это изучение, в свою очередь, оказалось весьма важным для чтения аэроснимков,
и потребовалось расширение исследований для большого числа как растительных
покровов, так и других земных образований. Эти исследования опять вернулись
на Марс и позволили понять особенности марсианской растительности в отношении
инфракрасных лучей. Далее, отсутствие у марсианской растительности главной полосы
поглощения хлорофилла сделало необходимым обратить специальное внимание на эту
полосу у земных растений.
Наконец, исследование земных цветов открыло неизвест-
21
ную до сих пор флуоресценцию цветов в инфракрасных лучах.
Таким образом, наши исследования несколько раз перебрасывались с Марса на Землю
и обратно и тесно связали астрономию с другими науками. Можно даже думать, что
астроботаника раньше или позже сможет давать некоторые указания и советы ученым,
изучающим науки, имеющие непосредственное практическое значение, каковы ботаника,
энтомология и т. п.
Для дальнейших успехов чисто астроботанического характера необходимо продолжать
изучение оптических свойств земных растений как на высоких горах, так и в Арктике,
а также тщательно изучать Марс как по многочисленным литературным данным, так
и производя новые наблюдения над ним при помощи сильных астрономических инструментов.
* * *
В настоящее время отработаны спектры, полученные в Субарктике
в 1948 г. Оказалось, что многие из субарктических зеленых растений полосы поглощения
хлорофилла не имеют. Таковы: багульник (Ledum palustre) , мытник (Pedicularis
palustris), кукушкин мох (Polytrichum communis), карликовая береза (Betula nana)
и другие.
Недавно опубликованы спектральные наблюдения Марса, произведенные в 1948 г.
на американской обсерватории Мак Дональда при помощи громадного 84-дюймового
зеркального телескопа. Спектр области, соответствующей растительному, покрову,
полосы поглощения хлорофилла не обнаружил.
В июне месяце 1949 г. в «астроботанический» сад прилетел рой пчел, и мы его
поместили в заготовленный заранее улей.
СОДЕРЖАНИЕ
Введение .............................................................................................................................................................................................. 3
§ 1. Корни астроботаники ................................................................................................................................................................ –
§ 2. Ботанические исследования, связанные с аэрофотосъемкой ......................................................................................... 4
§ 3. Лекции в Алма-Ата. Рождение астроботаники ................................................................................................................... –
§ 4. Наблюдения на высотах Заилийского Алатау ..................................................................................................................... 5
§ 5. Наблюдения 1946 года ............................................................................................................................................................... 6
§ 6. Наблюдения 1947 года ............................................................................................................................................................... 7
§ 7. Учреждение Сектора астроботаники .................................................................................................................................... –
§ 8. Высокогорные наблюдения 1948 года. Открытие флуоресценции цветов .................................................................. 8
§ 9. Субарктические наблюдения 1948 года ............................................................................................................................... 10
§ 10. Продолжение работ по высокогорным наблюдениям 1948 года ................................................................................ 15
§ 11. Исследование спектральных свойств растений способом светофильтров ............................................................... 16
§ 12. Еще два оптических свойства марсианской растительности, отличающих ее от растительности земной ........ 17
§ 13. Связь астроботаники с другими науками ......................................................................................................................... 20
Заключение ........................................................................................................................................................................................ 21
Технич. редактор Э. Д. Рогоза
Корректор П. Ф. АЛФЕРОВА
Подписано к печати 26.IX.49. Объем 2,5 п. л. Уч.-авт. л. 1.75.
УГ05050. Тираж 5000.
Тип. АН КазССР. Зак. 88

Фиг. 1. Кварцевый спектрограф.

Фиг. 2. Ворота в Туюк-Су.

Фиг. 3. Яблони и груши. Обыкновенный снимок.

Фиг. 4. Яблони и груши. Инфракрасный снимок.

Фиг. 5. Ели тяньшанские. Обыкновенный снимок.

Фиг. 6. Ели тяньшанские. Инфракрасный снимок.

Фиг. 7. Флуоресцентный ящик во время съемки.
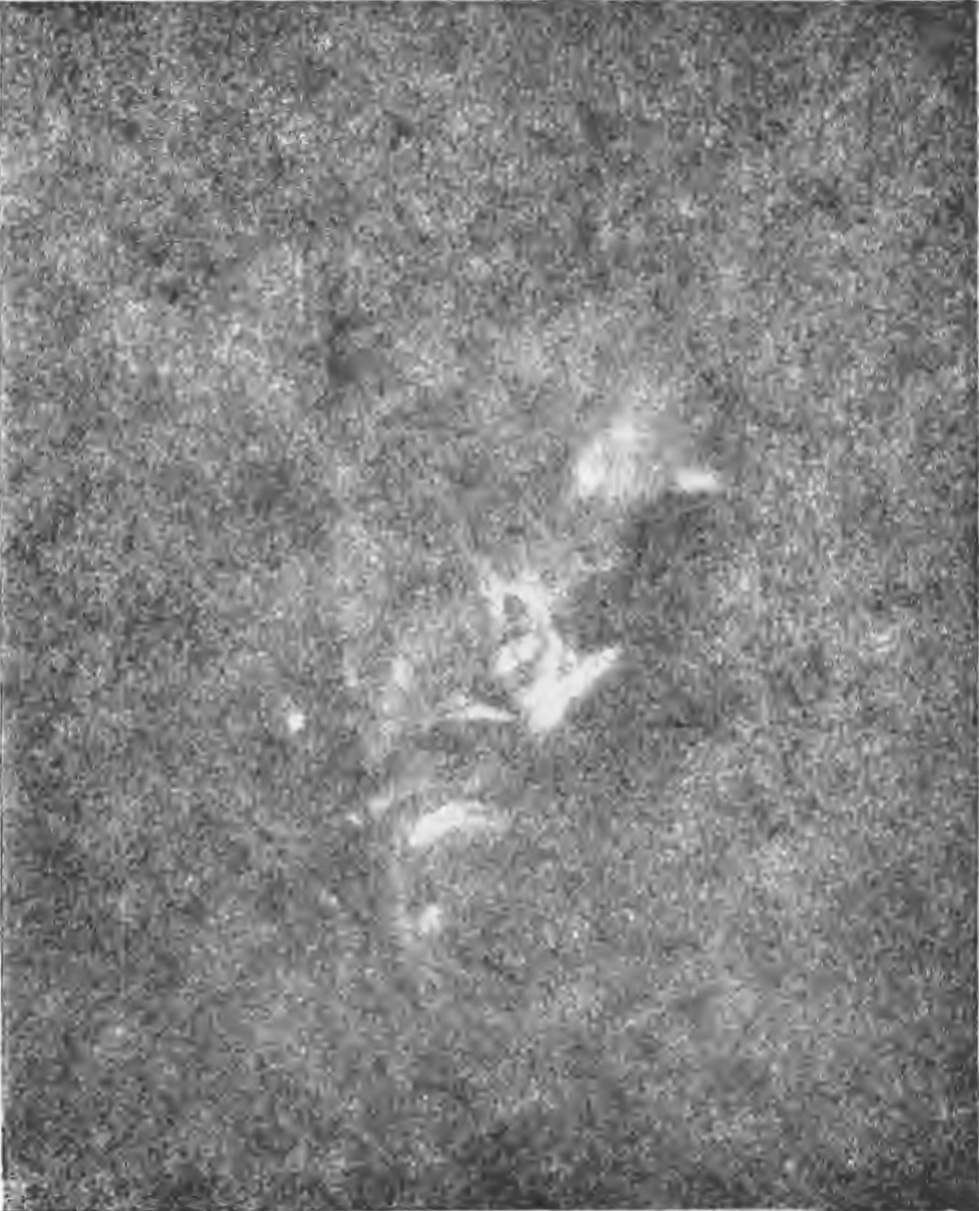
Фиг. 8. Колокольчик скученный.

Фиг. 9. Мелколепестник золотистый.

Фиг. 10. Девясил корнеголовый.

Фиг. 11. Жимолость при купоросном освещении.

Фиг. 12. Жимолость при красно-инфракрасном освещении.

Фиг. 13. «Борец» при купоросном освещении.

Фиг. 14. «Борец» при красно-инфракрасном освещении.

Фиг. 15. «Принцесса» в красных лучах.

Фиг. 16. «Принцесса» в ультрафиолетовых лучах.

Фиг. 17. Спектр сосны.

Фиг. 18. Спектр канадской ели.

Фиг. 19. График спектра канадской ели и сосны.
| Главная | Библиотека сайта | Форум | Гостевая книга |