Гербарий будущего


Гербарий будущего |
|
| В этом разделе собраны описания отдельных видов животных и растений, которые могли бы обитать на Земле в эпоху неоцена. Раздел будет обновляться по мере возникновения новых идей относительно возможных путей эволюции жизни. Если читатель не обнаружит здесь каких-то видов, про которые здесь было рассказано раньше, пусть не огорчается, а просто ищет новую главу "Путешествия в неоцен". | |
растения |
Цветковые
растения - двудольные
|
Лютик
разрезнолистный (Ranunculus lacinifolius)
Порядок: Лютикоцветные (Ranunculales)
Семейство: Лютиковые (Ranunculaceae)
Место обитания: горные ручьи и реки на востоке Северной Америки.
Будучи более старыми горами, чем Скалистые горы на западе, Аппалачи имеют относительно
неоднородную геологическую структуру – они сильнее разрушены временем, а в северной
части – ледниковым щитом голоцен-неоценового оледенения. В этих горах протекает
большее количество мелководных ручьёв и рек с более тихим и менее разрушительным
течением. Иногда, если местность имеет небольшой уклон, реки могут разливаться,
образуя заболоченные участки с медленным течением. В таких местах обильно разрастаются
влаголюбивые растения, задерживающие корнями ил и песок, и со временем образуются
питательные почвы. В таких местах складываются особые растительные сообщества,
и одно из типичных растений этих мест – лютик разрезнолистный.
Лютик разрезнолистный – это типичный представитель своего рода, травянистое
растение с ползучим стеблем, легко укореняющимся в узлах, и прямостоячими цветоносами
высотой около 15-20 см. Листья овальных очертаний, с сильно рассечённой листовой
пластинкой светло-зелёного цвета, покрыты множеством жёстких коричневатых волосков
– это способ защиты от гусениц, улиток и других травоядных беспозвоночных. Лист
разделён на 5-7 долей почти до места прикрепления черешка к листовой пластинке,
и каждая доля сильно рассечена менее глубокими вырезами. Черешки листьев поднимаются
вертикально, отчего лист напоминает миниатюрную пальму. Ползучие стебли и цветоносы
тоже покрыты волосками. Образовавшиеся под водой побеги имеют более редкие волоски.
Для защиты от травоядных позвоночных этот лютик вырабатывает разные вещества,
которые придают его зелени острый запах и делают его ядовитым, но местные виды
копытных поедают его без особого вреда для себя, правда в очень небольших количествах.
При помощи этого растения звери избавляются от значительной части кишечных паразитов.
Лютик разрезнолистный – обильно цветущее растение, но его цветы не очень яркие,
зеленовато-жёлтые, более тусклые чем у других видов местных цветов. Зато они
сильно и приятно пахнут, и многие насекомые отыскивают эти цветы, привлечённые
запахом лютика, а не окраской его цветов. Рядом с этим лютиком часто растут
другие виды растений, в том числе и насекомоядные, у которых тоже жёлтые цветы,
поэтому растение сделало свои цветы безошибочно узнаваемыми, отдав предпочтение
другому способу привлечения насекомых. У цветков этого вида 7-12 лепестков,
у отдельных экземпляров ещё больше. Лепестки округлой формы. На одном цветоносе
образуется один цветок на верхушке и 2-3 в пазухах листьев на коротких боковых
цветоносах.
Размножается лютик разрезнолистный семенами, которые часто опадают в воду и
плывут по течению вдоль берегов. Семена также могут распространяться травоядными
животными – они выносят действие их пищеварительного сока. Молодое растение
цветёт на втором году жизни. Основной способ размножения – вегетативное. У этого
растения длинные и цепкие корни, способные закрепиться в почве прибрежных мелководий.
Растение даёт значительное количество ползучих побегов, расселяясь над и под
водой, и способно образовывать даже небольшие заросли. На зиму лютик разрезнолистный
отмирает, а весной даёт новые побеги – сперва листья, а потом и цветы. Продолжительность
жизни этого растения – до 10-15 лет.
Этот вид растений открыл Bhut, участник форума.
Тихоокеанская
дицентра (Dicentra pacifica)
Порядок: Лютикоцветные (Ranunculales)
Семейство: Маковые (Papaveraceae), подсемейство: Дымянковые (Fumarioideae)
Место обитания: тихоокеанские дождевые леса.
В эпоху неоцена к западу от Скалистых гор Северной Америки сложилась очень необычная
экосистема: очень влажные леса с умеренным климатом и выраженной сменой времён
года. Там регулярно идут дожди (зимой – довольно холодные, иногда со снегом),
и уровень влажности очень высок. Там складывается особая фауна: высшие позвоночные,
птицы и особенно звери, предоставлены главным образом крупными видами, тогда
как среди мелких позвоночных доминируют рептилии и особенно амфибии. Беспозвоночные
представлены тоже в необычной пропорции: чешуекрылых, двукрылых и перепончатокрылых
насекомых там мало, но очень велико разнообразие жуков и наземных моллюсков
– слизней и улиток. Такие беспозвоночные – плохие опылители, поэтому растения,
которые процветают на северо-западном побережье Северной Америки, широко пользуются
услугами иных опылителей. Тихоокеанская дицентра, одно из характерных травянистых
цветковых растений этих лесов, использует местных птиц, а точнее – колибри.
Колибри – достаточно характерные обитатели этих лесов, но они живут в таких
лесах лишь сезонно, с весны до конца лета. Местные виды колибри в обмен на относительно
безопасную от хищников жизнь платят значительным однообразием рациона: в этих
лесах мало видов кормовых растений, поэтому здесь могут жить лишь неспециализированные
виды этих птиц.
Тихоокеанская дицентра процветает во влажных лесах. Это довольно крупное корневищное
растение – до полуметра в высоту, а розетка листьев может достигать 60-70 см
в диаметре. Внешне эта дицентра напоминает папоротник – у неё такие же широкие
и изрезанные листья зелёного цвета с сизоватым оттенком, который выгодно контрастирует
с цветами этой дицентры.
У разных видов дицентр цветки разной окраски; у тихоокеанского вида они сильно
вытянутые, белые или бело-жёлтые, с красным пятном внутри. Цветы образуются
на высоких цветоносах – больше метра высотой, дугообразно изогнутых и покачивающихся
от слабого ветерка. Цветонос окрашен в кораллово-красный цвет, привлекающий
опылителей, и на нём имеется несколько листочков такого же красного цвета в
основании соцветия. Колибри нормально видят красный цвет, слетаются к растениям
издалека и лезут своим клювом в ярко окрашенный зев цветка. Предковый вид цвёл
в основном с весны до начала лета, но у тихоокеанской дицентры цветы можно найти
большую часть года, кроме зимних месяцев. Зимой растение впадает в состояние
относительного покоя, сохраняя лишь небольшое количество листьев.
Семена этого вида созревают в стручковидных коробочках, которые внешне напоминают
стручки недотроги и подобным же образом лопаются от любого прикосновения, разбрасывая
семена достаточно далеко, чтобы они смогли прорасти. Эта дицентра не образует
протяжённых густых зарослей.
Дицентра тихоокеанская является многолетним растением и вполне может прожить
10-20 лет. Корневище этого вида часто поедают личинки жуков и улитки.
Этот вид растений открыл Bhut, участник форума.
Барбарис
южнейший (Berberis notalissimus)
Порядок: Лютикоцветные (Ranunculales)
Семейство: Барбарисовые (Berberidaceae)
Место обитания: кустарниковые заросли Антарктиды.

Рисунок Лягушки
В эпоху неоцена Антарктида всё ещё остаётся очень холодным
континентом с довольно бедной флорой даже по сравнению с южной оконечностью
Южной Америки. Одной из причин этого является изоляция материка: очень мало
растений смогло расселиться за сотни километров через море, чтобы заселить эту
негостеприимную землю. В этом плане некоторым ягодоносным травам и кустарникам
повезло больше, чем другим: их семена были принесены птицами вместе с помётом,
который одновременно становился своеобразным удобрением, давшим возможность
семенам прорасти и закрепиться на новом месте. Но чаще всего семена попадали
на материк на лапах птиц, присохшие к раздавленной мякоти – пищеварение мелких
птиц проходило слишком быстро, и семена чаще оказывались в море. Но крупные
птицы часто приносили семена в пищеварительном тракте.
Одним из ягодных растений-переселенцев стал потомок барбариса самшитолистного
(Berberis buxifolia), процветавшего на Огненной Земле в эпоху человека. Этот
кустарник был относительно поздним переселенцем на материк – он обосновался
там лишь после относительного потепления климата. За многие тысячи лет из его
потомков сформировалось несколько подвидов нового вида – барбариса южнейшего.
Они немного различаются между собой размером, формой листа и длиной колючек,
но обладают одним общим свойством – повышенной морозоустойчивостью. Это низкорослое
растение способно перезимовать под снегом даже в пятидесятиградусный мороз.
Антарктический барбарис южнейший – это стелющийся кустарник с длиной ствола
от 1 до 1,5 метров, с коричневыми опушёнными побегами. Ствол одревесневает,
тёмно-серая кора толстая и рыхлая, служит стволу своеобразной «шубой». Под корой
на поверхности ствола формируется значительное количество спящих почек – в случае
повреждения растения они быстро трогаются в рост и растение восстанавливается.
Кроме того, нижние ветви часто укореняются и при гибели или повреждении материнского
растения продолжают расти как самостоятельные растения. Иногда на месте гибели
старого куста образуются кольцевидные заросли его клонов, образовавшихся из
ветвей.
Из-за сокращения вегетационного сезона в новом месте обитания этот вид – вечнозелёное
растение, сохраняющее листву до весны. Листья эллиптические, у некоторых подвидов
гладкие, у других с мелкими зубцами по краю, к основанию листья клиновидные,
на верхушке с остриём, цельнокрайние, длиной от 1 до 5 см у разных подвидов.
Ближе к побережью обитают относительно крупнолистные формы, а в горах и на удалении
от океана встречаются главным образом мелколистные сильноветвистые формы с плотной
кроной, не продуваемой ветрами. Колючки тройчатые, длиной от 0,5 до 2 см у разных
подвидов. Их наличие – жизненная необходимость: листвой этого вида любят лакомиться
местные травоядные птицы луговые
лебеди.
Цветение начинается при установлении стабильной тёплой погоды и продолжается
почти до конца лета. Цветки жёлто-оранжевые, у некоторых подвидов красноватые,
собранные по 1-2 цветка, на тонких цветоножках длиной до 2,5 см. Цветы опыляются
жуками, мухами, комарами, местными бабочками и прилетающими пчёлами и даже птицами
мышевидками, которые выклёвывают
из цветков насекомых.
Плоды созревают на протяжении всей второй половины лета. Они почти шаровидные,
ярко-красные, диаметром 5-9 мм. Ягоды барбариса – прекрасная пища для многочисленной
местной орнито- и энтомофауны. В особенно урожайные годы опавшие осенью на землю
ягоды сохраняются относительно съедобными всю зиму и помогают местным птицам
пережить зиму под толщей снега.
Молодое растение развивается относительно медленно и начинает плодоносить только
на пятом году жизни. Медленное развитие компенсируется значительным долголетием
растения – до 150 лет и более. Кольцевидные клоны способны жить до 500 лет.
Этот вид растений открыл Wovoka, участник форума.
Дуб
змеествольный (Quercus serpenticaulis)
Порядок: Буковые (Fagales)
Семейство: Буковые (Fagaceae)
Место обитания: Средиземноморская котловина, склоны Альп.
Средиземноморская котловина относится к весьма неблагоприятным для жизни растений
областям. После высыхания Средиземного моря климат Южной Европы стал очень сухим.
Лишь весной снега, тающие в Альпах, образуют реки, которые вскоре теряются в
песках и среди солончаков, оставшихся от Средиземного моря. В этих местах растения
приспособились переносить неблагоприятные условия среды различными способами.
Одни из них запасают воду в стволах и листьях, другие переживают сухой сезон
в виде клубней или луковиц, а третьи стали эфемерами и успевают вырасти и дать
семена в течение короткой весны. В этой местности деревья принимают самые причудливые
формы.
Одно из деревьев Средиземноморской котловины не поднимается выше метра от поверхности
земли. Но его ствол достигает длины 30 – 40 метров и даже больше. Характерные
плоды-жёлуди указывают на принадлежность этого растения к дубам, но его ствол
напоминает скорее какую-то лиану, лишённую опоры. Это дуб змеествольный, одно
из характерных растений Южной Европы.
Ствол этого дерева достигает толщины полуметра, но изредка встречаются экземпляры
со стволами почти метровой толщины. Такого размера достигают растения, которые
произрастают в местах с достаточным увлажнением – вблизи русел пересыхающих
рек или в местах, где грунтовые воды стоят высоко. Ствол покрыт рыхлой тёмно-серой
корой, изборождённой множеством мелких трещин. Древесина этого вида очень твёрдая,
с большим количеством мелких годичных колец: растение со стволом полуметровой
толщины может иметь возраст около 300 лет. У дуба змеествольного ствол и ветви
очень легко укореняются. Основной ствол растёт, слегка приподнимаясь, но с возрастом
полегает и укореняется нижней стороной. В молодости ствол этого растения гибкий
– он огибает камни и разрастается, словно заполняя пустоты между ними. Контуры
растения напоминают типичную крону дерева, но расплющенную и разрастающуюся
в одной плоскости. Часто две или три крупных ветки у основания ствола достигают
примерно таких же размеров, как основной ствол. Укореняясь и удерживая камни,
дуб змеествольный укрепляет склоны гор. «Любовь» растения к камням имеет сугубо
практическую направленность – камни являются конденсаторами влаги из воздуха,
и растение получает достаточно влаги для роста, просто запустив корни прямо
под камень.
Листья дуба змеествольного не слишком отвечают стереотипному образу дуба. Они
узкие и длинные (длина около 10 см при ширине до 7 мм), с мелковолнистыми краями.
Такая форма листа помогает равномерно распределить свет и тень как по поверхности
самого листа, так и в глубине кроны. Почки и молодые листья покрыты густым белым
опушением; полностью сформировавшийся лист имеет кожистую тёмно-зелёную поверхность,
но сохраняет белое опушение на нижней стороне. В условиях достаточного увлажнения
растение сохраняет листья в течение лета, но экземпляры из засушливых местообитаний
могут сбрасывать листья во второй половине лета и остаются безлистными до весны.
Дерево зацветает ранней весной, с появлением первых листьев; пыльцу этого растения
переносит ветер. Мелкие жёлуди дуба змеествольного (диаметр около 6 мм), наполовину
скрытые в деревянистой плюске, созревают в начале лета. Их разносят птицы и
грызуны. Эти животные часто прячут жёлуди про запас, но не всегда находят их
потом. Из таких спрятанных желудей вырастают новые экземпляры растения.
Восточный
горный дуб (Quercus appalchiensis)
Порядок: Букоцветные (Fagales)
Семейство: Буковые (Fagaceae)
Место обитания: восток Северной Америки, склоны Аппалачей.

Рисунок Биолога
Ещё до эпохи неоцена разные виды дубов сумели приспособиться
к антропогенному прессингу и относительно успешно существовать в условиях изменения
природы человеком. Начало неоцена, ознаменовавшееся исчезновением человека,
нанесло определённый урон дубам, отбросив эти теплолюбивые деревья на юг из-за
наступления ледникового периода. Затем ледник отступил, и теплолюбивые растения,
в том числе деревья, вернулись на север и породили новые виды, приспособленные
к несколько изменившимся условиям жизни. Таков, к примеру, восточный горный
дуб, потомок дуба шарлахового (Quercus coccinea), одного из видов североамериканских
дубов эпохи голоцена.
Как и многие дубы, это довольно большое дерево, достигающее 10-20 м в высоту,
и образующее густые рощи на всём протяжении Аппалачей, но на севере ареала крупные
заросли этого вида встречаются реже. Листья этого вида имеют типичный «дубовый»
облик, но с глубокими вырезами полукруглой формы, с семью (реже – с пятью или
девятью) долями, слегка расширенными на концах. Листовая пластинка тёмно-зелёного
цвета, который осенью сменяется на столь же насыщенный красный, как у предкового
вида. Кора относительно ровная, мелкобороздчатая, отслаивается небольшими пластинками.
Дерево чаще всего растёт одним стволом, его толщина у основания достигает 150-180
см. Корневая система глубокая и мощная, позволяет дереву закрепляться даже в
трещинах скал. Растение предпочитает хорошо дренированный субстрат и бедную
каменистую почву.
Мелкие ветроопыляемые цветки типичного для дубов облика, жёлуди созревают в
течение двух лет, покрыты небольшой бугристой плюской. Спелые жёлуди держатся
на ветках относительно слабо; иногда их срывает ветер, и они скатываются вниз
по склонам. Часто они попадают в подходящие для прорастания и укоренения места
– в глубокие трещины среди камней. Сами жёлуди округлой формы, слегка вытянутые,
с желтовато-бурой оболочкой. Эти жёлуди являются одним из важных кормовых ресурсов
для грызунов и некоторых птиц.
Жёлуди восточного горного дуба довольно горькие на вкус, и большинство животных
их не ест, но один из видов соек, попугайная
сойка (Cyanocitta pecularia), питается главным образом желудями и орехами,
и восточный горный дуб является одним из её основных кормовых деревьев. В рационе
попугайной сойки доля желудей этого вида дубов пропорционально больше, чем у
многих других животных. Она гораздо чаще закапывает эти жёлуди про запас на
зиму, или натаскивает в щели между камнями, и часто забывает о них к весне.
И если эти запасы не будут обнаружены зимой другими животными, то они прорастут
через год, и если не погибнут в первые 10 зим, то смогут прожить долгие годы.
Средняя продолжительность жизни у восточного горного дуба – до 200-300 лет.
Этот вид растений открыл Bhut, участник форума.
Дуб
прорастающий (Quercus proliferans)
Порядок: Букоцветные (Fagales)
Семейство: Буковые (Fagaceae)
Место обитания: Евразия, умеренные районы Европы.
Дубы – характерные древесные растения Северного полушария, распространённые
главным образом в субтропических и тропических широтах: в умеренных широтах
их видовое разнообразие ниже. Однако в умеренных широтах Евразии и Северной
Америки обитают несколько видов, являющихся основными лесообразующими видами.
В европейских лесах умеренного климата фоновым видом дуба является дуб прорастающий
– вид, активно размножающийся вегетативно, который образует обширные заросли.
Подобно большинству дубов, данный вид является многолетним растением, крупным
деревом. Высота растения в благоприятных условиях достигает 25-30 метров. Кора
очень толстая (до 10 см), бежевато-серого цвета, опробковевшая, покрыта немногочисленными
глубокими трещинами. Дерево чаще всего растёт одним стволом; крона конических
очертаний с притупленной вершиной. При повреждении вершины появляются деревья,
состоящие из нескольких стволов. На стволе и корнях закладываются многочисленные
спящие почки, поэтому при гибели или повреждении основного ствола данный вид
даёт обильную корневую поросль (отсюда название). Ветви и стволы клонов в местах
трения часто срастаются друг с другом, образуя причудливые наплывы древесины.
Часто одно дерево со временем образует обширный клон, и лесной массив может
состоять всего из нескольких таких клонов, взаимно опыляющих друг друга. Самоопыление
приводит к появлению меньшего количества семян.
Листья дуба прорастающего одиночные, простые, кожистые, длиной около 20-25 см,
у северных популяций меньше. Очертания листа лодочковидные, края неглубоко вырезанные,
вершина притупленная. Окраска взрослого листа тёмно-зелёная, молодого и растущего
– розоватая, переходящая в оливковую. У отдельных экземпляров в южной части
ареала ювенильная розоватая окраска сохраняется у взрослого листа, что защищает
растение от солнечных ожогов. В южной части ареала растение вечнозелёное, в
умеренном климате листопадное. Осенняя окраска листьев ярко-оранжевая.
Жёлуди короткие и широкие, с плотной оболочкой, сохраняют всхожесть несколько
лет. Плюска толстая, деревянистая. Жёлуди обычно держатся на ветках по 2-3,
реже одиночные. В урожайные годы жёлуди этого дуба являются важным кормовым
ресурсом для европейской фауны травоядных млекопитающих и ряда видов птиц. Основными
распространителями вида являются птицы и грызуны.
Благодаря исключительно высокой способности к вегетативному размножению дуб
прорастающий заходит на север дальше остальных видов рода, но в условиях холодных
зим часто обмерзает до уровня снегового покрова. На северной оконечности ареала
жёлуди вызревают редко, но вид активно размножается корневой порослью, превращаясь
в кустарник высотой около 100-120 см. Кустовидная форма дуба прорастающего вечнозелёная,
старая листва опадает в начале лета, с развитием молодых побегов.
Продолжительность жизни – до 900-1000 лет. Первое плодоношение начинается в
возрасте 30-40 лет.
Степная
берёза (Betula endurabilis)
Порядок: Букоцветные (Fagales)
Семейство: Берёзовые (Betulaceae)
Место обитания: прерии Северной Америки.
На протяжении голоцена берёзы смогли пережить массовое вымирание, ставшее результатом
деятельности человека. Они выдержали климатические перемены раннего неоцена,
и среди них появились новые виды, многие из которых, однако, достаточно консервативны,
особенно внетропические виды, населяющие области умеренного климата в Евразии
и Северной Америке. На распространение берёз оказали значительное влияние ледники,
сдвинувшие их ареалы к югу. При отступлении ледников часть видов также отступила
на север, в более привычные условия, но некоторые виды остались в южных широтах
и приспособились к изменениям климата.
Одно из таких деревьев – североамериканская степная берёза. Её внешность типична
для берёз Северного полушария: кора очень светлого оттенка (белая или светло-серая)
с чёрными пятнами. Благодаря этому одни участки коры нагреваются под солнцем
гораздо быстрее, чем другие, и степная берёза одновременно получает необходимое
ей количество солнечного тепла, и не перегревается. Это очень низкорослое дерево
открытых пространств – высота взрослого дерева не превышает 2-3 метра. В типичном
случае молодое дерево быстро растёт одним стволом, после чего рост в высоту
тормозится, и начинается активное отрастание ветвей. На корнях формируется значительное
количество почек, которые трогаются в рост при повреждении ствола, и быстро
формируют заросль-клон диаметром до 10-15 метров.
Листья степной берёзы очень узнаваемые, овальной или яйцевидной формы с зазубренным
краем, на длинных тонких черешках. Для защиты от иссушения нижняя сторона листа
покрыта белым опушением, а на верхней стороне листа растут многочисленные волоски.
Осенью листья приобретают ярко-жёлтый цвет и опадают. В некоторых случаях часть
листвы опадает в особенно жаркое и засушливое лето, и отчасти восстанавливается
с осенними дождями.
Это ветроопыляемое растение, цветущее рано весной, и даже в конце зимы. Её соцветия
– длинные висячие серёжки. Из-за особенностей опыления, как и многие другие
деревья американских степей и прерий, эта берёза нередко образует одновидовые
рощи из нескольких крупных старых клонов собственного вида.
Семена этого вида тоже разносятся ветром. Они небольшие и мало для кого съедобны,
но некоторые степные виды певчих птиц их поедают, особенно перед зимой, когда
другого корма в североамериканских прериях маловато. Но гораздо большее число
птиц ищет в зарослях степной берёзы разных беспозвоночных, которые питаются
листвой, древесиной и другими частями этого дерева. Надземные части растения
поедают также более крупные травоядные – позвоночные. Степная берёза противостоит
им благодаря интенсивному росту, но пекари-бизоны
обладают достаточно сильными челюстями, чтобы объесть даже крупные ветви и обглодать
кору на стволе. В местах, где часто кормятся крупные травоядные, это дерево
встречается реже, чем другие виды деревьев североамериканских прерий.
Отдельное растение степной берёзы живёт не более 12-15 лет, но клон успешно
существует в среднем 40-70 лет и больше, особенно в условиях объедания крупными
травоядными и последующего омоложения зарослей.
Этот вид растений открыл Bhut, участник форума.
Патагонская
ложная лещина (Nothavellana antipodorum)
Порядок: Букоцветные (Fagales)
Семейство: Нотофаговые (Nothofagaceae)
Место обитания: юг Патагонии, Огненная Земля, леса.
Нотофагусы – характерные растения древней антарктической флоры, распространённые
на материках и островах южного полушария. Один из центров их видового разнообразия
– крайний юг Южной Америки, область умеренного климата с выраженной сезонностью.
Разные виды нотофагусов занимают различные экологические ниши в умеренных лесах
юга. В процессе эволюции среди них появился своеобразный «близнец» лещины (Coryllus)
Северного полушария – патагонская ложная лещина.
Этот вид – низкорослое древовидное растение с крупными ореховидными семенами.
Высота растения в благоприятных условиях не превышает 3-4 метра, чаще всего
немногим выше 2 метров. Обычно это одноствольное растение, но при повреждении
верхушки легко ветвится и растёт несколькими стволами, сохраняя шаровидные общие
очертания кроны. Это теневыносливый вид, произрастающий в подлеске. Нижние ветви
часто опускаются к земле, укореняются и продолжают рост, образуя новые вертикальные
стволы из боковых почек. Упавший ствол также может укорениться – тогда боковые
ветви начинают расти вверх, образуя густые заросли в подлеске. Листья широкие,
гладкие, округлых очертаний, с притупленным кончиком и крупнозубчатым краем,
длиной до 15-20 см. Этот вид – листопадное растение, осенью листва желтеет.
Цветки очень мелкие, ветроопыляемые. Растение цветёт весной при появлении листьев.
Семена созревают к середине лета; размер семени – до 15 мм, оно покрыто прочной
деревянистой скорлупой. Это один из основных видов корма для местных птиц и
млекопитающих, которые собирают значительные запасы семян этого вида и являются
их основными распространителями. Семена долго сохраняют всхожесть – до 3 лет.
Молодые растения начинают плодоносить в возрасте 5 лет, продолжительность жизни
достигает 200 лет, у клонов благодаря омолаживанию за счёт поросли возраст может
достигать 400 лет.
Настоящее
капустное дерево (Brassixylon crassus)
Порядок: Каперсовые (Capparales)
Семейство: Капустные (Brassicaceae)
Места обитания: сухие местности Юго-Западной Азии, Северной Африки и Южной Европы.

Рисунок Биолога
Много видов растений эпохи голоцена получило название «капустных
деревьев». Но это название давалось им исключительно за внешнее сходство некоторых
частей растения с домашней капустой, разводимой человеком. Зато дерево, появившееся
в неоцене в сухих районах Юго-Западной Азии (в предгорьях к востоку от Средиземноморской
низменности), и позже расселившееся по берегам Средиземноморских болот, носит
своё название более заслуженно: это один из ближайших родственников капусты,
относящийся к одному с ней ботаническому семейству.
Настоящее капустное дерево не составит конкуренции деревьям-великанам тропического
леса: его максимальная высота не превышает 13 – 15 метров. Но зато оно может
выживать там, где дуют сухие и жаркие ветра, а на почве оседает тонкий налёт
солей с солёных Средиземноморских болот.
В связи с очень сухим климатом в местах, где живёт это дерево, у него развился
ряд приспособлений, позволяющих выжить в экстремальных условиях. Настоящее капустное
дерево – стеблевой суккулент: для хранения запаса воды ему служит очень толстый
ствол, похожий на бутылку. Он сильно расширен в нижней трети, и иногда полегает:
кажется, словно дерево не выдержало тяжести своего «пивного живота». Древесина
этого растения непрочная и рыхлая, обильно насыщенная водой. Но пить её мало
кто отважится: дерево надёжно защищает себя с помощью горьких веществ, растворённых
в соке. Листья этого дерева также горькие на вкус. Они сочные и довольно широкие:
дерево активно фотосинтезирует большую часть года, впадая в стагнацию («оцепенение»)
только в самые жаркие летние месяцы. Но и тогда большая часть листьев сохраняется,
опадают лишь самые старые. Молодые листья образуют на кончиках ветвей дерева
своеобразные розетки. Черешки листьев могут совершать движения, и в зависимости
от окружающих условий дерево либо складывает листья в подобие кочанов, либо
раскрывает их. Обычно листья сложены вместе ночью и в самое жаркое время дня,
раскрываясь утром и вечером. Но в пасмурные дни розетки листьев могут быть открыты
целый день. Кроме того, для уменьшения потерь влаги листья покрыты тонкими восковым
налётом.
Настоящее капустное дерево добывает воду из глубинных слоёв почвы, поэтому оно
процветает там, где подземные воды близко подходят к поверхности. В редких оазисах
Северной Африки это дерево образует своеобразные разреженные леса совместно
с пальмами и некоторыми другими растениями.
Это растение, в отличие от капусты, многолетнее: предельный возраст его может
превышать 200 лет. И цветёт оно не раз в жизни, а ежегодно примерно с 5-летнего
возраста. Крупные розовые цветки с четырьмя лепестками в больших щитковидных
соцветиях раскрываются на самых верхних ветвях в середине короткого сезона дождей,
привлекая бабочек и
мух. Каждый цветок цветёт недолго – всего лишь два дня и одну ночь. Поэтому
многие цветки этого растения остаются не опылёнными. Плоды настоящего капустного
дерева – длинные, скрученные, словно сверло, стручки. При прикосновении (например,
если на соцветие села птица) спелый плод «взрывается»: створки моментально скручиваются
и далеко разбрасывают семена. Оболочка семян голая, но зато очень прочная и
яркая: красная с белыми крапинками. Птицы, привлечённые такой окраской, склёвывают
эти семена. Но в их желудке оболочка не переваривается, а лишь немного истончается
и становится более проницаемой. Выпав из кишечника птицы с порцией помёта, семена
имеют хорошие шансы на прорастание.
Первый год молодое растение имеет подземный клубнеобразный стебель («корнеплод»),
образованный подсемядольным коленом проростка, и розетку небольших листьев на
поверхности почвы. На второй год начинается рост ствола, хотя подземный толстый
стебель не исчезает ещё долгое время. Если ствол будет повреждён (чаще всего
попросту съеден), у растения есть шанс выжить за счёт подземной части.
Древопунция
ложнооблиственная (Equadopuntia pseudophyllata)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: Галапагосские острова, прибрежные районы.

Рисунок Лягушки, обработка Биолога
Галапагосские острова в эпоху человека отличались засушливым
климатом: на них не было постоянных водоёмов, и пресная вода встречалась только
в дождевых лужах. Поэтому на островах преобладала растительность трёх типов:
ксерофиты (выносящие значительную сухость почвы), суккуленты (запасающие воду
в стволах) и мангровая растительность (способная жить в морской воде).
Суккулентная растительность на Галапагосских островах была представлена несколькими
видами кактусов, среди которых наиболее типичными были опунции (Opuntia). Эти
кактусы с широкими лепёшковидными стеблями достигли на Галапагосах размеров
дерева. Единственными животными, которые поедали эти кактусы, были гигантские
черепахи. Завоз на острова коз пошёл даже на пользу кактусам: численность черепах
сократилась, а козы, выедавшие другие растения, лишали кактусы естественных
конкурентов. Поэтому опунции остались доминирующими растениями на Галапагосах.
Неоценовый потомок галапагосских опунций остался древовидным растением. Но изменения
климата заставили его несколько изменить облик, хотя основные черты кактусов
всё же сохранились. Галапагосская древопунция ложнооблиственная вырастает до
20 метров в высоту, а ствол старого дерева бывает до 6 метров толщиной. У этого
вида чётко проявляется дифференцировка побегов на два резко отличающихся типа.
Ствол и скелетные ветви этого дерева покрыты толстой, рыхлой, легко крошащейся
корой. Такая кора надёжно предохраняет ствол растения от перегрева, тем самым
сокращая потери влаги. Древесина древопунции рыхлая, водянистая, и относительно
непрочная. Во время ураганов некоторые деревья падают, но продолжают расти,
укореняясь в земле. Также легко укореняются отдельные ветви древопунции.
Побеги возрастом до трёх лет сохраняют на поверхности колючки, собранные в пучки
по несколько штук. На молодых скелетных ветвях вырастают побеги второго типа:
овальные однолетние «листья», зелёные и мягкие, почти без колючек. Только по
краям такого «листа» торчат короткие колючки. Многие «листья» этого растения
живут всего один год, опадая в сухой сезон. Лишь самые молодые «листья» могут
пережить его, и сохраниться два сезона подряд.
Древопунция цветёт почти круглый год, лишь в летнюю засуху может приостановить
цветение, одновременно сбрасывая «листья». Цветы этого растения ярко-красные,
с длинными лепестками, диаметром до 10 см. Они выделяют много нектара, но лишены
запаха: это растение опыляется птицами. У древопунции сладкие и сочные грушевидные
плоды, покрытые колючками. Местные птицы расклёвывают их, поедая мякоть и распространяя
семена, а упавшие плоды едят крупные местные черепахи.
Древопунция растёт относительно быстро, давая за год метровый прирост. Но это
дерево живёт недолго: не более 200 лет.
Идею о существовании этого растения высказал Семён, участник форума.
Коралловая
опунция (Rhodoopuntia corallina)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: южные склоны Альп.

Рисунок Биолога
Миллионы лет прошли после исчезновения человечества, но следы
человеческой деятельности в области интродукции разных видов животных и растений
сохранятся ещё долгое время в виде потомков этих видов, которые обитают там,
куда не могли бы попасть естественным путём. Средиземноморская котловина окружена
Альпами на севере и пустынями с юга, востока и отчасти с запада. Земли, которые
не отравлены солью высохшего Средиземного моря, отличаются сухостью, и лишь
немногие растения могут успешно расти на них. Среди тех, кому удалось прижиться
в этих местах, были кактусы, потомки завезённых человеком опунций (Opuntia spp.).
Некоторые виды этих растений приобрели причудливый облик под воздействием суровых
условий среды обитания.
Растения семейства кактусовых вырабатывают беталаины – красные пигменты, близкие
бетанину свеклы. Они придают цветкам кактусов характерный красный цвет. Эти
пигменты очень хорошо поглощают ультрафиолетовое излучение и защищают растение
от жгучих лучей солнца. В неоцене в Средиземноморской котловине сформировался
необычно выглядящий вид опунции, у которого способность к синтезу этих пигментов
резко усилена. Поэтому её стебли окрашены в свекольно-красный цвет (хлорофилл
присутствует, но замаскирован красным пигментом) и похожи на заросли благородного
красного коралла, словно он сохранился на своём месте с момента высыхания моря.
Это растение так и называется – коралловая опунция.
Из-за очень большого количества пигментов, синтезируемых растением, лепестки
его цветков окрашены в тёмно-пурпурный, почти черный цвет, как цветки чёрного
тюльпана, выведенного когда-то человеком. Изредка в популяции встречаются носители
редких аллелей генов с лимонно-жёлтой, чёрной и зелёной окраской стебля.
Коралловая опунция произрастает не в самой болотистой котловине, а на примыкающих
к ней южных склонах Альп. Краснолистность характерна для горных растений, поскольку
в горах солнечный ультрафиолет особенно опасен. На сравнительно малых высотах
(до 3000 – 3500 м над уровнем моря) эта опунция может принимать древовидную
форму и разрастается до размеров канделябрового кактуса (Carnegia gigantea)
эпохи человека, но ветвится значительно сильнее, образуя крону из плоских побегов.
На таких высотах в популяции встречаются особи с нетипичной окраской стеблей,
поскольку здесь интенсивность ультрафиолетового излучения не столь велика, и
мутанты не получают ожогов. Выше коралловая опунция сильно уменьшается в размерах,
теряет основной ствол и на больших высотах, немногим ниже снеговой линии, образует
стланик с ползучими многолетними побегами, от которых поднимаются вверх побеги
текущего и прошлого годов. Высоко в горах красная окраска усиливается. Стебли-лепёшки
этого вида похожи на побеги современных опунций. Они сохраняют острые иголки,
растущие пучками по 5 – 8 штук, а молодые побеги обладают недоразвитыми листьями,
похожими на рожки, и довольно густым опушением из тонких волосков, защищающих
растущие ткани от солнечного ожога и заморозков. Вызревшие побеги становятся
более выносливыми, и на них образуются цветки и плоды.
Корневая система приспособлена для закрепления на скалистых уступах. Стланиковая
форма в высокогорье легко укореняется нижней частью многолетних побегов, и играет
важную роль в предотвращении обвалов.
Этот вид растений не предъявляет строгих требований к опылителям; пыльцу этих
кактусов могут переносить различные животные – от жуков и бабочек до мелких
птиц и грызунов. Цветочные почки закладываются по краю прошлогодних побегов
зимой. Весной и в начале лета раскрываются необычные чёрные цветки этого растения.
Они сильно нагреваются на солнце, и привлекают мух и жуков, питающихся падалью.
А насекомые, собирающиеся на цветках, в свою очередь привлекают насекомоядных
птиц. Плоды коралловой опунции имеют очень тёмную окраску и покрыты сизоватым
слоем воска. Этот покров отражает ультрафиолетовые лучи, делая плоды более заметными
для птиц. Также плодами этого кактуса питаются грызуны и другие млекопитающие.
Упавшие с древовидных кактусов этого вида плоды охотно поедают хищники, которых
привлекает влага, содержащаяся в них. Мякоть плодов сочная, тёмно-красная, очень
сладкая; мелкие семена рассеяны по всей мякоти. В сухие годы мякоть плодов может
не достигать полной спелости, но семена нормально развиваются и могут даже прорастать
прямо внутри плода. Эта особенность даёт дополнительную гарантию выживания проростков.
В типичном случае семена распространяются птицами и млекопитающими.
Этот вид растений открыл Антон, участник форума.
Компасная опунция (Platyopuntia compassa)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: Средиземноморская котловина, Северная Африка, пустынные районы.
Экстремальные условия существования вызывают появление у обитающих в таких
условиях живых организмов разнообразных приспособлений для выживания, которые
зачастую придают их обладателям причудливый облик.
В засушливых районах на севере Африки и по окраинам Средиземноморской котловины
сложился комплекс растительности, включающий разнообразные виды, устойчивые
к экстремальным условиям существования. Одни из них растут и развиваются в
более благоприятное время года, пока солнце греет не так сильно, и переживают
летнюю засуху в виде клубней или луковиц. Другие успевают прожить всю жизнь
в течение нескольких недель и переживают трудные времена в виде семян. А третьи
никуда не прячутся и успешно противостоят иссушающей жаре.
Человек завёз в Средиземноморье американские кактусы опунции. Они успешно развивались
в этих местах и без человека, поэтому после вымирания человека они сохранились
и их потомки стали распространяться по пустынным районам Средиземноморской
котловины. Один из неоценовых видов опунций в Старом Свете – компасная опунция.
Это растение получило название за характерную особенность роста: его плоские
побеги всегда ориентированы ребром вдоль линии север – юг, поэтому жаркие лучи
полуденного солнца скользят вдоль их поверхности и не вызывают ожогов.
Компасная опунция сохраняет характерный облик своих предков: у неё плоские
стебли, состоящие из лепёшковидных сегментов. Сегменты имеют округлую форму,
жёсткие, хрящеватые, сильно сплющенные и волокнистые. Очертания растения вееровидные:
все побеги расположены практически в одной плоскости и не перекрывают друг
друга сбоку. У этого растения сохраняются несколько многолетних стеблей – его
«ствол» и «ветви», а значительная часть побегов не живёт более трёх лет. По
мере роста побеги меняют облик и функции. Однолетний побег покрыт тонким слоем
воска, предохраняющим нежные растущие ткани от солнечного ожога и придающим
побегу сизоватый цвет. На второй год побег вызревает и теряет восковой покров.
Его поверхность приобретает светло-зелёный цвет, и побег начинает активно фотосинтезировать.
Иголки компасной опунции очень крупные и редко расположенные, но деревянистые
и прочные. Травоядные животные встречаются в пустынях редко, но защита от них
всё равно необходима. Высота растения не превышает одного метра.
Некоторые побеги второго года образуют исключительно вегетативные почки, но
на большинстве закладываются цветочные почки. На третьем году жизни побега
в начале весны раскрываются цветки. Они белые, мелкие, с множеством притупленных
лепестков, расположены по краю сегмента побега группами по 4 – 7 штук. Лепестки
этих цветков покрыты слоем воска, благодаря чему они хорошо отражают солнечный
свет и заметны для редких насекомых-опылителей.
После опыления на растении вызревают мелкие ягодообразные плоды красно-розового
цвета, съедобные, с кисловатым вкусом и множеством мелких семян. При созревании
плодов сегмент, на котором они созревают, постепенно усыхает, отдавая в плоды
питательные вещества, накопленные в предыдущем году. Одновременно на таком
побеге опадают колючки. Высохшие сегменты стебля с плодами остаются на растении,
где их находят птицы, являющиеся главными распространителями семян компасной
опунции. После того, как все плоды оказываются съеденными, побеги постепенно
отсыхают и опадают. Так происходит естественное прореживание кроны растения.
Отломившиеся однолетние побеги, как правило, гибнут. Двухлетний побег, накопивший
достаточно питательных веществ, может жить, не укореняясь, до трёх лет в экстремально
засушливых условиях. Благодаря такой живучести он легко приживается в благоприятных
условиях – достаточно редкого дождя, чтобы он смог укорениться в земле. Постепенно
в течение трёх – пяти лет он образует полноценное растение и начинает цвести.
Баобабовая опунция (Adansopuntia obesa)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: саванны и полупустыни на севере Африки.
В эпоху человека в регион Средиземноморья были завезены кактусы рода опунция
(Opuntia). После исчезновения человека они пережили климатические изменения
и широко распространились в аридных районах Старого Света, дав начало различным
жизненным формам. Самым крупным из потомков опунции в Старом Свете является
баобабовая опунция. Это крупное дерево, внешним видом, размером и формой похожее
на баобаб, но при ближайшем рассмотрении оказывающееся кактусом. Этот вид сохранил
и усилил характерную для кактусов способность запасать в стволе воду, благодаря
чему может выдерживать длительную засуху и расти в условиях, когда другие древовидные
растения погибают.
Ствол в обхвате до 20 м, но высота дерева небольшая – тоже около 20 м. Древесина
этого вида рыхлая, с большим количеством водозапасающей паренхимы, внешний слой
укреплён сетью волокон. Нижняя часть ствола неветвистая, первые крупные ветви
начинаются на высоте около 10 метров. Многолетние ветви округлые в сечении,
прирост возрастом до трёх лет похож на типичные уплощенные стебли опунций. Молодые
ветви колючие; колючки длиной до 5 см, с зазубренными краями, собраны в группы
по 4 – 7 штук. Ствол молодых растений тоже колючий, с возрастом колючки исчезают.
Кора ствола взрослого растения гладкая, без иголок, но очень плотная и толстая
– до полуметра в основании ствола. Так растение защищается от редких пустынных
травоядных, желающих добраться в первую очередь до сочной древесины. Повреждения
коры это растение переносит очень легко, быстро восстанавливая утраченные участки
коры. Также толстый слой коры служит защитой от перегрева.
Цветки баобабовой опунции очень крупные – размером с тарелку, ярко-красные,
многолепестные. Они появляются на двухлетних побегах; почки закладываются в
осеннее-зимний период. Дерево в цвету представляет собой незабывемое зрелище.
Опыление производят насекомые и птицы, которых соблазняет водянистый нектар.
Плоды величиной с грейпфрут, с красно-фиолетовой мякотью, сладкие и сочные.
Ими питаются обезьяны, являющиеся одними из основных разносчиков семян. В засушливых
районах плодами питаются зайцелопы и птицы.
Проростки успешно развиваются только в редкие дождливые годы, когда успевают
запасти достаточно воды, чтобы пережить сухой сезон. Обычно критическими для
выживания являются первые два года жизни растения. Первый раз баобабовая опунция
зацветает в возрасте около 15 лет, достигнув 4-метровой высоты. Продолжительность
жизни более 1000 лет.
Идею о существовании этого вида растений высказал Антон, участник форума.
Утренний
диморфоцереус (Dimorphocereus matutinus)
Порядок: Гвоздикоцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: Большая Антигуа, Куба, Флорида, тропические леса.
Характерными растениями тропических лесов Нового Света были специализированные
лесные кактусы. В эпоху человека это были главным образом эпифиты и небольшие
полукустарники, в том числе лазающие. В неоцене кактусы в разных частях Земли
успешно вышли в крупноразмерный класс,
и среди лесных кактусов также появились крупные виды. Во влажных тропических
лесах островов Центральной Америки обитает один из таких видов – утренний диморфоцереус,
очень крупное лиановидное растение с одревесневающим стеблем. Начиная свою жизнь
в подлеске, он достигает вершин деревьев, а его побеги вытягиваются на 30-40
метров. Необычной особенностью облика этого растения являются побеги двух типов
– отсюда название рода. Многолетние одревесневающие скелетные побеги образуют
ствол и ветви растения. Они покрыты серой рыхлой корой и с возрастом утратили
функцию фотосинтеза, передав её побегам другого типа. Эти побеги плоские и плотно
прилегают к стволу дерева-опоры. Их сегментация почти не выражена, поскольку
они сильно разрослись по сравнению со своим первоначальным обликом. Эти побеги
обильно образуют воздушные корни, при помощи которых растение крепится к опоре
и получает дополнительную влагу. При разрыве и отмирании основного стебля растение
теряет связь с почвой, но может расти как гигантский эпифит, хотя скорость роста
при этом существенно замедляется.
Фотосинтезирующие побеги имеют совершенно иной облик. Они плоские, но по краям
есть лопасти, разделённые очень глубокими вырезами, из-за чего эти побеги напоминают
пальмовые листья. Они ярко-зелёного цвета, с гладкой поверхностью, длиной до
50-60 см при ширине до 30 см. Молодые фотосинтезирующие побеги красноватого
цвета. Такие побеги вырастают в большом количестве на молодых ветвях растения,
на уровне кроны дерева. При достаточном освещении они способны развиться из
спящих почек на стволе растения. Колючки на них сильно редуцированы. Отломившийся
фотосинтезирующий побег легко укореняется в земле и начинает расти, добираясь
до ближайшего дерева. С возрастом покровы этого побега покрываются корой, боковые
лопасти отмирают, и побег начинает расти в ширину и в толщину, превращаясь в
скелетный побег. Обычный срок жизни фотосинтезирующего побега – до 5 лет.
На фотосинтезирующих побегах на протяжении значительной части года образуются
цветки. Они ярко-красные с оранжеватым оттенком лепестков, диаметром до 20 см,
с длинной трубкой и многочисленными жёлтыми тычинками. Запах цветков сладковатый,
пряный. Они опыляются преимущественно колибри
и другими птицами, совершающими перелёты через острова Карибского моря между
Северной и Южной Америкой, а также бабочками, которые успевают полакомиться
нектаром, пока цветки ещё открыты.
Диморфоцереус утренний цветёт со второй половины ночи до утра; примерно через
три часа после рассвета цветки вянут. У растения две волны цветения – весенняя
и осенняя, совпадающие с миграциями опылителей этого вида. Зимой этот вид не
цветёт, летом образует небольшое количество цветков.
Плод этого вида – сочная многосемянная ягода. Это обычная пища древесных птиц,
но упавшими плодами лакомятся лесные четвероногие – мазамары
и другие травоядные млекопитающие, ящерицы и черепахи.
В тропических лесах этого региона произрастает близкий вид: вечерний
диморфоцереус (Dimorphocereus vespertinus), который зацветает вечером
и ночью. У него очень пахучие цветки, белоснежные лепестки и многочисленные
жёлтые тычинки. Это растение опыляется летучими мышами и бабочками-бражниками.
Благодаря особенностям опыления формируется надёжная репродуктивная изоляция
между видами: они могут произрастать на соседних деревьях, но не скрещиваться.
Гибридизация этих видов в природе крайне редка. Вечерний диморфоцереус предпочитает
расти как эпифит, его стебель быстро отмирает у основания, а скелетные побеги
прилегают к стволу и ветвям растения-опоры. Фотосинтезирующие побеги этого вида
длиной до 80 см, шириной около 20 см, имеют перистую форму.
Кротовый
кактус (Cryptocactus subterraneus)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: Мексиканская пустыня, засушливые районы с небольшим количеством
осадков.
Кактусы являются одними из наиболее приспособленных к засушливым условиям растений.
Почти все кактусы, кроме самых примитивных, являются стеблевыми суккулентами,
и их зелёные стебли берут на себя функцию и листьев, и запасающих воду органов.
Стебли кактусов бывают разнообразной формы – от небольших шаровидных до огромных
бочкообразных и даже древовидных.
Исключительное многообразие видов кактусов указывает на их успех в борьбе за
существование.
Как правило, стебель кактуса является многолетним, хотя в эпоху голоцена были
единичные исключения из этого правила. В неоцене среди кактусов появился вид,
который отошёл от общего правила: каждый его стебель фактически существует менее
двух лет. Этот кактус – очень своеобразное растение-«невидимка»: его можно увидеть
лишь в течение нескольких месяцев в году. Всё остальное время растение прячется
под землёй от жарких лучей солнца, и эта особенность послужила основанием для
названия растения – кротовый кактус.
Появление кротового кактуса на поверхности земли начинается с цветков. Незадолго
до сезона весенних дождей, когда зимняя прохлада постепенно уходит, на поверхности
земли раскрываются цветки – ярко-красные, заметные издалека, диаметром около
4 – 5 см. Обычно их бывает несколько. Насекомые замечают их благодаря особенности
строения лепестков – они сильно отражают ультрафиолетовый свет благодаря особой
текстуре лепестков. Этот вид опыляют разнообразные насекомые – даже муравьи
и жуки встречаются на его цветках. После опыления лепестки вянут, а под землёй
начинается созревание плода – небольшой округлой ягоды с многочисленными мелкими
семенами и сочной мякотью. Обычно на одном растении успешно развивается одна
или две ягоды, и только в исключительно благоприятных условиях их может быть
больше. Ягоды созревают к началу лета, и цветоножка, бурно разрастаясь, выносит
их на поверхность земли, где их поедают грызуны и птицы.
Когда цветок опылён или просто отцвёл, начинается активный рост стеблей этого
растения. Побеги кротового кактуса шарообразные, с многочисленными неглубокими
складками на поверхности. Окраска кожицы светло-зелёная с сизоватым восковым
налётом, на вершинах побегов и в ареолах имеется белое опушение. Иглы этого
растения крепкие, но немногочисленные. В каждой ареоле сидит одна длинная крючкообразная
иголка и несколько мелких прямых. С помощью крючкообразных игл побеги этого
кактуса зацепляются за шерсть и перья крупных обитателей пустыни. Они легко
отламываются от основания и таким образом расселяются на большие расстояния.
Побеги текущего года растут из прошлогоднего стебля, который находится под землёй
и постепенно отдаёт на рост новых побегов и созревание плодов все накопленные
питательные вещества. По мере роста побегов и созревания семян прошлогодний
побег постепенно сморщивается и отмирает, и к середине лета новые побеги уже
начинают расти независимо. Таким способом на протяжении ряда лет исходное растение
образует своего рода клон, разрастающийся в виде почти правильного кольца. Обычно
цветы и плоды появляются на внутреннем крае кольца, а новые побеги – на наружном.
Диаметр побегов кротового кактуса достигает 6 – 8 см.
В середине лета, когда прошлогодний побег отмирает окончательно, молодые растения
образуют по несколько длинных толстых корней, которые тянутся вниз и разветвляются
на небольшой глубине. Они являются своеобразными «якорями» для растения, но
их роль в его жизни намного важнее. Когда становится слишком жарко, корни начинают
подсыхать и скручиваться. При этом они образуют значительное усилие, которое
втягивает растение под землю. В течение одной – двух недель кактус скрывается
под землёй, и эти побеги больше не покажутся на свет. При этом колючки постепенно
ослабевают и отваливаются, ломаясь при втягивании побега под землю. В темноте
стебель постепенно теряет хлорофилл и становится похожим на клубень. В течение
зимы на нём закладываются почки, из которых на следующий год появятся цветы
и новые побеги.
Антигуария
трубкоцветная (Antiguaria tubiflora)
Порядок: Гвоздикоцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: горные районы Большой Антигуа.
В эпоху человека кактусы были типичными растениями тропических, субтропических
и умеренных областей Нового Света. Деятельность человека по-разному отразилась
на разных представителях этих растений. Одни из них значительно расширили ареал
и колонизировали тропические районы Старого Света. Другие виды стали крайне
редкими и в дальнейшем вымерли, не оставив потомков. В целом разнообразие неоценовых
кактусов сравнимо с таковым в эпоху человека. Они занимают различные экологические
ниши и встречаются в разных природных сообществах.
В горах острова Большая Антигуа растёт особый эндемичный кактус – антигуария
трубкоцветная. Избегая конкуренции с соснами, этот вид встречается на горных
склонах, на сухих каменистых почвах. Эти кактусы образуют обширные заросли,
легко узнаваемые издалека: каждый такой кактус выглядит серебристой колонной
шерсти. Тело растения покрыто слоем длинных прозрачных волосков, отражающих
часть солнечного излучения и проводящих к фотосинтезирующим тканям лишь небольшую
часть солнечного излучения. Молодое растение этого вида колонновидное и неветвистое.
С возрастом кактус ветвится от корня. Молодые побеги растут наклонно и полегают
на землю, укореняясь, после чего начинают расти вверх параллельно старому побегу.
У взрослых растений нижняя часть стволов одревесневает, теряет волоски и часть
колючек и покрывается рыхлой корой, предохраняющей от перегрева. Стебли ветвятся
редко, главным образом при повреждении точки роста. Стебель с многочисленными
рёбрами (до 20). Колючки желтоватые, длиной до 3-5 см, собраны пучками по 10-15
штук и почти не видны из-под покрова из волосков. Высота побегов взрослого растения
достигает 3-4 метров. Одно растение может образовывать до 20 вертикальных побегов.
Антигуария является птицеопыляемым кактусом. Её узкие красные цветки длиной
до 7-8 см развиваются на поверхности растения на прошлогоднем приросте. Цветки
растут группами по несколько десятков штук. Лепестки многочисленные, отгиб цветка
небольшой. Каждый цветок держится до 3 дней, в пасмурную погоду до 5 дней. Период
цветения делится на две волны – весеннюю и осеннюю, по времени совпадающие с
периодом миграции некоторых видов
колибри в Северную Америку через
Антильские острова и Кубу и обратно. После весенней волны цветения следует вегетационный
период, созревают плоды. После осеннего цветения и созревания плодов наступает
период покоя.
Этот вид кактусов размножается семенами и вегетативно. Первое цветение молодого
растения наступает в возрасте 3 лет. Продолжительность жизни – до 50 лет.
Кактус
«старая дева» (Megacephalocereus virgo-vetula)
Порядок: Гвоздикоцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: пустыни и полупустыни Мексики.
Кактусы эпохи человека демонстрировали интересное (хоть и не уникальное) приспособление
к жизни в засушливых регионах, а также стали излюбленными декоративными растениями:
некоторые виды имели весьма причудливую, необычную внешность. Это позволило
им значительно расширить ареал и сохранить значительное видовое разнообразие
к неоцену. Потомок одного из выживших видов к неоцену стал формировать своеобразные
растительные сообщества.
Кактус «старая дева» – потомок вида Cephalocereus senilis, живущий там же, где
и предок – в пустынях и полупустынях Мексики. Это очень крупное растение – высотой
до 10 м, с колонновидным стволом, толщина которого доходит до метра, и 10-15
крупными боковыми ветвями разной длины. Обликом он несколько напоминает гигантские
карнегии (Carnegia) эпохи человека. Рёбра на стволе и ветвях заметно выражены
в сухой сезон и частично разглаживаются при пополнении кактусом запаса воды.
Ареолы крупные, расположены правильными рядами, колючки отсутствуют. Зато из
каждой ареолы растут длинные, до 50-60 см, густо расположенные ниспадающие волоски,
что придает всему кактусу вид длинноволосой человеческой головы. Волоски бесцветные,
полупрозрачные, что обеспечивает доступ света к фотосинтезирующей зелёной ткани
под ними, при этом надёжно защищая поверхность растения от палящих солнечных
лучей и ветра.
Корневая система стержневая, главный корень часто утолщённый, водозапасающий.
Он достигает водоносных слоёв почвы на глубине до 10 метров. При цветении довольно
длинные цветоносные побеги вырастают сквозь «волосяной покров» на концах ствола
и ветвей, образуя подобие венка на седой голове (отсюда название вида). Цветки
крупные, до 20 см в диаметре, ярко-оранжевые или жёлтые. Лепестки и тычинки
многочисленные, пестик один. Цветки издают сладкий запах нектара и привлекают
опылителей – насекомых и мелких
птиц. Часть из них находит пристанище здесь же, среди покровов кактуса:
насекомые поселяются в «волосах» и откладывают там яйца, а птицы
живут в дуплах в стволе или крупных ветвях. Плоды – сочные ягоды лимонообразной
формы до 15 см длиной, кожура оранжевая, мякоть белая с чёрными семенами. Семена
распространяются животными, питающимися плодами этого кактуса.
Растение образует своеобразные редколесья в местах с неглубоким залеганием грунтовых
вод. Подлесок в лесах этих кактусов образуют другие суккулентные растения, в
том числе кактусы мелких видов. Они подавляют рост «старой девы» в молодом возрасте,
но примерно с 5-летнего возраста этот кактус обгоняет их в росте и начинает
нормально развиваться.
«Волосяной покров» кактуса «старой девы» даёт убежище многим сожителям: некоторые
насекомые живут среди «волос», словно вши, другие держатся под слоем волос на
поверхности ствола и ветвей Подобным образом поступают некоторые ящерицы. А
ряд видов птиц вьёт гнезда в «волосах» или селится в дуплах в стволе или основании
ветвей «старой девы». Под слоем опушения кактуса они укрываются от солнечного
зноя, взамен опыляя кактус и разнося его семена.
Этот вид растений: открыл Биолог, участник форума.
Зефирный
кактус (Echinocactus dulcedo)
Порядок: Гвоздикоцветные (Caryophyllales)
Семейство: Кактусовые (Cactaceae)
Место обитания: пустыни Мексики.
В эпоху человека эхинокактусы (Echinocactus) из мексиканских пустынь были примечательны
своей формой стебля – почти правильной шаровидной, с длинными острыми иглами
на многочисленных продольных рёбрах. Они часто выращивались человеком как декоративные
растения, что помогло им выжить в эпоху биологического кризиса. В неоцене их
потомки развили некоторые новые особенности.
Зефирный кактус получил своё название за внешний вид: его побеги имеют вид шаров
до 1 м и более в диаметре, с большим количеством рёбер – до 60-70. Вдоль рёбер
расположены многочисленные крупные ареолы с 6-7 длинными (до 20 см) колючками,
расположенными пучком, и густыми волосками около 10 см длиной. Волоски образуют
толстый термоизолирующий и светопроводящий покров, защищающий кактус от солнечного
зноя, но не препятствующий фотосинтезу. Его стебель тёмно-зелёный с красноватым
оттенком, колючки тёмно-красные, а волоски имеют нежно-розовую окраску. Розовые
волоски придают кактусу сходство с зефиром – отсюда название растения. Их окраска
обеспечивает дополнительную защиту кактуса от ультрафиолетового излучения.
Корневая система стержневая, главный корень водозапасающий, бывает сильно утолщён.
У основания главного побега на определённом этапе роста начинают развиваться
боковые побеги – такие же шаровидные, которые затем могут укореняться, даже
лёжа боком на земле, и давать начало вторичным побегам. Их число в заросли-клоне
может достигать многих десятков, и растение может распространяться на несколько
метров в стороны.
В период цветения на вершине стебля появляется множество малиновых цветков диаметром
до 12-13 см с жёлтым центром, содержащих множество тычинок и пестик. Днём цветки
издают сладкий аромат и привлекают насекомых для опыления – главным образом
ос и одиночных пчёл. Также на цветках растения охотно кормятся колибри.
Плоды растения – круглые (до 3-4 см диаметром) красные ягоды с многочисленными
мелкими чёрными семенами. Плоды служат пищей для многих животных (в основном
птиц и насекомых), которые распространяют семена в желудочно-кишечном тракте
или прилипшие к телу. Некоторые насекомые выстригают волоски зефирного кактуса,
пролезая между колючками. Паразитическое растение лжеомела
не встречается на нём, так как колючки и волоски надёжно защищают поверхность
побега от заноса животными семян паразита. Но гриб
трихоботритис может поражать этот вид пьедрой во время цветения, при этом
на «волосяном покрове» кактуса под колючками развивается серый ватообразный
налёт мицелия гриба. В зарослях зефирного кактуса находят укрытие многие животные
(змеи, ящерицы, мелкие млекопитающие) – колючки кактуса служат им надёжной защитой.
Этот вид растений открыл Биолог, участник форума.
Росянка-спрут
(Arbopendera nocturna)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Росянковые (Droseraceae)
Места обитания: степные рощи южной Европы.
В неоцене юг Европы изменился сравнительно мало: от средней полосы и до Балкан
раскинулись обширные степи, где произрастают травянистые растения разных семейств.
Лишь там, где существуют более-менее постоянные реки и другие крупные водоёмы,
растут и более крупные растения – кустарники и деревья.
Но растения адаптируются к среде обитания не хуже животных и грибов, и даже
там, где произрастают деревья и кусты, своё место смогли найти более мелкие
представители флоры.
Акациелистный клён является
одним из самых успешных и высоких деревьев европейских степей; он часто образует
густые заросли на берегах степных рек. Но на его ветвях, помимо его собственных
перистых листьев, встречаются и другие, не похожие на них – длинные, зелёные,
блестящие, свисающие вниз с ветвей дерева, как щупальца хищного спрута. Эти
листья принадлежат росянке-спруту, одному из местных видов хищных растений.
Росянка-спрут является розеточным растением: её стебель очень короткий, увенчанный
розеткой из длинных и широких листьев, которые свешиваются с ветвей акациелистного
клёна (или других степных деревьев) и свободно покачиваются в воздухе. Корневая
система растения представлена множеством длинных и тонких белых корней, которые
образуют под корой и в трещинах коры дерева-хозяина своеобразную сетку. Однако
росянка-спрут является не паразитом, а эпифитом: большую часть необходимых питательных
веществ она добывает самостоятельно, как и его сородичи.
Но у росянки-спрута есть уникальная особенность, которая, пожалуй, не встречается
у её сородичей: это строго ночное растение. Днём листья растения сворачиваются
в тугой комок и практически не заметны для разных травоядных зверей, которые
заходят в рощу, чтобы поглодать листву деревьев. Растение в таком положении
получает достаточно света для осуществления фотосинтеза. На закате, когда температура
окружающей среды несколько понижается, а влажность воздуха возрастает, листья
разворачиваются и свисают вниз с ветвей дерева, на котором растёт эта росянка.
Различные комары, мотыльки и другие ночные насекомые подлетают к этим листьям,
привлечённые каплями сладковатой жидкости, и прилипают к ним. Почувствовав,
что к листьям что-то прилипло, росянка сворачивает их и начинает поглощать пойманных
насекомых.
Кроме ловли насекомых, это растение добывает с помощью листвы необходимую влагу.
К утру на листьях росянки начинает оседать роса, и растение реагирует на неё
точно так же, поглощая её всей поверхностью листа. В степи дождь выпадает редко,
поэтому практически вся вода, доступная росянке – это роса. Свернувшиеся днём
листья помогают сберечь влагу на жарком солнце.
Росянка-спрут начинает цвести во второй половине весны. Её цветки, похожие на
мелкие колокольчики, распускаются по ночам и растут на длинных цветоносах, поднятых
вверх (листья свисают вниз, поэтому насекомые-опылители находятся в относительной
безопасности). Те же насекомые, которые служат росянке-спруту кормом, опыляют
её и переносят пыльцу с цветка на цветок. Плоды росянки-спрута представляют
собой мелкие невскрывающиеся коробочки с мясистыми стенками, похожие на белёсые
ягодки. Их охотно поедают местные певчие птицы; семена проходят через их пищеварительную
систему без особого для себя ущерба и успешно прорастают, если попадают в подходящую
для их роста среду (ветки клёна или другого степного дерева).
Этот вид растений открыл Bhut, участник форума.
Росянка
прорастающая (Drosera prolifera)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Росянковые (Droseraceae)
Место обитания: влажные тропические леса Флориды, побережья Мексиканского залива,
Куба.
В неоцене на территории Флориды и североамериканском побережье Мексиканского
залива сложились благоприятные условия для развития тропических лесов. Это мир
обширных болот и низменностей, царство влаголюбивых растений.
Среди травянистых растений этих мест часто встречаются насекомоядные растения
различных семейств. На суше и по краям болот часто встречаются различные виды
росянок. Один из их представителей – росянка прорастающая, довольно крупный
вид этого семейства.
Росянка прорастающая представляет собой многолетнее розеточное растение с коротким
клубневидным стеблем. Листья сидят на прочных длинных черешках; диаметр розетки
взрослого растения достигает 30 см. Этот вид может расти не только на земле:
росянка прорастающая является факультативным эпифитом. Если птицы или ветер
заносят семена этого растения на деревья, она нормально растёт, используя в
качестве субстрата древесную труху и мох. Обычно же это обитатель моховых зарослей
в подлеске.
Листья росянки прорастающей покрыты характерными ловчими щупальцами, выделяющими
капли липкой жидкости, к которой приклеиваются пойманные животные. Диаметр листьев
этого вида достигает 5 см. Лист имеет ложкообразную форму, а его верхняя сторона
слегка прогнута. Окраска листа красновато-коричневая, а щупальца ярко-красные
с белыми точками, которые образованы группами клеток, отражающими ультрафиолетовые
лучи. Так растение приманивает насекомых, но это не единственная его добыча.
Благодаря относительно крупным размерам росянка прорастающая способна ловить
и переваривать мелких лягушек и ящериц.
Благодаря тропическому климату в местах его обитания, это растение цветёт почти
круглый год. Цветонос высотой около метра, тонкий и упругий, пурпурного цвета.
У росянки прорастающей мелкие белые или жёлтые цветки с пятью лепестками.
Этот вид отличается лёгкостью вегетативного размножения, благодаря чему может
быстро расселяться. Лист этого растения легко отламывается в месте прикрепления
к нему листовой пластинки. Отломленный от растения лист сохраняет жизнеспособность
и двигательные реакции по отношению к попавшим на него животным. Иногда мелкая
лягушка может отломить лист, случайно попав на него и пытаясь освободиться.
Но отломившийся лист продолжает выделять пищеварительные ферменты и может убить
и переварить такую добычу самостоятельно. На изломе листа вскоре появляется
почка, из которой в дальнейшем развивается молодое растение. Также придаточная
почка есть на вершине листа. Если кончик нормально растущего листа касается
земли, почка начинает развиваться и образует молодое растение. Оно постепенно
укореняется и начинает расти самостоятельно рядом с материнским растением. Также
придаточная почка развивается на отломленном листе, но только при наличии дополнительного
питания.
Молодые растения зацветают уже на втором году жизни.
Росянка
непентолистная (Neodrosera nepenthifolia)
Порядок: Гвоздикоцветные (Caryophyllales)
Семейство: Росянковые (Droseraceae)
Место обитания: окраины Средиземноморской котловины.
В голоцене род росянок (Drosera) был одним из самых распространённых среди насекомоядных
растений. Росянки могли расти на разных почвах – от влажных болот до гор и песчаников.
Их ловчий аппарат устроен довольно просто: длинные железистые волоски с каплями
клейкого сока на конце покрывают листья различных форм и размеров. Раздражение
листа веществами белковой природы заставляет его скручиваться, а выделяемый
сок расщепляет эти вещества. В неоцене росянковые встречаются на всех материках,
даже в Антарктиде.
Один из потомков европейских росянок освоил Средиземноморскую котловину и обзавёлся
почти постоянными микроскопическими сожителями, позволяющими выживать в таких
суровых условиях. Непентолистная росянка живёт на сухих почвах окраин этой котловины,
избегая солончаков. Она сохранила множество признаков своих предков: прикорневую
розетку диаметром 20-30 см из 15-20 листьев и тонкий высокий (до 50 см) цветонос,
на конце которого развивается соцветие.
Корневая система непентолистной росянки отличается внушительной длиной – до
нескольких метров в глубину, что связано с необходимостью добычи пресной воды
из глубоких слоёв. Основной корень реповидно утолщается прямо под розеткой,
и на корневой шейке образуются почки, из которых вырастают новые побеги. Листья
по облику напоминают курительную трубку: на конце черешка находится преобразованная
листовая пластинка – небольшой (около 3 см глубиной) кувшинчик с крышечкой.
Лист сильно вогнут, и укороченные железистые волоски находятся на его внутренней
поверхности, покрывая две трети её площади. Одна сторона листа вытянута вверх
и загибается, образуя крышечку. Края кувшинчиков гладкие и скошенные внутрь,
способствуют падению насекомых внутрь ловушки. Волоски внутри кувшинчика выделяют
воду, сахара и пищеварительные ферменты для переваривания добычи. Кувшинчики
всегда заполнены жидкостью до уровня верхних волосков на их поверхности, и весь
кувшинчик изнутри покрыт плотной кутикулой. Испарение компенсируется выделениями
растения и при случае – дождями и росой. Жидкость издает сладкий запах, привлекающий
насекомых.
Все надземные части растения (стебель, черешки и наружная сторона листьев-ловушек)
светло-зелёные, края кувшинчиков тёмно-красные.
Цветёт непентолистная росянка летом, в период максимального лёта насекомых-опылителей.
Цветки около 1,5 см в диаметре собраны в колосовидное соцветие, в котором насчитывается
до 10-12 цветков. Строение цветка такое же, как у предков: пятичленная зелёная
чашечка, пятичленный ярко-красный венчик, пять тычинок и один пестик с верхней
многогнёздой завязью, у основания которой имеются нектарники, выделяющие душистый
нектар. Растение не специализировано к опылителям: цветки посещаются одиночными
пчёлами и осами, мухами, жуками и бабочками. Плод – дольчатая коробочка, вскрывающаяся
крышечкой с растрескиванием, с мелкими, как у мака, семенами, разносимыми ветром.
Всхожесть семян сохраняется около года, они обычно прорастают в период редких
весенних дождей.
Но самое необычное в жизни этого растения – его микроскопические симбионты.
В тканях непентолистной росянки живут легуминобактерии,
образующие клубеньки на корнях и черешках листьев, когда растение испытывает
дефицит азота из-за нехватки добычи. Растение выживает на бедных почвах благодаря
способности добывать азотные соединения при помощи симбионтов. Над жидкостью
и на плавающей там добыче часто встречаются корненожки Drosarcella
и Droseroflugia – потомки
голоценовых родов Arcella и Difflugia, соответственно. А в самой жидкости кувшинчика
обитают жгутиконосцы Endobodo
– потомки голоценового рода Bodo. Корненожки и жгутиконосцы – комменсалы у стола
росянки, находящие у неё приют, пропитание и влагу, которые непросто добыть
в солёной пустыне.
Этот вид растений открыл Биолог, участник форума.
Ложноягодник
маврикийский (Pseudobaccodendron mauritianus)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Маревые (Chenopodiaceae)
Место обитания: остров Маврикий, леса.
В эпоху человека растительный и животный мир острова Маврикий подвергся тяжёлому
испытанию – за несколько сотен лет на остров было завезено едва ли не больше
видов растений и животных, чем появилось на нём за миллионы лет естественной
эволюции острова. Вполне естественным результатом этого явилась деградация
исконной флоры и фауны острова, и её полное вымирание к неоцену.
В условиях, когда коренная маврикийская флора оказалась в угнетённом состоянии,
завезённые виды начали активно эволюционировать. В благоприятных тропических
условиях даже травянистые растения стали гигантами и превратились в деревья.
Одно из таких деревьев, ложноягодник маврикийский, широко расселилось по влажным
долинам и побережьям острова. Его предок – один из видов лебеды (Atriplex),
завезённой на Маврикий человеком. В процессе эволюции лебеда превратилась из
травянистого растения вначале в одревесневающий полукустарник, затем в кустарник,
и дальше в дерево. Семейство маревых на континентах представлено в основном
травянистыми растениями, но изредка среди них встречались древовидные формы.
Саксаул (Haloxylon) – один из таких видов. Он приспособлен к экстремальным
условиям пустыни, и показывает возможность появления древовидных растений в
семействе, представленном большей частью травами. В отличие от саксаула, ложноягодник
не привык к такой спартанской жизни: это растение тропического леса, любящее
влагу и обилие солнца.
Высота ложноягодника бывает до 20 – 25 метров при толщине ребристого ствола
до 3 метров у основания. Это дерево с мощной глубокой корневой системой (как
ответ на наличие в неоценовой фауне копытных на Маврикии) и твёрдой белой древесиной.
Кора ложноягодника толстая, покрытая глубокими продольными трещинами, светло-серая.
Крона этого дерева похожа на кипарисовую – она высокая, сужающаяся к вершине.
Листья ложноягодника внешне похожи на ивовые – они узкие и продолговатые, густо
покрывающие молодые побеги. Сверху каждый лист покрыт микроскопическими пузыревидными
волосками, защищающими от избытка солнечного излучения и придающими кроне дерева
заметный издали серебристый оттенок. Особенно много волосков на молодых листьях,
имеющих розоватый оттенок. Волоски защищают от солнечных ожогов, рассеивая
свет, доходящий до поверхности листа, но мешают испарять воду. Однако, у растения
есть приспособление, решающее такую проблему: на краю листа расположены гидатоды
– желёзки, выделяющие воду. Время от времени из них выделяется капля воды,
которая быстро скатывается с листа благодаря покрову из волосков. Поэтому,
бродя в рощице ложноягодника маврикийского, животное может быстро промокнуть
даже в сухой день. Произрастает это дерево в долинах рек и невдалеке от болот.
Ложноягодник цветёт на редкость невзрачно: у крошечных цветков, опыляемых ветром,
нет лепестков, и внешне цветение дерева можно определить, лишь специально изучив
его цветки. Очень небольшие цветки без лепестков – это общая особенность представителей
семейства. Зато плодоносящее дерево заметно издалека. Своё название это растение
получило за особенность плодов. У маревых плоды обычно сухие, плотно покрытые
обёрткой из чашелистиков, и отпадающие вместе с нею. Сочный плод – это скорее
исключение из правила. И ложноягодник – как раз одно из таких исключений. У
него чашечка к моменту созревания семян не усыхает, а разрастается, облекая
плод со всех сторон, и становится сочной и даже сладковатой. Зрелый плод ложноягодника
размером с крупную вишню. Меняется и его цвет: чашечка становится ярко-красной,
издали выделяясь на фоне серебристой листвы и привлекая птиц и обезьян, питающихся
плодами. Зрелые плоды легко осыпаются от ветра, и их охотно поедают местные
наземные звери: очень мелкие лесные
свиньи и дриокапреллы – карликовые потомки
коз.
Когда семечко дерева проходит через желудок животного, его оболочка становится
более проницаемой для воды, и оно легче прорастает. Множество сеянцев ложноягодника
гибнет, либо оказывается угнетёнными в тенистом подлеске. В тени их рост сильно
замедляется, и растения как бы останавливаются в росте на высоте не более метра.
Большинство таких сеянцев бывает съедено местными травоядными животными. Но,
стоит только какому-то старому дереву упасть, сеянцы ложноягодника начинают
быстро расти дальше. В первый год они отрастают на два метра, с минимальным
ущербом «проскочив мимо рта» местных карликовых травоядных. Далее темп роста
уменьшается, но зато ствол дерева начинает утолщаться и около его основания
отрастают мощные досковидные корни.
Ложноягодник доживает до 300 лет и более, и начинает плодоносить с 10-летнего
возраста, достигнув примерно половины максимальной высоты.
Пятикрылое
дерево (Pentalaminodendron fragilis)
Порядок: Гвоздичноцветные (Caryophyllales)
Семейство: Маревые (Chenopodiaceae)
Место обитания: тропические леса Юго-Восточной Азии.
Восстановление лесов Юго-Восточной Азии, сильнее всего пострадавших от деятельности
человека, сопровождалось множеством попыток травянистых растений колонизировать
эти леса. Часть видов травянистых растений так и осталась на вторых ролях, среди
пионерной растительности повреждённых местообитаний. Но некоторые из бывших
трав сделали удачную попытку превратиться в деревья. Одним из таких растений
была лебеда (Atriplex). На острове Маврикий одним из потомков этого растения
является ложноягодник маврикийский
(Pseudobaccodendron mauritianus). А в азиатских тропиках произрастает своеобразное
пятикрылое дерево – растение с очень узнаваемым обликом.
Среди множества стволов разных цветов и текстур коры этот вид деревьев распознаётся
безошибочно. Этот вид – высокое (до 30 метров в высоту) дерево с пятигранным
стволом. Уже у молодого растения ствол показывает характерную пятигранную форму
с рёбрами, оттянутыми в виде слегка волнистых гребней, с крупными одиночными
листьями, расположение которых соответствует этим рёбрам. Это быстрорастущее
и очень светолюбивое дерево – оказавшись в «окне» полога леса, оно даёт прирост
до 5 метров в год, и уже на втором году жизни начинает ветвиться. В течение
всей жизни этот вид сохраняет пять продольных гребней вдоль всего ствола, и
ствол на срезе выглядит пятилучевой звездой с тонкими лучами. Ширина этих рёбер
может достигать 2-3 метров. В нижней части ствола они плавно переходят в досковидные
корни, разрастающиеся в разные стороны приблизительно на ширину кроны дерева.
Крона пятикрылого дерева широкая, зонтиковидная. Ветви на большей части ствола
засыхают и опадают от недостатка света. Ветви ориентированы строго по направлениям
роста рёбер, и ребро продолжается вдоль нижней стороны ветви, усиливая её. Когда
дерево достигает верхнего яруса леса, его рост значительно замедляется.
Древесина этого вида твёрдая, но хрупкая, поэтому широкие рёбра на стволе служат
для придания дереву прочности и устойчивости, но одновременно позволяют не наращивать
огромную массу древесины, занимая верхний ярус леса. Листья этого вида широкие,
ромбовидные, с оттянутым в капельницу кончиком. Молодые листья с обеих сторон
покрыты белым мучнистым налётом, на взрослых листьях нижняя сторона листа покрыта
слоем серебристых волосков, а верхняя – гладкая, светло-зелёная.
Цветение этого вида протекает непримечательно – его цветки безлепестные, с пятью
кожистыми чашелистиками. Они неприятно пахнут и опыляются жуками, которые поедают
часть обильной пыльцы. Цветки собраны в небольшие кисти, растущие на стволе
и нижней части крупных ветвей, на узкой части ребра. Плоды пятикрылого дерева
– прочные сухие коробочки с твёрдыми семенами. Они вскрываются птицами и грызунами,
которые разносят семена. Семена этого вида унаследовали от предка-лебеды способность
долго сохранять всхожесть и неодновременное прорастание. Иногда семена могут
лежать в почве до 5 лет, прежде чем из них покажутся ростки. В пятилетнем возрасте
пятикрылое дерево начинает цвести, а продолжительность жизни может достигать
120 лет.
Виноград
плющевидный (Vitis hеderomorphus)
Порядок: Виноградоцветные (Vitales)
Семейство: Виноградовые (Vitaceae)
Место обитания: влажные леса Кавказского полуострова и Балкан.
После потепления климата в неоцене области тропического и субтропического климата
значительно расширились, и на Кавказе и Балканах установился климат, близкий
к тропическому, с относительно мягкой зимой. Это наложило отпечаток на внешний
вид растений, составляющие леса в этих регионах. В облике многих эндемичных
видов наблюдаются особенности, характерные больше для растений тропиков.
Характерным компонентом флоры этих мест являются дикие виды винограда – потомки
одомашненного винограда (Vitis vinifera), занимающие различные экологические
ниши в растительном сообществе.
Во влажных тенистых чащах лесов, на замшелых стволах древесных гигантов разрастается
множество видов эпифитных растений, в том числе мелкий потомок культурного винограда
– виноград плющевидный, карликовое растение с тонкими длинными побегами и небольшими
листьями. Как и все эпифиты, он прорастает на стволах деревьев, укореняясь в
складках коры и зарослях мха. Когда стебель растения касается подходящего субстрата,
на нём развиваются дополнительные корешки. На твёрдых субстратах растение даёт
многочисленные короткие корни, внедряющиеся в малейшие щели субстрата.
Лоза растения тонкая, светло-зелёная, слабо одревесневающая и обильно ветвящаяся.
Листья небольшие, пятиугольные, с мелкозубчатым краем и тонким кончиком-«капельницей».
С возрастом более старые части растения покрываются рыхлой шелушащейся корой.
Длина побегов достигает 4-5 метров, они густо покрывают нижнюю часть стволов
крупных деревьев.
Цветки мелкие, желтоватые, собраны в многоцветковую метёлку. Плоды в виде небольших,
не более пяти миллиметров, ягод фиолетового цвета, собранных в очень плотную
кисть, структурой и формой больше похожую на крупную ягоду малины. Растение
плодоносит два, а в тёплых регионах – три раза в год. Семена разносят птицы,
поедающие ягоды. Молодое растение зацветает уже на втором году жизни. Зимой
растение теряет часть листьев и впадает в состояние покоя. Продолжительность
жизни – до 15 лет.
Этот вид растений открыл Моргот, участник форума.
Листокоготник
мексиканский (Phyllonyx mexicanum)
Порядок: Бобовые (Fabales)
Семейство: Бобовые (Fabaceae)
Место обитания: пустыни Мексиканского нагорья.
В пустынях произрастают растения, способные противостоять суровым природным
условиям и выигрывать в жёсткой и бескомпромиссной борьбе за существование.
У них два главных врага – сухость и травоядные животные. И растения в процессе
эволюции вырабатывают различные стратегии. В пустыне, раскинувшейся на Мексиканском
нагорье, одно из растений приспособилось к тому, чтобы давать отпор травоядным
животным.
Когда в пустыне начинается весна, из земли появляются аккуратно сложенные перистые
листья. Они быстро растут, образуя компактные розетки. Каждый сложный перистый
лист длиной около 30 – 40 см состоит приблизительно из десяти пар овальных простых
листочков длиной до 3 см. На конце каждого листочка средняя жилка формирует
очень острую колючку. Эта особенность определила название растения – листокоготник
мексиканский. Колючки – это очень распространённое оружие пустынных растений.
Но листокоготник добавил к этой особенности строения ещё одно умение, характерное
для бобовых (впрочем, не только для них одних). В основании каждого простого
листочка есть прослойка ткани, состоящая из клеток, способных быстро накачивать
воду и освобождаться от неё. Это придаёт листу замечательные свойства.
Обычно розетки листокоготника свободно лежат на земле, и листочки раскрыты навстречу
лучам солнца. Но, если травоядное животное прикоснётся к ним, наступает удивительная
трансформация: листочки начинают складываться вдоль средней жилки, и одновременно
поднимаются вверх. Растение буквально на глазах «ощетинивается» колючками. Если
его не тревожат, через несколько минут листья снова принимают прежнее положение.
Это растение максимально полно использует благоприятное для роста время. Весной
листокоготник активно вегетирует, наращивая листву. Не менее бурно он нарастает
под землёй. Это растение запасает воду в подземных клубнях, покрытых толстой
кожурой. На кожуре имеются крупные бородавки – это не проявление болезни, а
совершенно необходимая для жизни растения особенность. В бородавках живут колонии
азотфиксирующих бактерий. Эти симбионты помогают растению получать необходимый
для роста азот прямо из воздуха, и благодаря им листокоготник может поселяться
на самых бедных почвах, и процветает там, где другие растения хиреют. Разрастаясь,
листокоготник образует дочерние клубни на длинных хрупких столонах. Когда крупное
травоядное животное (например, медвежий
дикобраз, характерный житель мексиканской пустыни) раскапывает почву и поедает
главный клубень, столоны разрываются, и дочерние клубни выживают, не замеченные
этим зверем.
Вскоре после развития молодых листьев растение зацветает. Цветки листокоготника
зигоморфные, с хорошо развитым лепестком-«парусом». Они ярко окрашены – сиреневые
с серебристыми полосками, сходящимися в зеве цветка. Волоски обладают свойством
отражать ультрафиолетовый свет, благодаря чему насекомые замечают их издалека.
Это растение опыляется преимущественно пчёлами и жуками. Нектар цветка спрятан
глубоко в зеве, и для того, чтобы достать его, насекомое должно пролезть в глубину
цветка. В этот момент тычинки касаются его спины, и насекомое оказывается обсыпанным
пыльцой. Одно из пустынных насекомых, бражник-шипун,
может просто «красть» нектар. Бабочка зависает в воздухе перед цветком, просовывает
длинный тонкий хоботок между лепестками, и высасывает нектар, не производя опыления.
Каждый цветок цветёт недолго – около четырёх дней. Но это растение образует
длинное соцветие, на котором цветки сидят гроздьями, поэтому в целом растение
цветёт на протяжении двух месяцев и более. Если опыление прошло успешно, примерно
из трети цветков формируются плоды. К моменту созревания плодов цветонос ложится
на траву. Плоды у листокоготника особые – каждый плод содержит всего одно крупное
(длиной до 2 см) плоское семя. Обычно плоды бобовых растений вскрываются из-за
неравномерного высушивания волокон, пронизывающих его створки. Но у листокоготника
плод не вскрывается, а на его конце развивается двойная загнутая колючка, покрытая
щетинками. Такой плод напоминает блесну с крючком, и на одном цветоносе насчитывается
до двух десятков таких плодов. К моменту созревания плод держится на цветоножке
очень слабо – он готов к встрече с животными, которые их распространяют. Крупная
равнинная сова, сумчатая
гиена или медвежий дикобраз – вот основные распространители плодов этого
растения. Прицепившись к их покровам, плод листокоготника переносится ими на
большое расстояние. Рано или поздно животное вычёсывает его из покровов тела,
и плод падает на землю. Возле укрытия сов или пыльных ванн медвежьего дикобраза
часто образуются скопления этого растения. Семя долго сохраняет всхожесть, а
из-за плотной оболочки прорастание обычно затягивается. Это целесообразная тактика
– растение не прорастёт во время случайного дождя, после которого может быть
ещё несколько недель жаркой сухой погоды, либо во время кратковременного зимнего
потепления. В первый год жизни листокоготник формирует только розетку коротких
листьев, на второй год листья достигают нормального размера, а с третьего года
жизни растение начинает цвести.
Охотничье дерево (Prolesrobinia venator)
Порядок: Бобовые (Fabales)
Семейство: Бобовые (Fabaceae)
Место обитания: степные области Евразии.
В XVII веке в качестве декоративных растений в Евразию были завезены различные
акации – под этим названием в эпоху человека были известны деревья нескольких
родов. Среди них была и робиния клейкая, иначе называемая робиния железистоопушенная
(Robinia viscosa). Отличительным признаком этого 12-метрового растения были
липкие волоски железистого опушения, густо покрывающие побеги, черешки листьев
и соцветий. После исчезновения человечества, в период ранненеоценового оледенения
этот вид оказался на грани вымирания, но экстремальные условия разбили единый
ареал вида на несколько изолированных участков и тем самым ускорили эволюцию
этого растения. И в неоценовых степях Евразии произрастает потомок робинии
клейкой – охотничье дерево. Подобно томату-мухоловке, охотничье дерево находится
на раннем этапе специализации к питанию насекомыми и другими мелкими беспозвоночными,
хотя эта специализация более выражена, чем у томата-мухоловки. Охотничье дерево
– самое крупное хищное растение неоцена, достигающее до полутора метров в высоту.
По внешнему виду это растение похоже на своего голоценового предка, а также
на другие акации эпохи голоцена. У него длинные непарноперистые листья, состоящие
из 8 – 11 пар мелких округлых листочков.
Как и у предкового вида, побеги, черешки листьев и соцветий охотничьего дерева
густо покрыты липкими волосками. Но, в отличие от своего предка, это приспособление
для охоты. На волосках выделяется много густой и клейкой жидкости с привлекательным
для насекомых запахом. Если насекомое садится на эту жидкость, то прилипает,
а потом волоски врастают в тело жертвы. Каждый волосок представляет собой гигантскую
клетку с прочной оболочкой. Когда насекомое садится на лист и прилипает к волоскам,
вещества, выделяемые его телом, стимулируют прорастание верхушки волоска в
ткани насекомого. Вершина волоска начинает активно делиться и прорастает в
тело жертвы в виде цепочки тонкостенных клеток, которые впитывают его жидкое
содержимое. После того, как добыча высосана, волоски, проросшие в неё, отмирают.
Старые листья перестают выделять жидкость, и некоторое время участвуют в фотосинтезе.
Охотничье дерево опасно даже для мелких позвоночных вроде небольших древесных
лягушек. Осенью рост листвы замедляется. Молодые листья перестают расти и выделять
клейкую жидкость, а старые опадают. Весной рост растения возобновляется.
Цветы охотничьего дерева крупные, красивые, розоватого оттенка. Они собраны
в узкие двурядные кисти и поднимаются над кроной на длинных цветоносах, чтобы
насекомые-опылители не сели случайно на ловчие волоски. Около цветков ловчие
волоски отсутствуют, чтобы не препятствовать процессу опыления. Цветение охотничьего
дерева проходит в середине лета в течение двух – трёх недель. Плоды этого вида
– длинные тонкие стручки. Их створки при высыхании скручиваются тугой спиралью
и разбрасывают семена далеко от материнского растения.
Охотничье дерево светолюбиво, но любит влажную почву. Оно произрастает по берегам
рек и стариц, часто на заболоченной почве. В течение первых лет жизни это растение
развивается за счёт симбиоза с клубеньковыми бактериями, но во взрослом состоянии
получает значительную часть азота из пойманной добычи, а клубеньки на корнях
уменьшаются. На третьем – пятом году жизни растение начинает цвести. Продолжительность
жизни не превышает 20 лет.
Этот вид растений открыл Odin, участник форума.
Степная
роза (Rosa aridophila)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: североамериканские прерии.
Вопреки популярному мнению, семейство розовых не ограничивается одним шиповником
и его домашними потомками. Среди его представителей есть и разные виды тундровых
растений дриад, и одна из них, маликаат,
стала в неоцене одним из характерных травянистых растений тундры, своеобразным
«спутником» местных травоядных зверей. Другой член семейства розовых – «настоящая»
роза, американский потомок шиповника эпохи голоцена.
Степная роза – засухоустойчивое кустарниковое растение прерий. Она напоминает
своего предка в общих чертах, но вырастает до 3,5-4 м в высоту. Отдельное растение
степной розы – это скорее куст с несколькими стволами, чем одноствольное дерево.
Его ветви длинные и колючие, и даже одиночное растение выглядит неприступным
для травоядных, но степные розы чаще растут небольшими рощами, поскольку дают
значительное количество корнеотпрысков.
Листья овальные (фактически, это сложный лист из одного листочка), с зубчатым
краем; листорасположение очередное. Цветки крупные, пятилепестные, типичного
для шиповников облика. Лепестки тёмно-розовые, но иногда встречается цветовая
морфа, у которой цветы, наоборот, белоснежные. Они опыляются разными насекомыми,
в первую очередь местными видами пчёл и шмелей. Цветение растения происходит
весной и в самом начале лета, пока почва достаточно хорошо снабжает растение
влагой.
Степные розы имеют прочные, длинные и колючие ветки; колючки плоско-треугольной
формы одревесневают и подолгу сохраняются на побегах. Крупные травоядные звери
с большой осторожностью объедают эти кусты, а от гусениц, улиток и других беспозвоночных
степную розу защищают местные мелкие певчие птицы, которые часто гнездятся в
гуще куста этого растения, защищённые от врагов. Летом растение сбрасывает часть
листвы, а в особенно сухое лето опадает также значительная часть недозрелых
плодов. При повреждении кроны растения у него образуется обильная корневая поросль,
в том числе на расстоянии нескольких метров от взрослого растения. Так образуются
обширные разреженные заросли этого вида.
Осенью, когда у степной розы созревают плоды, степные млекопитающие охотно приходит
к её зарослям и поедают опадающие плоды. Плод растения – шарообразный цинародий
диаметром около 20 мм со сладкой мякотью и крупными деревянистыми семенами.
Семенам степной розы пребывание в кишечнике травоядного идёт только на пользу
– они от этого лучше прорастают. Поэтому степная роза часто растёт вдоль миграционных
путей крупных травоядных. Молодое растение зацветает примерно на пятом году
жизни.
Если степную розу не съедят в первые годы жизни, когда она ещё маленькая и плохо
защищённая, то она вполне сможет прожить несколько десятков лет.
Этот вид растений открыл Bhut, участник форума.
Горный
шиповник, якорная роза (Ancorosa montana)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: Альпы, южные склоны.
Шиповники были типичными представителями европейской флоры в эпоху человека.
Некоторые из их видов были крайне редкими узкоареальными эндемиками и быстро
вымерли, тогда как другие были широко распространены и легко приспосабливались
к изменённым человеком ландшафтам. После исчезновения человека выжившие виды
шиповников начали активно эволюционировать, занимая новые экологические ниши.
Одна из своеобразных жизненных форм потомков шиповника – якорная роза, обитающая
на склонах Альп. Это типичное горное растение, образующее длинные «бороды» свисающих
побегов (длиной до 4-7 метров), отрастающих от короткого ползучего стебля. Крючковатые
колючки на коре побегов позволяют растению закрепляться в трещинах скал. Побеги
якорной розы легко укореняются в любых местах, где скапливается хоть какая-то
почва и развиваются мхи и лишайники. Корни растения прорастают в трещины скал,
заполняя их, и являются важными участниками процесса выветривания. Во время
обвалов побеги растения отрываются, но быстро укореняются на новом месте. Побеги
якорной розы укрепляют горные склоны, образуя в некоторых местах сплошной ковёр
из ползучих стеблей.
Якорная роза – вечнозелёное растение. У него сложные непарноперистые листья
из 3-5 листочков с кожистой верхней стороной коричневато-зелёного цвета. Такой
оттенок листу придаёт большое количество антоциана, защищающего растение от
интенсивного солнечного излучения. Молодые листья ярко-красного цвета. Изнанка
листьев бело-войлочная, способствует сохранению влаги.
Подобно своим предкам, якорная роза – насекомоопыляемое растение. В конце весны
и начале лета в пазухах листьев развиваются яркие цветки диаметром 4-6 см –
жёлтые или бело-розовые. Они опыляются пчёлами, шмелями и бабочками. Осенью
созревают съедобные плоды-цинародии с мясистыми стенками ярко-красного цвета,
мякоть которых имеет кисловато-сладкий вкус. Эти плоды содержат около десятка
крупных семян с прочной толстой оболочкой, которые сохраняют всхожесть до 10
лет. Распространителями плодов являются главным образом птицы и грызуны.
Молодое растение впервые зацветает на 3-м году жизни. Продолжительность жизни
достигает 30-40 лет.
Дендророза
съедобная (Dendrorosa esculenta)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: Балканы, Малая Азия, предгорья.
Эпоха неоцена отмечена появлением значительного количества видов растений, сделавших
успешную попытку освоить новые жизненные формы. Например, среди капустных,
бегоний и полыней
появились деревья, а среди ряда семейств растений, бывших в эпоху человека главным
образом травами, появились многолетние
одревесневающие лианы. Во флоре
субтропиков и теплоумеренных широт Евразии возникли своеобразные древовидные
растения семейства розовых – потомки одного из распространённых видов шиповника
эпохи человека.
В субтропических лесах южнее горного пояса Альп одним из обычных видов деревьев
является дендророза съедобная – дерево высотой до 20-25 метров с раскидистой
кроной и очередными сложными непарноперистыми листьями. Длина листа – около
50 см, он состоит из 11-17 листочков длиной 6-9 см. Это полулистопадное растение,
осенью старые листья желтеют и опадают. Молодые листья богаты антоцианом и имеют
яркую окраску – от ярко-розовой до пурпурной. По мере развития окраска меняется
на ярко-зелёную, характерную для зрелого листа. Растение очень светолюбиво.
Молодые побеги колючие, кора часто яркого цвета – красная или коричневатая.
На старых побегах шипы отпадают, крупные ветви и ствол покрывает рыхлая мелкотрещиноватая
кора.
Весной начинается интенсивный рост растения, и крона приобретает красивый розоватый
оттенок. После формирования нескольких листьев в их пазухах развиваются цветки
с белыми лепестками, сильно отражающими ультрафиолетовый свет и хорошо заметными
для насекомых. Эта окраска дополняется интенсивным сладковатым запахом.
Летом созревают плоды растения – съедобные яблоковидные цинародии диаметром
около 5 сантиметров, ярко-жёлтой или оранжеватой окраски, с высохшими чашелистиками
на вершине, которые легко опадают при созревании плода. Мякоть плода сочная,
иногда даже водянистая, особенно при хорошем увлажнении почвы. Семян в полости
плода может быть мало – на влажных питательных почвах развивается 5-6 семян
с прочной деревянистой оболочкой, а у растений с бедных почв сухих водоразделов
– всего лишь одно или два крупных семени. Распространителями семян чаще всего
бывают птицы, в том числе пещерные
голуби. Грызуны часто поедают сами семена, хотя также разносят их далеко
от материнского растения. При прохождении через кишечник позвоночного животного
оболочка семени частично растворяется и становится более проницаемой, что облегчает
прорастание семян.
Молодое растение впервые зацветает в возрасте 10 лет, достигнув высоты 11-13
метров. Продолжительность жизни достигает 80-100 лет.
В лесах вдоль южного берега Четвероморья произрастает небольшой быстрорастущий
вид дендророза
малая (Dendrorosa parva) – полулистопадное дерево высотой до 10 метров,
с колючими побегами и узкими непарноперистыми листьями. Избегая конкуренции
за опылителей, это растение зацветает ранней весной, до отрастания листьев,
и его плоды созревают уже в начале лета. Цветки розовато-белые, с интенсивным
запахом, а плоды-цинародии диаметром около 15 миллиметров содержат по 2-3 семени
и очень многочисленны. У них розовая окраска, интенсивный фруктовый запах и
водянистая кисло-сладкая мякоть.
Дендророза
крымско-кавказская (Dendrorosa taurocaucasica) имеет разорванный ареал:
она произрастает на острове Крым и на Кавказском полуострове. Распространение
явно связано с маршрутами перелётов птиц через Четвероморье, хотя не исключается
перенос плодов водой. Это дерево высотой до 20 метров (на острове Крым ниже,
до 10 м) с непарноперистыми листьями длиной до 30 см. Цветение летом, цветки
многочисленным, диаметром до 3-4 см, от розовых до красных, с узором из продольных
полос на лепестках, заметным в ультрафиолетовом свете. Плоды-цинародии красного
цвета созревают к концу лета, отличаются пряным ароматом и сладким вкусом, содержат
3-5 крупных деревянистых семян.
Дендророза
деликатесная (Dendrorosa delicatissima)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: Персидский хребет, горные долины в горах Центральной Азии к
югу от Четвероморья.
В эпоху человека различные виды шиповника (Rosa) были широко распространены
на территории Евразии. Среди них встречались как обычные и распространённые,
так и редкие узкоареальные виды. Пережив эпоху человека, один из видов шиповника
в Евразии эволюционировал в ряд видов древовидных растений, ставших частью субтропической
флоры неоцена. Один из этих видов – дендророза деликатесная, произрастающая
в горных долинах Центральной Азии и Персидского хребта.
Этот вид – вечнозелёное древовидное растение высотой до 10-15 метров, приспособленное
к жизни на бедных почвах в условиях жаркого солнца и нехватки влаги. Кора на
стволе и старых ветвях толстая, рыхлая, предохраняющая от перегрева. Верхний
слой коры отслаивается небольшими чешуйками. Ствол растения утолщённый, а пористая
непрочная древесина способна накапливать значительное количество воды. Обычно
крона дерева очень раскидистая, а ствол начинает ветвиться низко над землёй.
Ветви часто полегают и легко укореняются, продолжая рост, стелясь по поверхности
земли.
Листья очередные, сложные, тройчатые, с крупными быстро отсыхающими прилистниками.
Листочки расположены на конце короткого черешка. Верхняя сторона у них кожистая,
морщинистая, с многочисленными волосками, образующими редкое опушение. Нижняя
сторона листа покрыта мучнистым восковым налётом – насекомые часто соскребают
этот воск для постройки собственных гнёзд. Поверхность черешка покрыта многочисленными
острыми щетинками, ec
Молодые побеги покрыты крупными треугольными колючками красно-коричневого цвета
со слегка загнутым кончиком. Кора на таких побегах плотная, гладкая, вишнёво-красная;
антоциан защищает ткани растения от избытка солнечного света. С возрастом кора
на ветках утолщается, а колючки постепенно опадают.
Цветение дендророзы деликатесной происходит в начале весны. Цветки этого дерева
одиночные или собраны по 2-3 в небольших кистях. Они пятилепестковые, белые
или нежно-розовые, испускают сильный приятный запах. Этот вид опыляется перепончатокрылыми
и бабочками.
Плоды созревают к середине лета и представляют собой съедобные яблоковидные
цинародии с толстым слоем сочной мякоти и высохшими чашелистиками на вершине,
которые легко отваливаются. Кожица плодов бархатистая, покрытая многочисленными
волосками. Мякоть кисловатая, приятного вкуса, привлекательная для растительноядных
млекопитающих. Характерной особенностью является наличие всего лишь одного семени
внутри плода. Семя крупное, покрытое прочной оболочкой, способной противостоять
желудочному соку крупных млекопитающих. Распространение семян происходит при
помощи крупных местных травоядных – свиней, громорогов
и каркаданнов.
Молодое растение долгое время наращивает корневую систему, способную собирать
росу и конденсат на обширной площади, и достигающую глубоких водоносных горизонтов
почвы. Развитие происходит медленно: первое цветение происходит в возрасте 10-13
лет, продолжительность жизни составляет до 200 лет. Старое растение часто образует
клон из молодых побегов, нормально развивающихся после отмирания родительского
растения и живущих в общей сложности значительно дольше, чем одиночное растение.
Эглантерия
южная (Eglanteria notalis)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosidae)
Место обитания: Новая Зеландия, леса на равнинах и в предгорьях в области субтропического
климата.
Характерной особенностью эпохи человека стал масштабный обмен представителями
флоры и фауны между разными материками и островами, который позволил множеству
видов появиться в местах, недоступных им для расселения естественным путём.
Так в изолированном морями и расстояниями мире природы Новой Зеландии появился
шиповник красно-бурый (Rosa rubiginosa), завезённый людьми в историческую эпоху.
После исчезновения человека приобретённая в условиях материка конкурентоспособность
помогла потомкам этого вида занять видное место в экосистемах архипелага.
В неоценовых лесах Новой Зеландии растёт потомок этого шиповника – эглантерия
южная, дерево высотой до 20 метров. Это растение очень светолюбиво, и его раскидистая
крона является важной частью полога леса. У дерева маловетвистый ствол, достигающий
толщины около 120-150 см. Кора тёмно-бурая, рыхлая и волокнистая, на стволе
и старых ветвях шелушится небольшими пластинками. Молодые побеги колючие – эту
особенность растение унаследовало от предка. Колючки защищают молодые побеги
растения от травоядных, сохраняются на коре до 2-3 лет, далее постепенно опадают.
Листья этого вида сложные, состоят из 5 овальных листочков с остроконечной вершиной,
длиной до 20-30 см. Верхняя сторона листочков слегка морщинистая, кожистая,
тёмно-зелёная; изнанка листа белая с тонким войлочным опушением. Листочки сближены
на вершине черешка в виде зонтика. Черешок листа упругий, вздутый и красноватый
в основании, с прилистниками. Растение вечнозелёное, каждый лист существует
около 2 лет.
От своих предков эглантерия южная унаследовала склонность к опылению насекомыми.
Цветки этого вида одиночные, крупные, розового цвета; цветочные почки закладываются
на однолетних побегах в конце сезона роста. Цветки развиваются после зимнего
периода относительного покоя. Они пятилепестковые, диаметром около 10 см, с
многочисленными тычинками на длинных нитях. Характерной особенностью вида являются
крупные чашелистики неправильной формы в основании цветка. Основными опылителями
этого растения являются насекомые разных систематических групп – перепончатокрылые,
жуки, мухи и дневные бабочки. Иногда во время цветения это растение посещают
птицы, которые опыляют цветки во время поиска в них насекомых. Некоторые птицы
также слизывают нектар растения. Чашелистики в таких случаях служат насестом
для птиц.
Плод эглантерии южной – цинародий с сочными мясистыми стенками ярко-жёлтого
цвета, диаметром до 5 сантиметров. На плодах при созревании сохраняются чашелистики,
высыхающие к этому времени. Это зоохорное растение, опавшие плоды поедают наземные
животные. Семена обладают прочной оболочкой и лучше всходят после прохождения
через кишечник млекопитающего. Основные распространители плодов – олени новозеландские
ультрадамы и сумчатые медвежьи
кускусы, а также кролики-кастеляны.
Молодое растение лучше всего растёт в условиях хорошей освещённости, в тени
рост сильно замедляется. Первое цветение молодого растения происходит в возрасте
9-12 лет, продолжительность жизни – до 300 лет.
Родственным видом является эглантерия
мелкоплодная (Eglanteria microcarpa), произрастающая в горных лесах
Южного Острова и на островах близ его побережья. Плоды этого вида оранжевые
или красные, диаметром до 2-3 см, поедаются птицами, которые разносят семена.
Цветки этого растения розово-красные с белыми пятнами в основании лепестков
и с длинными тычинками, опыляются главным образом мелкими певчими птицами, мухами
и жуками. Листочки более узкие, длиной до 15-20 см при ширине около 2 см – это
позволяет им не травмироваться при сильном ветре. Зимой растение частично сбрасывает
листья.
Земляника
антарктическая (Xerofragaria antarctica)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: тундра и луга Антарктиды.
После потепления климата и отступления ледников флора Антарктиды формировалась
практически с нуля – исключительно за счёт расселения растений с других материков
– с помощью ветра и птиц. Основным источником поступления новых видов-поселенцев
стала Южная Америка: основной путь сезонных миграций птиц пересекал пролив Дрейка.
Попадая в новые места обитания с суровыми природными условиями, некоторые виды
выживали и давали начало новой антарктической флоре. Среди растений неоценовой
Антарктиды встречаются виды, приспособленные к распространению их семян многочисленными
местными птицами. К ним принадлежит земляника антарктическая – небольшое травянистое
растение, которое широко распространено на материке и произрастает вдоль всего
побережья Антарктики от песчаных пляжей до горных районов, достигая километровой
высоты над уровнем моря. Болотный подвид антарктической земляники обитает в
заболоченных местностях, в том числе вблизи края ледникового щита.
В отличие от многих других растений неоценовой Антарктики, антарктическая земляника
приживалась на континенте долго – несколько миллионов лет. В течение первых
миллионов лет неоцена земляника была редким видом в Антарктиде, который полностью
зависел от миграций птиц. Перед началом миграции птицы поедали ягоды земляники
на Огненной Земле и переносили с помётом или на лапах семена растения на антарктическую
почву. Здесь она прорастала, временно приживалась и начинала плодоношение, но
с приходом холодной зимы растение полностью вымерзало, и его популяция поддерживалась
только мигрирующими птицами. Лишь спустя несколько миллионов лет, с постепенным
потеплением климата, образовалась устойчивая популяция, и на континенте постепенно
появился вид местной антарктической земляники, которая оказалась способной переживать
холодную зиму.
Антарктическая земляника – многолетнее двудомное травянистое растение, характеризующееся
засухоустойчивостью и холодостойкостью цветков и завязей. Это розеточное растение
с укороченным многолетним стеблем и мочковатыми корнями, уходящими неглубоко
в почву. Всё растение густо опушено многочисленными белыми волосками – стебель,
черешки листьев и цветоносы, а также нижняя сторона листьев. Листья тройчатые,
с верхней стороны синевато-зелёные, листочки округлые со слегка зубчатым краем.
Листва этого вида земляники не отмирает и не сгнивает осенью, а сохнет на корню
и при этом скручивается в шар, защищая розетку молодых листьев и точку роста
в условиях холодной зимы. Весной при таянии снега прошлогодние листья намокают,
и шар раскрывается, подставляя точку роста под солнечный свет. При этом начинается
активный рост растения. Второй тип побегов растения – ползучие столоны, на которых
образуются молодые растения. За сезон вегетации взрослое растение даёт 2-4 столона
и образует до 5 дочерних растений.
Цветение начинается вскоре после формирования розетки из 3-4 полноценных листьев.
Цветки белые, на коротких цветоножках – не выступают над листьями. Плоды – характерные
многоорешки на сочном разросшемся цветоложе красного цвета, очень крупные (от
2 до 4 см в диаметре), с бугристой поверхностью. Плотные «ягоды» хорошо отделяются
от чашечки, по окраске варьируют от тускло-красных до жёлтых и белых, отличаются
сочной мякотью и сладким вкусом.
Цветы опыляются жуками, мухами, комарами, местными бабочками и одиночными осами,
и даже птицами мышевидками,
которые склёвывают на них насекомых. Ягодами питаются многие виды местных жуков
и птиц. Сок с надклёванной птицей ягоды могут пить мухи, комары, бабочки и пчелы.
Растение начинает цвести и плодоносить на третьем-четвёртом году жизни. Продолжительность
жизни – до 10 лет. Старое растение менее активно плодоносит и, как правило,
отмирает осенью, дав начало дочерним растениям.
Этот вид растений описал Wovoka, участник форума.
Шиповика
или Аидовы тернии (Rubus thornus-aidii)
Порядок: Розоцветные (Rosаles)
Семейство: Розовые (Rosaceae)
Места обитания: субтропические леса предгорий Кавказского хребта, Малая Азия,
Балканы.
Бурный рост теплолюбивых широколиственных лесов по берегам Четвероморья привёл
к появлению необычного облика у потомков растений, бывших обычными спутниками
человека в голоцене. Среди таких удачливых видов – обыкновенная ежевика (Rubus
fruticosus), процветавшая в культурных ландшафтах. Десятки миллионов лет эволюции
в дикой природе сформировали у одного из её потомков поистине демонический облик.
Этот потомок ежевики – шиповика, или аидовы тернии, представляющая собой гигантский
многолетний кустарник. Обычно она образует обширные заросли, занимающие большую
площадь. Побеги этого вида хлыстообразной формы, достигают длины до 10 метров
при толщине до 5 сантиметров. Они отрастают от ползучего подземного корневища
и в процессе роста дугообразно склоняются вершинами к земле. Часто старые побеги
ложатся верхушкой на землю и укореняются, давая начало новым побегам. В процессе
роста побеги ветвятся, и на концах ветвей вызревают гроздья очень сладких ягод
тёмно-фиолетового цвета, которые привлекают многочисленных животных. Птицы являются
основными распространителями семян этого вида.
Старые побеги одревесневают, но древесина этого растения ломкая и рыхлая, а
стволы пустотелые. Кора светло-коричневого цвета, отслаивается тонкими бумагоподобными
пластинками. Побеги шиповики покрыты многочисленными загнутыми шипами длиной
до 3 сантиметров. На самой старой части побега шипы со временем отваливаются
вместе с шелушащейся корой. Шипиками также покрыта и нижняя сторона листьев
вдоль главных жилок. Листья плотные, тёмно-зелёные сверху и серебристые из-за
опушения снизу, состоят из трёх листочков примерно одинакового размера и имеют
треугольные очертания. Их края зубчатые, а поверхность ребристая. Шиповика образует
такие густые заросли, что некоторые звери, по неосторожности проникнув в гущу
её зарослей в поисках ягод, попадают в смертельную ловушку. Молодые неодревесневшие
побеги вооружены прочными шипами, гибкие и прочные на разрыв. Они легко способны
зацепить и удержать неосторожное животное. Как правило, паникующее животное,
пытаясь освободиться, бросается в стороны и наносит себе ранения шипами больших
старых побегов и может погибнуть от потери крови. В глубине кустов очень часто
можно увидеть множество скелетов и полуразложившихся трупов животных, среди
которых немало останков детёнышей.
Плодоносят аидовы тернии почти весь вегетационный сезон – с мая по ноябрь. Почти
всё это время растение цветёт, являясь важным кормовым ресурсом для нектароядных
насекомых. Цветки этого растения белые, пятилепестные, собраны в кисти по несколько
десятков. Зимой на севере ареала и в горных районах растение сбрасывает листву
полностью, в тёплых регионах на юге ареала – лишь часть самых старых листьев.
В это время оно находится в состоянии относительного покоя, и на побегах закладываются
новые цветочные почки. Возраст многих крупных кустов достигает полувека, хотя
старые стволы существуют не более 6-8 лет.
Этот вид растений открыл Моргот, участник форума.
Маликаат
(Aromadryas malikaat)
Порядок: Розоцветные (Rosales)
Семейство: Розовые (Rosaceae)
Место обитания: североамериканская лесотундра и тундра.
Коротким полярным летом североамериканская тундра в неоцене буквально расцветает.
Несмотря на бедность видового разнообразия, тундровая растительность поражает
своим количеством. Едва стаял снег, как начинают раскрываться первые цветы,
а к мменту наступления устойчивой тёплой погоды тундра уже в цвету. Рассыпают
пыльцу заросли неарктической сосны, розоватые
цветки украшают зелёные заросли квижуктаата,
а между ними растут растения поменьше – например полярная камнеломка аупиллатингват
и ползучий кустарник кинникинник.
Одно из самых красочных цветковых растений тундры – это невысокий маликаат,
раскрывающий свои крупные белые цветки.
Маликаат является потомком дриады восьмилепестной (Dryas octopetala) – невысокого
травянистого растения эпохи голоцена. Родство этих видов показывают его цветки
– подобно цветам его предка, они большие (диаметром до 8 сантиметров), белые,
с жёлтой сердцевиной, которую образуют многочисленные тычинки и пестики. Окраской
они немного напоминают цветок ромашки, только более крупный: растение маликаата
достигает в среднем 15-20 см в высоту. Однако это не корзинка, как у ромашки,
а одиночный цветок: маликаат принадлежит к совершенно иному семейству растений.
На стеблях растения долгое время сохраняются остатки листовых черешков. Они
высыхают и служат импровизированными шипами, отчасти защищая растение от крупных
травоядных млекопитающих. Но листьями маликаата кормятся личинки разнообразных
местных бабочек. Зимой зелень этого растения поедают под снегом грызуны.
Маликаат – вечнозелёное морозостойкое растение, не страдает от полярных холодов,
как его соседи вроде кинникинника. В то же время этот вид размножается только
семенами. В отличие от своего предка, семена которого разносились ветром, маликаат
эволюционировал в сторону зоохории. Семена быстро теряют при созревании пушистые
выросты, но погружены в разрастающееся цветоложе, которое обладает приятным
запахом, привлекающим травоядных млекопитающих (отсюда родовое название, означающее
«ароматная дриада»), которые объедают растение, собирая с него семена. Пройдя
через пищеварительный тракт животного, семена выпадают вместе с навозом и благодаря
обработке желудочным соком быстрее трогаются в рост следующей весной. Из-за
этой особенности расселения маликаат обычно растёт в строго определённых местах
– вдоль троп миграций косорогов
и подобных зверей. Во время цветения этого вида тропы животных становятся особенно
заметными на фоне пейзажа тундры. Поскольку растение не образует длинных ползучих
побегов и расселяется исключительно на стадии семян, его популяция меньше, чем
популяции других полярных растений. Однако это компенсируется стойкостью растения
к вытаптыванию и значительным долголетием – отдельный куст может расти до 40-60
лет.
Этот вид растений открыл Bhut, участник форума.
Лихеноморфа
жизнестойкая (Lichenomorpha vivax)
Порядок: Мальпигиецветные (Malpighiales)
Семейство: Подостемовые (Podostemaceae)
Место обитания: западный Мадагаскар, сезонные проточные водоёмы.
Одними из самых специализированных водных растений являются подостемовые – своеобразные
водные травы, у которых значительная часть тела представлена таллусом. Это уплощённое
образование, напоминающее слоевище водорослей, составляет значительную часть
массы растения и прочно прикрепляется к субстрату. Подостемовые – теплолюбивые
растения, распространённые главным образом в тропиках обоих полушарий.
В западной части Мадагаскара пресные водоёмы отличаются непостоянством и пересыхают
на срок от нескольких недель до нескольких месяцев. Обитающие в них водные растения
в процессе эволюции выработали способность переживать засуху. Этими особенностями
обладают некоторые представители подостемовых, освоившие водоёмы такого типа.
Один из обычных видов в западной части острова – лихеноморфа жизнестойкая, растение,
способное переносить значительное осушение. Это сравнительно крупный вид: длина
таллуса взрослых экземпляров достигает 30-40 см. Таллус плотно прикреплён к
субстрату (камням) и покрыт толстой кожицей, а при высыхании водоёма на покровах
растения выделяется восковой налёт, благодаря чему этот вид способен переносить
нахождение на воздухе в течение нескольких месяцев. В сухой сезон лихеноморфа
жизнестойкая существует в виде сухой корочки на камнях, напоминающей лишайники
– отсюда название вида. Таллус имеет неправильно-листовидную форму и обычно
разделён на несколько нечётко отделённых друг от друга лопастей с волнисто изрезанным
краем. В сухой сезон края подсыхают и крошатся, но под водой быстро отрастают
и восстанавливают форму. Окраска таллуса в состоянии роста ярко-зелёная, на
ярком солнце края краснеют.
В тканях таллуса закладывается большое количество почек – вегетативных и цветочных.
В сезон дождей таллус быстро оживает, в течение нескольких дней пробуждаются
почки и разрастаются вегетативные побеги, похожие на слоевища водорослей – плоские,
полупрозрачные, сильно расширенные, листовидной формы с заострённым кончиком.
Первые побеги цельные, более поздние раздвоены 1-2 раза. Растение образует несколько
десятков вегетативных побегов. В ходе вегетации в таллусе накапливается значительное
количество крахмала и белков.
В конце влажного сезона, когда уровень воды падает, вегетативные побеги оказываются
на поверхности воды и начинают подсыхать. Это сигнал для начала цветения, и
растение выпускает множество мелких цветков с редуцированным околоцветником.
Высота цветоносов не превышает 3-4 см. Растение опыляется ветром, реже самоопыляется.
Семена покрыты липкой слизистой оболочкой. Они прилипают к ногам водных птиц
и переносятся ими в другие водоёмы. Молодое растение в первые 2-3 года жизни
развивает только вегетативные побеги и наращивает массу таллуса, после чего
начинает ежегодно цвести. Продолжительность жизни достигает 20 лет и более.
Наядины
волосы (Hydrocapillatura najadalis)
Порядок: Мальпигиецветные (Malpighiales)
Семейство: Подостемовые (Podostemaceae)
Место обитания: горные реки Южной Европы, на южных склонах Альп.
Изменения климата в эпоху неоцена изменили ареалы различных групп растений,
позволив теплолюбивым видам расселиться в более высокие широты. Специализированное
семейство водных растений подостемовые в неоцене значительно расширило ареал,
и его представители стали характерными водными растениями проточных водоёмов
Европы. Некоторые виды этих растений расселились даже в высокогорные районы
– очевидно, следуя маршрутами миграций водяных птиц, разносивших семена на лапах
и оперении. В новых местах обитания за миллионы лет формировались новые виды
этих растений. Одним из таких видов является альпийское растение наядины волосы.
Этот вид – типичный представитель подостемовых; значительная часть тела растения
превращена в ползучий таллус, прочно прикрепляющийся к субстрату. Ветви таллуса
напоминают плоские извилистые ленты со слегка волнистыми краями. На верхней
стороне таллуса формируются многочисленные вегетативные и цветочные почки. Растение
обладает явно выраженной сезонностью вегетации, связанной с уровнем воды в реках.
Этот вид встречается на прибрежных мелководьях, на глубине не больше 20-30 см.
Основной период вегетации – зима и весна, когда реки более полноводны за счёт
таяния ледников. В это время развиваются вегетативные почки и растение выпускает
длинные лентовидные вегетативные побеги, стелющиеся по течению и развевающиеся
в потоке воды, напоминая волосы зелёного цвета (отсюда название). Длина таких
побегов достигает 20-50 см при ширине не более 3 мм. В это время растение активно
фотосинтезирует и накапливает в таллусе питательные вещества, необходимые для
успешного цветения.
Летом, когда уровень воды в реках снижается и течение замедляется, часть таллусов
оказывается над водой. В это время на них начинают развиваться цветочные почки
и растение обильно цветёт. Цветки очень мелкие, с крайне редуцированным околоцветником,
на тонких ножках, бледно-розового цвета. Многочисленные одиночные цветки собраны
в пучки, торчащие на небольших бугорках над поверхностью таллуса. Опыление осуществляется
ветром или случайными насекомыми. Некоторое количество цветков поедают улитки
и жуки-водолюбы.
Мелкие семена покрыты липкой слизью и легко приклеиваются к оперению, ногам
и клювам водяных птиц, которые осуществляют расселение растения. Первые два
года проросток формирует таллус и захватывает жизненное пространство, на третьем
году жизни формирует несколько вегетативных побегов. Цветение молодого растения
происходит на 4-м году жизни.
Потамоматрикс
сахарский (Potamomatrix saharensis)
Порядок: Мальпигиецветные (Malpighiales)
Семейство: Подостемовые (Podostemaceae)
Место обитания: Африка, бассейн Сахарского Нила, сезонные водоёмы и пересыхающие
участки русла реки.
Подостемовые являются глубоко специализированными водными травами, и в процессе
эволюции претерпели глубокие изменения в строении по сравнению с наземными растениями.
В большинстве своём они населяют непересыхающие водоёмы, однако некоторые виды
смогли освоить местообитания, не слишком подходящие для жизни специализированных
водных растений. Одним из таких растений является потамоматрикс сахарский, населяющий
долину Сахарского Нила.
Это нетипичный вид подостемовых, приспособившийся к жизни в медленно текущей
воде и в пересыхающих реках. В процессе эволюции это растение приспособилось
выживать в пересыхающем водоёме, несмотря на специализацию к жизни под водой.
Таллус этого вида корковидный, развивается на твёрдом субстрате – обычно на
камне или затонувшем дереве – и плотно прикрепляется к нему. От него отрастают
многочисленные тонкие фотосинтезирующие побеги – лентовидные, полупрозрачные,
без листьев. Они достигают длины 30-80 см, и стелются по поверхности воды. В
тени они зелёного цвета, на ярко освещённых местах приобретают розоватый цвет.
В сезон дождей среди них быстро отрастают упругие цветоносы, возвышающиеся над
поверхностью воды и несущие скромные колосовидные соцветия. Семена очень мелкие,
липкие, распространяются на ногах и оперении околоводных птиц.
При пересыхании водоёма цветоносы и фотосинтезирующие побеги отмирают, и таллус
образует выводковые почки, покрытые многослойной оболочкой и способные переносить
высыхание. Покрывающая их слизь застывает в виде корочки, предохраняющей растение
от высыхания и гибели. Выводковые почки легко отрываются от таллуса и распространяются
ветром и птицами. При смачивании водой слизь быстро набухает и приклеивает почку
к субстрату. Почка задерживается в росте на несколько дней, одновременно впитывая
воду. Если водоём не пересыхает, и снабжение водой не прерывается, это становится
сигналом к продолжению роста. Во время разливов реки потамоматрикс быстро образует
новые побеги и активно разрастается. Молодое растение обычно зацветает на следующий
год, а при размножении семенами – на третьем году жизни. Продолжительность жизни
растения составляет до 5-6 лет.
Дендрофуксия,
фуксиевое дерево (Dendrofuchsia penduliflora)
Порядок: Миртоцветные (Myrtales)
Семейство: Кипрейные (Onagraceae)
Место обитания: умеренные области Южной Америки, северная часть Огненной Земли.

Рисунок Fanboyphilosopher (растение опыляется колибри тайином)
Растения рода фуксия (Fuchsia) в эпоху человека были характерным
компонентом флоры умеренно тёплых и субтропических областей Южной Америки. Они
представляли собой кустарники с характерными висячими цветками трубчатой формы,
которые опылялись бабочками и птицами. В неоцене один из видов этого рода сделал
удачную попытку освоить новую жизненную форму, и превратился в небольшое дерево.
Тем не менее, эта жизненная форма довольно изменчива: на севере ареала растение
представляет собой полулистопадное дерево высотой до 8 метров, но на юге ареала
резко уменьшается в росте и превращается вначале в многоствольный кустарник,
а затем в листопадную стелющуюся форму с ползучим стволом длиной около 10 метров,
которая зимует под снегом. Это дендрофуксия, или фуксиевое дерево.
Листья этого растения кожистые, тёмно-зелёные, яйцевидной формы с притупленным
кончиком. Поверхность листа между крупными жилками слегка вздутая. На севере
ареала в начале зимы опадают только старые листья, а на юге ареала растение
полностью теряет листву на зиму. Если молодые побеги подмерзают, растение быстро
восстанавливается из спящих почек. Из-за этого на юге ареала растение обычно
имеет густо разветвлённые побеги. На севере ареала побеги дендрофуксии длинные
и слегка повисшие. Древесина этого растения рыхлая и довольно хрупкая. На севере
ареала, где растения сохраняют листву на зиму, во время внезапных снегопадов
ствол дендрофуксии может не выдержать нагрузки, и разламывается. Но растение
не гибнет – его побеги легко укореняются, и растение продолжает развиваться
в виде лежачего куста. В первые годы жизни годовой прирост дендрофуксии достигает
1 – 2 метров.
В период цветения дендрофуксия – это настоящее украшение скудных на яркие краски
лесов крайнего юга Южной Америки. Весной, когда температура воздуха повышается,
а дни становятся длиннее, на дереве раскрывается множество цветков. Висячие
цветки трубчатой формы с четырьмя остроконечными чашелистиками и четырьмя округлыми
лепестками очень похожи на цветки обычной фуксии. Они образуются на побегах
прошлого года. На одном цветоносе развивается кисть из трёх-четырёх цветков.
Цветы изменчивы по окраске, но всегда окрашены в оттенки красного цвета, который
хорошо различают их опылители – птицы. В наиболее типичном случае чашечка цветка
тёмно-красная, а лепестки карминно-розовые. В период цветения дерево полностью
покрыто цветками, и листва с трудом различается среди массы цветов. Цветущую
дендрофуксию легко увидеть издалека – красная масса цветков выделяется на фоне
окружающих кустарников и листвы нотофагуса – доминирующего рода растений крайнего
юга Южной Америки. Это растение опыляется местными бабочками и перелётным колибри
тайином. Дендрофуксия опыляется
исключительно перекрёстно.
Плоды этого растения ягодообразные, сочные, с тонкой кожицей и мелкими семенами.
Их распространяют млекопитающие и птицы, питающиеся плодами. Продолжительность
жизни дендрофуксии не превышает 60 – 80 лет.
Фуксия
подснежная (Fuchsia subnivalis)
Порядок: Миртоцветные (Myrtales)
Семейство: Кипрейные (Onagraceae)
Место обитания: крайний юг Южной Америки, Антарктида (Антарктический полуостров),
открытые пространства, каменистые пустоши.
В эпоху человека различные виды и гибриды фуксий были популярными комнатными
растениями – быстрорастущими, обильно цветущими и вечнозелёными. В некоторых
районах Земли они какое-то время выживали после исчезновения человека, более
или менее успешно отвоёвывая себе место в экосистемах раннего неоцена. У себя
на родине, в Южной Америке, фуксии активно эволюционировали, приспосабливаясь
к меняющимся условиям на рубеже голоцена и неоцена. Некоторые их виды приспособились
к экстремальным условиям высокогорья и высоких широт с выраженным сезонным климатом
и снежной зимой. Одним из таких видов-экстремалов стала фуксия подснежная, которая
смогла приспособиться к холодному климату, преодолеть пролив Дрейка и заселить
Антарктиду.
Это растение – полукустарниковая стланцевая фуксия с одревесневающими многолетними
побегами, которые укореняются в почве между камнями и дают обильную поросль.
Длина побегов может достигать 150-200 см, но высота растения не превышает 6-10
см. Этот вид – вечнозелёное растение с очень мелкими округлыми листьями с коротким
черешком. Поверхность листа кожистая, гладкая, края мелкозубчатые. Листва накапливает
много гликозидов, поэтому на вкус она очень горькая и несъедобная для местных
травоядных птиц – луговых лебедей.
Растение успешно зимует под снегом, часто вместе с цветками.
Растение цветёт почти весь вегетационный сезон, но особенно обильно – весной,
после установления тёплой погоды и таяния снега. Её цветки очень яркие – лепестки
красные с белыми прожилками, чашелистики зелёные с красными пятнами и красные
изнутри. Прожилки отражают ультрафиолетовые лучи, и это гарантированно привлекает
редких местных бабочек, являющихся опылителями растения. Цветки поникшие, на
вертикальных цветоносах, поднимающихся на 9-12 см над землёй. Такое расположение
цветков привлекает также антарктических птиц-мышей,
которые вылизывают её цветки, стоя на земле. После опыления образуются ярко-красные
ягоды, которые становятся лакомством для местных птиц. Мякоть содержит множество
мелких семян, которые часто прилипают к ногам и оперению птиц, благодаря чему
растение распространяется даже на изолированные острова, где при отсутствии
опылителей размножается вегетативно. Молодое растение зацветает в возрасте 5-6
лет, продолжительность жизни до 80 лет.
Иван-чай
сибирский (Chamerion sibiricus)
Порядок: Миртоцветные (Myrtales)
Семейство: Кипрейные (Onagraceae)
Место обитания: Сибирь, участки лесов с нарушенным древостоем, «дороги обды».
В эпоху человека иван-чай узколистный (Chamerion angustifolium) был одним из
самых обычных видов в нарушенных человеком местообитаниях, где процветал, достигая
высокой численности. Благодаря способности расселяться и расти на бедных почвах
это растение сохранило высокую численность популяций в эпоху биологического
кризиса, и после стабилизации условий в раннем неоцене стало родоначальником
нескольких видов, обитающих в Северном полушарии. Один из потомков этого вида
– иван-чай сибирский, обитающий в биотопах, складывающихся в результате деятельности
местной мегафауны, нарушающей сплошные заросли деревьев.
Это довольно высокое травянистое растение, достигающее 50-150 см в высоту: на
влажной почве оно разрастается сильнее, образуя куст из десяти и больше стеблей.
Оно обычно растёт в сообществах травянистых растений вдоль «дорог обды», но
иногда появляется и вдали от них, на участках нарушенного древостоя. Стебель
растения прочный, четырёхгранный на срезе, красноватого цвета, особенно у основания.
Листья узкие, особенно отрастающие в конце и в начале сезона вегетации. Окраска
листа сверху ярко-зелёная, снизу более светлая. Это растение служит пищей обде,
местным видам зайцелоп,
шурге, разным грызунам и прочим
травоядным животным, а также насекомым. После поедания млекопитающими растение
относительно быстро восстанавливается, и успевает отцвести и дать семена, но
цветёт менее обильно.
Цветы растут густыми цилиндрическими кистями на верхушках стеблей и окрашены
в красный или розовый цвет; в разгар вегетации цвет лепестков более насыщенный.
Цветки четырёхлепестные, с округлыми лепестками. Этот вид опыляется разными
насекомыми, в основном двукрылыми и перепончатокрылыми, реже бабочками. В конце
лета созревают плоды - сухие коробочки, где находятся маленькие семена с длинным
шелковистым «парашютом», позволяющим им разлетаться. Оболочки семян довольно
прочные, легко могут пережить путешествие по пищеварительной системе крупных
травоядных зверей и суровые сибирские зимы. Иногда созревшие семена разносятся
местными травоядными – съев растение вместе с семенами, животное распространяет
его семена с навозом. Весной семена прорастают и довольно быстро достигают высоты
полуметра и больше. Обычно цветение начинается на втором году жизни.
Растение предпочитает хорошее освещение с притенением в жаркие полуденные часы.
На зиму иван-чай сибирский засыхает, но его сухие стебли сохраняются долго и
служат частью зимнего рациона травоядных. В Евразии растут родственные виды
(возможно, всего лишь местные формы), которые отличаются цветов цветков: белый,
жёлтый, пастельный и т.д.
Этот вид растений открыл Bhut, участник форума.
Иван-чай
выносливый (Chamerion endurabilis)
Порядок: Миртоцветные (Myrtales)
Семейство: Кипрейные (Onagraceae)
Место обитания: открытые местности – североамериканские прерии, тундра.
Одна из главных особенностей разных видов иван-чая (Chamerion) – скорость колонизации
этим растением открытых пространств, а также нетребовательность к климатическим
условиям и к среде обитания. Это помогало иван-чаю узколистному (Chamerion angustifolius)
пережить и эпоху антропогенного прессинга голоцена, и ледниковый период начала
неоцена. А в тёплое послеледниковое время растение продолжило процветать, и
его потомки заняли разнообразные экологические ниши в природе Северного полушария.
Иван-чай выносливый является типичным представителем флоры североамериканских
открытых пространств. Это высокое травянистое растение, обычно 100-150 см в
высоту, с тонкими стройными стеблями, узкими листьями и большими кистями розовых
или тёмно-розовых цветков на верхушке стебля. На севере своего ареала иван-чай
выносливый цветёт во второй половине полярного лета, добавляя красок короткому
полярному лету, а на юге, в более тёплом климате – во второй половине лета и
в начале осени. Это делает его очень важным источником корма для многих североамериканских
насекомых, которые активно опыляют его цветы перед зимними холодами. Часто этим
занимаются даже мигрирующие колибри, отправляющиеся на зимовку. Осеннее цветение
связано с засушливым летом, когда растение приостанавливает рост и экономит
влагу. В конце лета растение буквально «вспыхивает» яркими красками и отцветает
полностью за две недели. Крупные травоядные животные вроде азиногиппусов
и даже пекари-бизонов охотно
поедают этот вид – это растение произрастает на открытых пространствах большими
зарослями и является обильным источником корма.
Иван-чай выносливый размножается семенами, которые разносятся ветром; представители
этого вида, растущие вместе на небольшой территории, цветут и выпускают свои
семена очень синхронно. Семена очень выносливы, успешно перезимовывают на новом
месте под снегом, а весной начинают бурно расти. Во второй половине лета они
нередко уже взрослые и цветут. В тундре молодое растение набирает полную силу
и цветёт лишь на третьем году жизни, и цветение начинается раньше, чем на юге
ареала.
Этот вид – многолетнее растение, подземная часть с мясистыми корнями позволяет
быстро восстанавливаться после поедания травоядными животными. Продолжительность
существования зарослей – не менее двадцати лет, в тундре до 60 лет и больше.
Этот вид растений открыл Bhut, участник форума.
Приречный
ложный миндаль (Neoterminalia riparia)
Порядок: Миртоцветные (Myrtales)
Семейство: Комбретовые (Combretaceae)
Место обитания: галерейные леса в долинах рек в центральной и северо-восточной
части Южной Америки.
В голоцене человек перевозил с материка на материк множество видов животных
и растений в качестве украшения. Одним из таких интродуцированных видов был
индийский миндаль (Terminalia catappa). В середине 20-го века человеческой эры
это дерево широко использовалось в городском ландшафтном дизайне в Бразилии,
поскольку было редким тропическим листопадным деревом, и его опавшие листья
придавали улице «европейский» колорит. Этот вид очень хорошо приспособился к
новым местам обитания, и в неоцене дал начало роду Neoterminalia, типичным представителем
которого является приречный ложный миндаль. Совместно с лиловым
дудчатым деревом (Tabebuia violacea) и пальмой
дендем (Elaeis dendem) это дерево образует галерейные леса, произрастающие
вблизи русел рек, и распространено только в этих прибрежных районах.
Приречный ложный миндаль вырастает до 30 м в высоту, у дерева прямостоячая симметричная
крона и горизонтальные ветви. По мере взросления дерева его крона становится
более уплощённой и раскидистой, в форме вазы. Его ветви расположены отчётливо
выраженными ярусами. Листья крупные, 23 см длиной и 12 см шириной, яйцевидные,
глянцевые тёмно-зеленые, кожистые. Они опадают в сухой сезон, и перед опадением
приобретают жёлто-бурый цвет.
Деревья однодомные, с явно отличающимися друг от друга мужскими и женскими цветками
на одном дереве. И те, и другие диаметром 3 см, беловато-розовые, ароматные,
без лепестков; они образуются на пазушных или концевых колосках. Плод представляет
собой костянку длиной 6 см, вначале зелёную, а при созревании красную, внутри
которой находится одно семя. Плоды распространяются водой и животными. По сравнению
с плодами предкового вида, плоды более мясистые; таким образом, животное поедает
околоплодник, не трогая семя. Цветы опыляются насекомыми.
Другим видом этого рода является прибрежный
ложный миндаль (Neoterminalia atlantica). Этот вид, более крупный,
чем приречный ложный миндаль, вырастает до 40 м в высоту и входит в состав атлантических
лесов юго-востока Южной Америки. Его плоды длиной 5 см при созревании становятся
жёлтыми, а цветки белые и безлепестные. В засушливый сезон этот вид теряет меньше
листьев, чем приречный вид.
Этот вид растений открыл Жуан Витор Кутиньо, Бразилия.
Паранепентес
обезоруженный (Paranepenthes dearmatus)
Порядок: Непентовые (Nepenthales)
Семейство: Непентовые (Nepenthaceae)
Место обитания: Юго-Восточная Азия, Берег Джакарта, Земля Сунда; полог дождевого
леса.
Среди ветвей деревьев и побегов разнообразных лиан тропического дождевого леса
Юго-Восточной Азии встречаются различные виды плотоядных растений семейства
Непентовых – собственно непентес, и его родственник цветолист,
имеющий более специализированные ловчие листья, имитирующие цветки некоторых
растений. Но по соседству с этими растениями обитает их родственник, который,
в противоположность этим растениям, развил совершенно иную форму отношений с
мелкими обитателями полога леса. Это эпифитное растение называется паранепентес
обезоруженный. У него длинные ползучие стебли, покрытые воздушными корнями.
Он прикрепляется к зарослям мха и ползёт по коре, плотно прикрепившись к покрывающему
её слою мха очень короткими корнями-присосками. Стебель этого растения достигает
длины 3 – 5 метров и больше.
Листья паранепентеса имеют форму, характерную для представителей семейства:
они крупные, кувшинообразные, а их объём достигает двух литров. Края листьев
окрашены тускло, а крышечка широкая, с длинными выростами по краям. Черешки
листьев, так же, как у настоящего непентеса, взяли на себя роль фотосинтезирующих
органов – они прочные, листообразно расширенные, округлых очертаний.
«Стандартные» представители семейства используют кувшинообразные листья как
охотничьи снасти, но паранепентес не имеет нектарников и пищеварительных желез
на внутренней стороне листа-кувшинчика (отсюда название). В листьях просто накапливается
дождевая вода, и это очень привлекает разнообразных лазающих животных. Растение
вступило с ними в симбиоз, и использует их помёт и выделения как удобрение,
получая из них нужные для роста минеральные и органические вещества. Лазающие
лягушки и крабы, обитающие в пологе леса, используют эти листья как постоянное
укрытие. Кроме того, в листьях паранепентеса откладывают яйца различные комары.
Для того, чтобы использовать выделяемые «постояльцами» вещества, в нижней части
кувшинчиков у паранепентеса развиваются ворсинки, с помощью которых растение
всасывает выделения животных. Иногда в листья спускаются воздушные корни с вышележащих
побегов.
Взрослое растение паранепентеса обезоруженного несёт до 20 – 25 нормально развитых
листьев и превращается в «многоквартирный дом» для различных древесных лягушек
и других влаголюбивых животных.
Поскольку растение «не интересуется» насекомыми как источником пищи, оно приспособилось
к опылению бабочками и перепончатокрылыми. Его 4-лепестковые цветки диаметром
около 3 см собраны в большое колосовидное соцветие. Они окрашены в ярко-розовый
цвет и испускают сладковатый запах.
Аупиллатингват
(Saxifraga aupillatinguat)
Порядок: Камнеломкоцветные (Saxifragales)
Семейство: Камнеломковые (Saxifragaceae)
Место обитания: тундра Северной Америки, каменистые участки.
Ледники оказали значительное влияние на рельеф местности в арктических широтах
Евразии и Северной Америки. Распространяясь на юг, они сгребали слои отложений,
а после их таяния на местности оставались участки, практически лишённые гумусного
слоя. Лишь среди камней накапливалось небольшое количество почвы, и оно позволяло
сравнительно сносно существовать видам растений, привыкшим к скудным условиям
жизни в тундре.
В начале полярного лета каменистые местности, созданные ледниками, расцвечиваются
яркими красками. Бледно-розовые колокольчатые цветки покачиваются на концах
побегов квижуктаата, потомка кассиопеи
четырёхгранной (Cassiope tetragona) эпохи голоцена, а неподалёку из щелей между
камнями словно брызжет сиреневато-розовая пена: это зацвело растение аупиллатингват
– потомок камнеломки супротивнолистной (Saxifraga oppositifolia). Аупиллатингват
– эскимосское название камнеломки. Как и многие камнеломки голоцена и неоцена,
аупиллатингват не образует отдельных кустов, как это делает квижуктаат, а стелется
по земле, обильно ветвясь и образуя довольно плотный ковер 5-10 см в высоту.
Его однотонную зелень нарушают, а в период начала цветения полностью скрывают
звездообразные цветы фиолетового цвета с 6-8 крупными лепестками. Они большие
(диаметр до 30 мм) и заметные, в отличие от чешуек, которые аупиллатингвату
заменяют листья, как многим другим полярным растениям. Эти цветы опыляются разными
полярными насекомыми, и аупиллатингват является важным кормовым растением для
насекомых, привлекая их внимание «указателями мёда» – полосками и пятнами. Побеги
этого растения сильно укороченные и утолщённые, напоминают четырёхгранные зелёные
шишечки. Корневая система поверхностная, состоит из многочисленных коротких
корней.
Аупиллатингват не синтезирует смолистых веществ-«антифриза» в тканях, но размеры
растения позволяют ему полностью скрываться под снегом и проводить суровую полярную
зиму в относительном тепле. Крупные травоядные вроде косорогов
способны выкопать его из-под снега, но его защитой служит рост в «неудобных»
местах среди камней, где крупному животному не всегда удаётся его объесть. Разные
мелкие грызуны легко находят и поедают это растение под снегом, но аупиллатингват
очень жизнестоек и способен размножаться не только семенами, но и вегетативно
– прикорневыми отростками и легко укореняющимися побегами. Крупные животные
чаще всего поедают цветки этого растения – стебли бывает слишком сложно откусить
из-за их специфической формы. Семенные коробочки у этого вида подняты на длинных
ножках над уровнем цветов и легко раскрываются, а поверхность семян снабжена
многочисленными ворсинками. Поэтому косороги, когда пытаются съесть цветы, неизбежно
задевают их. При прикосновении семенные коробочки лопаются и цепкие семена прилепляются
к шерсти зверей, чтобы потом упасть где-нибудь в другой части тундры.
Продолжительность жизни аупиллатингвата составляет около 20 лет.
Этот вид растений открыл Bhut, участник форума.
Древесная
толстянка яйцелистная, «яичное дерево» (Dendrocrassulopsis ovifolium)
Порядок: Камнеломковые (Saxifragales)
Семейство: Толстянковые (Crassulaceae)
Места обитания: Юго-Западная Азия, горы Атлас.
Ледниковые периоды характеризуются иссушением климата: огромные массы воды связаны
в леднике, уровень океана снижается, и на месте мелководных морей возникают
участки суши. Испарение воды снижается, и, как следствие, выпадает меньшее количество
дождей. Расширение площади материков приводит к более резкому температурному
контрасту между побережьями и массивами суши вдали от моря. В таких условиях
сокращается численность влаголюбивых растений, зато процветают выносливые виды,
способные запасать воду в своих органах, и переносящие прохладный климат зимой.
В Старом Свете среди таких выносливых растений можно отметить семейство толстянковых,
отличающееся значительным разнообразием. Когда в ледниковый период на месте
Средиземного моря возникла солёная пустыня, на её краях, относительно свободных
от соли, процветали разнообразные толстянки. Позже, когда климат стал более
влажным, некоторые их представители приобрели совсем причудливый облик.
Древесная толстянка – это коренастое суккулентное дерево с ползучим стволом,
легко укореняющимся в почве. Её высота невелика: около 3 – 4 метров, но зато
ствол часто достигает метровой толщины. Корни этого растения поверхностные,
не уходят глубоко в почву, но зато распространяются на большие площади, позволяя
очень быстро собирать дождевую воду даже после самого слабого дождя, едва смочившего
пыль. При засухе часть корней может отмирать, но в сезон дождей они очень быстро
восстанавливаются. Поверхностная корневая система – плохая опора для ствола
растения, поэтому ветра часто валят массивное растение на землю. Однако это
не причиняет растению неудобств: поваленные побеги быстро дают корни, и дерево
продолжает жить.
Кора древесной толстянки очень толстая и рыхлая – она служит для термоизоляции,
предохраняет ствол дерева от солнечных ожогов и перегрева.
Листья этого растения толстые, удлинённо-шаровидной формы, покрыты тонким восковым
налётом, собраны на вершинах побегов. Их длина – около 5 см при толщине до 4
см. В листьях, поднятых над землёй и недоступных для большинства травоядных,
сконцентрирована значительная часть запаса влаги, накопленной этим растением.
Кроме того, каждый лист – залог своеобразного «бессмертия» растения: как и все
толстянки, это растение может размножаться укоренением отдельных листьев.
Живёт это своеобразное растение недолго: не более 10 лет, давая за это время
значительную биомассу за счёт укоренившихся на земле листьев и разросшихся в
разные стороны ветвей.
Древесная толстянка относится к монокарпическим цветковым растениям, то есть,
цветёт каждое растение этого вида лишь один раз в жизни, и после цветения быстро
отмирает. Цветение начинается в конце сухого сезона. На каждом побеге раскрываются
в большом количестве невзрачные белые цветки, испускающие сильный аромат. На
них слетаются в основном мухи и жуки, обитающие в пустыне. Цветение продолжается
долго: около месяца. За это время начинается необратимое истощение растения:
листья спадаются, становятся похожими на сморщенные мешочки, и в конце концов
отсыхают. Ствол растения заметно «худеет», на нём появляются продольные борозды,
а кора отваливается крупными кусками. Ко времени созревания плодов дерево совершенно
истощается, и в течение влажного сезона полностью сгнивает. Зато семена имеют
прекрасные шансы выжить: в течение сезона дождей они успевают прорасти и молодое
растение может накопить достаточно влаги для пережидания засухи.
Шерстяная
капуста (Platyaeonium lanatum)
Порядок: Камнеломковые (Saxifragales)
Семейство: Толстянковые (Crassulaceae)
Место обитания: Земля Зиндж, горы Усамбара.
В эпоху неоцена раскол Африки по линии Великой Рифтовой долины привёл к тому,
что все высочайшие вершины материка оказались на Восточно-Африканском субконтиненте
– Земле Зиндж. В некотором смысле высочайшие горные вершины подобны островам
в море: это островки, на которых складываются особые природные условия, окружённые
обширными территориями, где условия жизни совершенно иные. Изоляция этих территорий
приводит к формированию на горных вершинах тропических широт совершенно особой
флоры и фауны.
В горах Узамбара, в верхней части пояса облачных лесов, встречается особый вид
многолетних травянистых растений своеобразного облика – шерстяная капуста. Внешне
это растение действительно напоминает лежащую на земле кучу кочанов капусты
с листьями, покрытыми многочисленными белыми волосками. При ближайшем рассмотрении
оказывается, что это растение не имеет никакого отношения к капусте и лишь обладает
отдалённым внешним сходством с этим видом. Шерстяная капуста является потомком
одного из видов рода эониум (Aeonium), широко распространённого в Африке в эпоху
человека.
Приспособление к жизни в горных районах вызвало значительные изменения в облике
растения. Предки этого вида были кустистыми растениями с плоскими розетками
листьев на кончиках ветвей. Жизнь в горах превратила шерстяную капусту в ползучее
растение, стебель которого стелется по земле и обильно ветвится, укореняясь
по всей длине. Длина стебля может достигать 2 метров. Розетки листьев у этого
вида крупные – до 30 см в диаметре. Они располагаются на поверхности земли достаточно
плотно, образуя подушковидные заросли, и между ними могут расти лишь немногие
растения иных видов. Листья имеют ложковидную форму с расширенной вершиной,
загибаются к центру розетки, прикрывая точку роста. Нижняя сторона и край листа
обильно опушены белыми волосками, верхняя сторона покрыта воском – это защита
от интенсивного ультрафиолетового излучения. Пластинка листа суккулентная, имеет
ширину до 10 см и толщину около 1 см, хрящеватую консистенцию.
Стебель растения в период роста несёт небольшие языковидные листья, которые
быстро отмирают. Выбравшись из тени уже развившихся розеток, кончик стебля укореняется
и начинает формировать новую розетку. В первый год она достигает диаметра около
10 см, на третий год приобретает максимальный диаметр и начинает накопление
питательных веществ, необходимых для цветения. На пятом-шестом году существования
розетка зацветает.
Цветение шерстяной капусты происходит очень бурно. У основания каждой развитой
розетки быстро образуется стебель, достигающий двухметровой высоты. Цветки этого
вида имеют звёздчатую форму и узкие белые лепестки, собраны в крупную сложную
кисть конической формы. Гроздья цветков издают медовый запах и обильно выделяют
нектар, привлекающий летучих мышей и бабочек – основных опылителей этого растения.
Семена мелкие, снабжены волосками и разносятся ветром. После цветения розетки
растения отмирают, и от целого растения остаются лишь самые молодые розетки,
возрастом не более 2 лет. Они продолжают расти и со временем образуют клон материнского
растения.
Ниже по горным склонам произрастает близкий вид – восковой
зонтик (Platyaeonium umbellatum), обитающий на каменистых осыпях. У
него также ползучий стебель, и плоские зонтиковидные розетки диаметром до 50
см образуются на приподнимающихся стеблях. Листья имеют языковидную формы, сверху
покрыты слоем воска, особенно обильным у притупленной вершины листа. Растение
очень светолюбиво и благодаря плотным розеткам листьев затеняет своих конкурентов.
Цветы у этого вида карминно-красного цвета со слабым запахом, опыляются главным
образом птицами и осами, собирающими воск с листьев растения для постройки гнёзд.
Цветонос образуется у основания розетки листьев и прорастает между листьями
вверх.
Скальный
фикус (Ficus platycaulis)
Порядок: Крапивные (Urticales)
Семейство: Тутовые (Moraceae)
Место обитания: горы Южной и Юго-Восточной Азии, отвесные склоны.
В эпоху человека фикусы отличались большим разнообразием жизненных форм: среди
них можно было встретить деревья, кустарники и лианы любого размера. Именно
фикусы могут образовывать своеобразную жизненную форму баньян: одно дерево,
разрастаясь в стороны и давая многочисленные корни-подпорки, может образовывать
целый лес. Среди фикусов известны также «душители», которые сдавливают многочисленными
корнями ствол дерева, в развилке которого растут, и постепенно подавляют и
замещают его в пологе леса. В неоцене среди многообразных фикусов в тропиках
Старого Света появился лазающий вид, освоивший горные склоны.
Скальный фикус растёт в условиях обильных дождей и высокой температуры круглый
год. Это растение начинает свою жизнь как эпифит, когда его небольшое семечко
попадает в трещину скалы, где скапливается мох и растительный мусор. Проросток
начинает быстро расти, цепляясь густой щёткой придаточных корней за малейшие
трещинки скал. Постепенно тонкий стебель разрастается и достигает соседних
трещин, где закрепляется и продолжает тянуться вверх. Постепенно стебель утолщается,
одревесневает, и корни начинают глубже проникать в трещины скал, заполняя и
скрепляя их, словно цемент. Стебель взрослого растения нарастает не столько
в толщину, сколько в ширину: он плоский и лентовидный. Растение обильно ветвится,
покрывая собой огромные участки горных склонов. Длина стебля может достигать
100 м и более, а площадь, занятая растением – до нескольких тысяч квадратных
метров.
Разумеется, такая мощная зелёная масса растения не может существовать только
за счёт скудного минерального питания, находимого корнями в трещинах скал.
Достигнув определённого размера, скальный фикус спускает со скал длинные воздушные
корни вниз, к почве. Вначале это всего лишь один тонкий корешок, робко тянущийся
к земле, до которой может быть несколько десятков метров. Когда он достигает
почвы, дополнительный приток минерального питания стимулирует дальнейший рост
растения, и к почве тянется ещё несколько корней. Переплетаясь и разветвляясь,
они врастают в почву и утолщаются, становясь похожими на клубок исполинских
змей.
Скальный фикус образует на горных склонах и каменистых участках своеобразные
«оазисы», привлекая множество птиц и насекомых. Кроме того, растение укрепляет
склоны и препятствует обвалам. Конечно, иногда особенно сильные обвалы сносят
вниз заросли этого растения, но всё равно фикус довольно быстро восстанавливается
из оставшихся ветвей.
Листья скального фикуса мелкие, сердцевидные, кожистые. Они образуют сплошной
покров, под которым находят приют разнообразные насекомые и пауки, а также
обильно разрастаются мхи и лишайники. Кончик каждого листа сильно вытянут –
это своеобразная «капельница», характерная для тропических растений влажного
климата. Во влажную жаркую погоду кажется, что под зарослями скального фикуса
постоянно идёт дождь – капли непрерывно падают с листьев растения, и стекают
струйками с камней, на которых он растёт.
Соцветия этого вида невзрачны, как у всех фикусов. Это небольшие зелёные «ягоды»
– сиконии на стволе и крупных ветвях. Скальный фикус опыляется микроскопическими
осами, живущими только в его соцветиях. Личинки ос уничтожают часть семян,
но оставшихся хватит на воспроизводство растения. Зрелые плоды этого вида мелкие,
красно-коричневые, очень сладкие и мягкие. Птицы – основные распространители
семян этого растения. Они глотают плоды целиком, рассеивая семена скального
фикуса вместе с помётом.
Бегония гриболистная (Begonia fungifolia)
Порядок: Бегониевые (Begoniales)
Семейство: Бегониевые (Begoniaceae)
Место обитания: Юго-Восточная Азия, скалы; эпилитное растение.
Горные склоны в Юго-Восточной Азии представляют собой благоприятное место для
роста травянистых растений, которые привыкли довольствоваться небольшим количеством
почвы и влаги. Дожди обильно проливаются на скалы, но вода быстро стекает по
ним, и растения выработали приспособления, позволяющие удерживать и накапливать
влагу.
Среди зарослей, которые образуются на скалах, часто встречается вид растений,
который можно легко спутать с плодовыми телами каких-то грибов. Кажется, будто
среди мха выросло множество одинаковых грибов с зелёными кожистыми шляпками
и утолщёнными ножками, покрытыми волосками. Но среди зарослей этих «грибов»
поднимаются тонкие цветоносы, несущие кисти мелких розовых цветков. Эти цветки
очень характерные – у них четыре лепестка и трёхгранная «крылатая» завязь.
Они свидетельствуют о принадлежности их обладателя к обширному семейству бегониевых.
В неоцене некоторые представители этого семейства превратились в деревья, но
большинство осталось травянистыми растениями. Данный вид бегоний носит название
бегония гриболистная.
Это полусуккулентное растение, у которого развита водоносная ткань. Бегонии
тропического леса обладают крупными асимметричными листьями с тонкой листовой
пластинкой. У бегонии гриболистной лист, напротив, небольшой, округлый и кожистый.
Его диаметр не превышает 8 – 9 см. Листовая пластинка этого растения имеет
почти правильные округлые очертания, хрящеватую консистенцию и гладкую верхнюю
сторону со слегка вдавленными жилками. В противоположность большинству бегоний
с яркими листьями, листья бегонии гриболистной тусклые: они имеют тёмно-зелёный
цвет с более светлой каймой, а вдоль крупных жилок от центра листа тянутся
полоски светло-зелёного цвета, образующие звездообразный рисунок.
Черешок листа прикрепляется в центре щитовидной листовой пластинки и является
главным водозапасающим органом растения. Черешки листьев у этой бегонии вздутые,
похожи на ножку гриба. Они светло-зелёные и полупрозрачные; большую часть их
объёма составляет водозапасающая паренхима. Поверхность черешка покрыта волосками,
что снижает испарение.
Корневая система этой бегонии мочковатая, состоит из множества тонких корней.
На стебле растения развиваются многочисленные придаточные корни. Во время
дождя растение всасывает воду, которая быстро запасается в черешках листьев.
Стебель
растения представлен толстым ползучим корневищем; листья сохраняются только
в верхней части побегов. Черешки листьев неплотно прикрепляются к корневищу,
и при небольшом усилии отламываются. Отломленный лист долго сохраняет жизнеспособность
благодаря запасу влаги в черешке. На месте слома образуются придаточные почки,
из которых прорастают молодые побеги. Пока корневая система плохо развита,
они растут за счёт запасов влаги, сохраняющейся в черешке старого листа.
Бегония гриболистная цветёт почти круглый год, кроме нескольких недель, когда
количество осадков минимально. Она опыляется насекомыми разных видов – преимущественно
пчёлами и бабочками. Мелкие семена этого растения просто высыпаются из раскрывшихся
коробочек, и их разносит ветер.
Серебряная брахибегония (Brachybegonia argentimaculata)
Порядок: Бегониевые (Begoniales)
Семейство: Бегониевые (Begoniaceae)
Место обитания: Юго-Восточная Азия, полог влажного тропического леса.
Богатое видами семейство бегониевых в эпоху человека демонстрировало относительную
бедность на уровне родов – в это время существовало лишь четыре рода, из которых
три были представлены единичными видами, и только род Begonia был очень разнообразен
и широко распространен в тропиках. В неоцене среди бегоний появились особые
виды бегониевых деревьев (Begoniodendron) –
своеобразная попытка выйти в крупноразмерный класс и образовать новые
жизненные формы. На уровне травянистых
растений эволюция бегоний также продолжилась, и в неоцене на территории Юго-Восточной
Азии появился род близких к бегониям эпифитных растений брахибегония (Brachybegonia).
Эти растения освоили жизнь в пологе леса и представляют собой исключительно
эпифитные травянистые формы. Они сохраняют характерные несимметричные листья,
расположенные двурядно. Стебель брахибегоний сильно укорочен, и листья лежат
один на другом. Черешки листьев также настолько сильно укорочены, что листья
кажутся растущими прямо из стебля, образуя симметричную розетку. Корни этих
растений очень тонкие и нежные, корневая система мочковатая. Корни брахибегоний
обильно ветвятся и проникают в трещины на коре дерева, прочно закрепляя на
его ветвях даже крупные кусты.
Нижние листья в розетке, самые старые, отличаются по форме и цвету от верхних.
Они более округлые, короткие и широкие. Окраска таких листьев тусклая. Когда
растение развивается в полную силу, эти листья оказываются в тени. Но они не
отмирают, а служат своеобразными ловушками для органики. В них накапливаются
мусор, остатки коры, помёт птиц и насекомых. От стебля, и даже от выше расположенных
листьев сюда спускаются придаточные корни, и растение получает дополнительное
питание. Подобную роль играют стерильные вайи у папоротника «олений рог» (Platycerium),
который также сохранился в тропических лесах неоцена. Таким образом, брахибегонии
и папоротник «олений рог» представляют собой пример конвергенции.
Верхние листья брахибегоний сильно отличаются от нижних. Они начинают развиваться,
когда растение достаточно окрепнет. Эти листья бывают крупнее, у них длинный
кончик-«капельница», а поверхность листа бывает покрыта узором из полос и пятен,
похожим на узор листьев бегоний.
От бегоний брахибегонии унаследовали лёгкость вегетативного размножения. Для
них типично образование в пазухах листьев длинных побегов, на кончиках которых
развиваются розетки листьев. Такие «детки» легко отрываются от побегов и укореняются
в подходящих местах. На листьях некоторых видов развиваются выводковые почки,
дающие начало молодым растениям.
Семенное размножение играет важную роль в расселении растений на большие расстояния.
Взрослые растения выпускают из вершины главного побега цветонос, на котором
развивается соцветие – кисть из нескольких десятков, а у крупных видов даже
из сотен мелких цветков. Плод брахибегоний – коробочка, наполненная сухими
мельчайшими семенами. Они легко разносятся ветром на многие километры. После
цветения стебель отмирает, но в его основании из почек развивается один или
несколько боковых побегов, которые замещают родительское растение. Один побег
живёт до четырёх лет у крупных видов, и не больше двух лет у мелких.
Разные виды брахибегоний отличаются друг от друга формой и окраской листьев,
размерами взрослого растения и количеством цветков. Серебряная брахибегония
обитает в верхнем ярусе тропического леса. Её розетки достигают 50 – 60 см
в ширину и насчитывают до 10 – 15 листьев. Верхняя сторона листьев почти целиком
покрыта мелкими волосками, придающим поверхности листа серебристый блеск. Лишь
в середине листа ближе к месту прикрепления черешка волоски отсутствуют и лист
имеет типичный зелёный цвет и гладкую поверхность; нижняя часть листа всегда
покрыта белым опушением, которое со временем редеет. Молодые листья из-за густого
опушения выглядят почти белыми. Также опушены цветоносы растения. Соцветие
этого вида щитковидное. Цветки имеют окраску, резко контрастирующую с окраской
листьев – они тёмно-красные. Их опыляют преимущественно одиночные пчёлы и мухи
разных видов. Этот вид обитает в приречных тропических лесах Индокитая.
Живородящая брахибегония (Brachybegonia vivipara) отличается от прочих видов
этого рода повышенной способностью к вегетативному размножению. У неё крупные
листья с зубчатым краем, тёмно-зелёные с красноватой серединой, покрытые по
краю грубыми жёсткими волосками. На зубцах по краю листа образуются выводковые
почки, из которых развиваются мелкие дочерние растения. Соцветие представляет
собой раскидистую метёлку из нескольких десятков ярко-розовых цветков. Этот
вид распространён во влажных горных лесах на Берегу Джакарта.
Лазающая брахибегония (Brachybegonia scandens) избрала другую тактику вегетативного
размножения. Этот вид образует сравнительно короткие побеги-«усы», обладающие
способностью ветвиться и укореняться в узлах. От розетки материнского растения
в разные стороны расползаются усы. Часть из них укореняется на коре и в зарослях
мха, и продолжает рост вверх или вниз. Побеги, оставшиеся в воздухе, прекращают
рост и вскоре отмирают. Успешно растущий побег ветвится и продолжает рост в
длину. Добравшись до места, где скапливается перегной, ус укореняется и образует
молодое растение. Листья этого вида образуют розетку шириной около 50 см из
округлых листьев светло-зелёного цвета с мелкими зубцами по краям. Поверхность
листа покрыта короткими редкими волосками рыжего цвета. Этот вид цветёт высокой
остроконечной метёлкой из нескольких десятков мелких белых цветков. Лазающая
брахибегония произрастает в тёплых влажных лесах низинных районов Берега Джакарта
и близлежащих островов, предпочитая верхнюю часть полога леса.
Гигантская брахибегония (Brachybegonia gigantea) из тропических лесов Индостана
и Юго-Восточной Азии представляет собой самый крупный вид рода. Она образует
огромные листья – длиной до метра при ширине около 70 см. Листья этого вида
цельные, с небольшими зубцами по краю. Их окраска ярко-зелёная, по краю протягивается
узкая розовато-красная кайма. По гладкой поверхности листа разбросаны отдельные
пятна белого цвета, чаще всего расположенные в месте прикрепления черешка к
листовой пластинке. Цветонос достигает длины двух метров и несёт до тысячи
мелких красных цветков. Этот вид выпускает небольшое количество усов, но они
легко укореняются в узлах и тянутся по стволу и ветвям на расстояние до пяти
метров.
Изящная брахибегония (Brachybegonia elegans) является самым мелким видом рода
– ширина её розетки не превышает 10 – 12 см. Её округлые листья окрашены в
ярко-красный цвет с зелёной каймой неправильной формы – от отдельных «затёков»
и «брызг» до широкой каймы, достигающей половины радиуса листа. Она быстро
растёт и даёт обильные отпрыски. Взрослое растение начинает образовывать боковые
отростки даже до цветения. Цветонос этого вида очень длинный по сравнению с
размером самого растения – он достигает почти полуметровой длины. Соцветие
этого вида – кисть крупных розовых цветков, повисающих на длинных цветоножках.
Этот вид распространён во влажных лесах от восточной части Индостана до юга
Берега Джакарта.
Брахибегония рассечённая (Brachybegonia dissecta) имеет листья необычного облика.
Первые листья этого растения обычные, округлые. Но по мере наступления зрелости
они меняют форму – между жилками появляются вырезы, и самые крупные листья
имеют надрезы глубиной до трети ширины листа. Это приспособление помогает избежать
затенения нижних листьев верхними. Поверхность листьев гладкая, тёмно-зелёная
с одиночными тёмно-красными пятнами. Ширина розеток этого растения – около
40 см. Этот вид населяет нижнюю часть полога леса, где уже начинает сказываться
недостаток света. Рассечённые листья – приспособление, позволяющее компенсировать
это. Этот вид распространён в низинных лесах Берега Джакарта и Индокитая.
Ползучая тектобегония (Tectobegonia repens)
Порядок: Бегониевые (Begoniales)
Семейство: Бегониевые (Begoniaceae)
Место обитания: Юго-Восточная Азия, влажный тропический лес.
В эпоху человека бегонии были исключительно травянистыми растениями или полукустарниками.
В эпоху экологического кризиса, вызванного деятельностью человека, многие виды
этих растений стали редкими, и даже просто вымерли. Зато после исчезновения
человечества и восстановления тропических лесов бегонии смогли освоить иные
жизненные формы. Среди них появились бегониевые
деревья (Begoniodendron) и
эпифитные брахибегонии (Brachybegonia). А по стволам деревьев влажного тропического
леса Юго-Восточной Азии взбираются вверх их родственники. В них легко узнать
потомков бегоний: листья этих растений имеют характерную кососердцевидную форму.
Но внешний вид этого растения нетипичен для бегоний – листья располагаются
на стебле в два ряда, перекрывая друг друга, словно черепица. Отсюда название
этого растения – тектобегония (от лат. “tectus” – покрытый).
Стебель тектобегонии очень длинный – у старых экземпляров он достигает длины
6 – 8 метров. Стебель плотно прижимается к коре деревьев, и растение ползёт
по поверхности ствола дерева. Это корнелазающая лиана, подобно плющу (Hedera)
– в узлах стебля развивается большое количество белых прочных корешков, внедряющихся
в кору и моховые подушки. Стебель уплощенный, лентовидной формы. Он хрупкий
и легко обламывается; кроме того, растение ветвится и общая длина стеблей одной
особи может достигать 20 – 30 метров. Обычно растение начинает жизнь, укореняясь
в растительном мусоре. Первые недели жизни побег растёт вертикально, но затем
ложится и начинает ползти. Постепенно побеги приобретают форму, типичную для
данного вида: они начинают ползти по субстрату, стебли становятся лентовидными,
междоузлия сближаются, и листья начинают расти, перекрываясь черепицеобразно.
Тектобегония растёт во влажных лесах и такое листорасположение помогает отклонить
потоки дождевой воды. Растение быстро разрастается и выносит побеги в хорошо
освещённый полог леса. У старых экземпляров корни и основания стеблей отмирают,
но верхняя часть стебля не погибает. Она продолжает расти как эпифит, питаясь
за счёт перегноя, который скапливается в дуплах и среди ветвей. Это растение
хорошо приспособлено к вегетативному размножению: сломанный лист легко укореняется,
и в скором времени образует полноценное растение.
Листья ползучей тектобегонии тёмно-зелёные, округлые, с удлинённым кончиком,
вытянутым в виде капельницы. Длина листа – не более 5 см, ширина – до 3 см.
В тени листья выглядят невзрачно, но на солнце, когда растение оказывается
в верхней части кроны дерева, на листьях появляются серебристые пятна.
Во время цветения тектобегония образует в пазухах листьев короткие цветоносы,
на которых сидят густые шаровидные кисти мелких белых цветков. На молодых листьях
в основании цветоноса появляются большие серебристые пятна, привлекающие насекомых
издалека своим блеском. Обычно тектобегонии опыляются бабочками и мухами, реже
одиночными пчёлами.
Плод тектобегоний – крылатая коробочка. Мелкие семена этих растений (весом
всего несколько миллиграммов) разносятся ветром.
Бегониевое
дерево кислолистное (Begoniodendron oxyphyllus)
Порядок: Бегониевые (Begoniales)
Семейство: Бегониевые (Begoniaceae)
Место обитания: тропические леса Южной Азии. Близкие виды распространены от
Земли Зиндж до Юго-Восточной Азии.
В эпоху голоцена в семейство бегониевые входили исключительно многолетние травянистые
растения; у некоторых видов однолетние надземные побеги отмирали, но сохранялся
крупный многолетний клубень. Среди этих растений встречались наземные травы
(часто достигавшие крупных размеров), травянистые лианы и эпифиты. Но в эпоху
неоцена, когда состав древесной растительности в тропиках существенно поменялся,
среди бегоний возникли древовидные формы, представители рода бегониевых деревьев
Begoniodendron. Растения, входящие в этот род, отличаются признаками, характерными
ранее для травянистых бегоний. Они растут очень быстро – годовой прирост в
высоту достигает пяти метров и даже больше. Древесина бегониевых деревьев рыхлая,
очень сочная и хрупкая. Иногда во время сильных ветров ствол дерева ломается,
но это не приносит ему большого вреда: сказывается ещё одна черта, унаследованная
от травянистых бегоний – способность к вегетативному размножению. Поваленное
дерево легко укореняется, лёжа на земле. От ствола начинают расти вертикальные
побеги, а ветви изгибаются вверх. Так одно дерево превращается в густую заросль.
Через какое-то время часть стволов будет заглушена и погибнет, но несколько
стволов разовьются нормально и, постепенно утолщаясь, дорастут до высоты 25
– 30 метров – обычной высоты бегониевых деревьев.
Кроме этого бегониевые деревья сохранили способность размножаться укоренением
веток и даже отдельных листьев.
Благодаря скорости роста бегониевые деревья – пионерная растительность тропических
лесов. Они одними из первых поселяются на свободной земле, где упало старое
дерево, или на речных наносах (бегониевые деревья очень влаголюбивы). Они растут
быстро, но живут очень недолго: не больше 50 – 70 лет, если их не свалит буря.
Древесина бегониевых деревьев, сравнительно мягкая и сочная, была бы прекрасным
лакомством для всевозможных личинок насекомых. Но деревья рода Begoniodendron
выработали особую стратегию защиты от них, развив одну из особенностей их предков.
Сок бегониевых деревьев – настоящий химический «коктейль» из всевозможных органических
кислот – яблочной, лимонной, и даже муравьиной и уксусной. У разных видов состав
сока варьирует, но принцип его действия остаётся прежним: в очень кислой среде
практически не живут личинки насекомых, а развитие грибов и бактерий существенно
подавляется.
Как и травянистые бегонии, бегониевые деревья отличаются своеобразными косыми
листьями. Они окрашены не столь ярко, как у мелких травянистых бегоний, но
у ряда видов на листьях есть серебристые пятна и опушение, а некоторые виды
имеют изящные разрезные листья. Это вечнозелёные деревья.
Достигнув примерно десятилетнего возраста, бегониевые деревья начинают обильно
цвести. Небольшие розовые четырёхлепестковые цветки этих деревьев собраны в
крупные щитковидные соцветия. Их окружает пара ярко окрашенных листьев-прицветников.
Женские цветки располагаются на концах ветвей в средней и нижней части кроны,
а мужские распускаются в основном на вершине дерева. Если насекомые не очень
активно посещают цветки, то есть шанс опыления при помощи ветра. Цветут бегониевые
деревья практически круглый год. Лишь в местах с сезонными засухами цветение
прерывается на два – три месяца. Семена бегониевых деревьев очень мелкие. Они
переносятся ветром на больше расстояния, поэтому бегониевые деревья часто встречаются
на островах Индийского океана.
Бегониевое дерево кислолистное – типичный представитель рода. Его высота –
до 20 метров. Этот вид встречается в долинах рек, где образует иногда плотные
заросли. Это растение выносит затопление корней во время паводков, сохраняя
устойчивость с помощью хорошо развитых досковидных корней. Ствол этого дерева
получает дополнительную прочность от того, что имеет не цилиндрическую, а ребристую
форму – в виде пяти- или шестигранной призмы. Это связано с тем, что этот вид
деревьев растёт в местах, где часто дуют сильные ветры. Кора окрашена в серый
цвет – это предохраняет ствол дерева от перегрева.
Листья этого дерева тёмно-зелёные с многочисленными серебристыми пятнами, мелкозубчатые
по краю. Длина листьев – около 25 см, ширина – 7 – 10 см. Прицветники очень
крупные (длиной до 30 см), округлой формы, тёмно-красные с алыми пятнами. Поверхность
прицветников и листьев по-разному отражает ультрафиолетовые лучи, поэтому насекомые
легко замечают цветы растения. Цветки мелкие, бледно-розовые, собраны по 30
– 40 штук. Плод – трёхгранная коробочка.
В род бегониевых деревьев также входят виды:
Бегониевое
дерево «серебряная рука» (Begoniodendron chira-argentea) – распространено
на островах Индийского океана. Высота – до 15 – 20 метров. Ствол толстый, «бутылочный»,
приспособлен для хранения запаса пресной воды. Корневая система неглубокая
(дерево избегает засоленных грунтовых вод на островах), но представлена мощными
досковидными корнями, расходящимися далеко в стороны – они удерживают дерево
от падения во время штормовых ветров, проносящихся над островами. Листья длиной
около 20 см, округлых очертаний, разделены на пять – шесть долей, обильно покрыты
серебристыми волосками. Волоски служат для защиты от избыточного солнечного
излучения, а во время дождей абсорбируют воду. Кора тёмно-серая, толстая.
Цветки ярко-красные, прицветники розовые. Цветёт в сезон муссонов, при недостатке
влаги сбрасывает часть листвы.
Бегониевое
дерево отпрысковое (Begoniodendron proliferus) – обитает на Восточно-Африканском
субконтиненте, населяет тропические леса. Высота – около 25 метров.
Произрастает в лесу, избегая переувлажнённых почв. Листья косо-сердцевидные,
с крупными зубцами по краю, светло-зелёные с красными пятнами между
главными жилками. В месте прикрепления черешка на листовой пластинке
развивается придаточная почка. На ветках, особенно сильно освещённых
солнцем, она начинает развиваться в молодое растение. В это же самое
время прикрепление черешка к ветке ослабевает, и во время бури такие
листья с молодыми растеньицами отрываются от материнского дерева и
уносятся на многие десятки метров в сторону. Лист материнского растения
служит парусом, увеличивая дальность полёта молодых растений. Упав
на землю, такой отпрыск быстро укореняется и трогается в рост.
Цветки мелкие, розовые. Прицветники очень крупные, красного цвета.
Бегониевое
дерево ползучее (Begoniodendron repens) – произрастает в дождевых лесах Юго-Восточной
Азии, наиболее часто встречаясь в горных лесах и по обрывистым берегам рек.
У этого дерева нельзя однозначно сказать, что можно считать «высотой» - главный
ствол этого дерева вначале растёт вверх, затем быстро полегает и продолжает
рост в горизонтальном направлении. От него отходят многочисленные толстые вертикальные
стволы. Такая форма роста позволяет растению «вклиниваться» между крупными
деревьями, значительно опережая в росте сеянцы деревьев-конкурентов. Чтобы
«закрепить достижения», горизонтальный ствол активно растёт в толщину. Длина
основного ствола может достигать 50 метров и более, вертикальные стволы 2-го
порядка достигают 15 - 20-метровой высоты. На речных обрывах основной ствол
может свешиваться вниз, спускаясь вниз по склону до самой воды, образуя иногда
своеобразные естественные «мосты» через узкие реки.
Листья крупные (длина – до 40 см, ширина – около 30 см), косо-сердцевидные,
край листа волнистый. Кончик листа сильно вытянут в «капельницу» – это общая
черта растений влажного дождевого леса. Листья окрашены в тёмно-зелёный цвет
с прерывистыми серебристыми полосами вдоль крупных жилок.
Прицветники серебристые с несколькими зелёными пятнами; цветки крупные, розовые.
Пустынный цикламен (Aridocyclamen macrotuber)
Порядок: Первоцветные (Primulales)
Семейство: Первоцветные (Primulaceae)
Место обитания: пустыни Средиземноморской низменности.
На протяжении миллионов лет Африка двигалась на север, в сторону Европы. И
через миллионы лет после исчезновения человека это привело к значительным изменениям
природных условий в районе Средиземноморья. Когда закрылся Гибралтарский пролив,
Средиземное море очень быстро высохло, и на его месте образовалась суровая
солёная пустыня, окружающая болота с мёртвой сверхсолёной водой. Высыхание
Средиземноморья значительно изменило климат Южной Европы – он стал сухим и
суровым, с значительными сезонными и суточными перепадами температур. Пустыню
пересекает несколько небольших рек. Одни из них впадают в болота, другие теряются
в песках. Все они берут начало в альпийских ледниках и дают жизнь немногим
обитателям пустыни.
Сухие пустыни тянутся лентой по краям областей бывшего морского дна, покрытых
коркой соли. Эти места отличаются суровым климатом, но их всё же заселяют растения
способные запасать или сохранять влагу. Растений в пустыне значительно больше,
чем кажется на первый взгляд. Для пустыни очень характерны многолетники-эфемеры,
цветущие весной, когда почва достаточно увлажнена потоками воды, текущими с
гор.
Одно из самых красивых растений Средиземноморской пустыни – пустынный цикламен,
представитель эфемерной растительности. Этот вид – выходец с Альп, потомок
европейского цикламена (Cyclamen europaea). Он обитает по берегам временных
водоёмов, где дольше сохраняется влага в почве.
Это растение развивается в течение двух месяцев весной, пока почва достаточно
влажная. Лето, осень и зиму пустынный цикламен переживает в виде большого веретеновидного
клубня. На глубине около 40 см, где находится клубень, почва смягчает перепады
температур и защищает от большинства травоядных животных. Но во время вегетации
клубень буквально выползает наверх, ближе к солнцу. Это свойство растения связано
с особенностями его строения. Корни пустынного цикламена очень длинные. Они
надёжно удерживают растение в почве, и обладают способностью скручиваться при
высыхании. Весной корни напитываются влагой и выталкивают клубень ближе к поверхности
земли, а в начале засухи сокращаются и втягивают клубень глубоко под землю.
В предгорьях Альп встречаются популяции этого вида, которые могут вегетировать
значительно дольше, чем в пустыне. Но этот вид отличается пониженной конкурентоспособностью
– пустынный цикламен очень светолюбив и не терпит затенения.
Весной, когда почва становится достаточно влажной, растение трогается в рост.
Оно быстро, в течение нескольких дней, развивает 4 – 5 крупных округлых листьев
(диаметром до 15 см) на длинных черешках. Листья имеют сероватый оттенок из-за
слоя воска, который развивается на них. После этого пустынный цикламен зацветает.
Его цветонос быстро разрастается и достигает высоты 70 см. Одновременно на
растении держится не больше 2 – 3 крупных цветков с лепестками, характерным
образом отогнутыми назад. Цветки испускают сильный аромат, который становится
особенно интенсивным к вечеру. Это растение опыляется бабочками и одиночными
осами. Особенно часто на цветках пустынного цикламена встречаются осы. Эти
насекомые ищут на растении не только нектар, но и собирают с листьев воск,
нужный для постройки гнёзд. Цветки пустынного цикламена розовые с серебристыми
краями лепестков, отражающими ультрафиолетовые лучи – это помогает привлекать
насекомых.
К началу лета у пустынного цикламена созревают семена. В отличие от предкового
вида, семена которого очень быстро теряли всхожесть, семена этого вида могут
сохранять всхожесть до 3 лет – в изменчивом мире пустыни условия не всегда
благоприятны для прорастания, а часть всходов может просто погибнуть. Но в
почве всегда есть резерв семян пустынного цикламена. Семена имеют прочную оболочку,
на которой растёт съедобный маслянистый придаток. Благодаря ему их охотно склёвывают
птицы, с помощью которых этот вид расселяется. Пустынный цикламен часто встречается
в «оазисах» Средиземноморских болот.
Крылоклённик
пальчатолистный (Pteracer palmatus)
Порядок: Сапиндовые (Sapindales)
Семейство: Кленовые (Aceraceae)
Место обитания: тёплые влажные леса Южной Европы (Балканы), Кавказа и Малой
Азии (по южному берегу Четвероморья).
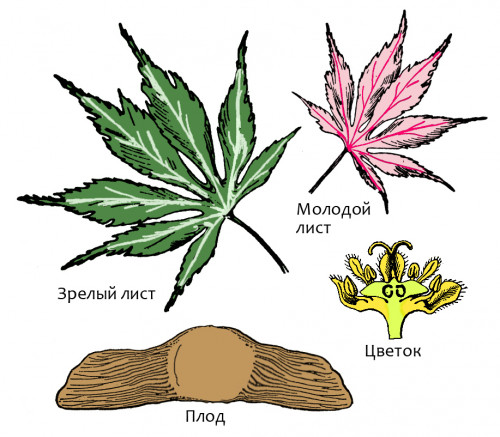
Рисунок Биолога
В голоцене лесная флора Европы подверглась суровым испытаниям
на выживаемость. Человек вырубил некогда почти сплошные и непроходимые леса,
а после этого испытания наступил ледниковый период. Когда Средиземное море пересохло,
и на его месте образовалась жаркая пустыня, леса Европы сохранились на узкой
полосе между северной тундрой и суровой солёной пустыней Средиземноморья. Леса
остались на юге Западной Европы, где их орошали тучи с Атлантики, а также по
горным хребтам Альп и Карпат. Достаточно большие лесные массивы образовались
на Балканах.
Когда в Европе началось потепление, леса начали активно распространяться дальше
на север. К тому времени человек уже исчез, и живая природа эволюционировала
по собственным законам. В неоцене область тёплого климата сдвинулась ближе к
полюсам, но это происходило не во всей Евразии. Евразия – огромный массив суши,
область резко континентального климата. Значительные территории этого континента
удалены от крупных водоёмов, делающих климат ровным и влажным, поэтому на северо-востоке
Евразии зимы по-прежнему холодны и снежны.
На юге и востоке Европы, где ощущается влияние Четвероморья, признаки, характерные
для тропических растений, появились у потомков видов умеренной флоры. Это широкие,
часто сложные и рассечённые листья с острыми кончиками-«капельницами». Орех,
каштан, клёны и платаны – типичные виды, составляющие европейские субтропические
леса эпохи неоцена. Острая конкуренция вызвала появление эффективных приспособлений
для размножения и расселения.
Ветер несёт над кронами крупное плоское семечко с двумя широкими полупрозрачными
крыльями. От каждого порыва оно взмывает выше и выше, и летит на большое расстояние.
Оно свалилось с высокого дерева, достигающего высоты 30 – 35 метров. Это дерево
средней величины в европейских субтропических лесах – оно из числа видов, которые
составляют полог леса и не «высовываются» над ним. У дерева зонтиковидная крона,
а на гибких молодых ветвях покачиваются гроздья таких же крылатых семян.
Семя этого дерева с двумя большими крыльями подсказывает происхождение этого
растения: это дерево – потомок клёна (Acer), разные виды которого были характерны
для Северного полушария. За характерную форму семян это растение называется
крылоклённик. Для клёна характерен плод двукрылатка – два семени, несущих по
одному крылышку. При созревании растения они распадаются и держатся в воздухе
благодаря вращению падающего семени вокруг своей оси. У крылоклённика на плодах
в двукрылатке один плод редуцирован и от него осталось только крылышко. Остаток
этого семени – небольшой рубчик с одной стороны плоского дисковидного семени
с двумя широкими крыльями. Порывы ветра относят такие семена далеко от материнского
растения. Семечко лёгкое и может долго находиться в воздухе, реагируя даже на
слабое дуновение ветра.
Если ему повезёт попасть в благоприятное для жизни место и прорасти, через восемьдесят
– сто лет из него разовьётся огромное дерево крылоклённика. Ствол взрослого
дерева этого вида прямой, по мере роста нижние ветви опадают. У взрослого дерева
первые крупные ветви находятся на высоте 20 метров от земли. Ствол крылоклённика
покрыт гладкой серой корой, которая постоянно отшелушивается небольшими чешуйками
– такая особенность не даёт дереву обрастать эпифитами, которые при благоприятных
условиях образуют «висячие сады» весом в десятки килограммов. Ему придают дополнительную
устойчивость несколько толстых досковидных корней-подпорок в нижней части ствола.
Такие корни начинают развиваться, когда дерево достигает приблизительно половины
нормальной высоты.
Лист крылоклённика типичен для растений влажного леса: он разрезной на 7 – 9
острых долей, разделённых по длине на 9/10 и расположенных на черешке веером.
Ткани листа довольно нежные. Листья сидят на молодых побегах попарно, и достигают
диаметра около 50 – 60 см. Молодые листья этого дерева розовые с ярко-малиновыми
полосами вдоль прожилок – это так называемая антоциановая окраска, защита от
солнечного излучения. Взрослый лист крылоклённика пальчатолистного светло-зелёный,
но вдоль жилок располагается более тёмная зона. Жилки взрослого листа белые.
Это растение полулистопадное – осенью, при похолодании крылоклённик сбрасывает
примерно треть старых листьев и проводит в относительном покое около 2 месяцев.
На ветках остаётся лишь самая молодая листва. На севере ареала это дерево может
терять почти все листья. Период покоя является очень важной особенностью крылоклённика:
в это время у дерева закладываются цветочные почки.
Листва мешает насекомым-опылителям находить цветы, поэтому тропические деревья
делают цветы более заметными с помощью сильного запаха или яркой окраски венчика.
У крылоклённика соцветие длинное, из множества мелких пятилепестных цветков.
Это дерево опыляется пчёлами и осами. Цветки располагаются гроздьями на длинном
цветоносе, который повисает с ветки вниз. Цветок этого дерева размером около
2 см. Он жёлтый, ароматный, с короткими лепестками и широким цветоложем, в середине
которого растут два плодолистика, образующие структуру, похожую на голову старинного
ключа. После опыления одна завязь редуцируется, а вторая развивается. Крыло
редуцированной завязи достигает такого же размера, как на нормальной завязи,
и в итоге получается одно крупное семечко с двумя широкими крыльями, которое
может долго держаться в воздухе. Оно прорастает весной, и начинает быстро расти.
Обычно в подлеске растёт много проростков этого растения, но большая часть их
гибнет, не найдя благоприятных условий для жизни. За первый год жизни молодое
растение достигает высоты около двух метров. В первые годы жизни крылоклённик
может расти в тени подлеска, конкурируя с другими растениями, но, если он не
получает достаточно света, он начинает чахнуть. Если же старое дерево, закрывающее
доступ света в подлесок, падает, молодой крылоклённик начинает быстро расти,
стремясь обогнать конкурентов. Это дерево зацветает первый раз в возрасте около
сорока лет, достигнув высоты примерно 20 метров. Предельная продолжительность
жизни этого растения достигает 400 лет.
Конский
каштан зонтичный (Aesculus umbelliferus)
Порядок: Сапиндовые (Sapindales)
Семейство: Конскокаштановые (Hippocastanaceae)
Место обитания: Балканы, Карпаты, Кавказский полуостров, южный и западный берега
Четвероморья.

Рисунок Биолога
Появление Четвероморья оказало значительное влияние на климат
окружающих территорий. На Балканах узкая полоса земли между горами и берегами
Четвероморья превратилась в область своеобразных субтропических лесов, образованных
потомками более северных видов деревьев. Произрастающие здесь виды деревьев
отличаются широкими листьями, на кончиках которых развиваются характерные «капельницы»
для удаления избытка влаги. Зимой деревья сбрасывают часть листвы и приостанавливают
рост, но этот период довольно короткий, и годичные кольца в их древесине выражены
слабо.
Один из характерных обитателей субтропических лесов по берегам Четвероморья
– конский каштан зонтичный. Это дерево легко узнаётся издалека по своеобразным
листьям, которые сильно отличаются от листьев других деревьев этих лесов. У
конского каштана зонтичного листья напоминают пальчатосложный лист его предка,
конского каштана обыкновенного (Aesculus hippocastanum), но отличаются от него
несколькими признаками: размером и количеством листочков. У конского каштана
зонтичного гигантские листья, похожие одновременно и на зонтики, и на веерные
листья пальм. Черешок каждого такого листа, длиной около метра и толщиной до
четырёх сантиметров, несёт листовую пластинку диаметром около полутора метров,
которая состоит из 15 – 19 приблизительно одинаковых листочков, расширенных
и скруглённых на концах. Средняя жилка в каждом листочке образует тонкую вытянутую
капельницу.
Обладая огромными листьями, конский каштан зонтичный очень требователен к наличию
влаги. Это дерево встречается по берегам рек, и изредка заходит даже на болота,
где произрастает на более сухих участках. Наиболее часто этот вид деревьев встречается
в лесах вдоль побережья Четвероморья.
Кора этого дерева гладкая, тёмно-серая. Конский каштан зонтичный может достигать
высоты 25 – 30 метров. Обычно это одноствольное дерево, но часто он даёт прикорневые
отпрыски и растёт несколькими стволами, образуя пышную крону. Однако, такие
деревья чаще разламываются на отдельные стволы и падают, поскольку у каждого
ствола образуется несимметричная односторонняя крона, и ствол может не выдержать
тяжести листвы и ветвей. Деревья с нормально развитой кроной могут расти до
300 лет и больше. С возрастом ствол дерева становится ребристым – на нём появляются
своеобразные «рёбра жёсткости», отдалённые подобия досковидных корней тропических
деревьев. Благодаря рыхлой коре обеспечивается дыхание корней и растение хорошо
выдерживает переувлажнённую почву.
Зимой конский каштан зонтичный лишается значительной части листвы. На ветках
остаются только самые молодые листья, которые не достигают своего полного развития
и вырастают лишь до 30 – 40 см в диаметре. А сильный заморозок может вовсе убить
их, и тогда огромное дерево стоит безлистным. Но весной, когда возвращается
тепло, дерево быстро одевается новой листвой. Лист этого растения развивается
очень быстро – за неделю он достигает своего обычного размера. В это же самое
время дерево зацветает, становясь истинным украшением субтропических лесов.
Соцветие конского каштана зонтичного – тирс высотой до метра. Цветоносы поднимаются
вертикально, как у его предка. Цветение начинается снизу соцветия. Цветки этого
дерева довольно крупные – их диаметр достигает 4 – 5 см. Они зигоморфные, белого
цвета с жёлтой серединой; в зеве цветка разбросаны чёрные пятнышки. Это дерево
не специализировано к определённым опылителям. На его цветках кормятся бабочки,
жуки и одиночные пчёлы. Очень часто на цветках этого дерева встречаются птицы,
которые играют немаловажную роль в опылении растения. По мере отцветания каждый
цветок этого вида меняет окраску с белой на жёлтую и розовую. Непосредственно
перед опадением лепестки приобретают на один или два дня пунцово-красный цвет.
Плоды конского каштана зонтичного похожи на плод его предка – они крупные, кожистые,
с оболочкой, покрытой шипиками. Однако, в каждом плоде созревает не одно, а
два семени, как это было, очевидно, у древних видов этого рода. Однако, второй
семязачаток у этого вида появился вторично. В процессе эволюции конский каштан
зонтичный приспособился к распространению его семян мелкими животными, и его
семена относительно мелкие – диаметром лишь около 2 см. Их распространяют лесные
грызуны и птицы. Осенью семенами этого растения питаются дятлы.
Многие семена этого дерева оказываются съеденными осенью и зимой, но некоторым
удаётся уцелеть и они прорастают. Первый раз это дерево зацветает в возрасте
около 30 лет, при высоте 10 – 15 метров.
Бальзамин
багряный (Impatiens cinnabarifolia)
Порядок: Верескоцветные (Ericales)
Семейство: Бальзаминовые (Balsaminaceae)
Место обитания: дождевые леса на севере Меганезии, участки нарушенного древостоя,
берега рек.
Симбиоз между растениями и животными чаще всего строится в отношении опыления
растений, к которому узко приспосабливается определённый вид опылителей, наличие
которого гарантирует воспроизводство растения. Реже растение предоставляет
убежище насекомым вроде муравьёв, которые защищают его от насекомых-вредителей.
Один из видов травянистых растений, бальзамин багряный, установил симбиотические
отношения с представителем позвоночных животных, что само по себе является
редкостью в растительном мире. Это растение находится в симбиозе с одной из
местных птиц, шалашником-садовником
(Hortulornis plantarum-cultor). Два обстоятельства
позволили установиться симбиотическим отношениям: потребность птицы в ярких
предметах с одной стороны, и красная антоциановая окраска молодых листьев растения,
с другой стороны. Возможно, предки данного вида шалашника первоначально просто
обрывали подобные яркие верхушки растений и раскидывали их вокруг своей постройки
для привлечения самок. Верхушки бальзамина легко укореняются, и это растение
во многих случаях приживалось возле построек шалашников. Постепенно эволюция
птицы пошла в направлении поддержания жизни таких «декоративных растений»:
птицы обрывали ростки других трав, чтобы сделать выращиваемый куст ещё более
заметным, и тем самым снижали конкуренцию бальзамина с другими растениями.
Кроме того, птицы старались выбирать для выращивания растения с самыми яркими
листьями. Подобные отношения сформировали характерный облик растения.
Этот вид растений хорошо разрастается при достаточном количестве света – в
участках нарушенного древостоя, а также по берегам рек. Листья бальзамина багряного
имеют яркую антоциановую окраску – они зелёные с многочисленными красными пятнами,
а самые молодые листья полностью рубиново-красные. Этот вид отличается быстрым
ростом и развитием: от прорастания семечка до цветения проходит менее четырёх
месяцев. Если растение развивается без заботы со стороны птицы, то за это время
оно вырастает до метра в высоту и сравнительно мало ветвится, а его листья
в нижней части стебля постепенно опадают. Зато при заботе со стороны шалашника-садовника
облик растения совершенно иной: птица выщипывает клювом верхушку растения,
и оно начинает обильно ветвиться, превращаясь в шаровидный куст высотой около
40 – 50 см. Благодаря ветвлению у него большое количество точек роста, и яркий
красный цвет молодых листьев хорошо заметен на фоне окружающей зелени.
Но симбиоз с шалашником имеет обратную сторону: бальзамины принадлежат к числу
насекомоопыляемых растений, а присутствие птицы-симбионта отпугивает насекомых-опылителей,
либо птица просто ловит и съедает их. Поэтому цветение бальзамина багряного
происходит исключительно по ночам. В пазухах верхних листьев появляются цветочные
бутоны. Они долго развиваются, прежде чем раскроются всего лишь на одну ночь.
Их цветоножка свёрнута в спираль, обернув собой бутон. Ночью цветоножка начинает
быстро раскручиваться, и за час выпрямляется полностью, вынося бутон над листьями.
Цветок с длинным шпорцем и ослепительно-белыми лепестками раскрывается, источая
сильный сладкий аромат с лёгким гнилостным оттенком, привлекая ночных бабочек.
Растение выделяет большое количество жидкого нектара, и бабочки охотно посещают
его цветы. Если цветок остался неопылённым, в течение дня он буквально расплывается
в слизь, и цветоножка отваливается. Часто птица, убирая мусор, выдёргивает
и выбрасывает такие цветоножки. Если опыление произошло, цветоножка снова скручивается,
и под прикрытием листьев начинает вызревать плод – коробочка с растрескивающимися
створками. К моменту созревания семян цветоножка выпрямляется и вытягивается
горизонтально. При случайном прикосновении птицы она с треском лопается и разбрасывает
семена на расстояние нескольких метров.
Брюхоствольник
безлистный (Gastrocaudex aphyllus)
Порядок: Зонтичные (Apiales)
Семейство: Зонтичные, или Сельдерейные (Apiaceae)
Место обитания: Юго-Восточная Европа, Балканы.
В неоцене Африка вплотную приблизилась к Европе. Это вызвало целый ряд изменений
в природе Средиземноморья. В первую очередь, итогом этого медленного, но неуклонного
движения было исчезновение Средиземного моря – Гибралтарский пролив закрылся
навсегда, и за несколько тысяч лет море полностью высохло. На западной части
солёной низменности, некогда бывшей дном Средиземного моря, раскинулись суровые
пустыни. Восточная часть Средиземноморья – это мир солёных болот, где могут
жить только существа, устойчивые к соли. Эти болота питаются ручьями, стекающими
весной с Альп. Второе последствие движения Африки на север – это вулканическая
активность в Южной Европе, и рост в высоту Альп. Им ещё далеко до Гималаев,
но первый шаг уже сделан, и в течение следующих миллионов лет Альпы могут стать
высочайшими горами планеты.
Узкая полоса между южными склонами Альп и Средиземноморскими болотами – засушливая
равнина. Соль отступающего моря осталась в болотах, а Альпы преграждают путь
влажным северным ветрам. Вода попадает в эти места зимой в виде туманов, весной
в виде талых вод с горных вершин. В это время на несколько недель здесь развивается
богатая эфемерная растительность, которая большую часть года проводит в виде
семян, клубней и луковиц. Но некоторые растения не боятся холодных туманов
и обжигающего летнего зноя.
На глинистой почве растёт гротескное растение, в чём-то похожее на вымершие
растения палеозоя и раннего мезозоя. Оно не собирается прятаться от солнца
и отличается высокой специализацией к экстремальным условиям обитания. Толстые
неветвистые стебли этого растения торчат из скрытого под землёй корневища,
достигают высоты 3 – 4 м при толщине у основания около 20 см. Стволы слабо
изогнуты и совершенно лишены листвы, а междоузлия вздуты. Это растение называется
брюхоствольник безлистный. Растение похоже на Ottoa oenanthoides, растение,
обитавшее в Андах (Венесуэла) на высоте более 4 километров над уровнем моря,
и относится к одному с ним семейству. Условия обитания обоих видов сходны в
одном – они отличаются сухостью. В горах сухость физиологическая – из-за холода
растения хуже всасывают воду. А в пустыне жарко, и ощущается фактическая нехватка
воды.
Чтобы экономить драгоценную влагу, листья брюхоствольника редуцированы. Они
представлены широкими черешками без листовой пластинки, с развитыми прилистниками,
и существуют весьма непродолжительное время. Черешки и прилистники тонкие,
свёрнутые в колпачок. Они служат для защиты точки роста, прикрывая её от палящих
лучей солнца. По мере нарастания стебля эти колпачки быстро отмирают, но под
ними постоянно образуются новые. Стволы брюхоствольника имеют прочную кору
и заполнены рыхлой водоносной тканью. В пустыне встречаются некоторые виды
травоядных животных, например, свистороги. Но растение надёжно защищено от
них популярным среди зонтичных оружием – жидкость, запасённая в его стебле,
ядовитая и очень горькая. Брюхоствольник не вооружился колючками, как кактусы
или молочаи, но химическая защита обеспечивает его выживание.
Стволы этого растения отрастают от короткого реповидного корневища. Сигналом
для роста служит увлажнение почвы. Весной с Альп стекают ручьи, и в пустыне
образуются небольшие временные водоёмы. Растение словно стряхивает с себя зимнее
оцепенение, и начинает бурный рост. Каждый стебель растёт несколько лет, постепенно
утолщаясь. В узлах стебель не утолщается, поэтому его стволы кажутся перетянутыми
поперёк. Молодой стебель имеет тонкую кору и рыхлую ткань внутри. Концентрация
ядовитых веществ в нём особенно велика: от коры молодого стебля исходит горьковатый
запах, предупреждающий травоядных о том, что растение защищено ядом. Активный
рост продолжается на протяжении трёх – четырёх лет жизни стебля. Когда он вызревает,
кора твердеет и становится более сухой, зато клетки сердцевины наливаются водой
и в них накапливаются ядовитые вещества. В первые годы жизни каждый стебель
накапливает питательные вещества, а на 6 – 8-й год существования зацветает.
Соцветие брюхоствольника характерно для всех зонтичных – это крупный (около
80 см в диаметре) зонтик, состоящий из множества мелких белых цветков. Он поднимается
над стеблями на невысоком толстом цветоносе. Цветки опыляются насекомыми. Они
не ядовиты, и от них исходит приятный аромат. Семена брюхоствольника колючие,
их оболочка покрыта многочисленными крючочками. Они разносятся птицами, прицепляясь
к их оперению. После созревания семян ствол высыхает и отламывается у основания.
Брюхоствольник безлистный живёт до 30 – 40 лет, зацветает на десятом году жизни
и цветёт ежегодно до самой смерти.
Несмотря на ядовитость, это растение даёт пищу и кров некоторым видам пустынных
насекомых. На молодых стволах брюхоствольника паразитирует водоносная
тля.
Она не боится ядов, содержащихся в растении, может нейтрализовывать их. Это
насекомое накапливает яд в теле и обильно выделяет почти чистую воду. Это насекомое
служит как бы естественным «фильтром» яда и источником воды для некоторых насекомых.
Популяции этой тли «пасёт» жук-пастушок. Он слизывает воду, выделяемую тлями,
и благодаря этому выживает в пустыне. Этот жук оберегает тлю от других насекомых,
кормится на соцветиях брюхоствольника и опыляет цветки.
Муравьиный
корень средиземноморский (Myrmiradix mediterraneus)
Порядок: Зонтичные (Apiales)
Семейство: Зонтичные, или Сельдерейные (Apiaceae)
Место обитания: пустыни Средиземноморской котловины, маквис Южной и Западной
Европы.
В процессе эволюции растения очень часто вступали в союз с насекомыми. Наиболее
распространённой формой отношений между растениями и насекомыми является приспособленность
того или иного вида растений исключительно к одному виду насекомых-опылителей.
Менее распространённым явлением является мирмекофилия – образование симбиоза
с муравьями. В обмен на кров и пищу, предоставляемые своим шестиногим постояльцам,
растения получают надёжную защиту.
Часто симбиоз растений и муравьёв возникает в тропических лесах, где конкуренция
и опасность со стороны травоядных животных особенно велики. Но изредка такое
содружество возникает в гораздо менее благоприятных условиях обитания. Один
из случаев такого симбиоза возник в засушливых районах Средиземноморской котловины,
где травоядных животных сравнительно немного, но повреждение побегов в очень
неблагоприятных условиях обитания может значительно ослабить растение. Это растение
– средиземноморский муравьиный корень, представитель зонтичных.
Внешность этого вида растений мало отличается от типичного облика его родственников
– высокие полые стебли, одиночные широкие листья и очень характерные соцветия-зонтики
мелких невзрачных цветков. Листья средиземноморского муравьиного корня имеют
толстую центральную жилку и листовую пластинку, разделённую на три округлых
доли с притупленным кончиком. Верхняя поверхность этого листа покрыта тонким
слоем воска, отражающего часть ультрафиолетовых лучей. Нижняя сторона листа
густо покрыта мелкими белыми волосками, что создаёт благоприятный микроклимат
в тонком слое воздуха возле самой поверхности листа. Стебли этого растения достигают
метровой высоты при толщине около 5 – 6 см у основания. При отмирании стеблей
их ткани быстро высыхают и сохраняются в вертикальном положении почти весь следующий
год. За год растение способно образовать до трёх полноценных цветущих стеблей.
Соцветие диаметром до 20 см состоит из 150 – 200 мелких цветков. Цветение начинается
ранней весной и продолжается около месяца. Когда плоды созревают, цветоножки
неоплодотворённых цветков выпрямляются и скручиваются в виде колючек, защищая
семена от поедания птицами. Семена средиземноморского муравьиного корня распространяются
ветром. Для этого каждое семечко имеет широкий кожистый парус, окружающий его
по краю.
Главная особенность этого растения – толстое подземное корневище. Оно достигает
длины свыше 1 метра при диаметре до 7 – 9 см, разветвляется на несколько отростков
и лежит в почве на глубине до полуметра. Оно пустотелое, разделено на камеры
внутренними поперечными перегородками. Растение само по себе не ядовито, но
от травоядных млекопитающих и насекомых у него есть другая защита: муравьи.
Средиземноморский муравьиный корень предоставляет место для жизни нескольким
видам муравьёв – в условиях, где обитает это растение, нецелесообразно становиться
слишком узким специалистом. Фактически, его корень и прошлогодние стебли представляют
собой готовое обиталище для муравьёв. Они лишь пробуравливают проходы в перегородках
и окружают корневище растения своими тоннелями, улучшая циркуляцию воздуха.
Тот факт, что средиземноморский муравьиный корень является настоящим мирмекофильным
растением, подтверждается ещё одной особенностью – растение постоянно снабжает
муравьёв некоторым количеством пищи. На внутренних стенках корневища время от
времени выступают капли жидкости, которую муравьи пьют, а по краям листьев образуются
крахмалистые выделения, которые собирают муравьи. Растение хорошо растёт без
муравьёв, но в этом случае его листья сильно повреждаются насекомыми.
Молодое растение впервые зацветает только на пятом году жизни. Продолжительность
жизни может достигать 20 лет и более.
Пустынная
морковь (Daucus aridophilus)
Порядок: Зонтичные (Apiales)
Семейство: Зонтичные, или Сельдерейные (Apiaceae)
Место обитания: пустыни Ближнего Востока.
Как правило, культурные растения, оставшиеся без попечения со стороны человека,
оказывались нежизнеспособными. Многовековая селекция привела к слишком сильной
зависимости большинства видов культурных растений от человека, и после его исчезновения
лишь немногие виды оказывались в состоянии конкурировать с дикими видами, которые
в эпоху человека были лишь нежелательными сорняками в культурных посадках. Но
с другой стороны, культурные растения благодаря человеку расселились по всей
Земле и смогли получить преимущества в выживании в некоторых местообитаниях.
Один из таких видов – морковь (Daucus carota). В пустынях по окраинам Средиземноморской
котловины обитает один из потомков этого вида – пустынная морковь.
Обитание в суровых условиях пустыни значительно повлияло на облик растений этого
вида. У пустынной моркови крупные корнеплоды (длиной около 80 см при толщине
до 10 см), глубоко уходящие в землю. Благодаря длинным корням растение достигает
уровня грунтовых вод, прочно заякоривается в почве и постепенно заглубляется
в неё с каждым годом, пока не приходит время цветения. В начале сухого сезона
корни подсыхают, слегка скручиваются и втягивают растение глубже в почву. В
первые два года жизни корень очень тонкий, но к моменту цветения сильно утолщается
и накапливает много питательных веществ. Сахар придаёт мякоти отчётливый сладковатый
вкус. Сама мякоть корня у этого растения бледнее, чем у культурного предкового
вида, и имеет бледно-жёлтый цвет. В сухой сезон почка защищена листом с сильно
редуцированной пластинкой и расширенным черешком.
До момента цветения растение ежегодно образует розетку сильно рассечённых листьев
на жёстких вертикальных черешках. В год, предшествующий цветению, размер листьев
максимальный – до метра в длину на черешке длиной около 50 см. Пустынная морковь,
подобно своему предку, является монокарпиком и зацветает в возрасте 4-6 лет.
Цветонос этого растения достигает высоты 3-4 метра, соцветие из мелких жёлтых
цветков – зонтик диаметром около 60 см. Семена этого вида покрыты мелкими крючками
и колючками, переносятся птицами и летучими мышами, останавливающимися на растении
на отдых. Семена долго сохраняют всхожесть и прорастают весной в период дождей.
У молодого растения быстро отрастает главный корень, повреждение которого в
это время приводит к гибели растения.
Морковь
прерий (Daucus praeriensis)
Порядок: Зонтичные (Apiales)
Семейство: Зонтичные, или Сельдерейные (Apiaceae)
Место обитания: прерии Северной Америки.
Подобно большинству одомашненных животных, одомашненные растения не пережили
человека. Многие из их видов и сортов исчезли без следа, другие оставили совсем
немного потомка. После миллионов лет жизни в дикой природе потомки бывших культурных
растений лишь отдалённо напоминают своих домашних предков: это справедливо как
для растений, так и для животных. Один из таких потомков – это степная морковь,
потомок моркови обыкновенной, широко распространённого культурного растения.
Степная морковь произрастает в прериях Северной Америки и представляет собой
розеточное травянистое растение средних размеров, около 50 см в высоту. Листья
этого вида удлинённые, перисторассечённые на тонкие доли. Растение светолюбиво,
раскидывает розетку своих листьев поверх окружающих трав. Зелень растения богата
эфирными маслами и имеет характерный острый запах.
Корнеплод этого растения значительно деградировал по сравнению с культурным
предком. Он гораздо тоньше, чем у предка, длинный, волокнистый и горький на
вкус. Тем не менее, разные звери охотно выкапывают и поедают это растение. Медленно
пережёвывая растение, животные размазывают разжёванную мякоть по шерсти, что
помогает отогнать некоторых кровососущих насекомых, или просто валяются по зарослям
растения.
Морковь прерий – типичное двулетнее растение, зацветающее на втором году жизни.
Цветы мелкие, собраны в широкие белые зонтики, опыляются в основном разными
двукрылыми и реже жуками и осами. Запах цветков достаточно неприятный. Тёмные
семена снабжены многочисленными крючковатыми щетинками и созревают в плодах-вислоплодниках.
Их распространяют крупные млекопитающие и птицы: семена цепляются за их покровы
тела и разносятся далеко от родительского растения.
Этот вид растений открыл Bhut, участник форума.
«Жабье
дерево» (Bufodendron toxicus)
Порядок: Зонтичные (Apiales)
Семейство: Аралиевые (Araliaceae)
Место обитания: Южная Европа, побережье бывшего Средиземного моря.
В неоцене Европа и Северная Африка стали составлять единое целое – Африка на
протяжении всего кайнозоя медленно дрейфовала на север, и постепенно соединилась
с Европой. Средиземное море высохло ещё в предыдущую ледниковую эпоху, и его
котловина, занятая солёными болотами, в неоцене постепенно сужается. Через миллионы
лет она может исчезнуть полностью, превратившись в горную страну, но в неоцене
эта местность довольно обширна.
Выше солёных болот располагается пояс сухой каменистой полупустыни с отдельными
участками солончаков. Пустыня отличается жарким и сухим климатом, но растения
освоили даже такую негостеприимную местность.
Кое-где в пересохших руслах ручьёв, стекающих с Альп весной, растёт необычное
растение. Это толстое дерево, похожее на представителей рода Adenium, живших
в эпоху человека. Ствол этого растения короткий – растение не превышает двух
метров в высоту. Зато он толстый и коренастый, достигает почти метровой толщины
в основании. В верхней части от ствола отходят несколько таких же толстых ветвей,
искривлённых вверх и в стороны. Корни этого растения также очень толстые, клубневидно
вздутые в основании. Основания корней, достигающие метровой ширины, окружают
ствол дерева, поэтому кажется, будто дерево «растаяло» от иссушающей жары на
каменистой почве, словно гигантская свеча. Это растение называется «жабье дерево».
Родственные связи этого вида с растениями эпохи голоцена необычны: «жабье дерево»
является суккулентным потомком типичного европейского вида – плюща (Hedera helix).
Столь резкий переход одной жизненной формы в другую демонстрировали представители
рода Cissus семейства виноградных. Большинство видов этих растений – лианы,
но Cissus macropus является ярко выраженным суккулентом из Южной Африки с клубнеобразным
стеблем.
«Жабье дерево» накапливает большое количество воды в рыхлой древесине и основаниях
корней. Но дерево вовсе не расположено делиться ею с кем-либо: сок «жабьего
дерева» ядовит, и это защищает растение от посягательства травоядных животных.
Кора этого растения очень рыхлая и мягкая, светло-серая с тёмными бугорками,
внешне напоминающая жабью кожу. Такое строение коры характерно для ряда растений,
обитающих в подобных местах – кора плохо проводит тепло и спасает ствол от перегрева,
а зимой спасает растение от редких заморозков. Растение образует побеги двух
типов: многолетние скелетные ветви и тонкие однолетние побеги, отмирающие и
опадающие с наступлением сухого сезона. Скелетные ветви нарастают медленно –
метровое растение превышает по возрасту 50 лет, а двухметровые гиганты прожили
уже по полтора века. Это дерево очень долговечно – его возраст может достигать
400 лет и больше.
Более явно родство «жабьего дерева» с плющом демонстрируют листья и цветы. Листья
этого растения отрастают с первыми весенними дождями и держатся на растении
всего около трёх месяцев в году. Листья «жабьего дерева» бывают двух типов:
они отличаются на цветоносах и вегетативных побегах. На вегетативных побегах
листья пальчато-расстечённые, с тонкими долями. Форма листа – это показатель
развития и самочувствия растения. На здоровом и сильном растении листья пяти-
и даже семилопастные. А молодое или очень старое растение обычно образует тройчатые
листья.
В возрасте около 20 лет растение впервые в жизни зацветает. «Жабье дерево» цветёт
зимой, до распускания листьев. В это время с Альп движутся холодные воздушные
массы, и по утрам на места произрастания этого растения оседает густой туман.
Холодный период стимулирует развитие цветочных почек, а туман служит стимулом
для цветения. Растение цветёт и плодоносит рано, чтобы к сухому сезону его плоды
успели созреть и проростки смогли достаточно развиться для того, чтобы пережить
засушливый сезон.
Присоцветные листья отличаются от обычных: они овальные с притупленным кончиком.
Они густо сидят на коротких цветоносных побегах, образуя обвёртку для соцветия,
похожую на корзинку сложноцветных. Невзрачные цветы с сильным запахом собраны
в густое щитковидное соцветие. Их пряный запах привлекает мелких жуков, сохраняющих
активность даже в зимнюю прохладу. В это время источники питания очень редки,
поэтому насекомые надолго задерживаются на соцветиях «жабьего дерева» и ночуют
на них же, забившись в глубину кисти. Плоды этого растения – круглые сизые ягоды,
покрытые восковым налётом. Они созревают весной, к моменту пролёта птиц из Африки.
Это растение специализировано к переносу семян птицами: воск отражает ультрафиолетовый
свет, и спелые плоды хорошо заметны для птиц. Кроме того, ягоды ядовиты для
зверей, изредка появляющихся в местах обитания этого растения. Птицы охотно
поедают ягоды и разносят семена по окрестностям. Пока в почве достаточно влаги,
семена прорастают. Молодое растение образует розетку остроконечных листьев и
даёт длинный стержневой корень. Оно растёт в высоту очень медленно: в первый
год образует небольшой реповидный клубень и розетку из 4 – 5 листьев. На пятом
году жизни начинается рост стебля в высоту.
Эпихедера
рассечённолистная (Aepyhedera dissecta)
Порядок: Зонтичные (Apiales)
Семейство: Аралиевые (Araliaceae)
Место обитания: Юго-Восточная Европа, Балканы.
Тёплые субтропические леса обязаны своим появлением влиянию Четвероморья. Дожди
с этого водоёма дают достаточно влаги для роста лесов, а само Четвероморье
смягчает климат. Леса по берегам этого водоёма имеют некоторые черты влажного
тропического леса. Их составляет широколиственная флора, и в таких лесах наблюдается
обилие лиан и эпифитов. Вместе с тем, эти леса приспособлены к выживанию в
условиях выраженного сезонного климата.
Гигантская горная цепь затрудняет связь субтропических лесов с тропическими
лесами Азии и Африки. Поэтому в субтропиках Четвероморья у местных растений
появились черты, характерные для настоящих субтропических растений южного происхождения
– крупные размеры, широкие или рассечённые листья, длинные кончики листьев
– «капельницы», приспособления для отвода избытка влаги.
Среди лиан европейских лесов доминируют потомки плюща нескольких видов. Одни
из них остались холодостойкими, как их предок, вечнозелёными лианами с прочными
кожистыми листьями. Они распространились далеко на север и обвивают стволы
деревьев практически по всей Европе, кроме Скандинавии и северо-востока. Северные
виды – это многолетние мелколистные лианы с тонким одревесневающим стеблем.
Гигантский потомок некогда небольшой лианы, плюща (Hedera helix), является
одной из доминирующих лиан в южных субтропических лесах Европы. Южный потомок
плюща, эпихедера рассечённолистная, отличается от мелких северян – это роскошная
вечнозелёная деревянистая лиана с толстым стволом, грубой корой и корнями-прицепками,
растущими на всём протяжении ствола. С их помощью толстый стебель этого растения
(“Aepyhedera” означает буквально «сверх-плющ») прочно удерживается на стволах
деревьев. Она является многолетним растением, и достигает возраста 200 лет
и более. Возраст отдельных лоз этого вида может быть лишь немногим меньше,
чем возраст окружающих деревьев. Эпихедера – гигант среди лиан: у взрослого
растения может развиваться до десяти побегов длиной более 70 метров, опутывающих
несколько соседних деревьев. Основания стволов лежат на земле, переплетаясь
с корнями деревьев и делая участки леса, где они растут, труднопроходимыми.
Они покрыты толстой корой, которая отщепляется отдельными пластинками. На стволе
развиваются не только короткие корни-прицепки, но и длинные воздушные корни,
которые свободно свисают вниз и достигают большой длины. Врастая в почву, они
утолщаются, древеснеют, и доставляют растению дополнительное питание.
Листья эпихедеры пальчатые, округлой формы. По мере развития растения форма
листьев меняется. У молодого растения листья мелкие, овальные. Затем они становятся
пятиугольными, напоминающими листья плюща. По мере роста листья постепенно
становятся лопастными, и выемки между долями листа увеличиваются. Далее увеличивается
размер листа, и появляются дополнительные доли. У полностью развившегося растения
лист достигает диаметра 70 см и несёт 7 – 9 долей. Кончик каждой доли вытянут
в «капельницу», помогающую избавиться от избытка влаги.
Достигнув стадии полной физиологической зрелости, эпихедера зацветает. Перед
цветением она вновь меняет форму листьев. На стволе начинают развиваться цветоносные
побеги, покрытые цельными листьями, имеющими удлинённую форму и «капельницу»
на конце. Цветы эпихедеры мелкие, белые с желтоватым оттенком, в широких щитковидных
соцветиях. Их запах густой и тяжёлый, может вызвать головную боль. Из-за этой
особенности лазающие и летающие млекопитающие не приближаются к цветкам этого
растения во время его цветения. Зато аромат цветов эпихедеры очень привлекателен
для крупных жуков и одиночных пчёл, которые опыляют это растение. С конца лета
эпихедера плодоносит. Её плоды – синевато-фиолетовые ягоды размером до 5 см,
покрытые тонким слоем воска. Они ядовиты для млекопитающих, но птицы без вреда
поедают их и разносят семена. Одни из основных распространителей семян эпихедеры
– пещерные голуби (Cavernicolumba
strigops), гнездящиеся в пещерах Балкан.
Женьшеневое
дерево (Megalopanax flabelliphyllum)
Порядок: Зонтичные (Apiales)
Семейство: Аралиевые (Araliaceae)
Место обитания: Юго-Восточная Азия, Берег Джакарта, дождевые леса.
В эпоху человека Юго-Восточная Азия была одним из центров разнообразия растений
семейства аралиевых. Эти растения образуют разнообразные жизненные формы – лианы,
многолетние травы, а также небольшие деревья. Когда область распространения
тропических лесов расширилась, некоторые представители аралиевых оказались в
условиях жёсткой конкуренции с растениями других групп. Среди аралиевых эпохи
неоцена не появилось крупных видов деревьев, достигающих возраста нескольких
сотен лет. Но они весьма преуспели в качестве пионерной растительности на повреждённых
участках леса.
Там, где упал подточенный временем лесной патриарх, или где река прорезает лес,
в пологе леса образуется обширное «окно», и солнечный свет достигает поверхности
земли. Здесь начинается рост крупных лесных трав, а также сеянцев деревьев различных
видов, которые конкурируют между собой за право развиться и захватить этот участок
леса. Среди этих видов, процветающих в течение немногих лет, встречается один
из представителей древовидных аралиевых – женьшеневое дерево.
Этот вид растений имеет довольно характерный облик. Женьшеневое дерево – быстрорастущее
дерево с длинными колючками на стволе, высотой около 15 метров. Такой высоты
дерево достигает в течение примерно шести лет. Ствол этого растения обычно не
ветвится; лишь при повреждении вершины начинают развиваться 2 – 3 боковых ветки.
Чаще всего одна наиболее сильная ветка продолжает расти, а остальные засыхают
и отламываются. Зато в основании ствола есть большое количество почек, из которых
развиваются отпрыски. Кора этого растения лаково-коричневая, блестящая, на молодых
побегах с красноватым оттенком. В нижней части ствола старая кора слоится и
шелушится длинными полосками между шипами. Шипы, служащие для защиты от травоядных
животных, чёрные с белыми кончиками. На нижней части ствола развивается большое
количество толстых придаточных корней, служащих для опоры.
Листья женьшеневого дерева также имеют очень характерные очертания. Они одиночные,
глубоко пальчаторассечённые на 15 – 19 длинных долей с округлым концом, резко
переходящим в тонкую «капельницу». Листья огромные: они достигают двух с половиной
метров в диаметре. Листовые пластинки сидят на длинных упругих черешках (длиной
до 5 метров) и образуют зонтиковидную крону, характерную для древовидных аралиевых.
Поверхность листьев прочная и кожистая, поэтому отмирающий лист надолго сохраняет
форму и разлагается сравнительно медленно. Полностью развитый лист имеет тёмно-зелёный
цвет с белыми жилками, молодой лист красноватый: это антоциановая окраска, защищающая
от избытка солнечного излучения.
Женьшеневое дерево относится к числу влаголюбивых растений. Оно произрастает
по берегам рек, где лесной полог нарушается естественным образом. Особенно часто
этот вид произрастает на речных отмелях и песчаных косах. Этот вид относится
к числу пионерной растительности, он светолюбив и малоконкурентоспособен. По
мере поселения других видов растений женьшеневое дерево деградирует и быстро
вытесняется другими деревьями. Главное преимущество этого вида в борьбе за существование
– его нетребовательность к питательным веществам. Семена женьшеневого дерева
прорастают, и в дальнейшем растение нормально развивается практически на песке,
довольствуясь минимальным количеством органических веществ.
В тканях женьшеневого дерева синтезируется большое количество биологически активных
веществ. Они обладают тонизирующим воздействием и повышают половую активность
у самцов многих млекопитающих. Поэтому заросли женьшеневого дерева постоянно
несут ущерб от различных животных, которых не останавливают даже крепкие шипы
на стволе дерева. Лазающие животные поедают листву дерева, а массивные наземные
звери обгладывают кору или сваливают молодые деревья, чтобы поесть листвы. Даже
хищники в брачный сезон не обходят стороной этот вид деревьев.
Женьшеневое дерево цветёт ежегодно начиная с третьего года жизни. В пазухах
листьев развиваются повисающие зонтиковидные соцветия, состоящие из невзрачных
бледных цветков, опыляемых мухами и жуками. Плоды этого растения – мелкие, но
многочисленные ягоды синего цвета с сизым восковым налётом. Их охотно поедают
птицы (особенно голуби и попугаи), а упавшие ягоды отыскивают звери. Ягоды также
обладают тонизирующим действием. Семена этого растения распространяются животными.
Сандал-душитель
(Odoroxylon scandens)
Порядок: Санталоцветные (Santalales)
Семейство: Санталовые (Santalaceae)
Место обитания: тропические леса Южной Азии.
Семейство санталовые наиболее богато представлено в тропических лесах. В него
входят растения, ведущие частично или полностью паразитический образ жизни.
Несколько видов этих растений пострадало от вырубки человеком (по крайней мере
один вид был полностью уничтожен) из-за ценной ароматической древесины. После
исчезновения на Земле человека и глобального потепления, когда площадь тропических
лесов выросла, растения из этого семейства вернули утраченные позиции, образовав
новые своеобразные виды.
Сандал-душитель – очень своеобразная лиана-полупаразит. Это растение достигает
внушительных размеров: длина его горизонтально растущего ствола может доходить
до 90-100 метров. Растение даёт множество боковых побегов и разрастается в ширину
на десятки метров, прирастая к большинству подходящих для роста деревьев в окрестностях.
Растение начинает свою жизнь как эпифит: птицы или насекомые заносят его семена
в развилку ветвей на каком-нибудь крупном дереве. Проросток первые несколько
дней развивается за счёт запасных питательных веществ, полученных от материнского
растения. Но его корни постепенно начинают буравить кору дерева, на котором
поселилось растение. Если дерево оказывается не подходящим на роль растения-хозяина,
сандал-душитель может долгое время развиваться как эпифитная лиана. Он формирует
побеги, тянущиеся к соседним деревьям. Достигнув соседнего дерева, побег образует
несколько придаточных корней, проникающих сквозь кору дерева.
Если же молодому растению повезло, либо если побег нашёл дерево, удовлетворяющее
потребностям сандала-душителя, в местах контакта с корой дерева образуются сосущие
придаточные корни, буравящие кору дерева и внедряющиеся в его древесину. Благодаря
выделяемым фитогормонам место срастания формирует своеобразный «узел», в котором
«переплетается» древесина дерева-хозяина и этой лианы. На срезах таких «узлов»
образуется интересный рисунок, особенно эффектный, если древесина паразита и
хозяина различается по цвету. У сандала-душителя древесина имеет очень тёмную,
почти свекольно-красную окраску. Она богата эфирными маслами и приятно пахнет.
Масла защищают растение от насекомых-древоточцев. Ствол сандала-душителя, получивший
щедрый приток дополнительного питания, начинает расти в длину и сильно утолщаться.
Боковые побеги тянутся к новым деревьям, и вскоре большой участок леса оказывается
оплетённым одним крупным растением. Его легко заметить среди ветвей: кора сандала-душителя
очень светлая, почти белая.
В отличие от некоторых своих родственников, полностью перешедших к паразитическому
существованию, сандал-душитель зависит от дерева-хозяина лишь частично. От него
он получает только воду и минеральные соли, которых обычно не хватает эпифитным
растениям. У этого вида листья не утрачены, растение способно к самостоятельному
фотосинтезу. От горизонтальных стволов сандал-душитель даёт многочисленные вертикальные
фотосинтезирующие побеги с характерными овальными листьями. В трёх-пятилетнем
возрасте сандал-душитель зацветает. На концах фотосинтезирующих побегов развиваются
вертикальные соцветия-кисти из множества мелких розоватых цветков с сильным
приятным запахом. Они опыляются насекомыми – в основном жуками и мухами, хотя
иногда их посещают осы и одиночные пчёлы. Растение однодомное, но мужские цветки
распускаются лишь после отцветания женских. Плоды – белые сладковатые ягоды
с тонкой кожицей и одним крупным семенем. Они распространяются птицами, иногда
опавшие ягоды поедают грызуны.
Аромоксилон
лазающий (Aromoxylon scandens)
Порядок: Санталоцветные (Santalales)
Семейство: Санталовые (Santalaceae)
Место обитания: Юго-Восточная Азия, тропические леса.
В эпоху человека сандаловые деревья имели важное значение как источник драгоценной
древесины, что привело к значительному сокращению их численности и к вымиранию
ряда видов, особенно узкоареальных эндемиков. Сокращение площадей тропических
лесов приводило к вымиранию узкоспециализированных полупаразитических видов
этих растений, и к выживанию преимущественно малоспециализированных форм, способных
паразитировать на большом количестве видов растений. В неоцене, при стабилизации
природных условий и расширении площади лесов, среди растений семейства сандаловых
проявились две стратегии: сужение специализации при повышении степени приспособленности
к паразитизму, и, напротив, расширение спектра видов-хозяев при сохранении относительно
слабой специализации к паразитическому образу жизни.
В тропиках Юго-Восточной Азии обитает полупаразитический вид сандаловых – аромоксилон
лазающий, очень крупная одревесневающая лиана, разрастающаяся в пологе леса.
Аромоксилон начинает свою жизнь как маленькое лиановидное растение подлеска.
Он не теряет зелёные листья, и способен много лет развиваться в подлеске, подобно
проросткам многих тропических деревьев. Ползучий стебель выбирает направление
роста, ориентируясь по химическим веществам, выделяемым в почву корнями и опавшими
листьями подходящих видов растений-хозяев. Добравшись до привлекательного дерева,
лиана начинает ползти по его стволу, прикрепляясь придаточными корнями к его
стволу и ветвям. В местах контакта корни начинают буравить древесину дерева-опоры,
и там со временем развиваться наплывы из смеси тканей дерева и лианы. Растение
получает от дерева-хозяина воду и минеральные вещества, и благодаря этому достигает
гигантских размеров. Взрослое растение может отмирать в нижней части и расти
в кронах деревьев исключительно как полупаразит, всецело завися от них в плане
снабжения водой и минеральными веществами. Несмотря на это, растение сохраняет
листву и нормальную способность к фотосинтезу. Срастаясь с проводящей системой
дерева-хозяина, растение снабжает его сахарами и другими продуктами фотосинтеза.
Стебель взрослого растения этого вида достигает толщины 15-20 см, покрыт тонкой
и гладкой серой корой. Древесина обладает сильно выраженным характерным приятным
ароматом. Листья простые, остроконечные, с кожистой поверхностью и ровным краем.
Цветки очень мелкие, невзрачные, собраны в кисти, развивающиеся на молодых ветвях.
Плод – крупная костянка с ярко-жёлтой кожурой и сочной мякотью, содержащая одно
крупное семя. Семена распространяются главным образом крупными наземными млекопитающими,
которых привлекает сильный аромат созревших и опавших плодов. Хотя плоды растения
охотно поедают птицы, они почти не играют роли как распространители плодов:
в желудках птиц оболочки семян растворяются.
Молодое растение зацветает примерно на 10-12-м году жизни. Продолжительность
жизни часто составляет до 200 лет и более. Часто растение переживает свои деревья-хозяева,
и существует за счёт соседних деревьев. При этом части растения, срастающиеся
со старым хозяином, отмирают вместе с ним, и дерево распадается на ряд клонов,
живущих в кронах новых деревьев-хозяев.
Кактусовая
лжеомела (Pseudoviscum cactorum)
Порядок: Санталоцветные (Santalales)
Семейство: Ремнецветниковые (Loranthaceae)
Место обитания: пустыни и полупустыни Мексики, паразит кактуса «старая дева».
В эпоху человека очень немногие кактусы были подвержены поражению растениями-паразитами.
В Южной Америке некоторые кактусы поражались растениями рода Tristerix из семейства
ремнецветниковых, но другие члены семейства жили на растениях других таксонов
в Южной и Центральной Америке. В ходе неоценового потепления один из таких неспециализированных
видов освоил новую нишу – он перешёл на паразитирование на центральноамериканских
кактусах, в том числе на кактусе
«старая дева».
Кактусовая лжеомела – узнаваемое и характерное растение-паразит, поражающий
значительную часть взрослых кактусов «старая дева». На поражённом кактусе видны
только проросшие сквозь слой «волос» пучки красных цветоносов (длиной около
10-15 см) с цветками, листья полностью отсутствуют. Стебель сильно укорочен,
а корни преобразованы в гаустории, находящиеся внутри ствола или ветки кактуса.
Цветки розовые, около 1,5 см в диаметре, у них 5-членная чашечка, 5 узких лепестков
раскрываются веером, в цветке 10 тычинок и 1 пестик. Цветки имеют развитые нектарники
и источают сладкий аромат.
Опыляется лжеомела насекомыми и птицами, питающимися нектаром. Большую роль
при этом играют колибри кактусовые
искорки. Плоды – круглые белые сочные костянки диаметром около 1 см, с очень
клейким соком. Семя заключено в плотную оболочку чёрного цвета. Семена разносятся
чаще всего птицами, реже другими животными, поедающими эти плоды. Клейкий сок
приклеивает семена к ногам и телу животных, когда они давят опавшие ягоды, и
они разносят их на себе на новые места. Если же птица (или иное древесное животное)
проглотит семя, плотная оболочка защитит его от переваривания, и оно выпадет
с экскрементами на новом месте.
Кактусовая лжеомела наносит растению-хозяину сравнительно небольшой вред и может
встречаться на нём в большом количестве: при обильном заражении кактус «старая
дева» обрастает паразитами и выглядит покрытым словно клочковатой красно-розовой
бахромой поверх своих «волос». «Волосы» защищают кактус лишь частично: семя
не прорастает на слое «волос», но животные часто придавливают его к самой поверхности
растения сквозь «волосы» или заносят под них. От одного места заражения гаустории
легко растут внутри кактуса в разные стороны, и новые пучки цветоносов пробиваются
по ходу их роста. Молодое растение зацветает уже на втором году жизни.
Этот вид растений открыл Биолог, участник форума.
Пламенеющее ядовитое дерево (Poinsettia arborea)
Порядок: Молочайные (Euphorbiales)
Семейство: Молочайные (Euphorbiaceae)
Место обитания: горные леса Гавайских островов.
В эпоху человека флора Гавайских островов понесла сильный ущерб. Ряд видов
растений вымер из-за исчезновения связанных с ними животных-опылителей. Некоторые
виды деревьев исчезли из-за вырубки, а другие пострадали из-за завезённых растительноядных
млекопитающих. Не меньший ущерб эндемичной флоре нанесло вселение на острова
экзотических растений с различных материков. «Закалённые» в более суровых условиях
жизни на материке, они быстро вытеснили значительное количество видов аборигенной
флоры. В неоценовой флоре Гавайских островов потомки завезённых человеком видов
растений занимают значительное место.
Склоны вулканов, образующих Гавайские острова, покрыты пышной растительностью.
Однообразие зелёного цвета листвы нарушается яркими красками цветущих растений.
Среди них особенно ярко выделяются кроны одного из видов неоценовой гавайской
флоры – красные с жёлтым, словно охваченные огнём. Это растение, вносящее яркие
краски в палитру тропического леса, является потомком пуансеттии, или «рождественской
звезды» (Euphorbia (Poinsettia) pulcherrima), завезённой человеком. Этот вид
называется пламенеющее ядовитое дерево, и его особенности оправдывают это название.
Пламенеющее ядовитое дерево – влаголюбивое растение, которое очень хорошо чувствует
себя во влажном воздухе ущелий гавайских гор. Оно особенно обильно во влажных
лесах восточных склонов гор. Обычно это полукустарник или невысокое дерево,
растущее в несколько стволов. Высота этого растения достигает 6 – 8 метров.
Кора пламенеющего ядовитого дерева серовато-коричневая, толстая и рыхлая. В
нижней части ствола она отслаивается длинными полосами. Листья этого растения
простые, длиной до 20 см, широкие, с крупными угловатыми боковыми лопастями.
Кончик листа вытянут в характерную «капельницу», которая помогает выводить
из тканей избыток воды. Сок этого дерева белый, как у большинства представителей
молочайных. Он содержит большое количество каучука, который при подсыхании
образует натёки и закупоривает пораненные места на стволе и ветвях. Сок не
так ядовит, как у некоторых материковых видов молочаев, но обладает очень жгучим
вкусом и сильным раздражающим действием. Это дерево защищается от древогрызущих
насекомых, заливая обильным млечным соком отверстия, которые они бурят в стволе.
Листва этого дерева также несъедобна, но некоторые животные Гавайских островов
специально едят её в небольших количествах, чтобы избавиться от паразитов.
Этот вид деревьев принадлежит к пионерной растительности островов, заселяющей
поля застывшей вулканической лавы после извержения. Когда вулканическая лава
затвердевает и среди неё разрастаются травянистые растения, пламенеющее ядовитое
дерево появляется одним из первых среди древесных растений. Оно очень быстро
растёт – молодое растение в течение года образует трёхметровый ствол и начинает
формировать крону. Ствол этого дерева в молодости четырёхрёберный, но с возрастом
рыхлая сердцевина разрушается, и в стволе появляются отверстия, часто сквозные.
При этом рост в толщину продолжается, края трещин покрываются корой, и ствол
более взрослых экземпляров напоминает несколько параллельно растущих стволов,
частично сросшихся друг с другом. У старых деревьев во время ураганов или просто
под тяжестью мокрой листвы после дождя ствол может разломиться на несколько
частей. Однако при этом дерево остаётся в живых, и части ствола, сохранившие
связь с корнями, продолжают расти – ветви изгибаются вверх и формируют придаточные
корни. Часто ствол ложится на склон, и дерево продолжает жить, давая новые
вертикальные побеги. Продолжительность жизни этого вида не превышает 100 лет.
Соцветия пламенеющего ядовитого дерева очень мелкие, как у всех типичных представителей
семейства. Соцветие – типичный для молочаев циатий: женский цветок с редуцированным
околоцветником окружён несколькими мелкими мужскими цветками. Но цветение этого
дерева не пройдёт незаметно для опылителей: соцветия окружены яркими присоцветными
листьями. Это растение опыляется различными птицами, которые, как и этот вид
растений, являются потомками видов, завезённых на Гавайи человеком. Присоцветные
листья пламенеющего ядовитого дерева определили название вида: они привлекают
птиц контрастной окраской, малиново-розовой с жёлтой каймой. Когда дует ветер,
присоцветные листья колышутся, создавая впечатление огня. Когда растение отцветает,
его прицветники теряют яркость – они становятся бурыми и постепенно приобретают
обычную зелёную окраску. Пламенеющее ядовитое дерево цветёт несколькими «волнами»,
одновременно развивая на вершинах молодых побегов большое количество соцветий.
Мелкие семена этого растения снабжены хохолком белых волосков. Они легко разносятся
ветром, и одними из первых попадают в трещины застывшей лавы на лавовых полях
островов. Это дерево быстро развивается, требуя минимальное количество гумуса,
а его корни глубоко проникают по трещинам в лаве. Но во взрослом состоянии
оно очень светолюбиво, и по мере зарастания лавовых полей этот вид постепенно
вытесняют другие растения. Также пламенеющее ядовитое дерево часто поселяется
на склонах гор, где конкуренция с другими растениями меньше, чем в долинах.
Эпимона
превосходная (Aepymona superba)
Порядок: Ясноткоцветные (Lamiales)
Семейство: Яснотковые (Lamiaceae)
Место обитания: восточная часть Северной Америки в области умеренного и теплоумеренного
климата.
Эпоха человека отмечена обширной селекционной работой с отдельными видами животных
и растений, которые приобрели новые свойства в одомашненном состоянии. Обычно
эти свойства оказывались вредными для выживания в дикой природе, поскольку их
следствием были изменения в физиологии и приобретение прочих неадаптивных признаков.
После краха промышленного сельского хозяйства и исчезновения человека многие
домашние растения и животные быстро вымерли. Однако в случае с декоративным
травянистым растением монардой (Monarda) селекционная работа подарила растению
ярко-красную окраску цветков, которая позволила потомкам садового растения приобрести
новых опылителей – колибри. Восстановление природных местообитаний позволило
одичавшим потомкам монарды расселиться, снабжая пищей мигрирующих колибри, и
эволюционировать в новые виды.
Потомком монарды в Северной Америке является эпимона превосходная – многолетнее
корневищное травянистое растение. Стебли мощные, четырёхгранные, покрыты короткими
белыми волосками, слегка одревесневающие снизу. В южной части ареала стебель
может продолжать рост до 2 лет подряд, достигая высоты 250-300 см. Отсюда название
растения: приставка aepy- означает превосходную степень, и -mona от названия
предка – Monarda. На севере ареала стебли однолетние, но высотой до 200 см.
Листья супротивные, длиной до 20 см, ярко-зелёные, удлинённые с сильно вытянутыми
кончиками. Жилки вдавленные, сильно выступают с нижней стороны листа.
Этот вид – влаголюбивое растение, которое предпочитает расти на влажных лугах
и слегка заболоченной почве. Во время наводнений растение нормально растёт и
цветёт, выдерживая затопление в течение нескольких недель.
Цветение начинается в конце весны и продолжается до середины лета; этот вид
является важным кормовым растением для мигрирующих колибри. Цветки крупные (длиной
до 8 см), трубчатые, узкие, слегка выгнутые, птицеопыляемые. Они ярко-красного
цвета снаружи, располагаются в верхней части стеблей мутовками в пазухах листьев.
В зеве цветков расположен узор из тёмных глазчатых пятен неправильной формы
с белой каймой, на нижней губе цветка тянутся продольные белые штрихи, направленные
вглубь зева цветка. Белые отметины образованы густыми прозрачными волосками,
отражающими ультрафиолетовые лучи – это метки-указатели пути к нектару. Резервными
опылителями растения являются шмели и крупные одиночные пчёлы, способные разглядеть
эти указатели и обладающие достаточно длинными хоботками, чтобы достать нектар
из глубины цветка.
Плод – сухая коробочка; семена лёгкие, способны плавать на поверхности воды.
Обычно они распространяются водой – во время летних наводнений или осенних дождей.
Молодое растение зацветает на третий год, достигает пика развития в возрасте
8-9 лет. Корневище может нормально развиваться до 15-20 лет.
Полярная
мята (Menthula densa)
Порядок: Ясноткоцветные (Lamiales)
Семейство: Яснотковые (Lamiaceae)
Место обитания: тундровые районы Евразии, Северная Европа, Полярный Урал, Таймыр,
острова Северного Ледовитого океана; каменистые пустоши.
В эпоху человека среди культивируемых растений появилось большое количество
гибридных форм. Многие из них вымерли после исчезновения человека, не выдержав
конкуренции с дикими видами растений, но некоторые гибридные растения оказались
достаточно жизнеспособными, чтобы выживать в дикой природе достаточно долгое
время и эволюционировать, становясь предками новых видов растений эпохи неоцена.
Среди флоры неоцена встречается достаточно много видов, предками которых были
гибриды, выведенные человеком в историческую эпоху. Один из таких видов – полярная
мята, потомок гибридной мяты перечной (Mentha piperita). Этот вид освоил жизнь
в негостеприимных районах тундры и лесотундры на севере Евразии, где достаточно
успешно конкурирует с прочими видами растений.
Полярная мята – это вечнозелёное травянистое растение с мелкими побегами, образующими
густую подушку. Растение является корневищным многолетником, подземные корневища
распространяются в разные стороны от материнского растения. Побеги высотой до
10 см, растут очень густо рядом друг с другом, ветвясь от основания. Листья
парные, округлые, диаметром до 2 см, морщинистые, с мелкозубчатым краем. Поверхность
листа покрыта короткими волосками, нижняя сторона обильно опушена. Надземная
часть растения богата ментолом, придающим зелени характерный мятный запах.
Цветки собраны в короткие шаровидные соцветия на концах побегов. Цветок трубчатой
формы, светло-сиреневый, покрыт волосками, отражающими ультрафиолетовый свет.
Данный вид растений опыляется одиночными пчёлами, шмелями, мухами и мелкими
бабочками.
Семена очень мелкие, покрыты крючковатыми щетинками, позволяющими зацепляться
за оперение птиц. Они вызревают относительно редко, и растение обычно размножается
при помощи ползучих корневищ. Зачастую все растения этого вида с какого-то острова
представляют собой всего лишь несколько крупных старых клонов.
Растение является важным кормовым ресурсом для травоядных животных – от насекомых
до млекопитающих. В некоторых местностях полярная мята – важнейшее медоносное
растение, поддерживающее существование популяции насекомых.
Руброкулярия
великолепная (Rubrocularia magnifica)
Порядок: Ясноткоцветные (Lamiales)
Семейство: Акантовые (Acanthaceae)
Место обитания: Центральная Америка, острова Карибского моря, Куба и Большая
Антигуа, тропические леса.
Для эпохи человека характерно значительное смешение представителей разных флористических
и фаунистических областей, и появление различных видов живых организмов в несвойственных
им областях Земли. Многие из переселенных людьми видов прижились на новом месте,
пережили эпоху глобального экологического кризиса и ледниковый период, и начали
эволюционировать в новых местах обитания, принимая участие в формировании новых
экосистем постчеловеческой эпохи.
В тропиках Центральной Америки произрастает потомок одного из сорных видов эпохи
человека – тунбергии крылатой (Thunbergia alata). Это растение – руброкулярия
великолепная, огромная лиана с широкими листьями и длинными трубчатыми цветками.
По сравнению с предком руброкулярия значительно изменилась внешне. Она превратилась
в мощную многолетнюю лиану, побеги которой достигают длины 30-40 метров. Нижняя
часть стебля этого растения сильно утолщается и одревесневает, покрываясь мелкотрещиноватой
серой корой, отпадающей чешуевидными пластинками. Растение обильно ветвится,
молодые побеги быстро растут. В процессе роста они совершают круговые движения
в воздухе, пока не наткнутся на опору. Коснувшись опоры, побег закрепляется
на ней, сделав несколько оборотов, и продолжает рост вверх. С возрастом побеги
утолщаются, и растение может медленно душить деревья с относительно мягкой древесиной.
Медленно растущие деревья с твёрдой древесиной, как правило, не испытывают ущерба
от роста этой лианы. На стебле активно развиваются воздушные корни, помогающие
растению прикрепляться к опоре и усваивать влагу из росы и дождей.
Листья крупные, длиной до 35-40 см, широкосердцевидные, с характерным тонким
кончиком-капельницей. Поверхность листа гладкая, тёмно-зелёная, с вдавленной
сетью крупных жилок.
Название растения описывает характерный облик его цветков, буквально означая
«красноглазка». Цветки этого растения трубчатые. Длина трубки около 10 сантиметров,
диаметр околоцветника – до 6-8 см. Околоцветник актиноморфный, пятилепестный.
Цветоносы развиваются в пазухах листьев на побегах старше полугода и несут по
2-3 цветка. Окраска околоцветника ярко-красная с оттенками от тёмно-розового
до почти бордового, внутренняя часть трубки чёрная. В зеве цветка между красной
и чёрной зонами располагается узкое белое кольцо – это указатель нектара для
опылителей. Руброкулярия опыляется птицами
и некоторыми бабочками,
цветение продолжается круглый год. Это важный кормовой ресурс для мигрирующих
колибри.
Плоды сухие, растрескивающиеся. При прикосновении створки плода скручиваются
и стреляют семенами на расстояние до 20 метров, а с вершин крупных деревьев
семена разлетаются на 30-40 метров. Всхожесть семян очень хорошая, но руброкулярия
– светолюбивое растение, и многие проростки гибнут в лесной тени. Растение особенно
хорошо разрастается на деревьях по берегам рек и на участках повреждённого древостоя.
Молодое растение достигает высоты 10 метров в первые 3 года жизни. В это же
время оно обычно зацветает. Продолжительность жизни растения редко превышает
15-20 лет.
Лиловое
дудчатое дерево (Tabebuia violacea)
Порядок: Ясноткоцветные (Lamiales)
Семейство: Бигнониевые (Bignoniaceae)
Место обитания: галерейные леса центральной и северо-восточной части Южной Америки.
Род Tabebuia семейства Bignoniaceae пережил голоцен, и в неоцене появился новый
вид – лиловое дудчатое дерево. Этот вид произрастает в саваннах, равнинах и
редколесьях Южной Америки, но встречается в большем количестве совместно с приречным
ложным миндалём (Neoterminalia riparia) и пальмой
дендем (Elaeis dendem) в галерейных лесах Южной Америки южнее области сельвы.
Лиловое дудчатое дерево – листопадное дерево, сбрасывающее листву в сухой сезон
и вырастающее до 20 м в высоту. Листья пальчатосложные, из шести листочков,
каждый из которых длиной 15 см, зелёные с серебристыми чешуйками сверху и снизу.
Крона дерева раскидистая, ветви отрастают от ствола беспорядочно, на разной
высоте; толстых ветвей очень немного. Кора тёмно-серая, может покрываться вертикальными
трещинами.
Цветение происходит в засушливые периоды, когда дерево совсем без листьев, или
их очень мало. Цветки ярко-фиолетовые, диаметром до 5 см, собраны по нескольку
штук в рыхлую метелку. Опылением занимаются птицы и насекомые. Длинные и тонкие
плоды-стручки могут вырастать до 30 см в длину. После высыхания из них выпадают
анемохорные крылатые семена. Они могут распространяться ветром на большие расстояния.
Этот вид растений открыл Жуан Витор Кутиньо, Бразилия.
Австралийская
кустарниковая маслина (Austrolea edibilis)
Порядок: Губоцветные (Lamiales)
Семейство: Маслинные (Oleaceae)
Место обитания: Южная Меганезия, зона кустарниковых зарослей (джарра), кустарниковая
саванна.
В эпоху человека в Австралии, как и во многих других местах на Земле, произрастало
множество трав и деревьев, которые были инвазивными «сорными» видами. В южной
части Австралии распространённым сорным видом деревьев была европейская маслина
(Olea europaea) – причиной этого было её повсеместное выращивание в этой местности
для получения масла благодаря благоприятному климату. В неоценовой Меганезии
она оставила нескольких потомков, хотя ни один из них не растёт в виде дерева,
каким иногда был его предок.
Это растение представляет собой кустарник среднего размера, вырастающий до 1,7
метров в высоту, хотя ширина его может достигать 3 метров в диаметре. Чаще всего
у него один ствол, который многократно ветвится, образуя густо растущие тонкие
ветви; форма этого кустарника обычно шарообразная или приземистая, но объедание
его травоядными животными может привести к тому, что растение будет выглядеть
иначе. Открытость его местообитания означает, что это растение способно выдерживать
лишь слабое затенение. Ветви опускаются до самой земли, со стороны его ствола
не видно, и мелкие животные часто прячутся и устраивают укрытия внутри этого
кустарника или под ним. Листья тонкие и узкие, похожи на листья его предка,
но несколько мельче и плотнее. Это вечнозелёное дерево, но она склонна к возгоранию
во время пожаров из-за масла, содержащегося в листьях и плодах. Кора очень тёмная,
серовато-бурая, а поверхность основного ствола имеет шероховатую текстуру. Корневая
система очень раскидистая; это дерево очень устойчиво к засухе.
Цветки мелкие, белые, собраны в кисти, образуют большое количество очень лёгкой
пыльцы; дерево ветроопыляемое. Цветение начинается в середине сезона дождей.
Плоды созревают в начале сухого сезона; плод представляет собой небольшую костянку,
которая значительно мельче, чем у её домашнего предка. Маленькое семя длиной
5 миллиметров заключено в оболочку из маслянистой мякоти, которая на вкус одновременно
и горькая, и сладкая. Весь плод округлый, до 1,7 см в диаметре, при созревании
тёмно-фиолетовый. Многие птицы и млекопитающие поедают плоды этого дерева и
распространяют его семена со своим помётом. Семена обладают толстой оболочкой
и способны выдерживать пожар, если он проходит достаточно быстро; таким образом,
опавшие семена могут выжить, даже если многие взрослые деревья погибнут из-за
пожаров. Молодые растения начинают давать семена уже в возрасте 5 лет. Отдельные
кусты могут увеличиваться в диаметре, раскидывая свою крону, на протяжении всей
своей жизни и доживать до ста лет.
Этот вид растений открыл Тимоти Моррис, Аделаида, Австралия.
Корнецветник
чуждый (Rhizoflosculus xenos)
Порядок: Норичниковые (Scrophulariales)
Семейство: Норичниковые (Scrophulariaceae)
Место обитания: леса Юго-Восточной Европы (южное и западное побережье Четвероморья)
В голоцене среди представителей семества норичниковых встречалось большое количество
видов, которые вели полупаразитический образ жизни – они прикреплялись слаборазвитыми
корнями к корневой системе других растений, и получали от них минеральные вещества
из почвы. Но они сохраняли зелёные листья, и выглядели внешне как самые обычные
травянистые растения. Также в семействе существовало небольшое количество настоящих
паразитических видов.
В неоцене тенденция перехода к паразитическому образу жизни усилилась. Этому
благоприятствовали изменения климата, которые привели к формированию новых продуктивных
экосистем. Влажные субтропические леса по берегам Четвероморья стали местом
обитания одного из новых видов паразитических растений.
На корнях высоких деревьев субтропических лесов Европы можно увидеть цветки
этого растения, за что оно получило название корнецветник. Это паразитическое
растение не имеет собственных зелёных листьев, и живёт практически полностью
за счёт дерева-хозяина. Его отношения с деревом-хозяином складываются своеобразно:
это паразитизм на грани симбиоза. Корнецветник большую часть года существует
в виде тканевых тяжей под корой и в толще древесины дерева-хозяина. Его присутствие
вызывает причудливые узловатые наплывы на корнях и нижней части ствола (не выше
метра над землёй). Но интересен тот факт, что у корнецветника очень хорошо развита
собственная корневая система. Эволюция сделала своеобразный шаг назад: вначале
растение «приросло» к дереву-хозяину и стало зависимым от него, а затем отношения
паразита и хозяина стали развиваться совсем в другую сторону. Под землёй корнецветник
развивает густую сеть корней, связанных с грибницей различных видов местных
грибов. Это приносит огромную пользу как паразиту, так и его хозяину: дерево-хозяин
получает от цветокоренника минеральные вещества из почвы, но взамен растение-паразит
забирает часть синтезированной хозяином органики. Сосудистая система растений
– хозяина и паразита – сплетается внутри узловидных наплывов на стволе, которые
со временем сильно разрастаются. Корнецветник может паразитировать на большом
количестве видов лиственных деревьев европейских лесов. Он избегает лишь растений,
на которых растут лианы семейства аралиевых – потомки плюща. Очевидно, вещества,
выделяемые этими растениями, угнетают проростки корнецветника на ранних стадиях
развития.
Летом на образованных тканями паразита наплывах, а также на корнях дерева-хозяина
образуются цветки. Два прочных хрящеватых прицветника пробивают кору растения-хозяина,
и раскрываются, выпуская наружу одиночный цветок. Цветки этого паразитического
растения крупные – диаметр цветка достигает 4 – 5 см. они многочисленные – на
одном квадратном метре раскрывается до нескольких десятков цветков. Цветок корнецветника
двугубый, с вытянутым в трубку околоцветником, ярко-жёлтый с сиреневым «глазком»
в центре. Верхний лепесток покрыт серебристыми волосками, отражающими ультрафиолетовый
свет, которые складываются в продольные полосы, сходящиеся в зеве цветка. Это
своеобразный «указатель» для насекомых-опылителей. Их также привлекает сильный
аромат с лёгким гнилостным оттенком. Цветки этого растения опыляются преимущественно
одиночными перепончатокрылыми, хотя изредка их посещают бабочки. Лепестки корнецветника
плотно смыкаются, и раздвинуть их может только сильное насекомое, поэтому мелкие
случайные опылители вроде мух не могут добраться до нектара. Бабочки с длинным
хоботком, крупные виды дневных бражников, могут просто просовывать тонкий хоботок
между плотно сомкнутыми лепестками, и пить нектар, не производя опыления.
Плод корнецветника очень своеобразен – в нём независимо развилось и повторилось
«конструктивное решение» растения из семейства тыквенных, известного как «бешеный
огурец» (Ecballium elaterium). Зрелый плод корнецветника – округлая коробочка
длиной около 3 см, заполненная жидкостью, в которой плавают зрелые семена. Жидкость
находится под высоким давлением. Плод вскрывается с помощью небольшой крышечки
на конце. Когда крупное животное случайно задевает плод, он «взрывается» и выстреливает
в животное клейкой жидкостью с семенами. Семена корнецветника мелкие, они легко
прилипают к шерсти и переносятся животным в другие места. Когда животное трётся
об деревья, семена остаются на коре и прорастают. Проросток корнецветника очень
слабый и чувствительный: его легко заглушают фитонциды, выделяемые другими растениями.
Поэтому корнецветник встречается в лесах Европы относительно редко. Если вид
дерева-хозяина подходит корнецветнику, и ничто не препятствует развитию паразита,
проросток внедряется под кору дерева и разрастается. Первые годы жизни он развивает
в почве тонкие корни, разыскивая грибницу и образуя микоризу. В это же самое
время на дереве-хозяине начинают формироваться характерные наросты. Достигнув
примерно десятилетнего возраста, корнецветник зацветает.
Виев цвет (Sublathraea wiijii)
Порядок: Норичниковые (Scrophulariales)
Семейство: Норичниковые (Scrophulariaceae)
Место обитания: леса Юго-Восточной Европы (южное и западное побережье Четвероморья)
В голоцене на территории Австралии люди обнаружили две разновидности орхидей,
отличающихся своеобразным способом цветения – криптантемис (Cryptanthemys
slateri) и ризантелла (Rhizanthella gardneri). Оба этих растения лишены хлорофилла.
Они росли и цвели под землёй. Даже на момент открытия оба этих растения были
крайне редкими. Орхидея криптантемис исчезла ещё в историческую эпоху, а
ризантелла
исчезла вслед за ним в эпоху антропогенного прессинга, несмотря на усилия
по охране.
В эпоху неоцена на Земле появилось ещё одно растение, ведущее подземную жизнь,
и даже цветущее под землёй. Это потомок петрова креста (Lathraea squamaria),
паразитического растения семейства норичниковых. Ещё в эпоху голоцена это
растение утеряло хлорофилл, а в неоцене специализация его потомков достигла
вершины.
Потомок петрова креста – полностью подземное цветковое растение, виев цвет.
Вий – подземное чудовище у славян, которое появлялось на земле лишь раз в
четыре года, 29 февраля, и было способно убивать взглядом. Всё остальное
время Вий,
по поверьям, проводил под землёй. Неоценовое растение, которое встречается
в лесах на побережье Четвероморья, ведёт столь же скрытный подземный образ
жизни.
Виев цвет – это паразитическое растение, которое большую часть года растёт
под землёй. Этот вид паразитирует на кустарниковых растениях семейства берёзовых
(Betulaceae), обитающих в подлеске. Виев цвет предпочитает рыхлую мягкую
почву, и встречается близ берегов рек или в долинах, где почва рыхлая и достаточно
влажная. Очевидно, это связано с оптимальными условиями роста его хозяев.
У виева цвета нет хлорофилла, и его листья превратились в толстые чешуи.
Тело растения – ползучее корневище длиной около метра, располагающееся на
глубине
20 – 30 см. Корневая система растения редуцирована до нескольких корней-присосок,
с помощью которых виев цвет прикрепляется к корням растений-хозяев. Это паразитическое
растение полностью зависит от своего хозяина.
Летом, когда почва тёплая и влажная, а растение-хозяин усиленно вегетирует,
виев цвет зацветает. Это событие нельзя увидеть, не прилагая особых усилий,
поскольку виев цвет даже цветёт под землёй. Но соцветия этого растения можно
обнаружить по характерному запаху, привлекающему животных-опылителей.
Виев цвет образует соцветия в пазухах чешуевидных листьев, плотно охватывающих
корневище. Соцветие развивается в обёртке из нескольких крепких сочных листьев,
расположенных попарно. Они защищают цветки от повреждений. Обёртка «пробуривает»
толщу земли и слегка раскрывается на конце, достигнув уровня почвы. При созревании
цветков обёртка соцветия разрастается в ширину, образуя камеру, в которую
попадают опылители растения.
Цветки виева цвета не отличаются изысканной формой – они привлекают опылителей
не внешним видом, а запахом. У них редуцированные лепестки и бледно-серая
окраска с розоватым оттенком. Каждый цветок окружён двумя большими прицветниками.
Цветки
плотно сидят продольными рядами на толстой оси соцветия. Аромат цветков этого
вида сильный, сладкий с гнилостным оттенком. Виев цвет опыляется наземными
жуками, которых очень привлекает такой запах. Часто на соцветиях виева цвета
можно найти мелких наземных улиток и слизней, которых также привлекает запах
цветущего растения. Они не играют существенной роли в опылении, но могут
поедать значительную часть завязей. От повреждений растение спасают его основные
опылители
– жуки: многие жуки, кормящиеся на соцветиях виева цвета, плотоядны и охотятся
на моллюсков.
После опыления, незадолго до созревания семян, виев цвет показывается на
поверхности земли. Ось соцветия сильно вытягивается и поднимает созревающие
плоды на высоту
около метра над землёй. Плоды виева цвета – сухие коробочки, вскрывающиеся
мелкими крышечками на концах. Когда ветер раскачивает растение, через отверстия
высыпаются мелкие семена, которые разносятся ветром. В одной коробочке может
находиться несколько тысяч мелких семян.
Семя виева цвета прорастает сразу, попав на землю. Проросток ищет химические
вещества, выделения корней растения-хозяина, и растёт в направлении этих
корней. Обычно значительная часть проростков гибнет, не найдя подходящего
места для
жизни. Виев цвет развивается медленно: от прорастания семени до первого цветения
проходит свыше 10 лет.
Снежнолист узамбарский (Niveophyllum usambaricus)
Порядок: Норичниковые (Scrophulariales)
Семейство: Геснериевые (Gesneriaceae)
Место обитания: горы Узамбара (Земля Зиндж).
Геснериевые – одно из характерных семейств растений тропической зоны. В эпоху
голоцена оно было представлено исключительно травянистыми растениями разного
облика – от клубневых многолетних растений и вечнозелёных эпифитов до однолетников
и монокарпических трав с единственным листом. В ледниковую эпоху видовое многообразие
этого семейства значительно пострадало из-за сокращения площади пригодных для
обитания мест тропического пояса. Но в неоцене, когда влажные вечнозелёные
леса широко распространились по Земле, эти растения получили новую возможность
для эволюции, и использовали её максимально полно.
Неоценовые геснериевые захватили даже такие места обитания, которые ранее были
недоступны для них. Один из таких нетипичных представителей семейства обитает
в горах Земли Зиндж. Когда Африканский континент разломился по линии рифта,
и его восточная часть откололась, самые высокие горы Африки перестали принадлежать
ей. Живые существа, оставшиеся на Земле Зиндж, стали эволюционировать независимо
от континентальных видов.
Это невысокое растение – до метра в высоту. Его жизненная форма нехарактерна
для семейства: у него толстый одревесневающий ствол. На вершине ствола раскинулась
розетка из широких листьев, покрытых белым опушением. Молодые листья опушены
особенно густо: кажется, что в середине растения лежит снег. За эту особенность
растение получило название снежнолист. Однако этот вид не произрастает там,
где выпадает настоящий снег: снежнолист сохранил относительную теплолюбивость,
характерную для семейства. Предком этого растения был вид, широко известный
в эпоху человека как сенполия, или «узамбарская фиалка» (Saintpaulia ionantha).
Облик этого растения несколько напоминает некоторые гигантские виды лобелий
(Lobelia), обитавших в горах Африки в человеческую эпоху. Ствол снежнолиста
покрыт листьями только в верхней части, где они образуют широкую розетку. Основание
ствола защищено от внешних воздействий рыхлой губчатой корой, которая смягчает
перепад между дневными и ночными температурами: ночью в местах обитания снежнолиста
бывает довольно прохладно, а изредка даже случаются заморозки. Поэтому такая
защита растению как нельзя кстати. Если заморозки слишком затягиваются, старые
листья отмирают, но не отваливаются, а остаются на стволе как дополнительная
теплоизоляция. Но несколько молодых листьев, наиболее густо покрытых серебристыми
волосками, всегда выживает, и растение быстро восстанавливается после холодов.
Если же заморозки оказались слишком сильными, растение всё равно не погибает:
у основания ствола снежнолиста из спящих почек развиваются молодые розетки
листьев. Со временем они разрастаются, образуя дополнительные стволы, и уже
сами образуют новые розетки. Так за несколько десятков лет одиночное растение
может превратиться в заросль с несколькими десятками стволов.
Почти круглый год снежнолист цветёт, делая перерыв только в самое холодное
время. Цветки снежнолиста собраны в большую кисть, насчитывающую до 20 цветков.
Цветок длинный, трубчатый, белый с сильным запахом. В зеве цветка есть рисунок
из мелких красных пятен, сливающихся в глубине цветка в одно пятно. Цветки
снежнолиста раскрыты постоянно, но их специфический пряный аромат днём почти
не ощущается, зато усиливается к ночи. В это время на кормёжку вылетают мелкие
летучие мыши, которые питаются исключительно нектаром растений. Им не приходится
далеко летать за кормом: они живут под листьями снежнолиста, где находят удобное
тёплое жилище, защищённое от ветра. Эти же животные опыляют растение. Цветонос
очень прочный, и летучие мыши, вылизывая нектар, могут просто перепрыгивать
с растения на растение.
Плод этого растения – небольшая коробочка с очень мелкими семенами, которые
распространяются ветром. Проростки снежнолиста очень слабые и нежные, и большая
часть их просто гибнет среди травы. Но некоторым проросткам удаётся выжить
и вырасти. Ещё один способ размножения снежнолиста – вегетативно, с помощью
листьев. Отломленный ветром или животными лист растения может легко укорениться
в почве и дать несколько молодых растений.
Асимметрокаликс правосторонний (Asymmetrocalyx dexter)
Порядок: Норичниковые (Scrophulariales)
Семейство: Геснериевые (Gesneriaceae)
Место обитания: влажные тропические леса Амазонии, полог леса.
Тропические леса Амазонии в эпоху оледенения пережили крайнюю степень упадка
– их область распространения на некоторых стадиях оледенения распадалась на
ряд изолированных островков, где сохранялась лишь небольшая доля былого разнообразия
их обитателей. После отступления ледников, когда климат вновь стал теплее,
дождевые леса образовали сплошной покров в северной половине Южной Америки.
Разнообразие обитателей дождевых лесов неоцена породило конкуренцию, и как
её следствие – узкую специализацию. Цветковые растения стали вырабатывать различные
способы привлечения опылителей, доводя их до разной степени изощрённости. Одни
цветы гостеприимно открыты всем опылителям, а другие делятся нектаром только
с теми, кто приспособился его добывать. К растениям, опыляемым птицами и насекомыми,
относятся представители семейства геснериевых. Представители одного из родов
этих растений длительное время эволюционировали совместно с особым родом колибри,
и итогом этой эволюции стала нерушимая связь между птицей и растением.
Цветки этого вида геснериевых имеют длинный трубчатый венчик с ярким отгибом,
который скрывает нектар от посторонних простым способом: он изогнут вбок. За
это растение получило название Asymmertocalyx («несимметричная чашечка цветка»).
Изгиб может быть как вправо, так и влево, и два вида этого рода растений успешно
реализовали оба варианта строения. Эти виды отличаются друг от друга местами
обитания, и их опылители также не встречаются друг с другом.
Оба вида этих растений – эпифиты, обитающие в пологе леса. В тропических лесах
Амазонии водится асимметрокаликс правосторонний, вид, у которого трубковидный
венчик изогнут вправо. Соцветие у этого растения – длинный колос на прочном
вертикальном цветоносе, причём ось соцветия винтообразно закручена. Цветки
сидят попарно на коротких ножках в пазухах широких округлых листочков. На одном
узле цветоноса растёт, таким образом, четыре цветка. Цветоносы этого вида покрыты
густыми белыми волосками – это «фамильная черта» геснериевых.
Длина трубки цветка – около 5 сантиметров, что примерно соответствует длине
клюва колибри, опыляющего этот вид. Для привлечения птицы-опылителя отгиб венчика
имеет ярко-красную расцветку с полосками серебристых волосков, тянущихся от
краёв в зев цветка. Диаметр цветка этого вида – около 2 сантиметров. Плод асимметрокаликса
– округлая сухая коробочка, растрескивающаяся случайным образом. Мельчайшие
семена снабжены хохолками-летучками.
Асимметрокаликсы – многолетники с прочными ползучими стеблями. Они прикрепляются
к коре дерева-опоры множеством тонких корней, а в воздух свисает борода белых
воздушных корней, собирающих росу и дождевую воду. Листья асимметрокаликса
правостороннего тёмно-зелёные, рельефные, с вдавленными жилками. Они не очень
крупные, кожистые, с небольшим количеством волосков на верхней стороне. Листья
собраны в плотные розетки, и во время дождя направляют дождевую воду к корням.
Это растение опыляется колибри бакороро, или левосторонним
кривоносым колибри (Asymmetrochilus bakororo), клюв которого приспособлен к добыче нектара из
таких цветков. Приспособленность асимметрокаликса к опылению колибри не абсолютна
– некоторые крупные бабочки также могут добывать нектар растения, повернувшись
боком. А пчёлы просто подгрызают трубку цветка, и добывают нектар, минуя изгиб
венчика цветка.
В прохладных туманных лесах на склонах гор обитает родственный вид – асимметрокаликс
левосторонний (Asymmetrocalyx sinister). Этот вид приспособился к жизни в условиях
высокой влажности и прохладного климата. Всё растение покрыто интенсивным белым
опушением, которое задерживает капли росы, защищая листья от загнивания. Многочисленные
короткие воздушные корни собирают влагу с поверхности растения. Они развиваются
даже на цветоносах, из одного узла с листьями. Цветки этого вида карминно-розовые,
более крупные, чем у вида из дождевого леса – диаметром до 5 см. Асимметрокаликс
левосторонний опыляется итубори,
правосторонним кривоносым колибри (Asymmetrochilus itubory). Но также это растение может эффективно расселяться с помощью вегетативного
размножения – на кончиках крупных листьев у него образуются выводковые почки
с укороченными листьями и волосками, видоизменёнными в крючки. Выводковая почка
цепляется за оперение птиц и легко отламывается от материнского растения, а
птицы разносят такие почки по лесу, оставляя в местах, которые могут быть пригодными
для роста.
Колериопсис
трубчатый (Kohleriopsis tubiflorus)
Порядок: Норичниковые (Scrophulariales)
Семейство: Геснериевые (Gesneriaceae)
Место обитания: Южная Америка, влажные тропические леса бассейна Амазонки и
Гипполиты.
Уничтожение тропических лесов в эпоху человека сопровождалось массовым вымиранием
видов животных и растений, для которых лес был средой обитания. Сильно пострадали
узкоареальные виды с невысокой численностью, сильно зависящие от локально сложившихся
условий среды обитания. После исчезновения человека и восстановления приемлемых
условий для жизни началась активная эволюция эпифитных растений, осваивающих
полог новых тропических лесов.
Колериопсис трубчатый является представителем эпифитных геснериевых. Благодаря
мелким семенам эти растения легко смогли освоить полог леса, где обитают различные
виды этого семейства. Колериопсис встречается в кронах деревьев на крупных горизонтальных
ветвях и стволах лиан. Это многолетнее травянистое растение; основным прикрепительным
органом является корневище, покрытое чешуевидными рудиментами листьев. Оно растёт
горизонтально, и врастает в трещины коры дерева, заполняя их по мере роста.
Изредка растение может «переползать» на ствол и разрастаться вверх и в стороны.
Растение прикрепляется к субстрату при помощи многочисленных мелких корней,
среди которых скапливается лесной мусор – из него растение извлекает минеральные
вещества. От корневища отрастают короткие вертикальные побеги, на которых образуются
цветки. Стебли и черешки листьев покрыты красновато-бурыми волосками, в узлах
развиваются воздушные корешки. Листья простые, супротивные, округлые, тёмно-зелёные,
с заострёнными кончиками и мелкозубчатым краем, длиной до 10 см. Нижняя сторона
листа серовато-белая. Поверхность листа покрыта множеством волосков, придающих
ему бархатистую фактуру. На побеге высотой до 15 см обычно развивается 4-6 пар
листьев; после этого рост побега замедляется, а на его верхушке начинают развиваться
цветки.
Цветоносы короткие, прочные, поднимаются над листьями на 5-8 см. Цветки колериопсиса
трубчатого отличаются очень характерным обликом – они трубчатые, длиной около
15 см при ширине трубки не более 8 мм. Из-за большой длины трубки кончик цветка
выступает за края листьев, и опылитель вынужден сосать нектар исключительно
в полёте. Диаметр пятилепесткового цветка не превышает 3 см. Трубка околоцветника
прямая, цветки птицеопыляемые, но могут эпизодически посещаться пчёлами крупного
размера. Трубка цветка окрашена в различные оттенки красного – от тёмно-розового
до вино-красного; одна из экологических рас этого вида обладает белыми цветками
и опыляется бабочками-бражниками. Лепестки пятнистые – по фоновому цвету разбросаны
тёмно-красные пятна (кроме белоцветковой расы), и на них имеются ряды волосков,
отражающих ультрафиолетовый свет. Волоски служат указателями нектара, и особенно
развиты у белоцветковой расы. На границах ареалов отдельных рас встречаются
формы с промежуточными характеристиками цветков.
Семена этого вида очень мелкие, переносятся ветром. От прорастания семени до
первого цветения молодого растения проходит около года.
Пузырчатка-химера
(Utricularia amphibiotica)
Порядок: Норичниковые (Scrophulariales)
Семейство: Пузырчатковые (Lentibulariaceae)
Место обитания: временные водоёмы в пампасах Южной Америки.
Многие группы растений сравнительно благополучно преодолели рубеж голоцена и
неоцена, и эволюционировали в новые виды в восстанавливающихся природных сообществах.
Хищные растения пузырчатки (Utricularia) были одними из широко распространённых
насекомоядных растений. В эпоху человека они обитали на всех материках, кроме
Антарктиды, и это положение дел не изменилось в неоцене.
Большинство из новых видов пузырчаток мало чем отличается от своих предков,
но есть и исключения; например, в болотах Сибири и Китая растёт исполинская
пузырчатка, настоящие великан среди своих сородичей, а на другом конце света,
в прериях Южной Америки, растёт её дальняя родственница, названная в честь античного
чудовища.
В древних мифах химера была трёхголовым чудовищем, символически объединявшим
три стихии – огонь, воздух и землю. Её южноамериканская тёзка способна обитать
сразу в трёх стихиях: корнями в земле, листьями и стеблями в воздухе, и, если
представится такая возможность, в воде. Это растение обитает на берегах неглубоких
рек и озёр в пампасной зоне Южной Америки, которые имеют тенденцию пересыхать
в сухой сезон. Большинство из пузырчаток, конечно, скорее сухопутные чем подводные
растения, и предпочитают влажные местообитания тропических лесов. Другие пузырчатки,
напротив, ведут строго водный образ жизни и не могут выживать на суше. Пузырчатка-химера
умудряется в какой-то мере использовать оба мира. Она произрастает на берегах
водоёмов и выпускает стебли двух видов. Само растение обитает на влажной почве
и образует прямостоячий стебель со сближенными междоузлиями и округлыми листьями
со слабонадрезанным краем. На вершине стебля во время влажного сезона образуется
высокое (до 50 см) неветвистое соцветие-кисть с очередными зигоморфными двугубыми
цветками. Каждый цветок диаметром до 2 см, белого цвета с крупным красно-коричневым
пятном на нижней губе. Цветок плотно закрыт, и пролезть в него может только
тяжёлое насекомое вроде жука, мухи или пчелы. Открыв зев цветка, оно высыпает
себе на спину пыльцу, которую насекомое перенесёт на следующий цветок. Также
на цветках кормятся комары и бабочки, но они обычно просто просовывают хоботок
между губами цветка и не производят опыления. Семенное размножение этого вида
происходит успешно – около четверти цветков завязывают семена.
В то время как основное растение живёт на суше, оно выпускает из пазух нижних
листьев длинные лианоподобные побеги. Если они растут в сторону воды, то начинают
изменяться, реагируя на влажность воздуха. Листья на них по-прежнему имеют округлые
очертания, но сильнее рассечённые. Под собственной тяжестью побег скоро опускается
в воду и начинает образовывать тонкий стебель с пушистыми листьями, рассечёнными
на нитевидные доли, на которых вырастают пузырьки-ловушки. С помощью этих пузырьков
растение ловит мелких водяных беспозвоночных, в то время как побеги наземного
типа образуют новые растения на суше вегетативным способом: конец длинного побега
укореняется и даёт начало новому растению. Если побег ложится на землю и укореняется
в узлах, из этих узлов отрастают новые надземные побеги.
Подводные побеги могут переноситься в другие водоёмы на ногах птиц. В сухой
сезон они гибнут, но образуют в пазухах листьев шишковидные почки в твёрдой
оболочке, которые сохраняются в мокрой грязи. В начале сезона дождей из этих
побегов могут вырасти как наземные, так и водные побеги, в зависимости от условий,
в которых прорастает почка. Семена легко переносят пересыхание и прорастают
вскоре после увлажнения почвы. В благоприятных условиях каждый наземный побег
растения живёт до трёх лет. Растение активно размножается вегетативно и образует
плотные куртины на берегах и мелководьях.
Этот вид растений открыл Bhut, участник форума.
Томат-мухоловка (Neolycopersicon muscicarpus)
Порядок: Паслёноцветные (Solanales)
Семейство: Паслёновые (Solanaceae)
Место обитания: Южная Америка, редколесья в тропической и субтропической областях.
В эпоху человека Южная Америка была признана центром многообразия томатов (род
Lycopersicon). Среди этих растений встречались виды, приспособленные к различным
местам обитания: например, на Галапагосских островах обитал один солеустойчивый
вид. Виды, не введённые человеком в культуру, оказались достаточно выносливыми,
чтобы пережить эпоху антропогенного прессинга и глобальный экологический кризис
на рубеже голоцена и неоцена. В неоцене некоторые потомки томатов приобрели
удивительные приспособления для выживания. Человеку удалось вывести сорта томата,
которые самостоятельно защищались от мелких насекомых, удерживая их с помощью
клейких волосков. Потомок одного из южноамериканских видов томатов независимо
от культурных сортов томата развил такую же способность, но продвинулся ещё
дальше. Клейкие волоски из простого защитного приспособления превратились у
этого растения в ловчее орудие – растение специально привлекает и ловит мелких
насекомых и клещей. Этот вид растений – томат-мухоловка.
Растение ловит насекомых не только для того, чтобы защитить себя от повреждений.
Этот вид приспособился к жизни на бедной почве, и от присутствия насекомых
зависит его благополучие. Томат-мухоловка высасывает пойманных животных, получая
таким способом необходимое количество минеральных и органических веществ. Волоски
на его листьях имеют особое строение. Каждый волосок представляет собой гигантскую
клетку с прочной оболочкой. На кончике волоска выделяется капля клейкого вещества.
Когда насекомое садится на лист и прилипает к волоскам, вещества, выделяемые
его телом, стимулируют прорастание верхушки волоска в ткани насекомого. Волосок
прорастает в тело жертвы в виде цепочки тонкостенных клеток, которые впитывают
его жидкое содержимое. Особенно успешно растение потребляет мелких мягкотелых
насекомых – тлей, трипсов, мелких гусениц и личинок двукрылых. После того,
как питательные вещества будут поглощены, волосок отмирает и отсыхает, а ветер
уносит прочь опустошённую оболочку насекомого. Наиболее успешно ловят насекомых
молодые листья, густо покрытые волосками. Они красноватые из-за пигмента антоциана,
защищающего их от солнечного ожога. Для привлечения насекомых на некоторых
волосках выделяются капли сладкой жидкости. Это растение находится на раннем
этапе специализации к питанию насекомыми и ещё не имеет приспособлений, позволяющих
активно удерживать добычу.
Первые листья молодого растения округлые, но на последующих листьях появляются
зубчики на краях, переходящие в глубокие вырезы. Полностью сформированные листья
перисторассечённые, тёмно-зелёные, слегка липкие и бархатистые на ощупь. На
взрослых листьях ловчие волоски остаются только на боковых лопастях и по краю
листа. Всё растение защищено от травоядных животных алкалоидами и имеет специфический
запах.
Томат-мухоловка – многолетнее растение. На первом году жизни его стебель вертикальный,
но с возрастом он полегает и укореняется в узлах. Боковые побеги и верхушка
приподнимаются вертикально, и растение образует целую заросль. Это растение
опыляется насекомыми, и, чтобы случайно не поймать собственных опылителей,
выносит цветки высоко вверх на длинном цветоносе. Цветки томата-мухоловки небольшие,
пятилепестковые, похожие на цветки большинства паслёновых. Венчик срастается
в короткую трубку, а белые остроконечные лепестки отогнуты назад. Плод этого
растения – маленькая красная ягода. В зрелом виде плод растения съедобен, и
его охотно поедают крупные наземные грызуны.
Некоторые виды клопов и жуков приспособились «обворовывать» это растение, схватывая
с его листьев прилипших насекомых.
Табак
длиннейшецветковый (Nicotiana longissimoflora)
Порядок: Паслёноцветные (Solanales)
Семейство: Паслёновые (Solanaceae)
Место обитания: Большая Антигуа, участки тропического леса.
Среда обитания любого вида живых организмов включает не только факторы неживой
природы (климат, количество осадков, особенности рельефа и почвы), но и окружающие
виды. Любой вид неизбежно взаимодействует с другими видами, которые выступают
в роли хищников, добычи или паразитов, опылителей или распространителей семян
и спор, комменсалов или симбионтов. И характер взаимодействия с этими видами
оказывает огромное влияние на эволюцию любого отдельно взятого вида.
У цветковых растений взаимодействие с животными в буквальном смысле наиболее
ярко проявляется на примере эволюции приспособлений к опылению. Венчики цветов
имеют различную форму и строение, а цветки издают определённый запах, чтобы
максимально соответствовать особенностям строения или поведения того или иного
опылителя и исключить появление каких-то случайных опылителей.
Один из примеров узкой специализации к опылителю демонстрирует растение, обитающее
в тропических лесах острова Большая Антигуа, в тропической Америке. Этот вид,
табак длиннейшецветковый, произошёл от одичавшего культурного табака (Nicotiana
tabacum), который широко культивировался в Новом Свете человеком. Его потомок
представляет собой крупное многолетнее растение с древовидным стеблем высотой
до 6 м. Табак длиннейшецветковый относится к светолюбивым и быстрорастущим видам.
В подлеске тропического леса растения этого вида сильно вытягиваются и быстро
гибнут из-за недостатка солнечного света. Этот вид растёт как пионерное растение
на участках леса, где древостой повреждён. Обычно заросли этого вида встречаются
там, где ураганы повалили старое дерево, и оно, падая, подмяло под себя ещё
несколько. Большие заросли этого вида встречаются также по берегам рек.
В строении табака длиннейшецветкового всё указывает на то, что растение в полной
мере пользуется кратковременными преимуществами роста на хорошо освещённых участках
повреждённого леса. Это очень быстрорастущее растение, которое способно за один
год дать прирост около трёх метров. Стебель изначально травянистый, с пучками
жёстких продольных волокон. Но по мере роста он одревесневает в нижней части,
быстро утолщается и становится значительно прочнее. Поверхность стебля покрывается
серой корой, которая постепенно отслаивается узкими лентами. Листья этого вида
очень крупные, в нижней части стебля их длина приближается к 1 метру. Листья
молодого растения цельные, овальные, с перисто-сетчатым жилкованием. Поверхность
листа между жилками слегка вздутая и складчатая. Края и верхняя сторона листа
опушены серебристыми волосками. Молодые листья имеют более густое опушение,
и верхушки побегов из-за этого кажутся серебристыми. Достигнув высоты около
пяти метров, растение теряет листья на нижней части стебля и начинает ветвиться.
Листья на ветвях мельче первичных – их длина не более 70 см. Но их форма отличается
от тех, что развивались в первый год жизни растения: в основании листа появляются
две заметных лопасти, которые постепенно увеличиваются к кончикам побегов. На
концах ветвей листья трёхлопастные, боковые лопасти составляют в длину примерно
половину от длины центральной лопасти листа. На кончиках листьев развивается
длинная узкая «капельница», помогающая избавляться от излишков воды.
Цветки этого растения начинают развиваться уже на втором году жизни. Они появляются
в виде кистей на концах веток. Цветение этого вида представляет собой великолепное
зрелище. Цветки табака длиннейшецветкового трубчатые, повислые, с пятью округлыми
лепестками. Диаметр венчика составляет около 10 см, но его длина уникальна для
растительного мира: около 50 см. Цветки ярко-красные, покрыты волосками, отражающими
ультрафиолетовое излучение. Они опыляются только одним видом бабочек – антигуанским
гигантским бражником. Хотя насекомые не различают красный цвет, волоски
на лепестках делают цветок хорошо заметным в ультрафиолетовом свете, и помогают
опылителю отыскать его. Кроме того, цветки этого растения очень душистые, и
их запах насекомое-опылитель может почувствовать на расстоянии около километра.
Самые большие соцветия развиваются на верхушке растения – бабочка-опылитель
не любит спускаться близко к земле. Изредка насекомые других видов могут добывать
нектар этого растения. Одиночные осы и пчёлы просто прогрызают трубку цветка
у основания и вылизывают нектар, не производя опыления.
Плод этого растения – коробочка. После цветения рост ветки в длину прекращается
и она начинает ветвиться. По количеству ветвлений можно приблизительно определить
возраст этого растения. Но табак длиннейшецветковый не живёт долго – максимальный
возраст этого растения составляет не более десяти лет. По мере роста этот вид
вытесняется порослью деревьев, составляющих тропический лес, и постепенно хиреет.
Для защиты от травоядных животных растение продуцирует смесь ядовитых алкалоидов
близкой химической структуры – никотин, анабазин, кониин, гигрин и т. д. Мало
кто из травоядных может съесть
его. Однако животные часто поедают старые и опавшие листья, содержащие остаточное
количество алкалоидов, чтобы избавиться от внутренних паразитов, или валяются
в листве, чтобы изгнать паразитов из шерсти или перьев. Лишь гусеницы бабочки-опылителя
могут безнаказанно поедать листья этого растения.
Идею о существовании этого вида растений высказал Антон, участник форума.
Бромелиевая
повилика (Baccuscuta bromeliophila)
Порядок: Паслёноцветные (Solanales)
Семейство: Вьюнковые (Convolvulaceae)
Место обитания: Южная Америка, полог влажного тропического леса.
Рисунок Биолога
Одной из выгодных стратегий выживания растений во влажном
тропическом лесу является паразитизм. Растения-паразиты способны освоить жизнь
в любом ярусе леса, а высокая степень приспособленности к паразитизму позволяет
им расти на любом ярусе леса, даже в самом тенистом подлеске, за счёт более
успешных растений-хозяев. Во влажных тропических лесах Южной Америки эпохи неоцена
обитает один из видов таких растений – крупная повилика, паразитирующая на деревьях
в пологе леса. Подобно всем своим родственникам, это бесхлорофилльное растение,
лишённое листьев, с длинными травянистыми нитевидными стеблями. Многолетний
стебель достигает 3-4 метров в длину, ветвится и обильно образует гаустории
при соприкосновении с растением-хозяином подходящего вида. Окраска побегов светло-оранжевая,
на кончиках побегов бледно-розовая.
Соцветия представляют собой клубочки из многочисленных мелких цветов на тонкой
длинной ножке. Они поднимаются из массы побегов вертикально и покачиваются на
слабом ветру, привлекая насекомых-опылителей. «Цветонос» фактически представляет
собой укороченный боковой побег – его кончик выступает из верхушки соцветия,
останавливая рост на этом этапе. Мелкие цветки пятилепестные, актиноморфные.
Растение не специализировано к определённым опылителям, и его пыльцу могут переносить
мелкие мухи, жуки, осы и ночные бабочки. В отличие от родственных видов эпохи
человека, плоды этого вида представляют собой невскрывающиеся коробочки. Их
стенки сочные, мясистые, по мере созревания срастаются вместе. Они приобретают
ярко-красный цвет и сладковатый вкус, имитируя ягоды – отсюда название рода,
означающее «ягодная повилика». Семена погружены в мякоть соплодия. Это приспособление
для жизни в условиях высокой конкуренции в тропическом лесу. Семена растения
разносятся птицами и млекопитающими, также поедаются мелкими игуанами. Благодаря
зоохории повышается вероятность доставки семян к потенциальному растению-хозяину.
Они очень долго сохраняют всхожесть, и прорастают в разное время.
Чаще всего это растение прорастает в эпифитных «корзинках», и первое время развивается
на самом эпифите – обычно это растение семейства бромелиевых, либо орхидея.
Побеги вылезают за пределы зарослей эпифитов, дотягиваются до коры дерева и
внедряются в неё гаусториями. Бромелиевая повилика обычно паразитирует на деревьях,
используя эпифитные растения в качестве промежуточного хозяина, но при росте
на неподходящем виде деревьев может проходить весь жизненный цикл на эпифите.
Дендровольвулюс
древодушитель (Dendrovolvulus arbrosphinga)
Порядок: Паслёноцветные (Solanales)
Семейство: Вьюнковые (Convolvulaceae)
Место обитания: острова Карибского моря, тропики Центральной и Северной Америки,
леса.
Восстановление биологического многообразия тропических лесов в раннем неоцене
сопровождалось многочисленными попытками растений из различных семейств занять
новые экологические ниши и образовать новые жизненные формы. В Новом Свете присутствие
колибри способствовало активной эволюции орнитофильных растений, получивших
преимущество за счёт использования надёжных опылителей из числа этих птиц. Один
из примеров успешной эволюции орнитофильных растений в тропиках Нового Света
– крупная лиана дендровольвулюс древодушитель.
Это растение – крупная многолетняя лиана с сильно разветвлённым одревесневающим
стеблем. Длина стебля взрослого растения может достигать 20 метров при толщине
до 5 см в основании. Вид является потомком травянистого растения калистегии
(Calystegia); название происходит от старого названия предкового рода – Volvulus.
Стебель вьющийся, на ранних этапах роста ползёт по лесной подстилке и не одревесневает.
Найдя опору, растение быстро обвивает её и начинает интенсивный рост в высоту.
Нижняя часть стебля с возрастом одревесневает; древесина упругая и прочная,
серая кора отшелушивается небольшими чешуйками. Одревесневающий стебель мешает
утолщению ствола растения-опоры и может буквально душить обвитые стволы деревьев
у основания, тем самым позволяя растению конкурировать с ними за свет. Особенно
часто дендровольвулюс удушает молодые активно растущие деревья. От предка это
растение унаследовало ползучее подземное корневище, которое тянется к соседним
деревьям и выпускает из земли новые побеги. На корневище развиваются многочисленные
спящие почки, прорастающие при его повреждении.
Листья этой лианы черешковые, стреловидных очертаний. Боковые лопасти в основании
листа значительно меньше средней и сильно отогнуты в стороны. У зрелого растения
на крупных листьях они отделяются от основной доли листа сужениями и могут быть
сильно выгнуты в стороны, отчего лист выглядит трёхлопастным.
Цветки этого вида характерны для семейства вьюнковых: они обладают узкой трубкой
и широким отгибом из пяти сросшихся лепестков, почти без запаха. Околоцветник
красного цвета, изредка с белыми пятнами в зеве цветка. Цветки развиваются на
молодом приросте по одному в пазухах листьев, держатся на цветоносах горизонтально
или слегка повислые. Цветение каждого отдельного цветка продолжается 3-4 дня,
на ночь и перед дождём они закрываются. Днём цветки широко раскрыты и обильно
выделяют жидкий нектар; они опыляются колибри, изредка крупными бражниками.
Некоторые осы прогрызают трубку цветка сбоку и вылизывают нектар через отверстие,
не производя опыления.
Взрослое растение этого вида очень светолюбиво, при затенении цветёт плохо или
вовсе не зацветает.
Плоды дендровольвулюса – деревянистые коробочки, вскрывающиеся продольным швом
и с силой разбрасывающие семена. Благодаря прочной оболочке семена легко выдерживают
пребывание в морской воде в течение нескольких дней. Эта особенность позволила
данному виду широко распространиться по островам Карибского моря и в тропиках
Центральной и Северной Америки. Проростки этого вида встречаются на берегах
Северной Америки в зоне умеренного климата, но молодые растения гибнут зимой.
Семена сохраняют всхожесть до 2-3 лет, молодое растение впервые зацветает на
третьем году жизни. Продолжительность жизни растения составляет до 30-40 лет.
Новоазорский
банановый подорожник, «азорский банан» (Neomusa plantagoidea)
Порядок: Подорожниковые (Plantaginales)
Семейство: Подорожниковые (Plantaginaceae)
Место обитания: Новая Азора, горные районы острова.
Флора островов в эпоху человека отличалась наличием множества эндемичных видов
травянистых растений, которые превратились из мелких видов в гигантов растительного
царства. На Гавайских островах обитали древовидные герани и лобелии, а на острове
Святой Елены ранее рос гигантский вид подорожника Plantago robusta. Это растение
даже получило название «банан острова Святой Елены». Но этим видам не суждено
было надолго пережить человечество. Многие виды, в том числе гигантские подорожники,
вымерли, несмотря на усилия человека по их сохранению. В неоценовой флоре островов
появились новые виды гигантских подорожников, но они не родственны видам, известным
в эпоху человека.
В горных районах Новой Азоры растёт один из таких видов растений. Это растение
немного похоже на банан, но отличается от него округлыми листьями с дуговым
жилкованием. За сходство с бананом растение получило название «азорский банан»,
и официально называется новоазорский банановый подорожник. Однако при ближайшем
знакомстве у этого растения заметно множество отличий от своего прототипа эпохи
человека. Если «ствол» банана был образован черешками листьев, то у новоазорского
бананового подорожника есть настоящий ствол – толстое корневище. Оно растёт
вертикально, может утолщаться с возрастом и ветвиться. Оно покрыто волокнистыми
остатками листьев и грубой корой, защищающей от повреждений. Под поверхностью
почвы тянутся тонкие корневища другого типа: они могут изгибаться вверх на некотором
расстоянии от материнского растения, и образовывать новые вертикальные стволики.
Стволы этого растения волокнистые, упругие и жёсткие. Высота новоазорского бананового
подорожника обычно не превышает 1 метра, но в труднодоступных горных районах
встречаются великаны-долгожители высотой около 2 метров.
Вершины стеблей-корневищ этого растения увенчаны розетками широких овальных
листьев тёмно-зелёного цвета на толстых черешках. Листья опушены короткими густыми
волосками серого цвета. Молодые листья имеют особенно густое опушение, отчего
средняя часть розетки выглядит серебристой. Длина сформировавшегося листа взрослого
растения достигает метра.
Примерно с пятого года жизни стебель начинает формировать соцветия. Соцветие
формируется в точке роста. Это характерный для подорожников початок, покрытый
многочисленными мелкими цветками. Высота соцветия может достигать метра, при
такой же длине цветоноса. Это растение опыляется насекомыми; особенно часто
на цветках новоазорского бананового подорожника можно встретить жуков и различных
ос. Мелкие птицы, а изредка даже летучие мыши, прилетают на эти соцветия, чтобы
покормиться нектаром, но они лишь эпизодически участвуют в опылении «азорского
банана».
Плодоношение новоазорского бананового подорожника очень интересно: когда цветки
опылены, ось соцветия начинает разрастаться в толщину, а цветонос изгибается
к земле. Завязи погружаются в разрастающуюся ось соцветия и покрываются плотной
оболочкой. Мякоть оси соцветия становится рыхлой и сладковатой на вкус, а кожица
приобретает яркий оранжевый цвет. Такое соцветие формирует соплодие, несколько
похожее на соплодие ананаса по принципу формирования. Семена этого растения
распространяют местные куриные птицы – скальные
азорские куропатки. Птицы склёвывают опускающиеся к земле соплодия. Семена,
проходя через желудок птицы, не только не гибнут, но даже повышают всхожесть:
от действия желудочного сока их оболочка истончается, что помогает зародышу
получать воду и минеральные соли, нужные для прорастания и развития. Помёт птиц
даёт дополнительную подкормку, нужную для роста. Но воспроизводство этого вида
тормозится тем, что мелкие птицы часто выклёвывают из растущих соплодий незрелые
завязи.
После плодоношения рост стебля на некоторое время останавливается. Когда начинает
созревать соплодие, около основания цветоноса образуется новая точка роста,
а сильные стебли изредка начинают ветвиться.
«Азорский банан» растёт медленно: крупное растение может достигать возраста
120 – 130 лет, а общая продолжительность жизни достигает 200 лет и более.
Цветолодочник
обыкновенный (Hydroplantago vulgaris)
Порядок: Подорожниковые (Plantaginales)
Семейство: Подорожниковые (Plantaginaceae)
Место обитания: реки, озёра и солоноватые воды Европы и Малой Азии.

Рисунок Nem’а
В эпоху человека различные растения семейства подорожниковых
начинали осваивать водные местообитания. После окончания эпохи человека эволюция
подорожниковых в этом направлении продолжилась, и в неоцене пресные и солоноватые
водоёмы Европы населены различными потомками гидрофильных подорожниковых, принадлежащими
к роду Hydroplantago. Эти растения имеют ряд общих характерных черт: они все
– укореняющиеся или плавающие формы с надводным соцветием в виде одностороннего
спирально закрученного колоса. Цветки очень мелкие, безлепестные, с липкими
рыльцами. Один из листьев в основании соцветия лодочковидный – отсюда название
рода. Он несёт функцию обёртки соцветия, помогая ему держаться на воде. Поверхность
присоцветного листа покрыта густыми тонкими волосками и не смачивается водой.
Остальные листья, как правило, подводные, узкие, расположены на дихотомически
ветвящихся побегах.
В водоёмах Центральной и Южной Европы, а также в близлежащих районах Малой Азии
обитает цветолодочник обыкновенный – один из самых распространённых видов рода.
У этого вида два типа побегов: укореняющиеся и плетевидные. Укореняющиеся побеги
розетковидные, с редуцированными чешуевидными листьями и многочисленными почками,
в пазухах которых развиваются удлинённые плетевидные побеги с длинными междоузлиями
и узкими ланцетовидными листьями светло-зелёного цвета. Этот вид населяет мелководные
участки рек и озёр, хорошо освещаемые и прогреваемые солнцем. Разрастание происходит
сразу и на дне, и в толще воды, в результате чего значительные участки побережья
оказываются заполненными зарослями этого вида. Размножение происходит главным
образом вегетативно.
Опыление у цветолодочника происходит при помощи ветра. Летом на поверхности
воды появляются соцветия, закрытые свёрнутым в трубку покрывалом. В процессе
созревания цветков покрывало отходит от соцветия и отгибается, ложась на поверхность
воды, словно лодка, а соцветие поднимается над поверхностью, где происходит
опыление. Семена мелкие, с твёрдой пористой оболочкой, прорастают неравномерно,
зачастую с задержкой до 2-3 лет.
В реках на юге Европы (за Альпами) обитает наименее специализированный вид –
цветолодочник
простой (Hydroplantago primitiva). Это укореняющийся на дне вид, значительно
отличающийся формой роста: у него образуются розетки с укороченными вегетативными
побегами, выходящими из пазух листьев, и подземные ползучие столоны, дающие
начало новым розеткам. Только цветоносы с присоцветным листом достигают поверхности
воды. Популяции этого вида сильно изолированы друг от друга.
В солоноватых водоёмах на побережье Атлантического океана встречается мелкий
плавающий вид цветолодочник
рдестовидный (Hydroplantago potamogetonis) – обитатель хорошо освещённых
участков водоёма. У него отсутствуют розеточные побеги, а листья очень мелкие,
покрытые водоотталкивающими волосками и густо расположенные на стеблях. Благодаря
быстрому росту этот вид образует на поверхности тихих водоёмов плавающие островки,
способные выдержать вес небольшой птицы или крысы. Соцветия короткие, с небольшим
присоцветным листом, который не ложится на поверхность воды.
Цветолодочник
плавучий (Hydroplantago natans) из рек юго-востока Европы также полностью
перешёл к водному образу жизни и утратил связь с субстратом. Это растение отличается
коротким стеблем с воздухоносными полостями, а его листья малочисленные и крупные,
с остроконечной верхушкой. Жилки листьев утолщены, в них развита воздухоносная
паренхима, играющая роль поплавка. В узлах побегов развиваются многочисленные
корни. Растение образует на поверхности воды сплошной покров из листьев. Покрывало
очень крупное (в 3-4 раза больше обычного листа), лодочковидное, с сильно развитыми
жилками, служит опорой для полегающего соцветия.
Этот вид растений открыл Nem, участник форума.
Кизил
дожделюбивый (Cornus pluviphilus)
Порядок: Кизилоцветные (Cornales)
Семейство: Кизиловые (Cornaceae)
Место обитания: тихоокеанские дождевые леса Северной Америки.
В эпоху неоцена на тихоокеанском побережье Северной Америки сформировался особый
тип растительного сообщества – дождевой лес умеренного климата. С океана постоянно
приходят дождевые облака, проливающиеся дождями на относительно узкой полосе
между океаном и Скалистыми горами. Флора этих мест представлена влаголюбивыми
растениями, обладающими приспособлениями для испарения избытка влаги. Одна из
характерных особенностей местных растений – широкие листья. Даже у местного
вида туи очень широкая хвоя.
В тенистом подлеске таких лесов разрастаются мхи и папоротники, а одним из характерных
видов кустарников является кизил дожделюбивый. Это довольно большое растение
с раскидистой кроной, 3-4 м в высоту, и до 5 м в диаметре. Обычно это растение
образует один главный ствол, хотя достаточно часто от его основания отрастают
боковые побеги, не уступающие ему в размерах. Внешне, несмотря на размеры, кизил
дожделюбивый напоминает скорее куст, и образует обширные заросли. Очень редко
это растение вырастает одноствольным деревом. Кора имеет очень характерную окраску.
Молодые побеги одеты красной гладкой корой, которая со временем темнеет; на
стволе кора тёмно-бурая, покрытая мелкими трещинами.
Листья растения крупные, тёмно-зелёные, эллиптические с оттянутым кончиком,
образующие «капельницу», более характерную для растений влажного тропического
леса. В этом месте у растения расположены гидатоды, выводящие избыток влаги
в виде капель чистой воды. Молодые растущие листья розоватые; с возрастом окраска
меняется на зелёную, а розоватый цвет сохраняется на жилках, но со временем
исчезает. Осенняя листва тёмно-красная, растение листопадное. Зимой становится
особенно хорошо заметной окраска молодых ветвей.
Кизил цветёт в начале лета и опыляется животными; его часто посещают колибри
и бабочки-бражники. У его голоценового предка (скорее всего, Cornus nuttallii)
цветы были относительно небольшими, тогда как у кизила дожделюбивого они выглядят
большими белыми звёздами, которые очень заметны на фоне тёмно-зелёных листьев
этого дерева. Головчатое соцветие из мелких цветков окружено 5-6 крупными прицветниками
белого или слегка розоватого цвета. Широкие и плоские соцветия кизила удобны
для жуков, которые садятся на них, поедают нектар и часть пыльцы, и переносят
пыльцу с одного цветка на другой.
Плоды кизила дожделюбивого – крупные белые ягоды, сплюснутые на вершине и у
плодоножки. При созревании они очень хорошо заметны на фоне осенней листвы растения.
Они не очень питательные, а их мякоть довольно водянистая. Но местный вид соек
охотно поедает их; мякоть переваривается, а семена распространяются с помётом.
Также ягоды поедают травоядные млекопитающие. Молодое растение впервые зацветает
на третьем-пятом году жизни. Кизил дожделюбивый может прожить несколько десятков
лет.
Этот вид растений открыл Bhut, участник форума.
Поддубник
североамериканский (Monotropa querciphila)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae), подсемейство: Вертляницевые (Monotropoidae)
Место обитания: дубовые леса на востоке Северной Америки.
Предком этого растения был подъельник обыкновенный (Monotropa hypopitys) – очень
необычное растение, паразит на грибнице разных грибов, лишившийся хлорофилла,
и выглядящий словно химически обесцвеченным. Несмотря на отсутствие хлорофилла,
подъельник оказался относительно выносливым растением, тенелюбивым и способным
выживать, пока были живы питающая его грибница и те деревья, с которыми состояла
в симбиозе пронизывающая это растение грибница.
В неоцене союз с грибами пригодился растению: этот предок породил несколько
новых видов, отчасти благодаря восстановлению местообитаний после исчезновения
людей. Эволюция этих растений пошла по пути специализации к паразитизму. Один
из видов рода, поддубник североамериканский, паразитирует на разных видах дубов,
которые растут в восточных лесах Североамериканского материка.
Это растение средних размеров, около 10 см в высоту. Как и у предкового вида,
у него нет хлорофилла и ткани окрашены в бледно-жёлтый цвет. Этот вид зависит
от грибов только в ранний период жизни, и по мере развития внедряется в корни
деревьев подходящих видов, чаще всего дубов (отсюда название), и продолжает
развиваться за счёт этих растений. Его корни глубоко внедряются в древесину
разных видов дубов, и если дуб слишком молод или стар, то поддубник иногда может
погубить всё дерево.
Исключением становится лишь восточный горный
дуб (Quercus appalchiensis). В процессе эволюции этот вид дерева выработал
своеобразный иммунитет к паразитированию поддубника: он вырабатывает вещества,
которые ограничивают рост и развитие поддубника, хотя и не убивают его. Таким
образом, это растение может жить на данном виде дуба в несколько «подавленном»
состоянии, не угрожая жизни дерева-хозяина.
Поддубник размножается при помощи мельчайших семян, которые распространяются
даже слабыми дуновениями ветра. Семена попадают на поверхность почвы и прорастают
лишь при попадании в них грибницы почвенных грибков. Окрепшие растения формируют
короткое подземное корневище и выпускают в разные стороны корешки, которые находят
корни деревьев-хозяев и прорастают в них; если это удаётся, растение быстро
развивается и формирует многочисленные придаточные почки в месте проникновения
в корень хозяина. Некоторое время спустя формируются первые цветки – обычно
в конце весны и начале лета. Их запах кисловато-пряный, и их опыляют разные
насекомые подлеска, чаще всего жуки или одиночные пчёлы. К зиме плоды успевают
сформироваться и распространить семена, после чего надземная часть растения
отмирает, и растение зимует в виде корневищ или тяжей тканей под корой дерева-хозяева.
Это растение поедается разными видами зайцев и кроликов, а также лесными видами
пекари.
Поддубник североамериканский отличается значительным долголетием и часто живёт
столько же, сколько и его дерево-хозяин.
Этот вид растений открыл Bhut, участник форума.
Эпигея
горная (Alloepigaea montana)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae)
Место обитания: горные леса на востоке Северной Америки.
Горные леса на востоке, и отчасти на северо-востоке Северной Америки несколько
отличаются от горных лесов на западе: они суше лесов тихоокеанского побережья
из-за особенностей розы ветров, а склоны гор более пологие, поэтому фауна и
флора более однородны, без резких различий на восточных и западных склонах,
как это наблюдается в Скалистых горах. В целом флора Аппалачей – это широколистные
деревья (хвойные встречаются реже), главным образом разные виды дубов (хвойные
более разнообразны), и тенелюбивые травянистые растения, которые процветают
в этих лесах. Некоторые из них превратились
в паразитов, которые обходятся без хлорофилла, но таковы лишь немногие виды.
Среди растений более традиционного образа жизни типичным обитателем лесов Аппалачей
является горная эпигея, потомок североамериканской эпигеи ползучей (Epigaea
repens). Однако этот вид имеет гибридное происхождение – некоторая часть генов
унаследована от азиатских видов и их садовых гибридов, культивировавшихся в
эпоху человека.
Горная эпигея стелется по земле, давая многочисленные одревесневающие побеги,
легко укореняющиеся в лесной подстилке. Предковый вид эпигеи был вечнозелёным
растением, и горная эпигея сохранила его черты: она нечувствительна к холоду
и спокойно зимует под снегом. Однако образ жизни этого вида заметно изменился
по сравнению с предками – это растение находится на пути приспособления к паразитическому
образу жизни. Зимой она сбрасывает старые листья, сохраняя их лишь на концах
побегов, и существует как полупаразит, поглощая питательные вещества из корней
деревьев, к которым присосалась корнями – обычно это разные виды дубов. Один
из видов деревьев, восточный горный дуб,
содержит в своём соке вещества, которые губят паразитические растения и заглушают
их рост, а другие виды деревьев могут и погибнуть, особенно молодые экземпляры.
Весной, как только растает снег, эпигея выпускает новые листья и переходит на
самостоятельное питание за счёт фотосинтеза. Её листья – непропорционально широкие,
округлой формы, тёмные. Они поглощают очень много солнца, особенно когда на
деревьях только начинает появляться листва. На это время приходится пик паразитизма
со стороны растения, которое активно наращивает зелень и вытесняет своих конкурентов
в подлеске. Рано весной эпигея уже выпускает собственные цветы – небольшие,
бледные бело-розовые звёздочки с пятью лепестками и более тёмной серединой,
которые будут цвести до начала лета, когда появятся другие горные цветы и разрастутся
травянистые растения. К тому времени разные насекомые уже успевают опылить эпигею,
и на месте цветов у неё появятся ягоды, отдалённо напоминающие по форме землянику.
Эти ягоды поедаются разными травоядными и всеядными зверями и птицами, но семена
эпигеи устойчивы к их желудочному соку. Пройдя через пищеварительную систему
животного, они только лучше прорастают.
Горная эпигея может успешно размножаться ползучими побегами и корнями, вернее
даже отростками корней, если часть корневой системы погибнет, выкопанная из
земли местными травоядными. На оторванных от растения корнях появляются придаточные
почки, образующие полноценные побеги. Растение способно долгое время существовать
самостоятельно, не паразитируя на чужих корнях, но при этом хуже цветёт. В присутствии
подходящих видов деревьев-хозяев эпигея горная легко переходит к паразитическому
образу жизни.
Этот вид – многолетний кустарник, способный прожить до 40-50 лет.
Этот вид растений открыл Bhut, участник форума.
Водяника
шилолистная (Empetrum subulatum)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae)
Место обитания: Антарктида, болота и заросли мхов у основания ледников, горы
ниже линии снегов.
Во время одного из плейстоценовых оледенений водяника из Северной Америки смогла
преодолеть экваториальные области Земли и расселиться в Южном полушарии. На
крайнем юге Южной Америки растение дало жизнеспособные популяции, превратившиеся
в полноценные обособленные виды в раннем неоцене, а после отступления ледников
в Антарктиде водяника с Огненной Земли оказалась одним из поселенцев на свободном
от ледников побережье этого материка. Благодаря неприхотливости она быстро расселялась
по освобождающимся по ледникового покрова землям, и в неоцене является широко
распространённым видом в Антарктиде.
Водяника шилолистная – это многолетнее растение с ползучими одревесневающими
стеблями и красными ягодами, которые при созревании привлекают птиц окраской.
На ягодах имеется тонкий слой воска, отражающий ультрафиолетовые лучи – это
помогает ягодам быть заметными для птиц издалека. Этот вид – вечнозелёное растение,
у которого очень узкие листья с подвёрнутыми краями, напоминающие хвою, длиной
до 20 мм. Специфическая форма листьев связана с необходимостью экономить влагу
при жизни на холодных болотах. Заросли этого растения напоминают низкорослый
злак и отличаются от него невзрачными цветками, появляющимися у самой земли,
на месте которых летом развиваются яркие ягоды. Мякоть ягод сладкая, но очень
водянистая.
Побеги этого растения достигают длины до 100 см. Одно растение образует несколько
крупных ветвистых побегов и занимает площадь до 3 квадратных метров. Лежащие
на субстрате побеги легко укореняются, закрепляясь в мельчайших неровностях
субстрата. Короткие корни покрыты узелками – в них поселяются азотфиксирующие
бактерии, помогающие растению добывать нужный для роста азот. Также на корнях
растения селятся почвенные грибки, снабжающие его питательными веществами. Благодаря
таким союзникам растение хорошо разрастается даже на самой скудной и бесплодной
почве и является важным видом пионерной растительности, способствующим почвообразованию.
При этом водяника шилолистная отличается низкой конкурентоспособностью и в смешанных
зарослях легко вытесняется другими видами растений.
Медленный рост этого растения компенсируется значительным долголетием – растение
способно прожить до 50 лет и больше. Часто при отмирании основного корня и гибели
центрального побега боковые ветви продолжают расти, образуя кольцевидный клон.
Квижуктаат
(Cassiope qijuktaat)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae)
Место обитания: североамериканская тундра.
Летом североамериканская Арктика в неоцене – довольно шумное место. Разные виды
гусей и уток, а также чистиковых птиц образуют там большие птичьи базары, которые
привлекают разных хищных зверей и птиц, которые пытаются на них охотиться. Множество
птиц спешит обзавестись потомством, используя все преимущества полярного лета
и долгого полярного дня. Жизнь обитателей тундры кипит на фоне местной растительности,
которая эволюционировала, реагируя на наступление и таяние ледников, и на сопутствовавшие
им изменения границ природных зон. Как и в эпоху человека, в неоцене в тундрах
преобладает низкорослая травянистая и полукустарниковая растительность.
Из-за малой населённости тундры людьми растения, обитавшие в этой природной
зоне, понесли наименьший ущерб от деятельности человека, и в неоцене в тундрах
Северного полушария произрастают их потомки. Один из таких видов – квижуктаат,
потомок кассиопеи четырёхгранной (Cassiope tetragona) эпохи голоцена: «квижуктаат»
– название кассиопеи у эскимосов. Это довольно высокое растение по меркам тундровой
растительности – примерно 25-30 см в высоту. Это кустарник с ползучими стволами
и крупными ветвями. Распростёртый по земле куст образует множество вертикальных
фотосинтезирующих побегов. Молодые ветви покрыты толстыми чешуевидными листьями
тёмно-зелёного цвета. На старых ветвях, лежащих на земле, листья засыхают, но
не отпадают и формируют защитный покров. Концы побегов с молодым приростом красноватые,
а вершины побегов окрашены в малиново-красный цвет, поэтому заросли растения
в фазе активного роста приобретают своеобразный вид.
Среди листьев прорастают вертикальные цветоносы, выносящие над кончиками побегов
цветы колокольчатой формы (как и у предкового вида), окрашенные в бледно-розовый
цвет (внутренняя сторона лепестков, пестик и тычинки более яркого оттенка).
Часто встречается белоцветковая морфа этого вида. Квижуктаат – важный медонос
и кормовое растение для многих насекомых североамериканской тундры; даже самцы
кровососущих насекомых охотно кормятся на этих цветках. Позвоночные животные
вроде диких гусей охотно поедают ветки с молодым приростом, а особенно листья-чешуйки
этого верескового растения. Кормящиеся птицы буквально «сбривают» листья, протягивая
побеги сквозь полусомкнутый клюв. Травоядные млекопитающие объедают фотосинтезирующие
побеги почти до земли.
Квижуктаат устойчив к полярной зиме и к холодам. Его сок содержит смолистые
вещества, которые действуют как антифриз и накапливаются к зиме, делая растение
менее съедобным, чем летом. Однако это не останавливает косорогов,
которые выкапывают квижуктаат с корнями и поедают нижние части растения, где
концентрация этих веществ понижена. Таким образом за зиму они наносят растению
большой ущерб, но это растение очень живуче: не съеденные ими побеги могут укорениться
весной, а остатки стеблей быстро обрастают новыми зелёными побегами. К концу
полярного лета растение часто восстанавливает прежнее изобилие, хотя в период
восстановления хуже цветёт. Квижуктаат успешно размножается семенами, а ползучие
ветви легко укореняются, что помогает ему выживать в присутствии крупных травоядных
животных.
Молодое растение впервые зацветает на третьем году жизни, а предельный возраст
квижуктаата может достигать 20-25 лет и более.
Этот вид растений открыл Bhut, участник форума.
Кинникинник
(Arctostaphylos kinnikinnick)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae)
Место обитания: североамериканская лесотундра и тундра.
Североамериканская тундра в неоцене – богатое жизнью место, особенно полярным
летом, когда с мест зимовки прилетают многочисленные птицы, а растения стараются
провести с пользой каждый день короткого и прохладного северного лета. На сухих
водоразделах находятся заросли неарктической сосны,
в других местах покачиваются зелёные волны зарослей квижуктаата,
украшенные розоватой пеной мелких цветков. Среди камней растут миниатюрные растения
– полярная камнеломка аупиллатингват
или краснеют ягоды кинникинника.
Кинникинник – это потомок толокнянки, или медвежьей ягоды (Arctostaphylos uva-ursi)
эпохи голоцена. Это достаточно высокое растение – ползучий полегающий вечнозелёный
кустарник около 30 см в высоту, часто намного ниже. На северной границе ареала
растение едва приподнимается над землёй. Побеги легко укореняются, корневая
система поверхностная, мочковатая. Листья этого растения округлые, кожистые
и тёмно-зелёные сверху, светло-зелёные шероховатые снизу. По краю тянется густая
бахрома из белых волосков. Цветки растения отдалённо напоминают цветки квижуктаата:
это тоже колокольчики, но более мелкие и узкие и с заметным розово-красным оттенком
внутри: насекомые-опылители чётко различают эти два вида цветов и не смешивают
их – одни больше опыляют квижуктаат, другие – именно кинникинник, однако строгой
специализации опылителей нет. Цветение начинается рано весной, вскоре после
таяния снега. Цветки легко переносят заморозки.
Как и предковый вид, кинникинник является ягодоносным растением: в период плодоношения
растения бывают сплошь покрыты ярко-красными ягодами с суховатой крахмалистой,
но сладкой мякотью. Это одно из немногих ягодных растений в североамериканской
тундре, поэтому в конце лета многие звери, и даже некоторые птицы вроде гуся-путешественника,
охотно и помногу поедают его ягоды – причём это делают не только такие травоядные
звери как косороги,
но и всеядные с уклоном в хищничество геопоссумы.
Причём эти сумчатые поедают не только ягоды, но и зелень растения: она содержит
вещества, укрепляющие здоровье этих сумчатых. В результате звери разных видов
нередко выедают заросли кинникинника подчистую, но семена этого растения, прошедшие
сквозь пищеварительную систему млекопитающих прорастают быстрее, чем без обработки
их желудочным соком. Кроме того, семена и молодые растения кинникинника легче
переносят зимний холод, чем взрослые экземпляры, вымерзающие в суровые малоснежные
зимы. По сравнению с другими полярными растениями кинникинник быстрее растёт
и легче переносит повреждения, нанесённые травоядными зверями. Молодое растение
плодоносит уже на третьем году жизни, а пик плодоношения наступает на десятом
году. Продолжительность жизни часто достигает 50-60 лет.
Этот вид растений открыл Bhut, участник форума.
Клубничная
клюква (Vaccinium fragarioides)
Порядок: Верескоцветные (Ericales)
Семейство: Вересковые (Ericaceae)
Место обитания: болота, тайга, лесотудра, тундростепь, долины рек и побережья
озёр Западной Сибири.
Клубничная клюква – это потомок клюквы обыкновенной (Vaccinium oxycoccus). И
если её предок был стелющимся вечнозеленым кустарничком, то в тайге и лесотундре
клубничная клюква значительно увеличилась в размерах, превратившись в одревесневший
кустарник, который в благоприятных условиях может вырастать до 2 метров высоту,
в тундростепи и в болотах одревесневшие стебли распростёрты по земле.
Корневая система стержневая. На корнях этого растения живёт гриб эндофома
симбиотическая, гифы которого плотно соединяются с клетками корня и образуют
микоризу. Гифы гриба активно всасывают из почвы питательные вещества и передают
их корням растения.
Листья мелкие, очерёдные, длиной до 20 мм, шириной до 10 мм продолговатые с
коротким черешком. Листовая пластинка кожистая, тёмно-зелёная, снизу, пепельно-серая,
остающаяся на зиму. Осенью опадает около четверти самой старой листвы. На нижней
поверхности листа находится воск, делающий нижнюю сторону листа несмачивающейся
и таким образом растение защищающий от нарушения его нормальных функций.
Растение цветёт в конце весны ярко-розовыми цветами. Цветки актиноморфные, поникшие,
на достаточно длинной цветоножке (до 7 см). Цветки, в отличие от предкового
вида, с пятью лепестками, лепестки отогнуты назад. Тычинок восемь, пестик один,
завязь нижняя. Продолжительность существования одного цветка – до 18 дней. Через
некоторое время появляются сладкие ярко-красные шаровидные ягоды диаметром до
25-30 мм (отсюда название: по размеру ягода похожа на ложный плод у клубники)
– это одни из самых крупных ягод из всех вересковых растений неоцена. Для клубничной
клюквы характерна зоохория: ягоды поедают многие звери (зайцелопы,
хандагаи, келпи и
др.) и птицы, которые переносят семена на большие расстояния и распространяют
их с помётом. Ежегодно одно растение образует несколько сотен ягод.
Ягоды клубничной клюквы представляют собой желанное лакомство для поедающих
их животных. Плоды богаты сахарами (глюкоза и фруктоза), органическими кислотами,
пектиновыми веществами, витаминами, макро- и микроэлементами, являясь важной
добавкой к рациону местных животных.
Молодое растение развивается очень медленно, давая в первые годы прирост длиной
несколько сантиметров. После развития грибницы симбионта рост растения ускоряется.
Первое плодоношение бывает в возрасте 13-16 лет, продолжительность жизни – до
100 лет и более.
Этот вид растений открыл Wovoka, участник форума.
Североамериканский
колокольчик (Campanula popularis)
Порядок: Астроцветные (Asterales)
Семейство: Колокольчиковые (Campanulaceae)
Место обитания: североамериканские прерии, луга, леса и лесотундра в области
умеренного и холодного климата.
Многие виды колокольчиков выглядят хрупкими и уязвимыми растениями, но это впечатление
обманчиво: на самом деле колокольчики являются очень выносливыми растениями,
легко образующими разнообразные жизненные формы. Эволюционная гибкость и культивирование
человеком помогли им пережить и антропогенный прессинг голоцена, и ледниковый
период начала неоцена, и продолжить процветать и эволюционировать.
Североамериканский колокольчик является одним из самых распространённых видов
этого семейства в Северной Америке. Он произрастает повсюду к востоку от Скалистых
гор – и на открытых пространствах, и в относительно тенистых, но сухих широколиственных
и смешанных лесах востока материка, и в более холодных лесах севера, и даже
в лесотундре.
Этот вид образует побеги двух типов. Летом во время цветения это растение образует
высокие прямостоячие побеги, типичные для колокольчиков, с упругими тонкими
стеблями, многочисленными голубыми цветками колокольчатой формы и длинными и
узкими листьями. Весной растение даёт розетку широких округлых листьев, которые
лучше поглощают солнечный свет и позволяют североамериканскому колокольчику
расти быстрее конкурентов. К середине лета растение выпускает длинные стебли
с «летними» листьями и цветами, которые охотно посещают разные насекомые – это
растение не специализировано к определённым видам опылителей. Крупные животные
поедают эти цветы, иногда вырывая и повреждая корни. Подземная часть растения
– короткое многолетнее корневище, от которого отрастают вертикальные побеги.
Этот вид не образует ползучих подземных побегов и разрастается в виде компактных
куртин.
На большие расстояния североамериканский колокольчик распространяется семенами.
Семена находятся во вскрывающихся коробочках, которые «взрываются», когда созревают,
разбрасывая семена на несколько метров в стороны. В результате этого данный
вид растения встречается локально и растёт многочисленными группами на небольшой
площади. Осенью стебли отмирают до уровня почвы, а под снегом зимуют только
корневища с корнями, которые прорастают весной после таяния снега. В северной
части ареала растения зимуют с прикорневой розеткой листьев.
Корневище растения часто поедают грызуны и личинки жуков. Если основное корневище
не будет съедено, то североамериканский колокольчик сможет прожить 8-10 лет,
а то и больше.
Этот вид растений открыл Bhut, участник форума.
Кобольдов
колокольчик (Campanula coboldiana)
Порядок: Астроцветные (Asterales)
Семейство: Колокольчиковые (Campanulaceae)
Место обитания: Альпы, каменистые участки лугов и горно-тундровый пояс у границы
снегов.
В эпоху человека Альпы были одним из центров видового разнообразия колокольчиков
в Европе – здесь встречалось большое количество редких узкоареальных и эндемичных
видов. В эпоху господства человека значительное количество видов этих растений
вымерло из-за разрушения и загрязнения мест обитания, изменений климата и уничтожения
самих растений. Особенно уязвимыми оказались узкоареальные виды, чувствительные
к изменениям среды обитания. В эпоху неоцена процессы горообразования в Альпах
продолжаются из-за сближения Европы с Африкой, и это привело к появлению новых
мест обитания, которые заселили специализированные эндемичные виды, появившиеся
уже после человека.
В поясе горных лугов и тундры немного ниже линии снегов обитает специализированный
высокогорный вид колокольчиков – кобольдов колокольчик. Это низкорослое травянистое
растение, образующее густую подушку укороченных побегов. Высота побегов не более
5 см, но они интенсивно ветвятся у основания и плотно прилегают друг к другу.
Листья округлые, очередные, часто расположены на стебле, образуя густые побеги-розетки.
Молодые листья и точки роста обильно опушены для защиты растущих тканей от ультрафиолетового
излучения; на зрелых листьях опушение сохраняется лишь на нижней стороне, предотвращая
потери влаги. Этот вид – многолетнее вечнозелёное растение, произрастающее на
каменистых почвах. Короткое подземное корневище находится в почве на глубине
1-2 см, часто прорастает между камнями.
Во время цветения растение становится лишь немного выше: маленькие одиночные
цветки кобальтово-синей окраски раскрываются поодиночке на тонких побегах высотой
6-8 см. Они не специализированы к определённому типу опылителей, на них кормятся
мухи, мелкие жуки и бабочки. Это растение является важным кормовым ресурсом
для насекомых: кобольдов колокольчик зацветает сразу после таяния снега, цветение
продолжается до середины лета.
Семена созревают на следующий год рано весной и разносятся потоками талой воды.
Плод – невскрывающаяся коробочка, которая служит поплавком и содержит 3-4 семени.
Оболочка плода постепенно впитывает воду, благодаря которой семена набухают
и прорастают. После прорастания самый сильный проросток заглушает рост остальных
и развивается во взрослое растение. Первое цветение наступает на третьем году
жизни; продолжительность жизни достигает 20 лет.
Стенокампана
лазающая (Stenocampana scandens)
Порядок: Астроцветные (Asterales)
Семейство: Колокольчиковые (Campanulaceae)
Место обитания: Южная Азия, тропические леса.
Эпоха восстановления лесов в раннем неоцене – это время активного видообразования
и интенсивной конкуренции между растениями, образующими природное сообщество.
В это время растения различных семейств пытались занять новые экологические
ниши, и в результате среди них возникали жизненные формы, неизвестные в эпоху
человека. Среди таких растений были, например, бегонии,
виноградовые и астровые,
образовавшие древовидные формы. Семейство колокольчиковых, наиболее разнообразное
во флоре Старого Света, было представлено преимущественно травянистыми растениями.
Но в эпоху восстановления лесов им также удалось вырваться в крупноразмерный
класс – но они стали не деревьями, а лианами. Один из крупнейших видов лиан
в этом семействе – стенокампана лазающая, которая представляет собой слабо одревесневающую
быстрорастущую лиану. Она встречается на участках повреждённого древостоя и
быстро разрастается, давая прирост до 10 метров за год. Стебель взрослого растения
достигает толщины 10-15 см, длина стеблей взрослого растения – до 40 метров.
Нижняя часть стебля у этого растения одревесневает, но древесина мягкая и непрочная.
Кора гладкая, растрескивается продольными трещинами и спадает со стебля полосками.
Стенокампана оплетает деревья и кустарники, прикрепляясь к ним короткими придаточными
корнями. Стебель ветвится преимущественно в верхней части, вблизи кроны дерева,
являющегося опорой для растения. Листья крупные, цельные, одиночные, на длинных
упругих черешках. Листовая пластинка достигает 20-25 см в длину и 15 см в ширину,
овальных очертаний. Кончик листа вытянут в узкую капельницу длиной 2-3 см. Поверхность
листа гладкая, светло-зелёная с тонким восковым налётом. В сухой сезон часть
листвы с нижней части стебля опадает, листья остаются только на молодых побегах.
В конце сухого сезона растение зацветает. На старом приросте просыпаются пазушные
почки, из которых прорастают укороченные побеги, образующие одиночные цветки
в пазухах листьев. У этого вида своеобразные длинные узкие трубчатые цветки
с небольшим отгибом – отсюда название, означающее «узкий колокол». Длина цветка
достигает 12-14 см, упругая цветоножка вырастает до 10 см. Околоцветник голубого
цвета с отражающими ультрафиолетовую часть спектра отметками в зеве, обозначающими
путь к нектару для насекомых-опылителей. Цветки повислые или направленные почти
горизонтально, покачивающиеся на цветоножках. Этот вид растений опыляется бражниками
и другими крупными бабочками с длинными хоботками.
Плод – невскрывающаяся ягода с суховатой сладкой мякотью и кожицей красного
цвета. Обычными распространителями плодов являются древесные и наземные птицы.
Прорастание семян происходит значительно успешнее после их прохождения через
пищеварительный тракт птицы. Молодое растение зацветает на втором году жизни,
достигает максимальных размеров в возрасте 10 лет. Продолжительность жизни –
свыше 50 лет.
Астрека
наиколючейшая (Astreca spinosissima)
Порядок: Астроцветные (Asterales)
Семейство: Астровые, или Сложноцветные (Asteraceae)
Места обитания: Северная Африка, граница Средиземноморских болот.
Высыхание в позднем голоцене Средиземного моря существенно повлияло на климат
местностей, окружавших его ранее. Территория бывшего Средиземноморья стала более
засушливой, а ветра из Средиземноморской котловины несут тучи солёной пыли,
оседающей далеко в глубине материков. В таких условиях мало какое растение сможет
нормально существовать, но некоторые виды, адаптировавшиеся к подобным условиям,
процветают.
На бывшем африканском берегу Средиземного моря среди чахлой травы кое-где поднимаются
группы интересных растений высотой около 2 метров. На первый взгляд они похожи
на небольшую пальму с колючим стволом, но, если подойти поближе, становится
очевидно, что это совсем не родственник пальмы.
Это причудливое растение – астрека наиколючейшая, один из обитателей засушливых
местностей, окружающих Средиземноморскую низменность. Название «астрека» составлено
из слов «астра», показывающего принадлежность растения к семейству астровых,
и «арека», то есть «пальма».
Толстый суккулентный стебель астреки имеет пять продольных острых рёбер, и его
поперечный срез похож на звезду. Каждое ребро обильно усажено острыми крепкими
колючками, надёжно оберегающими растение от крупных травоядных. Стебель слабо
ветвится, давая от основания два – три крупных боковых побега. Стволы астреки
покрыты плотной кожицей, в нижней части одревесневают.
Но колючки на стволе – это не главная черта астреки, придающая ей «пальмовый»
облик. Стебли этого растения увенчаны короной тёмно-зелёных перисторассечённых
листьев длиной около метра. Вдоль средней жилки листа проходит светлая серебристая
полоска. Под живыми листьями бахромой свисают остатки высохших прошлогодних
листьев – хотя они погибли, но всё ещё выполняют жизненно важную роль, предохраняя
стебель от солнечных ожогов. Каждый лист сверху и снизу вдоль главной и крупных
боковых жилок имеет ряды острых тонких колючек – надёжное оружие от зубов прожорливых
травоядных животных. Отмершие листья не опадают ещё и потому, что их колючки
столь же исправно участвуют в защите растения.
На нижней поверхности листьев густо растут серебристо-белые волоски, придавая
изнанке характерную войлокообразную структуру. Такое приспособление помогает
меньше испарять воду: устьица находятся на нижней стороне листа, и волоски смягчают
контраст между средой внутри листа и иссушающей жарой снаружи.
Растения семейства сложноцветных – чемпионы вегетативного размножения. Астрека
образует на метровой глубине под землёй обширную сеть корневищ, на которых образуются
отпрыски. Поэтому одно растение в подходящих условиях быстро разрастается в
своеобразную редкую рощицу.
Во время влажного сезона астрека быстро разрастается. За несколько месяцев растение
прибавляет в росте около 20 см, и образует около двух десятков крупных листьев.
В условиях обильного увлажнения на старых 7 – 10-летних стволах развиваются
цветоносы. Соцветия астреки – небольшие жёлтые корзинки обоеполых цветков, окружённые
сухими красно-коричневыми листочками обвёртки соцветия. Корзинки собраны в крупное
общее щитковидное соцветие на длинном цветоносе. Оно развивается на верхушке
старого ствола растения и видно издалека. Опыляют астреку в основном одиночные
осы и бабочки, хотя изредка на соцветиях растения можно увидеть жуков и мух.
К началу сухого сезона созревают семена астреки. После их созревания стебель
растения, давший цветонос, отмирает. Но возле него уже подрастает молодая поросль,
которая скоро окрепнет.
В пустыне не так уж много зверей, которые могли бы стать распространителями
семян, поэтому астрека избрала простую жизненную стратегию, характерную для
множества видов растений: её семена переносятся ветром. Крошечные семена астреки
имеют белые парашютики, с помощью которых переносятся на большие расстояния.
Таким способом астрека расселилась по всем берегам Средиземноморских болот,
и даже закрепилась на некоторых горах Средиземноморской низменности, образовав
несколько хорошо различающихся видов.
Семена астреки долго сохраняют всхожесть, годами дожидаясь благоприятных условий.
Во время дождя они быстро прорастают, и молодой росток начинает формировать
розетку листьев на поверхности, а его длинный корень тянется к влажным глубинным
слоям почвы. На второй год жизни начинает формироваться надземный ствол, достигающий
полного развития к 10-летнему возрасту. На третьем году жизни растение начинает
выбрасывать в разные стороны горизонтальные корневища, от которых отрастают
новые побеги.
В Северной Африке, на островах Средиземноморских болот и в Южной Европе обитает
ещё несколько родственных видов этого растения:
Астрека
астрофитумовидная (Astreca astrophytoides) – обитатель сухих горных
районов Средиземноморской низменности. Отличается коротким толстым стволом (высота
растения – около полуметра) с крупными мясистыми рёбрами, покрытыми короткими
колючками. Форма ствола этого вида астреки напоминает кактус астрофитум (Astrophytum).
Листья узкие, снизу густоопушённые, сверху покрыты многочисленными острыми волосками.
Цветонос очень короткий, соцветие состоит всего из 3 – 4 корзинок жёлтого цвета
с розовыми сухими листочками обвёртки.
Чаще всего этот вид встречается в трещинах скал небольшими группами, избегая
засоленных почв.
Астрека пальмовидная
(A. palmetto) – встречается в Северо-Западной Африке, на Гибралтарском
перешейке и в Южной Европе. Обитая в относительно более влажных условиях, этот
вид характеризуется высокими стволами (до 5 метров в высоту), большим количеством
длинных узких листьев (длиной около 1 метра при ширине примерно 10 см). Листья
перистонадрезанные, на конце каждого зубца торчит острая колючка. Ствол 5 –
6-рёберный, рёбра узкие, слегка волнистые, покрыты одиночными длинными колючками.
Цветонос длинный (до 1 метра), несёт большое количество мелких корзинок с белыми
листочками обвёртки.
Этот вид встречается отдельными группами вдоль русел рек и возле временных водоёмов,
предпочитая относительно увлажнённую почву.
Бесстебельник
толстокорневой (Taraxacaulis crassiradix)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteraceae)
Место обитания: полупустынные районы Средиземноморской низменности.
Суровые условия жизни приводят к появлению у животных и растений разнообразных
приспособлений, позволяющих им выживать. В эпохи, предшествующие неоцену, климат
на Земле был суровым и контрастным, вызвав к жизни множество видов живых организмов,
приспособленных к экстремальным условиям. В неоцене климат стал гораздо более
мягким, и виды, приспособленные к жёстким условиям среды обитания, начали вымирать.
Но даже в неоцене на Земле сохранились места, условия жизни в которых очень
суровые. Они стали своеобразными «заповедниками» и «полигонами» для растений,
обитающих в экстремальных условиях, и в этих местах эволюция продолжила «оттачивать»
их адаптацию. Один из таких районов – местность вокруг бывшего Средиземного
моря, превратившегося в скопление солёных болот и озёр в гигантской котловине,
бывшей некогда его дном.
В эпохи крупных геологических или климатических катаклизмов с большей вероятностью
выживают самые нетребовательные и широко распространённые виды. Поэтому неудивительно,
что предком одного из интересных растений полупустыни по краю Средиземноморских
болот был обычный одуванчик (Taraxacum officinalis). Впрочем, в эпоху человека
деление одуванчиков на виды было крайне неопределённым из-за большой изменчивости
этих растений. Очевидно, в этом и скрывался залог успеха этих растений в неоцене.
Потомок одуванчика, бесстебельник толстокорневой, обитает в засушливых районах
Азии и Северной Африки, предпочитая каменистые и глинистые почвы. Он представляет
собой розеточную траву, несколько напоминая своего предка. У этого растения
листья гораздо крупнее, чем у одуванчика: около 30 – 40 см длиной, с толстой
главной жилкой и перистонадрезанными краями. На угловатых лопастях листьев по
краю торчат тонкие острые колючки. Нижняя поверхность листа покрыта сероватым
войлочным налётом из волосков, верх глянцевый. Молодые листья густо покрыты
волосками даже сверху, но по мере развития листа опушение остаётся только на
нижней поверхности. Листья лежат на земле широкой розеткой, и прошлогодние листья
могут оставаться на растении ещё один – два года, защищая растение от травоядных
животных, а почву под ним – от перегрева.
На глубине примерно метра под поверхностью почвы располагается самая важная
часть растения – толстый главный корень, образующий своего рода «корнеплод».
У самых крупных экземпляров он может достигать длины полутора метров при толщине
более полуметра и весе около 100 кг. Он покрыт тонкой эластичной корой, и от
него отходит несколько боковых корней. Здесь находится запас воды, заботливо
собираемой растением во время редких дождей. В это время на поверхности почвы
появляются многочисленные всасывающие корешки, впитывающие дождевую воду и росу.
В засуху они отсыхают, но растение не испытывает недостатка в воде: его главный
корень тянется вглубь почвы на 20 – 30 метров и больше, достигая водоносных
слоёв. Даже если животное вроде насекомого или грызуна сумеет докопаться до
«корнеплода», ему от этого мало пользы: сок растения содержит большое количество
латекса, и густеет на воздухе. На вершине «корнеплода» находится святая святых
– точка роста. Если она окажется повреждённой, растение прекращает рост, но
на следующий год в начале сезона дождей развивает две, или даже три точки роста,
развивающиеся из спящих почек.
В начале сезона дождей сморщенный «корнеплод» постепенно впитывает воду, и растение
слегка «приподнимается», выпуская из-под земли порцию молодых листьев. А в сухой
сезон корни сжимаются, и втягивают «корнеплод» глубоко в почву, оставляя на
поверхности только отмирающие листья.
В краткий сезон дождей бесстебельник толстокорневой зацветает. Цветоносы этого
вида короткие, и жёлтые соцветия-корзинки почти лежат на земле в середине розетки,
образованной листьями: так растение меньше заметно издали для травоядных млекопитающих.
Но оно привлекает к себе немногочисленных опылителей (преимущественно жуков
и мух) специфическим запахом: смесью приятного цветочного и гнилостного ароматов.
Цветение длится недолго: не больше недели. После опыления семена быстро созревают,
а цветонос начинает расти. Он выносит созревающие семена на метр над почвой.
Семена бесстебельника толстокорневого похожи на семена одуванчика: они мелкие,
с толстой оболочкой и характерным «парашютиком», и так же разносятся ветром.
После плодоношения растение сохраняет листья недолго, и к концу лета у него
наступает период покоя: надземная часть отмирает, и растение проводит в покое
осень, зиму и начало весны, оставаясь зелёным менее полугода.
Этот вид развивается долго: растение впервые зацветает лишь в пятилетнем возрасте,
когда его «корнеплод» достигает веса 10 – 15 кг.
Бесстебельник легко расселяется семенами, поэтому он часто встречается в предгорьях
на островах-«оазисах» Средиземноморских болот. Но здесь его могут поедать крупные
местные травоядные – черепахи-антеи,
нанося большой ущерб медленно растущему растению.
Пазник
древовидный (Hypochaeris europaeus)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteraceae)
Место обитания: южная Европа, горные районы.
Мир флоры таит в себе не меньше неожиданностей, чем мир фауны. В эпоху человека
рядом с известными одуванчиками (Taraxacum) проживали и их близкие родственники
– пазники (Hypochaeris), которые экологически заменяли одуванчики в Южной Америке,
а также сосуществовали с ними в Евразии и Северной Африке, избегая при этом
серьёзной конкуренции. В неоцене эта ситуация несколько изменилась – пазники
по-прежнему доминируют в Южной Америке, одуванчики – в Северной, а в Европе
и Северной Африке некоторые виды пазников освоили древовидную жизненную форму,
в отличие от одуванчиков. Их типичный представитель – это пазник древовидный.
Это довольно крупное растение по меркам континентальных видов сложноцветных
– высотой до 3-4 метров. Этот вид растёт на склонах гор южной Европы и больших
одновидовых зарослей не образует – пазник древовидный предпочитает селиться
поодиночке или группами от 2 до 10 растений на расстоянии нескольких метров
друг от друга. Ствол растения относительно короткий, покрыт гладкой корой, шелушащейся
тонкими плёнками. Корневая система стержневая, с мощным главным корнем, уходящим
в трещины скал на глубину нескольких метров. Ветки образуют широкую раскидистую
крону и часто полегают на землю, укореняясь при этом. Листва напоминает листья
предковых видов, но с более широкой и изрезанной кромкой, чтобы производить
больше тени. Пазник древовидный довольно светолюбив и конкурентов вокруг своих
корней не терпит, подавляя их рост при помощи затенения. Листья одиночные, собраны
в розетки на концах ветвей. Зимой часть старой листвы опадает.
Цветы у пазника древовидного собраны в соцветия-корзинки жёлтого цвета, появляются
в центре листовых розеток на длинных цветоносах по 1-3 соцветия. Они опыляются
в основном местными видами пчёл, шмелей и похожих насекомых – в целом этот вид
не специализирован к конкретным видам опылителей. Семена с пушистыми хохолками-парашютиками,
заключены в обёртку из сухих присоцветных листочков. Они переносятся преимущественно
ветром, хотя иногда прицепляются к оперению разных мелких птиц, которые их охотно
поедают (обычно это разные виды вьюрков). Наружная оболочка семян переваривается,
а сами семена выходят вместе с испражнениями, и прорастают на новых местах значительно
легче по сравнению с теми, которые разносит ветер. В первые два года жизни молодые
растения очень уязвимы и часто гибнут от травоядных животных. В это время у
них развиваются колючие волокнистые ювенильные листья, которые сменяются взрослыми
при высоте растения около двух метров. На четвёртом-пятом году жизни растение
впервые зацветает. Отдельные особи пазника-дерева могут дожить до 100 лет и
больше, но многие не доживают даже до 40-50 лет.
Этот вид растений открыл Bhut, участник форума.
Ороматрика
серебряная (Oromatrica argenteofolia)
Порядок: Астроцветные (Asterales)
Семейство: Астровые, или Сложноцветные (Asteraceae)
Места обитания: Гималаи, высокогорные луга на южных склонах.
В неоцене Гималаи по-прежнему удерживают титул самых высоких гор на Земле.
Здесь, на огромной высоте, по-прежнему большую часть года царствуют холод и
снег, хотя ниже, в долинах и на южных склонах летом бывает очень жарко.
Растительность Гималаев разнообразна, хотя суровые условия жизни сделали многие
растения очень похожими друг на друга. Иногда, пока растения не зацветут, трудно
отличить мак от гвоздики или колокольчика – так похожи их листья и стебли.
Большинство травянистых растений Гималаев имеет ползучие или сильно укороченные
ветвистые стебли, и листья, образующие густые подушки между камней. Такая форма
роста защищает их от холодных зимних ветров: снег быстро закрывает эти низкорослые
растения, и они успешно перезимовывают, несмотря на зимние холода.
Но среди горной флоры есть не только низкорослые виды, жмущиеся к земле. Кое-где
на склонах, недоступных травоядным зверям, растёт необычное растение – ороматрика
(буквально «горная ромашка»). Она сильно отличается своим обликом от большинства
гималайских трав. У этого растения толстый прямостоячий ствол высотой около
полуметра, покрытый сухими остатками листьев. На вершине ствола растут живые
листья – они сильно разрезные, на нижней части имеют серебристое опушение.
Это главная защита растения от холодов: к зиме листья сворачиваются вверх,
закрывая точку роста и предохраняя её от мороза. Занесённое снегом, это растение
практически неотличимо от камней. Листья имеют неприятный запах и горький вкус,
защищающий их от разных травоядных. Но, несмотря на это, крупные травоядные
звери всё же объедают эти растения – зимой даже такой корм лучше, чем голодная
смерть.
Весной, когда снег оттаивает, начинается рост ороматрики. Живые листья на верхушке
растения раскрываются, и между ними появляются молодые листочки. Серебристые
волоски, покрывающие листья, защищают их от избыточного солнечного излучения
и отражают губительный ультрафиолет. В стеблях имеется запас питательных веществ,
поэтому растение может оправиться даже после того, как всю его верхнюю часть
съели звери или сорвала снежная лавина. Тогда на оставшейся части стебля прорастают
боковые побеги, и растение образует раскидистый куст. Молодые листья могут
переносить довольно сильные ночные заморозки – весна приходит в высокогорье
не сразу, и иногда зима может вернуться на несколько дней, и даже недель. Но
рано или поздно остатки снега тают, и наступает тёплая погода. Растение активно
образует новые побеги и восстанавливает корни. Корневая система ороматрики
в основном поверхностная и образует разветвлённую сеть, охватывающую крупные
камни – это помогает собирать росу, стекающую по ним утром. Обвалы и лавины
часто повреждают корни ороматрики, но они быстро восстанавливаются – медленный
рост в горах явно не способствует выживанию.
Когда окончательно устанавливается тёплая погода, и опасность ночных заморозков
уходит, ороматрика зацветает. Соцветия этого растения похожи на ромашки с короткими
белыми лепестками (точнее – краевыми язычковыми цветками в соцветиях). Они
возвышаются над растением на длинном разветвлённом цветоносе. Цветки приятно
пахнут, привлекая самых разнообразных насекомых – это растение нетребовательно
к опылителям. После опыления на соцветии созревают мелкие семена с пушистым
хохолком. Когда они полностью созреют, их хохолки вытягиваются и раскрываются.
Первый же порыв ветра разносит семена, но лишь немногие попадут в условия,
благоприятные для роста. Упав на землю, семена не прорастают сразу же: должна
пройти зима, прежде чем они пойдут в рост, а некоторые семена вообще прорастают
только через две зимы.
В течение первого лета жизни молодое растение формирует розетку листьев на
коротком стебле, а начинает цвести лишь со второго года. Продолжительность
жизни этого растения – около 13 – 16 лет.
Выше в горах произрастает близкий вид:
Ороматрика
капустовидная (Oromatrica pseudobrassica) – более низкорослое растение:
она образует «кочан» листьев на коротком стебле. В суровых условиях, когда
даже в разгар лета температура не поднимается выше +15°С, а по ночам возможны
заморозки, это растение ухитряется расти и цвести. Точка роста этого растения
постоянно закрыта пушистыми молодыми листьями, и заморозки ей не вредят. Цветоносы
этого растения короткие, а лепестки покрыты тонким слоем воска и отражают ультрафиолетовый
свет, привлекая редких в высокогорье насекомых.
Морская
безлистная астра (Aphyllaster littoralis)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteraceae)
Место обитания: литоральная зона от Северо-Западной Европы до Северной Африки,
Исландия, Новая Азора.
В неоцене береговая линия Северной Европы претерпела значительные изменения.
Когда тяжесть тысяч кубических километров ледников, сковывавших значительные
площади материка, исчезла, север Евразийской литосферной плиты начал подниматься.
Море отступало на значительных территориях, и бывшее морское дно превращалось
в равнины, на которых оставались солёные озёра, позже высыхавшие. Территория,
в эпоху человека занятая Северным морем, превратилась в обширные болотистые
равнины с отдельными возвышенностями, поросшими лесом. Балтийское море превратилось
в обширное солоноватое Венедское озеро, отрезанное от бассейна Атлантического
океана. Холодное течение Антигольфстрим определяет климат этих мест: прохладное
туманное лето, похожее на позднюю весну, плавно переходящую в раннюю осень.
Летом по утрам бывают густые туманы, оседающие на листьях прибрежных растений
обильной росой.
Побережье Северной Европы южнее Скандинавии представляет собой широкую полосу
солёных болот, где сильно ощущается действие приливов. Почва и вода здесь солоноваты,
и морские представители фауны встречаются в мелководных водоёмах на большом
расстоянии от океана. Цветковые растения активно осваивают эту область, образуя
множество эндемичных видов, приспособленных к существованию в условиях засоленности
почвы.
На небольшом расстоянии от океана в литоральной зоне встречается интересное
растение, выдерживающее дважды в сутки напор приливных волн – морская безлистная
астра. Это невысокое растение с толстыми ребристыми ветвями, разделёнными на
членики. Рёбра вытянуты в плоские широкие «крылья» с колючками по краю, и служат
для фотосинтеза. Настоящие листья растения редуцированы до плёнчатых обвёрток,
защищающих молодые побеги. Когда побег начинает развиваться, листья-обвёртки
быстро засыхают и опадают. Такое развитие листьев – необходимость: во время
прилива есть угроза повреждения растения, поэтому морская безлистная астра
утратила мягкие и нежные листья, превратившись в жёсткое растение, похожее
на коралл.
На молодом побеге рёбра небольшие – так меньше вероятность того, что прирост
будет поврежден приливной волной. По мере развития механических тканей они
разрастаются и упрочняются. Морская безлистная астра обитает в условиях избытка
соли, поэтому вдоль ребер стебля располагаются железы, выделяющие рассол в
виде густых капель. В отлив он засыхает на солнце в виде белых хлопьев на рёбрах,
и затем смывается приливом.
Для дополнительной защиты от солёной морской воды поверхность растения покрыта
тонким восковым налётом, особенно сильно развитым на молодых побегах. Из-за
этого они имеют ярко выраженный сизоватый оттенок. Смысл этого приспособления
состоит в том, что при отливе вода скатывается с растения и не образует на
нём корочку соли, затрудняющую фотосинтез.
Весной, когда становится достаточно тепло, морская безлистная астра зацветает.
Соцветия этого растения красивые – они белые с розоватой серединой, по форме
немного похожие на ромашку с «пушистой» серединой и широкими лепестками, диаметром
около 4 – 6 см. Язычковые цветки расположены в один ряд по краю соцветия. Серединные
цветки снабжены длинными розоватыми выростами околоцветника. Соцветия располагаются
на очень высоких прочных цветоносах, отрастающих выше уровня самого высокого
прилива. На одном растении может быть свыше 20 соцветий одновременно. Это растение
цветёт после прохладной зимовки: в период относительного покоя в пазухах прошлогоднего
прироста закладываются цветочные почки. Эта особенность ограничивает способность
морской безлистной астры к размножению семенами – растение нормально существует
только в условиях выраженных сезонных изменений климата. Это растение опыляется
разнообразными насекомыми – чаще всего жуками и пчёлами, но изредка на его
соцветиях кормятся бабочки. Цветение длится более двух месяцев. В это время
места произрастания морской безлистной астры выглядят очень красиво: цветы
вносят яркий акцент в тусклую зелёно-бурую палитру литорали.
Семена с пушистым хохолком созревают в конце лета. Когда они полностью развиваются,
ложе соцветия выгибается наружу, и они разносятся ветром. Благодаря этой особенности
растение широко расселилось по берегам Европы, захватив даже берега Исландии.
На Новой Азоре и африканском побережье это растение цветёт редко – только после
особенно прохладной зимы, когда закладываются цветочные почки.
Кроме семян, морская безлистная астра расселяется с помощью побегов. Приливные
волны часто отламывают ветви этого растения и уносят их в океан. Части растения
способны долгое время выживать, плавая в морской воде. Благодаря такой способности
это растение расселились ещё дальше на юг, но южные популяции этого вида не
цветут.
Бодяк
шарлаховый (Pachycirsium coccineus)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteraceae)
Место обитания: тихоокеанское побережье Северной Америки, влажные лиственные
леса.
Влажные леса умеренного климата, сформировавшиеся на тихоокеанском побережье
Северной Америки, представляют собой благоприятное место для жизни влаголюбивых
травянистых растений. Дожди и туманы с океана способствуют росту густого подлеска
в лесах из туи и влаголюбивых лиственных деревьев.
Местные растения приобретают причудливые формы, и в них бывает сложно узнать
скромно выглядевшие предковые формы эпохи человека.
Одно из характерных растений подлеска влажных тихоокеанских лесов – бодяк шарлаховый,
многолетнее корневищное травянистое растение с шипастыми стеблями и колючими
листьями. Этот вид произошёл от многолетнего бодяка полевого (Cirsium arvense)
– обычного и широко распространённого сорного растения. В условиях обилия влаги
растение достигает высоты 2-3 метров, а его листья вырастают до полуметра в
длину. Они перистонадрезанные, с крупными острыми колючками по краям и на верхней
стороне листа ближе к краю. Жилки очень толстые, рельефно выступают с нижней
стороны листа, вдоль средней жилки растут крупные колючки. Нижняя сторона листа
светло-зелёная. Стебли пятигранные, полые, однолетние; вдоль рёбер тянутся ряды
тонких острых колючек. После плодоношения стебли отмирают.
В начале вегетации на поверхности земли образуется розетка из небольших колючих
листьев. К концу весны отрастают крупные взрослые листья, и из середины розетки
вырастает цветущий побег.
У растения развивается массивная корневая система, накапливающая значительное
количество питательных веществ. Этот вид активно размножается ползучими корневищами,
которые достигают толщины до 2-3 см. У них плотная оболочка и рыхлая мякоть,
богатая крахмалом. На поверхности корневища формируются спящие почки, которые
при повреждении корневища трогаются в рост и формируют вертикальные побеги.
Вегетативное размножение позволяет растению формировать обширные клоны, тянущиеся
на десятки метров.
Это двудомное растение, мужские особи внешне неотличимы от женских. Цветочные
корзинки крупные – верхушечная достигает диаметра 6-8 см, пазушные – до 4 см.
В корзинках развивается множество узких трубчатых цветков ярко-красного цвета,
из-за чего корзинка приобретает полушаровидную форму. Этот вид цветёт практически
всё лето. Обильно выделяя нектар, предок этого вида привлёк нового опылителя
– колибри. Союз с этим опылителем определил внешний облик растения и окраску
его цветков, направив эволюцию растения в новую сторону. Красный цвет соцветий
привлекателен для птиц, однако на цветках имеются также участки, отражающие
ультрафиолетовый свет и делающие их заметными для резервных опылителей – дневных
и ночных бабочек.
Семена вызревают к концу лета. Они мелкие, летучие, снабжены хохолком из длинных
нитей, позволяющим держаться в воздухе. Семена быстро теряют всхожесть, поэтому
чаще всего этот вид размножается вегетативно, образуя крупные заросли-клоны.
Степная
ромашка (Leucanthemum americanum)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteraceae)
Место обитания: североамериканские прерии.
Человек разумный как вид просуществовал не так уж долго – всего лишь несколько
сотен тысячелетий. Но за этот относительно короткий срок он проявил себя как
сила, равная прочим силам природы: он уничтожил одни виды, искусственно создал
другие, и завёз третьи на другие материки, куда они ни за что не попали бы естественным
путём. Среди таких переселенцев был и нивяник обыкновенный (Leucanthemum vulgare),
или поповник, который, попав в Северную Америку, легко прижился там, пережив
людей и ледники без особых проблем, и породил за это время новые виды. Один
из этих видов – это степная ромашка, многолетнее травянистое растение американских
прерий.
Этот вид – многолетнее травянистое растение, образующее побеги двух типов. В
верхней части растение выглядит как хорошо узнаваемая ромашка около 10-20 см
в высоту, с большими соцветиями-корзинками с белыми краевыми цветками, а на
маловетвистых стеблях растут удлинённые листья – цельные, с неглубоко зубчатым
краем. В нижней части растения листья более широкие и округлые, чтобы лучше
улавливать солнечный свет. Нижняя поверхность листьев покрыта серебристым войлочным
опушением, на верхней стороне развиваются многочисленные полупрозрачные волоски,
слегка рассеивающие солнечный свет над поверхностью листа. Степная ромашка светолюбива,
избегает густых зарослей кустарников, растёт только на открытых пространствах
или в редколесье. Это растение вечнозелёное – осенью генеративные побеги отмирают,
и остаются лишь плотные прикорневые розетки некрупных листьев, которые дают
новые цветоносы на следующий год.
Размножается степная ромашка семенами и вегетативно. Её белые соцветия – одни
из самых обычных и привычных цветов североамериканских прерий. Её особенно любят
бабочки – и как источник нектара для них, и как пищевое растение их гусениц.
Травоядные позвоночные – от разных оленей
и азиногиппусов до пекари-бизонов,
тоже охотно поедают эти цветы, но степные ромашки от этого не страдают – обычно
прикорневая часть растения остаётся нетронутой. Это растение активно размножается
подземными корневищами – они у степной ромашки «ползучие», и способны давать
новые побеги, если старые будут съедены. Она образует новые кустики на расстоянии
до нескольких метров от родительского растения. Семена степной ромашки обладают
крохотными крючковидными щетинками, благодаря которым легко прилипают к мордам
и ногам разных травоядных зверей, которые потом разносят их по прериям, где
они и прорастают. В результате этого степная ромашка растёт чаще всего вдоль
кочевых путей разных травоядных зверей прерий. Молодое растение зацветает в
благоприятных условиях уже на следующий год, но чаще образует прикорневую розетку
листьев и начинает размножаться вегетативно, особенно на западном краю ареала,
в более засушливых условиях, где она зацветает лишь во влажные годы. В неблагоприятных
условиях растение размножается преимущественно вегетативно, давая крупные клоны,
распространяющие сеть побегов на сотни метров. Продолжительность существования
такого клона степной ромашки достигает 60-80 лет.
Этот вид растений открыл Bhut, участник форума.
Златобородник
степной (Aurobarba pratensis)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteridae)
Место обитания: Евразия, степи.
В эпоху человека разные виды растений существенно расширили ареал благодаря
деятельности человека. Многие из них расселялись целенаправленно как культурные
растения, в том числе декоративные. Культурные сорта многих декоративных растений
отличались низкой конкурентоспособностью и быстро исчезли, но некоторые виды
смогли одичать и вторгнуться в естественные экосистемы ещё в эпоху человека.
Одним из таких видов был североамериканский вид золотарник канадский (Solidago
canadensis). Пережив эпоху человека, этот вид продолжил эволюционировать в Старом
Свете, и его потомки стали частью экосистем неоцена.
В степях Евразии от юго-востока Европы до Центральной Азии встречается широко
распространённый потомок этого вида – златобородник степной. Подобно своему
предку, это многолетнее корневищное растение. Жизнь в степях определила внешний
облик растения – у него узкие очередные листья длиной до 30-35 см, и упругие
стебли высотой до 150-170 см. Верхняя сторона листьев тёмно-зелёная, гладкая;
изнанка листа покрыта тонким белым опушением. Растение засухоустойчиво, но очень
светолюбиво.
В середине лета на вершине стебля развивается кистевидное соцветие с длинными
поникающими ветвями, на которых расположены многочисленные мелкие корзинки из
4-8 жёлтых язычковых цветков. Массовое цветение златобородника длится не больше
двух недель. В это время развевающиеся на ветру жёлтые кисти соцветий придают
степям неповторимый облик. Растение хорошо выносит вытаптывание и поедание травоядными
животными, легко возобновляется от корневища.
К концу лета созревают многочисленные семена с пушистыми хохолками, которые
разносятся ветром на большие расстояния. Первое цветение наступает в возрасте
2 лет.
Златоголовник
узколистный (Chrysocephalus angustifolius)
Порядок: Астроцветные (Asterales)
Семейство: Астровые (Asteridae)
Место обитания: Евразия, лесная зона, луга.
Эпоха человека оставила свой след в фауне и флоре неоцена в виде многочисленных
потомков завезённых человеком видов, чужеродных для местности, в которой они
обитают. Предками некоторых из этих чужеродных видов были декоративные растения,
культивировавшиеся человеком. В лесной зоне Евразии одним из таких видов является
златоголовник узколистный, потомок завезённого в историческую эпоху североамериканского
вида – золотарника канадского (Solidago canadensis).
Подобно своему предку, этот вид является многолетним травянистым растением с
высокими неветвистыми однолетними стеблями, отрастающими от подземного многолетнего
корневища. Стебли достигают высоты 170-190 см, сочные, покрыты редкими короткими
волосками в основании. Вегетация начинается вскоре после таяния снега. Молодые
побеги быстро отрастают весной после установления тёплой погоды. Листья узкие,
длиной до 20 см при ширине около 15 мм. Вегетация продолжается до заморозков.
В конце лета златоголовник зацветает. Его соцветия – миниатюрные корзинки, содержащие
по 2-4 цветка. Они собраны в крупное верхушечное соцветие – кисть длиной до
20 см с мутовчатым расположением боковых ответвлений, густо покрытых корзинками.
Соцветие имеет облик пушистой ёлочки жёлтого цвета, цветение продолжается около
двух недель. В начале осени созревают мелкие семена, снабжённые хохолками, которые
разносятся ветром. После созревания семян стебель быстро усыхает и отмирает.
Семена прорастают ранней весной. Проростки устойчивы к заморозкам и быстро растут.
Молодое растение зацветает на втором году жизни, продолжительность жизни составляет
около 10 лет.
Цветковые
растения - однодольные
|
Тиарнеопсис
плодовитый (Tiarneopsis proliferus)
Порядок: Частухоцветные (Alismatales)
Семейство: Водокрасовые (Hydrocharitaceae)
Место обитания: юг Северной Америки, Центральная Америка, острова Карибского
моря.
После образования Панамского перешейка пресноводные растения Южной и Северной
Америки получили возможность расселяться на соседний континент – главным образом,
пользуясь услугами водоплавающих птиц, распространяющих их семена и отростки.
В дальнейшем изоляция Южной Америки привела к разделению ареалов этих видов.
Итогом независимой эволюции стало образование нового вида – тиарнеопсиса плодовитого,
потомка одного из видов рода Limnobium. Этот вид доминирует среди плавающих
растений тропических районов Нового Света к северу от Панамского пролива, и
даже вытеснил потомков пистии телорезовидной (Pistia stratiotes), завезённой
в Новый Свет человеком в историческую эпоху.
Успех в борьбе за существование у этого вида обусловлен способностью быстро
размножаться вегетативно. Взрослое растение образует розетку плавающих вздутых
листьев, заполненных губчатой паренхимой. Эти листья на коротких черешках легко
отламываются, если растение повреждает проплывающая птица. Новый лист вырастает
очень быстро: в течение 3-4 дней. Отломленный лист долго сохраняет жизнеспособность,
плавая по поверхности воды. Такое повреждение не приносит вреда растению, поскольку
оно размножается вегетативно при помощи листьев. На них в месте излома образуется
почка, из которой за счёт накопленных листом ресурсов быстро вырастает дочернее
растение.
Верхняя сторона листа гладкая, светло-зелёная, молодые листья розоватые. Нижняя
сторона листа серебристая, бугристая. Розетка взрослого растения диаметром до
10-15 см, содержит 4-6 листьев. Корни тонкие, короткие, свисают в толщу воды.
Значительную часть питательных веществ из воды растение всасывает через нижнюю
поверхность листьев. Ещё один способ вегетативного размножения – растение даёт
на поверхности воды усы с дочерними розетками. Цветки мелкие, растение двудомное.
Семена покрыты слизистой оболочкой, легко прилипают к ногам и оперению водяных
птиц и переносятся ими на большие расстояния, в том числе через морские проливы.
Сеянец впервые зацветает в возрасте 2-3 месяцев. Продолжительность жизни одной
розетки не превышает года, но вегетативное размножение продолжается круглый
год.
Пелагический
рдест (Potamogeton pelagicus)
Порядок: Частухоцветные (Alismatales)
Семейство: Рдестовые (Potamogetonaceae)
Место обитания: Четвероморье, верхние слои воды вдали от берегов.
Морские водоросли, которые произрастали в Чёрном море, вымерли задолго до того,
как высохло само море. Во время ледникового периода на рубеже голоцена и неоцена
огромные массы воды были связаны в ледниках, и сток воды в Чёрное море сильно
уменьшился. Постепенно солёность его вод возросла до океанической, а затем превысила
её и Чёрное море превратилось в несколько ультрагалинных водоёмов с отравленной
сероводородом водой. Такие условия были совершенно непригодными для жизни водорослей.
В неоцене на территории Евразии появилось Четвероморье – крупный внутриконтинентальный
водоём с солоноватой водой. Его флора представлена исключительно потомками пресноводных
и болотных растений, которые нашли здесь благоприятные условия для жизни. Большинство
растений Четвероморья теснится на узкой полосе прибрежных мелководий, и между
ними идёт суровая конкуренция за местообитания и солнечный свет. Зато вдали
от берега в открытой воде ничто не затеняет солнечные лучи. Здесь обильно развиваются
микроскопические водоросли, а из высших водных растений встречается лишь один
быстрорастущий вид – пелагический рдест.
Большинство рдестов представляет собой травы, укореняющиеся на дне. Пелагический
рдест освободился от связи с дном, и это позволило ему выйти в новые местообитания,
не занятые конкурентами. Он образует обширные плавающие заросли на поверхности
воды, тянущиеся на десятки метров и представляющие собой настоящие плоты из
переплетённых стеблей.
Пелагический рдест является длинностебельным растением. Длина его стебля достигает
4 – 5 метров, а ежедневный прирост составляет около 5 см у каждого стебля. Стебель
этого вида тонкий, красноватый, обильно ветвящийся в пазухах листьев и очень
прочный на разрыв. В узлах стебля развиваются длинные корни, свисающие вниз
и покрытые тонкими волосками.
Основными поплавками являются плавающие листья. Они овальные с притупленным
кончиком и длинным черешком, длиной около 10 см. Ткани листа богаты воздухоносной
паренхимой, а верхняя поверхность гладкая, серовато-зелёная, с тонким восковым
покровом. Благодаря этому покрытию листья не смачиваются водой и защищены от
избытка солнечных лучей. Листья очередные, и в их пазухах развиваются новые
побеги. Если во время шторма стебель оказывается разорванным, это не вредит
растению – у него быстро отрастают новые побеги. Заросли пелагического рдеста
бывают настолько густыми, что мелкие птицы могут ходить по ним, почти не проваливаясь.
Часто стаи птиц, занесённые ветром в море, спасаются именно на зарослях пелагического
рдеста, где могут отдохнуть и найти корм – мелких беспозвоночных, собирающихся
в зарослях.
Подводные листья этого вида мелкие, полупрозрачные, изумрудно-зелёные. Они развиваются
на самых молодых побегах и не обеспечивают полноценной плавучести. Лишь выпустив
первый плавающий лист, побег начинает устойчиво держаться на воде.
Цветки этого вида мелкие, невзрачные, четырёхлепестные, собранные в колосовидное
соцветие. Соцветия развиваются в пазухах листьев и поднимаются из-под воды вертикально.
Семенное размножение редко бывает успешным, поскольку насекомые лишь случайно
оказываются в местах обитания этого вида. Ветроопыление неэффективно, поскольку
крупная заросль этого растения может представлять собой клон, и фактически имеет
место самоопыление. Но этот недостаток вполне компенсируется активным вегетативным
размножением.
Осенью плавающие стебли и листья быстро отмирают и разлагаются. В это время
в пазухах листьев развиваются укороченные побеги с чешуевидными листьями – зимующие
почки. Они плавают в верхних слоях воды благодаря наличию полостей в сердцевине,
богатой крахмалом. Зимующие почки легко переносят промораживание и часто зимуют
в толще льда, покрывающего прибрежные акватории. При повышении температуры воды
они быстро разрастаются в новые побеги.
Дендронаяс
обыкновенный, морское дерево (Dendronajas vulgaris)
Порядок: Наядовые (Najadales)
Семейство: Дендронаясовые (Dendronajadaceae)
Место обитания: Атлантический океан, прибрежные воды и подводные отмели до глубины
20 метров.

Рисунок Nem’а, колоризация Биолога
Исходное изображение. Рисунок Nem’а |
В неоцене некоторые цветковые растения воспользовались последствиями
глобального экологического кризиса и сделали успешную попытку потеснить водоросли
в растительных сообществах морских мелководий. В раннем неоцене от семейства
цимодоцеевых (Cymodoceaceae) произошло обширное семейство дендронаясовых, бурно
эволюционировавшее на протяжении последующих 15 миллионов лет. Через 25 миллионов
лет после эпохи человека оно насчитывает 10 родов и более 100 видов, распространённых
в субтропической, тропической и бореальной зонах всех океанов.
Облик растений семейства дендронаясовых очень необычен. У них развиваются вертикально
стоящие корневища («стволы») – мощные, деревянистые, диаметром до 70 см и высотой
до 5-7 м, способные к вторичному росту за счёт камбиеподобной ткани. Внутри
стволов имеются сосуды, стенки старых сосудов в сердцевине ствола могут накапливать
известь. Размножение этих растений в основном вегетативное, но в благоприятных
условиях семена успешно завязываются.
Это двудомные растения; очень сильно редуцированные женские цветки развиваются
внутри спирально завёрнутого по краям покрывала (часто длинного и узкого), столбик
и рыльце длинные (иногда ветвистые), нитевидные. Мужские цветы развиваются на
особых коротких побегах, которые после цветения отмирают. В процессе созревания
прицветники вокруг семян становятся мясистыми, наружное покрывало раскрывается,
обнажая два ряда плодов, которые активно поедаются некоторыми рыбами. Семена
с плотной оболочкой, поэтому не перевариваются и с фекальными массами падают
на дно, где и прорастают. У некоторых родов оболочка плодов яркая, содержит
воздушные клетки; такие плоды всплывают на поверхность, где склевываются морскими
птицами.
Широко распространённый на атлантическом побережье Старого Света вид дендронаяс
обыкновенный, или морское дерево, образует в прибрежных морских водах обширные
заросли. Высота «стволов» достигает 3 – 4 метров, в субтропических водах до
5 метров. Старые части «стволов» видов этого рода покрыты специфическими корковыми
известковыми водорослями из рода Lithophaneroderma, которые в процессе роста
образуют толстую кору до 3-4 см толщиной. В ходе вторичного роста ствола эта
кора трескается, но водоросли в процессе роста закрывают образующиеся трещины.
Дендронаяс обыкновенный встречается строго на сублиторали, поскольку точки роста
этого растения не выносят обсыхания на воздухе. Если вершины обсохли, рост на
какое-то время приостанавливается, а под корой закладываются новые точки роста.
Новые растения отрастают также от основания сломанных «стволов». Кусок «ствола»,
перекатываемый морскими волнами, легко укореняется на дне в тихом месте и даёт
вертикальные побеги.
Листья морского дерева простые, лентовидные, тёмно-зелёного цвета, длиной до
3 метров и шириной около 3-4 см. Они держатся около 2 лет, постоянно нарастая
у основания. Концы листьев притупленные, но обычно вершина листа повреждается
морскими животными и крупные целые листья почти не встречаются.
Этот вид растений цветёт редко. На самой северной оконечности ареала дендронаяс
обыкновенный зацветает лишь в самые тёплые годы, хотя может расти даже в холодной
воде Антигольфстрима. В южной части ареала цветение этого вида происходит почти
ежегодно. Мужские растения отличаются от женских меньшей высотой «ствола» и
более короткими листьями. Они предпочитают расти на мелководных участках дна
и активно размножаются вегетативно, благодаря чему противостоят появлению в
зарослях женских растений этого вида. Мужские цветки с тремя крупными пыльниками,
покрыты кожистой чашечкой. Они образуют большое количество мелкой пыльцы, которая
вымывается водой по мере созревания. Женские соцветия развиваются в центре листовых
розеток. Длинное соцветие содержит до 10 пар мелких цветков с длинными рыльцами,
завёрнутых в края покрывала. Пестики высовываются наружу в виде длинных кисточек,
на которых оседает пыльца. Плоды мясистые, ягодообразные. В каждом плоде содержится
до 20 мелких семян, прорастающих примерно через две недели после прохождения
через кишечник рыбы.
Дендронаясовые рифовые леса стали наиболее высокопродуктивными прибрежными морскими
биоценозами неоцена. Благодаря многоярусной структуре этих зарослей в них находят
укрытие разнообразные рыбы и беспозвоночные.
Этот вид растений открыл Nem, участник форума.
Посейдонов
лук (Thalassoallium bulbosum)
Порядок: Наядовые (Najadales)
Семейство: Дендронаясовые (Dendronajadaceae)
Место обитания: юг Атлантического и Индийского океанов, прибрежные воды и подводные
отмели до глубины 10 метров.

Рисунок Nem’а, колоризация Биолога
Исходное изображение. Рисунок Nem’а |
Представители семейства дендронаясовых – это особое явление
в растительном мире: многолетние одревесневающие растения, способные расти в
воде океанической солёности. Большинство видов этих растений обладает высоким
стволом и населяет глубины до 20 метров, зачастую дотягиваясь листьями до поверхности
воды. Но представители одного рода этих растений эволюционировали в другом направлении,
освоив бурные прибрежные мелководья. Это представители рода Thalassoallium,
своеобразные «луковичные растения» морей южного полушария.
Эти растения отличаются очень укороченным и утолщённым «стволом» без ветвей.
Благодаря массивному стволу эти растения с лёгкостью выдерживают удары прибоя,
а длинные корни удерживают их на месте даже во время штормов.
Типичным представителем рода является посейдонов лук. Ствол этого растения достигает
высоты полуметра при почти таком же диаметре. У молодых растений ствол более
вытянут в высоту, но с возрастом он сильнее утолщается. Поверхность ствола покрыта
плотной корой, в верхней части сохраняющей рубцы от опавших листьев. На вершине
ствола растут пучком узкие линейные листья с притупленной верхушкой. Они короткие
– не более метра в длину даже у крупных экземпляров; кончик листа со временем
разрушается, но он долгое время нарастает от основания. Сердцевина ствола богата
крахмалом. У молодых растений ствол более губчатый, в сердцевине накапливаются
газы (обычно углекислый газ и кислород). Благодаря этой особенности молодые
растения, вырванные с корнем во время шторма, могут дрейфовать по волнам и таким
образом расселяются.
В основании ствола взрослого растения образуются почки. При повреждении точки
роста из них формируются новые вертикальные побеги, а в обычных условиях – ползучие
горизонтальные корневища. На некотором расстоянии от взрослого растения образуются
дочерние растения. У рода Thalassoallium вегетативное размножение преобладает
над семенным: иногда популяция, представленная тысячами растений, оказывается
клоном одного растения, разросшимся на протяжении нескольких веков.
Посейдонов лук относится к двудомным растениям; мужские и женские экземпляры
внешне сходны, и в равных условиях мужское растение лишь немного мельче женского,
но в большой популяции эта разница мало заметна. Семенное размножение происходит
достаточно редко: обычно популяция растений синхронно зацветает раз в несколько
лет. На женских растениях отрастают длинные цветоносы, несущие до 10 пар мелких
цветков, закрытых свёрнутым в трубку покрывалом. Из верхней части покрывала
высовываются длинные тонкие рыльца, разделённые на кончиках на множество нитей.
Благодаря прочному и гибкому цветоносу соцветие плавает на поверхности воды
и рыльца оказываются на плёнке поверхностного натяжения. Мужские растения образуют
кистевидные соцветия из множества невзрачных цветков без лепестков, производящих
пыльцу с гидрофобным слоем на поверхности. Пыльца всплывает на поверхность воды
в виде пятен желтоватого цвета и происходит опыление. Главное условие успешного
цветения – спокойная поверхность моря. После опыления цветонос скручивается
в спираль и семена созревают под водой.
В водах Южной Африки произрастает другой вид рода – пирамидальный
морской лук (Thalassoallium pyramidalis). Этот вид лучше приспособлен
к жизни в условиях штормовой погоды. Его стебель сильно утолщается, а старые
корни постепенно скручиваются, погружая его в толщу грунта. Поэтому видимая
на поверхности грунта часть стебля имеет коническую форму. Этот вид также размножается
преимущественно вегетативно, но способен цвести и опыляться даже в условиях
частых штормов. Его соцветия не достигают поверхности воды, а пыльца плавает
в толще воды.
Этот вид растений открыл Nem, участник форума.
Морской
баньян (Thallasoficus ramosus)
Порядок: Наядовые (Najadales)
Семейство: Дендронаясовые (Dendronajadaceae)
Место обитания: тропические воды Индийского океана, глубина до 10 метров.

Рисунок Nem’а
Исходное изображение. Рисунок Nem’а |
В неоцене цветковые растения осваивают океанские мелководья
в новом качестве – как многолетние одревесневающие растения, часто крупных размеров.
Благодаря крупным размерам и способности к вегетативному размножению они успешно
конкурируют с водорослями, а их заросли представляют собой основу для продуктивных
экосистем морских мелководий. Эти растения обладают разными формами роста, что
позволяет им успешно существовать в местах с различными условиями жизни: от
спокойных бухт до побережий, подвергающихся ударам волн во время шторма.
В тропических районах Индийского океана на побережьях Африки и Азии произрастает
представитель семейства дендронаясовых, отличающийся своеобразной формой роста:
«ствол» состоит из целой сети обильно ветвящихся и переплетённых корневищ, часто
срастающихся друг с другом в местах контакта. Заросли этого вида напоминают
по форме баньян – тропические виды фикусов, и само растение называется морской
баньян. Корневище растения начинает рост в вертикальном положении, но в дальнейшем
полегает, укореняется и начинает ветвиться. Побеги расползаются по участку дна,
а на них развивается множество вертикальных побегов, ветвящихся, переплетающихся
и срастающихся друг с другом. В итоге образуется трёхмерная ячеистая структура
высотой до 2 метров, состоящая из одревесневающих и постепенно утолщающихся
корневищ, покрытых серовато-зелёной эластичной корой. Среди стеблей этого растения
охотно поселяются рыбы и ракообразные. Диаметр зарослей может достигать 10 метров
и больше. Подземные корневища у этого вида не образуются.
Благодаря способности образовывать упругие и прочные заросли морской баньян
часто селится по окраинам рифа, где мелководье обрывается и сменяется склоном,
уходящим в глубину. В таких местах морской баньян образует «козырьки» длиной
до 3 метров, которые нависают над глубоководным участком. Во время штормов они
часто надламываются, но сохраняют жизнеспособность, а растение наращивает новый
слой корневищ сверху. Срывающиеся со склона заросли являются важным источником
пищи для животных, населяющих склоны подводных гор.
Листья морского баньяна простые, удлинённые, с округлой вершиной. Они растут
на верхушках побегов и покрывают сплошным ковром переплетение корневищ растения.
При повреждении зарослей, когда свет проникает вглубь переплетения корневищ,
на них быстро развиваются спящие почки, из которых образуются новые побеги.
Сломанное корневище быстро укореняется и вскоре образует заросли.
Морской баньян обильно цветёт ежегодно. Женские цветки мелкие и невзрачные,
без лепестков, расположены внутри свёрнутого покрывала по 3-4 пары. Длинные
разветвлённые рыльца похожи на кисточки, высовывающиеся из покрывала. Соцветия
на коротких стеблях образуются в пазухах листьев. Семена этого растения легко
переносятся водой на большие расстояния. В отличие от многих родственных видов,
морской баньян способен давать семена без оплодотворения. Мужские особи этого
вида встречаются намного реже женских. Они имеют меньшие размеры и предпочитают
произрастать в местах с постоянным слабым течением. Мужские цветки мелкие, с
тремя кожистыми чашелистиками, расположены в пазухах листьев по три. Пыльца
вымывается водой и находится в толще воды, подолгу сохраняя жизнеспособность.
Семена мелкие, тяжелее воды. Благодаря мясистой питательной оболочке они охотно
поедаются рыбами, которые распространяют их на большие расстояния вдоль побережья.
Вблизи островов, отделённых от берега открытым океаном, этот вид представлен
главным образом клонами немногих особей и размножается исключительно вегетативно.
Этот вид растений открыл Nem, участник форума.
Пневмоспермиум
птичий (Pneumospermium ornithochorus)
Порядок: Наядовые (Najadales)
Семейство: Дендронаясовые (Dendronajadaceae)
Место обитания: Тихий океан, повсеместно в тропической и субтропической зоне.

Рисунок Nem’а
Исходное изображение. Рисунок Nem’а |
В ходе бурной эволюции в раннем неоцене представители семейства
дендронаясовых выработали разнообразные стратегии выживания в морской среде
обитания. Обычно они были связаны с изменениями в росте и помогали противостоять
волнам и течениям. Но у большинства дендронаясовых семена распространялись исключительно
рыбами – они были заключены в питательную и привлекательную для них оболочку.
Род Pneumospermium из семейства дендронаясовых в процессе эволюции выработал
иной способ распространения семян – с помощью морских птиц.
Заросли пневмоспермиума птичьего являются очень характерной деталью тропических
мелководий: они как бы продолжают под водой мангровые заросли, удерживая грунт
и подготавливая почву для наступления мангрового леса на море. Пневмоспермиум
птичий образует крупные заросли, тянущиеся на десятки метров. Они могут быть
представлены как генетически разнородными растениями, так и клонами одного растения.
Высота стеблей этого растения не превышает одного метра. В отличие от большинства
дендронаясовых, этот вид редко образует ползучие грунтовые побеги. Обычно клон
образуется, когда старый куст растения начинает отмирать, а нижние ветви ложатся
на грунт. «Ствол» этого растения имеет древовидную форму, неправильно дихотомически
ветвистый. Поверхность корневищ покрыта эластичной буроватой корой, желтовато-коричневой
на молодых частях корневищ. На концах корневищ развиваются пучки удлинённых
кожистых листьев с губчатой внутренностью, среди которых развиваются невзрачные,
но довольно крупные соцветия. Обычно пневмоспермиум дорастает до уровня самого
низкого отлива. Растение достаточно легко переносит обсыхание вершин, поскольку
верхушка побега и молодые листья покрыты плотным слоем желеобразного защитного
вещества.
Корневая система глубокая, позволяет растению выдерживать удары штормовых волн.
Если растение будет повреждено штормом, оно быстро восстанавливается, а отломленные
части корневища уносятся волнами и укореняются на новом месте. Листья благодаря
своей губчатой структуре служат прекрасным поплавком, не давая тяжёлому корневищу
утонуть, а запасы крахмала в корневище позволяют растению долго сохранять жизнеспособность
и переноситься течением на большие расстояния.
Мужские и женские растения в нецветущем состоянии почти не отличаются друг от
друга: у мужских растений лишь немного мельче листья. Мужские цветки без лепестков,
с тремя кожистыми чашелистиками, развиваются в пазухах листьев густыми гроздьями.
Они образуют большое количество мелкой плавающей пыльцы, образующей желтоватые
пятна на поверхности воды. Цветонос короткий, покрывало напоминает свёрнутый
вдоль лист растения, но на нём двойным рядом вздутий обозначаются женские цветы.
Рыльца высовываются попарно из продольной щели в свёрнутом покрывале. Опыление
происходит во время отлива, когда рыльца достигают поверхности воды. Плоды ягодовидные,
с толстой и яркой оболочкой, которая содержит воздушные клетки. При созревании
плодов покрывало разрушается и оторвавшиеся от плодоножек плоды всплывают на
поверхность, где их склёвывают морские птицы. В желудке и кишечнике птицы оболочка
семян истончается и после этого они прорастают намного быстрее. Благодаря морским
течениям и птицам вид широко распространился по островам Тихого океана, не образуя
явно выраженных подвидов.
Этот вид растений открыл Nem, участник форума.
Огненная
лилия (Lilium igneum)
Порядок: Лилиецветные (Liliales)
Семейство: Лилейные (Liliaceae)
Место обитания: прерии Северной Америки.
Одна из основных проблем в выживании у растений из прерий Северной Америки в
неоцене – не зимний холод, а голодные животные. Многочисленные травоядные млекопитающие
этой природной зоны поедают растительный корм в большом количестве, и это обстоятельство
стало важным фактором, определяющим состав растительности прерий и их приспособления
для выживания. Многие растения этой экосистемы массово размножаются, а также
очень быстро растут; но если в случае некоторых видов деревьев и кустов это
способствует выживанию, то другие растения пошли несколько иным путём – например,
они стали ядовитыми, и травоядные млекопитающие их избегают.
Одно из таких ядовитых растений – огненная лилия североамериканских прерий.
Это высокое растение, до метра в высоту, с яркой зеленью, прочными стеблями
и длинными листьями. На верхушках стеблей раскрываются огненно-красные цветки
с шестью длинными лепестками с притупленной вершиной. Внутри цветка есть пятна
и полоски, покрытые волосками, отражающими ультрафиолетовый свет. Они являются
указателями пути к нектару для насекомых-опылителей. Семена образуются нечасто:
в среднем, лишь 1-2 коробочки на растение за весь вегетационный сезон. Цветение
продолжается с поздней весны до середины лета, и одиночные цветки успевают раскрыться
на молодых отпрысках в конце лета.
Окраска цветков делает огненную лилию довольно заметной в прериях, и это ей
нужно для выживания: зелень растения очень неприятна на вкус, а несколько съеденных
стеблей этого вида могут даже отравить такое большое животное, как пекари-бизон
или крупный олень. Многие
травоядные беспозвоночные также не едят это растение – есть лишь несколько видов
жуков, повреждающих зелень и подгрызающих луковицы этого вида.
Размножается огненная лилия в основном луковицами и подземными столонами, которые
образуются на нижней, подземной части стебля. Столоны расползаются в стороны,
образуя в разных местах новые луковицы, из которых прорастают новые растения,
зацветающие уже на следующий год. Большинство травоядных зверей избегает их
поедать, но пекари-бизоны иногда валяются на этих цветах, используя их ядовитый
сок для избавления от эктопаразитов.
Огненная лилия многолетнее растение, и отдельная особь может существовать до
20 лет и больше, при этом размножаясь вегетативно.
Этот вид растений открыл Bhut, участник форума.
Болотное
алоэ (Aloe multipoda)
Порядок: Спаржецветные (Asparagales)
Семейство: Асфоделовые (Asphodelaceae)
Место обитания: Земля Зиндж, саванны по берегам временных водоёмов.
Раскол Африки по линии Великой Рифтовой долины привёл к изоляции на Земле Зиндж
популяций восточноафриканских растений и животных, которые начали эволюционировать
независимо от материковых видов, порождая новые эндемичные формы. Среди видов
растений, оказавшихся на микроконтиненте, было несколько видов алоэ. Одним из
их потомков является болотное алоэ, обитающее в саваннах по берегам временных
водоёмов.
Болотное алоэ во взрослом состоянии представляет собой древовидное растение
высотой до 4-5 метров. Оно обильно ветвится и часто растёт несколькими стволами,
образуя раскидистую крону. Характерной особенностью этого растения является
способность образовывать ходульные корни в нижней части ствола, как у пандануса
– отсюда видовой эпитет «многоногое».
Стволы дерева покрыты серой корой с рубцами от старых листьев, на которых долго
сохраняются засохшие остатки проводящих пучков, образующие жёсткую щетину.
Листья толстые, языковидные, округлых очертаний с притупленной вершиной, длиной
до 35-40 см и шириной до 20-30 см. Они имеют тёмно-зелёный цвет, но покрыты
слоем воскового налёта, особенно молодые листья, приобретающие из-за этого почти
белую окраску. По краю листа растут плоские треугольные колючки красновато-бурого
цвета, верхняя поверхность листа покрыта небольшими бурыми колючками. Во влажный
сезон листья запасают воду и становятся значительно толще, чем в сухой сезон.
Это растение является суккулентом, однако устойчиво к затоплению и предпочитает
расти по берегам сезонных водоёмов, и во влажный сезон часто оказывается на
затопленной территории, на глубине до 1 метра. В течение влажного сезона растение
цветёт и аккумулирует воду. Цветение этого вида обильное, цветоносы появляются
в пазухах листьев прошлогоднего прироста. У растения крупные канделябровидные
соцветия светло-жёлтых трубчатых цветков длиной до 4-5 см с узкой трубкой. Растение
опыляется бабочками и крупными перепончатокрылыми, реже птицами. Плоды – округлые
вскрывающиеся коробочки с крупными семенами. Распространение семян происходит
при помощи грызунов, изредка при помощи воды.
Горный
рис (Oryza montana)
Порядок: Злакоцветные (Poales)
Семейство: Злаки (Poaceae)
Место обитания: Гималаи, тропические высокогорья.
Культурные виды и сорта риса происходят главным образом с Дальнего Востока и
из Африки. Этот род растений включает влаголюбивые виды, произрастающие на болотах
и в других влажных местах. В эпоху голоцена культурный рис был завезён во многие
страны тропического пояса и прижился на новых местах как культурное растение.
После исчезновения цивилизации ареал риса сильно сократился, большинство культурных
сортов вымерло, не оставив потомства, но кое-где на новых местах он прижился
– например, в южной части горного массива Гималаев. В процессе эволюции здесь
возник новый вид – горный рис, приспособленный к обитанию в местных водоёмах.
Горный рис не менее влаголюбив, чем его предки, но условия его произрастания
сильно отличаются от тёплых болот со стоячей водой, где жили его предки – это
бурные горные реки с сильным течением и более прохладной водой. Жизнь в таких
условиях значительно изменила растение. Взрослое растение этого вида больше
всего напоминает кустарник с многочисленными ветками, которые почти лишены листвы.
Ветви разных растений часто переплетаются друг с другом, закрепляя заросли этого
злака ещё больше. Эти побеги с прочными стеблями – генеративные: на них над
водой развиваются колосья, а сами побеги могут в определённой степени противостоять
течению воды. Листья у горного риса – тонкие ленты: над водой торчат только
цветки и колоски.
Растение образует ещё один вид побегов – вегетативные. Они отличаются более
слабым и гибким стеблем, прочным на разрыв, и длинными лентовидными листьями,
стелющимися по течению воды. Они легко образуют корни в узлах, а междоузлия
у них длинные, благодаря чему побеги достигают длины свыше 150 см. Они позволяют
растению захватывать новые территории, а на укоренившихся побегах вскоре начинается
рост генеративных побегов. Корни вегетативных побегов служат растению якорями,
они не дают ему быть унесённым течением, либо помогают ему закрепиться на новом
месте. С их помощью горный рис может размножаться вегетативно – его побеги,
оторванные от куста, могут пускать новые корни и прорастать на новом месте.
Корни обладают способностью прочно «прилипать» к шероховатой поверхности камней,
закрепляя растение на субстрате.
Невзрачные плёнчатые цветки горного риса собраны в рыхлые поникающие метёлки
и опыляются ветром. Его колоски довольно тёмные и невзрачные по сравнению с
его одомашненным предком. Семена растения долгое время остаются на материнском
растении – это наследие, доставшееся от культурного предка. Они довольно невзрачны
на вид и сохраняют плёнки, окружавшие цветок. Водные и околоводные птицы, и
даже некоторые рыбы охотно поедают их во множестве: эти зёрна богаты крахмалом
и питательны.
Горный рис приспособлен к жизни в текущей воде; в водоёмах с медленным течением
он проигрывает схватку за территорию другой растительности, поэтому его популяции
наиболее многочисленны в горных районах. Максимальная продолжительность жизни
растения горного риса – 6 лет. Обычно растение отмирает после массового цветения,
и значительная часть популяции бывает представлена проростками и молодыми растениями
в подводной фазе.
Этот вид растений открыл Bhut, участник форума.
Криобромелия
засыхающая (Cryobromelia arescens)
Порядок: Злакоцветные (Poales)
Семейство: Бромелиевые (Bromeliaceae)
Место обитания: Анды, высокогорье вблизи линии снегов.
В эпоху человека наибольшее разнообразие представителей семейства бромелиевых
приходилось на влажные тропические леса. Тем не менее, в различных экосистемах
Южной Америки встречались представители этого семейства, приспособленные к иным,
более суровым условиям жизни. Эта закономерность сохранилась также в неоцене:
некоторые представители семейства обитают в крайне суровых условиях, выработав
своеобразные приспособления для выживания.
На высокогорных лугах Анд встречается один из видов бромелиевых, произрастающий
практически до самой линии снегов – криобромелия засыхающая. Это растение приспособилось
к жизни в условиях физиологической сухости и повышенного уровня ультрафиолетового
излучения, а на юге ареала способно переживать зимние холода. Этот вид – розеточное
растение с толстым шарообразным стеблем и многочисленными узкими листьями, заострёнными
на кончиках в прочную одревесневшую колючку. Нижняя сторона листьев покрыта
опушением из серебристых волосков и многочисленными щетинкообразными колючками.
Верхняя сторона листьев гладкая, тёмно-зелёная, покрытая тонким слоем воска,
отражающим солнечные лучи. Зимой внешние листья у этого растения засыхают и
служат изолирующей обёрткой для розетки живых листьев, предохраняя их от холода
и иссушающего ветра. В это время растение похоже на шар или кактус – оно целиком
покрыто слоем волосков, среди которых торчат колючки, которые также спасают
от немногочисленных местных травоядных животных. Корневая система растения представлена
несколькими прочными волокнистыми корнями и служит исключительно для закрепления
растения на субстрате. Этот вид произрастает среди камней, на участках с истончённым
почвенным покровом, и даже просто в трещинах скал с минимальным количеством
почвы. Значительную часть питательных веществ этот вид получает из воздуха вместе
с пылью, которую удерживают его листья. Растение исключительно светолюбиво и
не растёт в местах, где появляются кустарники.
Весной растение переходит в фазу активного роста. С повышением влажности почвы
и воздуха обёртка из сухих листьев раскрывается и начинается активный рост новой
листвы, ведущей активный фотосинтез. Листья растения в фазе активного роста
образуют сильно распростёртую розетку: высота взрослого растения достигает 20
см, а диаметр розетки – до 50 см.
Этот вид растений – монокарпический, цветёт в возрасте 3-5 лет, набрав достаточное
количество питательных веществ в стебле. К этому моменту у основания материнского
растения уже образуется 2-4 молодых дочерних растения. Соцветие этого вида –
высокая кисть из колосков с ярко-красными прицветниками. Цветки трёхлепестковые,
трубчатые, с налётом растительного воска по краям лепестков. Растение опыляется
высокогорными видами колибри и бабочками бражниками. Семена мелкие, снабжены
хохолком, способны распространяться ветром на большие расстояния.
Криптобромелия
восконосная (Cryptobromelia cerosa)
Порядок: Злакоцветные (Poales)
Семейство: Бромелиевые (Bromeliaceae)
Место обитания: южные районы Южной Америки, Огненная Земля, открытые местности
и горные склоны.
В эпоху человека бромелиевые были известны преимущественно как тропические растения,
любящие влажность, хотя было также известно достаточно большое количество видов,
успешно выживающих в сухих местообитаниях. Во время голоцен-неоценового оледенения
среди этих растений стали появляться холодостойкие виды, которые по мере таяния
ледников отступали в высокогорье или на крайний юг материка. В процессе эволюции
у них появились приспособления, позволяющие выживать в условиях холодного климата.
На крайнем юге Южной Америки произрастает один из таких видов холодостойких
бромелиевых – криптобромелия восконосная. Этот вид сохранил типичную для представителей
семейства розеточную форму роста. Розетка диаметром около полуметра образована
примерно десятком кожистых языковидных листьев тёмно-зелёного цвета, покрытых
по краю тонкой корочкой растительного воска. Нижняя сторона листьев также покрыта
воском, придающим растению сизоватый оттенок. Нижняя часть листовой пластинки
толстая и мясистая, сохраняется живой до 2 лет и служит для запасания питательных
веществ. Верхняя часть листовой пластинки к зиме отмирает; мякоть листа в этой
части разрушается, и лист превращается в «щётку» волокон. Кроме того, к зиме
часть молодых листьев поднимается вертикально и сворачивается, закрывая точку
роста. По краям и на нижней стороне листьев обильно выделяется воск, «запечатывающий»
листья и предохраняющий точку роста от замерзания. В таком виде растение зимует,
выживая под снегом.
Цветение криптобромелии восконосной происходит весной и является одним из самых
красивых среди растений крайнего юга Южной Америки. Когда снег тает, листья,
предохраняющие точку роста, раскрываются, и вскоре из середины розетки поднимается
цветонос высотой до метра. На его конце образуется кисть «колосьев», образованных
яркими красными прицветниками с белым крапом по краю. Каждый прицветник прикрывает
несколько мелких белых трубчатых цветков, каждый из которых раскрывается примерно
на 2-3 дня. Криптобромелия восконосная опыляется местным колибри тайином
(Tayin migratorius), реже бабочками. Белые крапинки на прицветниках образованы
волосками, покрытыми тонким слоем воска и отражающими ультрафиолетовые лучи
– это сигнал для насекомых-опылителей. Воск с листьев этого вида собирают некоторые
одиночные пчёлы, которые также опыляют растение. Цветение этого вида длится
до месяца; по окончании цветения прицветники постепенно теряют яркость и буреют.
После опыления созревают плоды, содержащие мелкие летучие семена с хохолком,
которые разносятся ветром.
Это растение – монокарпик, после цветения розетка отмирает. В её основании закладываются
почки, из которых развиваются молодые растения. Также несколько мелких молодых
розеток появляется на цветоносе – их разносят крупные животные в своей шерсти.
Молодое растение зацветает на третьем году жизни.
Эйхорния
ныряющая, болотный арбуз (Eichhornia submersa)
Порядок: Коммелиноцветные (Commelinales)
Семейство: Понтедериевые (Pontederiaceae)
Место обитания: Северная Америка, реки и озёра в центральной и южной части материка.
В эпоху человека водяной гиацинт (Eichhornia crassipes) стал одним из самых
вредных инвазивных видов. Он наносил огромный ущерб природе, вытесняя аборигенные
виды, разрушая экосистемы и меняя гидрологический режим водоёмов. В эпоху человека
его численность удавалось сдерживать, но окончательной победы над этим видом
добиться не удалось, и он закрепился на новых территориях. Ледниковый период
заставил его отступать в места с более тёплым климатом, но по окончании ледникового
периода потомки этого вида вновь стали расселяться на север. В ледниковый период
ареал предкового вида оказался разделён на несколько изолированных участков,
что в дальнейшем привело к формированию разных видов. Один из них – эйхорния
ныряющая, обитающая в бассейнах крупных рек, в условиях субтропического климата.
Это растение, в отличие от теплолюбивого предка, приспособилось к выживанию
в условиях прохладных зим, и даже может некоторое время выживать под тонким
слоем льда. В связи с этим у него развились определённые особенности жизненного
цикла.
В тёплое время года этот вид – плавающее розеточное растение, похожее на предковый
вид. Основное отличие от предка в облике – крупные поплавки, образовавшиеся
из листовых черешков. Они так плотно прижаты друг к другу, что образуют шарообразную
структуру диаметром до 20 см. Листовая пластинка отделена от черешка тонким
слоем клеток, способным опробковевать. Сами листовые пластинки имеют овальную
форму, кожистые и тёмно-зелёные. Летом растение активно размножается усами,
давая в общей сложности до 30-40 молодых растений за сезон. Активно размножающиеся
растения образуют на поверхности водоёма плотный покров, способный выдержать
тяжесть крупной птицы. Цветки бело-розовые с жёлтым «глазком» на верхнем лепестке,
напоминают цветки предкового вида. Растение редко даёт семена, активно размножаясь
вегетативно. Зелень поедают различные травоядные млекопитающие, насекомые и
рыбы.
Корни длинные, с многочисленными волосками, активно всасывают растворённые в
воде питательные вещества.
К зиме листовые пластинки отмирают, но черешки листьев остаются зимовать. В
самых нижних черешках в процессе вегетации в паренхиме откладываются зёрна крахмала,
поэтому к осени они значительно тяжелеют и розетки растений погружаются в воду.
Листовые пластинки остаются только на самых молодых листьях, образуя своеобразный
«хохолок». Постепенно вместо воздуха в черешках листьев накапливается вода и
растение «ныряет» под воду. Зимующее растение имеет шарообразную форму, за что
получило второе название. В это время их поедают водные млекопитающие. Отломленный
черешок листа способен образовывать на месте слома выводковую почку, которая
за счёт запасов крахмала развивается в полноценное молодое растение. К весне
запасы крахмала истощаются и отломленные черешки листьев постепенно всплывают
на поверхность, где развивающиеся дочерние растения начинают самостоятельную
жизнь. На севере ареала молодые растения начинают цвести со второго года жизни,
после зимовки; на юге цветение почти круглогодичное, с перерывом на несколько
недель на зимовку. Продолжительность жизни отдельного растения не превышает
3-4 лет.
Неокаланта
пузыреносная (Neocalanthe pustulifera)
Порядок: Орхидные (Orchidales)
Семейство: Орхидные (Orchidaceae)
Место обитания: Юго-Восточная Азия, леса различных типов с постоянным или сезонным
увлажнением.
Среди растений, опыляемых животными, орхидеи демонстрируют высочайшую степень
специализации к такому симбиозу. В эпоху человека существовало большое количество
видов орхидей, демонстрировавших разнообразные способы привлечения насекомых
и использования их для опыления. На рубеже голоцена и неоцена большое количество
специализированных видов вымерло из-за разрушения местообитаний и вымирания
опылителей. В неоцене появилось большое количество новых видов, вновь выработавших
узкоспециализированные способы опыления.
На ветвях деревьев в вечнозелёных и сезонных тропических лесах Юго-Восточной
Азии обитает один из видов орхидей, вступивший в процессе эволюции в союз с
нектароядными летучими мышами – неокаланта пузыреносная, потомок одного из видов
рода каланта (Calanthe), существовавшего в эпоху человека. Это типичное эпифитное
растение, стебель которого превратился в крупную луковицеобразную псевдобульбу.
Он растёт в развилках ветвей, среди стеблей многолетних лиан с одревесневающим
стеблем, или в «эпифитных корзинах», образуемых папоротниками. На вершине этого
стебля ежегодно развивается пара крупных (длиной до 50 см) кожистых листьев
с притупленной вершиной, отмирающих перед наступлением периода покоя. Корни
растения прочно врастают в кору дерева, на котором оно развивается. Также в
период роста развивается густая «борода» воздушных корней белого цвета, покрытых
губчатой тканью.
В течение 1 – 2 месяцев (в сезонных лесах – до 4 месяцев) растение пребывает
в относительном покое. В это время в основании стебля формируются цветочные
почки. В сезонных лесах период покоя совпадает с сухим сезоном. В это время
воздушные корни растения полностью или частично отмирают.
По окончании периода покоя у основания псевдобульбы растения начинает развиваться
толстый прочный цветонос, направленный вбок, от ствола дерева. На нём развиваются
2-3 крупных цветка, распускающихся один за другим. Верхний чашелистик и боковые
лепестки цветка окрашены в белый цвет, в зеве есть красные мазки, указывающие
опылителю путь к нектару. Цветок распространяет очень сильный аромат ванили.
Нижние чашелистики сильно вытянутые, кожистые. Их концы свиты в спираль и покачиваются
по бокам цветка. Нижний лепесток (губа) розовато-белый, имеет небольшой отгиб,
но по его краям развиваются густые и прочные нитевидные выросты, а сам он вытянут
в длинный шпорец, на конце которого есть пузыревидное расширение. Внутренние
стенки этого пузырька богаты нектароносными железами.
Летучая мышь, привлечённая запахом цветка, садится на него, вцепляясь задними
лапами в «бороду» цветка, и хватается большими пальцами крыльев за крепкие хрящеватые
чашелистики. Затем летучая мышь просовывает в шпорец длинный язык и слизывает
нектар со стенок расширения. В этот момент зверёк касается головой середины
цветка. Поллинии приклеиваются на лоб летучей мыши, и она переносит их на следующий
цветок.
Плод – небольшая коробочка, заполненная микроскопическими семенами. После отцветания
у растения начинается рост новых листьев и период активной вегетации. В это
время у крупных старых растений образуются также боковые отростки-«детки».
Развитие молодого растения происходит достаточно медленно. В течение первого
года жизни проросток кормится исключительно за счёт гриба, с которым вступает
в симбиотические отношения, и лишь на второй год жизни развивает первый лист
длиной около 2-3 см. В шестилетнем возрасте растение может дать первый цветок.
Мирмиорхис
защищённый (Myrmiorchis tutus)
Порядок: Орхидные (Orchidales)
Семейство: Орхидные (Orchidaceae)
Место обитания: Южная Америка, дождевой лес. Эпифитное растение.
В эпоху человека было известно очень много видов растений, которые вступали
в симбиоз с муравьями. Растения образовывали в своём теле специальные полости,
в которых селились муравьи, и взамен не только пользовались выделениями и отходами
жизнедеятельности муравьёв, черпая из них питательные вещества, но и получали
надёжную защиту. Среди «зелёных друзей» муравьёв были акации, некоторые другие
деревья, а также многочисленные эпифиты. Среди многочисленных видов орхидей
растения родов Schomburgkia, Coryanthes и Gongora имели полые стебли, служившие
убежищем муравьям. В неоцене независимо от них подобные отношения с этими воинственными
насекомыми образовала ещё одна орхидея – мирмиорхис, эпифитное растение из
дождевых лесов Южной Америки.
Это растение имеет побеги двух типов. Плотные овальные листья орхидеи растут
на коротких вертикальных побегах. Основную массу растения составляет толстое
ползучее корневище, которое разрастается вдоль ветви, на которой поселилась
орхидея. Корневище особенно необычно тем, что на нём развиваются зоны вторичного
роста. Это явление крайне редко встречается не только у орхидей, но и вообще
у однодольных растений, к которым принадлежат орхидеи.
Молодая верхушечная часть корневища ровная. Она слегка приподнимается над субстратом
и покрыта листьями. Когда листья опадают, из почек начинают развиваться вторичные
побеги. Старая часть корневища начинает видоизменяться: появление зон вторичного
роста носит случайный характер, и корневище приобретает неправильную форму.
Оно становится шишковатым, в нём образуются отверстия и полости, а также дополнительные
точки роста. Старое корневище достигает длины около метра и полуметровой ширины.
В полостях корневища мирмиорхиса водятся муравьи различных видов, защищающие
орхидею от вредителей. Кроме того, помёт и остатки добычи муравьёв служат источником
органических веществ, нужных для роста этого эпифитного растения.
Но союз орхидеи и муравьёв имеет обратную сторону. Муравьи прекрасно защищают
орхидею от разных растительноядных насекомых, повреждающих листья и корневище
растения. Но они не менее успешно могли бы атаковать насекомых-опылителей,
жизненно необходимых для орхидеи. Поэтому мирмиорхис выработал приспособление,
позволяющее насекомым-опылителям посещать его цветки, не рискуя попасть в челюсти
муравьям. Растение просто препятствует доступу муравьёв на цветоносы. На середине
каждого цветоноса, ниже самого нижнего цветка, есть своеобразная «манжета»
из длинных жёстких щетинок разной длины, протяжённостью несколько сантиметров.
Муравей может преодолеть её лишь с очень большим трудом, и эти насекомые редко
появляются на цветках мирмиорхиса.
Цветы мирмиорхиса приспособлены для опыления бабочками. Соцветие этого растения
представляет собой рыхлую кисть из 15 – 20 цветков на высоком (около 60 см)
вертикальном цветоносе. Цветок мирмиорхиса небольшой, белый, с округлыми лепестками.
У него маленькая губа, переходящая в длинный шпорец, загнутый вниз и вперёд.
В губе есть несколько тонких полосок из пурпурных волосков, указывающих насекомому-опылителю
путь к нектару. Несколько точек такого же цвета может быть в основании лепестков.
Мирмиорхис развивается достаточно быстро – растение зацветает уже на пятом
году жизни. Эта орхидея отличается быстрым ростом, образуя к моменту цветения
лет куст весом до трёх килограммов.
Башмачок
бромелиелистный (Coronopedium bromeliaephyllum)
Порядок: Орхидные (Orchidales)
Семейство: Орхидеи (Orchidaceae)
Места обитания: острова Индийского океана.
Семейство орхидей – самое большое и богатое видами среди всех цветковых растений.
Его представители – травянистые растения, освоившие самые разнообразные места
обитания. Среди них больше всего эпифитов, но есть наземные виды, а также известны
орхидеи, живущие в болотах, и даже водные растения. В Австралии в эпоху человека
было известно несколько видов орхидей, постоянно растущих, и даже цветущих
под землёй. Одной из особенностей, обеспечивших успех орхидей в борьбе за существование,
является их симбиоз с грибами. Некоторые орхидеи довели его до крайности: они
утратили хлорофилл, и живут только за счёт органических веществ почвы, которые
усваивают с помощью грибов.
Семена орхидей очень мелкие, похожи на пыль. Они прорастают и развиваются лишь
тогда, когда в них прорастёт грибница определённого вида грибов.
Значительная часть орхидей обитает в тропиках, но в умеренном климате они также
довольно разнообразны. Эти растения можно встретить как во влажных тропических
лесах, так и в сравнительно сухих саваннах. В эпоху неоцена некоторые представители
орхидей благодаря мелким семенам, разносимым ветрами, попали из Индонезии на
острова Индийского океана, где заняли свою экологическую нишу в лесах.
Орхидея, обитающая на этих островах, весьма своеобразна даже тогда, когда не
цветёт. Это эпифит: как ни странно, на деревьях легче добыть нужную для жизни
воду, чем на земле: грунтовые воды на островах солоноватые из-за близости океана.
Поэтому единственный надёжный источник пресной воды на островах – дождь. Но
дождевую воду нужно собрать и сохранить, и орхидея с островов это с успехом
делает. У неё короткий стебель, а выгнутые листья формируют плотную розетку.
Во время дождей вода накапливается в розетке, образуя особые мини-водоёмы.
Такая же особенность характерна для южноамериканских растений семейства бромелиевых
(Bromeliaceae), поэтому из-за этой аналогии орхидея получила название башмачок
бромелиелистный. Сходство с бромелиями усиливается ещё и тем, что листья этого
башмачка покрыты пятнами, сливающимися в поперечные полоски. Для бромелиевых
также характерен похожий полосатый рисунок. Диаметр такой розетки достигает
полуметра, а глубина – до 20 см. В одной розетке взрослого растения накапливается
около двух литров дождевой воды. В этой воде часто находят приют личинки комаров
и мошек, а также более крупные животные: улитки и наземные раки, которые даже
выводят потомство, сидя в этих растениях.
Чтобы экономить воду, у этой орхидеи развилось ещё другое приспособление: листья
покрыты тонким слоем воска. Это предотвращает, с одной стороны, избыточное
испарение воды, а с другой стороны – листья не смачиваются водой, и нет опасности
их загнивания. Благодаря восковому налёту это растение может нормально расти
в сухом воздухе.
Цветоносы башмачка бромелиелистного не защищены восковым налётом, но это им
не нужно: цветонос прорастает у основания старой розетки. Вначале из-под розетки
прорастает толстый столон – побег с редуцированными листьями. Он некоторое
время растёт вбок, а затем изгибается вверх и образует цветонос, несущий один
крупный цветок.
Цветок у башмачка бромелиелистного опыляется жуками. Для этих насекомых растение
готовит особое «угощение» в нижнем лепестке (у орхидей он называется «губа»),
выгнутом в форме мешка. Это ароматная сладковатая жидкость, образующаяся из
обильно стекающего в лепесток нектара, разбавленного дождевой водой. Пытаясь
достать её, насекомые падают в лепесток, после чего начинают искать выход.
Края губы подвёрнуты внутрь, образуя «козырёк» над лужицей нектара, и выйти
из лепестка можно, лишь проползая около рыльца и получив после этого комочек
липкой пыльцы. Насытившись и вдоволь набарахтавшись в нектаре, жук покидает
цветок и летит… к следующему, где всё повторяется. Растение привлекает насекомых
контрастной окраской цветка: по винно-красному фону лепестков разбросаны белые
пятна, отражающие ультрафиолетовый свет. Верхний лепесток («парус») целиком
красный, но его край окаймлён белой полосой. Губа красно-коричневая, но для
насекомых есть специальные «указатели» - края губы покрыты тонкими белыми штрихами.
Взрослое растение образует до 3 – 5 цветков на одной розетке. Цветок держится
на цветоносе довольно долго: около 2 месяцев, но далеко не всегда на его месте
завязывается плод – узкая коробочка с микроскопическими семенами.
После цветения на столоне в месте прорастания цветоноса формируется новая розетка.
Но часто столоны после цветения просто засыхают.
Растение активно размножается отпрысками, но из семян возобновляется крайне
медленно. Первый год проросток существует за счёт проросшей в него грибницы
и представляет собой крошечный комочек клеток. Лишь на второй год жизни проросток
даёт первый лист, а зацветает только в восьмилетнем возрасте, и даже позже.
Зато с помощью семян растение расселилось на самые дальние острова Индийского
океана.
Башмачок
дубовый (Cypripedium quirinus)
Порядок: Орхидные (Orchidales)
Семейство: Орхидные (Orchidaceae)
Место обитания: дубовые леса на востоке Северной Америки.
Орхидеи – своеобразное семейство цветковых растений; значительная часть их видов
растёт в тропиках, и такие виды, как правило, принадлежат к числу самых красочных
и/или необычных видов орхидей. В эпоху человека многие из их видов оказались
под угрозой исчезновения или вымерли полностью. Но есть и такие виды орхидей,
которые растут в области умеренного климата, и даже проникают в приполярные
широты. Орхидеи из областей умеренного климата – это наземные цветущие растения,
которые, тем не менее, обладают достаточно яркими и красивыми цветками. По размерам
эти цветы всё равно уступают цветам тропических форм, но не лишены определённого
изящества и обладают привлекательной окраской.
Одна их таких орхидей – башмачок дубовый, потомок орхидеи башмачка мелкоцветного
(Cypripedium parviflorum), сравнительно многочисленного вида эпохи голоцена.
Это довольно высокое травянистое растение – около 40 см в высоту, с длинными
остроконечными листьями, расположенными на стебле поочерёдно. Листья в основании
стебля вырастают на 15-20 см в длину, с многочисленными рельефными жилками,
выступающими с нижней стороны листа; к вершине стебля размер листьев уменьшается.
Стебель несёт по несколько цветков-башмачков (обычно 2-3) бледно-жёлтого цвета,
которые опыляются различными насекомыми. Боковые лепестки покрыты в основании
бурыми точками. Специализация к опылителям невысокая – растение опыляется многими
видами одиночных и общественных перепончатокрылых, а также мелкими жуками.
Завязываемость семян низкая – плод завязывается в среднем на одном цветке из
десяти. Плод растения – узкая коробочка с пылевидными семенами. После плодоношения
растение «отдыхает» в течение двух лет, не образуя цветков. Не завязавшие семян
растения цветут ежегодно.
Это растение многолетнее, но не терпит холода, поэтому его ареал ограничивается
восточными широколиственными лесами и горными дубравами. В последних эта орхидея
находит особенно благоприятные условия для жизни – она часто «делится» с дубами
симбиотическими грибами, получая через них некоторое количество питательных
веществ. Эта орхидея скорее симбионт или комменсал, и не вредит деревьям. Под
землёй растение образует ползучее корневище, которое раз в несколько лет образует
боковые отростки и со временем образует крупные куртины, состоящие из десятков
крупных стеблей.
Темп роста башмачка дубового очень медленный: в первые три года жизни растение
питается исключительно за счёт грибницы и остаётся под землёй. На четвёртом
году жизни растения появляется первый надземный лист, а в десятилетнем возрасте
растение впервые зацветает. Эту орхидею изредка поедают разные млекопитающие
и насекомые. Соседство с дубами приносит растению некоторую пользу: орхидея
накапливает в теле дубильные вещества их опавшей листвы и даже из корней живого
дерева посредством почвенных грибов, и это делает растение менее съедобным для
травоядных животных.
Башмачок дубовый может достигать возраста нескольких десятков лет, нередко доживая
до векового возраста. Вегетативно размножающиеся клоны материнского растения
могут жить до 200 лет и более.
Этот вид растений открыл Bhut, участник форума.
Рапануйская
кокосовая пальма (Cocos paschalis)
Порядок: Пальмоцветные (Arecales)
Семейство: Пальмовые (Arecaceae)
Место обитания: остров Пасхи, прибрежные районы.
Потомок кокосовой пальмы (Cocos nucifera), которая была завезена на остров Пасхи
в голоцене. С исчезновением людей эти растения смогли не только выжить, но и
оставить потомков в неоцене.
Рапануйская кокосовая пальма – крупная пальма, достигающая 35 метров в высоту,
с перистыми листьями длиной 4-6 м, доли которых вырастают до 60-90 сантиметров
в длину; старые листья отпадают без следа, оставляя ствол гладким. Эти растения
дают свои первые плоды в возрасте шесть-десять лет, а для достижения пика плодоношения
требуется 15-20 лет.
Пальма образует и женские, и мужские цветки в одном и том же соцветии; таким
образом, эта пальма является однодомным растением. Женский цветок значительно
крупнее мужского. Цветение происходит непрерывно; растение не специализировано
к опылителям. Как и у её предков, плод у рапануйской кокосовой пальмы – это
костянка, а не настоящий орех. Подобно другим плодам, у него есть три слоя:
экзокарпий, мезокарпий и эндокарпий. Экзокарпий – это глянцевая внешняя оболочка,
обычно жёлто-зелёного или жёлто-бурого цвета. Экзокарпий и мезокарпий вместе
образуют «кожуру» кокоса, тогда как эндокарпий образует твёрдую кокосовую «скорлупу».
Толщина эндокарпия – около 4 миллиметров; в нём есть три характерных поры для
прорастания на дистальном конце. Две поры – заросшие, а одна функциональная.
Внутренняя часть эндокарпия полая и выстлана тонкой семенной оболочкой бурого
цвета, толщиной около 0,2 мм. Эндокарпий изначально заполнен многоядерным жидким
эндоспермом. По мере продолжения развития клеточные слои эндосперма откладываются
вдоль стенок эндокарпия толщиной до 11 мм, начиная с дистального конца. В конечном
итоге они образуют съедобный твердый эндосперм, который со временем затвердевает.
Маленький цилиндрический эмбрион погружён в твёрдый эндосперм прямо под функциональной
порой в эндокарпии. В ходе прорастания эмбрион выталкивается наружу через функциональную
пору и образует гаусторий внутри центральной полости. Гаусторий поглощает твёрдый
эндосперм для питания проростка.
Плод удлинённой формы, треугольный в сечении. Он также меньше, чем у предковых
видов, и достигает 35 см в длину.
Этот вид растений открыл Жуан Витор Кутиньо, Бразилия.
Пальма
дендем (Elaeis dendem)
Порядок: Пальмоцветные (Arecales)
Семейство: Пальмовые (Arecaceae)
Место обитания: тропики Южной Америки, в речных долинах.
В эпоху голоцена человек с различными намерениями завёз в новые места обитания
множество видов животных и растений. Африканскую масличную пальму (Elaeis guineensis)
завезли во многие регионы в целях производства растительного масла. Вид, интродуцированный
в Южную Америку, выжил в неоцене и стал предком пальмы дендем. Слово «dendém»
использовалось как обиходное название предка этого вида в Бразилии. Данное растение
произрастает в тропических областях Южной Америки, обычно встречается вблизи
рек в центральных и северо-восточных районах материка совместно с приречным
миндалем (Neoterminalia riparia) и фиолетовым
трубчатым деревом (Tabebuia violacea).
Взрослая пальма растёт одним стволом и достигает 20 м в высоту. Листья перистые,
длиной до 5 метров. Молодая пальма наращивает около 30 листьев в год. Закончившие
активный рост пальмы возрастом старше 10 лет дают примерно 20 листьев в год.
Цветки собраны в густые соцветия; каждый отдельный цветок небольшой, с тремя
чашелистиками и тремя лепестками.
Для созревания плодов пальмы от опыления до полной спелости требуется 6 месяцев.
Они красновато-жёлтые, размером 2,5 см, растут большими гроздьями. Каждый плод
состоит из маслянистого мясистого наружного слоя (околоплодника) с одним семенем.
Этими плодами питаются такие животные, как попугаи и обезьяны. Некоторые обезьяны
пользуются инструментами, чтобы раскалывать семена.
Этот вид растений открыл Жуан Витор Кутиньо, Бразилия.
Кротовый
цветок (Talpanthus subterraneus)
Порядок: Аронниковые (Arales)
Семейство: Аронниковые (Araceae)
Место обитания: лесные районы Европы.
Лиственные леса в области умеренного и субтропического климата – это место,
где смена сезонов вызывает значительные различия в условиях обитания растений
и животных. Летом листья деревьев практически полностью перехватывают солнечный
свет, и подлесок очень тенистый – в нём могут расти только теневыносливые растения.
Осенью значительная часть листвы опадает, и в течение примерно полугода освещённость
подлеска становится значительно лучше. Но зимние холода препятствуют росту травянистых
растений, поэтому время активной вегетации светолюбивых трав приходится на несколько
весенних недель. Представители этой экологической группы растений называются
эфемерами (однолетники) и эфемероидами (многолетники).
Весной цветов в лесу немного, и после зимовки насекомые активно ищут их. Любой
цветок, появившийся в начале весны, обязательно будет посещён насекомыми. Но
цвести слишком рано растениям мешают заморозки, которые могут убить цветки.
Тем не менее, одно из растений европейских лесов приспособилось цвести рано
весной.
Когда сходит снег и верхний слой почвы оттаивает, под землёй просыпается толстое
шишковатое корневище. С прошлой осени на нём развились крупные цветочные почки,
которые дожидались весеннего потепления. Когда почва достаточно прогревается,
цветочные почки прорастают. Их оболочка, представляющая собой видоизменённый
лист, покрывало, пробивает слой почвы и прошлогодней листвы, и вырастает на
высоту около 30 сантиметров. Покрывало плотно свёрнуто в трубку, которая снаружи
имеет желтовато-белый цвет. На вершине покрывало раскрывается, и становится
видна его изнанка – мясо-красная с сероватыми пятнами. Пятна покрыты волосками,
отражающими ультрафиолетовый свет, поэтому насекомые, даже не различая красного
цвета, замечают изнанку покрывала, и безошибочно направляются к нему. Когда
покрывало раскрывается, в воздухе распространяется гнилостный запах, на который
охотно собираются жуки – основные опылители этого растения.
Наиболее интересной особенностью этого растения является то, что его цветы находятся
ниже уровня почвы – за это растение получило название кротовый цветок. Мелкие
и невзрачные цветки этого вида собраны в соцветие-початок, которое не имеет
цветоноса и развивается прямо на корневище. Покрывало образует трубку, по которой
насекомые проникают к подземному соцветию, а волоски, направленные вниз, удерживают
насекомых возле соцветия, пока женские цветки не утратят восприимчивость к пыльце.
После оплодотворения верхняя часть покрывала отмирает, а нижняя часть служит
защитой развивающегося соплодия. Соплодие кротового цветка, представляющее собой
сильно разросшийся початок с крахмалистой сладковатой мякотью, в которую погружены
семена, созревает осенью. Нижняя часть початка начинает активно расти, и соплодие
выносится на поверхность земли, где его поедают птицы и звери – распространители
семян кротового цветка.
В середине весны, когда завершается цветение, кротовый цветок выносит на поверхность
земли крупную почку, в которой заключены несколько свёрнутых вместе листьев.
Лист кротового цветка имеет округлые очертания; он крупный (диаметром около
40 – 50 см), глубоко рассечённый на три, пять или семь долей. У молодых растений,
которые ещё не цвели, листья цельные или тройчатые, а у взрослых, с хорошим
развитым корневищем, они достигают полного развития. Поверхность листа кротового
цветка гладкая, тёмно-зелёная со светлыми жилками. Листья отмирают в первой
половине лета. За время вегетации растение успевает накопить достаточно питательных
веществ на образование новых цветочных почек и на созревание соплодия.
Молодое растение кротового цветка зацветает впервые на третьем году жизни, а
в северных частях ареала – на четвёртый, и даже на пятый год.
На севере Европы и в Альпах обитает близкий вид растений – поздний
кротовый цветок (Talpanthus tardus). У этого вида цветение и плодоношение
разделены зимними холодами, а вегетация происходит в течение всего лета. Особенностью
этого вида является то, что поздний кротовый цветок зацветает в начале осени.
Зимой растение находится в состоянии покоя, и прошлогоднее соплодие появляется
из земли поздней весной. У этого вида крупные и лишь слегка надрезанные по краю
листья тёмно-зелёного цвета.
Морское
таро (Paracolocasia transpacifica)
Порядок: Аронниковые (Arales)
Семейство: Ароидные (Araceae)
Место обитания: тропические районы Тихого океана – острова, побережья Азии,
Меганезии и Нового Света.
Таро (несколько видов и много разновидностей рода Colocasia) было одним из древнейших
пищевых растений, выращиваемых человеком. Ареал этой культуры в историческую
эпоху включал тропики обоих полушарий – материки и многие океанские острова.
После исчезновения человека многие культурные растения бесследно исчезли, однако
в некоторых местах таро успешно одичало и показало значительную конкурентоспособность
в формирующихся экосистемах раннего неоцена. В тропических районах Земли встречаются
различные потомки культурного таро. Один из их видов широко распространён на
островах тропической зоны Тихого океана – это морское таро.
Данный вид – крупное травянистое растение с крупным подземным клубнем весом
до 4-5 кг. Взрослое растение образует розетку из 8-10 листьев на черешках длиной
до 2 метров. Пластинка листа сердцевидная с вытянутым кончиком, тёмно-зелёная.
Поверхность листа голая, кожистая. Черешки листьев зелёные с серовато-белыми
пятнами – крупными в основании и мелкими ближе к пластинке листа.
У данного вида растений выработался интересный способ расселения, позволяющий
ему захватывать новые места обитания. Клубни морского таро легко выдерживают
путешествие в солёной воде благодаря плотной оболочке. Сердцевина старого клубня
полая, у мелких клубней заполнена рыхлой крахмалистой мякотью. Растения из прибрежных
популяций часто оказываются в море после сильных штормов. У плавающего клубня
быстро отмирают листья, а оболочка становится плотнее. Благодаря течениям и
штормам такие клубни способны преодолевать сотни километров по морю, благодаря
чему расселяются. Скорость роста у таких клубней довольно высокая. Попав на
берег, они прорастают за несколько часов, и уже через неделю обладают развитой
корневой системой и розеткой из 2-4 некрупных листьев. Для роста морское таро
предпочитает песчаные побережья при отсутствии затенения другими растениями,
поэтому данный вид часто оказывается в числе пионерных растений. Закрепившись
на берегу, растение активно размножается столонами и быстро образует заросли
на побережье. Часто клубни попадают с течениями далеко на север и юг, в умеренные
широты, где успешно развиваются летом, но обычно гибнут к зиме.
Этот вид – насекомоопыляемое растение. Покрывало соцветия снаружи зеленовато-белое,
изнутри мясо-красное. Соцветие – тонкий початок, как у предкового вида, испускает
неприятный запах. Растение опыляется мухами и мелкими жуками – самыми распространёнными
опылителями, встречающимися практически повсеместно. Семенное размножение обычно
происходит на крупных островах, где появляются расы, отличающиеся размером и
окраской листьев. Растения из береговых популяций разных островов более сходны.
Мадагаскарская
ксеноколоказия (Xenocolocasia madagascarensis)
Порядок: Аронниковые (Arales)
Семейство: Ароидные (Araceae)
Место обитания: Мадагаскар, пересыхающие болота в западной части острова.
В эпоху человека таро (Colocasia spp.) было одним из самых древних и распространённых
культурных растений, и широко культивировалось в тропиках Старого Света, в том
числе на отдалённых океанских островах. Значительная изменчивость и широкий
ареал позволили растению сохраниться в неоценовой флоре тропиков, и образовать
ряд полноценных видов. Несмотря на одомашнивание и сопутствующие ему изменения,
таро оказалось достаточно конкурентоспособным по отношению к островной флоре
Мадагаскара, и в раннем неоцене заняло свою экологическую нишу в местной флоре.
Потомок таро в болотах острова – мадагаскарская ксеноколоказия, крупное травянистое
влаголюбивое растение. Стебель этого вида представляет собой многолетний клубень
весом до 10 кг, развивающийся под землёй и покрытый тонкой прочной кожицей.
От материнского клубня отрастают столоны длиной до 2 метров, на концах которых
развиваются дочерние клубни. Благодаря активному вегетативному размножению растение
образует обширные заросли в благоприятных условиях.
На поверхности земли растение образует мощный ложный стебель, который состоит
из черешков листьев, сильно расширенных в основании и вложенных один в другой,
как у банана. Заросли этого вида немного похожи на банановые из-за крупных листьев
длиной до 3 метров, имеющих узко-стреловидную форму. Длина листа в 5-6 раз превышает
максимальную ширину. Листовая пластинка кожистая, гладкая, с двумя «ушками»
в месте прикрепления листовой пластинки к черешку и с тонкой длиной капельницей
на кончике. Окраска взрослого листа ярко-зелёная с коричневатой зоной вдоль
средней жилки; молодые листья розовые, по мере развития становятся коричневатыми,
далее зеленеют. Листья и клубни растения обладают жгучим вкусом, однако их поедают
местные растительноядные млекопитающие. В клубнях растения часто развиваются
личинки жуков.
Растение обладает чётко выраженной сезонностью вегетации с сезоном покоя в засуху.
При пересыхании болота листья растения быстро отмирают, а клубень втягивается
глубоко в почву за счёт усыхания корней. В новый вегетационный сезон молодая
часть клубня нарастает на вершине.
Цветение происходит в середине влажного сезона, когда растение образовало все
листья текущего сезона. Соцветие – тонкий початок, окружённый мясо-красным покрывалом.
Растение опыляется главным образом жуками. Плоды – сладкие красные ягоды; семена
распространяются птицами и млекопитающими.
Продолжительность жизни растения – до 10 лет.
«Речная
маргаритка» (Spongiolemna gigantissima)
Порядок: Аронниковые (Arales)
Семейство: Рясковые (Lemnaceae)
Место обитания: субтропические и тропические районы Дальнего Востока и Восточной
Азии, Японские острова.
Рясковые – самое специализированное и наиболее сильно изменившееся семейство
цветковых растений. Их тело превратилось в листец – своеобразный сплав листа
и стебля. У некоторых рясковых могут отсутствовать корни. Цветки рясковых редуцированы
до пестиков и тычинок, прикрытых тонкой обёрткой.
Несмотря на такую глубокую дегенерацию тела, рясковые являются процветающей
группой растений. Благодаря высокой скорости роста и лёгкости вегетативного
размножения рясковые образуют сплошной покров на поверхности водоёмов, а также
являются важным пищевым ресурсом для водных животных.
В голоцене рясковые были представлены миниатюрными растениями, и именно к этому
семейству принадлежало самое маленькое растение Земли – вольфиелла (Wolfiella),
которая имела длину всего лишь около 0,5 мм. В неоцене семейство также представлено
преимущественно миниатюрными растениями, мало изменившимися по сравнению с эпохой
человека. Но среди них появилось несколько специализированных форм, в том числе
один своеобразный гигант. Это «речная маргаритка», водное растение, плавающее
по поверхности рек и озёр Восточной Азии. На первый взгляд «речную маргаритку»
можно спутать с другими растениями – плавающими листьями кувшинок. Но при внимательном
рассмотрении разница становится очевидной. Округлые листецы этого вида, отдалённо
похожие на листья кувшинки, имеют диаметр около 15 – 20 см. Их края слегка изрезанные
и волнистые, лопасти немного загнуты вверх и листец растения напоминает стилизованное
изображение цветка. Тело растения в нижней части губчатое, с развитой воздухоносной
паренхимой. Из-за многочисленных пузырьков воздуха оно имеет серебристую окраску.
Верхняя сторона листеца плотная и кожистая, ярко-зелёного цвета. Одно взрослое
растение способно одновременно наращивать по краям несколько новых листецов.
Почки, из которых они образуются, развиваются по краям лопастей листеца. На
ярком солнце молодые листецы краснеют. На северном краю ареала осенью растение
продуцирует мелкие выводковые почки, похожие на листецы обычной ряски. Они плавают
по поверхности воды и собираются у берегов. Зимой взрослые растения отмирают,
а выводковые почки погружаются на дно, где успешно зимуют. Весеннее потепление
и усиление освещённости стимулирует их развитие.
Скорость роста этого растения очень велика. На развитие одного листеца от появления
до достижения полного размера уходит около недели, и на одном взрослом листеце
может одновременно развиваться до семи листецов нового поколения. На нижней
стороне листеца растёт «борода» из тонких неветвящихся корешков, достигающих
длины 10 см. Они активно всасывают растворённые в воде органические вещества,
благодаря чему это растение эффективно очищает воду.
Это растение двудомное. Мужские растения отличаются меньшим размером, но вегетативно
размножаются более интенсивно, и биомасса мужских и женских особей в популяции
приблизительно одинакова. Цветки «речной маргаритки» редуцированы, как у всех
рясковых, но отличаются особым строением. Пестики и тычинки у них очень длинные
и крепкие, они прикрыты у основания небольшим листочком-покрывалом, которое
быстро засыхает. Цветки развиваются между лопастями листеца. «Речная маргаритка»
является энтомофильным растением, но её отношения с насекомыми значительно более
просты, чем, например, у орхидей. На пестики и тычинки, торчащие над зарослями
«речной маргаритки», любят садиться для отдыха мелкие стрекозы. Они переносят
липкую пыльцу этого растения на ногах. Семена «речной маргаритки» мелкие, покрытые
липкой слизью. Они долго плавают по поверхности воды, и прилипают к перьям или
шерсти водных четвероногих, которые расселяют растение.
Идею о возможности появления таких растений высказал Nem, участник форума.
Озёрная
разнолистка (Heterophyllemna integra)
Порядок: Аронниковые (Arales)
Семейство: Рясковые (Lemnaceae)
Место обитания: Евразия, болота и озёра в умеренном и субтропическом поясе.
Семейство рясковые демонстрирует особое направление эволюции – тенденцию к крайнему
упрощению строения тела, которое у них превратилось в листец, образованный из
стебля и листа. Простота строения тела, высокая скорость роста и размножения,
а также простота расселения и способность к адаптации – эти качества оказались
залогом успешного выживания рясковых в эпоху голоцена. В неоцене эти растения
пережили новый виток эволюции – усложнение строения тела путём специализации
листецов и образования модульной конструкции единого тела растения. Модульные
ряски являются потомками ряски многокоренника (Spirodela), широко распространённой
в эпоху человека. Новая группа модульных рясок характерна для водоёмов Евразии
в различных климатических поясах, кроме самых холодных территорий. Подобно своим
предкам, это растения, плавающие на поверхности воды.
Озёрная разнолистка из пресных водоёмов Евразии демонстрирует характерное строение
тела растений этой группы. Это мелкое растение диаметром не больше 15-20 мм
с составным телом из листецов разного строения и назначения. Основной листец
является главным в составе растения. Он активно участвует в вегетативном размножении,
образуя почки, из которых развиваются новые листецы, и зимующие почки, способные
переживать зиму подо льдом. В нём также отлагается запас крахмала, расходуемый
в процессе зимовки. Этот листец живёт два года, развиваясь из почки. Из него
образуются листецы всех остальных типов. Он участвует в вегетативном размножении
в сезон роста.
Несколько листецов по краям основного являются специализированными органами
фотосинтеза. Они тонкие и полупрозрачные, лишены корней и приподняты над водой,
работая как паруса и органы фотосинтеза. Благодаря малой толщине фотосинтез
происходит во всей толще этих листецов.
Специализированным типом листеца является генеративный, или цветочный листец
– пузыревидный листец с прозрачной верхней стороной, внутри которого развиваются
группы цветков. Когда редуцированные цветки созревают, они прорывают прозрачную
плёнку и опыляются мелкими насекомыми. Этот листец снабжён запасом крахмала
в нижней части, после опыления он отрывается от материнского растения и расходует
все накопленные ресурсы на дозревание семян, после чего отмирает, и семена попадают
в воду. Они распространяются потоками воды или водными птицами, прилипая к их
оперению.
К зиме фотосинтезирующие листецы отмирают и рассасываются, а на основном листеце
второго сезона роста образуются зимующие почки, которые отрываются от него и
тонут. Ресурсы этого листеца полностью исчерпываются на их образование, и он
отмирает. Листецы первого года жизни зимуют под водой, образуя лишь единичные
цветочные почки.
Схизолемна
садовая (Schizolemna anabaenophila)
Порядок: Аронниковые (Arales)
Семейство: Рясковые (Lemnaceae)
Место обитания: Евразия и Северная Америка, озёра холодного климата.

Рисунок Биолога
Растения семейства рясковых успешно пережили эпоху человека
благодаря неприхотливости в выборе мест обитания, высокой скорости размножения
и лёгкости расселения. Упрощение строения тела позволило им сократить до минимума
расходы на поддержание жизнедеятельности, направив всю энергию на размножение.
В процессе эволюции в раннем неоцене среди рясковых появились виды, использующие
иные жизненные стратегии. Одним из удачных эволюционных ходов стал симбиоз одного
из видов ряски с сине-зелёной
водорослью анабеной (Anabaena). Этот шаг позволил ряске расти в олиготрофных
озёрах благодаря способности водоросли фиксировать азот из атмосферы и использовать
его для собственных нужд. Ряска схизолемна садовая предоставила симбиотической
водоросли благоприятные условия для жизни в обмен на часть полученного ею азота.
Специализация к симбиозу привела к изменению облика этого вида.
Взрослая особь схизолемны садовой – это дисковидный или овальный листец диаметром
около 10 мм. Средняя часть этого листеца полупрозрачная, и под слоем покровных
тканей растения лежит рыхлая паренхима, в которой живут и размножаются клетки
водорослей. В летнее время растение становится «теплицей» для водоросли (отсюда
видовой эпитет), и она обильно размножается внутри тела растения. Хлорофиллоносные
ткани самой ряски сохранились лишь в виде каймы по краю листеца. Летом схизолемна
размножается вегетативно путём образования боковых листецов. Процесс образования
нового листеца своеобразен: он отшнуровывается от края взрослого растения, захватывая
часть «теплицы» с симбиотическими водорослями. Также часто происходит простое
деление взрослого листеца путём образования поперечной перегородки, по которой
он просто разваливается надвое – отсюда название: «схизо-» означает «расщеплять».
Благодаря симбиотической водоросли схизолемна садовая способна расти в олиготрофных
озёрах за полярным кругом, в полной мере используя преимущества полярного дня.
Это растение является важным кормовым ресурсом для местных болотных птиц.
Осенью растение образует зимующие почки внутри старого листеца, который отмирает.
Почки сохраняют внутри себя споры водорослей, из которых развивается новая популяция
симбионтов. Весной после таяния льда почки всплывают; на ногах водоплавающих
птиц они расселяются по новым водоёмам, где начинается их рост.
Голосеменные
растения |
Неарктическая
сосна (Pinus nearctica)
Порядок: Сосновые (Pinales)
Семейство: Сосновые (Pinaceae)
Место обитания: североамериканская лесотундра.
Отношения между лесом и тундрой никогда не были однозначными. Когда мир охватывало
потепление, тундра отступала в высокие широты, а на её прежние владения наступал
лес; когда наступал ледниковый период, ситуация была противоположной. Начало
неоцена было отмечено резким отступлением лесов в низкие широты и обширным оледенением.
В те времена полоса лесотундры отсутствовала почти полностью.
Но ледниковый период завершился, климат потеплел, и тундра отступила опять.
Между ней и таёжными лесами снова появилась переходная зона лесотундры, заселённая
растениями обеих природных зон. В Северной Америке и особенно на северо-западе,
в Берингии, одно из характерных деревьев лесотундры – неарктическая сосна.
Неарктическая сосна является потомком сосны Банкса (Pinus banksiana) эпохи голоцена,
и это довольно небольшое дерево – примерно 4-5 м в высоту. Она растёт редкостойными
рощами на более сухих и защищённых от полярных ветров местах, избегая болот
и низин. На продуваемых холодными ветрами местах это дерево образует стланиковую
форму высотой не больше полуметра. Ствол неарктической сосны часто бывает неправильной
формы, сильно искривлённый, и выглядит почти как гигантский куст. Ветки у этой
сосны раскидистые, покрыты длинными парными иглами и часто отрастают от ствола
под разными углами. Нижние ветви растения часто могут догонять в росте основной
ствол, образуя очень широкую крону, поэтому кроны этих растений часто смыкаются
и в рощах неарктической сосны всегда довольно тенисто и сумрачно. Каменистая
почва в таких местах покрыта слоем опавших игл, а из-за недостатка света большинство
травянистых растений не может там расти. Рост этого дерева очень медленный,
часто не превышает нескольких сантиметров в год.
Несмотря на небольшую высоту неарктической сосны и отсутствие травянистых растений
в её зарослях, крупные полярные млекопитающие часто посещают эти странные леса.
Там всегда можно укрыться от полярного ветра и там значительно меньше снега.
Летом заросли этой сосны помогают животным спасаться от гнуса.
Как и большинство хвойных растений, неарктические сосны опыляются ветром. Размножение
этих растений происходит в начале полярного лета, и растения производят обильную
мелкую пыльцу. Созревание семян происходит долго: шишки могут развиваться до
пяти лет, прежде чем полностью сформируются семена – небольшие, лёгкие, снабжённые
крылышком. Они легко разносятся ветром на большие расстояния. В первые годы
жизни они растут очень медленно и гибнут от различных травоядных, но способны
противостоять холодам полярной зимы. В отличие от многих хвойных, это растение
легко переносит повреждение верхушки животными или холодом и со временем формирует
многоствольное дерево с раскидистой кроной.
Первые семена образуются на этом растении в возрасте около двадцати лет, а продолжительность
жизни может составлять до трёхсот лет и больше.
Этот вид растений открыл Bhut, участник форума.
Степная
сосна (Pinus cerifera)
Порядок: Сосновые (Pinales)
Семейство: Сосновые (Pinaceae)
Место обитания: североамериканские прерии.
В неоцене Северная Америка отличается достаточно необычным распределением растительности.
На западе, от Берингии на севере до Мексики на юге тянутся высокие Скалистые
горы, на склонах гор растут леса. На востоке располагаются старые и значительно
более низкие горы – Аппалачи, также поросшие лесами – на севере хвойными, южнее
смешанными и широколиственными. Существуют и другие типы лесов – на юго-востоке
это мангры, на северо-западе – влажный дождевой лес умеренного климата. В центральных
районах Северной Америки находится длинная полоса прерий, тянущаяся практически
от тундры на севере до полупустыни и пустыни на юге. Там также растут деревья,
но не образуют протяжённых лесов, а только отдельные рощи и редколесья. Один
из местных видов – степная сосна.
Сосны являются одними из самых древних по происхождению растений на Земле; они
успешно пережили голоцен и эпоху антропогенного прессинга, ранний неоцен и климатические
изменения. Эти растения смогли колонизировать даже суровую тундру – это сделала
неарктическая сосна. В прериях поселилась
её дальняя родственница – сосна степная.
В отличие от своей приземистой северной родственницы, степная сосна – это стройное
высокое дерево, около 40 м в высоту. У степной сосны крона легко узнаваема:
она узкая, коническая, с острой вершиной; её ветки относительно длинные, тонкие
и маловетвистые, всегда повисают. Ствол этого дерева также относительно тонкий:
обычно не больше 2.5-3 м в диаметре. При своей высоте дерево кажется очень тонким
и хрупким, что на самом деле не так: крона легко выдерживает натиск сухих ветров
прерий, и растение не падает, если пекари-бизон
решит почесаться об её ствол.
Кора растения гладкая, желтовато-серая, на молодых побегах сизоватая, отшелушивается
тонкими пластинками. Все ветки степной сосны растут в верхней трети её ствола.
У земли находятся ветви лишь у самых молодых растений. В рощах степной сосны
тени всегда мало, а растения растут на расстоянии 10-20 метров друг от друга.
Тем не менее, пекари-бизоны всё равно заходят в эти заросли: там всегда много
травянистых растений, которых можно съесть, а кора степной сосны достаточно
шершавая, чтобы об неё можно было почесаться.
Побеги растения имеют очень узнаваемый облик: хвоя длинная и тонкая (примерно
20 см в длину и 2-3 мм в ширину), с обильным восковым налётом, особенно на молодых
побегах и хвоинках. Из-за этого концы побегов растения очень светлые, и крона
дерева издалека кажется зеленовато-белой. Воск с хвои сосны часто используют
для строительства гнёзд местные осы.
Как и другие сосны, степная сосна опыляется ветром, и распространяется тоже.
Форма шишек этого вида своеобразна: у шишек широкие чешуи, накрывающие друг
друга в неправильном порядке. Длина шишки – до 15 см при ширине около 4 см.
У семян широкое крылышко, но они не такие мелкие и лёгкие, как семена некоторых
других североамериканских хвойных деревьев, и путешествуют на меньшие расстояния,
зато растут быстрее. Проростки теневыносливы и спокойно прорастают в зарослях
местной степной розы и других кустарников. Это довольно важно для сеянцев этого
вида, т.к. разные степные травоядные звери, вроде пекари-бизонов, охотно поедают
молодые растения степной сосны, пока те не станут достаточно большими, чтобы
быть в безопасности.
Если растение не будет съедено в первые годы своей жизни, то в возрасте 20 лет
оно сможет достичь высоты 15 метров и первый раз дать семена. Продолжительность
жизни этого вида достигает 200 лет.
Этот вид растений открыл Bhut, участник форума.
Болотная
лиственница (Larix paludiphila)
Порядок: Сосновые (Pinales)
Семейство: Сосновые (Pinaceae)
Место обитания: Западная Сибирь, болота.
В эпоху человека лиственницы (Larix spp.) были самыми распространёнными деревьями
на Земле и образовывали обширные лесные массивы на территории Евразии. В конце
эпохи человека их ареал сузился из-за массовой вырубки лесов, и в дальнейшем
эти деревья отступали к югу, теснимые ледниками во время голоцен-неоценового
оледенения. По окончании оледенения они одними из первых заселяли территории,
с которых отступали льды, и вновь образовали обширные лесные массивы. На территории
Западной Сибири им пришлось столкнуться с новыми для себя условиями жизни: равнины
заболачивались из-за медленного, но непрерывного поднятия северного края Евразийской
литосферной плиты, освободившейся от тяжести ледникового щита. В результате
часть популяции предкового вида оказалась на обширных заболоченных территориях,
где в процессе эволюции сформировался новый вид – болотная лиственница.
Этот вид деревьев – своеобразный морозоустойчивый аналог североамериканского
болотного кипариса (Taxodium), образующий протяжённые заросли на заболоченных
участках. Популяции этого вида распространены по всей Западно-Сибирской равнине,
за исключением самых северных районов. Болотная лиственница выдерживает подтопление
нижней части ствола и нормально растёт в водоёмах, глубина которых достигает
2 метров. Этот вид отличается исключительно твёрдой и стойкой к гниению древесиной:
даже у погибших растений стволы долгое время остаются стоять вертикально. В
источенной насекомыми древесине образуются дупла, в которых гнездятся многочисленные
лесные и болотные птицы.
Болотная лиственница достигает высоты 25-30 метров, диаметр ствола взрослого
дерева до 2 метров. Крона этого дерева относительно рыхлая, пропускает значительную
часть света вниз, к поверхности болота. Очертания кроны ширококонические, диаметр
её в нижней части может достигать 15 метров. Хвоя нежная, мягкая, опадающая
на зиму; длина хвоинок достигает 80-100 мм, они растут пучками по 5-8 штук.
Молодые женские шишки ярко-розового цвета, позже краснеют и буреют. Семена этого
растения созревают в течение года и высыпаются из шишек весной, во время половодья.
Благодаря значительному количеству жирного масла семена плавают по поверхности
воды. Они распространяются потоками воды или прилипают к лапам и клювам водных
птиц. Проростки этого вида легко выдерживают затенение и в первые годы жизни
растут очень медленно. Выживаемость и всхожесть семян сравнительно низкие: не
более 10% образовавшихся семян дают проростки.
Корневая система этого вида раскидывается на большое расстояние от растения
– отдельные корни на 10 метров и больше. В основании ствола имеется 4-5 мощных
досковидных корней, укрепляющих растение в вязком грунте. В отличие от большинства
хвойных деревьев, этот вид хорошо размножается вегетативно, активно давая корнеотпрыски
на расстоянии до 5 метров от материнского растения. Чаще всего корневая поросль
образуется на внешнем крае зарослей болотной лиственницы и растёт значительно
быстрее, чем растения из семян. Новые деревья могут подниматься к поверхности
воды с двухметровой глубины, проводя под водой до 5 лет и более, прежде чем
их верхушка покажется из воды. У формирующихся под водой побегов длинный прирост,
тонкие ветви и расширенные хвоинки. На поверхности воды такие растения начинают
формировать типичные для вида побеги, а ствол начинает быстро утолщаться.
Продолжительность жизни болотной лиственницы достигает 500 лет. Первые семена
дерево даёт в возрасте 20 лет, и далее образует их почти ежегодно. Годы наибольшего
урожая семян повторяются через 4-6 лет.
Восточная
горная ель (Picea appalachiensis)
Порядок: Сосновые (Pinales)
Семейство: Сосновые (Pinaceae)
Место обитания: склоны Аппалачей близ вершин гор.
Аппалачские горы Северной Америки во многом отличаются от Скалистых гор на западе
материка. Они гораздо ниже и их склоны более пологие, поскольку они значительно
старше. Вершины этих гор сильно разрушены выветриванием, на них часто образуются
плато. Деревья, растущие на склонах Скалистых гор – чаще хвойные, чем лиственные:
выше по склонам образуются сплошные хвойные леса. Горные леса Аппалачей смешанные,
гораздо более разнообразны по видовому составу. Типичными деревьями этих лесов
являются дубы, но хвойные также растут в этих лесах. Восточная горная ель –
один из видов хвойных деревьев востока материка.
Её внешность типична для представителей рода – это высокое, стройное дерево
около 20 м в высоту, растущее одним стволом, с конической кроной и ветками-лапами
до земли. В отличие от елей эпохи человека из-за «плакучих» ветвей она выглядит
непропорционально тонкой – ширина кроны в средней части около 100 см; вершина
остроконечная.
Ветки у горной ели длинные и узкие, но с многочисленными боковыми ответвлениями,
с широкими и длинными иглами, 20-40 мм в длину. Хвоя светло-зелёного цвета,
острая и густая. Ветви образуют довольно плотную крону, которая почти не пропускает
солнечный свет, поэтому корни горной ели не имеют соперников в добыче питательных
веществ и воды из почвы, зато мелкие звери часто устраивают под этим деревом
свои норы – под такой елью всегда относительно теплее и менее снежно даже в
ветреные горные зимы. Благодаря узкой кроне дерево легко сбрасывает с кроны
накапливающийся снег.
Горная ель опыляется при помощи ветра, как и другие хвойные, а её семена распространяются
животными. У этого дерева короткие толстые шишки – 5-10 см в длину – и пропорционально
большие и тяжёлые семена, лишённые крылышка. Они крупные, богатые питательными
веществами, главным образом жирами, и служат пищей разным грызунам и мелким
птицам. Благодаря им часть семян оказывается закопанной в землю в качестве запаса
на зиму. Если хозяин забыл о своих запасах или был пойман хищником, весной спрятанные
семена прорастают, обеспечивая воспроизводство вида.
Врагами этого вида являются главным образом насекомые и грибы, вызывающие заболевания
и гибель растений этого вида на больших территориях. Травоядные звери лишь изредка
поедают макушки этих растений во время зимней бескормицы, но питательная ценность
их очень мала. После повреждения вершины дерево долго болеет и восстанавливается.
Если горную ель в молодом возрасте не повредят травоядные животные, она вполне
сможет прожить несколько сотен лет. Между этой елью и горным
дубом происходит постоянная конкуренция – дуб более светолюбив и его проростки
не могут развиваться среди тенистых крон горной ели, но его поросль вытесняет
ель с более плодородных участков почвы.
Этот вид растений открыл Bhut, участник форума.
Тихоокеанская
туя (Thuja pacifica)
Порядок: Сосновые (Pinales)
Семейство: Кипарисовые (Cupressaceae)
Место обитания: тихоокеанские влажные леса Северной Америки.
В неоцене к западу от Скалистых гор Северной Америки сложилась очень необычная
экосистема: дождевой лес с умеренным климатом. С океана регулярно приходят дождевые
облака, обеспечивающие высокий уровень влажности воздуха и почвы. В этих лесах
складывается природное сообщество из влаголюбивых видов растений и животных.
Одним из фоновых видов деревьев, определяющих облик этих лесов, является тихоокеанская
туя – выносливое и влаголюбивое хвойное растение.
Тихоокеанская туя является потомком туи складчатой (Thuja plicata) эпохи голоцена.
Подобно своему предку, это очень высокое дерево, вырастающее до 60-65 метров
в высоту, с многочисленными корнями, которые врастают глубоко в почву и помогают
растению выдерживать удары стихии – ветра, наводнения, а на горных склонах –
схождение селевых потоков. Кроме того, корни этих деревьев играют важную роль
в укреплении горных склонов, а могучие стволы задерживают крупные камни, скатывающиеся
со склонов. Ствол у этого дерева ровный и прямой, покрыт красноватой мелкобороздчатой
корой; ветвление у взрослого растения начинается на значительной высоте – в
верхней трети ствола. Ветви образуют широкую коническую крону на самой макушке
дерева. Ветви длинные, относительно прямые; фотосинтезирующие побеги покрыты
широкими чешуеобразными листочками, на поверхности которых выделяется тончайший
слой воска, помогающий скатываться дождевой воде. Сами листочки значительно
шире, чем у любого другого вида туи в эпоху человека. Благодаря содержанию смолистых
веществ все части дерева приятно пахнут.
Тихоокеанская туя обильно образует пыльцу в начале лета, когда заканчиваются
весенние дожди и погода становится относительно сухой. Это дерево не только
опыляется ветром, но и распространяется с его помощью: её шишки пропорционально
небольшие, длиной до 10 см, из 4-5 пар крупных деревянистых чешуек. Семена небольшие
и лёгкие, снабжены широким крылом и распространяются ветром. Западное побережье
Северной Америки является ветреным местом, и хотя в подлеске тихоокеанских лесов
ветер ощущается слабо, на высоте крон тихоокеанской туи присутствие ветра ощущается
очень сильно, и пыльца, а также и семена этого дерева преодолевают очень большое
расстояние – достаточное для обмена генетическим материалом между отдалёнными
популяциями.
Этот вид – очень долговечное дерево, но платой за долголетие является низкая
выживаемость проростков. Её ростки изначально очень маленькие и растут медленно
– пока растение станет достаточно сильным, у него найдётся немало врагов из
числа грибков и насекомых. Молодые растения в подлеске часто оказываются в угнетённом
состоянии, находясь в тени взрослых экземпляров. Если молодая тихоокеанская
туя сможет выжить в первые десять-двадцать лет жизни и достичь в высоту хотя
бы 5-10 метров, есть большая вероятность, что она сможет вырваться в полог леса
и прожить несколько веков – возраст взрослых экземпляров доходит до 700-800
лет, и даже больше.
Этот вид растений открыл Bhut, участник форума.
Споровые растения
- папоротники
|
Листовик разнолистный (Epiphyllitis heteromorphus)
Порядок: Циатейные (Cyatheales)
Семейство: Асплениевые (Aspleniaceae)
Место обитания: тёплые влажные леса по западному и южному побережью Четвероморья.
Потепление климата привело к формированию на юге Европы широколиственных лесов,
несущих отдельные особенности, характерные для тропического леса. Но область
распространения этих лесов очень ограниченна – зажатое между Европой и Африкой
Средиземноморье превратилось в солёную пустыню, и влага поступает в эти леса
только с Четвероморья – огромного солоноватого озера-моря на юго-востоке Европы.
Тепло- и влаголюбивые леса покрывают его южные и западные берега. Растения,
составляющие эти леса, являются потомками местных европейских растений – горы
и Средиземноморская котловина надёжно изолировали их от «настоящих» тропических
лесов. Кроме особенностей деревьев, образующих эти леса, тропический облик
им придаёт наличие характерной жизненной формы – эпифита. В европейских лесах
тропического типа эпифитами являются мхи и папоротники.
Один из эпифитных папоротников этих мест – листовик разнолистный, потомок обычного
европейского папоротника листовика (Phyllitis scolopendrium) эпохи голоцена.
Этот вид встречается в развилках ветвей, где скапливается достаточное количество
растительных остатков для прорастания его спор.
Подобно своему предку, этот вид является розеточным растением, которое образует
плотную воронку листьев. У этого вида развиваются вайи двух типов – стерильные
и фертильные (отсюда видовой эпитет). Стерильные нижние вайи округлые, широкие
и кожистые. В них скапливается растительный мусор, птичий помёт и другие вещества,
из которых растение получает питание. Туда проникают придаточные корни, развивающиеся
на укороченном стебле растения. Будучи молодыми, стерильные вайи какое-то время
фотосинтезируют, но по мере их развития количество хлорофилла в них снижается
и развивается пробковая ткань. Эти вайи становятся жёсткими, и только на краях
ещё остаётся область растущей ткани. Похожий тип вай развивается у папоротника
«олений рог» (Platycerium), который успешно дожил до эпохи неоцена, но обитает
в Юго-Восточной Азии и Меганезии.
Верхние вайи листовика разнолистного более нежные. Они фотосинтезируют, и на
их нижней стороне вдоль боковых жилок, под углом к центральной жилке, развиваются
спорангии, собранные в линейные сорусы. Этот вид имеет удлинённые, цельные
и плотные вайи – эта особенность унаследована им от предка. Фертильные вайи
этого папоротника имеют слегка волнистые края и довольно нежную ткань. При
кратковременных зимних похолоданиях они могут отмирать. Но до этого времени
растение формирует в центре розетки несколько молодых вай, которые зимой не
разворачиваются и прикрывают уязвимую точку роста. Они выделяют богатую полисахаридами
слизь, которая защищает точку роста от повреждения холодом. Весной, когда становится
достаточно тепло, они раскрываются и новые фертильные вайи отрастают очень
быстро. Полностью развитое растение образует розетку диаметром около 50 – 60
см.
Листовик разнолистный прочно прикрепляется к коре деревьев корнями, которые
легко врастают в неровности коры и по мере её утолщения проникают глубже. На
старой части корневища отрастают новые побеги, и одно растение может образовывать
заросли из нескольких крупных розеток.
Это растение размножается спорами, которые легко распространяются ветром и
насекомыми. Но у листовика разнолистного есть ещё один способ размножения.
Фертильные вайи легко образуют выводковые почки, если повреждены. Обычно почка
быстро образуется на месте разрыва жилок, и обломленный кусочек фертильной
вайи тоже может дать хотя бы одну почку, из которой успешно развивается молодое
растение.
Споровые растения
- хвощи
|
Эпифитный
хвощ (Equisetum dendrophilus)
Порядок: Хвощовые (Equisetales)
Семейство: Хвощовые (Equisetaceae)
Место обитания: леса Балкан и Южной Европы.
Климат неоцена отличается теплотой и влажностью. Это привело к расширению тропического
и субтропического поясов, и смещению границ ареалов теплолюбивых растительных
сообществ в более высокие широты. В Европе теплолюбивые леса появились на юге,
в предгорьях Альп, на Балканах и в Малой Азии. Южнее, за Альпами, расстилается
котловина, ранее бывшая дном Средиземного моря. Это место, где господствуют
солёные болота и суровые солончаковые пустыни. Ещё южнее Средиземноморской низменности
расстилаются саванны Северной Африки, прорезаемые огромной рекой – Сахарским
Нилом. Обширные равнины оказались непреодолимым препятствием для теплолюбивой
растительности Африки, а горы воспрепятствовали проникновению в Европу азиатских
теплолюбивых растений. Поэтому субтропические леса Европы образованы потомками
местных растений, эволюционировавшими в благоприятных условиях: огромное озеро-море
Четвероморье обеспечивает эти леса влагой.
В многоярусных лесах юго-востока Европы появилось множество возможностей для
эволюции. Гигантские деревья увиты лианами, а на ветвях, как в настоящем тропическом
лесу, обитают эпифиты. В основном, это разные папоротники – от мелких и нежных
до очень крупных с кожистыми листьями. Но среди их листьев (вай) разнообразной
формы выделяются длинные, похожие на метёлки, побеги другого спорового растения
– эпифитного хвоща, эндемика юга Европы.
Спороносному растению легче занять нишу эпифита, нежели цветковому – споры легко
распространяются ветром (орхидеи, часто растущие как эпифиты, имеют микроскопические
семена с редуцированным зародышем). Но у споровых растений есть одна уязвимая
черта: из споры появляется не побег взрослого растения, а заросток, половое
поколение. На нём развиваются женские и мужские половые органы – архегонии и
антеридии. А для оплодотворения и развития взрослого растения (спорофита) нужна
вода в капельно-жидком виде. Поэтому особенного успеха споровые растения достигли
в лесах с высокой влажностью воздуха.
В раннем неоцене моря юго-востока Европы вернулись в свои берега после иссушающего
ледникового периода, а затем слились в Четвероморье. Это благоприятно отразилось
на климате Балкан – климат стал более влажным и дождливым. Предок эпифитного
хвоща, очевидно, поселялся в этих условиях в ковре мха, а потом стал лазающим
растением. Постепенно потомки этого вида стали проходить полный цикл развития
в кронах деревьев. Так появился неоценовый эпифитный хвощ.
Эпифитный хвощ представляет собой корневищное травянистое растение. Его корневища
ползут по стволу вверх и вниз, скрываясь от солнца в трещинах коры. Покровы
корневищ толстые и губчатые – это надёжная защита от излишнего испарения воды.
Стебли эпифитного хвоща тонкие и невысокие – не более 20 см. Но это растение
образует густые подушковидные заросли сочно-зелёного цвета. Как у всех видов
хвощей, у этого растения нет листьев, а боковые ветви сидят в узлах стебля мутовками
по десятку и более. Нижние узлы стеблей расположены ближе друг к другу, и боковые
ветви здесь гуще, чем на верхушках побегов – так внутри зарослей сохраняется
оптимальный микроклимат в зоне роста корневища и молодых побегов.
Корневища эпифитного хвоща двух типов. Основное корневище относительно короткое,
со сближенными междоузлиями. От него отрастают вегетативные фотосинтезирующие
побеги. От основного корневища через определённое расстояние отрастают боковые
корневища второго типа. Эти корневища очень тонкие, с длинными междоузлиями,
редуцированными корнями и вегетативными побегами. Они ползут по стволу, а иногда
свободно свисают вниз с веток. Ветер раскачивает такое корневище и перекидывает
на соседние деревья. Там оно закрепляется, быстро образуя корни, вцепляющиеся
в кору дерева, и начинает расти, образуя характерные зелёные «ёлочки». Так это
растение расселяется.
Спороносные побеги этого растения сильно отличаются от вегетативных – они длинные
(достигают полутораметровой длины), без боковых ветвей, с узкой длинной спороносной
шишечкой на конце. Такие побеги развиваются в начале лета. Стебель спороносного
побега упругий, поэтому побег дугообразно изгибается и повисает, раскачиваясь
на ветру. Растение попросту эксплуатирует одну поведенческую особенность мух
– эти насекомые любят садиться на свисающие тонкие предметы. Не только ветер,
но и насекомые разносят споры эпифитного хвоща. Споры с выростами (элатерами)
цепляются за лапки мух и переносятся насекомыми в места, куда веер не смог бы
занести споры. Этот способ расселения даёт эпифитному хвощу заметное преимущество
в расселении.
Лучше всего развиваются споры, попавшие во влажные места – например, на рыхлую
кору дерева, удерживающую влагу. Из споры вырастает небольшой заросток, похожий
на шляпу с изрезанными краями. На его выпуклой средней части располагаются мужские
половые органы (антеридии), а на краевых лопастях сидят женские архегонии. Оплодотворению
способствует форма заростка: капля росы собирается на антеридиях, в неё попадают
сперматозоиды, а затем капля просто скатывается вниз, к архегониям. Иногда из
сцепившихся вместе спор формируются несколько заростков, сидящих рядом. В этом
случае возможно даже перекрёстное оплодотворение, дающее более жизнеспособное
растение.
Молодое растение в первые годы жизни образует только стерильные побеги и не
даёт длинных корневищ, с помощью которых расселяется. Примерно на четвёртом
году жизни у него формируются первые спороносные побеги, а окрепшее растение
в возрасте около восьми лет начинает расселяться с помощью корневищ. Оно образует
множество зарослей-клонов на протяжении десятков метров полога леса.
Близкий вид, висячий
древесный хвощ (Equisetum pendulus), как бы сохраняет особенности предыдущей
стадии развития эпифитных хвощей. Этот вид растёт на коре и держится за неё
корнями, прорастающими по мере её нарастания глубже. Вегетативные побеги этого
растения свободно свисают вниз, достигая длины метра и больше. Их боковые побеги
тонкие и длинные, поэтому заросли этого хвоща выглядят более пушистым. Спороносные
колоски образуются на концах побегов, внешне не отличающихся от вегетативных.
Спороносный колосок висячего хвоща длинный, и споры этого вида также переносятся
не только ветром, но и мухами.
Сибирский
сладкий хвощ (Equisetum dulcis)
Порядок: Хвощовые (Equisetales)
Семейство: Хвощовые (Equisetidae)
Место обитания: болота Западной Сибири, долины рек.

Рисунок Биолога
Хвощи в эпоху человека представляли собой сравнительно консервативную
и бедную видами группу споровых растений. Тем не менее, у многих видов был достаточно
широкий ареал, а некоторые виды успешно сосуществовали с человеком, становясь
сорняками на возделываемых землях. Деятельность человека и климатические изменения
на рубеже голоцена и неоцена привели к масштабным изменениям в экосистемах,
итогом которых стало активное видообразование выживших видов, среди которых
были и хвощи.
На территории болот Сибири произрастает один из новых видов хвощей эпохи неоцена
– сибирский сладкий хвощ. Это травянистое многолетнее растение, редко достигающее
высоты 1 метра; обычная высота вегетативных побегов – около 50-60 см. Эти побеги
несут мутовки коротких боковых ветвей – до 15 штук в мутовке. Стебель ярко-зелёный,
в узлах коричневатый.
У этого вида ползучее подземное корневище, запасающее к зиме крахмал и другие
полисахариды. Оно узловатое и легко распадается, когда крупное млекопитающее
выкапывает его из земли, и некоторые части корневища при этом избегают поедания.
В сезон вегетации от корневища последовательно отрастают вертикальные побеги
двух типов. После таяния снега из земли появляются мясистые бесхлорофилльные
побеги бледно-розового цвета, несущие на концах крахмалистые спороносные шишки,
содержащие крупные споры с плотной оболочкой. Высота таких побегов – до 10-15
см. У спороносных шишек сочная крахмалистая мякоть с выраженным кисло-сладким
вкусом; весной их охотно поедают грызуны и куриные птицы, которые рассеивают
споры с помётом. После половодья начинается период активного роста вегетативных
побегов. Они однолетние, отмирающие на зиму. Ткани богаты кремнезёмом, поэтому
зелень плохо поедают травоядные млекопитающие.
Этот вид – многолетнее растение, активно размножающееся вегетативно. Благодаря
переносу спор птицами и млекопитающими в популяциях растения повышается вероятность
совместного произрастания генетически разнородных растений, и появления у них
жизнестойкого потомства.
Бурые водоросли |
Листоватая
морская ряска (Algolemna prolifera)
Отряд: Фукусовые (Fucales)
Семейство: Альголемновые (Algolemnaceae)
Место обитания: Тихий океан севернее экватора, поверхность воды.
Рисунок Биолога
В эпоху человека крупные бурые водоросли были представлены
донными формами, которые прикреплялись к твёрдому субстрату при помощи корнеподобных
выростов – ризоидов. Слоевище этих видов часто достигало гигантских размеров,
вытягиваясь на многие метры. В неоцене одна из групп бурых водорослей совершила
настоящий прорыв: они полностью отделились от дна и превратились в плавающие
на поверхности воды растения, составляющие самостоятельное семейство водорослей.
За отдалённое сходство с плавающими растениями пресных вод они получили название
«морская ряска», хотя заметно отличаются от них крупными размерами тела.
Слоевище морской ряски отчётливо разделено на две части: крупный округлый поплавок
с кожистой оболочкой, заполненной губчатой воздухоносной паренхимой, и фотосинтезирующую
часть – листовидные отростки слоевища, быстро отрастающие по бокам поплавка,
за что растение получило название листоватая морская ряска. Диаметр поплавка
этого вида достигает 40-70 мм, длина листовидных выростов – до 20 мм. Окраска
тканей растения красновато-бурая, выросты полупрозрачные. На краях поплавка
время от времени образуются «точки роста», где происходит формирование нового
дочернего поплавка. Их расположение не фиксировано, и рост дочернего экземпляра
может начинаться в любом месте. Взрослое растение может образовывать одновременно
до трёх поплавков. Старое растение может разломиться пополам: каждая часть достраивает
недостающие части в течение нескольких дней. Обычно растения этого вида встречаются
поодиночке или с не полностью сформированными отростками. Во время штормов растения
благодаря небольшому размеру и весу практически не повреждаются волнами – лишь
отламываются боковые отростки. В тихую погоду морская ряска быстро разрастается,
образуя обширные заросли. Размножение этих растений главным образом вегетативное
– отделением дочерних растений. Рост очень бурный: за три-четыре дня растение
удваивает биомассу.
Споровое размножение у морской ряски происходит регулярно. Жизненный цикл диплофазный:
гаплоидное поколение отсутствует. Антеридии и оогонии развиваются по краям поплавка
между отростками слоевища и высвобождают половые клетки в воду, где происходит
оплодотворение и развитие молодых растений.
Морская ряска является важным компонентом океанских биоценозов, обеспечивая
домом и пищей мелких морских животных. Растения являются субстратом для полипов,
сидячих ракообразных. Также на них откладывают икру пелагические рыбы и мелкие
головоногие моллюски. Наличие этих растений, распространяющихся с течениями,
облегчает трансокеанское расселение морских животных.
В Южном полушарии обитает близкий вид: бахромчатая
морская ряска (Algolemna marginalis), у которой боковые «листочки»
слоевища короткие, образующие лишь бахрому по краю поплавков. Диаметр поплавка
у этого вида достигает 30-40 мм. Она более холодостойка, в экваториальных водах
быстро разрастается, но со временем деградирует. Популяции этого вида встречаются
в водах течений, омывающих берега Антарктиды, но течения часто заносят их в
экваториальные воды.
В Индийском океане встречается русалочья
лепёшка, или плоская морская ряска (Algolemna platycystis)
– вид с широкими поплавками, сплющенными и сверху, и снизу, и заполненными губчатой
тканью. Диаметр такого поплавка достигает 130-150 мм при толщине до 12-20 мм.
Его поверхность кожистая сверху и бугристая снизу, желтовато-коричневого цвета.
Дочерние растения у этого вида держатся на материнском слоевище очень прочно,
поэтому у данного вида иногда образуются «коврики» из одного родительского растения,
нескольких дочерних и многочисленных внучатых, которые распадаются только во
время сильного шторма.
Зелёные водоросли |
Буллония
плавающая (Bullonia natans)
Порядок: Кладофоровые (Cladophorales)
Семейство: Валониевые (Valoniaceae)
Место обитания: поверхность воды в тропической зоне всех океанов.

Рисунок Биолога
В эпоху человека среди морских зелёных водорослей существовали
весьма необычные виды, выглядевшие как зеленые пузыри. Такими были представители
рода валония (Valonia), обитавшие в тёплых регионах всех океанов. Эти виды жили
на дне, часто на коралловых рифах, но в неоцене по мере потепления климата они
перешли к жизни на поверхности воды. Их потомком является плавающая морская
водоросль буллония.
Слоевище буллонии представляет собой зелёные блестящие пузыри диаметром до 15
см (обычно с теннисный мяч), плавающие на поверхности воды и частично из неё
выступающие. Весь пузырь – это фактически одна большая многоядерная клетка.
Поверхность пузыря покрыта блестящей желеобразной кутикулой, отражающей излишек
солнечного света и не дающей таллому высыхать. Стенки пузыря толщиной около
1,5-2 см заполнены фотосинтезирующей «паренхимой» – участками клетки, в которых
имеются обширные газовые вакуоли. Эта часть клетки отделена от остальной цитоплазмы
септами Полость таллома заполнена газом. По достижении размера около 8-10 см
в диаметре клетка пузыря делится способом, очень похожим на почкование, и на
материнском пузыре сбоку образуется новый, иногда на небольшой ножке, но чаще
сидячий. Таких пузырей образуется много, и зелёный плот неправильной формы может
вырастать до 2-3 м в поперечнике. Во время шторма буллониевые маты распадаются,
но быстро регенерируют.
Размножается буллония так же, как и её предки: на нижней стороне пузырей формируются
половые органы, вырабатывающие гаплоидные изогаметы. Изогаметы сливаются в воде
попарно и зиготы делятся на 10-15 диплоидных клеток, каждая из которых по отдельности
даёт начало новому таллому водоросли.
На талломах-плотах буллонии часто поселяются различные мелкие животные – рачки,
моллюски и другие, а некоторые даже проделывают ходы в стенках более крупных
пузырей, не нарушая их целости, и живут внутри таллома, питаясь им же.
Этот вид растений открыл Биолог, участник форума.
Филодюналиелла
средиземноморская (Filodunaliella mediterranea)
Порядок Хламидомонадные (Chlamydomonadales)
Семейство: Дюналиелловые (Dunaliellaceae)
Место обитания: гипергалинные воды Средиземноморской котловины.

Рисунок Биолога
В эпоху человека очень немногие водоросли могли жить в сверхсолёной
воде. Самым известным родом таких организмов были дюналиеллы (Dunaliella) –
одноклеточные подвижные зелёные водоросли, очень похожие на хламидомонад и часто
вызывавшие цветение воды. В неоцене их потомки претерпели заметные изменения,
освоив новые экологические ниши. Филодюналиелла освоила жизнь в гипергалинных
водоёмах Средиземноморской котловины, где развивается в изобилии, практически
не имея врагов и конкурентов.
Филодюналиеллы из одноклеточных стали нитчатыми водорослями – они образуют короткие
цепочки из 4-5 овальных клеток, заключённых в общую слизисто-маслянистую капсулу.
Клетки около 10 мкм в диаметре, нити длиной около 50 мкм, и одна из двух концевых
клеток нити несёт полярный пучок жгутиков, вращение которых тянет цепочку клеток
вперёд.
Каждая клетка, подобно предковым формам, содержит крупный хлоропласт со сферическим
пиреноидом и крахмальными зёрнами, а также большое количество глицерина, который
защищает клетку от осмотического действия рапы. У предковых форм хлорофилл в
хлоропласте обычно маскировался большим количеством каротиноидов для защиты
от ультрафиолета, придавая клетке красно-оранжевую окраску. У филодюналиеллы
их количество снижено – в своем обиталище она прикрыта пластинками бактерии
палочки соляной крови (Halobacterium
sanguinisalis) и часто с ними сцепляется, поэтому её клетки имеют золотисто-жёлтый
цвет.
Размножается филодюналиелла так же, как и её предки: митотическим делением.
В неблагоприятных условиях (высыхание, опреснение) клетки внутри цепочки попарно
образуют зигоспоры, а оставшиеся без пары клетки отмирают. Зигоспоры имеют толстую
стойкую оболочку, позволяющую долгое время ждать возвращения подходящих условий
для роста, и легко переносятся ветром и животными, чтобы в новых условиях прорасти
множеством новых клеток, которые делятся с образованием цепочек. Большое количество
зигоспор этого вида встречается по берегам солёных озёр и на дне высохших водоёмов.
Выделение филодюналиеллой больших количеств глицерина обеспечивает возможность
существования галобактерий (источник углерода). При бурном размножении вместе
с галобактериями наблюдается тёмное кроваво-красное окрашивание рассола, а без
галобактерий – золотисто-жёлтое. В обоих случаях может наблюдаться маслянистость
поверхности воды из-за глицерина.
Этот вид простейших открыл Биолог, участник форума.



