НОСОХОДКИ
НОСОХОДКИ
| Главная | Библиотека | Форум | Гостевая книга |
Герольф Штайнер
Формы и жизнь ринограденций
НОСОХОДКИ
Формы и жизнь ринограденций
Харальд Штумпке
Бывший куратор музея Дарвиновского института на Хай-Яй-Яй, Майрувили.
Эпилог Герольфа Штайнера
Перевод Ли Чедвик
Русский перевод Павла Волкова
«Носоходки» были первоначально опубликованы в Германии под названием Bau und Leben der Rhinogradentia издательством Gustav Fischer Verlag, Штутгарт в 1957 году.
Copyright © 1964 by Gustav Fischer Verlag, Штутгарт.
Главы из книги появились в журнале Natural History в апреле 1967 года и переизданы здесь с разрешения.
15 таблиц и 12 рисунков в тексте в этой книге были нарисованы Герольфом Штайнером.
The University of Chicago Press, Chicago 60637 The University of Chicago Press, Ltd., London
English translation copyright © 1967 by Doubleday & Company, Inc

Предисловие к русскому переводу
Введение
Общие сведения
Описание некоторых групп
Однорылы (Monorrhina)
Примитивные типы носоходок (Archirrhiniformes)
Мягконосые носоходки (Asclerorrhina)
Наземные носоходки (Epigeonasida)
Иловые носоходки (Hypogeonasida)
Роющие носоходки (Georrhinida)
Хоботоноги (Sclerorrhina)
Мордоскоки (Hopsorrhinida)
Многорылы (Polyrrhina)
Четырёхрылые носоходки (Tetrarrhinida)
Шестирылые носоходки (Hexarrhinida)
Долгоносые носоходки (Dolichoproata)
Эпилог
Библиография
Дополнение к русскому переводу
Носоходки Харальда Штумпке (Герольфа Штайнера) пока не слишком хорошо
известны российским любителям вымышленных существ. Из всего, что издано о них
на русском языке, можно упомянуть лишь статью из
журнала «Природа» от апреля 2000 года, которую легко можно найти в Интернете, и
ещё несколько статей компилятивного плана. Возможно, когда-нибудь появятся и
другие публикации на эту тему, но на момент, когда пишутся эти строки, можно
смело утверждать, что больше не известно ни одной оригинальной статьи по
данному вопросу.
Пожалуй, самое лучшее, что может сделать любознательный читатель – это припасть
непосредственно к первоисточнику. Но оригинальная книга о носоходках, изданная
на немецком языке, была переведена на английский, французский и даже японский
языки, но только не на русский. Теперь ситуация в какой-то мере исправлена.
Располагая английским переводом книги, который мне удалось получить благодаря
помощи одного очень хорошего человека, я смог сделать всё от меня зависящее,
чтобы читатели из России и люди за рубежом, говорящие по-русски, смогли не
только разглядывать картинки, но и окунуться в полный загадок мир таинственных
существ под названием «носоходки», обитавших где-то на далёком архипелаге
Хай-яй-яй в южных областях Тихого океана. Картинки – это, конечно, хорошо. Но
без их истолкования они кажутся лишь плодом не слишком здоровой фантазии. Самый
популярный вариант комментария, находимого в Интернете, в этом случае обычно
сводится к вопросу «Какую траву курил автор?» и его вариантам (явно отражая
широту интересов самого комментатора). Но, возвращённые в контекст работы,
рисунки помогают по-новому взглянуть на мир забавных вымышленных существ и
обретают дополнительный смысл.
Почему «носоходки»? Глядя на рисунки, изображающие этих существ, можно
заметить, что не все они ходят на носах. В английском варианте название этих
животных – “snouters” – означает буквально «рыльники» (от англ. “snout” – рыло,
морда животного). Тем не менее, во время перевода книги я предпочёл оставить за
этими существами название «носоходки». Во-первых, этим я признавал приоритет
авторов статей, уже опубликованных на русском языке, где употребляется именно
это название. Во-вторых, название «носоходки» совершенно точно отражает смысл
их латинского названия – Rhinogradentia.
За рубежом тема носоходок очень популярна – эти существа часто мелькают на
сайтах, посвящённых вымышленным существам. В оригинальном издании книги про
носоходок были только чёрно-белые графические иллюстрации. Цветные иллюстрации,
изображающие макеты носоходок, взяты с различных сайтов в Интернете, в
частности, с сайта
Tokiwa Takeshi. Фотографии и рисунки без соответствующей подписи об
авторстве найдены на других сайтах в Интернете. Можно лишь пожалеть, что в
России мало таких же энтузиастов интереснейшего направления «вымышленной
биологии».
Книга Герольфа Штайнера – это прекрасная пародия на научные работы.
Фантастические существа, описанные в ней, представлены вниманию читателя не
менее точно и подробно, нежели реальные животные, описанные в «Жизни животных»
или какой-либо другой литературе, посвящённой реальным живым существам. Но
образы этих существ построены в свете знаний о принципах и закономерностях
эволюции. Каждая черта их образа имеет определённое значение, и её биологический
смысл объясняется на страницах книги. Пусть существа, описанные в ней,
откровенно вымышленные. Зато эту книгу можно считать своеобразным упражнением в
понимании теории эволюции. Перенося знания об эволюционном процессе на
совершенно фантастических животных, автор добивается глубокой реалистичности их
описаний – пожалуй, даже большей, чем десятилетия спустя достиг шотландский
палеонтолог Дугал Диксон в своих книгах про животных
будущего и «новых динозавров».
В распоряжении автора перевода была не оригинальная бумажная книга, а её
электронная копия в формате PDF, уже распознанная. Но качество распознания
книги было не слишком хорошим, поэтому некоторые слова пришлось восстанавливать
по смыслу, а отдельные фрагменты точно восстановить не удалось. Также потеряна
связь между ссылками на страницы оригинального издания и конкретными местами в
тексте, к которым они относятся. Но я надеюсь, что отдельные неясности не
повлияли на общее содержание книги. Желаю всем приятного чтения! Добро
пожаловать на Хай-яй-яй!
Павел Волков
Владимир, апрель 2009 г.
Среди млекопитающих отряд НОСОХОДОК занимает уникальное положение, которое в
первую очередь объясняется тем фактом, что они являются животными наиболее
странного строения из числа открытых в последнее время. И до настоящего времени
они оставались неизвестными науке, потому что их родина, архипелаг Хай-яй-яй
(по-немецки пишется Heieiei) в Южных Морях, не была обнаружена до 1941 года, и даже
тогда цивилизованные европейцы впервые посетили её, воспользовавшись редким
шансом, который дала война на Тихом океане. И в дополнение к этому
обстоятельству эта группа животных имеет особо важное значение, потому что её
представители демонстрируют принципы строения, типы поведения и экологические
типы, которые неизвестны более нигде, не только среди млекопитающих, но и
вообще среди позвоночных.
Честь открытия архипелага должна быть отдана шведу Эйнару
Петтерссон-Скамтквисту, который – спасаясь из японского плена – потерпел
кораблекрушение на острове Хай-дуд-дай-фи (Хейдадайфи). Это остров, который в
противоположность многим островам Южных морей, имеет невулканическое
происхождение даже при том, что на нём есть действующий вулкан
(Котсобуси-Козобауси) весьма значительной высоты (5740 футов), протягивается
примерно на двадцать миль с севера на юг и приблизительно на десять миль с
востока на запад, и состоит преимущественно из известняка и метаморфических
сланцев, а его самая высокая вершина – двуглавая гора Шоу-ванунда (Шауанунда)
высотой 7316 футов.
Климат острова чрезвычайно ровный, как и положено островам центральной и
восточной частей Tихого океана. Тропическая растительность, ботаническая оценка
которой едва началась, демонстрирует наряду с родами, распространёнными по
всему миру, много эндемичных форм архаичного характера (таковы Maierales,
близкородственные Psilotales, и род Necolepidodendron, который классифицируется
как принадлежащий к Lepidodendrales; а также Schultzeales, образующие ряд
великолепных деревьев девственного леса, которые должны располагаться около
Ranunculaceae; и многие другие). Следовательно, архипелаг Хай-яй-яй, к которому
принадлежит Хай-дуд-дай-фи, должен иметь древнее происхождение, что также верно
в свете геолого-палеонтологических находок почти исключительно из палеозойских
отложений; см. работу Эцио Спуталаве «Классификация милиолидных песков из
верхнего горизонта D16 Майрувили». И наконец, группа островов, очевидно,
полностью изолировалась от других континентов в верхнемеловую эпоху;
аналогичным образом считается, что архипелаг в этом плане является остатком
континента значительных размеров, и с тех пор – в противоположность Новой
Зеландии – он демонстрирует несравненно большие разнообразие и специфичность
туземных групп организмов при суммарной площади островов лишь чуть больше 650
квадратных миль.
Туземцы, найденные Скамтквистом при его прибытии в 1941 году, называли себя
Хууака-Хатчи (Эуача-Хат-ши). С тех пор они вымерли, но, согласно Скамтквисту,
они, кажется, были полинезийцами европеоидного облика. Их язык было невозможно
исследовать, потому что насморк, занесённый первооткрывателем, уничтожил этих
детей природы изнутри в течение нескольких месяцев. Из их культурных артефактов
удалось сохранить только несколько деревянных изделий (см. также Дейтерих 1944
и Комбинаторе 1943).
|
ТАБЛИЦА I |
Оружие было неизвестно Хууака-Хатчи. Существование мирного племени
поддерживалось естественным изобилием среды обитания. Не было никакой
чрезмерной рождаемости; напротив, «с незапамятных времен» двадцать два вождя
управляли приблизительно семьюстами душами. Это было самое большее, что смог
установить Скамтквист. Этот порядок имел удачные побочные последствия для науки
в том плане, что, несмотря на присутствие человека, сохранился экстраординарный
животный мир архипелага, что ещё более поразительно, потому что почти все
наземные животные были бы истреблены, если бы на них охотились более
интенсивно.
Несмотря на тот факт, что их родина была неизвестна, до этого момента носоходки
упоминались в одном случае. Ни больше, ни меньше, чем поэт Христиан Моргенштерн
примерно шестьдесят лет назад заявил о существовании носоходок в своей
известной поэме:
«На своих носах шагаем
Hасобята с насобемами1...
Сами о себе не знаем,
Не знакомы даже Брему* мы.И не Майер** дал нас Миру,
Не включил в словарь Брокгауз**...
Моргенштерна злая лира
Извлекла на свет нас за нос.На своих носах шагают...
Пусть не будет странно тем,
Кто случайно повстречает
Hасобят и насобем.»***
* Классический немецкий специалист по зоологии, первое издание в 1863 г., четвёртое (последнее) издание в 1918 г.
** Названия широко известных немецких энциклопедий.
*** Русский перевод дан по статье «Загадочные носоходки» С. Д. Фелдоянца и А. Зельбстандера – В. П.
Это краткое, но всё же ясное описание, которое отражает специфическую
походку этой носоходки даже в ритме стиха, точнейшим образом соответствует
описанию Nasobema lyricum2. Следовательно, нельзя
предположить иначе, нежели что Моргенштерн имел перед глазами экземпляр этой
носоходки или обладал подробной информацией о нём. Блидкуп (1945) в Das
Nasobemproblem («О проблеме носоходок») полагает, что есть две
возможности этого. Или же Моргенштерн был с кратким визитом на Хай-яй-яй в
период с 1893 до 1897 года, или по воле того или иного случая он получил шкурку
Nasobema lyricum («хонатата» у туземцев). Но поскольку ни о
какой тропической экспедиции Моргенштерна не известно, то каким образом он смог
бы получить шкурку? Согласно устному сообщению от госпожи Кейт Циллер, с
которой Моргенштерн был хорошо знаком, он, как было сказано, вернулся домой в
один из вечеров 1894 года, будучи в очень большом волнении и бормоча много раз
«Хай-яй-яй! Хай-яй-яй!» Вскоре после этого, как говорит она, он сочинил
приведённую здесь поэму, которую он также показал её брату. Из этого Блидкуп
делает заключение, что Моргенштерн узнал про Хай-яй-яй по знакомству. Но
действительно ли лежала перед ним хонатата, или он с поэтической
интуицией просто сделал набросок облика животного, остаётся в тумане загадок.
Строки: «Моргенштерна злая лира / Извлекла на свет нас за нос» позволяют
сделать заключение, что он не видел её фактически, а знал только в соответствии
с описанием. Возможно ли также, что он желал скрыть острова с их древними
организмами от жадности европейцев и потому – в виде своего рода камуфляжа –
вплёл эти строки в свою поэму? Мы этого не знаем, так же, как мы не знаем, от
кого Моргенштерн получил свою информацию о Хай-яй-яй и его фауне. Фактически,
единственная возможная фигура в этом направлении – мореход торгового флота,
капитан Альбрехт Йенс Миспотт, который рано умер; Моргенштерн поддерживал с ним
тесную переписку. В 1894 году, после возвращения из долгого и необычного
путешествия, Миспотт умер в Гамбурге в состоянии умственного расстройства.
Возможно, он был тем, кто знал тайну Хай-яй-яй и унёс её с собой в могилу.
Столько удалось установить в ходе исследований Блидкупа.
В своём заслуживающем похвалы исследовании И. И. Шутливицкий занимался той же
самой проблемой. Он приходит почти к тем же выводам, что и Блидкуп, но с тем
отличием, что он считает возможным то, что в период между 1894-м и 1896-м
годами Моргенштерн получил из состояния Миспотта живую хонатату,
которую он содержал в коробке от сигар на протяжении нескольких недель. Но всё
же и эти данные противоречивы. Более того, это мог быть не больше, чем «детёныш
из сумки», так как хонататы дорастают до значительного размера (см.
стр. 55). Всё, что доподлинно известно – это то, что коробка из-под сигар была
весьма глубокой, с этикеткой “Los selectos hediondos de desecho”.
1 nasus Лат. = нос; Bema Греч. = ходить.
2 lyricus Греч. = имеющий отношение к лире.
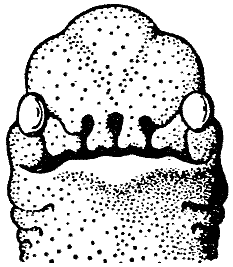
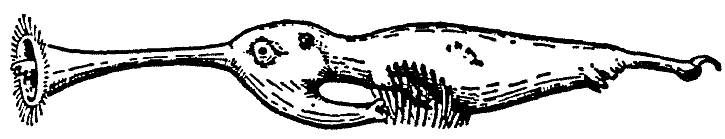
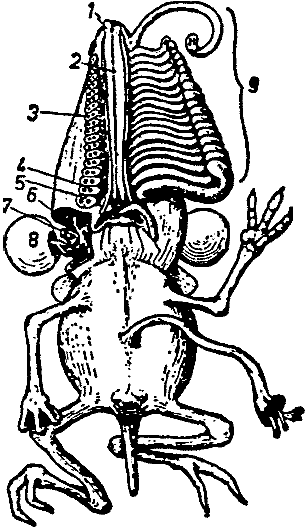
Носоходки, которые признаны особым отрядом млекопитающих, и которые нашли автора монографии о них в лице известного специалиста Бромьена де Бурла, все – как указывает название – отличаются тем, что их носы необычным образом развиты. Нос может присутствовать в единичном или в большем количестве. Последнее состояние уникально среди позвоночных. В настоящее время анатомические исследования (здесь мы опираемся на всесторонние исследования Бромьена де Бурла) показали, что у полиринных видов носовой зачаток расщепляется на ранней эмбриональной стадии таким образом, что зачаточные отдельные ноздри, которые на нём развиваются, имеют голоринный способ дифференциации, то есть, каждый [зачаток] образует рыло законченной формы (см. рис. 1). Вместе с ранней полириналлизацией происходят многочисленные обширные изменения всего плана строения головы. Специальные мускулы, производные лицевой мускулатуры (иннервированные N. Facialis или его ответвлением, N. nasuloambulacralis3, который здесь необычайно сильно развит), принимают участие в формировании мускульного аппарата рыла. Сверх того, в одной группе (Hopsorrhinida4 или мордоскоки) способность рыла развивать усилия ещё более усилена M. longissimus dorsi, который протягивается вперёд по черепу. Носовые пазухи и corpora spongiosa [губчатые тела – В. П.] подвергаются глубокому изменению и увеличению в размере, что сопровождается изменением функции. Так, например, почти у всех более продвинутых форм слёзный проток берёт на себя функции наружного дыхательного прохода. Такие специальные детали будут рассматриваться далее при обсуждении отдельных видов.
|
[Рис. l] Nasobema lyricum. Голова зародыша на ранней стадии развития, которая демонстрирует полиринное состояние (по Штультену, 1949). |
Поскольку у этих носоходящих животных – за исключением рода Archirhinus
(примитивная носоходка) nasarium5 служит средством
передвижения, другие придатки потеряли свои локомоторные функции.
Соответственно, придатки, знимающие более заднее положение, главным образом
более или менее редуцированы, в то время как придатки, занимающие более
переднее положение, видоизменились в хватательные органы, чтобы удерживать
пищу, или в маленькие лапки для ухода за телом. В роде Rhinostentor
(носоходки-трубачи) они участвуют в формировании аппарата для фильтрования
воды.
В то время как парные придатки – менее впечатляющие особенности общей
организации носоходок, хвост у этих животных занимает выдающееся место, и в его
строении развились многообразные и в целом отклоняющиеся типы. Таким образом,
здесь можно найти не только гибкие хвосты и хвосты, действующие подобно лассо,
но также у Sclerorrhina6 (хоботоногов) хвост служит более
примитивным формам для прыжков, а более продвинутым как хватательный орган (см.
стр. 35, 36 и 48).
У большинства носоходок тело покрыто довольно однородным волосяным покровом, в
котором никак нельзя различить остевые волосы и подшёрсток; это должно быть
отнесено не только на счёт климатических условий архипелага, но также, согласно
Бромьену де Бурла, должно расцениваться как примитивный признак. В пользу
такого взгляда говорит также правильность группировки волос. В одном роде,
кроме того, присутствуют прочные роговые чешуи (подобные чешуям у чешуйчатых
муравьедов [панголинов – В. П.]), которые в целом имеют ту же природу, что и
чешуи рептилий. В отдельных случаях окраска шерсти великолепна. Наиболее
известен экстраординарный блеск шкуры, вызванный особой структурой коркового
слоя волоса. Безволосые участки – голова, ноги, хвост, уши, гребешки кожи на
голове и особенно на рыле – в некоторых случаях также ярко окрашены. Несколько
водных видов и самые мелкие роющие виды, которые водятся на песчаном побережье,
совершенно лишены волос; то же самое относится к единственному паразитическому
виду (см. стр. 19).
Способ питания сильно варьирует среди различных семейств, и даже в пределах
одного и того же семейства или рода. Всё же это едва ли покажется удивительным,
если вспомнить, что, помимо одного вида водяной землеройки7,
носоходки – это единственные млекопитающие архипелага, и, таким образом, у них
была возможность захватить все экологические ниши. Большинство животных из
числа носоходок, средний размер которых действительно маленький, едят
насекомых. Но помимо них есть также травоядные – особенно фруктоядные виды – и
один хищный род. В заключение стоит упомянуть особо специализированные формы –
планктоноядные виды, которые живут в пресной воде, и роющие формы, среди
которых есть самые крошечные из известных позвоночных. Крабоядные формы среди
мордоскоков могут быть с лёгкостью выведены из насекомоядных форм. Странный
случай симбиоза будет обсуждаться в разделе, посвящённом систематике (стр. 15,
37).
Особенно примечателен тот факт, что среди носоходок есть один летающий род (с
единственным видом), и что имеются также сидячие и паразитические формы. При
рассмотрении привычек и структурной организации животных, однако, не будет
казаться удивительным, что число видов довольно велико. В этой связи
геологический интерес связан с превосходным исследованием М. О. Джестером и С.
П. Ассфаглом рода Dulcicauda8 (сладкохвосты). Этим авторам
удалось показать, что в течение различных отрезков времени между разными
островами архипелага должны были сохраняться сухопутные мосты, а также удалось
оценить даты их исчезновения (см. также Ludwig 1954). В целом, изучение этого
материала, касающегося Rassen-kreise и их эволюции (Rensch 1947)
выглядит особенно многообещающим даже при том, что во многих областях есть
большие пробелы, которые едва ли будут перекрыты даже при использовании
палеонтологического метода, поскольку подходящие ископаемые остатки заключены в
отложениях, которые залегают глубоко на дне моря.
В общем плане репродуктивный потенциал носоходок невысок, что позволяет сделать
заключение, что показатель смертности также низок. Насколько известно в
настоящее время, рождается лишь один детёныш, без вариантов (мордоскоки с
физиологической полиэмбрионией – исключение). Однако беременные самки
встречаются круглый год. Период беременности – опять-таки за исключением
мордоскоков – долгий и длится в среднем семь месяцев. Среди монорринных форм
молодняк настолько продвинут в развитии при рождении, что не нуждается в
молочном вскармливании. Соответственно, молочные железы этих носоходок
рудиментарны или демонстрируют, как у рода Columnifax9
(Столбоносы), выделение молока, независимое от гормона, стимулирующего лактацию
(см. стр. 15). В полирринных родах, у которых новорождённый находится в весьма
зависимом состоянии, есть единственная пара сосков (главным образом в
подмышечной области). Как правило, эти виды также обладают выводковой сумкой,
которая образуется из складок кожи на горле и поддерживается хрящевыми
полосками, растущими от кадыка.
Ринограденции вряд ли имеют каких-либо врагов. В фауне островов единственные
теплокровные существа, кроме уже упомянутой болотной землеройки (Limnogaloides)
– птицы рода Hypsiboas10 (птицы-мегафоны). Все они размером
с певчих птиц и занимают биотопы, весьма отличающиеся от биотопов Rhinogradentia.
Согласно Бюффону и Сотпримаршу, они происходят от буревестников, а именно от
форм, близких к Hydrobates. Рептилии отсутствуют. Имеется только единственный
примитивный вид земноводных (Urobombinator submersus11),
чьими гигантскими личинками объедаются Хууака-Хатчи на церемониальных пирах.
Медлительные виды Nasobema имеют врагов из их собственного числа в лице хищных
ринограденций рода Tyrannonasus12. Однако этот род
ограничен несколькими островами. Большей частью только океанские птицы, которые
в определённые сезоны гнездятся на некоторых малых островах, иногда добывают
ринограденций. Однако, те самые виды (например, сладкохвосты и столбоносы),
которые живут на берегу, защищены от нападений птиц отчасти ядовитыми
приспособлениями и отчасти тем, что они несъедобны; а мордоскоки настолько
проворны сами по себе, что эти птицы не могут их поймать.
Здесь внимание может быть также обращено на другую особенность хай-яй-яйской
фауны: насекомые демонстрируют большое количество очень примитивных форм. Так,
похожие на тараканов виды представлены многочисленными различающимися по
строению типами, большинство которых может быть помещено среди Blattadae. Кроме
них, есть также небольшое количество более продвинутых насекомых, прежде всего
Hymenoptera, тогда как Lepidoptera отсутствуют вообще. Следовательно, опыление
производится частично представителями Hymenoptera (прежде всего видами рода Pseudobombus,
которые внешне напоминают шмелей, но фактически связаны с ксилокопидами), и
частично ручейниками и тараканами. Муравьёв нет.
Как примечательная особенность должны быть упомянуты шестикрылые насекомые
(Hexaptera из надотряда Hexapteroidea13); они являются потомками
Palaeodictyoptera и имеют наземных личинок. Эти насекомые – главным образом
жители открытых мест; то есть, за исключением нескольких видов, они избегают
густых девственных лесов, которые одевают склоны гор на больших островах. Здесь
также мы можем особо отметить, что на крупных островах есть несколько
эндемичных видов. Эти примитивные формы полностью отсутствуют на более мелких
островах. Вероятно, это следует отнести на счёт того, что более мелкие острова
(например Овмвусса или Савабиси) являются коралловыми островами и,
следовательно, образовались недавно, или же потому что они не предоставляют
достаточной защиты от ветра для плохих летунов, и потому эндемичные виды там
вымерли, когда острова погружались в океан и уменьшались.
В отношении систематического положения носоходок высказаны следующие
соображения:
Как показывает единственный вид, который всё ещё ходит на всех четырёх лапах
(род Archirrhinos = примитивные носоходки), их следует выводить из
примитивных насекомоядных. В этой связи важно присутствие Limnogaloides
на Майрувили; это животное, которое, бесспорно, должно рассматриваться в числе
Insectivora, имеет много особенностей, общих с Archirrhinos, и потому
представляется вполне возможным проследить родословную обоих видов назад к
общему предку.
В отношении остальных видов систематическая классификация носоходок связана
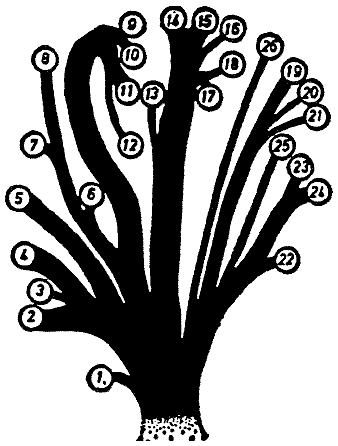
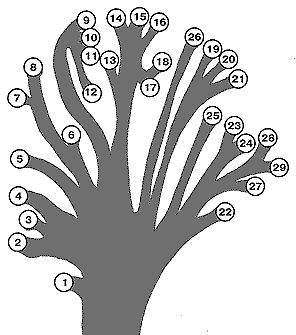
главным образом со степенью развития рыла. «Родословное древо» на рис. 2,
которое показывает систематические подразделения отряда, было предложено
Бромьеном де Бурла (1950). Следуя ему, он различает в качестве основных группы
однорылов, которые ходят на лапах = Monorrhina pedestria (с
единственным видом Archirhinus haeckelii), однорылов, которые ходят
на
носу = Monorrhina nasestria (включающих мягконосых носоходок = Asclerorrhina
и хоботоногов = Sclerorrhina), и многорылых носоходок = Polyrrhina
(включающих коротконосых носоходок = Brachyproata и долгоносых
носоходок = Dolichoproata). Хотя большинство родов может быть без
труда размещено в этой схеме, всё ещё остаются сомнения в отношении кротовых
носоходок = Rhinotalpiformes: неясно, можно ли их объединить в одну
группу с иловыми носоходками = Hypogeonasida14, или же они произошли
от Sclerorrhina
(хоботоногов) и обладают вторично разросшимся nasarium’ом.
|
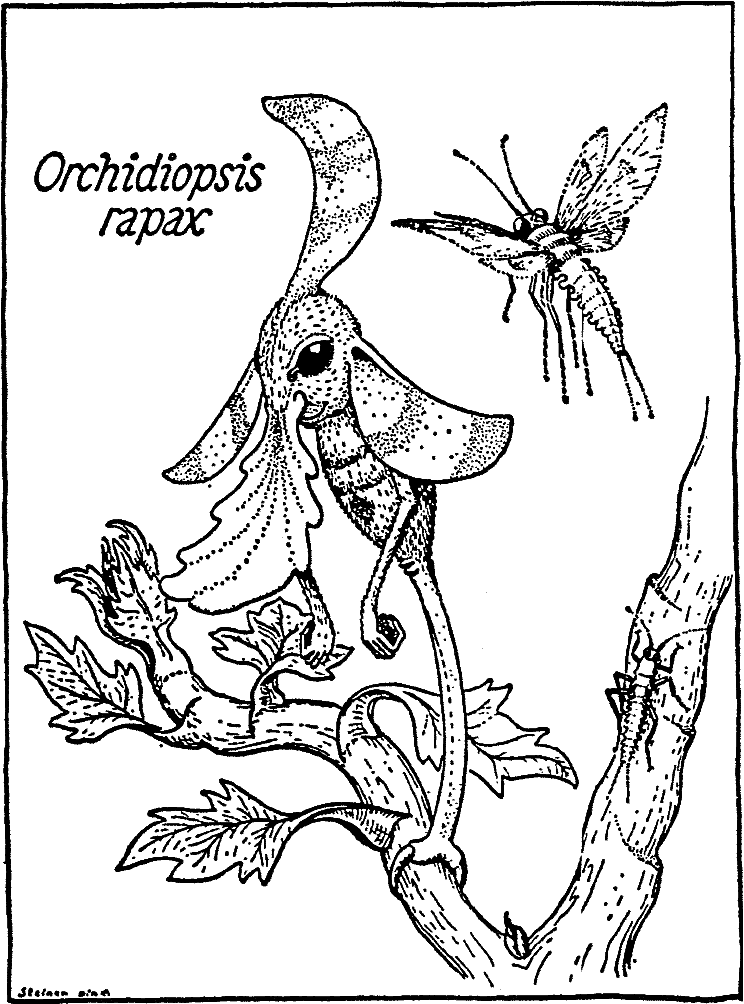
[Рис. 2] Предположительное родословное древо отдельных родов Rhinogradentia (по Бромьену де Бурла, с изменениями, с добавлением данных из Штультена). 1. Archirrhinos; 2. Nasolimaceus; 3. Emunctator; 4. Dulcicauda; 5. Columnifax; 6. Rhinotaenia; 7. Rhinosiphonia; 8. Rhinostentor; 9. Rhinotalpa; 10. Enterorrhinus; 11. Holorrhinus; 12. Remanonasus; 13. Phyllohoppla; 14. Hopsorrhinus; 15. Mercatorhinus; 16. Otopteryx; 17. Orchidiopsis; 18. Liliopsis; 19. Nasobema; 20. Stella; 21. Tyrannonasus; 22. Eledonopsis; 23 Hexanthus; 24. Cephalantus; 25. Mammontops; 26. Rhinochilopus. Толщина ветвей означает относительное количество видов некоторых родов. Здесь Dulcicauda и Dulcidauca, которые в других местах считаются отдельными родами, помещены вместе под названием Dulcicauda. |
Эти 14 семейств в целом содержат 189 видов; всё ещё существует вероятность того, что тот или иной неизвестный вид населяет какую-то изолированную область архипелага. Этого можно ожидать с большой вероятностью, потому что вышеупомянутая группа Rhinotalpiformes в последние годы дала удивительные открытия новых разновидностей. Также должны ожидаться некоторые систематические трудности, когда вопрос касается широко распространённого вида в плане того, когда мы имеем дело с истинными расами, т. е. генетически отличающимися популяциями, а когда просто с местными разновидностями. Пример Mammontops15 (мохнатолицей носоходки), которая первоначально обитала на Шау-вануунде, а позже содержалась Военно-Морской администрацией в зоопарке на экспериментальной станции на Шей-нэй-лакха (Шэйелаха), показывает высочайшую степень, до какой может измениться фенотип. Генетические эксперименты до настоящего времени оканчивались провалом из-за трудности выращивания животных (см. стр. XIX). Только Hopsorrhinus вновь представляет собой исключение. Однако здесь последний эксперимент показал, что различные островные формы являются истинными – хотя и близкородственными – видами. Только когда Hopsorrhinus aureus (золотомордый попрыгун) с Митадины был вытеснен Hopsorrhinus macrohopsus16 (щукомордым попрыгуном) с Хай-дуд-дай-фи, появилось лишь немногочисленное [гибридное] потомство с ограниченной способностью к воспроизводству. Hopsorrhinus mercator17 (= Mercatorrhinus galactophilus18, мордоскок Хили) также оказался подходящим для генетических экспериментов; обладая периодом беременности лишь в восемнадцать дней, он обычно приносит восемь голов молодняка одного пола и очень легко выкармливается имеющимся в продаже сгущённым молоком для детей (см. стр. 37).
3 nasulus Лат. = носик; ambulare Лат. = идти.
4 hopsos Греч. = скачок или прыжок (слово, встречающееся только у Златоуста из Массилии, которое, несомненно, должно отслеживаться до западногерманского корня); rhis, rhinos Греч. = нос.
5 Бромьен де Бурла определяет как nasarium всю совокупность носовых ambulacrum, независимо от происхождения его компонентов. Следовательно “nasarium” – это скорее функциональное, нежели морфологическое понятие. Поскольку этот термин закрепился в литературе и его использование позволяет избежать необходимости использования длинных описаний, то он будет использоваться в следующих частях книги,
6 skleros Греч. = твёрдый.
7 Limnogaloides mairuviliensis B. d. B. (майрувилийская болотная землеройка) – примитивное насекомоядное. То, что она принадлежит к настоящим землеройкам, недавно было оспорено. Также в связи с этим обстоятельством вышло из употребления более раннее название Limnosorex mairuviliensis. Зубная формула, хорошо развитая скуловая дуга, необычно маленький передний мозг и присутствие межпозвонковых мускулов по всей длине хвоста расцениваются как особенно примитивные признаки.
8 dulcis Лат. = сладкий; cauda Лат. = хвост.
9 columnar Лат. = столб; fax Лат. = изготовление.
10 hypsiboas Греч. = Громкий крикун (дорийская форма аттического hypsiboes).
11 ura Греч. = хвост; bombina Лат. = жаба; submersus Лат. = погруженный.
12 тиран-нос.
13 шестикрылые насекомые.
14 hypo Греч. = под; gea Греч. = земля.
15 mamont, Рус., заимствовано из палеосибирских языков = мамонт. Написание “mammonta” по сути ошибочно, но правильно согласно правилам номенклатуры, -ops Греч. = лицо. На рисунке (рис. XV) использовалось написание, принятое в 1952-56 годах.
16 makros Греч. = большой; hopsos см. сноску 4.
17 mercator Лат. = торговец.
18 galactophilus Греч. = любящий молоко
ПОДОТРЯД: Monorrhina (Однорылы),
СЕКЦИЯ: Pedestria (Стопоходящие),
ТРИБА: Archirrhiniformes (примитивные типы носоходок),
СЕМЕЙСТВО: Archirrhinidae (Примитивные носоходки в широком смысле),
1 РОД: Archirrhinos (собственно примитивные носоходки),
1 вид.
 Archirrhnos
haeckelii (архиррин Хекеля) – единственный ныне живущий представитель
примитивных носоходок*. Он всё ещё ходит на четырёх лапах, как остальные
млекопитающие, и всё ещё не имеет высокоспециализированного nasarium’а.
Таким образом, его рыло в целом не приспособлено для того, чтобы работать в
качестве локомоторного органа и просто служит животному как опора, когда оно
поедает схваченную добычу (см. табл. II, животное на заднем плане). Повадки
примитивных носоходок во многих отношениях напоминают повадки землеройки; если
днём она спит в простом гнезде под корнями, то в сумерках она отправляется на
поиски корма. Тогда и можно увидеть, как существо размером с мышь с крупной
головой и огромным рылом снуёт туда-сюда и гоняется за огромными тараканами,
которые часто собираются в большом количестве вокруг похожих на бананы ягод,
которые падают к подножию стволов дерева висолека. Поймав насекомое,
эта носоходка стремительно опрокидывается на морду, края который быстро
расширяются в стороны, образуя широкую опорную поверхность. Вязкая носовая
слизь обеспечивает прочное соединение с землёй. И теперь жадный зверь очень
быстро подносит пищу к своему рту всеми четырьмя лапами. Пируя, эти существа
выдают своё присутствие громким щебечущим писком, слышным издалека. Как только
с пищей покончено, они так же быстро выходят из своей стойки на голове, края
рыла снова заворачиваются внутрь и преследование добычи возобновляется. Пока
ещё мало что известно о размножении этих животных, поскольку они водятся только
в недоступных горных лесах Хай-дуд-дай-фи.
Archirrhnos
haeckelii (архиррин Хекеля) – единственный ныне живущий представитель
примитивных носоходок*. Он всё ещё ходит на четырёх лапах, как остальные
млекопитающие, и всё ещё не имеет высокоспециализированного nasarium’а.
Таким образом, его рыло в целом не приспособлено для того, чтобы работать в
качестве локомоторного органа и просто служит животному как опора, когда оно
поедает схваченную добычу (см. табл. II, животное на заднем плане). Повадки
примитивных носоходок во многих отношениях напоминают повадки землеройки; если
днём она спит в простом гнезде под корнями, то в сумерках она отправляется на
поиски корма. Тогда и можно увидеть, как существо размером с мышь с крупной
головой и огромным рылом снуёт туда-сюда и гоняется за огромными тараканами,
которые часто собираются в большом количестве вокруг похожих на бананы ягод,
которые падают к подножию стволов дерева висолека. Поймав насекомое,
эта носоходка стремительно опрокидывается на морду, края который быстро
расширяются в стороны, образуя широкую опорную поверхность. Вязкая носовая
слизь обеспечивает прочное соединение с землёй. И теперь жадный зверь очень
быстро подносит пищу к своему рту всеми четырьмя лапами. Пируя, эти существа
выдают своё присутствие громким щебечущим писком, слышным издалека. Как только
с пищей покончено, они так же быстро выходят из своей стойки на голове, края
рыла снова заворачиваются внутрь и преследование добычи возобновляется. Пока
ещё мало что известно о размножении этих животных, поскольку они водятся только
в недоступных горных лесах Хай-дуд-дай-фи.
|
ТАБЛИЦА II |
* В противоположность всем остальным носоходкам, известны ископаемые остатки Archirrhinos или близкородственных форм. На острове Оуса-доуса (Ауса-дауса), который занимает исключительное положение в архипелаге в тектоническом плане, есть раннетретичные отложения, которые, возможно, могли бы также оцениваться как верхнемеловые. При их раскопках были найдены зубы архиррина, который, очевидно, был размером с домашнюю кошку.
ПОДОТРЯД: Monorrhina (Однорылы),
СЕКЦИЯ: Nasestria (Мордолазы),
ТРИБА: Asclerorrhina (Мягконосые носоходки),
ПОДТРИБА: Epigeonasida19 (Наземные носоходки),
СЕМЕЙСТВО: Nasolimacidae20 (Улитконосые носоходки),
РОД: Nasolimaceus (Слизненосы),
4 вида,*
РОД: Rhinolimaceus (Сахарные мышки),
14 видов.
* Здесь, как и в последующих секциях, назван и описан только один особенно характерный представитель. Для более подробных сведений даётся ссылка на работу Бромьена де Бурла (1951) и на несколько более короткую монографию Дж. Д. Битбрейна (1950).
Одноносые Nasestria, включающие трибу Asclerorrhina, близкородственны
архирринам. Все они без исключения – животные, у которых модификации носовой
области, произошедшие в связи с изменившимся способом передвижения, остались
в узких границах. Существенные особенности включают расширение рыла и тех частей
черепа, которые служат для его поддержания. Однако в качестве новых событий
должны расцениваться следующие особенности строения: многократное разделение
мышц морды и носовых пазух, вследствие чего образовалась система воздушных
камер, которые сообщаются друг с другом, но которые могут быть изолированы
друг
от друга специальными мускулами; далее отмеченное развитие corpora
spongiosa, которые придают рылу упругость, существенно важную для его
функций. У большинства видов оно может произвольно менять форму. Кроме того,
мускулы тех частей морды, которые прилегают к рылу, частично протягиваются
по самому рылу и значительно дифференцируются, потому рыло (которое, согласно
Битбрейну, уже является истинным nasarium’ом) приобретает значительную
гибкость. Ещё одна особенность – значительное расширение секреторного эпителия;
продуцирование им слизи, регулируемое произвольно, важно для передвижения
животных и для их надёжного прикрепления на месте.
Как уже было сказано в разделе общих сведений, конечности редуцированы или
видоизменены. Задние ноги рудиментарны (но никогда не исчезают полностью) и
практически лишены функций. Передние конечности служат и для того, чтобы
схватывать корм, и для ухода за собой.
Nasolimaceus palustris21 (слизненос Фаделахи) будет описан
как типичный представитель улитконосых носоходок. Он также наиболее
близкородствен Archirrhinos. Это существо размером приблизительно с
мышь, с яркой золотисто-коричневой шкуркой, водится на Майрувили и там населяет
илистые берега Фаделахи. У него короткое, но широкое рыло, вентральная
поверхность которого занимает переднее положение и видоизменена в ползательную
ногу и функционирует во многом сходным образом с ногой Helix
[виноградной улитки – В. П.], с различием, однако, в том, что локомоторные
волны сокращений следуют друг за другом быстрее и направление их движения
обратимо. Скорость передвижения больше, чем можно ожидать от такого механизма
передвижения. Когда животное спасается бегством или кого-то преследует, оно покрывает
примерно 30 – 40 футов в минуту. В таких обстоятельствах кажется, будто оно
скользит с почти призрачной скоростью по гладкой влажной грязи; и точный способ
передвижения нельзя различить простым глазом, а можно зафиксировать только
высокоскоростной фотографией.
|
ТАБЛИЦА III |
Впечатляющая киносъёмка этого явления была недавно сделана Ф. Хайдеритчем из
Научной и Медицинской Кинематографической Компании, Блэк Гоатс. Улитки
эндемичного рода (Ankelella) являются единственным источником пищи для
видов Nasolimaceus; только Rhimolimaceus fodiens22,
известный морякам морской базы как «Счастливчик Пьер», выкапывает земляных
червей (которые частично принадлежат к тем же самым родам, что и эндемичные
земляные черви Новой Зеландии!). Прежде, чем потерять свои «молочные зубы»,
молодняк кормится главным образом личинками насекомых (комаров-звонцов),
поскольку он ещё не может справиться с твёрдыми раковинами улиток.
Спаривание, во время которого самец иногда издаёт звуки, как будто он
сморкается, происходит главным образом ночью. В это время самец описывает
тесные круги вокруг самки, которая также вращается. Время от времени она также
издаёт нежное «Гм-гм». Сцена для этих любовных игр – обычно большой плоский
камень, который покрыт тонким слизистым слоем диатомовых водорослей, и через
который иногда плещется вода. Сам по себе этот процесс напоминает гротескный
танец человеческой пары на коньках. Акт спаривания длится лишь несколько секунд,
и затем партнёры оставляют один другого, очень быстро расходясь в
противоположных направлениях со звуком лёгкого шмыганья носом. После
беременности, которая длится двадцать шесть (?) месяцев, самка приносит
единственного отпрыска, который уже похож на родителей во всех деталях, и
который без дальнейшей зависимости от них сразу начинает вести самостоятельную
жизнь.
Животные не охраняют постоянную территорию; они ведут строго уединённую жизнь и
терпимы по отношению друг к другу. В целом, поскольку улитконосые носоходки
приспособлены к илистым берегам пресных водоёмов, они привязаны к источникам
воды, где родились, хотя не умеют ни плавать, ни пересекать с большим желанием
протяжённые отрезки иного субстрата, нежели илистый или песчаный. Несмотря на это,
время от времени неполовозрелые экземпляры – чьи конечности на этой стадии
относительно сильнее развиты – медленно бродят по суше в поисках других
водоёмов. Животные не могут выживать в морской воде – как и многие другие
носоходки – и в свете этого ясно, что на различных островах существуют
многочисленные специфические разновидности.
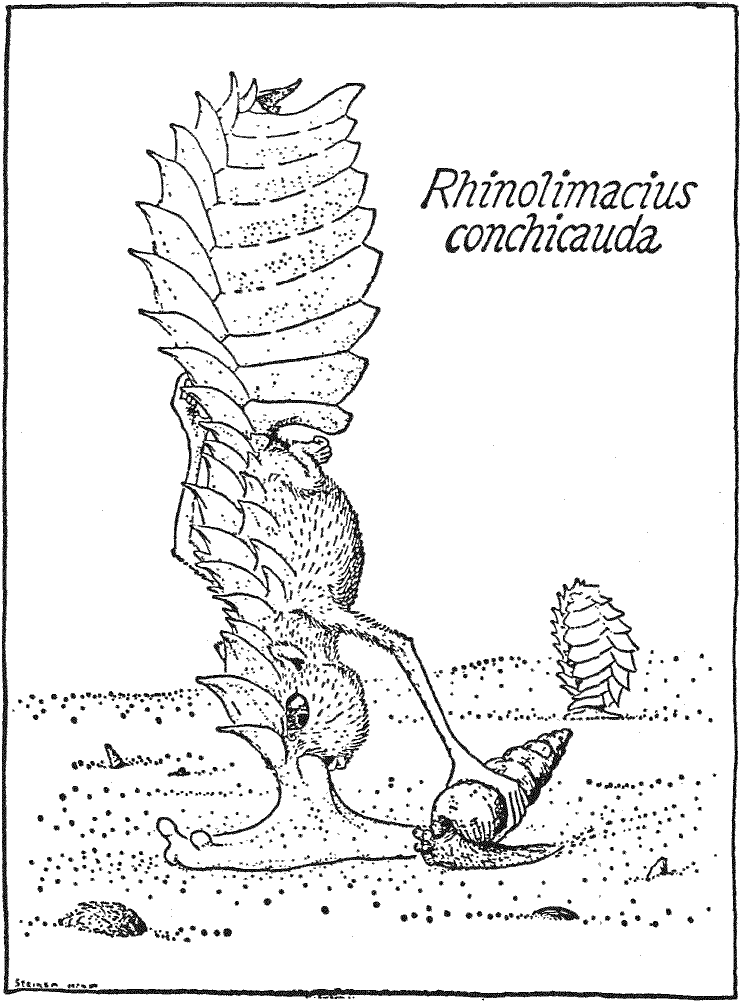
Близким родственником Nasolimaceus palustris является Nasolimaceus
conchicauda23 (бронехвостый слизненос). Это животное, как
говорит его название, обладающее бронированным хвостом, живёт на небольшом
вулканическом островке Ииза-зофа (Изазофа), на болоте с тем же самым названием.
Подгибая свой хвост вентрально под себя, животное может прятаться в нём, словно
в пляжном шезлонге. Важно то, что на Ииза-зофа обитает птица-мегафон, которая
представляет собой угрозу для носоходок. Это нелетающая птица Hypsiboas
fritschii (птица-мегафон Фритча), ловкий бегун и пловец размером с дрозда,
который питается практически любыми животными, которых может поймать и одолеть.
Сохранившиеся улитконосые носоходки хорошо защищены от врагов тем, что в
основании хвоста у них есть железа, которая выделяет сладкий секрет (отсюда
название «сахарные мышки»). Жидкость привлекает очень мелкий вид рода Pseudobombus,
который всегда находится в готовности жалить. «Сахарные мышки» посещаются
главным образом роями этих насекомых и, следовательно, защищены*.
*Согласно исследованиям Ширина Тафарруи, секрет железы содержит лишь небольшое количество глюкозы, но в дополнение к ней сладкое вещество, природа которого всё ещё полностью не определялась; структурно оно не является близким ни к дульцину, ни к сахарину, и в чистой форме обладает стимулирующим действием примерно в 200 раз сильнее, чем сахарин. Примечательно, что это вещество обладает приблизительно равной сладостью и для людей, и для насекомого.
19 Epi Греч. = на; gea см. сноску 14.
20 limax Лат. = улитка.
21 palustris Лат. = обитающий на болоте.
22 fodiens Лат. = роющий.
23 conche Греч. = раковина или щит; cauda Лат. см. сноску 8.
СЕМЕЙСТВО: Rhinocolumnidae
(Столбоносы в широком смысле),
РОД: Emunctator (Нюхачи)
1 вид,
РОД: Dulcicauda (Сладкохвосты),
19 видов,
РОД: Dulcidauca24 (Сахарнохвосты),
1 вид,
РОД: Columnifax (Собственно столбоносы)
11 видов.
Положение сахарных мышек в классификации всё ещё является предметом
дискуссии: хотя Спасман и Штультен ещё в 1947 году занимали позицию, согласно
которой они должны быть помещены в специальную секцию (Sedentaria),
противоположную Peripatetica, сегодня люди, главным образом следуя взглядам
Бромьена де Бурла, включают их в подтрибу Epigeonasida. Основная причина для
этого – открытие Emunctator sorbens (сопящего нюхача), который
занимает место, промежуточное между бродячими сахарными мышками и сидячими
столбоносами.
|
ТАБЛИЦА IV |
 С
другой стороны, нельзя исключать и того, что столбоносы являются
полифилетической группой. Бюффон (1954) только недавно еще раз указал, что
существуют радикальные различия между Emunctator и Dulcicauda
с одной стороны, и Columnifax на другой: 1. иннервация гипоринальной
мускулатуры в этих двух группах существенно различается; и 2. вещества,
найденные в основании колонны (опоры, на которой стоят Dulcicauda и Columnifax)
в некоторой части резко различны. Прикрепительные нити Emunctator и
опорная площадка Dulcicauda вместе содержат так называемый
«эмунктатор-муцин», который включает в себя пентозосодержащую слизистую серную
кислоту, которая отсутствует у Columnifax. С другой стороны, опора Columnifax
содержит псевдоринокератин, который не встречается у других родов.
С
другой стороны, нельзя исключать и того, что столбоносы являются
полифилетической группой. Бюффон (1954) только недавно еще раз указал, что
существуют радикальные различия между Emunctator и Dulcicauda
с одной стороны, и Columnifax на другой: 1. иннервация гипоринальной
мускулатуры в этих двух группах существенно различается; и 2. вещества,
найденные в основании колонны (опоры, на которой стоят Dulcicauda и Columnifax)
в некоторой части резко различны. Прикрепительные нити Emunctator и
опорная площадка Dulcicauda вместе содержат так называемый
«эмунктатор-муцин», который включает в себя пентозосодержащую слизистую серную
кислоту, которая отсутствует у Columnifax. С другой стороны, опора Columnifax
содержит псевдоринокератин, который не встречается у других родов.
Сопящий нюхач, Emunctator sorbens (см. таблицу IV) – это животное
размером с маленькую крысу. Он живёт на Хай-дуд-дай-фи вдоль берегов медленно
текущих ручьёв. Там он цепляется за стебли растений, которые поднимаются над
поверхностью воды. Его манера добывания пищи наиболее специфическая; с
удлинённого рыла свисают длинные, тонкие липкие нити, которые свисают в воду и
к которым приклеиваются мелкие водные животные. Добыча (главным образом
веслоногие рачки и личинки насекомых, а также равноногие рачки и бокоплавы, и
реже мелкая рыба) частью втягивается через хоаны, вместе со слизистыми нитями,
а частью слизывается с рыла чрезвычайно длинным языком.
Медлительные, неуклюжие животные обладают в качестве средства защиты длинным,
очень подвижным хвостом с ядовитой железой на его кончике; токсин выделяется
внутрь полого когтя (образовавшегося из видоизменённых волос). Поскольку Emunctator
живёт главным образом небольшими группами, мелкие существа могут защищать друг
друга, совместно раскачивая хвостами.
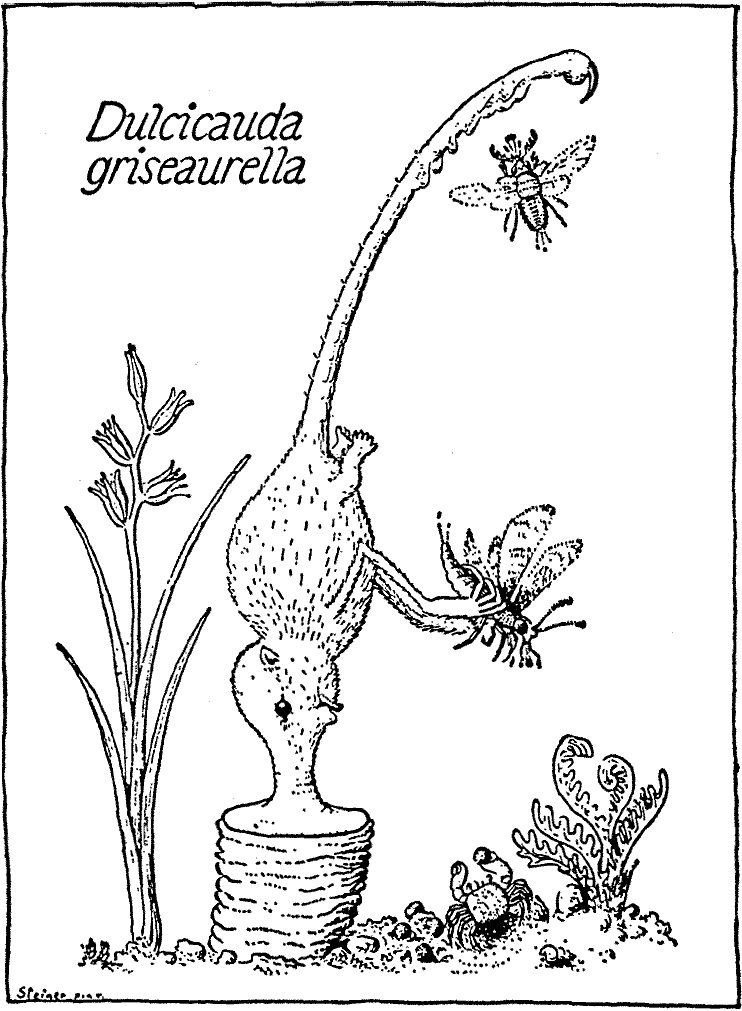
В качестве типичного представителя рода Dulcicauda, будет описан вид D.
griseaurella (серовато-золотой сладкохвост) (здесь и далее будут использоваться
названия, данные Бромьеном де Бурла, и потому имя автора обычно не приводится).
Этот вид, вместе с D. aromaturus (пахучим сладкохвостом) водится на
Митадине, D. griseaurella на восточной, а D. aromaturus на
западной половине острова.
|
|
Странная особенность этих животных состоит в том, что они являются истинными сидячими формами, которые прочно стоят на своих мордах и обычно никогда не оставляют место прикрепления, которое они выбрали, будучи молодыми. Таким образом, они остаются стоять на рыле, которое выделяет красновато-жёлтый секрет, со временем поднимающий животных (голова+тело примерно 3 дюйма длиной, хвост примерно 4 дюйма) на явно выраженной столбообразной опоре, известной как основа или sella (см. таблицу V). В особенности вблизи кончика, который несёт ядовитый коготь, хвост снабжён большим количеством кожных желёз, которые обильно выделяют липкое вещество. Насекомые, привлечённые к хвосту распространяющимся ароматом, садятся, приклеиваются к нему, и их отрывают передние лапы, чтобы поднести ко рту. Если же садящиеся насекомые большей частью мелкие, их не собирают с хвоста по одному: время от времени животное протягивает хвост через рот и облизывает их с него.
|
|
|
|
|
Dulcicauda (с сайта Токивы Такеши) |
Dulcicauda живёт колониями на каменистых склонах около берега. Эти
колонии, как правило, связаны с маленьким земляным крабом (Chestochele25
marmorata) который питается остатками пищи носоходок и уносит их
помёт.
В брачный период самцы слезают со своих пьедесталов и приближаются к самкам,
скользя и держа впереди свои передние конечности; когда спаривание завершено,
они возвращаются на свои пьедесталы. Они могут освободиться от основы своей
колонны путём частичного растворения её верхних слоёв с помощью ферментов,
выделяемых железами Пусдивы на discus nasalis (то же самое справедливо
в отношении растворения прикрепительной слизи у Archirrhinos).
Род Columnifax в общих чертах отличается сокращением в длину
хватательного хвоста. Из-за этого животные неспособны самостоятельно ловить
добычу. Только молодые экземпляры, не старше трёх месяцев, всё ещё имеют
относительно длинный, железистый хвост и всё ещё питаются таким же способом,
как Dulcicauda или Dulcidauca (для Dulcidauca
характерна утрата задних конечностей). У старших особей устанавливается очень
примечательный симбиоз с мордоскоками: каждый из одиннадцати видов Columnifaxсвязан с одним из одиннадцати подвидов Hopsorrhinos mercator
(мордоскока Хили). Два партнера тесно зависят друг от друга и заботятся о
питании друг друга: в прибрежной зоне, где они оба живут, преобладающая добыча Hopsorrhinus
mercator – это мелкие раки-отшельники, которых, однако, он не может
глотать из-за модификаций его рта (см. стр. 37). Затем он подаёт их столбоносу,
предварительно подавив защитные реакции последнего с помощью определённых
звуков и жестов. (Columnifax защищается, выбрызгивая секрет анальных
запаховых желёз, а благодаря своей очень подвижной морде способен
поворачиваться приблизительно на 180° вокруг продольной оси). Вслед за этим Columnifax
разрешает мордоскоку сосать молоко в рамках описанного симбиоза; молоко
производится особями обоих полов старше трёх месяцев, независимо от любой
связанной с полом функции (см. таблицу VII).
24 Dulcidauca: анаграмма от Dulcicauda, см. сноску 8.
25 cheston Греч. = экскременты (у Эвфемия Теретея из Александрии); chele Греч. = подвижный коготь, клешни краба.
ПОДТРИБА: Hypogeonasida (Иловые
носоходки),
СЕМЕЙСТВО: Rhinosiphonidae (Сифонные носоходки в широком смысле),
РОД: Rhinotaenia (Ленторылы),
2 вида
РОД: Rhinosiphonia (Собственно сифонные носоходки),
3 вида
СЕМЕЙСТВО: Rhinostentoridae (Носоходки-трубачи в широком смысле),
РОД: Rhinostentor (Собственно носоходки-трубачи),
3 вида
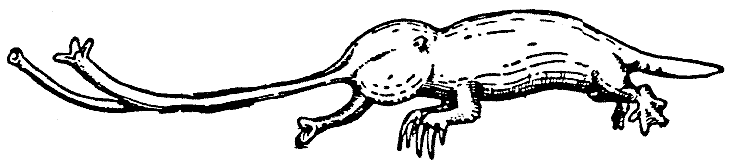
Hypogeonasida сами по себе также представляют относительно изолированную группу. Все они – мелкие невзрачные животные, которые исходно вели подземный образ жизни, который наиболее типично проявляется у рода Rhinotaenia. Далее как характерный представитель будет описан вид Rhinotaenia asymmetrica (трубконосый ленторыл).
|
[Рис. 3] Rhinotaenia asymmetrica. (ориг.) |
Это животное живет в иле нескольких маленьких озёр, а также в
супралиторальной зоне нескольких медленно текущих ручьёв. Там оно питается
преимущественно олигохетами и личинками насекомых, которых выкапывает и
заглатывает с помощью своего длинного хоботообразного рта. Занимаясь этим, Rhinotaenia
ежедневно прокапывает для себя тоннель длиной один или два ярда на глубине
около фута. Дыхание представляется возможным благодаря длинному носу, который
вытянут в виде сифона длиной до 16 дюймов – то есть, приблизительно в четыре
раза больше общей длины головы и туловища животного (см. рис. 3). Асимметрия
рыла – левая ноздря со своей конечной розеткой служит для вдоха, а правая для
выдоха – представляет собой прекрасный механизм снабжения воздухом, несмотря на
длину воздушных трубок.
В отношении спаривания и размножения не известно ничего. Беременные самки и
очень мелкий молодняк встречаются на протяжении всего года.
Бейлигу удалось выделить из отделённых рыл Rhinotaenia муцин, который
идентичен таковому у Emunctator. С позиций морфологии может быть
выдвинуто достаточно много аргументов в пользу представления о том, что иловые
носоходки происходят от Emunctator-подобных предков (см. Бромьена де
Бурла, 1952, а также Джеркера и Селиаццини, 1953).
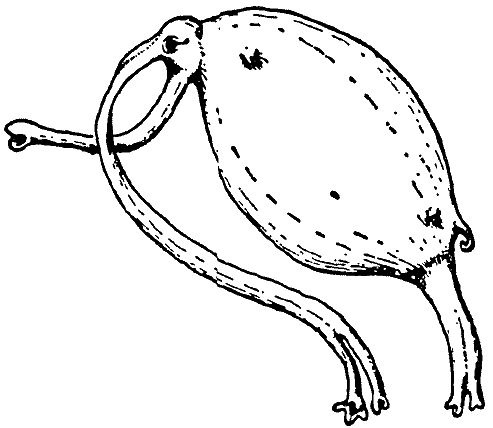
Род Rhinosiphonia отличается от Rhinotaenia прежде всего
более тонким строением рыла, но не показывает никаких дальнейших отличий от Rhinotaenia,
которые здесь нужно комментировать. Вместо этого мы можем обсудить здесь один
вид Rhinotaenia, который уникален своим специфическим паразитическим
образом жизни.
|
[Рис. 4] Rhinotaenia tridacnae, показанная
в возрасте половой зрелости. |
Rhinotaenia tridacnae (раковинный ленторыл; см. рис. 4) обитает в
приливно-отливной зоне повсюду на архипелаге. Молодые животные и самцы живут в
иле, который откладывается в спокойных участках лагун или накапливается в
небольших расселинах между коралловыми глыбами. Даже в большей степени, чем у
остальных Rhinogradentia, и особенно у иловых носоходок, гомойотермия
у Rhinotaenia tridacnae развита лишь в зачаточной степени. С этим
связан тот факт, что R. tridacnae может произвольным образом в течение
значительного времени выдерживать более или менее полное прерывание
окислительного метаболизма. Конечно, эти существа обитают в верхней части
приливно-отливной полосы, где грязь скрывается под водой лишь от четверти часа
до получаса подряд. Тем не менее, эти животные могут сохранять
жизнеспособность, отрезанные от доступа к атмосферному воздуху в течение целых
трёх часов. Тогда у них наступает своего рода припадок и они – голокожие, как
есть – приобретают синий цвет по всему телу и восстанавливают желтоватый
телесный цвет, как только вдыхают воздух ещё раз.
Во время высокого прилива зрелые самки R. tridacnae проникают в
открытые раковины рода Tridacna (гигантские двустворки) и стремительно
внедряются между раковиной и мантией. Там они вскоре вызывают вздутие мантии
размером с кулак или голову ребёнка, лишь часть которого, однако, выстлана
перламутром; в период низкого стояния воды этот нарост заполняется воздухом и
выступает, словно грыжа, в жаберную полость. Своим сосущим хоботком паразит
забирает у хозяина и гемолимфу, и часть половых продуктов. Спаривание с самцами
Rhinotaenia происходит во время ночного прилива. Очевидно, молодняк
также рождается ночью во время прилива.
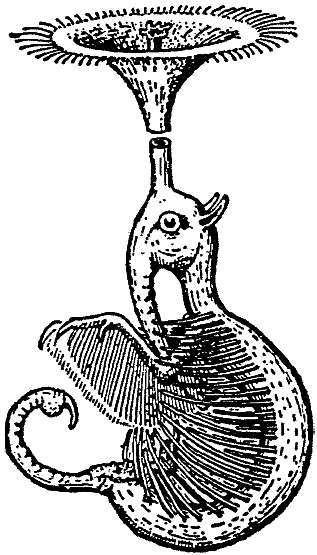
Носоходки-трубачи тесно связаны с Rhinotaenia, но приспособились к
погружённой жизни и потому подверглись некоторым модификациям*, которые
наиболее типично выражены у Rhinostentor submersus26
(рачковой носоходки-трубача).
* Здесь не делается попытки объяснения в смысле Boker, а приводится лишь запись наблюдений.
|
[Рис. 5] Rhinostentor submersus. (ориг.) |
Rhinostentor submersus живёт в различных озёрах внутри кратеров и в
солоноватых лагунах архипелага, и кормится там планктоном, который образован
преимущественно листоногими рачками Branchipusiops lacustris, который
большей частью встречается здесь в изобилии и только изредка его численность
сокращается из-за вездесущих Cladocera или Rotatoria. Здесь Rhinostentor
парит в толще воды на глубине 8 – 20 дюймов от носового сифона, который устроен
в общих чертах сходно с Rhinotaenia, но, сообразно с водным образом
жизни, демонстрирует расширение носовой розетки. Розетка разрастается в виде
воронки вокруг выводящего носового прохода, тогда как вводящий носовой проход
приподнят над ним, образуя маленькую вторичную розетку. Воронковидная или
трубообразная носовая розетка (см. рис. 5) бахромчатая, с водоотталкивающими
волосками, а по краям (видоизменённые восковые железы) выделяет
водоотталкивающую воскоподобную смазку, поэтому животное зависит от этого
раструба, который образует поплавок. Вдоль боков безволосого в иных отношениях
тела тянется ряд жётких, толстых щетинок, которые с вентральной стороны
образуют своего рода желобок, в пределах которого передние конечности, также
окаймлённые гребешками жёстких щетинок, выполняют гребущие движения. Как единое
целое всё приспособление работает наподобие фильтрующего аппарата Daphina
(= водяная блоха [так в оригинале, правильнее пишется “Daphnia” – В.
П.]), а своим ротовым хоботком Rhinostentor вытаскивает из этого
фильтровального приспособления планктонные организмы, которые попались туда,
занесённые током воды.
У Rhinostentor spumonasus (пенной носоходки-трубача) преобладают те же
самые отношения, за исключением того, что существо вместо своей носовой трубки
висит на пузырчатой массе, которую выпускает из последней; в эту массу
носоходка забирается для сна, спаривания, а также при первых признаках
опасности.
|
[Рис. 6] Rhinostentor foetidus. (Ориг.) |
Посетитель архипелага получит весьма большое впечатление от плавающей пены Rhinostentor
foetidus (вонючей носоходки-трубача; рис. 6), не только потому что они
часто плавают большими скоплениями в самых разных источниках пресной воды, но и
потому, что время от времени их невыносимый запах отравляет пребывание во
многих местах, которые без этого можно было бы считать идиллическими. R.
foetidus живёт скоплениями на плотах, которые строят эти животные. Его
вентральный собирающий аппарат редуцирован до пары маленьких гребешков, с
помощью которых существо, ползая по постоянным дорожкам на своём плоту и
подтягиваясь на частично редуцированной носовой трубке, собирает свой корм.
Этот корм состоит из личинок грибных комариков рода Spumalyophilus,
которые кормятся мицелием, который разрастается по плавающей слизи. Гриб,
который не был точно идентифицирован, хотя представляет собой один из видов
Eumycetis, питается отмирающими сине-зелёными водорослями, которые плавают в
толще плавающей слизи. Так образуется симбиотическая система:
Rhinostentor посредством питательных веществ, содержащихся в слизи и
выделяемых с мочой и фекалиями, снабжает водоросли подходящим субстратом,
который окружён грибковыми нитями. Водоросли усваивают эту пищу и затем
частично высасываются или перерабатываются грибком*. Гриб поедается личинками
грибного комарика. Часть этих личинок составляет пищу носоходки-трубача.



Rhinostentor с сайта Токивы Такеши
Интересно, что слизистые плоты Rhinostentor spumonasus и Rh. foetidus населяет целый ряд других животных: птица-мегафон Фритча регулярно использует их как место для гнездования. Шестикрылая стрекоза Hexapteryx handlirschii откладывает на них яйца, а выводящиеся личинки поедают личинок грибного комарика. Целый ряд ногохвосток (Colembola) среди множества других существ живёт в богатых кислородом воздушных пузырьках на верхней стороне плавучих масс.
* Является ли это видоизменённым лишайниковым симбиозом, пока неясно.
26 см. сноску 7.
ПОДТРИБА: Georrhinidae (Роющие
носоходки),
СЕМЕЙСТВО: Rhinotalpidae (Кротовидные носоходки),
РОД: Rhinotalpa (Кротовые носоходки),
4 вида,
РОД: Enterorrhinus (Кишечные носоходки),
5 видов,
СЕМЕЙСТВО: Holorrhinidae (Тольконосые),
РОД: Holorrhinus (Тольконосы),
18 видов,
РОД: Remanonasus27 (Карликовые носоходки),
1 вид
|
[Рис. 7] Rhinotalpa phallonasus. (Ориг.) |
Хотя роющие носоходки обладают совершенно примитивно выглядящим nasarium’ом,
который несомненный произошёл от nasarium’а предковых носоходок,
однако они примечательны тем, что в крайних случаях рыло значительно
преобладает над остальным телом. В этом отношении – а не только в сопутствующей
редукции всей их структурной организации – они уникальны не только среди
носоходок, но и среди всех позвоночных.
Отношения, ближайшие к тем, которые первоначально преобладали, демонстрирует
род Rhinotalpa (кротовые носоходки), и из них мы выбираем в качестве
типичного представителя Rhinotalpa phallonasus, фаллоносую носоходку,
которая является обитателем Майрувили (см. рис. 7). Это существо размером с
мышь, имеющее в большей или меньшей степени повадки крота; то есть, она живёт в
тоннелях, которые выкопала во влажной земле, и кормится почвенными насекомыми и
земляными червями. И передние, и задние конечности сильно редуцированы. Кисти и
ступни очень хорошо сохранились и вместе с крупными когтями служат и как
устройство для толкания, и как якорь в туннеле. Работа по рытью, однако,
выполняется рылом, которое снабжено большими эректильными органами, и в которое
протягиваются обширные воздушные мешки (развившиеся из носовых полостей);
последние также принимают участие в расширении рыла. Не только вокруг самой
толстой части рыла, но также и вокруг затылка и вентральнее нижних челюстей
расположены пояски сильных, направленных назад щетинок. Оба пояска могут
растопыриваться и задействованы в двигательных актах, которые делятся на
следующие стадии:
l. Поясок щетинок на горле растопыривается, то же самое делают когти конечностей;
2. Рыло раздувается, воздух набирается через рот и одновременно закрывается голосовая щель (к этому добавляется способность замыкать ноздри);
3. Носовой поясок растопырен; воздух выходит из рыла и животное продвигается вперед сокращением мышцы M. retractor nasarii, после чего может повториться стадия l.
Носовые эректильные органы вовлекаются в действие только в очень твёрдой
земле, и у них есть особая функция напряжения и расширения переднего конца
морды. Но вообще животное старается рыть как можно меньше и использует ходы,
которые уже есть под рукой; в результате тоннели, которые они прокопали, имеют
очень твёрдые стенки. По ним животное скользит с замечательной скоростью,
одновременно собирая гребешком, образовавшимся из chaetae submentales28,
земляных червей, найденных по пути; затем оно вытаскивает червей из гребешка
ротовым хоботком. Papillae basonasales [нижне-носовые сосочки – В. П.]
играют важную часть в распознавании добычи. Их иннервация, кроме чувствительных
лицевых нервов, производится также теми нервами, которые у близкородственных
форм пронизывают якобсонов орган, и потому сосочки являются органами как
химического, так и обонятельно-контактного чувства.
На заднем конце тела Rhinotalpa обладает железой над хвостом, которая
служит для защиты; это особенно важно для маленького существа, потому что оно
не способно развернуться, и потому что его туннели часто заселяет мелкий
агрессивный вид земляного краба (Chelygnathomachus29 altevogtii).
Между рудиментарными задними лапами также находятся соски; молодое животное
подталкивается к ним сразу после рождения. Этой особенностью Rhinotalpa
демонстрирует признаки примитивности, поскольку другие однорылы больше не
вскармливают молоком свой молодняк.
Даже у Rhinotalpa уже есть характерные анатомические особенности,
которые в гораздо большей степени реализовались у других представителей роющих
носоходок: тенденции к заполнению полости тела соединительной тканью. У Rhinotalpa
она занимает только плевральные области, поэтому лёгкие здесь прочно связаны с
плевральной стенкой. У Rhinotalpa phallonasus это проявляется в
меньшей степени, чем у более мелкого, близкородственного вида R.
angustinasus (узконосой кротовой носоходки), у которой также найдены
другие структурные особенности, которые неизвестны у кого-либо ещё среди
млекопитающих и связаны с мелким абсолютным размером животных (см. рис. 8).
Среди них должны быть отмечены следующие особенности: сокращение относительной
длины пищеварительного тракта; уменьшение ёмкости лёгких; исчезновение ноздрей;
утрата волос; расширение ресничного эпителия, который у Rh. phallonasus выстилает
только основные пазухи, а здесь достиг основания наружной части рыла; упрощение
мозга; редукция глаз; и, наконец, как наиболее поразительная физиологическая особенность,
полная потеря гомойотермности. Естественно, все эти структурные и
функциональные особенности тесно связаны с повадками животного; оно живёт не в
сплошной почве, а в полостях в осыпях грубого гравия. Там оно ползает взад и
вперёд, во многом подобно Rh. phallonasus, но с тем отличием, что, в
связи с его удлинённым телосложением, оно до некоторой степени способно также к
волнообразному движению. Также примечательно, что существо разыскивает не
только полости, которые заполнены воздухом, но также и те, которые заполнены
грунтовыми водами. В них оно набирает воду в лёгкие, которые редуцированы до
простых мешочков. Пазухи служат дополнительными органами дыхания. В течение
быстрого движения вперёд они также ритмично заполняются водой. Когда животное
находится в покое, кожного дыхания через богато снабжённый железами эпидермис
ему, кажется, достаточно.

Rhinotalpa phallonasus с сайта Токивы Такеши

Rhinotalpa angustinasus с сайта Токивы Такеши
Эволюционные направления, уже обозначенные у Rhinotalpa, гораздо
более полно обозначены у Enterorrhinus, кишечной носоходки.
Представители этого рода вырастают до максимальной длины 0,7 дюйма и
демонстрируют необычайно глубокую редукцию: от конечностей сохранились только
когти, а их мускулатура больше не может быть сопоставлена с определёнными
мускулами конечностей. Кишка вытянута в прямую линию. Лёгкие отсутствуют.
Сердце – простая циркуляционная трубка, и её состояние соответствует тому,
которое в иных случаях демонстрируют ранние эмбрионы млекопитающих. Вся
поверхность тела покрыта ресничками. Мозг полностью лишён отделов, по крайней
мере, внешних. В скелете можно различить только слабо развитый нотохорд; он
протягивается дорсальнее кишки и вперёд под мозгом в рыло. Ничего не известно о
половых органах. Почки – протонефридии, с единственной выстланной ресничным
эпителием воронкой на каждой стороне. Эта воронка входит в эндотелиальный
мешок, который находится в основании рыла внутри полости тела, которая
заполнена соединительной тканью. Мочеполового протока больше нет.
Род представляет по одному виду на каждом из пяти больших островов архипелага.
Там животные обитают на гравийных берегах дельт маленьких ручьёв в ограниченной
области, которая напрямую связана с содержанием соли в грунтовых водах
(приблизительно 0,6% – 1,4%). В пределах таких областей часто находят отдельные
рыла и экземпляры, которые обладают или необычно большим, или чрезвычайно
маленьким рылом относительно размеров их тела, что приводит к предположению,
что размножение происходит путём деления в основании рыла.
Если бы не кротовые носоходки, которые уже известны, и не факт несомненного
родства между Rhinotalpa и Enterorrhinus, никто бы и не
подозревал, что животные, ныне объединённые в семействе Holorrhinidae
(Тольконосы) – это носоходки. Это крошечные существа длиной несколько
миллиметров, чьё строение стало настолько «примитивным», что мысль о
классификации их среди хордовых никогда не пришла бы в голову.
В первом роде тольконосых, Holorrhinus (Тольконос), строение хордовых
проявляется лишь в следующих деталях: удлинённый нотохорд протягивается на
всю длину рыла и через всё сильно редуцированное тело. Присутствует также
замкнутая, хотя и сильно редуцированная циркуляторная система, которая, однако,
несёт черты, свойственные ранним этапам эмбрионального развития. Из почек на
каждом боку всё ещё есть трубка с ресничным эпителием, упомянутая выше; она
открывается в эндотелиальный пузырёк. Когда были найдены половые органы – хотя
только у самца – оказалось, что они лежат недалеко от задней части животного,
в области, где тяж чётко дифференцированных мускулов указывает на былое положение
задних конечностей, которое не проявляется иным образом. Мускулатура связана
со
слегка выступающими углами тела, которые играют некоторую роль при рытье. С
другой стороны, целый ряд структурных особенностей накладывает на признаки
организмов печать отличий от хордовых: удлинённые носовые пазухи участвуют
в пищеварении в дополнение к короткому прямому кишечнику. Они функционируют
как
придатки или таким же образом, как энтодермальные слепые кишки многих
беспозвоночных, поочерёдно заполняясь пищей и освобождаясь от неё. Соматическая
мускулатура больше не поперечно-полосатая, а гладкая. Мозг заметно редуцирован,
и на протяжении ранних стадий развития нервная трубка расщепляется на два
тканевых тяжа, из которой развиваются две прилегающих к нотохорду цепочки
ганглиев, которые связаны поперечными комиссурами. Полость тела заполнена
соединительной тканью. Наружная поверхность, как это уже произошло у Enterorrhius,
покрыта ресничным эпителием, по которому разбросаны слизевыделяющие клетки,
и который аналогичным образом происходит из носовых проходов. В дополнение
к этому
следует отметить, что в эндотелиальном пространстве надпочечников ряды
ресничных клеток несут очень удлинённые реснички, производя эффект наподобие
пламенных клеток.
18 видов рода Holorrhinus распространены по всему архипелагу, где
обитают частью в пойменных песках, а частью в солоноватых водах морских пляжей.
Передвижение обычно происходит хвостом вперёд. Два вида (Holorrhinus
variegatus = изменчивый тольконос и H. rhinenterus = тольконос
Пиноккио) обитают в ручьях; часто их успешно выкармливают из, очевидно, недавно
отложенной молоди, которая встречается в эпителиальном пузыре на стадии
нейрулы, и это дало интересную информацию о строении этих животных. Так, в ходе
этой работы было показано, что глаза (эти животные способны видеть) возникают
как впячивания на мешкообразной стадии развития мозга, но затем сохраняются как
простые пузырьковидные глаза, в то время как мозг вторично теряет свою
центральную полость и становится широким образованием в форме пряжки, главные
ганглиозные скопления которого расположены справа и слева от пищевода.
До настоящего времени найден лишь единственный вид рода Remanonasus30
в речных песках Виси-Виси, речки на острове Майрувили. Это червеобразное
существо, которое достигает максимальной длины 2 мм. Что отличает Remanonasus
menorrhinus31, похожую на турбеллярий карликовую носоходку, от
предыдущего рода, так это прежде всего потеря анального отверстия и кровеносной
системы. Аналогичным образом больше нельзя найти ни следа нотохорда. К
сожалению, до настоящего времени были обнаружены только самцы этих животных.
Почки больше не имеют никакого видимого образования, похожего на трубу с
ресничным эпителием, и оказываются устроенными как протонефридии, с
единственной огромной пламенной клеткой, несущей реснички на каждой стороне.
|
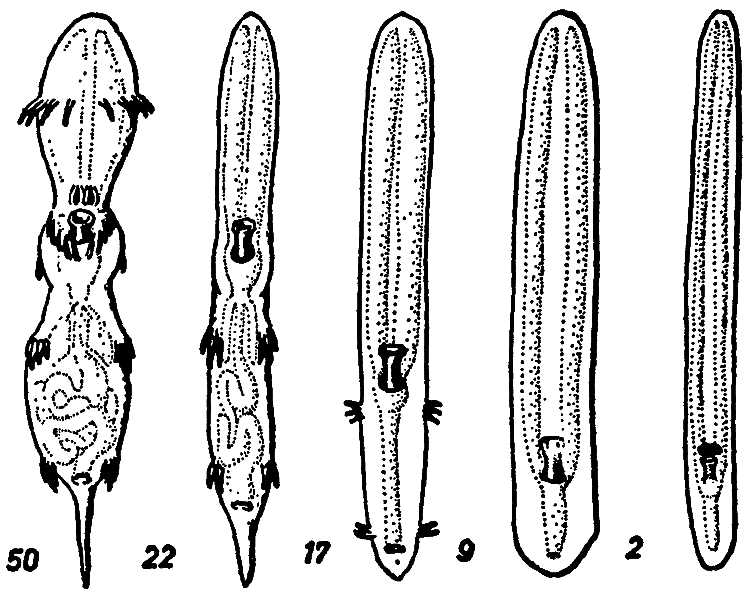
[Рис. 8] Анатомический ряд: Rhinotalpa phallonasus – Rhinotalpa angustinasus –Enterorrhinus dubius – Holorrhinus ammophilus – Remanonasus menorrhinus. Числа показывают общую длину в миллиметрах. Из внутренних органов выделен только пищеварительный тракт. (По Майер-Мейеру, 1949) |
Потому неудивительно, что разные учёные заключали, что эти животные вообще
не должны числиться среди Rhinogradentia. Мюллер-Гирмандинген (1948) описал их
как Dendrocoelopsis minutissima и считал верной их классификацию как
Tridadida; однако Майер-Мейер (1949) показал с помощью тщательных
гистологических исследований, что именно слизевыделяющие клетки не могут
рассматриваться как типичные для трикладид. Однако он был вынужден допустить,
что некоторые структурные характеристики несут столь большое сходство с
таковыми у трикладид из Turbellaria, что, по крайней мере, в целом не является
невозможным то, что трикладиды произошли от Remanonasus-подобных форм.
Так, видимое головное положение мужских половых желез у трикладид – которое
первоначально производит на любого непредубеждённого наблюдателя впечатление
странного – оказывается вполне доступным для понимания, если вспомнить, что
тольконосы ползают задом наперёд и, следовательно, их хвостовой конец стал
физиологически передним. Кроме того, форма пищеварительного тракта становится
полностью понятной тогда, когда – как это иллюстрируется переходными формами
морфологического ряда (см. рис. 8) – он выводится из кишечника + носовых пазух
тольконосых. Конечно, вопрос может быть наконец полностью разрешён лишь после
того, как будут обнаружены животные со зрелым женским половым аппаратом. Этот
самый аппарат у трикладид является очень сложным, и в целом для турбеллярий
позволяет предположить их происхождение от более высоко дифференцированных
животных. Ремейн (1954) также подчеркивает это, но предпочитает выводить
Turbellaria из кольчатых червей. Позже Штультен (1955) склонялся к мнению, что
Rhabdocoela могли бы происходить от кольчецов, но что у Tridadida (и
Polycladida, которые в любом случае должны выводиться из последних) были предки
из числа ринограденций31a.
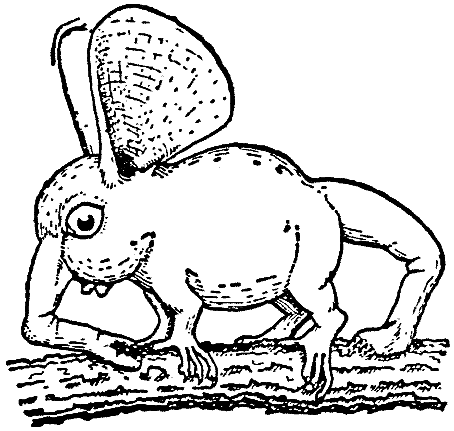
Триба Sclerorrhina (Хоботоноги) образует ряд самых странных и наиболее красивых
видов Rhinogradentia. Общим для них стал тот факт, что nasarium32
стал ведущим органом, носовой ногой, с помощью которой животное может делать
сильные прыжки; однако, из-за действия гравитации (см. таблицу V) они
направлены назад.
Наиболее примитивные состояния встречаются среди древесных мордоскоков,
перихопсид33, чьи конечности всё ещё показывают некоторое подобие
конечностям Archirrhiniformes. Однако с локомоторной точки зрения наиболее
типичными должны быть признаны Hopsorrhinidae, мордоскоки в узком смысле, у
которых задние лапы исчезли, за исключением небольших остатков бедра и голени,
и чьё рыло служит единственным органом передвижения. Наконец, у Orchidiopsidae
(Орхидорылов) рыло вторично стало мягким, в соответствии с их более сидячим
образом жизни.
27 remanere Лат. = оставаться; nasus см. сноску 3
28 Была сделана попытка связать происхождение chaetae submentales с собирающей корзиной носоходок-трубачей и таким образом вывести роющих носоходок из иловых носоходок (Naquedai 1948). Однако все остальные детали организации этих двух подтриб свидетельствуют против этой концепции.
29 chele см. сноску 25; gnathos Греч. = челюсть; maMmenos или -machos Греч. = воин.
30 см. сноску 27.
31 menein Греч. = оставаться; rhis см. сноску 4.
31a Ради объективности следует отметить, что Гриссе в своём предисловии к Anatomie et biologie de l’Rhino grades (Masson & Cie, Париж, 1962 год) оспаривает тот факт, что Remanonasus является носоходкой. Он предпочитает считать его скорее обитающим в толще песка плоским червём, нежели деградировавшей носоходкой.
32 см. сноску 5.
33 peri Греч. = вокруг; hopsos см. сноску 4.
ПОДТРИБА: Hopsorrhinida (Мордоскоки в
широком смысле),
СЕМЕЙСТВО: Amphihopsidae (Мордоскоки-тянитолкаи, или Древесные мордоскоки);
РОД: Phyllohopla (Листопрыги),
2 вида,
СЕМЕЙСТВО: Hopsorrhinidae (Мордоскоки в узком смысле),
РОД: Hopsorrhinus (Зубастые мордоскоки),
14 видов,
РОД: Mercatorrhinus (Сосущие мордоскоки),
11 видов,
РОД: Otopteryx (Крылоухи),
1 вид,
СЕМЕЙСТВО: Orchidiopsidae (Орхидорылы),
РОД: Orchidiopsis (Орхидорылы),
5 видов,
РОД: Liliopsis (Лилиерылы),
3 вида.
Мордоскоки-тянитолкаи – это существа девственного леса; они живут в кронах
деревьев, где проворно прыгают с ветки на ветку или спокойно ползают по
прутикам. Они имеют компактное телосложение и, подобно большинству
Rhinogradentia, являются насекомоядными существами размером примерно с мышь.
Хотя их туловище и конечности сохранили много особенностей, характерных для
Archirrhiniformes, на их большой голове с крупными глазами явственно заметно
суставчатое рыло, которое заканчивается на дистальном конце дорсально
расположенной подошвенной пластинкой, и которое движется с помощью сильных
лицевых мускулов и сильного вытягивающего мускула (протрактора), M.
extensor nasipodii longus (= M. longissimus nasarii). По
Штультену, M. extensor nasipodii longus развился из удлинённого в
переднем направлении M. longissimus dorsi или М. l. thoracis,
на что указывает его иннервация через спинномозговые нервы грудного отдела тела
(см. также рис. 11). Хвост имеет такую же странную форму, как и рыло; он также
исключительно мускулистый и сильный, и несёт конечную подошвенную пластинку,
крепкие щетинки которой позволяют хвосту прочно закрепляться на неровных
участках субстрата (рис. 9). В дополнение к метамерным хвостовым мускулам,
которые как основной признак всегда сохранялись у Rhinogradentia – примитивная
особенность, на которую уже обратили внимание Труфагура (1948) и Изеха (1949) –
поверх них протягивается M. iliocaudalis, который действует как
разгибатель хвоста. И так, с помощью морды и хвоста периопсиды способны прыгать
с невероятным проворством взад и вперёд через чащу лиан, когда передом, когда
задом наперёд, а затем боком, и поэтому их очень трудно ловить. Это их
проворство в первом приближении трудно понять, поскольку у них фактически нет
врагов. Однако они живут вместе маленькими группами, внутри которых происходит
постоянное движение туда-сюда, преследование и избегание, что, без сомнения,
имеет отношение к их иерархии, социологическое значение которой ещё не
полностью объяснено. Кроме того, эти способности, конечно, полезны для животных
при сборе корма; они питаются почти исключительно летающими насекомыми, которых
схватывают во время прыжков.
|
[Рис. 9] Phyllohopla bambola. (Orig). |
Hopsorrhinidae, в противоположность предыдущей группе носоходок, обитают на
земле. Как уже было сказано, их задние конечности рудиментарны и больше не
видны снаружи. Рыло ещё более дифференцировано, чем у Perihopsidae, в данном
случае оно подверглось разделению на три доли; рядом с головой (см. рис. 10) – nasur,
который связан суставом с nasibia, к которому на конце прикрепляются ринангии.
Выпрямление nasur и nasibia происходит с помощью двух
раздельных тяжей M. extensor nasipodii, тогда как ринангии
движутся с помощью лицевой мускулатуры и M. flexores longi и breves
назиподиума (который составляют вместе зигоназиум и автоназиум), которые также
являются производными лицевой мускулатуры.
В результате сращения позвонков и из-за вентрального ребра жёсткости,
образованного грудиной и processus styliformis34 лобковой
кости, туловище оказывается заключённым в твёрдую капсулу. Передние конечности
– подвижные хватательные органы. Хвост больше не способствует движению вперёд,
но используется для защиты от пищевых объектов, в качестве которых выступают
прежде всего бокоплавы, равноногие ракообразные и мелкие раки-отшельники,
обитающие в верхней зоне литорали. В связи с этим одиночная опорная хвостовая
пластинка видоизменилась, превратившись в пару хватательных клещей, когти
которых образовались из видоизменённых или слившихся волос, и на поперечном
срезе демонстрируют гистологическую картину, напоминающую ткань рога носорога.
Благодаря такому хвосту мордоскоки весьма преуспели в способности вытаскивать
свою добычу из самых узких расщелин и укрытий. Их прыжки, которые при обычной
скорости движения приблизительно в полтора раза превышают длину тела, а при
спасении бегством или преследовании брачного партнёра или конкурента могут быть
и в десять раз длиннее, обычно делаются задом наперёд (см. табл VI). Прыжок
может в некоторой степени управляться движениями огромных ушей.

Hopsorrhinus с сайта Токивы Такеши
Мордоскоки принадлежат к наиболее часто встречающимся представителям
Rhinogradentia. Повсюду на побережье они населяют каменистые коралловые глыбы,
пойменные пески или гравийные россыпи вулканического или осадочного
происхождения. Очевидно, более сильные самцы содержат небольшие гаремы и
отгоняют более слабых самцов. Однако, различия между полами настолько невелики,
что полевые наблюдения ещё не позволяют анализировать детали поведения внутри
групп.
Штультен поместил первые два рода семейства Hopsorrhinidae в единый род Hopsorrhinus.
Однако позднее  Бромьен
де Бурла пришёл к выводу, что род Mercatorrhinus35
определённо заслуживает выделения; тогда как все виды Hopsorrhinus
питаются вышеописанным способом и в связи с этим обладают исходным типом зубной
системы (отсюда также обиходное название «зубастые мордоскоки»), все виды Mercatorrhinus
более не способны проглатывать твёрдую пищу и полностью зависят от описанных
выше (стр. 15) симбиотических отношений со столбоносыми носоходками рода Columnifax.
Это явно заметно не только в строении их ротовых частей – отсутствии зубов,
редукции M. masseter и M. Temporalis – но также в
рудиментарной природе передних конечностей. Вместе с этими признаками редукции
животные, в прямой связи с их симбиозом, приобрели некоторые способности,
которых нет у самостоятельно питающихся форм. Так, они могут присаживаться на
хвосте, который они сворачивают под собой, и регулярно делают это каждый раз,
когда, передав их добычу, садятся рядом с Columnifax, ожидая питания.
(Для составителя этих примечаний [Штайнера], однако, аргументы де Бурла не
имеют полной убедительности. Поэтому, по примеру самого Штумпке, вопрос о роде Mercatorrhinus
оставлен открытым.)
Бромьен
де Бурла пришёл к выводу, что род Mercatorrhinus35
определённо заслуживает выделения; тогда как все виды Hopsorrhinus
питаются вышеописанным способом и в связи с этим обладают исходным типом зубной
системы (отсюда также обиходное название «зубастые мордоскоки»), все виды Mercatorrhinus
более не способны проглатывать твёрдую пищу и полностью зависят от описанных
выше (стр. 15) симбиотических отношений со столбоносыми носоходками рода Columnifax.
Это явно заметно не только в строении их ротовых частей – отсутствии зубов,
редукции M. masseter и M. Temporalis – но также в
рудиментарной природе передних конечностей. Вместе с этими признаками редукции
животные, в прямой связи с их симбиозом, приобрели некоторые способности,
которых нет у самостоятельно питающихся форм. Так, они могут присаживаться на
хвосте, который они сворачивают под собой, и регулярно делают это каждый раз,
когда, передав их добычу, садятся рядом с Columnifax, ожидая питания.
(Для составителя этих примечаний [Штайнера], однако, аргументы де Бурла не
имеют полной убедительности. Поэтому, по примеру самого Штумпке, вопрос о роде Mercatorrhinus
оставлен открытым.)
Все сосущие мордоскоки могут легко содержаться в неволе, как уже было отмечено
в начале (стр. xxiv), и успех
зависит от того, насколько легко можно достать искусственную пищу-заменитель:
поскольку молоко Columnifax относительно богато сахаром и в нём низкое
содержание жира, оно, таким образом, очень напоминает женское молоко, животных
можно без труда выкормить сгущённым молоком, используемым для питания. Тем не
менее, это можно успешно сделать только с помощью приспособления, которое было
открыто путём утомительного изучения поведения животных.
|
Таблица VI |
Когда такой сосущий мордоскок хочет есть, задействуется его механизм поиска пищи. Животное бродит по разным местам и засовывает свой хвост в расщелины и трещины, чтобы захватить вышеупомянутых крабов. Поймав добычу, он осторожно приближается к столбоносой носоходке и с помощью характерных прыжков привлекает к себе внимание последнего. Лишь когда ближайший Columnifax издаёт хрюкающий звук, сосущий мордоскок подходит ближе, и всякий раз он приближается к брюшной стороне Columnifax. Со своей стороны, последний продолжает постоянно поворачиваться вдоль своей продольной оси при приближении любого животного, всегда готовый выбрызнуть секрет вонючих желёз в потенциального нападающего. Но теперь Mercatorrhinus подходит ближе таким способом, совершая повторяющиеся боковые прыжки вправо и влево, чтобы Columnifax мог ясно увидеть его. Также в перерывах он стоит на своей морде и хвостом держит добычу над собой, энергично и призывно махая ею из стороны в сторону. Только когда Columnifax прекращает своё продольное вращение и слышится испускаемое им долгое гортанное сопение, Mercatorrhinus вплотную приближается к его брюшной стороне и передаёт ему предмет обмена, свою добычу. Сразу после этого Columnifax изучает её, чтобы убедиться, что она совсем свежая. Если она не такая, столбоносая носоходка сразу принимает защитную позу и обрызгивает вонючими выделениями всего пришельца, если последний тем временем не поторопился отскочить подальше на безопасное расстояние. Только когда «товары» безупречны, Columnifax предлагает свою грудь Mercatorrhinus, который тогда делает небольшой подскок на своём свёрнутом пружиной хвосте и начинает сосать.
|
ТАБЛИЦА VII |
Теперь, при наблюдении за содержащимися в неволе сосущими мордоскоками,
стало ясно, что их нельзя содержать без Columnifax. С другой стороны,
подходящую добычу можно было собрать лишь с большими трудностями. Но как только
случайные наблюдения показали, что Mercatorrhinus хорошо чувствует
себя на упомянутой искусственной диете, было испытано принудительное кормление.
Однако это утомительно и часто в процессе этого активные животные получают
смертельные повреждения. Решение дало только тщательное наблюдение за жёстко
закреплённым «ритуалом заключения сделки»: было отмечено, что Mercatorrhinus,
даже если не нашёл никакого подходящего корма для Columnifax, тем не
менее, иногда всё ещё пытался сосать грудью. Описанная выше «устрашающая»
реакция защиты Columnifax не будет проявляться, если Columnifax
всё ещё испытывает ощущение сытости от предыдущей трапезы, а также в избытке
образует молоко, уменьшение количества которого приносит чувство удовольствия.
При таких обстоятельствах Mercatorrhinus-«обманщик», который
предложил, например, просто пустую раковину улитки без Pagurus внутри,
может получить данное молоко. С другой стороны, Mercatorrhinus не
сможеть пить, если сперва сам не принесёт что-либо, без полного ритуала обмена.
Он должен захватить «добычу» и подарить её, совершив танцующий подход,
описанный выше, прежде чем будет пить. Помимо этого, имеют значение следующие
особенности со стороны Columnifax: клиновидная форма, расширенная
сверху; жёлтый цвет; пятна глаз в нижней трети; шипящий звук; и надлежащая
форма вымени. Помимо этого, он должен взять «добычу» у кормильца. Один из
коллег Битбрейна добился успеха, создав относительно простой управляемый с
помощью электроники прибор, который отвечал обозначенным требованиям. При
максимальной скорости работы он способен кормить грудью 80 экземпляров Mercatorrhinus
в час. В качестве «добычи» используются пустые раковины улиток; после того, как
модель Columnifax принимает их, они падают вниз на фальшдно клетки, в
трещины, из которых Mercatorrhinus снова могут вытаскивать их.
Численность специфичных для Mercatorrhinus блох, которые вначале
наносили большой ущерб экспериментам, может эффективно контролироваться с
помощью липкой бумаги на нижней стороне верхнего дна клетки с двойным дном (ДДТ
и другие инсектициды слишком ядовиты для Mercatorrhinus).
Ухокрыл Otopteryx volitans B. d B. (= Hopsorrhinus viridiauratus36STU.),
единственный представитель своего рода, по своему строению чётко определяется
как видоизменившийся мордоскок (таблица VIII). Фактически это животное
отличается от своих кузенов только огромным размером ушей и дифференциацией и
усилением, в соответствии с его способностью к полёту, мускулатуры наружного
уха. Ещё одно отличие, его рудиментарный хвост, является малозначительной
особенностью строения. Во всех других отношениях Otopteryx – это
типичный мордоскок, потому Штультен даже сомневался в обоснованности отделения
его от других родов. Однако в дополнение к сказанному в пользу установления для
него отдельного рода должны быть приведены следующие положения: nasarium
чрезвычайно удлинённый и грацильного сложения. Мускулы, которые двигают ринангии,
частично редуцированы, поэтому животное не способно бегать по неровной земле с
проворством мордоскоков. С другой стороны, отводящие мышцы ринангиев
особенно сильные; они служат для того, чтобы растопыривать autonasium,
который работает как рулящий хвост. На голове далее следует упомянуть развитие
специальных костных гребней – мест прикрепления ушной мускулатуры – а также os
alae auris, которая, однако, не кость, а обызвествлённый волокнистый хрящ;
и в дополнение к этому образование заполненных воздухом боковых носовых пазух
под и рядом с упомянутыми гребнями. Вместе с мордоскоками Otopteryx
демонстрирует инверсию направления роста волос на значительной части
поверхности огузка.
|
ТАБЛИЦА VIII |
У Otopteryx переливчатый блеск кожи, который придаёт другим представителям хоботоногов блеск металлических поверхностей или драгоценных камней, достигает своей наивысшей степени проявления, поэтому животных можно сравнить лишь с тропическими бабочками или колибри. Потому это великолепный вид, когда с помощью быстрых взмахов своих ушей животное стремительно мчится совсем невысоко над украшенными цветами горными лугами, преследуя стрекоз или Hexaptera, или резко взмывает вверх в синее небо, кружась там в играх с другими особями своего вида. Наиболее очаровательны среди них – новорождённые, всё ещё едва способные протянуть свои уши – которые зигзагами летают над цветами, словно стрекозы-красотки, в поисках мелких насекомых. Самая странная вещь изо всех – это то, что Otopteryx летает задом наперёд, хотя это также является вполне доступным для понимания, если вспомнить, что полёт Otopteryx развился из планирующих прыжков мордоскоков, которые прыгают задом наперёд.
|
|
|
|
|
|
|
|
|
|
|
Otopteryx с сайта Токивы Такеши |

Особенно специфичны и характерны взлёт и приземление ухокрылов. Животное, стоящее на своём подогнутом рыле, сперва «взводит» свои уши, то есть, поднимает их вертикально таким образом, чтобы они коснулись друг друга; затем сгибает дейтероназальный сустав даже ещё сильнее, чем это делает Hopsorrhinus (см. таблицу VI выше); после этого дальнейшие стадии следуют, как у последнего, с отличием в том, что прыжок имеет более вертикальное направление. Незадолго до того, как прыжок достигнет предельной высоты, уши с силой опускаются. Полностью выпрямленное рыло широко расправляется в автоназальной области и животное летит. Эти отдельные стадии, конечно, могут быть проанализированы только с помощью высокоскоростной фотосъёмки. Процесс самого полёта чрезвычайно разнообразен: когда преследуется искусно летающее насекомое, или во время игровых лётных манёвров, большие расстояния преодолеваются на бешеной скорости, когда уши непрерывно совершают движения вверх и вниз со скоростью примерно десять ударов в секунду. Во время поискового полёта взмахи ушей той же частоты, но маленькой амплитуды, чередуются с короткими периодами планирующего полёта. Otopteryx также способен парить вдоль склонов, в потоках бризовых островных ветров. Во всех случаях он редко взлетает высоко в воздух и большей частью держится на высоте не более двадцати ярдов. Его способ приземления специфичен, будучи затруднённым тем фактом, что рыло должно исполнять двойные обязанности, и как нога, и как рулящий хвост. Когда ухокрыл хочет опуститься на землю, он главным образом приближается к месту посадки крутым планирующим полётом, с ушами, отведёнными в несколько дорсальное и назальное положение. Сильно приблизившись к земле, он резко принимает вертикальное положение, когда задний конец несколько приподнят, которое завершается резкой, временно восходящей дугой, когда рулящий хвост – то есть, кончник рыла – почти касается земли. В этом положении, когда уши сильно выгнуты (М. inarcantes auris37), животное скользит вперёд почти к самой земле, теряя в это время высоту и скорость. Затем оно резко складывает носовой рулевой аппарат, подгибает рыло вентрально и после того, как уши полностью подняты, позволяет себе пружинисто сесть на рыло, которое в данный момент отведено далеко в каудальном направлении. Эта последняя стадия процесса посадки вновь демонстрирует большое сходство с приземлением мордоскоков после прыжка (см. таблицу VI выше, стадии 6 – 8).
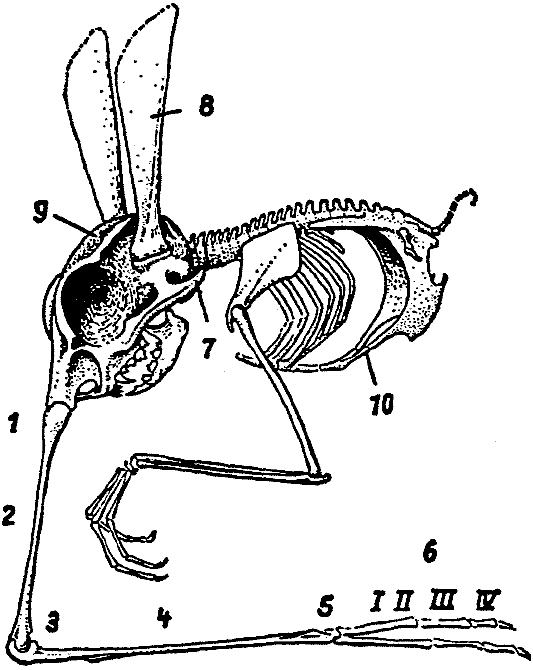
[Рис. 10] Otopteryx volitans, скелет. 1. Articulatio nasofrontalis; 2. Nasur; 3. Articulatio deutonasalis; 4. Nasibia; 5. Articulatio carponasalis; 6. Ринангии (= Назалгии) I – IV; 7. Processus jugalauris; 8. Os alae auris (= Cartiago aeroplana), 9. Christa temporalis; 10. Processus pubici (Ориг.)

(Фотография включена в электронное издание)
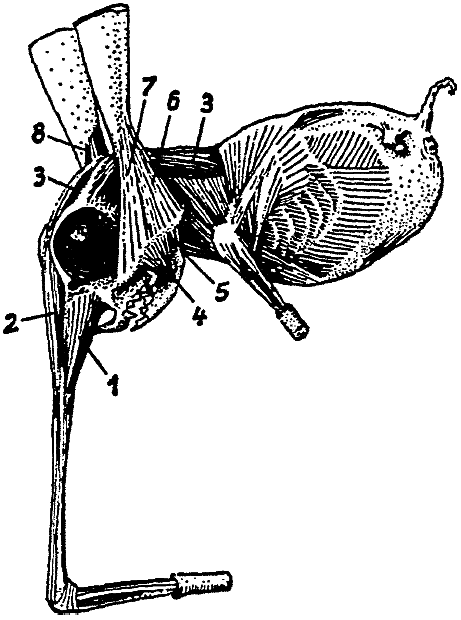
[Рис. 11] Otopteryx volitans, мускулатура.
1. M. lacrymonasuralis; 2. M. extensor nasipodii superficialis; 3. M. extensor nasipodii longus; 4. M. masseter; 5. M. depressor mandibulae; 6. M. aeroplano-jugalairis posterior; 7. M. aeroplano-jugalauris anterior; 8. Levator aeroplanae.Справа от 3, M. extensor nasipodii был открыт путём частичного удаления M. trapezius cervicalis. (Ориг.)
Способ, которым Otopteryx решил проблему передвижения – наиболее экстраординарное решение с морфологической позиции – требует сравнения с другими летающими формами животного царства. Помимо самих Rhinogradentia, истинные летающие формы в целом появились лишь четыре раза: насекомые, летающие рептилии, птицы и летучие мыши. Среди них насекомые, чьи летательные органы являются дополнением, сделанным не за счёт органов передвижения по земле, фактически нашли наиболее законченное решение. Двуногая походка птиц аналогичным образом даёт им большую подвижность и на земле, и в воздухе, даже при том, что в действительности крылья были «украдены» у механизма наземного передвижения. Среди летающих рептилий и летучих мышей способность к полёту возникла за счёт передвижения на ногах; и потому эти две группы имеют недостаточно навыков, чтобы конкурировать с вышеупомянутыми. Но в случае Otopteryx ситуация такая же благоприятная, как у насекомых, то есть уши – фактически дополнительные инструменты для полёта. Тем не менее, Otopteryx с высокой степенью редукции конечностей, предшествовавшей полёту, происходит от животных, которые были сильно специализированы в единственном направлении – хотя из-за их носовой «монополии» их достаточно легко сравнить с прыгающими птицами. В любом случае Otopteryx обладает заметным преимуществом перед летающими рептилиями или летучими мышами, поскольку он более искусный прыгун, а участие рыла в полёте не ограничило его пригодности для наземного передвижения в той же степени, как это произошло у представителей двух предыдущих групп. Вопрос о том, смог бы Otopteryx выжить в более суровой конкуренции с материковыми животными, не разрешён. При любом раскладе у него вряд ли были какие-либо враги на островах. Ни туземные птицы-мегафоны, ни морские птицы, которые время от времени собираются многочисленными стаями на побережье, не могут поймать его в полёте. С этим согласуется тот факт, что беременные экземпляры встречаются редко. Период беременности, по крайней мере, такой же короткий, как у мордоскоков. Каждый раз вынашивается только единственный зародыш (Харрокерриа и Ирри-Эгингам). Есть предположение, что самки приносят двух детёнышей в год.
Ухокрылы не живут в неволе, потому что они остаются слишком
пугливыми, отбивают зад в своих диких прыжках и попытках взлететь, и вскоре
становятся жертвами
заразных болезней, которые возникают в ранах.
Родословная орхидорылов (Orchidiopsidae) может быть отслежена назад к их
предкам из числа мордоскоков, которые сменили наземный образ жизни на
древесный, но которые уже обладали различными признаками редукции, характерными
для мордоскоков, и в частности, исчезновением задних конечностей. Поэтому
из них больше не могли развиться умелые древолазы. Вместо этого стало развиваться
эволюционное направление, при котором животные больше не прыгали с места
на
место, а скорее медленно лазали с помощью передних конечностей и хвоста.
Представителей с функциональной носовой ногой в любом случае больше не
известно, и по своим повадкам и строению виды Orchidiopsis и Liliopsis в
настоящее время столь высоко специализированы, что первоначально их происхождение
от мордоскоков даже не рассматривалось (см. Гокари-Судур, Бюффон
и Пейгниопулос). Однако в то же время были получены некоторые данные, связанные
с развитием, которые имеют эволюционное значение, и из них без возможных
противоречий ясно, что nasur и nasibia, равно как ринангии,
вначале закладываются на эмбриональной стадии рыла Orchidiopsis,
но позже резорбируются, поэтому рыло полностью развитого животного следует
рассматривать как вторично упрощенное. (Бюффон и Запартегингарри, 1953, пишут: “Les
embryons des Orchidiopsides ne manquent point m de nasur ni de nasibie, mais
pendant le develop fement [longueur de l’embryon eniiron 15 a 18 mm] il y a
lieu un ramollusement progressif de ceux-ci, de telle facon, que le nouveau-ne
ne montre plus ancune trace d'ossification dans son nasarlum epiati et
petaloide.”*) В особенности Бюффону и его школе мы обязаны дальнейшим
объяснением особенностей организации Orchidiopsidae. Так, Бюффон и
Ло-Ибилатц-Судур смогли показать, что секретируемый аттрактант («слизистое
облачение») видов Orchidiopsis не выделяется наружной носовой
поверхностью, которая фактически лишена любых железистых клеток, которые
могли быть задействованы в этом. Он скорее производится большими железистыми
областями внутренней поверхности, вдоль носовых проходов, и после этого
распределяется вручную по наружной поверхности рыла. Кроме того, тем же самым
авторам удалось показать, что хватательные механизмы хвоста являются
гомологичными образованиями у Orchidiopsis и у Hopsorrhinus.
И, наконец, Астеид, ученик Бюффона, смог представить доказательство того,
что в рыле орхидорылов есть остатки M. extensor nasipodii, который
в процессе эмбрионального развития находится в положении, характерном для
Rhinogradentia из числа мордоскоков.
*Перевод: (Бюффон и Запартегингарри, 1953, пишут: «Эмбрионы орхидорылов не имеют ни nasur, ни nasibia, поскольку в ходе развития [длина эмбриона от 15 до 18 мм] они подвергаются прогрессивному размягчению таким образом, что новорождённое животное более не показывает никаких следов окостенения в своём гладком, похожем на лепесток nasarium».)
|
|
Ванильный орхидорыл, Orchidiopsis rapax38, наиболее известный представитель семейства, живёт в лесах Митадины, в особенности на уровне более высоких древесных крон, а также на меньших высотах в местах, где ветровалы или наводнения оставили в лесу открытые места (таблица IX). Обычно животное неподвижно стоит на своём хвосте и потому при взгляде издалека имеет некоторое сходство с большим цветком39. Причина подобия состоит в том, что огромные уши, медиальный кожный гребень и уплощенное рыло, торчат на голове, сверкая яркими красками, словно лепестки цветка, тогда как неприметное зелёное туловище незаметно на первый взгляд. Уже упомянутый выделяемый аттрактант на рыле пахнет ванилью и действует как пахучая приманка. Насекомые, которые садятся на рыло или летают близко к нему, оказываются схваченными с молниеносной быстротой цепкими пальцами на концах длинных тонких передних лап, и отправляются в рот. Редкие смены местоположения происходят с медлительностью, достойной хамелеона, с помощью передних лап и двухкоготного хватательного хвоста. Относительно взаимоотношений между отдельными особями ничего не известно; однако, до настоящего момента было добыто около дюжины беременных экземпляров, носящих эмбрионы на различных стадиях развития (см. выше).

Orchidiopsis с сайта Токивы Такеши
Среди трёх видов Liliopsis, которые отличаются от Orchidiopsis положением ушей и головных гребней, есть один, который спит днём и «цветёт», то есть, принимает свою хищную позу, ночью. Это животное, обычно называемое по-английски «пламенной лилией» и по-немецки “Wundernase” (= чудо-нос) (Liliopsis thaumatonasus), необычно тем, что его слизистый аттрактант испускает свет. Как и в случае с люминесцентной слизью других животных (см. Бюхнера), у «пламенной лилии» свет, по всей вероятности, также производится симбиотическими бактериями. При всём при этом крошечные организмы из слизи, которые отвечают за это явление, ещё не культивировались, и увеличение с помощью электронного микроскопирования не смогло подтвердить их клеточную природу.
34 P. styliformis – структурное новшество, которое не имеет никакого отношения к пред- или надлобковой кости, а также не показывает никакого отношения к os marsupialis однопроходных или дидельфид [опоссумов – В. П.].
35 см. сноску 17.
36 viridi-auratus Лат. = зеленовато-золотой.
37 inarcare Лат. = выгибать в виде дуги.
38 rapax Лат. = хищный.
39 Поскольку орхидеи не встречаются ни на одном острове в архипелаге, название «Orchidiopsis» фактически представляет собой неудачный выбор; в этой ситуации нет никакого подражания орхидее. Во всех случаях те цветы (Rochemontia renatellae ST.) которые являются моделями для покровительственного сходства Orchidiopsis, на первый взгляд очень похожи и по внешнему виду, и по запаху на орхидеи, хотя они близкородственны Ranuiculaceae [семейству лютиковых – В. П.].
ПОДОТРЯД: Polyrrhina (Многорылы),
КОГОРТА: Brachyproata (Короткорылые носоходки),
ТРИБА: Tetrarrhinida (Четырёхрылые носоходки),
СЕМЕЙСТВО: Nasobemidae (Носоходки в широком смысле),
РОД: Nasobema (Собственно носоходки),
5 видов,
РОД: Stella (Крохотные носоходки),
l вид,
СЕМЕЙСТВО: Tyrannonasidae (Хищные носоходки в широком смысле)
РОД: Tyrannonasus (Хищная носоходка),
l вид.
 Как
говорит название, Polyrrhina занимают обособленное место из-за обладания
несколькими рылами. Этой деталью они иллюстрируют особенность, выглядящую
элементом, чуждым для систематики млекопитающих, но, тем не менее – если
рассматривать в общем смысле – повторяющуюся в других филогенетических рядах
животных, а именно увеличение количества органов, которые у более или менее
близкородственных форм возникают в единичном количестве или в виде единственной
пары, или которые присутствуют только у немногих из этих родственных форм. Как
известно, можно – хотя бы мысленно – вывести всех Articulata из многочленных
существ, прототипы или более ранние предки которых олигомерны; подобным же
образом здесь может быть упомянуто изменчивое число жаберных щелей среди
сравнительно близкородственных низших позвоночных. И теперь многорылость у
многорылых Rhinogradentia оказывается – чисто формально – лишь не более чем
простым увеличением количества органов на ранней эмбриональной стадии (см. рис.
1). В любом случае было бы неточностью упоминать об этом просто как о примере
многократного уродства или неправильной модификации и проводить параллели между
этим явлением и тем, что известно о морфогенезе аберрантных мутантов Drosophila,
мутантов, как это попытались сделать Кнаддл и Кишерлинг. Миддлстед и Хассенштин
весьма справедливо отметили, что при простом дублировании независимое движение
нескольких рыл неизбежно было бы невозможным, поскольку из исследований П.
Вайсса среди прочего известно, что такие дубликаты получают идентичные
двигательные импульсы. Таким образом, среди Rhinogradentia многорылость
сопровождается соответствующей централизованной нервной координацией столь же
высокой степени дифференциации. С эволюционной точки зрения эта ситуация
содержит в себе существенные трудности, если вспомнить, что Rhinogradentia,
очевидно, возникли, самое раннее, в верхнем мелу. Согласно Ремайну (1954), как
свидетельство относительной примитивности должнен быть принят тот факт, что
среди многорылов существуют по крайней мере три группы в отношении состояния
полиринии: с четырьмя, с шестью и с 38 рылами, помимо различных типов
дифференцировки рыла внутри каждой из этих групп. Также следует считать,
разделение этих групп произошло очень рано; равным образом и отделение
многорылов от однорылов должно было пройти очень рано. На настоящем этапе
исследований трудно даже отследить родословную многорылов назад к Archirrhinos
или к примитивным Rhinogradentia из числа архирриниформ. Также ошибочно
пытаться отследить происхождение многорылов назад к носоходящим однорылам, как
это попытался сделать д’Эпп (Штультен 1949; Бромьен де Бурла 1949). Одна из
главных причин для этого суждения – это то, что строение nasarium –
помимо вопроса многорылости – является в целом различным, и что редукция задних
конечностей происходит весьма различным образом; кроме того, число рёбер и
развитие зигапофизов позвонков у однорылов совершенно отклоняющееся (в
направлении состояния, наблюдаемого у Xenarthra, но, конечно, как проявление
конвергенции), тогда как многорылы более полно сохранили исходные признаки.
Как
говорит название, Polyrrhina занимают обособленное место из-за обладания
несколькими рылами. Этой деталью они иллюстрируют особенность, выглядящую
элементом, чуждым для систематики млекопитающих, но, тем не менее – если
рассматривать в общем смысле – повторяющуюся в других филогенетических рядах
животных, а именно увеличение количества органов, которые у более или менее
близкородственных форм возникают в единичном количестве или в виде единственной
пары, или которые присутствуют только у немногих из этих родственных форм. Как
известно, можно – хотя бы мысленно – вывести всех Articulata из многочленных
существ, прототипы или более ранние предки которых олигомерны; подобным же
образом здесь может быть упомянуто изменчивое число жаберных щелей среди
сравнительно близкородственных низших позвоночных. И теперь многорылость у
многорылых Rhinogradentia оказывается – чисто формально – лишь не более чем
простым увеличением количества органов на ранней эмбриональной стадии (см. рис.
1). В любом случае было бы неточностью упоминать об этом просто как о примере
многократного уродства или неправильной модификации и проводить параллели между
этим явлением и тем, что известно о морфогенезе аберрантных мутантов Drosophila,
мутантов, как это попытались сделать Кнаддл и Кишерлинг. Миддлстед и Хассенштин
весьма справедливо отметили, что при простом дублировании независимое движение
нескольких рыл неизбежно было бы невозможным, поскольку из исследований П.
Вайсса среди прочего известно, что такие дубликаты получают идентичные
двигательные импульсы. Таким образом, среди Rhinogradentia многорылость
сопровождается соответствующей централизованной нервной координацией столь же
высокой степени дифференциации. С эволюционной точки зрения эта ситуация
содержит в себе существенные трудности, если вспомнить, что Rhinogradentia,
очевидно, возникли, самое раннее, в верхнем мелу. Согласно Ремайну (1954), как
свидетельство относительной примитивности должнен быть принят тот факт, что
среди многорылов существуют по крайней мере три группы в отношении состояния
полиринии: с четырьмя, с шестью и с 38 рылами, помимо различных типов
дифференцировки рыла внутри каждой из этих групп. Также следует считать,
разделение этих групп произошло очень рано; равным образом и отделение
многорылов от однорылов должно было пройти очень рано. На настоящем этапе
исследований трудно даже отследить родословную многорылов назад к Archirrhinos
или к примитивным Rhinogradentia из числа архирриниформ. Также ошибочно
пытаться отследить происхождение многорылов назад к носоходящим однорылам, как
это попытался сделать д’Эпп (Штультен 1949; Бромьен де Бурла 1949). Одна из
главных причин для этого суждения – это то, что строение nasarium –
помимо вопроса многорылости – является в целом различным, и что редукция задних
конечностей происходит весьма различным образом; кроме того, число рёбер и
развитие зигапофизов позвонков у однорылов совершенно отклоняющееся (в
направлении состояния, наблюдаемого у Xenarthra, но, конечно, как проявление
конвергенции), тогда как многорылы более полно сохранили исходные признаки.
Однако и одно- и многорылы совместно демонстрируют одну особенность;
расширенный слёзный проток, который во многих случаях служит в качестве
дыхательных путей. Бромьен де Бурла рассматривает это как результат
конвергенции в строении, что, вероятно, связано со сменой функции рыл (р) и их
полостей. Так случается, что дистальные носовые отверстия у тех форм, у которых
они образуются, главным образом содействуют специальным функциям, которые не
связаны с дыханием: исследование запахов, потребление пищи и, наконец, даже
участие в издавании голоса у животного (см. стр. 83).
Детали, касающиеся устройства nasarium’а, невозможно изложить в этом
кратком сообщении; для них мы даём ссылку на исследования Бромьена де Бурла,
Штультена и Бюффона, а также на всесторонний обзор этих вопросов Х. Штумпке.
|
ТАБЛИЦА X |
 Большая
моргенштернова носоходка, хонатата туземцев (Nasobema lyricum; таблица
X), является лучше всего известным представителем многорылов, и этот пример
будет обсуждаться с несколько большими подробностями. Будучи типичным
многорылом, она несёт на своей короткой толстой голове четыре рыла равной
длины, которые довольно длинные, и на которых – как уже описанно Моргенштерном
– она ходит. Она может это делать, несмотря на несответствующий носовой скелет,
потому что рыла становятся весьма твёрдыми благодаря сильному тургору их corpora
spongiosa. Кроме того, в рыла проникают сильно разветвлённые воздушные
ходы, заполнение которых регулируется с помощью ampullae choanales (образования,
дифференцировавшиеся из мягкого нёба по краям носовых раковин, которые
простираются далеко в вентро-каудальном направлении), поэтому тургор рыла обеспечивается
двумя системами: гидравлической системой corpora spongiosa, которая
ответственна прежде всего за долговременное сохранение жёсткости, необходимой
для ходьбы, и пневматической системой, которая обеспечивает походку и
пружинистые движения, что уменьшает опасность повреждения, когда неожиданно
возникают твёрдые препятствия. В дополнение к роли, которую играют ampullae
choanales, в этом принимают участие также ampullae pneumonasales,
которые образовались из сильно разросшихся носовых пазух. Их по три на каждой
стороне, и они распределяют воздух, закачанный под давлением ampullae
choanales. Canales ramosi самих пазух пока имеют ещё одно orificium
externum [наружное отверстие – В. П.] близ кончика рыла; оно большей
частью бывает закрыто, но может очень быстро открываться рефлекторно, если на
рыло оказаны сильные механические воздействия, поэтому последнее может
мгновенно сжаться. Упомянутая система иннервирована N. trigeminus,
тогда как N. facialis обслуживает прежде всего периназальную кольцевую
и продольную мускулатуру. Подобно всем многорылам, Nasobema также
лишена os nasale; она даже не закладывается у эмбриона.
Большая
моргенштернова носоходка, хонатата туземцев (Nasobema lyricum; таблица
X), является лучше всего известным представителем многорылов, и этот пример
будет обсуждаться с несколько большими подробностями. Будучи типичным
многорылом, она несёт на своей короткой толстой голове четыре рыла равной
длины, которые довольно длинные, и на которых – как уже описанно Моргенштерном
– она ходит. Она может это делать, несмотря на несответствующий носовой скелет,
потому что рыла становятся весьма твёрдыми благодаря сильному тургору их corpora
spongiosa. Кроме того, в рыла проникают сильно разветвлённые воздушные
ходы, заполнение которых регулируется с помощью ampullae choanales (образования,
дифференцировавшиеся из мягкого нёба по краям носовых раковин, которые
простираются далеко в вентро-каудальном направлении), поэтому тургор рыла обеспечивается
двумя системами: гидравлической системой corpora spongiosa, которая
ответственна прежде всего за долговременное сохранение жёсткости, необходимой
для ходьбы, и пневматической системой, которая обеспечивает походку и
пружинистые движения, что уменьшает опасность повреждения, когда неожиданно
возникают твёрдые препятствия. В дополнение к роли, которую играют ampullae
choanales, в этом принимают участие также ampullae pneumonasales,
которые образовались из сильно разросшихся носовых пазух. Их по три на каждой
стороне, и они распределяют воздух, закачанный под давлением ampullae
choanales. Canales ramosi самих пазух пока имеют ещё одно orificium
externum [наружное отверстие – В. П.] близ кончика рыла; оно большей
частью бывает закрыто, но может очень быстро открываться рефлекторно, если на
рыло оказаны сильные механические воздействия, поэтому последнее может
мгновенно сжаться. Упомянутая система иннервирована N. trigeminus,
тогда как N. facialis обслуживает прежде всего периназальную кольцевую
и продольную мускулатуру. Подобно всем многорылам, Nasobema также
лишена os nasale; она даже не закладывается у эмбриона.

Nasobema lyricum с сайта Токивы Такеши
Парные конечности относительно хорошо развиты. Очень молодые животные
демонстрируют лишь небольшую их редукцию. У более старших особей, то есть, у
таких, которые достигли 2/3 максимальной длины тела, задние конечности
практически неподвижны и также не функционируют. Передние конечности – это
хватательные органы, которые эффективно дополнены длинным, лассоподобным
хвостом.
Хвост в высшей степени специализирован, и его организация может быть понята
только по отношению к повадкам носоходок. Он служит этим фруктоядным животным,
чтобы доставать их пищу с большой высоты. Способ, которым это происходит,
заключается в том, что хвост, лишь проксимальная часть которого пронизана
позвоночным столбом, содержит наполненный газом канал, который связан со слепой
кишкой; поэтому, после того, как расслабляется sphincter gasotubalis,
канал может быстро заполниться кишечными газами, и из-за этого хвост резко
надувается и взлетает вверх на высоту более четырёх ярдов. Благодаря очень
мускулистой ampulla (поперечно-полосатые волокна, производные от
подвздошно-хвостовой мускулатуры) в основании хвоста это происходит с такой
силой, что в мгновение ока хвост выпрямляется вверх на всю длину с тихим
хлопком. Как только кончик хвоста, богато снабжённый осязательными сосочками,
захватывает плод, газ выпускается из хвоста с нежным свистом, он снова
становится уплощенной лентой и сокращается. Затем сорванный плод схватывается
передними лапами и подносится ко рту. Интересно, что выделение газа в целом
достаточно хорошо адаптировано к этому механизму: чем голоднее животное, тем
более сильно раздута его толстая кишка и, соответственно, ampulla
gasomotorica. С этим связан тот факт, что очень голодные животные, даже
когда в поле зрения нет ничего подходящего, выполняют действия по добыче плодов
«вхолостую» или выбрасывают хвост вверх к объектам любой природы, которые
напоминают плоды. Это особенно заметно у Nasobema aeolus40
(голубоносой носоходки).
Один раз в год Nasobema рождает одного детёныша, который сначала
вынашивается в горловой сумке, которая открывается в каудальном направлении, и
кормится из подмышечных сосков матери. Горловая сумка имеется только у самок и
поддерживается хрящами, происходящими из хрящей кадыка. Животные моногамны,
представители обоих полов очень нежны друг к другу. Самку после родов кормит
самец. Только на самых больших островах архипелага у носоходок есть враги в
лице хищных носоходок (Tyrannonasus imperator B. d. B. = Nasobema
tyrannonasus STU.), которые водятся там. Следует отметить, что
Хууака-Хатчи во время своих празднеств на равноденствие, которые происходили
каждую осень и весну, ели на этих ритуальных пиршествах носоходок, жареных в
листьях. Они считали животное священным и не охотились на него, кроме случаев,
связанных с этими религиозными обрядами.
Род Stella был установлен Бромьеном. Штультен всё ещё относит Stella
matutina41 B. d. B. к Nasobema, называя её Nasobema
morgensternii. Различия между двумя родами действительно невелики и имеют
отношение к механизму выбрасывания хвоста, который значительно менее
дифференцирован у мелкой моргенштерновой носоходки; эта ситуация, несомненно,
связана с тем фактом, что Stella matutina питается почти исключительно
ягодами, которые растут около земли.
В отличие от миролюбивых носоходок, единственный ныне живущий представитель
тиранноназид – их хищный родич, который питается почти исключительно ими. По
своему строению хищная носоходка Херберера, Tyrannonasus imperator B.
d. B. (= Nasobema tyrannonasus Stulten), действительно во всех
основных отношениях напоминает прочих носоходок, но отличается развитием
хвоста, который, как и у столбоносов – несёт на кончике ядовитый коготь; и
кроме того, конечно, своими хищными зубами, острые кончики которых предназначены
для того, чтобы разрывать плоть жертвы. В дополнение к этому примечателен тот
факт, что задние конечности развиты удивительно хорошо для носоходящего вида,
что связано с их участием в схватывании добычи. И в конце можно упомянуть, что
шерсть этого вида не текстурирована и создает впечатление плюшевой, несколько
напоминающей шерсть крота.
Tyrannonasus imperator особенно примечателен по двум причинам: подобно
всем видам многорылов, животное передвигается на носу не слишком быстро, но всё
равно движется более быстрым шагом, чем носоходки. Но, поскольку все виды
многорылов из-за их внутрирыльного пневматического аппарата во время ходьбы
издают свистящее шипение, которое можно услышать издалека, Tyrannonasus
не может тихо подползти к своей жертвы; но – поскольку они спасаются бегством,
когда он находится на расстоянии – он должен сначала тихо лежать в засаде, а
затем быстрым шагом настигать их. В процессе этой погони, которая из-за
производимого сильного шума, но, увы, такой малой скорости, создаёт у наблюдателя
комическое впечатление, Tyrannonasus часто должен часами преследовать
намеченную жертву, чтобы догнать её, в то время как Nasobema, спасаясь
бегством, также использует хвост, выбрасывая его вверх, обвивая им ветки и
таким способом преодолевая канавы или маленькие ручьи. Даже когда хищник
вплотную приблизился к объекту своего преследования, Nasobema также
часто успешно использует хвост как последнее средство спасения: подвешиваясь
хвостом на ветке, она раскачивается над землёй взад-вперёд круговыми или
широкими маятникообразными движениями, пока хищник в своих постоянных попытках
схватить добычу, не испытает в конце концов головокружение и не отступит.
|
|
Тогда, во время этого периода дезориентации хищника, Nasobema часто
находит возможность убежать.
Но если Tyrannonasus смог захватить свою добычу, у последней нет
никаких дальнейших надежд на спасение: она отравлена с помощью ядовитого когтя
и вскоре
гибнет в страданиях, пока хищник наносит ей смертельный удар, тащит её в
тенистое место и там пожирает её в тишине, оставляя лишь крупные кости. Тогда
как наиболее поразительной особенностью Tyrannonasus является его
высокая выносливость в преследовании добычи, вторая особенность – это
дополнительная способность к быстрому движению в течение необычно долгого для
млекопитающего времени. Эта способность связана и с удивительно низким уровнем
метаболизма в целом, и со способностью запасать гликоген не только в печени,
но и в подкожных депо. С точки зрения гистологии последние сложены из производных
тех же самых эмбриональных клеток, какие дают начало жировой ткани, которая
занимает это местоположение у других организмов. Запасание энергии в форме
гликогена,
а не жира, выглядит более экономичным, по крайней мере, у Tyrannonasus.
Поскольку этот процесс происходит после питания, тело сытого животного в
течение пары дней весьма заметно деформируется; сразу после обильной трапезы
животное залегает в месте, защищённом от дождя, и дремлет там, пока подкожные
депо гликогена не исчерпываются, на что уходит несколько недель. Только после
того, как животное снова становится стройным, но всё ещё сохраняет в печени
достаточные запасы для будущей охоты, оно снова становится активным и выходит
на охоту.
Тот примечательный факт, что в неволе носоходка плачет, имеет психологический
интерес, поскольку это предполагает, что животное обладает пониманием и
способностью к отражению. Если принять во внимание значительные размеры и
степень дифференциации мозга, такая возможность не исключена (см. в связи с
этим Х. В. Грале 1947).
|
|
|
|
|
|
Tyrannonasus imperator с сайта Токивы Такеши
По физиологии питания Tetrarrhinida (четырёхрылые носоходки) полностью
выпадают из пределов нормы для отряда и в этом отношении, несомненно, являются
вторично дивергировавшими формами. По Бюффону (1953) дело здесь в следующем:
Rhinogradentia – исходно насекомоядны по своему поведению, и в целом небольшой
размер животных соотносится с этим. В случаях, когда происходила специализация
поведения и способа питания, которое отличает животных от типичного
насекомоядного типа, они, тем не менее, должны быть его производными; все
крабоядные Hopsorrhinidae (мордоскоки) и их симбиотические формы, питающиеся
молоком близки к этому насекомоядному типу, и аналогичным образом Hypogeonasidae
(иловые носоходки) и Georrhinidae (роющие носоходки) могут выводиться из них. В
случае четырёхрылых носоходок, чьи более примитивные представители
исключительным образом фруктоядные, такое установление связей выглядит более
трудным, хотя, например, зубная система в главных чертах имеет полностью
насекомоядный тип. Помимо всего остального, пищеварительный трактат
демонстрирует обширную специализацию, не говоря уже о слепой кишке, выделяющей
газ. Но главный фактор, который отличает четырёхрылых носоходок от большинства
других Rhinogradentia – значительный размер тела. Носоходки достигают почти 3
футов в высоту! Происхождение хищной носоходки Tyrannonasus на первый
взгляд кажется более простым, поскольку, по-видимому, простое увеличение во
всех размерах превратило бы насекомоядное существо в хищника. Но Бюффон,
посредством вскрытия показал, что Tyrannonasus происходит от
фруктоядного вида. Это видно прежде всего в устройстве пищеварительного тракта
и хвоста, который на стадии половой зрелости всё ещё сохраняет признаки,
характерные для носоходок. Бюффон считает, что преобразование в хищника – и
фактически в хищника-монофага – должна пройти через стадию хищнического
комменсализма. Некоторые особенности в поведении хищника дают дальнейшие
указания в этом направлении, а именно: он жадно пожирает плоды, брошенные
убегающими носоходками, и только тогда атакует последних, когда он может
броситься на них, пока они едят плоды. С этим согласуется тот факт, что молодые
особи Tyrannonasus не хищничают, но просто нападают на кормящихся
носоходок, чтобы отнять у них плоды, или же кормятся остатками их пищи.
Это явление, как также указывает Бюффон, ни в коем случае не уникально в мире
животных; переход от насекомоядного к фруктоядному поведению часто наблюдался,
например, среди родственных дроздам певчих птиц, у самих насекомоядных, а также
у обитающих в Южной Америке рукокрылых, полуобезьян и широконосых обезьян.
40 Aeolus Греч. = бог ветра [Эол – В. П.].
41 Stella matutina Лат. = Morgenstern (= утренняя звезда).
ТРИБА: Hexarrhinida (Шестирылые
носоходки в широком смысле),
СЕМЕЙСТВО: Isorrhinidae (Равнорылые носоходки),
РОД: Eledonopsis42 (Лентонос),
5 видов,
РОД: Hexanthus (Шестицветковые носоходки),
3 вида,
РОД: Cephalanthus43 (Цветолицая носоходка),
7 видов,
СЕМЕЙСТВО: Anisorrhinidae* (Неравнорылые носоходки),
РОД: Mammontops44 (Мохнатолицая носоходка),
1 вид
Триба Hexarrhinida (шестирылые носоходки в широком смысле) включает два
сильно различающихся семейства; тогда как равнорылые носоходки (Isorrhinidae) –
это мелкие насекомоядные животные относительно примитивной организации,
единственный вид в семействе неравнорылых носоходок (Anisorrhinidae) обладает
типом строения, который на первый взгляд кажется гораздо более напоминающим
собственно носоходок, однако наряду с этим демонстрирует целый ряд
особенностей, которые отделяют его от этого семейства. Поэтому Бюффон считает,
что триба Hexarrhinida, выделенная Бромьеном де Бурла, не может быть принята,
или что она является полифилетической. Это будет рассматриваться далее, при
обсуждении Mammontops ursulus45.
Равнорылые носоходки, как уже говорилось, это животные, которые – помимо их
полиринии – должны считаться примитивными; парные конечности находятся на
ранней стадии редукции и всё ещё хорошо приспособлены к передвижению, даже при
том, что животные лишь немного используют их. Дифференциация рыла аналогичным
образом всё ещё примитивна*. С другой стороны, прогрессивные роды Hexanthus
и Cephalanthus Br. d. B. (= Ranunculonasus и Corbulonasus
STU.) отличаются высокоразвитым проявлением миметизма, который изменил
внешность этих животных весьма специфическим способом. [Примечание переводчика:
Br. d. B. – это просто B. d. B. в других местах.]
В качестве представителя более примитивного рода Eledonopsis будет
описан Eledonopsis terebellum (червеносый лентонос).
В маленьких норах в земле, под камнями и корнями, на Майрувили часто
встречается маленькое животное размером с землеройку, которая спит там,
свернувшись клубочком, в течение дня и на первый взгляд выглядит совсем как
небольшая землеройка, с серовато-коричневым мехом и розовыми лапками. Маленькое
существо не пытается убежать и позволяет себе оставаться в своём логове. Если
отметить положение такой норы и ночью сделать с помощью вспышки фотографию её
входа и окружения, то на картинке можно заметить, что в стороны от входа
тянется от четырёх до шести лентообразных структур. Эти окрашенные в розовый
цвет ленты шириной около 2 – 3 мм и длиной около фута. На верхней поверхности
они несут два узких, влажно поблёскивающих углубления, к которым приклеиваются,
как можно заметить, некоторые мельчайшие насекомые, главным образом подуры
(ногохвостки) и сеноеды (Corrodentia). Если попробовать рассмотреть эти
структуры поближе с помощью прожектора, то они быстро втягиваются назад в нору.
На протяжении долгого времени это явление не предоставляло возможности для
более точного наблюдения. На заспиртованных образцах ленты действительно были
без дальнейших разговоров идентифицированы как рыла, но их функция стала
объяснимой только, когда животных держали при постоянном освещении (метод
Шаллера). Теперь стало очевидно, что фактически ленты – это рыла, а эти две
канавки – растянутые ноздри, которые развёрнуты вверх; кроме того, было обнаружено,
что ресничный эпителий носовых полостей протягивается вдоль ноздрей, и что
вместе с носовой слизью он используется, чтобы транспортировать мелких
насекомых, которые приклеились к нему; они засасывались в носовые проходы и
через хоаны попадали в пищеварительный тракт. Кроме того, они развились таким
образом, что Eledonopsis также может поймать в ловушку более крупных
насекомых – вплоть до размера тли – и перемещать их таким способом к себе,
тогда как лентовидное рыло сокращается и образует жёлоб, по которому добыча
доставляется к основанию рыла, отчасти с помощью тока, образуемого ресничками и
отчасти посредством перистальтических движений желобка. Там добыча или
захватывается языком, или выбирается передними лапами из носовых частей,
прилегающих к голове, и поедается. Также интересно то, что большие, слишком
крупные для этого способа перемещения животные – прежде всего, пауки семейств
Heieiatypidae и Lycosodromidae – вначале попадаются в носовую слизь, а затем
оплетаются несколькими рылами и подтягиваются к голове. Решение относительно
того, когда и какие механизмы ловли добычи приводить в действие, принимается
отчасти на основании осязательных ощущений, а затем также на основании
химического чувства, которое, несомненно, распространяется до кончиков рыл.
(Это задача рецепторов и нервов, которые образуют такие же отношения к
непосредственному обонятельному органу, как якобсонов орган у многих других
позвоночных.)
|
|
У Eledonopsis забота о потомстве ничем не отличается от таковой у
других Placentalia. Выводковой сумки нет. Молодняк становится независимым в
очень раннем возрасте. Спаривание, очевидно, происходит ночью. До настоящего
времени попытки вырастить Eledonopsis в неволе оказывались неудачными.
Поведение молодых экземпляров шестицветковых носоходок (Hexanthus)
очень похоже на поведение Eledonopsis. Они также живут в норах под
землёй или под листьями и оттуда вытягивают свои рыла, чтобы ловить добычу. Но
это справедливо лишь в отношении самых молодых животных, которые только что
начали добывать пищу самостоятельно, освободившись от материнской заботы. Позже
проявляются следующие отличия от Eledonopsis: носовая канавка
срастается в проксимально-дистальном направлении таким образом, что в конечном
счёте отверстия остаются только у основания и на кончике рыла, тогда как
остальная часть рыла образует закрытую трубку. Затем на кончике каждого рыла
вырастают четыре широких, заострённых дермальных лопасти. В зависимости от вида
они окрашены по-разному и способны, кроме того, довольно сильно изменять цвета
в пределах, установленных характерной для вида основной окраской46.
В конечном счёте, каждое рыло становится похожим на цветок на длинном стебле. В
процессе этого изменения своих рыл животные вообще едва меняют свои повадки.
Они продолжают протягивать свои рыла дальше от места, где прячутся, но обвивают
ими выше и выше стебли растений47, и добывают свой корм так, как уже
было сказано выше. При всём этом самое большое изменение произошло в рационе. С
этого момента они ловят главным образом летающих насекомых, которые, обманутые
цветоподобной формой и окраской, садятся на кончики рыл. Перемещение мелких
жертв происходит способом, уже описанным выше. Также, как объяснялось выше,
более крупные экземпляры переносятся в сторону головы перистальтическими
движениями в носовых трубках. Однако ещё более крупные объекты, которые не
могут пройти сквозь них, не добываются целиком – в отличие от Eledonopsis.
Вместо этого сильно растяжимые, похожие на венчик открывающиеся лопасти рыл
складываются вокруг насекомого так, чтобы оно фактически оказалось полностью
завёрнутым в них. Вслед за этим Hexanthus отрыгивает [пищеварительный
сок – В. П.] в соответствующее рыло и переваривает экземпляр до состояния,
когда он может быть высосан через носовую трубку.
Зрелые особи Hexanthus более не скрываются в норах; они скорее лежат
на скалистых склонах среди зелени травянистых участков и низкорослых трав. Их
собственная зеленоватая окраска делает их незаметными в тех местах, а их рыла
почти всё время обвивают стебли тех цветов, чьи цвет и форму они могут
имитировать. Кроме того, эта адаптация содержит визуальный элемент; если перед Hexanthus
поставить синие цветки, приклеенные к листу картона, а за ним рыла находятся
среди жёлтых цветов, то носовые цветкоподобные лопасти станут синими, и
наоборот. Кроме того, различные виды Hexanthus обладают различным
фотопериодическим поведением; так, круглоцветковая носоходка (Hexanthus
ranunculonasus = Ranunculonasus pulcher48) полностью
дневная, а большей частью фиолетовые рыла носоходки «царица ночи» (Hexanthus
regma-noctis49) расцветают главным образом ночью. В отличие от
носоходок рода Ranunculonasus, которые обладают просто кисловатым
запахом, похожим на запах кислого молока, ночные носоходки испускают сильный
ванильный аромат, который привлекает ночных шестикрылых насекомых.
|
|
К роду цветолицых носоходок (Cephalanthus = Corbulonasus)
принадлежит целый ряд самых красивых Rhinogradentia из ныне известных.
Все они отличаются тем, что короткие, широкие, похожие на лепесток рыла сидят
близко ко рту и снабжены очень простой эпи- и гипоназальной мускулатурой,
которая позволяет животным очень быстро сжимать вместе тонически раскрытые рыла
всякий раз, когда насекомое садится в ротовой области.
Ещё одна особенность – эти очень ограниченные в умственном плане животные
испускают сильные запахи изо рта, что, несомненно, также служит для привлечения
насекомых. Помимо этого, странной особенностью является то, что, в отличие от
других многорылов, представители этого рода никак не заботятся о своём
потомстве и не кормят его молоком.
В качестве типичного представителя рода мы выбрали чудесную цветолицую
носоходку (Cephalanthus thaumasios50 = Corbulonasus
longicauda51), которая колониями живёт на Митадине на лугах
из Ranunculaceae в возвышенных горных областях. Скамтквист описывает облик
таких
колоний как самую красивую вещь, которую он видел на Хай-яй-яй. Яркость
расцветок и блеск рыл описывают как что-то весьма экстраординарное, а
характерный вид животных, покачивающихся на своих хвостах в порывах свежего
морского бриза, просто зачаровывает. Вполне очевидно, то, что доставляет нам
наибольшее удовольствие в этих любопытных существах, c точки зрения насекомых,
разыскивающих цветы, представляет собой не более, чем демонстрацию сверхэффективных
стимулов; в этом же ключе
также должен быть
истолкован похожий на молочную сыворотку запах, который исходит из открытых
ртов прячущихся цветолицых носоходок.
|
|
В дополнение к nasarium’у, который уже обсуждался, одна из их поразительных особенностей – жёсткий хвост, который может достигать целых двадцати дюймов в длину. Интерес представляет то, как меняется строение хвоста, когда животное растёт; новорождённые животные, которые уже обладают полностью развитым nasarium’ом, падают на землю и поднимаются вверх по стеблям растущих рядом цветов. Добравшись до верхушки, они откусывают все почки, разворачивают nasarium и начинают ловить добычу так же, как делают более старшие особи. Их всё ещё мягкий хвост на тот момент примерно равен длине тела и ни в каком отношении не отличается от нормального хвоста млекопитающего. Но он быстро растёт в длину путём удлинения позвонков; поскольку межпозвонковые сочленения окостеневают, а межпозвонковые сухожилия и связки также становятся жёстче, хвостовая мускулатура дегенерирует, поэтому от M. ischiocaudalis, M. iliocaudails и M. depressor caudae остаются только связки, которые прикрепляются к хвостовому отделу позвоночника и к его отвердевшим объединяющим структурам. На кончике хвоста имеется большое количество ороговевшего эпителия, который в конечном счёте образует своего рода заострённый вилкообразный роговой наконечник. Как только кончик хвоста достигает земли, животное, которое забралось по стеблю растения, начинает бурить этим хвостовым наконечником землю, в которую через четыре – шесть дней он проникает на глубину целых 6 дюймов. Тогда животное отпускает стебель растения и после этого держится только на своём собственном хвосте, который продолжает удлиняться. Степень увеличения длины хвоста зависит от снабжения животного пищей и происходит медленнее, когда последнее хорошо питается. Животное, однажды крепко «посаженное» таким образом, больше не может изменить своё местоположение, и просто ждёт добычу со сложенными руками и открытым ртом. Как уже говорилось, умственные способности невелики. Спаривание происходит в ветреную погоду; когда особи раскачиваются взад-вперёд на своих хвостах и соприкасаются, ищущие встречи самцы крепко вцепляются в самок. Отмечается, что период беременности длится всего лишь три недели, а полная продолжительность жизни оценивается максимально в восемь месяцев. Рост от рождения до зрелости составляет, возможно, два месяца; от рождения до внедрения в землю хвостом проходит от восемнадцати до двадцати двух дней.

Cephalanthus с сайта Токивы Такеши
Нередко можно найти колонии, которые производят жалкое впечатление;
рыла выглядят слабыми, обесцвечиваются и покрываются корочкой. Существа худеют,
а их
тихое хныканье слышно издалека. Такие колонии были поражены носовой чесоткой,
которая вызвана видом клеща, который близкородствен гамазовым клещам. Небольшое
заражение едва ли причиняет значимый вред. Но когда массовое размножение клещей
делает рыла неэффективными для поимки добычи, это, естественно, становится
катастрофой для Cephalanthus. Голодные, измученные существа постоянно
ковыряются в своих инфицированных рылах и таким образом просто усиливают свои
страдания. В конце концов остаются только крошечные трупы, висящие на длинных
хвостовых стеблях; и во многих местах можно найти торчащими на лугах группы от
шестидесяти до сотни скелетированных хвостов, под которыми лежат только
разлагающиеся остатки костей и шкурок. Однако, изначальная причина эпизоотий
–
не местные клещи, а скорее вирусные болезни, развитию которых
благоприятствовали изменения погоды; они снижают естественную сопротивляемость
клещам, и при этом особи, страдающие от вирусной болезни, не могут оказывать
постоянный уход своим рылам, поддерживая их должным образом увлажнёнными.
Большинство видов Cephalanthus живёт описанным образом. Только Cephalanthus
ineps52 (= Corbulonasus ineps) и Cephalanthus
piger53 (= Corbulonasus acaulis54) обладают
редуцированными хвостами и просто лежат на своих спинах на солнечных местах
среди камней и цветов. Согласно взглядам Бромьена де Бурла, их выделение из
прочих цветолицых носоходок в качестве нового рода уже оценивалось и не
выглядит оправданным.
|
|
Как уже упоминалось на стр. 66, неравнорылая носоходка Mammontops
ursulus (Медведеобразная мохнатолицая носоходка), которая также
встречается на горных лугах Митадины, в значительной степени выпадает из рядов
шестирылых носоходок. Она представляет собой довольно внушительное животное,
которое достигает полной высоты 4 футов 3 дюймов у самцов и 3 футов 7 дюймов у
самок; и она травоядна.
Её рыла дифференцированы несколько иным образом, нежели у четырёхрылых
носоходок, и это причина неуверенности в отношении её точного систематического
положения; тогда как Штультен склонен помещать Mammontops в
непосредственной близости от четырёхрылых носоходок, в пользу чего он считает
более весомым аргументом строение рыл, нежели их количество, Бромьен де Бурла
придерживается мнения, что число рыл заслуживает большего внимания для целей
систематики, тогда как степень дифференциации рыл должна относиться на счёт
конвергенции. В пользу этой точки зрения он привлекает исследования группы
французских ученых (Бюффон, Ирри-Эгингарри и Шабли), которые показали, что
иннервация нескольких групп мускулов у четырёхрылых весьма отличается от
таковой у Mammontops. Здесь она кажется производным дальнейшей
дифференцировки эпи- и гипоринальных мускулов равнорылых носоходок; а отсутствие
corpora spongiosa, характерных для рыл современных равноносых, больше
не кажется таким примитивным, как считалось некогда. Бюффон и Гокари-Судур
выдвигают для шестирылых носоходок общих четырёхрылоподобных предков, из
которых с одной стороны развились равнорылые носоходки наших дней, а с другой –
неравнорылые. В этой связи, однако, любопытен тот факт, что равнорылые
показывают явно выраженные примитивные особенности, касающиеся их парных
конечностей, в то время как в линии неравнорылых они особенно сильно
редуцированы. Кроме того, примечательно, что завитки волос, которые характерны
для многорылых носоходок, отсутствуют у равнорылых. Будучи противником того,
чтобы уделять слишком большое внимание этой особенности, Бромьен де Бурла
выдвигает точку зрения, что даже явно вторично эволюционировавший Orchidiopsis
не показывает инверсию направления роста волос, хотя такая инверсия наиболее
заметна у гораздо более примитивных мордоскоков. В любом случае проблему
положения Mammontops в настоящее время следует оставить открытой, пока
не будут выполнены более обширные и детальные исследования.
Mammontops живёт маленькими стадами, во главе которых стоят старые
самцы. Животные кормятся почти исключительно сложноцветным растением Mammontopsisitos
dauciradix55, которое они выдёргивают с корнем своими двумя
хватательными рылами. Зубная система демонстрирует наиболее высокую
специализацию, известную среди Rhinogradentia (если не считать полной потери
зубов у сосущих мордоскоков); резцы редуцированы, клыки маленькие и притупленные,
предкоренные и коренные зубы расширены и похожи на пластины.
Мохнатолицая носоходка кормит молоком своего детёныша, который цепляется своими
рылами за густую шерсть матери, а также за её жесткие соски, расположенные в
паховой области. Темп размножения низкий. Животные, очевидно, доживают до
глубокой старости. Более взрослые самцы отличаются от однотонных
шоколадно-коричневых молодых самцов и самок серебристо-серым хвостом, взмахи
которого вызывают ответную реакцию со стороны стада. Например, Тассино ди
Кампотасси удалось, побелив хвост молодой самки, вызывать ответную реакцию
стада, в которое он её помещал. Побеленный хвост был особенно эффективным в
качестве супернормального стимула ответной реакции у более молодых самцов.
*но см. страницу 76.
42 eledone Греч. = животное, родственное осьминогу [название одного из родов осьминогов – В. П.].
43 kephale Греч. = голова; anthos Греч. = цветок.
*Примечание переводчика в отношении Anisorrhinidae; большинство обстоятельств говорит в пользу варианта названия Mammontopsidae, так как Mammontops – это единственный род; однако есть ситуации, в которых правила номенклатуры допустили бы название семейства Anisorrhinidae.
44 см. сноску 15.
45 ursulus Лат. = маленький медведь.
46 Фреддуриста и Периччерци показали, что красные цвета регулируются расширением капилляров, жёлтые лежащей у поверхности жировой тканью (которая находится, однако, под субэпителиальной капиллярной сетью), а голубые чёрным пигментом, расположенным в сократимых меланофорах.
47 постоянно влево, все рыла как с правой стороны тела, так и с левой (см. Людвиг 1932).
48 ranunculus Лат. = лютик (цветок); pulcher Лат. = красивый.
49 regina noctis Лат. = королева ночи
50 thaumasios Греч. = изумительный, редкий.
51 longi-cauda Лат. = длиннохвостый.
52 ineps Лат. = тугодум.
53 piger Лат. = ленивый.
54 akaulos Греч. = бесстебельный.
55 sitos Греч. = пища; dauci-radix Лат. = корень, похожий на морковь.
КОГОРТА: Dolichoproata (Длинноносые
носоходки),
СЕМЕЙСТВО: Rhinochilopidae56 (Носоходки-многоноски),
РОД: Rhinochilopus (Многоноски [англ. “Tasselsnouters” – В. П.]*),
2 вида.
Род Rhinochilopus со своими двумя видами, Rh. ingens57 (Гигантская многоноска) и Rh. musicus (Органная многоноска), демонстрирует наиболее впечатляющую полиринию: у обоих этих животных голова вытянута в длинную proa, или рострум. Нижняя сторона этого образования поддерживается верхнечелюстными, предчелюстными и нёбными костями, верхняя – верхнечелюстными, предчелюстными и рыльными, а также частью носовой кости. Нижняя сторона (см. рис. 12) демонстрирует удлинение ротовой щели, так называемый желобок proa (2), который достигает губ. На переднем конце proa у самца растут два асимметричных резца. Справа и слева от этого желобка proa находится девятнадцать пар рыл, называемых здесь назидиями [букв. «рыльцами» – В. П.] (3, или 9). Первая пара служит в качестве усиков, остальные – как органы передвижения. (О более детальном строении nasarium’а, см. ниже.) Парные конечности в значительной степени редуцированы. Задние ноги функционируют исключительно как антенны, используемые при движении назад. Передние лапы не касаются земли, а также не играют никакой роли при сборе пищи. У самок они служат для того, чтобы держать детёныша (всегда только одного). Хвост – это также не больше, чем орган осязания. Животные достигают внушительного размера (у Rh. musicus почти 5 футов от proa до основания хвоста, у Rh. ingens более 7 футов). Они всеядны, но предпочитают насекомых, улиток и грибы, а также ягодообразные плоды. Иногда они также поедают молодые листья. Rhinochilopus – одиночное животное, неторопливо бродящее по девственному лесу, особенно по менее густым участкам и по краю леса, где у него есть определённые маршруты и протоптанные тропинки. Однако у животных нет постоянных участков, и они беспрепятственно используют тропы сородичей.
|
[Рис. 12] Rhinochilopus musicus; эмбрион на поздних сроках развития. 1. Зачатки резцов (только у самцов); 2. Субпроальный желобок; 3. Ductulus musicus одного удалённого рыльца; 4. Ductulus osmaticus, аналогично; 5. Corpus spongiosum рылец; 6. Ductus musicus; 7. Слёзный проток; 8. Vesica inflatrix organi; 9. Рыльца. Медиальнее расположенный ductus osmaticus не пронумерован. Обратите внимание на отличающееся строение первого рыльца, заметное даже у эмбриона. (По Бюффону и Гокари-Судуру 1952) |
Однако наиболее примечательной особенностью этих двух видов, в особенности Rh. musicus, является процедура ухаживания и соответствующая специализация рылец, которую вкратце опишем: как и большинство Rhinogradertia, Rhinochilopus больше не дышит исключительно через ноздри, но большей частью через слёзный проток, который соответствующим образом значительно расширен, как это имеет место почти у всех видов (см. также рис. 4). Теперь с одной стороны слёзный проток (рис. 12, 7) связан непосредственно с глоткой, от которой проток – ductus osmaticus – входит в proa и там разделяется на ductuli osmatici (4) в рыльцах. С другой стороны, слёзный проток также связан посредством ductus inflatorius с vesica inflatrix organi (8), который со своей стороны соединяется посредством ductus vesico-gularis с горлом. Вторую связь (6) между слёзным протоком и vesica inflatrix organi и рыльцами поддерживает ductus musicus через ductuli musici, которые расположены в рыльцах латеральнее ductuli osmatici. В покое vesica inflatrix organi лежит под кожей щеки, и при активации носового органного аппарата раздувается до размера детской головы. Аппарат в целом представляет собой дифференцированную часть слёзного протока и хоанальной области носа, что может быть различимо у ранних зародышей. В качестве дополнительных компонентов органный аппарат включает corpora spongiosa (5) из рылец, а также кольцевую и продольную мускулатуру рылец.

Rhinochilopus с сайта Токивы Такеши
В целом аппарат работает следующим образом: во время передвижения ductus musici и ductuli musici закрыты и вместе с corpora spongiosa придают рыльцам необходимый тургор, достаточный для того, чтобы кольцевая и продольная мускулатура могли двигаться. Во время вдоха воздух проникает через ductuli osmatici. Так воздух постоянно анализируется. Первая пара рылец, работающая как усики, особенно тесно связана с чувством обоняния. Направленное обонятельное исследование стало возможным благодаря разветвлению обонятельного нерва внутри proa таким образом, что каждая пара рылец обладает собственным обонятельным эпителием. Пища захватывается рыльцами: схватывается пальцеобразными выростами кончика рыльца, или окружается дистальной третью всего рыльца и затем переносится в желобок proa. Затем добыча перемещается в сторону рта сильно вытянутыми вперёд губами. Музыкальный аппарат nasarium’а используется только в период ухаживания. В это время самцы лежат пластом на земле. Ductuli osmatici прекращают функционировать, так же, как и corpora spongiosa. Вначале мускулатура nasarium’а полностью расслабляется. Только сфинктеры, которыми завершаются ductuli musici, слегка сокращаются. Sphincteres glossiformes, расположенные в основании каждого ductulus musicus, расслаблены. Потом с помощью интенсивного непрерывного дыхания животное раздувает vesicae inflatores organi, а также нагнетает давление в ductus musici. Когда приоткрываются sphincteres glossiformes, рыльца оказываются раздутыми, и хотя кольцевая мускулатура полностью расслаблена, продольная мускулатура может сделать рыльце длиннее или короче. Если теперь sphincter glossiformis внезапно откроется шире, короткие дуновения воздуха проходят в соответствующее рыльце и заставляют его отражать звук, поскольку вибрацию воздушного столба, который в нём находится, вызывают края сфинктера. Так как каждое рыльце может удлиняться или укорачиваться, быстро чередуя эти состояния, оно работает как духовой инструмент по принципу игры на тромбоне с той особенностью, что не могут проигрываться длительные тоны, а только последовательности коротких. Благодаря восемнадцати парам рылец, которые работают таким образом (первая пара лишена sphincter glossiformis58), животное, если можно так выразиться, снабжено оркестром из 36 духовых инструментов, которые могут работать независимо. То, как они используются в ухаживании, было живо описано Скамтквистом:
«В это время – это было весеннее равноденствие – Хууака-Хатчи праздновали пир хонататы, на котором подавались копчёные хонататы, под аккомпанемент ритуальных песнопений в деревенском длинном доме. Это происходило вечером, в сумерках. Церемониальная трапеза продолжалась не более двух часов. Затем деревенская компания поднялась и направилась прямо к ближайшей лесной поляне, на западном краю которой они все расселись. Полная луна уже виднелась над вершинами деревьев на горе напротив, когда Мостада Датсавима («Владыки многоножек») показались из лесной темноты и вышли на поляну. Большие животные перемещались бесшумно, словно плыли. В призрачном лунном свете их ноги (рыла) были неясно различимы. Был различим только блеск длинных морд и спин. Всего было четырнадцать или шестнадцать животных, которые сначала шествовали по кругу пару раз, прежде чем шесть особенно огромных самцов легли на землю и вытянули все свои рыла, пока самки продолжали кружиться вокруг них. И затем там начался самый замечательный концерт, который я когда-либо слышал: он начался с глухого, ритмичного мычания одного из животных. Вначале оно было медленным, но затем стало быстрее. Вскоре к нему присоединился второй, мыча на несколько нот выше; наконец в этом приняли участие все шестеро. Ритм менялся, но строго синхронно поддерживался всеми животными, в то время как число участвующих голосов росло и росло. Внезапно настала тишина, а затем резкое, блеющее тремоло, многоголосое и prestissimo, перекрыло этот ритмичный, частью урчащий, мычащий или мурлыкающий глухой шум. Это составило вторую часть. Временами это «соло» одного самца, сконцентрированного на его исполнении, усиливалось вплетением в него, в придачу к блеянию и пассажам «стаккато», нечётких и размытых тональных переходов. Животное, которое исполняло сольную партию, можно было отличить по тому, что его рыла, которые сейчас без труда различались, потому что они были вытянуты в одну сторону, поочерёдно раздувались и становились короче или длиннее. Внезапно снова настала тишина; затем низкий основной фон всего музыкального хора заиграл ещё раз, после чего начал солировать второй самец. В течение этого «концерта» самки кружились в медленном, правильном темпе вокруг поющих самцов, пока последний из них не закончил свою сольную партию. Затем самцы поднялись, и вся призрачная компания исчезла так же медленно, как они появились, в тёмном лесу. Жители деревни встали и низко поклонились месту, где скрылись Мостада Датсавима, и ещё раз низко – полной луне. Потом мы пошли назад в деревню, где, несмотря на поздний час, звучали трубы и барабаны, бледное эхо музыки, услышанной ранее...»
Грустно сознавать, что детальное изучение этих животных более невозможно, так как они – точно так же, как Хууака-Хатчи – вскоре стали жертвами насморка, завезённого Скамтквистом. Скамтквиста, однако, ждал успех в поимке и приручении одного из самцов. Животное казалось очень смышленым, что стало понятным в свете информации о весе его мозга, который был определён позже. Оно стало очень ручным, и Скамтквисту даже удалось научить его двум органным фугам Баха, которые он знал наизусть, так, что оно могло безошибочно исполнять их. Лишь его неспособность производить долго звучащие ноты составляла некоторую сложность. Здесь животное сбивалось на очень быстрые tremoli, используя четыре рыльца, отрегулированных на одну и ту же высоту тона.
56 chilopus Греч. = тысяченожка.
*Примечание переводчика: “Tatzel” означает скорее «лапа», чем «кисточка», но английское слово “Tasselsnouters” выглядит более подходящим для этих животных.
57 ingens Лат. = огромный.
58 sphincter glossiformis [язычковый сфинктер – В. П.] – это не простой кольцевой мускул. Целостный дозирующий механизм, описываемый этим термином, образован настоящим сфинктером, который простирается на протяжении трёх четвертей внутренней окружности рыльца, и твёрдой подушкой соединительной ткани, которая занимает оставшуюся четверть. На этой подушке соединительной ткани находится V-образная пара выростов, которые работают как голосовые связки. Голосовой аппарат мог бы с точки зрения логики называться “narynx”, так как он представляет собой полностью сопоставимый аналог гортани (larynx) и евстахиевой трубы (syrinx).
Рукопись ХАРАЛЬДА ШТУМПКЕ ожидала публикации, когда стало известно, что во
время секретных ядерных испытаний (о которых ничего не было известно даже
прессе) весь архипелаг Хай-яй-яй был уничтожен из-за оплошности какого-то
подчинённого. В результате тектонических напряжений, появления которых никто не
ожидал, целая группа островов погрузилась в море, когда на расстоянии около 125
миль был произведён взрыв.
В это самое время международная комиссия по изучению архипелага была на сессии
на Майрувили. Среди неё присутствовало большинство учёных, названных в этой
работе. С ними утонул Дарвиновский Институт Хай-яй-яй, расположенный в
прекрасном восточном заливе острова, а с ним и невосполнимый фотографический
материал, различные препараты и отчёты о наблюдениях и экспериментах, которые
должны были составлять формальное ядро большого, всеохватного трактата,
касающегося архипелага и его геологических, исторических, зоологических и
социологических особенностей.
Наиболее удачным во всём этом было то, что незадолго до своего путешествия
Штумпке взял на себя труд составить короткое описание строения и повадок
Rhinogradentia. С целью подготовки рисунков он снабдил меня также кое-каким
материалом, который он – можно лишь сказать «увы!» – взял с собой назад на
Хай-яй-яй. И всё сложилось таким образом, что по крайней мере часть дела всей
жизни этого скромного и заслуженного искателя Истины удалось сохранить как
единое целое и для науки, и для более широкой аудитории, а вместе с этим и
знание о ныне исчезнувшем мире.
ГЕРОЛЬФ ШТАЙНЕР
Гейдельберг, октябрь 1957
ASTEIDES, S. (1954); Le nez d’Orchidiopsis, son anitomie, son developpement.
C. r. Soc. biol. Rh. 516; 28.
BEILIG, W. (1954); Ein vanadiumhal tiger Eiweisssymilex aus den nasalen
Fangfaden von Emunctator. S. H. Z. physiol. Chem. 884; 55.
BITBRAIN, J. D. (1946); Anatomical and histological sudy of the nose of a
Rhinogradent, Rhinolimacius. J. gen. Amt. Soyi 18.
---- (1950) The Rhinogradents. Univ. Press S. Aidrews.
BLEEDKOOP, FR. (1945); Das Nasobemproblem. Z. v. Lit. 34; 205.
BOKER, H. (1935 and 1937); Einfuhrung in die vergleichende Anatomie der
Wirbelriere. Fischer, Jena.
BOUFFON, L. (1953); A propos du systeme nutritif des Rhinogradents. Bull.
Darwin Inst. Hi. f; Suppl. 2.
---- (1954); A propos du groupe polyphyletique de Rhinocolumnides. Bull. Darwin
Inst. Hi. 8; 12.
BOUFFON, L., and GAUKAM-SUDUR, O. (1952); L’anatomic comparee des Polyrrhines.
Bull. Darwin Inst. H. 6; 33.
BOUFFON, L., IRM-EGINGARRI, J., and CHAIBUN, F. (1953); A propos de
l’innervation du nasoire des Polyrrhines. C. r. Soc. Biol. Rh. JJJ; 24.
BOUFFON, L., and LO-IBILATZE-SUDUR, Ch. (1954); Comment Orchidiopsis
attire-t-elle sa proie? La nature (P) v; 311.
BOUFFON, L., and SCHPRIMARSCH, J. (1950); Corcernant la question de la
descendance du genus endemique Hypsiboas. Bull. Darwin Inst. Hi. 4; 441.
BOUFFON, L., and ZAPARTEGINGARRI, V. (1953); Fur Fembryologie des
Orchidiopsides. Bull. Darwin Inst. H. 7; 16.
BROMEANTE DE BURLAS Y TONTERIAS, J. (1948); A systematica dos Rhinogradentes.
Bull. Darwin Inst. Hi. 2; 4j.
---- (1948); Systematic studies on the new wder of the Rhinogradents. Am. Nat.
F. 374; 1498.
---- (1949); Os Polyrrhines e a derivacao d'elles Boll. Braz. Rhin. i; 77.
---- (1950); A derivacao e a arvore genealogica dos Rhinogradentes. Boll. Braz.
Rhin. 2; 1203.
---- (1951); The Rhinogradents. Bull. Darwin Inst. Hi. SS Suppl.
---- (1952); The Hypogeonasidae. Bull. Darwin Inst. Hi. 6; 120.
---- (1954); The hides of Rhinogradents and their grain. Nature
(Danuddlesborough) 92; 2.
BROWN, A. B., and BITBRAIN, J. D. (1948); A simple electronically controlled
substitute for feeding Mercatorrhinus. J. psych, a. neur. contr. 181; 23.
BUCHNER, P. (1953); Endosymbiose der Tiere mit pflanzlichen Mikroorganismen.
Birkhauser, Basel.
COMBINATORE, M. (1943); Un pezzo di legno appurtato, trovato sulla spiaggia di
Owsuddowsa. Lav. preist. (Milmo) 74; 19.
D’EPP, FR. (1944); La descendance des Polyrrhinei. C. r. Soc. biol. Rh. so6i
403.
DEUTERICH, T. (1944); Em holzemer Suppenloffel von Haidadaifi. Z. f. v.
Prahist. 22; 199.
---- (l944a); Grundsatzliches fiber die Esslestecke der Huacha-Hatschi, eines
ausgestorbenen polynesisch-bajuwarischen Mischvolkes. ibid. 2 4;-312.
FREDDHRISTA, P., and PERISCHERZI, N. (1948); L’amblamento di colore fssiologico
nei mammiferi, specialemente nei generi Hexanthus e Cephalanthus (Polyrrhina,
Rhinogradentia) Arch. di fisiol. comp. ed. irr. 34; 222.
GAUKARI-SUDUR, O., BOUFFON, L., and PAIGNDPOULOS, A. (1950); L’anatomie
comparee des Sclerorrhine. C. r. Soc. Biol. Rh. 512; 39.
GRUHLE, H. W. (1947); Ursache, Gnind, Motif, Auslosung. Festschr. f. KURT
SCHNEIDER, Heidelberg, Sclerer.
HARROKERRIA, J., and IRRI-EGINGARRI, J. (1949); Note sur la biologie
d’Otopteryx volitans. C. r. Soc. Biol. Ih. 511; 56.
HYDERITSCH, F. (1948); The slug which was a mammal. Sci.a. med. cinemat. Cie,
Black Goats.
IZECHA, F. (1949); La primitividad de la cola de los Rhinogradentes. Boll. Arg.
Rhin. 2; 66.
JERKER, A. W., and CELIAZZINI, S. (1953): The ancestors of the Hypogeonasidae,
were they Emunctators? Evolution (Littletown) JJ; 284.
JESTER, M. O., and ASSFUGL, S. P. (1949); The genus Dulcicauda and the problem
of “Rassenkreis”. Bull. Darwin Inst. Hi. 3; 2il.
LUDWIG, W. (1932); Das Rechts-Links-Problem in Tierreich und beim Menschen.
Berlin.
---- (1954); Die Selektionstheorie. In: Die Evolution der Organismen. Hrsgeg.
v. G. HEBERER. Fischer, Stittgart.
MAYER-MEIER, R. (1949); Les “Triclades” de MUELLER-GIRMADINGEN, sont ils des
mammiferes? Bull. bnl. mar. St. V. H. 171 i.
MORGENSTERN, CHR. (1905); Galgenlieder. B. Cassrer-Berlin.
MULLER-GIRMADINGEN, P. (1948); Les triclades do sables du Wisi-Wisi. Acta
Helvetica Nas. Ser. B. IS! 210.
NAQUEDAI, BR. B. (1948); Georrhinida et Hypogeoiasida, deux subtribes parentes.
C. r. Soc. biol. Rh. 5101 64.
PETTERSSON-SKAMTKVIST, E. (1943); The discovery of the Hi-lay-Archipelago. J.
A. geogr. 322; 187.
---- (1946); Aventyrer pa Haiaiai-oema. Nystrims Forlag och Bokhandel, Lilleby.
PUSDIVA, FR. (1953); Uber die Schleimdrusen und de proteolytischen Prozesse in
der Sellarscheibe von Dulcicauda grise-aurella. S. H. Z. physiol. Chemie 822;
1443.
REMANE, A. (1954); Die Geschichte der Tiere. Ii; Die Evolution der Organismen,
hrsgeg. v. G. HEBERER. Fiseher, Stuttgart.
REMSCH, B. (1947); Neuere Probleme der Abstunmungslehre. Stuttgart.
SCHUTLIWITZKIJ, I. I. (1947); Hat Morgenstern die Rhinogradentier gekannt?
(Russisch mit dtsch. Zusanmenfassung.) Lit. prom. N. S. 27; 81.
SHIRIN TAFARUJ (1954); A propos du chimisme du sue attractif des Nasolimacides.
J. physiol. irr. u; 74.
SPASMAN, O., and STULTEN, D. (1947); Rhinogndenternas systemet. Acta Scand.
Rhin. 4; I.
SPUTALAVE, E. (1946); Le sabbie miliolidiche del orizzonte D 16 superiore del
l’isola Miruveely. G. geogr. frdd. Ital. Jp<? 12.
STULTEN, D. (1949); The descendency of the PolTrhines. Bull. Darwin Inst. Hi.
3; 31.
---- (1950); The anatomy of the nasarium of Hopsorrhinus. Bull. Darwin Inst.
Hi. 4; 511.
---- (1955); The evolution of turbellarians, a leview of new aspects. Piltdown
Univ. press.
STUMPKE, H. (1956); Das Nasarium der Poljrrhinen, erne Zusammenfassung der
bisherigen Ergebnisse, unrer besonderer Berucksichtigung der neueren
Untersuchungen iber die Innervierung. Zool. Jahrb. Abt. XX-XI, 43; 497.
TASSINO DI CAMPOTASSI, I. (1955): Un “releaser” sopranormale in Mammontops. G.
psicol. comp. e corn. 2: 714
TRUFAGURA, A. (1948); La cola de los Rhinogradentes. Boll. Arg. Rhin. I; I.
История о носоходках не была бы полной, если бы не две заметки в различных журналах, продолжающие данную тему. Наиболее широко известна и доступна в Интернете статья из журнала «Природа», где описаны дальнейшие находки представителей группы носоходок, сделанные уже после того, как архипелаг Хай-яй-яй погрузился на дно океана. И уж тем более она была бы неполной без рассказа об авторе этих существ. Поэтому я включаю в качестве приложения в электронную версию книги статью «Загадочные носоходки», опустив лишь её первую часть, где просто коротко пересказывается содержание самой книги. Ещё одна заметка - статья из журнала «Биология моря» за 2004 год, где также рассказывается о новом представителе этой группы животных. Данная статья приводится полностью.
Павел Волков.
С. Д. Фелдоянц,
кандидат биологических наук
Зоологический институт РАН
Санкт-Петербург
А. Зельбстандер
Рурский университет
Бохум (Германия)
Вторжение в биологию точных знаний оттеснило ее традиционные направления, в частности, описательную и классификационную зоологию. Между тем, представители именно этих разделов науки первыми встречают неведомое – ранее неизвестные ученым живые организмы. Так уж вышло, что спрятавшиеся в своем "затерянном мире" ринограденции пока не привлекли к себе должного внимания генетиков, молекулярных биологов и т.п., но уж сейчас, пусть с запозданием, они должны стать объектом тщательного изучения и применения генно-инженерных технологий.
Какая благодарная область для молодых дарований, тем более, что грядущие исследования, разумеется, будут тотчас поддержаны европейскими, американскими и японскими научными фондами! Уже данная пионерская работа может служить прекрасным примером плодотворности международного делового сотрудничества.
Лишь традиционным пренебрежением к беспозвоночным можно объяснить молчание наших популяризаторов о носоходках, особенно, на фоне их безответной любви к снежному человеку и динозаврам. Прошли десятилетия со времени первой публикации о ринограденциях в "Науке и жизни", однако лишь теперь массовый читатель может познакомиться с их безукоризненно научным, но вполне доступным описанием.VIVOS VOCO!
Non cuicunque datum est habere nasum
Martialis
[начало статьи опущено, поскольку в нём даётся лишь краткий пересказ содержания книги - В. П.]
До последнего времени наука не располагала информацией, сохранились ли
отдельные виды Rhinogradentia в акватории затонувших островов. Нам
посчастливилось убедиться в том, что фауна Rhinogradentia не вымерла
полностью, и науке еще предстоят открытия, связанные с таинственными
носоходками. Но все по порядку…
В 1990 г. во время экспедиции на борту научно-исследовательского судна
“Bipolarmond” в материале, который собрал один из авторов этих строк (А.
Зельбстандер) в северо-западной части моря Уэдделла, у побережья о. Кювета, был
найден неизвестный колониальный организм. Первоначально эта маленькая колония,
обнаруженная в сублиторали на глубине 4—5 м, была идентифицирована как
водоросль по наличию у нее ризоидов и ветвящегося столона. Главное отличие этой
колонии от всех ранее известных — сильно утолщенная базальная часть, что
характерно для молочаев — наземных растений рода Euphorbia. Позже, в
результате более детального изучения колонии, было решено, что она принадлежит
к одной из наиболее известных групп морских беспозвоночных — Hydrozoa,
а точнее — к неизвестному роду отряда Athecata, у которых есть
бульбовидные щупальца кормящего полипа и нет перисарка (скелетной оболочки)
вокруг головки полипа.
Примерно в это же время другой автор статьи (С. Фелдоянц) занималась исследованием коллекции, собранной русскими биологами-аквалангистами А. Ганнибаловым и Е. Карговым в северной части моря Дейвиса, рядом с о. Гуднайт. В коллекции был обнаружен странный овальной формы организм, покрытый напоминающими чешую пластинками и дающий небольшие столоны с овальными почками. Кроме того, проба, взятая в планктоне у берегов острова, содержала маленьких медузок сходной морфологии. Идентифицировать эти организмы оказалось столь же трудно, что и находки немецкого коллеги.
Спустя несколько лет, во время совместной работы с антарктическими коллекциями в университете BPG, мы сравнили имеющиеся у каждого непонятные морские организмы. К тому времени коллекции пополнились аналогичным новым материалом. Совместное исследование позволило сделать смелое предположение: найденные на мелководье у островов Кювета и Гуднайт непонятные организмы принадлежат к группе Rhinogradentia, открытой 50 лет назад на островах Хей-Эй и описанной Штюмпке. Обнаруженные у берегов Антарктиды животные выделили в два новых семейства — Hydroidopsidae и Scyphoidopsidae.
Для подтверждения правильности наших выводов мы обратились за консультацией к известному таксономисту профессору В. Ричворду, работающему в Лейдене. Первоначально профессор высказал сомнение по поводу подобной интерпретации, но позднее согласился с нашим мнением и продемонстрировал свою коллекцию с близкими по морфологии организмами, которые были собраны во время экспедиции “Onion Performance” у берегов о. Клозет. Ричворд любезно предоставил свой материал в наше распоряжение. Итак, попытаемся дать таксономические диагнозы трем новым видам Rhinogradentia, описать их поведение и экологию, воспроизвести жизненные циклы, а также определить место описываемых семейств в естественной системе группы.
Отряд Rhinogradentia подотряд Polyrrhina
триба Hexarhinida
1. Семейство Hydroidopsidae fam.n.
род Rhizoidonasus gen.n.
вид Rhizoidonasus euphorbiformis sp.n. [6]
Типовой материал: голотип (N1/7843) хранится в коллекции BPG университета; паратипы (N2/1011 и N3/1012) — в коллекции PZR института. Материал собран в феврале—марте 1990 и 1991 гг. в море Уэдделла у о. Кювета на глубине 5 м, грунт — песок с камнями.
Описание. Колонии до 20 см в высоту. Материнский организм прикреплен к грунту с помощью шести утолщенных в основании носов, каждый из которых имеет разветвленный ризоид, покрытый тонким роговым скелетом. Ризоид заканчивается присоской, обращенной к грунту. Голова с двумя большими глазами и удлиненными ушами. Относительно короткое тело заужено в противоположном голове направлении. Тело и базальная часть каждого носа покрыты волосами. Передние конечности (“руки”) и задние конечности (“ноги”) имеют по два очень длинных пальца, каждый с когтем. “Руки” и “ноги” в верхней части опушены волосками, а в нижней — заключены в наружный скелет. Чрезвычайно длинный полисифонный хвост разветвлен, на ветвях которого сидят молодые организмы обоих полов. Hосы некоторых из них хорошо развиты, снабжены ризоидами и готовы к закреплению на грунте, тогда как носы самых молодых еще лишены ризоидов, но имеют по одной присоске.
Замечания. Материнский организм прикреплен к каменисто-песчаному грунту всеми четырьмя толстыми, прижатыми один к другому носами, ризоиды которых простираются на расстояние нескольких сантиметров и могут оплетать талломы водорослей, присасываться к камням или раковинам. Чтобы удерживать равновесие, материнский организм прижимает уши к телу и как бы поддерживает себя вытянутыми вдоль него “руками”, при этом “ноги” животного цепко держатся когтями за собственный хвост.
Пока не ясно, как происходит репродукция описываемой колонии, не известно, могут ли ее образовывать взрослые мужские особи. Замечено, что заботится о потомстве материнский организм — молодые особи обоих полов висят у нее на хвосте. Как долго тянется эта связь, пока не установлено.
 |
Колония
Rhizoidonasus
euphorbiformis,
образованная материнским организмом,
на ветвях хвоста которого висят молодые особи обоих полов.
2. Семейство Hydroidopsidae
род Larvanasus gen.n.
вид Larvanasus haleciformis sp.n. [7]
Типовой материал: голотип (N1/10113) в коллекции PZR института; паратипы (N2/2985) в PLN музее и (N3/7844, N4/7845) в BPG университете. Материал собран в январе 1980 г. в Индийском океане у о. Клозет на глубине 2 м на заиленном песке с камнями.
Описание. Вытянутое и изогнутое в виде знака вопроса тело (до 5 см в высоту) очень напоминает по форме половые зооиды на колониях представителей рода Halecium (Hydrozoa). Оно покрыто тонким прозрачным наружным скелетом, сквозь который просвечивает внутренняя камера с 9—10 разновозрастными и сильно отличающимися по облику личинками — от червеобразных, покрытых нежным “мехом”, до рыбообразных или тюленевидных и, наконец, типичных для Larvanasus организмов. Это — прекрасный пример рекапитуляции органов в процессе онтогенеза. Камера связана с внешней средой воронкой — ларватекой. Полностью сформированные молодые ларваносы покидают материнскую камеру через ларватеку. Голова, шея и проксимальная часть трехпалых конечностей покрыты нежными волосками. Широкая голова материнского организма снабжена шестью короткими, узкими и так же опушенными носами, крупными ушами, глазами на стебельках и непомерно большим ртом с широкими губами.
Hа том же месте был пойман другой взрослый экземпляр (скорее всего мужской) того же вида, но меньших размеров (3 см), с редуцированными камерой и ларватекой, погруженной под наружный скелет. Поверхность тела и края губ этого экземпляра покрыты диатомовыми водорослями. Кроме того, на грунте было обнаружено несколько крошечных особей (до 2 см), которые отличались от взрослых лишь длинными носами с присосками.
Замечания. В начале февраля на том же месте, где были встречены взрослые ларванасусы, обнаружено множество червеобразных организмов около 1 см длиной. Первоначально они были приняты за плоских червей турбеллярий, но в конечном итоге идентифицированы как личинки ларванасусов. Интересно, что эти личинки концентрировались вокруг взрослых материнских особей и были найдены в каналах их носов, по которым, видимо, проникают в ларвальную камеру. Мужская особь ларванасуса представляет собой явно редуцированную форму (редуцированы ларвальная камера и ларватека). После вымета в воду половых продуктов мужские особи, видимо, ведут пассивный образ жизни, прячась в свою наружную скелетную оболочку. При этом питаются они тоже пассивно: диатомовые водоросли, обильно покрывающие поверхность их тел, проникают через нос в желудок с током воды, так же как основная пища ларванасусов — мелкие планктонные организмы. Есть основания полагать, что виду свойственно наружное оплодотворение, главенствующую же роль в жизненном цикле ларванасусов играют женские особи, вынашивающие молодь в ларвальных камерах. Найденные возле материнского организма на камешках и песке детеныши передвигались на всех своих шести носиках и четырех ногах. Это означает, что редукция двигательной функции носа происходит лишь на взрослой стадии развития ларванасусов.
 |
Семья ларванасусов. Через прозрачную стенку внутренней камеры женской особи (в центре) видны личинки на разных стадиях развития. У другой, скорее всего мужской (слева), — редуцированы камера и ларватека. Вокруг материнского организма концентрируются червеобразные личинки.
3. Семейство Scyphoidopsidae
род Nudirhinus gen.n.
вид Nudirhinus medusiformis sp.n. [8]
Типовой материал: голотип (N1/10114) хранится в коллекции PZR института, паратипы (N2/7845) — в BPG университете и (3/2986) — в RLN музее. Материал собран в феврале 1981 г. в море Дейвиса у о. Гуднайт на глубине 4 м на скальном грунте.
Описание. Маленький (2.5 см в диаметре), овальной формы организм покрыт толстым наружным скелетом в виде пластинок с тончайшими выростами, подобными биссусу у моллюсков. У этой специфической “раковины” нудиринуса есть четыре отверстия для конечностей, два — для ушей и еще одно — для хвоста. Как правило, голова находится внутри раковины, снаружи можно видеть лишь очень длинные уши, крупные овальные глаза и шесть носов с присоской на каждом. Взрослые организмы стоят на грунте и передвигаются по скале с помощью носов. Глаза светятся в темноте голубым светом, что характерно для некоторых морских беспозвоночных (например, гидроидов рода Obelia). Природа свечения, возможно, связана с наличием в тканях животного особых клеток фотоцитов, содержащих Ca2+-зависимый фотопротеин. Если эта версия будет подтверждена и позже удастся обнаружить места скопления нудиринусов, возможно, эти животные будут использованы в медицине для ранней диагностики злокачественных заболеваний, подобно препаратам “экворину” и “обелину”, получаемым из некоторых морских гидроидных [9]. Передние конечности (“руки”) заканчиваются типичными клешнями для взрыхления грунта, задние (“ноги”) — четырьмя пальцами. Такое строение конечностей обеспечивает животным передвижение по скальному грунту.
Самая интересная особенность нудиринусов — образование почек на их хвостах и ушах. Как показали дальнейшие исследования, эти почки развиваются в пелагические формы. Hаличие вегетативной стадии — феномен, абсолютно не свойственный ринограденциям. Прикрепленные почки можно видеть на хвостиках и ушках даже личинок. Лапки и носики также становятся различимыми у пелагических личинок в момент их оседания на грунт. Пелагические личинки нудиринусов очень похожи на сцифомедуз: желеобразное тело напоминает умбреллу, носы — ротовые лопасти медузы [10].
Замечания. Репродукция нудиринусов неизвестна. Пол трех найденных экземпляров установить не удалось. Возможно, они — гермафродиты. Но есть и другая версия, согласно которой пелагическая стадия — носитель пола животного. В таком случае правомерен вопрос, что считать эволюционно первичным — донный или пелагический образ жизни группы ринограденций. По-видимому, эволюционно пелагическая стадия — не что иное, как трансформированный донный организм [11].
 |
Обнаружение трех новых представителей Rhinogradentia, принадлежащих к трем новым родам и двум новым семействам, ставит перед наукой очень важные теоретические вопросы.
1. Насколько реально существование и поныне фауны Rhinogradentia? Для решения этого вопроса возможно использование нескольких подходов, в том числе и нумерического — например, статистической конструкции для определения потенциального родового разнообразия группы [12].
2. Каким может оказаться современный ареал этой группы? Как известно, первые находки Rhinogradentia сделаны на затонувших ныне островах южной части Тихого океана, наши — в холодных водах Южного океана. По-видимому, Rhinogradentia — биполярная группа животных, обитающих в холодных водах океана, в том числе и на глубинах Центральной Пацифики, в Южном океане, и, вполне возможно, они будут обнаружены в холодных или умеренных водах Северного полушария [13].
3. Каково филогенетическое положение этой группы? В монографии Штюмпке дано эволюционное древо родов Rhinogradentia. Мы попытались найти место на этом древе описанных выше новых таксонов. Несомненно, что все три рода должны находиться на правой ветви, между родами Eledonopsis (22) и Cephaloanthus (24). Эти роды принадлежат к группе Hexarrhinida (шесть носов). Как известно, Eledonopsis suavis имеют каналы в носу, сквозь которые пища проходит в желудок животного. Носы Larvanasus haleciformis имеют каналы сходной конструкции. Таким образом, мы помещаем наш род Larvanasus в основании ветви. В то же время хорошо известны колониальные Hexanthus и Cephalanthus (Ranunculonasus и Corbulonasus). Мы помещаем наш род Rhizoidonasus (28) рядом с этими троль-цветоносами (23 и 24). Наконец, наш Nudirhinus (29) может быть помещен рядом с Rhizoidonasus, поскольку представители обоих родов имеют в той или иной форме выраженные колонии. К сожалению, отсутствие живого материала лишает нас возможности использовать в наших филогенетических построениях молекулярно-биологические методики.
|
Филогенетическое древо Rhinogradentia.
|
1 — Archerrhinos, |
1. Pettersson-Skamtkvist E. The discovery of the Hi-Iay-Archipelago // J. A. geogr. 1943. №3. Р.322.
2. Morgenstern Ch. Galgenlieder. Berlin, 1905. (Пер. с нем. С.Степаньянц.)
3. Bromeante de Burlas y Tonterias J. A systematica dos Rhinogradentas //Bull. Darwin Inst. 1948. Р.2.
4. Combinatore M. Un pezzo di legno, appuntato trovato sulla spiaggia di Owsuddowsa. Milano, 1943; Schutliwitzkij I. I. Hat Morgenstern die Rhinogradentier gekannt? (Russisch mit deutsch. Zusammenfassung.). Berlin, 1947; Trufagura A. La cola de los Rhinogradentes // Boll. Arg. Rhin. 1948; Stulten D. The anatomy of the nasarium of Hopsorrhinus // Bull. Darwin. Inst. 1950. Р.4 etс.
5. Stumpke H. Bau und Leben der Rhinogradentia. Stuttgart; Jena, 1960.
6. Родовое название дано в связи с наличием ризоидов на носу, видовой эпитет означает сходство носов каждого зооида с корнями растения Euphorbia.
7. Родовое название дано в связи с тем, что нос выполняет двигательную функцию лишь у личинок этого вида, видовое — по сходству формы тела этих организмов с гонотеками у гидроидов рода Halecium.
8. Родовое название дано по голым, непокрытым волосками, носам; видовое — означает сходство со сцифомедузами.
9. Highotsky E. Ca2+-dependent photoproteins and bioluminiscence of marine invertebrata. Proc. Internation. Invertebr. Conference. 1999.
10. Greyavin L. Evolution of the scyphomedusae behaviour // Proc. Behaviour Internation. Conf. 1989.
11. Offerro F. Life history of some low invertebrates // Phil.-Evol. J. 1969. №6. Р.61.
12. Nonsensevitch A. How many genera will be described during the closest future // Phil.-Evol. J. 1994. №3. Р.105.
13. Fieldojants S., Selbstander A. Problem of bipolarity on the marine invertebrate example. Phil.-Evol. J. (in press).
Вольный перевод строки римского поэта
Марциала (43—104 г.)
" Non cuicunque datum est habere nasum"
Дабы не вводить в заблуждение несведущего и не разгневать серьезного, хорошо
разбирающегося в зоологии читателя, раскроем карты и напомним, что номер, в
котором опубликована наша (под псевдонимами С.Д.Фелдоянц, А.Зельбстандер)
статья "Загадочные носоходки", был апрельским. Тем более, однажды это
уже случилось с известным таксономистом, названным в статье профессором
В.Ричвордом. Получив в подарок рисунки наших носоходок, профессор помрачнел и
сказал: "Не признаю шуток в науке!"
"Открытия" носоходок и других не менее фантастических животных
принадлежат известному немецкому зоологу Герольфу Штайнеру. Еще в студенческие
времена в Гейдельбергском университете (Германия) он вместе с друзьями
развлекался выдумыванием разных мифических животных, описания и рисунки которых
служили основой для развития фантастических эволюционных идей, не лишенных
рационального зерна, поскольку горячие дискуссии способствовали формированию
филогенетического мышления.

Автопортрет Герольфа Штайнера
(он же Харольд Штюмпке, он же Карл Д.С. Геесте),
известного немецкого зоолога,
автора загадочных носоходок.
Еще во времена Отто Бючли (1898) здесь выходило многотомное издание
"Stammbuch des Zoologischen Institut", впоследствии переименованное в
"Schwarzes Buch" ("Черная книга"), в котором, между прочим,
принимали участие и русские исследователи В. В. Редикорцев, Н. А. Касьянов и
др. Героями одного из томов, вышедшего в 1929 г., были Balhurner (рогатые шары)
— гипотетические млекопитающие, которые обитают на болотах, в степях, пустынях
и на холмах. Особое внимание уделялось адаптациям, закрепляющимся в ходе
эволюции.
До второй мировой войны Штайнер работал ассистентом на кафедре зоологии в
Гейдельбергском университете. Профессор этой кафедры, начальник Штайнера,
в то время наблюдал за стайным поведением озерных гольянов (Phoxinus phoxinus).
Его эксперименты сводились к изучению последствий удаления части мозга
у этих рыбок. Оказалось, что в результате такой операции рыбки теряли стайный
инстинкт
и стремились к лидерству (Fuhrerschaft). Вывод о том, что удаление мозга
приводит к "фюрерству", стоил профессору свободы. На его место
был приглашен Штайнер. Работая со студентами, он продолжал выдумывать новых
животных и рисовать их черно-белые и цветные изображения. Во время лекций
Штайнер демонстрировал эти рисунки. После войны города Гейдельберг и Дармштадт,
в котором находился другой, более молодой Дармштадтский технический университет
(между ними — не более 50 км), оказались в американской зоне оккупации.
Дом,
где была квартира Штайнера, разбомбили, но ему удалось спрятать свои рукописи,
рисунки и акварельные краски в подвале разрушенного дома, а позже перенести
в пустующий дом коллеги и поселиться там. Университеты не работали. Жители
города
сидели по домам и лишь днем на час-другой выходили на улицу, пытаясь раздобыть
еду. По воспоминаниям Штайнера, чувство голода было доминантой, и день
считался удачным, если на обеденном столе оказывались улитки, майские жуки,
крапива или,
если повезет, мясо кролика. Студентка Штайнера приносила ему на обед аспарагус
из своего сада, а он дарил ей рисунки. Но дубликаты оставались. Часть из
них он пытался продавать, чтобы заработать на еду... В 1946 г. вновь открылся
Дармштадтский университет и Штайнер начал там работать. Но он не забывал
своих
коллег-зоологов и друзей из находящегося неподалеку Гейдельберга. Профессор
Вернер Раух помог сделать c рисунков Штайнера цветные слайды, которые другой
коллега, Эрих фон Холст, использовал на семинарах, посвященных эволюции
животных.
Популярность рисунков Штайнера росла, и вслед за рогатыми шарами на свет появились носоходки (Rhinogradentia), изображения которых также демонстрировались на семинарах. Иллюстрации постепенно стали обрастать текстом, и в 1950 г. Штайнер сделал первую попытку издать книгу о Rhinogradentia c собственными цветными иллюстрациями. Но тогда это оказалось ему не по карману. В 1957 г. он написал послесловие к "Манускрипту Харальда Штюмпке" о фауне островов, затонувших в результате атомных испытаний.
Первое издание книги Х.Штюмпке (псевдоним Г. Штайнера) "Bau und Leben der Rhinogradentia" вышло в 1960 г. в самом известном и серьезном немецком издательстве "Gustav Fischer Verlag" (Stuttgard; Jena), специализирующемся на выпуске руководств, учебников и теоретических монографий. Книга имела настолько ошеломляющий успех, что в конце того же года была переиздана. Первоначально монография, естественно, вызвала шок: зоологи долго не могли понять подвоха — слишком высоко было доверие к издательству со знаменитой эмблемой — рыбкой, машущей хвостом... Когда же шутка оказалась раскрыта, Штайнера простили за профессиональность и то удовольствие, которое испытывает каждый читающий и от текста, и от юмора, и от великолепных иллюстраций. Не удивительно, что книга была несколько раз переиздана на немецком (последнее издание — в 1998 г., с цветными иллюстрациями) и переведена на французский, английский и японский языки. Не так давно издательство "Gustav Fischer Verlag" выпустило монографию "Stumpke's Rhinogradentia. Versuch einer Analyse", где на 119 страницах анализируется роль группы носоходок в эволюции животных. Автор монографии Karl D. S. Geeste — все тот же известный профессор зоологии Герольф Штайнер, сейчас ему 91 год.
С. Степаньянц,
кандидат биологических наук,
Зоологический институт РАН, С.-Петербург
А. Свобода,
Рурский университет, Бохум (Германия)
Журнал «БИОЛОГИЯ МОРЯ», 2004, т. 30, №2, стр. 163 - 164.
В 1987 г. в составе небольшой группы студентов я проходила летнюю
практику по морской зоологии на о-ве Путятина близ г. Владивостока. Большую
часть
своего времени мы посвящали прижизненным наблюдениям, вскрытию и зарисовке
различных
представителей морской фауны, но иногда случалось сунуть кое-что интересное
в банку с формалином, чтобы рассмотреть эти объекты позднее. По возвращении
в С.-Петербург (тогда он еще назывался Ленинградом) из-за всевозможных
текущих дел руки до этой банки не дошли. Лишь недавно, наводя порядок в лаборатории,
я наткнулась на забытую банку и решила в нее заглянуть. При этом среди
морских
червей, моллюсков и асцидий, хорошо известных науке и потому не представляющих
особого интереса, мне попался кусок полусгнившей древесины, к которому
была прикреплена небольшая колония неизвестного вида животных, состоящая всего
их трех зооидов и двух почек. Два зооида были сильно смяты и разрушены
из-за того,
что по неопытности я слишком плотно набила банку и находящиеся в ней нежные
объекты оказались в непосредственном соседстве с более грубыми (например,
раковинами моллюсков). Довольно хорошо сохранился лишь один зооид. Каково
же было мое
удивление и восхищение, когда при внимательном рассмотрении я увидела,
что
передо мной представитель крайне своеобразной группы - отряда носоходок
(Rhinogradentia), открытого немецким зоологом Г. Штюмпке (Stumpke; первые сообщения
об этих
удивительных существах были опубликованы им еще в 50-х гг., но я пользовалась
монографией,
изданной в 1993 г.). Носоходки замечательны тем, что их нос очень легко
приспосабливается к выполнению различных функций: многие виды используют его
для передвижения
(откуда происходит название группы), другие — для рытья в почве, а некоторые
- для прикрепления к субстрату, как и оказавшийся в моей банке вид. Г.
Штюмпке описал 26 видов носоходок, совсем недавно С. Фелдоянц и А. Зельбстандер
(2000)
описали еще 3 вида, а теперь мне посчастливилось найти еще один (30-й!)
вид, которому я решила дать название Dendronasus – «древовидоносый».
Описание. Колония дендроназуса состоит из осевого столбика, или стебелька,
который, по всей вероятности, прикрепляется к нижней поверхности судов
и деревянных сооружений. На апикальном свисающем вниз конце стебелька располагается
предположительно
самый старший зооид, являющийся основателем колонии; к сожалению, именно
этот зооид оказался почти полностью разрушенным, но все же удалось рассмотреть,
что стебелек является непосредственным продолжением носа зооида. Где-то
в
средней
части осевого стебелька находится утолщение, от которого отходят веточки,
тянущиеся к носам двух других зооидов и к двум еще очень слабо развитым
почкам (см. рисунок,
А). Очевидно, это утолщение представляет собой зону почкования и содержит
резервные элементы различных тканей, необходимые для развития почек. Общая
длина колонии
не превышает 2 см.
Строение хорошо развитого зооида представлено на рисунке (Б). Очевидно,
зооиды дендроназуса должны висеть ногами вниз, но, следуя обычаю, установившемуся
еще в классической монографии Г. Штюмпке, я изобразила его в положении
вниз
носом. Довольно компактное тело зооида (без стебелька) составляет приблизительно
5-6 мм (возможно, это самые мелкие представители класса млекопитающих).
Оно имеет желтоватую окраску и покрыто нежными золотистыми волосками. Голова
слабо отграничена от туловища. В основании стеблеобразного носа находятся
ноздри,
но никаких сведений о наличии у дендроназуса легких, способных служить
для
дыхания воздухом, у нас нет; очень вероятно, что ноздри играют роль лишь
обонятельных ямок. Рот слегка приоткрыт, и в нем хорошо видны зубы. Очень
крупные выпуклые
глаза свидетельствуют о том, что в жизни этого животного зрение имеет большое
значение, а также указывают на пребывание в слабо освещенных местах, что
хорошо согласуется с предположением о прикреплении колоний дендроназуса
к нижней поверхности
различных подводных предметов. Уши маленькие, козелки тесно прижаты к дну
ушных раковин и закрывают отверстия, ведущие в наружный слуховой канал,
из чего следует,
что слух у этих животных сильно понижен или вовсе отсутствует.
Непосредственно позади головы к туловищу примыкают два лопастевидных придатка,
от которых отходят пучки нежных разветвленных нитей. Внешне эти придатки
очень похожи на наружные жабры. Но то обстоятельство, что непосредственно
от каждой
лопасти отходит по 5 таких нитей, которые затем ветвятся, наводит на мысль,
что перед нами модифицированные передние конечности, которые стали функционировать
как органы водного дыхания (причем ногти бесследно исчезли).
Dendronasus sp. A - общий вид колонии (зооиды пронумерованы по старшинству), Б - отдельный зооид при большом увеличении.
Задние ноги дендроназуса имеют строение хватательных конечностей
и очень напоминают человеческие руки – большой палец противостоит остальным
четырем. Поэтому я
сначала приняла их за передние конечности, оказавшиеся на заднем конце тела
из-за какой-то фантастической гетеротопии. Ошибочность этой идеи стала ясна
лишь только после того, как я поняла истинную природу жаберных придатков.
По-видимому, задние ноги служат для схватывания различных планктонных организмов,
которыми
питаются зооиды. Хвост очень длинный и гибкий, с утолщением и большим роговым
когтем на конце. У описанного Г. Штюмпке Emunctator sorbens хвост имеет
сходное строение, причем в концевом утолщении содержится ядовитая железа.
Возможно,
это относится и к дендроназусу.
На брюшной стороне туловища имеются две выпуклости, соответствующие положению
млечных желез, из чего можно заключить, что рассматриваемый зооид является
особью женского пола (но нельзя исключить и гермафродитизм - явление широко
распространенное у животных, ведущих прикрепленный образ жизни).
Приведенное выше описание относится только к внешнему виду дендроназуса (впрочем,
так же поверхностно описаны и все прочие носоходки), но я надеюсь, что изучение
остатков двух других зооидов позволит получить сведения о внутреннем строении
этого вида, которые составят содержание моего следующего сообщения. После завершения
работы все изготовленные препараты и типовой экземпляр дендроназуса будут переданы
в музей Института мифозоологии.
Обсуждение. Как известно, многие зоологи, в том числе и проф. А.И. Лягушенков
(под руководством которого мне посчастливилось много лет работать), отнеслись
к открытию д-ра Г. Штюмпке весьма скептически. Проф. Лягушенков даже заявил,
что ринограденции созданы «из глубины немецкого гения» (см. Иванова, 2001).
С большим сомнением отнесся он и к сообщению С. Фелдоянц и А. Зельбстандера,
но теперь, увидев чудесного дендроназуса, был вынужден признать ошибочность
своих прежних высказываний.
К настоящему времени известно, что отряд Rhinogradentia содержит два подотряда:
более примитивный подотряд одноносых, имеющих, как и большинство животных,
только один нос, и подотряд многоносых, у которых произошла полимеризация
этого органа. Наш Dendronasus, бесспорно, должен быть отнесен к
одноносым. Из всех
известных видов носоходок ближе всего он стоит к Dulcicauda griseaurella и Columnifax
lactans, которые используют нос для прикрепления к субстрату,
причем
последний вид к тому же является обитателем моря, хотя адаптации к водному
образу жизни у него выражены не так сильно, как у дендроназуса.
Самыми замечательными особенностями дендроназуса следует признать превращение
передних конечностей в жабры и бесполое размножение. Позвоночным животным
бесполое размножение практически несвойственно, и до недавнего времени единственное
исключение представляла полиэмбриония броненосцев, которая осуществляется
очень своеобразно - путем изменения механизма эмбрионального развития. Но
носоходки дали нам еще два примера бесполого размножения. Помимо дендроназуса
оно обнаружено
у Rhizoidonasus euphorbiformis (Фелдоянц, Зельбстандер, 2000) — морской носоходки
из подотряда многоносых, у которой почки образуются на концах ризоидов (тонких
корнеобразных отростков, отходящих от кончиков носов). Принадлежность этих
двух видов к разным подотрядам не оставляет сомнения в том, что бесполое
размножение
возникло у них независимо. Интересно, что в обоих случаях базовым органом
бесполого размножения служит нос — орган, который, как уже отмечалось, легко
изменяет
свою форму и функцию и обеспечивает возможность приспособления животных к
совершенно различным условиям существования.
Было бы очень важно изучить не только внутреннее строение дендроназуса (что,
может быть, мне удастся), но и его индивидуальное развития при половом и
бесполом размножении. Однако последнее - дело почти безнадежное, так как
мы не знаем,
где искать дендроназуса, из какой части Мирового океана принесли нам волны
щепку с колонией этого вида. Можно рассчитывать только на счастливый случай.
Иванова О. М. Почти все о русалках. СПб: Лисс. 2001. 157
с.
Фелдоянц С. Д., Зельбстандер А. Загадочные носоходки //
Природа. 2000. № 4. С. 46-54.
Stumpke H. Bau und Leben der Rhinogradentia.
Stuttgart; Jena. 1993. 83 S.