THE SNOUTERS
THE SNOUTERSForm and life of the Rhinogrades
Harald Stumpke
Quondam curator of the museum of the Darwin Institute of Hy-Yi-Yi, Mairuwili.
| Main | Site library | Forum | Guestbook |
Gerolf Steiner
THE SNOUTERS
Form and life of the Rhinogrades
THE SNOUTERS
Form and life of the Rhinogrades
Harald Stumpke
Quondam curator of the museum of the Darwin Institute of Hy-Yi-Yi, Mairuwili.
Epilogue by Gerolf Steiner
Translated by Leigh Chadwich
The Snouters was originally published in Germany, under the title Bau und Leben der Rhinogradentia, by Gustav Fischer Verlag Stuttgart in 1957. Copyright ® 1964 by Gustav Fischer Verlag Stuttgart.
Portions of the book appeared in Natural History magazine in April 1967 and is reprinted here with permission.
The 15 plates and 12 text figures in this volume were drawn by Gerolf Steiner.
The University of Chicago Press, Chicago 60637 The University of Chicago Press, Ltd., London
English translation copyright ® 1967 by Doubleday & Company, Inc

Introduction
General
Description of the several groups
Uni-Snouters (Monorrhina)
Primitive Snouter Types
(Archirrhiniformes)
Soft-nosed Snouters (Asclerorrhina)
Surface Snouters (Epigeonasida)
Mud Snouters (Hypogeonasida)
Burrowing Snouters (Georrhinida)
Proboscipedes (Sclerorrhina)
Snout Leapers (Hopsorrhinida)
Multi-Snouters (Polyrrhina)
Four-Snouters (Tetrarrhinida)
Six-Snouters (Hexarrhinida)
Long-nosed Snouters (Dolichoproata)
Epilogue
Bibliography
Snouters: Russian version
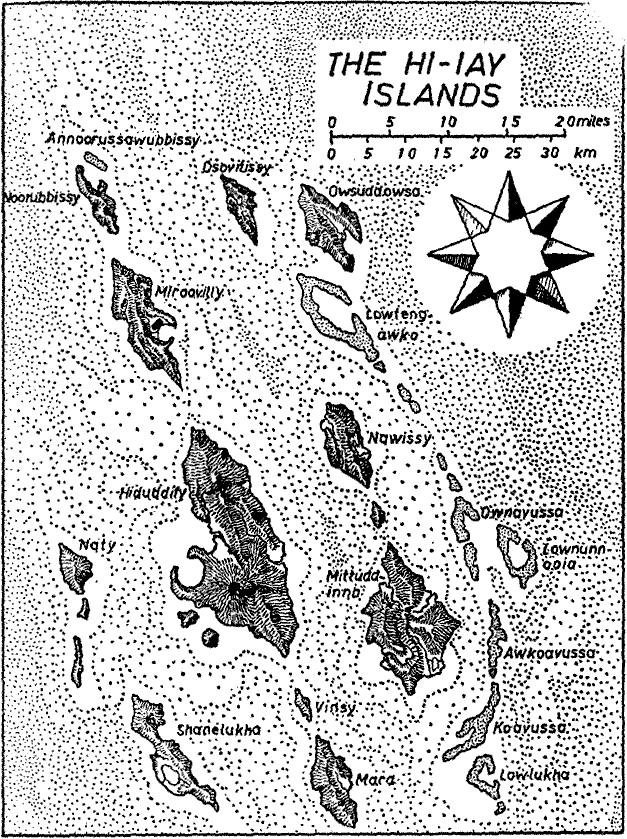
Among mammals the order SNOUTERS holds an unique position,
to be explained first of all by the fact that these most oddly constructed
animals have been
discovered only very recently. That hitherto they have remained unknown
to science is because their native land, the South Sea Archipelago Hy-yi-yi
(written Heieiei in German), was not discovered until the year 1941 and
even
then was visited for the first time by civilized Europeans through a rare
chance connected with the Pacific war. But in addition this group of animals
has particular significance because among them are found principles of
structure, modes of behavior, and ecological types that are unknown elsewhere,
not just
among mammals but among vertebrates in general.
Discovery of the archipelago is to be credited to the Swede Einar Pettersson-Skamtkvist
who ¢ escaping from Japanese imprisonment ¢ was wrecked on the island Hy-dud-dye-fee
(Heidadaifi). This island, which in contrast to many islands of the South
Seas is not of volcanic origin even though it does not lack an active volcano
(Kotsobousi-Kozobausi)
of respectable height (5740 feet), extends some twenty miles from north to
south and about ten miles east to west, consists predominantly of limestone
and metamorphic shales, and has as its highest elevation Shou-wunoonda (Schauanunda),
a twin-peaked mountain 7316 feet high.
The climate of the island is extremely equable, as is customary of islands
of the central and eastern Pacific. The tropical vegetation, the botanical
evaluation of which has scarcely begun, displays alongside genera of worldwide
distribution many endemic forms of archaic character (thus the Maierales,
closely related to the Psilotales; and the genus Necolepidodendron, to be
classed among the Lepidodendrales; and likewise the Schultzeales, that form
a series of magnificent virgin forest trees that are to be ranked near the
Ranunculaceae; and many more). The Hy-yi-yi Archipelago, to which Hy-dud-dye-fee
belongs, must hence be of ancient origin, as is true also in respect to the
geological-palaeontological findings almost exclusively palaeozoic deposits;
cf. Classification of the Miliolid Sands from the Upper Horizon D16 of
Mairuvili by Ezio Sputalave. At the very latest, the island group must have become
completely isolated from other continents in the Upper Cretaceous; likewise
it is to be assumed that the archipelago, for its part, is the remnant of
a fair-sized continent, since - in contrast to New Zealand - it contains
an incomparably greater variety and peculiarity of native groups of organisms
on a total island area of only slightly more than 650 square miles.
The natives found by Skamtkvist on his arrival in 1941 called themselves
Hooakha-Hutchi (Euacha-Hat-schi). They have since become extinct, but according
to Skamtkvist seem to have been polynesian-europaeoid. It was impossible
to investigate their language because a headcold introduced by the discoverer
destroyed these children of nature inside of a few months. Of their cultural
artifacts only a few wooden objects could be saved (cf. also Deuterich 1944
and Combinatore 1943).
PLATE I |
Weapons were unknown to the Hooakha-Hutchi. The peaceful tribe
was supported by the natural bounty of the surroundings. There was no excess
of births; ratter, ōfrom time immemorialö twenty-two chiefs had held sway over
approximately seven hundred souls. This much Skamtkvist was able to ascertain.
These arrangements had the fortunate scientific byproduct that despite the
presence of humankind the extraordinary archipelago world of organisms was
preserved, as is the more astounding because almost all the terrestrial animals
would have been destroyed if they had been hunted more intensively.
Despite the fact that their native home was unknown, Snouters had been mentioned
on one previous occasion. No lesser personage than the poet Christian Morgenstern
some sixty years ago announced the existence of the Snouters in his well-known
poem;
| ōAlong on its proboscis there goes the nasobame1 accompanied by its young one. It is not found in Brehm,* It is not found in Meyer,** Nor in the Brockhaus** anywhere. 'Twas only through my lyre we knew it had been there. Thenceforth on its probosces (above I've said the same) accompanied by its offspring there goes the nasobame.ö |
* Classical German treatise on zoology, first edition 1863, Fourth (last) edition 1918.
** Names of well-known German encyclopedias.
This concise and yet clear description, that expresses the
peculiar gait of this Snouter even in the rhythm of the verse, answers to a
hair that of Nasobema lyricum2. Hence one cannot think
other than that Morgenstern must have had a specimen of this Snouter before
him or have
had detailed information
about it. Bleedkoop 1945 in Das Nasobemproblem (The Nasobame Problem) considers
that there are two possibilities. Either Morgenstern was briefly in Hy-yi-yi
during the years 1893 to 1897, or through some chance or other he received
a hide of Nasobema lyricum (the honatata of the natives).
Yet no tropical expedition of Morgenstern's is known, and how might he have
obtained a hide? According
to a verbal communication from Mrs. Kathe Ziller, with whom Morgenstern was
well acquainted, he is said to have come home one evening in 1894 in the greatest
excitement, muttering over and over ōHy-yiyi, Hy-yi-yi!ö Soon thereafter,
it is said, he composed the poem in question, which he also showed her brother.
Bleedkoop concludes from this that Morgenstern had learned of Hy-yi-yi from
an acquaintance. But whether he actually had the honatata before him, or with
a poets intuition simply sketched a picture of the animal, mist remain enigmatic.
The lines: ō'Twas only through my lyre we knew it had been thereö would
permit the deduction that he didn't actually see it, but knew of it only by
description. Perhaps too he wished to veil the islands with their ancient organisms
from European greed and hence-to an extent as camouflage-wove these lines into
his poem? We do not know, just as we do not know from whom Morgenstern got
his information about Hy-yi-yi and its fauna. Actually the only possibility
in this direction is a merchant sailor, Captain Albrecht Jens Miespott, who
died at an early age; Morgenstern maintained a considerable correspondence
with him. In 1894, after returning from a long and extraordinary journey, Miespott
died in a state of mental derangement in Hamburg. Perhaps it was he who knew
the secret of Hy-yi-yi and carried it with him to the grave. So much for Bleedkoop's
investigations.
In a meritorious study I. I. Schutliwitzkij occupied himself with the same
problem. He comes to about the same conclusion as Bleedkoop, but with the difference
that he considers it possible that between the years 1894 and 1896 Morgenstern
received from Miespott's estate a living honatata, that he kept in a cigar
box for some weeks. Yet here too the data are contradictory. Moreover, it could
have been no more than a ōpouch-babyö, since honatatas grow to a respectable
size (cf. p. 55). All that is certain is that the cigar box was quite a deep
one, labeled ōLos selectos hediondos de desechoö.
1 nasus Lat. = nose; Bema Grk. = to walk.
2 lyricus Grk. = pertaining to the lyre.
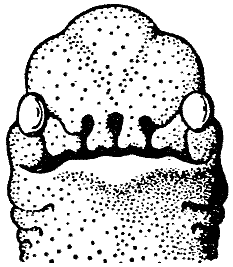
The Snouters, which are regarded as a special order of the mammals, and which have found a monographer in the well-known specialist Bromeante de Burlas, are ¢ as the name indicates ¢ all distinguished by the fact that the snout is extraordinarily developed. It may be present singly, or in a greater number. The latter condition is unique among the vertebrates. Now, anatomical investigations (here we follow Bromeante de BurlasÆ discussions) have shown that in the polyrrhine species the nasal rudiment is cleft at an early embryonic stage, so that the rudimentary individual nostrils that develop from it have a holorrhinous differentiation, i.e. each forms a complete snout (cf. Fig. 1). Together with the early polyrrhinallization there occur numerous extensive alterations of the entire structural plan of the head. Special muscles, derived from the facial musculature (innervated by the N. Facialis or by a branch, the N. nasuloambulacralis3 that is unusually well-developed here) take part in the musculation of the snout. Beyond this, in one group (the hopsorrhines4 or Snout Leapers) the capacity of the snout to develop energy is increased further by M. longissimus dorsi, which is drawn out forwards over the skull. The nasal sinuses and the corpora spongiosa undergo far-reaching alteration and an increase in size, which are accompanied by a shift in function. Thus for instance in almost all of the more advanced forms the tear duct takes on the functions of an outer respiratory passage. Such special details will be considered further in discussing the individual species.
[Fig. l] Nasobema lyricum. Head of a
young embryo, to show the polyrrhine
condition (after Stulten 1949). |
Inasmuch as in these rhinograde animals ¢ with exception
of the genus Archirhinus (a Primitive Snouter) the nasarium5 serves
as a means of progression, the other appendages have lost their locomotory
functions.
Correspondingly, the posterior appendages are mostly more or less reduced,
while the anterior appendages have been modified as grasping organs for holding
the food or as little hands for grooming. In the genus Rhinostentor (Trumpet
Snouters) they participate in the formation of a water-filtering apparatus.
Whereas, then, the paired appendages are less impressive features of the general
organization of the Snouters, with these animals the tail occupies an outstanding
place and in its construction has developed manifold and altogether aberrant
types. Thus one finds not only coiling tails and lasso-like tails, but also
in the sclerorrhines6 (the Proboscipedes) the tail serves the
more primitive forms for jumping and the more advanced as a grasping organ
(cf. pp. 35, 36,
and 48).
In most Snouters the body is covered with a fairly uniform coat, in which no
distinction can be made between underfur and guard hairs; this is to be attributed
not only to the climatic conditions of the archipelago, but according to Bromeante
de Burlas is to be regarded as a primitive character. This view is favored
also by the regular way in which the hairs are grouped. In one genus there
are in addition strong horny scales (similar to those of scaly anteaters),
that are altogether of the nature of reptilian scales. At times the coloration
of the coat is magnificent. Famous above all is the extraordinary luster of
the pelt, caused by the special structure of the hair cortex. The bare spots
too ¢ head, feet, tail, ears, combs of skin on the head and especially on the
snout ¢ are brilliantly colored at times. A few aquatic species, and the very
small burrowing species that are found on the sandy shore, are completely bare;
likewise a single parasitic species (cf. p. 19).
The manner of feeding varies greatly among the different families, and even
within the same family or genus. Yet this is hardly surprising when one recalls
that besides a single aquatic shrew7 the Snouters are the only
mammals of the archipelago and have thus been able to take possession of all
ecological niches.
Most of the rhinograde animals, the average size of which is indeed small,
eat insects. But in addition there are also herbivores-especially fructivorous
species-and one predacious genus. To be mentioned finally as particularly specialized
forms are the plankton feeders that live in fresh water, and the burrowing
forms, among which are the tiniest vertebrates known. The crab-eaters among
the hopsorrhines can be derived readily from the insectivorous forms. A strange
case of symbiosis will be discussed in the systematic section (pp. 15, 37).
It is particularly remarkable that among the Snouters there is one flying genus
(with a single species), and that there are also sessile and parasitic forms.
Considering the habits and the structural organization of the animals, however,
one is not astonished that the number of species is rather large. In this connection
geological interest is attached to the excellent study by M. O. Jester and
S. P. Assfugl of the genus Dulcicauda8 (the Honeytails). These
authors were able to show that land bridges must have persisted for various
lengths
of time
between the different islands of the archipelago, and were able to estimate
the dates of their interruption (cf. also Ludwig 1954). All in all, the study
with this material of Rassen-kreise and their evolution (Rensch 1947) is especially
promising, even though in many areas there are great gaps that are hardly to
be closed even by the palaeontological approach, since the pertinent fossils
lie in deposits that have sunk beneath the surface of the sea.
In general the reproductivity of the Snouters is not high, which permits the
conclusion that the death rate also is low. So far as now is known there is
invariably a single young at a birth (the Snout Leapers with physiological
polyembryony are an exception). However, pregnant females occur all year long.
The period of gestation ¢ again with exception of the Snout Leapers ¢ is long
and lasts seven months on the average. Among the monorrhine forms the young
are so advanced in development at birth that they do not have to be suckled.
Correlatively, the mammary glands of these Snouters are vestigial or display,
in the genus Columnifax9 (Pillar Snouters), lactation
independent of a lactation hormone (cf. p. 15). In the polyrrhine genera, in
which the newborn are in
quite a dependent condition, there is a single pair of (mostly axillary) teats.
As a rule these species also possess a brood pouch, that is formed of skin
folds on the throat and is supported by cartilaginous bars descending from
the AdamÆs apple.
The rhinogrades have scarcely any enemies. In the interior of the islands the
only warm-blooded creatures besides the already mentioned swamp shrew (Limnogaloides)
are birds of the genus Hypsiboas10 (Megaphone Birds).
All these are of the size of songbirds and have occupied biotopes quite distinct
from those of the
Rhinogradentia. According to Bouffon and Sotprimarsch they are descended
from petrels, and in fact from forms close to Hydrobates. Reptiles are lacking.
There occurs only a single primitive amphibian species (Urobombinator submersus11),
whose gigantic larvae are devoured by the Hooakha-Hutchi at ceremonial feasts.
The slow-moving Nasobema species have enemies from their own ranks in the predacious
rhinogrades of the genus Tyrannonasus12. However, this genus
is confined to a few islands. For the most part, only the oceanic birds that
at certain seasons
breed on some of the smaller islands occasionally take a rhinograde. However,
those very species (for instance the Honeytails and the Pillar Snouters) that
live on the shore are protected from the birds' attacks, in part by poison
devices and in part by being inedible; and the Snout Leapers are in general
so agile that they are not to be caught by these birds.
Here attention may be called to yet another peculiarity of the Hy-yi-yi-an
fauna: the insects display a great number of very primitive forms. Thus the
cockroach-like kinds are represented by numerous structurally distinct types,
most of which can be placed among the Blattadae. Besides these there are also
a few more advanced insects, above all Hymenoptera, whereas Lepidoptera are
absent altogether. Hence, pollination is accomplished partly by Hymenoptera
(above all by the Pseudobombus species, that outwardly resemble bumble-bees
but that actually are related to the xylocopids), and partly by caddisflies
and cockroaches. There are no ants.
As a marked peculiarity the six-winged insects (Hexaptera of the superorder
Hexapteroidea13) should be mentioned; they are descendants of the
Palaeodictyoptera and have terrestrial larvae. These insects are mostly animals
of the open country;
i.e. except for a few species they avoid the thick virgin forests that clothe
the mountain slopes of the larger islands. Here too we may record the peculiarity
that the larger islands have some endemic species. These primitive forms are
wholly lacking in the smaller islands. Probably this is to be attributed to
the fact that the smaller islands (for instance Owmvussa or Sawabisi) are coral
islands and hence of recent formation, or because they do not afford sufficient
protection against the wind for inept fliers, so that an endemic species there
died out as the islands sank and grew smaller.
With respect to the systematic arrangement of the Snouters, the following considerations
apply:
As shown by the single species that still walks on all four legs (genus Archirrhinos = Primitive Snouters), they must have been derived from primitive insectivores.
In this connection the presence on Mairuvili of Limnogaloides is significant;
for this animal, that unquestionably is to be reckoned among the Insectivora,
has many features in common with Archirrhinos, so that it is not impossible
that both species may be traced back to a common ancestry.
For the rest, the systematic classification of the Snouters follows predominantly
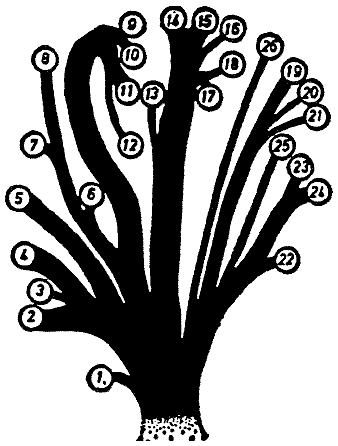
the degree of development of the snout. The ōfamily treeö in Fig. 2, showing
the systematic subdivisions of the order, was proposed by Bromeante de Burlas
(1950). According to it he distinguishes as principal groups Uni-Snouters that
walk on legs = Monorrhina pedestria (with Archirhinus haeckelii as
the sole species), Uni-Snouters that walk on the snout = Monorrhina nasestria (with
Soft-nosed Snouters = Asclerorrhina and Proboscipedes = Sclerorrhina),
and Multi-Snouters = Polyrrhina (with Short-nosed Snouters = Brachyproata and
Long-nosed Snouters = Dolichoproata). Whereas most genera can be accommodated
without difficulty in this scheme, it is still uncertain with regard to the
Mole Snouters = Rhinotalpiformes whether they can be united in a single
group with the Burrowing Snouters= Hypogeonasida14, or
whether they have been derived from the Sclerorrhina (Proboscipedes) and possess
a secondarily expanded nasarium.
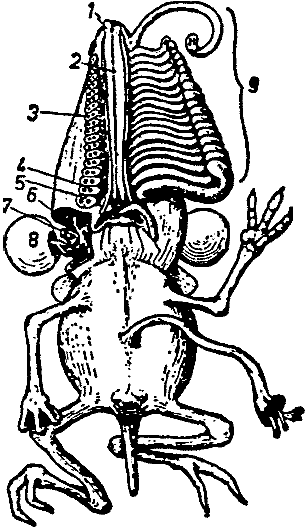
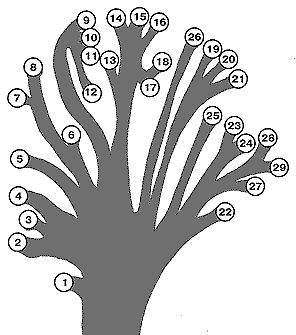
[Fig. 2] Proposed family tree of the individual genera of the Rhinogradentia (modified from Bromeante de Burlas, with the help of StultenÆs data). 1. Archirrhinos; 2. Nasolimaceus; 3. Emunctator; 4. Dulcicauda; 5. Columnifax; 6. Rhinotaenia; 7. Rhinosiphonia; 8. Rhinostentor; 9. Rhinotalpa; 10. Enterorrhinus; 11. Holorrhinus; 12. Remanonasus; 13. Phyllohoppla; 14. Hopsorrhinus; 15. Mercatorhinus; 16. Otopteryx; 17. Orchidiopsis; 18. Liliopsis; 19. Nasobema; 20. Stella; 21. Tyrannonasus; 22. Eledonopsis; 23 Hexanthus; 24. Cephalantus; 25. Mammontops; 26. Rhinochilopus. The thickness of the branches indicates the relative numbers of species of the several genera. Here Dulcicauda and Dulcidauca that elsewhere are regarded as separate genera are placed together under Dulcicauda. |
The 14 families contain altogether 189 species; there is still
a possibility that one or another unknown species inhabits some isolated region
of the archipelago. This may be anticipated the more because the just-mentioned
group of the Rhinotalpiformes has yielded surprising discoveries of new types
in recent years. Some systematic difficulties are to be expected also when
the question is clarified for broadly distributed species as to when we are
dealing with true races, i.e. genetically distinct populations, and when merely
with local modifications. The example of Mammontops15 (Shaggy-faced
Snouters), that originally was native to Shou-wunoonda and later was kept by
the Naval
Administration in the zoo at the experimental station on Shay-nay-lukha (Schaielacha),
demonstrated the extraordinary degree to which the phenotype could be modified.
Genetic experiments have failed hitherto because of the difficulty of rearing
the animals (cf. p. xix). Only Hopsorrhinus once again affords an
exception. However, here experiment last shown that the different island forms
are true
¢ though closely related ¢ species. Only when Hopsorrhinus aureus (the
Golden Snout Leaper) from Mitadina was erased with Hopsorrhinus macrohopsus16 (PikeÆs
Snout Leaper) from Hy-dud-dye-fee were there a few of the offspring with a
limited capacity for reproduction. Hopsorrhinus mercator17 (=
Mercatorrhinus galactophilus18, HealeyÆs Snout Leaper) too has
proved favorable for genetic
experimentation; with a gestation period of only eighteen days it usually bears
eight young of a single sex and is very readily maintained on commercially
obtainable condensed milk for babies (cf. p. 37).
3 nasulus Lat. = a little nose; ambulare Lat. = to walk.
4 hopsos Grk. = a jump or leap (a word, found only in Chrysostomos of Massilia, that apparently is to be traced back to a West Germanic root); rhis, rhinos Grk. = a nose.
5 Bromeante de Burlas designates as nasarium the entirety of the rhinal ambulacrum, irrespective of the origin of its components. Hence ōnasariumö is a functional rather than a morphological concept. Since this term has become entrenched in the literature and its use avoids the necessity of lengthy descriptions, it will be employed throughout the passages that follow,
6 skleros Grk. = hard.
7 Limnogaloides mairuviliensis B. d. B. (the Mairuvilian swamp shrew) is a primitive insectivore. That it belongs to the true shrews has recently been contested. On this account too the earlier name Limnosorex mairuviliensis has come into disuse. The dental formula, the well-developed zygomatic arch, the unusually small forebrain, and the presence of intervertebral muscles along the entire length of the tail are regarded as especially primitive characters.
8 dulcis Lat. = sweet; cauda Lat. = tail.
9 columnar Lat. = pillar; fax Lat. = making.
10 hypsiboas Grk. = Loud crier (Doric form of Attic hypsiboes).
11 ura Grk. = tail; bombina Lat. = toad; submersus Lat. = submerged.
12 tyrant-nose.
13 six-winged insects.
14 hypo Grk. = under; gea Grk. = earth.
15 mamont, Russ., borrowed from Palaeoslberian languages = mammoth. The spelling mammonta is intrinsically erroneous, but correct according to the rules of nomenclature, -ops Grk. = face. In the figure (Fig. a) the spelling accepted in the years 1952-56 has been used.
16 makros Grk. = big; hopsos cf. Footnote 4.
17 mercator Lat. = a merchant.
18 galactophilus Grk. = fond of milk.
SUBORDER: Monorrhina (Uni-Snouters),
SECTION: Pedestria (Foot-Walkers),
TRIBE : Archirrhiniformes (Primitive Snouter Types),
FAMILY: Archirrhinidae (Primitive Snouters sens. lat.),
1 GENUS: Archirrhinos (Primitive Snouters proper),
1 Species.
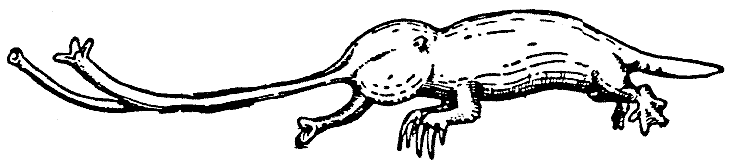
 Archirrhnos
haeckelii (HaeckelÆs Primitive Snouter) is the
only living representative of the Primitive Snouters.* It still walks on four
legs like other mammals
and still lacks a highly differentiated nasarium. Thus the snout
is altogether unadapted to function as a locomotory organ and merely serves
the animal
as a support when it is devouring captured prey (cf. Plate II, animal in
background)
. The habits of the Primitive Snouter resemble in many respects those of
a shrew; whereas in the daytime it sleeps in a simple nest under roots,
at twilight
it emerges in search of food. Then the mouse-sized creature with its thick
head and enormous snout may be seen running about and chasing the huge
cockroaches, large numbers of which often are gathered around the banana-shaped
berries
that have fallen at the foot of the Wisoleka trunks. When a Snouter
has caught an insect, it dives swiftly onto the snout, the margins of which
spread
quickly
apart to form a broad supporting surface. Viscous nasal mucus provides
firm hold of the ground. And now the greedy beast brings the food very rapidly
to its mouth with all four feet. From afar the feasting creatures betray
their
presence with loud chattering squeaks. Once the meal is done they reverse
from their headstand just as quickly, the margins of the snout are rolled
inward
again, and the chase is resumed. Little is known as yet about the reproduction
of these animals, since they occur only in the inaccessible mountain forests
of Hy-dud-dye-fee.
Archirrhnos
haeckelii (HaeckelÆs Primitive Snouter) is the
only living representative of the Primitive Snouters.* It still walks on four
legs like other mammals
and still lacks a highly differentiated nasarium. Thus the snout
is altogether unadapted to function as a locomotory organ and merely serves
the animal
as a support when it is devouring captured prey (cf. Plate II, animal in
background)
. The habits of the Primitive Snouter resemble in many respects those of
a shrew; whereas in the daytime it sleeps in a simple nest under roots,
at twilight
it emerges in search of food. Then the mouse-sized creature with its thick
head and enormous snout may be seen running about and chasing the huge
cockroaches, large numbers of which often are gathered around the banana-shaped
berries
that have fallen at the foot of the Wisoleka trunks. When a Snouter
has caught an insect, it dives swiftly onto the snout, the margins of which
spread
quickly
apart to form a broad supporting surface. Viscous nasal mucus provides
firm hold of the ground. And now the greedy beast brings the food very rapidly
to its mouth with all four feet. From afar the feasting creatures betray
their
presence with loud chattering squeaks. Once the meal is done they reverse
from their headstand just as quickly, the margins of the snout are rolled
inward
again, and the chase is resumed. Little is known as yet about the reproduction
of these animals, since they occur only in the inaccessible mountain forests
of Hy-dud-dye-fee.
PLATE II |
* In contrast with all other Snouters, fossils are known of Archirrhinos or of closely related forms. On the island Owsa-dowsa (Ausa-dausa), that is tectonically exceptional in the archipelago there are Early Tertiary deposits that might perhaps be assigned in part to the Upper Cretaceous also. In excavations of these there have been found teeth of an archirrhine that must have been the size of a house cat.
SUBORDER: Monorrhina (Uni-Snouters),
SECTION: Nasestria (Snout-Walkers),
TRIBE: Asclerorrhina (Soft-nosed Snouters),
SUBTRIBE: Epigeonasida19 (Surface Snouters),
FAMILY: Nasolimacidae20 (Snail-like Snouters),
GENUS: Nasolimaceus (Slime Snouters),
4 Species,*
GENUS: Rhinolimaceus (Sugarmice),
14 Species.
* Here, as in subsequent sections, only a single especially typical representative is named and described. For detailed date reference is made to the work by Bromeante de Burlas (1951) and to the somewhat shorter monograph by J. D. Bitbrain (1950).
The monorrhine Nasestria, with the tribe Asclerorrhina, are
closely related to the archirrhines. Without exception they are animals
in which the
modifications of the nasal region made in connection with an
altered mode of locomotion
have remained within narrow limits. The significant features
comprise enlargement of the snout and of those parts of the skull that serve
to support it.
However, the following structural peculiarities are to be regarded
as new developments:
multiple subdivision of the muscles of the snout, and of the
nasal
sinuses, whereby is formed a system of air chambers that intercommunicate
but
that can
be closed off from one another by special muscles; further the
marked development of the corpora spongiosa, that provide the
snout with
the turgor essential
for its functions. In most species this can be varied at will.
Besides, the muscles of those portions of the face adjacent to
the snout are
in part extended
over the snout itself and differentiated extensively so that
the snout (which according to Bitbrain is already a true nasarium)
has gained
considerable flexibility. A further characteristic is the great
expansion of the secretory
epithelium,
the voluntarily regulated mucus production of which is important
for the locomotion of the animals and for their fastening themselves
securely
in
place.
As already mentioned in the General Section, the appendages
are reduced or modified. The hindlegs are vestigial (but are never lacking
entirely) and are
practically without function. The anterior appendages serve both for seizing
the food and for grooming.
Nasolimaceus palustris21 (the Fadelacha Slime Snouter) will be
described as a typical representative of the nasolimacids. It is also most
closely related
to Archirrhinos. This approximately mouse-sized creature, with a vivid golden-brown
pelt, occurs on Mairuvili and there inhabits the muddy banks of the Fadelacha.
It has a short but broad snout, the anteriorly directed ventral surface of
which is modified as a creeping foot and functions much like the foot of Helix,
with the difference however that the locomotory waves of contraction succeed
one another more rapidly and are reversible. The speed of translation is greater
than would be expected from the mechanism of movement. When the animal is in
flight or is chasing something it will cover some 30-40 ft/min. Under such
circumstances it seems to glide with almost ghostly speed over the smooth damp
mud; and the precise mode of progression is no longer to be followed by eye
but can be recorded only by high-speed photography.
An impressive motion picture
of this has been made recently by F. Hyderitsch of the Scientific and Medical
Cinematographic Company, Black Goats. Snails of an endemic genus (Ankelella)
furnish the sole food of the Nasolimaceus species; only Rhimolimaceus fodiens22,
known to the sailors of the marine base as ōLucky Pierreö, digs out earthworms
(that in part belong to the same genera as the endemic earthworms of New Zealand
!). Before they lose their ōbaby teethö, the young feed mostly on
larval insects (chironomids), inasmuch as they are not yet able to cope with
the hard snail shells.
Mating, during which the male occasionally utters sounds as though he were
blowing his nose, takes place mostly at night. Then the male dashes in tight
curves around the female, who also is rotating. From time to time she too emits
a gentle ōHm-hmö. The stage for these games of love is usually a large flat
stone that is coated with a thin, slimy layer of diatoms and ever which the
water occasionally purls besides. The affair as a whole has a grotesque resemblance
to the dancing of a human couple on ice skates. The act of mating lasts but
a few seconds, and then the partners forsake one an-other, going very rapidly
in opposite directions with a sound as of gentle nose-blowing. After gestation
of twenty-six (?) months the female bears a single offspring that already resembles
the parents in all details and that, without further dependence on them, undertakes
a life of its own.
The animals do not patrol a constant range; they lead strictly solitary lives
and are tolerant of one another. In general, as Snail-like Snouters that are
adapted to muddy fresh-water beaches, they are confined to the bodies of water
where they were born, since they can neither swim nor will willingly cross
extended stretches of other than muddy or sandy substrates. Despite this, from
time to time one comes upon immature specimens-whose appendages at this stage
still are relatively more strongly developed-wandering slowly overland in search
for other bodies of water. The animals canÆt withstand sea water ¢ like many
other Snouters ¢ and on this account it is understandable that there are numerous
specific variations on different islands.
Closely related to Nasolimaceus palustris is Nasolimaceus conchicauda23 (the
Armor-tailed slime Snouter). This animal that as its name implies has an armored
tail, lives on the little volcanic islet Eeza-zofa (Isasofa), on the marsh
of the same name. By bending its tail ventral beneath it the animal can conceal
itself in it as though in a beach chair. It is significant that Eeza-zofa is
inhabited by a hypsiboant that poses a threat to the Snouter. This is the flightless
Hypsiboas fritschii (FritschÆs Megaphone Bird), an adept runner
and swimmer the size of a thrush, that feeds upon practically all animals it
can catch
and overcome.
The remaining nasolimacids are well protected against enemies by the fact that
at the base of the tail they have a gland that gives off a sweet secretion
(whence the name ōSugarmouseö). The fluid attracts a very small Pseudobombus species,
that is extremely ready to sting for the most part the Sugarmice are attended
by swarms of these insects and hence are protected.*
*According to studies by Shirin Tafarruj the glandular secretion contains only small amounts of glucose but in addition to them a sweet substance the constitution of which has still not been fully determined; structurally this is close neither to dulcin nor to saccharin and in pure form is some 200-times more stimulating than saccharin. It is noteworthy that this material has approximately equal sweetness for people and for the insect.
19 Epi Grk.= upon; gea cf. Footnote 14.
20 limax Lat. = a snail.
21 palustris Lat. = marsh-inhabiting.
22 fodiens Lat. = burrowing.
23 conche Grk. = a shell or shield; cauda Lat. cf. Footnote 8.
FAMILY: Rhinocolumnidae (Pillar-nosed Snouters sens. lat.),
GENUS: Emunctator (Snufflers)
1 Species,
GENUS: Dulcicauda (Honeytails),
19 Species,
GENUS: Dulcidauca24 (Sugartails),
1 Species,
GENUS: Columnifax (Pillar-nosed Snouters proper)
11 Species.
The position of the rhinolimacids in classification is still disputed: whereas Spasman and Stulten as late as 1947 still maintained that they were to be placed in a special Section (Sedentaria) opposite the Peripatetica, today people mostly follow Bromeante de Burlas and include them in the subtribe Epigeonasida. The principal reason for this is the discovery of Emunctator sorbens (the Snuffling Sniffler), that occupies a place intermediate between the errant rhinolimacids and the sedentary rhinocolumnids.
 On
the other hand it is not impossible that the rhinocolumnids are a polyphyletic
group. Bouffon (1954) only recently has once
again pointed out that then
are radical
differences between Emunctator and Dulcicauda on the one
hand and Columnifax on the other: 1. the innervation of the hyporrhinal
musculature is
fundamentally different in the two groups; and 2. the substances
found in the column
base (the pedestal on which Dulcicauda and Columnifax stand)
are
in part very
distinct. The prehensile threads of Emunctator and the supporting
base of Dulcicauda
both contain the so-called ōEmunctator-mucinö, that has in
it a pentose-containing mucoid sulphuric acid which is lacking
in
Columnifax.
On the other
hand, the Columnifax pedestal contains a pseudorhinokeratin
that does not occur
in the
other genera.
On
the other hand it is not impossible that the rhinocolumnids are a polyphyletic
group. Bouffon (1954) only recently has once
again pointed out that then
are radical
differences between Emunctator and Dulcicauda on the one
hand and Columnifax on the other: 1. the innervation of the hyporrhinal
musculature is
fundamentally different in the two groups; and 2. the substances
found in the column
base (the pedestal on which Dulcicauda and Columnifax stand)
are
in part very
distinct. The prehensile threads of Emunctator and the supporting
base of Dulcicauda
both contain the so-called ōEmunctator-mucinö, that has in
it a pentose-containing mucoid sulphuric acid which is lacking
in
Columnifax.
On the other
hand, the Columnifax pedestal contains a pseudorhinokeratin
that does not occur
in the
other genera.
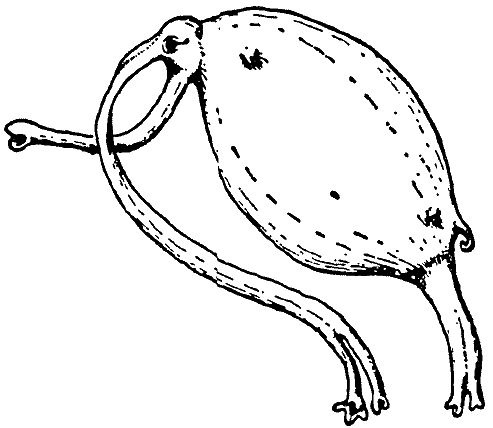
The Snuffling Sniffler, Emunctator sorbens (cf. Plate IV),
is an animal about the size of a small rat. It lives on Hy-dud-dye-fee
along the
banks of slowly
flowing brooks. There it clings to the stalks of plants that
rise
above the surface. Its manner of obtaining nourishment is most
peculiar; from the elongate
snout it blows long, fine prehensile threads that hang down
into
the
water and to which little aquatic animals get stuck. The prey
(mainly copepods
and larval insects, but also isopods and amphipods, and less
often small fish)
is in part ingested choanally by pulling up the slimy threads,
and is in part licked off the snout by the extremely long tongue.
The sluggish, dull animals possess, as a means of defense,
a long, very mobile tail with a poison gland at the tip; the
toxin
is secreted
into
a hollow
claw (derived from modified hairs). Since Emunctator lives
for the most part in
small groups, the little creatures can protect one another
by mutual tail-wagging.
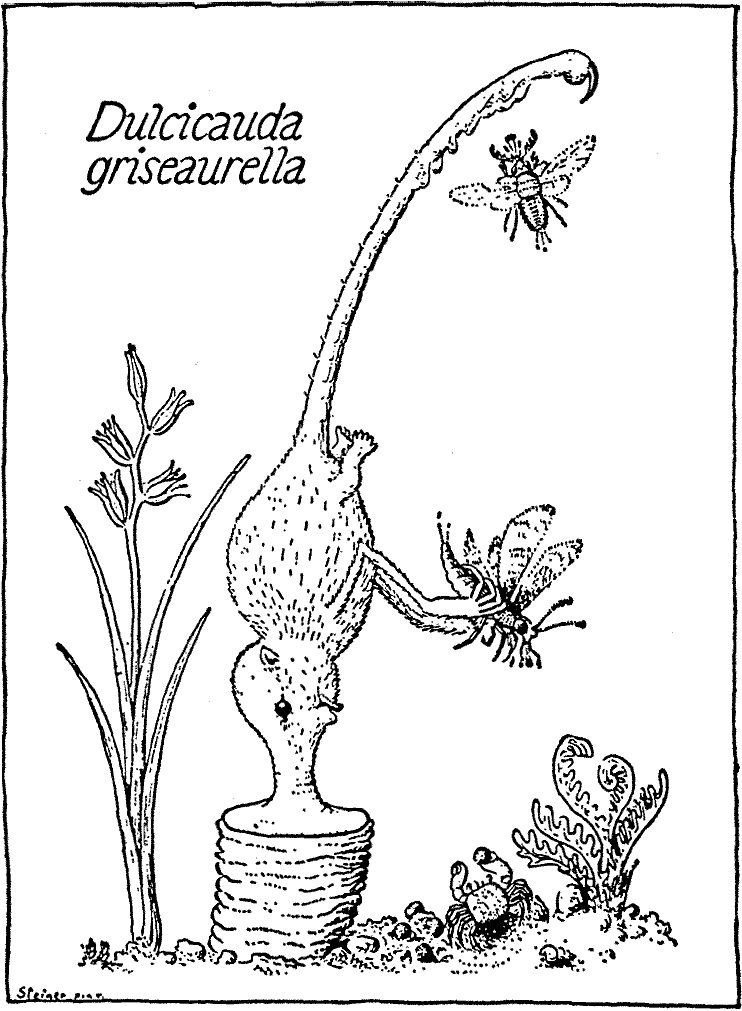
As a typical representative of the genus Dulcicauda, D. griseaurella (the
Greyish-Golden Honeytail) will be described (here as in what follows the
names assigned by
Bromeante de Burlas will be used, and hence the name of the
original author usually is not given). This species, together with D. aromaturus (the Perfumed Honeytail) is found on Mitadina,
D. griseaurella in the eastern
and D. aromaturus in the western half of the island.
The odd thing about these animals is that they are true sedentary forms that stand fast on their snouts and normally never forsake the place of attachment that they have chosen when young. Thus they remain supported on the snout, that gives off a reddish-yellow secretion which in time elevates the creatures (head+torso ca. 3 in. long, tail about 4 in.) on a conspicuous, pillar-shaped pedestal, known as the base or sella (cf. Plate V). Especially toward the tip, that bears a poison-claw, the tail has a wealth of dermal glands, that secrete a fruity, sticky product. Insects attracted to settle on the tail by the fragrance thus given out, stick to it and are picked off by the forepaws and brought to the mouth. At times, when the insects landing mostly are small they are not collected individually from the tail, but from time to time the animal pulls the tail through the mouth and licks them off.
|
|
|
|
|
Dulcicauda (from Tokiwa TakeshiÆs site) |
Dulcicauda lives in colonies on rubbly slopes near the shore.
These colonies regularly are associated with a small land crab
(Chestochele25 marmorata)
that feeds on scraps from the SnoutersÆ meals and carries
away their droppings.
At mating time the males descend from their pedestals and approach
the females, slipping and sliding along with their forelegs
in advance; once
copulation
has been accomplished they return to their pedestals. They
are en-abled to free themselves from the base of their column
through
a partial
dissolution of its uppermost layers by means of enzymes given
out by PusdivaÆs glands
of the discus nasalis (the same is true in regard to the dissolution
of the
fastening
mucus in Archirrhinos).
The genus Columnifax is distinguished in general by a reduction
in length of the prehensile tail. On this account the animals
are unable
to catch
their own prey. Only young specimens, not yet three months
old, still have a relatively
long, secretory tail and still nourish themselves in the same
way as Dulcicauda or Dulcidauca (Dulcidauca is characterized
by loss
of the
posterior appendages).
With older individuals there is established a most noteworthy
symbiosis with a hopsorrhinid: each of the eleven Columnifax species is associated
with
one
of the eleven subspecies of Hopsorrhinos mercator (HealeyÆs
Snout Leaper). The two partners are rigidly dependent on one
another,
and see to each
othersÆ nourishment: in the littoral zone where the two live
the predominant booty
of Hopsorrhinus mercator is small hermit crabs which, however,
it is unable to ingest because of its oral modifications (cf.
p. 37).
Hence
it turns
them over to the Pillar Snouter after it has suppressed the
latterÆs defense reactions
by means of definite sounds and gestures. (Columnifax defends
itself by squirting out a secretion from anal stink glands,
and with its
very mobile
snout is
able to twist some 180░ about the long axis). Thereupon Columnifax permits the hopsorrhinid
to suckle in connection with the described symbiosis, milk
is produced by individuals of both sexes, when over three months
old, without
reference to any sexual
function (cf. Plate VII).
24 Dulcidauca: an anagram of Dulcicauda, cf. Footnote 8.
25 cheston Grk.= dung (In Euphemios Thereutea of Alexandria); chele Grk. = a deft claw, crabÆs pincers.
SUBTRIBE: Hypogeonasida (Mud Snouters),
FAMILY: Rhinosiphonidae (Siphon Snouters Sens. Lat.),
GENUS: Rhinotaenia (Ribbon Snouters),
2 Species
GENUS: Rhinosiphonia (Siphon Snouters Proper),
3 Species
FAMILY: Rhinostentoridae (Trumpet Snouters Sens. Lat.),
GENUS: Rhinostentor (Trumpet Snouters Proper),
3 Species
The Hypogeonasida too are in themselves a relatively isolated group. They are all small unimpressve animals with originally a subterranean way of life that is most typically developed in the genus Rhinotaenia. As a characteristic representative Rhinotaenia asymmetrica (the Snorkeling Ribbon Snouter) will next be described.
[Fig. 3] Rhinotaenia asymmetrica. (Orig.) |
This animal lives
in the mud of a few small lakes as in the supralittoral regions of a few
gently flowing
streams.
There
it feeds principally
upon oligochaetes
and larval insects, that it digs out and gulps down
with its long, proboscis-like mouth. In doing this, Rhinotaenia daily
digs itself
forward one or two
yards it a depth of about a foot. Respiration is
made possible by the long nose,
that is extended siphon-like as much as 16 in.- i.e.
up to some four times the length of the animalÆs
head and
body
(cf. Fig.
3). The
asymmetry of
the snout - the left nostril with its terminal rosette
serves for inhalation and
the right for exhaling-affords a perfect supply of
air, despite the length of the air tubes.
Nothing is known regarding mating and reproduction.
Pregnant females and very small young are to be found
all year
long.
Beilig was able to extract from isolated Rhinotaenia snouts a mucin that is identical with that of Emunctator.
Morphologically
too
much can be
advanced in favor of the view that the hypogeonasids
derive from
Emunctator-like ancestors (cf. Bromeante de Burlas
1952 as well as Jerker and Celiazzini
1953).
The genus Rhinosiphonia differs from Rhinotaenia primarily in the finer construction of the snout
but does not
offer any further
distinctions from Rhinotaenia that require comment here. Instead we may discuss
here one Rhinotaenia species that
is unique in its peculiar parasitic habits.
[Fig. 4] Rhinotaenia tridacnae, shown at the age of sexual maturity. |
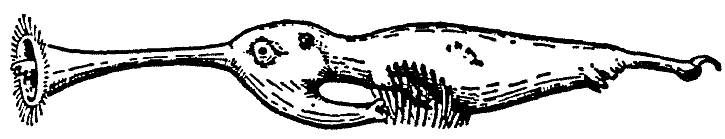
Rhinotaenia tridacnae (the Shell-inhabiting Ribbon
Snouter; cf. Fig. 4) is found in the tidal zone
throughout the
archipelago. The young
animals and
the males live in mud that is deposited in still
portions of the lagoons or that
collects in little clefts between the blocks of
coral. Even more
than with
the rest of the Rhinogradentia and especially with the hypogeonasids,
homiothermy is only very incompletely developed with Rhinotaenia tridacnae.
Connected with this is the fact that
R. tridacnae will for a considerable time withstand a more or less complete
interruption of oxidative metabolism Of course the creatures live in the upper
tidal zone, the mud of which is inundated for only from a quarter- to half-an-hour
at a time. Yet these animals can withstand being cut off from atmospheric air
for as much as three hours. They then fall into a kind of fit and ¢ bareskinned
as they are ¢ turn blue over the whole body, to regain a yellowish flesh color
as soon as they inhale air once more.
Now at high tide the mature R. tridacnae females enter the open shells of the
genus Tridacna (Giant Mussels) and burrow very swiftly down between the shell
and the mantle. There they soon cause a swelling of the mantle about as large
as a fist or a childÆs head, only a part of which, however, delaminates mother-of-pearl;
at low tide this growth is filled with air by the mussel and projects like
a hernia into the gill space. With its suctorial proboscis the parasite takes
in from the host both hemolymph and a part of the sexual products. Mating with
the Rhinotaenia males takes place at flood tide at night. Apparently the young
also are born at flood tide at night.
The rhinostentorids are closely related to Rhinotaenia, but have become adapted
to a submerged life and hence have undergone some modifications* that are expressed
most typically in Rhinostentor submersus26 (the Cladoceran-like
Trumpet Snouter).
* Here there is no attempt at an explanation in the sense of Boker, but merely a record of the observations.
[Fig. 5] Rhinostentor submersus. (Orig.) |
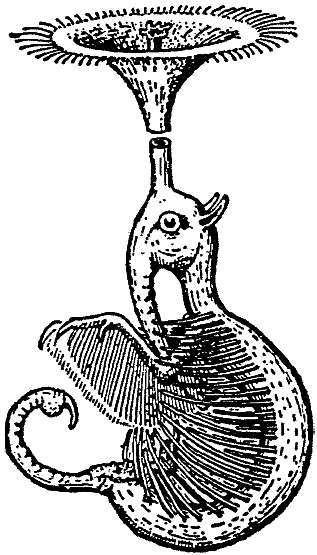
Rhinostentor submersus lives in various crater
lakes and brackish lagoons in the archipelago, and feeds there on plankton,
to wit principally on the leaf-footed
crab, Branchipusiops lacustris, that for the most part occurs here in numbers
that only occasionally are reduced by the ubiquitous Cladocera or Rotatoria.
Here Rhinostentor hangs, at a depth of 8-20 in., from a nasal
siphon that is constructed in general like that of Rhinotaenia but
that, in correlation with
an aquatic habit, exhibits an expansion of the nasal rosette. The rosette
grows funnel-like around the outgoing nasal passage while the afferent nasal
passage
is raised above it to form a small secondary rosette. The funnel-shaped
or trumpet-shaped
nasal rosette (cf. Fig. 5) is fringed with water-repellent hairs and at
the margins (modified wax glands) gives off a water-repellent, wax-like
coating,
so that the animal depends from this trumpet
as from a buoy. Along the sides of the otherwise naked body, there is a
row of stiff, thick bristles
that
ventrally form a kind of groove within which the anterior appendages, likewise
covered
with combs of stiff bristles, carry out rowing motions. The whole arrangement
works like the filtering apparatus of a Daphina (= water flea),
and with its oral proboscis the Rhinostentor removes from this filtration
device
the planktonic
organisms that are wafted in and caught there.
In Rhinostentor spumonasus (the Frothing Trumpet Snouter) the same relationships
prevail, with the exception that the creature hangs not from its nasal
trumpet but instead from a bubbly mass floated of from the latter; into
this mass
the Snouter withdraws for sleeping, for mating, and at signs of
danger.
[Fig. 6] Rhinostentor foetidus. (Orig.) |
A visitor to the archipelago is provided with a very impressive
experience by the foam floats of Rhinostentor foetidus (the Stinking
Trumpet Snouter; (Fig. 6), not only because they often float about in great
masses on the
most varied bodies of fresh water but because at times their unbearable
odor ruins
a sojourn at many of these otherwise so idyllic spots. R. foetidus lives
altogether within these rafts that it has made. Its ventral collecting
apparatus is reduced to a pair of small rakes with which the creature, creeping
in
regular paths about its raft and pulling itself along by the somewhat reduced
nasal
trumpet, gathers its food. This food consists of larval fungus gnats of
the genus Spumalyophilus, that browse on the mycelia that ramify
throughout the
floating mucus. The fungus, that has not been positively identified although
it is one of the Eumycetis, is nourished by the dying blue-green algae
that swam throughout the floating mucus. Thus there is a symbiotic system:
Rhinostentor, via the nutrients contained in the
mucus and excreted in the urine and feces, supplies the algae with a suitable
substrate, that
is enclosed
by the fungal strands. The algae assimilate this food and are then partly
sucked dry or metabolized by the fungus.* The fungus is eaten by the
larval fungus
gnats. A part of these larvae furnish the rhinostentorianÆs food.



Rhinostentor from Tokiwa TakeshiÆs site
It is of interest that the mucoid rafts of Rhinostentor spumonasus and Rh. foetidus are inhabited by a whole series of other animals; FritschÆs Megaphone Bird regularly makes use of them as a nesting site. The six-winged damselfly, Hexapteryx handlirschii deposits its eggs on them, and the larvae that hatch eat the larval fungus gnats. A whole series of Springtails (Colembola), among many other creatures, lives in the oxygen-rich air bubbles on the upper surface of the floats.
*Whether this constitutes a modified lichenaceous symbiosis is uncertain.
26 cf. Footnote 7.
SUBTRIBE: Georrhinidae (Burrowing Snouters),
FAMILY: Rhinotalpidae (Mole-Like Snouters),
GENUS: Rhinotalpa (Mole Snouters),
4 Species,
GENUS: Enterorrhinus (Gut Snouters),
5 Species,
FAMILY: Holorrhinidae (Allsnouters),
GENUS: Holorrhinus (Wholesnouters),
18 Species,
GENUS: Remanonasus27 (Dwarf Snouters),
1 Species
Although the Burrowing Snouters have a thoroughly primitive-looking nasarium, that doubtless has been derived from that of the ancestral Snouters, nevertheless they are remarkable in that in extreme cases the snout predominates heavily over the rest of the entire body. In this respect ¢ quite apart from the concomitant reduction of their whole structural organization ¢ they are unique, not only among Snouters but among all vertebrates.
[Fig. 7] Rhinotalpa phallonasus. (Orig.) |
Relationships closest to those that originally prevailed are shown by the genus Rhinotalpa (Mole Snouters); and from these we choose as a typical representative Rhinotalpa phallonasus, the Glansnosed Mole Snouter, which is native to Mairuvili (cf. Fig. 7). This is a creature the size of a mouse and has more or less the habits of a mole; i.e. it lives in passageways it has dug in the humid ground and feeds on soil insects and earthworms. Both fore- and hind-appendages are greatly reduced. The autopodium is very well preserved and together with the large claws serves both as a pushing device and as an anchor in the tunnel. The labor of digging, however, is carried out by the snout, that is equipped with large erectile organs and into which spacious air sacs (derived from the nasal sinuses) are incorporated; these last also contribute to the expansion of the snout. Not only around the thickest portion of the snout but also around the occiput and ventral to the lower jaws are wreaths of strong, posterioriy directed bristles; both wreaths can be extended and are concerned in locomotion, that occurs in the following phases:
l. the wreath of bristles on the throat is spread out, as are the claws of the appendages;
2. the snout is inflated by taking air in through the mouth and simultaneously closing the glottis (contributory is the ability to close off the nares);
3. the nasal wreath is extended; the air is blown out of the snout and the animal pulled ahead by contraction of M. retractor nasarii, after which Phase-l may be repeated.
The nasal erectile organs come into action
only in very hard ground and they have the special function of stiffening
and
expanding the anterior
end of
the snout. But in general the animal does as little digging
as possible and uses
passageways that are already at hand; as a result of
the way in
which they were excavated they are very firm-walled.
Along them the animal
glides
with remarkable speed, in the process gathering up, with
a rake formed by the
chaetae submentales28, the earthworms found
on the way; it then removes the worms from
the rake with the oral proboscis. The papillae basonasales play an important part in the perception of the prey.
Their innervation,
apart from that
from the sensory facial nerves, comes also from those
nerves that in related forms
supply JacobsonÆs organ, and hence the papillae are organs
both of
the chemical and of the contact-olfactory sense.
At the posterior end, Rhinotalpa possesses above the
tail a gland that serves for defense; this is especially
important
to the
little creature
because
it is not able to turn around and because its tunnels
frequently are inhabited by a small, aggressive species
of land crab
(Chelygnathomachus29 altevogtii).
Between the rudimentary hindlegs are also the teats;
the young ere pushed back
to them briefly after birth. In this particular, Rhinotalpa displays signs of primitiveness, inasmuch as other monorrhines
no longer
suckle their
young.
Even in Rhinotalpa there is already indicated a structural
peculiarity that has been realized to a far greater extent
in other representatives
of the
georrhinids: a tendency for filling the body cavity with
connective tissue. In Rhinotalpa this holds only for the pleural regions, so that here
the lungs are connected firmly with the pleural wall.
In Rhinotalpa
phallonasus this is less
the case than in the smaller, closely related R.
angustinasus (the Narrow-nosed Mole
Snouter), in which are to be found yet other structural
features that are
unknown elsewhere among mammals and that are connected
with the small
absolute size
of the creatures (cf. Fig. 8). Among these the following
peculiarities should be stressed: reduction of the relative
length of the
digestive tract; decrease
in pulmonary capacity; disappearance of the nares; lack
of hair; expansion of the ciliated epithelium, that in
Rh. phallonasus clothes only the
larger sinuses, so that here it reaches the basal external
region
of the snout;
simplification of the brain; reduction of the eyes; and
finally, as the most striking physiological
characteristic, complete loss of homoiothermy. Naturally
all these structural and functional peculiarities are
connected closely with
the animalÆs
habits; it does not live in solid ground, but in cavities
in
beds of coarse gravel.
Therein it goes back and forth much like Rh. phallonasus,
but with the difference that, in correlation with its
longish build
it is
able also
to undulate to
some extent. Significantly too, the creature does not
seek out only clefts that contain air but also those
that are
filled with
ground
water. In
them it takes water into the lungs, that are reduced
to simple sacs. The sinuses
serve as additional respiratory organs. During rapid
progression they too are filled rhythmically with water.
When the animal
is at rest,
surface respiration through the richly glandular epidermis
appears to suffice.

Rhinotalpa phallonasus from Tokiwa TakeshiÆs site

Rhinotalpa angustinasus from Tokiwa TakeshiÆs site
Developments already indicated in the Rhinotalpa are
much more fully emphasized in Enterorrhinus, the Gut
Snouter.
Representatives
of
this genus grow to
a maximal length of 0.7 inches and display extraordinarily
extensive reduction: of the extremities only the claws
are retained, and
their musculature can
no
longer be homologized with definite limb muscles. The
gut is linear. Lungs are lacking. The heart is a simple
circulatory
tube, and
its condition corresponds to that displayed elsewhere
by young
mammalian
embryos. The
entire body surface
is ciliated. The brain completely lacks subdivisions,
at least outwardly. Of the skeleton, only a weakly
developed notochord
can be distinguished;
this
extends dorsal to the gut and forward beneath the brain
into the snout. As to the sexual organs nothing is
known. The
kidneys are
protonephridia,
with
a single ciliated funnel on each side. This funnel
sticks out into an endothelial
sack that lies at the base of the snout within the
body cavity, which is filled with connective tissue. A genito-urinary
sinus no longer
is present.
The genus is represented by a single species on each
of the five big islands of the archipelago. There the
animals
live
on the
gravelly beaches of the
deltas of small streams in a restricted region that
is united narrowly by the salt
content of the ground water (ca. 0.6% - 1.4%). Within
such areas one often comes upon isolated snouts and
upon specimens
that,
in relation
to their
body size,
have either an extraordinarily large or exceptionally
small snout, so that one is led to suspect that multiplication
takes place
by fission
at the
base of the snout.
Were it not that the Mole-like Snouters were already
known and the relationship between Rhinotalpa and Enterorrhinus beyond question, never would one
have suspected that the animals now grouped together
in
the family
Holorrhinidae (Allsnouters) are Snouters. For these
are tiny organisms, a few millimeters
in size, whose structure has become so ōprimitiveö
that the thought of classifying them among the chordates
would
never
have occurred.
In the first holorrhine genus, Holorrhinus (Whole-snouters),
chordate structure has been retained in the following
particulars: a slender
notochord extends
the whole length of the snout and through the entire,
greatly reduced trunk. Present is a closed though reduced
circulatory
system that,
however, bears
early embryonic traits. Of the kidneys there is still
on each side the ciliated funnel mentioned above; it
opens
into an
endothelial ampulla. So far as sexual
organs have been found ¢ as yet only in the male ¢
they lie not far from
the rear of the animal, in a region when a band of
well-differentiated muscle indicates
the former position of the posterior limbs, that otherwise
an no
longer demonstrable. The musculature is inserted on
bluntly protruding body
angles, that play
some part in digging activity. On the other hand, a
whole series of structural features
give the organisms a stamp divergent from that of the
chordates: the lengthy nasal sinuses participate in
digestion, in
addition to the
short, straight
gut. They function as appendices or in a manner similar
to that of the endodermal caeca of many invertebrates,
and are
alternately
filled
with
or emptied of
food. The somatic muscles no longer are striated, but
smooth. The brain is markedly reduced, and during early
developmental
stages
the nerve
tube becomes
split into two strands of tissue from which there develop,
adjacent to the notochord, two chains of ganglia that
are connected by
cross-commissures. The body cavity is filled with connective
tissue. The outer surface,
as is
already
the case in Enterorrhius is covered with a ciliated
epithelium, within which are scattered mucous cells
that likewise
have originated from
the
nasal passages.
Additionially it is noteworthy that in the endothelial
spaces of the adrenals rows of ciliated cells bear
greatly elongated
cilia,
producing
an effect
like that of flame cells.
The 18 species of the genus Holorrhinus are distributed
over the entire archipelago, living there partly in
the sands
of the flood
plains and
partly in the brackish
waters of the coastal beaches. Locomotion normally
is caudad. Two species (Holorrhinus variegatus = the
Variable
Wholesnouter
and
H. rhinenterus = PinnochioÆs Wholesnouter)
live in streams; frequently they have been reared successfully
from apparently freshly deposited young, found as neurulae
in an epithelial
bladder,
and this has provided interesting information about
the structure of these
animals. Thus, in the course of this work it has been
shown that the eyes (these animals
are able to see) arise as evaginations of the sac-like
brain, but then persist as simple vesicular eyes while
the brain
is secondarily
losing
its central
cavity and is becoming a broad, buckle-shaped structure
the main ganglionic masses of which are disposed to
right and
left of
the esophagus.
Up till now only a single species of the genus Remanonasus30 has
been found, from the river sands of the Wisi-Wisi, a stream on
Mairuvili island. This
is a worm-like creature that reaches a maximal length
of 2 mm. What distinguishes Remanonasus menorrhinus31,
the
Turbellarian-like Dwarfsnouter,
from the
preceding genus, is above all the loss of the anus and of the system
of blood vessels. Likewise there is no longer a trace
of a notochord to be found. Unfortunately only males
of these animals have been discovered
up to the present. The kidneys no longer have any demonstrable ciliated funnel-like
formation, but appear to be constructed like protonephridia, with a single
huge cilia-bearing flame cell on each side.
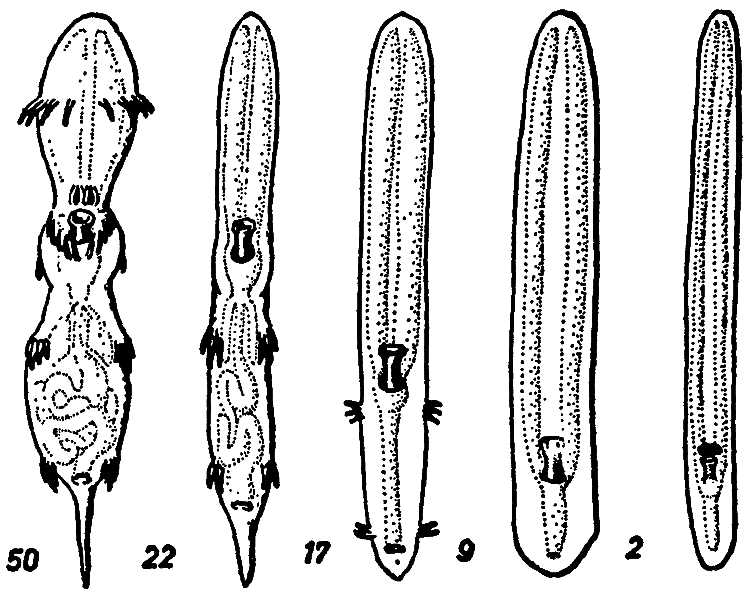
[Fig. 8] The anatomical series; Rhinotalpa phallonasus ¢ Rhinotalpa angustinasus ¢ Enterorrhinus dubius ¢ Holorrhinus ammophilus ¢ Remanonasus menorrhinus. The numbers show the over-all length in mm. Of the internal organs only the digestive tract is outlined. (After Mayer-Meier 1949) |
Hence it is not surprising that various scientists have
concluded that these animals are not to be counted among the Rhinogradentia
at all. Muller-Girmadingen
(1948) described them as Dendrocoelopsis minutissima and
sought to classify them as Tridadida; however, Mayer-Meier (1949) showed with
the help of careful histological investigations that particularly the mucus
cells could not be looked on as typically tricladid. Nevertheless he was forced
to grant that certain structural characteristics bore such great similarity
to those of the tricladid Turbellaria that at least it was not altogether impossible
that the tricladids had been derived from Remanonasus-like forms.
Thus, the seemingly cephalic position of the testicles in the tricladids ¢
that initially
strikes any unbiased observer as odd ¢ is comprehensible without difficulty
when one recalls that the holorrhinids crawl backwards and that hence their
caudal pole has become physiologically the anterior one. Furthermore, the form
of the digestive tract first becomes completely understandable when ¢ as illustrated
by the transitional forms of the morphological series (cf. Fig. 8) ¢ one derives
it from the gut+nasal sinuses of the georrhinids. Of course the entire question
can only be settled finally after animals with mature female sexual apparatus
have been found. That this apparatus is so complex in the tricladids and in
the turbellarians generally does of course suggest their derivation from more
highly differentiated animals. Remane (1954) too emphasizes this, but would
prefer to derive the Turbellaria from annelids. Stulten (1955) more recently
is inclined to the opinion that the Rhabdocoela might have descended from annelids,
but that the Tridadida (and the Polycladida, that in any case are to be derived
from the latter) possessed rhinogradentiate ancestors31a.
The tribe of the sclerorrhines (the Proboscipedes) constitutes a series of
the strangest and most beautiful species of the Rhinogradentia. Common to them
is the fact that the nasarium32 has become a leading organ, a
nasal leg, with which the animal can make mighty jumps; however, in view of
the gravitational
relationships (cf. Plate V), these are directed backward.
The most primitive conditions are found among the Arboreal Snout Leapers, the
perihopsids33, whose appendages still bear a certain resemblance
to those of the Archirrhiniformes. However, locomotorily to be regarded as
the most typical
are the Hopsorrhinidae, the Snout Leapers sens. strict., in which the hind
legs have disappeared except for slight remnants of the femur and tibia, and
whose snout serves as the sole organ of progression. Finally, in the Orchidiopsidae
(Orchidsnouter-like Snouters) the snout has become softened secondarily, in
correlation with their more sessile habits.
27 remanere Lat. = to remain; nasus cf. Footnote 3
28 An attempt has been made to trace the chaetae submentales back to the collecting basket of the rhinostentorids, and thus to derive the georrhinids from the hypogeonasids (Naquedai 1948). However, everything else in the organization of the two subtribes speaks against this conception.
29 chele cf. Footnote 25; gnathos Grk. = dentition; maMmenos or -machos Grk. = a warrior.
30 cf. Footnote 27.
31 menein Grk. = to remain; rhis cf. Footnote 4.
31a For the sake of objectivity, it should be noted that Grisse, in his foreword to Anatomie et biologie de lÆRhino grades (Masson & Cie, Paris 1962), contests that Remanonasus is a snouter. He prefers to take it as an endopsammic flatworm rather than a degraded Rhinograde.
32 cf. Footnote 5.
33 peri Grk. = around; hopsos cf. Footnote 4.
SUBTRIBE: Hopsorrhinida (Snout Leapers sens. lat.),
FAMILY: Amphihopsidae (Two-way Snout Leapers or Arboreal Snout Leapers);
GENUS: Phyllohopla (Leaf Leapers),
2 Species,
FAMILY: Hopsorrhinidae (Snout Leapers sens. strict.),
GENUS: Hopsorrhinus (Toothed Snout Leapers),
14 Species,
GENUS: Mercatorrhinus (Suctorial Snout leapers),
11 Species,
GENUS: Otopteryx (Earwings),
1 Species,
FAMILY: Orchidiopsidae (Orchidsnouter-like Snouters),
GENUS: Orchidiopsis (Orchidsnouters),
5 Species,
GENUS: Liliopsis (Lilysnouters),
3 Species.
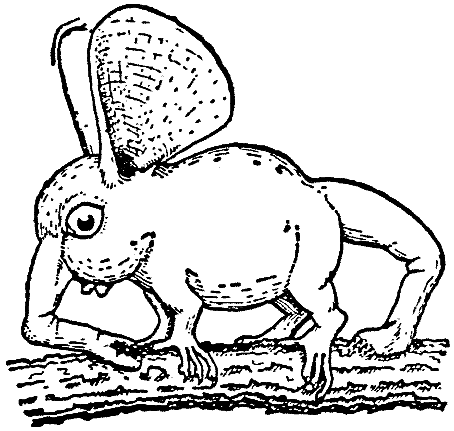
The Two-way Snout Leapers are creatures of the virgin forest
and live in the crowns of the trees, where they leap nimbly from branch to
branch
or creep
at leisure along the twigs. They are compactly built, and like most
Rhinogradentia are insectivores about the size of a mouse.
Whereas their torso and limbs have retained many archirrhiniform features,
immediately conspicuous on the large head with its great eyes is the
jointed snout, which ends distally in a dorsally located sole-plate
and which is
moved by powerful facial muscles and by a strong protractor, M.
extensor nasipodii
longus ( = M. longissimus nasarii). According to Stulten, M.
extensor nasipodii longus has been derived from the anteriorly
lengthened M.
longissimus dorsi or M. l. thoracis, as it indicated
by its innervation via thoracic spinal nerves (cf. also Fig. 11). Just
as oddly shaped as the snout
is the tail;
it too is
exceptionally muscular and powerful and bears a terminal sole-plate,
the strong bristles of which permit the tail to be braced firmly against
roughened
areas
of the substrate (Fig. 9). In addition to the metameric caudal muscles,
that as a primary character have always been retained among the Rhinogradentia
¢ a primitive feature to which Trufagura (1948) and Izecha
(1949) have already drawn attention ¢ it is above all M. iliocaudalis that
acts as an extensor of the tail. Now, by means of the snout and the tail the
periopsids are able to leap with improbable celerity back and forth through
the thicket of vines, now frontwards, now backwards, and then sidewise, so
that they are very hard to catch. This agility of theirs is at first hard to
comprehend, since they are practically without enemies. However, they live
together in small groups, within which there IS a perpetual rushing to and
fro, a chasing and a fleeing, that no doubt has to do with contests over their
pecking order, the sociological significance of which has not yet been fully
explained. In addition, their skill is of course useful to the animals also
in their gathering of nourishment; for they feed almost exclusively on flying
insects, that are snatched in the course of a leap.
[Fig. 9] Phyllohopla bambola. (Orig.) |
The Hopsorrhinidae, in contrast with the preceding group of Snouters, live
on the ground. As already mentioned, their hindlimbs are vestigial and no longer
visible externally. The snout is even further diferentiated than in the Perihopsidae,
in that it has undergone a division into three segments; next to the head (cf.
Fig. 10) is the nasur, which is connected by a joint with the nasibia, to which
the rhinanges finally are autonasally attached. Extension of the nasur and
the nasibia is by means of two separate bellies of M. extensor
nasipodii, while
the rhinanges are moved by the facial musculature and M. flexores longi and
breves of the nasipodium (that is by those both of the zygonasium and the autonasium),
which also are derived from the facial musculature.
As a result of ankylosis of the vertebrae and because of ventral stiffening
provided by the sternum and the processus styliformis34 of the
pubis, the torso is firmly encapsulated. The forelimbs are mobile grasping
organs. The tail
no longer subserves progression but is used for securing the food, that consists
primarily of amphipods, isopods, and little hermit crabs from the high tide
level. In consequence, the supporting caudal sole-plate has been modified to
constitute a par of grasping tongs, the claws of which have been formed from
modified or fused hairs and in cross-section present a histological picture
resembling that of rhinoceros horn tissue. With this tail the Snout Leapers
are most adept at pulling their prey out of the narrowest clefts and hiding
places. Their leaps, that at ordinary speeds of progression cover about one-and-a-half
body lengths but when fleeing or pursuing a mate or rival may be ten times
as great, regularly are rearward (cf. Plate VI). The jump can be guided to
some extent by slight movements of the huge ears.

Hopsorrhinus from Tokiwa TakeshiÆs site
The Snout Leapers belong to the most frequently encountered Rhinogradentia.
Everywhere on the shore they inhabit the stony coral patches, the sands of
the flood plains, or the volcanic or sedimentary gravel beds. It seems that
the stronger males maintain small harems and drive off weaker males. However,
the differences between the sexes are so slight that field observation has
not yet permitted analysis of details of behavior within the packs.
 Stulten
placed the first two genera of the Hopsorrhinidae under the single genus Hopsorrhinus,
However, more recently Bromeante de Burlas has reached the conclusion that
the genus Mercatorrhinus35 definitely deserves separation;
whereas the Hopsorrhinus species all feed in the manner described above and
in this connection possess the original type of dentition (whence also the
common name ōToothed Snout Leapersö), all Mercatorrhinus species no longer
are able to ingest solid food and are fully dependent on the symbiotic relationship
described above (p. 15) with the Pillar-nosed Snouters of the genus Columnifax.
This is apparent not only in the oral formation ¢ absence of teeth, reduction
of M. masseter and M. temporalis ¢ but also in the vestigial nature of the
forelimbs. Together with these reductions the animals, in correlation with
their symbiosis, have acquired some capacities that are lacking in the idiotrophic
forms. Thus, they are able to squat on the tail, which they coil beneath
them, and do so regularly whenever, after having handed over their prey,
they sit
down next to a Columnifax to await their nourishment. (For the collator [Steiner]
of these notes, however, De BurlasÆ arguments do not carry full conviction.
Therefore, following StumpkeÆs own example, the question of the genus Mercatorrhinus is left open.)
Stulten
placed the first two genera of the Hopsorrhinidae under the single genus Hopsorrhinus,
However, more recently Bromeante de Burlas has reached the conclusion that
the genus Mercatorrhinus35 definitely deserves separation;
whereas the Hopsorrhinus species all feed in the manner described above and
in this connection possess the original type of dentition (whence also the
common name ōToothed Snout Leapersö), all Mercatorrhinus species no longer
are able to ingest solid food and are fully dependent on the symbiotic relationship
described above (p. 15) with the Pillar-nosed Snouters of the genus Columnifax.
This is apparent not only in the oral formation ¢ absence of teeth, reduction
of M. masseter and M. temporalis ¢ but also in the vestigial nature of the
forelimbs. Together with these reductions the animals, in correlation with
their symbiosis, have acquired some capacities that are lacking in the idiotrophic
forms. Thus, they are able to squat on the tail, which they coil beneath
them, and do so regularly whenever, after having handed over their prey,
they sit
down next to a Columnifax to await their nourishment. (For the collator [Steiner]
of these notes, however, De BurlasÆ arguments do not carry full conviction.
Therefore, following StumpkeÆs own example, the question of the genus Mercatorrhinus is left open.)
That all the Suctorial Snout Leapers can be cultured readily was already
noted at the beginning (p.xxiv), and depends on the ease with which artificial
substitute
food can be obtained: inasmuch as Columnifax milk is relatively
rich in sugar and low in fat content and thus greatly resembles human milk,
the animals
can without difficulty be fed on condensed milk used for tables. Yet this
is carried
out successfully only with the help of an artifice that was discovered by exhaustive
studies of the behavior of the animals.
When such a mercatorrhine is hungry, its drive to search for food is activated. The animal wanders about and sticks its tail into clefts and cracks in order to seize the aforementioned crabs. If it has taken prey, it then cautiously approaches a Pillar-nosed Snouter, and with characteristic leaps calls itself to the latterÆs attention. Not until a nearby Columnifax utters a grunt does the mercatorrhine come closer, and in fact approaches from the ventral side of the Columnifax. For its part, the latter keeps twisting constantly about its long axis at the approach of any animal whatever, always ready to squirt the secretion of its stink glands toward the potential attacker. But now Mercatorrhinus comes nearer in such a way that it makes repeated sidewise leaps to left and right, so that Columnifax can see it clearly. At intervals too it stands on its snout and with the tail holds aloft the prey, which it waves vigorously and vibrantly from side to side. Only when Columnifax ceases its lengthwise writhing and lets a long, guttural snuffling be heird, does Mercatorrhinus close in from the ventral aspect and hand over its medium of barter, its prey. Thereupon Columnifax examines this, to see that it is thoroughly fresh. If it is not, the Pillar-nosed Snouter at once assumes a defensive stance and squirts stinking secretion all over the bearer, unless the latter has meanwhile leaped hastily away to a safe distance. Only when the ōgoodsö are beyond reproach does Columnifax offer its breast to Mercatorrhinus, that then gives a little hop onto its coiled-up tail and begins to suck.
Now, with captured mercatorrhines it became apparent
that they could not be maintained without Columnifax. On the other hand, adequate
prey was to be
gathered only with difficulty. But since incidental observations had shown
that Mercatorrhinus did well on the artificial diet mentioned, forced feeding
was then tried. However, this is tedious and often the way active animals
were injured fatally in the process. Only meticulous observation of the rigidly
determined ōbargaining riteö provided the solution: it was noted that Mercatorrhinus,
even if it had found no food suitable for Columnifax, nevertheless would
occasionally still attempt to suckle. The above-mentioned ōhorrifiedö defense
reaction of Columnifax fails to occur when Columnifax is still sated from
previous meals and also has an excess supply of milk, the reduction of which
has a pleasurable aura. Under such circumstances the ōdeceitfulö Mercatorrhinus,
that for instance has proffered merely an empty snail shell with no Pagurus inside, may be given milk. On the other hand, Mercatorrhinus will not drink
without having first carried, out the entire ritual of exchange. It has to
have captured ōpreyö and have presented this, after having made the dancing
approach outlined above, before it will drink. Beyond this, the following
features on the part of Columnifax are essential: the shape of a wedge, thicker
at the top; yellow color; eye spots in the lower third; hissing sound; and
proper udder form. Besides it must take the ōpreyö from the donor. One of
BitbrainÆs colleagues succeeded in making a relatively simple electronically
operated device that met the requirements indicated. At maximal speed of
operation it is able to suckle 80 Mercatorrhinus specimens an hour. Empty
snail shells are used as ōpreyö; after they have been accepted by the Columnifax model they gravitate beneath the false bottom of the cage into cracks from
which they can again be fetched by Mercatorrhinus.
The peculiar Mercatorrhinus-fleas, that did great damage to the experiments
in the beginning, can be controlled effectively by means of sticky paper on
the underside of the upper layer of the double-bottomed cage (DDT and other
insecticides are too toxic to Mercatorrhinus).
The Earwing, Otopteryx volitans B. d B. (= Hopsorrhinus viridiauratus36 STU.), the only representative of its genus, is by its structure readily to
be recognized
as a modified hopsorrhine (Plate VIII). Actually this animal is distinguished
from its cousins only by the enormous size of the ears and the differentiation
and strengthening, in correlation with its flight ability, of the musculature
of the external ear. The one other difference, its vestigial tail, is a structural
detail of little import. In all other respects, Otopteryx is a typical hopsorrhine,
so that Stulten even hesitated to split it off from the other genera. However,
in addition to what has been said, the following points are to be cited in
favor of establishing a separate genus: the nasarium is extremely slender and
gracefully constructed. The muscles that move the rhinanges are in part reduced,
so that the animal is not able to run over uneven ground with the agility of
the hopsorrhines. On the other hand, the abductors of the rhinanges are especially
powerful; they serve to expand the autonasium, that functions as a steering
tail. On the head there should be mentioned further the development of special
bony ridges ¢ the seats of attachment of the aural musculature ¢ as well as
the os alae auris, which however is not a bone but a calcined fibrous
cartilage; and in addition the formation of air-filled lateral nasal sinuses
beneath and
within the bony ridges mentioned. In common with the hopsorrhines, Otopteryx displays inversion of the course of the hairs over large parts of the surface
of the rump.
In Otopteryx the iridescence of the pelt, that gives other representatives of the sclerorrhines the brilliance of metallic surfaces or of jewels, attains its highest expression, so that the animals can be compared only with tropical butterflies or hummingbirds. Hence it is a magnificent sight when with rapid strokes of its ears the animal dashes close above the flower-bedecked mountain meadows in pursuit of dragonflies or of Hexaptera, or climbs abruptly aloft into the blue sky, there to wheel in play with others of its kind. Most enchanting of all are the newborn ¢ still scarcely able to hold their ears out ¢ that stagger calopteryx-like about the flowers, on the search for small insects. The strangest thing about all this is that Otopteryx flies backward, though this too is comprehensible when one recalls that the flight of Otopteryx has been derived from the gliding of the hopsorrhines, that leap backward.
|
|
|
|
|
|
|
|
|
|
|
Otopteryx from Tokiwa TakeshiÆs site |

Especially peculiar and characteristic is the take-off and landing of the Earwings. The animal, standing on its flexed snout, first ōcocksö its ears, i.e. raises them vertically so that they touch one another; then flexes the deutonasal joint even more strongly, is in Hopsorrhinus (cf. Plate VI above); after which the several phases ensue as in the latter, with the difference that the leap is more vertical. Shortly before the jump reaches peak height, the ears are powerfully depressed. The fully-extended snout is spread wide in the autonasal region, and the animal flies. These individual phases can of course be analyzed only by high-speed photography. The process of flight itself is extremely rich in variety: when an adept insect is being pursued, or during playful flight maneuvers, great distances are traversed at blinding speed, with the ears beating uninterruptedly up and down at a rate of about ten strokes/sec. During searching flight, earbeats of equal frequency but of a small amplitude alternate with short periods of gliding. Along the slopes, in the usually breezy island winds, Otopteryx also is able to soar at length. At all events it does not often go high into the air, and for the most part remains at altitudes not above twenty yards. Its method of landing is peculiar, being rendered difficult by the fact that the snout must perform in a double capacity, both as a foot and as a steering tail; if an Earwing wishes to come to earth, it mostly approaches the landing place in a steep glide, with the ears held somewhat dorsad and nasad. When close above the ground it suddenly adopts a vertical stance with the hind end somewhat elevated, which results in an abrupt, temporarily upward arc during which the steering tail ¢ i.e. the tip of the snout ¢ almost touches the earth. In this position, in which the ears are strongly arched (M. inarcantes auris37) the animal glides forward for yet a short stretch close above the ground, losing altitude and velocity the while. Then it suddenly folds the nasal steering mechanism together, curves the snout ventrad, and after elevating the ears to their full extent lets itself settle elastically onto the snout, which by now is stretched far caudad. This last phase of the alighting process again bears a great resemblance to the landing of the hopsorrhines after a leap (cf. Plate VI above, phases 6-8).
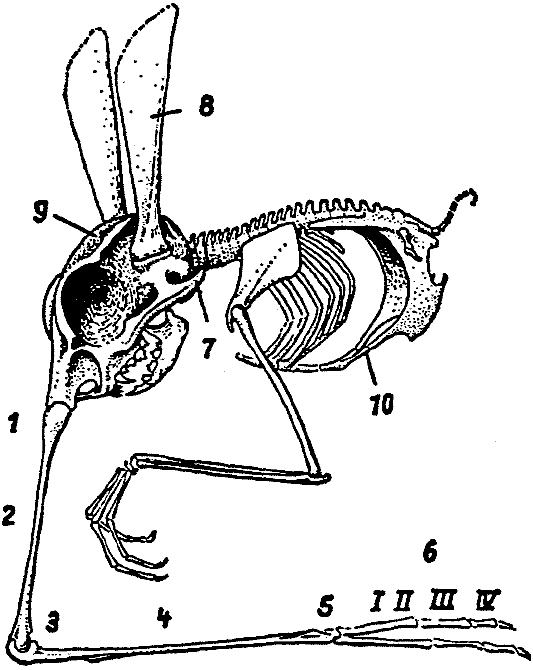
[Fig. 10] Otopteryx volitans, skeleton. 1. Articulatio nasofrontalis; 2. Nasur; 3. Articulatio deutonasalis; 4. Nasibia; 5. Articulatio carponasalis; 6. Rhinanges (= Nasalges) I ¢ IV; 7. Processus jugalauris; 8. Os alae auris (= Cartiago aeroplana), 9. Christa temporalis; 10. Processus pubici (Orig.)

(photo inserted for electronic edition)
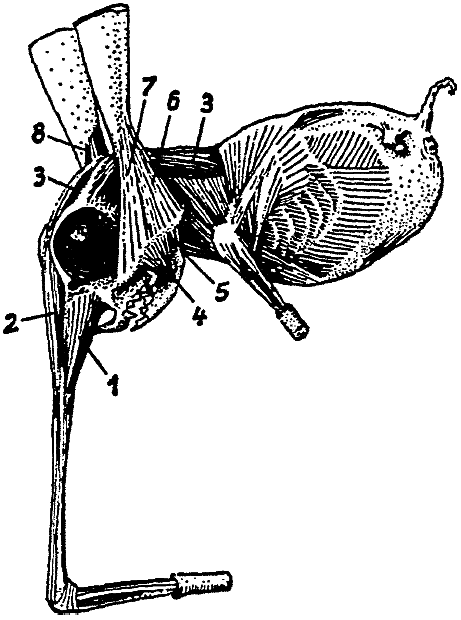
[Fig. 11] Otopteryx volitans, musculature.
1. M. lacrymonasuralis; 2. M. extensor nasipodii superficialis; 3. M. extensor nasipodii longus; 4. M. masseter; 5. M. depressor mandibulae; 6. M. aeroplano-jugalairis posterior; 7. M. aeroplano-jugalauns anterior; 8. Levator aeroplanae.To the right of 3, M. extensor nasipodii has been bared by partial removal of M. trapezius cervicalis. (Orig.)
The way in which Otopteryx has solved the problem of locomotion ¢ from the morphological standpoint a most extraordinary solution ¢ calls for a comparison with the other flying forms of the animal kingdom. With exception of the Rhinogradentia themselves, true fliers have appeared only four times altogether: the insects, the flying reptiles, the birds, and the bats. Among these, the insects, whose organs of flight are supplementary additions not made at the expense of terrestrial locomotion, actually have found the most complete solution. The bipedal gait of birds likewise permitted great mobility both on the ground and in the air, even though in actuality the wings were ōstolenö from the mechanism for progression on land. Among the flying reptiles and the bats the power of flight arose at the cost of locomotion on foot; and therefore the two group were not and are not fully qualified to compete with the others just mentioned. But in Otopteryx now the situation is just as favorable as in insects, i.e. the ears are actually supplementary instruments of flight. Nonetheless, Otopteryx, with the high degree of limb reduction that preceded flight, has been derived from animals that were strongly specialized in a single direction ¢ yet in their nasal ōmonopolyö they are quite comparable to hopping birds. In any case Otopteryx enjoys a clear advantage over flying reptiles or bats; for it is a most skillful leaper, and the participation of the snout in flight has not limited its usefulness for terrestrial progression to the same extent as has occurred with the anterior limbs of these other groups. Whether Otopteryx could hold its own in sharper competition with continental animals is questionable. At all events it has scarcely any enemies in the islands. Neither the indigenous Megaphone Birds nor the seabirds, that are numerous from time to time along the coast, can catch it in flight. In agreement with this is the fact that one rarely finds pregnant specimens. The period of gestation is at least as short as with the hopsorrhines. Only a single foetus is carried at a time (Harrokerria and Irri-Egingam). It is suspected that the females bear two young per annum.
The Earwings are not to be kept in captivity, because they remain too easily
frightened, bruise the buttocks in their wild leaps and attempts at flight,
and soon succumb to the infections that arise in the wounds.
The Orchidsnouters (Orchidiopsidae) can be traced back to hopsorrhine ancestors
that exchanged the terrestrial way of life for an arboreal one, but that already
possessed the various hopsorrhine reductions ¢ in particular the disappearance
of the hindlimbs. Hence, adept climbers could no longer be developed from them.
Instead there must have been initiated an evolutionary course in which the
animals no longer leaped from place to place but rather climbed slowly about
with the help of the forelimbs and the tail. Representatives with a functional
nasal leg are at all events no longer known, and in their habits and construction
the Orchidiopsis and Liliopsis species of today are so highly specialized that
initially their derivation from Snout Leapers was not even considered (cf.
Gaukari-Sudur, Bouffon and Paigniopoulos). However, in the meantime some developmental
data of evolutionary significance have been gathered, and from them it is clear,
without possibility of contradiction, that a nasur and a nasibia as well as
rhinanges are at first laid down in the embryonic Orchidiopsis snout but are
later resorbed, so that the snout of the fully developed animal must be regarded
as having been softened secondarily. (Bouffon and Zapartegingarri 1953 write:
ōLes embryons des Orchidiopsides ne manquent point m de nasur ni de nasibie,
mais pendant le develop fement [longueur de lÆembryon eniiron 15 a 18 mm] il
y a lieu un ramollusement progressif de ceux-ci, de telle facon, que le nouveau-ne
ne montre plus ancune trace dÆossification dans son nasarlum epiati et petaloide.ö*)
It is especially to Bouffon and his school that we are indebted for the further
explanation of the organizational peculiarities of the Orchidiopsidae. Thus
Bouffon and Lo-Ibilatze-Sudur were able to show that the secretory attractant
(ōmucus attirantö) of the Orchidiopsis species is not formed by the outer nasal
surface, which in fact lacks any glandular cells that could be thus involved.
Rather, it is produced by the great glandular fields of the inner surface,
along the nasal passages, and is thereafter spread manually over the outer
surface of the snout. Furthermore, the same authors succeeded in demonstrating
that the grasping mechanisms of the tail are homologous structures in Orchidiopsis and
in Hopsorrhinus. And finally Asteiides, a pupil of BouffonÆs, was able to bring
proof that in the snout of orchidiopsids there are vestiges of M. extensor
nasipodii, that during embryonic development have the location characteristic
of the hopsorrhine Rhinogradentia.
*Translation: (Bouffon and Zapartegingarri 1953 write; ōEmbryonic orchidiopsids lack neither a nasur nor a nasibia, but during development [length of the embryo from 15 to 18 mm] these undergo a progressive softening such that the new-born animal no longer shows any trace of ossification in its flattened, petal-like nasarium.ö)
Ankers Vanilla-perfumed Orchidsnouter, Orchidiopsis rapax38, the best-known representative of the family, lives in the forests of Mitadina, especially at the level of the higher tree crowns, but also at more moderate elevations where windfalls or floods have left openings in the woods (Plate IX). Ordinarily the animal stands motionless on its tail and hence, seen from afar, bears a certain resemblance to a big flower39. The genesis of the similarity is that the huge ears, the medial dermal comb, and the flattened snout, protrude with vivid colors like floral petals from the head, whereas the inconspicuous green torso is at first inapparent. The already mentioned attractant secretion on the snout smells of vanilla and acts as an odorous lure. Insects that settle on the snout or that flutter close above it are snatched like lightning with the prehensile paws at the ends of the long, thin arms, and are conveyed to the mouth. Occasional changes of location are made with chameleon-like slowness, with the help of the forepaws and the two-clawed grasping tail. As to the mutual interrelationship among the several individuals of a species nothing is known; however, about a dozen pregnant specimens carrying embryos in various stages of development have been taken up to now (cf. above).

Orchidiopsis from Tokiwa TakeshiÆs site
Among the three Liliopsis species, that are distinguished from Orchidiopsis by the position of the ears and the cephalic combs, there is one that sleeps by day and ōbloomsö, i.e. maintains its predacious pose, at night. This animal, commonly called the ōglowing lilyö in English and the ōWundernaseö (= Miracle Snouter) in German (Liliopsis thaumatonasus), is extraordinary in that its mucous attractant gives off light. As with the luminescent slime of other animals (cf. Buchner), in the ōglowing lilyö too the light evidently is produced by symbiotic bacteria. At all events the tiny slime organisms that are held responsible have not yet been cultured, and electronmicrographic magnifications have failed to confirm their cellular nature beyond question.
35 cf. Footnote 17.
36 viridi-auratus Lat. = greenish-gold.
37 inarcare Lat. = to arch.
38 rapax Lat. = rapacious.
39 Since orchids do not occur on any island in the archipelago, the name ōOrchidiopsisö actually is an unfortunate choice; there is in fact no orchid-mimesis in the situation. At all events, thise flowers (Rochemontia renatellae ST.) that are the models for the simesis of Orchidiopsis are superficially very similar both in appearance and odor to orchids, although they are closely related to the Ranuiculaceae.
SUBORDER: Polyrrhina (Multi-Snouters),
PHALANX: Brachyproata (Short-nosed Snouters),
TRIBE: Tetrarrhinida (Four-Snouters),
FAMILY: Nasobemidae (Snout Walkers sens. lat.),
GENUS: Nasobema (Snout Walkers),
5 Species,
GENUS: Stella (The Small Snout Walker),
l Species,
FAMILY: Tyrannonasidae (Predacious Snouters sens. lat.)
GENUS: Tyrannonasus (The Predacious Snouter),
l Species.
 As
the name indicates, the Polyrrhina are set apart by the possession of several
snouts. In this feature they exemplify a peculiarity
that, though
it appears
as an element foreign to mammalian systematics, nevertheless
¢ if considered generally ¢ is repeated in other phylogenetic
series of animals,
i.e.
a multiplication of organs that in more or less closely related
forms occur
singly or in single
pairs, or that are present in only a few of these related
forms. As is well-known, one can ¢ at least in thought ¢ derive all the
Articulata from polymerous
creatures whose counterparts or whose earlier ancestors are
oligomerous; similarly there
may be mentioned here the variable number of gill slits among
the
relatively
closely related lower vertebrates. Now, the multiplicity
of the snout in the polyrrhine Rhinogradentia appears ¢ purely formally
¢as the
mere
multiplication
of an organ, at an early embryonic stage (cf. Fig. 1). At
all events it is not proper to cite it simply as an example of multiple
malformation
or mismodification
and to draw a parallel between it and what is known from
the morphogenesis of aberrant Drosophila mutants, as Knaddle and Kicherling
have
tried to
do. Middlestead and Hassenstine very rightly have emphasized
that with simple
duplications
the independent movement of the several snouts would necessarily
be impossible, since one knows through the investigations of
P.
Weiss among
others that
such duplications receive identical motor impulses. Thus
among the Rhinogradentia polyrrhiny is accompanied by a corresponding
central nervous
coordination
of
an equally great degree of differentiation. From the evolutionary
standpoint this situation holds within it significant difficulties,
when one recalls
that the Rhinogradentia must have arisen at the earliest
in the Upper Cretaceous. According to Remane (1954) it must be taken as
an
indication of relative primitiveness that among the polyrrhines
there exist at least three groups with respect to the condition
of polyrrhiny:
those
with four, those with six, and those with 38 snouts, quite
apart from the various
types of nasal differentiation within these groups. Also
it is to be assumed that the separation of these groups took
place
very early;
just as the
separation of the polyrrhines from the monorrhines must have
occurred very early. In
the current state of investigation it is even difficult to
trace
the
polyrrhines back to Archirrhinos or to archirrhiniform
primitive Rhinogradentia. It
is altogether erroneous to try to trace the polyrrhines back
to nasestrian monorrhines,
as dÆEpp has attempted to do (Stulten 1949; Bromeante de
Burlas 1949). One of the main reasons for this judgment is that
the
structure of
the nasarium ¢ quite apart from the matter of polyrrhiny
¢ is altogether different, and that the reduction of the
hindlimbs is accomplished
in quite
a different manner,
moreover that the number of ribs and the development of the
zygapophyses of the vertebrae in the monorrhines is completely
aberrant (in
the direction of
the conditions seen in the Xenarthra, but of course as a
manifestation of
convergence), whereas the polyrrhines have more nearly maintained
the original relationships.
As
the name indicates, the Polyrrhina are set apart by the possession of several
snouts. In this feature they exemplify a peculiarity
that, though
it appears
as an element foreign to mammalian systematics, nevertheless
¢ if considered generally ¢ is repeated in other phylogenetic
series of animals,
i.e.
a multiplication of organs that in more or less closely related
forms occur
singly or in single
pairs, or that are present in only a few of these related
forms. As is well-known, one can ¢ at least in thought ¢ derive all the
Articulata from polymerous
creatures whose counterparts or whose earlier ancestors are
oligomerous; similarly there
may be mentioned here the variable number of gill slits among
the
relatively
closely related lower vertebrates. Now, the multiplicity
of the snout in the polyrrhine Rhinogradentia appears ¢ purely formally
¢as the
mere
multiplication
of an organ, at an early embryonic stage (cf. Fig. 1). At
all events it is not proper to cite it simply as an example of multiple
malformation
or mismodification
and to draw a parallel between it and what is known from
the morphogenesis of aberrant Drosophila mutants, as Knaddle and Kicherling
have
tried to
do. Middlestead and Hassenstine very rightly have emphasized
that with simple
duplications
the independent movement of the several snouts would necessarily
be impossible, since one knows through the investigations of
P.
Weiss among
others that
such duplications receive identical motor impulses. Thus
among the Rhinogradentia polyrrhiny is accompanied by a corresponding
central nervous
coordination
of
an equally great degree of differentiation. From the evolutionary
standpoint this situation holds within it significant difficulties,
when one recalls
that the Rhinogradentia must have arisen at the earliest
in the Upper Cretaceous. According to Remane (1954) it must be taken as
an
indication of relative primitiveness that among the polyrrhines
there exist at least three groups with respect to the condition
of polyrrhiny:
those
with four, those with six, and those with 38 snouts, quite
apart from the various
types of nasal differentiation within these groups. Also
it is to be assumed that the separation of these groups took
place
very early;
just as the
separation of the polyrrhines from the monorrhines must have
occurred very early. In
the current state of investigation it is even difficult to
trace
the
polyrrhines back to Archirrhinos or to archirrhiniform
primitive Rhinogradentia. It
is altogether erroneous to try to trace the polyrrhines back
to nasestrian monorrhines,
as dÆEpp has attempted to do (Stulten 1949; Bromeante de
Burlas 1949). One of the main reasons for this judgment is that
the
structure of
the nasarium ¢ quite apart from the matter of polyrrhiny
¢ is altogether different, and that the reduction of the
hindlimbs is accomplished
in quite
a different manner,
moreover that the number of ribs and the development of the
zygapophyses of the vertebrae in the monorrhines is completely
aberrant (in
the direction of
the conditions seen in the Xenarthra, but of course as a
manifestation of
convergence), whereas the polyrrhines have more nearly maintained
the original relationships.
Nevertheless the mono- and polyrrhines do exhibit one feature
in common; the expanded tear duct, that in many instances
serves as
a respiratory
passage. Bromeante de Burlas regards this as a result of
structural convergence, that
probably is related to the change in function of the snout
(s) and their cavities.
So it comes about that the distal nasal openings, in those
forms where they occur, mostly subserve special functions
that are
not connected
with respiration:
olfactory examination, the intake of food, and finally even
participation in the production of the animalÆs voice (cf.
p. 83).
Details regarding the construction of the nasarium cannot
be given in this concise report; for them we make reference
to
the studies
by Bromeante
de Burlas, Stulten, and Bouffon, as well as to the comprehensive
exposition of these matters
by H. Stumpke.
 The
Great MorgensternÆs Nasobame, the honatata of the natives (Nasobema lyricum;
Plate X), is the best-known representative of the polyrrhines
and on this
account will be discussed in somewhat more detail. As a
typical polyrrhine it has on
its short, fat head four equal snouts, that are fairly
long and on which ¢ as already
described by Morgenstern ¢ it walks. It is enabled to do
this, despite an inadequate nasal skeleton, because the
snouts are
rendered quite
rigid by
the strong turgor
of their corpora spongiosa. Besides there ramify through
the snouts highly branched air-passages, the filling of
which is
regulated
by the ampullae
choanales (differentiations of the soft palate at the margins
of the turbinallas that
extend far ventro-caudad), so that the turgor of the snout
is ensured by two systems; the hydraulic system of the
corpora spongiosa,
that is responsible
primarily for the lasting rigidity required in walking,
and the pneumatic system,
that endows the gait and the movements with elasticity
and that reduces the danger of damage when hard objects
are encountered
unexpectedly.
In addition
to the part played by the ampullae choanales a contribution
is
made by the ampullae pneumonasales, that are formed from
the powerfully
developed
nasal
sinuses. There are three of them on each side and they
distribute the air delivered under pressure by the ampullae
choanales.
The canales
ramosi of the sinuses
themselves have yet another orificium externum below the
tip of the snout; mostly it is kept closed, but it can
be opened
reflexly
very
rapidly
if
strong mechanical stimuli impinge on the snout, so that
the latter can collapse
instantaneously. The. system mentioned is innervated from
N. trigeminus, while N. facialis supplies
primarily the perinasal circular and longitudinal musculature.
Like all polyrrhines, Nasobema lacks the os
nasale altogether;
it is not
even
laid down in the
embryo.
The
Great MorgensternÆs Nasobame, the honatata of the natives (Nasobema lyricum;
Plate X), is the best-known representative of the polyrrhines
and on this
account will be discussed in somewhat more detail. As a
typical polyrrhine it has on
its short, fat head four equal snouts, that are fairly
long and on which ¢ as already
described by Morgenstern ¢ it walks. It is enabled to do
this, despite an inadequate nasal skeleton, because the
snouts are
rendered quite
rigid by
the strong turgor
of their corpora spongiosa. Besides there ramify through
the snouts highly branched air-passages, the filling of
which is
regulated
by the ampullae
choanales (differentiations of the soft palate at the margins
of the turbinallas that
extend far ventro-caudad), so that the turgor of the snout
is ensured by two systems; the hydraulic system of the
corpora spongiosa,
that is responsible
primarily for the lasting rigidity required in walking,
and the pneumatic system,
that endows the gait and the movements with elasticity
and that reduces the danger of damage when hard objects
are encountered
unexpectedly.
In addition
to the part played by the ampullae choanales a contribution
is
made by the ampullae pneumonasales, that are formed from
the powerfully
developed
nasal
sinuses. There are three of them on each side and they
distribute the air delivered under pressure by the ampullae
choanales.
The canales
ramosi of the sinuses
themselves have yet another orificium externum below the
tip of the snout; mostly it is kept closed, but it can
be opened
reflexly
very
rapidly
if
strong mechanical stimuli impinge on the snout, so that
the latter can collapse
instantaneously. The. system mentioned is innervated from
N. trigeminus, while N. facialis supplies
primarily the perinasal circular and longitudinal musculature.
Like all polyrrhines, Nasobema lacks the os
nasale altogether;
it is not
even
laid down in the
embryo.
The paired appendages are relatively well-developed. Very
young animals display but little reduction of them. With
older specimens,
i.e.
with those that
have attained about 2/3 of the maximal body length, the
posterior limbs are practically
immovable and are also without function. The forelimbs
are prehensile organs that are supplemented effectively
by the
long, lasso-like
tail.

Nasobema lyricum from Tokiwa TakeshiÆs site
The tail is specialized to an extreme degree, and its organization
is to be understood only in relation to the habits of
the Nasobemes. It
serves
these
fructivorous animals for fishing down their food from
great heights. The way in which this is accomplished is that
the tail, only
the proximal portion
of
which is penetrated ay the vertebral column, incorporates
a gas-filled channel that is connected with the coecum;
so that,
after the
sphincter gasotubalis has been relaxed, the
canal can be filled suddenly with belly-gases, whereupon
the tail is abruptly blown taut
and hurled upward
to a height of more than
four yards. Because of a strongly musculated ampulla (cross-striated
fibers, derived from the ilio-caudal musculature) at
the base of the tail, this
occurs with such violence that in the twinkling of an
eye
the tail is projected with a slight pop to its full length.
No
sooner has
the tip
of the tail,
richly
supplied with tactile papillae, seized the fruit than
the gas is released with a gentle whistle from the tail, that
again
becomes a flattish
ribbon and contracts.
The fruit that has been pulled down is then seized by
the forepaws and brought
to the mouth. It is interesting that the production of
gas in general
is rather well adapted to this mechanism: the hungrier
an animal is the more
strongly
inflated is its colon and hence the ampulla gasomotorica.
Related is the fact that very hungry animals, even when
no reward is
in sight, will carry
out a
ōdry runö of the fruit-catching performance or will hurl
the tail aloft
toward all sorts of objects that resemble fruits. This
is especially noticeable with Nasobema aeolus40 (The
Bluenosed Nasobame).
Once a year Nasobema bears a single young that at first
is carried about in the throat-pouch, which opens caudad,
and
that nourishes
itself from
the motherÆs
axillary teats. The throat-pouch occurs only in females
and is supported by cartilages derived from those of
the AdamÆs
apple.
The animals
are monogamous and the two sexes are very tender toward
one another. The
post-partum female
is fed by the male. Only on the largest islands of the
archipelago do the Nasobames
have enemies, in the predacious Nasobames (Tyrannonasus
imperator B. d. B. = Nasobema tyrannonasus STU.)
that occur there. It
is remarkable that
the Hooakha-Hutchi,
at their equinoctial ceremonies that took place each
fall and spring, ate Nasobames roasted in leaves at these
ritual
feasts.
They regarded
the animal
as holy,
and did not hunt it except in connection with these religious
rites.
The genus Stella was erected by Bromeante. Stulten still
assigns Stella matutina41 B. d. B.
to Nasobema,
as Nasobema morgensternii.
The differences
between
the two genera are indeed small, and have to do with
the tail-hurling mechanism, that is considerably less
highly
differentiated
in the Little MorgensternÆs-Nasobame,
a situation that no doubt is related to the fact that
Stella matutina lives almost exclusively on berries that
grow
near the ground.
In contrast with the peaceable Nasobames, the only extant
representative of the tyrannonasids is a predacious fellow
that feeds almost
exclusively upon
them. In its organization, HebererÆs Predatory Nasobame,
Tyrannonasus imperator B. d. B. (= Nasobema
tyrannonasus Stulten) indeed
resembles the Nasobames
in all major respects, but differs in the development
of the tail, that ¢ as in
the rhinocolumnids ¢ bears a poison claw at the tip;
and besides of course because of its predatory teeth,
the sharp
points
of which are
suited
for ripping the flesh from the victim. Additionally remarkable
is the fact
that the posterior
limbs are astonishingly well-developed for a nasestrian
species, as is related to their employment in seizing
the prey. Finally
it may
be mentioned
that
there is no grain to the coat of this species, whose
pelt gives rather a plushy impression,
somewhat like that of a mole.
Tyrannonasus imperator is especially noteworthy for two
reasons: like all polyrrhine species the animal is not
particularly
swift on nose,
and yet
it travels at
a more rapid pace than the nasobemids. But now, since
all polyrrhine species, because of their intranasal pneumatic
apparatus, when
walking give out
a whistling hiss that can be heard from afar, Tyrannonasus is unable to creep
silently
upon his victims; but ¢ since they flee while he is still
at a distance ¢ must first lie quietly in wait and then
stride after. In this process
of
flight
and pursuit that, because of the considerable expenditure
of
noise
and the yet, so moderate velocity, at first strikes the
onlooker as comical,
Tyrannonasus often must trail the intended victim for hours in order
to catch up with him, since Nasobema uses the tail in
flight
also, by
hurling it
aloft,
encircling branches with it, and thus swinging across
ditches or small streams. Even
when
the predator has come very near the object of his pursuit,
Nasobema often also employs the tail successfully as
a last
resort hanging
by the tail
from a branch,
it swings back and forth in circles or with broad pendular
movements close above the ground until the predator,
in his constant efforts
to grab the
prey, finally gets dizzy and throws up.
Then, during this interval of the predatorÆs disorientation,
Nasobema frequently is able to make off.
But once Tyrannonasus has actually taken hold of his
victim, the latter has no further hope of escape: by
means of the
toxic claw
he is poisoned
and
soon collapses in tears, while the predator gives him
the coup de grace, hauls him
to a shady spot, and there at leisure devours him down
to the larger bones. Whereas a most striking characteristic
of Tyrannonasus thus
is his tough
endurance in pursuit, a second peculiarity is the additional
ability to fast for an extraordinarily
long time for a mammal. This capacity is related both
to an astonishingly low basal metabolic rate and to the
ability
to
store glycogen
not only in the liver
but in sub epidermal depots. Histologically the latter
are
composed of derivatives of the same embryonic cells as
those that give
rise to the
fatty tissue that
occupies this location in other organisms. Storage of
energy in the form of glycogen rather than fat seems
to be more
economical, at
least in
Tyrannonasus. As this process progresses after feeding,
the engorged animal becomes quite
misshapen in a couple of days; immediately after its
ample repast it lies down
in a spot protected from the rain and dozes there until
the sub epidermal store of glycogen is depleted, which
takes
several weeks.
Only after
the animal has
grown slender again, but still retains in the liver sufficient
reserves for possible predatory pursuits, does it again
become active go out
on the hunt.
The noteworthy fact that the captured Nasobame weeps
has psychological interest, for it presupposes that the
animal
possesses insight
and the power of reflection.
In view of the considerable volume and degree of differentiation
of the brain, such a possibility is not excluded (cf.
in connection H.
W. Gruhle
1947).
|
|
|
|
|
|
Tyrannonasus imperator from Tokiwa TakeshiÆs site
In their nutritional physiology, the tetrarrhines (Four-Snouters)
fall entirely outside the framework of the order and
in this respect surely
are secondarily
divergent forms. According to Bouffon (1953) matters
here are as follows: the Rhinogradentia are primarily
insectivorous
in habit,
and the generally
small
size of the animals is correlated with this. In instances
where
there has been a specialization of the habits and way
of nourishment that
distinguishes the
animals from the typical insectivorous type, they nevertheless
are to be
derived from it; all the crab-eating hopsorrhines (Snout
Leapers) and their milk-symbiotic
forms are close to this insectivorous type, and likewise
the hypogeonasids (Mud Snouters) and the georrhinids
(Burrowing Snouters) can be
derived therefrom. With the tetrarrhines, those more
primitive members are
out-and-out fructivores,
such a derivation seems more difficult, although for
example the dentition fundamentally is wholly of the
insectivorous
kind. Above
everything
else, the digestive tract displays extensive specialization,
not to mention
the gas-producing
coecum. But the main factor that distinguishes the
tetrarrhines from most other Rhinogradentia is the considerable body
size. The Nasobames
get to
be nearly
3 feet high! The derivation of the Predacious Nasobame,
Tyrannonasus, at first appears easier, inasmuch as
seemingly a simple increase
in all dimensions
would
convert an insectivorous into a predacious type. But
Bouffon, by means of penetrating studies, has shown
that Tyrannonasus has descended
from
the fructivorois
species.
This is revealed primarily in the organization of the
digestive tract and of the tail, which during puberty
is still nasobematous.
Bouffon
believes
that
the modification into a predator ¢ and in fact into
a monophagous predator ¢ came to pass via a predatory commensalism.
Certain
peculiarities in the behavior of the predator give
further indications in this
direction:
namely,
it greedily
devours fruits thrown aside by the fleeing Nasobames,
and only
attacks the latter when it can fall upon than while
they are eating fruit.
In
agreement with this is the fact that young Tyrannonasus specimens are not predacious
but merely pounce upon the feeding Nasobame in order
to snatch their fruits away, or to feed upon the remnants
of their
meals.
This phenomenon, as Bouffon too points out, is by no
means unique in the world of animals; the transition
from the
insectivorous to the
fructivorous habit
has frequently been observed, thus for example among
the thrush-like songbirds,
in the insectivores themselves, is well as in the South
American Chiroptera, prosimiais and catarrhine monkeys.
40 Aeolus Grk. = the wind god.
41 stella matutina Lat = Morgenstern (= morning star).
TRIBE: Hexarrhinida (Six-Snouters sens. lat.),
FAMILY: Isorrhinidae (Equal-snouted Snouters),
GENUS: Eledonopsis42 (Ribbon Snouters),
5 Species,
GENUS: Hexanthus (Six-flowered Snouters),
3 Species,
GENUS: Cephalanthus43 (Flower-faced Snouters),
7 Species,
FAMILY: Anisorrhinidae* (Unequal-snouted Snouters),
GENUS: Mammontops44 (Shaggy-faced Snouters),
1 Species
The tribe Hexarrhinida (Six-Snouters sens. alt.) comprises
two very different families; whereas the Equal-snouted
Snouters (Isorrhinidae)
are small
insectivorous animals of relatively primitive
organization, the sole
species in the family
of Unequal-snouted Snouters (Anisorrhinidae)
is of a type that at first glance seems much more reminiscent
of the
Nasobemidae but that
also
displays a whole
series of features that separate it from this
family. Bouffon feels therefore that the tribe Hexarrhinida
erected
by
Bromeante de Burlas
cannot be
upheld or that it is polyphyletic. This will
be considered later when Mammontops
ursulus45 is discussed.
The Equal-snouted Snouters, as already said,
are animals that ¢ apart from their polyrrhiny
¢ are
to be regarded
as primitive;
the paired
appendages are scarcely reduced and still are
well adapted for progression, even
though
the
animals make but little use of then. The differentiation
of the snout likewise is still primtive.* On
the other hand, the
progressive
genera
Hexanthus and Cephalanthus Br. d. B. (= Ranunculonasus and Corbulonasus STU.)
are distinguished
by highly developed mimesis, that has modified
the exterior of tht animals in a most peculiar
fashion.
[TranslatorÆs
note; Br.
d. B.
is just B.
d. B. elsewhere.]
As representative of the more primitive genus
Eledonopsis, Eledonopsis terebellum (The Tubeworm-snouted
Ribbon
Snouter) will be described.
In little holes in the ground, beneath stones
and roots, on Mairuvili one frequently finds
a small
animal the
size of a
shrew, that
sleeps there
rolled up during
the daytime and on first glance looks just
like a little shrew, with grayish-brown fur
and rosy
paws. The little
creature does
not try
to run away and lets
itself be put back into its den. If one marks
the position of such a hole and at night
makes a flash-photograph of its entrance and
the surroundings, then one sees on the picture
that
four to six ribbon-like
structures are
stretched
forth
from the hole. These rose-colored ribbons are
some 2 ¢ 3 mm wide and as much as a foot long.
On the
upper surface
they
bear two
narrow, damply glistening
grooves to which are stuck, one sees, some
tiny insects, mostly podurids (springtails)
and barklice
(Corrodentia).
If one tries
to view these
structures
more closely
with a flashlight they are then pulled quickly
back into
the hole. For a long time the phenomenon failed
to yield to more
precise
observation. In
fixed specimens
the ribbons were indeed to be identified without
further ado as snouts, but their function became
explicable
only when the
animals
were kept
permanently in the light (SchallerÆs method).
Now it became apparent that the ribbons
actually are the snouts, and the two grooves
the drawn-out nares that
have been swiveled
upward; furthermore it was found that the ciliated
epithelium of the nasal cavities was continued
throughout the nares
and that,
together with the
nasal
mucus, served to transport the small insects
that stuck there;
these were swept into the nasal passages and
were carried choanally to
the
digestive
tract.
Besides, it developed that Eledonopsis also
is able to entrap larger insects ¢ up to the
size
of woodlice
¢
and move them
proximad in
this fashion,
while the ribbon-like snout contracts and forms
a gutter within which the prey
is brought to the base of the snout, partly
by means of the current set
up by
the cilia and partly by means of peristaltic
movements of the trough. There the prey either
is seized
with the tongue
or
dug with the
hands out of
the nasal parts directed toward the head, and
devoured. Interesting too is it
that large, unwieldy animals ¢ primarily spiders
of the families Heieiatypidae and
Lycosodromidae ¢ are first entrapped in the
nasal slime and then entwined by several snouts
and
drawn toward
the head.
As to when
the various
prey-catching mechanisms are set in motion,
the decision is made in part by the very
acute tactile sense and then too by the chemical
sense, that extends clear to the tips of the
snouts. (Here it is a matter of receptors and
nerves that
bear the same
relationship to
the actual
olfactory
organ as JacobsonÆs organ in many other vertebrates.)
In Eledonopsis care of the brood, differs in
no wise from that of other Placentalia. A marsupial
pouch is
not present.
The
young become
independent
in a very
early age. Mating seems to take place at night.
Up till now attempts to rear Eledonopsis in captivity have been unsuccessful.
The behavior of young specimens of the Six-flowered
Snouters (Hexanthus) is very similar
to that of Eledonopsis. They
too live in holes
in the ground or
under leaves and from there stretch out their
snouts in order to catch prey. But this is
true only of
the very
young animals
that
have just
begun to get
their own food after having been released from
their motherÆs nursery. Later, the following
differences from Eledonopsis
come about; the
nasal groove grows
together, in a proximal to distal direction,
in such
a way that ultimately openings remain only
at the base and
the
tip of the
snout while the
rest of the snout forms a covered tube. Then
four broad, pointed dermal lobes
grow
out at the tip of each snout. According to
the species, these are variously colored and
are
capable besides
of fairly strong
color
changes within
the limits set by the specific basic coloration46.
Eventually each snout comes to look like a
long-stalked flower. During this modification
of their snouts
the animals scarcely
change their
habits at
all. They continue to stretch their snouts
forth from their hiding place, but curl
them higher and higher about the stems of plants47,
and get their nourishment as described above.
At all events
there
now takes
place a constantly
greater change in the menu. For now they catch
primarily flying insects that, deceived
by the flower-like shape and color, settle
at the tips of the snouts. Transportation of
small
victims
continues
in
the manner
already
described. As also explained
above, somewhat larger specimens are borne
cephalad by peristaltic movements in the nasal
tubes.
Yet larger objects, that could
not pass through,
are ¢ in contrast with Eledonopsis ¢ not taken
in entire. Instead, the very
expansible, corolla-like flaps of the snout
are folded about the insect so that it is
practically
wholly enwrapped; thereupon the Hexanthus vomits
through the appropriate snout and digests the
specimen to the
point where
it can be sucked
back through the
nasal tube.
Mature Hexanthus do not continue to lurk in
holes; rather, they lie on rocky slopes amid
the greenery
of grassy
patches and small
herbs.
Their
own greenish
coloration makes them inconspicuous there,
and their snouts are for the most part twined
about
the stalks
of those
flowers whose
color
and shape
they
can simulate. Moreover, this adaptation contains
a visual element; if Hexanthus is presented
with blue
flowers
pasted to a sheet
of cardboard behind which
the snouts are lying among yellow flowers,
then the rhinal corolla-like lobs turn blue
¢ and
vice versa.
Furthermore,
the various Hexanthus species
have
differing photoperiodic behavior whereas the
Globeflower Snouter (Hexanthus ranunculonasus =
Ranunculonasus
pulcher48) is thoroughly
diurnal, the
predominantly violet snouts of the Gorgeous
Night-Snouter (Hexanthus regma-noctis49)
blossom mainly at night. In contradistinction
to the
Ranunculonasus Snouters, that have merely a
slightly acidic odor, like that of sour milk,
these Night Snouters emit a strong fragrance
of vanilla, that attracts nocturnal six-winged
insects.
To the genus of the Flower-faced Snouters (Cephalanthus = Corbulonasus) belongs
a whole series of the most beautiful Rhinogradentia that are known. All of
them are distinguished by the fact that the short, broad, petal-like snouts
are set close about the mouth and are supplied with a very simple epi- and
hyponssal musculature, that permits the animals to clap together the tonically
outspread snouts very rapidly whenever an insect settles in the oral field.
A further peculiarity is that these mentally very limited animals emit strong
oral fragrances, that apparently serve likewise for the attraction of insects.
An additional oddity is that ¢ in contrast with other polyirhines ¢ representatives
of this genus neither care for the brood in any way nor suckle them.
As a typical representative of the genus we choose the Miraculous Flower-faced
Snouter (Cephalanthus thaumasios50 = Corbulonasus longicauda51),
that lives colonially on Mitadina in the meadows of Ranunculaceae of the more
elevated
mountainous regions. Skamtkvist describes the sight of such colonies as the
most beautiful thing he saw on Hi-yi-yi. The vividness of the colors and the
luster of the snouts is said to be quite extraordinary, and the characteristic
spectacle of the animalsÆ swaying on their tails in the fresh sea breezes
enchanting.
Evidently what pleases us most in these curious creatures is for the flower-seeking
insects nothing more than the presentation of hypereffective stimuli; and in
this sense too is to be interpreted the buttermilk-like odor
that emanates from the open mouths of the lurking cephalanthids.
In addition to the nasarium, which has already been discussed, one of their striking features is the stiffened tail, that may be as much as twenty inches long. An interesting thing is how the caudal structure changes as the animal grows; the newly born animals, that already possess a fully developed nasarium, fall to the ground and climb up on the stems of nearby flowers. Having reached the top, they bite off all the buds, unfold the nasarium, and begin to capture prey just as older specimens do. Their still soft tail is then just about the length of the body and differs in no respect from a normal mammalian tail. But it promptly grows in length, and in fact by means of elongation of the vertebrae; as the intervertebral articulations ankylose and the intervertebral sinews and ligaments also stiffen, the caudal musculature degenerates, so that of M. ischiocaudalis, M. iliocaudails and M. depressor caudae there remain only ligamentous bundles that are attached to the caudal vertebral column and to its rigidified uniting structures. The tip of the tail bears a much cornified epithelium, that eventually forms a sort of pointed, plugshaped horny cap. As soon as the tip of the tail reaches the ground, the animal that had climbed up the plant stem starts to bore this caudal plug into the ground, which within four to six days it penetrates to a depth of as much as 6 inches. Then the animal lets go of the plant stem and henceforth is supported by its own tail, that continues to lengthen. The degree of caudal extension attained depends on the nutritional state of the animal, and proceeds more slowly when the latter is well fed. An animal once firmly ōplantedö in this way can no longer move from its location, but simply waits there for prey with folded arms and open mouth. As mentioned, the mental capacities are slight. Mating takes place when it is windy; as the specimens are blown back and forth on their tails and come into contact the desirous males take firm hold of the females. The period of gestation is said to be but three weeks, and the total duration of life is estimated maximally at eight months. Growth from birth to maturity requires perhaps two months; from birth to caudal implantation eighteen to twenty-two days.

Cephalanthus from Tokiwa TakeshiÆs site
Not infrequently one finds colonies that make a pitiful impression;
the snouts look limp and are discolored and incrusted. The creatures are losing
weight,
and their soft whimpering is audible from afar. Such colonies have been attacked
by a nasal scab, that is caused by a species of mite that is closely related
to the gamasids. A slight infestation scarcely causes noticeable harm. But
when mass increase of the mites renders the snouts ineffective for taking
prey, this is naturally a catastrophe for Cephalanthus. The starving, tortured
creatures
that dig constantly at their diseased snouts and thereby merely increase
their suffering. At the end there are only the little corpses hanging on their
long
caudal stalks; and in many places one finds standing in the meadows groups
of sixty to a hundred skeletonized tails, beneath which there lie only the
decaying remains of bones and hides. However, the primary cause of the epizootics
is not the endemic mites, but rather viral diseases favored by variations
in the weather; they reduce the natural resistance to the mites, in that individuals
suffering from viral disease are not able to give regular care to their snouts
aid to keep them properly lubricated.
The majority of Cephalanthus species live in the manner described. Only Cephalanthus
ineps52 (= Corbulonasus ineps) and Cephalanthus piger53 (=
Corbulonasus acaulis54) have reduced tails and simply lie
on their backs in sunny spots between
stones and flowers. According to Bromeante de Burlas, their separation from
other cephalanths as a new genus, already weighed above does not seem justified.
As has been mentioned on p. 66, the anisorrhine Mammontops
ursulus (The Bearlike Shaggyfaced Snouter), that likewise occurs in the mountain
meadows
of Mitadina,
falls quite outside the hexarrhine sens. It is a relatively majestic animal,
that attains an overall height of 4 feet 3 inches in the male and 3 feet
7 inches in the female; and is herbivorous.
Its snouts are differentiated somewhat in the tetrarrhine manner, and this
is what causes uncertainty as to the proper systematic arrangement; whereis
Stulten favors placing Mammontops in the immediate vicinity of the tetrarrhines,
in that he lays more weight on the nasal structure than on the number of
snouts, Bromeante de Burlas holds the opinion that the number of snouts
deserves the
greater systematic consideration, while the degree of nasal differentiation
is to be regarded as due merely to convergence. In support of this view
he draws upon the investigations of the group of French scientists (Bouffon,
Irri-Egingam, and Chaiblin), who have shown that the innervation of the
several
groups of
muscles in the tetrarrhines is quite different from that of Mammontops.
Here it seems to be a matter of a further differentiation of the epi- and
hyporrhinal
muscle sips of the isorrhines; and the absence of corpora spongiosa from
the snouts of recent isorrhines no longer appears so primitive as was once
assumed. Bouffon and Gaukari-Sudur postulate for the hexarrhines common
tetrarrhine-like ancestors from which the isorrhines of today have developed
on the one hand
and the anisorrhines on the other. In this connection it is nevertheless
a curious fact that the isorrhines display emphatically primitive features
in
respect to their paired appendages, while in the anisorrhinous line these
are especially reduced. Moreover it is noteworthy that the hair whorls
that are
characteristic of the polyrrhines are lacking in the isorrhines. Against
the placement of too much stress on this feature, Bromeante de Burlas brings
up
the point that even the surely secondarily evolved Orchidiopsis does not
exhibit an inversion in the course of the hairs although such an inversion
is most
conspicuous in the much more primitive hopsorrhines. In any case the problem
of the position of Mammontops will have to be left open for the present,
until more extensive detailed investigations have been made.
Mammontops occurs in small herds that are led by older males.
The animals feed almost exclusively on a composite, Mammontopsisitos dauciradix55,
that they
tear up by the roots with their two grasping snouts. The dentition is the
most highly specialized known among the Rhinogradentia (aside from the
total loss
of teeth in the mercatorrhines); the incisors are reduced, the canines
small and blunt, the premolars and molars broad and slab-like.
The Shaggyfaced Snouter suckles its offspring, that clings with its snouts
to the thick fur of the mother and also to her stiff, inguinal teats. The
reproductory rate is low. The animals seem to grow quite old. Older males
are distinguished
from the uniformly chocolate-brown younger males and the females by a silvery
gray tail, the waving of which releases a following response on the part
of the herd. For example, Tassino di Campotassi was able, by whitening
the tail
of a young female, to provoke a following reaction from the herd into which
he put her. The whitened tail was particularly effective as a supernormal
releaser of the following response in the younger males.
*but cf. page 76.
42 eledone Grk. = an animal, related to the octopus.
43 kephale Grk. = a head; anthos Grk. = a flower.
* TranslatorÆs note re Anisorrhinidae; under most circumstances this should be Mammontopsidae, since Mammontops is the sole genus; however there are situations in which the rules of nomenclaure would allow the family name Anisorrhinidae.
44 cf. Footnote 15.
45 ursulus Lat. = a little bear.
46 Freddurista and Perischerzi have shown that reds are regulated by expansion of the capillaries, yellows by the superficial fatty tissue (that lies, however, below the subepithelial capillary net), and blues by black pigment situated in contractile melanophores.
47 constantly to the left, both the snouts of the right side of the body as well as those of the left (cf. Ludwig 1932).
48 ranunculus Lat. = a crowfoot (flower); pulcher Lat. = beautiful.
49 regina noctis Lat. = queen of the night
50 thaumasios Grk. = marvelous, rare.
51 longi-cauda Lat. = longtailed.
52 ineps Lat. = mentally sluggish.
53 piger Lat. = lazy
54 akaulos Grk. = stalkless.
55 sitos Grk. = food; dauci-radix Lat. = a carrot-like root.
PHALANX: Dolichoproata (Long-nosed Snouters),
FAMILY: Rhinochilopidae56 (Tasselsnouter-like
Snouters),
GENUS: Rhinochilopus (Tasselsnouters*),
2 Species.
The genus Rhinochilopus with its two species, Rh. ingens57 (The Giant Tasselsnouter) and Rh. musicus (The Pipeorgan Tasselsnouter), possesses the most impressive polyrrhiny: in both these animals the head has been protracted into a long proa or rostrum. The underside of this structure is supported by the maxillary, the premaxillary and the palatine, the upperside by the maxillary, premaxillary and nasulary, as well as by a part of the nasal bone. The lower side (cf. Fig. 12) exhibits a lengthening of the oral cleft, the so-called proal groove (2), that is enclosed by the lips. At the anterior end of the proa the male bears two asymmetric incisors. To the right and left of this proal groove are the nineteen pairs of snouts, here termed nasidi (3, or 9). The first pair serves as tentacles, the rest as organs of locomotion. (As to the more detailed structure of the nasarium, see below.) The paired appendages are much reduced. The hindlegs function solely as antennae for use during rearward movement. The forelegs do not touch the ground and also play no part in the uptake of food. In the female they serve to clasp the invariably single offspring. The tail too is no more than an organ of touch. The animals attain a respectable size (in Rh. musicus nearly 5 ft. from proa to the base of the tail, in Rh. ingens more than 7 ft.). They are omnivorous but prefer insects, snails, and fungi, and also berry-like fruits. Occasionally they also consume young leaves. Rhinochilopus is solitary, wandering in leisurely fashion through the virgin forest and especially through the less dense portions and the margins of the woods, where it has definite routes and stamping spots. However, the animals have no fixed beats, and make un-contested use of the paths of conspecific individuals.
[Fig. 12] Rhinochilopus musicus; advanced embryo. 1. Incisor rudiments (only in the male); 2. Subproal groove; 3. Ductulus musicus of one excised nasulus; 4. Ductulus osmaticus, same; 5 Nasular corpus spongiosum; 6. Ductus musicus; 7. Tear duct; 8. Vesica inflatrix organi; 9. Nasuli. The more medially situated ductus osmaticus is not numbered. Note the distinct structure of the first nasulus, apparent even in the embryo. (After Bouffon and Gaukari-Sudur 1952) |
The most noteworthy peculiarities of the two species, especially of Rh. musicus, are however the wooing procedure and the correlated nasarial specialization, that will be described briefly: like most Rhinogradertia, Rhinochilopus no longer respires exclusively through the nares, but predominantly through the tear duct, that likewise is expanded significantly, as it is in almost all species (cf. also Fig. 4). Now, on the one hand the ear duct (Fig. 12, 7) is connected immediately with the throat, from which a passage ¢ the ductus osmaticus ¢ runs into the proa and there gives off the ductuli osmatici (4) into the nasuli. On the other hand, the tear duct also is connected via the ductus inflatorius with the vesica inflatrix organi (8), which again for its part communicates by way of the ductus vesico-gularis with the throat. A second connection (6) between tear duct and vesica inflatrix organi and the nasuli the ductus musicus, supplies the ductuli musici, that are located in the nasuli laterad of the ductuli osmatici. When at rest the vesica inflatrix organi lies beneath the skin of the cheek, and on activation of the nasal pipeorgan is inflated to the size of a childÆs head. The apparatus as a whole constitutes a differentiation of the tear duct and the choanal portion of the nose, as may be determined in young embryos. As additional components the pipeorgan includes the corpora spongiosa (5) of the nasuli, as well as the nasilar circular and longitudinal musculature.

Rhinochilopus from Tokiwa TakeshiÆs site
In its entirety the apparatus operates as follows; during progression the ductus musici and ductuli musici are closed and together with the corpora spongiosa afford the nasuli the turgor required if the circular and longitudinal musculature is to suffice to move then. During inspiration, air flows in through the ductuli osmatici. Thus the air is constantly being tested. The first pair of nasuli, that function as tentacles, are especially concerned with olfactory examination. Directional olfactory testing is rendered possible by the division, in the proa, of the olfactory nerve in such a manner that each pair of nasuli comes to have a distinct olfactory epithelium. Food is taken in through the nasuli by being grasped by a digit-shaped lengthening of the nasular tip or by being encircled by the distal third of the entire nasulus and then brought into the proal groove. The prey is then impelled toward the mouth by the strongly protracted lips. The musical apparatus of the nasarium is made use of only during courtship. Then the males lie flat on the ground. The ductuli osmatici cease to function, as do the corpora spongiosa. Initially the nasalar musculature relaxes completely. Only the sphincteres terminates of the ductuli musici are slightly contracted. The sphincteres glossiformes, situated at the base of each ductulus musicus, are relaxed. Now, by means of violent, persistent breathing, the animal inflates the vesicae inflatores organi and thereby also puts the ductus musici under pressure. By opening slightly the sphincteres glossiformes the nasuli now are inflated, and though the circular musculature is fully relaxed the longitudinal musculature can make the nasulus longer or shorter. If now a sphincter glossiformis is suddenly opened more widely, brief puffs of air pass into the corresponding nasulus and cause it to reverberate, inasmuch as the air column it contains is set into vibration by the lips of the sphincter. Since each nasulus can be lengthened or shortened in rapid alternation, it works like a wind instrument on the principle of a slide trombone, with the peculiarity that long-lasting tones cannot be produced, but only successions of short ones. With eighteen pairs of nasuli that function in this wise (the first pair lacks a sphincter glossiformis58), the animal has command, so-to-speak, of 36 wind instruments that can be activated independently. How they are employed in courtship has been described graphically by Skamtkvist:
ōAt that time-it was the vernal equinox-the Hooakha-Hutchi were celebrating the honatata feast, at which the smoked honatatas were consumed to the accompaniment of ritual chants in the village long house. It was in the evening, at dusk. The ceremonial meal lasted not more than two hours. Then the village company arose and betook themselves to a nearby forest glade, at the western edge of which they all sat down. The full moon was already visible above the treetops on the mountain opposite when the Mostada Datsawima (the æLords of the MyriapodsÆ) made their appearance from the dark of the woods and stepped out into the glade. Silently, as if they were floating, the great beasts moved. In the uncertain moonlight their legs (snouts) were not to be clearly made out. Only the luster of the long heads and backs could be discerned. There were fourteen or sixteen animals, that first paraded in a circle a couple of times before the six especially huge males lay down and extended all their snouts, while the females continued to circle about them. And now there began the most peculiar concert that I had ever heard: it began with the hollow, rhythmic lowing of one of the animals. Slow at first and then growing faster. Soon a second one joined in, lowing a few notes higher; and finally all six were taking part. The rhythm varied, but was maintained in rigid synchrony by all the animals, while the number of participating voices grew and grew. All at once there was a silence, and then a sharp, bleating tremolo, many-voiced and prestissimo, was superimposed on this rhythmic, half-rumbling, lowing or thrumming, hollow outcry. This constituted the second movement. In time, this æsoloÆ by the individual male concerned in its production was heightened by the weaving into it, in addition to the bleating and æstaccatoÆ passages, of slurred and smudged tonal transitions. The beast that had the solo part could be distinguished by the fact that its snouts, that now were readily discerned inasmuch as they were extended to one side, were alternately inflated and made shorter and longer. Suddenly there was silence once again; and then the hollow basic pattern of the whole musical chorus started up once more, whereupon a second male gave solo voice. During this æconcertoÆ the females circled in slow, regular tempo about the performing males, until the last of these had finished his solo contribution. Then the males arose, and the whole ghostly company vanished slowly, as they had come, into the dark forest. The inhabitants of the village stood up, and bowed deeply toward the place into which the Mostada Datsawima had disappeared, and deeply once again toward the full moon. Then we walked back into the village, where until the hour was late the pipes and drums resounded, a pale echo of the music heard earlier ...ö
Sad to relate, a thorough study of these animals was no longer possible, since they too-just like the Hooakha-Hutchi-soon fell victims to the head cold introduced by Skamtkvist. Skamtkvist did succeed, however: in capturing and taming one of the males. The animal seemed very intelligent, as is understandable in view of the weight of its brain, which was determined later. It grew very tame, and Skamtkvist was even able to teach it two of BachÆs organ fugues that he knew by heart, so that it could perform them perfectly. Only its inability to produce long-held notes caused any difficulty. Here the animal made shift with very rapid tremoli, by using four nasuli adjusted to the same pitch.
56 chilopus Grk. = a myriapod.
*TranslatorÆs note: ōTatzelö signifies ōpawö rather than ōtasselö, but the English term ōTasselsnoutersö seems more suitable for these animals.
57 ingens Lat. = enormous.
58 The sphincter glosiformis is not a simple circular muscle. The entire dosing mechanism summarized in the term is composed of the actual sphincter, that extends around three-quarters of the arcumference of the nasulus, and a stout cushion of connective tissue, that occupies the remaining quarter. On this cushion of connective tissue there sits a V-shaped pair of outgrowths that act as vocal cords. The vocal apparatus might logically be called a ōnarynxö, since it constitutes a wholly comparable analog of the larynx and syrinx.
HARALD STUMPKEÆS manuscript was awaiting publication when
it became known that, during secret tests of atomic explosions (about which
even the Press had
heard nothing), the entire Hi-yi-yi Archipelago had been annihilated through
the oversight of some subordinate. As a result of tectonic tensions that
had not been anticipated, the whole group of islands sank beneath the sea
when the burst was set off some 125 miles distant.
At the time in question, an international commission for study of the archipelago
was in session on Mairuvili. Among those present were most of the scientists
named in this work. With them there went down the Darwin Institute of Hi-yi-yi,
situated on the lovely eastern bay of the islands, and in it the irreplaceable
photographic material, the various preparations and observational and experimental
records that were to have formal the nucleus of a great, comprehensive treatise
concerning the archipelago and its geological, historical, zoological, and
sociological peculiarities.
Thus it was most fortunate that, shortly before his voyage, Stumpke had undertaken
to compose a short exposition of the structure and habits of the Rhinogradentia.
Toward preparation of drawings he had supplied me also with some material
that he ¢ one can only say, Alas! ¢ took back to Hi-yi-yi with him. Still
it was
in this way that at least a part of the lifework of this modest and deserving
seeker after Truth could be preserved as a balanced whole for science and
for a broader audience, and with it the knowledge of a now-vanished world.
Heidelberg, October 1957
GEROLF STEINER
ASTEIDES, S. (1954); Le nez dÆOrchidiopsis, son anitomie,
son developpement. C. r. Soc. biol. Rh. 516; 28.
BEILIG, W. (1954); Ein vanadiumhal tiger Eiweisssymilex aus den nasalen Fangfaden
von Emunctator. S. H. Z. physiol. Chem. 884; 55.
BITBRAIN, J. D. (1946); Anatomical and histological sudy of the nose of a Rhinogradent,
Rhinolimacius. J. gen. Amt. Soyi 18.
---- (1950) The Rhinogradents. Univ. Press S. Aidrews.
BLEEDKOOP, FR. (1945); Das Nasobemproblem. Z. v. Lit. 34; 205.
BOKER, H. (1935 and 1937); Einfuhrung in die vergleichende Anatomie der Wirbelriere.
Fischer, Jena.
BOUFFON, L. (1953); A propos du systeme nutritif des Rhinogradents. Bull. Darwin
Inst. Hi. f; Suppl. 2.
---- (1954); A propos du groupe polyphyletique de Rhinocolumnides. Bull. Darwin
Inst. Hi. 8; 12.
BOUFFON, L., and GAUKAM-SUDUR, O. (1952); LÆanatomic comparee des Polyrrhines.
Bull. Darwin Inst. H. 6; 33.
BOUFFON, L., IRM-EGINGARRI, J., and CHAIBUN, F. (1953); A propos de lÆinnervation
du nasoire des Polyrrhines. C. r. Soc. Biol. Rh. JJJ; 24.
BOUFFON, L., and LO-IBILATZE-SUDUR, Ch. (1954); Comment Orchidiopsis attire-t-elle
sa proie? La nature (P) v; 311.
BOUFFON, L., and SCHPRIMARSCH, J. (1950); Corcernant la question de la descendance
du genus endemique Hypsiboas. Bull. Darwin Inst. Hi. 4; 441.
BOUFFON, L., and ZAPARTEGINGARRI, V. (1953); Fur Fembryologie des Orchidiopsides.
Bull. Darwin Inst. H. 7; 16.
BROMEANTE DE BURLAS Y TONTERIAS, J. (1948); A systematica dos Rhinogradentes.
Bull. Darwin Inst. Hi. 2; 4j.
---- (1948); Systematic studies on the new wder of the Rhinogradents. Am. Nat.
F. 374; 1498.
---- (1949); Os Polyrrhines e a derivacao d'elles Boll. Braz. Rhin. i; 77.
---- (1950); A derivacao e a arvore genealogica dos Rhinogradentes. Boll. Braz.
Rhin. 2; 1203.
---- (1951); The Rhinogradents. Bull. Darwin Inst. Hi. SS Suppl.
---- (1952); The Hypogeonasidae. Bull. Darwin Inst. Hi. 6; 120.
---- (1954); The hides of Rhinogradents and their grain. Nature (Danuddlesborough)
92; 2.
BROWN, A. B., and BITBRAIN, J. D. (1948); A simple electronically controlled
substitute for feeding Mercatorrhinus. J. psych, a. neur. contr. 181; 23.
BUCHNER, P. (1953); Endosymbiose der Tiere mit pflanzlichen Mikroorganismen.
Birkhauser, Basel.
COMBINATORE, M. (1943); Un pezzo di legno appurtato, trovato sulla spiaggia
di Owsuddowsa. Lav. preist. (Milmo) 74; 19.
DÆEPP, FR. (1944); La descendance des Polyrrhinei. C. r. Soc. biol. Rh. so6i
403.
DEUTERICH, T. (1944); Em holzemer Suppenloffel von Haidadaifi. Z. f. v. Prahist.
22; 199.
---- (l944a); Grundsatzliches fiber die Esslestecke der Huacha-Hatschi, eines
ausgestorbenen polynesisch-bajuwarischen Mischvolkes. ibid. 2 4;-312.
FREDDHRISTA, P., and PERISCHERZI, N. (1948); LÆamblamento di colore fssiologico
nei mammiferi, specialemente nei generi Hexanthus e Cephalanthus (Polyrrhina,
Rhinogradentia) Arch. di fisiol. comp. ed. irr. 34; 222.
GAUKARI-SUDUR, O., BOUFFON, L., and PAIGNDPOULOS, A. (1950); LÆanatomie comparee
des Sclerorrhine. C. r. Soc. Biol. Rh. 512; 39.
GRUHLE, H. W. (1947); Ursache, Gnind, Motif, Auslosung. Festschr. f. KURT SCHNEIDER,
Heidelberg, Sclerer.
HARROKERRIA, J., and IRRI-EGINGARRI, J. (1949); Note sur la biologie dÆOtopteryx
volitans. C. r. Soc. Biol. Ih. 511; 56.
HYDERITSCH, F. (1948); The slug which was a mammal. Sci.a. med. cinemat. Cie,
Black Goats.
IZECHA, F. (1949); La primitividad de la cola de los Rhinogradentes. Boll.
Arg. Rhin. 2; 66.
JERKER, A. W., and CELIAZZINI, S. (1953): The ancestors of the Hypogeonasidae,
were they Emunctators? Evolution (Littletown) JJ; 284.
JESTER, M. O., and ASSFUGL, S. P. (1949); The genus Dulcicauda and the problem
of ōRassenkreisö. Bull. Darwin Inst. Hi. 3; 2il.
LUDWIG, W. (1932); Das Rechts-Links-Problem in Tierreich und beim Menschen.
Berlin.
---- (1954); Die Selektionstheorie. In: Die Evolution der Organismen. Hrsgeg.
v. G. HEBERER. Fischer, Stittgart.
MAYER-MEIER, R. (1949); Les ōTricladesö de MUELLER-GIRMADINGEN, sont ils des
mammiferes? Bull. bnl. mar. St. V. H. 171 i.
MORGENSTERN, CHR. (1905); Galgenlieder. B. Cassrer-Berlin.
MULLER-GIRMADINGEN, P. (1948); Les triclades do sables du Wisi-Wisi. Acta Helvetica
Nas. Ser. B. IS! 210.
NAQUEDAI, BR. B. (1948); Georrhinida et Hypogeoiasida, deux subtribes parentes.
C. r. Soc. biol. Rh. 5101 64.
PETTERSSON-SKAMTKVIST, E. (1943); The discovery of the Hi-lay-Archipelago.
J. A. geogr. 322; 187.
---- (1946); Aventyrer pa Haiaiai-oema. Nystrims Forlag och Bokhandel, Lilleby.
PUSDIVA, FR. (1953); Uber die Schleimdrusen und de proteolytischen Prozesse
in der Sellarscheibe von Dulcicauda grise-aurella. S. H. Z. physiol. Chemie
822; 1443.
REMANE, A. (1954); Die Geschichte der Tiere. Ii; Die Evolution der Organismen,
hrsgeg. v. G. HEBERER. Fiseher, Stuttgart.
REMSCH, B. (1947); Neuere Probleme der Abstunmungslehre. Stuttgart.
SCHUTLIWITZKIJ, I. I. (1947); Hat Morgenstern die Rhinogradentier gekannt?
(Russisch mit dtsch. Zusanmenfassung.) Lit. prom. N. S. 27; 81.
SHIRIN TAFARUJ (1954); A propos du chimisme du sue attractif des Nasolimacides.
J. physiol. irr. u; 74.
SPASMAN, O., and STULTEN, D. (1947); Rhinogndenternas systemet. Acta Scand.
Rhin. 4; I.
SPUTALAVE, E. (1946); Le sabbie miliolidiche del orizzonte D 16 superiore del
lÆisola Miruveely. G. geogr. frdd. Ital. Jp<? 12.
STULTEN, D. (1949); The descendency of the PolTrhines. Bull. Darwin Inst. Hi.
3; 31.
---- (1950); The anatomy of the nasarium of Hopsorrhinus. Bull. Darwin Inst.
Hi. 4; 511.
---- (1955); The evolution of turbellarians, a leview of new aspects. Piltdown
Univ. press.
STUMPKE, H. (1956); Das Nasarium der Poljrrhinen, erne Zusammenfassung der
bisherigen Ergebnisse, unrer besonderer Berucksichtigung der neueren Untersuchungen
iber die Innervierung. Zool. Jahrb. Abt. XX-XI, 43; 497.
TASSINO DI CAMPOTASSI, I. (1955): Un ōreleaserö sopranormale in Mammontops.
G. psicol. comp. e corn. 2: 714
TRUFAGURA, A. (1948); La cola de los Rhinogradentes. Boll. Arg. Rhin. I; I.
The history about snouters would not be complete without the mention of the article from ōPrirodaö (ōThe Natureö) magazine where the further siscoveries of representatives of Rhinogradentia group are described, made already after with archipelago Hi-Yi-Yi has plunged to the ocean floor. And it would be especially incomplete without the story about the author of these creatures. Therefore I include to the HTML version of the book as the appendix the article ōMysterious snoutersö, having omitted only its first part where the content of the book is simply retold shortly. Sorry for any grammar mistakes in translation; I hope my work is understandable for English readers.
Pavel I. Volkov.
S. D. Fieldojants
Cand. Biol. Sci.
Zoological institute of the Russian Academy of Science
Saint Petersburg
A. Selbstander
The Ruhr university
Bochum (Germany)
Intrusion of exact knowledge into biology has pushed aside its traditional directions, in particular, the descriptive and the classification zoology. Meanwhile, just the representatives of these sections of science are first to meet unknown things ¢ the live beings unknown to the scientist earlier. So has left, that rhinogradentians hidden in their ōlost worldö yet have not involved in itself due attention of genetic scientists, molecular biologists etc., but now, though late a bit, they should become objects of careful studying and application of genetic engineering technologies.
What a grateful area for young talents as the future researches, certainly, will be immediately maintained by the European, American and Japanese scientific funds! The present pioneer work can already serve as a fine example of fruitfulness of the international business cooperation.
Only by traditional neglect to invertebrates it is possible to explain silence of our popular writers about snouters, especially on the background of their unreciprocated love to the yeti and dinosaurs. Decades had passed from time of the first publication about Rhinogradentia in ōScience and lifeö magazine, but only now the usual reader can get acquainted with their irreproachably scientific, but quite accessible description.VIVOS VOCO!
Non cuicunque datum est habere nasum
Martialis
[The beginning of the article is omitted because it is only short re-telling of the book contents. - P. I. V.]
Until recently the science had no information, whether separate species of
Rhinogradentia were kept in water area of the sunken islands. We were
lucky to be sure that Rhinogradentia fauna had not died out completely, and
the discoveries
connected with mysterious snouters still are coming a science. But all under
the order ģ
In 1990 during the expedition onboard a research ship ōBipolarmondö
in a material collected by one of authors of these lines (A. Selbstander) in
the northwest
part of Weddell Sea, at the coast of Cuvette Island, the unknown colonial organism
had been found. Primarily this small colony found out in sublittoral zone at
the depth of 4 ¢ 5 m was identified as a kind of seaweed because of the presence
of rhizoids and branching runner at this one. The main difference of this colony
from all things known before is strongly thickened basal part that is typical
for spurges ¢ terrestrial plants of Euphorbia genus. Later, as a result of
more detailed studying of a colony, it was concluded that it belongs to one
of the most known groups of sea invertebrates ¢ Hydrozoa, and to be exact ¢
to unknown genus of Athecata order which has bulb-like tentacles of
feeding polyp and are lack of perisarc (a skeletal environment) around the
head of
polyp.
Approximately at the same time another author of this article (S. Fieldojants) was engaged in research of the collection gathered by Russian skin-diver biologists A. Gannibalov and E. Kargov in northern part of Davis Sea, near the Goodnight Island. In collection the strange oval-shaped organism covered of plates resembling scales and producing small runners was found out. Besides the sample taken in plankton at the coast of island contained tint jellyfishes of similar morphology. It was as difficult to identify these organisms, as finds of German colleague.
After some years, during the teamwork with the Antarctic collections at BPG University, we have compared mysterious sea organisms being in hands of each other. By then collections had been replenished with a similar new material. Joint research has allowed to make the courageous assumption: strange organisms found at shoaliness of Cuvette and Goodnight islands belong to Rhinogradentia group discovered 50 years ago at Hy-Yi-Yi islands and described by Stumpke. Animals discovered at the coast of Antarctica had been classified in two new families ¢ Hydroidopsidae and Scyphoidopsidae.
For confirmation of correctness of our conclusions we had addressed for consultation to known taxonomist, professor V. Richword working in Leiden. At first professor had stated doubt relatively to similar interpretation, but later he had agreed with our opinion and demonstrated the collection with organisms closed in morphology collected during the expedition of ōOnion Performanceö at the coast of Closet Island. Richword had kindly given the material in our order. So, we shall try to give taxonomic diagnoses to three new species of Rhinogradentia to describe their behaviour and ecology, to reproduce life cycles, and also to determine the place of described families in natural system of group.
Ordo Rhinogradentia subordo Polyrrhina
Tribe Hexarhinida
1. Family Hydroidopsidae fam. n.
Genus Rhizoidonasus gen. n.
Species Rhizoidonasus euphorbiformis sp. n. [6]
Typical material: holotype (N1/7843) is preserved in collection of BPG University; paratypes (N2/1011 and N3/1012) are in collection of PZR Institute. The material is collected in February ¢ March 1990 and 1991 in Weddell Sea near Cuvette Island at the depth of 5 m; ground ¢ sand and stones.
Description. Colonies are up to 20 sm in height. The parent organism is attached to the ground with the help of six noses thickened in the basis, each of which has ramified rhizoid covered with thin corneous skeleton. Rhizoid is terminated with a sucker inverted to the ground. Head has two big eyes and lengthened ears. Rather short body is narrowed in a direction opposite to the head. Body and basal part of each nose are covered with hair. Forelimbs (ōarmsö) and rear limbs (ōlegsö) have two very long fingers/toes, every one of them has a claw. ōArmsö and ōlegsö at the top part are indumented, and at the bottom are included in external skeleton. Extremely long polysiphon tail is ramified, on which branch young organisms of both genders are located. Noses of some of them are well advanced, supplied with rhizoids and ready to attaching on the ground whereas noses of the youngest ones are still lack of rhizoids, but each has one sucker.
Notes. The parental organism is attached to stony-sandy ground by all four thick, nestled to one another noses, rhizoids of which reach the distance of several centimeters and can braid thalli of sea algae, stick to stones or shells. In order to keep balance, the parental organism presses ears to the body and as if supports itself by ōarmsö extended along it, thus ōlegsö of animal keep tenaciously by claws against its own tail.
It is not evident yet how the reproduction of a described colony proceeds; it is not known, whether adult male individuals can produce it. It is noticed that the parental organism cares of posterity ¢ young individuals of both genders hang at its tail. It is not established yet how much time this connection lasts.
 |
Colony of Rhizoidonasus euphorbiformis formed by parental organism,
on which tail branches young individuals of both genders hang.
2. Family Hydroidopsidae
Genus Larvanasus gen. n.
Species Larvanasus haleciformis sp. n. [7]
Typical material: holotype (N1/10113) in collection of PZR Institute; paratypes (N2/2985) in PLN museum and (N3/7844, N4/7845) in BPG University. The material is collected in January, 1980 in Indian Ocean near Closet Island at the depth of 2 m on silty sand with stones.
Description. Extended and bent like a question-mark body (up to 5 sm in height) resembles very much by its shape the sexual zooids in colonies of representatives of genus Halecium (Hydrozoa). It is covered with thin transparent exoskeleton through which the internal chamber appears containing 9 ¢ 10 larvae of different ages and strongly distinguished by shape ¢ from worm-like ones covered with delicate ōfurö, up to fish-shaped and seal-shaped, and, at last, to organisms typical for Larvanasus. It is a fine example of reconciliation of organs till the onthogenesis process. The chamber is connected to an environment with a funnel ¢ larvatheca. Completely developed young larvanoses leave the parental chamber through the larvatheca. Head, neck and proximal part of three-fingered (three-toed) limbs are covered with fine hairs. The wide head of parental organism is supplied with six short, narrow and likewise haired noses, large ears, eyes on small stalks and unreasonably big mouth with wide lips.
At the same place another adult specimen (most likely the male) of the same species had been caught, but it has smaller size (3 cm), reduced chamber and larvatheca placed under the external skeleton. The surface of body and edge of lips of this specimen are covered with diatoms. Besides on the ground some tiny individuals (up to 2 cm) differed from adults only in long noses with suckers were revealed.
Notes. In the beginning of February at the same place where adult Larvanasus individuals had been met, the set of worm-like organisms about 1 cm long was revealed. At first they had been misidentified as flat turbellarian worms, but finally they had been identified as the larvae of Larvanasus. It is interesting that these larvae concentrated around of adult feminine individuals and were found in channels of their noses along which, probably, they penetrate into larval chamber. The masculine individual of Larvanasus represents obviously reduced form (larval chamber and larvatheca are reduced). After the spawning of sexual products in water male individuals, probably, lead a passive way of life, hiding in their external skeletal environment. Thus they feed passively too: diatoms plentifully covering a surface of their bodies penetrate through the nose into stomach with the current of water, the same way as the basic food of Larvanasus ¢ tiny planktonic organisms. There are reasons to believe that the external fertilization is peculiar to this species; the female individuals bearing young ones in larval chambers have a predominating role in life cycle of Larvanasus. Cubs found near parental organism on pebbles and sand moved on all six noses and four legs. It means that the reduction of locomotor function of a nose takes place only at adult stage of Larvanasus development.
 |
Family of Larvanasus. Through the transparent wall of the internal chamber of female individual (center) larvae at various stages of development are visible. At another one, most likely male (at the left), chamber and larvatheca are reduced. Around of parental organism worm-like larvae concentrate.
3. Family Scyphoidopsidae
Genus Nudirhinus gen. n.
Species Nudirhinus medusiformis sp. n. [8]
Typical material: holotype (N1/10114) is stored in collection of PZR Institute, paratypes (N2/7845) ¢ in BPG University and (3/2986) ¢ in RLN Museum. The material is collected in February, 1981 in Davis Sea near the Goodnight Island at the depth of 4 m on rocky ground.
Description. Small (2.5 cm in diameter) oval-shaped
organism is covered with thick external skeleton looking like plates with very
thin outgrowths similar
to byssus of molluscs. This specific ōshellö of Nudirhinus has four apertures
for the limbs, two for ears and one more for tail. As a rule, the head is inside
the shell, only very long ears, large oval eyes and six noses with a sucker
on each one are possible to be seen from outside. Adult organisms stand on
the ground and move on rock with the help of noses. Eyes shine in darkness
with blue light, that it is typical of some sea invertebrates (for example,
for hydroids of Obelia genus). The nature of the luminescence is probably connected
to the presence of special photocyte cells containing Ca2+-dependent photoprotein
in tissues of animal. If this version will be confirmed and it will be possible
later to find out places of Nudirhinus congestion, it may well be that these
animals will be used in medicine for early diagnostics of malignant diseases,
similarly to ōaequorineö and ōobelineö preparations received from some sea
hydroids [9]. Forelimbs (ōarmsö) terminate by typical claws for ground loosening,
rear limbs (ōlegsö) ¢ by four toes. Such anatomy of limbs assures to the animal
the movement on rocky ground.
The most interesting feature of Nudirhinus is the formation of buds on their
tails and ears. As the further researches had shown, these buds develop to
pelagic forms. The presence of vegetative stage is a phenomenon absolutely
not peculiar to rhinogrades. The attached buds can be seen on tales and ears
even at larvae. Paws and noses also become distinct at pelagic larvae at the
moment of their subsidence on a ground. Pelagic larvae of Nudirhinus are very
similar to scyphoid jellyfishes: the jelly-like body resembles umbrella, noses
are like oral lobes of jellyfish [10].
Notes. The reproduction of Nudirhinus is unknown. It was not possible to determine the gender of three found specimens. Probably, they are hermaphrodites. But there is also another version, according to which the pelagic stage is the gender carrier of the animal. In that case it appears the natural question: what habit of life of Rhinogradentia group must be initial from evolutional point of view ¢ ground or pelagic one. Apparently, from evolutional point of view the pelagic stage is nothing else than the transformed ground organism [11].
 |
The discovery of three new representatives of Rhinogradentia belonging to three new genera and two new families, puts very important theoretical questions to the science.
1. How real is the existence of Rhinogradentia fauna until now? It is probable to use several approaches for the solution of this question, including numerical ¢ for example, a statistical construction for definition of potential generic variety of group [12].
2. What can be a modern area of this group? As it is known, first finds of Rhinogradentia had been made on the islands of a southern part of Pacific Ocean which have sunk nowadays, our ones ¢ in cold waters of Southern Ocean. Apparently, Rhinogradentia is the bipolar group of animals living in cold waters of the ocean including the depths of Central Pacific, in Southern Ocean, and, quite probably, they will be found out in cold or temperate waters of Northern hemisphere [13].
3. What is the phylogenetic position of this group? In StumpkeÆs monography the evolutionary tree of Rhinogradentia genera is present. We have tried to find a place for new taxons described above on this tree. It is undoubtfully that all three genera should be placed on the right branch, between genera Eledonopsis (22) and Cephaloanthus (24). These genera belong to the Hexarrhinida (six noses) group. As it is known, Eledonopsis suavis has channels in nose through which the food passes into the stomach of animal. Noses of Larvanasus haleciformis have channels of similar structure. Thus, we place our genus Larvanasus in the basis of the branch. At the same time there are well-known colonial Hexanthus and Cephalanthus (Ranunculonasus and Corbulonasus) ones. We place our genus Rhizoidonasus (28) near to these hell-flowernoses (23 and 24). And finally, our Nudirhinus (29) may be placed near to Rhizoidonasus as representatives of both genera form the colonies of one shape or another. Unfortunately, the absence of live material deprives us with an opportunity to use molecular-biological techniques in our phylogenetic constructions.
|
Phylogenetic tree of Rhinogradentia.
|
1 Ś Archerrhinos, |
1. Pettersson-Skamtkvist E. The discovery of the Hi-Iay-Archipelago // J. A. geogr. 1943. ╣ 3. ą.322.
2. Morgenstern Ch. Galgenlieder. Berlin, 1905. (Transl. from Germ. S. Stepanyantz.)
3. Bromeante de Burlas y Tonterias J. A systematica dos Rhinogradentas // Bull. Darwin Inst. 1948. ą.2.
4. Combinatore M. Un pezzo di legno, appuntato trovato sulla spiaggia di Owsuddowsa. Milano, 1943; Schutliwitzkij I. I. Hat Morgenstern die Rhinogradentier gekannt? (Russisch mit deutsch. Zusammenfassung.). Berlin, 1947; Trufagura A. La cola de los Rhinogradentes // Boll. Arg. Rhin. 1948; Stulten D. The anatomy of the nasarium of Hopsorrhinus // Bull. Darwin. Inst. 1950. ą.4 et±.
5. Stumpke H. Bau und Leben der Rhinogradentia. Stuttgart; Jena, 1960.
6. The generic name is given due to the presence of rhizoids on the nose, the specific epithet means similarity of noses of every zooid with roots of Euphorbia plant.
7. The generic name is given due to that the nose has locomotive function only at larvae of this species, specific name becaues of similarity of the body shape of these organisms with gonothecae of hydroids of genus Halecium.
8. The generic name is given due to naked noses lack of hairs; specific name means the similarity with scyphoid jellyfishes.
9. Highotsky E. Ca2 +-dependent photoproteins and bioluminiscence of marine invertebrata. Proc. Internation. Invertebr. Conference. 1999.
10. Greyavin L. Evolution of the scyphomedusae behaviour // Proc. Behaviour Internation. Conf. 1989.
11. Offerro F. Life history of some low invertebrates // Phil.-Evol. J. 1969. ╣6. ą.61.
12. Nonsensevitch A. How many genera will be described during the closest future // Phil.-Evol. J. 1994. ╣3. ą.105.
13. Fieldojants S., Selbstander A. Problem of bipolarity on the marine invertebrate example. Phil.-Evol. J. (in press).
Free translation of a line of Roman poet Martialus (43 ¢
104)
ōNon cuicunque datum est habere nasumö
Not to mislead ignorant reader and not to anger the serious
one well understanding the zoology, we shall lay our cards on the table and
remind that the volume,
in which our (under the pseudonyms S. D. Fieldojants, A. Selbstander) article
ōMysterious snoutersö is published, was from April. Even more so, it already
happened once with the known taxonomist, professor V. Richword mentioned
in article. Having received as a gift pictures of our snouters, professor
had
grown dark and had said: ōI do not admit jokes in science!ö
The ōdiscoveryö of snouters and other not less fantastic animals belong
to known German zoologist Gerolf Steiner. Even in student time at Heidelberg
University (Germany) he had a good time in common with friends with inventing
of various
mythical animals, which descriptions and pictures formed a basis for development
of fantastic evolutionary ideas having a rational share, because hot discussions
promoted formation of phylogenetic thinking.

Self-portrait of Gerolf Steiner
(also known as Harald Stumpke
and Karl D. S. Geeste),
the known German zoologist,
the author of mysterious
Rhinogrades.
Even in times of Otto Butschli (1898) the multivolume edition
ōStammbuch des Zoologischen Institutö had been issued here, renamed subsequently
to ōSchwarzes
Buchö (ōBlack bookö) in which also Russian researchers V. V. Redikortzev,
N. A. Kasyanov etc. took part, by the way. The heroes of one volume issued
in
1929, were Balhurner (horned balls) ¢ hypothetical mammals living on bogs,
in steppes, deserts and on hills. The special attention was given to the
adaptations fixed during the evolution.
Before World War II Steiner worked as the assistant at the zoology faculty
at Heidelberg University. The professor of this faculty, SteinerÆs chief,
at that time observed of gregarious behaviour of minnows (Phoxinus phoxinus).
His experiments came to studying of consequences of removal of the part of
brain at these fishes. It was proved that as a result of such operation fishes
lost a gregarious instinct and aspired to leadership (Fuhrerschaft). The
conclusion
that removal of brain results in ōfuhreringö cost of freedom to the professor.
Steiner was invited to replace him. Working with students, he continued to
invent new animals and to draw their black-and-white and color pictures.
During the lectures Steiner showed these pictures. After the war cities Heidelberg
and Darmstadt, in which there was another, younger Darmstadt Technical University
(it is no more than 50 kms between them), appeared in American occupation
zone.
The house where there was SteinerÆs apartment, had been destroyed by bombing,
but he managed to hide his manuscripts, pictures and water colour paints
in a cellar of the destroyed house, and later to transfer to the empty colleagueÆs
house and to lodge there. Universities did not work. City dwellers stayed
at
home and only in the afternoon left on street for hour or two, trying to
find food. According SteinerÆs memoirs the feeling of famine was a dominant,
and
day was considered successful if on a lunch table snails, may-bugs, nettle,
or, if to be lucky, rabbit meat appeared. SteinerÆs student brought to him
for dinner asparagus from the garden, and he presented her with the pictures.
But duplicates remained. He tried to sell a part of them to earn for food...
In 1946 Darmstadt University had inaugurated again and Steiner started to
work there. But he did not forget the colleagues ¢ zoologists and friends
from Heidelberg
located nearby. Professor Werner Rauh had helped to make color slides from
SteinerÆs pictures, which another his colleague, Erich von Holst, used at
the seminars dedicated to evolution of animals.
Popularity of SteinerÆs pictures grew, and after horned balls snouters (Rhinogradentia) had came to light, which images also had been shown at seminars. Illustrations began to acquire the text gradually, and in 1950 Steiner had made the first attempt to issue the book about Rhinogradentia with his own color illustrations. But then it appeared too expensive to him. In 1957 he had written an epilogue to ōHarald StumpkeÆs manuscriptö about the fauna of the islands sunk as a result of nuclear tests.
The first edition of the H. StumpkeÆs (G. SteinerÆs pseudonym) book ōBau und Leben der Rhinogradentiaö was issued in 1960 in the most known and serious German publishing house ōGustav Fischer Verlagö (Stuttgard; Jena) specializing in publishing of guides, textbooks and theoretical monographies. The book made so stunning success that at the end of the same year it was republished. Originally the monography had caused a shock, of course: zoologists could not understand a dirty trick for a long time ¢ the trust to publishing house with a well-known emblem ¢ tail-shaking fish ¢ was too high... When the joke appeared revealed, Steiner had been forgiven for professionalism and that pleasure which every reader gets both from the text, and from humour, and from magnificent illustrations. It is not so surprising that the book was republished some times in German (last edition was in 1998 with color illustrations) and was translated to French, English and Japanese languages. Not so long time ago ōGustav Fischer Verlagö publishing house had published the monography ōStumpkeÆs Rhinogradentia. Versuch einer Analyseö, where in 119 pages the role of snouters group in evolution of animals is analyzed. The author of monography, Karl D. S. Geeste, is the same known professor of zoology Gerolf Steiner, now he is 91 year old.
S. Stepanjants
Cand. Biol. Sci.
Zoological institute of the Russian Academy of Science,
Saint Petersburg
└. Svoboda
The Ruhr University, Bochum (Germany)
ōBiologya moryaö (ōMarine biologyö) magazine, 2004, vol. 30, #2, pp. 163 - 164.
In 1987 in staff of small group of students I had summer
practice on marine zoology on Putyatin Island near Vladivostok. We dedicated
the most
part of time to lifetime supervision, dissection and sketch of various
representatives
of marine fauna, but sometimes it happened to put something interesting
in
jar with formalin in order to consider these objects later. After
returning to St.-Petersburg (then it still was named as Leningrad) because
of
every possible current affairs it was no time to examine the contents
of this
jar. Only recently,
putting things in order in laboratory, I had found the forgotten
jar and had decided to glance in it. Thus among sea worms, molluscs and sea
squirts, well-known
to a science and consequently not representing special interest,
I
found a piece of semi-rotten wood to which the small colony of a unknown
species
of
animals was attached, consisting only of three zooids and two buds.
Two zooids were strongly crumpled and destroyed, because by inexperience
I
had
filled
the jar too densely and gentle objects in it appeared in immediate
vicinity with rougher ones (for example, mollusc shells). Only one zooid
was kept
rather good. I had been really deeply surprised and admired when
at close examination
I had seen, that I have the representative of the extremely original
group ¢ the snouter order (Rhinogradentia) discovered by German zoologist
H.
Stumpke (the first reports about these surprising creatures were published
by him
in 50th, but I used the monography issued in 1993). Snouters are
remarkable because
their nose very easily adapts to performance of various functions:
many species use it for movement (hence the name of group), others for
digging
in ground,
and some for an attachment to substratum, as well as the species
appeared in my jar. H. Stumpke had described 26 species of snouters, more recently
S. Feldojants
and A. Selbstander (2000) described 3 more species, and now I have
the luck to discover one more (30-th!) species which I have decided to
denominate
as Dendronasus ¢ ōtreelike-nosedö.
Description. The colony of Dendronasus consists
of an axial column, or stalk which, most likely, is attached to the
bottom surface of
ships and wooden
constructions. On apical end of stalk hanging downwards the oldest
zooid,
presumably being
the founder of colony, is located; unfortunately, this zooid appeared
almost completely destroyed, but nevertheless it was possible to
see that the
stalk is the direct continuation of the nose of zooid. Approximately
in middle
part of axial stalk there is the thickening, from which the branches
grow reaching
the noses of two others zooids and to two still very poorly advanced
buds (see figure └). Obviously, this thickening represents a zone
of budding
and contains
the reserve elements of various tissues necessary for development
of buds. The general length of a colony does not exceed 2 cm.
The
structure of well advanced zooid is present in figure (B). Obviously,
zooids of Dendronasus should hang legs downwards, but, following the custom
which has established even in the classical monography of H. Stumpke, I have
represented it in nose downwards position. Rather compact zooidÆs body (not
including stalk) is approximately 5 ¢ 6 mm long (possible, these ones are the
tiniest representatives of mammal class). It has yellowish colouring and is
covered with fine golden hairs. Head is slightly bordered from the body. In
the basis of stalk-like nose there are nostrils, but we have no any data on
presence of lungs capable to serve for air breathing at Dendronasus; it is
very probable that nostrils only play a part of olfactory holes. The mouth
is slightly opened, and teeth are well visible in it. Very large convex eyes
indicate that sight sense has great importance in life of this animal, and
also specify the residence in poorly lighted places that is well coordinated
to the assumption of attachment of Dendronasus colonies to the bottom surface
of various underwater objects. Ears are small; hircuses are closely pressed
to the bottom of auricles and close the apertures leading to the external acoustic
duct; it follows from it that the hearing at these animals is strongly lowered
or lack at all.
Right behind the head two blade-like appendages adjoin to the body, and bunches
of delicate ramified strings grow from them. These appendages are very similar
externally to external gills. But the circumstance, that directly from each
blade 5 of such strings grow, which branch then, suggests that we look at modified
forelimbs which began to function as underwater breathing organs (and nails
have completely disappeared).
Dendronasus sp. A ¢ a general view of a colony (zooids are numbered according their age), B ¢ separate zooid at the enlargement.
Rear legs of Dendronasus have an anatomy of prehensile limbs and resemble
human hands very much ¢ the thumb opposites to four fingers. Therefore in the
beginning I had misidentified them as the forelimbs appeared on the back end
of the body because of any fantastic heterotopia. The impropriety of this idea
became clear only after I had understood the true nature of branchiate appendages.
Apparently, back legs serve for grasping of various planktonic organisms which
zooids eat. Tail is very long and flexible, with a thickening and large corneous
claw on the tip. At Enunctator sorbens described by H. Stumpke the tail has
a similar structure, and the tip thickening contains poison gland. Possible,
it is characteristic also for Dendronasus.
On the belly side of body
there are two swellings appropriate to position of milk glands from what it
is possible to conclude that examined zooid is
a female individual (but it is incorrect to exclude also the possibility of
hermaphroditism ¢ the phenomenon widespread at the animals leading the sessile
way of life).
The above mentioned description belongs only to appearance of Dendronasus (however
all other snouters are also described only superficially), but I hope that
studying of the rests of two others zooids will allow to receive data on internal
structure of this species which will make the contents of my next message.
After the end of work all made preparations and typical specimen of Dendronasus will be handed to the museum of Mythozoology Institute.
Discussion. As is known, many zoologists, including Prof. A. I. Lyagushenkov
(under the direction of which I had a luck to work till many years), have regarded
about the discovery of Dr. H. Stumpke rather skeptically. Prof. Lyagushenkov
even had declared that Rhinogradentias are descended ōfrom depth of the German
geniusö (see Ivanova, 2001). He also had regarded with the great doubt about
the message of S. Fieldojants and A. Selbstander, but now, having seen wonderful
Dendronasus, he was compelled to recognize an inaccuracy of the former statements.
To the present time it is known that Rhinogradentia order includes two suborders:
more primitive suborder Uni-Snouters, having only one nose, as well as the
majority of animals, and the suborder Multi-Snouters which have polymerization
of this organ. Our Dendronasus, indoubtedly, should be classified as Uni-Snouter.
From all known species of Rhinogradentia it is related closely of all to Dulcicauda
griseaurella and Columnifax lactans which use the nose for attachment to substratum,
and the latter species besides is the sea inhabitant sea though its adaptations
to aquatic habit of life are expressed not so evidently as at Dendronasus.
The transformation of forelimbs into gills and asexual breeding must be recognized
as the most remarkable features of Dendronasus.
Asexual breeding is practically alien for vertebrate animals, and until recently
the only exception was the
polyembryony of armadilloes which proceeds by very original way ¢ by change
of the embryonal development mechanism. But snouters present us with two
more examples of asexual breeding. In addition to Dendronasus it
is discovered at Rhizoidonasus euphorbiformis (Fieldojants, Selbstander,
2000) ¢ sea snouter
from Multi-Snouter suborder, at which buds are formed on tips of rhizoids
(thin root-like outgrowths branching from tips of noses). The belonging
of these two species to different suborders does not leave doubt that asexual
breeding has arisen independently at them. It is interesting that the nose
serves in both cases as base organ of asexual breeding ¢ it is the organ
which, as it was already marked, easily changes shape and function and
provides
an opportunity of the adaptation of animals to absolutely different conditions
of existence.
It would be very important to study not only the internal structure of Dendronasus (that,
maybe, will be possible for me), but also its individual development at
sexual and asexual breeding. However the last one is almost hopeless business,
because we do not know where to search Dendronasus, from what part
of World Ocean waves have brought to us a chip with a colony of this species.
It is
possible to expect only for a fortune.
╚ŌÓĒŅŌÓ ╬. ╠. ŽŅ„“Ķ Ō±Õ Ņ ¾±ÓļĻÓ§. čŽß: ╦Ķ±±. 2001. 157 ±. (Ivanova
O. M. ōAlmost all about mermaidsö SPb; Liss. 2001. 157 pp.)
įÕļõŅ Ē÷ č. ─., ŪÕļ³ß±“ÓĒõÕ └. ŪÓŃÓõŅ„Ē¹Õ ĒŅ±Ņ§ŅõĻĶ // ŽĶŅõÓ. 2000.
╣ 4. č. 46-54. (Fieldojants S. D., Selbstander A. ōMysterious snoutersö//
ōThe
natureö.
2000. #4 pp. 46-54.)
Stumpke H. Bau und Leben der Rhinogradentia. Stuttgart; Jena. 1993. 83
S.