| Главная | Библиотека сайта | Форум | Гостевая книга |
Если не вдаваться в таксономические неясности, то выяснить расселение ушастых
тюленей по земному шару как будто не должно составить труда. Стоит лишь нанести
на карту известные всем специалистам береговые лежбища десятка ли, полутора
ли десятков форм, выделяемых наиболее новой системой группы (см. рис. 23).
Ареалы одних ушастых тюленей дополнят или перекроются с ареалами других, и
получится зона, освоенная всем семейством. Тут же всплывают трудности.
Кроме береговых лежбищ, есть акватория, а распределение особей в море более
или менее целенаправленно изучалось лишь для северных морских котов в связи
с проблемами морского промысла, причем степень изученности хорошо показывает
удивление от обнаружения в середине нашего века зимующих у Камчатки зверей.
Для всего же Южного полушария, где котов на воде бить не было надобности, происходящее
с ушастыми тюленями в море покрыто густым туманом. В этом тумане малочисленные
сейчас стада выглядят вполне изолированными друг от друга, пока меченые звери,
иммунологические пробы или записи прошлых веков не обнаружат вдруг какой-нибудь
любопытной связи (например, неожиданной идентичности австралийских и южноафриканских
морских котов).
Впрочем, сегодня и эти трудности скорее технические, чем принципиальные. Можно
с помощью биохимических тестов определить степень разобщенности (или общности)
всех сегодняшних стад ушастых тюленей. Можно разместить по океанским берегам
сеть квалифицированных наблюдателей и наладить сбор и обработку получаемой
от них информации. Постоянно совершенствуемая техника учетов животных с помощью
авиации не сегодня, так завтра позволит картировать размещение отариид в океане.
А если к этому добавить экипировку некоторого числа особей данного стада (разного
пола и возраста) портативными передатчиками с надежными и долговечными батареями,
сигналы которых будут приниматься на один из многочисленных спутников, что
опять же активно обсуждается в рамках различных проектов мониторинга ресурсов
в последнее десятилетие, то можно будет уже через несколько лет уверенно говорить
о закономерностях размещения этих зверей во всех трех океанах. В пределах освоенного
данной формой ушастых тюленей (или всем семейством в целом) пространства вычленяются
несколько зон:
1. Зона, охватывающая все сегодняшние точки побережий, где появляются на свет
щенки данного вида ушастых тюленей и где соответственно, сегодняшние условия
пригодны как для длительного и регулярного пребывания на берегу взрослых особей,
так и для выживания приплода.
2. Одна или несколько зон плотных скоплений особей в море, т. е. зон, доступных
для многих особей и обеспечивающих их стабильным и обильным запасом кормов.
3. Зона, очерченная самыми дальними от мест размножения точками, посещаемыми
особями от случая к случаю, т. е. пригодная по сегодняшним условиям и доступная
для сегодняшних обитателей.
Выйдя за пределы предыдущей зоны, мы окажемся в местах, либо не пригодных по
сегодняшним условиям и потому недоступным сегодняшним обитателям рассматриваемого
ареала (т. е. выйдем за пределы экологической ниши владельцев этого ареала),
либо пригодных по сегодняшним условиям и недоступных из-за нехватки рекрутов,
т. е. в запустелой части данной экологической ниши. Эти две внешне неотличимые
ситуации (зверя нет, потому что не может быть его там; зверя нет, потому что
он пропал, но может еще вернуться) хорошо бы было различать, но делать это
труднее, чем картировать распределение сегодняшних обитателей. Уже вчерашнее
распределение зверей, не говоря уже о позавчерашнем, мешает вспомнить и короткая
память, и недолговечность свидетельств.
Насколько трудно вспомнить, что же жило несколько веков назад даже не в океане,
а в более доступных для наблюдения водоемах, показывает существование очень
интересного пласта свидетельств о неожиданных зверях в неожиданных местах.
Авторитетность очевидцев не позволяет вроде бы просто отмахнуться от их наблюдений,
а несуразность фактов не позволяет использовать эти факты без предъявления
добавочных доказательств. Вот, к примеру, три евразийских тюленя.
Не так широко известно неспециалистам, но не вызывает сомнения у зоологов,
что в, казалось бы, давно обжитом Черном море дотянул почти до наших дней один
из экзотических тюленей — средиземноморский тюлень-монах. Этому есть множество
очевидцев, и до сих пор предпринимаются попытки найти у болгарских и турецких
берегов последних из оставшихся в живых зверей. Но если открыть опубликованную
чуть меньше века назад книгу авторитетного петербургского профессора А. М.
Никольского «Позвоночные животные Крыма» (СПб, 1891), то в ней, кроме черноморского
монаха, окажется и другой вид тюленей — Phoca vitulina. Этот тюлень водится,
по словам квалифицированных очевидцев, в Черном и Азовском морях, везде так
же редок, как и монах, почти не выходит на берег, но довольно часто заходит
в Севастопольскую бухту. Чуть позже крупнейший российский специалист по ластоногим
Н. А. Смирнов в обзоре распространения ластоногих Северного полушария для бассейна
Средиземного моря называл обоих тюленей, хотя и писал, что «последний вид —
под сильным сомнением». Можно было бы отмахнуться от этих свидетельств как
от ошибочных (просто не узнали тюленя-монаха). Но почему бы, собственно, форме,
родственной каспийскому тюленю (он из того рода, что названный А. М. Никольским
зверь) и обитавшей в едином прежде Каспийско-Черноморском бассейне, не сохраниться
в обособившейся Азово-Черноморской части. Тем более, что остатки каспийского
тюленя известны из нижнеплейстоценовых отложений азовского побережья. К тому
же, как выяснилось в последние годы, современный каспийский тюлень освоил не
только северные, но и южные районы Каспия, принося щенков недалеко от Красноводска.
Так что климат Черного моря и его плейстоценовые перемены не должны были бы
быть серьезной помехой.
Еще менее можно предполагать тюленей в соленом озере Кукунор вблизи верховьев
Хуанхэ в северо-восточных предгорьях Тибета. Но вот недавно опубликовано письмо
исследователя Центральной Азии геолога В. А. Обручева из Су-Чжоу географу Г.
Н. Потанину, написанное 24 августа 1893 г. (опубликовано в журнале «Природа»
(1983, № 2. С. 67)): «Озеро Кукунор, несомненно, усыхает, представляя собой
остаток внутреннего азиатского моря, как и озера западного Цайдана... Мнение
о реликтовом характере Кукунора подтверждается рассказами монголов о существовании
в этом озере тюленя; рассказы правдоподобные — людей, видевших убитого тюленя
(которого они называют морской собакой) и описывающих его наружность вполне
верно; это факт новый и не менее интересный для зоолога, чем для геолога, Байкал
нашел себе соперника, еще перещеголявшего его своей абсолютной высотой...».
Если оставить в стороне занимавшую В. А. Обручева гипотезу о недавнем центральноазиатском
море, останутся неподтвержденные ни ископаемыми остатками, ни письменными источниками
рассказы охотников о тюлене (морской собакой называют тюленя жители всей Восточной
Азии), ближайшими соседями которого (если, конечно, он сам реален) были бы
байкальская нерпа и гипотетический тюлень, якобы виденный в прошлом веке вблизи
устья Хуанхэ. Что касается байкальских ластоногих, то вот что писал сосланный
в те края в середине XVII в. протопоп Аввакум (по тексту кн.: Изборник. М.:
Худ. лит-ра, 1969. С. 650): «Поехали из Даур... и до Байкалова моря доплыли...
Погода окинула на море и мы гребми перегреблись: не больно в том месте широко
— или со сто или с осьмдесят веръст... Около ево горы высокие, утесы каменные
и зело высоки... Птиц зело много, гусей и лебедей,— по морю, яко снег плавают.
Рыба в нем — осетры и таймени, стерьледи и омули, и сиги и прочих родов много.
Вода пресная, а нерпы и зайцы великия в нем; во окияне море большом, живучи
на Мезени, таких не видал». Существует несколько списков его «Жития» со многими
разночтениями, но во всех говорится как об общеизвестных сейчас байкальских
нерпах, так и о других тюленях, которых он называет поморским названием — морским
зайцем (или лахтаком), виденных триста лет назад в байкальских водах. Конечно,
мог и протопоп Аввакум ошибиться, несмотря на опыт своих многолетних скитаний
по Поморью и Сибири, могли напутать безымянные переписчики, донесшие до нас
его записи, но отбросить это свидетельство без проверки не хотелось бы.
Эти три полумифических, полуреальных зверя хорошо иллюстрируют, с какими проблемами
приходится сталкиваться при попытке восстановить облик промысловой фауны не
самых глухих мест и не самой глубокой древности. В любом случае эти сведения
заставляют при анализе сегодняшнего расселения ластоногих искать их следы и
в незаселенных сегодня местах. Если духи тюленей бродят по Центральной Азии,
то уж в океанах они заполняют все, казалось бы, непригодные для тюленей районы.
В Океании на севере до сих пор сохранились последние остатки гавайских тюленей-монахов
(еще и сегодня одиночки достигают атолла Джонстон на 17° с. ш.), жители центральной
Полинезии говорят о каком-то «тюлене» на расположенном на экваторе острове
Ошен. На о-ве Новой Ирландии, вблизи о-ва Новая Гвинея, местные жители верят
в существование водяного существа «ри» с человечьими руками и двумя ластами
сзади. Тур Хейердал в книге «Искусство острова Пасхи» приводит изображение
тюленеобразных статуэток.
В тропической части Индийского океана европейские мореходы конца XVIII в. видели
каких-то полуфантастических тюлене- или дюгонеобразных зверей у нескольких
из островов группы Сейшельских. В эту зону и сейчас заходят скитающиеся по
океану молодые морские слоны, так что легко допустить несколько большее число
тех же визитеров в позапрошлом веке и списать на них все наиболее правдоподобные
встречи. Но и эта, хотя и правдоподобная, версия висит в воздухе из-за недостатка
фактов.
В Атлантике сейчас пустуют по крайней мере две тюленьих вотчины. На Канарских
островах какие-то тюлени дожили до времен Колумба, о чем напоминают и хроники,
и названия островков, бухт и мысов. Этот зверь заполнял отчасти брешь между
тюленями-монахами Вест-Индии, исчезнувшими лишь в последние десятилетия, и
побережья Северо-Западной Африки, где все еще тлеют одна или несколько их колоний.
И в этом случае систематическая принадлежность канарских обитателей определяется
простым соображением, что, кроме монаха, быть больше некому, хотя ни рисунков,
ни костного материала не найдено. К югу же от экватора несомненно были тюлени
на о-ве Святой Елены, На нем добывали каких-то «морских коров» и в 1682 г.,
и в 1739 г., а последнюю прикончили в 1810 г. Может быть, это были те же морские
слоны, одного из которых, к счастью, догадался нарисовать удивленный его видом
Питер Мёнди в середине XVII в. Этот рисунок — один из немногих дошедших до
нас однозначно толкуемых документов о фауне малообжитых земель.
Приведенные примеры, во всяком случае, показывают, что ластоногие не приурочены
только к шельфовым зонам и приполярным районам и что отсутствие в каком-либо
районе следов ластоногих в памяти местных жителей и исследователей не исключает
возможности их обитания здесь даже в недавнее (век или несколько веков назад)
время. Наконец, получается, что уверенно утверждать невозможность присутствия
каких-нибудь ластоногих нельзя практически ни для одного района мирового океана.
Эти же примеры, однако, показывают, насколько труден сегодня сбор фактов о
прошлом распространении зверей. Анализ устных сведений, передаваемых из поколения
в поколение местными жителями, для опознания и привязки к месту действующих
там разнообразных зверей требует специальной подготовки или хотя бы более тесных,
чем сейчас, контактов между фольклористами и зоологами. Анализ сведений о зверях,
сохраненных в старых записках и отчетах, а также в названиях различных деталей
местности (мыс, скала в море, бухта), требует каталогизации разбросанных по
разным странам архивов и возможности извлечь нужное. Результативность же зоологических
исследований архивов была прекрасно показана исследованиями С. В. Кирикова
недавней промысловой фауны российской лесостепи.
Но наиболее значимы, конечно, костные остатки ушедших обитателей. А кость плохо
сохраняется почти на всех побережьях, да и редко привлекает там квалифицированное
внимание. Крайне важны костные скопления у поселений и мест промысловой активности
прежних насельников. Однако археологические сборы редко получают полноценную
зоологическую обработку, так же рассредоточены, как зоологические и палеонтологические
коллекции. Информация об остатках фауны в них, как о не самом важном для археолога-владельца,
еще труднее доступна для исследователей других специальностей и мест обитаний.
В итоге проблема сбора фактов о прошлом распространении зверей упирается в
специфику современных исследований (наверное, в любой области), состоящую в
резком увеличении числа исследователей, их узкой специализации с одновременным
уменьшением самостоятельности в проведении работ и постановке вопросов и соответственно
и контактов не только между специалистами в разных областях, но и между близкими
соседями. И все это — в условиях резкого роста технической оснащенности и расширения
возможностей в коммуникациях и технических перспективах.
Для выяснения степени близости существующих сегодня стад отариид нужна хорошо
оснащённая иммунологическая лаборатория, имеющая в своем распоряжении сыворотки
крови из нескольких десятков точек в трех океанах и заинтересованная в решении
конкретной таксономической задачи. Для изучения морской жизни каждого стада
нужна практически глобальная система слежения за несколькими десятками снабженных
радиопередатчиками особей каждого стада, занимающаяся этим делом из месяца
в месяц на протяжении по крайней мере нескольких лет. Для работы с накопившимися
за несколько веков и продолжающими поступать сообщениями о встречах отариид
(живых или костных остатков) вдалеке от сегодняшних лежбищ нужна предварительная
каталогизация (с составлением предметных и географических указателей) материалов
архивов, музеев, картохранилищ, охотничьих и рыболовных инспекций, метеослужб
и т. д. многих стран мира. Необходимо создание соответствующих поисковых систем.
То есть нужно то, что еще далеко не достигнуто даже в отношении библиотечных
фондов. Кроме всего этого, нужны целенаправленные поиски следов островных фаун,
существовавших на освоенных человеком островах при начале колонизации, а такие
поиски на океанических островах практически только начинаются. Об их плодотворности
говорят работы С. Олсона по исчезнувшим птицам Гавайских островов и о-ва Св.
Елены.
Однако пока возможность получения новых фактов только маячит впереди, даже
то немногое, что есть в руках сейчас, позволяет увидеть ареалы сегодняшних
отариид несколько по-иному, чем на приведенной выше схеме (см. рис. 23).
Сначала посмотрим по отдельности каждую из трех свободно перекрывающихся групп.
У морских котов (Callorhinus) линия, связывающая северные гаремные лежбища,
тянется от юго-восточной оконечности Сахалина (о-в Тюлений) к Командорским
и еще дальше на северо-восток к островам Прибылова. К северу от этой линии
в Охотском море лежит о-в Ионы, на котором в конце прошлого века, по отчетам
зверобоев, было взято две с лишним тысячи котовых шкур и лежбище иссякло. Но
в существование здесь всего сто лет назад гаремного котового лежбища трудно
поверить и из-за суровости условий, и из-за отсутствия подтверждений от более
поздних и более ранних наблюдателей, проще списать все на ошибку в отчетности
(сивучьи шкуры — а сивучиное лежбище существует на острове и сейчас — по ошибке
были означены в отчете котовыми).
К северу от северной границы зоны береговых лежбищ тянется зона, в которой
котов регулярно видят на плаву и иногда их небольшие группы выходят на берег
в разных участках азиатского и американского берегов, вплоть до южных подходов
к Берингову проливу. Наконец, еще севернее отмечены лишь отдельные встречи
котов, но эти встречи очерчивают большую акваторию в Северном Ледовитом океане
от Колючинской губы на западе до восточных берегов моря Бофорта на востоке.
Говоря о редкости встреч котов в окрестностях Берингова пролива, надо помнить
о малой плотности наблюдателей и о том, что, несмотря на это, например, узко
локальный язык одной из групп азиатских эскимосов (науканцы, живущие вблизи
мыса Дежнева) имеет отдельное название для морского кота, чего не приобрел,
скажем, никто из латиноамериканцев.
Рассматривая южную границу береговых лежбищ северного кота, видим сильно ломанную
линию, идущую от того же Тюленьего острова через центральную часть Курильской
гряды (сейчас лежбища только на островах Ловушки и Среднева, а в прошлом веке
— и на островах Райкоке и Броутона) к центральной части Алеутской гряды, где
на одном из Крысьих островов (о-в Булдырь), по словам алеутов, были лежбища.
И затем — круто на юго-запад к о-ву Сан-Мигель у калифорнийских берегов, смыкаясь
с зоной, заселенной южными морскими котами (Arctocephalus).
Южнее границы береговых лежбищ идут хорошо освоенные акватории до 38-й параллели
у корейских берегов и до 36-й параллели у восточных японских берегов. А затем
их южная граница пересекает океан на уровне той же параллели (о регулярных
встречах котов к северу от Гавайских островов писал еще сто лет назад Л. Стейнегер,
а в наши дни — китобои А. А. Ровнин и Л. М. Вотрогов), опускаясь на подходе
к калифорнийским берегам до 33-й параллели. То есть при явно неравномерном
распределении лежбищ вдоль азиатского и американского берегов Тихого океана
глубина проникновения на юг примерно одинакова и там и там.
Не ясно, приведет ли дальнейший рост лежбищ Сан-Мигеля к проникновению его
резидентов за сегодняшние морские границы на юг, во владения южных морских
котов. Пока же нет никаких свидетельств выхода рода Callorhinus к южной оконечности
о-ва Кюсю, северным Гавайским островам или к о-ву Гваделупе, хотя, кажется,
доплыть туда при котовых способностях ничего не стоит.
Гваделупский морской кот, самый северный из рода Arctocephalus, прежде, возможно,
заселял острова вблизи американского континента между 34° и 18° с.ш. Он и сейчас,
при сохранении единственной небольшой колонии на Гваделупе, контактирует с
северным собратом на Сан-Мигеле, т. е. звери могут уходить от лежбищ не меньше
чем на 500 км. Но сведения о проникновении на острова Ревилья-Хихедо (юг предполагаемого
ареала) содержатся лишь в записках капитана Моррелла (1832 г.), а их правдивость
сейчас находится под сомнением. На южных границах этого вида начинается пяти-
(или четырех-, если верить Морреллу) тысячекилометровый разрыв в ареале рода
вплоть до экваториальных Галапагосских островов. Расстояние между краями разрыва
не больше расстояния, преодоленного прибыловскими котами при колонизации Сан-Мигеля,
но острова на этом пути пустуют (правда, на одном из них, Клиппертоне, Моррелл
кого-то добывал, но это все тот же подозрительный Моррелл), и большинство современных
исследователей уверены, что для преодоления разрыва нужны глобальные климатические
перестройки.
Зато ареал галапагосского морского кота, охватывая лишь этот архипелаг с малочисленным
и на первый взгляд малоподвижным населением (в разгар промыслового бума в 1816
г. взято лишь 8 тыс. шкур; а сегодняшние встречи в море приурочены к 50-километровой
зоне вокруг островов), примыкает к ареалу южноамериканского морского кота,
лежбища которого охватывают континент от перуанского мыса Паракас вдоль берега,
включая Фолклендские (Мальвинские) острова, до уругвайских островов Лобос.
На юге южноамериканец отделен только проливом Дрейка от котов Субантарктики,
на западе к нему примыкает океанский форпост — хуан-фернандесский морской кот,
прежде насчитывавший в обоих своих очагах (архипелаги Сан-Феликс и Хуан-Фернандес)
более миллиона особей. Тысяча километров между двумя архипелагами вполне преодолима,
а в двух тысячах километров на запад лежит о-в Пасхи с его легендами, но без
каких-либо подтверждений присутствия ушастых тюленей когда-нибудь. Субарктические
острова южной Пацифики (Петра I, Скотта и Баллени), по-видимому, непригодны
для котов из-за ледовой обстановки. Так что между южноамериканскими и новозеландскими
лежбищами оказывается порядка 8 тысяч километров, лишенных каких-либо следов
присутствия не только морских котов, но и вообще ластоногих (впрочем, на о-ве
Пасхи Т. Хейердал все же нашел зуб морского леопарда).
Западнее начинается чересполосица. Новозеландский морской кот плодится в зоне
от о-ва Маккуори (55° ю. ш.) до островов Чатем на востоке, а на запад — до
юго-западного края Австралии, включая все южное побережье материка западнее
западного входа в Бассов пролив. К северу от этой зоны проникают одиночки,
достигающие на востоке ареала Новой Каледонии, а на западе, вероятно, тоже
заходят севернее гаремных лежбищ, но наблюдений нет. Между новозеландской и
австралийской частями ареала этого вида морских котов вклинивается кусок ареала
южноафриканских котов, непонятно как оказавшихся разделенными Индийским океаном,
но тем не менее благополучно заселившими Тасманию, острова на востоке Бассова
пролива и юго-восток Австралии, проникая на север вдоль побережья, по крайней
мере до 27-й параллели. Разрыв в 9 тысяч километров без каких-либо следов южноафриканских
котов на островах Индийского океана отделяет западный кусок видового ареала
от восточного. При этом, почти не проникая на восточное побережье Африки (нет
находок ни на берегу, ни в море севернее 33-й параллели), африканский морской
кот распространяет зону береговых лежбищ по западному (атлантическому) побережью
до 21-й параллели, а в море его видят до 11-й параллели. Хотя коты этого вида
уходят на несколько сот километров от берега, свидетельств прошлого или сегодняшнего
присутствия за пределами материкового шельфа нет.
Напротив, два других индоатлантических вида морских котов явно не сторонятся
глубоководных районов: один идет узкой и длинной широтной полосой к северу
от границы антарктической конвергенции, размножается на островах Тристан-да-Куньи,
Марион, Сен-Поль, Амстердам и Маккуори (т. е. вклинивается между восточными
и западными южноафриканцами и смыкается с новозеландцами); другой примыкает
к ареалу южноамериканца, идя от Южных Шетлендских островов (Ливингстон и Кинг-Джордж),
где гаремные лежбища немного не доходят до Полярного круга, по островной дуге
к Южной Георгии с ее стадами, сравнимыми при открытии с прибыловскими, а затем
к островам Буве, Марион (смешиваясь с предыдущим видом), Кергелену и Хёрду,
где упирается во что-то, устанавливающее видовую границу здесь, посреди Индийского
океана.
Получается, что среди девяти видов морских котов (если действительно число
и граьицы видов таковы) наименее приурочены к ограниченным участкам шельфов
наиболее южные и наиболее северная формы. В целом же вся зона, освоенная морскими
котами, простирается от летней границы льдов в Чукотском и Бофортовом морях
на юг примерно до 30-й параллели. Места зимовок и летовок вполне могут совпадать,
но зона, пригодная для береговых лежбищ, не заходит так далеко в приполярье,
как освоенная, и заключена между 60-й и той же 30-й параллелями. Особи любого
стада могут достигнуть практически любой точки видового ареала, включая центральные
районы океана. К этому массиву на юго-востоке прилеплен изолированный сейчас
форпост южных котовых стад.
В Южном же полушарии морские коты в продвижении на юг также ограничены морскими
льдами, а на север могут проникать до Анголы и Новой Каледонии. Хотя зона береговых
лежбищ, как и в северной Пацифике, зажата между 60-й и 30-й параллелями, но
ее язык в восточной Пацифике выходит за пределы зоны на 30° к северу, достигая
и осваивая экваториальные Галапагосские острова. Подвижность особей внутри
родового ареала, во всяком случае, в его приполярной части, высока, и коты
с о-ва Амстердам выходят на о-в Маккуори, пройдя по открытому океану не менее
7 тыс. км, т. е. всего в полтора раза меньше, чем отделяет о-в Маккуори от
берегов Южной Америки.
Так что, несмотря на трудности в истолковании разрывов между новозеландскими
и южноамериканскими (как и между галапагосскими и гваделупскими в Северном
полушарии) стадами, можно считать членов группы морских котов (и северных,
и южных) достаточно подвижными, чтобы совершать трансокеанские миграции и обживать
районы от Полярного круга до экватора.
Из пяти форм морских львов по крайней мере две — сивуч и южноамериканец — образуют
хорошую группу дополняющих друг друга ареалов. Сивуч дублирует распространение
северного морского кота: наиболее северные береговые лежбища у мыса Наварин
и на островах Прибылова. Так же как и коты, сивучи обжили не только район Берингова
пролива, но и проникают вдоль берега моря Бофорта до 139° з. д. Обжиты лежбищами
острова северной части Охотского моря — обе островных дуги (Курильская и Алеутская).
Самые южные гаремы с американской стороны на том же острове Сан-Мигель, а с
азиатской стороны сивуч, судя по топонимике, заселял Японские острова до 37-й
параллели со стороны Японского моря и до 35-й, если не до 33-й, со стороны
Тихого океана. О заходах сивучей южнее сан-мигельской параллели нет сведений,
так что от самых северных (изредка наблюдаемых на Галапагоссах) южноамериканских
морских львов отделяет около четырех тысяч километров американского побережья.
С западной стороны океана японских сивучей отделяет от австрало-новозеландских
морских львов около восьми тысяч километров. Расселение южноамериканского морского
льва тоже в общих чертах дублирует соответствующего морского кота: лежбища
— от северной части Перу на юг до о-ва Эстадос и с атлантической стороны, включая
Фолклендский (Мальвинский) архипелаг, до уругвайских островов Лобос. Одиночки
с одного края ареала доходят до Галапагосских островов, а с другого — до южной
Бразилии, где щенок пойман на 32-й параллели. Распределение лежбищ на Фолклендских
(Мальвинских) островах показывает, что что-то делает неприятной для зверей
повернутую в сторону Антарктиды юго-восточную сторону архипелага. А на лежащих
всего в 1000 км к востоку островах Южной Георгии морских львов не нашли и в
момент открытия. Что такое расстояние не помеха само по себе, показывают визиты
на Галапагоссы. Климатических различий между заселенными районами и Южной Георгией
не заметно, но что-то мешает львам идти к западу от Фолклендов. Что-то сделало
для этой группы недоступной Африку и Субантарктику. Можно было бы постулировать
для группы тесную привязку к материковым прибрежьям и непригодность к трансокеанским
переходам, если б не Галапагоссы да не разорванные островные дуги северной
Пацифики.
Заведомо отделен от предыдущей пары форм калифорнийский морской лев, чей ареал
перекрывается и с сивучей, и с южноамериканцем. Сегодня ареал калифорнийца
разорван на три неравные части, одна из которых погибла на наших глазах. Основной
массив, давший видовое название, охватывает береговые лежбища от 36° с.ш. (наиболее
северные встречи самок чуть севернее, вблизи Сан-Франциско) до 21° с.ш., острова
Лос-Трес-Мариас, на юге, захватив и Калифорнийский залив. Заходы к югу от этой
зоны неизвестны, если не считать сомнительных свидетельств Морелла о каких-то
отариидах на Клиппертоне (как раз на полпути до Галапагосс). Но зато на север
и к тому же как раз зимой эти, по общему мнению, теплолюбивые звери регулярно
уходят до о-ва Ванкувер, хотя делают это сегодня только самцы, оставляя самок
за санфранцисской параллелью. О выходах в открытый океан ничего не известно
и считается, что звери ограничиваются каботажными плаваниями.
Второй угол треугольника дает галапагосское стадо, ограниченное только этим
архипелагом, несущее как будто морфологические следы (им придают статус подвида)
длящейся изоляции. Достоверно отмеченных встреч зверей этого стада дальше 100—200
км от береговых лежбищ не было. По всем признакам получается типичный изолят
в трех тысячах километрах от основного калифорнийского стада.
Третье стадо, существовавшее еще в начале нашего века, было тогда отделено
от двух других девятью и двенадцатью тысячами километров соответственно. Звери,
не отличимые по сохранившимся немногим музейным черепам от номинального подвида,
заселяли берега Хонсю, Кюсю и юга Корейского полуострова, а также острова в
Японском море. Наиболее южные лежбища топонимика отметила на 33-й параллели
(Кюсю), а наиболее северные заходы одиночек в этом веке были отмечены у южного
Сахалина и на южных Курилах, т. е. на 45-й параллели. Судя по немногим ископаемым
находкам, калифорнийцы жили в этих районах уже в плиоцене. Очевидно, что азиатский
кусок ареала сжат чем-то внешним, не дающим освоить тот же набор условий, который
освоен у берегов Северной Америки. В противном случае азиатское стадо должно
было доходить на юг до о-ва Тайвань, а на север — до Камчатки, но свидетельств
этого нет.
Получается занятное противоречие между способностью зверя жить в широком спектре
условий от зимнего Ванкувера до летнего тропика, приживаться на новых местах
(беглые из зверинцев калифорнийские морские львы освоили атлантическое побережье
Северной Америки от Ньюфаундленда до Луизианы) и сохранять морфологическое
единство популяций с двух противоположных берегов океана, а с другой стороны,
ограниченностью каждого стада сравнительно узким районом расселения и отсутствием
видимых контактов между сегодняшними (да и вчерашними) стадами. И наиболее
простым способом убрать противоречие представляется предположение, что контакты
между стадами, если и прекратились, то совсем недавно, а прежде либо ареал
был шире, захватывая все три стада, либо трансокеанские миграции позволяли
стадам контактировать достаточно свободно.
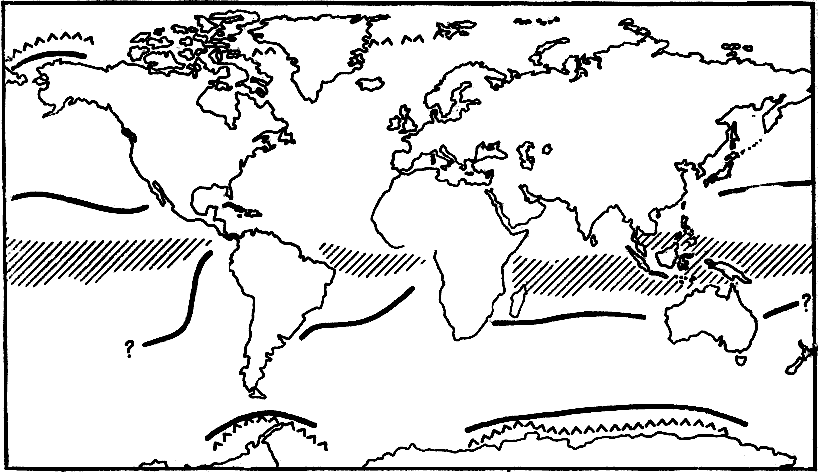
Рис. 24. Границы освоенных в обоих полушариях ушастыми тюленями зон с учетом морского распределения и контактов показаны толстой сплошной линией
Видно их хорошее совпадение и для Арктики, и для Антарктики с летней границей плавучих льдов (короткие зигзаги). Штриховка вдоль экватора показывает выделяемую климатологами зону, в которую сегодня не проникают ушастые тюлени. Хорошо виден зазор между полосой неосвоенного климата и реальным сегодняшним расселением
Последняя пара дополняющих расселение друг друга морских львов — австралиец
и новозеландец — может быть и является единой ареальной группой с сивучем-отарией
или с калифорнийцем, но и до тех и до других плыть надо весь Тихий океан поперек
(либо 8, либо 10 тысяч километров). Так что до появления новых материалов пара
остается особняком. Лежбища австралийца еще в историческое время были от Тасмании
и островов Бассова пролива вдоль всего южного и юго-западного побережья континента
и, возможно, до скал Хаутмена (на 28° ю.ш., в 70 км от западного побережья),
куда и сейчас при стаде, не превышающем 5 тысяч, заходят небольшие группы зверей.
Дальние выходы в океан неизвестны, и вид считается оседлым. Напротив, новозеландец
заселяет удаленные друг от друга океанские острова (Окленд, Кемпбелл, Снэрс)
примерно в пятистах километрах к югу от Новой Зеландии, а одиночки посещают
и Новую Зеландию на севере, где есть костные остатки тысячелетней давности,
и о-в Маккуори на юге. То есть пятисоткилометровые вояжи в открытом океане
нормальны и сейчас, а освоена меридиональная полоса между 55° и 45° ю. ш.,
несмотря на общую численность всего стада менее пяти тысяч.
Все ушастые тюлени вместе (рис. 24), таким образом, осваивают в обоих полушариях
акваторию от северной полярной шапки до южной полярной шапки, за исключением
Атлантики к северу от линии южная Бразилия—Ангола, половины Индийского океана
к северу от южного тропика и экваториального клина в Тихом океане между линиями
южная Япония—Гавайи—Коста-Рика и Коста-Рика—о-в Пасхи—северовосточная Австралия.
Сразу возникает вопрос, давно ли так стало и что, собственно, мешает сегодняшнему
ушастому тюленю последовать примеру белого прибыловского кота из сказки Р.
Киплинга и обшарить оба полушария в поисках безлюдного и безопасного места
для лежбища или хотя бы просто немного расширить существующие границы на юг
и на север. То есть не ясны соотношения потенциально возможного сегодня ареала
и того, который реализован в данный момент. С реализуемым ареалом сравнительно
просто — собирай сведения и наноси на карту. Вот только побольше бы наблюдений
в океане и получше бы знать связи между отдельными стадами, было бы на карте
меньше тумана. А как узнать потенциальный ареал?
Многочисленные связи соединяют каждый из живущих на планете видов (соответственно
и каждого из десятка или полутора десятков ушастых тюленей) с широким кругом
видов, поставляющих ему необходимые компоненты атмосферы, доступную воду, пищу
и разнообразную информацию о происходящем вокруг, а также с видами, для которых
он сам — пища или источник чего-либо другого, необходимого. Одновременно морфологические
и поведенческие возможности зверя лимитируют набор пригодных для него климатов
и рельефов.
Поскольку то же можно сказать о любом из нескольких миллионов существующих
ныне видов, то в целом получается сложное, тесное и прочное переплетение нескольких
миллионов компонентов друг с другом и геосферой в единую целостную конструкцию,
где каждый вид имеет свое четко очерченное и устойчивое место, видовую экологическую
нишу.
Это многомерное место в системе, проецируясь на двумерную поверхность, очерчивает
потенциально пригодную для данного вида зону (т. е. то, что он мог бы заселить,
если бы перед расселяющимися особями не вставали бы барьерами непроходимые
или бескормные места, заслоны из хищников и нехватка собственных рекрутов для
заселения пустот). Ко всему и с самой нишей может случиться что-либо, меняющее
ее размеры.
Все же очень заманчиво исходя из видовых характеристик оценить экологическую
нишу данного вида и, прикинув сегодняшние факторы в климате и структуре земной
поверхности, рассчитать, где именно и до каких пределов должен наш зверь обитать
сегодня. А коль его нет в предназначенных районах, то на наших глазах происходит
либо разрушение популяций, либо разрушение экологической ниши.
Посчитав все это, можно было бы заранее знать, куда расселятся в Евразии ондатра
или колорадский жук, где еще сохранились места, потенциально пригодные для
исчезающих мадагаскарских лемуров, и какой уровень промысла трески еще не повреждает
ее ареала. Но быстро оказывается, что для таких прогнозов нужен другой уровень
знаний. Для ушастых тюленей же все расчеты осложняются тем, что они в отличие
от медведя и дельфина живут в двух мирах.
Но попробуем прикинуть, что может остановить ушастого тюленя в воде, кроме
сплошных льдов, слишком горячей (или слишком холодной) воды, бескормного пространства,
невыносимого давления хищников и отсутствия рекрутов для освоения далеких от
дома акваторий.
Плавучие льды (родной дом для отариидной родни — моржей), по-видимому, непреодолимы
для ушастых тюленей. Об этом говорят и прямые наблюдения, и совпадение самых
полярных встреч этих зверей в водах обоих полушарий с означенными на картах
летними границами льдов. Поскольку теплоизоляция ушастых тюленей настолько
надежна, что позволяет месяцами (а то и годами) находиться в неограниченном
по теплоемкости теплопроводнике, Мировом океане, без переохлаждения, то трудно
представить, что это можно делать только при температурах воды в 10—20° и нельзя
в акваториях, остуженных до 0°. Так что даже для тех отариид, которые явно
не идут в воды, близкие по температуре к точке замерзания, как, например, калифорнийский,
южноамериканский, австралийский и новозеландский морские львы, это нельзя списать
на переохлаждение в воде. Тем более, что наиболее теплолюбивый (или теплоустойчивый)
калифорниец как раз на зиму откочевывает от согретой Калифорнии к холодному
Ванкуверу.
Скорее можно ожидать в теплой воде препятствия к расселению хорошо изолированных
от рассеивания тепла зверей. Тем более, что прямые наблюдения как будто однозначно
показывают приуроченность заходов в теплые воды к ветвям соответствующих холодных
течений. Легко представить себе трудности, возникающие у преследующего рыбу
на большой скорости в воде, близкой по температуре к температуре тела, если
тело укрыто надежной шубой. Но неразрешимы они лишь при наличии в конструкции
чего-то, мешающего перегревшемуся в воде зверю снизить мышечную активность
и соответственно теплопродукцию или, наоборот, увеличить кроветок в больших
по поверхности и лишенных теплозащиты четырех ластах и благополучно сбросить
избыток тепла в безграничный по теплоемкости океан, если, конечно, окружающая
вода хоть на несколько градусов холоднее тела. Такого препятствия в конструкции
как будто нет хотя бы у калифорнийского морского льва. Он, по всей видимости,
вполне благоденствует в водах Калифорнийского залива, где поверхностный слой
воды прогревается до 30° и не остывает ниже 15°. То есть общая для всех членов
семейства конструкция тела позволяет достаточно длительные теплые ванны. Как
будто об этом же свидетельствуют морские львы и коты в теплой летней воде зверинцев
и океанариумов в тропических и субтропических странах.
Бескормное пространство, способное остановить зверя, который может проплыть
в день километров двести и долго обходиться без еды, должно быть достаточно
большим и лишенным широкого набора кормовых объектов, так как ушастый тюлень
не слишком привередлив в выборе и готов проглотить все соразмерное глотке.
Акватория должна быть настолько бедной, чтобы при скоростях плавания 10—15
км/ч и освоенной стометровой толще нельзя было собрать пяти-десятикилограммовый
суточный рацион из рыб, кальмаров или планктонных беспозвоночных.
Низкопродуктивные зоны Мирового океана представляют собой пятна в центральных,
далеких от континентальных шельфов районах трех океанов и в общем приурочены
к обоим тропикам, примыкая с севера и юга к высокопродуктивным экваториальным
районам. Как будто все эти голодные места можно обойти по сытным шельфам, да
и сами голодные зоны низкопродуктивны по сравнению с высокопродуктивными, обильными
жизнью Субарктикой и Субантарктикой.
В 1852 г. появилась «Меркаторская карта пяти частей света, изображающая: ход,
временное и постоянное местопребывание различного рода китов, с показанием
удобнейшего времени года для их ловли. Издана Российско-Американскою Компанией
по новейшим сведениям». На этой карте отчетливо видно, что еще во времена Г.
Мелвилла интересовавшие промысловиков скопления усатых китов и кашалотов заходили
глубоко в эти низкопродуктивные районы. Сегодняшние ареалы океанских дельфинов,
например, такой обычной и сходной по размерам и разнообразию кормовых объектов
с ушастыми тюленями формы, как дельфин-белобочка, вполне равномерно охватывают
просторы всех трех океанов от умеренной полосы одного полушария до умеренной
полосы другого полушария (см. карты видовых ареалов океанских дельфинов во
II томе сводки «Млекопитающие Советского Союза» под редакцией В. Г. Гептнера
(М., 1976)). Ресурсы низкопродуктивных океанских зон оказываются достаточными
и для некоторого количества обитающих там рыбоядных птиц, т. е. и на бескормицу
нельзя списать полное отсутствие любых отариид в тех или иных местах, если,
конечно, не говорить о вторичной бескормице в результате нерегламентированной
человеческой активности.
Крупные морские хищники, скажем, акулы, несомненно могут как-то влиять на численность
и плотность распределения тех или иных видов ушастых тюленей в море. Но полностью
блокировать освоение какого-либо района им не под силу из-за надежной обратной
связи между численностью добычи и потребителя. Это под силу, однако, совсем
мелким хищникам. Так, муха це-це блокирует проникновение рогатого скота во
многие районы экваториальной Африки (но не блокирует зебр и крупных антилоп),
но и на это у природы есть свои обходные пути. Быстрый рост возможностей человека
с его оружием и зверобойными шхунами может сделать то, что было не под силу
львам и акулам. Но развитый морской промысел был лишь в северной части Тихого
океана и начал довольно рано подвергаться регламентации. Теперь, правда, появилась
другая опасность: неумеренным промыслом или загрязнением вод подорвать численность
служивших отариидам кормом объектов. Но это уж проблемы сегодняшнего, а не
вчерашнего дня.
Остаются внутренние проблемы стада, привязанность к месту рождения, склонность
к оседлости и нехватка рекрутов для выхода в дальние воды. Но привязанность
к своему лежбищу не мешает уходить на несколько тысяч километров, и даже принимаемый
обычно за оседлого калифорнийский морской лев идет зачем-то па зиму на две
тысячи километров к северу. Можно предполагать, что запечатление на местность
происходит еще в раннем детстве, а на подвижность молодых влияет степень легкости
их вписывания в структуру стада.
В качестве предварительной гипотезы можно было бы предположить, что чем меньше
и компактнее стадо, тем меньше у него шансов выделить рекрутов для освоения
дальних окрестностей, тем теснее (особенно рано взрослеющие самки) и раньше
его молодежь приковывается к месту рождения. Можно полагать, что оседание молодежи
более надежно в стадах с менее выраженной сезонностью событий. Тогда более
приближенные к тропикам стада одного и того же вида отариид будут существенно
более оседлы, чем удаленные к полярным широтам. Но пока это только спекуляции,
для проверки которых нужно длительное наблюдение меченых зверей. Крайне важно
было бы узнать характер миграций (если они есть) наиболее южного из наличных
стад северных морских котов — сан-мигельского.
Таким образом, сейчас можно лишь говорить, что если и существуют факторы, препятствующие
ушастым тюленям плавать от одного полярного круга до другого, как дельфины
или белый кот из сказки, то это скорее особенности структуры стад, привязывающие
потенциальных первопроходцев к месту, чем гипотетическая возможность перегрева
в воде или малокормные акватории.
Посмотрим теперь на помехи в освоении побережий, перебрав тот же набор из риска
перегрева и переохлаждения, бескормицы, напора хищников и неспособности стада
продуцировать достаточное число колонистов.
На первый взгляд трудно представить, как может ушастый тюлень, избегая переохлаждения
в воде, опасаться его на воздухе. Но вот — неравномерность распределения береговых
лежбищ и морских львов и морских котов на приполярных островах. Так, котовые
лежбища на обоих Командорских островах последние два века располагались таким
образом, как если бы прятались от чего-то неблагоприятного, идущего с севера,
или, напротив, ориентировались на что-то благоприятное, идущее с юга. Так же
неравномерно освоены берега и более северных островов Прибылова, и на одном
из них, действительно, северной и западной сторон коты избегают, но на другом
что-то меняет ситуацию на прямо противоположную. В Южном полушарии отсутствие
котовых лежбищ на обращенных в сторону Антарктиды побережьях можно увидеть,
скажем, на о-ве Херд (53° ю. ш.) и на острове Элефант (61° ю. ш.), но вот на
Южной Георгии все нынешние лежбища открыты на юг. Зато вся юго-восточная сторона
Фолклендских (Мальвинских) островов свободна от лежбищ южноамериканских морских
львов.
При всей противоречивости этих наблюдений несомненно одно: в приполярных стадах
различных видов ушастых тюленей действует какой-то четко ориентированный по
сторонам света фактор, надежно исключающий образование береговых лежбищ на
как будто вполне пригодных для этого и легко доступных пляжах. Неизвестный
фактор ориентирован по-разному в разных местах и для разных видов. Это вряд
ли опасность перегрева, так как речь идет о приполярных, а не о субтропических
частях соответствующих ареалов. Скорее можно подозревать опасность переохлаждения
(не с ней ли, кстати, связано отчетливое убегание взрослых морских котов от
дождя в воду), хотя бы для щенков с их несовершенной теплоизоляцией (их часто
можно видеть сбившимися в кучу и дрожащими), или опасность появления способных
блокировать подходы к береговому лежбищу плавучих льдов.
Напротив, если возможность переохлаждения на суше проблематична для взрослого
ушастого тюленя, риск перегрева и теплового шока висит над ним летом даже на
нежарких Командорских островах, где в ясный день не выдерживают и сходят в
воду даже стойкие гаремные секачи. Та же проблема и на других островах Субарктики
и Субантарктики. Но универсальный способ — сойти при перегреве в воду или только
зайти в лужу холодной воды и намочить ласты — позволяет жить и на раскаленных
берегах Калифорнийского залива, где даже вода залива прогревается до 30°. Еще
показательнее Галапагоссы с морскими котами и двумя формами морских львов.
Обычно вспоминают о холодном течении, омывающем берега архипелага и влияющем
на микроклимат пляжа, но забывают о ясной погоде и экваториальном солнце в
зените.
Получается рискованная игра: легко получить смертельный тепловой удар на берегу
(треть сан-мигельских котовых щенков погибает в отдельные годы от перегрева,
хотя это только 34° с. ш. и песок при воздухе в 19° нагревается до 35°) или
его избежать, найдя возможность в критический момент остудиться в воде. Можно
прятаться не только в воду, а просто поселиться в прохладных гротах, как гваделупские
морские коты. Нужны бы специальные исследования тепловыносливости ушастых тюленей,
но и так ясно, что поведенческими находками можно многое компенсировать. Так
что, глядя на климатическую карту океанов и видя необжитой отариидами лишь
одну климатическую зону (см. рис. 24), было бы преждевременно списывать ее
неосвоенность только на невыносимую жару на суше. Но зато можно уверенно говорить
об отсутствии климатических барьеров для рассредоточения береговых лежбищ в
пределах зоны, включающей климат Галапагоссов, т. е. зона неосвоенного сегодня
климата оказывается неширокой экваториальной полосой.
Естественно, что миллион котов может существовать на одном острове только при
наличии в ближайших окрестностях, в одном-двух дневных переходах, угодий, обеспечивающих
на протяжении хотя бы летних месяцев стабильный ежедневный сбор примерно 5
тысяч тонн корма, учитывая, что основной потребитель — кормящая самка, холостяки
могут сплавать и в дальний район, секач постится, щенок — на молочной диете.
Полагая, что колония из тысячи голов обеспечивает каждому виду ушастых тюленей
достаточно прочную структуру лежбища, получим величину ежедневного сбора урожая
в три-пять тонн, т. е. столько, сколько нужно для прокорма примерно пятитысячной
колонии некрупных морских птиц (олуша, баклан, кайра), а такие колонии есть
или еще недавно были в каждом районе Мирового океана. То есть бескормица, делая
тот или иной остров непригодным для отариид, одновременно сделала бы его непригодным
для рыбоядных птиц. Так что вроде и кормность наличных в океане угодий может
лишь ограничить численность прокармливаемых отариид, а не исключить их из сообщества
нацело.
Наземные хищники и эпизоотии должны бы разгуляться в переуплотненных береговых
скоплениях ушастых тюленей. Но роль паразитарных болезней не изучена и для
больших стад. Правда, одна из форм гельминтоза признается ответственной за
существенную детскую смертность на благоприятных для ее распространения песчаных
пляжах, но истребить полностью какое-либо лежбище, а тем более целое стадо
паразитам вряд ли под силу.
Неизвестно, как происходило (если происходило) взаимодействие лежбищ ушастых
тюленей с крупными и средними наземными хищниками африканских, азиатских и
американских побережий. Насколько бы могли повредить структуру лежбищ медведи
или стаи собак при малой связи щенка с родителями и неспособности ушастых тюленей
к групповой защите или нападению на берегу. Ведь даже командорский песец время
от времени одолевает то одного, то другого котового щенка на глазах у взрослых
обитателей лежбища. Сейчас контакт ушастых тюленей с наземными хищниками сведен
к минимуму из-за сосредоточения лежбищ на труднодоступных островах и островках.
Исконна ли эта изоляция или приобретена недавно, может ли колониальный морской
зверь вписаться в материковое сообщество с малоспециализированными стайными
хищниками и противостоять их напору, нельзя сказать без прямых наблюдений таких
контактов или их результатов. А тем временем как раз такой эксперимент происходит
на побережье Южной Африки, где морские коты начинают с мелких прибрежных островков
заселять оставленные несколько веков назад берега континента в запретных для
людей районах. Роль наземных хищников в динамике стад ушастых тюленей, вероятно
все же, вполне сравнима с ролью морских хищников в морском расселении.

Рис. 25. На этой фотографии из журнала прошлого века хорошо видна техника забоя морских котов на берегу и экипировка промысловиков, вполне доступная и в каменном веке и сохранившая рациональность до наших дней
Обратите внимание на поведение диких сильных зверей, не делающих никаких попыток группового отпора
Эффективности же человеческого морского и берегового нерегламентированного
промысла различаются радикально. Хорошо известна и документирована способность
небольшой группы людей, вооруженных только палками, т. е. с экипировкой возрастом
по крайней мере в сотни тысяч лет, очистить лежбище за ограниченное время (см.
рис. 25). Вспомните морского волка из предисловия, инструктирующего новичка-зверобоя,
как за два-три сезона полностью вычистить найденный остров. Это ставило судьбу
каждого конкретного лежбища в прямую зависимость от его доступности для людей
и способности этих людей соразмерять промысел с необходимостью сохранять ресурсы.
Чем разнообразнее ресурсы острова и чем меньше зависят прибывшие на остров
от отдельного продукта, тем меньше у тамошних ушастых тюленей шансов пережить
встречу с пришельцами.
Через какое-то время о прошлых стадах, возможно, пожалеют и попытаются сохранить
остатки. Но произойдет это не при первом знакомстве, а позднее, и то, если
не появится волна новых колонистов, не имеющих местного опыта.
Эти простые доводы как будто легко объясняют приуроченность котовых лежбищ
в момент их обнаружения европейцами к необитаемым, необжитым индейцами, алеутами
или курильцами островам. Опасность новой волны колонистов наглядно показали
происходившие затем события. Первобытные морские охотники к этому времени использовали
добычу очень полно. Так, шкура убитого сивуча шла на обтяжку байдары, мясо
и жир — в пищу, кишки — на водонепроницаемую одежду, зубы и кости — на поделки.
Новая волна переселенцев-европейцев интересовалась больше товарной стоимостью
меха и ворвани, емкость рынка была большой, и лежбища быстро пустели одно за
другим. Понадобилось почти двести лет, чтобы осознать необходимость выработать
и внедрить простейшие правила регламентации промысла. И теперь нам приходится
по крупицам восстанавливать картину размещения и жизни отариид до начала широкого
промысла. Действенность человеческого пресса на современное распространение
морских львов и котов несомненна, надо лишь оценить его реальное действие.
Остается поискать, нет ли и в структуре берегового лежбища чего-либо способного
замкнуть его население и воспрепятствовать продуцированию переселенцев в соседние
и дальние страны. С одной стороны, чтобы сделать расстояние между двумя пригодными
пляжами непреодолимым препятствием для колонизации, нужно либо удалить их друг
от друга дальше, чем на пять тысяч километров (расстояние между источником
колонистов — островами Прибылова и местом колонизации — о-вом Сан-Мигель),
либо вводить дополнительные ограничения вроде предположения о большей изолированности
лежбищ в более теплых районах ареала вида. С другой же стороны, вероятнее всего,
именно особенности связей внутри стада мешают подвижным и не самым малочисленным
фолклендским морским львам преодолеть тысячу километров по течению и вдоль
той же параллели и присоединиться к морским котам Южной Георгии.
Существование замкнутого стада должно предполагать, что приплод только покрывает
убыль в стаде, и вся молодежь, по крайней мере все самки, легко вписывается
в существующую структуру лежбищ данного стада, не имея возможностей и позывов
покидать надолго родимые берега. Рост крупного лежбища, по-видимому, с неизбежностью
должен сопровождаться чередованием фаз изоляционизма и открытых дверей. Но
возможно, в монотонных субэкваториальных условиях при стабильном и ограниченном
количестве доступного корма и при ограниченности термально безопасных мест
на берегу можно, сократив рождаемость и увеличив оседлость, получить небольшие
замкнутые и стабильные группировки. Тогда, если по каким-нибудь причинам погибнет
одна или несколько таких группировок, в видовом ареале надолго образуется прореха
с пригодными землями и отсутствием желающих туда вселиться.
Таким образом, помех в расселении ушастых тюленей по берегам заметно больше,
чем преград в океане, и они существенно заметнее. При этом в субполярных частях
ареалов граница зоны береговых лежбищ значительно не доходит до границ освоенных
акваторий, в субтропических же частях обе границы совпадают. Резкое возрастание
теплового риска по мере приближения лежбищ к экватору сокращает количество
пригодных пляжей и ставит возможность существования береговых лежбищ в зависимость
от поведенческих уловок, т. е. придает еще больше значения внутренней структуре
стада в вопросе о возможности заселения того или иного теплого района. Наконец,
очевидным и очень мощным ограничителем в береговом расселении отариид служит
активность людей, способных перемещаться от острова к острову.
Получается, что потенциальным ареалом семейства ушастых тюленей в первом приближении
можно считать всю акваторию Мирового океана, от одной полярной шапки до другой,
в пределах которой распределение определяется возможностью наличной формы ушастых
тюленей образовывать береговые лежбища. А для них существует зона повышенного
риска в области экватора, которая в сочетании с большей оседлостью субэкваториальных
стад по сравнению с субполярными превращает экватор в преодолимый, но действенный
барьер между отариидами Северного и Южного полушарий.
Так что проблемы конфликта белого котика, могущего проплыть от Берингова пролива
до мыса Горн, и его соплеменников, не желающих сдвинуться с места, не ограничиваются
только сказочной ситуацией, придуманной Р. Киплингом.
В пределах потенциального ареала семейства возможно несколько вариантов соединения
отдельных стад в единый, реализуемый видовой ареал: 1) члены каждого стада
рассеиваются по всему видовому ареалу в те или иные периоды индивидуальной
жизни, разделяются же стада лишь привязанностью особей к разным лежбищам; 2)
члены каждого стада рассеиваются лишь в ближних окрестностях своего лежбища,
ограничивая контакты с соседними стадами своего вида не только привязанностью
к разным лежбищам, но и использованием разных акваторий.
Первый вариант наглядно показывают сегодняшние северные морские коты, второй
вариант вероятен, но нуждается в подтверждении прямыми наблюдениями. Если реальны
оба варианта, то сочетать их можно либо как видоспецифичные и приуроченные
к разным видам, либо как приполярный и притропический варианты в пределах одного
вида, либо как отдельные фазы в истории каждого стада одного вида (время прохождения
может либо совпадать, либо не совпадать у разных стад одного вида).
Вернемся снова к карте (см. рис. 24) с очерченной зоной, в которой сегодня
можно встретить ушастого тюленя. По самым умеренным прикидкам получается, что
климат, кормовые ресурсы и миграционные способности допускают значительно более
широкое расселение ушастых тюленей, чем то, кажущееся сейчас золотым веком
отариид, с которым столкнулись землепроходцы позапрошлого века. Особенно удивительна
пустота пригодной по всем параметрам Северной Атлантики, чью пригодность для
заселения подтверждают все более частые встречи беглых из неволи калифорнийских
морских львов, прижившихся на воле. То есть реализуемый ареал семейства не
только сегодня, но и до начала массового европейского промысла оставлял в пределах
потенциального ареала много места незаполненным, и этот зазор не удается списать
на достоверно известные барьеры расселению.
А если попробовать поискать причины сегодняшних проблем с расселением ушастых
тюленей глубже тех очень недавних времен, когда европейские зверобои начали
обшаривать Мировой океан. Ушастые тюлени современного облика, хорошо вписывающиеся
в современные рамки этого семейства, обитали у берегов Калифорнии, т. е. в
пределах современного ареала семейства, еще 10 млн лет назад и, следовательно,
имели некоторое время пообжиться в пределах ниши семейства. Для неспециалиста
рискованно спускаться на зыбкую почву реконструкций даже близкого геологического
прошлого (разные варианты изменений объема Земного шара, разные модели перемещений
тектонических плит и уж совсем разные у разных авторов реконструкции климата
и географии конкретных эпох). Но все же на фоне меняющейся географии и перераспределяющихся
климатов большинство исследователей видят некоторое количество вех, отмечающих
радикальные и глобальные перестройки, хотя степень радикальности и глобальности
всегда дискуссионна.
Для разговора о наших зверях надо помнить уж во всяком случае три последние
такие вехи. Порядка 60 млн лет назад какие-то катаклизмы с заметным похолоданием,
расползанием ледников и полярных шапок и понижением уровня Мирового океана
сочетались с исчезновением со сцены многих групп чудовищных рептилий и широким
расселением по Земле разнообразных крупных млекопитающих, в том числе и не
менее чудовищных. Следующие тридцать миллионов лет все вроде выравнялось, если
не считать тектонических подвижек и миграций полюсов, полярные шапки отступили.
Уровень Мирового океана приблизился к современному. На месте нынешней Прибалтики
среди сосновых лесов можно было встретить среди насекомых современные виды,
но о ластоногих известные нам куски геологической летописи молчат (о китообразных
базилозаврах и морских коровах они же кое-что рассказывают).
25 млн лет назад — снова похолодание с сопряженными переменами в Мировом океане
и распределении климатических поясов. О существенности этой вехи говорит геологическая
терминология, именующая время до нее палеогеном, после нее — неогеном. Двадцать
с лишним миллионов лет последующих событий (миоцен—плиоцен) проходили, по-видимому,
на фоне климатической обстановки, близкой к современной, и вывели на сцену
в начале этого промежутка медведеобразных эналиарктид и тюленеобразных аллодесмид,
а в середине — у калифорнийских берегов видны предшественники командорской
морской коровы и несомненные ушастые тюлени, в Северной Атлантике разнообразны
настоящие тюлени, а где-то в Африке уже ходят (или вот-вот пойдут) двуногие
приматы.
Полтора миллиона лет назад — последняя веха, начало протянувшейся до нашего
времени череды ледниковых и межледниковых эпох с последним отступлением холода
всего лишь десять тысяч лет назад. Эти события в отличие от миоценовых и плиоценовых
даже таких важных для расселения ушастых тюленей, как образования 5—7 млн лет
назад сплошного ледяного панциря зеленой до этого Антарктиды с сопутствующим
50-метровым понижением уровня океана, захватывали циркуляцию атмосферы и гидросферы
всей планеты. Размах этих глобальных событий определяется по-разному разными
авторами, но большинство склоняется к тому, что в периоды максимального развития
полярных шапок они покрывали до трети поверхности Земли (против примерно одной
шестой сейчас), понижая океан, по разным оценкам, на 80— 130 м к современному
уровню.
Сложнее с температурным режимом, особенно между тропиками, так как по одним
расчетам в тропической Пацифике средние температуры опускались на 3° (т. е.
тропические условия заменялись на субтропические), а в тропической Атлантике
— даже на 5—6°. По другим же, не менее авторитетным (реконструированные на
август 18 тысяч лет назад климатические карты, международный проект «CLIMAP»)
раскладкам выходит, что максимальное развитие полярных шапок больше влияло
на климат умеренных зон по сравнению с тропическими и материковых областей
по сравнению с океаническими, хотя и в океанских тропиках сказывается глобальное
охлаждение поверхностных вод и локальные языки холодных течений проникают к
экватору у западных берегов Африки, Австралии и Южной Америки.
Так или иначе, сто тысяч лет начавшейся полтора миллиона лет назад ледниковой
эпохи сменились двумя сотнями тысяч лет межледниковья с условиями, близкими
к современным. Потом сто двадцать пять тысяч лет следующей ледниковой эпохи
— и новый возврат к современным климатам на шестьсот с лишним тысяч лет, новое
похолодание на пятьдесят тысяч лет и новое трехсоттысячелетнее потепление.
Последнее перед нашим временем пятидесятитысячелетнее похолодание сменяет наша
оттепель, длящаяся уже (или еще только) десять тысяч лет.
Такую историю читают по океанским осадкам, и трудно представить, чтобы ушастые
тюлени не заметили происходивших на планете перемен.
Но перемены происходили не только в географии, менялось и разнообразие форм
млекопитающих. Простой подсчет числа семейств млекопитающих, выходящих на сцену
и уходящих с нее в каждом действии, сделанный Н. Н. Каландадзе и А. С. Раутианом,
обнаружил довольно хорошее постоянство общего известного палеонтологам числа
участников на протяжении всего послемелового времени (130 семейств в палеоцене,
150 — в миоцене, 120 — в плейстоцене). За это время отмечено появление 250
новых семейств и исчезновение 160 старых, причем если исчезновение выглядит
довольно равномерным и постоянным, то появление новых семейств, достаточно
бурное в начале (в эоцене половина семейств неизвестна для предшествующего
периода), редеет в середине (в миоцене новых — меньше трети) и практически
прекращается в последние десять миллионов лет. То есть нас, гоминид, объединяет
с отариидами и то, что обе группы успели на сцену как раз перед тем, как выход
новых актеров прекратился. Да и со старыми участниками не все ладно: после
плейстоцена их осталось на треть меньше, чем было в миоцене (а ведь мы заведомо
знаем не всех миоценовых). А среди оставшихся положение многих семейств такое
же, как у некогда процветавших слонов, жирафов и носорогов. Так что Красные
книги лишь отмечают современное состояние процесса, начавшегося для млекопитающих
по крайней мере в плейстоцене, если не раньше. И, вероятно, это звериное отступление
нельзя списать на климатические перемены в ледниковые эпохи последних 60 млн
лет (это ведь не первые глобальные перестройки климата на их веку).
Опять частный вопрос о расселении ушастых тюленей оказывается пробным камнем
наших знаний, на этот раз представлений о глобальных перестройках климатов
и животного мира в преддверии наших дней, в плейстоцене, и степени взаимосвязанности
этих событий.
Поскольку на расселение отариид с несомненностью влияют климат, распределение
и продуктивность пастбищ и давление, оказываемое человеком, а прямых свидетельств
плейстоценового ареала ушастых тюленей вне сегодняшних границ не найдено, объяснить
это равновероятно могут по крайней мере три модели.
I. Существуют незамечаемые нами, но вполне отчетливые для каждой конкретной
формы ушастых тюленей барьеры, надежно удерживающие зверей в их сегодняшних
границах. Эти границы соответственно должны чувствительно реагировать на изменения
климатов, меняясь вслед за ними. Следовательно, отсутствие отариид, например
в Северной Атлантике (где найдены обильные ископаемые настоящие тюлени), однозначно
показывает, что плиоценовый проход на месте сегодняшнего Панамского перешейка
между Атлантикой и Пацификой и сегодняшний экваториальный барьер, отделяющий
северную Атлантику от южной, были всегда непроходимы для ушастых тюленей. Так
же непроходим был еще более надежный барьер в Индийском океане. Тихоокеанский
барьер вел себя в рамках этой модели более странным образом, оставаясь непроницаемым
в западной Пацифике и приоткрываясь в восточной, образуя проход между Галапагоссами
и Калифорнией, пустивший южных морских котов и калифорнийских морских львов
(не смогших с Галапагосс попасть к перуанским берегам) и не пустивший северных
морских котов, сивучей и южноамериканских морских львов.
То есть плейстоценовые похолодания практически не затрагивали экваториальной
зоны ни одного из трех океанов, кроме локальных и краткосрочных событий у берегов
Центральной Америки. Влияние человеческого промысла сказывалось только на численности
отдельных видов ушастых тюленей, не меняя существенно границ их расселения,
так как даже интенсивный европейский промысел XVIII и XIX вв. как будто достоверно
не уничтожил ни одной из самостоятельных форм ушастых тюленей, а очищенные
промыслом архипелаги вновь заселялись через несколько десятилетий после прекращения
промысла из сохранившихся в пределах ареала очажков. Кроме того, даже в прошлом
веке промысловики, поселившиеся на Курилах, долго не находили котовых лежбищ
на островках в центре островной дуги, т. е. и технические возможности прошлого
века не позволяли даже в давно осваиваемой области полностью взять наличное
поголовье промыслового вида (в данном случае северных морских котов). При этом
менее оснащенный и интенсивный доевропейский промысел можно просто не принимать
в расчет.
Получается экологическая ниша семейства чуть больше — на Северную Атлантику
и узкие полосы вблизи 30-х параллелей — реализуемого сегодня ареала, а история
последних миллионов лет — череда пульсаций границ всех трех вложенных друг
в друга ареалов (потенциальный, реализуемый полный и реализуемый репродуктивный)
вблизи современных пределов. Повреждение структуры стад не сказывалось на структуре
ареалов:
В пользу этой модели свидетельствуют неспособность калифорнийских морских львов
галапагосского стада колонизовать перуанское побережье и еще более показательное
отсутствие фолклендских морских львов на Южной Георгии при ее первых посещениях
мореплавателями, дальние заходы южноафриканских морских котов вдоль западного
побережья Африки при полном отсутствии их наблюдений к востоку от 30° в. д.
и прочие свидетельства стабильности сегодняшних ареалов.
Против — идентичность привязанных к шельфам калифорнийцев из Калифорнии и Японии
и морских котов из Африки и Австралии.
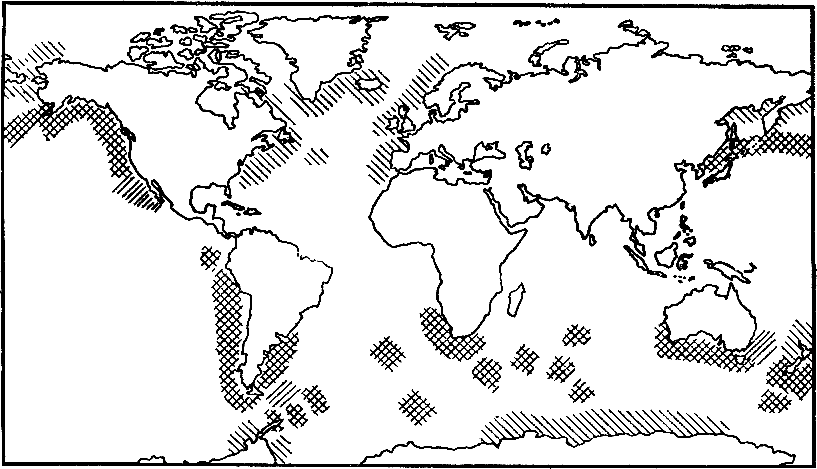
Рис. 26. Сравнение распределения сегодняшних береговых лежбищ ушастых тюленей (штриховка вправо) и сегодняшних гнездовых колоний морских птиц (чистиковых птиц северного и пингвинов южного полушарий) (штриховка влево)
Совпадение этих двух групп колоний (звериных и птичьих) дает крестообразную штриховку. Отчетлива большая терпимость рыбоядных птиц ко льдам и достаточность ресурсов Северной Атлантики
За говорит надежность климатических барьеров, угрожающих риском теплового
шока на берегу и жестко сковывающих расселение холоднокровных кормовых объектов
в море (а через них и едоков), и наличие аргументированной климатической модели,
полагающей сохранение экваториального океанского барьера в плейстоцене. За
говорит и хорошее совпадение ареалов отариид с ареалами пингвинов в Южном полушарии
и чистиковых птиц вместе с серыми китами в северной Пацифике (см. рис. 26).
Против — сегодняшнее обитание чистиковых птиц и вчерашнее (по крайней мере
до XVII в.) обитание изолированного стада серых китов в Северной Атлантике.
Не заходящие в тропики и привязанные к мелководьям серые киты как-то преодолели
сдерживающие их сегодня барьеры. Против — и реконструкции плейстоценовых внедрений
холодных вод в тропики.
Наконец, основной довод в пользу рассматриваемой модели — отсутствие встреч
отариид вдали от современных береговых лежбищ (северного морского кота можно
бы посчитать исключением) — требует наличия специальных механизмов блокировки
расселения в структуре стада. Он дискредитируется несомненным присутствием
ушастых тюленей в пределах значительной акватории Ледовитого океана вопреки
полному молчанию об этом первопроходцев, топонимики, археологии и палеонтологии,
при крупных (сравнительно) размерах ближайших отариидных стад, небольших размерах
акватории и довольно большом числе наблюдателей (по сравнению, конечно, с огромными
пространствами, свободными ото льда в трех остальных океанах). При всех трудностях,
оставляемых в стороне этой моделью, ее кажущаяся очевидность обеспечивает ей
сегодня наибольшую популярность.
II. Отсутствие ясности в понимании механизмов, способных исключить расселение
ушастых тюленей в многочисленные пустующие сейчас области, и возможность компенсировать
поведенческими уловками неудобства на берегу и в океане позволяют допустить
проницаемость экваториальной зоны для всех или, по крайней мере некоторых,
членов семейства. Тогда потенциальный ареал захватывает всю акваторию трех
океанов от одной полярной шапки до другой, а реализуемый расползается на всю
ширь потенциального, следуя за его колебаниями при изменениях климата. То есть
калифорнийским морским львам издавна был открыт доступ в южную Пафицику, южноафриканским
морским котам и австралийским морским львам — в северную часть Индийского океана,
а Северная Атлантика должна была принять морских котов и по обоим своим берегам,
и по островам Срединно-Атлантического хребта да еще южноамериканских морских
львов в придачу (а может быть, и калифорнийских морских львов — через плиоценовый
пролив между Америками). Плейстоценовые похолодания, какими бы они ни были,
могли лишь облегчить трансэкваториальные связи.
С другой стороны, образование компактных и жестко структурированных береговых
поселений, ежегодно скучивающих зверей в одном месте, даже при сильных неблагоприятных
воздействиях делает членов семейства, одних больше, других меньше, но всех
уязвимыми к перегреву на берегу, ухудшению кормовых условий в районе лежбища
и появлению людей, способных достигнуть океанских островов. Соответственно
наиболее уязвимыми оказываются наиболее жаркие и заселенные людьми острова
Океании и Индийского океана, а в послеколумбовые времена — и острова между
Старым и Новым Светом. Наименее уязвимы осваиваемые лишь с конца XVIII в. острова
Субарктики (недаром крупнейшие стада северных морских котов оказались на необжитых
в момент открытия Командорских и Прибыловых островах) и Субантарктики, где
периферийные для зоны береговых лежбищ условия сочетают высокую кормность акваторий
с сезонными переменами благоприятности лежбищ и риском прихода непредвиденной
обитателями катастрофы (блокирование лежбища плавучими льдами, мор массового
кормового объекта).
Возможность пребывания сегодняшних ушастых тюленей в более теплых местах показывает
их благоденствие (сохранение хорошей спортивной формы) в тропических и субтропических
океанариумах и то, что приуроченность их в океане к холодным фронтам можно
списать не на предпочтение холодной воды, а на предпочтение привязанных к этой
холодной воде кормовых объектов. Решили бы вопрос удачные опыты по разведению,
скажем, калифорнийского морского льва в тропических водах или специальные исследования
физиологической и поведенческой тепловыносливости. Пока этого нет, остаются
сильные доводы против (легкость доведения ушастых тюленей до теплового шока
и отсутствие заходов в тропики сейчас, в противоположность редко, но встречаемым
морским леопардам, слонам и монахам).
Доводом против доступности сегодняшних тропиков остаются климатические карты,
где галапагосский район — еще не экваториальная зона. Доводом за служит схема
районирования растительности земного шара, построенная С. М. Разумовским. Полагая
растительность более тонким индикатором климатической ситуации, чем расселение
теплокровных отариид, и видя освоенность ушастыми тюленями только части единого
ботанического района, можно полагать пригодность и прочих его частей, а тогда
от экваториальных барьеров остается немного.
Вторая половина модели, деформация ареалов промысловых видов промыслом, более
бесспорна. Опыт зверобойного промысла показывает, что дюжина человек с крепкими
палками за несколько приемов выбивает все население крупного лежбища (см. рис.
25). А дальше все зависит от доступности лежбища и способа хозяйствования:
либо судьба япономорского стада калифорнийских морских львов, гибель которого
сократила видовой ареал сразу на треть, либо судьба калифорнийского стада морских
котов, ожившего через полтора века после уничтожения за счет сохранившегося
на противоположном конце ареала источника реколонизации.
Насколько плотно осваивали острова мореплаватели каменного века, показывают
открытия многочисленных эндемиков среди птиц Гавайских островов и Новой Зеландии,
переживших все плейстоценовые перемены, но сметенных полинезийской колонизацией.
Насколько эффективно влияли на географию животного населения еще не слишком
технически оснащенные (парусные суда и ручной гарпун) люди железного века,
показывает общеизвестный пример с локальным стадом командорских сирен, потеря
которого превратила всю группу из широко распространенной в ограниченную только
тропиками, так же известен пример с истреблением (полным за несколько веков)
широко распространенной по северной Атлантике еще во времена Колумба бескрылой
(но прекрасно плававшей по океану) гагарки и менее известный пример полного
уничтожения к концу XVII в. североатлантического стада серых китов, что сократило
видовой ареал его с двух океанов до одного. Причем коров успешно добывали простейшим
гарпуном предельно ослабленные цингой люди, гагарку истребили с помощью палки
и сбора яиц, а серых китов добили гарпунщики на весельных ботах.
Эти бесспорные примеры дают настолько простую и наглядную связь между ходом
расселения людей с экипировкой еще охотников каменного века и гибелью многочисленных
форм крупных птиц и млекопитающих, что трудно удержаться от экстраполяции этих
событий и на плейстоценовое вымирание крупных млекопитающих. Попытки оценить
опустошения, причиненные появлением на сцене и перемещениями по ней плейстоценовых
охотников на хоботных и копытных, делались И. Г. Пидопличкой для Евразии, а
позже оформились в гипотезу перепромысла П. Мартина. Получается достаточно
складная картина единого плейстоценово-голоценового процесса изымания набирающим
силу человечеством одного промыслового (или косвенно связанного с хозяйственной
активностью человека) вида за другим из богатого, доставшегося ему в наследство
миоценово-плиоценового мира. В рамках этого общего процесса рассматриваемая
модель (отступание отариид из осваиваемых человеком районов с сохранением к
нашим дням лишь на окраинах прежнего ареала) выглядит частными, несколько запоздавшими,
но хорошо укладывающимися в общие рамки событиями. Европейские же зверобойные
шхуны лишь завершали и чуть было не завершили процесс, начатый палеолитическими
ордами, но тут начали действовать новые силы, пытающиеся упорядочить использование
ресурсов оказавшейся вдруг такой маленькой и уязвимой Земли.
Слабые места модели перепромысла (или промысла на износ) так же очевидны, как
и сильные. Ход колонизации побережий континентов, а тем более островов, археология
представляет не вполне отчетливо. Свидетельства плейстоценовых взаимоотношений
с ластоногими если и сохранились, то у тогдашней береговой линии, оказавшейся
сейчас под стометровым слоем воды. Исследования демографии и взаимоотношения
с ресурсами наших палеолитических предков только начинается, и опереться здесь
почти не на что. Исчезновение после, казалось бы, ничем не омраченного расцвета
многочисленных семейств млекопитающих (не говоря уже о гигантских рептилиях)
началось задолго до выхода человека, и раз происходило без его участия прежде,
то и по новым обвинениям надо его судить осторожно.
Наконец, гибель от преследования человеком, коснувшись одних, надолго миновала
других. Стеллерова корова не протянула и полувека после открытия, дюгони (то
же семейство) служили регулярной добычей жителей давно обжитого Персидского
залива еще во времена Гильгамеша и дожили в малом числе почти до наших дней.
Сохранение курильских лежбищ северных морских котов до конца прошлого века,
несмотря на близость человеческих поселений и активный промысел, говорит о
мозаичности использования угодий, дающей ресурсам шанс оправиться. На заселенном
сразу после открытия в 1502 г. острове Св. Елены какие-то «морские коровы»
протянули до начала прошлого века, а на Гавайских островах с четвертью миллиона
жителей (перед появлением Кука) погибли бескрылые гуси и ибисы, но сохранились
до наших дней тюлени-монахи.
III. Компромиссный вариант модели, исходя из наличия сегодняшних экваториальных
барьеров и барьеров, привязывающих к окрестностям лежбища, вместе с наличием
плейстоценовых перемен в уровне океана и распределении климатов, понуждающих
отариид к перемещениям со старых лежбищ и открывающих на время какое-то число
ворот в экваториальных барьерах, должен также с неизбежностью рисовать картину
расселения ушастых тюленей, отличную от сегодняшнего. В пределах модели возможны
варианты, приближающие ее либо к модели I, либо к модели II, но в любом случае
остается взаимодействие идущих, помимо человека, событий и человеческого вклада.
После миоценовых перетрясок и выхода последних, еще не известных по предыдущим
листам геологической летописи семейств млекопитающих из небытия наступает,
по-видимому, какая-то стабилизация в распределении ролей в системе и мест на
поверхности земного шара. Климатическая ситуация и уровень океана, вероятно,
сравнимы с современными. Отарииды, по крайней мере в Пацифике, преодолели экватор
(есть находки в Перу с датировкой 5 млн лет). Если они заселяли и берега Антарктиды,
на них должно было произвести впечатление и сдвинуть с насиженных мест образование
сплошного антарктического панциря 5—7 млн лет назад. Если же не заселяли, то
непонятно, что отгораживало Антарктику от побережья нынешнего Перу. Сейчас
из отложений этого времени известны богатые захоронения морских зверей (кроме
калифорнийских находок) из Южной Африки, Европы и атлантического побережья
Северной Америки. Но в них во всех при большом разнообразии сирен, китообразных
и настоящих тюленей нет никаких следов ушастых тюленей, что можно либо списать
на неполноту летописи, либо на неизвестные барьеры, которые, соответственно,
надо изобретать.
В то же время правдоподобной представляется картина широкого расселения тех
групп млекопитающих, которые сегодня рассматриваются как приуроченные самой
своей конструкцией к одной климатической зоне (так, носороги, сирены, хоботные,
крупные пантеры, быки широко расселены вне сегодняшних ареалов соответствующих
групп) с одновременным существованием высоко эндемичных островных фаун, замкнутых
территорией своего островка или архипелага и давших миру поразительных и разнообразных
нелетающих птиц (бескрылая гагарка и разнообразные пастушки атлантических островов,
дронты и эпиорнисы островов Индийского океана; нелетающие гуси, бакланы, ибисы
и нелетающие птицы Новой Зеландии в Тихом океане).
Небезразличные для судьбы ушастых тюленей Южная Америка, Антарктида (пока ее
совсем не прикрыла ледяная шапка) и Австралия дают своим обитателям больше
свободы, чем океанские острова, но меньше, чем громадные полигоны материков
Северного полушария. Кстати, одно время всплывала идея определить отариид в
уроженцы Антарктиды, но потом заглохла из-за отсутствия надежных доводов.
Третий независимый мир, заполняющий пространство между большими и маленькими
кусочками суши и населенный не вовлеченными в сухопутные проблемы китообразными,
как будто все-таки достаточно един (родня белух и нарвалов — у центральноамериканских
берегов, а родня лаплатских речных дельфинов — у калифорнийских) последние
10 миллионов лет, судя по анализу Л. Барнса наличных находок.
Получается, что роль связок между независимыми ойкуменами приходится на летающих
птиц, морских черепах да ластоногих. К этой же роли связки отдельных частей
воедино история подводит, наконец, и одну из форм гоминид, хотя когда именно
человек получил возможность преодолевать океанские преграды, не вполне ясно.
Пока гоминиды копят силы в Афро-Евразии, плиоценовым сообществам надо тесниться
под напором климатических встрясок, делающих положение вблизи полярных шапок
неустойчивым и экстремальным, а экватор преодолимым и для тех, кому он раньше
мешал в расселении, т. е. неизбежно усиление обмена между и теплокровными обитателями
океанов Северного и Южного полушарий и проникновение каких-то отариид в экваториальную
зону с выходом в северные части Индийского и Атлантического океанов (если их
там не было прежде).
С концом ближнего к нам ледниковья, поднятием уровня океана на десятки метров
и освобождением из подо льда новых земель и акваторий следы старых лежбищ уходят
глубоко под воду. Их обитатели переходят на новые места, а популяции, слишком
близкие к экватору, оказываются в положении заметно ухудшенном невыносимым
климатом или бескормицей. Единый до этого ареал семейства около десяти тысяч
лет назад должен был распасться на сплошной ареал Южного полушария и оторванные
от него куски в северной Атлантике, северной части Индийского океана и северной
Пацифике. В тропиках и субтропиках еще долго могли сохраняться небольшие, изолированные
друг от друга малыми размерами и большей оседлостью стада.
Вот тут-то и должно было вступить начатое еще людьми каменного века освоение
все новых и новых побережий на континентах и на все более и более удаленных
островах. Там люди сталкивались с остатками прежних фаун, которые должны были
казаться пришельцам сказочно изобильными. Острова северной Пацифики населяли
морские выдры, ушастые тюлени, нелетающие птицы разных групп, а у берегов плавали
морские коровы. Вероятно, столь же богаты были острова Северной Атлантики и
океанов Южного полушария. Для попавших на остров людей это была легкая добыча
и незаменимое подспорье на первых, самых тяжелых фазах колонизации. Первыми
должны были лишиться своих лежбищ побережья континентов, а затем и острова.
К началу европейских великих географических открытий острова северной части
Индийского океана (включая Мадагаскар), Полинезии, Японии и Филиппин были очищены
и заселены. Возможно, последние отарииды Северной Атлантики дожили до прихода
испанцев и португальцев на Канарские и Азорские острова, где и пошли на шкуры,
смешавшись под общим названием «морских псов» с последними тюленями-монахами.
Еще больше ужали ареал семейства промышленники последних веков, планомерно
очищавшие остров за островом и донесшие до нас смутные рассказы о зверях на
островах Св. Елены, Реюнион и Клиппертон. Но это было лишь продолжение задолго
до них начатого дела, приведшего к сохранению отариид лишь в краевых частях
ареала. Так же лишь в нескольких краевых участках оставались доведенные до
грани уничтожения остатки ареалов зубра или бамбукового медведя.
Деструкция касалась не только отдельных видов, но и их комплексов, поэтому
сегодняшнее совпадение границ распространения береговых лежбищ отариид и гнездовых
колоний пингвинов и чистиковых птиц (см. рис. 26) надежнее связывать не с одними
лишь климатическими границами или определяемыми ими деталями распределения
кормовых объектов, а и с прессом человеческого населения.
Аргументированность всех трех рассмотренных моделей плейстоценово-голоценовой
истории береговых поселений ушастых тюленей примерно одинакова, пока новые
находки не увеличат шансы какой-либо из них или не приведут к построению новой
модели.
Проекция многомерной экологической ниши на поверхность планеты, потенциальный
ареал, может Сжиматься от наступающих льдов, усиливающейся жары или терять
куски, отхваченные человеческой экспансией. Наполняющее потенциальный ареал
живое содержимое может вести себя по-разному: с разной скоростью заполнять
пустоты в потенциальном ареале ареалом реализуемым и с неодинаковой скоростью
восполнять утраты в разных участках единого видового ареала. При этом наиболее
значимыми для понимания происходящего с ареалом любого таксона будут события
в краевой зоне реализуемого в данный момент ареала.
Варианты поведения реализуемого ареала (характер блочности целого и возможности
роста) так же не бесконечны, как и варианты потенциального ареала (характер
расчлененности барьерами и степень стабильности) , и, по-видимому, не очень
большой цифрой будет число моделей, надежно описывающих систему ареал—экологическая
ниша. Так что при некотором минимуме наблюдений над размещением особей и их
группировок в пределах сегодняшнего ареала, скажем, того или иного ушастого
тюленя, можно было бы вполне надежно отвечать не только на вопрос, где живет
данный вид, но и где он жил вчера и, что важнее, где для него останется место
завтра.
В этом отношении отарииды изучены не хуже других промысловых видов и могут
служить хорошим индикатором приближения того времени, когда события с видовыми
ареалами будут прогнозироваться так же, как сейчас прогнозируются метеорологами
перемещения холодных и теплых масс воздуха в атмосфере.
Пока накапливаются необходимые для прогнозов наблюдения, остается сделать еще
один шаг в иерархии систем и перейти от видового ареала с вмещающей нишей к
системе, объединяющей отдельные виды организмов воедино, и попытаться оценить
место отариид в ней.
Если каждый из нескольких десятков видов ушастых тюленей, прошедших, сменяя
друг друга, или, вернее, проплывших по поверхности нашей планеты с миоцена
до сегодняшнего дня, занимал свое неповторимое место в системе связей между
видами, то как отражалось на системе его исчезновение и как вписывался в систему
новый вид? Где была видовая экологическая ниша сивуча до его выхода на арену,
и что случилось с видовой экологической нишей миоценовой имаготарии после гибели
последнего зверя этого вида? Наконец, что происходит с надвидовой системой
с утратой того же сивуча или даже всего семейства отариид (утрата дюжины видов
близких родственников из нескольких миллионов наличных): оказывается ли эта
утрата незаметной, или она повреждает только локус системы, или она служит
началом длинной и все более ветвящейся цепи сопряженных событий?
Эксперименты по изъятию вида из системы, правда, с сохранением некоторого количества
старых связей, и перевода его в статус домашнего и полудомашнего проводятся
человеком с разной степенью обдуманности уже не одно тысячелетие и накопили
достаточно большой материал по сопровождающим этот процесс, обычно достаточно
постепенный, деструктивным изменениям морфологических, поведенческих и социальных
структур. Много материала и по обратным экспериментам: введению в надвидовую
систему одичалых домашних зверей или специально выбранных для завоза и акклиматизации
видов. Здесь одновременно с нормализацией внутривидовых структур вселенца деструкции
подвергается сама система (опустошения от одичалых собак, коз и кроликов хорошо
известны).
Изъятие нескольких компонентов системы, например замена диких копытных в саваннах
стадами одного-двух видов домашнего скота, и последующие экологические катастрофы
хорошо смоделированы всей практикой монокультур в животноводстве и земледелии,
показывая рост риска деструкции системы при уменьшении числа составляющих ее
компонентов.
Получилась ситуация, когда и общие проблемы ведения человеческого хозяйства
на планете, и попытки понять происходящее с маленькой группой ушастых тюленей
упираются в необходимость исследовании структуры биосферы, с тесным переплетением
и взаимопроникновением результатов рассмотрения конкретных проблем.
Известное об отариидах достаточно надежно укладывается в схему, где каждый
отдельный вид является узлом, соединяющим воедино три независимые системы:
систему дискретного таксономического разнообразия; систему дискретных и одновременно
сопряженных друг с другом своими пространственными структурами видовых ареалов;
систему функциональных связей между видами.
Появление нового вида, так же как и исчезновение старого, невозможно без сопряженных
перестроек во всех трех системах. Хотя неизвестно, насколько каждая из систем
способна предотвратить расползание локальных событий на всю конструкцию в целом,
можно говорить о плейстоценовом и продолжающем его голоценовом вымирании млекопитающих
как об утратах разнообразия, дестабилизирующих все три сопряженные системы.
При этом независимо от причин исчезновения конкретного вида зверей разрушение
проходит ряд фаз, в которых вмешательство человека может либо сделать происходящее
необратимым еще скорее, либо затормозить или повернуть вспять.
В первой фазе происходит либо запустевание видовой экологической ниши с разрушением
тканы видового ареала, либо разрушение самой ниши, либо оба процесса одновременно.
Следующая фаза разрушает уже не просто куски ареала, а внутривидовые связи,
формируя различные патологические состояния оставшегося ареала или его частей.
При этом состоящее из внешне здоровых зверей стадо оказывается неспособным
найти оптимальную стратегию использования ресурсов (кормов, убежищ), неспособным
быстро восстанавливать утраченные после неблагоприятных воздействий поселения,
неспособным обеспечить оптимальный режим воспроизводства и возрастной смены
ролей отдельной особи.
При увеличении силы или длительности давления на вид (или того и другого вместе)
наступает третья фаза, когда распавшийся на очажки ареал теряет их один за
другим, лишаясь даже при благоприятном раскладе дальнейших событий возможности
вернуть свое недавнее место полноценно работающего компонента биосферы. А поскольку
биосфера не только наш дом, но и машина, достаточно давно производящая для
нас еду, питьевую воду и кислород, то, скажем, съеденная морским львом рыба
не просто пропала с нашего стола, а обеспечила работу одной из деталей системы
нашего жизнеобеспечения.
Применительно к биосфере риск потери вида для работы системы несколько выше,
чем риск испортить телевизор или сходный по сложности аппарат, выдернув из
него деталь наугад. Разумнее было бы не допускать утери видов из биосферы,
а уже выпавшие оттуда, но сохранившиеся, скажем в зоопарке Дж. Даррелла, вернуть
обратно, не забывая одновременно собирать необходимый человеку урожай с биосферы,
а не только с десятка видов взятых в культуру животных. Но чтобы снимать урожай
не годится ни музейный экспонат, сохраняющий только анатомию или детали анатомии
особей, ни обитатели зверинца, сохраняющие, в лучшем случае, кроме анатомии
и физиологии, еще и крошечную выборку из бывших в распоряжении вида геномов,
ни заповедник с куском более или менее поврежденной ткани вида. Лишь видовой
ареал, соизмеримый с потенциально возможным, определяемым видовой экологической
нишей, дает полноценно работающий компонент биосферы.
Этот компонент можно использовать по-разному: либо собирать без разбора то,
что дает какая-то территория, рискуя потерять кусок видового ареала (первая
фаза деструкции); либо проводить выборочный выбой отдельных возрастных или
половых групп, рискуя получить патологические изменения внутривидовых структур
на данной территории (вторая фаза деструкции); либо взять вид целиком в культуру,
для чего надо донести до последней фазы деструкции многочисленные сопряженные
с ним виды, попадающие при этом в категорию «вредных», но и для оберегаемого
вида та или иная степень патологии внутривидовых структур (т. е. вторая фаза
деструкции) неизбежна.
Эта неопределенность хорошо отражает сегодняшние проблемы распутья, на которое
вывела человека эволюция его взаимоотношений с биосферой. Нечто подобное, по-видимому,
происходило во времена так называемой неолитической революции, когда человек
стал ориентироваться на культивирование немногих перспективных с его точки
зрения видов вместо прежнего использования рассредоточенных, разнообразных,
но ограниченных ресурсов (еще недавно жители Алеутских островов использовали
в качестве пищевых объектов несколько сотен видов животных и растений) . При
этом громадное большинство видов неизбежно попадает в число жизненно не необходимых
и соответственно не заслуживающих какого-либо оберегания или в число мешающих
культивированию жизненно необходимых немногих видов-избранников и подлежащих
соответственно изъятию из обживаемых человеком мест.
Наконец, число не заселенных человеком мест сократилось до некоторого количества
каменистых, дремучелесных и ледяных пустынь, ожидающих своего освоения со дня
на день. Уже сегодняшнее население (человеческое) земного шара дает человек
тридцать на квадратный километр суши (или десять, если считать и акваторию
океанов). Глобальной стала проблема, с которой прежде закономерно сталкивались
жители любого острова через некоторое время после его колонизации (опять взаимоотношения
человека с отариидами хорошо моделируют общечеловеческие проблемы): наличные
ресурсы не есть манна небесная, подаваемая в нужном количестве в нужное время.
С выходом в околоземное пространство человек приобрел более реалистичный взгляд:
планета стала представляться не очень крупным космическим кораблем с ограниченными
ресурсами и необходимостью считаться с возможностями систем жизнеобеспечения.
Нужно выбирать долговременную стратегию, а вариантов, как всегда, три (как
издавна известно из похождений героев сказок).
1. Либо посчитать роль биосферы выполненной, как исполнившей свое предназначение
плаценты, и заменить ее некоторым подобием искусственной системы жизнеобеспечения
космического корабля, легко управляемой и с предельно упрощенным разнообразием
используемых компонентов, т. е. идти путем людей, перешедших от охоты к животноводству,
а потом и к промышленным методам производства пищевых объектов.
2. Либо, напротив, совершенствуя технологии и локализуя производства в местах
наименьшего контакта с природными комплексами, превратить всю очищенную от
загрязнений и реставрированную от повреждений биосферу в заповедник или национальный
парк.
3. Либо найти способ снимать урожай со всего наличного ассортимента видов,
не повреждая при этом ни деталей системы, ни связей между ними и максимально
используя при этом как получаемое сырье, так и работу системы из одного-двух
миллионов видов организмов, т. е. попробовать стратегию альтернативную к выбранной
несколько тысячелетий назад.
Возвращаясь теперь к началу рассказа и вопросу о том, может ли разговор о
проблемах изучения строения, биологии и расселения небольшой группы связанных
очевидным родством зверей — семейства ушастых тюленей — иметь интерес для не
имеющих с этой группой дела людей, я бы рискнул ответить на этот вопрос утвердительно.
Во-первых, довольно наглядно очерчивается соотношение известного и неизвестного
в обследованиях давно известных групп животного мира. То, что в данном примере
прорехи в знаниях связаны с распределением особей в море, степенью близости
отдельных видов и популяций в пределах каждого вида, плейстоценово-голоценовой
истории ареалов, приуроченностью палеонтологических находок лишь к двум-трем
районам, физиологией зверя в экстремальных условиях и изменчивостью (индивидуальной
и групповой) анатомических, физиологических и поведенческих характеристик,
представляется довольно общим положением.
Соответственно общей представляется и неизбежная необходимость оперировать
несколькими равновероятными моделями при истолковании почти каждой группы фактов:
недавнее отхождение отариидной ветви от ствола наземных хищных и давнее их
расхождение с последующим параллельным ходом; разные варианты плейстоценовой
истории; большая дробность субтропических частей ареала по сравнению с более
монолитными субполярными и равная степень дискретности разных частей ареала
вида; определение срока задержки имплантации, конца лактации и начала полового
созревания внутренними часами или, напротив, внешними факторами; различные
оценки тепло- и холодоустойчивости на берегу.
Однозначность других построений показывает сходство событий у далеких друг
от друга животных при несходстве близких родственников. Жесткие рамки, сковывающие
изменчивость, и конструктивные варианты, блочно построенные и могущие работать
в режиме минимального обеспечения, укладывают преобразования организма зверя
в рамки системы запретных и дозволенных путей. Сам организм со всеми своими
локальными перестройками вовлекается в иерархию систем следующих порядков.
По общим схемам для животных разных групп из особей строится лежбище, то стягивающее
обитателей к себе, то рассеивающее их по акватории (рис. 27 и 28). В свою очередь,
из этих единиц (лежбище с освоенной акваторией) строится новая система — реализуемый
видовой ареал, а из нескольких миллионов видовых ареалов — система следующего
порядка — биосфера. Так очерчивается место изучения маленькой группы зверей
в деле изучения глобальных событий.
Во-вторых, сам поворот в видении, превращающий вчерашнюю экзотическую, любопытную
(и только) зверушку в элемент сложной жизненно необходимой глобальной системы,
ставит новую общеобразовательную проблему. А в спорах об актуальности тех направлений
исследований и роли таких традиционных наук, как зоология или анатомия, в общем
продвижении системы наших званий вперед достаточно очевидно проступает злободневность
(наряду с астрофизическими и субмолекулярными исследованиями) исследований
структуры биосферы с прозаической инвентаризацией ее быстро исчезающих компонентов.
Рис. 27. Еще раз взгляните на степень скученности ушастых тюленей на береговом лежбище
Каждая из трех групп населения живет своей достаточно отдельной жизнью: слева — территориальный секач придавил к земле нетерриториального секача-вторженца, рядом с ними — сбившиеся на свободном от взрослых месте щенки, на заднем плане — толпа лежащих бок о бок самок
Рис. 28. Контрастом к предыдущему рисунку выглядят те же звери, покинувшие лежбище и рассредоточившиеся но прилежащей акватории
Характерна поза крупного, отдыхающего на тихой воде зверя с поднятыми вверх задними и одним из передних ластов
В-третьих, конкретная история взаимоотношений человека с дюжиной промысловых
видов ушастых тюленей ставит нас перед необходимостью обдуманного выбора долгосрочной
стратегии взаимоотношений с этими и другими компонентами целостной биосферы.
При этом очевидно, что ценность морских львов и котов, помимо разнообразного
традиционного и нетрадиционного сырья (шкура, жир, мясо, витаминное сырье),
состоит и в той работе, которую они выполняют в системе жизни на планете, и
в тех знаниях, которыми они нас снабжают, и в красоте живого и вольного зверя.
И еще несколько слов о прорехах в нашей системе знаний о жизни на Земле. Неожиданно,
на первый взгляд, но неизвестное об ушастых тюленях покрывается одним, казалось
бы, обветшавшим и не привлекающим внимания в наш век словом — анатомия. Только-только
собирается из кусочков нормальная анатомия особи для промысловых зверей. Появляются
лишь первые наметки анатомии поселений. Все еще бесформенным и текучим кажется
видовой ареал, хотя уже давно все попытки прогнозировать происходящее с ареалом
(даже на уровне прогнозов погоды) разбиваются о что-то невидимое, но реально
существующее и диктующее ход событий, разбиваются о жесткие внутривидовые структуры,
об анатомию видового ареала. Еще хуже обстоит дело с знаниями анатомии надвидовых
построек (биогеоценозов, биот и самой биосферы), где после обнадеживающего
начала, положенного работами академика В. Н. Сукачева, наметившиеся анатомические
структуры исчезли в клубах многочисленных информационных и заимствованных из
экономического прогнозирования моделей. Т. е. пока мы радуемся находкам в анатомии
субмолекулярных и внутриклеточных образований, анатомия особи все еще далеко
не ясна, а об анатомии надиндивидуальных образований мы знаем непозволительно
мало.
К счастью, трудно представить, скажем хирурга, при подготовке которого было
бы обойдено изучение нормальной и патологической анатомии особи. Но вот зоологи
как-то обходились до сих пор без знания нормальной анатомии особей большинства
видов, с которыми опи имели дело. И уже нельзя восполнить многое упущенное
в изучении реализованного разнообразия живого. Анатомию вчера еще жившего австралийского
сумчатого волка (единственного дотянувшего до нас крупного сумчатого хищника)
мы так и будем знать хуже, чем анатомию мамонта. До сих пор незнание возможностей
перестроек нормальной индивидуальной анатомии дает пищу произвольным истолкованиям
роли рассматриваемых деталей строения особи, заставляя исследователя удовлетворяться
подобием диалога доверчивой Красной Шапочки и Волка; большие глазки — чтобы
лучше видеть... и т. д.
Незнание анатомии надиндивидуальных образований позволяет нам поступать с ними
вольно. Можно пытаться переселением беременных самок морского кота на необжитый
участок берега создать новое лежбище, можно безгаремных секачей продуцирующего
лежбища объявить излишними и отстрелять. И неудачи обоих экспериментов будут
лишь побуждать хвататься за первое подвернувшееся объяснение (любое, кроме
единственного: надо выявлять анатомию и, выявив, сообразовываться с ней). Можно
любое увеличение численности нужного зверя или появление его в обжитых местах
объявить результатом успехов хозяйствования, а потом ненадолго изумиться разрушениям,
причиняемым деструктурированными стадами лосей пли обживающими с голодухи помойки
полярных поселков белыми медведями. Изумиться лишь для того, чтобы объявить
вчерашних подопечных сегодняшними «вредителями» и начать их отстреливать, так
и не увидев те анатомические структуры видовых ареалов, о которые разбились
наши ожидания.
Только нежеланием увидеть анатомию ценозов и биосферы можно объяснить легкость
в списывании разрушения ценозов на деятельность местных видов — «вредителей»
при одновременном самодовольстве «обогащением» флоры и фауны вселением чужеродных
видов. Легко объявить виновными в нескладывании диалога между биосферой и техносферой
индивидуальных браконьеров, труднее увидеть разрушительные последствия собственного
хозяйствования, но даже увидев тот и другой вред, но не поняв, какие именно
анатомические структуры повреждаются, мы остаемся в положении хирурга, не знающего
анатомии, перед постелью тяжелобольного.
Рис. 29. В зажатой между полярными плейстоценовыми шапками ойкумене складывались начальные фазы диалога современного человека и биосферы

Рис. 30. Становление глобальной техносферы обострило диалог человечества и биосферы
Небрежение анатомией биосферы имеет и другую сторону, поскольку структурированность
биосферы неотделима от структурированности таксономического разнообразия в
населении нашей планеты, представлению об аморфной биосфере соответствует непредсказуемое
и не знающее запретов филогенетическое дерево с размытыми, перетекающими вольно
друг в друга таксонами, где потеря любого таксона не безвозвратна, а легко
восполнима соседями. При таком раскладе действительно временную последовательность
любых похожих форм легко счесть одновременно и чередой генетически преемственных
форм, где соответственно любой таксон — плод творчества систематика, вольно
трансформирующийся в другой такой же плод.
Сумеем ли мы соотнести стратегию нашего поведения с осознанием прорех в системе
знаний, пугающей быстротой накапливаемых в живом мире планеты разрушений и
стремлением сохранить жизненные стандарты? Наш пробный камень — мир ушастых
тюленей, выйдя из передряг ледниковых эпох (рис. 29), оказался зажатым в щелях
техносферы (рис. 30). Оба рисунка, скомпонованные Е. В. Сыроечковским, естественно,
несколько усугубляют ситуацию. Но вот заметка из газеты «Советская Россия»
от 3 июля 1986 г. в разделе любопытных фактов, где корреспондент С. Штейнберг
пишет: «Он появился среди бела дня в самом центре Владивостока — в Спортивной
гавани. Взобрался на одну яхту, полежал, осмотрелся, переполз на другую. Устроившись
поудобнее, довольно рявкнул, к восторгу мальчишек... Что привело в город дальневосточного
тюленя (речь идет о морском льве-сивуче.— Прим. автора)!... Кто следующий наведается
в богатый рыбой залив Петра Великого, где в свое время гостили и летучие рыбки,
и морские черепахи, и змеи, и акулы, и даже киты?»
Подумаем не спеша над заключительным вопросом газетчика и над неизбежно стоящим
рядом вопросом: почему общительному пришельцу (так озаглавлена заметка) не
нашлось места в заливе Петра Великого, где почему-то перестали гостить и летучие
рыбки, и морские черепахи, и даже киты. Вспомним и о лосях, забредших в город,
и о медведях на помойках полярных станций, и о натыкающихся на самолеты птицах,
и о прочих общительных пришельцах, которым не находится места в преобразованном
нами мире.
На этом самое время закончить с надеждой на то, что доброжелательного читателя
этот далекий от совершенства текст может натолкнуть на плодотворные размышления
о темных местах картины, и с благодарностью людям, поощрявшим работу, и в первую
очередь герпетологу Б. Д. Васильеву и орнитологам В. М. Гаврилову и Е. В. Сыроечковскому,
без участия которых она не была бы написана.
Адам-Смит П. Охотники на лунных птиц. М.: Наука, 1983. 184 с.
Берх В. Н. Известие о меховой торговле, производимой россиянами, при островах
Курильских, Алеутских и на северо-западном берегу Америки // Сын отечества,
1823. Книги 39, 43 и 44.
Бляхер Л. Я. Проблемы морфологии животных. М.: Наука, 1977. 360с.
Боумен У. Графическое представление информации. М.: Мир. 1971. 225 с.
Бычков В. А. Охрана и рациональное использование ластоногих. М.: ВНИИТЭИСХ,
1976. 62 с.
Ваксель С. Вторая Камчатская экспедиция В. Беринга. М.; Л.: Главсевморпуть,
1940.
Вениаминов И. А. Записки об островах Уналашкинского отдела. СПб, 1840.
Вернадский В. И. Живое вещество. М.: Наука, 1978. 358 с.
Дуброво И. А. Новое подсемейство ископаемых тюленей//ДАН СССР. 1981. Т. 256.
№ 4. 970—974 с.
Канаев И. И. Фрэнсис Гальтон. Л.: Наука, 1972. 133 с.
Карус К. Г. Основания краниологии. СПб, 1844.
Крушинская Н. Л., Лисицина Т. Ю. Поведение морских млекопитающих.
М.: Наука, 1983. 336 с.
Меркаторская карта пяти частей света, изображающая: ход, временное и постоянное
пребывание различного рода китов, с показанием удобнейшего времени года для
их ловли. Изд. Российско-Американскою компанией по новейшим сведениям. 1852.
Млекопитающие Советского Союза/Под ред. В. Г. Гептнера. М.: Высш. шк. 1976.
Т. 2. Ч. 3. 718 с.
Мужчинкин В. Ф. Пространственное распределение особей и индивидуальное пространство
животных. Биогеография /Итоги науки/. М.: ВИНИТИ АН СССР, 1976. Вып. 1. С.
219— 247.
Мужчинкин В. Ф. О береговых лежбищах ушастых тюленей // Экология, 1977. № 1.
С. 83—91.
Мужчинкин В. Ф. Блочность конструкции млекопитающих па примере семейства ушастых
тюленей // Журн. общей биологии. 1978. Т. 39. № 5. С. 777—782.
Разумовский С. М. Ботанико-географическое районирование Земли как предпосылка
успешной интродукции растений // Интродукция тропических и субтропических растений.
М.: Наука, 1980. 318 с.
Соколов В. Е. Редкие и исчезающие животные; Млекопитающие. М.: Высш. шк. 1986.
519 с.
Федорова С. Г. Русское население Аляски и Калифорнии. М.: Наука. 1971. 275
с.
Хенниг Р. Неведомые земли. М.: Иностр. лит-ра, 1961—1963. Т. 1. 515 с.; Т.
2. 518 с.; Т. 3. 471 с.; Т. 4. 547 с.
Хлебников К. Т. Русская Америка в неопубликованных записках К. Т. Хлебникова.
Л.; М.: Наука, 1979. Т. 1, 279 с; 1984. Т. 2. 302 с.
Шелихов Г. И. Российского купца Гр. Шелихова странствования из Охотска по Восточному
океану к Американским берегам. Хабаровск, 1971. 176 с.
Antarctic Pinnipedia/Burt W. (ed.) //Antarctic Research Ser., 1971. Vol. 18.
Barnes L., Damning D., Ray C. Status of studies on fossil marine mammals //
Marine Mammals Science. 1985. Vol. 1 (1). P. 15—53.
Bering Sea Biology. An evaluation of the enviromental data base related to
Bering sea oil and gas exploration and development/Lewbel G. ed. Sohio, Anchorage,
1983. 180 p.
Banner N. The fur seal of South Georgia // Sci. Rep. British Antarctic Survey.
1968, Vol. 56. P. 1—81.
CLIMAP. The Surface of Ice-age Earth//Science. 1976. Vol. 191. N 4232. P. 1131—1137.
Craig A. Histology of reproduction and the astrus cycle in the female fur seal
// Fishery Res. Board of Canada. 1964. Vol. 21. N 4. P. 773—811.
Fetcher E. The water balance in marine mammals // Quart. Review of Biol. 1939.
Vol. 14. N 4. P. 451—453.
Flenley J., King S. Late quaternary pollen records from Easter island // Nature.
1984. Vol. 307. P. 47—49.
Hamilton J. The Southern Sea Lion//Discovery Report, 1934—1939. 1934. Vol.
8. N 2. P. 269—318; 1939. Vol. 19. N 2. P. 121—164.
Harrison R., Tomlinson D. Observations on diving seals and certain other animals
// Symp. Zool. Soc. L. 1964. Vol. 13. P. 59—70.
Howell A. Contribution on the comparative anatomy of the eared and earless
seals//Proc. U. S. Mat. Museum. 1929. Vol. 73. N 15. P. 1—142.
Irving L., Peyton L, Bahn C. Regulation of temperature in fur seal//Physiol.
Zoology. 1962. Vol. 35. N 4. P. 275—284.
King J. Seals of the world. L. 1964.
Mammals in the seas. FAO Fisheries series. Rome. 1978. Vol. 5. N 1. P. 1—268.
Quarternary Extinction (a prehistoric Revolution)/Martin P., Klein eds. Tacspn:
Univ. of Arizona Press, 1984. P. 1—896.
Ray C. Locomotion in the pinniped//Natural History. 1963. Vol. 72. N 3. P.
11—21.
Reppening C., Tedford R. Otarioid seals of the Neogene // U. S. Geol. Survey
Prof. 1977. Paper 992.
Sachet M.-H. Monographic physique et biologique de l’ile Clipperton // Annales
de l’institut Oceanografique. 1962. T. 40. P. 1— 108.
Sarich V. Pinniped origin and the rate of evolution of the carnivore albumins
// Systematic Zoology. 1969. Vol. 18. N 3. P. 286—295.
Scheffer V. Pelage and surface topography of the northern fur seal. Washington,
1962.
Seals//IUCN Publications New Series. 1973. Suppl. Paper 39.
Stelneger L. How the great northern sea cow became exterminated //Amer. Naturalist.
1888. Vol. 21. P. 1047—1054.
Stumpke H. Bau und Leben der Rhinogradentien. Stuttgart, 1962. S. 1—82.
Taylor J., Braithwaite C., Peake J., Arnold E. Terrestrial faunas and habitats
of Aldabra during late Pleistocene // Philosoph. Transac. Roy. Soc. London
(B). 1979. Vol. 286. N. 1011. P. 17— 66.
Thompson D’Arcy. On growth and form. 1917. Cambridge, 1917.
Watanabe H. The Ainu Ecosystem: Enviroment and group Structure. Tokio,
1972.
| Чем интересен ушастый тюлень ........................................... |
3 |
| Как складывалось знакомство с ушастыми тюленями ....... | 4 |
| Строение тела ушастого тюленя и освоение океана ........... | 14 |
| Составные портреты Фрэнсиса Гальтона и возможность увидеть размах изменчивости .............................................. |
29 |
| Когда появились на сцене ушастые тюлени ........................ |
63 |
| Как живет ушастый тюлень .................................................. |
71 |
| Как из отдельных особей строится скопление .................... |
79 |
| Нормированы ли события индивидуальной жизни ... | 105 |
| Сколько видов ушастых тюленей живет сейчас ................. | 111 |
| Где живут ушастые тюлени .............................................. |
118 |
| Где место ушастым тюленям в современном мире ..... | 159 |
| Что дает изучение ушастых тюленей ............................. |
163 |
| Литература ........................................................................... | 170 |
 Вадим Федорович
Вадим Федорович
Мужчинкин
МОРСКИЕ ЛЬВЫ И КОТИКИ
Строение тела,
биология, место в биосфере
и сопряженные проблемы
Утверждено к печати
Редколлегией серии
научно-популярных изданий
Академии наук СССР
Редактор издательства
Г. М. Орлова
Художник
М. Л. Блох
Художественный редактор
Б. Ю. Кученков
Технический редактор
М. Ю. Соловьева
Корректоры Н. Г. Васильева,
Е. В. Шевченко
И Б № 35134
Сдано в набор 17.11.86
Подписано к печати 22.01.87
Т-05316. Формат 84х108 1/32
Бумага типографская М 1
Гарнитура обыкновенная
Печать высокая
Усл. печ. л. 8,24. Усл. кр. стт. 9,45. Уч.-изд. л. 9,6
Тираж 62 000 экз. Тип. зак. 3147
Цена 60 коп.
Ордена Трудового Красного Знамени
издательство «Наука»
117864, ГСП-7, Москва, В-485
Профсоюзная ул., 90.
2-я типография издательства «Наука».
121099, Москва, Г-99, Шубинский пер., 6
![]()
В издательстве
«НАУКА»
готовятся к печати:
В. Д. Ильичев, В. Т. Бутьев, В. М. Константинов
ПТИЦЫ МОСКВЫ И ПОДМОСКОВЬЯ
15 л.
В популярной форме описана история изучения птиц Москвы и Подмосковья. Приводятся
справочные материалы о птицах, гнездящихся или встречающихся на пролете на
территории данного региона, включая их географическое распространение, излюбленные
места обитания, численность, поведение, гнездование, питание и хозяйственное
значение. Рассказывается о видах, встречающихся в черте Москвы в различные
сезоны года, их численности в зависимости от характера городского ландшафта.
Рассматриваются вопросы охраны птиц.
Для зоологов, зоогеографов и любителей природы.
М. И. Шатуновский, Е. П. Огнев, Л. И. Соколов, Е. А. Цепкий
РЫБЫ ПОДМОСКОВЬЯ
10 л.
Дается характеристика водоемов Московской области. В популярной форме излагаются
сведения по биологии обитающих в них рыб. Характеризуются признаки каждого
вида, дана таблица для определения рыб. Рассматриваются изменения в ихтиофауне
за последние десятилетия под влиянием зарегулирования стока рек и других антропогенных
факторов. Подчеркивается необходимость проведения широкой природоохранительной
пропаганды среди населения.
Для широкого круга любителей природы.
Для получения книг почтой заказы просим направлять по одному из адресов: 117192 Москва, Мичуринский проспект, 12, магазин «Книга — почтой» Центральной конторы «Академкнига»; 197345 Ленинград, Петрозаводская ул., 7, магазин «Книга — почтой» Северо-Западной конторы «Академкнига» или в ближайший магазин «Академкнига», имеющий отдел «Книга — почтой».
480091 Алма-Ата, 91, ул. Фурманова, 91/97;
370005 Баку, 5, Коммунистическая ул., 51;
690088 Владивосток, Океанский проспект, 140;
320093 Днепропетровск, проспект Ю. Гагарина, 24;
734001 Душанбе, проспект Ленина, 95;
664033 Иркутск, ул. Лермонтова, 289;
252030 Киев, ул. Пирогоиа, 4;
277012 Кишинев, проспект Ленина, 148;
343900 Краматорск, Донецкой области, ул. Марата, 1;
443002 Куйбышев, проспект Ленина, 2;
220012 Минск, Ленинский проспект, 72;
630090 Новосибирск, Академгородок, Морской проспект, 22;
620151 Свердловск, ул. Мамина-Сибиряка, 137;
700185 Ташкент, ул. Дружбы народов, 6;
450059 Уфа, 59, ул. Р. Зорге, 10;
720000 Фрунзе, бульвар Дзержинского, 42;
310078 Харьков, ул. Чернышевского, 87.

