 Рис. 20. Возрастные перемены в поведении ушастых тюленей
Рис. 20. Возрастные перемены в поведении ушастых тюленей| Главная | Библиотека сайта | Форум | Гостевая книга |
Появление нового персонажа на сцене в разгар действия вызывает у зрителя понятное
желание узнать, откуда он взялся и не родной ли его брат стоит сейчас рядом
с ним на сцене, и не его ли родителей мы видели в предыдущем акте. И вот уже
складно выстраивается в голове его родословная и не хочется обращать внимание
на мелкие, назойливые неувязки и думать о том, что тот, кого мы прочим в родители,
мог быть и непричастен к этому, а за момент появления героя на сцене мы приняли
просто переход из темного угла громадной сцены на ее освещенный пятачок.
Ископаемые находки последних десятилетий показывают, что уже 10—15 миллионов
лет назад у калифорнийских берегов плескались ластоногие звери, хорошо вписывающиеся
в сегодняшние рамки семейства ушастых тюленей. Настоящие тюлени в это время
освоили европейские берега, а по евразийскому континенту были широко расселены
члены медвежьего семейства. Дальнейшие палеонтологические поиски, если повезет
с находками, покажут, существовали ли более древние медведи, фоциды и отарииды
и как они были расселены. Поскольку ясно, что геологическая летопись не очень
склонна сохранять океанских ластоногих (шансов на то, что костяк океанского
зверя будет захоронен раньше, чем разрушится, а захороненный минерализуется
и сохранится, гораздо меньше, чем для костяка медведя или тигра), а вероятность
найти их чудом сохранившиеся останки мала (нужно, чтобы океанские отложения
оказались на суше, в обследованном районе, в поле зрения заинтересованных людей),
то присутствие сегодняшних находок в миоценовых слоях Калифорнии не свидетельствует
об отсутствии ушастых тюленей в других районах и в более раннее время. Миоценовые
океанские млекопитающие до сих пор неизвестны ни с одного континента Южного
полушария, хотя никто, видимо, не сомневается в его заселенности. Так или иначе
сегодняшним рамкам, ограничивающим отариидную конструкцию (хотя бы только черепа),
не меньше 10—15 миллионов лет.
Само по себе освоение океана оказалось, по-видимому, не слишком сложным для
многих групп млекопитающих. В него в разное время внедрялись и выдры, и морские
коровы, и различные группы китообразных, и малопонятные звери десмостилии.
А уж пресноводные формы можно найти чуть не в каждом отряде.
Для всех трех групп современных ластоногих и строение тела, и сходство белков
сыворотки крови тесно связывают их со стволом наземных арктоидных хищных, объединяющем
вместе медведей, енотов, куниц и собак. То есть любое ластоногое ближе к собаке,
чем кошка.
При этом ушастые тюлени оказываются теснее связанными с медведями, а настоящие
— с куньими. Так сошлось старое немецкое название морских котов (морские медведи,
Seebaren) с сегодняшними представлениями о родственных связях. Несходство ушастых
и настоящих тюленей, а особенно их взаимоисключающие способы плавания, как
будто указывают на возможность расхождения их путей еще до ухода в океан и
независимого его освоения, как, скажем, различны пути сегодняшних потомков
наземных хищных — морских «бобров» и сегодняшних потомков других групп наземных
хищных — ластоногих. Напротив, сегодняшние моржи по всем признакам тесно примыкают
к ушастым тюленям.
Как выяснилось за последние двадцать лет, в миоценовой Северной Пацифике жило
еще две группы хищных зверей, история которых смыкается с историей отариид.
Около 15 миллионов лет назад одновременно или почти одновременно с ранними
ушастыми тюленями жили крупные (величиной с крупного сивуча, т. е. не меньше
тонны весом) ластоногие, по-видимому, с отариидной моделью поясов конечностей,
с черепом, отличным в равной степени от фоцидного и отариидного и вдобавок
еще с хоботом, как у морского слона (см. рис. 17). Звери эти — аллодесмиды
— были найдены еще в двадцатые годы (как и многие другие ископаемые североамериканские
морские звери) Р. Келлоггом, но детальное описание их анатомии было опубликовано
лишь в шестидесятых годах Э. Митчеллом. Чуть позднее тот же Митчелл издал описание
другой группы — водных или околоводных (скелет их конечностей неизвестен) —
эналиарктид — зверей размером со среднюю собаку, с вполне медвежьим черепом,
но с несколько увеличенной черепной коробкой и мелкими и более простыми зубами
(см. рис. 15). Их заметно различающиеся формы жили по обоим берегам Тихого
океана (в конце семидесятых годов одну из них нашла И. А. Дуброво на западном
побережье Камчатки) между 20 и 25 миллионами лет назад. Наличный черепной материал
не противоречит тесной связи эналиарктид с современными медведями, не позволяя
ни достаточно аргументированно обособить их друг от друга, ни пристегнуть миоценовых
жителей тихоокеанских берегов к какой-либо другой группе.
Если верно представление о жестких рамках, сковывающих изменчивость конструкции
в пределах хорошо очерченных семейств, то замкнутая в пределах семейства группа
медведей имела своих представителей на берегах Тихого океана уже почти 25 миллионов
лет назад (они немного старше наиболее древних из найденных материковых форм).
Такая же замкнутая группа несомненных ластоногих (аллодесмид) какое-то время
благоденствует в этом районе 10 миллионами лет позднее, а потом исчезает. Чуть
позднее на той же сцене уже действует хорошо очерченная и разнообразная группа
ушастых тюленей. Наиболее древние из известных членов семейства ушастых тюленей
моложе наиболее древних из известных членов семейства настоящих тюленей. Обе
эти группы имеют около себя каждая по отчетливо сателлитной группе гигантских
родственников, претендующих по своей обособленности на статус самостоятельных
семейств: для ушастых тюленей — моржи (до 4 м длиной и 2 т массой), для настоящих
— морские слоны (до 6 м длиной, 5 т).
На протяжении последних примерно 10 миллионов лет все перечисленные семейства,
кроме выбывших из игры аллодесмид, в уже оформившихся рамках двигались параллельным
курсом, сохраняя неизменными интервалы между собой. Но их отношения в предыдущие
15 миллионов лет можно моделировать по-разному.
Сходство каждой пары сравниваемых групп может быть истолковано трояко: либо
одна получила наследство другой (родительская и дочерняя группы), либо обе
несут наследство общего ближнего или дальнего предка (тогда встает проблема
определения давности их разделения), либо к обеим группам общие признаки пришли
независимо при ограниченности набора реализуемых вариантов (как, вероятно,
оказались сходными пропорции черепа у ленивца и морского кота).
При рассмотрении самого простого перехода, от родительской группы к дочерней,
возможны два варианта: либо сам переход между двумя стабильными состояниями
быстр и краток (тогда почти нет шансов, что его уловит летопись), либо он достаточно
долог и идет через цепь стабильных промежуточных состояний, но тогда нужны
свидетельства перехода от звена к звену, без чего вместо цепи окажется лишь
еще некоторое количество изолированных группок с проблематичными связями между
ними. При этом хорошо бы не упускать из виду ограничений, налагаемых возможностями
родительской и дочерней конструкции на предполагаемый переход от первой ко
второй. Соответственно гипотетическому предку ушастого тюленя нужны следующие
условия.
1. Предпосылки для сосредоточения теплоотдачи на поверхности кистей и ступней,
увеличение которых не должно иметь конструктивных препятствий.
2. Предпосылки для приобретения отариидного способа плавания, т. е. кандидат
в ушастые тюлени должен быть средних размеров, коротколапым, стопоходящим зверем
с длинной шеей и непригодным для плавания тонким или укороченным хвостом. Последнее
обстоятельство сразу исключает из числа претендентов выдрообразных зверей (вроде
калана или еще недавно претендовавшего на роль предка позднемиоценового семантора).
Тот же, отличный от фоцидного способ плавания свидетельствует в пользу расхождения
путей предков настоящих и ушастых тюленей еще на берегу.
3. Свидетельства водного или околоводного образа жизни, помимо устройства конечностей
(редко известного для ископаемых зверей). Широкое использование палеонтологами
в качестве маркера океанской или сухопутной жизни соотношения деталей среднего
и внутреннего уха не слишком надежно из-за неоднозначности этих особенностей.
Да и сухопутное млекопитающее — человек, ныряющий на стометровую глубину, вполне
обходится без этих усовершенствований. А именно на однозначно толкуемых деталях
ушной области черепа основывается причисление эналиарктид к морским зверям.
4. Нужны простые конические щечные зубы, а владельцы таких зубов перевелись
среди млекопитающих задолго до миоцена, либо нужно доказывать возможность постепенного
или скачкообразного преобразования сложного звериного коренного зуба в простой
конус. Среди аномальных вариантов зубной системы у современных хищных можно
найти примеры усложнения зубной коронки, по неизвестны случаи уподобления сложного
зуба тюленьему. Пока нет ни экспериментальной, ни какой-либо другой аргументации
в пользу таких преобразований, эналиарктиды остаются еще одной интересной группой
хищных обитателей морских побережий, если и связанных с ушастыми тюленями,
то скорее окольно. Проще допустить происхождение эналиарктидной зубной системы
из отариидной, чем наоборот. Та же проблема должна быть преодолена и в отношении
сегодняшних претендентов на роль предков зубатых китов.
5. Нужны предпосылки для приобретения еще одного маркера отариидного черепа
— загнутого внутрь углового отростка нижней челюсти. Этот признак ставит его
владельцев особняком среди плацентарных млекопитающих (у сумчатых этот отросток
тоже загнут внутрь, но иначе). Но неясно, когда его приобрели ушастые тюлени
(или их предки) и какую специфику он дает работе челюстного аппарата.
6. Нужны предпосылки для приобретения эмбриональной диапаузы и рождения необычно
зрелых для наземных хищных детенышей.
7. Нужны предпосылки в возможностях головного мозга и рецепторов для выхода
в трехмерный мир и более или менее длительная изоляция районов обитания предполагаемых
предков от контакта с континентальными животными сообществами. И сейчас ушастые
тюлени в отличие от настоящих лучше вписываются в сообщества небольших океанских
островков, чем океанских побережий материков.
Если теперь соотнести ограничения, накладываемые современным материалом, с
известным в настоящее время палеонтологическим материалом (из северной части
Тихого океана за последние 25 миллионов лет) и со схемой эволюции сывороточных
альбуминов, то вместо красиво ветвящегося единственно верного филогенетического
древа у нас в лучшем случае окажется некоторое количество шероховатых грубо
сколоченных заготовок, сильно отличающихся друг от друга, но вероятных в равной
степени.
Предполагая скорость изменений альбуминов постоянной на протяжении всей послемеловой
истории (около 65 миллионов лет) В. Сэрич (Sarich), тот самый, который определил
дату расхождения путей предков человека и шимпанзе в 5 миллионов лет назад,
посчитал иммунологические дистанции для 7 современных наземных хищных и 11
современных ластоногих по 7 антисывороткам (антисобака, антикошка, антимедведь,
антиенот, антикуница, антинерпа и антиморской лев). Он прикинул время расхождения
путей различных групп. Получилось, что от общего хищного ствола около 60 миллионов
лет назад отделилась ветка с будущими кошками, около 30 миллионов лет назад
оставшийся ствол развалился на три ветки (к будущим псам, ко всем ластоногим,
к енотам), около 15 миллионов лет назад расходятся пути настоящих и ушастых
тюленей. От настоящих тюленей около 10 миллионов лет назад отходят тюлени-монахи,
а чуть позднее расходятся пути нерп и морских слонов. В это же время в другой
ветви ушастые тюлени отходят от моржей. Схема красива, но исходит из сомнительной
предпосылки о постоянстве скорости изменений и явно искажает распределение
игроков по игровому полю использованием меньшего числа мерок (антисывороток),
чем число участников. Одновременно ощущается боязнь автора вступить в противоречие
с сегодняшней зоологической системой.
Активно работающие палеонтологи Ч. Репенинг (Reppening) и Р. Тедфорд (Tedford),
выведшие из небытия многих североамериканских ископаемых зверей, представляют
схему Сэрича растянутой по своему материалу: где-то в конце олигоцена от арктоидного
ствола наземных хищных отщепляется один сук о двух ветвях (одна — медведи,
другая — эналиарктины, будущие ушастые тюлени) и другой сук тоже о двух ветвях
(одна — куньи, другая — семантор, предок настоящих тюленей). На протяжении
миоцена эналиарктины выходят из игры, но оставляют после себя по крайней мере
две дочерние группы — специализированных аллодесмид, протянувших почти до конца
миоцена, и собственно отариид, которые в конце миоцена вычленили еще внешне
неотличимых от ушастых тюленей членов моржовой ветви (тех самых имаготарию,
айвукуса и других с пропорциями черепов, неотличимыми от современных морских
львов). А в плиоцене отариидная ветвь распалась на современные роды ушастых
тюленей. Четыре миллиона лет назад отошла веточка северных морских котов, два
миллиона лет назад оставшийся стволик разделился на южных морских котов и всех
остальных (кучкой — сивуч и калифорниец и другой кучкой — южноамериканский,
новозеландский и австралийский морские львы).
Схема тоже красивая и показывает, что для палеонтолога иммунологические свидетельства
не абсолютны, но зато важна убежденность в трех вещах: 1) эволюция может манипулировать
конструкцией зверя без ограничений (медвежий зуб свободно превращается в тюлений),
2) прежний жилец данной квартиры — почти несомненно предок сегодняшнего (если
в миоценовой Калифорнии аллодесмидам предшествовали эналиарктины, то они и
предки), 3) по деталям черепа можно однозначно определить образ жизни зверя
при отсутствии прочих частей скелета, а значит, можно постулировать принадлежность
эналиарктид к ластоногим, не дожидаясь находки их костяка.
В прошлом веке Ж. Кювье приписывали уверенность в возможности по одной кости
восстановить весь скелет. Сейчас мало кто верит и в эту возможность, и в то,
что Кювье говорил подобное, а в своем фольклоре сами палеонтологи шутят над
слишком категоричными реконструкциями: «На основе копролита голова слона отлита».
Но, по-видимому, сам характер ископаемого материала и работы с ним понуждает
к уверенности, что сегодняшние запреты тогда не действовали, а найденное решение
единственно. В результате каждая новая находка заставляет перерисовывать красивое
филогенетическое древо предыдущей публикации. Что это относится не только к
ластоногим, показывают постоянно меняющиеся (с каждым новым черепом и даже
зубом, по которым также уверенно строится облик владельца) схемы происхождения
человека. Если исключить из рассмотрения неисследованный вопрос о гомологичных
изменениях целостной конструкции и ограниченном наборе стабильных вариантов
конструкции, а оставить очень мягкие допущения, что сходство альбуминов падает
у потомков общего предка не обязательно всегда с постоянной скоростью и что
чуть раньше появившаяся на арктоидном стволе ветка не обязательно родительская
по отношению к чуть более молодой ветви, то не следует отбрасывать и третий
вариант возможной истории: пути медведей и ушастых тюленей разошлись еще в
момент формирования обеих этих групп, так же как пути фоцид и какой-то из групп
арктоидных хищных. Семантор — водный вариант куньих, а эналиарктины — водный
или околоводный вариант медвежьих. Каждый из них имеет отношение к событиям
внутри своего семейства, но не к событиям в соседних семействах. Аллодесмиды
же — независимая океанская ветвь арктоидного ствола, равноудаленная и от отариид,
и от фоцид. То есть для миоцена арктоидный ствол — совокупность равноудаленных
друг от друга и сохраняющих эту равноудаленность с течением времени семейств
наземных (собачьи, куньи, медвежьи, енотовые) и океанских (отарииды, фоциды
и аллодесмиды) хищных. Равноудаленность друг от друга семейств арктоидного
ствола не противоречит иммунологическим дистанциям Сэрича, так же как и допущение
их равновозрастности. Схема не противоречит и палеонтологическому материалу.
Очевидны и ее прорехи: она не решает проблемы появления новой ветви, а просто
отодвигает ее назад и оперирует возможным, но недоказуемым допущением о существовании
трех групп ластоногих раньше их самых древних сегодняшних находок (ведь нет
пока сведений о третичных морских зверях ни одного из южных континентов).
То есть наличный анатомический, палеонтологический и биохимический материал,
не давая однозначного, единственно верного ответа на вопрос, когда и откуда
появились на сцене владельцы отариидной конструкции, предполагает равновероятность
нескольких сценариев. Какие из этих схем придется отбросить и какие новые взять
в рассмотрение, покажут только новые находки — не только среди ископаемых,
но и среди современного материала.
Знать историю наших современников ушастых тюленей не менее важно для представлений
о мире и не менее полезно для ведения хозяйства, чем интересоваться собственной
историей, понять которую опять же можно лишь в рамках единой системы, биосферы.
Но понять историю владельцев отариидной конструкции можно лишь в той степени,
в какой будут преодолены барьеры между исследователями современного и ископаемого
материала, говорящих пока на существенно различающихся языках, т. е. нужно
построение единой системы знаний об уже вымерших и еще не вымерших зверях.
Представив устройство и возможности отариидной конструкции, владельцы которой
уже не меньше десяти миллионов лет притираются к месту среди прочих компонентов
биосферы и противостоят переменам в океане и на суше, попытаемся представить
основные вехи жизни ушастого тюленя, умудренного таким историческим опытом.
Щенок появляется на свет примерно через год после оплодотворения зрячим, подвижным,
с весом в одну десятую материнского (от 5 кг у морских котов до 20 кг у сивуча).
Прямая противоположность, скажем, медвежонку, родившемуся через полгода после
зачатия слепым и весящим около полукилограмма (одна пятисотая матери). Из этого
года, потраченного самкой Ушастого тюленя, на собственно беременность приходится
лишь около двух третей, т. е. примерно восемь месяцев, а четыре месяца — на
отсрочку в установлении контактов зародыша (на этой стадии — комочка клеток
с полостью между ними) со стенкой матки. Такая отсрочка, эмбриональная пауза,
достаточно обычна для млекопитающих различных групп, и хищных, и копытных,
и неполнозубых, а степень ее обязательности и продолжительность неодинаковы
даже в пределах одного вида зверей. Так, у косули от зачатия до родов может
проходить и шесть, и десять месяцев. Ускорение развития зародыша после паузы
происходит на фоне событий, которые охотоведы описывают как псевдотечку и псевдогон.
Такое же осеннее оживление половой активности через несколько месяцев после
основного весеннего пика демонстрируют многие звери и птицы (вспомним хотя
бы осеннее появление поющих скворцов у пустовавших с лета скворечников). Не
вполне ясно, кто участвует во втором пике: то ли отдохнувшие за лето участники
первого раунда, то ли пропустившие его взрослые или только вступающие в игру
молодые. Не ясно также, насколько велика реальность оплодотворения в это предзимнее
время. Но ясно, что какой-то будильник будоражит даже зверей и птиц с четко
очерченным сезоном размножения дважды в году, а не однажды. Для всех морских
котов и львов обычно называют годичным срок, за который оплодотворенная яйцеклетка
становится новорожденным. Но у северных морских котов и у южноамериканских
морских львов найдены осенние и зимние случаи родов, что может свидетельствовать
и о неурочном спаривании, и о вмешательстве чего-то, управляющего паузой в
ходе развития зародыша.
Переход из теплой материнской утробы на переполненное зверями, негостеприимное
лежбище ставит и пятикилограммового котового щенка и двадцатикилограммового
сивучонка в очень жесткие условия. Нехорошо быть затоптанным или загрызенным,
но нехорошо и утонуть.
«Не суйся в воду, пока тебе не исполнилось шесть недель, не то голова перетянет
пятки. И летние штормы и киты-косатки скверны для маленьких котиков, так скверны,
что хуже и не придумаешь. Но плескайся и набирайся сил, и ты не ошибешься,
Дитя морских просторов». Так не вполне последовательно наставляет своего сына
котиха в сказке Р. Киплинга «Белый тюлень».
Действительно, в воде новорожденный щенок неуклюж, хотя и может проплыть несколько
метров, высоко задрав голову, и старается как можно скорее выскочить из нее.
Но виноваты здесь скорее не пропорции тела, которые у него как раз уже родительские
(за исключением крупной детской головы), а неумение окунать голову под воду
и промокающий насквозь мех. Под промокшим мехом кожа уже после 15-минутного
купания остывает с 38° до 25°, а сам мех даже после отряхивания щенка на берегу
удерживает на себе около полулитра воды, медленное испарение которой еще долго
будет студить детеныша. Остуживание мокрым мехом мало подходит для промозглых
летних дней на островах Берингова моря, зато оно же спасает от теплового шока
на о-ве Сан-Мигель, где в жаркие дни температура воздуха на лежбище доходит
до 35°, а песок раскаляется до 50°.
У сивучонка проблемы другие, он добровольно идет в воду сразу после рождения,
и, возможно, этому способствует меньшая зрелость скелета его конечностей по
сравнению с менее крупными отариидными щенками (окостенение элементов кисти
и стопы к рождению опережает у котика на месяц ход у сивучонка), делая плавание
более легким, чем хождение. Детский мех его ненамного уступает котиковому,
но соотношение поверхность тела—объем заметно отличается. Однако котик, нарастивший
ко второму месяцу жизни на лежбище, вне материнской утробы, запас сала и более
густой мех (после отряхивания на берегу мех удерживает на себе не больше стакана
воды), уже уверенно лезет в воду при любой возможности (помните: «не суйся
в воду, пока тебе не исполнилось шесть недель»), хотя вес его все еще не больше
половины веса новорожденного сивучонка. Но особых дел в воде у недавно родившегося
щенка нет, зато на берегу он должен найти свое место в структуре лежбища. Он
может достаточно свободно перемещаться по личным территориям секачей, так как
на него еще не распространяются территориальные ограничения, жестко регламентирующие
передвижения взрослых самок и самцов. К тому же связь его с матерью из-за ее
многодневных отлучек между кормлениями не привязывает к одной-единственной
точке лежбища.
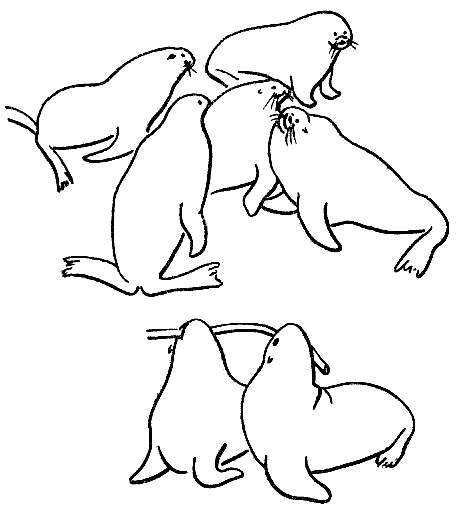
Рис. 20. Возрастные перемены в поведении ушастых тюленей
Внизу — два щенка новозеландского морского льва (по фотографии Б. Марлоу) пытаются завладеть куском стебля морской травы, демонстрируя элементы будущего взрослого территориального поведения. Вверху — разгар территориального конфликта между секачами северных морских котов (в окружении четырех законных владельцев своих территорий незадачливый вторженец, блокированный собственниками, каждый из которых, в свою очередь, не может выйти за пределы своего крошечного удела, не будучи атакован потревоженным соседом)
Блуждая по окрестностям места своего рождения, щенки скапливаются на местах,
свободных от взрослых зверей. К месячному возрасту котик осваивает уже участок
до 75 м радиусом от места появления на свет. Все больше и больше времени взрослеющие
щенки проводят в играх, в которых можно найти элементы будущего взрослого поведения
(рис. 20). В играх же осваивается вода. Время первого знакомства с водой сильно
меняется в зависимости от того, насколько близко к урезу воды находился гарем
с его матерью. Во время игр же проглатываются первые твердые предметы — чаще
несъедобные камешки, перышки и кусочки дерева.
Высокая жирность молока (20—40%) позволяет кормить реже, но, по-видимому, не
ускоряет рост и не сокращает срок выкармливания. Большинство ушастых тюленей
кормят щенка до рождения следующего, а в случае его гибели — и дольше, т. е.
год-полтора. Однако уже пяти-шестимесячного щенка можно перевести на чисто
рыбную диету. Такая пятимесячная лактация стала нормой у северных морских котов
Берингова и Охотского морей, где необходимость и щенкам и самкам покидать острова
на зиму с неизбежностью ведет к разлуке в море и разным местам зимовок для
взрослых и щенков. То же происходит и в другом полушарии у морских котов субантарктической
Южной Георгии. Осенний отрыв щенка от кормилицы на субарктических и субантарктических
лежбищах, связанный с необходимостью ухода в море, показывает не только зависимость
длительности лактации от внешней ситуации, но и поразительную слабость связей
между особями (даже такой сильной, как связь мать—дитя) у этих зверей, Эта
слабость связей разительно контрастирует с прочностью и длительностью личных
привязанностей у ближайших родственников отариид — моржей, где, несмотря на
почти круглогодичные скитания матерей среди льдов в крайне неблагоприятных
для сохранения родительских привязанностей условиях, самку могут сопровождать
(кроме выкармливаемого в данный момент щенка) ее потомки прошлого и даже позапрошлого
приплода, получающие свою долю молока.
Можно ожидать, что если убрать внешние помехи продолжению молочного кормления
у ушастых тюленей с необычно укороченной лактацией, ее срок достигнет обычного
для других года. К сожалению, нет сведений о длительности выкармливания щенков
северных котов на теплом калифорнийском острове Сан-Мигель, но на Утришской
биостанции беринговоморская котиха продолжала кормить своего щенка и глубокой
осенью, яростно обороняя его от любой угрозы (на лежбище мать обычно бросает
щенка на произвол судьбы при первой угрозе).
Независимо от степени предопределенности отлучения в первую осень оно достаточно
болезненно и стоит жизни многим младенцам. Порвав с кормилицей в возрасте полугода
или года с лишним и не имея тесной связи с местом рождения, ушастый тюлень
получает свободу в перемещениях по громадной акватории. Скитания могут длиться
год и дольше (некоторые беринговоморские коты появляются на лежбищах лишь в
двух-, а то и в трехлетнем возрасте). По мере роста зверя добыча, заглатываемая
целиком, должна оставаться соразмерной глотке, и соответственно набор кормов
будет неодинаков у особей разной величины, определяя распределение в море сеголетков,
взрослых самок, молодых самцов и секачей. Ежегодная линька постепенна и не
нарушает водонепроницаемости меха, а для морских львов проблемы с мехом вообще
не должно существовать, но как раз морские львы выглядят более оседлыми.
Малая доля неполовозрелых зверей, не считая щенков, среди толпящихся на лежбище
в летний сезон а большая примесь пришельцев из разных точек видового ареала
среди вернувшихся домой трехлеток говорят о том, что запечатление образа родного
лежбища у молодых отариид не очень жестко и может быть легко позабыто. Те холостяки
(трех-четырехлетние самцы), которые составляют основу промысловой добычи на
лежбищах северных морских котов, проводят на берегу за три летних месяца не
больше 20 дней (не подряд, а с перерывами). Тем не менее приближающееся созревание
заставляет все большую и большую часть выживших в море ровесников собираться
каждое лето к родному или по каким-то причинам выбранному чужому лежбищу. Но
даже в разгар летней береговой жизни примерно треть любого стада остается рассеянной
по океану, путая карты береговым наблюдателям и прогнозистам. Самки ушастых
тюленей созревают в два-три года и в этом же возрасте могут оказаться оплодотворенными,
если нет каких-либо существенных нарушений в структуре взаимоотношений на лежбище.
Такие нарушения могут отсрочить вступление самки в размножение на несколько
лет.
Со следующего после оплодотворения года жизнь самки будет регламентироваться
ежегодно до 20— 30-летнего возраста необходимостью выходить на берег, часто
из года в год на один и тот же его участок, за несколько дней до родов, оставаться
несколько дней при новорожденном, зачинать следующего, а затем раз в неделю
возвращаться из моря и отыскивать среди тысяч слоняющихся по всему лежбищу
и около, орущих и пытающихся сосать любую самку щенков именно своего, накормить
и снова уйти надолго в море. И только у тех немногих отариид, чьи самки кормят
меньше года, получается перерыв между концом лактации и очередными родами,
да гибель щенка (если возникшая пустота не заполняется выкармливанием приемыша)
или нарушение в сроках прихода течки разрывает непрерывность этой цепи событий.
Самцы отариид как будто способны к оплодотворению почти в том же возрасте,
что и самки, но обычно надежно устраняются пространственной изоляцией от контакта
с находящимися в течке самками. Вступив в физиологическую зрелость, но не достигнув
социальной, они продолжают быстро расти, обгоняя почти остановившихся в росте
самок за несколько лет в три—пять раз по весу и в полтора раза по длине тела.
Лишь в 5—8 лет (у разных форм ушастых тюленей) самец приобретает статус секача,
претендующего на собственную территорию на гаремном лежбище.
Следующие 10 лет, а у редких удачников даже 15 он будет каждую весну, не дожидаясь
прихода самок, участвовать в переделе лежбища между владельцами, стремясь обосноваться
из года в год в одном и том же месте, караулить занятый плацдарм несколько
дней или несколько недель независимо от присутствия или отсутствия самок на
нем, многократно покидать свой клочок берега и с трудом пробиваться на него.
И так до конца лета, а когда уж все как будто затихло на лежбище, вдруг начинается
поздней осенью второй пик активности, совпадающий по времени с концом задержки
развития зародышей у оплодотворенных самок и связанный то ли с восстановлением
сил у ветеранов, то ли с приходом только подготовившихся к размножению особей
обоего пола. Бывает ли осеннее оплодотворение или это действительно лишь ложное
оживление, не известно ни применительно к отариидам, ни применительно к поющим
осенью скворцам.
По мере приближения конца лета не связанные больше с лежбищем секачи, так же
как и нелактирующие самки, рассеиваются по океану, скапливаясь в благоприятных
районах различных для разных полов и возрастов, чтобы через некоторое время
снова рассеяться по акватории и успеть к концу весны на свое лежбище.
Если теперь разметить жизненный путь ушастого тюленя основными вехами: продолжительность
беременности после окончания эмбриональной паузы, степень зрелости и относительные
размеры новорожденного, срок созревания (и физиологического, и социального),
длительность участия в размножении, период лактации и продолжительность жизни,
то обнаружится, что жизнь отариид до деталей повторяет схему жизни далеких
от них и по биологии, и по положению в системе класса млекопитающих — копытных
(сходных размеров, конечно), отличаясь одновременно по распределению тех же
вех на жизненном пути от варианта своей медвежьей родни.
Морской котик родится через 8 месяцев после окончания эмбриональной диапаузы
массой в 5—7 кг, кормит его мать 4—5 месяцев, половой зрелости он достигает
в 3 года, когда самка и начинает размножаться при массе в 50 кг, но у самца
начало размножения откладывается до восьмилетнего возраста и массы в 300 кг.
Самки доживают, сохраняя способность к размножению, до 25 лет, самцы не достигают
двадцатилетнего возраста. Бурый медвежонок появляется на свет после 6 месяцев
в утробе, весит при рождении полкило, сосет мать полгода, созревают оба пола
к 10 годам, когда самка весит 300 кг, а самец — 500, плодятся до 30—35-летнего
возраста. Зубренок вынашивается 8 месяцев и при рождении весит около 20 кг,
в пятимесячном возрасте может быть переведен на взрослую диету, созревает на
третьем году, но самцы участвуют в размножении лишь после 7 лет (масса самки
500, самца — до 1000 кг), самки могут плодиться до 25-летнего возраста, самцы,
обычно не доживают до 20 лет. Внешние события могут изменить некоторые детали,
но в целом каждый вариант достаточно жесток.
Еще раз взглянем на расписание жизни ушастого тюленя, чтобы представить себе
наиболее уязвимые места этой схемы. Первое — задержка установления контакта
бластоцисты со стенкой матки повышает риск потери зародыша и ставит длительность
беременности в зависимость от конкретной обстановки. Промысловики предполагают,
например, значительные потери зародышей у самок южноафриканских морских котов
во время осенних отгонов на лежбищах.
После имплантации зародыша идут восемь относительно безопасных месяцев его
роста в теплом и покойном плодном пузыре, подводящие его ко второму кризису
— переходу на холодное и негостеприимное лежбище, где нерасторопному или потерявшему
мать — гибель. Миновав этот кризис, щенок входит в ровную жизнь среди сверстников
на детских площадках, размеренную редкими, но регулярными визитами матери с
молоком. Структура лежбища еще не сковывает щенка так жестко, как взрослого.
Затем приходит третий кризис — рвется связь с матерью. Очевидно, что чем раньше
это происходит, тем острее ломка и вероятнее гибель.
Пережив и это, ушастый тюлень заработал следующие несколько лет спокойной жизни
(если не считать морских хищников и возможности лишиться шкуры на берегу) в
океане с недолгими визитами на побережье. Четвертый кризис связан с необходимостью
для взрослеющего зверя найти свое место в сложившейся структуре лежбища, которая
определяет и срок наступления социальной зрелости у самок и самцов данного
стада, и степень трудности перехода в категорию размножающихся особей, и успех
размножения в дальнейшем. Ясно, что это четвертое испытание мягче дается самкам,
чем самцам, и секачам калифорнийских морских львов при более рыхлой структуре
их лежбища, чем секачам северных морских котов, у которых костяк лежбища жестче.
Собственно, для секачей этот кризис, хоть и в несколько смягченной, привычной
форме, повторяется ежегодно при возобновлении структуры лежбища.
Последний, пятый кризис определяет конец жизни зверя, избежавшего гибели в
четырех предыдущих ломках, не съеденного в море и не погибшего во время промысла.
Наступает он у секачей раньше (у северных котов на десять лет раньше), чем
у самок, но в разное время в разных стадах одного вида ушастых тюленей и у
разных их видов.
Легко заметить, что все четыре перелома на протяжении жизни особи связаны с
переменой места в системе: мать—зародыш, мать—щенок—лежбище, щенок—океан, океан—особь—лежбище.
Лежбище и освоенная его членами акватория представляют собой хорошо очерченную
единицу, которая связана с другими такими же единицами совсем слабым или, напротив,
сильным обменом особями, а также эксплуатацией общих ресурсов. В результате
система таких блоков образует единый и дробный одновременно ареал, освоенный
сегодня данным видом ушастых тюленей. Но прежде чем говорить об ареале, посмотрим
на одну из составляющих его структур — структуру берегового лежбища.
На первый взгляд на берегу без всякого порядка, явно мешая друг другу, копошатся звери, согнанные сюда нехваткой места для более просторной жизни. Английское название этого скопища — грачевник (rookery) объединяет в одну категорию облепленные галдящими птицами и их гнездами деревья и кишащий столь же крикливыми ушастыми тюленями участок на берегу океана. Структурирован ли этот беспорядок, и если структурирован, то что общего и что отличает конструкцию лежбища и птичьей колонии?
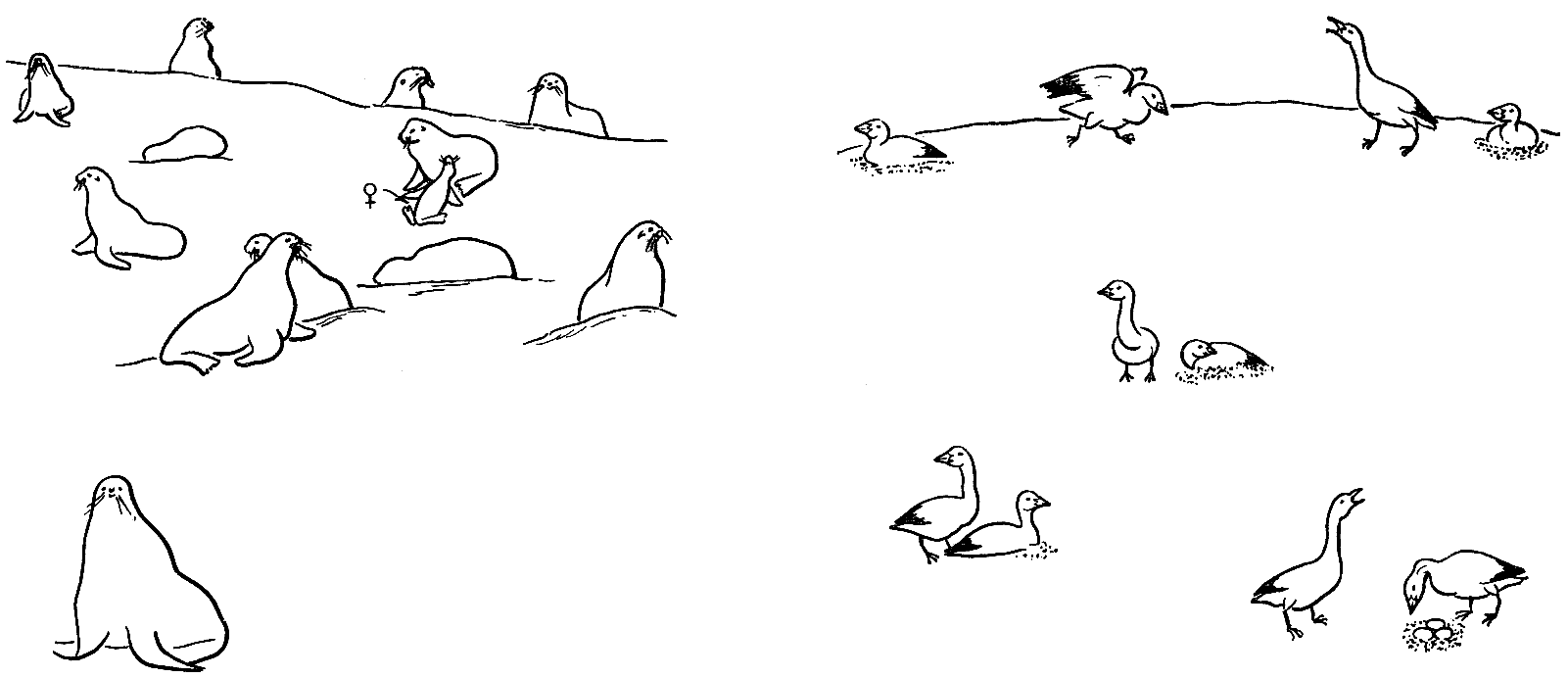 |
|
|
|
|
|
Для упрощения сравнения заменим трехмерный грачевник развернутой на плоскости,
удобной для наблюдения колонией белых гусей в центре острова Врангеля. Маленький
кусок ее нарисован рядом с таким же маленьким куском колонии северных морских
котов (рис. 21). Любой внимательный наблюдатель быстро найдет элементы порядка
на обоих рисунках. «Здесь каждый одинок и равнодушен, покамест сам внезапно
не укушен, не сдвинут с места, не поддет клыком»,— такой не слишком гуманистичный
порядок увидел писатель Ф. Искандер на котовом лежбище. Этот порядок, обеспечивающий
одиночество в толпе, прочен и основан на простой структурной единице: у значительной
части секачей каждый владеет участком в несколько метров поперечником, который
недоступен для самцов любого возраста старше щенячьего. Любая же самка будет
приветствоваться на любом участке, и владелец приложит все усилия, чтобы ее
удержать. Отдельные владения, плотно смыкаясь, образуют единый монолитный костяк
любого лежбища. Среди ушастых тюленей наиболее отличен от котового вариант,
осуществляемый калифорнийскими морскими львами, у которых индивидуальное владение
секача разрастается до нескольких десятков метров в поперечнике и включает
не только кусок берега, но и прилежащую акваторию, а кроме того, оно доступно
и для не достигших полных размеров самцов на условиях их беспрекословного подчинения
владельцу.
После весеннего раздела территории между удачливыми секачами новым претендентам
остается либо пристраиваться со своими участочками с краю обжитой территории,
либо ждать вакансии на лежбище, занимая любой безнадзорный клочок. В разгар
сезона поделенной оказывается даже литораль, обнажающаяся во время отлива перед
лежбищем, которая через несколько часов вновь окажется под водой. Так как секач,
хотя и может просидеть на своем береговом владении активным и голодным десять
и больше дней, но почти никогда не высиживает весь летний сезон безвылазно,
то в течение сезона происходит постепенное обновление набора владельцев. Секачи,
не имеющие в данный момент претензий на владение собственным куском пляжа и
не достигшие статуса секача самцы, не могущие соответственно претендовать на
что либо, оказываются вне каркаса лежбища и могут образовывать самостоятельные
залежки, иногда примыкающие вплотную к гаремному лежбищу, а иногда удаленные
на десятки и сотни километров от основного лежбища.
Эти холостяковые залежки не имеют структуры и объединяют секачей, покинувших
свои владения на лежбище (после отдыха некоторым из них удастся вновь прорваться
туда), с молодыми самцами и немногочисленными самками, забредшими сюда. Присутствие
самок поблизости от секачей приводит к тому, что то один, то другой из них
вдруг вспоминает о своей роли и начинает преследовать самку, отгоняя подвернувшихся
соседей. Однако такого запала хватает обычно ненадолго, и вскоре все снова
успокаивается.
Тем временем на гаремное лежбище после возникновения каркаса, образованного
территориальными секачами, стали выходить самки (выходят самки незадолго до
родов) и располагаться группами вплотную друг к другу в пределах этого каркаса,
примыкая к пришедшим раньше товаркам. Вскоре каркас оказывается заполненным
более подвижной и плотно упакованной массой самок. Но одновременно идет и развал
структуры.
Родившие и оплодотворенные самки все больше времени проводят в море, возвращаясь
на берег лишь для того, чтобы отыскать и накормить своего щенка. Щенки же,
чуть окрепнув, перебираются на места, свободные от взрослого зверя. Секачи
по-прежнему несут вахту на своих местах, но место выбывших уже не всегда занимается
новыми рекрутами из-за постоянного уменьшения числа претендентов. То есть в
каркасе появляются все новые и новые бреши, и к концу лета он окончательно
разваливается. Лежбище становится таким же скопищем разнополого и разновозрастного
зверя, как и холостяковая лежка. Лишь иногда можно видеть одного или двух секачей,
пытающихся удержать около себя каждый свою облюбованную самку.
Число таких секачей увеличивается во время осеннего пика активности, ложного
гона. Все больше зверей покидает лежбище, рассеиваясь по океану. Однако при
благоприятных погодных и кормовых условиях в окрестностях часть зверя может
досидеть на лежбище до следующей весны, когда быстро начнет восстанавливаться
его структура. С другой стороны, и в разгар летней жизни часть стада остается
рассеянной по океану.
Это сильно схематизированное описание событий на лежбищах северных морских
котов в Северной Пацифике может быть приспособлено для любого ушастого тюленя
путем поправок в трех местах: 1) меняется размер индивидуального пространства
секача, т. е. пространства вокруг особи, куда не допускаются особи одного с
ним ранга, и набор допускаемых внутрь владения особей (только самки, или и
молодежь, или и взрослые субординатные самцы впридачу); 2) меняется степень
плотности (или рыхлости) упаковки соседних владений секачей в единый массив;
3) меняется длительность существования описанных структур па протяжении года
(от весенне-летней до круглогодичной) . При этом одним крайним вариантом будет
галапагосское лежбище калифорнийского морского льва, где секач атакует только
близкого по размерам самца, беспрепятственно пропуская всех более мелких. Включение
акватории во владения каждого территориального секача исключает для всех обитателей
опасность перегрева и обеспечивает беспрепятственный уход в воду и выход из
воды. Участки секачей могут и смыкаться друг с другом, и быть разделены неудобными
кусками берега. Сезон размножения растянут на полгода на Галапагоссах, а па
северных калифорнийских лежбищах сокращен до трех месяцев. Секача-владельца
легко сгоняет с места и человек, и вышестоящий в иерархии секач.
У прочих морских львов секачи владеют меньшими участками, соответственно недоступными
для субординатных самцов, но вне гаремных территорий между самцами устанавливается
иерархия. В сезон размножения секачи и половозрелые самки присутствуют не на
всех лежбищах (у сивучей А. Н. Белкин нашел даже отдельные лежбища молодняка,
годовиков и двухлеток). Сезон размножения длится несколько месяцев, и на его
протяжении набор территориальных секачей обновляется за счет запаса претендентов.
Из морских котов галапагосские и гваделупские похожи на калифорнийских львов
большими владениями секачей, рыхло разбросанными среди неблагоприятных местообитаний,
но, по-видимому, так же мало терпимы даже к субординатным самцам, как и прочие
морские коты.
Завершает этот ряд вариантов уже знакомая модель лежбища северных морских котов,
повторяющаяся в общих чертах почти у всех южных и диаметрально противоположная
наблюдаемому у калифорнийских морских львов.
Внутривидовая изменчивость конструкции лежбища должна оперировать теми же ее
характеристиками, что и межвидовая изменчивость, но, к сожалению, остается
совершенно неизученной.
Свободное перекрывание ареалов калифорнийца, прочих морских львов и морских
котов сопровождается перекрыванием, хотя и неполным, сезонов размножения и
предпочитаемых мест скопления на берегу. Таким образом, создаются предпосылки
для совместного залегания двух и даже трех видов ушастых тюленей и образования
конгломератных лежбищ. Такие химеры известны для сивучей и морских котов на
Командорах, для калифорнийских морских львов и тех же котов — на о-ве Сан-Мигель
и для сивучей и калифорнийцев — на о-ве Аньо-Нуэво. При этом в индивидуальные
владения секачей калифорнийских морских
львов допускаются нетерриториальные звери других видов, чего не допускают территориальные
секачи морских котов, которые, в свою очередь, оттесняются территориальными
секачами сивучей.
И все это происходит на фоне того, что так же, как особи одного вида тяготеют
к обществу друг друга, стремясь сбиться в одновидовую кучу, так особи разных
видов отариид явно предпочитают совместное залегание раздельному. В результате
гаремные территории одного вида либо оттесняются на место холостяковых залежек
другого вида отариид, либо территориальные секачи одного вида располагаются
мозаично среди территориальных секачей другого вида. На холостяковых же залежках
свободно смешиваются нетерриториальные особи любых видов ушастых тюленей.
Несмотря на многочисленные темные места в жизни отариидного лежбища (не ясно,
что ограничивает размеры гаремного лежбища при обилии претендентов во владельцы
участков, чем определяется место первого в жизни данного секача участка, чем
определяется степень привязанности данной особи к данному лежбищу, чем определяется
место и время выхода на лежбище неоплодотворенных самок), можно в общих чертах
представить историю лежбища с момента его возникновения. Поскольку у всех ушастых
тюленей взрослые самки и секачи из года в год стремятся выйти не только на
одно и то же лежбище, но и на один и тот же небольшой его участок (облюбованный
то ли в детстве, то ли в зрелости, если не навсегда, то надолго), можно полагать
основным контингентом для основания новых колоний самцов и самок, только вступающих
в пору социальной зрелости. Так как самкам легче вписаться в существующую структуру
любого лежбища ушастых тюленей, доля непристроенных самцов будет существенно
выше, хотя и у самок есть свои трудности, на что указывает иногда очень значительное
запаздывание первой беременности (самки прибыловских котов, например, созревая
к трем годам, приносят первого щенка в пяти-шестилетнем возрасте).
Для всех отариид известны не только скопления холостяков вокруг гаремных территорий,
но и залежки неразмножающихся зверей в десятках, сотнях и даже тысячах километров
от ближайшего гаремного лежбища. Так, в нескольких тысячах километров от родительских
лежбищ залегали холостяки северных морских котов на еще не продуцировавших
лежбищах о-ва Сан-Мигель. Известно также, что на некоторых из этих залежек
со временем появляются размножающиеся особи и превращают их в продуцирующие
лежбища. Наконец, возвращение молодых на родное лежбище после ухода с него
щенками за год или несколько лет до возвращения, притом, что в море связи между
особями слабы и молодые держатся отдельно от взрослых, однозначно говорит о
запечатлении образа дома еще в детстве. Остается допустить, что запечатлевается
ограниченный участок берега в пределах, скажем, ста метров вокруг места рождения,
где были сосредоточены впечатления первого месяца жизни, и станет понятной
судьба лежбища.
Небольшое по размерам лежбище может принимать всех своих щенков по мере их
взросления, давая им возможность прирастить его территорию собственным участком
с краю, который при малых размерах лежбища все еще оказывается в пределах их
запечатленного дома, или надеяться на освобождение кем-нибудь из старожилов
места в центре.
Каждое новое приращение будет раздвигать края лежбища, и если процесс идет
быстро, то скоро подросшие щенки из центральных районов обнаружат, что они
могут претендовать только на краевые участки, которые уже настолько отодвинулись
от центра, что вышли за границы запечатленного как дом. Попасть домой можно,
только ожидая вакансии на обнажающейся в отлив литорали, и таких претендентов
с каждым годом все больше, а шансов все меньше. То есть появляются особи, для
которых дом их детства в момент приобретения физиологической зрелости и подхода
социальной окажется недоступным несколько сезонов подряд. Связи с домом рвутся
все у большего числа взрослеющих особей, они либо должны проводить жизнь в
море, либо искать любые пригодные по рельефу и климату места для временных
залежек. А поскольку при утрате связей с домом сохраняется тяга к выходу в
места, где уже лежат звери своего или близких видов, то место выхода на берег
одного зверя будет привлекать и других скитальцев.
В результате растущее лежбище будет продуцировать не только молодежь, оттесненную
на края лежбища и ждущую своего срока, но и все большее количество бездомных
скитальцев, залегающих сначала только осенью, а потом и летом в одной или нескольких
точках, удаленных от родительского лежбища. Тем временем исходное лежбище все
еще растет с краев, но его середина уже продуцирует только мигрантов, а естественная
убыль резидентов в центральных районах компенсируется уже не своими, а периферийными
особями. Лежбище оказывается трехслойным: центр дает выселенцев за пределы
лежбища, все реже оседающих здесь же, и принимает все больше вселенцев с краев
лежбища. Примыкающая к центру зона продуцирует выселенцев и достаточное количество
резидентов, исключающих вселение посторонних. Периферия лежбища все еще обеспечивает
прирост территории, но он уже замедлен оттоком переселенцев к центру лежбища.
На холостяковых лежках тем временем взрослеют пришедшие прежде других самцы
и появляются первые самки, которых рост родительского лежбища тоже лишил дома.
Гаремная территория вмещает плотно упакованных самок больше, чем рыхло упакованных
секачей, но самок становится больше, чем может вместить и плотная упаковка.
Остается один шаг до оплодотворения самок на холостяковой залежке, появления
там первых щенков и превращения бывшей холостяковой залежки в маленькое продуцирующее
лежбище.
Место не каждой холостяковой залежки может обеспечить условия (на берегу и
вокруг), необходимые для выживания приплода, и соответственно лишь некоторые
из них могут превратиться в продуцирующие лежбища. Но движение в этом направлении
неизбежно.
Итак, в пределах акватории, известной обитателям исходного разбухшего лежбища,
появляется одно или несколько сателлитных лежбищ, гаремные территории которых
будут еще более притягательны для переселенцев, чем холостяковые залежки на
их месте. И отток, и приток особей обладают некоторой инерцией. Так что молодые
лежбища еще некоторое время будут расти ускоренно за счет переселенцев и собственной
продукции, пока сначала переселенцы, а потом и часть собственных потомков не
будут блокированы разбуханием территории уже дочернего лежбища. Из-за той же
инерции отток с родительского лежбища не прекратится сразу, как будут выравнены
продукция и выселение, и лежбище не только остановится в росте, но и начнет
сокращаться в размерах, либо сжимаясь к центру, либо распадаясь на несколько
самостоятельных кусков.
Уменьшение размеров неизбежно позднее пресечет выселение, и начнется вначале
очень медленный рост за счет только своего небольшого поголовья. Цикл замкнулся.
От освоения нового необжитого места под холостяковую залежку, на которой спустя
десять-двадцать лет появятся первые беременные самки и еще лет через десять
начнется медленный рост гаремной территории, через фазу ускоряющегося роста
к продуцированию выселенцев и образованию удаленных холостяковых лежек (будущих
дочерних лежбищ), а затем и к обособлению от событий на прочих лежбищах в пределах
собственной резко сократившейся территории должно проходить не меньше полувека,
если не век.
В этой упрощенной схеме не учтены по крайней мере три группы факторов, влияющих
на смену и длительность фаз: ограниченность пригодных мест для береговых лежбищ
(кусок пригодного берега может быть длиной и сто метров, и в несколько километров)
и кормовых ресурсов в море; включение регулятора — детской смертности при повышении
плотности населения и влияние выборочного (или невыборочного) промысла на структуру
данного лежбища. Но сам ход событий как будто не противоречит известному об
отариидных лежбищах сейчас.
Рассмотрев структуру отариидной колонии, возможности многовидовых конгломератных
поселений и историю отдельной колонии от детства к зрелости, попробуем примерить
отариидную схему к на первый взгляд похожей второй половине рис. 21, где Е.
В. Сыроечковский нарисовал часть гусиной колонии.
Вся наличная площадь ее поделена между отдельными владельцами — гусаками, изгоняющими
все постороннее, каждый со своего участка. На колонии остаются только территориальные
самцы и взрослые самки, участвующие в размножении в этом сезоне. Если вычленить
основной элемент конструкции колонии, то обнаружатся прочные многолетние пары,
в которых место для гнезда выбирает гусыня, а привязанный к ней на невидимый
поводок гусак старается не отстать и одновременно очистить ближайшую округу
радиусом от одного до пятнадцати метров (в зависимости от обстоятельств радиус
меняется) от попадающих туда гусей одного с ним ранга, т. е. от любого гусака.
Невольно вспомнишь секача, преследующего на залежке среди разнополого и разновозрастного
зверя избранную самку и очищающего свое индивидуальное пространство вокруг
от конкурентов.
К месту гнездования весной гуси приходят уже парами, сбившимися в стаи по нескольку
десятков особей, вместе с некоторыми родителями держатся их прошлогодние птенцы
(числом от 1 до 5). Уже через неделю после появления первых гусей на острове
место будущей колонии распределено между отдельными парами, рассеянными с интервалами
от нескольких до нескольких десятков метров. Взаимное расположение гнезд определяется
притягательностью места колонии для многих гусынь и невозможностью для привязанных
к ним гусаков располагаться близко друг к другу. Место выбирает самка по своим
неясным для нас меркам, с учетом качеств биотопа и привязанности к местам своих
прошлых гнезд.
После выбора точки на местности самка, а следовательно, и самец с индивидуальным
обороняемым пространством вокруг него прочно закреплены в ней. Различные интервалы
между гнездами могут быть связаны как с непригодностью отдельных участков,
неотличимых на первый взгляд от соседних, занятых, так и с личными качествами
гусаков. Закрепленный на местности гусак как будто ничем не отличается от территориального
секача. Оба они будут защищать свою землю от посторонних и пытаться спариваться
с любой попавшей туда самкой своего вида. То, что гусыня вместе с гусаком нападает
на вторженцев, показывает, что и у нее в это время есть свое охраняемое пространство.
В то же время гусак, по крайней мере к концу насиживания, покидая надолго гнездо
с гусыней, как бы выходит на это время из своего индивидуального пространства,
и ничто не показывает наличие неприкосновенной зоны вокруг него (опять же полная
аналогия секачу, покинувшему на время свое место на гаремном лежбище).
Таким образом сформировавшаяся колония представляет собой местами плотное,
местами диффузное скопление закрепленных на местности индивидуальных пространств
гусаков. Прошлогодние птенцы и не участвующие в этом году в гнездовании одиночки
и не закрепившиеся пары активно изгоняются резидентами (в том числе и собственными
родителями) и в разгар сезона уже кочуют группами по острову.
Отклонения от схемы двояки: гусак, потерявший гусыню или ушедший от нее достаточно
далеко, пытается спариваться с любой гусыней, независимо от того, в паре она
или нет; часть гусынь в парах либо по реальной непригодности незанятых другими
участков, либо по индивидуальным особенностям поведения не находит места для
гнезда и откладывает яйца в чужие гнезда или около них.
За гусыней, прорывающейся к чужому гнезду, следует ее гусак, что неизбежно
ведет к конфликту с хозяином, который то гонит вторженцев, то пытается спариваться
с ними независимо от их половой принадлежности. В результате таких попыток
некоторая часть территориальных гусаков успевает за сезон покрыть по нескольку
прошедших через их индивидуальное пространство самок, т. е. проигрывает нормальный
для территориальных секачей ушастых тюленей вариант.
Первыми колония отторгает прошлогодков, изгнанных из родительских индивидуальных
пространств. Затем уже в тундре, вдали от колонии, к ним присоединяются не
участвовавшие в размножении или потерявшие кладку (или партнера) особи, державшиеся
до этого в пределах колонии. Птицы сбиваются в крупные (до 700 птиц) табуны,
где индивидуальные пространства особей, если и сохраняются, то радиус их минимален.
У гусаков на гнездовье индивидуальные пространства прикреплены к гнездам до
вылупления гусят. Якорь, державший территорию самца на месте, оказывается поднятым,
и гусак уходит из колонии, не отставая от гусыни с гусятами и отгоняя вторженцев
от своей передвижной территории.
Выводки один за другим покидают гнездовье. Уже двух-, трехнедельные гусята
участвуют в защите от вторженцев (сначала от мелких, потом и от более крупных)
общего для семьи индивидуального пространства гусака. Попыток раздела пространства
внутри семьи не видно. К подъему выводка на крыло индивидуальное пространство
оказывается сжавшимся до предела, как уже случилось раньше у незагнездившихся
особей.
Сначала вылинявшие годовики, потом взрослые неудачники и, наконец, вылинявшие
выводки начинают группами покидать остров, двигаясь к местам зимовок. При этом
места зимовок разных пар одной колонии могут оказаться существенно различными.
Как обстоит дело с индивидуальным пространством особей на зимовках, неясно.
Но именно на зимовках происходит образование новых пар, а также иногда восстановление
старых прерванных связей. Это и определяет поведение гусака — носителя индивидуального
пространства — на следующий год или часто на многие годы (т. е. репродуктивное
перемешивание происходит на местах зимовок и затруднено на местах гнездовий).
В то же время на зимовке сохраняются еще связи родителей с последним выводком.
Образование конгломератных колоний с совместным гнездованием разных видов гусей
не редко и связано, помимо привлекательности одних и тех же угодий для разных
гусей, еще и с отчетливым предпочтением компании (даже других видов) одиночеству
и малочисленности. При этом дистанция между гнездами разнопле-менников меньше,
чем между гнездами единоплеменников, т. е. индивидуальное пространство гусака,
закрытое для своих, частично открыто для чужих.
Наиболее занятным результатом влечения гусиных пар к иноплеменникам, да еще
очень далеким, пожалуй, будет гнездование в пределах активно охраняемого индивидуального
пространства гнездящейся пары белых сов (или других крупных хищников). Исследовавший
это рискованное сожительство Е. В. Сыроечковский полагает, что именно оно позволяет
первым десяткам пар вселиться и окрепнуть в месте, защищенном присутствием
гнезда хищников от прочих охотников до гусиных яиц, положив тем самым начало
новой растущей колонии.
Вероятно, начало гусиной колонии может быть и другим (например, на островке
или в другом защищенном месте), но несомненно, что начало кладут не одиночки,
а немногочисленные пары, по каким-то причинам порвавшие со своим местом рождения
и нуждающиеся в годах спокойного гнездования на новом месте, при постоянном
приращении площади гнездовья и сохранении плотной упаковки индивидуальных пространств
гусаков. Лишь после этого спокойного роста за счет туземного приплода и переселенцев
из других мест будет сформирован достаточно прочный костяк из достаточного
количества достаточно прочно закрепленных и достаточно скученных гнезд, который
может противостоять разрушительному действию непогоды и хищников.
Выйдя из-под опеки и продолжая расти, гнездовье должно пройти, по-видимому,
те же фазы, что и отариидная колония, с неодинаковостью работы разных зон колонии,
неизбежным (при отсутствии помех) продуцированием выселенцев, усиленным оттоком
их в новые места гнездования, сокращением территории гнездовья и временной
стабилизацией перед повторением цикла. Разумеется, и здесь ограничение ресурсов
(кормовых и территориальных), изменение смертности приплода и выборочное (или
без разбора) разрушение гнезд будет определять ход и длительность отдельных
фаз.
Сравнение гусиной и отариидной колонии показывает, что в основе
образования обеих лежит стремление большого количества особей закрепиться на
ограниченном
участке и претензии взрослых самцов не допускать в ближнее пространство вокруг
себя (индивидуальное пространство) особей своего ранга. Эта мерка — недоступное
для других пространство — позволяет довольно надежно описывать геометрию взаимного
размещения особей в сообществе, ту основу, на которой из отдельных особей одного
вида возникает постройка следующего порядка. Кроме того, введение этой мерки
позволяет заменить ей одной многочисленные частные характеристики, прижившиеся
в зоологической литературе, где отдельно называют дистанцию, активно поддерживаемую
между особями в стаде (индивидуальная дистанция), отдельно — охраняемое вокруг
движущейся особи пространство (передвижное пространство), отдельно — охраняемое
пространство вокруг оседлой особи (территория), хотя все три — лишь характеристики
единого механизма, препятствующего сбиванию в кучу. Индивидуальное пространство
либо перемещается вместе с владельцем,
либо оказывается закрепленным в какой-то точке местности. Кроме того, варьируют
величина владения, его проницаемость и степень скученности владений. То есть
для достаточно полного описания можно обойтись ограниченным количеством параметров,
комбинация которых даст любую конкретную конструкцию поселения.
Радиус индивидуального пространства и у гусаков, и у ушастых тюленей не превышает
десятка длин тела, хотя обычно гораздо меньше (1—2 длины). Он может меняться
в каких-то пределах у одной и той же особи, позволяя, например, после ухода
соседей по колонии оставшимся несколько расширить свои владения, так же как
и гусаку в момент перехода его гусыни к оседлости (до этого момента радиус
не превышает 10 м, после — может достигнуть 16 м).
Внутрь индивидуального пространства самца и у гусей, и у морских котов допускаются
лишь самки и сеголетки, но у морских львов к ним добавляются и субординатные
самцы более старших возрастов. Основным контингентом владельцев индивидуального
пространства и у гусей, и у отариид стали взрослые самцы в сезон размножения.
Но его можно найти и у самок (гусыня защищает пространство вокруг кладки и
выводка, но только в присутствии гусака, а котиха при благоприятных условиях
долго защищает пространство вокруг щенка в одиночку), и у детенышей (у щенков
— в играх со сверстниками, а у гусят — при защите родительского индивидуального
пространства от гусят других выводков).
Необходимость смыкать края индивидуальных пространств соседей можно предполагать
с большей уверенностью для морских котов, с меньшей — для морских львов и гусей
(но и здесь одиночное гнездование пары — скорее результат утраты прежде бывших
соседей).
И у гусей, и у отариид четко наблюдается фантастичная на первый взгляд ситуация
присутствия индивидуального пространства в отсутствие владельца, если он перед
уходом успел оставить достаточно следов в памяти соседей.
Напротив, владелец может обнаруживать свое индивидуальное пространство либо
только в присутствия определенной особи, за которой и следует неотрывно (гусак
с еще незагнездившейся гусыней, секач с одиночной самкой вне гаремного лежбища,
котиха со щенком) , либо даже в отсутствие полового партнера (секач до прихода
самок, ищущий место на будущем гаремном лежбище). А кроме того, возможны два
внешне похожих варианта: 1) владелец пространства привязан только к точке на
местности (гусыня у еще пустой гнездовой ямки и территориальный секач на лежбище)
и 2) владелец привязан к особи, закрепленной на местности (гусак при загнездившейся
гусыне, гусыня у кладки, котиха у только что родившегося и неспособного к перемещениям
щенка).
Можно еще больше упростить описание и построить таблицу, расположив по горизонтали
характеристики индивидуального пространства, а по вертикали — способы ограничения
перемещения владельца, и, получив в ее клетках возможные варианты, сопоставить
их с реализуемыми в наблюдаемой модели.
Из сравнения же гусиной и котовой моделей обнаруживается не только нехватка
прямых наблюдений (событий вне сезона размножения; индивидуальной, возрастной
и сезонной изменчивости поведения), но и возможность увидеть некоторые частные
и более общие закономерности.
Так, для понимания возникновения гнездового паразитизма у птиц (наиболее известна
обыкновенная кукушка) важна наблюдаемая в гусиной колонии ситуация, когда часть
пар оказывается незакрепленной на местности и гусыни либо бросают свои яйца
поблизости от чужих гнезд, либо с риском для жизни из-за активной обороны хозяевами
своего индивидуального пространства откладывают яйца в чужие гнезда, т. е.
практикуют гнездовой паразитизм внутри вида. Неясно, насколько это определяется
качествами гусыни и насколько — конкретной ситуацией в колонии.
Несколько иную окраску получает и такая, казалось бы, простая проблема, как
отличие полигама от моногама. Гусака-моногама и секача-полигама различают не
столько количество оплодотворенных за сезон самок (через территорию гусака
нередко проходит несколько гусынь, а многие секачи, долго просидевшие на гаремном
лежбище, так и не увидели самок на своих территориях), сколько привязанность
владельца индивидуального пространства к конкретной самке или конкретному месту.
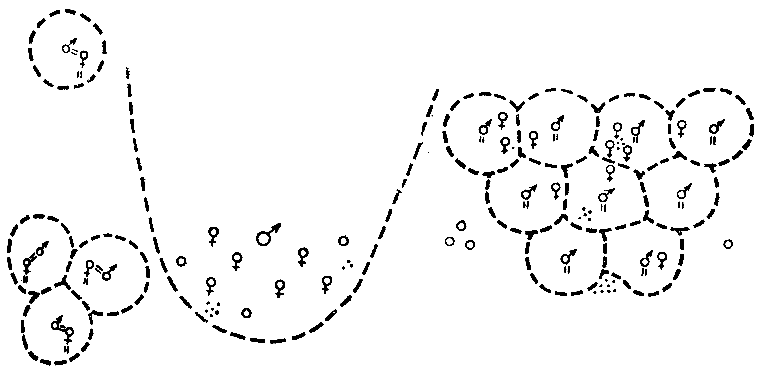
Рис. 22. Три схематизированные модели пространственного размещения особей
Слева — белые гуси, посредине — калифорнийские морские львы, справа — морские коты. Самки и территориальные самцы обозначены обычными для полов значками, субординатный самец — кружком, детеныши — точками, границы индивидуального владения очерчены прерывистой линией, а привязанность особи к другой особи или точке на местности показана короткими парными штрихами. Обращает внимание различный способ ограничения перемещений особи в гусиной и котовой моделях, а также большие размеры и неотчетливость части пограничной линии в индивидуальном владении калифорнийского морского льва
Но главное, что у таких далеких таксономически и биологически, как сравниваемые,
просматривается общая тенденция к уравновешиванию скучивания многих особей
на ограниченном участке проявлением у некоторых групп населения (здесь у взрослых
самцов) владения индивидуальными пространствами. Набор вариантов индивидуального
пространства и способов взаимодействия их владельцев достаточно ограничен,
чтобы поддаваться классификации (рис. 22). Особи, в норме не обнаруживающие
наличия индивидуального пространства (здесь самки и молодняк), сохраняют потенциальную
способность к его проявлению, а владельцы в разных ситуациях демонстрируют
разные варианты индивидуального пространства.
Так же и в пределах обоих таксонов потенциально возможны разные варианты индивидуальных
пространств, т. е. набор их общ, а употребляемость различных вариантов неодинакова
и норма для одного таксона оказывается редкостью в другом, В результате при
явном несходстве способов ограничения подвижности носителей индивидуального
пространства можно найти у морских котов типичные гусиные варианты (секач на
холостяковой лежке, привязанный к избранной самке и неотвязно следующий за
ней), а у гусей — типично котовые, когда через закрепленное на местности индивидуальное
пространство гусака проходят свободно перемещающиеся по местности самки (хоть
и в сопровождении собственных гусаков).
Обе колонии сходны и тем, что собраны из индивидуальных пространств взрослых
самцов и недоступны для всех, кроме полового партнера владельца и последнего
приплода. Различие же моделей — в противоположных способах закрепления владельцев
индивидуальных пространств на местности, разной степени их скученности (индивидуальные
пространства смыкаются краями у котов и более дисперсны у гусей) и в большей
отчетливости намеков на владение индивидуальными пространствами у гусят и гусынь
по сравнению с котовыми щенками и котихами.
Итак, на двух половинках рисунка 21 — похожие постройки, но сделанные из разного
строительного материала. Набор стройматериалов небезграничен, и, так же как
пропорции черепа ленивца повторяют пропорции черепа ушастого тюленя, так и
строение котового лежбища оказывается и в целом, и в деталях повторяющим скопление
собравшихся для размножения североамериканских лягушек-быков, с разделением
участка между территориальными самцами с последующим прохождением через их
сплоченные индивидуальные пространства позднее приходящих готовых к икрометанию
самок.
Сравнение колоний разных групп животных помогает увидеть общие для всех законы
и специфику каждого варианта, но не восполняет недостаток прямых наблюдений.
А даже для наиболее изученных северных морских котов неизвестно, где и как
включаются в структуру лежбища впервые размножающиеся самцы и самки, чем определяется
интенсивность обмена особями внутри лежбища и между соседними лежбищами, как
влияет более чем вековой выборочный выбой молодых самцов на окраинах лежбищ
на расселение зверей и структуру лежбищ. Без этих знаний приходится постоянно
сталкиваться с несовпадением реального положения в стаде с прогнозами.
Изучение пространственного распределения животных (да и растений) долгое время
проводилось лишь на видовом уровне, отмечая наличие или отсутствие данного
вида в том или ином месте и его численность. Позже оно стало различать отдельные
популяции, составляющие видовое население, а в последние десятилетия неизбежно
распространилось и на события, происходящие с отдельными особями.
Каждая особь на протяжении своей жизни определенным образом перемещается в
пространстве, формируя тем самым и структуру своего вида, и структуры включающих
его многовидовых сообществ. Достаточно очевидно, что, не зная характера взаимного
размещения и подвижности именно отдельных особей, нельзя сколько-нибудь обоснованно
говорить ни о продуктивности угодий, ни об эффективности природоохранных мероприятий.
Не вызывает сомнений, что, говоря о территориальных самцах, гаремах и скоплениях
гаремов у кашалотов, морских слонов и морских котов (так же как при сравнении
морских котов и ближайших их родственников калифорнийских морских львов), мы
общими словами называем разные вещи и обсуждаем различные системы землепользования.
Но, как показывают гусаки и секачи, при самых разных системах землепользования
можно пользоваться общей меркой и ограниченным набором параметров для достаточно
полного описания пространственного распределения особей. Для некоторых из характеристик
видна их взаимозависимость.
Так, при относительно большом поперечнике индивидуального пространства внутрь
допускается широкий набор особей, включающий, кроме полового партнера, неполовозрелых
особей и субординатных взрослых одного пола с владельцем. При сокращении же
поперечника уменьшается и терпимость к вторженцам, в крайнем варианте допускается
лишь краткое пребывание полового партнера. Владеть индивидуальным пространством
могут либо все особи данного вида на протяжении всей жизни, либо только взрослые
обоего пола, либо только часть взрослых одного из полов на протяжении незначительной
части года. Максимальная скученность сопряжена с терпимостью (если не с влечением)
к присутствию соседа того же ранга настолько близко, насколько позволяют сомкнувшиеся
края индивидуальных пространств, минимальная — предполагает активное избегание
контактов с какими-либо следами присутствия соседа.
Большая привязанность к местности, чем к значимой особи своего вида, определяет
необходимость привязать дом к определенной точке земной поверхности, большая
привязанность к значимой особи — и дом в любом месте, где эта избранная особь.
И та и другая привязка (иногда меняясь в жизни одной особи) не позволяют владельцу
унести свое индивидуальное пространство неизвестно куда.
Наличие индивидуального пространства с той или иной комбинацией перечисленных
характеристик легко прослеживается в опубликованных описаниях распределения
особей в многочисленных группах морских и наземных позвоночных и беспозвоночных
животных, не говоря уже о млекопитающих. Так, владеющего небольшим индивидуальным
пространством самца, следующего за одной и той же самкой и допускающего внутрь
своего пространства последний и предпоследний ее приплод, легко увидеть в описанных
Георгом Стеллером — единственным натуралистом, видевшим это,— скоплениях командорской
морской коровы. Морской слон (взрослый самец) владеет большим индивидуальным
пространством, внутрь которого допускаются даже субординатные взрослые самцы,
довольно свободно перемещается по побережью от одной группы неоплодотворенных
самок к другой. Секач же калифорнийского морского льва при сходных характеристиках
индивидуального пространства скован привязанностью к определенной точке побережья.
А секач морского кота при малых размерах индивидуального пространства, доступного
лишь самкам и новорожденным щенкам, скован еще взаимоотношениями между соседними
владельцами. Большие стада дельфинов, судя по описаниям, распадаются на отдельные
подвижные группы, состоящие из территориального самца, нескольких самок, полувзрослых
самцов и детенышей, близко соответствуя модели павианов.
Эти модели, не исчерпывая и не упраздняя поведенческой специфики, позволяют
увидеть общую геометрию в построении населения разных видов из отдельных особей.
Широкий же набор групп многоклеточных животных с наличием свидетельств об индивидуальных
пространствах особей заставляет поискать аналогии и в других сообществах. Но
в первую очередь интересны лишенные полового размножения протисты и сидячие
колониальные животные. Есть ли индивидуальное пространство при отсутствии полового
размножения и как в случае его наличия образуется колония сальп и кораллов?
Затем: так ли уж далека от зоологических проблем с пространственным распределением
особей проблема пространственных структур растущей колонии бактерий, если бактериологи
считают систематическим признаком внешний вид колонии на питательной среде?
Сидячие животные ставят вопрос о совместимости наличия индивидуального пространства
с отсутствием активного перемещения особи, подводя к рассматриваемому кругу
многоклеточные растения. Чем достигается рассредоточение экземпляров растений
при слишком плотном высеве? И, наконец, неизбежно вовлекается в рассмотрение
тот вид приматов, к которому мы принадлежим, несмотря на наши качественные
отличия, и чья высшая нервная деятельность не исключает безусловных рефлексов.
Интерес территориальных аспектов человеческого поведения вполне очевиден, но
не менее интересным представляется попытка пространственного истолкования архаичных
систем исчисления родства, проводящих границу между доступными и недоступными
соплеменниками. С другой стороны, качественная специфика человеческой природы
позволяет ставить вопрос о пространственном структурировании не только в географическом
пространстве, но и в том, где разворачивается специфическая человеческая творческая
активность.
Частный вопрос о пространственном распределении особей ушастых тюленей и его
закономерностях так же быстро выводит обсуждение на широкий круг актуальных
проблем, как и рассмотрение частных проблем изменчивости плана строения тела
ушастого тюленя. Но определение применимости найденного к другим группам организмов
не исчерпывает неизбежных вопросов.
Во-первых, встает вопрос о природе сил, распределяющих особи в пространстве,
стягивая их в скопления и разгоняя в стороны. И рассеивание, и скучивание не
совсем хаотичны. Рассеивание вызывается как ослаблением привязанности особи
к месту своего рождения (это ослабление географических привязок может быть
и однократным, и многократно повторяющимся на протяжении жизни), так и избеганием
контактов с другими особями своего вида, т. е. следствием некоторого взаимного
отталкивания. Скучивание же зависит как от стремления многих особей занять
одну географическую точку независимо от наличия пли отсутствия там соплеменников
(например, стремление вернуться в точку рождения), так и от влечения особей
к соплеменникам, что ведет не только к встречам половых партнеров, но и к сбиванию
в кучу однополых зверей на лежбище или членов одной стаи рыб или саранчи. Это
влечение гонит к соплеменникам независимо от локализации и перемещений последних.
То есть положение особи в пространстве будет определяться действием на нее
двух независимых пар сил: притягивание к определенной географической точке—собственная
подвижность особи и притягивание к определенной особи (или группе особей)—отталкивание
от особей своего вида. Сфера вокруг особи — индивидуальное пространство — своими
размерами и проницаемостью для соплеменников будет менять результаты взаимодействия
перечисленных четырех сил. Хорошо бы, конечно, иметь возможность измерять тягу
к скитаниям и тягу к возвращению домой, стремление изолироваться от соплеменников
и стремление контактировать с ними.
С другой стороны, взаимодействие этих сил формирует сами характеристики индивидуального
пространства: наличие или отсутствие охраняемой зоны определяется наличием
или отсутствием взаимного отталкивания особей; диаметр охраняемого пространства—
степенью отталкивания; набор допускаемых внутрь пространства особей — набором
неподверженных отталкиванию особей; необходимость контакта с соседними владельцами
индивидуальных пространств — степенью взаимного притягивания особей определенной
категории; привязка к точке на местности или к определенной особи — степенью
преобладания тяги к одному над тягой к другому; степень подвижности владельца
индивидуального пространства — результатом взаимодействия хоминга1 и трех
остальных сил, да и значимость даже пустого места ушедшего соседа — мера инерционности
отталкивания от соседей. То есть характеристиками
индивидуального пространства можно косвенно померить четыре действующие на
данную особь в данный момент силы.
1 Хоминг — стремление особи к дому. |
Поскольку понятно, что реакция на любой раздражитель происходит лишь при приближении
его на какое-то пороговое расстояние, то в чем причина наличия именно этого,
а не другого, порогового расстояния? Очень заманчива аналогия с биологическими
полями клеток, определяющими их пространственное размещение, а также возможная
сопричастность наличия индивидуального пространства особи и фундаментального
отличия клетки как части организма от особи как части ценоза. Во всяком случае,
понятие поля все чаще применяется при описании агрессивных и коммуникационных
взаимодействий особей (вспомним сигнальные поля в описаниях коммуникации зверей
Н. П. Наумовым).
Наличие индивидуального пространства позволяет двигаться по крайней мере в
трех направлениях.
1. В меру возможностей организма и наличной модели индивидуального пространства
можно укреплять и усложнять владение, создавая некоторое подобие наружного
скелета из нор, гнезд, убежищ и системы сигнальных меток.
2. Дальше на основании характерного для данного вида способа оперирования с
индивидуальным пространством при сохранении наряду с ним добавочных, редких
и краткосрочных вариантов (основных для других таксонов) можно строить из них
замкнутые системы — популяции, заполняя ими все доступное особям данного вида
пространство. Сейчас основными, по крайней мере для позвоночных животных, представляются
три модели (см. рис. 22): скопление одноранговых владельцев индивидуальных
пространств, свободных от субординатных особей; раздутое индивидуальное пространство
владельца, заполненное разнородными подчиненными ему особями; переносное индивидуальное
пространство, привязанное запечатленном к особи своего (а иногда чужого) вида,
перемещение которой и определяет перемещения владельца. Число реализуемых моделей
разумеется больше трех, но существенно меньше количества таксонов, что ведет
к частому использованию систематически далекими формами организмов общей модели
пространственного размещения особей.
3. Если принять за исходное сообщество, состоящее из особей, владеющих индивидуальными
пространствами и равных по рангу, то возможен либо путь к установлению иерархии
среди владельцев при росте владений у доминантов с поглощением владений субординатов,
либо путь к установлению сопряженности собственных перемещений с перемещениями
других особей. Оба пути, вероятно, осуществлялись многократно и независимо
в разных ветвях и веточках эволюционного древа.
Популяция, таким образом, оказывается не просто скопищем свободно скрещивающихся
особей в кольце непригодных или неприятных мест, изолирующих от других таких
же скопищ, а собранием жестко регламентированных структурой связей между особями
участников единого сообщества, изолированного от других таких сообществ не
столько географией, сколько трудностью внедрения в чужую постройку.
Соответственно для белых гусей репродуктивно изолированным от соседей комплексом
будет место зимовки, где формируются брачные пары и комплекс (иногда сокращающийся
до одной) принимающих их гнездовых колоний, где могут бок о бок находиться
пары с разных зимовок. То есть гнездовая колония окажется конгломератом репродуктивно
изолированных кусков, как было недавно показано для врангелевской колонии Е.
В. Сыроечковским. Для морских же котов картина будет прямо противоположной.
И центром, перемешивающим партнеров, окажется именно колония — лежбище, сохраняющее
способность извлекать свой контингент из удаленных друг от друга точек океана
и одновременно могущее быть надежно изолированным от расположенного в нескольких
километрах побережья соседнего лежбища.
Единое поселение членов одного вида может представлять конгломерат репродуктивно
изолированных кусков. В то же время конгломерат разных видов, привязанных к
общей географической точке, несмотря на специфику их видовых экологических
ниш, может быть конструктивно единым.
Зависимость пространственных структур вида-хищника от системы землепользования
у особей кормового объекта широко известна. Однако хищник не только деформирует
пространственное распределение особей кормового объекта, по и обеспечивает,
как показывает пример белой совы и белого гуся, начальные этапы колонизации
другим видам. Взаимное проникновение пространственных структур, нескольких
даже не связанных пищевой цепью видов (многовидовые лежбища, многовидовые стаи)
дает разнообразные пространственные композиции. То есть внутривидовые закономерности
пространственного размещения особей формируют те шипы и пазы, которые сшивают
отдельные, даже не едящие друг друга виды в единую многовидовую постройку.
Вот и снова частный эпизод с отариидным лежбищем помог увидеть один из общих
интегрирующих механизмов надындивидуальных систем.
Различные системы землепользования при разных моделях пространственного размещения
особей дают неодинаковый ответ на неблагоприятные воздействия. Из трех подробно
рассмотренных моделей наиболее защищенное детство будет обеспечивать гусиный
вариант, но он же будет давать наибольшую нагрузку на самок. Котовый вариант
кладет тяжкое бремя на территориальных самцов и оставляет приплод почти беззащитным,
но не слишком перегружает самок. И вариант калифорнийского морского льва секачам
дает меньшую нагрузку, чем котовый, но большую, чем гусиный, давление на самок
меньше, чем в двух других, щенкам же легче, чем в котовом, но тяжелее, чем
в гусином варианте.
Соответственно демографические пирамиды будут терпеть больше ущерба снизу (щенки)
и сверху (секачи) в котовом варианте, нежели в двух других. Наложение добавочной
нагрузки ухудшением кормовых или климатических условий, по-видимому, сильней
ударит по секачам и щенкам котового варианта, резко увеличив их смертность
по сравнению с другими группами населения, такое же увеличение смертности при
гусином варианте более равномерно распределится между всеми группами населения.
Что касается хищников, то для тех, кому доступны лишь детеныши, котовая модель
дает больше возможностей поживы, чем гусиная, для тех же, кому доступны взрослые,
будут привлекательны центральные наиболее плотно заселенные зоны скоплений
территориальных самцов любого из трех типов.
При всех этих напастях урон демографический (потери в определенном поколении
и потери в определенных возрастных и половых группах стада) будет сопровождаться
менее очевидным уроном для остающихся в живых в условиях сообщества с нарушенной
структурой. К чему ведет нарушенная структура пространственного размещения
особей и регламентированных контактов между особями, показывают, например,
наблюдения над обитателями зоопарков Десмонда Моррисса. Неспособность вступать
в нормальные контакты с особями своего вида, неспособность выходить потомство
и убийство собственных детенышей, неспособность сохранить собственную жизнь
оказываются следствиями лишения хорошо ухоженных обитателей зверинца всего
лишь возможности свободно перемещаться и в нужное время вступать в контакт
с нужными особями своего вида.
Если теперь повреждаемость структур в трех наших моделях попробовать оценить,
то придется признать, что гусиный вариант с его мелкими ячейками (пара взрослых
и последний приплод) позволяет скорее разбить колонию на составляющие ее ячейки
и уничтожить каждую поодиночке, чем деформировать отдельную единицу. Вероятно,
наиболее разрушительным для внутренних связей здесь будет увеличение диспропорции
между смогшими осесть в колонии парами и парами, лишенными этой возможности,
что ведет к патологическому росту числа подкидышей с последующим ростом гибели
всего приплода.
В противоположность гусиному котовый вариант из-за сравнительной легкости повышения
выборочной гибели щенков и территориальных секачей будет сохранять привязку
к месту колонии, когда уж нет секачей, обеспечивающих структурирование населения
(а перед этой последней стадией будет держать структуру недостаточным количеством
секачей), превращая колонию в бесформенное скопление, если не свалку, разнополого
и разновозрастного зверя, не способного исполнять свои роли.
Вариант калифорнийского морского льва должен запять промежуточное положение
и по повреждаемости, и по способности к компенсаторным перестройкам между котовой
и гусиной моделями. Таким образом, неблагоприятные воздействия в зависимости
от того, на какой вариант структуры они действуют, могут но только влиять па
численность данного вида, но и продуцировать больные сообщества. То есть рациональное
использование ресурсов должно предполагать не только слежение за динамикой
освоенных данным видом животных площадей и за динамикой его численности, но
и за сохранением (или несохранением) структурных связей между особями. Соответственно
оценка эффективности промысла, особенно если он выборочно захватывает лишь
одну группу видового населения (как выборочный промысел молодых самцов северного
морского кота), должна идти с учетом и его влияния на структуру поселений.
А мероприятия по улучшению ресурсов должны включать не только хирургические
мероприятия по отрезанию, консервации и пересадке кусков ткани видового населения
(акклиматизация, реакклиматизация и удаление ненужных поселений), но и терапевтические
— по восстановлению нормально работающих структурных связей между особями поселения.
Попытка соотнести устройство особи, устройство ее жизненного пути и устройство
того, что из отдельных особей строится, и поискать нечто целостное в событиях,
происходящих на разных уровнях, трудна. При ставшем, к несчастью, традиционным
обособлении специалистов, обследующих разные уровни организации (жизнь и устройство
зародыша, взрослого, популяции, ареала, биосферы), переход с уровня на уровень
оказывается ничьей землей. Попадая в эту нейтральную полосу, исследователь
рискует оказаться чужеродным и для коллег слева, и для коллег справа.
В то же время вроде бы ясно, что события верхнего уровня накладывают некоторые
ограничения на события на нижнем, а то, что происходит на нижнем уровне, строит
верхние этажи. Отсюда — двойственность объектов, которые, с одной стороны,
самостоятельны и независимы в поведении, а с другой — отчетливо обнаруживают
скованность рамками и вовлеченность в построение целого следующего порядка.
На разном материале и на разных уровнях наряду с многочисленными факторами,
определяющими широкую индивидуальную и групповую изменчивость хода онтогенеза,
постоянно обнаруживается действие противоположных сил, налагающих на изменчивость
достаточно жесткие рамки, ограничивающие набор вариантов как в онтогенезе,
так и в филогенезе.
То, что для каждой конкретной формы организмов размах сегодняшней изменчивости
ограничен видовыми рамками, выглядит банальным. Но представляется перспективным
для понимания исторических преобразований попытаться понять механизмы, удерживающие
муху Fannia scalaris, известную из эоценовых балтийских янтарей и широко распространенную
сегодня, в одних и тех же видовых рамках на протяжении по крайней мере 50 миллионов
лет (т. е. одно и то же воспроизводится из поколения в поколение не меньше
50 миллионов раз). Известная долговечность таксонов млекопитающих существенно
меньше. И хотя о видовой принадлежности зуба или куска звериной челюсти судить
труднее, чем о насекомом в янтаре, возможно, ход онтогенеза в нашем классе
более подвержен изменениям.
Целостность особи аргументировать не нужно. Однако и сохранение во взрослом
черепе следов расчлененности по продольной оси на крупные блоки (метамеры),
нормальной для ранних зародышей, не столь очевидно, но все же выявляется довольно
простым графическим способом. То есть то, что, действуя в жестких рамках осуществления
наследственной программы, обособляет друг от друга массы множащихся клеток,
замыкая их в блоках-метамерах, продолжает до старости зверя держать границы
между блоками несглаженными и обеспечивать некоторую независимость событий
в разных, даже соседних, блоках.
События с блочной особью разворачиваются по крайней мере на двух различных
полигонах: там, где происходят таксономические превращения с появлением и расцветом
новых таксонов, и в географическом пространстве, где формируются и живут совокупности
особей данного вида.
На таксономическом поле индивидуальные варианты распределяются не равномерно,
а скапливаются группами, и из всей многоступенчатой, детально разработанной
систематиками иерархии особую значимость обнаруживает ступень, обозначаемая
обычно как семейство. Рамки, замыкающие формы одного семейства, по-видимому,
не растягиваются с появлением каждой новой модели, а, напротив, лимитируют
число членов семейства. Во всяком случае, размах изменчивости пропорций черепа
в пределах таксона не имеет прямой зависимости от числа входящих в него таксонов
следующего порядка. Локальность отличий пропорций при переходе от семейства
к семейству позволяет лишь несколькими надстройками превратить череп ушастого
тюленя в череп представителя другого семейства — моржа (см. рис. 8). Локальные
преобразования лишь предглазничной области черепа обеспечивают большое разнообразие
среди хоботных. В то же время локальные выросты громадных оленьих рогов не
нуждаются в сопряженных перестройках, а рогатые и безрогие формы легко умещаются
в рамках одного семейства. К тому же размах отличий форм одного семейства не
превосходит размаха индивидуальной внутривидовой изменчивости любого из членов
семейства.
Все эти неожиданности складываются в схему некоторой единой совокупности (семейства),
в пределах которой различия между рангом членов не столь существенны, а число
членов небезгранично. Выход за пределы этих рамок требует, по-видимому, принципиально
отличных перестроек по сравнению с событиями внутри рамок. Среди нескольких
сотен семейств млекопитающих, доживших и не доживших до нашего времени, имеют
место как «профессиональные союзы» (крупные и мелкие травоядные, крупные и
мелкие ловцы мелкой подвижной добычи, крупные и мелкие собиратели малоподвижной
добычи и т. д.), так и объединения владельцев одинакового имущества (общая
модель черепа у кита и утконоса или у морского кота и ленивца, общая модель
зуба у зубатых китов и неполнозубых, общая модель конечностей у литоптерн и
лошадей). Представить себе временную развертку событий на этом поле, увидеть
всю крону зеленеющего древа жизней, образом которого (des Lebens goldner Baum)
гетевский Мефистофель дразнил школяра, мешает и темнота геологической летописи,
и невозможность отойти от нее.
Одновременно с жизнью на таксономическом поле блочная особь живет на доступной
прямым наблюдениям географической поверхности. И здесь как раз временная развертка
вполне доступна. Основные вехи жизни (рождение, отлучение от матери, фазы физиологической
и социальной зрелости, срок окончания жизни) особей каждого вида представляются
обычно зависящими в первую очередь от комплекса сиюминутных факторов, меняющихся
в широких пределах. В то же время все эти события, отмечая созревание и включение
в работу различных систем развивающегося организма (предмет разработок П. К.
Анохина и его школы), являются существенными характеристиками онтогенеза, а
перемены в расположении этих вех обеспечивают филогенетические приобретения
и утраты. На зыбкой почве хронологии, оказывается, тоже существуют достаточно
четкие границы, и выбор возможен лишь между ограниченным числом моделей с чередованием
жестких и гибких звеньев в каждой. Ни разница в размерах зверя, ни диаметрально
противоположная биология, ни филогенетическая дистанция не мешают зубру и морскому
коту делить общую хронологическую модель. Так же как сходство по этим признакам
медведя и морского кота не понуждает их следовать общей модели.
Фиксированные вехи на жизненном пути в рамках любой модели легко увязываются
с переходом особи на новое место в системе (зародыш—мать, мать—младенец—сообщество,
особь—сообщество—живое и неживое окружение сообщества), что после появления
особи на свет с каждым новым сезоном в жизни все больше связывается и с перемещениями
(или избеганием их) по земной поверхности. А эти перемещения нормируются контактами
особи с ближайшими соседями, что, в свою очередь, определяется наличием и характеристиками
индивидуального пространства вокруг особи, куда не допускаются соплеменники
одного с ней ранга.
Получается, что тело животного вписано в целостную постройку (той или другой
из некоторого числа моделей), как рак-отшельник в свою раковину, которую можно
на время снять, можно заменить на раковину другой модели, но без которой нельзя
обойтись. Характеристики этой внешней по отношению к особи постройки, направляющие
агрессивность и коммуникабельность особи, изменчивы и на протяжении жизни особи,
и в разных ветвях филогенетического древа, где каждая из моделей постройки
многократно и независимо вводится в употребление, так как число моделей существенно
меньше числа таксонов.
Таким образом, вместо совокупности особей получается совокупность построек,
нормирующих и структуру скоплений особей одного вида и образование конгломератов
особей разных видов, объединяемых географией. То есть здесь онтогенетические
события выходят на два уровня: система своих и система, объединяющая своих
с чужими.
Оперируя на уровне особи, приходишь к выводу, что все три рассмотренных типа
структур (анатомические, хронологические и пространственных связей особи с
соплеменниками) в своих перестройках следуют ограниченному числу моделей, большему
для анатомических структур и меньшему для структур индивидуального пространства.
Разнообразие увеличивается локальностью перестроек в рамках единой конструкции,
которая представляется системой блоков с относительной независимостью событий
в каждом из них, даже если внешне они столь же консолидированы, как в черепе
взрослого млекопитающего. Блочность позволяет конструкции сохранять одновременно
гибкость и жесткость при разновременности изменений в разных блоках. Представляется
перспективным пытаться определить потенциальные возможности конструкции и характер
присущих ей ограничений, что позволило бы вычислять возможные, но не реализуемые
п данный момент формы (и анатомии, и жизненного пути, и надындивидуальных структур).
Переход к надындивидуальным структурам нормируется возможностями анатомических
структур особи и возможностями структур межиндивидуальных связей. Анатомия
зверя определяет набор приемлемых для него характеристик неживого окружения,
возможности перемещения в пространстве, набор доступных кормовых объектов и
способ (и возможности) противостояния врагам. А следовательно, налагает рамки
на его расселение, очерчивая потенциально пригодные для него территории или
акватории, лимитируя одновременно плотность населения и возможности контактов
между особями. Но контакты между особями дополняют или урезают спектр определенных
анатомией пригодных условий, так как скопление может и согреть особь, там где
в одиночку она уже не согреется, и перегреть ее там, где в одиночку она еще
не перегреется, может сделать доступной недоступную для одного добычу и лишить
доступной для одиночки пищи. А контакты между особями соплеменников зависят
от анатомических возможностей и географии, складываются из структур индивидуального
пространства. Индивидуальные пространственные блоки, сочетаясь в блоки следующего
порядка, строят замкнутые стабильные или меняющие объем, открытые для вселенцев
(или выселенцев) поселения, нормированные следованием определенной структурной
модели, и заполняют ими доступное пространство.
Потенциально пригодное для всего единого комплекса соплеменников место в системе
таких же комплексов организмов других видов, экологическая ниша данного вида,
очерчивается анатомией особи, хронологией онтогенеза и структурой межиндивидуальных
связей. Все три составные представляют собой системы жестких и гибких звеньев,
нормированные каждая своими жесткими рамками. И экологическая ниша, заключая
все события, происходящие с соплеменниками, налагает прочную узду на строение
особи и ее онтогенетические преобразования, и на межиндивидуальные связи. В
пределах ниши живет, однако, не особь, а сотканная из особей живая ткань вида,
стремящаяся, если она здорова, расползтись на все доступное пространство, от
одного непреодолимого барьера до другого.
В принципе любая особь крупного подвижного зверя может в пределах соответствующей
экологической ниши попасть в любую точку и оставить там потомство, если, конечно,
порвет связь с местом рождения и его населением, сумеет добраться до этой точки,
найти там полового партнера и вписаться в структуру тамошнего населения. Однако
и для особи, не прикованной на всю жизнь к своему локальному блоку населения,
перемещения нормируются двойным подчинением: особь—экологическая ниша и особь—структура
видового населения. Двуединая конструкция — экологическая ниша и живущий внутри
нее вид — стоит, в свою очередь, на двух якорях: многомерную нишу держат соседние
такие же образования и связь с географией, а конструкцию из особей одного вида
— тоже привязка к географии и образование конгломератных поселений с другими
топографически связанными видами. То есть прослеживаются в многоступенчатой
иерархии систем от особи до биосферы как регулирующее влияние биосферы через
видовую экологическую нишу и структуру видового населения на особь, так и обратное
воздействие особи и ее индивидуальной жизни наверх, по крайней мере до уровня
вида и его экологической ниши.
В итоге проблема нормировки онтогенеза оказывается одной из сторон проблемы
построения из частей целостной постройки и вопроса о целом как о многоступенчатой
иерархии соподчиненных систем с прямыми и обратными связями между уровнями.
Оторвавшись от особи ушастого тюленя, перейдем к их видам, начав с неизбежной
инвентаризационной проблемы — сколько видов ушастых тюленей живет на земном
шаре.
Вопрос о том, сколько же видов и как они очерчены в пределах исследуемой группы
(любых организмов), неизбежно встает при первых же попытках упорядочить наличный
материал. Скоро обнаруживается, что уже двум долго и независимо работающим
исследователям трудно получить сходные ответы, даже если группа мала, хорошо
изучена и четко очерчена.
Систематика набрала силу уже достаточно давно, и многие виды растений и животных
благополучно живут в тех рамках, которые им определил в позапрошлом веке Карл
Линней. А для ушастых тюленей крупные размеры, расселение по трем океанам,
удобство для наблюдения и интерес как к промысловым объектам создают, казалось
бы, максимально благоприятные условия для однозначной интерпретации разнообразия
и взаимоотношений членов семейства (рис. 23). И тут трудно удержаться от того,
чтобы привести рядом пять вариантов системы отариид (см. таблицу), четыре из
которых почти одновременны.
Соотношение видов ушастых тюленей в пяти вариантах системы
Gray, 1874 |
Siversten, 1954 |
Scheffer, 1958 |
King, 1964 |
Reppening et al., 1971 |
| Euotaria compressa Е. schisthyperoes |
Ar. pusillus |
Ar. pusillus |
Ar. pusillus |
Ar. (1) pusillus |
| Ar. tasmanicus | ||||
| Eu. cinerea | Ar. forsteri | Ar. forsteri | Ar. forsteri | Ar. (2) forsteri |
| Gypsophoca tropicalis Ar. antarcticus |
Ar. gazella | Ar. gazella | Ar. tropicalis | Ar. (3) gazella Ar. (4) tropicalis |
| Euotaria nigriscens Eu. latirostris |
Ar. australis | Ar. australis | Ar. australis | Ar. (5) australis |
| Ar. galapagoensis Arctophoca townsendi Arctophoca philippii |
Ar. philippii | Ar. philipii | Ar. (6) galapagoensis Ar. (7) townsendi Ar. (8) philippii |
|
| Callorhinus ursinus | C. ursinus | C. ursinus | С. (9) ursinus | |
| Zalophus gilliespii |
Z. californianus Zalophus wollebaeki |
Z. californianus |
Z. (10) californianus |
|
| Phocarctos elongatus Eumetopias stelleri | Eumetopias jubatus |
Eumetopias jubatus |
Eumetopias (11) jubatus |
|
| Otaria jubata Otaria minor Otaria ulloae |
Otaria byronia | Otaria byronia | Otaria (12) byronia | |
| Neophoca lobata Phocarctos hookeri | Neophoca cinerea |
Neophoca cinerea Neophoca hookeri |
Neophoca (13) cinerea Phocarctos (14) hookeri |
Рис. 23. Размещение сегодняшних береговых лежбищ 14 форм, образующих семейство ушастых тюленей (по Дж. Кинг, 1964, с изменениями)
Номера соответствуют тем же формам в табл. 1: 1 — южноафриканский морской кот, 2 — новозеландский морской кот, 3 — антарктический морской кот, 4 — субантарктический морской кот, 5 — южноамериканский морской кот, 6 — галапагосский морской кот, 7 — гвадалупский морской кот, 8 — хуан-фернандесский морской кот, 9 — северный морской кот, 10 — калифорнийский морской лев, 11 — сивуч, 12 — южноамериканский морской лев, 13 — австралийский морской лев, 14 — новозеландский морской лев
Удобно было бы отнести противоречия за счет недостаточности морфологического
материала от мелких, поставленных на грань уничтожения локальных стад. Так,
одно время опытные систематики находили достаточно признаков для обособления
в отдельные виды трех стад северных морских котов (курильского, командорского
и прибыловского). По мере роста стад и накопления сведений об обменах между
ними исчезали сомнения в принадлежности их к единой форме Callorhinus ursinus.
Исчезли в новом материале и многие формы, вычлененные более ста лет назад Грэем.
На нехватку материала можно списать чередование слияния и расчленения новозеландских
и австралийских морских львов (у Джудит Кинг они в разных родах, у В. Шеффера
— в двух видах единого рода, а у Э. Сиверстен они просто составляют единый
вид) или гвадалупских, галапагосских и хуан-фернандесских морских котов, которые
у Кинг образуют единый вид, а у Сиверстен — три вида двух разных родов. Но
об ином говорит неожиданное признание единым видом легко доступных, многочисленных
и, казалось, хорошо изученных и надежно разобщенных австралийских и южноафриканских
морских котов.
Выходит, что обилие материала не спасает от диаметрально противоположных выводов
при наличии нескольких интерпретаторов. Более того, обилие материала как будто
понуждает одного и того же автора менять истолкование с течением времени. Кроме
приуроченности таксономических неясностей к небольшим островкам и группам островков,
таблица (порядковый номер вида на рис. 23 и таблице совпадает) показывает и
смену волн дробительства и укрупнительства. На смену дробителю Дж. Грэю приходит
укрупнитель В. Шеффер, а его почти сразу сменяет неодробитель Ч. Рененинг.
Причем эти волны захватывают как будто не только самих создателей систем. Можно
довольно уверенно ожидать прихода следующей волны укрупнительства, но было
бы заманчиво запастись к ее приходу дополнительной аргументацией, помимо обычной
интуитивной оценки значимости того или иного признака и достаточности локальных
отличий для придания им видовой специфики. Злободневность проблемы усиливается
тем, что для многих малочисленных стад ближайшее время не принесет заметного
увеличения численности, а многочисленные стада не сегодня завтра легко могут
потерять это свое преимущество.
Без повидовых кадастров ни о рациональном использовании ресурсов, ни о составлении
Красных книг говорить не приходится, так что вопрос о числе и границах видов
ушастых тюленей или любого другого семейства, а главное, о том, как их надежно
сосчитать, не только любопытен. В этих условиях хорошо бы вначале рассмотреть
характеристики самих ящиков, по которым мы раскладываем наличный материал,
а не полагать эти ящики, как принято на практике, безразмерными. Если реальна
дискретность совокупностей особей со сходными характеристиками, отделяющая
такие совокупности друг от друга, то столь же реально и единство построенной
из этих дискретных единиц земной биосферы. Условиями стабильности совокупности
особей должны быть тогда постоянное и четко очерченное место ее в системе (экологическая
ниша), некоторый общий набор признаков конструкции и отсутствие размывания
и помех при передаче генома от поколения к поколению. То есть там, где совпадают
дискретная морфологическая единица с дискретной экологической нишей и с дискретной
совокупностью потенциально скрещиваемых друг с другом, но репродуктивно изолированных
(не только сейчас, но и потенциально) от не входящих в совокупность особей,
мы получим стабильную на протяжении ряда поколений совокупность, надежно изолированную
от других таких же совокупностей — видов. Попробуем теперь примерить эти простые
соображения к наличному материалу об ушастых тюленях и посмотреть на вытекающие
возможности.
Дискретность вариантов конструкции довольно хорошо выявляется при сравнении
пропорций черепов, показывая одновременно и жесткость рамок, заключающих изменчивость
в пределах таксона как всего семейства, так и каждого из семи родов системы
Джудит Кинг. Неясной остается ситуация внутри рода Arctocephalus из-за отсутствия
соответствующего материала. Диагностика по особенностям зубной системы малоубедительна,
пока не доказано, что принимаемые за видоспецифичные варианты отсутствуют в
пределах нормальной изменчивости соседних видов. Не хватает материала по анатомии
ушной области черепа, а уж о привлечении для опознания видовых отличий строения
чего-либо в теле, кроме черепа, все еще не может быть и речи из-за отсутствия
знаний об изменчивости. Тем не менее обнаружение блочности конструкции и соответствующей
локальности возможных перестроек, а также предположение наличия конструктивно
запретных вариантов позволяют надеяться на выявление морфологической дискретности
у отариид даже на сегодняшнем материале. Однако работа крайне трудоемка и почти
не начата.
Дискретная экологическая ниша, определяющая место ее обитателей в системе биосферы,
труднее всего поддается очерчиванию. Но несомненно, что именно ее проекцией
является потенциальный ареал вида. А даже с изучением потенциального ареала
дело обстоит настолько плохо (хотя и с реализуемым ареалом много неясностей),
что все изменения, происходящие с сегодняшним ареалом на наших глазах, оказываются
неожиданными. Достаточно назвать сенсацию, вызванную обнаружением колонии прибыловских
котов на калифорнийском острове Сан-Мигель, и удивление от вселения калифорнийских
морских львов, сбежавших из различных зверинцев, в биоту атлантического побережья
Северной Америки.
На другие параметры ниши будут указывать энергетика обитателей (траты на пребывание
в воде и на суше, траты на поиск и сбор корма, общая стратегия трат и приобретений)
и набор связей их с другими членами ценоза. И здесь разграничить эти характеристики,
хотя бы для двух форм отариид и хотя бы в отношении лишь набора кормов, не
позволяет наличный материал. В то же время попытки разграничить видовые стратегии
сбора корма, найти видовую специфику энергетики и рассчитать потенциальные
ареалы отдельных форм ушастых тюленей представляются вполне возможными и уместными
при сегодняшнем уровне исследований. Ведь даже реализуемые сейчас, после нескольких
видов нерегламентированного промысла, ареалы отариид содержат свидетельства
степени их совместимости для отдельных форм, в одних комбинациях дополняя один
другой, в других — свободно перекрываясь.
То есть и вычленение дискретных экологических ниш возможно уже на сегодняшнем
уровне исследований при некоторой координации рассредоточенных по глобусу исследователей
и разработке методики сбора рассеянной по архивам, музеям и библиотекам информации
о вчерашнем и позавчерашнем расселении исследуемых зверей.
Дискретность населения по потенциальной способности к скрещиванию и оставлению
плодовитого потомства, без учета которой мы еще долго будем описывать под разными
названиями популяции одного вида, разобщенные общим сокращением численности
или гибелью промежуточных популяций, как будто доступнее для наблюдения, чем
морфологическая дискретность и тем более дискретность экологических ниш.
Можно попробовать разные варианты скрещиваний в неволе (сейчас ушастые тюлени
плодятся во многих зоопарках и океанариумах), но нет заинтересованности у владельцев
приплода. Так что даже судьбу случайно полученных гибридов калифорнийского
морского льва и южноафриканского морского кота (т. е. межродовых гибридов,
редких среди млекопитающих) проследить не удалось. Дожили ли они до повзросления,
были плодовиты или стерильны? Еще доступнее и проще по методике посмотреть
совместимость гамет разных отариид в культуре, тем более, что благодаря успехам
скотоводства есть надежные способы длительного хранения и транспортировки яйцеклеток
и спермиев. Но и здесь проблема упирается в необходимость сосредоточить материал
с громадной акватории в одних заинтересованных и технически оснащенных руках.
Наконец, анализ сходства сывороточных белков крови помог бы оценить дистанции
между популяциями одного или предположительно разных форм ушастых тюленей.
Но снова иммунологи не имеют еще интереса к таксономическим проблемам и вполне
удовлетворяются обнаружением сходства медвежьей и тюленьей сывороток, а зоологи
не владеют методикой и обычно имеют доступ к материалу лишь из одного региона.
То есть потенциальная репродуктивная дискретность может быть оценена с помощью
разных методик при наличии заинтересованных учреждений и решении организационных
проблем. Пока же эту изоляцию показывает лишь свободное перекрывание ареалов,
сезонов размножения и лежбищных структур отдельных форм ушастых тюленей.
Как же быть сейчас, пока эти проблемы еще не решены, с разбивкой четко очерченного
семейства ушастых тюленей на отдельные аргументированно выделенные виды, обособленные
друг от друга репродуктивно, конструктивно и по месту в биосфере? Можно попытаться
подытожить то положительное, что уже накоплено.
Не вызывающее сомнений в своей самостоятельности семейство членится на биосферном
уровне по крайней мере на три части (калифорниец, морские коты и морские львы).
На уровне репродуктивных изолятов единиц пока меньше (пока есть сомнения в
надежности изоляции, скажем, калифорнийца и южноафриканского кота), чем позволяет
опознать морфология. А на уровне строения особи есть не меньше семи равноудаленных
друг от друга вариантов (две формы котов и пять форм львов) при значительных
неясностях в пределах того, что называется родом Arctocephalus. И так же как
и в случае с моделями филогенетических связей всего семейства, для сегодняшнего
членения содержимого семейства приходится допустить равновероятность нескольких
моделей таксономии. Эта неоднозначность, по-видимому, неизбежна, хотя и представляется
далекой от оптимальности, т. е. и таксономические проблемы, поначалу, казалось
бы, ограниченные лишь одним семейством, при ближайшем рассмотрении оказались
достаточно общими.
| Чем интересен ушастый тюлень ........................................... |
3 |
| Как складывалось знакомство с ушастыми тюленями ....... | 4 |
| Строение тела ушастого тюленя и освоение океана ........... | 14 |
| Составные портреты Фрэнсиса Гальтона и возможность увидеть размах изменчивости ............................................... |
29 |
| Когда появились на сцене ушастые тюлени
................. |
63 |
| Как живет ушастый тюлень ............................................. |
71 |
| Как из отдельных особей строится скопление
.............. |
79 |
| Нормированы ли события индивидуальной жизни ...... | 105 |
| Сколько видов ушастых тюленей живет сейчас .......... | 111 |
| Где живут ушастые тюлени ................................................... |
118 |
| Где место ушастым тюленям в современном мире ............. | 159 |
| Что дает изучение ушастых тюленей .................................... |
163 |
| Литература .............................................................................. | 170 |